Embed Size (px)
Citation preview

www.elsevier.com/locate/meegid
Infection, Genetics and Evolution 7 (2007) 520–534
The relative frequency of intraspecific lateral gene transfer of penicillin
binding proteins 1a, 2b, and 2x, in amoxicillin resistant
Streptococcus pneumoniae
Michael J. Stanhope a,*, Stacey L. Walsh b, Julie A. Becker b, Linda A. Miller b,Tristan Lefebure a, Ping Lang a, Paulina D. Pavinski Bitar a, Heather Amrine-Madsen c
a Department of Population Medicine and Diagnostic Sciences, College of Veterinary Medicine,
Cornell University, Ithaca, NY 14853, United Statesb GlaxoSmithKline, 1250 South Collegeville Road, Collegeville, PA 19426, United States
c GlaxoSmithKline, 5 Moore Drive, Research Triangle Park, NC 27709, United States
Received 28 December 2006; received in revised form 21 March 2007; accepted 23 March 2007
Available online 30 March 2007
Abstract
Evidence exists for both interspecific and intraspecific recombination (lateral gene transfer; LGT) involving Streptococcus pneumoniae pbp
(penicillin binding protein) loci. LGT of capsular genes, or serotype switching, is also know to occur between S. pneumoniae of different serotype.
It is not clear whether intraspecific pbp LGT is relatively common, whether there is a difference in the relative frequency of intraspecific LGT of
different pbps, and whether serotype switching is more or less frequent than pbp LGT. The purpose of this study was to use comparative
evolutionary biology analysis of 216 international clinical S. pneumoniae isolates, from the Alexander Project collection, to gain insight on these
issues, as well as the possible role they might be playing in spreading amoxicillin resistance. All 216 isolates were genotyped using MLST and
complete or nearly complete sequences for pbp1a, pbp2b, and pbp2x were determined. Amoxicillin MICs were available for each isolate. pbps
were genotyped using phylogenetics and two or more pbp types within a MLST sequence type (ST) or clonal complex were taken as putative cases
of pbp LGT; these hypotheses were statistically evaluated using the approximately unbiased (AU) test. Serotypes were determined for 171 of these
isolates and the minimum number of switching events necessary to explain the serotype phenotypes for each of the STs and clonal complexes were
evaluated. The majority (78%) of the amoxicillin resistant isolates were comprised in 5 clonal complexes. The relative frequency of pbp LGT was
greatest for pbp2b and 2x (minimum of 10.2 and 7.8%, respectively, of the isolates consistent with the LGT hypothesis), followed by 1a (3.9%).
Serotype switching was more frequent than intraspecific pbp LGT (33% of isolates consistent with serotype switching hypothesis). Although
intraspecific LGT of pbps is occurring and has played a role in the spread of amoxicillin resistance in S. pneumoniae, clonal dissemination appears
to be more significant.
# 2007 Elsevier B.V. All rights reserved.
Keywords: Streptococcus pneumoniae; Penicillin binding proteins; Amoxicillin resistance; Lateral gene transfer
1. Introduction
Beta-lactam resistance in Streptococcus pneumoniae has
emerged through the development of altered penicillin binding
proteins which have a decreased affinity for penicillin and other
beta-lactam antibiotics. Mutations within the transpeptidase
domain of these proteins are implicated in conferring this
* Corresponding author. Tel.: +1 607 256 5638; fax: +1 607 253 3440.
E-mail address: [email protected] (M.J. Stanhope).
1567-1348/$ – see front matter # 2007 Elsevier B.V. All rights reserved.
doi:10.1016/j.meegid.2007.03.004
decreased affinity. S. pneumoniae contains six pbps: the high
molecular weight pbp1a, pbp1b, pbp2a, pbp2b, and pbp2x and
the low molecular weight pbp3. Alterations in pbp2b and pbp2x
are the primary resistance determinants, conferring low-level
resistance to sensitive strains, whereas alterations in pbp1a
result in an increase in resistance levels (Grebe and Hakenbeck,
1996; Hakenbeck et al., 1999).
Although the efficacy of beta-lactam antibiotics has been
compromised through such alterations in pbps this has been less
so for amoxicillin than for penicillin. For example, in a
summary of the Alexander Project for the period 1998–2000

M.J. Stanhope et al. / Infection, Genetics and Evolution 7 (2007) 520–534 521
(the Alexander Project is a surveillance program established in
1992, examining the antimicrobial susceptibility of several key
respiratory pathogens to a range of compounds), Jacobs et al.
(2003) reported that 18.2% of the isolates are penicillin
resistant and 2.1% of the isolates are amoxicillin resistant.
There have been several recent studies reporting on compara-
tive pbp sequence analysis and/or issues of clonality involving
amoxicillin resistant S. pneumoniae (e.g. Doit et al., 1999;
Perez-Trallero et al., 2003; Kosowska et al., 2004; Cafini et al.,
2006). These studies have generally either included a relatively
small number of isolates, been focused on a particular country,
or did not include comparative pbp sequence analysis along
with the isolate genotyping. Additional information on issues of
clonality of amoxicillin resistant S. pneumoniae, concomitant
with comparative molecular evolution analyses of their pbp
sequences, could be helpful in elucidating the details under-
lying the development and spread of amoxicillin resistance in S.
pneumoniae.
Important alterations in pbps appear to have emerged
through recombination events involving S. pneumoniae pbp
genes and their homologues in human commensal streptococcal
species (Dowson et al., 1989, 1990; Potgieter and Chalkley,
1995; Reichmann et al., 1997). Some of these mosaic pbps
appear to have evolved through several recombination events
involving different donors (Dowson et al., 1997). In addition to
these interspecific recombination events there is also evidence
that intraspecific recombination of pbps is also occurring,
between different clones, suggesting an important possible
means of resistance spread (e.g. Coffey et al., 1991, 1995; Sa-
Leao et al., 2002). Intraspecific recombination is not surprising
given that recombination frequency in S. pneumoniae increases
with decreasing sequence divergence between donor and
recipient (Majewski et al., 2000). However, the relative
frequency with which such intraspecific lateral gene transfer
(LGT) or recombination events is occurring amongst clinical
isolates of different S. pneumoniae clones is not well
understood or thoroughly explored. In addition, we are not
aware of any study which specifically attempts to assess this
with regard to amoxicillin resistant isolates. This focus on LGT
of pbps from amoxicillin resistant isolates could shed light on
the spread of amoxicillin resistance in S. pneumoniae.
Estimates of the relative frequency of LGT involving resistance
conferring loci should prove beneficial in developing models
designed to predict the spread of resistance. Our intentions here
are to use a comparative evolutionary biology approach to
provide insight on the relative frequency of lateral gene transfer
of pbp1a, 2b, and 2x genes in S. pneumoniae using nearly
complete pbp gene sequences derived from a large set of
clinical isolates including amoxicillin resistant isolates as well
as other resistance profiles and sampled from a variety of
geographic areas.
It has been clear for some time that S. pneumoniae clinical
isolates of the same clone or sequence type often express
different serotypes (e.g. Coffey et al., 1991) indicating that
lateral gene transfer of capsular genes takes place between S.
pneumoniae lineages (sometimes referred to as serotype
switching), emulating the early transformation experiments
on S. pneumoniae involving capsular loci (reviewed in Yother,
2004). Analyses of recombinational junctions and sequence
polymorphisms within introduced capsular genes led Coffey
et al. (1998) to conclude that 19F variants of the Spain 23F-1
clone arose on multiple independent occasions, from which
they suggested that changes in capsular type by recombination
may be relatively frequent in pneumococci. MLST databases
provide ample evidence supporting a diversity of genotypes for
individual serotypes and the most parsimonious hypothesis to
explain this is horizontal gene transfer of the capsular gene
locus (e.g. Spratt et al., 2004). There is also evidence that
capsular genes and pbp loci (particularly pbp1a and pbp2x) can
be co-transferred (Coffey et al., 1999; Trzcinski et al., 2004). It
is presently unclear whether serotype switching is more
frequent than lateral gene transfer of pbps and the degree, or
frequency, with which pbps and capsular loci are co-
transferred. An additional purpose of this study was to provide
insight on these issues through a comparative evolutionary
biology approach involving isolates for which serotype,
housekeeping genotypes, and pbp gene sequences have been
obtained. Although there are numerous studies supporting the
importance of clonal dissemination in the spread of beta-lactam
resistance in S. pneumoniae (reviewed in Klugman, 2002),
these studies generally do not have as an additional goal the
simultaneous assessment of the role of intraspecific pbp LGT.
We argue that without concomitantly evaluating the clonal
hypothesis with knowledge of the intraspecific evolutionary
history of the pbps one cannot adequately disentangle the
relative importance of clonal dissementation versus pbp LGT in
the spread of resistance. We have recently adopted a similar
approach to assess the role of intraspecific LGT of fluoroqui-
nolone target loci in the spread of fluoroquinolone resistance in
S. pneumoniae (Stanhope et al., 2005), concluding that a small
proportion of the isolates were consistent with the intraspecific
LGT hypothesis. Layering on top of this an attempt to better
understand the relative frequency of serotype switching is
important because of the potential role serotype switching
could play in avoiding newly developed vaccines (e.g. PCV7)
and possibly the simultaneous development of beta-lactam
resistance and vaccine avoidance through co-transformation of
the capsular locus and pbps.
2. Materials and methods
2.1. Isolates
Isolates included in this study come from the Alexander
Project collection. Susceptibility data and descriptions of the
tests for the Alexander Project have been published elsewhere
(Felmingham and Washington, 1999; Jacobs et al., 2003). The
Alexander Project collection contains a total of 172 isolates of
non-US origin that are amoxicillin resistant (MIC � 8 mg/ml).
We made an effort to include all of these but quite a number
could not be cultured, or one or another of the pbp loci
amplified, resulting in a total of 120 resistant isolates from non-
US locations. Within the Alexander Project collection there are
a total of 340 isolates from non-US locations that are of

M.J. Stanhope et al. / Infection, Genetics and Evolution 7 (2007) 520–534522
intermediate amoxicillin resistance (MIC = 4 mg/ml) and we
randomly chose 20 from this set for inclusion in our analysis;
two of these could not be cultured. These non-susceptible
isolates of non-US origin come from the following countries:
South Africa (ZA), United Kingdom, Spain, France, Israel (IS),
Portugal (PR), Slovakia (SL), Switzerland, Belgium, Czech
Republic (CR), Japan, Italy, Poland, and China (Hong Kong).
