Embed Size (px)
Citation preview

Sex Plant Reprod (1993) 6:176-182 Sexual Plant Reproduction �9 Springer-Verlag 1993
Abortion and determination of stages for embryo rescue in crosses between sweet-potato, Ipomoea batatas Lam. ( 2 n = 6 x - - 9 0 ) and its wild relative, L trifida (H. B. K.) G. Don. (2n= 2x = 30) Jos~ Mont, Masaru Iwanaga*, Gisella Orjeda, and Kazuo Watanabe**
International Potato Center (CIP), P.O. Box 5969, Lima, Peru
Summary. The frequency of aborted fruits and the changes and abnormalities that occur during the embryo development in intraspecific crosses of sweet-potato Ipo- moea batatas (2n = 6x = 90) and interspecific crosses be- tween I. batatas and I. trifida (2n = 2x = 30) were investi- gated in order to study the causes of the low seed produc- tion. Three genotypes of L batatas and 18 genotypes of L trifida were intermated. The frequency of aborted fruits was below 25% in the intraspecific crosses and over 90% in the interspecific crosses. Paraffin sections were used to examine the developmental stages of fruits and seeds. Embryos in different developmental stages were observed to determine the stage of abortion. These observations permitted the identification of developmen- tal stages of embryo rescue in interspecific crosses. There were no significant differences in the frequency of em- bryo abortion before the early globular stage among female sweet-potato progenitors for the intraspecific and interspecific crosses. The frequency of the late occur- rence of embryo abortion (when embryo abortion occurs after the pre-globular stage) was higher in interspecific crosses (19.1%) than in intraspecific crosses (5.5%). The frequency of the late occurrence of embryo abortion in interspecific crosses was higher at the globular stage (9.6%) than at the heart stage (4.3%). Providing that embryo rescue is conducted in interspecific crosses, the estimated number of potentially viable embryos could be increased: 30 times with embryos at the globular stage; 20 times with embryos at the heart stage; and 11 times if embryos at the torpedo stage were used for the rescue with respect to the seed set. The results sug- gested that the appropriate time for embryo rescue in interspecific crosses is at the globular stage. If embryos
* Present address: International Plant Genetic Resources Institute, C/O FAO United Nations, Via delle Sette Chiese 142, 00145 Rome, Italy; this research was initiated during sabbatical of M.I. at the Asian Vegetable Research and Development Center (AVRDC) in Taiwan ** Present address: 252 Emerson Hall, Cornell University, Ithaca, NY 14853-1902, USA.
Correspondence to: K. Watanabe
could be rescued at the globular stage, it would be possi- ble to increase the number of surviving embryos up to 30 times in interspecific crosses and 0.02 times in intrasp- ecific crosses with respect to natural conditions without embryo rescue.
Key words: Embryo abortion Interspecific crosses - Interploidy crosses - Ipomoea batatas - Ipomoea trifida
Introduction
The sweet-potato ovary is bicarpelar and bilocular. There are two ovules in each cavity, and they are sepa- rated by a pseudoseptum. With normal fertilization and embryogenesis, four seeds can be expected. Kokubu et al. (1982) reported that even in intraspecific compati- ble crosses in which a large number of pollen grains germinate on the stigma, the seed set per pollination is usually low (one seed/capsule). Consequently, the low production of botanical seed is considered to be one of the bottlenecks for breeding.
Several causes such as absence of egg fecundity, in- compatibility, sterile ovules and embryonic imbalances have been suggested as the main factors that lower seed production (Nishiyama and Inomata 1966; Jones and Kobayashi 1969; Kokubu et al. 1982). Moreover, it ap- pears that embryonic problems in higher plants occur with greater frequency at the heart stage of embryo de- velopment (Phillips et al. 1982; Sharma 1980; Smith and Desborough 1986).
Low seed set is a constraint to obtaining enough progeny in sweet-potato breeding programs. This prob- lem is especially acute in interspecific crosses (Freyre et al. 1991 ; Kobayashi 1978, 1981). In other genera, low set seed was also found in interploidy crosses (Ahn and Hartmann 1978; Cooper and Brink 1945; Blakeslee et al. 1923).
There are two alternative methods for obtaining more seeds: (1) increasing the number of pollinations and (2)
rescuing the e m b r y o s at a p p r o p r i a t e stages. The logist ics for the first a l t e rna t ive are m o r e c o m p l i c a t e d than the second. The second a l t e rna t ive requi res i m p r o v e d tech- no logy a n d i n f o r m a t i o n on e m b r y o rescue, on which ava i lab le knowledge is l imi ted.
We s tud ied the f requency o f a b o r t e d frui ts a n d abno r - mal i t ies occur r ing du r ing e m b r y o d e v e l o p m e n t to iden- tify the m o s t a p p r o p r i a t e s tage for e m b r y o rescue.
Materials and methods
Three genotypes of hexaploid Ipomoea batatas (RCB2117H, DLP627 and EEC251) from collection of the International Potato Center (CIP) and 18 genotypes of diploid L trifida from the Kyushu National Agricultural Experimental Station (KNAES), Japan, were used. The L batata accessions were selected, based on their high flowering ability and self-incompatibility and the L trifida acces- sions because of their compatibility with L batatas and high pollen stainability percentages (Orjeda et al. 1991).
