Embed Size (px)
Citation preview

Research ArticleThe Pathogenicity of Anti-1205732GP1-IgG AutoantibodiesDepends on Fc Glycosylation
Christoph Fickentscher1 Iryna Magorivska12 Christina Janko13
Mona Biermann1 Rostyslav Bilyy2 Cecilia Nalli4 Angela Tincani4
Veronica Medeghini5 Antonella Meini5 Falk Nimmerjahn6 Georg Schett1
Luis E Muntildeoz1 Laura Andreoli4 and Martin Herrmann1
1Department for Internal Medicine 3 University Hospital Erlangen Friedrich-Alexander-Universitat Erlangen-Nurnberg (FAU)Ulmenweg 18 91054 Erlangen Germany2Institute of Cell Biology National Academy of Sciences of Ukraine Drahomanov Street 1416 Lviv 79005 Ukraine3Department of Otorhinolaryngology Head and Neck Surgery Section for Experimental Oncology and Nanomedicine (SEON)University Hospital Waldstraszlige 1Gluckstraszlige 10 91054 Erlangen Germany4Rheumatology and Clinical Immunology Department of Clinical and Experimental Sciences Spedali Civili and University of BresciaPiazzale Spedali Civili 1 25123 Brescia Italy5Paediatric Immunology and Rheumatology Childrenrsquos Hospital Brescia Department of Clinical and Experimental SciencesSpedali Civili and University of Brescia Piazzale Spedali Civili 1 25123 Brescia Italy6Department of Biology Institute of Genetics Friedrich-Alexander-Universitat Erlangen-Nurnberg (FAU) Erwin-Rommel Str 391058 Erlangen Germany
Correspondence should be addressed to Luis E Munoz luismunozuk-erlangende
Received 11 February 2015 Accepted 12 May 2015
Academic Editor Kurt Blaser
Copyright copy 2015 Christoph Fickentscher et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
To analyze the glycosylation of anti-1205732GP1 we investigated purified IgG from healthy children patients with APS andasymptomatic adult carriers of antiphospholipid antibodies We observed that in the sera of healthy children and of patients withAPS IgG3 and IgG2 were predominant respectively The potentially protective anti-1205732GP1-IgM was lower in the sera of healthychildren Although anti-1205732GP1-associated C1q did not differ between children and patients with antiphospholipid syndrome theassociated C3c was significantly higher in the sera of healthy children This indicates a more efficient clearance of anti-1205732GP1immune complexes in the healthy children This clearance is not accompanied by inflammation or coagulatory events It is likelythat the most important pathogenic factor of the anti-1205732GP1-IgG is related to the different glycosylation observed in healthy anddiseased individuals We detected a significantly higher sialylation of anti-1205732GP1-IgG isolated from the sera of healthy childrenand asymptomatic adults when compared with that of patients with clinically apparent antiphospholipid syndrome Low sialylatedIgG reportedly ameliorates inflammation and inflammation promotes hyposialylation Thus both reactions create a vicious circlethat precipitates the pathology of the antiphospholipid syndrome including thrombus-formation We conclude that the increasedsialylation of anti-1205732GP1-IgG of sera of healthy individuals limits their pathogenicity
1 Introduction
Theantiphospholipid syndrome (APS) is an autoimmune dis-ease affecting the coagulation system Its diagnosis requiresat least one clinical criterion and one serological criterion asdefined during conferences held in 1998 and in 2006 [1 2]
The serological criteria include the detection of so-calledantiphospholipid antibodies (aPL) which comprise the lupusanticoagulant (LA) anticardiolipin autoantibodies (aCL)and anti-1205732-glycoprotein 1 autoantibodies (anti-1205732GP1) [1]
Thrombosis as one major symptom in APS is mainlyassociated with anti-1205732GP1 [3] 1205732GP1 is a plasma protein
Hindawi Publishing CorporationJournal of Immunology ResearchVolume 2015 Article ID 638129 12 pageshttpdxdoiorg1011552015638129
2 Journal of Immunology Research
composed of five domains [4] and it is known to be aninhibitor of the contact activation of the intrinsic coagulationpathway [5] Additionally aPL can cause a prothromboticstate in individuals by activating platelets and endothelialcells resulting in a higher expression of tissue factor or in theupregulation of proinflammatory cytokines and cell adhesionmolecules [6 7]
Patients with APS can be classified into two groupsdepending on the occurrence of further diseases (I) patientsmerely suffering from APS (primary antiphospholipid syn-drome PAPS) and (II) those with an underlying systemiclupus erythematodes (SLE) The latter condition is referredto as secondary antiphospholipid syndrome (SAPS) [8]
Up to now a variety of hypotheses about possible triggersfor the induction of anti-1205732GP1 have been proposed Pub-lished studies suggest that their production can be provokedduring infectious diseases like parvovirus B19 HIV CMVor HCV [3 9ndash11] Certain molecular structures of theseinfectious agents may resemble structures of 1205732GP1 and thuscause the induction of cross-reacting antibodies [7 12ndash16]Another one claims that nutritional exposure to 1205732GP1 couldalso trigger the production of transient and harmless anti-1205732GP1 in children [4 17ndash19]
Interestingly anti-1205732GP1 can be detected in a quite largeproportion of the healthy population who do neither sufferfrom APS nor another autoimmune disease [3 18 20]As complication-free periods of patients with APS occuralthough the aPL are persistently detectable in their circula-tion [15] it seems to be likely that the mere presence of theseantibodies is not sufficient for the development of clinicalactive APS complications
For this reason a two-hit hypothesis has been proposedto provide a putative explanation for this observation (I)the production of anti-1205732GP1 represents an initial hit thatincreases the risk for thrombotic events and (II) infectiousagents can then serve as the second hit provoking the typicalmanifestations of APS by activating toll-like receptors orcomplement [9 15 21 22] This two-hit hypothesis surelyprovides a good first model for the etiopathogenesis of APSYet it is not completely satisfying and does not explain whysome people harbouring aPL remain healthy Some studyresults indicate that this could be grounded in structuraldifferences of anti-1205732GP1 found in patients with APS andhealthy children [4 23]
Comparing obviously harmless aPL from healthy chil-dren with putatively pathogenic aPL from adults with APSwe suggested a difference in the epitope specificity as differ-entiating factor Anti-1205732GP1 from the sera of the childrenspecifically bound to the domain IVV of 1205732GP1 whereasanti-1205732GP1 from adults with APS preferentially targeteddomain I [24] These results were consistent with thosedescribed in other studies [19 25] Besides distinct epitopebinding specificities a different composition of the spectrumof the anti-1205732GP1 IgG subclasses between patients with APSand healthy carriers of this antibody has also been described[19 23 26 27]
The differences in glycosylation of antibodies which areaccompanied by crucial modifications of their effector func-tions [28] provide another promising approach in performing
structural analyses of anti-1205732GP1 IgG molecules possess abiantennary oligosaccharide attached to the asparagine atposition 297 of the Fc portion (Asn-297) Various glycoformscontaining distinct sugar moieties have been described [2829] Interestingly it has been observed that IgG moleculeswhose glycans terminate with sialic acid residues showa decreased affinity to activating Fc120574-receptors (Fc120574Rs)while simultaneously provoking an upregulation of theinhibitory Fc120574RIIB [29ndash31] These effects lead to a ratheranti-inflammatory activity andmight consequently be able toprevent potentially harmful immune responses
In particular in children aPL can readily be detectedalthough they do not show any clinical signs of APS In astudy carried out by Avcin et al the prevalence of anti-1205732GP1 in 61 healthy children was 66 [18] Based on thisobservation we intended to find out why these circulatingautoantibodies do not cause harm in their hosts We deter-mined the IgG-subclass distribution in the samples fromhealthy children and patients with APS and examined thebinding of the complement factors C1q and C3c to anti-1205732GP1 immune complexes since an important involvementof the complement system in the pathogenesis of APS hasbeen reported [21 22 32] Finally we analysed the sialylationstatus of anti-1205732GP1-IgG isolated from healthy childrenasymptomatic adult carriers and patients with APS
Our results revealed that compared to adults with APSthe healthy children showed a higher content of IgG3 at thecost of IgG2 of the anti-1205732GP1 Furthermore anti-1205732GP1-IgGof healthy children showed a higher sialylation and displayedmore bound C3c Hyposialylated IgG and inflammationmutually influence the pathogenicity of the antiphospholipidautoantibodies and form a vicious pathogenic circle
2 Materials and Methods21 Patients For the investigation of anti-1205732GP1-IgG itssubclasses C1q- and C3c-binding and anti-1205732GP1-IgM westudied sera of 46 adult patients (24 patientswith PAPS and 22with SAPS) and 16 healthy children harbouring anti-1205732GP1without any clinical manifestations associated with APS fromour partner hospital in Brescia Italy As a control groupwe examined sera of 17 age and sex matched adult normalhealthy donors (NHD) without aPL Inclusion criteria for thechildren were the presence of anti-1205732GP1-IgG no infectiousdiseases in the course of three months before blood takingand no chronic pathology Additionally it was excluded thattheir parents suffered from any autoimmune disease At thetime of blood taking the mean age of the children was about9 months (median age 85 months range 3 months and 5daysndash19 months and 10 days)
Since sugar residues attached to molecules can be dam-aged by repeated thawing and freezing we received newsera from our partner hospital in Brescia Italy to analysethe sialylation of anti-1205732GP1-IgG We obtained sera from 12patients with PAPS and 10 patients with SAPS 15 patientswith SLE and aPL but without clinical evidence of APSand sera from 6 asymptomatic carriers of aPL Furthermorewe obtained sera from 16 healthy children showing nosymptoms of APS although harbouring anti-1205732GP1-IgG in
Journal of Immunology Research 3
Table 1 Antibodies employed for each ELISA
ELISA Detection-antibody DilutionAnti-1205732GP1-IgG HRP-conjugated rabbit F(ab1015840) 2 anti-human-IgGlowast 1 20000
Anti-1205732GP1-IgG subclasses HRP-conjugated mouse anti-human IgG1 2 3 and 4 (Clones HP6001HP6002 HP6050 amp HP6025)lowast 1 20000
Anti-1205732GP1-IgM HRP-conjugated goat anti-human-IgMlowast 1 20000Anti-1205732GP1-bound C3c HRP-conjugated sheep anti-C3clowastlowast 1 500Anti-1205732GP1-bound C1q HRP-conjugated sheep anti-C1qlowastlowast 1 500lowast purchased from Southern Biotech Birmingham Alabama USAlowastlowast purchased from Abcam Cambridge United KingdomHRP = horseradish peroxidase
their circulation (mean age 95 months median age 10months range 3 months and 5 daysndash19 months and 10 days)
22 Blood Drawing Heparinised venous blood was drawnby venipuncture and processed within two hours Serum wasobtained by centrifugation at 1000 g for 15 minutes of clottedblood then aliquoted frozen and stored at minus20∘C until usefor investigation
Informed consent was obtained from all blood donorsand the childrenrsquos parents respectively and the study wasapproved by the ethics committees of the University HospitalErlangen and the Spedali Civili Brescia
23 Isolation of 1205732GP1 For 1205732GP1-isolation from dilutedsera of normal healthy donors (in 015MNaCl) percloric acidwas slowly added until the mixture got a creamy aspect Aftercentrifugation at 10000 g the supernatant was neutralizedwith NaOH and dialyzed against a buffer of 002M Tris and003MNaCl pH8Then itwas given into columns containingHiTrap heparin and contaminants of the supernatant wereeluted by the addition of 002M Tris and 015M NaCl pH 8In order to elute semipurified 1205732GP1 a buffer of 002M Trisand 035MNaCl pH 8 was injected into the columns and thefractions have been pooled and dialyzed against a solutioncontaining 005M acetate buffer 005M NaCl pH 48 Thismaterial was filtrated with 02120583membranes and injected intoresin columns equilibrated with 005M acetate buffer 005MNaCl pH 48 After washing with 005M acetate buffer065M NaCl pH 52 the fractions were pooled and dialyzedagainst PBS pH 72 To verify the purity of isolated 1205732GP1a sodium dodecyl sulfate polyacrylamide gel electrophoresis(SDS-PAGE) has been performed
24 Purification of Anti-1205732GP1-IgG For the analysis of thesialylation of anti-1205732GP1-IgG we isolated IgG antibodiesfrom the sera to exclude possible interactions of the detectionlectin with anti-1205732GP1-IgM in the samples Purification ofanti-1205732GP1-IgG was performed as follows 20120583L of serumwas added to 50120583L magnetic Dynabeads Protein G (Novexby life technologies Carlsbad CA USA) and incubated for45 minutes at room temperature After washing with 1x TBSglycin-HCl pH 26 was added for 5 minutes to elute the anti-1205732GP1-IgG The supernatant now containing the IgG anti-bodies was neutralized with Tris-HCl pH 88 and all sam-ples were centrifuged using 100K Amicon Ultra-centrifugal
filter devices (Millipore Tullagreen Ireland) to transfer theminto 300 120583L TBS-buffer
25 Serum ELISAs Anti-1205732GP1-IgG -IgG1 -IgG2 -IgG3-IgG4 and -IgM and C3c and C1q Bound to Anti-1205732GP1To quantify the amount of anti-1205732GP1-IgG and its foursubclasses of C3c and C1q associated with anti-1205732GP1 andof anti-1205732GP1-IgM in the sera of patients with PAPS andSAPS NHD and healthy children we performed enzyme-linked immunosorbent assays (ELISA) In detail the surfaceof F 96NUNCmaxisorpmicrotiter plates (Thermo ScientificRoskilde Denmark) was coated with 50 120583L of a 2 120583gmLsolution of 1205732GP1 in coating buffer (100mM Na
2CO3
100mM NaHCO3 pH 96) at 4∘C overnight After washing
the plates three times with washing buffer (WB 005 Tween20 01mM CaCl
2 01mM MgCl
2in TBS buffer (150mM
NaCl 50mM TrisHCl pH = 74)) the wells of the coatedplates were blocked with 100 120583Lwell of a 2 BSA solutionfor two hours at 37∘C After three further washing steps withWB 50 120583Lwell of diluted serum (1 200 in WB) was addedand the plates were incubated at 37∘C for two further hoursThen the plates were washed three times with WB again
Development of the plates based on an enzymaticreaction of horseradish peroxidase (HRP) with TMB aschromogenic substrate Therefore the appropriate HRP-conjugated detection-antibody was added as a next step(Table 1) After incubation at 37∘C for another hour andthree washing steps with WB the plates were devel-oped by the addition of 50120583Lwell of freshly preparedTMB-solution (1mL of 001 g 331015840551015840-tetramethylbenzidine(Sigma-Aldrich Steinheim Germany) dissolved in 10mLdimethyl sulfoxide mixed with 9mL of 01M Na
2HPO4
005M citric acid pH 45ndash55 and 2120583L of H2O2) The
reaction was stopped by adding 50120583Lwell of 25 sulfuricacid (H
2SO4) Finally optical density (OD) was measured
at 450 nm and 650 nm as correction factor using an infiniteF200 pro ELISA-reader (Tecan)
After subtraction of the corresponding blank values of theplates experimental values were expressed as optical densityor as mean optical density of each serum when ELISA testswere performed in monoplicates or duplicates respectively
26 IgG-ELISA SNA In order to examine the sialylationof anti-1205732GP1-IgG we slightly modified the previouslydescribed ELISA protocol Before coating the 1205732GP1 antigenwas treated with 1 of periodic acid (PIA Sigma-Aldrich
4 Journal of Immunology Research
Table 2 Detection reagents for lectin ELISA
SNA-ELISAPrimary detectionreagent
Biotinylated SNA-lectin(Galab Technologies Geesthacht Germany)
Secondarydetection reagent
HRP-conjugated streptavidin(Jackson ImmunoResearch laboratoriesNewmarket Suffolk United Kingdom)
Dilution 1 10000SNA = Sambucus nigra agglutinin HRP = horseradish peroxidase
Steinheim Germany) for 48 hours at 43∘C to remove itssugar residues to exclude unspecific binding of Sambucusnigra lectin (SNA) to naive 1205732GP1 After that we centrifugedthe 1205732GP1-solution with 10K Amicon Ultra-centrifugal fil-ter devices (Millipore Tullagreen Ireland) to transfer thetreated 1205732GP1 into coating buffer (100mMNa
2CO3 100mM
NaHCO3 pH 96)
Then the plates were coated with 50 120583Lwell of a 2120583gmLsolution of the PIA-treated 1205732GP1 at 4∘C overnight andsubsequently blocked for two hours at 37∘C with 100 120583Lwellof PIA-treated (05) 3 gelatine (Sigma-Aldrich SteinheimGermany) in TBST buffer with 1mM CaCl
2and 1mM
MgCl2 The subsequent steps were performed as described
in the previous ELISA protocol with two exceptions Insteadof serum we added 50 120583Lwell of purified IgG-samples toexclude possible binding of SNA to anti-1205732GP1-IgM And thedevelopment of the plate required the addition of a seconddetection reagent (Table 2) which was incubated for onehour at room temperature and demanded three preliminarywashing steps We calculated the SNAanti-1205732GP1-IgG ratiofrom the OD values we obtained in the SNA ELISA and asimultaneously conducted anti-1205732GP1-IgG ELISA
27 Statistical Analysis Differences between the cohorts wereassessed using the one-way analysis of variance (ANOVA)test In the case of multiple testing the significance valueswere adjusted by the Newman-Keuls posttest correction
3 Results31 Analysis of Anti-1205732GP1-IgG Wemeasured the OD valuesof total anti-1205732GP1-IgG in the sera of 24 patients with PAPSand 22 patients with SLEAPS 16 healthy children and17NHD as controls As shown in Figure 1 the lowest IgGvalues were to be observed in the cohort of healthy donors(308mOD) whereas the values of the healthy children(970mOD) even exceeded that of the patients with APS(PAPS 686mOD 119901 = 0010 SLEAPS 546mOD 119901 lt0001) All differences between NHD and the other cohortsas well as between the children and the other cohorts weresignificant Patients with PAPS and SLEAPS did not differsignificantly
We calculated the 975 quantile of the anti-1205732GP1-IgGOD values obtained from the NHD cohort and set this valueas cut-off (738mOD) Sera exceeding this value were consid-ered seropositive for anti-1205732GP1-IgG Applying this cut-offvalue we obtained 31 seropositive samples (1224 PAPS 622SAPS 1316 healthy children) which were further analysed in
APS SLEAPS Healthy childrenNHDAPSSLEAPS ns
ns
NHD APS SLEAPS Children0
200
400
600
800
1000
1200
1400
1600
Ant
i-1205732
GP1
-IgG
(mO
D)
p values
mdashmdash
lt00001 lt001 lt00001
lt0001
lt00001
Figure 1 Total anti-1205732GP1 IgG in the sera of NHD PAPSSLEAPS and healthy children and the corresponding 119901 valuesNHD showed significantly lower mOD values than all other cohortswhereas healthy children represented the highest mOD valuesShort horizontal lines mean mOD values of all samples of thecohort Long broken line 975 quantile of NHD (=738mOD)NHD = normal healthy donor PAPS = primary antiphospholipidsyndrome SLEAPS = antiphospholipid syndrome with systemiclupus erythematosus OD = optical density
our study (Supplementary Figure 1 in Supplementary Mate-rial available online at httpdxdoiorg1011552015638129)
32 Analysis of the IgG-Subclasses of Anti-1205732GP1 Analysingthe absolute OD values of the anti-1205732GP1-IgG subclasseswe observed that the children showed significantly higherlevels of the anti-1205732GP1-IgG1 (69mOD) and -IgG3 subclasses(212mOD) compared to NHD PAPS and SAPS (Figures2(a) and 2(c) Table 3(a)) The differences between healthychildren and NHD regarding these two subclasses showeda higher level of significance than the comparison betweenhealthy children and patients with APS However healthychildren did not show significant differences for anti-1205732GP1-IgG2 and -IgG4 when compared to NHD PAPS and SAPS(Figures 2(b) and 2(d) Table 3(b))
Patients with APS (PAPS and SAPS) showed significantlyhigherODvalues for anti-1205732GP1-IgG1 -IgG2 and -IgG3 thanNHD (Figures 2(a) 2(b) and 2(c) Table 3(b)) As in theanti-1205732GP1-IgG ELISA we observed no significant differencebetween the patients with PAPS and SAPS
To summarize patients with APS showed higher ODvalues than NHD in IgG1 IgG2 and IgG3 IgG4 did notshow any significant differenceThe children in turn had thehighest OD values of the IgG1 and IgG3 subclasses
These results were consistent with the analysis of thenumber of seropositive samples as shown in Table 4 Todetermine seropositivity we calculated the mean OD and thestandard deviation (SD) of the NHD group for the differentIgG subclasses IgG values exceeding the NHDrsquos mean OD
Journal of Immunology Research 5
0
50
100
150
200
250
300
350
NHD APS SLEAPS Children
IgG
1 (m
OD
)
(a)
NHD APS SLEAPS Children0
50
100
150
200
250
300
350
IgG
2 (m
OD
)
(b)
NHD APS SLEAPS Children
940483 mOD
0
50
100
150
200
250
300
350
IgG
3 (m
OD
)
(c)
NHD APS SLEAPS Children
508 mOD 482 mOD 665 mOD 1753 mOD
0
50
100
150
200
250
300
350
IgG
4 (m
OD
)
(d)
0
50
100
150
200
250
300
350
(mO
D)
Children NHD APS
IgG4IgG3
IgG2IgG1
(e)
0
10
20
30
40
50
60
70
80
90
100
()
APSChildren
IgG4IgG3
IgG2IgG1
(f)
Figure 2mODvalues of anti-1205732GP1-IgG1 (a) anti-1205732GP1-IgG2 (b) anti-1205732GP1-IgG3 (c) and anti-1205732GP1-IgG4 (d) inNHD PAPS SLEAPSand childrenThe horizontal lines represent the cut-off for seropositivity of the corresponding anti-1205732GP IgG-subclass (gtmeanmOD
[NHD] +2 times SD
[NHD]) NHD = normal healthy donor PAPS = primary antiphospholipid syndrome SLEAPS = antiphospholipid syndrome withsystemic lupus erythematosus OD = optical density Median OD values of the anti-1205732GP1-IgG subclasses in the study cohorts (e) Regardingthe anti-1205732GP1-IgG subclass-distribution within the cohorts children showed a significantly higher contribution of IgG3 than IgG1 andIgG2 Patients with antiphospholipid syndrome (APS) showed a higher content of anti-1205732GP1-IgG2 than -IgG1 and -IgG3 NHD showed ahigher contribution of anti-1205732GP1-IgG2 than -IgG1 -IgG3 and -IgG4The columns represent the median OD values of IgG1 IgG2 IgG3 andIgG4 in the cohorts OD = optical density NHD = normal healthy donor Percentage distribution of anti-1205732GP1-IgG subclasses in childrenand patients with APS (f) Children harbour significantly less anti-1205732GP1-IgG2 than patients with APS but more anti-1205732GP1-IgG3 APS =antiphospholipid syndrome
6 Journal of Immunology Research
0
50
100
150
200
250
300
350
400
Children APS
C3c (
mO
D)
(a)
0
200
400
600
800
1000
1200
Children APS
IgM
(mO
D)
(b)
Figure 3 OD values of C3c (a) bound to anti-1205732GP1 and OD values of anti-1205732GP1-IgM (b) in sera of patients with APS and healthy childrenAnti-1205732GP1 of healthy children displayed significantly more bound C3c than that of patients with APS (119901 = 000007) This suggests a moreefficient clearance of 1205732GP1-immune complexes in the healthy children Anti-1205732GP1-IgM OD values of patients with APS were significantlyhigher than those of healthy children Horizontal lines represent the median OD value of the cohort APS = antiphospholipid syndrome OD= optical density
Table 3 (a) OD values of the anti-1205732GP1-IgG subclasses in theinvestigated cohorts (b) Comparison (119901 values) of the OD valuesof the anti-1205732GP1-IgG subclasses
(a)
IgG1 [mOD] IgG2 [mOD] IgG3 [mOD] IgG4 [mOD]NHD 24 64 27 67Children 69 77 212 210PAPS 30 85 45 72SAPS 28 74 55 173APS 29 81 48 106
(b)
IgG1 IgG2 IgG3 IgG4Children versus NHD lt00001 ns lt00001 nsChildren versus PAPS lt00001 ns lt0001 nsChildren versus SAPS lt0001 ns lt0001 nsAPS versus NHD ns ns ns nsPAPS versus SAPS ns ns ns nsOD = optical density 450 nm PAPS = primary antiphospholipid syndromeSAPS = secondary antiphospholipid syndrome APS = PAPS and SAPSNHD = normal healthy donors ns = not significant
plus two times the SD of the considered subclass were definedas seropositive (Supplementary Figure 1) Applying this cut-off we observed that the healthy children had high levels ofthe IgG1 and IgG3 subclasses with 13 (100) and 8 (62)seropositive samples respectively In contrast only 7 (39)and 0 (0) sera of patients with APS were seropositive forIgG1 and IgG3 respectively APS showed seropositivity ofanti-1205732GP1-IgG2 in 7 cases (39) whereas the cohort ofhealthy children contained just 2 seropositive sera (15) Just
few samples were seropositive for anti-1205732GP1-IgG4 in eachcohort representing a range from 8 (PAPS) to 17 (SAPS)
To determine the subclass distributionwithin the cohortswe calculated the contribution of themeanOD values of eachindividual subclass to the sum
As shown in Figures 2(e) and 2(f) we saw that for thehealthy children IgG3 was the most abundant subclass witha contribution of 40 (IgG3 gt IgG1 (119901 lt 0001) and IgG3 gtIgG2 (119901 lt 001)) In contrast IgG2 dominated the subclassprofile for NHD (46 IgG2 gt IgG1 IgG2 gt IgG3 and IgG2 gtIgG4 (119901 lt 00001)) and patients with APS (38 IgG2 gt IgG1(119901 lt 00001) IgG2 gt IgG3 (119901 = 0001)) The contributionof IgG1 ranged from 12 (SAPS) to 17 (NHD) and thatof IgG4 from 21 (PAPS) to 35 (SAPS) Comparing thecontribution of the individual subclasses between the studycohorts we observed that children showed significantly lessIgG2 than patients with APS and NHD (119901 lt 0001 119901 lt00001) and a higher IgG3 content than NHD and patientswith APS (119901 lt 00001 119901 lt 0001) No significant differencebetween the cohorts was to be seen for the contributions ofIgG1 and IgG4
33 Analysis of IgM C1q and C3c Associated to Anti-1205732GP1-IgG Next we analysed potential 1205732GP1-binding proteins inthe sera of healthy children and adults with APS The signalsfor C1q were low and did not differ significantly between thecohorts
However the ELISAs detecting C3c bound to anti-1205732GP1and anti-1205732GP1-IgM showed highly significant differencesbetween healthy children and adults with APSWhereas C3c-anti-1205732GP1 adducts were increased in healthy children (119901 lt00001 Figure 3(a)) anti-1205732GP1-IgM was decreased (119901 lt0001 Figure 3(b))
Journal of Immunology Research 7
Table 4 Number (frequency) of seropositive samples of the anti-1205732GP1-IgG subclasses
Number of seropositive samples (gtmOD[NHD] + 2 times SD[NHD])IgG1
(gt29mOD)IgG2
(gt79mOD)IgG3
(gt85mOD)IgG4
(gt319mOD)NHD (119899 = 17) 0 (0) 0 (0) 1 (6) 1 (6)Children (119899 = 13) 13 (100) 2 (15) 8 (62) 2 (15)PAPS (119899 = 12) 6 (50) 5 (42) 0 (0) 1 (8)SAPS (119899 = 6) 1 (17) 2 (33) 0 (0) 1 (17)APS (119899 = 18) 7 (39) 7 (39) 0 (0) 2 (11)OD = optical density 450 nm PAPS = primary antiphospholipid syndrome SAPS = secondary antiphospholipid syndrome APS = PAPS and SAPS NHD =normal healthy donorsThe cut-off for seropositivity was calculated from the mean OD of the corresponding anti-1205732GP1-IgG subclass of the NHD cohort andits standard deviation (SD) according to the formula mean OD[NHD] + 2 times SD[NHD] The respective cut-off OD values are indicated in parentheses below theanti-1205732GP1-IgG subclasses
Table 5 Cohorts studied with the lectin ELISA
Cohort Abbreviation Number of samplesPatients with primary APS PAPS 12Patients with APS and SLE as underlying disease SAPS 10Patients with SLE without symptoms of APS harbouring circulating aPL SLE + aPL 15Asymptomatic carriers of aPL aaPL 6Healthy children with circulating anti-1205732GP1 mdash 16APS = antiphospholipid syndrome aPL = antiphospholipid antibodies SLE = systemic lupus erythematosus
34 Analysis of the Sialylation of Anti-1205732GP1 We analysedthe glycosylation of anti-1205732GP1-IgG focusing on its sialy-lation since highly sialylated Fc-parts of IgG are known tocause rather anti-inflammatory responses [30] Wemeasuredthe binding of the sialic acid specific lectin SNA to the anti-1205732GP1-IgG As shown in Table 5 we employed five cohorts ofindividuals 12 patientswith PAPS 10 patientswith SAPS (SLEas primary disease) 15 patients with SLE who did not showclinical sign of APS but with aPL in their sera (SLE + aPL)6 asymptomatic adult carriers of aPL (aaPL) and 16 healthychildren harbouring aPL
Analysing the SNAIgG-ratios (Figure 4) we identifiedthe highest sialylation level in the sera of the healthy childrenThe significances were rather high and decreased from PAPS(119901 lt 00001) to SAPS (119901 lt 00001) to SLE + aPL (119901 lt00001) and to aaPL (119901 lt 0001)
4 Discussion
We analysed two distinct sets of sera One was used toinvestigate anti-1205732GP1-IgG its subclasses and its associatedproteins C1q and C3c For the analysis of the sialylation ofanti-1205732GP1-IgG we had to collect fresh sera from patientswith APS and the healthy children since the stability of theantibody-glycosylation is unknown In order to avoid itspossible decay by processes like enzymatic degradation thisperiod has been kept as short as possible and was followed byimmediate investigation of the glycosylation
The antiphospholipid syndrome is characterized by clin-ical manifestations such as arterial and venous thrombosisor pregnancy-related complications and the detection of spe-cific autoantibodies (lupus anticoagulant anti-1205732GP1- and
anticardiolipin-autoantibodies) [1] In particular anti-1205732GP1sare considered responsible for provoking the pathologicalcomplications [3]We firstmeasured the total amount of anti-1205732GP1-IgG in the sera of 16 healthy children without clinicalmanifestations of APS and 46 adult patients with APS toidentify individual cohorts seropositive for anti-1205732GP1 17adult normal healthy donors served as negative control
The samples of the patientswithAPS showedhigher levelsof anti-1205732GP1-IgG than those of NHD We also observeda higher concentration of anti-1205732GP1-IgG in the sera ofhealthy children compared to NHD This is due to the factthat the detection of these antibodies has been the inclusioncriterion for the healthy children examined in this studyInterestingly the children showed even higher OD values forthe putatively pathogenic anti-1205732GP1-IgG than the patientswith APS This finding was surprising since the childrenwere clinically healthy and did not show any signs of APSsuch as thrombosis or thrombocytopenia complications inwhich the existence of aPL especially anti-1205732GP1 is maderesponsible for [7 33]
High levels of circulating aPL in 61 healthy children whencompared to healthy adults have already been described byAvcin and colleagues [18] The anti-1205732GP1-IgG was report-edly higher in preschool children with a mean age of 5years than in adolescents with a mean age of 135 years[18] The strongly elevated circulating anti-1205732GP1-IgG in ourcohort of healthy children may be explained by the fact thatthey had an even lower age (mean age was nine months)One possible explanation for this result is the inductionof anti-1205732GP1 by viral and bacterial infections [10 11] ornutritional exposure to 1205732GP1 [17] As the immune systemin children is still to evolve they are more susceptible to