The majority of these non-susceptible isolates (97/138) arose
from France and Spain. There are many thousands of non-US
isolates that are amoxicillin susceptible in the Alexander
Project collection. From these susceptible non-US isolates we
randomly chose 50; 6 of these would not culture. Countries in
this non-US susceptible category included: Germany, Greece,
Spain, France, Portugal, Slovakia, Switzerland, Belgium, Italy,
and Poland. Large numbers of amoxicillin resistant and
intermediate isolates from the US are included in the Alexander
Project and we intend to focus on the US in a subsequent
analysis. However, in order to provide some preliminary
perspective on relationships between European and US isolates
in the present study we included a small number of US isolates.
Our focus in this instance involved the state of Ohio because our
preliminary analysis of the Alexander Project database
indicated that it was one of the states with relatively high
numbers of amoxicillin resistant isolates. We included 32
isolates from Ohio, 9 of which were amoxicillin non-
susceptible. Two further US isolates (from California and
New York) of non-susceptibility were also included as
incidental to the entire set simply because sequence data
was already available for these particular isolates. In total 216
isolates were included.
Serotyping was performed at G.R. Micro Ltd. (London, UK)
using the Neufeld Quellung reaction and Statens Serum
Institute (SSI) antisera (SSI, Copenhagen, Denmark). SSI was
used as the reference laboratory for quality assurance and
identification of rare serotypes.
2.2. Sequence data collection
Sequence data for all 216 isolates were obtained for all 7
MLST housekeeping loci using previously described PCR
primers (Enright and Spratt, 1998) and for nearly complete
pbp1a, 2b and 2x genes. PCR primers for the pbp loci included
the following (the number in the primer name refers to the
nucleotide position of the 30 end of the primer in the R6
reference sequence): pbp1a-F26: 50 TGAACAAACCAAC-
GATTCTGCGCCT 30; pbp1a-R2140: 50 ATGTTATGGT-
TGTGCTGGTTGAGG 30; pbp2b-F53: 50 AACAGCCATTC-
GATTCCGATTCGGCT 30; pbp2b-R1997: 50 TCACGCG-
CAATGGAAGGTCCTA 30; pbp2x-F31: 50 GAAGTGGAC-
AAAAAGAGTAATCCGTTATG; pbp2x-R2362: 50 CTGATG-
CTGTTTGACATCCTCATGC 30 or pbp2x-R2265: 50 ATGT-
CACAATTCCAGCACTGATGG 30. The PCR products were
purified with a QIAGEN QIAquick PCR purification kit
according to the manufacturer’s instructions. The products
were sequenced on both strands using Applied Biosystems
machines 3730, 3700, and 3100 and Applied Biosystems Big
Dye. The resulting comparative data sets included a sequence
alignment for pbp1a that was missing 55 bp from the 5 prime
end and 57 bp from the 3 prime end of this particular locus
(2.0 kb fragment). For pbp2b the resulting sequence alignment
excluded 150 bp from the 5 prime end of the gene and 108 bp
from the 3 prime end (1.8 kb fragment). For pbp2x only 54 bp
from the 5 prime end were excluded; the 3 prime end was
complete (2.2 kb fragment).
2.3. Data analysis
MLST housekeeping sequence data were compared to the
MLST S. pneumoniae database (http://spneumoniae.mlst.net/)
to assign STs (sequence types). Sequence types not present on
the database were assigned unique identifiers for our analysis.
Groups of related STs differing in not more than one of the
seven loci that were thought to be descendants from a common
ancestor were grouped into clonal complexes using eBURST
(Feil et al., 2004). Singleton isolates were defined as individual
isolates of unique ST that did not group into a clonal complex.
Singleton clones were defined as unique STs that did not group
with a clonal complex and which were comprised of more than
one isolate.
Homologous DNA sequence alignments of pbp gene
sequences were generated by using ClustalX, version 1.83
(Thompson et al., 1994), with any necessary manual editing in
GeneDoc, version 2.6.0.2. The best-fitting model of sequence
evolution and the corresponding values for the rate matrix,
shape of the gamma distribution, and proportion of invariant
sites were estimated by the program MODELTEST (Posada and
Crandall, 1998). Phylogenies of the pbp sequences were
reconstructed by the maximum likelihood (ML) method
implemented in PAUP*4.0b. Starting trees for the ML analyses
were obtained by neighbor joining with nearest neighbor
interchange used as the branch-swapping algorithm. Bootstrap
support values were obtained with 500 ML replicates using the
program PHYML (Guindon and Gascuel, 2003).
The pbp phylogenies were used to determine pbp types,
where a type was considered to be a monophyletic group (two
or more sequences) comprised of sequences with little or no
sequence divergence. This means that singleton lineages were
considered ‘‘non-typeable’’ under this criterion. pbp types were
then compared to the MLST results demarcating STs and clonal
complexes. Isolates of the same ST, or clonal complex,
harboring the same pbp type were considered to share that pbp
type because of common ancestry and dissent (clonal
dissemination). In contrast, isolates of the same ST (or clonal
complex) with different pbp types were judged to have acquired
those different pbps through lateral gene transfer (LGT). To be
considered as putative LGT, different pbp types must be
separated by a minimum of three nodes on the pbp tree. This
subjective criterion was determined after reconstructing and
visually inspecting each phylogeny and its conservative
intention was to avoid the inclusion of moderately divergent,
neighboring sequences (particularly sister groups), with well-
defined pbp types. In order to provide a more rigorous statistical
perspective on whether such putative LGT events had a history
that was significantly different from that of clonal dissemina-

M.J. Stanhope et al. / Infection, Genetics and Evolution 7 (2007) 520–534 523
tion (common ancestry and descent), pbp ML trees were
constrained to support the clonal hypothesis for individual LGT
sequences and these constrained trees were assessed for
significant difference against the highest likelihood trees using
the AU test implemented in CONSEL (Shimodaira and
Hasegawa, 2001; Shimodaira, 2002). Rejection of the null
hypothesis of no difference between the constrained tree and
the highest likelihood tree, and acceptance of the alternative
hypothesis, was taken as phylogenetic statistical evidence for
LGT of that pbp involving that isolate. We refer to these two
different means of evaluating pbp LGT as the ‘‘subjective’’ and
‘‘statistical’’ criteria respectively; we only consider instances
that satisfy both criteria as putative cases of LGT.
The principle of parsimony was employed in evaluating the
occurrence and number of putative LGT and serotype switching
events. This means, for example, that if we have 5 isolates of a
particular ST and three of them are serotype 11 and two of them
are serotype 14, we consider this to be a single serotype switching
event from 11 to 14 (or vice versa); the additional serotype 14 (or
11) isolate is assumed to be the consequence of clonal
dissemination from a serotype 14 (or 11) ancestor. Because
we do not have an evolutionary history for isolates comprising a
clone it is not possible to definitively derive the directionality of
such switches. Nonetheless, it is often the case that our data, and/
or the MLST database, indicate that a particular serotype is
clearly more representative of a clone and thus suggest a possible
directionality of the serotype switch. In any event, the minimum
number of switching events necessary to explain the array of
serotype phenotypes is taken as the number of serotype switches
for that clone. Similarly, if we have a STor clonal complex which
has 20 isolates of a particular pbp type and 5 of another pbp type,
this is assumed to involve a single LGT event from the more
numerous type to the less numerous type, followed by clonal
multiplication of that particular LGT event. This conservative
approach will have the consequence of underestimating the
actual number of LGT or serotype switching events.
The nucleotide sequences obtained in this study have been
deposited in GenBank under the following accession numbers:
EF499539–EF501698.
3. Results and discussion
3.1. Clonality of amoxicillin resistance
The 216 isolates were comprised of 114 sequence types (ST),
including 49 STs listed on the S. pneumoniae MLST database.
The remaining 65 STs were single, occasionally double, and one
triple locus variant of existing STs (Appendix A). A total of 38
STs were unique because of combinations of existing alleles and
27 STs were unique because of unrecorded alleles. The 114 STs
comprised 13 clonal complexes (eBURST determination of STs
and their associated SLVs), plus 5 singleton clones (two or more
isolates of same ST) and 39 singleton isolates (individual isolates
of unique ST which could not be assigned to a clone or clonal
complex). Amoxicillin non-susceptible isolates were distributed
among 7 clonal complexes and the majority of amoxicillin
resistant isolates (98/125; 78.4%) were contained in 5 clonal
complexes. These five clonal complexes included representatives
of the PMEN (Pneumococcal Molecular Epidemiology Net-
work) clones Spain 23F-1, Spain 6B-2, Spain 9V-3, and Poland
23F-16, as well as one additional group without a specific PMEN
clonal member. This latter group did include, however, a ST (13)
which was a SLV of the England 14-9 clone. The three Spanish
clones have been mentioned elsewhere as major contributors to
amoxicillin resistance (Perez-Trallero et al., 2003); we are not
aware of Poland 23F-16 being similarly implicated. This clonal
complex included 2 isolates with amoxicillin MICs of 16 mg/L.
The isolates were sampled from the years 1996–2000 and were
all from Poland. Other PMEN clones represented in our set of
isolates included the following: Taiwan 23F-15, Sweden 1–28,
The Netherlands 3–31, The Netherlands 8–33, The Netherlands
14–35, The Netherlands 18C-36, The Netherlands 15B-37, The
Netherlands 7F-39, and Portugal 6A-41. With the exception of
the Taiwan 23F-15 isolates, these were all amoxicillin
susceptible isolates.
There were 11 amoxicillin non-susceptible isolates from the
USA, grouped into 6 STs; within this group there was a set of
clearly resistant isolates from Ohio, all ST 13. The clonal
complex containing ST 13 has 1 isolate collected from France
in 1999, with all other isolates from Ohio, USA in 2000. This
same sequence type has been reported previously in the US (e.g.
Sa-Leao et al., 2001), although we are not aware of a report
linking it to amoxicillin resistance. The majority of the
members of this clonal complex (and indeed all of the ST 13
isolates) have the same unique pbp types (Figs. 1 and 2). This
could be an important clone to watch in the USA for the spread
of amoxicillin and other types of resistance, and it is perhaps
fortunate that all these isolates were serotype 14, which is one
of the serotypes included in the PCV7 vaccine.
