Embed Size (px)
Citation preview

CHAPTER ELEVEN
Modification of the ColonicMicrobiotaJuliet Ansell*,1, Shanthi Parkar†, Gunaranjan Paturi‡,Douglas Rosendale*, Paul Blatchford**The New Zealand Institute for Plant & Food Research Limited, Private Bag 11600, Palmerston North,New Zealand†The New Zealand Institute for Plant & Food Research Limited, Private Bag 3123, Hamilton, New Zealand‡The New Zealand Institute for Plant & Food Research Limited, Private Bag 92169, Auckland, New Zealand1Corresponding author: e-mail address: [email protected]
Contents
1.
AdvISShttp
Introduction
ances in Food and Nutrition Research, Volume 68 # 2013 Elsevier Inc.N 1043-4526 All rights reserved.://dx.doi.org/10.1016/B978-0-12-394294-4.00011-0
205
2. Modification of the Colonic Microbiota by Kiwifruit Polyphenolic Compounds 206 3. Modification of the Colonic Microbiota by Kiwifruit Carbohydrates 207 4. Kiwifruit Fermentation Offers Functional Benefits to the Gut 215 5. Conclusion 215 References 216Abstract
It is becoming clear that the ecology and functionality of the human gut microbiota areextremely diverse and complex. The microbiota have coevolved with us metabolicallyto live symbiotically and to share the workload of extracting nutrients and energy fromthe diet. It is also clear that a diet rich in fruit, vegetables, and whole grain cereals is goodfor general health and gut health and that this is due partly to the phytochemicals andpartly to the nondigestible carbohydrates (or dietary fiber) that are present in plants.Kiwifruit contain polyphenolics and nondigestible carbohydrates in the form of pectic,hemicellulosic, and cellulosic polysaccharides, all of which can be degraded by variousmembers of the gut microbiota and result in beneficial effects. This chapter summarizeshow kiwifruit act tomodify the colonic microbiota and the resultant beneficial effects onhuman health.
1. INTRODUCTION
A healthy balance of bacteria colonizing the adult gastrointestinal tract
is essential for the breakdown and metabolism of foods as well as for the pro-
duction of microbially synthesized vitamins. From a health perspective, the
205

206 Juliet Ansell et al.
gut microbes play a crucial role in the competitive inhibition of pathogens
and enhanced immunity through secretory and excretory substances that in-
teract with other microbes and the gut lining. Conversely, diet can have a
profound influence on the composition of the microbiota in the adult gut
(Macfarlane, Bahrami, & Macfarlane, 2011). What we eat can modulate
the colonic flora directly and influence resulting gut health.
The modern diet is said to be energy rich but nutrient poor with many
foods comprising “empty calories” rather than healthful nutrients. Although
there is still some debate around the definition of a nutrient-dense food,
fruits and kiwifruit in particular are accepted as being naturally nutrient rich
(Drewnowski, 2005), containing vitamin C, carotenoids, polyphenols,
flavonoids, fiber, folate, and zinc. Some of these nutrients are absorbed in
the upper gastrointestinal tract, as described elsewhere in this volume, but
others pass through to the colon and interact with the microbiota that reside
there. Polyphenols, mainly procyanidins and flavonol glycosides (Dawes &
Keene, 1999), as well as fiber fractions such as pectic polysaccharides rich in
arabinose, galactose, and galactose (Dawson & Melton, 1991), have been
characterized in green and gold kiwifruit and these are likely to transit to
the large intestine intact. Once in the large intestine, there are several ways
in which components of kiwifruit may modulate the colonic microbiota:
– Polyphenolics reaching the colon may directly stimulate the growth of
beneficial bacteria or act as antimicrobial agents against pathogens.
– Nondigestible polysaccharides fermented by the microbiota may induce
selective growth of beneficial bacteria.
The modulation of microbial growth is just the start of a cascade of events, as
microbial growth is associated with an increase in microbial metabolism,
which in turn releasesmoremetabolites into the gut environment.Highermi-
crobial growth may also translate to a higher turnover of bacteria, with some
breakdown products of lysis (bacterial cell wall fragments or DNA) potentially
also having an impact on the gut environment and mucosal immunity.
2. MODIFICATION OF THE COLONIC MICROBIOTABY KIWIFRUIT POLYPHENOLIC COMPOUNDS
Promotion of a healthy colonic environment is partly about selectively
enhancing the growth of beneficial bacteria but also involves the competitive
inhibition of potentially pathogenic bacteria. Both green (Actinidia deliciosa
‘Hayward’) and gold (A. chinensis ‘Hort 16A’) kiwifruit extracts have demon-
strable antimicrobial activity against potential gut pathogens while increasing

207Modification of the Colonic Microbiota
the growth of lactic acid bacteria. Aqueous extracts of the edible portion
inhibited monocultures of Staphylococcus aureus, Streptococcus mutans, Salmonella
enterica serovarTyphimurium, andEscherichia coli, whereas growth of lactic acid
bacteria Lactobacillus rhamnosus, Lactobacillus acidophilus, and Bifidobacterium breve
were increased (Molan, Kruger, De, & Drummond, 2007).