Frui t abort ion
This experiment was carried out from January to March (the sum- mer season in Peru) in 1988 and 1989. Flowers were induced by using short day treatment. Hand-pollination was carried out in the morning from 8 to 10 o'clock under greenhouse conditions. About 100 flowers of the three L batatas genotypes were pollinated daily. During the first 3 days of every week 2 x L trifida was used as male, and during the next 2 days L batatas was used. The intraspecific crosses were used as checks. All aborted fruits were collected daily for calculation of the fruit abortion frequency.
E m b r y o l o g i c a l analysis
Seven fruits were collected every 2 days until the 18th day after pollination (DAP). They were fixed in CRAF V solution (7 parts
177
of 10% acetic acid: 3 parts of 37% formaldehyde: 10 parts of 1% chromic acid). After 24 h, these samples were processed using the standard paraffin-embedding procedure. The samples taken 2 and 4 DAP were sectioned at 10 pro. The samples taken 6-18 DAP were sectioned at 12 gin. When tissues were hard, the sample was put into a beaker with water and safranin (several drops) at 40~ for 24 h (Torres 1985). This procedure was used until the samples were easily sectioned. The sections were mounted and stained using 50% safranin, 1% aqueous crystal violet, and 96% fast green, and were then observed under the microscope. Eleven developmental stages were considered, and the last nine were grouped into three shapes : bicell; preglobular; early globular, glob- ular, late globular stages (for the globular shape); early heart, heart, late heart stages (for the heart shape); and torpedo, mature and late mature stages (for the mature shape).
Results and discussion
Frui t abort ion
To es t imate the f rui t a b o r t i o n f requency, 524 in t raspeci f - ic a n d 805 in terspeci f ic po l l i na t i ons were m a d e (Table 1). The to ta l f ru i t a b o r t i o n percen tages in in t raspeci f ic crosses r anged f rom 1.0% to 47 .3%, and the m e a n was less than 24% at the 34 D A P . In in terspeci f ic crosses, the to ta l pe rcen tages in f ru i t a b o r t i o n r anged f rom 85.7% to 97 .0% wi th an average o f m o r e than 90% at the 34 D A P . Beyond the 4th D A P , a c o m m o n m o d a l p e a k o f f rui t - fa l l in b o t h in t raspeci f ic a n d in terspeci f ic crosses was observed . A n o t h e r m o d a l p e a k o f f rui t - fa l l was obse rved be tween the 8th a n d 12th D A P on ly in the in terspeci f ic crosses. This p e a k di f fered a m o n g fe- males . E m b r y o l o g i c a l ana lyses were p e r f o r m e d to deter- mine the causes o f the b i m o d a l d i s t r ibu t ion .
Table 1. Numbers of pollinations, and cumulative number and percentage of aborted fruits from 2nd to 18th days after pollination (DAP) and at harvest (34 DAP) in intraspecific crosses in Ipomoea batatas (2n = 6x = 90) and in interspecific crosses between L batatas and L trifida (2n = 2x = 30)
DAP Cross combination
RCB2117Ha DLP627 a EEC251 ~ RCB2117H DLP627 EEC251 x DLP627 x EEC251 x RCB2117H • L trifida x L trifida x L trifida
2 16 b (11.0) ~ 2 (1.0) 52 (28.3) 57 (24.0) 0 (0.0) 13 (4.6) 4 19 (13.0) 8 (4.1) 72 (39.1) 73 (30.7) 5 (i.8) 51 (18.1) 6 19 (13.0) 9 (4.7) 81 (44.0) 80 (33.6) 12 (4.2) 73 (25.9) 8 21 (14.4) 10 (5.2) 87 (47.3) 85 (35.7) 21 (7.4) 121 (42.9)