3.2. Lateral transfer of pbp loci
Pbp1a was divided into 12 different phylogenetic groups, or
pbp types (Fig. 3), with 10 isolates considered as non-typeable
based on our criteria. One of the distinguishing features of this
treewas that the different pbp1a types were relatively distinct, yet
showed little or no variation within a type. pbp1a type 1 was the
most common, including a total of 73 isolates, encompassing two
clonal complexes (4 and 7; Figs. 2 and 3) and one singleton
isolate. Clonal complex 4 includes ST 81, which is the Spain 23F-
1 clone, and clonal complex 7 includes ST 156, which is the
Spain 9V-3 clone. These two clones have previously been
reported to possess the same pbp1a genotype, based on pbp RFLP
analysis (e.g. McGee et al., 2001). Clonal complex 4 includes 13
different sequence types, and all of these isolates have our pbp1a
type 1. Clonal complex 7 includes 21 different sequence types
and 9 of these sequence types possess pbp1a type 1. The most
parsimonious explanation of this evolutionary pattern is that
pbp1a type 1 has been common to clonal complex 4 for some
time, since all 13 different sequence types possess it, and at some
point this sequence was passed to a member of clonal complex 7,
via a lateral gene transfer event, and through common ancestry
and descent it subsequently diversified into other STs within this
other group. This suggests that two intraspecific LGT events are

Fig. 1. Summary of pbp LGT and serotype switching events, as well as
amoxicillin (amox.) MIC values, at the level of sequence type. pbp type and
serotypes are indicated along the top of each figure and sequence type is
indicted along the vertical axis. A horizontal red line indicates a LGT event for
M.J. Stanhope et al. / Infection, Genetics and Evolution 7 (2007) 520–534524
sufficient to explain the observed phylogenetic distribution for
this sequence in this set of 216 isolates; once from clonal
complex 4 to 7 and once from either 4 or 7 into the singleton
lineage identified by isolate 517.1FR95.
In general there was a very high degree of association between
pbp1a type and ST or clonal complex, suggesting that the
distribution of the vast majority of these pbp sequences amongst
these 216 isolates could be explained by clonal dissemination. Of
the 27 sequence types represented by more than one isolate (a
total of 128 isolates), only three of these STs showed evidence for
lateral transfer of pbp1a (Fig. 1); ST 156 supported 3 cases of
LGT, and there was one example in each of STs 1624, and SLV
156 (ddl-4). In the case of ST 156, two of the LGT cases were
correlated with an increase in amoxicillin MIC (from 8 to 16 mg/
L) and were associated with isolates from 1999 to 2000, with the
other members of the clone dated at 1995, suggesting a possible
time frame for the LGT events within this clone. All of these
putative cases of LGT were supported by both the ‘‘subjective’’
(different pbps must be separated by at least three nodes on the
pbp tree) and ‘‘statistical’’ (phylogenetic AU test) criterion.
Thus, the frequency of LGT for pbp1a, using the disrupted
monophyly of ST as the criterion, was 5/128 (3.9%). This
association was also quite apparent in a comparison of clonal
complex and pbp1a type (Fig. 2). Within the 160 isolates
distributed into 13 clonal complexes, there were 10 examples of
pbp1a LGT (10/160; 6.3%). This slight increase in number of
examples of LGT, when the criterion is disruption of clonal
complex monophyly, is precisely what would be expected with
the expansion of the comparative framework to a consideration of
groups of closely related clones, and not just single STs, since this
represents an expansion of the evolutionary timeframe for the
detection of such events.
The origins of many of these pbp sequences may have
involved recombination events with commensal Streptococcus
species such as Streptococcus mitis (e.g. Dowson et al., 1989),
after which antibiotic selection of particular recombinants,
clonal dissemination, and intraspecific LGT may have been the
more significant factors. The degree or frequency to which
ongoing interspecific LGT involving S. pneumoniae pbps and
commensal flora takes place is not well understood. A
phylogenetic comparison of these S. pneumoniae pbp1a
sequences to orthologs from S. mitis and Streptococcus oralis
available on GenBank (three sequences from S. mitis and one S.
oralis), indicates that one of these S. pneumoniae isolates
(675.1PL96) possesses a pbp1a sequence that is much more
closely related to S. mitis, and thus supports an instance of
interspecific LGT. The pbp1a sequence for 675.1PL96 was
listed as non-typeable in our analysis, and was not implicated in
an intraspecific LGT event. Additional comparative data on
pbps for commensal species such as S. mitis could eventually
lead to an assessment of the relative importance of interspecific
that sequence type. Non-typeable pbps are indicated with their isolate ID; non-
typeable pbps that were not implicated in a LGT event are indicated as an
addition to the pbp type with which they were judged not significantly different
(e.g. 5 + 675.1PL96).

Fig. 2. Summary of pbp LGT and serotype switching events, as well as
amoxicillin (amox.) MIC values, at the level of clonal complex. pbp type
and serotypes are indicated along the top of each figure and clonal complex is
indicted along the vertical axis. A horizontal red line indicates a LGT event for
Fig. 3. Unrooted maximum likelihood (ML) tree of pbp1a sequences derived
from the set of 216 S. pneumoniae isolates. pbp1a sequence type designations
are listed adjacent to the various clades comprising each type; *ML bootstrap
support in excess of 80%; isolate IDs for non-typeable pbp1a sequences are
indicated for those sequences implicated in a LGT event.
M.J. Stanhope et al. / Infection, Genetics and Evolution 7 (2007) 520–534 525
versus intraspecific LGT in development and spread of beta-
lactam resistance in S. pneumoniae.
One of the remarkable aspects of the evolutionary history of
pbp1a (and indeed to an extent, also pbp2b and 2x) is the
pronounced lack of sequence variation in many of the major
types. del Campo et al. (2006) have recently pointed out similar
such sequence conservation for pbps and MurM in penicillin
resistant isolates. Earlier RFLP studies suggested similar
homogeneity amongst certain pbps amplified from different
isolates, which formed the basis of PCR/RFLP typing schemes,
such as the one adopted by the Pneumococcal Molecular
Epidemiology Network (Gherardi et al., 2000). RFLP digests
though, are not as specific as complete DNA sequences. The
increased specificity for each of the pbp loci resulting from our
approach, results in distinguishing some pbps that the RFLP
scheme lumps into a single category. For example, the PMEN
lists the pbp1a type for ST 81 and ST 90 as both type 1,
that clonal complex. Non-typeable pbps are indicated with their isolate ID; non-
typeable pbps that were not implicated in a LGT event are indicated as an
addition to the pbp type with which they were judged not significantly different
(e.g. 5 + 675.1PL96).

M.J. Stanhope et al. / Infection, Genetics and Evolution 7 (2007) 520–534526
however, our sequence analysis clearly indicates that these
clones possess distinct pbp types (Figs. 1 and 3). Further more
subtle distinctions are apparent for some of the ST 156 isolates,
which also are listed on PMEN as pbp1a type 1; although the
majority of the ST 156 isolates have the same type as ST 81,
some isolates carry a pbp1a sequence that is clearly different,
and which we categorize as 1b (Figs. 1 and 3). Although such
distinctions may seem trivial at first glance, they result in very
different interpretations regarding the evolutionary history of
the pbp gene. For example, ST 81 and ST 90 with the same pbp
sequence, suggests recent lateral transfer of this pbp between
clones, however, different pbp types suggests something
different; it could still involve a history of lateral transfer,
followed by some period of change in the pbp, but in this case it
does not suggest a recent event.
The distinct lack of sequence variation for the majority of
pbp1a types is true even though several of these types, such as
pbp1a type 1 (common to the Spain 23F-1 clone), are distributed
amongst different clones and clonal complexes. Sequence types
that are members of the clonal complex defined by Spain 23F-1
(clonal complex 4 in our analysis) are generally single locus
variants of ST 81, or more occasionally, one of the other STs in
this complex. Current evidence suggests different STs are formed
by recombination of their housekeeping loci (Spratt, 2004).
Although there is undoubtedly selection pressure against certain
recombinants of such housekeeping loci, many such recombi-
nants will not possess sequences that will result in significant
negative selection pressure. Similar recombination events of
pbp1a type 1 are likely to be at a significant disadvantage, if type
1 is playing a role in conferring amoxicillin and perhaps beta-
lactam resistance in general. The majority (43/73; 59%) of the
isolates possessing pbp1a type 1, have amoxicillin MICs � 8 and
only 15% (11/73) have MICs � 2. Thus, a likely explanation is
that pbp1a type 1 was present in the ancestor of our clonal
complex 4, conferred a selective advantage to isolates that
retained it, and was passed onto other STs, through common
ancestry and descent, while those STs differentiated through
occasional recombination events of their housekeeping loci.
Spratt et al. (2004) suggested that SLVof ST81 have arisen over a
period of about 30 years, and amoxicillin was first introduced in
1972 (White et al., 2004). Thus, the history of this apparent
example of strong directional selection favoring a particular
pbp1a allele can be roughly dated as occurring over the course of
the last 30 years. The distinct nature of the majority of the pbp1a
types, concomitant with the pronounced lack of variation for
most of these groups, as well as their tendency to be clonally
disseminated, suggests a similar history for most of these pbp1a
types.
The phylogeny for pbp2b (Fig. 4) divided the sequences into
21 types, several of which were very closely related and thus
were designated as subtypes of a larger group (e.g. 1, 1b, 1c).
This tree differed from pbp1a in that the types tended not to be
as clearly distinct and there were many more sequences not
falling into clearly distinguishable clades, yielding more in the
non-typeable category. Ppb2b type 10 was the most common,
and included 54 isolates, all but four of which came from clonal
complex 4 or 7. Unlike pbp1a, where all of the clonal complex 4
isolates carried exactly the same pbp1a sequence, for pbp2b, 7
out of a total of 39 isolates were not type 10. In the case of
clonal complex 7, 37 out of a total of 55 isolates were not pbp2b
type 10. Overall, like pbp1a, pbp2b also showed a high degree
of association between pbp2b genotype and ST or clonal
complex. Out of the 128 isolates comprising the 27 STs
consisting of more than one isolate, there were 13 examples of
pbp2b LGT (13/128; 10.2%; Fig. 1). When one expands the
comparison to the level of clonal complex, there were a total of
30 examples of pbp2b LGT (30/160; 18.8%; Fig. 2).