8 Journal of Immunology Research
0
05
10
15
20
25SN
AIg
G
ChildrenPAPS SAPS aaPLSLE + aPL
lowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowast
Figure 4 SNAanti-1205732GP1-IgG ratio in PAPS SAPS SLE + aPLaaPL and children Healthy children show a significantly higherSNAanti-1205732GP1-IgG ratio than PAPS SAPS SLE + aPL and aaPL(lowastlowast119901 lt 00001 lowastlowastlowast119901 lt 000001) This suggests higher sialylation ofthe oligosaccharides attached to Fc fragments of the childrenrsquos anti-1205732GP1No significant differenceswere foundbetween the remainingcohorts Horizontal lines indicate the mean SNAIgG ratio of thecohort SNA = Sambucus nigra agglutinin APS = antiphospholipidsyndrome PAPS = patients with primary APS SAPS = patients withAPS and SLE as underlying disease SLE + aaPL = patients withSLE without symptoms of APS harbouring circulating aPL aaPL =asymptomatic carriers of aPL aPL = antiphospholipid antibodiesSLE = systemic lupus erythematosus
infections than adults and their immune system has to copewith a plethora of viral and bacterial pathogens for the firsttime in their life This scenario is prone to result in thestimulation of autoantibody-production Molecular mimicryof certain pathogens can lead to the production of antibodieswhich then cross-react with 1205732GP1 [7 12ndash15] When infantsdiversify their dietary habits and start to consume productslike milk and meat they can get into contact with nutritional1205732GP1 [19] This is a further condition capable triggeringthe production of anti-1205732GP1 Exposure to pathogen-derivedand dietary 1205732GP1 could explain the high level of circulatinganti-1205732GP1-IgG in healthy childrenThis low affine reactivityis consecutively lost during the maturation of the immunesystem and replaced by the emergence in adults of high affineantibodies that more specifically recognize pathogens Apartfrom possible origins of these autoantibodies it is obviousthat they do not provoke any of the hallmark symptoms ofAPS In contrast to their adult counterparts anti-1205732GP1-IgGscirculating in the plasma of healthy children are obviouslynot pathogenic So it seems decent to hypothesize thatanti-1205732GP1 autoantibodies in healthy children biochemicallydiffer from those in adults
To test this hypothesis we examined the anti-1205732GP1-IgG subclass distribution in the seropositive sera of the anti-1205732GP1-IgG ELISA The healthy children showed higher ODvalues for the IgG1- and IgG3-subclasses than patients withAPS and NHD This was consistent with the number ofseropositive samples In the anti-1205732GP1-IgG1-subclass every
tested serum of the healthy children surpassed the thresholdset for seropositivity (gtmean OD
[NHD] + 2 times SD[NHD]) and
8 samples (62) did so in the IgG3-subclass In the seraof patients with APS just 7 (39) and no samples wereseropositive for IgG1 or IgG3 In the sera of patients with APScompared to NHD we observed higher OD levels of anti-1205732GP1 in all subclasses except for IgG4This result is consis-tent with the findings of Guerin et al who reported elevatedlevels in all anti-1205732GP1-IgG subclasses even including IgG4in patients with APS compared to a control group of healthyadults [23]
It might be that the elevated absolute OD values of theanti-1205732GP1-IgG subclasses are the logical consequence ofthe generally higher level of anti-1205732GP1-IgG in the patientswith APS and healthy children who were selected for furthertesting Therefore we also analysed the subclass distributionof this antibody We first analysed the anti-1205732GP1-IgG sub-class distribution within the individual cohorts and observedthat in the healthy children the IgG3 antibody was mostabundant with a contribution of 40 to the total reactivityThis is consistent with the finding that all sera of the healthychildren were seropositive in this subclass and that theiranti-1205732GP1-IgG3 OD values were the highest of all cohortsThis indicates a predominance of this subclass in the healthychildren The remaining anti-1205732GP1-IgG subclasses in thesera of the children ranged from 15 (IgG1) over 20 (IgG2)to 25 (IgG4) but the differences between their proportionswere not statistically significant In contrast in the sera ofthe patients with APS the anti-1205732GP1-IgG2 subclass had thehighest contribution with 38
The finding of an elevated proportion of the IgG2 subclassin the patients with APS confirms former published datawhich also describe a skewing towards anti-1205732GP1-IgG2 inAPS [19 23 26 27] The authors detected an even highercontribution of anti-1205732GP1-IgG2 (702 [26] and 87 [27]resp) In contrast to our results the IgG1 subclass wasmore prominent in the published studies described aboveImportantly in the cohort of the healthy infants IgG2 wasrelatively lowTherefore onemay argue that anti-1205732GP1-IgG2is not just a marker of the disease but is also involved inthe pathogenic action of the aPL autoantibodies When wecompared the anti-1205732GP1-IgG subclass distribution betweenthe three cohorts we noticed a significant difference in thedistribution concerning the children on one hand and NHDand patients with APS on the other Healthy children showeda higher proportion of anti-1205732GP1-IgG3 and a lower one ofanti-1205732GP1-IgG2 In NHD and patients with APS it was theother way round IgG1 and IgG4 did not significantly differbetween healthy children and patients with APS orNHDThepredominance of anti-1205732GP1-IgG3 in the healthy children isa hint to an immune response elicited by a proteinaceousantigen [26 27 34] This supports the hypothesis thatthe harmless anti-1205732GP1 in children are infection-inducedwith pathogen-derived proteins serving as trigger for theirproduction However it is still elusive if the IgG3-dominatedanti-1205732GP1 response found in the sera of healthy childrencontributes to its low pathogenic potential Alternativelyit can just be ascribed to the proceeding evolution of theinfantrsquos immune system typically associated with an elevated
Journal of Immunology Research 9
proportion of IgG3 antibodies [34 35] During childhoodthe proportion of IgG3 continuously decreases to adult rangesusually reached at the age of 10ndash12 years However the aboveaverage elevated proportion of the anti-1205732GP1-IgG3 (40) inthe sera of the healthy childrenmay thus not just be due to theinfant immune system In particular when bearing in mindthat normal adult levels of the IgG3 subclass are just around6 [34 36]
IgG3 is the IgG-subclass exerting the most effectiveactivation of the complement cascade through the classicalpathway [34] Since complement activation is associated withthe pathogenesis of APS [21 32 37] the high proportionof complement-activating anti-1205732GP1-IgG3 in the childrenseems to contradict a possible protective effect against APS ofthese kinds of antibodies However the role of complement inthe etiopathogenesis of autoimmune diseases is manifold asthe complement components C1q C2 and C4 contribute tothe clearance of postapoptotic secondary necrotic cells andtheir deficiency is therefore a risk factor for autoimmunityand autoinflammation [38] In a similar manner studiesin mouse in vivo models of various autoimmune diseasesdemonstrated that IgG mediated tissue inflammation wasblocked in mice deficient in activating Fc120574Rs although thecomplement component C3 was still abundantly depositedin the tissue [39ndash41] Thus the contribution of complementdeposits in tissue with respect to tissue inflammation remainsto be established
To obtain new insights into the involvement of thecomplement system in the pathogenesis of APS we analysedthe C1q- and C3c-binding to anti-1205732GP1 We observed thatanti-1205732GP1 in the sera of the healthy children and in patientswith APS similarly bound C1q the first molecule of theclassical pathway of complement activation [42] Performinganti-1205732GP1-IgM-ELISAs we detected significantly higherIgM values in the patients with APS One would expecta higher C1q-binding of the IgM positive sera since thisimmunoglobulin binds and activates C1q more strongly thanIgG However the children mainly harbour IgG3 autoanti-bodies the most potent subclass for the activation of theclassical complement pathway [34]This may compensate forthe lower IgM level in their sera and thus be responsiblefor the similar C1q-binding of both cohorts This suggeststhat preconditions for complement activation via the classicalpathway are comparable in both cohorts We conclude thatclassical complement activation has a minor impact on thedifferential pathogenesis of aPL in healthy children andpatients with APS
What then is the culprit that induces their pathogenicpotential
Nextwe quantified the contribution ofC3c to the immunecomplexes bound to 1205732GP1 C3c is a domain in the C3bmolecule and one of its cleavage products created aftercleavage by factor I of C3b [42 43] C3c not just representsa marker of complement activation but also serves as anopsonin improving the clearance of bound targets Weobserved significantly more C3c bound to anti-1205732GP1 inthe sera of the healthy children than in those of patientswith APS This reflects a better opsonization by C3c ofthe potentially harmful anti-1205732GP1 immune complexes
anti-1205732GP1-opsonized cells or cellular fragments in thesera of the healthy children Consequently the anti-1205732GP1complexes can be cleared more efficiently by the powerfulcomplement system of the healthy children In patients withchronic inflammatory autoimmune diseases complement isusually low which may result from null alleles [44] or fromcomplement consumption [38]
But why do these processes not cause inflammation andaberrant coagulation in the childrenApart fromanti-1205732GP1-IgG subclass distribution and distinct ability to activate thecomplement system it is likely that other properties of theanti-1205732GP1 in the sera of children contribute to their lowpathogenicity We hypothesized that an increased sialylationof the anti-1205732GP1-IgG in healthy individuals renders themharmless for their hosts IgG is usually N-glycosylated at theCH2-domain of its Fc-part This N-glycan crucially modifiesthe effector functions the IgG molecule is endowed with[45ndash47] This biantennary core-fucosylated oligosaccharidecarries a variable amount of sugar monomers attached toboth arms [28 48] IgG antibodies terminating with sialicacids on one or both antennae have been shown to behaveanti-inflammatorily after infusion of IVIG [29 31]The signalexerted by the terminal sialic acid is dominant since thefraction of sialylated IgG in healthy plasma is just 4 to 14[28 46 49]
In mice these effects are mainly ascribed to decreasedbinding of sialylated IgG to activating Fc120574-receptors andan upregulation of the inhibitory Fc120574RIIB [29 31] Weobserved that anti-1205732GP1-IgG isolated from sera of healthychildren was significantly more sialylated than that fromsymptomatic patients with APS One may predict that theseautoantibodies in complex with 1205732GP1 show low affinity foractivating Fc120574Rs and therefore do not provoke proinflam-matory and procoagulant effector functions Fc120574RIIA is anexample for an activating receptor It is located on a plethoraof cells On the surfaces of platelets it mediates activationby immune complexes The activation is accompanied byplatelet aggregation and the release of mediators such asthe prothromboticThromboxaneA2 [50] Consecutively thiscascade can result in thrombus formation [51 52] Sialylatedanti-1205732GP1-IgG is unable to bind to and to activate plateletsvia Fc120574RIIA
Further mechanisms involved in thrombus formation inAPS comprise the interaction of anti-1205732GP1 with variouspattern recognition receptors [53] and its interference withregulatory functions ofmolecules involved in the coagulationprocess (protein C prothrombin or tissue factor [52 54])Direct binding to 1205732GP1 on endothelial cells (EC) shiftstheir phenotype towards coagulation and inflammation [54]Considering that Fc120574R-binding is involved in many of thesemechanisms it seems likely that a high sialylation of anti-1205732GP1-IgG leads to a low pathogenicity of these autoan-tibodies and does not cause APS Therefore we concludethat the higher sialylation of anti-1205732GP1-IgG in the seraof the healthy children plays a major role in protectingthem from typical complications of APS This hypothesis isfurther supported by the observation that antiphospholipidantibodies from sera of asymptomatic adult carriers aremore sialylated than those from patients with APS A high
10 Journal of Immunology Research
sialylation status of antiphospholipid antibodies seems toprevent the development of symptoms of APS also in theseindividuals
We conclude that a plethora of factors contributes tothe low pathogenicity of anti-1205732GP1-IgG in the sera ofhealthy children and asymptomatic adults We observed adistinct spectrum of IgG-subclasses compared to patientswith APS and a better binding of C3c indicating a moreefficient clearance of potentially harmful anti-1205732GP1 Thehypersialylation of the anti-1205732GP1-IgG is prone to amelioratetheir inflammatory and procoagulatory effects in healthycarriers whereas hyposialylation and inflammationmutuallyprecipitate the antiphospholipid syndrome
Disclosure
Thepresent workwas performed in fulfillment of the require-ments for obtaining the degree ldquoDr medrdquo for ChristophFickentscher
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Laura Andreoli andMartin Herrmann contributed equally tosenior authorship
Acknowledgments
This project was supported by the German Research Foun-dation (training research programme of the collaborativeresearch center 643) by the Emerging Fields Initiative (EFI)of the FAU Erlangen-Nuremberg and by the K und RWucherpfennig-Stiftung
References
[1] S Miyakis M D Lockshin T Atsumi et al ldquoInternationalconsensus statement on an update of the classification criteriafor definite antiphospholipid syndrome (APS)rdquo Journal ofThrombosis and Haemostasis vol 4 no 2 pp 295ndash306 2006
[2] W A Wilson A E Gharavi T Koike et al ldquoInternationalconsensus statement on preliminary classification criteria fordefinite antiphospholipid syndrome report of an internationalworkshoprdquo Arthritis and Rheumatism vol 42 no 7 pp 1309ndash1311 1999
[3] P G de Groot and R T Urbanus ldquoThe significance of autoan-tibodies against beta2-glycoprotein Irdquo Blood vol 120 no 2 pp266ndash274 2012
[4] C Nalli S Piantoni L Andreoli M Motta and A TincanildquoAntiphospholipid syndrome and antiphospholipid antibodiesin children the two sides of the coinrdquo Israel Medical AssociationJournal vol 14 no 5 pp 310ndash312 2012
[5] I Schousboe ldquo1205732-glycoprotein I a plasma inhibitor of thecontact activation of the intrinsic blood coagulation pathwayrdquoBlood vol 66 no 5 pp 1086ndash1091 1985
[6] J George M Blank Y Levy et al ldquoDifferential effects of anti-1205732-glycoprotein I antibodies on endothelial cells and on themanifestations of experimental antiphospholipid syndromerdquoCirculation vol 97 no 9 pp 900ndash906 1998
[7] R Willis and S S Pierangeli ldquoAnti-1205732-glycoprotein I antibod-iesrdquo Annals of the New York Academy of Sciences vol 1285 no1 pp 44ndash58 2013
[8] M J Fischer J Rauch and J S Levine ldquoThe antiphospholipidsyndromerdquo Seminars in Nephrology vol 27 no 1 pp 35ndash462007
[9] H Amital M Govoni R Maya et al ldquoRole of infectious agentsin systemic rheumatic diseasesrdquo Clinical and ExperimentalRheumatology vol 26 no 1 supplement 48 pp S27ndashS32 2008
[10] T Avcin ldquoAntiphospholipid syndrome in childrenrdquo CurrentOpinion in Rheumatology vol 20 no 5 pp 595ndash600 2008
[11] M Garcıa-Carrasco C Galarza-Maldonado C Mendoza-Pinto R O Escarcega and R Cervera ldquoInfections and theantiphospholipid syndromerdquo Clinical Reviews in Allergy ampImmunology vol 36 no 2-3 pp 104ndash108 2009
[12] P Cruz-Tapias M Blank J-M Anaya and Y ShoenfeldldquoInfections and vaccines in the etiology of antiphospholipidsyndromerdquo Current Opinion in Rheumatology vol 24 no 4 pp389ndash393 2012
[13] P von Landenberg Y Doring S Modrow and K J LacknerldquoAre antiphospholipid antibodies an essential requirement foran effective immune response to infectionsrdquoAnnals of the NewYork Academy of Sciences vol 1108 pp 578ndash583 2007
[14] M Blank I Krause M Fridkin et al ldquoBacterial induction ofautoantibodies to 1205732-glycoprotein-I accounts for the infectiousetiology of antiphospholipid syndromerdquo Journal of ClinicalInvestigation vol 109 no 6 pp 797ndash804 2002
[15] Y Shoenfeld M Blank R Cervera J Font E Raschi and P-LMeroni ldquoInfectious origin of the antiphospholipid syndromerdquoAnnals of the Rheumatic Diseases vol 65 no 1 pp 2ndash6 2006
[16] A E Gharavi S S Pierangeli R G Espinola X Liu MColden-Stanfield and E N Harris ldquoAntiphospholipid antibod-ies induced in mice by immunization with a cytomegalovirus-derived peptide cause thrombosis and activation of endothelialcells in vivordquo Arthritis and Rheumatism vol 46 no 2 pp 545ndash552 2002
[17] M Motta G Chirico C B Rebaioli et al ldquoAnticardiolipin andanti-beta2 glycoprotein I antibodies in infants born to motherswith antiphospholipid antibody-positive autoimmune diseasea follow-up studyrdquoAmerican Journal of Perinatology vol 23 no4 pp 247ndash251 2006
[18] T Avcin A Ambrozic M Kuhar T Kveder and B RozmanldquoAnticardiolipin and anti-1205732 glycoprotein I antibodies in seraof 61 apparently healthy children at regular preventive visitsrdquoRheumatology vol 40 no 5 pp 565ndash573 2001
[19] A Ambrozic ldquoAnti-beta2-glycoprotein I antibodies in children
with atopic dermatitisrdquo International Immunology vol 14 no 7pp 823ndash830
[20] J A Giron-Gonzalez E Garcıa Del Rıo C Rodrıguez JRodrıguez-Martorell and A Serrano ldquoAntiphospholipid syn-drome and asymptomatic carriers of antiphospholipid anti-body prospective analysis of 404 individualsrdquo Journal ofRheumatology vol 31 no 8 pp 1560ndash1567 2004
[21] K A Breen P Seed K Parmar GWMoore S E Stuart-Smithand B J Hunt ldquoComplement activation in patients with isolatedantiphospholipid antibodies or primary antiphospholipid syn-dromerdquo Thrombosis and Haemostasis vol 107 no 3 pp 423ndash429 2012
Journal of Immunology Research 11
[22] F Fischetti P Durigutto V Pellis et al ldquoThrombus formationinduced by antibodies to 1205732-glycoprotein I is complementdependent and requires a priming factorrdquo Blood vol 106 no7 pp 2340ndash2346 2005
[23] J Guerin E Casey C Feighery and J Jackson ldquoAnti-Beta 2-glycoprotein I antibody isotype and IgG subclass in antiphos-pholipid syndrome patientsrdquo Autoimmunity vol 31 no 2 pp109ndash116 1999
[24] L Andreoli C Nalli M Motta et al ldquoAnti-1205732-glycoprotein I
IgG antibodies from 1-year-old healthy children born to moth-ers with systemic autoimmune diseases preferentially targetdomain 45 might it be the reason for their lsquoinnocentrsquo profilerdquoAnnals of the Rheumatic Diseases vol 70 no 2 pp 380ndash3832011
[25] B de Laat R H W M Derksen M van Lummel M T T Pen-nings and P G deGroot ldquoPathogenic anti-beta2-glycoprotein Iantibodies recognize domain I of beta2-glycoprotein I only aftera conformational changerdquo Blood vol 107 no 5 pp 1916ndash19242006
[26] M Samarkos K A Davies C Gordon M J Walport and SLoizou ldquoIgG subclass distribution of antibodies against 1205732-GP1and cardiolipin in patients with systemic lupus erythematosusand primary antiphospholipid syndrome and their clinicalassociationsrdquo Rheumatology vol 40 no 9 pp 1026ndash1032 2001
[27] J Arvieux B Roussel D Ponard and M G Colomb ldquoIgG2subclass restriction of anti-1205732 glycoprotein 1 antibodies inautoimmune patientsrdquo Clinical and Experimental Immunologyvol 95 no 2 pp 310ndash315 1994
[28] J N Arnold M R Wormald R B Sim P M Rudd and R ADwek ldquoThe impact of glycosylation on the biological functionand structure of human immunoglobulinsrdquo Annual Review ofImmunology vol 25 pp 21ndash50 2007
[29] Y Kaneko F Nimmerjahn and J V Ravetch ldquoAnti-inflamma-tory activity of immunoglobulin G resulting from Fc sialyla-tionrdquo Science vol 313 no 5787 pp 670ndash673 2006
[30] RM Anthony and F Nimmerjahn ldquoThe role of differential IgGglycosylation in the interaction of antibodies with FcgammaRsin vivordquo Current Opinion in Organ Transplantation vol 16 no1 pp 7ndash14 2011
[31] R M Anthony and J V Ravetch ldquoA novel role for the IgGFc glycan the anti-inflammatory activity of sialylated IgG FcsrdquoJournal of Clinical Immunology vol 30 no 1 pp S9ndashS14 2010
[32] K Oku T Atsumi M Bohgaki et al ldquoComplement activationin patients with primary antiphospholipid syndromerdquoAnnals ofthe Rheumatic Diseases vol 68 no 6 pp 1030ndash1035 2009
[33] H P McNeil R J Simpson C N Chesterman and S AKrilis ldquoAnti-phospholipid antibodies are directed against acomplex antigen that includes a lipid-binding inhibitor of coag-ulation 1205732-Glycoprotein I (apolipoprotein H)rdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 87 no 11 pp 4120ndash4124 1990
[34] R Jefferis and D S Kumararatne ldquoSelective IgG subclassdeficiency quantification and clinical relevancerdquo Clinical andExperimental Immunology vol 81 no 3 pp 357ndash367 1990
[35] P H Schur F Rosen and M E Norman ldquoImmunoglobulinsubclasses in normal childrenrdquo Pediatric Research vol 13 no3 pp 181ndash183 1979
[36] M A H French and G Harrison ldquoSerum IgG subclass concen-trations in healthy adults a study using monoclonal antiserardquoClinical and Experimental Immunology vol 56 no 2 pp 473ndash475 1984
[37] M Odorczuk L B Keil and V A DeBari ldquoBinding of com-plement component C1q to anti-beta2 glycoprotein I antibodiesfrom patients with antiphospholipid syndromerdquo InflammationResearch vol 48 no 10 pp 524ndash526 1999
[38] L Truedsson A A Bengtsson and G Sturfelt ldquoComplementdeficiencies and systemic lupus erythematosusrdquoAutoimmunityvol 40 no 8 pp 560ndash566 2007
[39] S Aschermann C H K Lehmann S Mihai G Schett DDudziak and F Nimmerjahn ldquoB cells are critical for autoim-mune pathology in Scurfy micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 110 no47 pp 19042ndash19047 2013
[40] Y Kaneko F Nimmerjahn M P Madaio and J V RavetchldquoPathology and protection in nephrotoxic nephritis is deter-mined by selective engagement of specific Fc receptorsrdquo TheJournal of Experimental Medicine vol 203 no 3 pp 789ndash7972006
[41] D Sylvestre R Clynes M Ma H Warren M C Carrolland J V Ravetch ldquoImmunoglobulin G-mediated inflammatoryresponses develop normally in complement-deficient micerdquoJournal of ExperimentalMedicine vol 184 no 6 pp 2385ndash23921996
[42] G E Ritchie B E Moffatt R B Sim B P Morgan R A Dwekand P M Rudd ldquoGlycosylation and the complement systemrdquoChemical Reviews vol 102 no 2 pp 305ndash320 2002
[43] Y Palarasah K Skjodt J Brandt et al ldquoGeneration of aC3c specific monoclonal antibody and assessment of C3c as aputative inflammatorymarker derived from complement factorC3rdquo Journal of ImmunologicalMethods vol 362 no 1-2 pp 142ndash150 2010
[44] D G Doherty J A Underhill P T Donaldson et al ldquoPoly-morphism in the human complement C4 genes and geneticsusceptibility to autoimmune hepatitisrdquo Autoimmunity vol 18no 4 pp 243ndash249 1994
[45] M-H Tao and S L Morrison ldquoStudies of aglycosylatedchimeric mouse-human IgG Role of carbohydrate in thestructure and effector functions mediated by the human IgGconstant regionrdquoThe Journal of Immunology vol 143 no 8 pp2595ndash2601 1989
[46] R Jefferis ldquoGlycosylation of recombinant antibody therapeu-ticsrdquo Biotechnology Progress vol 21 no 1 pp 11ndash16 2005
[47] Y Mimura S Church R Ghirlando et al ldquoThe influence ofglycosylation on the thermal stability and effector functionexpression of human IgG1-Fc properties of a series of truncatedglycoformsrdquoMolecular Immunology vol 37 no 12-13 pp 697ndash706 2001
[48] C Huhn M H J Selman L R Ruhaak A M Deelder and MWuhrer ldquoIgG glycosylation analysisrdquo Proteomics vol 9 no 4pp 882ndash913 2009
[49] J Lund N Takahashi J D Pound M Goodall and R JefferisldquoMultiple interactions of IgG with its core oligosaccharide canmodulate recognition by complement and human Fc gammareceptor I and influence the synthesis of its oligosaccharidechainsrdquo The Journal of Immunology vol 157 no 11 pp 4963ndash4969 1996
[50] R Forastiero M Martinuzzo L O Carreras and J MacloufldquoAnti-1205732 glycoprotein I antibodies and platelet activation inpatients with antiphospholipid antibodies association withincreased excretion of platelet-derived thromboxane urinarymetabolitesrdquoThrombosis and Haemostasis vol 79 no 1 pp 42ndash45 1998
12 Journal of Immunology Research
[51] J-C Reverter and D Tassies ldquoMechanism of thrombosis inantiphospholipid syndrome binding to plateletsrdquo in HughesSyndrome M A Khamashta Ed pp 403ndash414 Springer Lon-don UK 2006
[52] G Espinosa R Cervera J Font and Y Shoenfeld ldquoAntiphos-pholipid syndrome pathogenic mechanismsrdquo AutoimmunityReviews vol 2 no 2 pp 86ndash93 2003
[53] T Shi B Giannakopoulos X Yan et al ldquoAnti-1205732-glycoprotein
I antibodies in complex with 1205732-glycoprotein I can activate
platelets in a dysregulated manner via glycoprotein Ib-IX-VrdquoArthritis amp Rheumatism vol 54 no 8 pp 2558ndash2567 2006
[54] P LMeroni E RaschiMCamera et al ldquoEndothelial activationby aPL a potential pathogenetic mechanism for the clinicalmanifestations of the syndromerdquo Journal of Autoimmunity vol15 no 2 pp 237ndash240 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Journal of Immunology Research
composed of five domains [4] and it is known to be aninhibitor of the contact activation of the intrinsic coagulationpathway [5] Additionally aPL can cause a prothromboticstate in individuals by activating platelets and endothelialcells resulting in a higher expression of tissue factor or in theupregulation of proinflammatory cytokines and cell adhesionmolecules [6 7]
Patients with APS can be classified into two groupsdepending on the occurrence of further diseases (I) patientsmerely suffering from APS (primary antiphospholipid syn-drome PAPS) and (II) those with an underlying systemiclupus erythematodes (SLE) The latter condition is referredto as secondary antiphospholipid syndrome (SAPS) [8]
Up to now a variety of hypotheses about possible triggersfor the induction of anti-1205732GP1 have been proposed Pub-lished studies suggest that their production can be provokedduring infectious diseases like parvovirus B19 HIV CMVor HCV [3 9ndash11] Certain molecular structures of theseinfectious agents may resemble structures of 1205732GP1 and thuscause the induction of cross-reacting antibodies [7 12ndash16]Another one claims that nutritional exposure to 1205732GP1 couldalso trigger the production of transient and harmless anti-1205732GP1 in children [4 17ndash19]
Interestingly anti-1205732GP1 can be detected in a quite largeproportion of the healthy population who do neither sufferfrom APS nor another autoimmune disease [3 18 20]As complication-free periods of patients with APS occuralthough the aPL are persistently detectable in their circula-tion [15] it seems to be likely that the mere presence of theseantibodies is not sufficient for the development of clinicalactive APS complications
For this reason a two-hit hypothesis has been proposedto provide a putative explanation for this observation (I)the production of anti-1205732GP1 represents an initial hit thatincreases the risk for thrombotic events and (II) infectiousagents can then serve as the second hit provoking the typicalmanifestations of APS by activating toll-like receptors orcomplement [9 15 21 22] This two-hit hypothesis surelyprovides a good first model for the etiopathogenesis of APSYet it is not completely satisfying and does not explain whysome people harbouring aPL remain healthy Some studyresults indicate that this could be grounded in structuraldifferences of anti-1205732GP1 found in patients with APS andhealthy children [4 23]
Comparing obviously harmless aPL from healthy chil-dren with putatively pathogenic aPL from adults with APSwe suggested a difference in the epitope specificity as differ-entiating factor Anti-1205732GP1 from the sera of the childrenspecifically bound to the domain IVV of 1205732GP1 whereasanti-1205732GP1 from adults with APS preferentially targeteddomain I [24] These results were consistent with thosedescribed in other studies [19 25] Besides distinct epitopebinding specificities a different composition of the spectrumof the anti-1205732GP1 IgG subclasses between patients with APSand healthy carriers of this antibody has also been described[19 23 26 27]
The differences in glycosylation of antibodies which areaccompanied by crucial modifications of their effector func-tions [28] provide another promising approach in performing
structural analyses of anti-1205732GP1 IgG molecules possess abiantennary oligosaccharide attached to the asparagine atposition 297 of the Fc portion (Asn-297) Various glycoformscontaining distinct sugar moieties have been described [2829] Interestingly it has been observed that IgG moleculeswhose glycans terminate with sialic acid residues showa decreased affinity to activating Fc120574-receptors (Fc120574Rs)while simultaneously provoking an upregulation of theinhibitory Fc120574RIIB [29ndash31] These effects lead to a ratheranti-inflammatory activity andmight consequently be able toprevent potentially harmful immune responses
In particular in children aPL can readily be detectedalthough they do not show any clinical signs of APS In astudy carried out by Avcin et al the prevalence of anti-1205732GP1 in 61 healthy children was 66 [18] Based on thisobservation we intended to find out why these circulatingautoantibodies do not cause harm in their hosts We deter-mined the IgG-subclass distribution in the samples fromhealthy children and patients with APS and examined thebinding of the complement factors C1q and C3c to anti-1205732GP1 immune complexes since an important involvementof the complement system in the pathogenesis of APS hasbeen reported [21 22 32] Finally we analysed the sialylationstatus of anti-1205732GP1-IgG isolated from healthy childrenasymptomatic adult carriers and patients with APS
Our results revealed that compared to adults with APSthe healthy children showed a higher content of IgG3 at thecost of IgG2 of the anti-1205732GP1 Furthermore anti-1205732GP1-IgGof healthy children showed a higher sialylation and displayedmore bound C3c Hyposialylated IgG and inflammationmutually influence the pathogenicity of the antiphospholipidautoantibodies and form a vicious pathogenic circle
