Embed Size (px)
Citation preview

ORIGINAL PAPER
Recovery of citrus triploid hybrids by embryo rescue and flowcytometry from 2x 3 2x sexual hybridisation and its applicationto extensive breeding programs
P. Aleza • J. Juarez • J. Cuenca • P. Ollitrault •
Luis Navarro
Received: 22 March 2010 / Revised: 27 May 2010 / Accepted: 14 June 2010 / Published online: 6 July 2010
� Springer-Verlag 2010
Abstract Seedlessness is one of the most important
characteristics for mandarins for the fresh-fruit market and
mandarin triploid hybrids have this trait. Citrus triploid
plants can be recovered by 2x 9 2x sexual hybridisations as
a consequence of the formation of unreduced gametes at
low frequency. Triploid embryos are found in small seeds
that do not germinate under greenhouse conditions.
Extensive breeding programs based on this type of
hybridisation require very effective methodologies for
embryo rescue and ploidy evaluation. In this work, we
describe an effective methodology to recover triploid
hybrids from 2x 9 2x hybridisations based on in vitro
embryo rescue and ploidy level determination by means of
flow cytometry. The influence of parents and environmental
conditions on obtaining triploid hybrids has been analysed.
The strongest effect was associated with the genotype of the
female parent while a strong interaction was found between
the male parent genotype and environmental conditions.
The effect of the female parent genotype on the length of the
juvenile phase was also demonstrated by observing a large
number of progenies over the last 10 years. The method-
ology described here has enabled us to obtain over 4,000
triploid hybrids so far, of which 13 have been protected in
the European Union and two are being extensively planted
by citrus growers to establish new commercial plots. These
triploid hybrids have been analysed with simple sequence
repeats markers to differentiate all the new triploid varieties
and their parents, and thus molecular identification will help
defend plant breeders’ rights.
Keywords Triploid � Embryo rescue � Flow cytometry �2x 9 2x sexual hybridisation � Juvenility � Mandarin
Introduction
Citrus are the most extensively produced fruit-tree crop in
the world (FAO 2009). In the Mediterranean area, citrus
fruits are primarily produced for the fresh-fruit market.
Spain is the main producer in the area with a surface of
330,000 ha and a production about 6.3 million tons.
Seedlessness is one of the most important characteristics
for mandarins sold on the fresh-fruit market as consumers
do not accept seedy fruits. The creation of triploid hybrids
is an important breeding strategy to develop new seedless
citrus commercial varieties (Ollitrault et al. 2008). Cyto-
genetic studies have shown that during meiosis of citrus
triploid hybrids, trivalent, bivalent and univalent associa-
tions are formed (Cameron and Frost 1968) producing
sterile gametes. Moreover, Fatta Del Bosco et al. (1992)
have shown that the abortion of megasporogenesis from the
first divisions of the embryo sac in the fertilised egg cell is
frequent. For this reason, citrus triploid hybrids are gen-
erally sterile, although they can occasionally produce fruits
with very few seeds. Triploid plants are generally consid-
ered as an evolutionary dead-end, since they generally give
rise to aneuploid gametes with very low fertility (Otto and
Communicated by W. Harwood.
P. Aleza � J. Juarez � J. Cuenca � P. Ollitrault � L. Navarro (&)
Centro de Proteccion Vegetal y Biotecnologıa,
Instituto Valenciano de Investigaciones Agrarias (IVIA),
Ctra. Moncada-Naquera km 4.5, 46113 Moncada,
Valencia, Spain
e-mail: [email protected]
P. Ollitrault
UPR amelioration genetique des especes a multiplication
vegetative, Centre de Cooperation Internationale en Recherche
Agronomique pour le Developpement (CIRAD), Avenue
Agropolis TA A-75/02, 34398 Montpellier Cedex 5, France
123
Plant Cell Rep (2010) 29:1023–1034
DOI 10.1007/s00299-010-0888-7

Whitton 2000). However, horticultural methods of grafting
allow the clonal vegetative propagation of triploid citrus.
During the past few years, new triploid genotypes have
been obtained from 2x 9 4x and 4x 9 2x hybridisations in
USA, Japan and Italy (Williams and Roose 2004; Toku-
naga et al. 2005; Reforgiato-Recupero et al. 2008), and
some of these cultivars are now starting to be commercially
propagated.
Citrus triploid hybrids can be recovered by 2x 9 2x
(Cameron and Frost 1968; Esen and Soost 1971a; Geraci
et al. 1975), 2x 9 4x (Esen and Soost 1971b; Oiyama et al.
1981; Starrantino and Recupero 1981) and 4x 9 2x
(Cameron and Burnett 1978; Esen et al. 1978; Aleza et al.
2009) sexual hybridisations. In the case of 2x 9 2x
hybridisation, triploid hybrids arise from the union of a 2n
megagametophyte with haploid pollen (Esen and Soost
1971a, 1973; Geraci et al. 1975). The frequency of such
events is generally low (Cameron and Frost 1968; Esen and
Soost 1971a; Geraci et al. 1975) and extensive breeding
programs based on this type of hybridisation require very
effective methodologies for embryo rescue and ploidy
evaluation of large progenies.
Esen and Soost (1971a) were the first to indicate that
triploid embryos are preferentially found in seeds that are
between 1/3 and 1/6 smaller than normal seeds; moreover,
these small seeds generally do not germinate in conven-
tional greenhouse conditions. The in vitro culture of whole
seeds with their integuments can improve germination
rates, although still at relatively low germination percent-
ages (Ollitrault et al. 1996). In rare cases, triploid hybrids
can be found in conventional greenhouse seedlings, as was
the case for ‘Winola’ mandarin (Vardi et al. 1991) and
‘A-12’ mandarin (Bono, personal communication).
In the past, the implementation of extensive triploid
projects based on 2x 9 2x hybridisation was also ham-
pered by the triploid selection step among the progenies.
Indeed, ploidy level analysis by cytogenetic methods is a
slow and inadequate process when large populations of
plants have to be analysed. By contrast, techniques for
ploidy level analysis by flow cytometry (Ollitrault and
Michaux-Ferriere 1992; Ollitrault et al. 1996; Navarro
et al. 2002a) enabled ploidy level to be determined both
accurately and very rapidly.
In this work, we present the methodological improve-
ments that have enabled us to establish an optimised pro-
cess for extensive triploid citrus breeding from 2x 9 2x
hybridisations, with the aim to produce new high-quality,
late-maturing and seedless triploid mandarins. The differ-
ent types of seeds obtained from 2x 9 2x hybridisations
have been characterised and small seed in vitro culture was
compared with embryo rescue method. We have analysed
the different factors that influence the behaviour of
embryos cultured in vitro, the ploidy level of plants
regenerated from each type of seed and how parents
influence triploid hybrid recovery using this technology.
Our breeding program gave rise to over 4,000 triploid
hybrids, from which 13 varieties have been protected and
fingerprinted by simple sequence repeats (SSR) markers,
thus contributing to the protection of breeders’ rights.