10 21 (14.4) 11 (5.7) 87 (47.3) 112 (47.1) 120 (42.1) 193 (68.4) 12 21 (14.4) 11 (5.7) 87 (47.3) 135 (56.7) 199 (69.8) 245 (86.9) 14 23 (15.7) 11 (5.7) 87 (47.3) 163 (68.5) 229 (80.3) 256 (90.8) 16 23 (15.7) 12 (6.2) 87 (47.3) 187 (78.6) 237 (83.1) 263 (93.3) 18 23 (15.7) 12 (6.2) 87 (47.3) 198 (83.2) 243 (85.2) 264 (93.6) 34 24 (16.4) 12 (6.2) 87 (47.3) 204 (85.7) 255 (89.4) 274 (97.1)
Total no. of 146 194 184 238 285 282 pollinations
524 805
a Ipomoea batatas b Number of aborted fruits ~ Percentage of aborted fruits
178
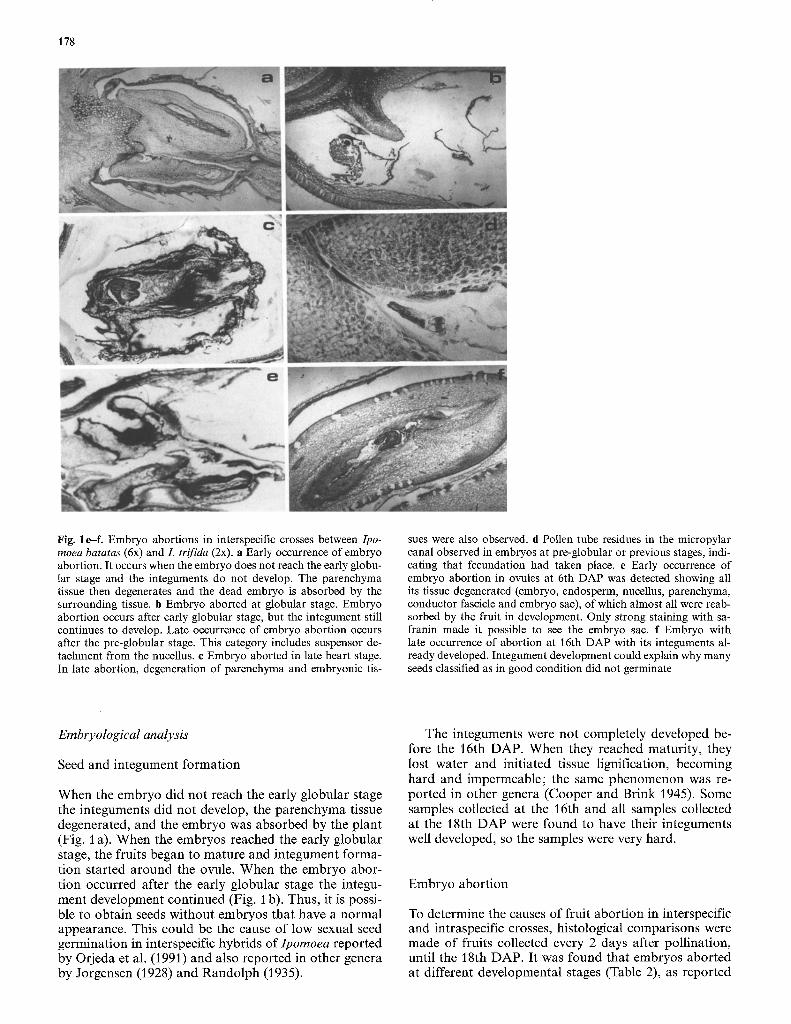
Fig. 1 e-f. Embryo abortions in interspecific crosses between Ipo- moea batatas (6x) and L trifida (2x). a Early occurrence of embryo abortion. It occurs when the embryo does not reach the early globu- lar stage and the integuments do not develop. The parenchyma tissue then degenerates and the dead embryo is absorbed by the surrounding tissue, b Embryo aborted at globular stage. Embryo abortion occurs after early globular stage, but the integument still continues to develop. Late occurrence of embryo abortion occurs after the pre-globular stage. This category includes suspensor de- tachment from the nucellus, c Embryo aborted in late heart stage. In late abortion, degeneration of parenchyma and embryonic tis-
sues were also observed, d Pollen tube residues in the micropylar canal observed in embryos at pre-globular or previous stages, indi- cating that fecundation had taken place, e Early occurrence of embryo abortion in ovules at 6th DAP was detected showing all its tissue degenerated (embryo, endosperm, nucellus, parencbyma, conductor fascicle and embryo sac), of which almost all were reab- sorbed by the fruit in development. Only strong staining with sa- franin made it possible to see the embryo sac. f Embryo with late occurrence of abortion at 16th DAP with its integuments al- ready developed. Integument development could explain why many seeds classified as in good condition did not germinate
Embryological analysis
Seed and integument format ion
When the embryo did not reach the early globular stage the integuments did not develop, the parenchyma tissue degenerated, and the embryo was absorbed by the plant (Fig. 1 a). When the embryos reached the early globular stage, the fruits began to mature and integument forma- tion started around the ovule. When the embryo abor- tion occurred after the early globular stage the integu- ment development continued (Fig. 1 b). Thus, it is possi- ble to obtain seeds without embryos that have a normal appearance. This could be the cause of low sexual seed germination in interspecific hybrids of Ipomoea reported by Orjeda et al. (1991) and also reported in other genera by Jorgensen (1928) and Randolph (1935).
The integuments were not completely developed be- fore the 16th DAP. When they reached maturity, they lost water and initiated tissue lignification, becoming hard and impermeable; the same phenomenon was re- ported in other genera (Cooper and Brink 1945). Some samples collected at the 16th and all samples collected at the 18th DAP were found to have their integuments well developed, so the samples were very hard.