Similarly, using a fluorescent bacteria adhesion assay,wehave examined the
abilityof total kiwifruit polyphenolextracts toprevent theadhesionofS. enterica
(serovar Typhimurium; Parkar et al., 2010), by incubating antibiotic-free
14-dayCaco-2cell cultureswithpolyphenol extractsobtained fromtheaqueous
phase of green and gold kiwifruit (Hunter, Greenwood, Zhang, &
Skinner, 2011) at concentrations of 50 mg/mL for 1 h. This resulted in
a modest decrease in the adhesion of S. enterica (serovar Typhimurium)
to Caco-2 cells (14% and 9% respectively; S.G. Parkar, unpublished data).
Green kiwifruit contain the polyphenolics caffeic acid, catechin, chlo-
rogenic acid, epicatechin, p-coumaric acid, and quercetin derivates (Dawes &
Keene, 1999; Latocha, Krupa, Wolosiak, Worobiej, & Wilczak, 2010;
Sun-Waterhouse et al., 2009), and as described in Chapter 6. Commercial
preparations of these polyphenols have also been shown to generate a
dose-dependent decrease in S. enterica (serovar Typhimurium) adhesion to
Caco-2 cells (Parkar, Stevenson, & Skinner, 2008), thereby indicating a likely
role of these compounds in the previously observed inhibition by kiwifruit.
While relative to other fruit, kiwifruit are not particularly rich in total
phenolics (Hunter et al., 2011), they are rich in vitamin C. To explore
interactions between kiwifruit polyphenols and vitamin C against Salmonella
adhesion, quercetin was chosen as a representative kiwifruit polyphenol
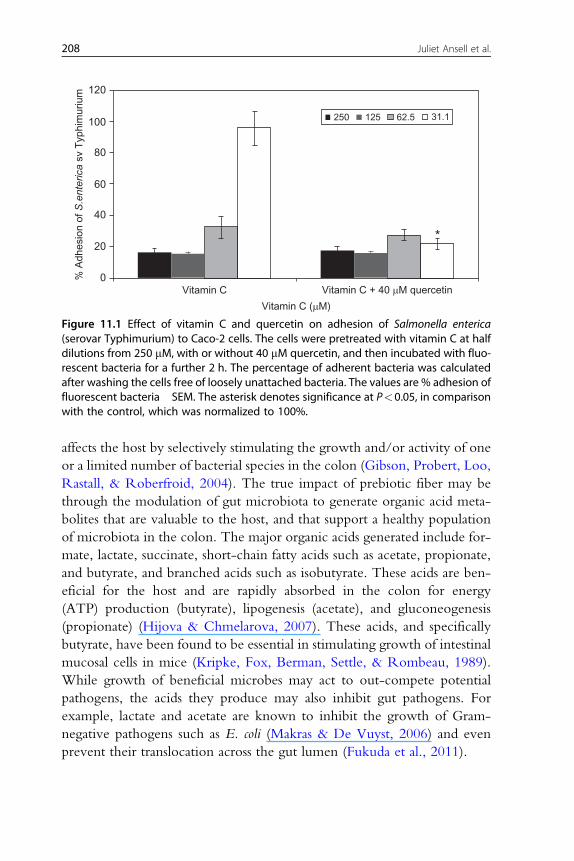
(Dawes & Keene, 1999). As shown in Fig. 11.1, vitamin C was found to
inhibit the adhesion of S. enterica (serovar Typhimurium to Caco-2) cells
at concentrations of up to 62.5 mM. However, in the presence of 40 mMquercetin, vitamin C was protective even at 31.25 mM. Thus, polyphenols
and vitamin C may combine to generate synergistic inhibitory effects on the
adhesion of pathogenic bacteria to the gut wall.
3. MODIFICATION OF THE COLONIC MICROBIOTABY KIWIFRUIT CARBOHYDRATES
Kiwifruit carbohydrates that escape digestion may act as prebiotics.
A prebiotic is defined as a food ingredient that is not hydrolyzed by the
human digestive enzymes in the upper gastrointestinal tract and beneficially
Vitamin C (mM)
Vitamin C
% A
dhes
ion
of S
.ent
eric
a sv
Typ
him
uriu
m
Vitamin C + 40 mM quercetin
20
0
40
60
250
*
125 62.5 31.1
80
100
120
Figure 11.1 Effect of vitamin C and quercetin on adhesion of Salmonella enterica(serovar Typhimurium) to Caco-2 cells. The cells were pretreated with vitamin C at halfdilutions from 250 mM, with or without 40 mM quercetin, and then incubated with fluo-rescent bacteria for a further 2 h. The percentage of adherent bacteria was calculatedafter washing the cells free of loosely unattached bacteria. The values are % adhesion offluorescent bacteria �SEM. The asterisk denotes significance at P<0.05, in comparisonwith the control, which was normalized to 100%.