Phylogenies that include pbp2b sequences taken from
GenBank for S. mitis, S. oralis, and Streptococcus sanguis (16
sequences from S. mitis, 2 S. oralis, and 1 S. sanguis), support
interspecific LGT events involving S. pneumoniae isolates
595.5IT97 and 640.6IT98 (ST 321), as well as for 595.2FR99;
the implicated species is S. mitis. The former two sequences
were pbp2b type 8, and were not implicated in an intraspecific
LGT event in our analysis; the latter sequence was non-typeable
and was implicated in a LGT event involving the clonal
complex 7 comparison.
The phylogeny for pbp2x (Fig. 5) divided the sequences into
25 types, including several subtypes. pbp2x type 3 was the most
common and with the exception of 5 isolates (four of which
were singleton isolates), was found only in clonal complex 4 or
7. However, ten isolates within clonal complex 4 were not
pbp2x type 3 and 37 isolates within clonal complex 7 were not
type 3. This set of 37 isolates from clonal complex 7, not
carrying pbp2x type 3, were the same isolates not carrying
pbp2b type 10. This in turn suggests a possible evolutionary
history in which a clonal complex 4 member (or members),
acted as a donor of all three pbps (either simultaneously or
independently) to a clonal complex 7 isolate(s). These pbps
were then passed on by common ancestry and descent within
the clonal complex 7 group, eventually being distributed
amongst 8 different STs within this complex. Concomitant with
this diversification there were further events of LGT involving
clonal complex 7 and other lineages, resulting in some isolates
within the various clones harboring different pbp types than
their ancestors. For example, the majority of the ST 156 (Spain
9V-3) isolates have the same pbp types, and these in turn are the
same as those of the majority of the clonal complex 4 members,
including ST 81 (Spain 23F-1). This similarity in pbps between
Spain 23F-1 and Spain 9V-3 and the hypothesis of LGT
involving these clones and their pbps was first noted by Coffey
et al. (1991). After the original transfer event we suggest there
were several subsequent LGT events, changing the pbp types of
some ST 156 members. Isolates 405.7PR00S, 405.3BE99, and
405.16SW99, all ST 156, each have different pbp1a, 2b and 2x
genes suggesting a minimum of 3 independent LGT events. In
the TIGR S. pneumoniae genome sequence (Tettelin et al.,
2001), pbp1a and pbp2x are separated by about 42 kb and
pbp2b is in excess of 1 Mb removed. Thus, although it is
possible that 1a and 2x are co-transferred, it is unlikely that 2b
would be transferred with either of the other loci. Our data
suggest there is less transfer involving 1a than 2x (and 2b as
well), and thus, even though 1a and 2x are relatively proximal,
we favor the hypothesis that each apparent LGT event is

Fig. 4. Unrooted maximum likelihood (ML) tree of pbp2b sequences derived from the set of 216 S. pneumoniae isolates. pbp2b sequence type designations are listed
adjacent to the various clades comprising each type; *ML bootstrap support in excess of 80%; isolate IDs for non-typeable pbp2b sequences are indicated for those
sequences implicated in a LGT event.
M.J. Stanhope et al. / Infection, Genetics and Evolution 7 (2007) 520–534 527
independent of the other pbps. In our recent LGT analysis of
fluoroquinolone target loci we concluded that Spain 23F-1 is a
frequent donor of fluoroquinolone resistance conferring loci to
other lineages (Stanhope et al., 2005). In the present case it
would appear that Spain 23F-1, or at least a member(s) of that
clonal complex, has been a likely donor of pbp genes as well.
However, the evidence in this case tends to support this LGT
event with one or two recipient lineages, rather than multiple
independent lineages as in our fluoroquinolone example. Of
course it is possible that a broader sampling of isolates might
have picked up other examples and we are currently working
towards making this assessment involving a more globally
complete sample of beta-lactam resistant isolates from the
Alexander Project collection. Using the criterion of disrupted
ST monophyly, there were 10 examples of pbp2x LGT (10/128;
7.8%; Fig. 1), and with the broader consideration of disrupted
clonal complex monophyly, there were 29 (29/160; 18.1%;
Fig. 2) examples of pbp2x LGT in this data set.
Phylogenies that include pbp2x sequences taken from
GenBank for S. mitis and S. oralis (12 sequences from S. mitis,
2 from S. oralis), do not provide clear support for interspecific
LGT events involving any of our S. pneumoniae isolates. Many
of the S. mitis and S. oralis sequences do fall within the S.
pneumoniae group, but none of the resulting branching
arrangements provide clear evidence for donor/recipient
relationships. Nonetheless, since the S. mitis sequences tend
to be arranged as small paraphyletic groups scattered
throughout the S. pneumoniae tree, it does suggest the
possibility of a complex history of recombination involving
pbp2x from S. mitis and S. pneumoniae.
3.3. Serotype switching
The phenomenon of serotype switching in Streptococcus
pneumoniae via capsular locus recombination between clinical
isolates, has been known for some time (Coffey et al., 1991,

Fig. 5. Unrooted maximum likelihood (ML) tree of pbp2x sequences derived from the set of 216 S. pneumoniae isolates. pbp2x sequence type designations are listed
adjacent to the various clades comprising each type; *ML bootstrap support in excess of 80%; isolate IDs for non-typeable pbp2x sequences are indicated for those
sequences implicated in a LGT event.
M.J. Stanhope et al. / Infection, Genetics and Evolution 7 (2007) 520–534528
1998; Barnes et al., 1995; Nesin et al., 1998; Ramirez and
Tomasz, 1999) and there is evidence, at least for Spain 23F-1, that
this may occur relatively frequently (Coffey et al., 1998; Spratt
et al., 2004). Not all of the isolates in the present dataset were
serotyped, however, we do have serotype data for 171 of them,
allowing an assessment of the relative frequency of capsular
switching versus pbp LGT in the same isolates. Of the total
isolates serotyped, 103 fall into STs with more than a single
serotyped representative (total of 22 STs). Using disrupted ST
monophyly as the criterion of capsular switching, there were a
minimum of 40 isolates within this set of 103 (40/103; 38.8%)
expressing serotypes consistent with an evolutionary history of
capsular switching (Fig. 1). For example, ST 62 includes three
isolates, two of which express serotype 11, and one serotype 14.
Whether one regards serotype 11 or 14 as the ancestral state, we
still consider this to be a single serotype switching event. In this
particular example our data supports more cases of serotype 11
(albeit only by one) for this clone and if one examines the isolates
for ST 62 on the MLST database, all six are either serotype 11 or
11A, suggesting 11 may be the ancestral state for this clone, and
that there was a single capsular switching event, from 11 to 14.
Only three STs, out of the total of 22 STs with multiple serotyped
representatives, do not support evidence for capsular switching.
If one examines pbp LGT for the same serotyped isolates
considered for possible capsular switching, there were 5
examples of pbp1a, 11 for pbp2b, and 10 for pbp2x. Thus,
our data suggest that capsular switching is roughly 3–7 times
more frequent than LGT of pbps. Furthermore, of the cases of
apparent pbp LGT (based on the criterion of disrupted ST
monophyly) involving the set of serotyped isolates, only a small
number of these involve LGT events that can be unambiguously
identified as independent of a serotype switch (1 case for pbp1a, 4
for pbp2b, and 3 for pbp2x). S. pneumoniae genome sequence
data indicate that pbp2x and pbp1a are located within 10 and
14 kb of the capsular biosynthesis operon, and it has been
demonstrated that capsular genes and pbp loci can be transferred

M.J. Stanhope et al. / Infection, Genetics and Evolution 7 (2007) 520–534 529
in a single transformation step (Trzcinski et al., 2004). Coffey
et al. (1999), presented evidence indicating that serotype 14
variants of the Spain 9V clone have arisen on multiple occasions
by large recombinational replacements that include the majority
of the pbp1a gene. This co-transformation of capsular loci and
pbps suggests a possible explanation for why there is a
correlation between serotype and resistance to beta-lactam
antibiotics, with some serotypes, such as 9Vand 19A, correlated
with high resistance, and other serotypes, such as 1–4, generally
associated with low resistance (McCormick et al., 2003).
Although our results involving possible co-transfer of capsular
genes and pbps are circumstantial, and thus should be regarded
with certain skepticism (e.g. often not possible to unambiguously
determine the ancestral state of serotype), it is also true that the
majority of pbp LGT cases (at least for the serotyped isolates)
cannot be disentangled from possible co-transfer with the
capsular locus. Thus, our results are certainly consistent with the
possibility of relatively frequent co-transformation of capsular
genes and pbp loci.