2 Materials and Methods21 Patients For the investigation of anti-1205732GP1-IgG itssubclasses C1q- and C3c-binding and anti-1205732GP1-IgM westudied sera of 46 adult patients (24 patientswith PAPS and 22with SAPS) and 16 healthy children harbouring anti-1205732GP1without any clinical manifestations associated with APS fromour partner hospital in Brescia Italy As a control groupwe examined sera of 17 age and sex matched adult normalhealthy donors (NHD) without aPL Inclusion criteria for thechildren were the presence of anti-1205732GP1-IgG no infectiousdiseases in the course of three months before blood takingand no chronic pathology Additionally it was excluded thattheir parents suffered from any autoimmune disease At thetime of blood taking the mean age of the children was about9 months (median age 85 months range 3 months and 5daysndash19 months and 10 days)
Since sugar residues attached to molecules can be dam-aged by repeated thawing and freezing we received newsera from our partner hospital in Brescia Italy to analysethe sialylation of anti-1205732GP1-IgG We obtained sera from 12patients with PAPS and 10 patients with SAPS 15 patientswith SLE and aPL but without clinical evidence of APSand sera from 6 asymptomatic carriers of aPL Furthermorewe obtained sera from 16 healthy children showing nosymptoms of APS although harbouring anti-1205732GP1-IgG in
Journal of Immunology Research 3
Table 1 Antibodies employed for each ELISA
ELISA Detection-antibody DilutionAnti-1205732GP1-IgG HRP-conjugated rabbit F(ab1015840) 2 anti-human-IgGlowast 1 20000
Anti-1205732GP1-IgG subclasses HRP-conjugated mouse anti-human IgG1 2 3 and 4 (Clones HP6001HP6002 HP6050 amp HP6025)lowast 1 20000
Anti-1205732GP1-IgM HRP-conjugated goat anti-human-IgMlowast 1 20000Anti-1205732GP1-bound C3c HRP-conjugated sheep anti-C3clowastlowast 1 500Anti-1205732GP1-bound C1q HRP-conjugated sheep anti-C1qlowastlowast 1 500lowast purchased from Southern Biotech Birmingham Alabama USAlowastlowast purchased from Abcam Cambridge United KingdomHRP = horseradish peroxidase
their circulation (mean age 95 months median age 10months range 3 months and 5 daysndash19 months and 10 days)
22 Blood Drawing Heparinised venous blood was drawnby venipuncture and processed within two hours Serum wasobtained by centrifugation at 1000 g for 15 minutes of clottedblood then aliquoted frozen and stored at minus20∘C until usefor investigation
Informed consent was obtained from all blood donorsand the childrenrsquos parents respectively and the study wasapproved by the ethics committees of the University HospitalErlangen and the Spedali Civili Brescia
23 Isolation of 1205732GP1 For 1205732GP1-isolation from dilutedsera of normal healthy donors (in 015MNaCl) percloric acidwas slowly added until the mixture got a creamy aspect Aftercentrifugation at 10000 g the supernatant was neutralizedwith NaOH and dialyzed against a buffer of 002M Tris and003MNaCl pH8Then itwas given into columns containingHiTrap heparin and contaminants of the supernatant wereeluted by the addition of 002M Tris and 015M NaCl pH 8In order to elute semipurified 1205732GP1 a buffer of 002M Trisand 035MNaCl pH 8 was injected into the columns and thefractions have been pooled and dialyzed against a solutioncontaining 005M acetate buffer 005M NaCl pH 48 Thismaterial was filtrated with 02120583membranes and injected intoresin columns equilibrated with 005M acetate buffer 005MNaCl pH 48 After washing with 005M acetate buffer065M NaCl pH 52 the fractions were pooled and dialyzedagainst PBS pH 72 To verify the purity of isolated 1205732GP1a sodium dodecyl sulfate polyacrylamide gel electrophoresis(SDS-PAGE) has been performed
24 Purification of Anti-1205732GP1-IgG For the analysis of thesialylation of anti-1205732GP1-IgG we isolated IgG antibodiesfrom the sera to exclude possible interactions of the detectionlectin with anti-1205732GP1-IgM in the samples Purification ofanti-1205732GP1-IgG was performed as follows 20120583L of serumwas added to 50120583L magnetic Dynabeads Protein G (Novexby life technologies Carlsbad CA USA) and incubated for45 minutes at room temperature After washing with 1x TBSglycin-HCl pH 26 was added for 5 minutes to elute the anti-1205732GP1-IgG The supernatant now containing the IgG anti-bodies was neutralized with Tris-HCl pH 88 and all sam-ples were centrifuged using 100K Amicon Ultra-centrifugal
filter devices (Millipore Tullagreen Ireland) to transfer theminto 300 120583L TBS-buffer
25 Serum ELISAs Anti-1205732GP1-IgG -IgG1 -IgG2 -IgG3-IgG4 and -IgM and C3c and C1q Bound to Anti-1205732GP1To quantify the amount of anti-1205732GP1-IgG and its foursubclasses of C3c and C1q associated with anti-1205732GP1 andof anti-1205732GP1-IgM in the sera of patients with PAPS andSAPS NHD and healthy children we performed enzyme-linked immunosorbent assays (ELISA) In detail the surfaceof F 96NUNCmaxisorpmicrotiter plates (Thermo ScientificRoskilde Denmark) was coated with 50 120583L of a 2 120583gmLsolution of 1205732GP1 in coating buffer (100mM Na
2CO3
100mM NaHCO3 pH 96) at 4∘C overnight After washing
the plates three times with washing buffer (WB 005 Tween20 01mM CaCl
2 01mM MgCl
2in TBS buffer (150mM
NaCl 50mM TrisHCl pH = 74)) the wells of the coatedplates were blocked with 100 120583Lwell of a 2 BSA solutionfor two hours at 37∘C After three further washing steps withWB 50 120583Lwell of diluted serum (1 200 in WB) was addedand the plates were incubated at 37∘C for two further hoursThen the plates were washed three times with WB again
Development of the plates based on an enzymaticreaction of horseradish peroxidase (HRP) with TMB aschromogenic substrate Therefore the appropriate HRP-conjugated detection-antibody was added as a next step(Table 1) After incubation at 37∘C for another hour andthree washing steps with WB the plates were devel-oped by the addition of 50120583Lwell of freshly preparedTMB-solution (1mL of 001 g 331015840551015840-tetramethylbenzidine(Sigma-Aldrich Steinheim Germany) dissolved in 10mLdimethyl sulfoxide mixed with 9mL of 01M Na
2HPO4
005M citric acid pH 45ndash55 and 2120583L of H2O2) The
reaction was stopped by adding 50120583Lwell of 25 sulfuricacid (H
2SO4) Finally optical density (OD) was measured
at 450 nm and 650 nm as correction factor using an infiniteF200 pro ELISA-reader (Tecan)
After subtraction of the corresponding blank values of theplates experimental values were expressed as optical densityor as mean optical density of each serum when ELISA testswere performed in monoplicates or duplicates respectively
26 IgG-ELISA SNA In order to examine the sialylationof anti-1205732GP1-IgG we slightly modified the previouslydescribed ELISA protocol Before coating the 1205732GP1 antigenwas treated with 1 of periodic acid (PIA Sigma-Aldrich
4 Journal of Immunology Research
Table 2 Detection reagents for lectin ELISA
SNA-ELISAPrimary detectionreagent
Biotinylated SNA-lectin(Galab Technologies Geesthacht Germany)
Secondarydetection reagent
HRP-conjugated streptavidin(Jackson ImmunoResearch laboratoriesNewmarket Suffolk United Kingdom)
Dilution 1 10000SNA = Sambucus nigra agglutinin HRP = horseradish peroxidase
Steinheim Germany) for 48 hours at 43∘C to remove itssugar residues to exclude unspecific binding of Sambucusnigra lectin (SNA) to naive 1205732GP1 After that we centrifugedthe 1205732GP1-solution with 10K Amicon Ultra-centrifugal fil-ter devices (Millipore Tullagreen Ireland) to transfer thetreated 1205732GP1 into coating buffer (100mMNa
2CO3 100mM
NaHCO3 pH 96)
Then the plates were coated with 50 120583Lwell of a 2120583gmLsolution of the PIA-treated 1205732GP1 at 4∘C overnight andsubsequently blocked for two hours at 37∘C with 100 120583Lwellof PIA-treated (05) 3 gelatine (Sigma-Aldrich SteinheimGermany) in TBST buffer with 1mM CaCl
2and 1mM
MgCl2 The subsequent steps were performed as described
in the previous ELISA protocol with two exceptions Insteadof serum we added 50 120583Lwell of purified IgG-samples toexclude possible binding of SNA to anti-1205732GP1-IgM And thedevelopment of the plate required the addition of a seconddetection reagent (Table 2) which was incubated for onehour at room temperature and demanded three preliminarywashing steps We calculated the SNAanti-1205732GP1-IgG ratiofrom the OD values we obtained in the SNA ELISA and asimultaneously conducted anti-1205732GP1-IgG ELISA
27 Statistical Analysis Differences between the cohorts wereassessed using the one-way analysis of variance (ANOVA)test In the case of multiple testing the significance valueswere adjusted by the Newman-Keuls posttest correction
3 Results31 Analysis of Anti-1205732GP1-IgG Wemeasured the OD valuesof total anti-1205732GP1-IgG in the sera of 24 patients with PAPSand 22 patients with SLEAPS 16 healthy children and17NHD as controls As shown in Figure 1 the lowest IgGvalues were to be observed in the cohort of healthy donors(308mOD) whereas the values of the healthy children(970mOD) even exceeded that of the patients with APS(PAPS 686mOD 119901 = 0010 SLEAPS 546mOD 119901 lt0001) All differences between NHD and the other cohortsas well as between the children and the other cohorts weresignificant Patients with PAPS and SLEAPS did not differsignificantly
We calculated the 975 quantile of the anti-1205732GP1-IgGOD values obtained from the NHD cohort and set this valueas cut-off (738mOD) Sera exceeding this value were consid-ered seropositive for anti-1205732GP1-IgG Applying this cut-offvalue we obtained 31 seropositive samples (1224 PAPS 622SAPS 1316 healthy children) which were further analysed in
APS SLEAPS Healthy childrenNHDAPSSLEAPS ns
ns
NHD APS SLEAPS Children0
200
400
600
800
1000
1200
1400
1600
Ant
i-1205732
GP1
-IgG
(mO
D)
p values
mdashmdash
lt00001 lt001 lt00001
lt0001
lt00001
Figure 1 Total anti-1205732GP1 IgG in the sera of NHD PAPSSLEAPS and healthy children and the corresponding 119901 valuesNHD showed significantly lower mOD values than all other cohortswhereas healthy children represented the highest mOD valuesShort horizontal lines mean mOD values of all samples of thecohort Long broken line 975 quantile of NHD (=738mOD)NHD = normal healthy donor PAPS = primary antiphospholipidsyndrome SLEAPS = antiphospholipid syndrome with systemiclupus erythematosus OD = optical density
our study (Supplementary Figure 1 in Supplementary Mate-rial available online at httpdxdoiorg1011552015638129)
32 Analysis of the IgG-Subclasses of Anti-1205732GP1 Analysingthe absolute OD values of the anti-1205732GP1-IgG subclasseswe observed that the children showed significantly higherlevels of the anti-1205732GP1-IgG1 (69mOD) and -IgG3 subclasses(212mOD) compared to NHD PAPS and SAPS (Figures2(a) and 2(c) Table 3(a)) The differences between healthychildren and NHD regarding these two subclasses showeda higher level of significance than the comparison betweenhealthy children and patients with APS However healthychildren did not show significant differences for anti-1205732GP1-IgG2 and -IgG4 when compared to NHD PAPS and SAPS(Figures 2(b) and 2(d) Table 3(b))
Patients with APS (PAPS and SAPS) showed significantlyhigherODvalues for anti-1205732GP1-IgG1 -IgG2 and -IgG3 thanNHD (Figures 2(a) 2(b) and 2(c) Table 3(b)) As in theanti-1205732GP1-IgG ELISA we observed no significant differencebetween the patients with PAPS and SAPS
To summarize patients with APS showed higher ODvalues than NHD in IgG1 IgG2 and IgG3 IgG4 did notshow any significant differenceThe children in turn had thehighest OD values of the IgG1 and IgG3 subclasses
These results were consistent with the analysis of thenumber of seropositive samples as shown in Table 4 Todetermine seropositivity we calculated the mean OD and thestandard deviation (SD) of the NHD group for the differentIgG subclasses IgG values exceeding the NHDrsquos mean OD
Journal of Immunology Research 5
0
50
100
150
200
250
300
350
NHD APS SLEAPS Children
IgG
1 (m
OD
)
(a)
NHD APS SLEAPS Children0
50
100
150
200
250
300
350
IgG
2 (m
OD
)
(b)
NHD APS SLEAPS Children
940483 mOD
0
50
100
150
200
250
300
350
IgG
3 (m
OD
)
(c)
NHD APS SLEAPS Children
508 mOD 482 mOD 665 mOD 1753 mOD
0
50
100
150
200
250
300
350
IgG
4 (m
OD
)
(d)
0
50
100
150
200
250
300
350
(mO
D)
Children NHD APS
IgG4IgG3
IgG2IgG1
(e)
0
10
20
30
40
50
60
70
80
90
100
()
APSChildren
IgG4IgG3
IgG2IgG1
(f)
Figure 2mODvalues of anti-1205732GP1-IgG1 (a) anti-1205732GP1-IgG2 (b) anti-1205732GP1-IgG3 (c) and anti-1205732GP1-IgG4 (d) inNHD PAPS SLEAPSand childrenThe horizontal lines represent the cut-off for seropositivity of the corresponding anti-1205732GP IgG-subclass (gtmeanmOD
[NHD] +2 times SD
[NHD]) NHD = normal healthy donor PAPS = primary antiphospholipid syndrome SLEAPS = antiphospholipid syndrome withsystemic lupus erythematosus OD = optical density Median OD values of the anti-1205732GP1-IgG subclasses in the study cohorts (e) Regardingthe anti-1205732GP1-IgG subclass-distribution within the cohorts children showed a significantly higher contribution of IgG3 than IgG1 andIgG2 Patients with antiphospholipid syndrome (APS) showed a higher content of anti-1205732GP1-IgG2 than -IgG1 and -IgG3 NHD showed ahigher contribution of anti-1205732GP1-IgG2 than -IgG1 -IgG3 and -IgG4The columns represent the median OD values of IgG1 IgG2 IgG3 andIgG4 in the cohorts OD = optical density NHD = normal healthy donor Percentage distribution of anti-1205732GP1-IgG subclasses in childrenand patients with APS (f) Children harbour significantly less anti-1205732GP1-IgG2 than patients with APS but more anti-1205732GP1-IgG3 APS =antiphospholipid syndrome
6 Journal of Immunology Research
0
50
100
150
200
250
300
350
400
Children APS
C3c (
mO
D)
(a)
0
200
400
600
800
1000
1200
Children APS
IgM
(mO
D)
(b)
Figure 3 OD values of C3c (a) bound to anti-1205732GP1 and OD values of anti-1205732GP1-IgM (b) in sera of patients with APS and healthy childrenAnti-1205732GP1 of healthy children displayed significantly more bound C3c than that of patients with APS (119901 = 000007) This suggests a moreefficient clearance of 1205732GP1-immune complexes in the healthy children Anti-1205732GP1-IgM OD values of patients with APS were significantlyhigher than those of healthy children Horizontal lines represent the median OD value of the cohort APS = antiphospholipid syndrome OD= optical density
Table 3 (a) OD values of the anti-1205732GP1-IgG subclasses in theinvestigated cohorts (b) Comparison (119901 values) of the OD valuesof the anti-1205732GP1-IgG subclasses
(a)
IgG1 [mOD] IgG2 [mOD] IgG3 [mOD] IgG4 [mOD]NHD 24 64 27 67Children 69 77 212 210PAPS 30 85 45 72SAPS 28 74 55 173APS 29 81 48 106
(b)
IgG1 IgG2 IgG3 IgG4Children versus NHD lt00001 ns lt00001 nsChildren versus PAPS lt00001 ns lt0001 nsChildren versus SAPS lt0001 ns lt0001 nsAPS versus NHD ns ns ns nsPAPS versus SAPS ns ns ns nsOD = optical density 450 nm PAPS = primary antiphospholipid syndromeSAPS = secondary antiphospholipid syndrome APS = PAPS and SAPSNHD = normal healthy donors ns = not significant
plus two times the SD of the considered subclass were definedas seropositive (Supplementary Figure 1) Applying this cut-off we observed that the healthy children had high levels ofthe IgG1 and IgG3 subclasses with 13 (100) and 8 (62)seropositive samples respectively In contrast only 7 (39)and 0 (0) sera of patients with APS were seropositive forIgG1 and IgG3 respectively APS showed seropositivity ofanti-1205732GP1-IgG2 in 7 cases (39) whereas the cohort ofhealthy children contained just 2 seropositive sera (15) Just
few samples were seropositive for anti-1205732GP1-IgG4 in eachcohort representing a range from 8 (PAPS) to 17 (SAPS)
To determine the subclass distributionwithin the cohortswe calculated the contribution of themeanOD values of eachindividual subclass to the sum
As shown in Figures 2(e) and 2(f) we saw that for thehealthy children IgG3 was the most abundant subclass witha contribution of 40 (IgG3 gt IgG1 (119901 lt 0001) and IgG3 gtIgG2 (119901 lt 001)) In contrast IgG2 dominated the subclassprofile for NHD (46 IgG2 gt IgG1 IgG2 gt IgG3 and IgG2 gtIgG4 (119901 lt 00001)) and patients with APS (38 IgG2 gt IgG1(119901 lt 00001) IgG2 gt IgG3 (119901 = 0001)) The contributionof IgG1 ranged from 12 (SAPS) to 17 (NHD) and thatof IgG4 from 21 (PAPS) to 35 (SAPS) Comparing thecontribution of the individual subclasses between the studycohorts we observed that children showed significantly lessIgG2 than patients with APS and NHD (119901 lt 0001 119901 lt00001) and a higher IgG3 content than NHD and patientswith APS (119901 lt 00001 119901 lt 0001) No significant differencebetween the cohorts was to be seen for the contributions ofIgG1 and IgG4
33 Analysis of IgM C1q and C3c Associated to Anti-1205732GP1-IgG Next we analysed potential 1205732GP1-binding proteins inthe sera of healthy children and adults with APS The signalsfor C1q were low and did not differ significantly between thecohorts
However the ELISAs detecting C3c bound to anti-1205732GP1and anti-1205732GP1-IgM showed highly significant differencesbetween healthy children and adults with APSWhereas C3c-anti-1205732GP1 adducts were increased in healthy children (119901 lt00001 Figure 3(a)) anti-1205732GP1-IgM was decreased (119901 lt0001 Figure 3(b))
Journal of Immunology Research 7
Table 4 Number (frequency) of seropositive samples of the anti-1205732GP1-IgG subclasses
Number of seropositive samples (gtmOD[NHD] + 2 times SD[NHD])IgG1
(gt29mOD)IgG2
(gt79mOD)IgG3
(gt85mOD)IgG4
(gt319mOD)NHD (119899 = 17) 0 (0) 0 (0) 1 (6) 1 (6)Children (119899 = 13) 13 (100) 2 (15) 8 (62) 2 (15)PAPS (119899 = 12) 6 (50) 5 (42) 0 (0) 1 (8)SAPS (119899 = 6) 1 (17) 2 (33) 0 (0) 1 (17)APS (119899 = 18) 7 (39) 7 (39) 0 (0) 2 (11)OD = optical density 450 nm PAPS = primary antiphospholipid syndrome SAPS = secondary antiphospholipid syndrome APS = PAPS and SAPS NHD =normal healthy donorsThe cut-off for seropositivity was calculated from the mean OD of the corresponding anti-1205732GP1-IgG subclass of the NHD cohort andits standard deviation (SD) according to the formula mean OD[NHD] + 2 times SD[NHD] The respective cut-off OD values are indicated in parentheses below theanti-1205732GP1-IgG subclasses
Table 5 Cohorts studied with the lectin ELISA
Cohort Abbreviation Number of samplesPatients with primary APS PAPS 12Patients with APS and SLE as underlying disease SAPS 10Patients with SLE without symptoms of APS harbouring circulating aPL SLE + aPL 15Asymptomatic carriers of aPL aaPL 6Healthy children with circulating anti-1205732GP1 mdash 16APS = antiphospholipid syndrome aPL = antiphospholipid antibodies SLE = systemic lupus erythematosus
34 Analysis of the Sialylation of Anti-1205732GP1 We analysedthe glycosylation of anti-1205732GP1-IgG focusing on its sialy-lation since highly sialylated Fc-parts of IgG are known tocause rather anti-inflammatory responses [30] Wemeasuredthe binding of the sialic acid specific lectin SNA to the anti-1205732GP1-IgG As shown in Table 5 we employed five cohorts ofindividuals 12 patientswith PAPS 10 patientswith SAPS (SLEas primary disease) 15 patients with SLE who did not showclinical sign of APS but with aPL in their sera (SLE + aPL)6 asymptomatic adult carriers of aPL (aaPL) and 16 healthychildren harbouring aPL
Analysing the SNAIgG-ratios (Figure 4) we identifiedthe highest sialylation level in the sera of the healthy childrenThe significances were rather high and decreased from PAPS(119901 lt 00001) to SAPS (119901 lt 00001) to SLE + aPL (119901 lt00001) and to aaPL (119901 lt 0001)
4 Discussion
We analysed two distinct sets of sera One was used toinvestigate anti-1205732GP1-IgG its subclasses and its associatedproteins C1q and C3c For the analysis of the sialylation ofanti-1205732GP1-IgG we had to collect fresh sera from patientswith APS and the healthy children since the stability of theantibody-glycosylation is unknown In order to avoid itspossible decay by processes like enzymatic degradation thisperiod has been kept as short as possible and was followed byimmediate investigation of the glycosylation
The antiphospholipid syndrome is characterized by clin-ical manifestations such as arterial and venous thrombosisor pregnancy-related complications and the detection of spe-cific autoantibodies (lupus anticoagulant anti-1205732GP1- and
anticardiolipin-autoantibodies) [1] In particular anti-1205732GP1sare considered responsible for provoking the pathologicalcomplications [3]We firstmeasured the total amount of anti-1205732GP1-IgG in the sera of 16 healthy children without clinicalmanifestations of APS and 46 adult patients with APS toidentify individual cohorts seropositive for anti-1205732GP1 17adult normal healthy donors served as negative control
The samples of the patientswithAPS showedhigher levelsof anti-1205732GP1-IgG than those of NHD We also observeda higher concentration of anti-1205732GP1-IgG in the sera ofhealthy children compared to NHD This is due to the factthat the detection of these antibodies has been the inclusioncriterion for the healthy children examined in this studyInterestingly the children showed even higher OD values forthe putatively pathogenic anti-1205732GP1-IgG than the patientswith APS This finding was surprising since the childrenwere clinically healthy and did not show any signs of APSsuch as thrombosis or thrombocytopenia complications inwhich the existence of aPL especially anti-1205732GP1 is maderesponsible for [7 33]
High levels of circulating aPL in 61 healthy children whencompared to healthy adults have already been described byAvcin and colleagues [18] The anti-1205732GP1-IgG was report-edly higher in preschool children with a mean age of 5years than in adolescents with a mean age of 135 years[18] The strongly elevated circulating anti-1205732GP1-IgG in ourcohort of healthy children may be explained by the fact thatthey had an even lower age (mean age was nine months)One possible explanation for this result is the inductionof anti-1205732GP1 by viral and bacterial infections [10 11] ornutritional exposure to 1205732GP1 [17] As the immune systemin children is still to evolve they are more susceptible to
8 Journal of Immunology Research
0
05
10
15
20
25SN
AIg
G
ChildrenPAPS SAPS aaPLSLE + aPL
lowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowast
Figure 4 SNAanti-1205732GP1-IgG ratio in PAPS SAPS SLE + aPLaaPL and children Healthy children show a significantly higherSNAanti-1205732GP1-IgG ratio than PAPS SAPS SLE + aPL and aaPL(lowastlowast119901 lt 00001 lowastlowastlowast119901 lt 000001) This suggests higher sialylation ofthe oligosaccharides attached to Fc fragments of the childrenrsquos anti-1205732GP1No significant differenceswere foundbetween the remainingcohorts Horizontal lines indicate the mean SNAIgG ratio of thecohort SNA = Sambucus nigra agglutinin APS = antiphospholipidsyndrome PAPS = patients with primary APS SAPS = patients withAPS and SLE as underlying disease SLE + aaPL = patients withSLE without symptoms of APS harbouring circulating aPL aaPL =asymptomatic carriers of aPL aPL = antiphospholipid antibodiesSLE = systemic lupus erythematosus
infections than adults and their immune system has to copewith a plethora of viral and bacterial pathogens for the firsttime in their life This scenario is prone to result in thestimulation of autoantibody-production Molecular mimicryof certain pathogens can lead to the production of antibodieswhich then cross-react with 1205732GP1 [7 12ndash15] When infantsdiversify their dietary habits and start to consume productslike milk and meat they can get into contact with nutritional1205732GP1 [19] This is a further condition capable triggeringthe production of anti-1205732GP1 Exposure to pathogen-derivedand dietary 1205732GP1 could explain the high level of circulatinganti-1205732GP1-IgG in healthy childrenThis low affine reactivityis consecutively lost during the maturation of the immunesystem and replaced by the emergence in adults of high affineantibodies that more specifically recognize pathogens Apartfrom possible origins of these autoantibodies it is obviousthat they do not provoke any of the hallmark symptoms ofAPS In contrast to their adult counterparts anti-1205732GP1-IgGscirculating in the plasma of healthy children are obviouslynot pathogenic So it seems decent to hypothesize thatanti-1205732GP1 autoantibodies in healthy children biochemicallydiffer from those in adults
To test this hypothesis we examined the anti-1205732GP1-IgG subclass distribution in the seropositive sera of the anti-1205732GP1-IgG ELISA The healthy children showed higher ODvalues for the IgG1- and IgG3-subclasses than patients withAPS and NHD This was consistent with the number ofseropositive samples In the anti-1205732GP1-IgG1-subclass every
tested serum of the healthy children surpassed the thresholdset for seropositivity (gtmean OD
[NHD] + 2 times SD[NHD]) and
8 samples (62) did so in the IgG3-subclass In the seraof patients with APS just 7 (39) and no samples wereseropositive for IgG1 or IgG3 In the sera of patients with APScompared to NHD we observed higher OD levels of anti-1205732GP1 in all subclasses except for IgG4This result is consis-tent with the findings of Guerin et al who reported elevatedlevels in all anti-1205732GP1-IgG subclasses even including IgG4in patients with APS compared to a control group of healthyadults [23]
It might be that the elevated absolute OD values of theanti-1205732GP1-IgG subclasses are the logical consequence ofthe generally higher level of anti-1205732GP1-IgG in the patientswith APS and healthy children who were selected for furthertesting Therefore we also analysed the subclass distributionof this antibody We first analysed the anti-1205732GP1-IgG sub-class distribution within the individual cohorts and observedthat in the healthy children the IgG3 antibody was mostabundant with a contribution of 40 to the total reactivityThis is consistent with the finding that all sera of the healthychildren were seropositive in this subclass and that theiranti-1205732GP1-IgG3 OD values were the highest of all cohortsThis indicates a predominance of this subclass in the healthychildren The remaining anti-1205732GP1-IgG subclasses in thesera of the children ranged from 15 (IgG1) over 20 (IgG2)to 25 (IgG4) but the differences between their proportionswere not statistically significant In contrast in the sera ofthe patients with APS the anti-1205732GP1-IgG2 subclass had thehighest contribution with 38
The finding of an elevated proportion of the IgG2 subclassin the patients with APS confirms former published datawhich also describe a skewing towards anti-1205732GP1-IgG2 inAPS [19 23 26 27] The authors detected an even highercontribution of anti-1205732GP1-IgG2 (702 [26] and 87 [27]resp) In contrast to our results the IgG1 subclass wasmore prominent in the published studies described aboveImportantly in the cohort of the healthy infants IgG2 wasrelatively lowTherefore onemay argue that anti-1205732GP1-IgG2is not just a marker of the disease but is also involved inthe pathogenic action of the aPL autoantibodies When wecompared the anti-1205732GP1-IgG subclass distribution betweenthe three cohorts we noticed a significant difference in thedistribution concerning the children on one hand and NHDand patients with APS on the other Healthy children showeda higher proportion of anti-1205732GP1-IgG3 and a lower one ofanti-1205732GP1-IgG2 In NHD and patients with APS it was theother way round IgG1 and IgG4 did not significantly differbetween healthy children and patients with APS orNHDThepredominance of anti-1205732GP1-IgG3 in the healthy children isa hint to an immune response elicited by a proteinaceousantigen [26 27 34] This supports the hypothesis thatthe harmless anti-1205732GP1 in children are infection-inducedwith pathogen-derived proteins serving as trigger for theirproduction However it is still elusive if the IgG3-dominatedanti-1205732GP1 response found in the sera of healthy childrencontributes to its low pathogenic potential Alternativelyit can just be ascribed to the proceeding evolution of theinfantrsquos immune system typically associated with an elevated
Journal of Immunology Research 9
proportion of IgG3 antibodies [34 35] During childhoodthe proportion of IgG3 continuously decreases to adult rangesusually reached at the age of 10ndash12 years However the aboveaverage elevated proportion of the anti-1205732GP1-IgG3 (40) inthe sera of the healthy childrenmay thus not just be due to theinfant immune system In particular when bearing in mindthat normal adult levels of the IgG3 subclass are just around6 [34 36]
IgG3 is the IgG-subclass exerting the most effectiveactivation of the complement cascade through the classicalpathway [34] Since complement activation is associated withthe pathogenesis of APS [21 32 37] the high proportionof complement-activating anti-1205732GP1-IgG3 in the childrenseems to contradict a possible protective effect against APS ofthese kinds of antibodies However the role of complement inthe etiopathogenesis of autoimmune diseases is manifold asthe complement components C1q C2 and C4 contribute tothe clearance of postapoptotic secondary necrotic cells andtheir deficiency is therefore a risk factor for autoimmunityand autoinflammation [38] In a similar manner studiesin mouse in vivo models of various autoimmune diseasesdemonstrated that IgG mediated tissue inflammation wasblocked in mice deficient in activating Fc120574Rs although thecomplement component C3 was still abundantly depositedin the tissue [39ndash41] Thus the contribution of complementdeposits in tissue with respect to tissue inflammation remainsto be established
To obtain new insights into the involvement of thecomplement system in the pathogenesis of APS we analysedthe C1q- and C3c-binding to anti-1205732GP1 We observed thatanti-1205732GP1 in the sera of the healthy children and in patientswith APS similarly bound C1q the first molecule of theclassical pathway of complement activation [42] Performinganti-1205732GP1-IgM-ELISAs we detected significantly higherIgM values in the patients with APS One would expecta higher C1q-binding of the IgM positive sera since thisimmunoglobulin binds and activates C1q more strongly thanIgG However the children mainly harbour IgG3 autoanti-bodies the most potent subclass for the activation of theclassical complement pathway [34]This may compensate forthe lower IgM level in their sera and thus be responsiblefor the similar C1q-binding of both cohorts This suggeststhat preconditions for complement activation via the classicalpathway are comparable in both cohorts We conclude thatclassical complement activation has a minor impact on thedifferential pathogenesis of aPL in healthy children andpatients with APS
What then is the culprit that induces their pathogenicpotential
Nextwe quantified the contribution ofC3c to the immunecomplexes bound to 1205732GP1 C3c is a domain in the C3bmolecule and one of its cleavage products created aftercleavage by factor I of C3b [42 43] C3c not just representsa marker of complement activation but also serves as anopsonin improving the clearance of bound targets Weobserved significantly more C3c bound to anti-1205732GP1 inthe sera of the healthy children than in those of patientswith APS This reflects a better opsonization by C3c ofthe potentially harmful anti-1205732GP1 immune complexes