Embryo abort ion
To determine the causes of fruit abort ion in interspecific and intraspecific crosses, histological comparisons were made of fruits collected every 2 days after pollination, until the 18th DAP. It was found that embryos aborted at different developmental stages (Table 2), as reported
179
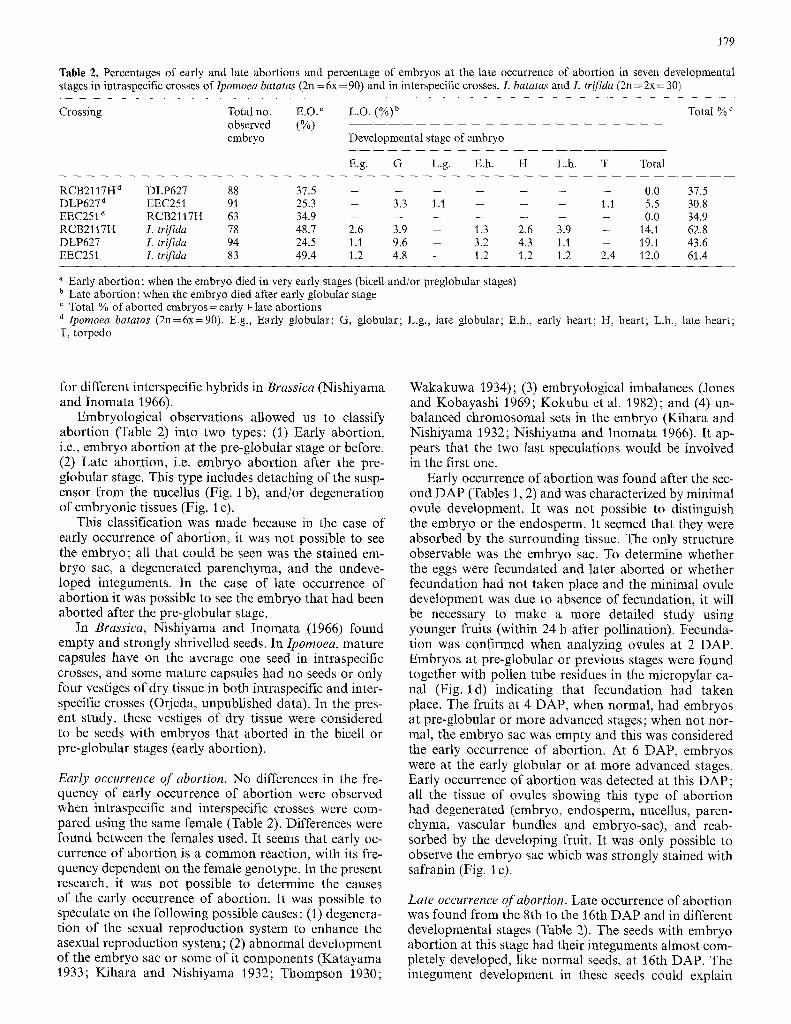
Table 2. Percentages of early and late abortions and percentage of embryos at the late occurrence of abortion in seven developmental stages in intraspecific crosses of Ipomoea batatas (2n = 6x = 90) and in interspecific crosses. L batatas and L trifida (2n = 2x = 30)
Crossing Total no. E.O. ~ L.O. (%)b Total % c observed (%) embryo Developmental stage of embryo
E.g. G L.g. E.h. H L.h. T Total
RCB2117H d DLP627 88 37 .5 . . . . . . . 0.0 37.5 DLP627 a EEC251 91 25.3 - 3.3 1.1 - - - 1.1 5.5 30.8 EEC251 d RCB2117H 63 34 .9 . . . . . . . 0.0 34.9 RCB2117H I. trifida 78 48.7 2.6 3.9 - 1.3 2.6 3.9 - 14,1 62.8 DLP627 L trifida 94 24.5 1.1 9.6 - 3.2 4,3 1.1 - 19,1 43.6 EEC251 I. trifida 83 49.4 1.2 4.8 - 1.2 1.2 1.2 2.4 12.0 61.4
" Early abortion: when the embryo died in very early stages (bicell and/or preglobular stages) b Late abortion: when the embryo died after early globular stage
Total % of aborted embryos = early + late abortions d Ipomoea batatas (2n=6x=90). E.g., Early globular; G, globular; L.g., late globular; E.h., early heart; H, heart; L.h., late heart; T, torpedo
for different interspecific hybrids in Brassica (Nishiyama and Inomata 1966).
Embryological observations allowed us to classify abortion (Table 2) into two types: (1) Early abortion, i.e., embryo abortion at the pre-globular stage or before. (2) Late abortion, i.e. embryo abortion after the pre- globular stage. This type includes detaching of the susp- ensor from the nucellus (Fig. 1 b), and/or degeneration of embryonic tissues (Fig. 1 c).
This classification was made because in the case of early occurrence of abortion, it was not possible to see the embryo; all that could be seen was the stained em- bryo sac, a degenerated parenchyma, and the undeve- loped integuments. In the case of late occurrence of abortion it was possible to see the embryo that had been aborted after the pre-globular stage.
In Brassica, Nishiyama and Inomata (1966) found empty and strongly shrivelled seeds. In Ipomoea, mature capsules have on the average one seed in intraspecific crosses, and some mature capsules had no seeds or only four vestiges of dry tissue in both intraspecific and inter- specific crosses (Orjeda, unpublished data). In the pres- ent study, these vestiges of dry tissue were considered to be seeds with embryos that aborted in the bicell or pre-globular stages (early abortion).