208 Juliet Ansell et al.
affects the host by selectively stimulating the growth and/or activity of one
or a limited number of bacterial species in the colon (Gibson, Probert, Loo,
Rastall, & Roberfroid, 2004). The true impact of prebiotic fiber may be
through the modulation of gut microbiota to generate organic acid meta-
bolites that are valuable to the host, and that support a healthy population
of microbiota in the colon. The major organic acids generated include for-
mate, lactate, succinate, short-chain fatty acids such as acetate, propionate,
and butyrate, and branched acids such as isobutyrate. These acids are ben-
eficial for the host and are rapidly absorbed in the colon for energy
(ATP) production (butyrate), lipogenesis (acetate), and gluconeogenesis
(propionate) (Hijova & Chmelarova, 2007). These acids, and specifically
butyrate, have been found to be essential in stimulating growth of intestinal
mucosal cells in mice (Kripke, Fox, Berman, Settle, & Rombeau, 1989).
While growth of beneficial microbes may act to out-compete potential
pathogens, the acids they produce may also inhibit gut pathogens. For
example, lactate and acetate are known to inhibit the growth of Gram-
negative pathogens such as E. coli (Makras & De Vuyst, 2006) and even
prevent their translocation across the gut lumen (Fukuda et al., 2011).

209Modification of the Colonic Microbiota
Kiwifruit contain about 2–3% cell wall nonstarch polysaccharides
(NSPs), comprising soluble pectic polysaccharides (homogalacturonans and
rhamnogalacturonans substituted with galactan and arabinogalactan side
chains), insoluble trifluoroacetate–soluble hemicellulosic polysaccharides
(xyloglucan, glucuronoarabinoxylan, and galacto-(gluco)-mannan), and insolu-
ble trifluoroacetate–insoluble cellulose (Carnachan,Bootten,Mishra,Monro,&
Sims, 2012). These are described in more detail in Chapter 5. The chemical
composition and structure of these NSPs remain largely unchanged after
digestion, albeit with a decrease in the degree of methyl esterification of
galacturonic acid residues (Carnachan et al., 2012); so they pass intact to the large
intestine, where they become substrates for microbial fermentation by the resi-
dent microbiota, and can serve to enrich fractions of the microbiota known to
confer probiotic benefits on the host. As an example, pectinwith a lowdegree of
polymerization has been shown to promote butyrate production while pectin
with a high degree of polymerization promotes propionate production
(Nilsson &Nyman, 2005). Thus, the quality and quantity of fiber released after
digestion may play a major role in determining its prebiotic potential.
The ability of the gut microbiota to catabolize carbohydrates and gener-
ate organic acids and other fermentation by-products has been the subject of
review (Louis, Scott, Duncan, & Flint, 2007). A subsequent review covers
the mechanism of carbohydrate utilization in more detail (Flint, Bayer,
Rincon, Lamed, & White, 2008). Essentially, the ability of members of
the microbiota to utilize kiwifruit carbohydrates for energy is dictated by
their arsenal of degradative enzymes (glycoside hydrolases, polysaccharide
lyases, and carbohydrate esterases), adhesion factors, and transport systems
for each substrate (Flint et al., 2008). This results in an array of
carbohydrate-degrading mechanisms employed by so-called primary feeders
(those members of the microbiota able to ferment the substrate directly) and
subsequent secondary feeders that make use of released oligosaccharides or
fermentation by-products from the primary feeders.
Twowell-studied systemsexemplifyextremeparadigmsofcarbohydrateus-
age employed bymembers of the gut microbiota: the extracellular cellulosome
paradigm exemplified by members of the Gram-positive Ruminococcus genus,
whichwould be required for the degradation of insoluble kiwifruit xyloglucan,
glucuronoarabinoxylan, galacto-(gluco)-mannan and cellulose, and the largely
cell-associated sequestration paradigm exemplified by the Gram-negative
Bacteroides thetaiotaomicron, which would be required for the utilization of
soluble kiwifruit homogalacturonans and rhamnogalacturonans.