anti-1205732GP1-opsonized cells or cellular fragments in thesera of the healthy children Consequently the anti-1205732GP1complexes can be cleared more efficiently by the powerfulcomplement system of the healthy children In patients withchronic inflammatory autoimmune diseases complement isusually low which may result from null alleles [44] or fromcomplement consumption [38]
But why do these processes not cause inflammation andaberrant coagulation in the childrenApart fromanti-1205732GP1-IgG subclass distribution and distinct ability to activate thecomplement system it is likely that other properties of theanti-1205732GP1 in the sera of children contribute to their lowpathogenicity We hypothesized that an increased sialylationof the anti-1205732GP1-IgG in healthy individuals renders themharmless for their hosts IgG is usually N-glycosylated at theCH2-domain of its Fc-part This N-glycan crucially modifiesthe effector functions the IgG molecule is endowed with[45ndash47] This biantennary core-fucosylated oligosaccharidecarries a variable amount of sugar monomers attached toboth arms [28 48] IgG antibodies terminating with sialicacids on one or both antennae have been shown to behaveanti-inflammatorily after infusion of IVIG [29 31]The signalexerted by the terminal sialic acid is dominant since thefraction of sialylated IgG in healthy plasma is just 4 to 14[28 46 49]
In mice these effects are mainly ascribed to decreasedbinding of sialylated IgG to activating Fc120574-receptors andan upregulation of the inhibitory Fc120574RIIB [29 31] Weobserved that anti-1205732GP1-IgG isolated from sera of healthychildren was significantly more sialylated than that fromsymptomatic patients with APS One may predict that theseautoantibodies in complex with 1205732GP1 show low affinity foractivating Fc120574Rs and therefore do not provoke proinflam-matory and procoagulant effector functions Fc120574RIIA is anexample for an activating receptor It is located on a plethoraof cells On the surfaces of platelets it mediates activationby immune complexes The activation is accompanied byplatelet aggregation and the release of mediators such asthe prothromboticThromboxaneA2 [50] Consecutively thiscascade can result in thrombus formation [51 52] Sialylatedanti-1205732GP1-IgG is unable to bind to and to activate plateletsvia Fc120574RIIA
Further mechanisms involved in thrombus formation inAPS comprise the interaction of anti-1205732GP1 with variouspattern recognition receptors [53] and its interference withregulatory functions ofmolecules involved in the coagulationprocess (protein C prothrombin or tissue factor [52 54])Direct binding to 1205732GP1 on endothelial cells (EC) shiftstheir phenotype towards coagulation and inflammation [54]Considering that Fc120574R-binding is involved in many of thesemechanisms it seems likely that a high sialylation of anti-1205732GP1-IgG leads to a low pathogenicity of these autoan-tibodies and does not cause APS Therefore we concludethat the higher sialylation of anti-1205732GP1-IgG in the seraof the healthy children plays a major role in protectingthem from typical complications of APS This hypothesis isfurther supported by the observation that antiphospholipidantibodies from sera of asymptomatic adult carriers aremore sialylated than those from patients with APS A high
10 Journal of Immunology Research
sialylation status of antiphospholipid antibodies seems toprevent the development of symptoms of APS also in theseindividuals
We conclude that a plethora of factors contributes tothe low pathogenicity of anti-1205732GP1-IgG in the sera ofhealthy children and asymptomatic adults We observed adistinct spectrum of IgG-subclasses compared to patientswith APS and a better binding of C3c indicating a moreefficient clearance of potentially harmful anti-1205732GP1 Thehypersialylation of the anti-1205732GP1-IgG is prone to amelioratetheir inflammatory and procoagulatory effects in healthycarriers whereas hyposialylation and inflammationmutuallyprecipitate the antiphospholipid syndrome
Disclosure
Thepresent workwas performed in fulfillment of the require-ments for obtaining the degree ldquoDr medrdquo for ChristophFickentscher
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Laura Andreoli andMartin Herrmann contributed equally tosenior authorship
Acknowledgments
This project was supported by the German Research Foun-dation (training research programme of the collaborativeresearch center 643) by the Emerging Fields Initiative (EFI)of the FAU Erlangen-Nuremberg and by the K und RWucherpfennig-Stiftung
References
[1] S Miyakis M D Lockshin T Atsumi et al ldquoInternationalconsensus statement on an update of the classification criteriafor definite antiphospholipid syndrome (APS)rdquo Journal ofThrombosis and Haemostasis vol 4 no 2 pp 295ndash306 2006
[2] W A Wilson A E Gharavi T Koike et al ldquoInternationalconsensus statement on preliminary classification criteria fordefinite antiphospholipid syndrome report of an internationalworkshoprdquo Arthritis and Rheumatism vol 42 no 7 pp 1309ndash1311 1999
[3] P G de Groot and R T Urbanus ldquoThe significance of autoan-tibodies against beta2-glycoprotein Irdquo Blood vol 120 no 2 pp266ndash274 2012
[4] C Nalli S Piantoni L Andreoli M Motta and A TincanildquoAntiphospholipid syndrome and antiphospholipid antibodiesin children the two sides of the coinrdquo Israel Medical AssociationJournal vol 14 no 5 pp 310ndash312 2012
[5] I Schousboe ldquo1205732-glycoprotein I a plasma inhibitor of thecontact activation of the intrinsic blood coagulation pathwayrdquoBlood vol 66 no 5 pp 1086ndash1091 1985
[6] J George M Blank Y Levy et al ldquoDifferential effects of anti-1205732-glycoprotein I antibodies on endothelial cells and on themanifestations of experimental antiphospholipid syndromerdquoCirculation vol 97 no 9 pp 900ndash906 1998
[7] R Willis and S S Pierangeli ldquoAnti-1205732-glycoprotein I antibod-iesrdquo Annals of the New York Academy of Sciences vol 1285 no1 pp 44ndash58 2013
[8] M J Fischer J Rauch and J S Levine ldquoThe antiphospholipidsyndromerdquo Seminars in Nephrology vol 27 no 1 pp 35ndash462007
[9] H Amital M Govoni R Maya et al ldquoRole of infectious agentsin systemic rheumatic diseasesrdquo Clinical and ExperimentalRheumatology vol 26 no 1 supplement 48 pp S27ndashS32 2008
[10] T Avcin ldquoAntiphospholipid syndrome in childrenrdquo CurrentOpinion in Rheumatology vol 20 no 5 pp 595ndash600 2008
[11] M Garcıa-Carrasco C Galarza-Maldonado C Mendoza-Pinto R O Escarcega and R Cervera ldquoInfections and theantiphospholipid syndromerdquo Clinical Reviews in Allergy ampImmunology vol 36 no 2-3 pp 104ndash108 2009
[12] P Cruz-Tapias M Blank J-M Anaya and Y ShoenfeldldquoInfections and vaccines in the etiology of antiphospholipidsyndromerdquo Current Opinion in Rheumatology vol 24 no 4 pp389ndash393 2012
[13] P von Landenberg Y Doring S Modrow and K J LacknerldquoAre antiphospholipid antibodies an essential requirement foran effective immune response to infectionsrdquoAnnals of the NewYork Academy of Sciences vol 1108 pp 578ndash583 2007
[14] M Blank I Krause M Fridkin et al ldquoBacterial induction ofautoantibodies to 1205732-glycoprotein-I accounts for the infectiousetiology of antiphospholipid syndromerdquo Journal of ClinicalInvestigation vol 109 no 6 pp 797ndash804 2002
[15] Y Shoenfeld M Blank R Cervera J Font E Raschi and P-LMeroni ldquoInfectious origin of the antiphospholipid syndromerdquoAnnals of the Rheumatic Diseases vol 65 no 1 pp 2ndash6 2006
[16] A E Gharavi S S Pierangeli R G Espinola X Liu MColden-Stanfield and E N Harris ldquoAntiphospholipid antibod-ies induced in mice by immunization with a cytomegalovirus-derived peptide cause thrombosis and activation of endothelialcells in vivordquo Arthritis and Rheumatism vol 46 no 2 pp 545ndash552 2002
[17] M Motta G Chirico C B Rebaioli et al ldquoAnticardiolipin andanti-beta2 glycoprotein I antibodies in infants born to motherswith antiphospholipid antibody-positive autoimmune diseasea follow-up studyrdquoAmerican Journal of Perinatology vol 23 no4 pp 247ndash251 2006
[18] T Avcin A Ambrozic M Kuhar T Kveder and B RozmanldquoAnticardiolipin and anti-1205732 glycoprotein I antibodies in seraof 61 apparently healthy children at regular preventive visitsrdquoRheumatology vol 40 no 5 pp 565ndash573 2001
[19] A Ambrozic ldquoAnti-beta2-glycoprotein I antibodies in children
with atopic dermatitisrdquo International Immunology vol 14 no 7pp 823ndash830
[20] J A Giron-Gonzalez E Garcıa Del Rıo C Rodrıguez JRodrıguez-Martorell and A Serrano ldquoAntiphospholipid syn-drome and asymptomatic carriers of antiphospholipid anti-body prospective analysis of 404 individualsrdquo Journal ofRheumatology vol 31 no 8 pp 1560ndash1567 2004
[21] K A Breen P Seed K Parmar GWMoore S E Stuart-Smithand B J Hunt ldquoComplement activation in patients with isolatedantiphospholipid antibodies or primary antiphospholipid syn-dromerdquo Thrombosis and Haemostasis vol 107 no 3 pp 423ndash429 2012
Journal of Immunology Research 11
[22] F Fischetti P Durigutto V Pellis et al ldquoThrombus formationinduced by antibodies to 1205732-glycoprotein I is complementdependent and requires a priming factorrdquo Blood vol 106 no7 pp 2340ndash2346 2005
[23] J Guerin E Casey C Feighery and J Jackson ldquoAnti-Beta 2-glycoprotein I antibody isotype and IgG subclass in antiphos-pholipid syndrome patientsrdquo Autoimmunity vol 31 no 2 pp109ndash116 1999
[24] L Andreoli C Nalli M Motta et al ldquoAnti-1205732-glycoprotein I
IgG antibodies from 1-year-old healthy children born to moth-ers with systemic autoimmune diseases preferentially targetdomain 45 might it be the reason for their lsquoinnocentrsquo profilerdquoAnnals of the Rheumatic Diseases vol 70 no 2 pp 380ndash3832011
[25] B de Laat R H W M Derksen M van Lummel M T T Pen-nings and P G deGroot ldquoPathogenic anti-beta2-glycoprotein Iantibodies recognize domain I of beta2-glycoprotein I only aftera conformational changerdquo Blood vol 107 no 5 pp 1916ndash19242006
[26] M Samarkos K A Davies C Gordon M J Walport and SLoizou ldquoIgG subclass distribution of antibodies against 1205732-GP1and cardiolipin in patients with systemic lupus erythematosusand primary antiphospholipid syndrome and their clinicalassociationsrdquo Rheumatology vol 40 no 9 pp 1026ndash1032 2001
[27] J Arvieux B Roussel D Ponard and M G Colomb ldquoIgG2subclass restriction of anti-1205732 glycoprotein 1 antibodies inautoimmune patientsrdquo Clinical and Experimental Immunologyvol 95 no 2 pp 310ndash315 1994
[28] J N Arnold M R Wormald R B Sim P M Rudd and R ADwek ldquoThe impact of glycosylation on the biological functionand structure of human immunoglobulinsrdquo Annual Review ofImmunology vol 25 pp 21ndash50 2007
[29] Y Kaneko F Nimmerjahn and J V Ravetch ldquoAnti-inflamma-tory activity of immunoglobulin G resulting from Fc sialyla-tionrdquo Science vol 313 no 5787 pp 670ndash673 2006
[30] RM Anthony and F Nimmerjahn ldquoThe role of differential IgGglycosylation in the interaction of antibodies with FcgammaRsin vivordquo Current Opinion in Organ Transplantation vol 16 no1 pp 7ndash14 2011
[31] R M Anthony and J V Ravetch ldquoA novel role for the IgGFc glycan the anti-inflammatory activity of sialylated IgG FcsrdquoJournal of Clinical Immunology vol 30 no 1 pp S9ndashS14 2010
[32] K Oku T Atsumi M Bohgaki et al ldquoComplement activationin patients with primary antiphospholipid syndromerdquoAnnals ofthe Rheumatic Diseases vol 68 no 6 pp 1030ndash1035 2009
[33] H P McNeil R J Simpson C N Chesterman and S AKrilis ldquoAnti-phospholipid antibodies are directed against acomplex antigen that includes a lipid-binding inhibitor of coag-ulation 1205732-Glycoprotein I (apolipoprotein H)rdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 87 no 11 pp 4120ndash4124 1990
[34] R Jefferis and D S Kumararatne ldquoSelective IgG subclassdeficiency quantification and clinical relevancerdquo Clinical andExperimental Immunology vol 81 no 3 pp 357ndash367 1990
[35] P H Schur F Rosen and M E Norman ldquoImmunoglobulinsubclasses in normal childrenrdquo Pediatric Research vol 13 no3 pp 181ndash183 1979
[36] M A H French and G Harrison ldquoSerum IgG subclass concen-trations in healthy adults a study using monoclonal antiserardquoClinical and Experimental Immunology vol 56 no 2 pp 473ndash475 1984
[37] M Odorczuk L B Keil and V A DeBari ldquoBinding of com-plement component C1q to anti-beta2 glycoprotein I antibodiesfrom patients with antiphospholipid syndromerdquo InflammationResearch vol 48 no 10 pp 524ndash526 1999
[38] L Truedsson A A Bengtsson and G Sturfelt ldquoComplementdeficiencies and systemic lupus erythematosusrdquoAutoimmunityvol 40 no 8 pp 560ndash566 2007
[39] S Aschermann C H K Lehmann S Mihai G Schett DDudziak and F Nimmerjahn ldquoB cells are critical for autoim-mune pathology in Scurfy micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 110 no47 pp 19042ndash19047 2013
[40] Y Kaneko F Nimmerjahn M P Madaio and J V RavetchldquoPathology and protection in nephrotoxic nephritis is deter-mined by selective engagement of specific Fc receptorsrdquo TheJournal of Experimental Medicine vol 203 no 3 pp 789ndash7972006
[41] D Sylvestre R Clynes M Ma H Warren M C Carrolland J V Ravetch ldquoImmunoglobulin G-mediated inflammatoryresponses develop normally in complement-deficient micerdquoJournal of ExperimentalMedicine vol 184 no 6 pp 2385ndash23921996
[42] G E Ritchie B E Moffatt R B Sim B P Morgan R A Dwekand P M Rudd ldquoGlycosylation and the complement systemrdquoChemical Reviews vol 102 no 2 pp 305ndash320 2002
[43] Y Palarasah K Skjodt J Brandt et al ldquoGeneration of aC3c specific monoclonal antibody and assessment of C3c as aputative inflammatorymarker derived from complement factorC3rdquo Journal of ImmunologicalMethods vol 362 no 1-2 pp 142ndash150 2010
[44] D G Doherty J A Underhill P T Donaldson et al ldquoPoly-morphism in the human complement C4 genes and geneticsusceptibility to autoimmune hepatitisrdquo Autoimmunity vol 18no 4 pp 243ndash249 1994
[45] M-H Tao and S L Morrison ldquoStudies of aglycosylatedchimeric mouse-human IgG Role of carbohydrate in thestructure and effector functions mediated by the human IgGconstant regionrdquoThe Journal of Immunology vol 143 no 8 pp2595ndash2601 1989
[46] R Jefferis ldquoGlycosylation of recombinant antibody therapeu-ticsrdquo Biotechnology Progress vol 21 no 1 pp 11ndash16 2005
[47] Y Mimura S Church R Ghirlando et al ldquoThe influence ofglycosylation on the thermal stability and effector functionexpression of human IgG1-Fc properties of a series of truncatedglycoformsrdquoMolecular Immunology vol 37 no 12-13 pp 697ndash706 2001
[48] C Huhn M H J Selman L R Ruhaak A M Deelder and MWuhrer ldquoIgG glycosylation analysisrdquo Proteomics vol 9 no 4pp 882ndash913 2009
[49] J Lund N Takahashi J D Pound M Goodall and R JefferisldquoMultiple interactions of IgG with its core oligosaccharide canmodulate recognition by complement and human Fc gammareceptor I and influence the synthesis of its oligosaccharidechainsrdquo The Journal of Immunology vol 157 no 11 pp 4963ndash4969 1996
[50] R Forastiero M Martinuzzo L O Carreras and J MacloufldquoAnti-1205732 glycoprotein I antibodies and platelet activation inpatients with antiphospholipid antibodies association withincreased excretion of platelet-derived thromboxane urinarymetabolitesrdquoThrombosis and Haemostasis vol 79 no 1 pp 42ndash45 1998
12 Journal of Immunology Research
[51] J-C Reverter and D Tassies ldquoMechanism of thrombosis inantiphospholipid syndrome binding to plateletsrdquo in HughesSyndrome M A Khamashta Ed pp 403ndash414 Springer Lon-don UK 2006
[52] G Espinosa R Cervera J Font and Y Shoenfeld ldquoAntiphos-pholipid syndrome pathogenic mechanismsrdquo AutoimmunityReviews vol 2 no 2 pp 86ndash93 2003
[53] T Shi B Giannakopoulos X Yan et al ldquoAnti-1205732-glycoprotein
I antibodies in complex with 1205732-glycoprotein I can activate
platelets in a dysregulated manner via glycoprotein Ib-IX-VrdquoArthritis amp Rheumatism vol 54 no 8 pp 2558ndash2567 2006
[54] P LMeroni E RaschiMCamera et al ldquoEndothelial activationby aPL a potential pathogenetic mechanism for the clinicalmanifestations of the syndromerdquo Journal of Autoimmunity vol15 no 2 pp 237ndash240 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Immunology Research 3
Table 1 Antibodies employed for each ELISA
ELISA Detection-antibody DilutionAnti-1205732GP1-IgG HRP-conjugated rabbit F(ab1015840) 2 anti-human-IgGlowast 1 20000
Anti-1205732GP1-IgG subclasses HRP-conjugated mouse anti-human IgG1 2 3 and 4 (Clones HP6001HP6002 HP6050 amp HP6025)lowast 1 20000
Anti-1205732GP1-IgM HRP-conjugated goat anti-human-IgMlowast 1 20000Anti-1205732GP1-bound C3c HRP-conjugated sheep anti-C3clowastlowast 1 500Anti-1205732GP1-bound C1q HRP-conjugated sheep anti-C1qlowastlowast 1 500lowast purchased from Southern Biotech Birmingham Alabama USAlowastlowast purchased from Abcam Cambridge United KingdomHRP = horseradish peroxidase
their circulation (mean age 95 months median age 10months range 3 months and 5 daysndash19 months and 10 days)
22 Blood Drawing Heparinised venous blood was drawnby venipuncture and processed within two hours Serum wasobtained by centrifugation at 1000 g for 15 minutes of clottedblood then aliquoted frozen and stored at minus20∘C until usefor investigation
Informed consent was obtained from all blood donorsand the childrenrsquos parents respectively and the study wasapproved by the ethics committees of the University HospitalErlangen and the Spedali Civili Brescia
23 Isolation of 1205732GP1 For 1205732GP1-isolation from dilutedsera of normal healthy donors (in 015MNaCl) percloric acidwas slowly added until the mixture got a creamy aspect Aftercentrifugation at 10000 g the supernatant was neutralizedwith NaOH and dialyzed against a buffer of 002M Tris and003MNaCl pH8Then itwas given into columns containingHiTrap heparin and contaminants of the supernatant wereeluted by the addition of 002M Tris and 015M NaCl pH 8In order to elute semipurified 1205732GP1 a buffer of 002M Trisand 035MNaCl pH 8 was injected into the columns and thefractions have been pooled and dialyzed against a solutioncontaining 005M acetate buffer 005M NaCl pH 48 Thismaterial was filtrated with 02120583membranes and injected intoresin columns equilibrated with 005M acetate buffer 005MNaCl pH 48 After washing with 005M acetate buffer065M NaCl pH 52 the fractions were pooled and dialyzedagainst PBS pH 72 To verify the purity of isolated 1205732GP1a sodium dodecyl sulfate polyacrylamide gel electrophoresis(SDS-PAGE) has been performed
24 Purification of Anti-1205732GP1-IgG For the analysis of thesialylation of anti-1205732GP1-IgG we isolated IgG antibodiesfrom the sera to exclude possible interactions of the detectionlectin with anti-1205732GP1-IgM in the samples Purification ofanti-1205732GP1-IgG was performed as follows 20120583L of serumwas added to 50120583L magnetic Dynabeads Protein G (Novexby life technologies Carlsbad CA USA) and incubated for45 minutes at room temperature After washing with 1x TBSglycin-HCl pH 26 was added for 5 minutes to elute the anti-1205732GP1-IgG The supernatant now containing the IgG anti-bodies was neutralized with Tris-HCl pH 88 and all sam-ples were centrifuged using 100K Amicon Ultra-centrifugal
filter devices (Millipore Tullagreen Ireland) to transfer theminto 300 120583L TBS-buffer
25 Serum ELISAs Anti-1205732GP1-IgG -IgG1 -IgG2 -IgG3-IgG4 and -IgM and C3c and C1q Bound to Anti-1205732GP1To quantify the amount of anti-1205732GP1-IgG and its foursubclasses of C3c and C1q associated with anti-1205732GP1 andof anti-1205732GP1-IgM in the sera of patients with PAPS andSAPS NHD and healthy children we performed enzyme-linked immunosorbent assays (ELISA) In detail the surfaceof F 96NUNCmaxisorpmicrotiter plates (Thermo ScientificRoskilde Denmark) was coated with 50 120583L of a 2 120583gmLsolution of 1205732GP1 in coating buffer (100mM Na
2CO3
100mM NaHCO3 pH 96) at 4∘C overnight After washing
the plates three times with washing buffer (WB 005 Tween20 01mM CaCl
2 01mM MgCl
2in TBS buffer (150mM
NaCl 50mM TrisHCl pH = 74)) the wells of the coatedplates were blocked with 100 120583Lwell of a 2 BSA solutionfor two hours at 37∘C After three further washing steps withWB 50 120583Lwell of diluted serum (1 200 in WB) was addedand the plates were incubated at 37∘C for two further hoursThen the plates were washed three times with WB again
Development of the plates based on an enzymaticreaction of horseradish peroxidase (HRP) with TMB aschromogenic substrate Therefore the appropriate HRP-conjugated detection-antibody was added as a next step(Table 1) After incubation at 37∘C for another hour andthree washing steps with WB the plates were devel-oped by the addition of 50120583Lwell of freshly preparedTMB-solution (1mL of 001 g 331015840551015840-tetramethylbenzidine(Sigma-Aldrich Steinheim Germany) dissolved in 10mLdimethyl sulfoxide mixed with 9mL of 01M Na
2HPO4
005M citric acid pH 45ndash55 and 2120583L of H2O2) The
reaction was stopped by adding 50120583Lwell of 25 sulfuricacid (H
2SO4) Finally optical density (OD) was measured
at 450 nm and 650 nm as correction factor using an infiniteF200 pro ELISA-reader (Tecan)
After subtraction of the corresponding blank values of theplates experimental values were expressed as optical densityor as mean optical density of each serum when ELISA testswere performed in monoplicates or duplicates respectively
26 IgG-ELISA SNA In order to examine the sialylationof anti-1205732GP1-IgG we slightly modified the previouslydescribed ELISA protocol Before coating the 1205732GP1 antigenwas treated with 1 of periodic acid (PIA Sigma-Aldrich
4 Journal of Immunology Research
Table 2 Detection reagents for lectin ELISA
SNA-ELISAPrimary detectionreagent
Biotinylated SNA-lectin(Galab Technologies Geesthacht Germany)
Secondarydetection reagent
HRP-conjugated streptavidin(Jackson ImmunoResearch laboratoriesNewmarket Suffolk United Kingdom)
Dilution 1 10000SNA = Sambucus nigra agglutinin HRP = horseradish peroxidase
Steinheim Germany) for 48 hours at 43∘C to remove itssugar residues to exclude unspecific binding of Sambucusnigra lectin (SNA) to naive 1205732GP1 After that we centrifugedthe 1205732GP1-solution with 10K Amicon Ultra-centrifugal fil-ter devices (Millipore Tullagreen Ireland) to transfer thetreated 1205732GP1 into coating buffer (100mMNa
2CO3 100mM
NaHCO3 pH 96)
Then the plates were coated with 50 120583Lwell of a 2120583gmLsolution of the PIA-treated 1205732GP1 at 4∘C overnight andsubsequently blocked for two hours at 37∘C with 100 120583Lwellof PIA-treated (05) 3 gelatine (Sigma-Aldrich SteinheimGermany) in TBST buffer with 1mM CaCl
2and 1mM
MgCl2 The subsequent steps were performed as described
in the previous ELISA protocol with two exceptions Insteadof serum we added 50 120583Lwell of purified IgG-samples toexclude possible binding of SNA to anti-1205732GP1-IgM And thedevelopment of the plate required the addition of a seconddetection reagent (Table 2) which was incubated for onehour at room temperature and demanded three preliminarywashing steps We calculated the SNAanti-1205732GP1-IgG ratiofrom the OD values we obtained in the SNA ELISA and asimultaneously conducted anti-1205732GP1-IgG ELISA
27 Statistical Analysis Differences between the cohorts wereassessed using the one-way analysis of variance (ANOVA)test In the case of multiple testing the significance valueswere adjusted by the Newman-Keuls posttest correction
3 Results31 Analysis of Anti-1205732GP1-IgG Wemeasured the OD valuesof total anti-1205732GP1-IgG in the sera of 24 patients with PAPSand 22 patients with SLEAPS 16 healthy children and17NHD as controls As shown in Figure 1 the lowest IgGvalues were to be observed in the cohort of healthy donors(308mOD) whereas the values of the healthy children(970mOD) even exceeded that of the patients with APS(PAPS 686mOD 119901 = 0010 SLEAPS 546mOD 119901 lt0001) All differences between NHD and the other cohortsas well as between the children and the other cohorts weresignificant Patients with PAPS and SLEAPS did not differsignificantly
We calculated the 975 quantile of the anti-1205732GP1-IgGOD values obtained from the NHD cohort and set this valueas cut-off (738mOD) Sera exceeding this value were consid-ered seropositive for anti-1205732GP1-IgG Applying this cut-offvalue we obtained 31 seropositive samples (1224 PAPS 622SAPS 1316 healthy children) which were further analysed in
APS SLEAPS Healthy childrenNHDAPSSLEAPS ns
ns
NHD APS SLEAPS Children0
200
400
600
800
1000
1200
1400
1600
Ant
i-1205732
GP1
-IgG
(mO
D)
p values
mdashmdash
lt00001 lt001 lt00001
lt0001
lt00001
Figure 1 Total anti-1205732GP1 IgG in the sera of NHD PAPSSLEAPS and healthy children and the corresponding 119901 valuesNHD showed significantly lower mOD values than all other cohortswhereas healthy children represented the highest mOD valuesShort horizontal lines mean mOD values of all samples of thecohort Long broken line 975 quantile of NHD (=738mOD)NHD = normal healthy donor PAPS = primary antiphospholipidsyndrome SLEAPS = antiphospholipid syndrome with systemiclupus erythematosus OD = optical density
our study (Supplementary Figure 1 in Supplementary Mate-rial available online at httpdxdoiorg1011552015638129)
32 Analysis of the IgG-Subclasses of Anti-1205732GP1 Analysingthe absolute OD values of the anti-1205732GP1-IgG subclasseswe observed that the children showed significantly higherlevels of the anti-1205732GP1-IgG1 (69mOD) and -IgG3 subclasses(212mOD) compared to NHD PAPS and SAPS (Figures2(a) and 2(c) Table 3(a)) The differences between healthychildren and NHD regarding these two subclasses showeda higher level of significance than the comparison betweenhealthy children and patients with APS However healthychildren did not show significant differences for anti-1205732GP1-IgG2 and -IgG4 when compared to NHD PAPS and SAPS(Figures 2(b) and 2(d) Table 3(b))
Patients with APS (PAPS and SAPS) showed significantlyhigherODvalues for anti-1205732GP1-IgG1 -IgG2 and -IgG3 thanNHD (Figures 2(a) 2(b) and 2(c) Table 3(b)) As in theanti-1205732GP1-IgG ELISA we observed no significant differencebetween the patients with PAPS and SAPS
To summarize patients with APS showed higher ODvalues than NHD in IgG1 IgG2 and IgG3 IgG4 did notshow any significant differenceThe children in turn had thehighest OD values of the IgG1 and IgG3 subclasses
These results were consistent with the analysis of thenumber of seropositive samples as shown in Table 4 Todetermine seropositivity we calculated the mean OD and thestandard deviation (SD) of the NHD group for the differentIgG subclasses IgG values exceeding the NHDrsquos mean OD
Journal of Immunology Research 5
0
50
100
150
200
250
300
350
NHD APS SLEAPS Children
IgG
1 (m
OD
)
(a)
NHD APS SLEAPS Children0
50
100
150
200
250
300
350
IgG
2 (m
OD
)
(b)
NHD APS SLEAPS Children
940483 mOD
0
50
100
150
200
250
300
350
IgG
3 (m
OD
)
(c)
NHD APS SLEAPS Children
508 mOD 482 mOD 665 mOD 1753 mOD
0
50
100
150
200
250
300
350
IgG
4 (m
OD
)
(d)
0
50
100
150
200
250
300
350
(mO
D)
Children NHD APS
IgG4IgG3
IgG2IgG1
(e)
0
10
20
30
40
50
60
70
80
90
100
()
APSChildren
IgG4IgG3
IgG2IgG1
(f)
Figure 2mODvalues of anti-1205732GP1-IgG1 (a) anti-1205732GP1-IgG2 (b) anti-1205732GP1-IgG3 (c) and anti-1205732GP1-IgG4 (d) inNHD PAPS SLEAPSand childrenThe horizontal lines represent the cut-off for seropositivity of the corresponding anti-1205732GP IgG-subclass (gtmeanmOD
[NHD] +2 times SD
[NHD]) NHD = normal healthy donor PAPS = primary antiphospholipid syndrome SLEAPS = antiphospholipid syndrome withsystemic lupus erythematosus OD = optical density Median OD values of the anti-1205732GP1-IgG subclasses in the study cohorts (e) Regardingthe anti-1205732GP1-IgG subclass-distribution within the cohorts children showed a significantly higher contribution of IgG3 than IgG1 andIgG2 Patients with antiphospholipid syndrome (APS) showed a higher content of anti-1205732GP1-IgG2 than -IgG1 and -IgG3 NHD showed ahigher contribution of anti-1205732GP1-IgG2 than -IgG1 -IgG3 and -IgG4The columns represent the median OD values of IgG1 IgG2 IgG3 andIgG4 in the cohorts OD = optical density NHD = normal healthy donor Percentage distribution of anti-1205732GP1-IgG subclasses in childrenand patients with APS (f) Children harbour significantly less anti-1205732GP1-IgG2 than patients with APS but more anti-1205732GP1-IgG3 APS =antiphospholipid syndrome
6 Journal of Immunology Research
0
50
100
150
200
250
300
350
400
Children APS
C3c (
mO
D)
(a)
0
200
400
600
800
1000
1200
Children APS
IgM
(mO
D)
(b)
Figure 3 OD values of C3c (a) bound to anti-1205732GP1 and OD values of anti-1205732GP1-IgM (b) in sera of patients with APS and healthy childrenAnti-1205732GP1 of healthy children displayed significantly more bound C3c than that of patients with APS (119901 = 000007) This suggests a moreefficient clearance of 1205732GP1-immune complexes in the healthy children Anti-1205732GP1-IgM OD values of patients with APS were significantlyhigher than those of healthy children Horizontal lines represent the median OD value of the cohort APS = antiphospholipid syndrome OD= optical density
Table 3 (a) OD values of the anti-1205732GP1-IgG subclasses in theinvestigated cohorts (b) Comparison (119901 values) of the OD valuesof the anti-1205732GP1-IgG subclasses
(a)
IgG1 [mOD] IgG2 [mOD] IgG3 [mOD] IgG4 [mOD]NHD 24 64 27 67Children 69 77 212 210PAPS 30 85 45 72SAPS 28 74 55 173APS 29 81 48 106
(b)
IgG1 IgG2 IgG3 IgG4Children versus NHD lt00001 ns lt00001 nsChildren versus PAPS lt00001 ns lt0001 nsChildren versus SAPS lt0001 ns lt0001 nsAPS versus NHD ns ns ns nsPAPS versus SAPS ns ns ns nsOD = optical density 450 nm PAPS = primary antiphospholipid syndromeSAPS = secondary antiphospholipid syndrome APS = PAPS and SAPSNHD = normal healthy donors ns = not significant
plus two times the SD of the considered subclass were definedas seropositive (Supplementary Figure 1) Applying this cut-off we observed that the healthy children had high levels ofthe IgG1 and IgG3 subclasses with 13 (100) and 8 (62)seropositive samples respectively In contrast only 7 (39)and 0 (0) sera of patients with APS were seropositive forIgG1 and IgG3 respectively APS showed seropositivity ofanti-1205732GP1-IgG2 in 7 cases (39) whereas the cohort ofhealthy children contained just 2 seropositive sera (15) Just
few samples were seropositive for anti-1205732GP1-IgG4 in eachcohort representing a range from 8 (PAPS) to 17 (SAPS)
To determine the subclass distributionwithin the cohortswe calculated the contribution of themeanOD values of eachindividual subclass to the sum