Materials and methods
Plant material
All the genotypes used (Table 1) are included in the Citrus
Germplasm Bank of pathogen-free plants of the Instituto
Valenciano de Investigaciones Agrarias (IVIA, Moncada,
Spain) (Navarro et al. 2002b). All the genotypes used as
female parents are self-incompatible and non-apomictic.
The hybridisations were done over a 10-year period (from
1996 to 2006) and are part of the breeding program carried
Table 1 Genotypes used in 2x 9 2x hybridisations
Scientific name
Female parents
Clementines
‘Clemenules’
‘Fina’
‘Hernandina’
‘Marisol’ Citrus clementina Hort. ex Tan.
‘Fortune’ mandarin C. clementina 9 C. tangerina
‘Moncada’ mandarin C clementina 9 (C. unshiu 9 C. nobilis)
Male parents
Clementines
‘Clemenules’
‘Fina’
‘Hernandina’ C. clementina
‘Chandler’ pummelo C. grandis 9 C. grandis
‘Comuna’ sweet orange C. sinensis (L.) Osb.
‘Fortune’ mandarin C. clementina 9 C. tangerina
‘Kara’ mandarin C. unshiu 9 C. nobilis
‘Moncada’ mandarin C clementina 9 (C. unshiu 9 C. nobilis)
‘Nova’ mandarin C. clementina 9 (C. paradisi 9
C. tangerina)
‘Willow leaf’ mandarin C. deliciosa Ten.
‘Chandler’ pummelo C. grandis 9 C. grandis
‘Pink’ pummelo Citrus grandis (L.) Osb.
Tangors
‘Ellendale’
‘Murcott’
‘Nadorcott’
‘Ortanique’ C. reticulata 9 C. sinensis
1024 Plant Cell Rep (2010) 29:1023–1034
123

out since 1995 (Navarro et al. 2005). In addition, we made
a hybridisation between ‘Clemenules’ clementine (Citrus
clementina Hort. ex Tan.) female parent with ‘Pink’
pummelo (C. grandis (L.) Osb.) male parent to describe
and characterise seeds obtained in 2x 9 2x hybridisations.
Pollination, seed extraction and characterisation
Pollinations were carried out in trees grown in a large
screenhouse and in the field. Anthers of the male parents
were removed from flowers collected in pre-anthesis and
dried in Petri dishes over silica gel in a desiccator. Dried
dehisced anthers were stored in small Petri dishes at
-20�C. Flowers were hand-pollinated.
Fruits were collected when ripe and seeds were extracted
and surface sterilised with a sodium hypochlorite solution
(0.5% active chlorine for 10 min). Seeds were classified by
size and developmental stage. Size was evaluated by mea-
suring the area (mm2) and developmental stage was eval-
uated by morphological parameters. Seeds were considered
developed when they had a normal appearance, totally filled
out, and without any malformation. Seeds were considered
undeveloped when had incomplete development, not totally
filled out, wrinkled and with split outer integument (Fig. 1).
Seeds were washed, dried and uniformly distributed in
a 9-cm Petri dishes, making sure that seeds did not
touch each other. Petri dishes were scanned at 150 pp in an
Epson Perfection 4870 Photo scanner. Images were ana-
lysed with the Matrox software, which gives automatic
exact measurements.
Embryo rescue and plant recovery
In experiments to determine the influence of integuments
on germination, intact small seeds and embryos isolated
with the aid of a stereoscopic microscope from small seeds
were cultured under aseptic conditions in Petri dishes
containing the Murashige and Skoog (1962) culture media
with 50 g/L sucrose, 500 mg/L malt extract and supple-
mented with vitamins (100 mg/L myo-inositol, 1 mg/L
pyridoxine hydrochloride, 1 mg/L nicotinic acid, 0.2 mg/L
thiamine hydrochloride, 4 mg/L glycine) and 8 g/L Bacto
agar (MS culture media). After germination, plants were
transferred to 25 9 150 mm test tubes with the same cul-
ture media without malt extract. Cultures were maintained
at 24 ± 1�C, 60% humidity and 16 h daily exposure to
40 lE m-2 s-1 illumination.
Ploidy level analysis
Ploidy level was determined by flow cytometry according
to the methodology described by Aleza et al. (2009). Each
sample consisted of a small piece of leaf (*0.5 mm2)
collected from each test-tube plant with a similar leaf piece
taken from a diploid control plant. Samples were chopped
together using a razor blade in the presence of a nuclei
isolation solution (High Resolution DNA Kit Type P,
solution A; Partec�, Munster, Germany). Nuclei were fil-
tered through a 30-lm nylon filter and stained with a DAPI
(4,6-diamine-2-phenylindol) (High Resolution DNA Kit
Type P, solution B; Partec�) solution. Following a 5-min
incubation period, stained samples were run in a Ploidy
Analyzer (Partec�, PA) flow cytometer equipped with a
HBO 100-W high-pressure mercury bulb and both KG1
and BG38 filter sets. Histograms were analysed using the
dpac v2.0 software (Partec�), which determines peak
position, coefficient of variation (CV) and the relative
ploidy index of the samples.
Transfer to soil
Triploid plants were transferred to pots containing steam-
sterilised artificial soil mix suitable to grow citrus (40%
black peat, 29% coconut fibre, 24% washed sand and 7%
perlite). Composition was developed in our group to grow
citrus in the greenhouse. Pots were enclosed in polyethyl-
ene bags that were closed with rubber bands and placed in a
shaded area in a temperature-controlled greenhouse set at
18–25�C. After 8–10 days, the bags were opened, and after
another 8–10 days, the bags were removed and the plants
grown under regular greenhouse conditions (Navarro and
Juarez 2007).
Field evaluation
Triploid plants were cultured in the greenhouse for
approximately 1 year to produce quality budwood to graft
in the field. Buds were grafted onto ‘Carrizo’ citrange
Fig. 1 Different types of seeds obtained in ‘Clemenules’ clementine
by ‘Pink’ pummelo hybridisation. a Developed small seeds,
b undeveloped seeds, and c developed seeds (normal seeds)
Plant Cell Rep (2010) 29:1023–1034 1025
123

rootstock (C. sinensis 9 P. trifoliata) for field evaluation at
IVIA plots. Flowering was recorded for each hybrid and
when they started fruiting, a sample of ten mature fruits
was collected and data were taken on diameter, weight,
fruit shape, rind adherence to flesh, pulp texture, flavour of
the juice and a general final evaluation. Triploid hybrids
selected during the first evaluation were later described
following the guidelines to conduct tests for distinctness,
uniformity and stability for Citrus L. Group 1 mandarins,
from the International Union for the Protection of New
Varieties of Plants (UPOV 2009).
Molecular characterisation
Thirteen triploid plants obtained by 2x 9 2x hybridisations
were selected for their high quality (Table 2). These trip-
loid plants and their parents were analysed with 13 SSR
markers (Kijas et al. 1997; Froelicher et al. 2008; Luro
et al. 2008) heterozygotic for clementine and ‘Fortune’
mandarin. The extraction of genomic DNA was done
according to Dellaporta and Hicks (1983) with slight
modifications. After a M13 tailed PCR reaction (Schuelke
2000), genetic analysis was performed in a capillary-array
sequencer CEQTM 800 System (Beckamn Coulter� Inc.,
Fullerton, CA) and results were analysed with Genome-
LabTM GeXP Genetic Analysis System software. A cluster
analysis was done with Darwin V.5.0.155 program (Perrier
et al. 2003; Perrier and Jacquemoud-Collet 2006) accord-
ing to the Weighted Neighbour-Joining method, using Dice
dissimilarity coefficient.