210 Juliet Ansell et al.
The Gram-positive, extracellular system features a multienzyme com-
plex tethered to a structural protein scaffold. This complex may incorporate
a binding module, glycosidases, and elements to modulate the number of
components comprising the cellulosome. Oligosaccharides released from
the hemicellulosic and cellulosic polysaccharides by the cellulosomal degra-
dation are taken up by transporters on the single cell membrane of the Gram-
positive bacterium. By-products expected from this fermentation include
hydrogen, carbon dioxide, and the organic acids formate, acetate, and suc-
cinate. Solubilized carbohydrates thus produced can act as a nutrient source
for Bacteroides (which in turn generate fermentation products, also including
propionate) and clostridial members of the Firmicutes genus such as Roseburia
spp. and Butyrivibrio, while the fermentation by-products themselves benefit
methanogens, acetogens, and the clostridial members. This latter group, in
turn, generates butyrate in addition to hydrogen, carbon dioxide, formate,
and acetate.
The Gram-negative, cell-associated system features “starch utilization”
(Sus) binding molecules on the outer membrane of the Bacteroides genera
which bind the pectic polysaccharide molecules to the cell surface, where-
upon the action of degradative enzymes releases and exposes oligosaccharides
to further degradation by additional glycosidases as they transit the periplasm,
cytoplasmic membrane and, via transporters, the cytoplasm. Waste products
include hydrogen, carbon dioxide, formate, acetate (which acts as a nutrient
for later members of the food chain), succinate, and propionate.
Structural differences in the responsible bacteria (single envelope vs.
double envelope enclosing the periplasmic space) and differences in the
physicochemistry of their preferred substrates (insoluble or soluble)maydictate
these different (extracellular breakdown vs. sequestration) approaches to kiwi-
fruit carbohydrate utilization. However, for both the systems described above,
successful carbohydrate-degrading bacteria possess multiple redundancies in
their degradative arsenals, as the loss of an “only-copy” would be catastrophic
to the survival of their possessors. Further, the glycosidase component of these
degradative systems shows considerable similarity in primary sequence, struc-
tural topology, and catalytic behavior across all bacteria. This enables the activ-
itiesof these enzymes tobedetermined in simple assay systems, such as theuseof
colorimetric enzyme substrates (Riley & Wertz, 2002).
A recent attempt to quantify some of these processes involved in kiwi-
fruit degradation used an in vitro batch fermentation system, minimizing the
confounding influence of host carbohydrates. Fresh whole green kiwifruit
were subjected to simulated gastric-small intestinal digestion, thus simulating

211Modification of the Colonic Microbiota
the physicochemical modifications that kiwifruit undergo as they transit
through the upper gut. The large polysaccharides remaining after dialysis
of the kiwifruit digesta were fermented for 48 h with fecal slurries obtained
from healthy donors. By assessing changes in the abundance of key microbial
phyla and determining glycosidase activities and fermentation by-product
(organic acid) generation, the degradation of kiwifruit polysaccharide by
the complex food web of the microbiota was investigated.
Real-time quantitative PCR was used to demonstrate that both gold
and green kiwifruit altered specific human microbial populations
over 48 h. Both kiwifruit substrates increased the total bacteria, Bacte-
roides–Prevotella–Porphyromonas group, Lactobacillus, and Bifidobacterium spp.
compared with the control (Parkar et al., 2012). Similar increases in bacterial
populations were reported by Han, Balan, Molist Gasa, and Boland (2011),
where a higher abundance of total bacteria and members of the Bacteroides
group, and an increase in the ratio of Lactobacillus to Enterobacteria were
observed in the colon of growing pigs fed green kiwifruit fiber compared
with those fed a control diet.
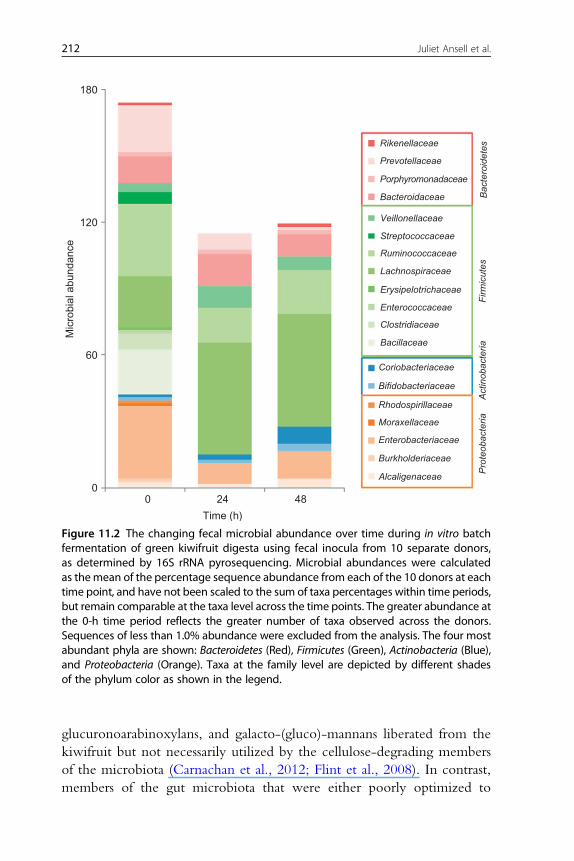
These prebiotic effects have since been explored in greater detail, using
16S rRNA pyrosequencing (Fig. 11.2) of the microbial DNA prepared from
the green kiwifruit fermenta from that same study (Parkar et al., 2012). Batch
fermentation with green kiwifruit led to an immediate and lasting reduction
in the average percentage abundance of all members of the Proteobacteria
phyla; Bacillaceae and Clostridiaceae members of the Firmicutes phyla; and
Prevotellaceae from the Bacteroidetes phyla, which were present in the original
fecal inocula from 10 donors. The abundance of Ruminococcaceae (Firmicutes)
was halved after 24 h. In contrast, Bacteroidaceae, Lachnospiraceae, and
Veillonella members of the Firmicutes phyla and Coriobacteriaceae of the
Actinobacteria phyla increased relatively following 24 h of exposure to green
kiwifruit. The increases in Lachnospiraceae remained sustainable after 48 h.