As shown in Figures 2(e) and 2(f) we saw that for thehealthy children IgG3 was the most abundant subclass witha contribution of 40 (IgG3 gt IgG1 (119901 lt 0001) and IgG3 gtIgG2 (119901 lt 001)) In contrast IgG2 dominated the subclassprofile for NHD (46 IgG2 gt IgG1 IgG2 gt IgG3 and IgG2 gtIgG4 (119901 lt 00001)) and patients with APS (38 IgG2 gt IgG1(119901 lt 00001) IgG2 gt IgG3 (119901 = 0001)) The contributionof IgG1 ranged from 12 (SAPS) to 17 (NHD) and thatof IgG4 from 21 (PAPS) to 35 (SAPS) Comparing thecontribution of the individual subclasses between the studycohorts we observed that children showed significantly lessIgG2 than patients with APS and NHD (119901 lt 0001 119901 lt00001) and a higher IgG3 content than NHD and patientswith APS (119901 lt 00001 119901 lt 0001) No significant differencebetween the cohorts was to be seen for the contributions ofIgG1 and IgG4
33 Analysis of IgM C1q and C3c Associated to Anti-1205732GP1-IgG Next we analysed potential 1205732GP1-binding proteins inthe sera of healthy children and adults with APS The signalsfor C1q were low and did not differ significantly between thecohorts
However the ELISAs detecting C3c bound to anti-1205732GP1and anti-1205732GP1-IgM showed highly significant differencesbetween healthy children and adults with APSWhereas C3c-anti-1205732GP1 adducts were increased in healthy children (119901 lt00001 Figure 3(a)) anti-1205732GP1-IgM was decreased (119901 lt0001 Figure 3(b))
Journal of Immunology Research 7
Table 4 Number (frequency) of seropositive samples of the anti-1205732GP1-IgG subclasses
Number of seropositive samples (gtmOD[NHD] + 2 times SD[NHD])IgG1
(gt29mOD)IgG2
(gt79mOD)IgG3
(gt85mOD)IgG4
(gt319mOD)NHD (119899 = 17) 0 (0) 0 (0) 1 (6) 1 (6)Children (119899 = 13) 13 (100) 2 (15) 8 (62) 2 (15)PAPS (119899 = 12) 6 (50) 5 (42) 0 (0) 1 (8)SAPS (119899 = 6) 1 (17) 2 (33) 0 (0) 1 (17)APS (119899 = 18) 7 (39) 7 (39) 0 (0) 2 (11)OD = optical density 450 nm PAPS = primary antiphospholipid syndrome SAPS = secondary antiphospholipid syndrome APS = PAPS and SAPS NHD =normal healthy donorsThe cut-off for seropositivity was calculated from the mean OD of the corresponding anti-1205732GP1-IgG subclass of the NHD cohort andits standard deviation (SD) according to the formula mean OD[NHD] + 2 times SD[NHD] The respective cut-off OD values are indicated in parentheses below theanti-1205732GP1-IgG subclasses
Table 5 Cohorts studied with the lectin ELISA
Cohort Abbreviation Number of samplesPatients with primary APS PAPS 12Patients with APS and SLE as underlying disease SAPS 10Patients with SLE without symptoms of APS harbouring circulating aPL SLE + aPL 15Asymptomatic carriers of aPL aaPL 6Healthy children with circulating anti-1205732GP1 mdash 16APS = antiphospholipid syndrome aPL = antiphospholipid antibodies SLE = systemic lupus erythematosus
34 Analysis of the Sialylation of Anti-1205732GP1 We analysedthe glycosylation of anti-1205732GP1-IgG focusing on its sialy-lation since highly sialylated Fc-parts of IgG are known tocause rather anti-inflammatory responses [30] Wemeasuredthe binding of the sialic acid specific lectin SNA to the anti-1205732GP1-IgG As shown in Table 5 we employed five cohorts ofindividuals 12 patientswith PAPS 10 patientswith SAPS (SLEas primary disease) 15 patients with SLE who did not showclinical sign of APS but with aPL in their sera (SLE + aPL)6 asymptomatic adult carriers of aPL (aaPL) and 16 healthychildren harbouring aPL
Analysing the SNAIgG-ratios (Figure 4) we identifiedthe highest sialylation level in the sera of the healthy childrenThe significances were rather high and decreased from PAPS(119901 lt 00001) to SAPS (119901 lt 00001) to SLE + aPL (119901 lt00001) and to aaPL (119901 lt 0001)
4 Discussion
We analysed two distinct sets of sera One was used toinvestigate anti-1205732GP1-IgG its subclasses and its associatedproteins C1q and C3c For the analysis of the sialylation ofanti-1205732GP1-IgG we had to collect fresh sera from patientswith APS and the healthy children since the stability of theantibody-glycosylation is unknown In order to avoid itspossible decay by processes like enzymatic degradation thisperiod has been kept as short as possible and was followed byimmediate investigation of the glycosylation
The antiphospholipid syndrome is characterized by clin-ical manifestations such as arterial and venous thrombosisor pregnancy-related complications and the detection of spe-cific autoantibodies (lupus anticoagulant anti-1205732GP1- and
anticardiolipin-autoantibodies) [1] In particular anti-1205732GP1sare considered responsible for provoking the pathologicalcomplications [3]We firstmeasured the total amount of anti-1205732GP1-IgG in the sera of 16 healthy children without clinicalmanifestations of APS and 46 adult patients with APS toidentify individual cohorts seropositive for anti-1205732GP1 17adult normal healthy donors served as negative control
The samples of the patientswithAPS showedhigher levelsof anti-1205732GP1-IgG than those of NHD We also observeda higher concentration of anti-1205732GP1-IgG in the sera ofhealthy children compared to NHD This is due to the factthat the detection of these antibodies has been the inclusioncriterion for the healthy children examined in this studyInterestingly the children showed even higher OD values forthe putatively pathogenic anti-1205732GP1-IgG than the patientswith APS This finding was surprising since the childrenwere clinically healthy and did not show any signs of APSsuch as thrombosis or thrombocytopenia complications inwhich the existence of aPL especially anti-1205732GP1 is maderesponsible for [7 33]
High levels of circulating aPL in 61 healthy children whencompared to healthy adults have already been described byAvcin and colleagues [18] The anti-1205732GP1-IgG was report-edly higher in preschool children with a mean age of 5years than in adolescents with a mean age of 135 years[18] The strongly elevated circulating anti-1205732GP1-IgG in ourcohort of healthy children may be explained by the fact thatthey had an even lower age (mean age was nine months)One possible explanation for this result is the inductionof anti-1205732GP1 by viral and bacterial infections [10 11] ornutritional exposure to 1205732GP1 [17] As the immune systemin children is still to evolve they are more susceptible to
8 Journal of Immunology Research
0
05
10
15
20
25SN
AIg
G
ChildrenPAPS SAPS aaPLSLE + aPL
lowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowast
Figure 4 SNAanti-1205732GP1-IgG ratio in PAPS SAPS SLE + aPLaaPL and children Healthy children show a significantly higherSNAanti-1205732GP1-IgG ratio than PAPS SAPS SLE + aPL and aaPL(lowastlowast119901 lt 00001 lowastlowastlowast119901 lt 000001) This suggests higher sialylation ofthe oligosaccharides attached to Fc fragments of the childrenrsquos anti-1205732GP1No significant differenceswere foundbetween the remainingcohorts Horizontal lines indicate the mean SNAIgG ratio of thecohort SNA = Sambucus nigra agglutinin APS = antiphospholipidsyndrome PAPS = patients with primary APS SAPS = patients withAPS and SLE as underlying disease SLE + aaPL = patients withSLE without symptoms of APS harbouring circulating aPL aaPL =asymptomatic carriers of aPL aPL = antiphospholipid antibodiesSLE = systemic lupus erythematosus
infections than adults and their immune system has to copewith a plethora of viral and bacterial pathogens for the firsttime in their life This scenario is prone to result in thestimulation of autoantibody-production Molecular mimicryof certain pathogens can lead to the production of antibodieswhich then cross-react with 1205732GP1 [7 12ndash15] When infantsdiversify their dietary habits and start to consume productslike milk and meat they can get into contact with nutritional1205732GP1 [19] This is a further condition capable triggeringthe production of anti-1205732GP1 Exposure to pathogen-derivedand dietary 1205732GP1 could explain the high level of circulatinganti-1205732GP1-IgG in healthy childrenThis low affine reactivityis consecutively lost during the maturation of the immunesystem and replaced by the emergence in adults of high affineantibodies that more specifically recognize pathogens Apartfrom possible origins of these autoantibodies it is obviousthat they do not provoke any of the hallmark symptoms ofAPS In contrast to their adult counterparts anti-1205732GP1-IgGscirculating in the plasma of healthy children are obviouslynot pathogenic So it seems decent to hypothesize thatanti-1205732GP1 autoantibodies in healthy children biochemicallydiffer from those in adults
To test this hypothesis we examined the anti-1205732GP1-IgG subclass distribution in the seropositive sera of the anti-1205732GP1-IgG ELISA The healthy children showed higher ODvalues for the IgG1- and IgG3-subclasses than patients withAPS and NHD This was consistent with the number ofseropositive samples In the anti-1205732GP1-IgG1-subclass every
tested serum of the healthy children surpassed the thresholdset for seropositivity (gtmean OD
[NHD] + 2 times SD[NHD]) and
8 samples (62) did so in the IgG3-subclass In the seraof patients with APS just 7 (39) and no samples wereseropositive for IgG1 or IgG3 In the sera of patients with APScompared to NHD we observed higher OD levels of anti-1205732GP1 in all subclasses except for IgG4This result is consis-tent with the findings of Guerin et al who reported elevatedlevels in all anti-1205732GP1-IgG subclasses even including IgG4in patients with APS compared to a control group of healthyadults [23]
It might be that the elevated absolute OD values of theanti-1205732GP1-IgG subclasses are the logical consequence ofthe generally higher level of anti-1205732GP1-IgG in the patientswith APS and healthy children who were selected for furthertesting Therefore we also analysed the subclass distributionof this antibody We first analysed the anti-1205732GP1-IgG sub-class distribution within the individual cohorts and observedthat in the healthy children the IgG3 antibody was mostabundant with a contribution of 40 to the total reactivityThis is consistent with the finding that all sera of the healthychildren were seropositive in this subclass and that theiranti-1205732GP1-IgG3 OD values were the highest of all cohortsThis indicates a predominance of this subclass in the healthychildren The remaining anti-1205732GP1-IgG subclasses in thesera of the children ranged from 15 (IgG1) over 20 (IgG2)to 25 (IgG4) but the differences between their proportionswere not statistically significant In contrast in the sera ofthe patients with APS the anti-1205732GP1-IgG2 subclass had thehighest contribution with 38
The finding of an elevated proportion of the IgG2 subclassin the patients with APS confirms former published datawhich also describe a skewing towards anti-1205732GP1-IgG2 inAPS [19 23 26 27] The authors detected an even highercontribution of anti-1205732GP1-IgG2 (702 [26] and 87 [27]resp) In contrast to our results the IgG1 subclass wasmore prominent in the published studies described aboveImportantly in the cohort of the healthy infants IgG2 wasrelatively lowTherefore onemay argue that anti-1205732GP1-IgG2is not just a marker of the disease but is also involved inthe pathogenic action of the aPL autoantibodies When wecompared the anti-1205732GP1-IgG subclass distribution betweenthe three cohorts we noticed a significant difference in thedistribution concerning the children on one hand and NHDand patients with APS on the other Healthy children showeda higher proportion of anti-1205732GP1-IgG3 and a lower one ofanti-1205732GP1-IgG2 In NHD and patients with APS it was theother way round IgG1 and IgG4 did not significantly differbetween healthy children and patients with APS orNHDThepredominance of anti-1205732GP1-IgG3 in the healthy children isa hint to an immune response elicited by a proteinaceousantigen [26 27 34] This supports the hypothesis thatthe harmless anti-1205732GP1 in children are infection-inducedwith pathogen-derived proteins serving as trigger for theirproduction However it is still elusive if the IgG3-dominatedanti-1205732GP1 response found in the sera of healthy childrencontributes to its low pathogenic potential Alternativelyit can just be ascribed to the proceeding evolution of theinfantrsquos immune system typically associated with an elevated
Journal of Immunology Research 9
proportion of IgG3 antibodies [34 35] During childhoodthe proportion of IgG3 continuously decreases to adult rangesusually reached at the age of 10ndash12 years However the aboveaverage elevated proportion of the anti-1205732GP1-IgG3 (40) inthe sera of the healthy childrenmay thus not just be due to theinfant immune system In particular when bearing in mindthat normal adult levels of the IgG3 subclass are just around6 [34 36]
IgG3 is the IgG-subclass exerting the most effectiveactivation of the complement cascade through the classicalpathway [34] Since complement activation is associated withthe pathogenesis of APS [21 32 37] the high proportionof complement-activating anti-1205732GP1-IgG3 in the childrenseems to contradict a possible protective effect against APS ofthese kinds of antibodies However the role of complement inthe etiopathogenesis of autoimmune diseases is manifold asthe complement components C1q C2 and C4 contribute tothe clearance of postapoptotic secondary necrotic cells andtheir deficiency is therefore a risk factor for autoimmunityand autoinflammation [38] In a similar manner studiesin mouse in vivo models of various autoimmune diseasesdemonstrated that IgG mediated tissue inflammation wasblocked in mice deficient in activating Fc120574Rs although thecomplement component C3 was still abundantly depositedin the tissue [39ndash41] Thus the contribution of complementdeposits in tissue with respect to tissue inflammation remainsto be established
To obtain new insights into the involvement of thecomplement system in the pathogenesis of APS we analysedthe C1q- and C3c-binding to anti-1205732GP1 We observed thatanti-1205732GP1 in the sera of the healthy children and in patientswith APS similarly bound C1q the first molecule of theclassical pathway of complement activation [42] Performinganti-1205732GP1-IgM-ELISAs we detected significantly higherIgM values in the patients with APS One would expecta higher C1q-binding of the IgM positive sera since thisimmunoglobulin binds and activates C1q more strongly thanIgG However the children mainly harbour IgG3 autoanti-bodies the most potent subclass for the activation of theclassical complement pathway [34]This may compensate forthe lower IgM level in their sera and thus be responsiblefor the similar C1q-binding of both cohorts This suggeststhat preconditions for complement activation via the classicalpathway are comparable in both cohorts We conclude thatclassical complement activation has a minor impact on thedifferential pathogenesis of aPL in healthy children andpatients with APS
What then is the culprit that induces their pathogenicpotential
Nextwe quantified the contribution ofC3c to the immunecomplexes bound to 1205732GP1 C3c is a domain in the C3bmolecule and one of its cleavage products created aftercleavage by factor I of C3b [42 43] C3c not just representsa marker of complement activation but also serves as anopsonin improving the clearance of bound targets Weobserved significantly more C3c bound to anti-1205732GP1 inthe sera of the healthy children than in those of patientswith APS This reflects a better opsonization by C3c ofthe potentially harmful anti-1205732GP1 immune complexes
anti-1205732GP1-opsonized cells or cellular fragments in thesera of the healthy children Consequently the anti-1205732GP1complexes can be cleared more efficiently by the powerfulcomplement system of the healthy children In patients withchronic inflammatory autoimmune diseases complement isusually low which may result from null alleles [44] or fromcomplement consumption [38]
But why do these processes not cause inflammation andaberrant coagulation in the childrenApart fromanti-1205732GP1-IgG subclass distribution and distinct ability to activate thecomplement system it is likely that other properties of theanti-1205732GP1 in the sera of children contribute to their lowpathogenicity We hypothesized that an increased sialylationof the anti-1205732GP1-IgG in healthy individuals renders themharmless for their hosts IgG is usually N-glycosylated at theCH2-domain of its Fc-part This N-glycan crucially modifiesthe effector functions the IgG molecule is endowed with[45ndash47] This biantennary core-fucosylated oligosaccharidecarries a variable amount of sugar monomers attached toboth arms [28 48] IgG antibodies terminating with sialicacids on one or both antennae have been shown to behaveanti-inflammatorily after infusion of IVIG [29 31]The signalexerted by the terminal sialic acid is dominant since thefraction of sialylated IgG in healthy plasma is just 4 to 14[28 46 49]
In mice these effects are mainly ascribed to decreasedbinding of sialylated IgG to activating Fc120574-receptors andan upregulation of the inhibitory Fc120574RIIB [29 31] Weobserved that anti-1205732GP1-IgG isolated from sera of healthychildren was significantly more sialylated than that fromsymptomatic patients with APS One may predict that theseautoantibodies in complex with 1205732GP1 show low affinity foractivating Fc120574Rs and therefore do not provoke proinflam-matory and procoagulant effector functions Fc120574RIIA is anexample for an activating receptor It is located on a plethoraof cells On the surfaces of platelets it mediates activationby immune complexes The activation is accompanied byplatelet aggregation and the release of mediators such asthe prothromboticThromboxaneA2 [50] Consecutively thiscascade can result in thrombus formation [51 52] Sialylatedanti-1205732GP1-IgG is unable to bind to and to activate plateletsvia Fc120574RIIA
Further mechanisms involved in thrombus formation inAPS comprise the interaction of anti-1205732GP1 with variouspattern recognition receptors [53] and its interference withregulatory functions ofmolecules involved in the coagulationprocess (protein C prothrombin or tissue factor [52 54])Direct binding to 1205732GP1 on endothelial cells (EC) shiftstheir phenotype towards coagulation and inflammation [54]Considering that Fc120574R-binding is involved in many of thesemechanisms it seems likely that a high sialylation of anti-1205732GP1-IgG leads to a low pathogenicity of these autoan-tibodies and does not cause APS Therefore we concludethat the higher sialylation of anti-1205732GP1-IgG in the seraof the healthy children plays a major role in protectingthem from typical complications of APS This hypothesis isfurther supported by the observation that antiphospholipidantibodies from sera of asymptomatic adult carriers aremore sialylated than those from patients with APS A high
10 Journal of Immunology Research
sialylation status of antiphospholipid antibodies seems toprevent the development of symptoms of APS also in theseindividuals
We conclude that a plethora of factors contributes tothe low pathogenicity of anti-1205732GP1-IgG in the sera ofhealthy children and asymptomatic adults We observed adistinct spectrum of IgG-subclasses compared to patientswith APS and a better binding of C3c indicating a moreefficient clearance of potentially harmful anti-1205732GP1 Thehypersialylation of the anti-1205732GP1-IgG is prone to amelioratetheir inflammatory and procoagulatory effects in healthycarriers whereas hyposialylation and inflammationmutuallyprecipitate the antiphospholipid syndrome
Disclosure
Thepresent workwas performed in fulfillment of the require-ments for obtaining the degree ldquoDr medrdquo for ChristophFickentscher
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Laura Andreoli andMartin Herrmann contributed equally tosenior authorship
Acknowledgments
This project was supported by the German Research Foun-dation (training research programme of the collaborativeresearch center 643) by the Emerging Fields Initiative (EFI)of the FAU Erlangen-Nuremberg and by the K und RWucherpfennig-Stiftung
References
[1] S Miyakis M D Lockshin T Atsumi et al ldquoInternationalconsensus statement on an update of the classification criteriafor definite antiphospholipid syndrome (APS)rdquo Journal ofThrombosis and Haemostasis vol 4 no 2 pp 295ndash306 2006
[2] W A Wilson A E Gharavi T Koike et al ldquoInternationalconsensus statement on preliminary classification criteria fordefinite antiphospholipid syndrome report of an internationalworkshoprdquo Arthritis and Rheumatism vol 42 no 7 pp 1309ndash1311 1999
[3] P G de Groot and R T Urbanus ldquoThe significance of autoan-tibodies against beta2-glycoprotein Irdquo Blood vol 120 no 2 pp266ndash274 2012
[4] C Nalli S Piantoni L Andreoli M Motta and A TincanildquoAntiphospholipid syndrome and antiphospholipid antibodiesin children the two sides of the coinrdquo Israel Medical AssociationJournal vol 14 no 5 pp 310ndash312 2012
[5] I Schousboe ldquo1205732-glycoprotein I a plasma inhibitor of thecontact activation of the intrinsic blood coagulation pathwayrdquoBlood vol 66 no 5 pp 1086ndash1091 1985
[6] J George M Blank Y Levy et al ldquoDifferential effects of anti-1205732-glycoprotein I antibodies on endothelial cells and on themanifestations of experimental antiphospholipid syndromerdquoCirculation vol 97 no 9 pp 900ndash906 1998
[7] R Willis and S S Pierangeli ldquoAnti-1205732-glycoprotein I antibod-iesrdquo Annals of the New York Academy of Sciences vol 1285 no1 pp 44ndash58 2013
[8] M J Fischer J Rauch and J S Levine ldquoThe antiphospholipidsyndromerdquo Seminars in Nephrology vol 27 no 1 pp 35ndash462007
[9] H Amital M Govoni R Maya et al ldquoRole of infectious agentsin systemic rheumatic diseasesrdquo Clinical and ExperimentalRheumatology vol 26 no 1 supplement 48 pp S27ndashS32 2008
[10] T Avcin ldquoAntiphospholipid syndrome in childrenrdquo CurrentOpinion in Rheumatology vol 20 no 5 pp 595ndash600 2008
[11] M Garcıa-Carrasco C Galarza-Maldonado C Mendoza-Pinto R O Escarcega and R Cervera ldquoInfections and theantiphospholipid syndromerdquo Clinical Reviews in Allergy ampImmunology vol 36 no 2-3 pp 104ndash108 2009
[12] P Cruz-Tapias M Blank J-M Anaya and Y ShoenfeldldquoInfections and vaccines in the etiology of antiphospholipidsyndromerdquo Current Opinion in Rheumatology vol 24 no 4 pp389ndash393 2012
[13] P von Landenberg Y Doring S Modrow and K J LacknerldquoAre antiphospholipid antibodies an essential requirement foran effective immune response to infectionsrdquoAnnals of the NewYork Academy of Sciences vol 1108 pp 578ndash583 2007
[14] M Blank I Krause M Fridkin et al ldquoBacterial induction ofautoantibodies to 1205732-glycoprotein-I accounts for the infectiousetiology of antiphospholipid syndromerdquo Journal of ClinicalInvestigation vol 109 no 6 pp 797ndash804 2002
[15] Y Shoenfeld M Blank R Cervera J Font E Raschi and P-LMeroni ldquoInfectious origin of the antiphospholipid syndromerdquoAnnals of the Rheumatic Diseases vol 65 no 1 pp 2ndash6 2006
[16] A E Gharavi S S Pierangeli R G Espinola X Liu MColden-Stanfield and E N Harris ldquoAntiphospholipid antibod-ies induced in mice by immunization with a cytomegalovirus-derived peptide cause thrombosis and activation of endothelialcells in vivordquo Arthritis and Rheumatism vol 46 no 2 pp 545ndash552 2002
[17] M Motta G Chirico C B Rebaioli et al ldquoAnticardiolipin andanti-beta2 glycoprotein I antibodies in infants born to motherswith antiphospholipid antibody-positive autoimmune diseasea follow-up studyrdquoAmerican Journal of Perinatology vol 23 no4 pp 247ndash251 2006
[18] T Avcin A Ambrozic M Kuhar T Kveder and B RozmanldquoAnticardiolipin and anti-1205732 glycoprotein I antibodies in seraof 61 apparently healthy children at regular preventive visitsrdquoRheumatology vol 40 no 5 pp 565ndash573 2001
[19] A Ambrozic ldquoAnti-beta2-glycoprotein I antibodies in children
with atopic dermatitisrdquo International Immunology vol 14 no 7pp 823ndash830
[20] J A Giron-Gonzalez E Garcıa Del Rıo C Rodrıguez JRodrıguez-Martorell and A Serrano ldquoAntiphospholipid syn-drome and asymptomatic carriers of antiphospholipid anti-body prospective analysis of 404 individualsrdquo Journal ofRheumatology vol 31 no 8 pp 1560ndash1567 2004
[21] K A Breen P Seed K Parmar GWMoore S E Stuart-Smithand B J Hunt ldquoComplement activation in patients with isolatedantiphospholipid antibodies or primary antiphospholipid syn-dromerdquo Thrombosis and Haemostasis vol 107 no 3 pp 423ndash429 2012
Journal of Immunology Research 11
[22] F Fischetti P Durigutto V Pellis et al ldquoThrombus formationinduced by antibodies to 1205732-glycoprotein I is complementdependent and requires a priming factorrdquo Blood vol 106 no7 pp 2340ndash2346 2005
[23] J Guerin E Casey C Feighery and J Jackson ldquoAnti-Beta 2-glycoprotein I antibody isotype and IgG subclass in antiphos-pholipid syndrome patientsrdquo Autoimmunity vol 31 no 2 pp109ndash116 1999
[24] L Andreoli C Nalli M Motta et al ldquoAnti-1205732-glycoprotein I
IgG antibodies from 1-year-old healthy children born to moth-ers with systemic autoimmune diseases preferentially targetdomain 45 might it be the reason for their lsquoinnocentrsquo profilerdquoAnnals of the Rheumatic Diseases vol 70 no 2 pp 380ndash3832011
[25] B de Laat R H W M Derksen M van Lummel M T T Pen-nings and P G deGroot ldquoPathogenic anti-beta2-glycoprotein Iantibodies recognize domain I of beta2-glycoprotein I only aftera conformational changerdquo Blood vol 107 no 5 pp 1916ndash19242006
[26] M Samarkos K A Davies C Gordon M J Walport and SLoizou ldquoIgG subclass distribution of antibodies against 1205732-GP1and cardiolipin in patients with systemic lupus erythematosusand primary antiphospholipid syndrome and their clinicalassociationsrdquo Rheumatology vol 40 no 9 pp 1026ndash1032 2001
[27] J Arvieux B Roussel D Ponard and M G Colomb ldquoIgG2subclass restriction of anti-1205732 glycoprotein 1 antibodies inautoimmune patientsrdquo Clinical and Experimental Immunologyvol 95 no 2 pp 310ndash315 1994
[28] J N Arnold M R Wormald R B Sim P M Rudd and R ADwek ldquoThe impact of glycosylation on the biological functionand structure of human immunoglobulinsrdquo Annual Review ofImmunology vol 25 pp 21ndash50 2007
[29] Y Kaneko F Nimmerjahn and J V Ravetch ldquoAnti-inflamma-tory activity of immunoglobulin G resulting from Fc sialyla-tionrdquo Science vol 313 no 5787 pp 670ndash673 2006
[30] RM Anthony and F Nimmerjahn ldquoThe role of differential IgGglycosylation in the interaction of antibodies with FcgammaRsin vivordquo Current Opinion in Organ Transplantation vol 16 no1 pp 7ndash14 2011
[31] R M Anthony and J V Ravetch ldquoA novel role for the IgGFc glycan the anti-inflammatory activity of sialylated IgG FcsrdquoJournal of Clinical Immunology vol 30 no 1 pp S9ndashS14 2010
[32] K Oku T Atsumi M Bohgaki et al ldquoComplement activationin patients with primary antiphospholipid syndromerdquoAnnals ofthe Rheumatic Diseases vol 68 no 6 pp 1030ndash1035 2009
[33] H P McNeil R J Simpson C N Chesterman and S AKrilis ldquoAnti-phospholipid antibodies are directed against acomplex antigen that includes a lipid-binding inhibitor of coag-ulation 1205732-Glycoprotein I (apolipoprotein H)rdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 87 no 11 pp 4120ndash4124 1990
[34] R Jefferis and D S Kumararatne ldquoSelective IgG subclassdeficiency quantification and clinical relevancerdquo Clinical andExperimental Immunology vol 81 no 3 pp 357ndash367 1990
[35] P H Schur F Rosen and M E Norman ldquoImmunoglobulinsubclasses in normal childrenrdquo Pediatric Research vol 13 no3 pp 181ndash183 1979
[36] M A H French and G Harrison ldquoSerum IgG subclass concen-trations in healthy adults a study using monoclonal antiserardquoClinical and Experimental Immunology vol 56 no 2 pp 473ndash475 1984
[37] M Odorczuk L B Keil and V A DeBari ldquoBinding of com-plement component C1q to anti-beta2 glycoprotein I antibodiesfrom patients with antiphospholipid syndromerdquo InflammationResearch vol 48 no 10 pp 524ndash526 1999
[38] L Truedsson A A Bengtsson and G Sturfelt ldquoComplementdeficiencies and systemic lupus erythematosusrdquoAutoimmunityvol 40 no 8 pp 560ndash566 2007
[39] S Aschermann C H K Lehmann S Mihai G Schett DDudziak and F Nimmerjahn ldquoB cells are critical for autoim-mune pathology in Scurfy micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 110 no47 pp 19042ndash19047 2013
[40] Y Kaneko F Nimmerjahn M P Madaio and J V RavetchldquoPathology and protection in nephrotoxic nephritis is deter-mined by selective engagement of specific Fc receptorsrdquo TheJournal of Experimental Medicine vol 203 no 3 pp 789ndash7972006
[41] D Sylvestre R Clynes M Ma H Warren M C Carrolland J V Ravetch ldquoImmunoglobulin G-mediated inflammatoryresponses develop normally in complement-deficient micerdquoJournal of ExperimentalMedicine vol 184 no 6 pp 2385ndash23921996
[42] G E Ritchie B E Moffatt R B Sim B P Morgan R A Dwekand P M Rudd ldquoGlycosylation and the complement systemrdquoChemical Reviews vol 102 no 2 pp 305ndash320 2002
[43] Y Palarasah K Skjodt J Brandt et al ldquoGeneration of aC3c specific monoclonal antibody and assessment of C3c as aputative inflammatorymarker derived from complement factorC3rdquo Journal of ImmunologicalMethods vol 362 no 1-2 pp 142ndash150 2010
[44] D G Doherty J A Underhill P T Donaldson et al ldquoPoly-morphism in the human complement C4 genes and geneticsusceptibility to autoimmune hepatitisrdquo Autoimmunity vol 18no 4 pp 243ndash249 1994
[45] M-H Tao and S L Morrison ldquoStudies of aglycosylatedchimeric mouse-human IgG Role of carbohydrate in thestructure and effector functions mediated by the human IgGconstant regionrdquoThe Journal of Immunology vol 143 no 8 pp2595ndash2601 1989
[46] R Jefferis ldquoGlycosylation of recombinant antibody therapeu-ticsrdquo Biotechnology Progress vol 21 no 1 pp 11ndash16 2005
[47] Y Mimura S Church R Ghirlando et al ldquoThe influence ofglycosylation on the thermal stability and effector functionexpression of human IgG1-Fc properties of a series of truncatedglycoformsrdquoMolecular Immunology vol 37 no 12-13 pp 697ndash706 2001
[48] C Huhn M H J Selman L R Ruhaak A M Deelder and MWuhrer ldquoIgG glycosylation analysisrdquo Proteomics vol 9 no 4pp 882ndash913 2009
[49] J Lund N Takahashi J D Pound M Goodall and R JefferisldquoMultiple interactions of IgG with its core oligosaccharide canmodulate recognition by complement and human Fc gammareceptor I and influence the synthesis of its oligosaccharidechainsrdquo The Journal of Immunology vol 157 no 11 pp 4963ndash4969 1996
[50] R Forastiero M Martinuzzo L O Carreras and J MacloufldquoAnti-1205732 glycoprotein I antibodies and platelet activation inpatients with antiphospholipid antibodies association withincreased excretion of platelet-derived thromboxane urinarymetabolitesrdquoThrombosis and Haemostasis vol 79 no 1 pp 42ndash45 1998