The vast majority of the amoxicillin non-susceptible isolates
in this dataset expressed serotypes included in the PCV-7
vaccine. The majority of putative switching events involved a
change from a vaccine serotype to either another vaccine
serotype, or a vaccine related serotype. There were, however, at
least six exceptions, which parsimoniously supported a change
from vaccine or vaccine related serotype to a non-vaccine
serotype. For example, with ST 838, we have serotype data for
16 isolates, 15 of which express vaccine or vaccine-related
serotypes, and one of them is serotype 3, leading to the most
parsimonious hypothesis of a switch from vaccine or vaccine
related, to serotype 3. It is important to realize that our
estimates of capsular switching are undoubtedly an under-
estimate of the actual number of switching events that have
occurred amongst these isolates. Coffey et al. (1998) concluded
from an examination of the differences in recombinational
junctions and sequence polymorphisms of the capsular genes of
19F variants of the Spain 23F-1 clone, that of the eight variants
they examined, a minimum of four separate evolutionary events
were responsible. In our parsimony based assessment of
serotype changes we are evaluating such changes as a single
evolutionary event. The pbp assessment on the other hand,
although conservative, is not as likely to be as much of an
underestimate of LGT as the capsular genes since in this case
we are examining nearly complete pbp sequences and not a
phenotype. Thus, it would seem that for each of these clones
and clonal complexes, there have been many more examples of
capsular gene recombination than pbp lateral transfer. It is
interesting to note that for clonal complex 4, which includes
Spain 23F-1 (ST 81), there is considerable variation in capsular
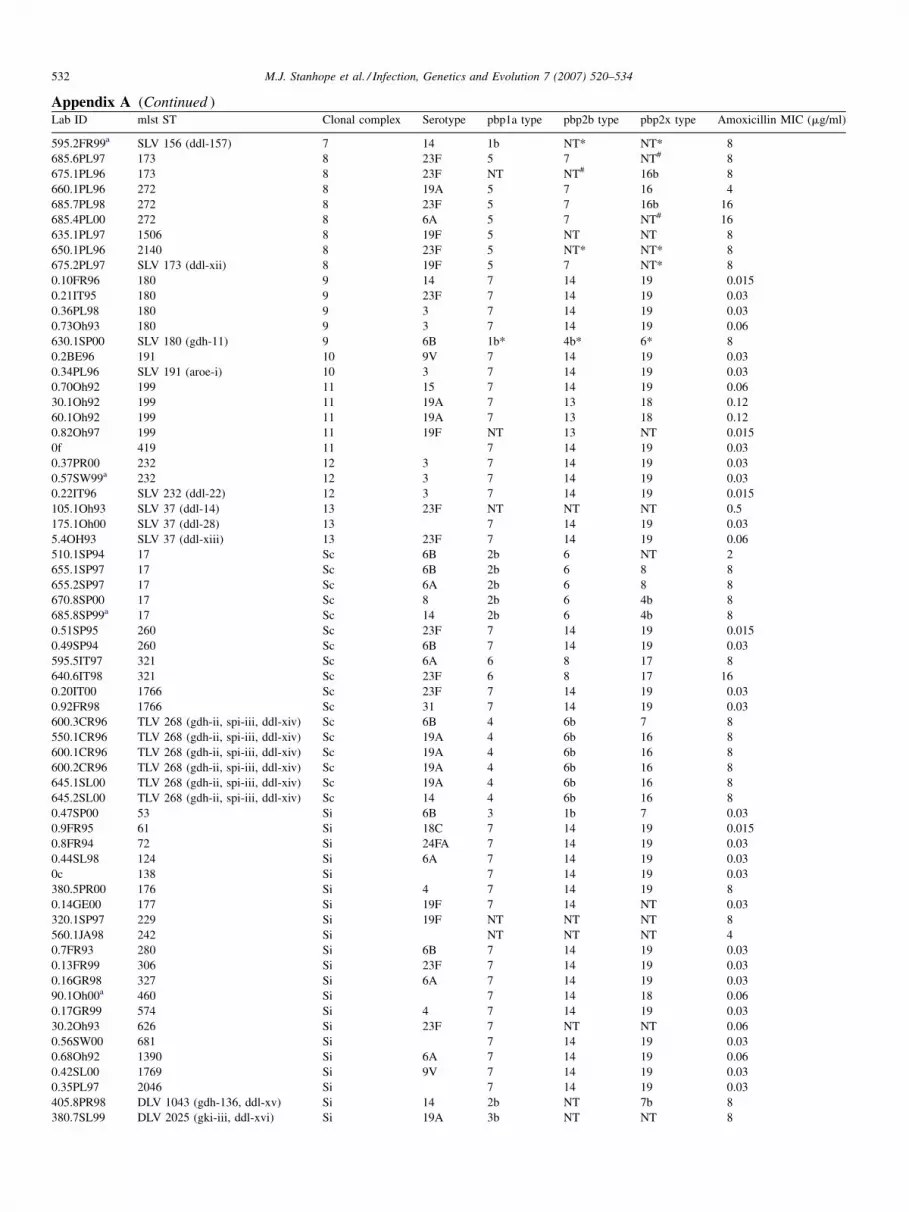
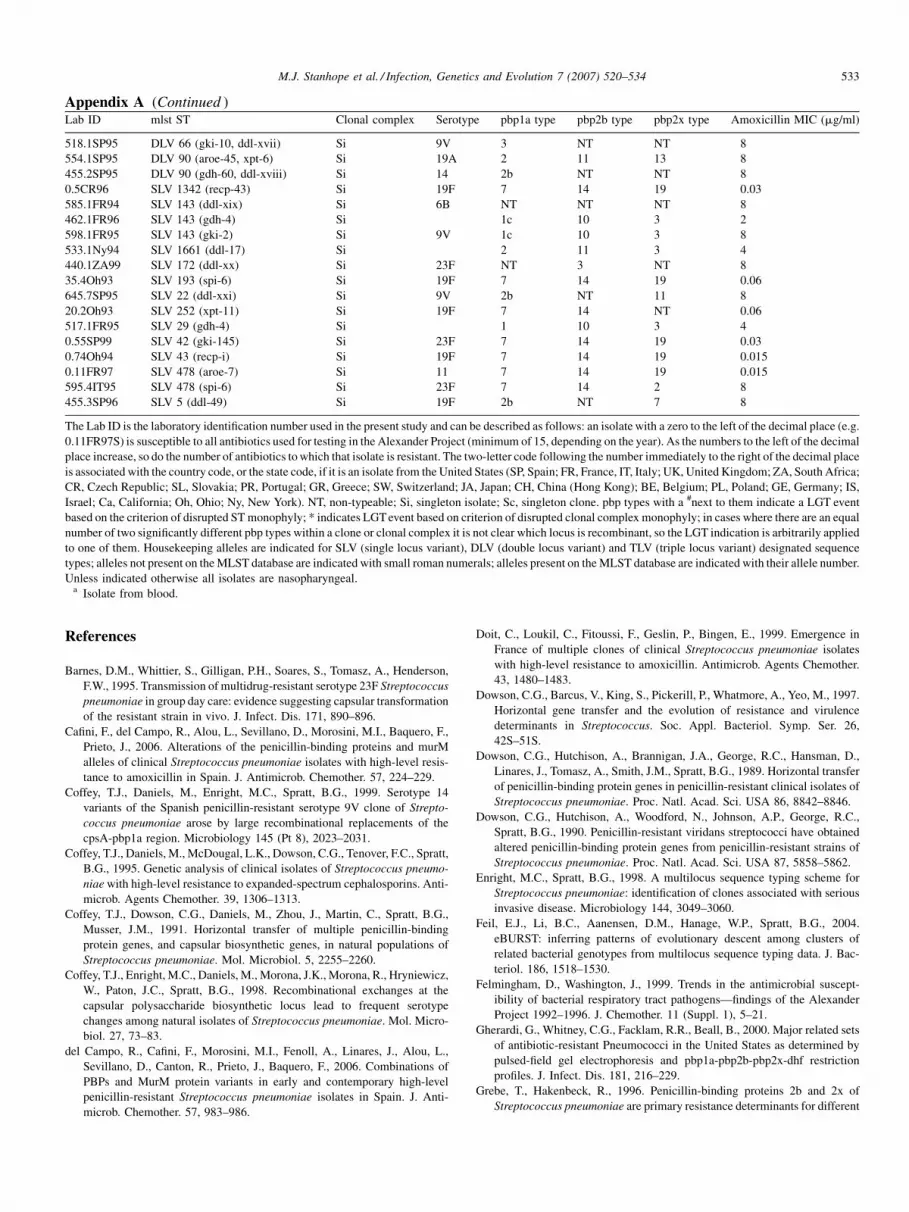
Appendix A. Clonal composition, pbp type, and amoxicillin Mincluded in this study
Lab ID mlst ST Clonal complex Serotyp
525.12Oh00a 13 1
525.14Oh00 13 1
525.4Oh00 13 1
genes, and housekeeping sequences of the various clones
comprising this complex obviously differ, yet their pbp1a
sequences are exactly the same.
It is possible that the reason we detect more examples of
capsular gene transfer than pbp lateral transfer is related to the
strong selection pressure against isolates that recombine any of
these resistance conferring pbps. If this were the case then one
might expect that the relative frequency of capsular switching
versus pbp lateral transfer would not be so extreme for
susceptible isolates. Although phylogenetic trees reconstructed
on just the susceptible pbps tend not to have a great deal of
support on many of the nodes, there are nonetheless several
clades that are strongly supported with either very limited or no
internode branch lengths and these tend to correspond with ST
and clonal complex, suggesting very little pbp lateral transfer
even amongst these susceptible isolates. For these same groups
there are multiple examples of serotype switching. For
example, in this data set there are 7 STs that are comprised
of more than one isolate, are serotyped, and are amoxicillin
susceptible. Within this set of 7 STs, including a total of 19
isolates, there are eight examples of capsular switching and two
examples of LGT for pbp1a, two for pbp2b, and two for pbp2x.
In summary, our results suggest that clonal dissemination is
playing a more significant role than is intraspecific lateral gene
transfer in the development and spread of amoxicillin resistance
in S. pneumoniae. Nonetheless, intraspecific lateral gene transfer
of pbps is occurring and has played a role in the spread of
resistance. The relative frequency of these transfer events
appears to be greatest for pbp2x and 2b, followed by 1a. Serotype
switching appears to be much more frequent than pbp LGT, in
this pre-vaccine era dataset (all isolates in the current study
collected prior to PCV-7 vaccine introduction), implying that S.
pneumoniae might relatively easily escape conjugatevaccines. In
this regard we would certainly concur with the suggestions of
others (e.g. Spratt and Greenwood, 2000) that current evidence
supporting serotype switching between vaccine serotypes is
likely a reflection of the fact these serotypes are most commonly
found in the nasopharynx of children. However, with develop-
ment of serotype replacement the potential donors of capsular
genes will be increasingly of non-vaccine serotype, potentially
leading to vaccine avoidance. Finally, our results are consistent
with occasional, to relatively frequent, cases of co-transforma-
tion of capsular genes and pbp loci.
Acknowledgements
This work was supported by the Alexander Network, and
Cornell University start-up funds, granted to MJS.