Results
Seed description and ploidy level of recovered plantlets
Two hundred and ninety-nine seeds from 40 fruits were
obtained from the pollination between ‘Clemenules’
clementine and ‘Pink’ pummelo. From 299 seeds, 252 were
developed (Fig. 1a, c) and 47 were undeveloped (Fig. 1b).
In developed seeds, we clearly identified two different
groups, 12 small seeds (Fig. 1a) and 240 normal seeds
(Fig. 1c).
Average area of normal seeds was 78 ± 11 mm2,
whereas area of small seeds was 34 ± 8 mm2, between 52
and 62% smaller than normal seeds. Average area of
undeveloped seeds was 56 ± 17 mm2, 18–42% smaller
than normal seeds (Table 3).
Normal seeds were germinated in a greenhouse and all
regenerated plantlets were diploid. Embryos were rescued
from the 12 small seeds, and cultured in vitro. Twelve
plantlets were obtained, of which three were diploid and
nine triploid. Only 13 of the 47 undeveloped seeds con-
tained embryos (27.7%) and only five plantlets, all diploid,
were recovered after embryo culture (Table 3). Embryos
contained in small seeds were well developed, with two
cotyledons and a well-formed embryonary axis (Fig. 2a).
This characteristic was observed systematically in all 44
hybridisations performed in the breeding program, and
triploid plants were recovered exclusively from embryos
rescued from the small developed seeds.
Recovery of citrus triploid plants
Comparison between in vitro germination of intact seeds
and embryo rescue
Small seeds obtained in fruits of open pollinated ‘Fortune’
mandarin were cultured in vitro with and without tegu-
ments. The germination percentage of intact small seeds
with teguments was 42.4% (25/59) slightly higher than the
33% obtained by Ollitrault et al. (1996) for small clem-
entine seeds, whereas germination of isolated embryos was
84% (42/50). Henceforth, in vitro embryo rescue was used
systematically in the breeding program.
Fruit set, plant regeneration, ploidy level
and transfer to soil
Results of embryo rescue, plant regeneration, ploidy level
and transfer to soil of plants recovered in 2x 9 2x
hybridisations are shown in Table 4. The data correspond
to more than 8,800 pollinations done between 1996 and
2006.
Table 2 Selected triploid hybrids obtained in 2x 9 2x hybridisations
and its parents
Female parent Male parent Triploid hybrids
‘Fortune’
mandarin
‘Murcott’ tangor ForMur 96-019
ForMur 96-023
ForMur 96-081
(Garbı’ mandarin)
ForMur 96-086
‘Ellendale’ tangor ForEll 96-023
ForEll 96-058
ForEll 96-060
Open pollination For? 95-087
For? 95-097
‘Kara’ mandarin ForKar 96-037
(‘Safor’ mandarin)
ForKar 96-073
‘Willow leaf’ mandarin ForMco 96-082
‘Willits Newcom’
clementine
Open pollination Cwn? 95-001
1026 Plant Cell Rep (2010) 29:1023–1034
123

Fruit set average was 59%. Average number of small
seeds per fruit varied between 0.23 with ‘Fina’ clementine
and 3.0 with ‘Fortune’ mandarin. In total, 4,400 small
seeds were obtained and 4,110 (93%) of them contained
only one embryo per seed (Fig. 2a), whereas the other
seeds were aborted. From the 4,110 embryos cultured in
vitro, 3,816 plants were recovered. Average germination
percentage was 93% using the embryo rescue technique,
fluctuating between 83% for ‘Marisol’ clementine and 98%
for ‘Fina’ clementine (Fig. 2b, c).
The average number of plants obtained from small seeds
per fruit was 0.22 for ‘Fina’ clementine and ‘Moncada’
mandarin, whereas for ‘Fortune’ mandarin it was 2.6 plants
per fruit.
From the 3,816 plants obtained, 315 were diploid (8.3%)
and 3,497 were triploid (91.6%). Also four pentaploid
plants were obtained, two from ‘Clemenules’ clementine
and two from ‘Fortune’ mandarin female parents.
The average survival percentage in the transplant phase
was as high as 90% and the plants obtained were generally
robust and vigorous (Fig. 2e).
Factors affecting recovery efficiency of citrus
triploid hybrids
Female parent
Recovery efficiency in citrus triploid plants was calculated
as the number of triploid plants per harvested fruit. Triploid
hybrids were obtained most efficiently with ‘Fortune’
mandarin (Table 4), with an average of 2.54 triploid
hybrids per fruit. The efficiency in clementines varied
between 0.32 with ‘Hernandina’ and 0.17 with ‘Fina’.
‘Moncada’ mandarin gave a low efficiency, with 0.12
triploid hybrids per fruit. The efficiency of ‘Fortune’
mandarin was eight times higher and more than 20 times
Table 3 Type and size of seeds
obtained in ‘Clemenules’
clementine and ‘Pink’ pummelo
hybridisation and number and
ploidy level of plants recovered
Type of seeds No. of seeds Average area (mm2) Obtained plants Ploidy level
Diploid Triploid
Developed seeds 96 78 ± 11 96 96 0
Developed small seeds 12 34 ± 8 12 3 9
Undeveloped seeds 47 56 ± 17 5 5 0
Fig. 2 a Small seed of
‘Fortune’ mandarin with one
embryo. b, c Germinated
embryos of ‘Fortune’ mandarin
rescued from small seeds.
d In vitro triploid plant obtained
from a hybridisation between
‘Fortune’ mandarin and
‘Hernandina’ clementine.
e Triploid plants obtained from
2x 9 2x hybridisations
transplanted and cultivated in
the greenhouse
Plant Cell Rep (2010) 29:1023–1034 1027
123
higher than ‘Hernandina’ clementine and ‘Moncada’
mandarin, respectively.
Male parent and environmental conditions
The triploid production efficiency was estimated for ‘For-
tune’ mandarin and three clementine parents crossed with
four male parents over 8 years (Table 5). Among the
clementine varieties and ‘Fortune’ mandarin, differences
were observed concerning the male parents. ‘Kara’ man-
darin male parent gave the lowest number of triploid
hybrids per fruit with all female parents, whereas the
highest number of triploid hybrids per fruit was obtained
with ‘Willow leaf’ mandarin and ‘Comuna’ sweet orange
male parents for all female parents. Inter-annual variances
have been estimated for the parental combinations for
which data of at least 3 years were available. This variance
is important and masks a potential male effect for the
crosses with clementine varieties. For example, in the
hybridisation between ‘Clemenules’ clementine and ‘Kara’
mandarin, inter-annual variance was ±0.02, whereas in the
hybridisation between ‘Hernandina’ clementine and ‘Kara’
mandarin was ±0.18 triploid hybrids per fruit.