Coriobacteriaceae continued to increase relatively, accompanied by increases
in Bifidobacteraceae (Actinobacteria) together with a slight resurgence in
Ruminococcaceae and Enterobacteraceae (Proteobacteria). After 48 h, the increased
levels of other bacteria observed at 24 h were not sustainable and had
decreased in relative abundance. These changes appeared to correlate
with the expected roles and carbohydrate utilization capabilities of the
various members of the gut microbiota. Known dietary carbohydrate uti-
lizers such as Lachnospiraceae and Bacteroidaceae were able to make early use
of the kiwifruit substrate, particularly soluble pectic homogalacturonans
and rhamnogalacturonans, and of the insoluble hemicellulosic xyloglucans,
Rikenellaceae
Prevotellaceae
Porphyromonadaceae
Bacteroidaceae
Veillonellaceae
Streptococcaceae
Ruminococcaceae
Lachnospiraceae
Erysipelotrichaceae
Enterococcaceae
Clostridiaceae
Bacillaceae
Coriobacteriaceae
Bifidobacteriaceae
Rhodospirillaceae
Moraxellaceae
Enterobacteriaceae
Burkholderiaceae
AlcaligenaceaeP
rote
obac
teria
Act
inob
acte
riaF
irmic
utes
Bac
tero
idet
es
00
60
Mic
robi
al a
bund
ance
120
180
24 48
Time (h)
Figure 11.2 The changing fecal microbial abundance over time during in vitro batchfermentation of green kiwifruit digesta using fecal inocula from 10 separate donors,as determined by 16S rRNA pyrosequencing. Microbial abundances were calculatedas themean of the percentage sequence abundance from each of the 10 donors at eachtime point, and have not been scaled to the sum of taxa percentages within time periods,but remain comparable at the taxa level across the time points. The greater abundance atthe 0-h time period reflects the greater number of taxa observed across the donors.Sequences of less than 1.0% abundance were excluded from the analysis. The four mostabundant phyla are shown: Bacteroidetes (Red), Firmicutes (Green), Actinobacteria (Blue),and Proteobacteria (Orange). Taxa at the family level are depicted by different shadesof the phylum color as shown in the legend.
212 Juliet Ansell et al.
glucuronoarabinoxylans, and galacto-(gluco)-mannans liberated from the
kiwifruit but not necessarily utilized by the cellulose-degrading members
of the microbiota (Carnachan et al., 2012; Flint et al., 2008). In contrast,
members of the gut microbiota that were either poorly optimized to

213Modification of the Colonic Microbiota
compete for dietary carbohydrate in a closed system (Proteobacteria), or
known to exhibit a preference for host (mucin) carbohydrate
(Prevotellaceae and, to some extent, Ruminococcaceae), declined relative to
the more numerous bacteria. After 48 h, it is expected that only the most
robust or specialized primary carbohydrate degraders would be able to make
use of remaining, poorly accessible kiwifruit sugar residues, such as poorly
fermented or underutilized cellulosic carbohydrates. This may explain the
observed resurgence of Ruminococcaceae, which possess the extracellular
cellusomal degradative system discussed above. Those degraders with a
cell-associated soluble carbohydrate-utilizing system, such asBacteroidaceae de-
clined in numbers relative to the more abundant or increasing Firmicutes and
Actinobacteria. It is likely that secondary metabolizers such as Bifidobacteriaceae
and perhaps Enterobacteraceae could continue to make use of metabolic
by-products and also obtain nutrients from dying bacteria.