ICs for the 216 Streptococcus pneumoniae isolates
e pbp1a type pbp2b type pbp2x type Amoxicillin MIC (mg/ml)
3c 2b 1 8
3c 2b 1 8
3c 2b 1 8

Appendix A (Continued )Lab ID mlst ST Clonal complex Serotype pbp1a type pbp2b type pbp2x type Amoxicillin MIC (mg/ml)
525.5Oh00a 13 1 3c 2b 1 8
540.2Oh00 SLV 13 (recp-2) 1 3c 2b 5* 8
5.3Oh92 33 2 23F 7 14 19 0.06
215.1Oh00 439 2 7 14 19 <0.015
0.6FR00 62 3 14 7 14 19 0.03
80.1Oh93 62 3 11 7 14 NT# 0.06
365.1SP94S 62 3 11 7 14 19 4
442.1SP95 1760 3 6B 7 14 19 8
275.1Oh93 81 4 23F 1 10 3 0.5
305.1Oh93 81 4 23F 1 10 3 1
520.1Oh00 81 4 1 10 3 2
521a.1FR96 81 4 1 10 3 2
535.1Oh94 81 4 23F 1 10 3 2
643.1CH99 81 4 1 10 3 2
450.1Oh93 81 4 19F 1 10 14# 2
552.1FR95 81 4 1 10 3 4
651.1FR98 81 4 1 10 3 4
705.4CH00 81 4 1 10 3 4
720.1CH00 81 4 1 10 3 4
720.5CH00 81 4 1 10 3 4
533.1SP95 81 4 1 12# 3 4
615.1Oh00 81 4 1 10 14 4
615.5Oh00 81 4 1 10 14 4
615.6Oh97 81 4 19F 1 10 14 4
464.1SP95 81 4 14 1 10 3 8
645.8FR95 81 4 6B 1 10 3 8
674.1FR95 81 4 19A 1 10 3 8
670.4FR99 81 4 23F 1 NT 3 8
670.7SL00 81 4 19A 1 NT# 3 8
415.1SP93 81 4 23F 1 5# 4 8
500.1FR95 81 4 3 1 NT# 4 8
688.1CH99 1275 4 1 10 3 4
705.2CH00 1275 4 1 10 3 4
498.1SP95 1623 4 6B 1 10 NT* 8
439.1BE96 SLV 1275 (gki-10) 4 1 10 3 2
522a.1FR94 SLV 1623 (xpt-4) 4 1 10 3 2
599.1FR95 SLV 81 (aroe-95) 4 23F 1 10 3 8
595.1FR00 SLV 81 (ddl-i) 4 14 1 3* NT* 8
645.9FR98 SLV 81 (ddl-ii) 4 23F 1 5* NT* 16
705.3CH00 SLV 81 (gdh-5) 4 1 10 15* 4
553.1FR95 SLV 81 (gdh-i) 4 19A 1 10 3 8
486.1FR95 SLV 81 (gki-10) 4 1 10 3 2
393.2FR95 SLV 81 (gki-10) 4 23F 1 10 3 8
498.3SP95 SLV 81 (gki-10) 4 21 1 10 3 8
601.1FR95 SLV 81 (gki-10) 4 23F 1 10 3 8
599.3FR95 SLV 81 (recp-15) 4 23F 1 10 3 8
674.3FR95 SLV 81(gki-4) 4 9V 1 10 3 8
460.1SW00a 90 5 14 2 4 11 4
670.10SP94 90 5 9V 2 5 12 8
670.9SP94 90 5 6B 2 5 12 8
685.3JA00 90 5 2 NT# 13 8
445.1SP95 90 5 9V 2 NT# 8b# 8
464.2SP95 90 5 6B 2 NT# 8b 8
590.7SP97 1624 5 23F 3 1 7 8
575.1SP95 1624 5 23F 3 1b 7 8
590.6SP97 1624 5 11 3 1b 7 8
590.1FR98 1624 5 6B 3* 1 7 16
595.6SP00 1624 5 6B 7# 14# 19# 16
605.1FR93 DLV 90 (gki-i, ddl-iii) 5 13 2 2 7 8
690.1FR98 SLV 90 (ddl-iv) 5 6B 2 NT NT* 8
605.2FR96 SLV 90 (ddl-v) 5 9V 2 2* 7 8
670.3FR99 SLV 90 (ddl-v) 5 6B 2 2 7 8
670.5FR99 SLV 90 (ddl-v) 5 6B 2 2 7 8
665.1FR97 SLV 90 (ddl-v) 5 19F 2 2 3b# 8
655.3FR97 SLV 90 (ddl-v) 5 19F 2 2 NT# 8
M.J. Stanhope et al. / Infection, Genetics and Evolution 7 (2007) 520–534530

Appendix A (Continued )Lab ID mlst ST Clonal complex Serotype pbp1a type pbp2b type pbp2x type Amoxicillin MIC (mg/ml)
640.4FR98a SLV 90 (ddl-v) 5 6B 2 2 7 16
530.2SP93 SLV 90 (ddl-vi) 5 6B 2 5 NT* 8
670.1FR94 SLV 90 (ddl-vii) 5 14 2 3b* 10* 8
670.2FR94 SLV 90 (ddl-vii) 5 14 2 3b 10 8
553.1SP95 SLV 90 (gki-10) 5 9A NT* NT* 3 8
674.1SP95 SLV 90 (gki-2) 5 23F 2 10* 13 8
0.53SP96 113 6 6B 7 14 19 0.03
0.71Oh93 113 6 18C 7 14 19 0.06
0a 121 6 7 14 19 0.03
0.4BE99 SLV 113 (spi-i) 6 18C 7 14 19 0.03
606.2FR98 144 7 1 10 3 4
336.1ZA98 156 7 1 10 3 4
393.3FR95 156 7 23F 1 10 3 8
393.3SP95 156 7 14 1 10 3 8
393.4FR95 156 7 23F 1 10 3 8
393.4SP95 156 7 9V 1 10 3 8
393.5SP95 156 7 9V 1 10 3 8
393.6SP95 156 7 6B 1 10 3 8
405.16SW99 156 7 14 1b NT# NT 8
380.3IS99 156 7 14 1b# 10 3 8
405.7PR00a 156 7 14 2b# NT# 9# 16
405.3BE99 156 7 23F 3# 4c# 7b# 16
0h 162 7 7 14 19* 0.015
0i 162 7 7 14 19 0.015
0g 162 7 7* 14 19 0.015
347.1SP95 333 7 14 NT 10 NT* 8
365.2SP96 838 7 6A 1 1* 3b 4
380.9SP94 838 7 9V 1 1b 6* 8
355.1SP96 838 7 14 1 1b 19 8
380.11SP95 838 7 6B 1 1 3b 8
380.16SP99 838 7 9A 1 1 3b 8
380.12SP96 838 7 19F 1 1b 3b 8
380.13SP97 838 7 1 1b 3b 8
380.15SP97 838 7 9A 1 1b 3b 8
405.10SP94 838 7 6B 1 1b 3b 8
405.11SP94 838 7 9A 1 1b 3b 8
405.13SP94 838 7 3 1 1b 3b 8
405.14SP95 838 7 14 1 1b 3b 8
335.2SP99 838 7 9A 1 1c 3b 8
335.3SP99a 838 7 9A 1 1c 3b 8
380.10SP95 838 7 9V 1 1 3c 8
335.1SP99 838 7 9V 1 1b 3b 16
380.1FR00a 838 7 19A 1 1b 3b 16
615.4Oh00 DLV 156 (gdh-4, gki-2) 7 1 10 3 4
685.2FR99 DLV 156 (gki-ii, ddl-119) 7 14 2b* 9* 2 8
530.1FR96 SLV 156 (ddl-119) 7 4 2b 9 7* 8
405.1BE96 SLV 156 (ddl-22) 7 23F NT 9 7b 8
347.2SP95 SLV 156 (ddl-4) 7 3 1b# 12* NT 8
435.1SP95 SLV 156 (ddl-4) 7 23F 2b NT# 15* 8
405.15SP95 SLV 156 (ddl-47) 7 23F 2b 6* 15 8
405.5FR94 SLV 156 (ddl-ix) 7 6B NT NT* 8* 8
380.8SP00 SLV 156 (ddl-viii) 7 14 1b 4b* 6 8
405.9PR99 SLV 156 (ddl-x) 7 18C 2b NT* 9 8
640.2FR94 SLV 156 (ddl-xi) 7 14 2b 4 5* 8
640.3FR96 SLV 156 (ddl-xi) 7 19F 2b 4 5 8
640.5IS99 SLV 156 (ddl-xi) 7 19F 2b 4 5 8
489.1Ca99 SLV 156 (gdh-16) 7 1 10 3 4
405.2BE97 SLV 156 (gdh-8) 7 6B 1b NT 7b 8
327.1FR95 SLV 156 (gki-1) 7 1 10 3 2
393.1FR95 SLV 156 (gki-1) 7 23F 1 10 3 8
336.1UK94 SLV 156 (gki-2) 7 1 10 3 4
393.2SP95 SLV 156 (gki-2) 7 3 1 10 3 8
348.1SP95 SLV 156 (gki-29) 7 11 1 10 3 8
393.1SP95 SLV 156 (gki-7) 7 14 1 10 3 8
405.4FR94 SLV 156 (spi-ii) 7 14 2b NT* NT 8
M.J. Stanhope et al. / Infection, Genetics and Evolution 7 (2007) 520–534 531
Appendix A (Continued )Lab ID mlst ST Clonal complex Serotype pbp1a type pbp2b type pbp2x type Amoxicillin MIC (mg/ml)
595.2FR99a SLV 156 (ddl-157) 7 14 1b NT* NT* 8
685.6PL97 173 8 23F 5 7 NT# 8
675.1PL96 173 8 23F NT NT# 16b 8
660.1PL96 272 8 19A 5 7 16 4
685.7PL98 272 8 23F 5 7 16b 16
685.4PL00 272 8 6A 5 7 NT# 16
635.1PL97 1506 8 19F 5 NT NT 8
650.1PL96 2140 8 23F 5 NT* NT* 8
675.2PL97 SLV 173 (ddl-xii) 8 19F 5 7 NT* 8
0.10FR96 180 9 14 7 14 19 0.015
0.21IT95 180 9 23F 7 14 19 0.03
0.36PL98 180 9 3 7 14 19 0.03
0.73Oh93 180 9 3 7 14 19 0.06
630.1SP00 SLV 180 (gdh-11) 9 6B 1b* 4b* 6* 8
0.2BE96 191 10 9V 7 14 19 0.03
0.34PL96 SLV 191 (aroe-i) 10 3 7 14 19 0.03
0.70Oh92 199 11 15 7 14 19 0.06
30.1Oh92 199 11 19A 7 13 18 0.12
60.1Oh92 199 11 19A 7 13 18 0.12
0.82Oh97 199 11 19F NT 13 NT 0.015
0f 419 11 7 14 19 0.03
0.37PR00 232 12 3 7 14 19 0.03
0.57SW99a 232 12 3 7 14 19 0.03
0.22IT96 SLV 232 (ddl-22) 12 3 7 14 19 0.015
105.1Oh93 SLV 37 (ddl-14) 13 23F NT NT NT 0.5
175.1Oh00 SLV 37 (ddl-28) 13 7 14 19 0.03
5.4OH93 SLV 37 (ddl-xiii) 13 23F 7 14 19 0.06
510.1SP94 17 Sc 6B 2b 6 NT 2
655.1SP97 17 Sc 6B 2b 6 8 8
655.2SP97 17 Sc 6A 2b 6 8 8
670.8SP00 17 Sc 8 2b 6 4b 8
685.8SP99a 17 Sc 14 2b 6 4b 8
0.51SP95 260 Sc 23F 7 14 19 0.015
0.49SP94 260 Sc 6B 7 14 19 0.03
595.5IT97 321 Sc 6A 6 8 17 8
640.6IT98 321 Sc 23F 6 8 17 16
0.20IT00 1766 Sc 23F 7 14 19 0.03
0.92FR98 1766 Sc 31 7 14 19 0.03
600.3CR96 TLV 268 (gdh-ii, spi-iii, ddl-xiv) Sc 6B 4 6b 7 8
550.1CR96 TLV 268 (gdh-ii, spi-iii, ddl-xiv) Sc 19A 4 6b 16 8
600.1CR96 TLV 268 (gdh-ii, spi-iii, ddl-xiv) Sc 19A 4 6b 16 8
600.2CR96 TLV 268 (gdh-ii, spi-iii, ddl-xiv) Sc 19A 4 6b 16 8