To obtain a better estimation of the environmental effect
and its interaction with the male parent, we evaluated the
number of triploid plants recovered per fruit of all the
crosses done with ‘Fortune’ mandarin over 5 years
(Fig. 3). A strong variability was observed between years
for the same parental combinations. For example, in the
cross between ‘Fortune’ mandarin and ‘Willow leaf’
mandarin, the variation in the number of triploid hybrids
per fruit varied from 1.5 in 1996 to 6.6 in 1998. Variability
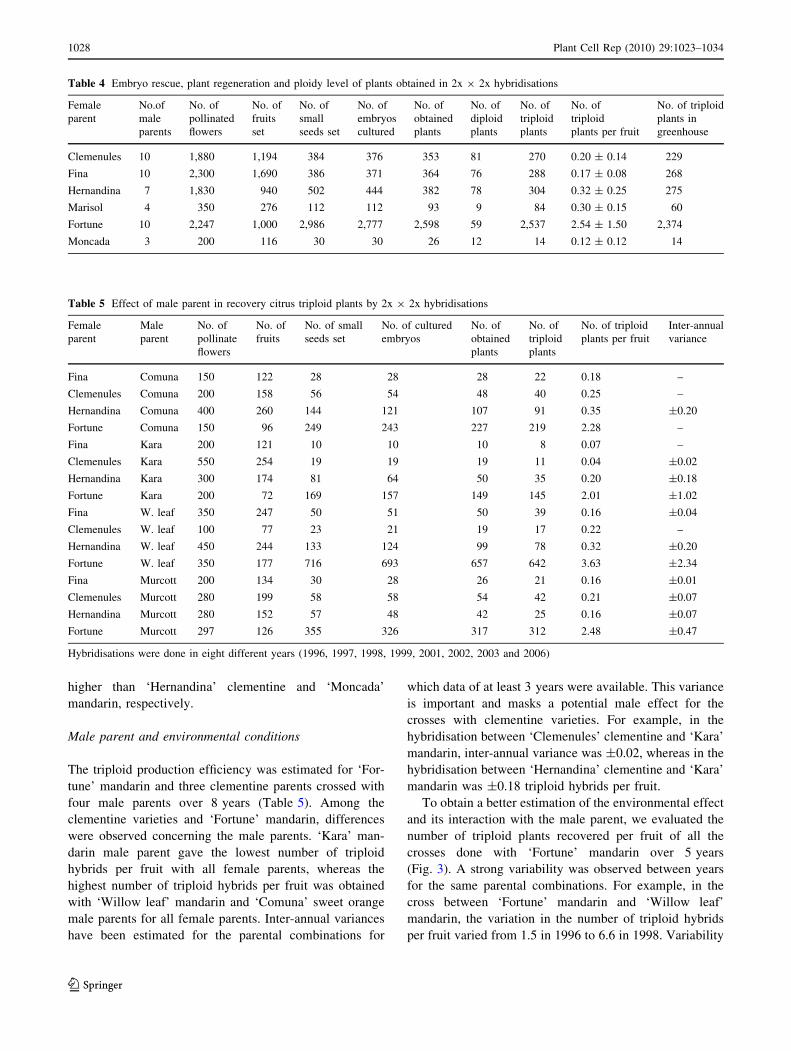
Table 4 Embryo rescue, plant regeneration and ploidy level of plants obtained in 2x 9 2x hybridisations
Female
parent
No.of
male
parents
No. of
pollinated
flowers
No. of
fruits
set
No. of
small
seeds set
No. of
embryos
cultured
No. of
obtained
plants
No. of
diploid
plants
No. of
triploid
plants
No. of
triploid
plants per fruit
No. of triploid
plants in
greenhouse
Clemenules 10 1,880 1,194 384 376 353 81 270 0.20 ± 0.14 229
Fina 10 2,300 1,690 386 371 364 76 288 0.17 ± 0.08 268
Hernandina 7 1,830 940 502 444 382 78 304 0.32 ± 0.25 275
Marisol 4 350 276 112 112 93 9 84 0.30 ± 0.15 60
Fortune 10 2,247 1,000 2,986 2,777 2,598 59 2,537 2.54 ± 1.50 2,374
Moncada 3 200 116 30 30 26 12 14 0.12 ± 0.12 14
Table 5 Effect of male parent in recovery citrus triploid plants by 2x 9 2x hybridisations
Female
parent
Male
parent
No. of
pollinate
flowers
No. of
fruits
No. of small
seeds set
No. of cultured
embryos
No. of
obtained
plants
No. of
triploid
plants
No. of triploid
plants per fruit
Inter-annual
variance
Fina Comuna 150 122 28 28 28 22 0.18 –
Clemenules Comuna 200 158 56 54 48 40 0.25 –
Hernandina Comuna 400 260 144 121 107 91 0.35 ±0.20
Fortune Comuna 150 96 249 243 227 219 2.28 –
Fina Kara 200 121 10 10 10 8 0.07 –
Clemenules Kara 550 254 19 19 19 11 0.04 ±0.02
Hernandina Kara 300 174 81 64 50 35 0.20 ±0.18
Fortune Kara 200 72 169 157 149 145 2.01 ±1.02
Fina W. leaf 350 247 50 51 50 39 0.16 ±0.04
Clemenules W. leaf 100 77 23 21 19 17 0.22 –
Hernandina W. leaf 450 244 133 124 99 78 0.32 ±0.20
Fortune W. leaf 350 177 716 693 657 642 3.63 ±2.34
Fina Murcott 200 134 30 28 26 21 0.16 ±0.01
Clemenules Murcott 280 199 58 58 54 42 0.21 ±0.07
Hernandina Murcott 280 152 57 48 42 25 0.16 ±0.07
Fortune Murcott 297 126 355 326 317 312 2.48 ±0.47
Hybridisations were done in eight different years (1996, 1997, 1998, 1999, 2001, 2002, 2003 and 2006)
1028 Plant Cell Rep (2010) 29:1023–1034
123

was also observed between hybridisations performed in a
same year with different male parents. For example, for the
1996 hybridisations, the number of triploid hybrids per
fruit oscillated between 1.4 and 3.3 triploids per fruit for
hybridisations with ‘Comuna’ sweet orange and ‘Ellendale’
tangor, respectively. Moreover, the rank of the different
male parents was not conserved between years which
would indicate an important interaction between the geno-
type of the male parent and the environmental conditions.
Selection of citrus triploid hybrids
From over 4,000 triploid hybrids obtained in the IVIA
triploid program based on 2x 9 2x hybridisations (Navarro
et al. 2005), 3,600 triploid hybrids have already flowered
and 1,998 have been evaluated. In this work, we have
analysed more than 730 triploid hybrids arising from
hybridisations between ‘Fortune’ mandarin as female parent
and ‘Willow leaf’ mandarin, ‘Murcott’ and ‘Ellendale’
tangors and ‘Comuna’ sweet orange as male parents as well
as hybridisations between ‘Clemenules’ and ‘Hernandina’
clementines with ‘Comuna’ sweet orange.