12 Journal of Immunology Research
[51] J-C Reverter and D Tassies ldquoMechanism of thrombosis inantiphospholipid syndrome binding to plateletsrdquo in HughesSyndrome M A Khamashta Ed pp 403ndash414 Springer Lon-don UK 2006
[52] G Espinosa R Cervera J Font and Y Shoenfeld ldquoAntiphos-pholipid syndrome pathogenic mechanismsrdquo AutoimmunityReviews vol 2 no 2 pp 86ndash93 2003
[53] T Shi B Giannakopoulos X Yan et al ldquoAnti-1205732-glycoprotein
I antibodies in complex with 1205732-glycoprotein I can activate
platelets in a dysregulated manner via glycoprotein Ib-IX-VrdquoArthritis amp Rheumatism vol 54 no 8 pp 2558ndash2567 2006
[54] P LMeroni E RaschiMCamera et al ldquoEndothelial activationby aPL a potential pathogenetic mechanism for the clinicalmanifestations of the syndromerdquo Journal of Autoimmunity vol15 no 2 pp 237ndash240 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

4 Journal of Immunology Research
Table 2 Detection reagents for lectin ELISA
SNA-ELISAPrimary detectionreagent
Biotinylated SNA-lectin(Galab Technologies Geesthacht Germany)
Secondarydetection reagent
HRP-conjugated streptavidin(Jackson ImmunoResearch laboratoriesNewmarket Suffolk United Kingdom)
Dilution 1 10000SNA = Sambucus nigra agglutinin HRP = horseradish peroxidase
Steinheim Germany) for 48 hours at 43∘C to remove itssugar residues to exclude unspecific binding of Sambucusnigra lectin (SNA) to naive 1205732GP1 After that we centrifugedthe 1205732GP1-solution with 10K Amicon Ultra-centrifugal fil-ter devices (Millipore Tullagreen Ireland) to transfer thetreated 1205732GP1 into coating buffer (100mMNa
2CO3 100mM
NaHCO3 pH 96)
Then the plates were coated with 50 120583Lwell of a 2120583gmLsolution of the PIA-treated 1205732GP1 at 4∘C overnight andsubsequently blocked for two hours at 37∘C with 100 120583Lwellof PIA-treated (05) 3 gelatine (Sigma-Aldrich SteinheimGermany) in TBST buffer with 1mM CaCl
2and 1mM
MgCl2 The subsequent steps were performed as described
in the previous ELISA protocol with two exceptions Insteadof serum we added 50 120583Lwell of purified IgG-samples toexclude possible binding of SNA to anti-1205732GP1-IgM And thedevelopment of the plate required the addition of a seconddetection reagent (Table 2) which was incubated for onehour at room temperature and demanded three preliminarywashing steps We calculated the SNAanti-1205732GP1-IgG ratiofrom the OD values we obtained in the SNA ELISA and asimultaneously conducted anti-1205732GP1-IgG ELISA
27 Statistical Analysis Differences between the cohorts wereassessed using the one-way analysis of variance (ANOVA)test In the case of multiple testing the significance valueswere adjusted by the Newman-Keuls posttest correction
3 Results31 Analysis of Anti-1205732GP1-IgG Wemeasured the OD valuesof total anti-1205732GP1-IgG in the sera of 24 patients with PAPSand 22 patients with SLEAPS 16 healthy children and17NHD as controls As shown in Figure 1 the lowest IgGvalues were to be observed in the cohort of healthy donors(308mOD) whereas the values of the healthy children(970mOD) even exceeded that of the patients with APS(PAPS 686mOD 119901 = 0010 SLEAPS 546mOD 119901 lt0001) All differences between NHD and the other cohortsas well as between the children and the other cohorts weresignificant Patients with PAPS and SLEAPS did not differsignificantly
We calculated the 975 quantile of the anti-1205732GP1-IgGOD values obtained from the NHD cohort and set this valueas cut-off (738mOD) Sera exceeding this value were consid-ered seropositive for anti-1205732GP1-IgG Applying this cut-offvalue we obtained 31 seropositive samples (1224 PAPS 622SAPS 1316 healthy children) which were further analysed in
APS SLEAPS Healthy childrenNHDAPSSLEAPS ns
ns
NHD APS SLEAPS Children0
200
400
600
800
1000
1200
1400
1600
Ant
i-1205732
GP1
-IgG
(mO
D)
p values
mdashmdash
lt00001 lt001 lt00001
lt0001
lt00001
Figure 1 Total anti-1205732GP1 IgG in the sera of NHD PAPSSLEAPS and healthy children and the corresponding 119901 valuesNHD showed significantly lower mOD values than all other cohortswhereas healthy children represented the highest mOD valuesShort horizontal lines mean mOD values of all samples of thecohort Long broken line 975 quantile of NHD (=738mOD)NHD = normal healthy donor PAPS = primary antiphospholipidsyndrome SLEAPS = antiphospholipid syndrome with systemiclupus erythematosus OD = optical density
our study (Supplementary Figure 1 in Supplementary Mate-rial available online at httpdxdoiorg1011552015638129)
32 Analysis of the IgG-Subclasses of Anti-1205732GP1 Analysingthe absolute OD values of the anti-1205732GP1-IgG subclasseswe observed that the children showed significantly higherlevels of the anti-1205732GP1-IgG1 (69mOD) and -IgG3 subclasses(212mOD) compared to NHD PAPS and SAPS (Figures2(a) and 2(c) Table 3(a)) The differences between healthychildren and NHD regarding these two subclasses showeda higher level of significance than the comparison betweenhealthy children and patients with APS However healthychildren did not show significant differences for anti-1205732GP1-IgG2 and -IgG4 when compared to NHD PAPS and SAPS(Figures 2(b) and 2(d) Table 3(b))
Patients with APS (PAPS and SAPS) showed significantlyhigherODvalues for anti-1205732GP1-IgG1 -IgG2 and -IgG3 thanNHD (Figures 2(a) 2(b) and 2(c) Table 3(b)) As in theanti-1205732GP1-IgG ELISA we observed no significant differencebetween the patients with PAPS and SAPS
To summarize patients with APS showed higher ODvalues than NHD in IgG1 IgG2 and IgG3 IgG4 did notshow any significant differenceThe children in turn had thehighest OD values of the IgG1 and IgG3 subclasses
These results were consistent with the analysis of thenumber of seropositive samples as shown in Table 4 Todetermine seropositivity we calculated the mean OD and thestandard deviation (SD) of the NHD group for the differentIgG subclasses IgG values exceeding the NHDrsquos mean OD
Journal of Immunology Research 5
0
50
100
150
200
250
300
350
NHD APS SLEAPS Children
IgG
1 (m
OD
)
(a)
NHD APS SLEAPS Children0
50
100
150
200
250
300
350
IgG
2 (m
OD
)
(b)
NHD APS SLEAPS Children
940483 mOD
0
50
100
150
200
250
300
350
IgG
3 (m
OD
)
(c)
NHD APS SLEAPS Children
508 mOD 482 mOD 665 mOD 1753 mOD
0
50
100
150
200
250
300
350
IgG
4 (m
OD
)
(d)
0
50
100
150
200
250
300
350
(mO
D)
Children NHD APS
IgG4IgG3
IgG2IgG1
(e)
0
10
20
30
40
50
60
70
80
90
100
()
APSChildren
IgG4IgG3
IgG2IgG1
(f)
Figure 2mODvalues of anti-1205732GP1-IgG1 (a) anti-1205732GP1-IgG2 (b) anti-1205732GP1-IgG3 (c) and anti-1205732GP1-IgG4 (d) inNHD PAPS SLEAPSand childrenThe horizontal lines represent the cut-off for seropositivity of the corresponding anti-1205732GP IgG-subclass (gtmeanmOD
[NHD] +2 times SD
[NHD]) NHD = normal healthy donor PAPS = primary antiphospholipid syndrome SLEAPS = antiphospholipid syndrome withsystemic lupus erythematosus OD = optical density Median OD values of the anti-1205732GP1-IgG subclasses in the study cohorts (e) Regardingthe anti-1205732GP1-IgG subclass-distribution within the cohorts children showed a significantly higher contribution of IgG3 than IgG1 andIgG2 Patients with antiphospholipid syndrome (APS) showed a higher content of anti-1205732GP1-IgG2 than -IgG1 and -IgG3 NHD showed ahigher contribution of anti-1205732GP1-IgG2 than -IgG1 -IgG3 and -IgG4The columns represent the median OD values of IgG1 IgG2 IgG3 andIgG4 in the cohorts OD = optical density NHD = normal healthy donor Percentage distribution of anti-1205732GP1-IgG subclasses in childrenand patients with APS (f) Children harbour significantly less anti-1205732GP1-IgG2 than patients with APS but more anti-1205732GP1-IgG3 APS =antiphospholipid syndrome
6 Journal of Immunology Research
0
50
100
150
200
250
300
350
400
Children APS
C3c (
mO
D)
(a)
0
200
400
600
800
1000
1200
Children APS
IgM
(mO
D)
(b)
Figure 3 OD values of C3c (a) bound to anti-1205732GP1 and OD values of anti-1205732GP1-IgM (b) in sera of patients with APS and healthy childrenAnti-1205732GP1 of healthy children displayed significantly more bound C3c than that of patients with APS (119901 = 000007) This suggests a moreefficient clearance of 1205732GP1-immune complexes in the healthy children Anti-1205732GP1-IgM OD values of patients with APS were significantlyhigher than those of healthy children Horizontal lines represent the median OD value of the cohort APS = antiphospholipid syndrome OD= optical density
Table 3 (a) OD values of the anti-1205732GP1-IgG subclasses in theinvestigated cohorts (b) Comparison (119901 values) of the OD valuesof the anti-1205732GP1-IgG subclasses
(a)
IgG1 [mOD] IgG2 [mOD] IgG3 [mOD] IgG4 [mOD]NHD 24 64 27 67Children 69 77 212 210PAPS 30 85 45 72SAPS 28 74 55 173APS 29 81 48 106
(b)
IgG1 IgG2 IgG3 IgG4Children versus NHD lt00001 ns lt00001 nsChildren versus PAPS lt00001 ns lt0001 nsChildren versus SAPS lt0001 ns lt0001 nsAPS versus NHD ns ns ns nsPAPS versus SAPS ns ns ns nsOD = optical density 450 nm PAPS = primary antiphospholipid syndromeSAPS = secondary antiphospholipid syndrome APS = PAPS and SAPSNHD = normal healthy donors ns = not significant
plus two times the SD of the considered subclass were definedas seropositive (Supplementary Figure 1) Applying this cut-off we observed that the healthy children had high levels ofthe IgG1 and IgG3 subclasses with 13 (100) and 8 (62)seropositive samples respectively In contrast only 7 (39)and 0 (0) sera of patients with APS were seropositive forIgG1 and IgG3 respectively APS showed seropositivity ofanti-1205732GP1-IgG2 in 7 cases (39) whereas the cohort ofhealthy children contained just 2 seropositive sera (15) Just
few samples were seropositive for anti-1205732GP1-IgG4 in eachcohort representing a range from 8 (PAPS) to 17 (SAPS)
To determine the subclass distributionwithin the cohortswe calculated the contribution of themeanOD values of eachindividual subclass to the sum
As shown in Figures 2(e) and 2(f) we saw that for thehealthy children IgG3 was the most abundant subclass witha contribution of 40 (IgG3 gt IgG1 (119901 lt 0001) and IgG3 gtIgG2 (119901 lt 001)) In contrast IgG2 dominated the subclassprofile for NHD (46 IgG2 gt IgG1 IgG2 gt IgG3 and IgG2 gtIgG4 (119901 lt 00001)) and patients with APS (38 IgG2 gt IgG1(119901 lt 00001) IgG2 gt IgG3 (119901 = 0001)) The contributionof IgG1 ranged from 12 (SAPS) to 17 (NHD) and thatof IgG4 from 21 (PAPS) to 35 (SAPS) Comparing thecontribution of the individual subclasses between the studycohorts we observed that children showed significantly lessIgG2 than patients with APS and NHD (119901 lt 0001 119901 lt00001) and a higher IgG3 content than NHD and patientswith APS (119901 lt 00001 119901 lt 0001) No significant differencebetween the cohorts was to be seen for the contributions ofIgG1 and IgG4
33 Analysis of IgM C1q and C3c Associated to Anti-1205732GP1-IgG Next we analysed potential 1205732GP1-binding proteins inthe sera of healthy children and adults with APS The signalsfor C1q were low and did not differ significantly between thecohorts
However the ELISAs detecting C3c bound to anti-1205732GP1and anti-1205732GP1-IgM showed highly significant differencesbetween healthy children and adults with APSWhereas C3c-anti-1205732GP1 adducts were increased in healthy children (119901 lt00001 Figure 3(a)) anti-1205732GP1-IgM was decreased (119901 lt0001 Figure 3(b))
Journal of Immunology Research 7
Table 4 Number (frequency) of seropositive samples of the anti-1205732GP1-IgG subclasses
Number of seropositive samples (gtmOD[NHD] + 2 times SD[NHD])IgG1
(gt29mOD)IgG2
(gt79mOD)IgG3
(gt85mOD)IgG4
(gt319mOD)NHD (119899 = 17) 0 (0) 0 (0) 1 (6) 1 (6)Children (119899 = 13) 13 (100) 2 (15) 8 (62) 2 (15)PAPS (119899 = 12) 6 (50) 5 (42) 0 (0) 1 (8)SAPS (119899 = 6) 1 (17) 2 (33) 0 (0) 1 (17)APS (119899 = 18) 7 (39) 7 (39) 0 (0) 2 (11)OD = optical density 450 nm PAPS = primary antiphospholipid syndrome SAPS = secondary antiphospholipid syndrome APS = PAPS and SAPS NHD =normal healthy donorsThe cut-off for seropositivity was calculated from the mean OD of the corresponding anti-1205732GP1-IgG subclass of the NHD cohort andits standard deviation (SD) according to the formula mean OD[NHD] + 2 times SD[NHD] The respective cut-off OD values are indicated in parentheses below theanti-1205732GP1-IgG subclasses
Table 5 Cohorts studied with the lectin ELISA
Cohort Abbreviation Number of samplesPatients with primary APS PAPS 12Patients with APS and SLE as underlying disease SAPS 10Patients with SLE without symptoms of APS harbouring circulating aPL SLE + aPL 15Asymptomatic carriers of aPL aaPL 6Healthy children with circulating anti-1205732GP1 mdash 16APS = antiphospholipid syndrome aPL = antiphospholipid antibodies SLE = systemic lupus erythematosus
34 Analysis of the Sialylation of Anti-1205732GP1 We analysedthe glycosylation of anti-1205732GP1-IgG focusing on its sialy-lation since highly sialylated Fc-parts of IgG are known tocause rather anti-inflammatory responses [30] Wemeasuredthe binding of the sialic acid specific lectin SNA to the anti-1205732GP1-IgG As shown in Table 5 we employed five cohorts ofindividuals 12 patientswith PAPS 10 patientswith SAPS (SLEas primary disease) 15 patients with SLE who did not showclinical sign of APS but with aPL in their sera (SLE + aPL)6 asymptomatic adult carriers of aPL (aaPL) and 16 healthychildren harbouring aPL
Analysing the SNAIgG-ratios (Figure 4) we identifiedthe highest sialylation level in the sera of the healthy childrenThe significances were rather high and decreased from PAPS(119901 lt 00001) to SAPS (119901 lt 00001) to SLE + aPL (119901 lt00001) and to aaPL (119901 lt 0001)
4 Discussion
We analysed two distinct sets of sera One was used toinvestigate anti-1205732GP1-IgG its subclasses and its associatedproteins C1q and C3c For the analysis of the sialylation ofanti-1205732GP1-IgG we had to collect fresh sera from patientswith APS and the healthy children since the stability of theantibody-glycosylation is unknown In order to avoid itspossible decay by processes like enzymatic degradation thisperiod has been kept as short as possible and was followed byimmediate investigation of the glycosylation
The antiphospholipid syndrome is characterized by clin-ical manifestations such as arterial and venous thrombosisor pregnancy-related complications and the detection of spe-cific autoantibodies (lupus anticoagulant anti-1205732GP1- and
anticardiolipin-autoantibodies) [1] In particular anti-1205732GP1sare considered responsible for provoking the pathologicalcomplications [3]We firstmeasured the total amount of anti-1205732GP1-IgG in the sera of 16 healthy children without clinicalmanifestations of APS and 46 adult patients with APS toidentify individual cohorts seropositive for anti-1205732GP1 17adult normal healthy donors served as negative control
The samples of the patientswithAPS showedhigher levelsof anti-1205732GP1-IgG than those of NHD We also observeda higher concentration of anti-1205732GP1-IgG in the sera ofhealthy children compared to NHD This is due to the factthat the detection of these antibodies has been the inclusioncriterion for the healthy children examined in this studyInterestingly the children showed even higher OD values forthe putatively pathogenic anti-1205732GP1-IgG than the patientswith APS This finding was surprising since the childrenwere clinically healthy and did not show any signs of APSsuch as thrombosis or thrombocytopenia complications inwhich the existence of aPL especially anti-1205732GP1 is maderesponsible for [7 33]
High levels of circulating aPL in 61 healthy children whencompared to healthy adults have already been described byAvcin and colleagues [18] The anti-1205732GP1-IgG was report-edly higher in preschool children with a mean age of 5years than in adolescents with a mean age of 135 years[18] The strongly elevated circulating anti-1205732GP1-IgG in ourcohort of healthy children may be explained by the fact thatthey had an even lower age (mean age was nine months)One possible explanation for this result is the inductionof anti-1205732GP1 by viral and bacterial infections [10 11] ornutritional exposure to 1205732GP1 [17] As the immune systemin children is still to evolve they are more susceptible to
8 Journal of Immunology Research
0
05
10
15
20
25SN
AIg
G
ChildrenPAPS SAPS aaPLSLE + aPL
lowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowast
Figure 4 SNAanti-1205732GP1-IgG ratio in PAPS SAPS SLE + aPLaaPL and children Healthy children show a significantly higherSNAanti-1205732GP1-IgG ratio than PAPS SAPS SLE + aPL and aaPL(lowastlowast119901 lt 00001 lowastlowastlowast119901 lt 000001) This suggests higher sialylation ofthe oligosaccharides attached to Fc fragments of the childrenrsquos anti-1205732GP1No significant differenceswere foundbetween the remainingcohorts Horizontal lines indicate the mean SNAIgG ratio of thecohort SNA = Sambucus nigra agglutinin APS = antiphospholipidsyndrome PAPS = patients with primary APS SAPS = patients withAPS and SLE as underlying disease SLE + aaPL = patients withSLE without symptoms of APS harbouring circulating aPL aaPL =asymptomatic carriers of aPL aPL = antiphospholipid antibodiesSLE = systemic lupus erythematosus
infections than adults and their immune system has to copewith a plethora of viral and bacterial pathogens for the firsttime in their life This scenario is prone to result in thestimulation of autoantibody-production Molecular mimicryof certain pathogens can lead to the production of antibodieswhich then cross-react with 1205732GP1 [7 12ndash15] When infantsdiversify their dietary habits and start to consume productslike milk and meat they can get into contact with nutritional1205732GP1 [19] This is a further condition capable triggeringthe production of anti-1205732GP1 Exposure to pathogen-derivedand dietary 1205732GP1 could explain the high level of circulatinganti-1205732GP1-IgG in healthy childrenThis low affine reactivityis consecutively lost during the maturation of the immunesystem and replaced by the emergence in adults of high affineantibodies that more specifically recognize pathogens Apartfrom possible origins of these autoantibodies it is obviousthat they do not provoke any of the hallmark symptoms ofAPS In contrast to their adult counterparts anti-1205732GP1-IgGscirculating in the plasma of healthy children are obviouslynot pathogenic So it seems decent to hypothesize thatanti-1205732GP1 autoantibodies in healthy children biochemicallydiffer from those in adults
To test this hypothesis we examined the anti-1205732GP1-IgG subclass distribution in the seropositive sera of the anti-1205732GP1-IgG ELISA The healthy children showed higher ODvalues for the IgG1- and IgG3-subclasses than patients withAPS and NHD This was consistent with the number ofseropositive samples In the anti-1205732GP1-IgG1-subclass every
tested serum of the healthy children surpassed the thresholdset for seropositivity (gtmean OD
[NHD] + 2 times SD[NHD]) and
8 samples (62) did so in the IgG3-subclass In the seraof patients with APS just 7 (39) and no samples wereseropositive for IgG1 or IgG3 In the sera of patients with APScompared to NHD we observed higher OD levels of anti-1205732GP1 in all subclasses except for IgG4This result is consis-tent with the findings of Guerin et al who reported elevatedlevels in all anti-1205732GP1-IgG subclasses even including IgG4in patients with APS compared to a control group of healthyadults [23]
It might be that the elevated absolute OD values of theanti-1205732GP1-IgG subclasses are the logical consequence ofthe generally higher level of anti-1205732GP1-IgG in the patientswith APS and healthy children who were selected for furthertesting Therefore we also analysed the subclass distributionof this antibody We first analysed the anti-1205732GP1-IgG sub-class distribution within the individual cohorts and observedthat in the healthy children the IgG3 antibody was mostabundant with a contribution of 40 to the total reactivityThis is consistent with the finding that all sera of the healthychildren were seropositive in this subclass and that theiranti-1205732GP1-IgG3 OD values were the highest of all cohortsThis indicates a predominance of this subclass in the healthychildren The remaining anti-1205732GP1-IgG subclasses in thesera of the children ranged from 15 (IgG1) over 20 (IgG2)to 25 (IgG4) but the differences between their proportionswere not statistically significant In contrast in the sera ofthe patients with APS the anti-1205732GP1-IgG2 subclass had thehighest contribution with 38
The finding of an elevated proportion of the IgG2 subclassin the patients with APS confirms former published datawhich also describe a skewing towards anti-1205732GP1-IgG2 inAPS [19 23 26 27] The authors detected an even highercontribution of anti-1205732GP1-IgG2 (702 [26] and 87 [27]resp) In contrast to our results the IgG1 subclass wasmore prominent in the published studies described aboveImportantly in the cohort of the healthy infants IgG2 wasrelatively lowTherefore onemay argue that anti-1205732GP1-IgG2is not just a marker of the disease but is also involved inthe pathogenic action of the aPL autoantibodies When wecompared the anti-1205732GP1-IgG subclass distribution betweenthe three cohorts we noticed a significant difference in thedistribution concerning the children on one hand and NHDand patients with APS on the other Healthy children showeda higher proportion of anti-1205732GP1-IgG3 and a lower one ofanti-1205732GP1-IgG2 In NHD and patients with APS it was theother way round IgG1 and IgG4 did not significantly differbetween healthy children and patients with APS orNHDThepredominance of anti-1205732GP1-IgG3 in the healthy children isa hint to an immune response elicited by a proteinaceousantigen [26 27 34] This supports the hypothesis thatthe harmless anti-1205732GP1 in children are infection-inducedwith pathogen-derived proteins serving as trigger for theirproduction However it is still elusive if the IgG3-dominatedanti-1205732GP1 response found in the sera of healthy childrencontributes to its low pathogenic potential Alternativelyit can just be ascribed to the proceeding evolution of theinfantrsquos immune system typically associated with an elevated
Journal of Immunology Research 9
proportion of IgG3 antibodies [34 35] During childhoodthe proportion of IgG3 continuously decreases to adult rangesusually reached at the age of 10ndash12 years However the aboveaverage elevated proportion of the anti-1205732GP1-IgG3 (40) inthe sera of the healthy childrenmay thus not just be due to theinfant immune system In particular when bearing in mindthat normal adult levels of the IgG3 subclass are just around6 [34 36]
IgG3 is the IgG-subclass exerting the most effectiveactivation of the complement cascade through the classicalpathway [34] Since complement activation is associated withthe pathogenesis of APS [21 32 37] the high proportionof complement-activating anti-1205732GP1-IgG3 in the childrenseems to contradict a possible protective effect against APS ofthese kinds of antibodies However the role of complement inthe etiopathogenesis of autoimmune diseases is manifold asthe complement components C1q C2 and C4 contribute tothe clearance of postapoptotic secondary necrotic cells andtheir deficiency is therefore a risk factor for autoimmunityand autoinflammation [38] In a similar manner studiesin mouse in vivo models of various autoimmune diseasesdemonstrated that IgG mediated tissue inflammation wasblocked in mice deficient in activating Fc120574Rs although thecomplement component C3 was still abundantly depositedin the tissue [39ndash41] Thus the contribution of complementdeposits in tissue with respect to tissue inflammation remainsto be established
To obtain new insights into the involvement of thecomplement system in the pathogenesis of APS we analysedthe C1q- and C3c-binding to anti-1205732GP1 We observed thatanti-1205732GP1 in the sera of the healthy children and in patientswith APS similarly bound C1q the first molecule of theclassical pathway of complement activation [42] Performinganti-1205732GP1-IgM-ELISAs we detected significantly higherIgM values in the patients with APS One would expecta higher C1q-binding of the IgM positive sera since thisimmunoglobulin binds and activates C1q more strongly thanIgG However the children mainly harbour IgG3 autoanti-bodies the most potent subclass for the activation of theclassical complement pathway [34]This may compensate forthe lower IgM level in their sera and thus be responsiblefor the similar C1q-binding of both cohorts This suggeststhat preconditions for complement activation via the classicalpathway are comparable in both cohorts We conclude thatclassical complement activation has a minor impact on thedifferential pathogenesis of aPL in healthy children andpatients with APS
What then is the culprit that induces their pathogenicpotential
Nextwe quantified the contribution ofC3c to the immunecomplexes bound to 1205732GP1 C3c is a domain in the C3bmolecule and one of its cleavage products created aftercleavage by factor I of C3b [42 43] C3c not just representsa marker of complement activation but also serves as anopsonin improving the clearance of bound targets Weobserved significantly more C3c bound to anti-1205732GP1 inthe sera of the healthy children than in those of patientswith APS This reflects a better opsonization by C3c ofthe potentially harmful anti-1205732GP1 immune complexes
anti-1205732GP1-opsonized cells or cellular fragments in thesera of the healthy children Consequently the anti-1205732GP1complexes can be cleared more efficiently by the powerfulcomplement system of the healthy children In patients withchronic inflammatory autoimmune diseases complement isusually low which may result from null alleles [44] or fromcomplement consumption [38]
But why do these processes not cause inflammation andaberrant coagulation in the childrenApart fromanti-1205732GP1-IgG subclass distribution and distinct ability to activate thecomplement system it is likely that other properties of theanti-1205732GP1 in the sera of children contribute to their lowpathogenicity We hypothesized that an increased sialylationof the anti-1205732GP1-IgG in healthy individuals renders themharmless for their hosts IgG is usually N-glycosylated at theCH2-domain of its Fc-part This N-glycan crucially modifiesthe effector functions the IgG molecule is endowed with[45ndash47] This biantennary core-fucosylated oligosaccharidecarries a variable amount of sugar monomers attached toboth arms [28 48] IgG antibodies terminating with sialicacids on one or both antennae have been shown to behaveanti-inflammatorily after infusion of IVIG [29 31]The signalexerted by the terminal sialic acid is dominant since thefraction of sialylated IgG in healthy plasma is just 4 to 14[28 46 49]
In mice these effects are mainly ascribed to decreasedbinding of sialylated IgG to activating Fc120574-receptors andan upregulation of the inhibitory Fc120574RIIB [29 31] Weobserved that anti-1205732GP1-IgG isolated from sera of healthychildren was significantly more sialylated than that fromsymptomatic patients with APS One may predict that theseautoantibodies in complex with 1205732GP1 show low affinity foractivating Fc120574Rs and therefore do not provoke proinflam-matory and procoagulant effector functions Fc120574RIIA is anexample for an activating receptor It is located on a plethoraof cells On the surfaces of platelets it mediates activationby immune complexes The activation is accompanied byplatelet aggregation and the release of mediators such asthe prothromboticThromboxaneA2 [50] Consecutively thiscascade can result in thrombus formation [51 52] Sialylatedanti-1205732GP1-IgG is unable to bind to and to activate plateletsvia Fc120574RIIA
Further mechanisms involved in thrombus formation inAPS comprise the interaction of anti-1205732GP1 with variouspattern recognition receptors [53] and its interference withregulatory functions ofmolecules involved in the coagulationprocess (protein C prothrombin or tissue factor [52 54])Direct binding to 1205732GP1 on endothelial cells (EC) shiftstheir phenotype towards coagulation and inflammation [54]Considering that Fc120574R-binding is involved in many of thesemechanisms it seems likely that a high sialylation of anti-1205732GP1-IgG leads to a low pathogenicity of these autoan-tibodies and does not cause APS Therefore we concludethat the higher sialylation of anti-1205732GP1-IgG in the seraof the healthy children plays a major role in protectingthem from typical complications of APS This hypothesis isfurther supported by the observation that antiphospholipidantibodies from sera of asymptomatic adult carriers aremore sialylated than those from patients with APS A high
10 Journal of Immunology Research
sialylation status of antiphospholipid antibodies seems toprevent the development of symptoms of APS also in theseindividuals
We conclude that a plethora of factors contributes tothe low pathogenicity of anti-1205732GP1-IgG in the sera ofhealthy children and asymptomatic adults We observed adistinct spectrum of IgG-subclasses compared to patientswith APS and a better binding of C3c indicating a moreefficient clearance of potentially harmful anti-1205732GP1 Thehypersialylation of the anti-1205732GP1-IgG is prone to amelioratetheir inflammatory and procoagulatory effects in healthycarriers whereas hyposialylation and inflammationmutuallyprecipitate the antiphospholipid syndrome
Disclosure
Thepresent workwas performed in fulfillment of the require-ments for obtaining the degree ldquoDr medrdquo for ChristophFickentscher
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Laura Andreoli andMartin Herrmann contributed equally tosenior authorship
Acknowledgments
This project was supported by the German Research Foun-dation (training research programme of the collaborativeresearch center 643) by the Emerging Fields Initiative (EFI)of the FAU Erlangen-Nuremberg and by the K und RWucherpfennig-Stiftung
References
[1] S Miyakis M D Lockshin T Atsumi et al ldquoInternationalconsensus statement on an update of the classification criteriafor definite antiphospholipid syndrome (APS)rdquo Journal ofThrombosis and Haemostasis vol 4 no 2 pp 295ndash306 2006
[2] W A Wilson A E Gharavi T Koike et al ldquoInternationalconsensus statement on preliminary classification criteria fordefinite antiphospholipid syndrome report of an internationalworkshoprdquo Arthritis and Rheumatism vol 42 no 7 pp 1309ndash1311 1999
[3] P G de Groot and R T Urbanus ldquoThe significance of autoan-tibodies against beta2-glycoprotein Irdquo Blood vol 120 no 2 pp266ndash274 2012
[4] C Nalli S Piantoni L Andreoli M Motta and A TincanildquoAntiphospholipid syndrome and antiphospholipid antibodiesin children the two sides of the coinrdquo Israel Medical AssociationJournal vol 14 no 5 pp 310ndash312 2012