645.1SL00 TLV 268 (gdh-ii, spi-iii, ddl-xiv) Sc 19A 4 6b 16 8
645.2SL00 TLV 268 (gdh-ii, spi-iii, ddl-xiv) Sc 14 4 6b 16 8
0.47SP00 53 Si 6B 3 1b 7 0.03
0.9FR95 61 Si 18C 7 14 19 0.015
0.8FR94 72 Si 24FA 7 14 19 0.03
0.44SL98 124 Si 6A 7 14 19 0.03
0c 138 Si 7 14 19 0.03
380.5PR00 176 Si 4 7 14 19 8
0.14GE00 177 Si 19F 7 14 NT 0.03
320.1SP97 229 Si 19F NT NT NT 8
560.1JA98 242 Si NT NT NT 4
0.7FR93 280 Si 6B 7 14 19 0.03
0.13FR99 306 Si 23F 7 14 19 0.03
0.16GR98 327 Si 6A 7 14 19 0.03
90.1Oh00a 460 Si 7 14 18 0.06
0.17GR99 574 Si 4 7 14 19 0.03
30.2Oh93 626 Si 23F 7 NT NT 0.06
0.56SW00 681 Si 7 14 19 0.03
0.68Oh92 1390 Si 6A 7 14 19 0.06
0.42SL00 1769 Si 9V 7 14 19 0.03
0.35PL97 2046 Si 7 14 19 0.03
405.8PR98 DLV 1043 (gdh-136, ddl-xv) Si 14 2b NT 7b 8
380.7SL99 DLV 2025 (gki-iii, ddl-xvi) Si 19A 3b NT NT 8
M.J. Stanhope et al. / Infection, Genetics and Evolution 7 (2007) 520–534532
Appendix A (Continued )Lab ID mlst ST Clonal complex Serotype pbp1a type pbp2b type pbp2x type Amoxicillin MIC (mg/ml)
518.1SP95 DLV 66 (gki-10, ddl-xvii) Si 9V 3 NT NT 8
554.1SP95 DLV 90 (aroe-45, xpt-6) Si 19A 2 11 13 8
455.2SP95 DLV 90 (gdh-60, ddl-xviii) Si 14 2b NT NT 8
0.5CR96 SLV 1342 (recp-43) Si 19F 7 14 19 0.03
585.1FR94 SLV 143 (ddl-xix) Si 6B NT NT NT 8
462.1FR96 SLV 143 (gdh-4) Si 1c 10 3 2
598.1FR95 SLV 143 (gki-2) Si 9V 1c 10 3 8
533.1Ny94 SLV 1661 (ddl-17) Si 2 11 3 4
440.1ZA99 SLV 172 (ddl-xx) Si 23F NT 3 NT 8
35.4Oh93 SLV 193 (spi-6) Si 19F 7 14 19 0.06
645.7SP95 SLV 22 (ddl-xxi) Si 9V 2b NT 11 8
20.2Oh93 SLV 252 (xpt-11) Si 19F 7 14 NT 0.06
517.1FR95 SLV 29 (gdh-4) Si 1 10 3 4
0.55SP99 SLV 42 (gki-145) Si 23F 7 14 19 0.03
0.74Oh94 SLV 43 (recp-i) Si 19F 7 14 19 0.015
0.11FR97 SLV 478 (aroe-7) Si 11 7 14 19 0.015
595.4IT95 SLV 478 (spi-6) Si 23F 7 14 2 8
455.3SP96 SLV 5 (ddl-49) Si 19F 2b NT 7 8
The Lab ID is the laboratory identification number used in the present study and can be described as follows: an isolate with a zero to the left of the decimal place (e.g.
0.11FR97S) is susceptible to all antibiotics used for testing in the Alexander Project (minimum of 15, depending on the year). As the numbers to the left of the decimal
place increase, so do the number of antibiotics to which that isolate is resistant. The two-letter code following the number immediately to the right of the decimal place
is associated with the country code, or the state code, if it is an isolate from the United States (SP, Spain; FR, France, IT, Italy; UK, United Kingdom; ZA, South Africa;
CR, Czech Republic; SL, Slovakia; PR, Portugal; GR, Greece; SW, Switzerland; JA, Japan; CH, China (Hong Kong); BE, Belgium; PL, Poland; GE, Germany; IS,
Israel; Ca, California; Oh, Ohio; Ny, New York). NT, non-typeable; Si, singleton isolate; Sc, singleton clone. pbp types with a #next to them indicate a LGT event
based on the criterion of disrupted ST monophyly; * indicates LGT event based on criterion of disrupted clonal complex monophyly; in cases where there are an equal
number of two significantly different pbp types within a clone or clonal complex it is not clear which locus is recombinant, so the LGT indication is arbitrarily applied
to one of them. Housekeeping alleles are indicated for SLV (single locus variant), DLV (double locus variant) and TLV (triple locus variant) designated sequence
types; alleles not present on the MLST database are indicated with small roman numerals; alleles present on the MLST database are indicated with their allele number.
Unless indicated otherwise all isolates are nasopharyngeal.a Isolate from blood.
M.J. Stanhope et al. / Infection, Genetics and Evolution 7 (2007) 520–534 533
References
Barnes, D.M., Whittier, S., Gilligan, P.H., Soares, S., Tomasz, A., Henderson,
F.W., 1995. Transmission of multidrug-resistant serotype 23F Streptococcus
pneumoniae in group day care: evidence suggesting capsular transformation
of the resistant strain in vivo. J. Infect. Dis. 171, 890–896.
Cafini, F., del Campo, R., Alou, L., Sevillano, D., Morosini, M.I., Baquero, F.,
Prieto, J., 2006. Alterations of the penicillin-binding proteins and murM
alleles of clinical Streptococcus pneumoniae isolates with high-level resis-
tance to amoxicillin in Spain. J. Antimicrob. Chemother. 57, 224–229.
Coffey, T.J., Daniels, M., Enright, M.C., Spratt, B.G., 1999. Serotype 14
variants of the Spanish penicillin-resistant serotype 9V clone of Strepto-
coccus pneumoniae arose by large recombinational replacements of the
cpsA-pbp1a region. Microbiology 145 (Pt 8), 2023–2031.
Coffey, T.J., Daniels, M., McDougal, L.K., Dowson, C.G., Tenover, F.C., Spratt,
B.G., 1995. Genetic analysis of clinical isolates of Streptococcus pneumo-
niae with high-level resistance to expanded-spectrum cephalosporins. Anti-
microb. Agents Chemother. 39, 1306–1313.
Coffey, T.J., Dowson, C.G., Daniels, M., Zhou, J., Martin, C., Spratt, B.G.,
Musser, J.M., 1991. Horizontal transfer of multiple penicillin-binding
protein genes, and capsular biosynthetic genes, in natural populations of
Streptococcus pneumoniae. Mol. Microbiol. 5, 2255–2260.
Coffey, T.J., Enright, M.C., Daniels, M., Morona, J.K., Morona, R., Hryniewicz,
W., Paton, J.C., Spratt, B.G., 1998. Recombinational exchanges at the
capsular polysaccharide biosynthetic locus lead to frequent serotype
changes among natural isolates of Streptococcus pneumoniae. Mol. Micro-
biol. 27, 73–83.
del Campo, R., Cafini, F., Morosini, M.I., Fenoll, A., Linares, J., Alou, L.,
Sevillano, D., Canton, R., Prieto, J., Baquero, F., 2006. Combinations of
PBPs and MurM protein variants in early and contemporary high-level
penicillin-resistant Streptococcus pneumoniae isolates in Spain. J. Anti-
microb. Chemother. 57, 983–986.
Doit, C., Loukil, C., Fitoussi, F., Geslin, P., Bingen, E., 1999. Emergence in
France of multiple clones of clinical Streptococcus pneumoniae isolates
with high-level resistance to amoxicillin. Antimicrob. Agents Chemother.
43, 1480–1483.
Dowson, C.G., Barcus, V., King, S., Pickerill, P., Whatmore, A., Yeo, M., 1997.
Horizontal gene transfer and the evolution of resistance and virulence
determinants in Streptococcus. Soc. Appl. Bacteriol. Symp. Ser. 26,
42S–51S.
Dowson, C.G., Hutchison, A., Brannigan, J.A., George, R.C., Hansman, D.,
Linares, J., Tomasz, A., Smith, J.M., Spratt, B.G., 1989. Horizontal transfer
of penicillin-binding protein genes in penicillin-resistant clinical isolates of
Streptococcus pneumoniae. Proc. Natl. Acad. Sci. USA 86, 8842–8846.