All the triploid hybrids displayed juvenile characteris-
tics. Triploid hybrids transplanted to pots in the greenhouse
took approximately 1 year to produce quality budwood that
could be grafted in the field. One year after field grafting,
none produced flowers, after 2 years 17% flowered, after
3 years 51% flowered and after 5 years 81% of the triploid
hybrids flowered (Fig. 4). Furthermore, clear differences were
observed between the female parents (Fig. 5). After 3 years of
grafting, more than 50% of the triploid hybrids obtained with
‘Fortune’ mandarin had already flowered, whereas only
25–30% of the triploid hybrids from clementines had done so.
This advantage of ‘Fortune’ mandarin was observed in the
following years, and after 7 years flowering percentages were,
respectively, 90 and 65% for ‘Fortune’ mandarin and clem-
entines. Moreover, the observations for the progenies obtained
from hybridisation between ‘Fortune’ mandarin and four male
parents suggest that a slight effect was exerted by the male
parent on the length of the juvenile phase. ‘Willow leaf’
mandarin seems to promote earlier flowering, while ‘Murcott’
tangor progenies appear to flower the latest.
From the 1,998 triploid hybrids evaluated for fruit
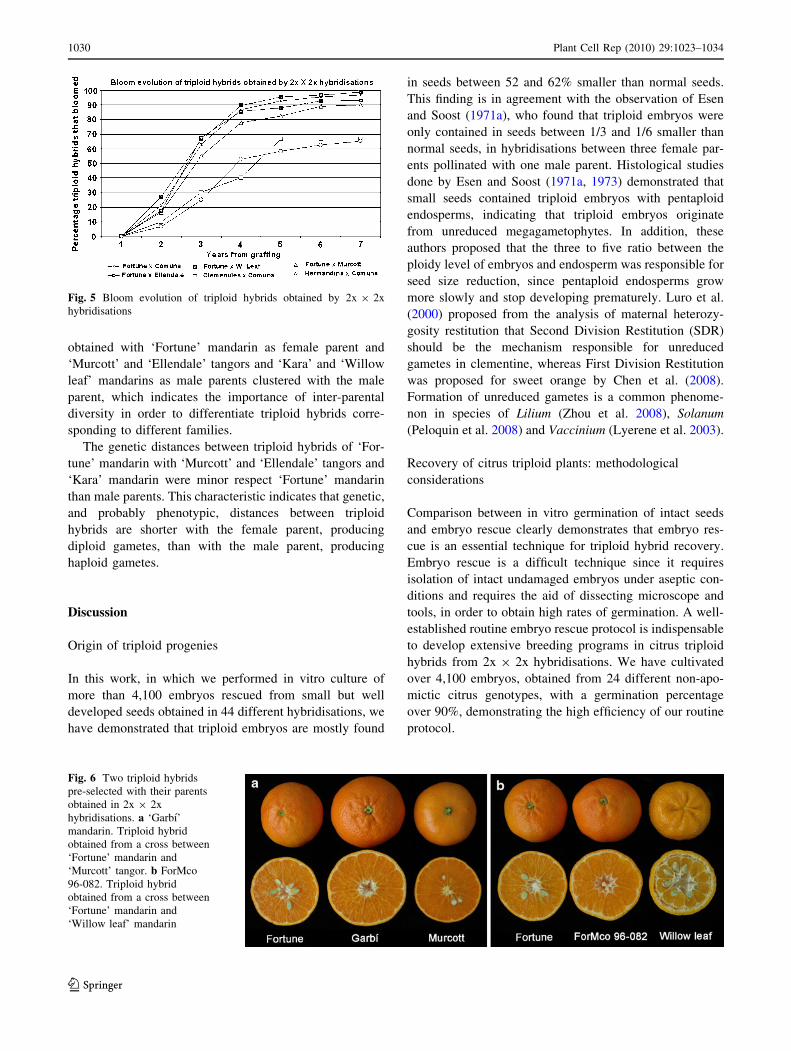
quality, 99 were pre-selected and 13 (Table 2; Fig. 6) were
finally selected for their high quality and excellent flavour
and breeder’s rights have been protected.
Genetic analysis
Genetic analysis with the SSR markers differentiated all
protected triploid hybrids and their parents, and confirmed
the origin of all hybrids analysed (Fig. 7). Triploid hybrids
Fig. 3 Triploid hybrids
recovered by fruit in 5 years
with ‘Fortune’ mandarin as
female parent and ten different
male parents
Fig. 4 a Triploid hybrid of
‘Fortune’ mandarin 9 ‘Willow
leaf’ mandarin that flowered
after 4 years grafted in the field.
b Triploid hybrid of ‘Fortune’
mandarin 9 ‘Willow leaf’
mandarin that not produced
flowers and c Control ‘Fortune’
mandarin at the same flowering
time
Plant Cell Rep (2010) 29:1023–1034 1029
123
obtained with ‘Fortune’ mandarin as female parent and
‘Murcott’ and ‘Ellendale’ tangors and ‘Kara’ and ‘Willow
leaf’ mandarins as male parents clustered with the male
parent, which indicates the importance of inter-parental
diversity in order to differentiate triploid hybrids corre-
sponding to different families.
The genetic distances between triploid hybrids of ‘For-
tune’ mandarin with ‘Murcott’ and ‘Ellendale’ tangors and
‘Kara’ mandarin were minor respect ‘Fortune’ mandarin
than male parents. This characteristic indicates that genetic,
and probably phenotypic, distances between triploid
hybrids are shorter with the female parent, producing
diploid gametes, than with the male parent, producing
haploid gametes.
Discussion
Origin of triploid progenies
In this work, in which we performed in vitro culture of
more than 4,100 embryos rescued from small but well
developed seeds obtained in 44 different hybridisations, we
have demonstrated that triploid embryos are mostly found
in seeds between 52 and 62% smaller than normal seeds.
This finding is in agreement with the observation of Esen
and Soost (1971a), who found that triploid embryos were
only contained in seeds between 1/3 and 1/6 smaller than
normal seeds, in hybridisations between three female par-
ents pollinated with one male parent. Histological studies
done by Esen and Soost (1971a, 1973) demonstrated that
small seeds contained triploid embryos with pentaploid
endosperms, indicating that triploid embryos originate
from unreduced megagametophytes. In addition, these
authors proposed that the three to five ratio between the
ploidy level of embryos and endosperm was responsible for
seed size reduction, since pentaploid endosperms grow
more slowly and stop developing prematurely. Luro et al.
(2000) proposed from the analysis of maternal heterozy-
gosity restitution that Second Division Restitution (SDR)
should be the mechanism responsible for unreduced
gametes in clementine, whereas First Division Restitution
was proposed for sweet orange by Chen et al. (2008).
Formation of unreduced gametes is a common phenome-
non in species of Lilium (Zhou et al. 2008), Solanum
(Peloquin et al. 2008) and Vaccinium (Lyerene et al. 2003).
Recovery of citrus triploid plants: methodological
considerations
Comparison between in vitro germination of intact seeds
and embryo rescue clearly demonstrates that embryo res-
cue is an essential technique for triploid hybrid recovery.