These microbial data correspond with the glycosidase activities recorded
from these fermenta (Table 11.1 and Parkar et al., 2012). Statistically
significant increases in activity were observed with almost all substrates
(fucose, galactose, glucose, uronic acid, arabinose, rhamnose, and xylose)
used. The corresponding sugar residues comprise the pectic andhemicellulosic
fractions of the kiwifruit (Carnachan et al., 2012); these are the substrates the
abundantBacteroidaceae andLachnospiraceae are expected to utilize or, in the case
of hemicellulosic sugars, theRuminococcaceae to release for utilization by others.
For example, a decrease inmannosidasewas noted to correspond to a rapid de-
cline inavailablemannoseof yeastorigin in the fermentationmedia, reinforcing
that the changes in activities observed tended to correspond to microbial car-
bohydrate utilization over time.
The microbial fermentation by-products (namely the organic acids ace-
tate, propionate, and butyrate) were observed to increase in the presence of
kiwifruit fermenta, while lactate and succinate concentrations declined after
24 h, lactate remained low at 48 h, and succinate increased (Table 11.1 and
Parkar et al., 2012). A decline in lactate and succinate would be expected in a
batch fermentation. They may act as metabolic intermediates or cross-
feeding substrates for the further generation of acetate and propionate,
respectively. Most of the genera observed by pyrosequencing are acetate
producers. Propionate is a product of Bacteroidetes fermentation, and also
produced by Veillonella and other members of the Firmicutes phyla. Butyrate
is produced by members of the Firmicutes subset which features members of
the Lachnospiraceae family, among others (Louis et al., 2007). While it would
be an oversimplification to state that these acids specifically correlate with
those taxa observed to increase with kiwifruit fermentation, all acid
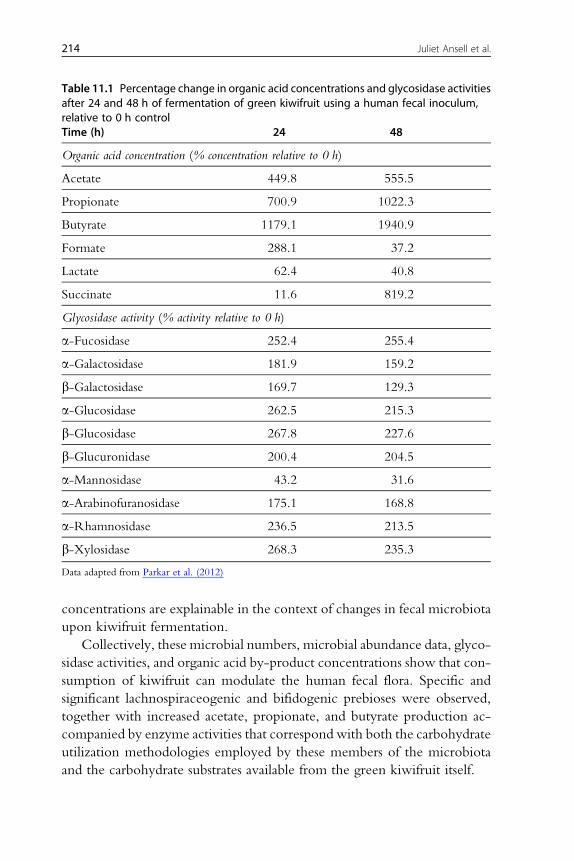
Table 11.1 Percentage change in organic acid concentrations and glycosidase activitiesafter 24 and 48 h of fermentation of green kiwifruit using a human fecal inoculum,relative to 0 h controlTime (h) 24 48
Organic acid concentration (% concentration relative to 0 h)
Acetate 449.8 555.5
Propionate 700.9 1022.3
Butyrate 1179.1 1940.9
Formate 288.1 37.2
Lactate 62.4 40.8
Succinate 11.6 819.2
Glycosidase activity (% activity relative to 0 h)
a-Fucosidase 252.4 255.4
a-Galactosidase 181.9 159.2
b-Galactosidase 169.7 129.3
a-Glucosidase 262.5 215.3
b-Glucosidase 267.8 227.6
b-Glucuronidase 200.4 204.5
a-Mannosidase 43.2 31.6
a-Arabinofuranosidase 175.1 168.8
a-Rhamnosidase 236.5 213.5
b-Xylosidase 268.3 235.3
Data adapted from Parkar et al. (2012)
214 Juliet Ansell et al.
concentrations are explainable in the context of changes in fecal microbiota
upon kiwifruit fermentation.
Collectively, these microbial numbers, microbial abundance data, glyco-
sidase activities, and organic acid by-product concentrations show that con-
sumption of kiwifruit can modulate the human fecal flora. Specific and
significant lachnospiraceogenic and bifidogenic prebioses were observed,
together with increased acetate, propionate, and butyrate production ac-
companied by enzyme activities that correspond with both the carbohydrate
utilization methodologies employed by these members of the microbiota
and the carbohydrate substrates available from the green kiwifruit itself.