Dowson, C.G., Hutchison, A., Woodford, N., Johnson, A.P., George, R.C.,
Spratt, B.G., 1990. Penicillin-resistant viridans streptococci have obtained
altered penicillin-binding protein genes from penicillin-resistant strains of
Streptococcus pneumoniae. Proc. Natl. Acad. Sci. USA 87, 5858–5862.
Enright, M.C., Spratt, B.G., 1998. A multilocus sequence typing scheme for
Streptococcus pneumoniae: identification of clones associated with serious
invasive disease. Microbiology 144, 3049–3060.
Feil, E.J., Li, B.C., Aanensen, D.M., Hanage, W.P., Spratt, B.G., 2004.
eBURST: inferring patterns of evolutionary descent among clusters of
related bacterial genotypes from multilocus sequence typing data. J. Bac-
teriol. 186, 1518–1530.
Felmingham, D., Washington, J., 1999. Trends in the antimicrobial suscept-
ibility of bacterial respiratory tract pathogens—findings of the Alexander
Project 1992–1996. J. Chemother. 11 (Suppl. 1), 5–21.
Gherardi, G., Whitney, C.G., Facklam, R.R., Beall, B., 2000. Major related sets
of antibiotic-resistant Pneumococci in the United States as determined by
pulsed-field gel electrophoresis and pbp1a-pbp2b-pbp2x-dhf restriction
profiles. J. Infect. Dis. 181, 216–229.
Grebe, T., Hakenbeck, R., 1996. Penicillin-binding proteins 2b and 2x of
Streptococcus pneumoniae are primary resistance determinants for different

M.J. Stanhope et al. / Infection, Genetics and Evolution 7 (2007) 520–534534
classes of beta-lactam antibiotics. Antimicrob. Agents Chemother. 40, 829–
834.
Guindon, S., Gascuel, O., 2003. A simple, fast, and accurate algorithm to
estimate large phylogenies by maximum likelihood. Syst. Biol. 52, 696–704.
Hakenbeck, R., Grebe, T., Zahner, D., Stock, J.B., 1999. Beta-lactam resistance
in Streptococcus pneumoniae: penicillin-binding proteins and non-penicil-
lin-binding proteins. Mol. Microbiol. 33, 673–678.
Jacobs, M.R., Felmingham, D., Appelbaum, P.C., Gruneberg, R.N., 2003. The
Alexander Project 1998–2000: susceptibility of pathogens isolated from
community-acquired respiratory tract infection to commonly used antimi-
crobial agents. J. Antimicrob. Chemother. 52, 229–246.
Klugman, K.P., 2002. The successful clone: the vector of dissemination of
resistance in Streptococcus pneumoniae. J. Antimicrob. Chemother. 50
(Suppl. S2), 1–5.
Kosowska, K., Jacobs, M.R., Bajaksouzian, S., Koeth, L., Appelbaum, P.C.,
2004. Alterations of penicillin-binding proteins 1A, 2X, and 2B in Strep-
tococcus pneumoniae isolates for which amoxicillin MICs are higher than
penicillin MICs. Antimicrob. Agents Chemother. 48, 4020–4022.
Majewski, J., Zawadzki, P., Pickerill, P., Cohan, F.M., Dowson, C.G., 2000.
Barriers to genetic exchange between bacterial species: Streptococcus
pneumoniae transformation. J. Bacteriol. 182, 1016–1023.
McCormick, A.W., Whitney, C.G., Farley, M.M., Lynfield, R., Harrison, L.H.,
Bennett, N.M., Schaffner, W., Reingold, A., Hadler, J., Cieslak, P., Samore,
M.H., Lipsitch, M., 2003. Geographic diversity and temporal trends of
antimicrobial resistance in Streptococcus pneumoniae in the United States.
Nat. Med. 9, 424–430.
McGee, L., McDougal, L., Zhou, J., Spratt, B.G., Tenover, F.C., George, R.,
Hakenbeck, R., Hryniewicz, W., Lefevre, J.C., Tomasz, A., Klugman, K.P.,
2001. Nomenclature of major antimicrobial-resistant clones of Streptococ-
cus pneumoniae defined by the pneumococcal molecular epidemiology
network. J. Clin. Microbiol. 39, 2565–2571.
Nesin, M., Ramirez, M., Tomasz, A., 1998. Capsular transformation of a
multidrug-resistant Streptococcus pneumoniae in vivo. J. Infect. Dis.
177, 707–713.
Perez-Trallero, E., Marimon, J.M., Gonzalez, A., Garcia-Rey, C., Aguilar, L.,
2003. Genetic relatedness of recently collected Spanish respiratory tract
Streptococcus pneumoniae isolates with reduced susceptibility to amox-
icillin. Antimicrob. Agents Chemother. 47, 3637–3639.
Posada, D., Crandall, K.A., 1998. MODELTEST: testing the model of DNA
substitution. Bioinformatics 14, 817–818.
Potgieter, E., Chalkley, L.J., 1995. Relatedness among penicillin-binding
protein 2b genes of Streptococcus mitis, Streptococcus oralis, and Strepto-
coccus pneumoniae. Microb. Drug Resist. 1, 35–42.
Ramirez, M., Tomasz, A., 1999. Acquisition of new capsular genes among
clinical isolates of antibiotic-resistant Streptococcus pneumoniae. Microb.
Drug Resist. 5, 241–246.
Reichmann, P., Konig, A., Linares, J., Alcaide, F., Tenover, F.C., McDougal, L.,
Swidsinski, S., Hakenbeck, R., 1997. A global gene pool for high-level
cephalosporin resistance in commensal Streptococcus species and Strepto-
coccus pneumoniae. J. Infect. Dis. 176, 1001–1012.
Sa-Leao, R., Tomasz, A., de Lencastre, H., 2001. Multilocus sequence typing of
Streptococcus pneumoniae clones with unusual drug resistance patterns:
genetic backgrounds and relatedness to other epidemic clones. J. Infect. Dis.
184, 1206–1210.
Sa-Leao, R., Vilhelmsson, S.E., de Lencastre, H., Kristinsson, K.G., Tomasz,
A., 2002. Diversity of penicillin-nonsusceptible Streptococcus pneumoniae
circulating in Iceland after the introduction of penicillin-resistant clone
Spain(6B)-2. J. Infect. Dis. 186, 966–975.
Shimodaira, H., 2002. An approximately unbiased test of phylogenetic tree
selection. Syst. Biol. 51, 492–508.
Shimodaira, H., Hasegawa, M., 2001. CONSEL: for assessing the confidence of
phylogenetic tree selection. Bioinformatics 17, 1246–1247.
Spratt, B.G., 2004. Exploring the concept of clonality in bacteria. Meth. Mol.
Biol. 266, 323–352.
Spratt, B.G., Greenwood, B.M., 2000. Prevention of pneumococcal disease by
vaccination: does serotype replacement matter? Lancet 356, 1210–1211.
Spratt, B.G., Hanage, W.P., Brueggemann, A.B., 2004. Evolutionary and
population biology of Streptococcus pneumoniae. In: Tuomanen , E.I.,
Mitchell, T.J., Morrison, D.A., Spratt, B.G. (Eds.), The Pneumococcus.
ASM Press, Washington, DC, pp. 119–136.
Stanhope, M.J., Walsh, S.L., Becker, J.A., Italia, M.J., Ingraham, K.A., Gwynn,
M.N., Mathie, T., Poupard, J.A., Miller, L.A., Brown, J.R., Amrine-Madsen,
H., 2005. Molecular evolution perspectives on intraspecific lateral DNA
transfer of topoisomerase and gyrase loci in Streptococcus pneumoniae,
with implications for fluoroquinolone resistance development and spread.
Antimicrob. Agents Chemother. 49, 4315–4326.
Tettelin, H., Nelson, K.E., Paulsen, I.T., Eisen, J.A., Read, T.D., Peterson, S.,
Heidelberg, J., DeBoy, R.T., Haft, D.H., Dodson, R.J., Durkin, A.S., Gwinn,
M., Kolonay, J.F., Nelson, W.C., Peterson, J.D., Umayam, L.A., White, O.,
Salzberg, S.L., Lewis, M.R., Radune, D., Holtzapple, E., Khouri, H., Wolf,
A.M., Utterback, T.R., Hansen, C.L., McDonald, L.A., Feldblyum, T.V.,
Angiuoli, S., Dickinson, T., Hickey, E.K., Holt, I.E., Loftus, B.J., Yang, F.,
Smith, H.O., Venter, J.C., Dougherty, B.A., Morrison, D.A., Hollingshead,
S.K., Fraser, C.M., 2001. Complete genome sequence of a virulent isolate of
Streptococcus pneumoniae. Science 293, 498–506.
Thompson, J.D., Higgins, D.G., Gibson, T.J., 1994. CLUSTAL W: improving
the sensitivity of progressive multiple sequence alignment through sequence
weighting, position-specific gap penalties and weight matrix choice. Nucl.
Acids Res. 22, 4673–4680.
Trzcinski, K., Thompson, C.M., Lipsitch, M., 2004. Single-step capsular
transformation and acquisition of penicillin resistance in Streptococcus
pneumoniae. J. Bacteriol. 186, 3447–3452.
White, A.R., Kaye, C., Poupard, J., Pypstra, R., Woodnutt, G., Wynne, B., 2004.
Augmentin (amoxicillin/clavulanate) in the treatment of community-
acquired respiratory tract infection: a review of the continuing development
of an innovative antimicrobial agent. J. Antimicrob. Chemother. 53 (Suppl.
1), i3–i20.
Yother, J., 2004. Capsules. In: Tuomanen, E.I., Mitchell, T.J., Morrison, D.A.,
Spratt, B.G. (Eds.), The Pneumococcus. ASM Press, Washington, DC, pp.
30–49.























![Câu 36. Giá trị lớn nhất của hàm số y=log,(2x+1) trên [0;1] là](https://img.dokumen.tips/doc/110x75/63142c7015106505030b3a71/cau-36-gia-tri-lon-nhat-cua-ham-so-ylog2x1-tren-01-la.jpg)