Embryo rescue is a difficult technique since it requires
isolation of intact undamaged embryos under aseptic con-
ditions and requires the aid of dissecting microscope and
tools, in order to obtain high rates of germination. A well-
established routine embryo rescue protocol is indispensable
to develop extensive breeding programs in citrus triploid
hybrids from 2x 9 2x hybridisations. We have cultivated
over 4,100 embryos, obtained from 24 different non-apo-
mictic citrus genotypes, with a germination percentage
over 90%, demonstrating the high efficiency of our routine
protocol.
Fig. 5 Bloom evolution of triploid hybrids obtained by 2x 9 2x
hybridisations
Fig. 6 Two triploid hybrids
pre-selected with their parents
obtained in 2x 9 2x
hybridisations. a ‘Garbı’
mandarin. Triploid hybrid
obtained from a cross between
‘Fortune’ mandarin and
‘Murcott’ tangor. b ForMco
96-082. Triploid hybrid
obtained from a cross between
‘Fortune’ mandarin and
‘Willow leaf’ mandarin
1030 Plant Cell Rep (2010) 29:1023–1034
123

Obtained plants had diploid (8.3%), triploid (91.6%) and
very occasionally pentaploid ploidy levels (0.1%). Oiyama
and Kobayashi (1991) also obtained some pentaploid plants
from small seeds in 2x 9 2x hybridisations and indicated that
they might have arisen from the union of double unreduced
female gametes with normal reduced male gametes.
Survival percentage of transplanted triploid plants pro-
duced in 2x 9 2x hybridisations was higher than 90%
following our methodology. It is essential to master this
acclimatisation phase for the implementation of large
triploid breeding programs.
Recovery of citrus triploid hybrids; biological
and environmental considerations
The results obtained in this work clearly demonstrate that
the main factor determining the frequency of recovery
citrus triploid hybrids is the genotype of the female parent.
In this context, ‘Fortune’ mandarin displayed higher fre-
quency of triploid hybrids per fruit than clementines and
‘Moncada’ mandarin. It confirms the results of Geraci et al.
(1975) who observed average values of triploid hybrids per
fruit between six and zero from nine non-apomictic citrus
genotypes pollinated with the same male parent. Frequency
of unreduced gametes is an intrinsic characteristic of each
genotype. In peach cultivars, frequency can range from 0 to
50% (Dermen 1938) and in potato, it is under genetic
control (Mok and Peloquin 1975). The frequency of unre-
duced gametes can be quantified by screening genotypes
before starting large-scale breeding programs with 2x 9 2x
hybridisations. At a practical level, this characteristic
implies that a different number of pollinations must be
performed for each genotype to recover a progeny of a
given number. For example, to obtain 100 triploid hybrids
in the greenhouse, more than 650 pollinations would have
to be performed with ‘Hernandina’ clementine, whereas
Fig. 7 Dendogram obtained by
Dice distance and tree
constructed by Weighted
Neighbour-Joining method
using 13 SSR markers
corresponding to registered
triploid plants obtained by
2x 9 2x hybridisations and
their parents
Plant Cell Rep (2010) 29:1023–1034 1031
123

with ‘Fortune’ mandarin the pollination of just 96 flowers
would be enough.
The use of in vitro embryo rescue and ploidy level
assessment by flow cytometry gave highly efficient
recovery of citrus triploid hybrids from small seeds and
enabled low unreduced gamete frequencies, such as ‘Fina’
clementine and ‘Moncada’ mandarin (0.17 and 0.12 trip-
loid hybrids per fruit, respectively), to be exploited for the
development of triploid breeding programs.
Geraci et al. (1975) observed that the production of
diploid female gametes was significantly affected by the
male parent. However, our observations over 10 years of
hybridisation suggest an important interaction between
male parent and environmental conditions on triploid pro-
duction. Luro et al. (2004) also showed that environmental
conditions dramatically affect the frequency of recovered
triploids. It could be hypothesised that for the same number
of 2x gametophytes potentially available, the production of
triploids will be influenced by the ability of the pollen to
perform successful fecundation. This ability is dependent
on (1) the quality of pollen determined by the male geno-
type, but also environmental conditions; (2) the compati-
bility level between female and male parents; and (3)
environmental effects on pollen tube germination. Studies
in a few species have shown that environmental conditions
affect pollen development (Young and Stanton 1990;
Johannsson and Stephenson 1998), as well as pollen tube
growth (Stephenson et al. 1992). One of the most important
environmental factors that could affect pollen performance
is the temperature during the pregamic phase. Indeed,
temperature has been shown to affect pollen germination
(Elgersma et al. 1989; Shivanna et al. 1991), and pollen
tube kinetics in the style (Lewis 1942; Jefferies et al. 1982;
Elgersma et al. 1989). Bono et al. (2006) demonstrated
that, within the mandarin group, there are great differences
in the number of seeds per fruit depending on the parents
used for the crosses. For example, the average number of
seeds per fruit in ‘Clemenules’ clementine was around two
when pollinated with ‘Ortanique’ tangor, and 30 with
‘Moncada’ mandarin. These observations could be related
with the gametophytic incompatibility system identified in
citrus (Soost 1965, 1969).
Juvenile characteristics of triploid progenies
Juvenile traits in citrus plants originating from seeds are
very prominent and often persist for a long time. They
include thorniness, vigorous growth, alternate bearing in
early years, physical differences in fruit characters and
slowness to flower and to bear fruit (Cameron and Frost
1968). The long juvenile period of citrus trees obtained
from seeds is the major impediment for citrus breeding
programs based on sexual hybridisations.
In this work, we demonstrate for the first time the effect
of the female and male parents on the length of the juvenile
period in progenies. This characteristic is of great interest,
since the triploid hybrids can be evaluated more quickly,
allowing extensive triploid breeding programs based on the
2x 9 2x strategy, since it determines the number of years
required to produce new varieties. In this context, it is very
important to evaluate the juvenile period, as well as the
frequency of unreduced gametes of the genotypes included
in Germplasm Banks. This information will allow for a
much better selection of parents for triploid breeding
programs.
Selection of citrus triploid hybrids
We have protected breeder’s rights in Spain for 13 new
triploid varieties for their high fruit quality. Two of them,
‘Garbı’ mandarin (Aleza et al. 2010) and ‘Safor’ mandarin
(Cuenca et al. 2010), have also been protected in the
European Union (EU), Morocco, Egypt, South Africa and
Turkey; moreover, a US Plant Patent has been requested.
Pathogen-free plants of ‘Garbı’ and ‘Safor’ mandarins have
been obtained by shoot-tip grafting in vitro, according to
the methodology described by Navarro et al. (1975) and
healthy budwood has been released to 22 Spanish nurser-
ies, which have signed propagation agreements and are
already producing certified plants. These are the first
varieties recovered in genetic breeding programs from the
2x 9 2x controlled hybridisation strategy.
Complete differentiation between the 13 new triploid
varieties was obtained using 13 SSR markers. The effi-
ciency of such markers for hybrid identification in sexual
diploid progeny had been demonstrated previously.
Molecular identification should contribute to defending
plant breeders’ rights, preventing possible fraud, and pro-
vides traceability from the field to sale points. In addition,
it is a very useful tool in certification programs as illegal
propagations can be detected and possible mistakes or
confusions avoided in nursery identification.