215Modification of the Colonic Microbiota
4. KIWIFRUIT FERMENTATION OFFERS FUNCTIONALBENEFITS TO THE GUT
In addition to modulating microbial numbers and by-products,
fermentation of both green and gold kiwifruit has been shown to influence
gut microecology bymodulating the adhesion of different bacteria to the gut,
which is the first step in microbial colonization of the gut wall (Bavington &
Page, 2005; Parkar et al., 2010). These fermenta were found to increase the
proliferation of Bifidobacterium longum (Parkar et al., 2012), one of the probi-
otic organisms most commonly found in human feces (Malinen et al., 2002)
and further increased their adhesion to human intestinal epithelial cells. In the
gut, this would increase their chances of stable colonization. These adhesion
effects are consistent with previous work that showed that a pectin fraction
rich in galactose, arabinose, and galacturonic acid enhanced the adhesion
of Lactobacillus rhamnosus to Caco-2 cells in vitro, but inhibited the adhesion
of S. enterica (serovar Typhimurium; Parkar et al., 2010).
As well as organic acids, the microbiota are known to directly produce
antimicrobial substances, such as bacteriocins, which exhibit bactericidal ac-
tivity toward closely related strains (Riley & Wertz, 2002). Although no
work has yet linked the production of bacteriocins with the consumption
of kiwifruit, we know that kiwifruit consumption increases the numbers
of appropriate probiotic strains, and this can increase beneficial bacteriocins.
This, in turn, can augment the host resistance to allochthonous pathogens
(Tuohy, Rouzaud, Bruck, & Gibson, 2005).
In addition to an elevated production of bacteriocins to challenge
pathogen growth, increases in organic acids andmucin-degrading enzyme ac-
tivities associated with Bacteroidetes (Xu et al., 2003) and Bifidobacteria (Turroni
et al., 2010) in the kiwifruit-fermenting microbiota (Parkar et al., 2012) are
associated with either increased host antimicrobial peptide production
(organic acids; described in Chapter 17), competitive exclusion through
outcompeting for mucin colonization sites (Bacteroidetes, Bifidobacteria), or
outcompeting for epithelial binding sites (Bifidobacteria).
5. CONCLUSION
There is growing evidence that consumption of kiwifruit beneficially
modulates the colonic microbiota. Kiwifruit carbohydrates, vitamins, and
phytochemicals that escape host digestion are fermented by the microbiota,

216 Juliet Ansell et al.
changingmicrobial growth and by-product formation; and affecting binding
to colonization sites in the gut and the ability to directly or indirectly stim-
ulate immune cells. Therefore, through modulation of the microbiota,
kiwifruit consumption can affect many of the predominant factors which
collectively comprise the large intestinal system. This is likely to ultimately
result in increased large intestinal health and overall wellness.
REFERENCESBavington, C., & Page, C. (2005). Stopping bacterial adhesion: A novel approach to treating
infections. Respiration, 72, 335–344.Carnachan, S. M., Bootten, T. J., Mishra, S., Monro, J. A., & Sims, I. M. (2012). Effects of
simulated digestion in vitro on cell wall polysaccharides from kiwifruit (Actinidia spp.).Food Chemistry, 133, 132–139.
Dawes, H. M., & Keene, J. B. (1999). Phenolic composition of kiwifruit juice. Journal ofAgricultural and Food Chemistry, 47, 2398–2403.
Dawson, D. M., & Melton, L. D. (1991). Two pectic polysaccharides from kiwifruit cellwalls. Carbohydrate Polymers, 15, 1–11.
Drewnowski, A. (2005). Concept of a nutritious food: Toward a nutrient density score.The American Journal of Clinical Nutrition, 82, 721–732.
Flint, H. J., Bayer, E. A., Rincon, M. T., Lamed, R., &White, B. A. (2008). Polysaccharideutilization by gut bacteria: Potential for new insights from genomic analysis. NatureReviews. Microbiology, 6, 121–131.
Fukuda, S., Toh, H., Hase, K., Oshima, K., Nakanishi, Y., Yoshimura, K., et al. (2011).Bifidobacteria can protect from enteropathogenic infection through production of ace-tate. Nature, 469, 543–547.
Gibson, G. R., Probert, H. M., Loo, J. V., Rastall, R. A., & Roberfroid, M. B. (2004).Dietary modulation of the human colonic microbiota: Updating the concept of prebi-otics. Nutrition Research Reviews, 17, 259–275.
Han, K. S., Balan, P., Molist Gasa, F., & Boland, M. (2011). Green kiwifruit modulates thecolonic microbiota in growing pigs. Letters in Applied Microbiology, 52, 379–385.