[5] I Schousboe ldquo1205732-glycoprotein I a plasma inhibitor of thecontact activation of the intrinsic blood coagulation pathwayrdquoBlood vol 66 no 5 pp 1086ndash1091 1985
[6] J George M Blank Y Levy et al ldquoDifferential effects of anti-1205732-glycoprotein I antibodies on endothelial cells and on themanifestations of experimental antiphospholipid syndromerdquoCirculation vol 97 no 9 pp 900ndash906 1998
[7] R Willis and S S Pierangeli ldquoAnti-1205732-glycoprotein I antibod-iesrdquo Annals of the New York Academy of Sciences vol 1285 no1 pp 44ndash58 2013
[8] M J Fischer J Rauch and J S Levine ldquoThe antiphospholipidsyndromerdquo Seminars in Nephrology vol 27 no 1 pp 35ndash462007
[9] H Amital M Govoni R Maya et al ldquoRole of infectious agentsin systemic rheumatic diseasesrdquo Clinical and ExperimentalRheumatology vol 26 no 1 supplement 48 pp S27ndashS32 2008
[10] T Avcin ldquoAntiphospholipid syndrome in childrenrdquo CurrentOpinion in Rheumatology vol 20 no 5 pp 595ndash600 2008
[11] M Garcıa-Carrasco C Galarza-Maldonado C Mendoza-Pinto R O Escarcega and R Cervera ldquoInfections and theantiphospholipid syndromerdquo Clinical Reviews in Allergy ampImmunology vol 36 no 2-3 pp 104ndash108 2009
[12] P Cruz-Tapias M Blank J-M Anaya and Y ShoenfeldldquoInfections and vaccines in the etiology of antiphospholipidsyndromerdquo Current Opinion in Rheumatology vol 24 no 4 pp389ndash393 2012
[13] P von Landenberg Y Doring S Modrow and K J LacknerldquoAre antiphospholipid antibodies an essential requirement foran effective immune response to infectionsrdquoAnnals of the NewYork Academy of Sciences vol 1108 pp 578ndash583 2007
[14] M Blank I Krause M Fridkin et al ldquoBacterial induction ofautoantibodies to 1205732-glycoprotein-I accounts for the infectiousetiology of antiphospholipid syndromerdquo Journal of ClinicalInvestigation vol 109 no 6 pp 797ndash804 2002
[15] Y Shoenfeld M Blank R Cervera J Font E Raschi and P-LMeroni ldquoInfectious origin of the antiphospholipid syndromerdquoAnnals of the Rheumatic Diseases vol 65 no 1 pp 2ndash6 2006
[16] A E Gharavi S S Pierangeli R G Espinola X Liu MColden-Stanfield and E N Harris ldquoAntiphospholipid antibod-ies induced in mice by immunization with a cytomegalovirus-derived peptide cause thrombosis and activation of endothelialcells in vivordquo Arthritis and Rheumatism vol 46 no 2 pp 545ndash552 2002
[17] M Motta G Chirico C B Rebaioli et al ldquoAnticardiolipin andanti-beta2 glycoprotein I antibodies in infants born to motherswith antiphospholipid antibody-positive autoimmune diseasea follow-up studyrdquoAmerican Journal of Perinatology vol 23 no4 pp 247ndash251 2006
[18] T Avcin A Ambrozic M Kuhar T Kveder and B RozmanldquoAnticardiolipin and anti-1205732 glycoprotein I antibodies in seraof 61 apparently healthy children at regular preventive visitsrdquoRheumatology vol 40 no 5 pp 565ndash573 2001
[19] A Ambrozic ldquoAnti-beta2-glycoprotein I antibodies in children
with atopic dermatitisrdquo International Immunology vol 14 no 7pp 823ndash830
[20] J A Giron-Gonzalez E Garcıa Del Rıo C Rodrıguez JRodrıguez-Martorell and A Serrano ldquoAntiphospholipid syn-drome and asymptomatic carriers of antiphospholipid anti-body prospective analysis of 404 individualsrdquo Journal ofRheumatology vol 31 no 8 pp 1560ndash1567 2004
[21] K A Breen P Seed K Parmar GWMoore S E Stuart-Smithand B J Hunt ldquoComplement activation in patients with isolatedantiphospholipid antibodies or primary antiphospholipid syn-dromerdquo Thrombosis and Haemostasis vol 107 no 3 pp 423ndash429 2012
Journal of Immunology Research 11
[22] F Fischetti P Durigutto V Pellis et al ldquoThrombus formationinduced by antibodies to 1205732-glycoprotein I is complementdependent and requires a priming factorrdquo Blood vol 106 no7 pp 2340ndash2346 2005
[23] J Guerin E Casey C Feighery and J Jackson ldquoAnti-Beta 2-glycoprotein I antibody isotype and IgG subclass in antiphos-pholipid syndrome patientsrdquo Autoimmunity vol 31 no 2 pp109ndash116 1999
[24] L Andreoli C Nalli M Motta et al ldquoAnti-1205732-glycoprotein I
IgG antibodies from 1-year-old healthy children born to moth-ers with systemic autoimmune diseases preferentially targetdomain 45 might it be the reason for their lsquoinnocentrsquo profilerdquoAnnals of the Rheumatic Diseases vol 70 no 2 pp 380ndash3832011
[25] B de Laat R H W M Derksen M van Lummel M T T Pen-nings and P G deGroot ldquoPathogenic anti-beta2-glycoprotein Iantibodies recognize domain I of beta2-glycoprotein I only aftera conformational changerdquo Blood vol 107 no 5 pp 1916ndash19242006
[26] M Samarkos K A Davies C Gordon M J Walport and SLoizou ldquoIgG subclass distribution of antibodies against 1205732-GP1and cardiolipin in patients with systemic lupus erythematosusand primary antiphospholipid syndrome and their clinicalassociationsrdquo Rheumatology vol 40 no 9 pp 1026ndash1032 2001
[27] J Arvieux B Roussel D Ponard and M G Colomb ldquoIgG2subclass restriction of anti-1205732 glycoprotein 1 antibodies inautoimmune patientsrdquo Clinical and Experimental Immunologyvol 95 no 2 pp 310ndash315 1994
[28] J N Arnold M R Wormald R B Sim P M Rudd and R ADwek ldquoThe impact of glycosylation on the biological functionand structure of human immunoglobulinsrdquo Annual Review ofImmunology vol 25 pp 21ndash50 2007
[29] Y Kaneko F Nimmerjahn and J V Ravetch ldquoAnti-inflamma-tory activity of immunoglobulin G resulting from Fc sialyla-tionrdquo Science vol 313 no 5787 pp 670ndash673 2006
[30] RM Anthony and F Nimmerjahn ldquoThe role of differential IgGglycosylation in the interaction of antibodies with FcgammaRsin vivordquo Current Opinion in Organ Transplantation vol 16 no1 pp 7ndash14 2011
[31] R M Anthony and J V Ravetch ldquoA novel role for the IgGFc glycan the anti-inflammatory activity of sialylated IgG FcsrdquoJournal of Clinical Immunology vol 30 no 1 pp S9ndashS14 2010
[32] K Oku T Atsumi M Bohgaki et al ldquoComplement activationin patients with primary antiphospholipid syndromerdquoAnnals ofthe Rheumatic Diseases vol 68 no 6 pp 1030ndash1035 2009
[33] H P McNeil R J Simpson C N Chesterman and S AKrilis ldquoAnti-phospholipid antibodies are directed against acomplex antigen that includes a lipid-binding inhibitor of coag-ulation 1205732-Glycoprotein I (apolipoprotein H)rdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 87 no 11 pp 4120ndash4124 1990
[34] R Jefferis and D S Kumararatne ldquoSelective IgG subclassdeficiency quantification and clinical relevancerdquo Clinical andExperimental Immunology vol 81 no 3 pp 357ndash367 1990
[35] P H Schur F Rosen and M E Norman ldquoImmunoglobulinsubclasses in normal childrenrdquo Pediatric Research vol 13 no3 pp 181ndash183 1979
[36] M A H French and G Harrison ldquoSerum IgG subclass concen-trations in healthy adults a study using monoclonal antiserardquoClinical and Experimental Immunology vol 56 no 2 pp 473ndash475 1984
[37] M Odorczuk L B Keil and V A DeBari ldquoBinding of com-plement component C1q to anti-beta2 glycoprotein I antibodiesfrom patients with antiphospholipid syndromerdquo InflammationResearch vol 48 no 10 pp 524ndash526 1999
[38] L Truedsson A A Bengtsson and G Sturfelt ldquoComplementdeficiencies and systemic lupus erythematosusrdquoAutoimmunityvol 40 no 8 pp 560ndash566 2007
[39] S Aschermann C H K Lehmann S Mihai G Schett DDudziak and F Nimmerjahn ldquoB cells are critical for autoim-mune pathology in Scurfy micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 110 no47 pp 19042ndash19047 2013
[40] Y Kaneko F Nimmerjahn M P Madaio and J V RavetchldquoPathology and protection in nephrotoxic nephritis is deter-mined by selective engagement of specific Fc receptorsrdquo TheJournal of Experimental Medicine vol 203 no 3 pp 789ndash7972006
[41] D Sylvestre R Clynes M Ma H Warren M C Carrolland J V Ravetch ldquoImmunoglobulin G-mediated inflammatoryresponses develop normally in complement-deficient micerdquoJournal of ExperimentalMedicine vol 184 no 6 pp 2385ndash23921996
[42] G E Ritchie B E Moffatt R B Sim B P Morgan R A Dwekand P M Rudd ldquoGlycosylation and the complement systemrdquoChemical Reviews vol 102 no 2 pp 305ndash320 2002
[43] Y Palarasah K Skjodt J Brandt et al ldquoGeneration of aC3c specific monoclonal antibody and assessment of C3c as aputative inflammatorymarker derived from complement factorC3rdquo Journal of ImmunologicalMethods vol 362 no 1-2 pp 142ndash150 2010
[44] D G Doherty J A Underhill P T Donaldson et al ldquoPoly-morphism in the human complement C4 genes and geneticsusceptibility to autoimmune hepatitisrdquo Autoimmunity vol 18no 4 pp 243ndash249 1994
[45] M-H Tao and S L Morrison ldquoStudies of aglycosylatedchimeric mouse-human IgG Role of carbohydrate in thestructure and effector functions mediated by the human IgGconstant regionrdquoThe Journal of Immunology vol 143 no 8 pp2595ndash2601 1989
[46] R Jefferis ldquoGlycosylation of recombinant antibody therapeu-ticsrdquo Biotechnology Progress vol 21 no 1 pp 11ndash16 2005
[47] Y Mimura S Church R Ghirlando et al ldquoThe influence ofglycosylation on the thermal stability and effector functionexpression of human IgG1-Fc properties of a series of truncatedglycoformsrdquoMolecular Immunology vol 37 no 12-13 pp 697ndash706 2001
[48] C Huhn M H J Selman L R Ruhaak A M Deelder and MWuhrer ldquoIgG glycosylation analysisrdquo Proteomics vol 9 no 4pp 882ndash913 2009
[49] J Lund N Takahashi J D Pound M Goodall and R JefferisldquoMultiple interactions of IgG with its core oligosaccharide canmodulate recognition by complement and human Fc gammareceptor I and influence the synthesis of its oligosaccharidechainsrdquo The Journal of Immunology vol 157 no 11 pp 4963ndash4969 1996
[50] R Forastiero M Martinuzzo L O Carreras and J MacloufldquoAnti-1205732 glycoprotein I antibodies and platelet activation inpatients with antiphospholipid antibodies association withincreased excretion of platelet-derived thromboxane urinarymetabolitesrdquoThrombosis and Haemostasis vol 79 no 1 pp 42ndash45 1998
12 Journal of Immunology Research
[51] J-C Reverter and D Tassies ldquoMechanism of thrombosis inantiphospholipid syndrome binding to plateletsrdquo in HughesSyndrome M A Khamashta Ed pp 403ndash414 Springer Lon-don UK 2006
[52] G Espinosa R Cervera J Font and Y Shoenfeld ldquoAntiphos-pholipid syndrome pathogenic mechanismsrdquo AutoimmunityReviews vol 2 no 2 pp 86ndash93 2003
[53] T Shi B Giannakopoulos X Yan et al ldquoAnti-1205732-glycoprotein
I antibodies in complex with 1205732-glycoprotein I can activate
platelets in a dysregulated manner via glycoprotein Ib-IX-VrdquoArthritis amp Rheumatism vol 54 no 8 pp 2558ndash2567 2006
[54] P LMeroni E RaschiMCamera et al ldquoEndothelial activationby aPL a potential pathogenetic mechanism for the clinicalmanifestations of the syndromerdquo Journal of Autoimmunity vol15 no 2 pp 237ndash240 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
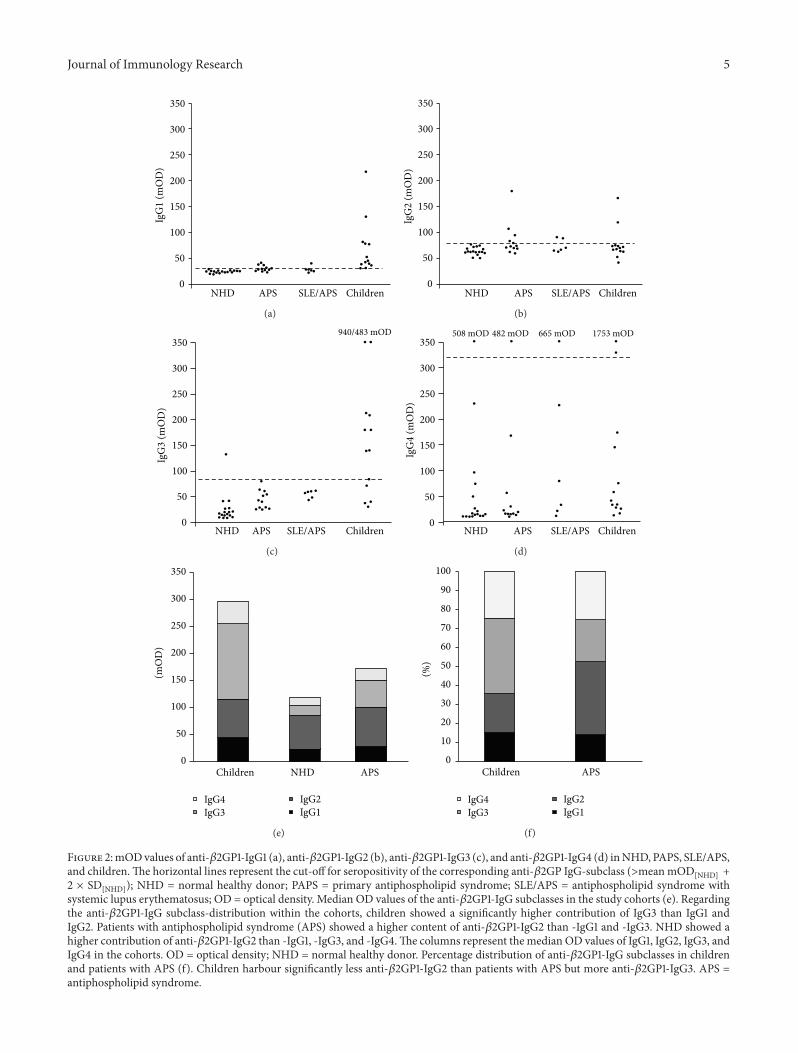
Journal of Immunology Research 5
0
50
100
150
200
250
300
350
NHD APS SLEAPS Children
IgG
1 (m
OD
)
(a)
NHD APS SLEAPS Children0
50
100
150
200
250
300
350
IgG
2 (m
OD
)
(b)
NHD APS SLEAPS Children
940483 mOD
0
50
100
150
200
250
300
350
IgG
3 (m
OD
)
(c)
NHD APS SLEAPS Children
508 mOD 482 mOD 665 mOD 1753 mOD
0
50
100
150
200
250
300
350
IgG
4 (m
OD
)
(d)
0
50
100
150
200
250
300
350
(mO
D)
Children NHD APS
IgG4IgG3
IgG2IgG1
(e)
0
10
20
30
40
50
60
70
80
90
100
()
APSChildren
IgG4IgG3
IgG2IgG1
(f)
Figure 2mODvalues of anti-1205732GP1-IgG1 (a) anti-1205732GP1-IgG2 (b) anti-1205732GP1-IgG3 (c) and anti-1205732GP1-IgG4 (d) inNHD PAPS SLEAPSand childrenThe horizontal lines represent the cut-off for seropositivity of the corresponding anti-1205732GP IgG-subclass (gtmeanmOD
[NHD] +2 times SD
[NHD]) NHD = normal healthy donor PAPS = primary antiphospholipid syndrome SLEAPS = antiphospholipid syndrome withsystemic lupus erythematosus OD = optical density Median OD values of the anti-1205732GP1-IgG subclasses in the study cohorts (e) Regardingthe anti-1205732GP1-IgG subclass-distribution within the cohorts children showed a significantly higher contribution of IgG3 than IgG1 andIgG2 Patients with antiphospholipid syndrome (APS) showed a higher content of anti-1205732GP1-IgG2 than -IgG1 and -IgG3 NHD showed ahigher contribution of anti-1205732GP1-IgG2 than -IgG1 -IgG3 and -IgG4The columns represent the median OD values of IgG1 IgG2 IgG3 andIgG4 in the cohorts OD = optical density NHD = normal healthy donor Percentage distribution of anti-1205732GP1-IgG subclasses in childrenand patients with APS (f) Children harbour significantly less anti-1205732GP1-IgG2 than patients with APS but more anti-1205732GP1-IgG3 APS =antiphospholipid syndrome
6 Journal of Immunology Research
0
50
100
150
200
250
300
350
400
Children APS
C3c (
mO
D)
(a)
0
200
400
600
800
1000
1200
Children APS
IgM
(mO
D)
(b)
Figure 3 OD values of C3c (a) bound to anti-1205732GP1 and OD values of anti-1205732GP1-IgM (b) in sera of patients with APS and healthy childrenAnti-1205732GP1 of healthy children displayed significantly more bound C3c than that of patients with APS (119901 = 000007) This suggests a moreefficient clearance of 1205732GP1-immune complexes in the healthy children Anti-1205732GP1-IgM OD values of patients with APS were significantlyhigher than those of healthy children Horizontal lines represent the median OD value of the cohort APS = antiphospholipid syndrome OD= optical density
Table 3 (a) OD values of the anti-1205732GP1-IgG subclasses in theinvestigated cohorts (b) Comparison (119901 values) of the OD valuesof the anti-1205732GP1-IgG subclasses
(a)
IgG1 [mOD] IgG2 [mOD] IgG3 [mOD] IgG4 [mOD]NHD 24 64 27 67Children 69 77 212 210PAPS 30 85 45 72SAPS 28 74 55 173APS 29 81 48 106
(b)
IgG1 IgG2 IgG3 IgG4Children versus NHD lt00001 ns lt00001 nsChildren versus PAPS lt00001 ns lt0001 nsChildren versus SAPS lt0001 ns lt0001 nsAPS versus NHD ns ns ns nsPAPS versus SAPS ns ns ns nsOD = optical density 450 nm PAPS = primary antiphospholipid syndromeSAPS = secondary antiphospholipid syndrome APS = PAPS and SAPSNHD = normal healthy donors ns = not significant
plus two times the SD of the considered subclass were definedas seropositive (Supplementary Figure 1) Applying this cut-off we observed that the healthy children had high levels ofthe IgG1 and IgG3 subclasses with 13 (100) and 8 (62)seropositive samples respectively In contrast only 7 (39)and 0 (0) sera of patients with APS were seropositive forIgG1 and IgG3 respectively APS showed seropositivity ofanti-1205732GP1-IgG2 in 7 cases (39) whereas the cohort ofhealthy children contained just 2 seropositive sera (15) Just
few samples were seropositive for anti-1205732GP1-IgG4 in eachcohort representing a range from 8 (PAPS) to 17 (SAPS)
To determine the subclass distributionwithin the cohortswe calculated the contribution of themeanOD values of eachindividual subclass to the sum
As shown in Figures 2(e) and 2(f) we saw that for thehealthy children IgG3 was the most abundant subclass witha contribution of 40 (IgG3 gt IgG1 (119901 lt 0001) and IgG3 gtIgG2 (119901 lt 001)) In contrast IgG2 dominated the subclassprofile for NHD (46 IgG2 gt IgG1 IgG2 gt IgG3 and IgG2 gtIgG4 (119901 lt 00001)) and patients with APS (38 IgG2 gt IgG1(119901 lt 00001) IgG2 gt IgG3 (119901 = 0001)) The contributionof IgG1 ranged from 12 (SAPS) to 17 (NHD) and thatof IgG4 from 21 (PAPS) to 35 (SAPS) Comparing thecontribution of the individual subclasses between the studycohorts we observed that children showed significantly lessIgG2 than patients with APS and NHD (119901 lt 0001 119901 lt00001) and a higher IgG3 content than NHD and patientswith APS (119901 lt 00001 119901 lt 0001) No significant differencebetween the cohorts was to be seen for the contributions ofIgG1 and IgG4
33 Analysis of IgM C1q and C3c Associated to Anti-1205732GP1-IgG Next we analysed potential 1205732GP1-binding proteins inthe sera of healthy children and adults with APS The signalsfor C1q were low and did not differ significantly between thecohorts
However the ELISAs detecting C3c bound to anti-1205732GP1and anti-1205732GP1-IgM showed highly significant differencesbetween healthy children and adults with APSWhereas C3c-anti-1205732GP1 adducts were increased in healthy children (119901 lt00001 Figure 3(a)) anti-1205732GP1-IgM was decreased (119901 lt0001 Figure 3(b))
Journal of Immunology Research 7
Table 4 Number (frequency) of seropositive samples of the anti-1205732GP1-IgG subclasses
Number of seropositive samples (gtmOD[NHD] + 2 times SD[NHD])IgG1
(gt29mOD)IgG2
(gt79mOD)IgG3
(gt85mOD)IgG4
(gt319mOD)NHD (119899 = 17) 0 (0) 0 (0) 1 (6) 1 (6)Children (119899 = 13) 13 (100) 2 (15) 8 (62) 2 (15)PAPS (119899 = 12) 6 (50) 5 (42) 0 (0) 1 (8)SAPS (119899 = 6) 1 (17) 2 (33) 0 (0) 1 (17)APS (119899 = 18) 7 (39) 7 (39) 0 (0) 2 (11)OD = optical density 450 nm PAPS = primary antiphospholipid syndrome SAPS = secondary antiphospholipid syndrome APS = PAPS and SAPS NHD =normal healthy donorsThe cut-off for seropositivity was calculated from the mean OD of the corresponding anti-1205732GP1-IgG subclass of the NHD cohort andits standard deviation (SD) according to the formula mean OD[NHD] + 2 times SD[NHD] The respective cut-off OD values are indicated in parentheses below theanti-1205732GP1-IgG subclasses
Table 5 Cohorts studied with the lectin ELISA
Cohort Abbreviation Number of samplesPatients with primary APS PAPS 12Patients with APS and SLE as underlying disease SAPS 10Patients with SLE without symptoms of APS harbouring circulating aPL SLE + aPL 15Asymptomatic carriers of aPL aaPL 6Healthy children with circulating anti-1205732GP1 mdash 16APS = antiphospholipid syndrome aPL = antiphospholipid antibodies SLE = systemic lupus erythematosus
34 Analysis of the Sialylation of Anti-1205732GP1 We analysedthe glycosylation of anti-1205732GP1-IgG focusing on its sialy-lation since highly sialylated Fc-parts of IgG are known tocause rather anti-inflammatory responses [30] Wemeasuredthe binding of the sialic acid specific lectin SNA to the anti-1205732GP1-IgG As shown in Table 5 we employed five cohorts ofindividuals 12 patientswith PAPS 10 patientswith SAPS (SLEas primary disease) 15 patients with SLE who did not showclinical sign of APS but with aPL in their sera (SLE + aPL)6 asymptomatic adult carriers of aPL (aaPL) and 16 healthychildren harbouring aPL
Analysing the SNAIgG-ratios (Figure 4) we identifiedthe highest sialylation level in the sera of the healthy childrenThe significances were rather high and decreased from PAPS(119901 lt 00001) to SAPS (119901 lt 00001) to SLE + aPL (119901 lt00001) and to aaPL (119901 lt 0001)
4 Discussion
We analysed two distinct sets of sera One was used toinvestigate anti-1205732GP1-IgG its subclasses and its associatedproteins C1q and C3c For the analysis of the sialylation ofanti-1205732GP1-IgG we had to collect fresh sera from patientswith APS and the healthy children since the stability of theantibody-glycosylation is unknown In order to avoid itspossible decay by processes like enzymatic degradation thisperiod has been kept as short as possible and was followed byimmediate investigation of the glycosylation
The antiphospholipid syndrome is characterized by clin-ical manifestations such as arterial and venous thrombosisor pregnancy-related complications and the detection of spe-cific autoantibodies (lupus anticoagulant anti-1205732GP1- and
anticardiolipin-autoantibodies) [1] In particular anti-1205732GP1sare considered responsible for provoking the pathologicalcomplications [3]We firstmeasured the total amount of anti-1205732GP1-IgG in the sera of 16 healthy children without clinicalmanifestations of APS and 46 adult patients with APS toidentify individual cohorts seropositive for anti-1205732GP1 17adult normal healthy donors served as negative control
The samples of the patientswithAPS showedhigher levelsof anti-1205732GP1-IgG than those of NHD We also observeda higher concentration of anti-1205732GP1-IgG in the sera ofhealthy children compared to NHD This is due to the factthat the detection of these antibodies has been the inclusioncriterion for the healthy children examined in this studyInterestingly the children showed even higher OD values forthe putatively pathogenic anti-1205732GP1-IgG than the patientswith APS This finding was surprising since the childrenwere clinically healthy and did not show any signs of APSsuch as thrombosis or thrombocytopenia complications inwhich the existence of aPL especially anti-1205732GP1 is maderesponsible for [7 33]
High levels of circulating aPL in 61 healthy children whencompared to healthy adults have already been described byAvcin and colleagues [18] The anti-1205732GP1-IgG was report-edly higher in preschool children with a mean age of 5years than in adolescents with a mean age of 135 years[18] The strongly elevated circulating anti-1205732GP1-IgG in ourcohort of healthy children may be explained by the fact thatthey had an even lower age (mean age was nine months)One possible explanation for this result is the inductionof anti-1205732GP1 by viral and bacterial infections [10 11] ornutritional exposure to 1205732GP1 [17] As the immune systemin children is still to evolve they are more susceptible to
8 Journal of Immunology Research
0
05
10
15
20
25SN
AIg
G
ChildrenPAPS SAPS aaPLSLE + aPL
lowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowast
Figure 4 SNAanti-1205732GP1-IgG ratio in PAPS SAPS SLE + aPLaaPL and children Healthy children show a significantly higherSNAanti-1205732GP1-IgG ratio than PAPS SAPS SLE + aPL and aaPL(lowastlowast119901 lt 00001 lowastlowastlowast119901 lt 000001) This suggests higher sialylation ofthe oligosaccharides attached to Fc fragments of the childrenrsquos anti-1205732GP1No significant differenceswere foundbetween the remainingcohorts Horizontal lines indicate the mean SNAIgG ratio of thecohort SNA = Sambucus nigra agglutinin APS = antiphospholipidsyndrome PAPS = patients with primary APS SAPS = patients withAPS and SLE as underlying disease SLE + aaPL = patients withSLE without symptoms of APS harbouring circulating aPL aaPL =asymptomatic carriers of aPL aPL = antiphospholipid antibodiesSLE = systemic lupus erythematosus
infections than adults and their immune system has to copewith a plethora of viral and bacterial pathogens for the firsttime in their life This scenario is prone to result in thestimulation of autoantibody-production Molecular mimicryof certain pathogens can lead to the production of antibodieswhich then cross-react with 1205732GP1 [7 12ndash15] When infantsdiversify their dietary habits and start to consume productslike milk and meat they can get into contact with nutritional1205732GP1 [19] This is a further condition capable triggeringthe production of anti-1205732GP1 Exposure to pathogen-derivedand dietary 1205732GP1 could explain the high level of circulatinganti-1205732GP1-IgG in healthy childrenThis low affine reactivityis consecutively lost during the maturation of the immunesystem and replaced by the emergence in adults of high affineantibodies that more specifically recognize pathogens Apartfrom possible origins of these autoantibodies it is obviousthat they do not provoke any of the hallmark symptoms ofAPS In contrast to their adult counterparts anti-1205732GP1-IgGscirculating in the plasma of healthy children are obviouslynot pathogenic So it seems decent to hypothesize thatanti-1205732GP1 autoantibodies in healthy children biochemicallydiffer from those in adults
To test this hypothesis we examined the anti-1205732GP1-IgG subclass distribution in the seropositive sera of the anti-1205732GP1-IgG ELISA The healthy children showed higher ODvalues for the IgG1- and IgG3-subclasses than patients withAPS and NHD This was consistent with the number ofseropositive samples In the anti-1205732GP1-IgG1-subclass every
tested serum of the healthy children surpassed the thresholdset for seropositivity (gtmean OD
[NHD] + 2 times SD[NHD]) and
8 samples (62) did so in the IgG3-subclass In the seraof patients with APS just 7 (39) and no samples wereseropositive for IgG1 or IgG3 In the sera of patients with APScompared to NHD we observed higher OD levels of anti-1205732GP1 in all subclasses except for IgG4This result is consis-tent with the findings of Guerin et al who reported elevatedlevels in all anti-1205732GP1-IgG subclasses even including IgG4in patients with APS compared to a control group of healthyadults [23]
It might be that the elevated absolute OD values of theanti-1205732GP1-IgG subclasses are the logical consequence ofthe generally higher level of anti-1205732GP1-IgG in the patientswith APS and healthy children who were selected for furthertesting Therefore we also analysed the subclass distributionof this antibody We first analysed the anti-1205732GP1-IgG sub-class distribution within the individual cohorts and observedthat in the healthy children the IgG3 antibody was mostabundant with a contribution of 40 to the total reactivityThis is consistent with the finding that all sera of the healthychildren were seropositive in this subclass and that theiranti-1205732GP1-IgG3 OD values were the highest of all cohortsThis indicates a predominance of this subclass in the healthychildren The remaining anti-1205732GP1-IgG subclasses in thesera of the children ranged from 15 (IgG1) over 20 (IgG2)to 25 (IgG4) but the differences between their proportionswere not statistically significant In contrast in the sera ofthe patients with APS the anti-1205732GP1-IgG2 subclass had thehighest contribution with 38
The finding of an elevated proportion of the IgG2 subclassin the patients with APS confirms former published datawhich also describe a skewing towards anti-1205732GP1-IgG2 inAPS [19 23 26 27] The authors detected an even highercontribution of anti-1205732GP1-IgG2 (702 [26] and 87 [27]resp) In contrast to our results the IgG1 subclass wasmore prominent in the published studies described aboveImportantly in the cohort of the healthy infants IgG2 wasrelatively lowTherefore onemay argue that anti-1205732GP1-IgG2is not just a marker of the disease but is also involved inthe pathogenic action of the aPL autoantibodies When wecompared the anti-1205732GP1-IgG subclass distribution betweenthe three cohorts we noticed a significant difference in thedistribution concerning the children on one hand and NHDand patients with APS on the other Healthy children showeda higher proportion of anti-1205732GP1-IgG3 and a lower one ofanti-1205732GP1-IgG2 In NHD and patients with APS it was theother way round IgG1 and IgG4 did not significantly differbetween healthy children and patients with APS orNHDThepredominance of anti-1205732GP1-IgG3 in the healthy children isa hint to an immune response elicited by a proteinaceousantigen [26 27 34] This supports the hypothesis thatthe harmless anti-1205732GP1 in children are infection-inducedwith pathogen-derived proteins serving as trigger for theirproduction However it is still elusive if the IgG3-dominatedanti-1205732GP1 response found in the sera of healthy childrencontributes to its low pathogenic potential Alternativelyit can just be ascribed to the proceeding evolution of theinfantrsquos immune system typically associated with an elevated
Journal of Immunology Research 9
proportion of IgG3 antibodies [34 35] During childhoodthe proportion of IgG3 continuously decreases to adult rangesusually reached at the age of 10ndash12 years However the aboveaverage elevated proportion of the anti-1205732GP1-IgG3 (40) inthe sera of the healthy childrenmay thus not just be due to theinfant immune system In particular when bearing in mindthat normal adult levels of the IgG3 subclass are just around6 [34 36]
IgG3 is the IgG-subclass exerting the most effectiveactivation of the complement cascade through the classicalpathway [34] Since complement activation is associated withthe pathogenesis of APS [21 32 37] the high proportionof complement-activating anti-1205732GP1-IgG3 in the childrenseems to contradict a possible protective effect against APS ofthese kinds of antibodies However the role of complement inthe etiopathogenesis of autoimmune diseases is manifold asthe complement components C1q C2 and C4 contribute tothe clearance of postapoptotic secondary necrotic cells andtheir deficiency is therefore a risk factor for autoimmunityand autoinflammation [38] In a similar manner studiesin mouse in vivo models of various autoimmune diseasesdemonstrated that IgG mediated tissue inflammation wasblocked in mice deficient in activating Fc120574Rs although thecomplement component C3 was still abundantly depositedin the tissue [39ndash41] Thus the contribution of complementdeposits in tissue with respect to tissue inflammation remainsto be established
To obtain new insights into the involvement of thecomplement system in the pathogenesis of APS we analysedthe C1q- and C3c-binding to anti-1205732GP1 We observed thatanti-1205732GP1 in the sera of the healthy children and in patientswith APS similarly bound C1q the first molecule of theclassical pathway of complement activation [42] Performinganti-1205732GP1-IgM-ELISAs we detected significantly higherIgM values in the patients with APS One would expecta higher C1q-binding of the IgM positive sera since thisimmunoglobulin binds and activates C1q more strongly thanIgG However the children mainly harbour IgG3 autoanti-bodies the most potent subclass for the activation of theclassical complement pathway [34]This may compensate forthe lower IgM level in their sera and thus be responsiblefor the similar C1q-binding of both cohorts This suggeststhat preconditions for complement activation via the classicalpathway are comparable in both cohorts We conclude thatclassical complement activation has a minor impact on thedifferential pathogenesis of aPL in healthy children andpatients with APS
What then is the culprit that induces their pathogenicpotential
Nextwe quantified the contribution ofC3c to the immunecomplexes bound to 1205732GP1 C3c is a domain in the C3bmolecule and one of its cleavage products created aftercleavage by factor I of C3b [42 43] C3c not just representsa marker of complement activation but also serves as anopsonin improving the clearance of bound targets Weobserved significantly more C3c bound to anti-1205732GP1 inthe sera of the healthy children than in those of patientswith APS This reflects a better opsonization by C3c ofthe potentially harmful anti-1205732GP1 immune complexes
anti-1205732GP1-opsonized cells or cellular fragments in thesera of the healthy children Consequently the anti-1205732GP1complexes can be cleared more efficiently by the powerfulcomplement system of the healthy children In patients withchronic inflammatory autoimmune diseases complement isusually low which may result from null alleles [44] or fromcomplement consumption [38]