Nowadays, SSR markers are being used as a legally
recognised technique in the defence of breeders’ rights.
Furthermore, our group has performed more in-depth and
detailed genetic studies that can analyse the genetic and
phenotypic structures of the triploid hybrids obtained from
2x 9 2x, 2x 9 4x and 4x 9 2x hybridisations.
Conclusions
This work explores the different factors influencing the
recovery of citrus triploid hybrids from 2x 9 2x hybridi-
sations. Triploid hybrids are found in developed seeds that
are between 52 and 62% smaller than normal seeds.
1032 Plant Cell Rep (2010) 29:1023–1034
123

Embryo rescue and flow cytometry are two indispensable
techniques for extensive citrus triploid breeding programs
based on 2x 9 2x hybridisation. The frequency of unre-
duced megagametophyte production depends on the
maternal genotype, and should be known before starting
extensive breeding programs. ‘Fortune’ mandarin displays
a higher frequency of this phenomenon than clementines
and ‘Moncada’ mandarin. An unclear male parent effect
has been observed in triploid production hybrids, as well
as effects of environmental conditions. In this work, we
have demonstrated for the first time the effect of the
female and male parents on the length of the juvenile
period of the progenies. At IVIA, an extensive breeding
program of citrus triploid hybrids based on such hybrid-
isation has been running since 1995. To date, we have
obtained over 4,000 hybrids and 13 new triploid varieties
have been protected; furthermore, triploid ‘Garbı’ and
‘Safor’ mandarins, obtained in this program, are being
planted extensively by citrus growers to establish new
commercial plots.
Acknowledgments This work is jointly financed by the AGL2008-
00596-MCI and Prometeo 2008/121 Generalidad Valenciana projects.
We thank C. Ortega, A. Navarro, V. Ortega, C. Martı, J.M. Arregui
and M. Hernandez for their technical assistance in the laboratory and
J.A Pina, V. Lloris, J.M. Conchilla, F. Ahuir, D Conchilla, A. Con-
chilla, R. Lopez and F.J. Martı for growing plants in the greenhouse
and in the field.
References
Aleza P, Juarez J, Ollitrault P, Navarro L (2009) Production of
tetraploid plants of non apomictic citrus genotypes. Plant Cell
Rep. doi:10.1007/s00299-009-0783-2
Aleza P, Cuenca J, Juarez J, Pina JA, Navarro L (2010) ‘Garbı’
mandarin: a new late-maturing triploid hybrid. HortScience
45(1):139–141
Bono R, Soler J, Buj A, Villalba D, Salvia J, Bellver R, Mares M,
Mendez JV, Gavila L (2006) El problema de la presencia de
semillas en los frutos cıtricos. Conselleria de Agricultura Pesca y
Alimentacion, Ficha Tecnica serie citricultura No. 5
Cameron JW, Burnett RH (1978) Use of sexual tetraploid seed
parents for production of triploid citrus hybrids. HortScience
13:167–169
Cameron JW, Frost HB (1968) Genetic, breeding and nucellar
embryony. In: Reuther W, Batchelor LD, Webber HJ (eds)
The citrus industry, vol 1. University of California, Riverside,
pp 325–370
Chen C, Lyon MT, O’Malley D, Federici CT, Gmitter J, Grosser JW,
Chaparro JX, Roose ML, Gmitter FG (2008) Origin and
frequency of 2n gametes in Citrus sinensis 9 Poncirus trifoliataand their reciprocal crosses. Plant Sci 174:1–8
Cuenca J, Aleza P, Juarez J, Pina JA, Navarro L (2010) ‘Safor’
mandarin: a new citrus mid-late triploid hybrid. HortScience
45(6) (in press)
Dellaporta J, Hicks JB (1983) A plant DNA minipreparation: version
II. Plant Mol Biol Rep 1:19–21
Dermen H (1938) Detection of polyploidy by grain size: investigation
with peaches and apricots. Proc Am Soc Hortic Sci 35:96–103
Elgersma A, Stephenson AG, Nijs APM (1989) Effects of genotype
and temperature on pollen tube growth in perennial ryegrass
(Lolium perenne L.). Sex Plant Reprod 2:225–230
Esen A, Soost RK (1971a) Unexpected triploids in citrus: their origin,
identification and possible use. J Hered 62:329–333
Esen A, Soost RK (1971b) Tetraploid progenies from 2x–4x crosses
of citrus and their origin. J Am Soc Hortic Sci 97:410–414
Esen A, Soost RK (1973) Precocious development and germination of
spontaneous triploid seeds in citrus. J Hered 64:147–154
Esen A, Soost RK, Geraci G (1978) Seed set, size and development
after 4x–2x and 4x–4x crosses in citrus. Euphytica 27:283–293
FAO, Food and Agriculture Organization (2009) http://faostat.fao.
org/site/567/default.aspx#ancor
Fatta Del Bosco S, Matranga G, Geraci G (1992) Micro and macro-
sporogenesis of two triploid hybrids of citrus. In: Proceedings of
7th international citrus congress, vol 1, International Society of
Citriculture, Acireale, Italy, pp 122–124
Froelicher Y, Dambier D, Bassene JB, Costantino G, Lotfy S, Didout
C, Beaumont V, Brottier P, Risterucci AM, Luro F, Ollitrault P
(2008) Characterization of microsatellite markers in mandarin
orange (Citrus reticulata Blanco). Mol Ecol Resour 8(1):
119–122
Geraci G, Esen A, Soost RK (1975) Triploid progenies from 2x–2x
crosses of citrus cultivars. J Hered 66:177–178
Jefferies CJ, Brain P, Stott KG, Belcher AR (1982) Experimental
systems and mathematical model for studying temperature
effects on pollen-tube growth and fertilization in plum. Plant
Cell Environ 5:231–236
Johannsson MH, Stephenson AG (1998) Effects of temperature
during microsporogenesis on pollen performance in Cucurbitapepo L. (Cucurbitaceae). Int J Plant Sci 159:616–626
Kijas JMH, Thomas MR, Fowler JCS, Roose ML (1997) Integration
of trinucleotide microsatellites into a linkage map of citrus.
Theor Appl Genet 94:701–706
Lewis D (1942) The physiology of incompatibility in plants. I. Effect
of temperature. Proc R Soc London Ser B Biol Sci 131:13–26
Luro F, Maddy F, Ollitrault P, Rist D (2000) Identification of 2ngamete parental origin and mode of nuclear restitution of
spontaneous triploid citrus hybrids. In: Proceedings of 9th
international citrus congress, Orlando, pp 168–169
Luro F, Maddy F, Jacquemond C, Froelicher Y, Morillon R, Rist D,
Ollitrault P (2004) Identification and evaluation of diplogyny in
clementine (Citrus clementina) for use in breeding. In: XI
Eucarpia symposium on fruit breeding and genetics. Acta Hortic
663:841–847
Luro F, Costantino G, Terol JF, Argout X, Allario T, Wincker P,
Talon M, Ollitrault P, Morillon R (2008) Transferability of the
EST-SSRs developed on Nules clementine (Citrus clementinaHort. ex Tan.) to other citrus species and their effectiveness for
genetic mapping. BMC Genom 9(287):1–13
Lyerene PM, Vorsa N, Ballington JR (2003) Polyploidy and sexual
polyploidization in the genus Vaccinium. Euphytica 133:27–36
Mok DWS, Peloquin JS (1975) The inheritance of three mechanisms
of diplandroid (2n pollen) formation in diploid potatoes.