Early occurrence o f abortion. No differences in the fre- quency of early occurrence of abortion were observed when intraspecific and interspecific crosses were com- pared using the same female (Table 2). Differences were found between the females used. It seems that early oc- currence of abortion is a common reaction, with its fre- quency dependent on the female genotype. In the present research, it was not possible to determine the causes of the early occurrence of abortion. It was possible to speculate on the following possible causes: (1) degenera- tion of the sexual reproduction system to enhance the asexual reproduction system; (2) abnormal development of the embryo sac or some of it components (Katayama 1933; Kihara and Nishiyama 1932; Thompson 1930;
Wakakuwa 1934); (3) embryological imbalances (Jones and Kobayashi 1969; Kokubu et al. 1982); and (4) un- balanced chromosomal sets in the embryo (Kihara and Nishiyama 1932; Nishiyama and Inomata 1966). It ap- pears that the two last speculations would be involved in the first one.
Early occurrence of abortion was found after the sec- ond DAP (Tables 1, 2) and was characterized by minimal ovule development. It was not possible to distinguish the embryo or the endosperm. It seemed that they were absorbed by the surrounding tissue. The only structure observable was the embryo sac. To determine whether the eggs were fecundated and later aborted or whether fecundation had not taken place and the minimal ovule development was due to absence of fecundation, it will be necessary to make a more detailed study using younger fruits (within 24 h after pollination). Fecunda- tion was confirmed when analyzing ovules at 2 DAP. Embryos at pre-globular or previous stages were found together with pollen tube residues in the micropylar ca- nal (Fig. l d) indicating that fecundation had taken place. The fruits at 4 DAP, when normal, had embryos at pre-globular or more advanced stages; when not nor- mal, the embryo sac was empty and this was considered the early occurrence of abortion. At 6 DAP, embryos were at the early globular or at more advanced stages. Early occurrence of abortion was detected at this DAP; all the tissue of ovules showing this type of abortion had degenerated (embryo, endosperm, nucellus, paren- chyma, vascular bundles and embryo-sac), and reab- sorbed by the developing fruit. It was only possible to observe the embryo sac which was strongly stained with safranin (Fig. I e).
Late occurrence o f abortion. Late occurrence of abortion was found from the 8th to the 16th DAP and in different developmental stages (Table 2). The seeds with embryo abortion at this stage had their integuments almost com- pletely developed, like normal seeds, at 16th DAP. The integument development in these seeds could explain
180
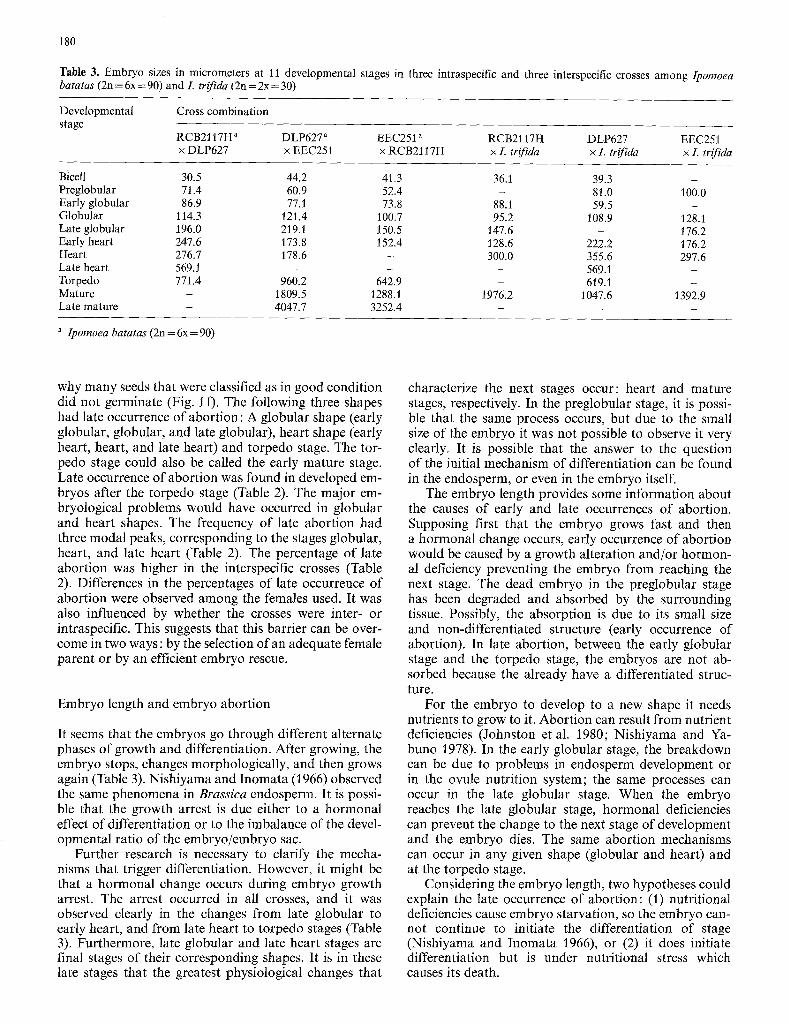
Table 3. Embryo sizes in micrometers at 11 developmental stages in three intraspecific and three interspecific crosses among Ipomoea batatas (2n = 6x = 90) and L trifida (2n = 2x = 30)
Developmental Cross combination stage
RCB2117H" DLP627 a EEC251 " RCB2117H DLP627 EEC251 x DLP627 x EEC251 x RCB2117H x L trifida x L trifida x L trifida
Bicell 30.5 44.2 41.3 36.1 39.3 - Preglobular 71.4 60.9 52.4 - 81.0 100.0 Early globular 86.9 77.1 73.8 88.1 59.5 - Globular 114.3 121.4 I00.7 95.2 108.9 128.1 Late globular 196.0 219. l 150.5 147.6 - 176.2 Early heart 247.6 173.8 152.4 128.6 222.2 176.2 Heart 276.7 178.6 - 300.0 355.6 297.6 Late heart 569.1 - - - 569.1 - Torpedo 771.4 960.2 642.9 - 619.1 - Mature - 1809.5 1288.1 1976.2 1047.6 1392.9 Late mature - 4047.7 3252.4 - - -
" Ipomoea batatas (2n = 6x = 90)