Hijova, E., & Chmelarova, A. (2007). Short chain fatty acids and colonic health. BratislavaMedical Journal, 108, 354–358.
Hunter, D. C., Greenwood, J., Zhang, J. L., & Skinner, M. A. (2011). Antioxidant and ‘nat-ural protective’ properties of kiwifruit. Current Topics in Medicinal Chemistry, 11,1811–1820.
Kripke, S. A., Fox, A. D., Berman, J. M., Settle, R. G., &Rombeau, J. L. (1989). Stimulationof intestinal mucosal growth with intracolonic infusion of short-chain fatty-acids. Journalof Parenteral and Enteral Nutrition, 13, 109–116.
Latocha, P., Krupa, T., Wolosiak, R., Worobiej, E., & Wilczak, J. (2010). Antioxidantactivity and chemical difference in fruit of different Actinidia sp. International Journal ofFood Sciences and Nutrition, 61, 381–394.
Louis, P., Scott, K. P., Duncan, S. H., & Flint, H. J. (2007). Understanding the effects of dieton bacterial metabolism in the large intestine. Journal of Applied Microbiology, 102,1197–1208.
Macfarlane, S., Bahrami, B., & Macfarlane, G. T. (2011). Mucosal biofilm communities inthe human intestinal tract. In A. I. Laskin, S. Sariaslani & G. M. Gadd (Eds.), Advances inapplied microbiology (pp. 111–143). San Diego: Academic Press.

217Modification of the Colonic Microbiota
Makras, L., & De Vuyst, L. (2006). The in vitro inhibition of Gram-negative pathogenic bac-teria by bifidobacteria is caused by the production of organic acids. International DairyJournal, 16, 1049–1057.
Malinen, E., Matto, J., Salmitie, M., Alander, M., Saarela, M., & Palva, A. (2002). PCR-ELISA: II: Analysis of Bifidobacterium populations in human faecal samples from a con-sumption trial with Bifidobacterium lactis Bb-12 and a galacto-oligosaccharide preparation.Systematic and Applied Microbiology, 25, 249–258.
Molan, A. L., Kruger, M. C., De, S., & Drummond, L. N. (2007). The ability of kiwifruit topositively modulate key markers of gastrointestinal function. In: Proceedings of the Nutri-tion Society of New Zealand, 32, 66–71.
Nilsson, U., & Nyman, M. (2005). Short-chain fatty acid formation in the hindgut of rats fedoligosaccharides varying in monomeric composition, degree of polymerisation and sol-ubility. The British Journal of Nutrition, 94, 705–713.
Parkar, S. G., Redgate, E. L., Wibisono, R., Luo, X., Koh, E. T. H., & Schroder, R. (2010).Gut health benefits of kiwifruit pectins: Comparison with commercial functional poly-saccharides. Journal of Functional Foods, 2, 210–218.
Parkar, S. G., Rosendale, D., Paturi, G., Herath, T. D., Stoklosinski, H., Phipps, J. E., et al.(2012). In vitro utilization of gold and green kiwifruit oligosaccharides by human gut mi-crobial populations. Plant Foods for Human Nutrition, 67, 200–207.
Parkar, S. G., Stevenson, D. E., & Skinner, M. A. (2008). The potential influence of fruitpolyphenols on colonic microflora and human gut health. International Journal of Food Mi-crobiology, 124, 295–298.
Riley, M. A., & Wertz, J. E. (2002). Bacteriocins: Evolution, ecology, and application. An-nual Review of Microbiology, 56, 117–137.
Sun-Waterhouse, D., Chen, J., Chuah, C., Wibisono, R., Melton, L. D., Laing, W., et al.(2009). Kiwifruit-based polyphenols and related antioxidants for functional foods: Kiwi-fruit extract-enhanced gluten-free bread. International Journal of Food Sciences and Nutrition,60, 251–264.
Tuohy, K. M., Rouzaud, G. C. M., Bruck, W. M., & Gibson, G. R. (2005). Modulation ofthe human gut microflora towards improved health using prebiotics—Assessment of ef-ficacy. Current Pharmaceutical Design, 11, 75–90.
Turroni, F., Bottacini, F., Foroni, E., Mulder, I., Kim, J.-H., Zomer, A., et al. (2010).Genome analysis of Bifidobacterium bifidum PRL2010 reveals metabolic pathways forhost-derived glycan foraging. Proceedings of the National Academy of Sciences of the UnitedStates of America, 107, 19514–19519.
Xu, J., Bjursell, M. K., Himrod, J., Deng, S., Carmichael, L. K., Chiang, H. C., et al. (2003).A genomic view of the human-Bacteroides thetaiotaomicron symbiosis. Science, 299,2074–2076.





