Heredity 35:295–302
Murashige T, Skoog F (1962) A revised medium for rapid growth and
bioassays with tobacco tissue cultures. Physiol Plant 15:473–479
Navarro L, Juarez J (2007) Shoot-tip grafting in vitro. In: Khan IA
(ed) Citrus genetics, breeding and biotechnology. CABI Head
Office, Wallingford, pp 353–364
Navarro L, Roistacher CN, Murashige T (1975) Improvement of
shoot-tip grafting in vitro for virus-free citrus. J Am Soc Hortic
Sci 100:471–479
Navarro L, Juarez J, Aleza P, Pina JA (2002a) Recovery of triploid
seedless mandarin hybrids from 2n 9 2n and 2n 9 4n crosses by
embryo rescue and flow cytometry. In: Proceedings of the 10th
Plant Cell Rep (2010) 29:1023–1034 1033
123

IAPTC&B congress, plant biotechnology 2002 and beyond,
Orlando, FL, pp 541–544
Navarro L, Pina JA, Juarez J, Ballester-Olmos JF, Arregui JM, Ortega
C, Navarro A, Duran-Vila N, Guerri J, Moreno P, Cambra M,
Zaragoza S (2002b) The citrus variety improvement program in
Spain in the period 1975–2001. In: Proceedings of the 15th
conference of the international organization of citrus virologists,
IOCV, Riverside, pp 306–316
Navarro L, Juarez J, Aleza P, Pina JA, Olivares-Fuster O, Cuenca J,
Julve JM (2005) Programa de obtencion de hıbridos triploides de
mandarino en Espana. Phytoma 170:36–41
Oiyama I, Kobayashi S (1991) Citrus pentaploids from small seeds of
diploid 9 diploid crosses. HortScience 26(3):292–293
Oiyama I, Okudai N, Takahara T (1981) Ploidy levels of seedlings
obtained from 2x 9 4x crosses in citrus. In: Proceedings of 4th
international citrus congress, vol 1, International Society of
Citriculture, Tokyo, Japan, pp 32–34
Ollitrault P, Michaux-Ferriere N (1992) Application of flow cytom-
etry for citrus genetic and breeding. In: Proceedings of the 7th
international citrus congress, vol 1, International Society of
Citriculture, Acireale, Italy, pp 193–198
Ollitrault P, Dambier D, Allent V, Luro F, Jacquemond C (1996) In
vitro embryo rescue and selection of spontaneous triploid by
flow cytometry for easy peeler citrus breeding. In: Proceedings
of the 8th citrus international congress, vol 1, International
Society of Citriculture, Sun City, South Africa, pp 254–258
Ollitrault P, Dambier D, Luro F, Froelicher Y (2008) Ploidy
manipulation for breeding seedless triploid citrus. Plant Breed
Rev 20:323–354
Otto SP, Whitton J (2000) Polyploid incidence and evolution. Annu
Rev Genet 34:401–437
Peloquin SJ, Boiteaux LS, Simon PW, Jansky SH (2008) A
chromosome-specific estimate of transmission of heterozygosity
by 2n gamotes in potato. J Hered. doi:10.1093/jhered/esm110
Perrier X, Jacquemoud-Collet JP (2006) DARwin software. http://
darwin.cirad.fr/darwin
Perrier X, Flori A, Bonnot F (2003) Data analysis methods. In:
Hamon P, Seguin M, Perrier X, Glaszmann JC (eds) Genetic
diversity of cultivated tropical plants. Enfield, Science Publish-
ers, Montpellier, pp 43–76
Reforgiato-Recupero G, Russo G, Recupero S (2008) Mandarin tree
named ‘Top mandarin Seedless’. United States Patent, Patent
number: US PP18,568 P
Schuelke M (2000) An economic method for the fluorescent labeling
of PCR fragments. Nat Biotechnol 18:233–234
Shivanna KR, Linskens HF, Cresti M (1991) Response of tobacco
pollen to high humidity and heat stress: viability and germina-
bility in vitro and in vivo. Sex Plant Reprod 4:104–109
Soost RK (1965) Incompatibility alleles in the genus Citrus. Proc Am
Soc Hortic Sci 87:176–180
Soost RK (1969) The incompatibility gene system in citrus. In:
Chapman HD (ed) Proceedings of the first international citrus
symposium, vol 1. University of California, Riverside, pp 189–
190
Starrantino A, Recupero G (1981) Citrus hybrids obtained in vitro
from 2x females 9 4x males. In: Proceedings of 4th interna-
tional citrus congress, vol 1. International Society of Citriculture,
Tokyo, Japan, pp 31–32
Stephenson AG, Lau TC, Quesada M, Winsor JA (1992) Factors that
affect pollen performance. In: Wyatt R (ed) Ecology and
evolution of plant reproduction. Chapman & Hall, New York,
pp 119–134
Tokunaga T, Nii M, Tsumura T, Yamao M (2005) Production of
triploids and breeding seedless cultivar ‘Tokushima 3X No.1’
from tetraploid 9 diploid crosses in sudachi (Citrus sudachiShirai). Jpn Soc Hortic Sci 4(1):11–15
UPOV, International Union for the Protection of New Varieties
of Plants (2009) http://www.upov.int/en/publications/tg-rom/
tg201/tg_201_1.pdf
Vardi A, Spiegel-Roy P, Elchanati A (1991) Mandarin tree named
‘Winola’. United States Patent, Patent number: 8,216
Williams TE, Roose ML (2004) ‘TDE2’ Mandarin hybrid (Shasta Gold�
Mandarin), ‘TDE3’ Mandarin hybrid (Tahoe Gold� Mandarin) and
‘TDE4’ Mandarin hybrid (Yosemite Gold� Mandarin): three new
mid and late-season triploid seedless mandarin hybrids from
California. In: Proceedings of 10th international citrus congress,
vol 1, International Society of Citriculture, Agadir, Marruecos,
pp 394–398
Young HJ, Stanton ML (1990) Influence of environmental quality on
pollen competitive ability in wild radish. Science 248:1631–
1633
Zhou S, Ramana MS, Visser RGF, Van Tuyl JM (2008) Genome
composition of triploid lily cultivars derived from sexual
polyploidization of Longiflorum 9 Asiatic hybrids (Lilium).
Euphytica 160:207–215
1034 Plant Cell Rep (2010) 29:1023–1034
123



























![Câu 36. Giá trị lớn nhất của hàm số y=log,(2x+1) trên [0;1] là](https://img.dokumen.tips/doc/110x75/63142c7015106505030b3a71/cau-36-gia-tri-lon-nhat-cua-ham-so-ylog2x1-tren-01-la.jpg)