why many seeds that were classified as in good condition did not germinate (Fig. J f). The following three shapes had late occurrence of abort ion: A globular shape (early globular, globular, and late globular), heart shape (early heart, heart, and late heart) and torpedo stage. The tor- pedo stage could also be called the early mature stage. Late occurrence of abortion was found in developed em- bryos after the torpedo stage (Table 2). The major em- bryological problems would have occurred in globular and heart shapes. The frequency of late abortion had three modal peaks, corresponding to the stages globular, heart, and late heart (Table 2). The percentage of late abortion was higher in the interspecific crosses (Table 2). Differences in the percentages of late occurrence of abortion were observed among the females used. It was also influenced by whether the crosses were inter- or intraspecific. This suggests that this barrier can be over- come in two ways: by the selection of an adequate female parent or by an efficient embryo rescue.
Embryo length and embryo abortion
It seems that the embryos go through different alternate phases of growth and differentiation. After growing, the embryo stops, changes morphologically, and then grows again (Table 3). Nishiyama and Inomata (1966) observed the same phenomena in B r a s s i c a endosperm. It is possi- ble that the growth arrest is due either to a hormonal effect of differentiation or to the imbalance of the devel- opmental ratio o f the embryo/embryo sac.
Further research is necessary to clarify the mecha- nisms that trigger differentiation. However, it might be that a hormonal change occurs during embryo growth arrest. The arrest occurred in all crosses, and it was observed clearly in the changes from late globular to early heart, and from late heart to torpedo stages (Table 3). Furthermore, late globular and late heart stages are final stages of their corresponding shapes. It is in these late stages that the greatest physiological changes that
characterize the next stages occur: heart and mature stages, respectively. In the preglobular stage, it is possi- ble that the same process occurs, but due to the small size of the embryo it was not possible to observe it very clearly. It is possible that the answer to the question of the initial mechanism of differentiation can be found in the endosperm, or even in the embryo itself.
The embryo length provides some information about the causes of early and late occurrences of abortion. Supposing first that the embryo grows fast and then a hormonal change occurs, early occurrence of abortion would be caused by a growth alteration and/or hormon- al deficiency preventing the embryo from reaching the next stage. The dead embryo in the preglobular stage has been degraded and absorbed by the surrounding tissue. Possibly, the absorption is due to its small size and non-differentiated structure (early occurrence of abortion). In late abortion, between the early globular stage and the torpedo stage, the embryos are not ab- sorbed because the already have a differentiated struc- ture.
For the embryo to develop to a new shape it needs nutrients to grow to it. Abortion can result from nutrient deficiencies (Johnston et al. 1980; Nishiyama and Ya- buno 1978). In the early globular stage, the breakdown can be due to problems in endosperm development or in the ovule nutrition system; the same processes can occur in the late globular stage. When the embryo reaches the late globular stage, hormonal deficiencies can prevent the change to the next stage of development and the embryo dies. The same abortion mechanisms can occur in any given shape (globular and heart) and at the torpedo stage.
Considering the embryo length, two hypotheses could explain the late occurrence of abortion: (1) nutritional deficiencies cause embryo starvation, so the embryo can- not continue to initiate the differentiation of stage (Nishiyama and Inomata 1966), or (2) it does initiate differentiation but is under nutritional stress which causes its death.
181
Table 4. Recommended number of DAP for embryo rescue in the three most important developmental stages in intraspecific and interspeci- fic crosses between Ipornoea batatas (2n-- 6x = 90) and I. trifida (2n = 2x = 30)
Developmental Cross combination stage
RCB2117H ~ DLP627 ~ EEC251 ~ RCB2117H DLP627 EEC251 x DLP627 x EEC251 x RCB2117H x L trifida x I. trifida x I. trifida
Globular 6 4 4 4 4 4 Heart 10 8 8 10 8 8 Torpedo 12 12 10 12 12 10
Ipomoea batatas (2n = 6x = 90)
Table 5. Estimated percentage of embryos at harvest (34 DAP) and estimated percentage of viable embryos at three principal developmental stages in intraspecific and interspecific crosses between L batatas (2n = 6x = 90) and L trifida (2n = 2x = 30). Chi-square test for goodness of fit was applied to test significance
Developmental Cross combination stage
RCB2117H ~ DLP627 ~ EEC251" RCB2117H DLP627 EEC251 x DLP627 x EEC251 x RCB2117H x I. trifida • L trifida x L trifida
34 DAP 52.5 66.3 34.3 6.4 6.6 1.3
Embryo rescue Torpedo 53.5 "s 66.7 ns 34.3 n~ 19.2" 18.7" 14.1 * Heart 53.5 nS 67.8 n~ 34.3 n~ 24.5" 58.0" 26.5" Globular 54.4 "S 69.3 ns 39.6 "~ 33.3" 66.4" 39.0'
a Ipomoea batatas (2n = 6x = 90) ns Not significant at 5% * Significantly different at 5%
In summary, nutritional barriers such as deficient en- dosperm tissue development by unbalanced chromosom- al sets (Katayama 1933; Kihara and Nishiyama 1932; Nishiyama and Inomata 1966; Thompson 1930; Wakak- uwa 1934), or the deficient suspensor development (Nishiyama and Inomata 1966) may be the main causes of embryo abortion in I p o m o e a interspecific hybrids. As newer concepts, Endosperm Balanced Number (EBN) (Johnston 1980) and the Polar-Nuclei Activation Index hypothesis (Nishiyama and Yabuno 1979) could be con- sidered. These barriers can be overcome with embryo rescue at an appropriate developmental stage.
Poten t ia l f o r e m b r y o rescue
Physio-embryonic barriers
Two physio-embryological barriers have been considered that decrease the botanical seed production in distant crosses: early occurrence and late occurrence of abor- tion. However, it may be possible to establish embryo rescue at three levels corresponding to each barrier: (1) keep fruits and ovules viable until the embryos reach an embryological stage that can be rescued; (2) improve the culture media and embryo extraction technique, so embryos can be rescued before the early globular stage; and (3) traditional embryo rescue in torpedo, heart and/ or globular stages.
Embryo and fruit abortion
The present research attempts to relate embryological development with daily fallen fruit after pollination in inter- and intraspecific crosses. The aim is to recommend not only the most appropriate stage but also the most efficient stage from the point of view of available em- bryos to be rescued.
Table 4 indicates the most appropriate DAP for em- bryo rescue in the different cross combinations used and their corresponding developmental stages. There are two interesting points: (1) The recommended DAP depends on the female parent, i.e., no matter what the male par- ent, the recommended DAP is the same for the same female except for RCB2117H (globular at 6 vs 4 DAP for other females); probably this is due to uniform em- bryo development in each female genotype. (2) Embryo development varies with the genotype but develops uni- formity time proceeds, no matter whether the cross com- bination is intra- or interspecific.
The results suggest the advisability of carrying out embryo rescue at the globular and heart stages because they would have a complete histological differentiation and hormonal equilibrium. Embryo rescue is not usually successful before the torpedo stage because it is difficult to handle small embryos and a more complex nutrient medium is required for these early stages (Salinas 1988).

182
Embryo rescue analysis
W h e n the capsules were mature (34 DAP) , it was esti- ma ted tha t a m a x i m u m o f 66.3% o f available embryos in intraspecific crosses and 6.6% in interspecific crosses were obta ined (Table 5). It would be possible to obta in a higher percentage o f embryos if embryo rescue were carried out.
Table 5 indicates that in intraspecific crosses there are no significant differences between the est imated em- bryo percentage at harvest time (66.3%) and the estimat- ed embryo rescue percentage at the three stages to rpedo (66.7%), heart (67.8%) and globular (69.3%). Conse- quently, for interspecific crosses, embryo rescue is no t r ecommended at any stage. On the o ther hand, signifi- cant differences were found at the torpedo, heart , and globular stages for interspecific crosses. Max ima esti- mated for available embryos were 18.7% in torpedo, 58.0% in heart, and 66.4% in globular stages. These percentages are significantly higher than the number o f embryos in mature capsules. These results point to the advisabili ty o f rescuing embryos at the globular stage, bu t if there are technical l imitat ions involved, we recom- mend carrying it out at the hear t stage. For embryos in early developmenta l stages more complex nutr i t ional media are required, so it would be more difficult to rescue embryos as successfully as in the intermediate or final developmenta l stages.
In summary , there are three phys io-embryologica l barriers to obta in ing viable seeds: fruit abor t ion, plus early and late occurrences o f abort ion. Wi th the present knowledge o f embryo rescue it is possible to avoid late occurrence o f abort ion. Frui t abor t ion could be avoided by using younger embryos , bu t unless an appropr ia te med ium and technique are developed to culture preglo- bular embryos , early abor t ion would no t be avoided. To overcome the latter two barriers fur ther research is needed into the origins o f these types o f abor t ion.
Acknowledgements. The authors would like to thank Dr. E. Carey, and Mrs. Alexandra Garcia-Bryce, CIP editor, for their sugges- tions. KW would like to thank several professors at Cornell Univer- sity for improvement of language expression and content.
References
Ahn CS, Hartmann RW (1978) Interspecific hybridization between mung bean (Vigna radiata L. Wilczek) and adzuki bean (K angularis Willd. Ohwi & Ohashi). J Am Soc Hort Sci 103:3-6
Blakeslee AF, Belling J, Farnham ME (1923) Inheritance in tetra- ploid Daturas. Bot Gaz 76:329-373
Cooper DC, Brink RA (1945) Seed collapse following matings be- tween diploid and tetraploid races of Lycopersicon pimpinelli- folium. Genetics 30 : 376
Freyre R, Iwanaga M, Orjeda G (1991) Use of Ipomoea tr~'da (H.B.K.) G. Don. germ plasm for sweet potato improvement.
2. Fertility of synthetic hexaploids and triploids with 2n gametes of L trifida, and their interspecific crossability with sweet-pota- to. Genome 34:209214
Johnston SA, den Nijs TPM, Peloquin S J, Hanneman RE Jr (1980) The significance of genic balance to endosperm development in interspecific crosses. Theor Appl Genet 57: 5-9
Jones A, Kobayashi M (1969) Interspecific hybrids of Ipomoea pescaprae and L erassicaulis. Bot Gaz 130:264-266
Jorgensen CA (1928) The experimental formation of heteroploid plants in the genus Solanum. J Genet 19:133-211
Katayama Y (1933) Crossing experiments in certain cereals, with special reference to different compatibility between the recipro- cal crosses. Mem Coll Agric, Kyoto Imp Univ 27:1-75
Kihara H, Nishiyama I (1932) Different compatibility in reciprocal crosses of Arena, with special reference to tetraploid hybrids between hexaploid species. Jpn J Bot 6:245-305
Kobayashi M (1978) Sweet potato breeding method using wild relatives in Japan. In: Symposium on Tropical Agriculture Re- search, October 1977, Tsukuba, Ibaraki, Japan. Ministry of Agriculture, Forestry and Fishery, Japan
Kobayashi M (1981) Origin and variety differentiation in sweetpo- taw (in Japanese). Yokenda, Tokyo. Rec Adr Plant Breed 22:107-103
Kokubu T, Murata T, Endo F (1982) Anatomical observations on the fertilization and embryogenesis in sweet potato, Ipomoea batatas (L.) Lam.. Jpn J Breed 32:239-246
Nishiyama I, Inomata N (1966) Embryological studies on cross- incompatibility between 2 x and 4 x in Brassica. Jpn J Genet 41 : 27-42
Nishiyama I, Yabuno T (1979) Triple fusion of the primary endo- sperm nucleus as a cause of interspecific cross incompatibility in Arena. Euphytiea 28 : 57-65
Orjeda G, Freyre R, Iwanaga M (1991) Use of Ipomoea trifida germplasm for sweet potato improvement. 3. Development of 4 x interspecific hybrids between Ipomoea batatas (L.) Lain. (2n=6x=90) and L trifida (H.B.K.) G. Don. (2n=2X=30) as storage-root initiators for wild species. Theor Appl Genet 83:159-163
Phillips GC, Collins GB, Taylor NL (1982) Interspecific hybridiza- tion of red clover (Trifolium pratense L.) with T. sarosiense Hazsl. using in vitro embryo rescue. Theor Appl Genet 62:17- 24
Randolph LF (1935) Cytogenetics of tetraploid maize. J Agric Res 50:591-605
Salinas R (1988) Cultivo in vitro de embriones de camote (Ipomoea batatas Lam). Thesis, La Molina National Agricultural Univer- sity, Lima (available at the International Potato Center)
Sharma DR (1980) Interspecific hybridization in genus Solarium: A cross between S. melongena and S. khasianum through em- bryo culture. Z Pflanzenzficht 85:248-253
Smith JA, Desborough SL (1986) An interpretation of genomic balance in seed formation in interspecific hybrids of Solanum. Theor Appl Genet 72:346-352
Thompson WP (1930) Shrivelled endosperm in species crosses in wheat, its cytological causes and genetical effects. Genetics 15:99-113
Torres F (1985) Anatomia comparativa de estructuras vegetativas de especies cultivadas de papa desarrolladas en dos ambientes contrastantes. Thesis, La Molina National Agricultural Univer- sity, Lima, (available at the International Potato Center)
Wakakuwa S (1934) Embryological studies on the different seed- development in reciprocal interspecific crosses of wheat. Jpn J Bot 7:151-185




























![A genetic linkage map of sweetpotato [Ipomoea batatas (L.) Lam.] based on AFLP markers](https://img.dokumen.tips/doc/110x75/63330e96b6829c19b80c3d31/a-genetic-linkage-map-of-sweetpotato-ipomoea-batatas-l-lam-based-on-aflp-markers.jpg)