But why do these processes not cause inflammation andaberrant coagulation in the childrenApart fromanti-1205732GP1-IgG subclass distribution and distinct ability to activate thecomplement system it is likely that other properties of theanti-1205732GP1 in the sera of children contribute to their lowpathogenicity We hypothesized that an increased sialylationof the anti-1205732GP1-IgG in healthy individuals renders themharmless for their hosts IgG is usually N-glycosylated at theCH2-domain of its Fc-part This N-glycan crucially modifiesthe effector functions the IgG molecule is endowed with[45ndash47] This biantennary core-fucosylated oligosaccharidecarries a variable amount of sugar monomers attached toboth arms [28 48] IgG antibodies terminating with sialicacids on one or both antennae have been shown to behaveanti-inflammatorily after infusion of IVIG [29 31]The signalexerted by the terminal sialic acid is dominant since thefraction of sialylated IgG in healthy plasma is just 4 to 14[28 46 49]
In mice these effects are mainly ascribed to decreasedbinding of sialylated IgG to activating Fc120574-receptors andan upregulation of the inhibitory Fc120574RIIB [29 31] Weobserved that anti-1205732GP1-IgG isolated from sera of healthychildren was significantly more sialylated than that fromsymptomatic patients with APS One may predict that theseautoantibodies in complex with 1205732GP1 show low affinity foractivating Fc120574Rs and therefore do not provoke proinflam-matory and procoagulant effector functions Fc120574RIIA is anexample for an activating receptor It is located on a plethoraof cells On the surfaces of platelets it mediates activationby immune complexes The activation is accompanied byplatelet aggregation and the release of mediators such asthe prothromboticThromboxaneA2 [50] Consecutively thiscascade can result in thrombus formation [51 52] Sialylatedanti-1205732GP1-IgG is unable to bind to and to activate plateletsvia Fc120574RIIA
Further mechanisms involved in thrombus formation inAPS comprise the interaction of anti-1205732GP1 with variouspattern recognition receptors [53] and its interference withregulatory functions ofmolecules involved in the coagulationprocess (protein C prothrombin or tissue factor [52 54])Direct binding to 1205732GP1 on endothelial cells (EC) shiftstheir phenotype towards coagulation and inflammation [54]Considering that Fc120574R-binding is involved in many of thesemechanisms it seems likely that a high sialylation of anti-1205732GP1-IgG leads to a low pathogenicity of these autoan-tibodies and does not cause APS Therefore we concludethat the higher sialylation of anti-1205732GP1-IgG in the seraof the healthy children plays a major role in protectingthem from typical complications of APS This hypothesis isfurther supported by the observation that antiphospholipidantibodies from sera of asymptomatic adult carriers aremore sialylated than those from patients with APS A high
10 Journal of Immunology Research
sialylation status of antiphospholipid antibodies seems toprevent the development of symptoms of APS also in theseindividuals
We conclude that a plethora of factors contributes tothe low pathogenicity of anti-1205732GP1-IgG in the sera ofhealthy children and asymptomatic adults We observed adistinct spectrum of IgG-subclasses compared to patientswith APS and a better binding of C3c indicating a moreefficient clearance of potentially harmful anti-1205732GP1 Thehypersialylation of the anti-1205732GP1-IgG is prone to amelioratetheir inflammatory and procoagulatory effects in healthycarriers whereas hyposialylation and inflammationmutuallyprecipitate the antiphospholipid syndrome
Disclosure
Thepresent workwas performed in fulfillment of the require-ments for obtaining the degree ldquoDr medrdquo for ChristophFickentscher
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Laura Andreoli andMartin Herrmann contributed equally tosenior authorship
Acknowledgments
This project was supported by the German Research Foun-dation (training research programme of the collaborativeresearch center 643) by the Emerging Fields Initiative (EFI)of the FAU Erlangen-Nuremberg and by the K und RWucherpfennig-Stiftung
References
[1] S Miyakis M D Lockshin T Atsumi et al ldquoInternationalconsensus statement on an update of the classification criteriafor definite antiphospholipid syndrome (APS)rdquo Journal ofThrombosis and Haemostasis vol 4 no 2 pp 295ndash306 2006
[2] W A Wilson A E Gharavi T Koike et al ldquoInternationalconsensus statement on preliminary classification criteria fordefinite antiphospholipid syndrome report of an internationalworkshoprdquo Arthritis and Rheumatism vol 42 no 7 pp 1309ndash1311 1999
[3] P G de Groot and R T Urbanus ldquoThe significance of autoan-tibodies against beta2-glycoprotein Irdquo Blood vol 120 no 2 pp266ndash274 2012
[4] C Nalli S Piantoni L Andreoli M Motta and A TincanildquoAntiphospholipid syndrome and antiphospholipid antibodiesin children the two sides of the coinrdquo Israel Medical AssociationJournal vol 14 no 5 pp 310ndash312 2012
[5] I Schousboe ldquo1205732-glycoprotein I a plasma inhibitor of thecontact activation of the intrinsic blood coagulation pathwayrdquoBlood vol 66 no 5 pp 1086ndash1091 1985
[6] J George M Blank Y Levy et al ldquoDifferential effects of anti-1205732-glycoprotein I antibodies on endothelial cells and on themanifestations of experimental antiphospholipid syndromerdquoCirculation vol 97 no 9 pp 900ndash906 1998
[7] R Willis and S S Pierangeli ldquoAnti-1205732-glycoprotein I antibod-iesrdquo Annals of the New York Academy of Sciences vol 1285 no1 pp 44ndash58 2013
[8] M J Fischer J Rauch and J S Levine ldquoThe antiphospholipidsyndromerdquo Seminars in Nephrology vol 27 no 1 pp 35ndash462007
[9] H Amital M Govoni R Maya et al ldquoRole of infectious agentsin systemic rheumatic diseasesrdquo Clinical and ExperimentalRheumatology vol 26 no 1 supplement 48 pp S27ndashS32 2008
[10] T Avcin ldquoAntiphospholipid syndrome in childrenrdquo CurrentOpinion in Rheumatology vol 20 no 5 pp 595ndash600 2008
[11] M Garcıa-Carrasco C Galarza-Maldonado C Mendoza-Pinto R O Escarcega and R Cervera ldquoInfections and theantiphospholipid syndromerdquo Clinical Reviews in Allergy ampImmunology vol 36 no 2-3 pp 104ndash108 2009
[12] P Cruz-Tapias M Blank J-M Anaya and Y ShoenfeldldquoInfections and vaccines in the etiology of antiphospholipidsyndromerdquo Current Opinion in Rheumatology vol 24 no 4 pp389ndash393 2012
[13] P von Landenberg Y Doring S Modrow and K J LacknerldquoAre antiphospholipid antibodies an essential requirement foran effective immune response to infectionsrdquoAnnals of the NewYork Academy of Sciences vol 1108 pp 578ndash583 2007
[14] M Blank I Krause M Fridkin et al ldquoBacterial induction ofautoantibodies to 1205732-glycoprotein-I accounts for the infectiousetiology of antiphospholipid syndromerdquo Journal of ClinicalInvestigation vol 109 no 6 pp 797ndash804 2002
[15] Y Shoenfeld M Blank R Cervera J Font E Raschi and P-LMeroni ldquoInfectious origin of the antiphospholipid syndromerdquoAnnals of the Rheumatic Diseases vol 65 no 1 pp 2ndash6 2006
[16] A E Gharavi S S Pierangeli R G Espinola X Liu MColden-Stanfield and E N Harris ldquoAntiphospholipid antibod-ies induced in mice by immunization with a cytomegalovirus-derived peptide cause thrombosis and activation of endothelialcells in vivordquo Arthritis and Rheumatism vol 46 no 2 pp 545ndash552 2002
[17] M Motta G Chirico C B Rebaioli et al ldquoAnticardiolipin andanti-beta2 glycoprotein I antibodies in infants born to motherswith antiphospholipid antibody-positive autoimmune diseasea follow-up studyrdquoAmerican Journal of Perinatology vol 23 no4 pp 247ndash251 2006
[18] T Avcin A Ambrozic M Kuhar T Kveder and B RozmanldquoAnticardiolipin and anti-1205732 glycoprotein I antibodies in seraof 61 apparently healthy children at regular preventive visitsrdquoRheumatology vol 40 no 5 pp 565ndash573 2001
[19] A Ambrozic ldquoAnti-beta2-glycoprotein I antibodies in children
with atopic dermatitisrdquo International Immunology vol 14 no 7pp 823ndash830
[20] J A Giron-Gonzalez E Garcıa Del Rıo C Rodrıguez JRodrıguez-Martorell and A Serrano ldquoAntiphospholipid syn-drome and asymptomatic carriers of antiphospholipid anti-body prospective analysis of 404 individualsrdquo Journal ofRheumatology vol 31 no 8 pp 1560ndash1567 2004
[21] K A Breen P Seed K Parmar GWMoore S E Stuart-Smithand B J Hunt ldquoComplement activation in patients with isolatedantiphospholipid antibodies or primary antiphospholipid syn-dromerdquo Thrombosis and Haemostasis vol 107 no 3 pp 423ndash429 2012
Journal of Immunology Research 11
[22] F Fischetti P Durigutto V Pellis et al ldquoThrombus formationinduced by antibodies to 1205732-glycoprotein I is complementdependent and requires a priming factorrdquo Blood vol 106 no7 pp 2340ndash2346 2005
[23] J Guerin E Casey C Feighery and J Jackson ldquoAnti-Beta 2-glycoprotein I antibody isotype and IgG subclass in antiphos-pholipid syndrome patientsrdquo Autoimmunity vol 31 no 2 pp109ndash116 1999
[24] L Andreoli C Nalli M Motta et al ldquoAnti-1205732-glycoprotein I
IgG antibodies from 1-year-old healthy children born to moth-ers with systemic autoimmune diseases preferentially targetdomain 45 might it be the reason for their lsquoinnocentrsquo profilerdquoAnnals of the Rheumatic Diseases vol 70 no 2 pp 380ndash3832011
[25] B de Laat R H W M Derksen M van Lummel M T T Pen-nings and P G deGroot ldquoPathogenic anti-beta2-glycoprotein Iantibodies recognize domain I of beta2-glycoprotein I only aftera conformational changerdquo Blood vol 107 no 5 pp 1916ndash19242006
[26] M Samarkos K A Davies C Gordon M J Walport and SLoizou ldquoIgG subclass distribution of antibodies against 1205732-GP1and cardiolipin in patients with systemic lupus erythematosusand primary antiphospholipid syndrome and their clinicalassociationsrdquo Rheumatology vol 40 no 9 pp 1026ndash1032 2001
[27] J Arvieux B Roussel D Ponard and M G Colomb ldquoIgG2subclass restriction of anti-1205732 glycoprotein 1 antibodies inautoimmune patientsrdquo Clinical and Experimental Immunologyvol 95 no 2 pp 310ndash315 1994
[28] J N Arnold M R Wormald R B Sim P M Rudd and R ADwek ldquoThe impact of glycosylation on the biological functionand structure of human immunoglobulinsrdquo Annual Review ofImmunology vol 25 pp 21ndash50 2007
[29] Y Kaneko F Nimmerjahn and J V Ravetch ldquoAnti-inflamma-tory activity of immunoglobulin G resulting from Fc sialyla-tionrdquo Science vol 313 no 5787 pp 670ndash673 2006
[30] RM Anthony and F Nimmerjahn ldquoThe role of differential IgGglycosylation in the interaction of antibodies with FcgammaRsin vivordquo Current Opinion in Organ Transplantation vol 16 no1 pp 7ndash14 2011
[31] R M Anthony and J V Ravetch ldquoA novel role for the IgGFc glycan the anti-inflammatory activity of sialylated IgG FcsrdquoJournal of Clinical Immunology vol 30 no 1 pp S9ndashS14 2010
[32] K Oku T Atsumi M Bohgaki et al ldquoComplement activationin patients with primary antiphospholipid syndromerdquoAnnals ofthe Rheumatic Diseases vol 68 no 6 pp 1030ndash1035 2009
[33] H P McNeil R J Simpson C N Chesterman and S AKrilis ldquoAnti-phospholipid antibodies are directed against acomplex antigen that includes a lipid-binding inhibitor of coag-ulation 1205732-Glycoprotein I (apolipoprotein H)rdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 87 no 11 pp 4120ndash4124 1990
[34] R Jefferis and D S Kumararatne ldquoSelective IgG subclassdeficiency quantification and clinical relevancerdquo Clinical andExperimental Immunology vol 81 no 3 pp 357ndash367 1990
[35] P H Schur F Rosen and M E Norman ldquoImmunoglobulinsubclasses in normal childrenrdquo Pediatric Research vol 13 no3 pp 181ndash183 1979
[36] M A H French and G Harrison ldquoSerum IgG subclass concen-trations in healthy adults a study using monoclonal antiserardquoClinical and Experimental Immunology vol 56 no 2 pp 473ndash475 1984
[37] M Odorczuk L B Keil and V A DeBari ldquoBinding of com-plement component C1q to anti-beta2 glycoprotein I antibodiesfrom patients with antiphospholipid syndromerdquo InflammationResearch vol 48 no 10 pp 524ndash526 1999
[38] L Truedsson A A Bengtsson and G Sturfelt ldquoComplementdeficiencies and systemic lupus erythematosusrdquoAutoimmunityvol 40 no 8 pp 560ndash566 2007
[39] S Aschermann C H K Lehmann S Mihai G Schett DDudziak and F Nimmerjahn ldquoB cells are critical for autoim-mune pathology in Scurfy micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 110 no47 pp 19042ndash19047 2013
[40] Y Kaneko F Nimmerjahn M P Madaio and J V RavetchldquoPathology and protection in nephrotoxic nephritis is deter-mined by selective engagement of specific Fc receptorsrdquo TheJournal of Experimental Medicine vol 203 no 3 pp 789ndash7972006
[41] D Sylvestre R Clynes M Ma H Warren M C Carrolland J V Ravetch ldquoImmunoglobulin G-mediated inflammatoryresponses develop normally in complement-deficient micerdquoJournal of ExperimentalMedicine vol 184 no 6 pp 2385ndash23921996
[42] G E Ritchie B E Moffatt R B Sim B P Morgan R A Dwekand P M Rudd ldquoGlycosylation and the complement systemrdquoChemical Reviews vol 102 no 2 pp 305ndash320 2002
[43] Y Palarasah K Skjodt J Brandt et al ldquoGeneration of aC3c specific monoclonal antibody and assessment of C3c as aputative inflammatorymarker derived from complement factorC3rdquo Journal of ImmunologicalMethods vol 362 no 1-2 pp 142ndash150 2010
[44] D G Doherty J A Underhill P T Donaldson et al ldquoPoly-morphism in the human complement C4 genes and geneticsusceptibility to autoimmune hepatitisrdquo Autoimmunity vol 18no 4 pp 243ndash249 1994
[45] M-H Tao and S L Morrison ldquoStudies of aglycosylatedchimeric mouse-human IgG Role of carbohydrate in thestructure and effector functions mediated by the human IgGconstant regionrdquoThe Journal of Immunology vol 143 no 8 pp2595ndash2601 1989
[46] R Jefferis ldquoGlycosylation of recombinant antibody therapeu-ticsrdquo Biotechnology Progress vol 21 no 1 pp 11ndash16 2005
[47] Y Mimura S Church R Ghirlando et al ldquoThe influence ofglycosylation on the thermal stability and effector functionexpression of human IgG1-Fc properties of a series of truncatedglycoformsrdquoMolecular Immunology vol 37 no 12-13 pp 697ndash706 2001
[48] C Huhn M H J Selman L R Ruhaak A M Deelder and MWuhrer ldquoIgG glycosylation analysisrdquo Proteomics vol 9 no 4pp 882ndash913 2009
[49] J Lund N Takahashi J D Pound M Goodall and R JefferisldquoMultiple interactions of IgG with its core oligosaccharide canmodulate recognition by complement and human Fc gammareceptor I and influence the synthesis of its oligosaccharidechainsrdquo The Journal of Immunology vol 157 no 11 pp 4963ndash4969 1996
[50] R Forastiero M Martinuzzo L O Carreras and J MacloufldquoAnti-1205732 glycoprotein I antibodies and platelet activation inpatients with antiphospholipid antibodies association withincreased excretion of platelet-derived thromboxane urinarymetabolitesrdquoThrombosis and Haemostasis vol 79 no 1 pp 42ndash45 1998
12 Journal of Immunology Research
[51] J-C Reverter and D Tassies ldquoMechanism of thrombosis inantiphospholipid syndrome binding to plateletsrdquo in HughesSyndrome M A Khamashta Ed pp 403ndash414 Springer Lon-don UK 2006
[52] G Espinosa R Cervera J Font and Y Shoenfeld ldquoAntiphos-pholipid syndrome pathogenic mechanismsrdquo AutoimmunityReviews vol 2 no 2 pp 86ndash93 2003
[53] T Shi B Giannakopoulos X Yan et al ldquoAnti-1205732-glycoprotein
I antibodies in complex with 1205732-glycoprotein I can activate
platelets in a dysregulated manner via glycoprotein Ib-IX-VrdquoArthritis amp Rheumatism vol 54 no 8 pp 2558ndash2567 2006
[54] P LMeroni E RaschiMCamera et al ldquoEndothelial activationby aPL a potential pathogenetic mechanism for the clinicalmanifestations of the syndromerdquo Journal of Autoimmunity vol15 no 2 pp 237ndash240 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
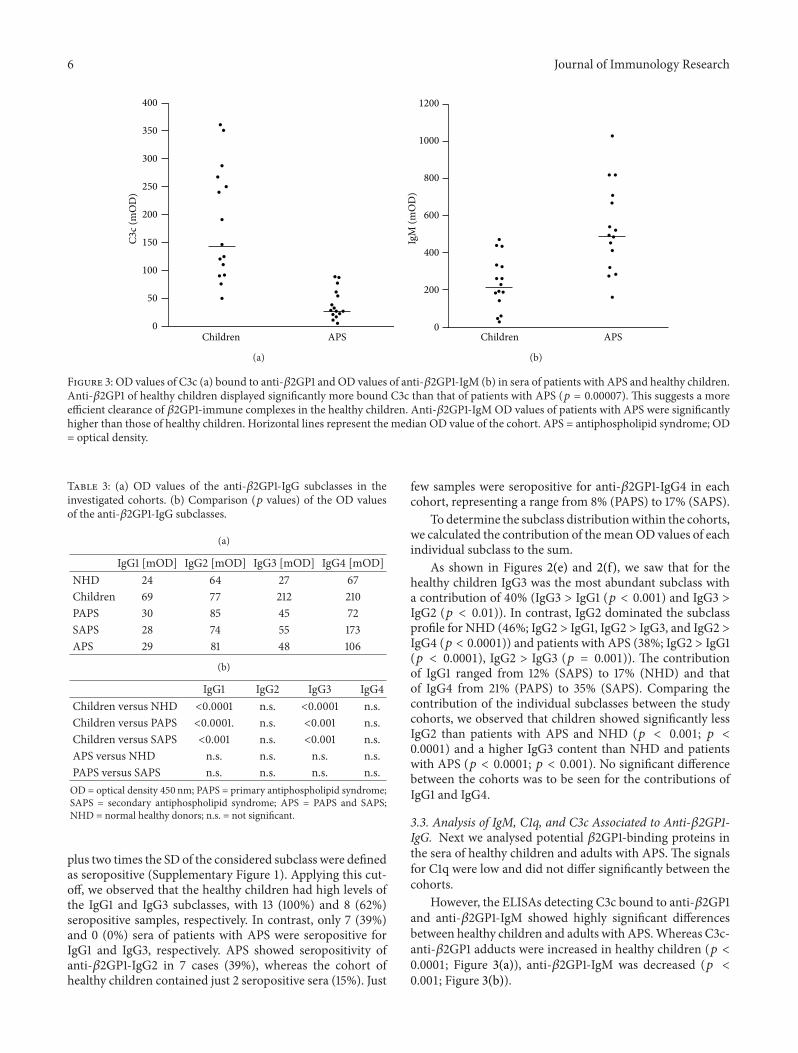
6 Journal of Immunology Research
0
50
100
150
200
250
300
350
400
Children APS
C3c (
mO
D)
(a)
0
200
400
600
800
1000
1200
Children APS
IgM
(mO
D)
(b)
Figure 3 OD values of C3c (a) bound to anti-1205732GP1 and OD values of anti-1205732GP1-IgM (b) in sera of patients with APS and healthy childrenAnti-1205732GP1 of healthy children displayed significantly more bound C3c than that of patients with APS (119901 = 000007) This suggests a moreefficient clearance of 1205732GP1-immune complexes in the healthy children Anti-1205732GP1-IgM OD values of patients with APS were significantlyhigher than those of healthy children Horizontal lines represent the median OD value of the cohort APS = antiphospholipid syndrome OD= optical density
Table 3 (a) OD values of the anti-1205732GP1-IgG subclasses in theinvestigated cohorts (b) Comparison (119901 values) of the OD valuesof the anti-1205732GP1-IgG subclasses
(a)
IgG1 [mOD] IgG2 [mOD] IgG3 [mOD] IgG4 [mOD]NHD 24 64 27 67Children 69 77 212 210PAPS 30 85 45 72SAPS 28 74 55 173APS 29 81 48 106
(b)
IgG1 IgG2 IgG3 IgG4Children versus NHD lt00001 ns lt00001 nsChildren versus PAPS lt00001 ns lt0001 nsChildren versus SAPS lt0001 ns lt0001 nsAPS versus NHD ns ns ns nsPAPS versus SAPS ns ns ns nsOD = optical density 450 nm PAPS = primary antiphospholipid syndromeSAPS = secondary antiphospholipid syndrome APS = PAPS and SAPSNHD = normal healthy donors ns = not significant
plus two times the SD of the considered subclass were definedas seropositive (Supplementary Figure 1) Applying this cut-off we observed that the healthy children had high levels ofthe IgG1 and IgG3 subclasses with 13 (100) and 8 (62)seropositive samples respectively In contrast only 7 (39)and 0 (0) sera of patients with APS were seropositive forIgG1 and IgG3 respectively APS showed seropositivity ofanti-1205732GP1-IgG2 in 7 cases (39) whereas the cohort ofhealthy children contained just 2 seropositive sera (15) Just
few samples were seropositive for anti-1205732GP1-IgG4 in eachcohort representing a range from 8 (PAPS) to 17 (SAPS)
To determine the subclass distributionwithin the cohortswe calculated the contribution of themeanOD values of eachindividual subclass to the sum
As shown in Figures 2(e) and 2(f) we saw that for thehealthy children IgG3 was the most abundant subclass witha contribution of 40 (IgG3 gt IgG1 (119901 lt 0001) and IgG3 gtIgG2 (119901 lt 001)) In contrast IgG2 dominated the subclassprofile for NHD (46 IgG2 gt IgG1 IgG2 gt IgG3 and IgG2 gtIgG4 (119901 lt 00001)) and patients with APS (38 IgG2 gt IgG1(119901 lt 00001) IgG2 gt IgG3 (119901 = 0001)) The contributionof IgG1 ranged from 12 (SAPS) to 17 (NHD) and thatof IgG4 from 21 (PAPS) to 35 (SAPS) Comparing thecontribution of the individual subclasses between the studycohorts we observed that children showed significantly lessIgG2 than patients with APS and NHD (119901 lt 0001 119901 lt00001) and a higher IgG3 content than NHD and patientswith APS (119901 lt 00001 119901 lt 0001) No significant differencebetween the cohorts was to be seen for the contributions ofIgG1 and IgG4
33 Analysis of IgM C1q and C3c Associated to Anti-1205732GP1-IgG Next we analysed potential 1205732GP1-binding proteins inthe sera of healthy children and adults with APS The signalsfor C1q were low and did not differ significantly between thecohorts
However the ELISAs detecting C3c bound to anti-1205732GP1and anti-1205732GP1-IgM showed highly significant differencesbetween healthy children and adults with APSWhereas C3c-anti-1205732GP1 adducts were increased in healthy children (119901 lt00001 Figure 3(a)) anti-1205732GP1-IgM was decreased (119901 lt0001 Figure 3(b))
Journal of Immunology Research 7
Table 4 Number (frequency) of seropositive samples of the anti-1205732GP1-IgG subclasses
Number of seropositive samples (gtmOD[NHD] + 2 times SD[NHD])IgG1
(gt29mOD)IgG2
(gt79mOD)IgG3
(gt85mOD)IgG4
(gt319mOD)NHD (119899 = 17) 0 (0) 0 (0) 1 (6) 1 (6)Children (119899 = 13) 13 (100) 2 (15) 8 (62) 2 (15)PAPS (119899 = 12) 6 (50) 5 (42) 0 (0) 1 (8)SAPS (119899 = 6) 1 (17) 2 (33) 0 (0) 1 (17)APS (119899 = 18) 7 (39) 7 (39) 0 (0) 2 (11)OD = optical density 450 nm PAPS = primary antiphospholipid syndrome SAPS = secondary antiphospholipid syndrome APS = PAPS and SAPS NHD =normal healthy donorsThe cut-off for seropositivity was calculated from the mean OD of the corresponding anti-1205732GP1-IgG subclass of the NHD cohort andits standard deviation (SD) according to the formula mean OD[NHD] + 2 times SD[NHD] The respective cut-off OD values are indicated in parentheses below theanti-1205732GP1-IgG subclasses
Table 5 Cohorts studied with the lectin ELISA
Cohort Abbreviation Number of samplesPatients with primary APS PAPS 12Patients with APS and SLE as underlying disease SAPS 10Patients with SLE without symptoms of APS harbouring circulating aPL SLE + aPL 15Asymptomatic carriers of aPL aaPL 6Healthy children with circulating anti-1205732GP1 mdash 16APS = antiphospholipid syndrome aPL = antiphospholipid antibodies SLE = systemic lupus erythematosus
34 Analysis of the Sialylation of Anti-1205732GP1 We analysedthe glycosylation of anti-1205732GP1-IgG focusing on its sialy-lation since highly sialylated Fc-parts of IgG are known tocause rather anti-inflammatory responses [30] Wemeasuredthe binding of the sialic acid specific lectin SNA to the anti-1205732GP1-IgG As shown in Table 5 we employed five cohorts ofindividuals 12 patientswith PAPS 10 patientswith SAPS (SLEas primary disease) 15 patients with SLE who did not showclinical sign of APS but with aPL in their sera (SLE + aPL)6 asymptomatic adult carriers of aPL (aaPL) and 16 healthychildren harbouring aPL
Analysing the SNAIgG-ratios (Figure 4) we identifiedthe highest sialylation level in the sera of the healthy childrenThe significances were rather high and decreased from PAPS(119901 lt 00001) to SAPS (119901 lt 00001) to SLE + aPL (119901 lt00001) and to aaPL (119901 lt 0001)
4 Discussion
We analysed two distinct sets of sera One was used toinvestigate anti-1205732GP1-IgG its subclasses and its associatedproteins C1q and C3c For the analysis of the sialylation ofanti-1205732GP1-IgG we had to collect fresh sera from patientswith APS and the healthy children since the stability of theantibody-glycosylation is unknown In order to avoid itspossible decay by processes like enzymatic degradation thisperiod has been kept as short as possible and was followed byimmediate investigation of the glycosylation
The antiphospholipid syndrome is characterized by clin-ical manifestations such as arterial and venous thrombosisor pregnancy-related complications and the detection of spe-cific autoantibodies (lupus anticoagulant anti-1205732GP1- and
anticardiolipin-autoantibodies) [1] In particular anti-1205732GP1sare considered responsible for provoking the pathologicalcomplications [3]We firstmeasured the total amount of anti-1205732GP1-IgG in the sera of 16 healthy children without clinicalmanifestations of APS and 46 adult patients with APS toidentify individual cohorts seropositive for anti-1205732GP1 17adult normal healthy donors served as negative control
The samples of the patientswithAPS showedhigher levelsof anti-1205732GP1-IgG than those of NHD We also observeda higher concentration of anti-1205732GP1-IgG in the sera ofhealthy children compared to NHD This is due to the factthat the detection of these antibodies has been the inclusioncriterion for the healthy children examined in this studyInterestingly the children showed even higher OD values forthe putatively pathogenic anti-1205732GP1-IgG than the patientswith APS This finding was surprising since the childrenwere clinically healthy and did not show any signs of APSsuch as thrombosis or thrombocytopenia complications inwhich the existence of aPL especially anti-1205732GP1 is maderesponsible for [7 33]
High levels of circulating aPL in 61 healthy children whencompared to healthy adults have already been described byAvcin and colleagues [18] The anti-1205732GP1-IgG was report-edly higher in preschool children with a mean age of 5years than in adolescents with a mean age of 135 years[18] The strongly elevated circulating anti-1205732GP1-IgG in ourcohort of healthy children may be explained by the fact thatthey had an even lower age (mean age was nine months)One possible explanation for this result is the inductionof anti-1205732GP1 by viral and bacterial infections [10 11] ornutritional exposure to 1205732GP1 [17] As the immune systemin children is still to evolve they are more susceptible to
8 Journal of Immunology Research
0
05
10
15
20
25SN
AIg
G
ChildrenPAPS SAPS aaPLSLE + aPL
lowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowast
Figure 4 SNAanti-1205732GP1-IgG ratio in PAPS SAPS SLE + aPLaaPL and children Healthy children show a significantly higherSNAanti-1205732GP1-IgG ratio than PAPS SAPS SLE + aPL and aaPL(lowastlowast119901 lt 00001 lowastlowastlowast119901 lt 000001) This suggests higher sialylation ofthe oligosaccharides attached to Fc fragments of the childrenrsquos anti-1205732GP1No significant differenceswere foundbetween the remainingcohorts Horizontal lines indicate the mean SNAIgG ratio of thecohort SNA = Sambucus nigra agglutinin APS = antiphospholipidsyndrome PAPS = patients with primary APS SAPS = patients withAPS and SLE as underlying disease SLE + aaPL = patients withSLE without symptoms of APS harbouring circulating aPL aaPL =asymptomatic carriers of aPL aPL = antiphospholipid antibodiesSLE = systemic lupus erythematosus
infections than adults and their immune system has to copewith a plethora of viral and bacterial pathogens for the firsttime in their life This scenario is prone to result in thestimulation of autoantibody-production Molecular mimicryof certain pathogens can lead to the production of antibodieswhich then cross-react with 1205732GP1 [7 12ndash15] When infantsdiversify their dietary habits and start to consume productslike milk and meat they can get into contact with nutritional1205732GP1 [19] This is a further condition capable triggeringthe production of anti-1205732GP1 Exposure to pathogen-derivedand dietary 1205732GP1 could explain the high level of circulatinganti-1205732GP1-IgG in healthy childrenThis low affine reactivityis consecutively lost during the maturation of the immunesystem and replaced by the emergence in adults of high affineantibodies that more specifically recognize pathogens Apartfrom possible origins of these autoantibodies it is obviousthat they do not provoke any of the hallmark symptoms ofAPS In contrast to their adult counterparts anti-1205732GP1-IgGscirculating in the plasma of healthy children are obviouslynot pathogenic So it seems decent to hypothesize thatanti-1205732GP1 autoantibodies in healthy children biochemicallydiffer from those in adults
To test this hypothesis we examined the anti-1205732GP1-IgG subclass distribution in the seropositive sera of the anti-1205732GP1-IgG ELISA The healthy children showed higher ODvalues for the IgG1- and IgG3-subclasses than patients withAPS and NHD This was consistent with the number ofseropositive samples In the anti-1205732GP1-IgG1-subclass every
tested serum of the healthy children surpassed the thresholdset for seropositivity (gtmean OD
[NHD] + 2 times SD[NHD]) and
8 samples (62) did so in the IgG3-subclass In the seraof patients with APS just 7 (39) and no samples wereseropositive for IgG1 or IgG3 In the sera of patients with APScompared to NHD we observed higher OD levels of anti-1205732GP1 in all subclasses except for IgG4This result is consis-tent with the findings of Guerin et al who reported elevatedlevels in all anti-1205732GP1-IgG subclasses even including IgG4in patients with APS compared to a control group of healthyadults [23]
It might be that the elevated absolute OD values of theanti-1205732GP1-IgG subclasses are the logical consequence ofthe generally higher level of anti-1205732GP1-IgG in the patientswith APS and healthy children who were selected for furthertesting Therefore we also analysed the subclass distributionof this antibody We first analysed the anti-1205732GP1-IgG sub-class distribution within the individual cohorts and observedthat in the healthy children the IgG3 antibody was mostabundant with a contribution of 40 to the total reactivityThis is consistent with the finding that all sera of the healthychildren were seropositive in this subclass and that theiranti-1205732GP1-IgG3 OD values were the highest of all cohortsThis indicates a predominance of this subclass in the healthychildren The remaining anti-1205732GP1-IgG subclasses in thesera of the children ranged from 15 (IgG1) over 20 (IgG2)to 25 (IgG4) but the differences between their proportionswere not statistically significant In contrast in the sera ofthe patients with APS the anti-1205732GP1-IgG2 subclass had thehighest contribution with 38
The finding of an elevated proportion of the IgG2 subclassin the patients with APS confirms former published datawhich also describe a skewing towards anti-1205732GP1-IgG2 inAPS [19 23 26 27] The authors detected an even highercontribution of anti-1205732GP1-IgG2 (702 [26] and 87 [27]resp) In contrast to our results the IgG1 subclass wasmore prominent in the published studies described aboveImportantly in the cohort of the healthy infants IgG2 wasrelatively lowTherefore onemay argue that anti-1205732GP1-IgG2is not just a marker of the disease but is also involved inthe pathogenic action of the aPL autoantibodies When wecompared the anti-1205732GP1-IgG subclass distribution betweenthe three cohorts we noticed a significant difference in thedistribution concerning the children on one hand and NHDand patients with APS on the other Healthy children showeda higher proportion of anti-1205732GP1-IgG3 and a lower one ofanti-1205732GP1-IgG2 In NHD and patients with APS it was theother way round IgG1 and IgG4 did not significantly differbetween healthy children and patients with APS orNHDThepredominance of anti-1205732GP1-IgG3 in the healthy children isa hint to an immune response elicited by a proteinaceousantigen [26 27 34] This supports the hypothesis thatthe harmless anti-1205732GP1 in children are infection-inducedwith pathogen-derived proteins serving as trigger for theirproduction However it is still elusive if the IgG3-dominatedanti-1205732GP1 response found in the sera of healthy childrencontributes to its low pathogenic potential Alternativelyit can just be ascribed to the proceeding evolution of theinfantrsquos immune system typically associated with an elevated
Journal of Immunology Research 9
proportion of IgG3 antibodies [34 35] During childhoodthe proportion of IgG3 continuously decreases to adult rangesusually reached at the age of 10ndash12 years However the aboveaverage elevated proportion of the anti-1205732GP1-IgG3 (40) inthe sera of the healthy childrenmay thus not just be due to theinfant immune system In particular when bearing in mindthat normal adult levels of the IgG3 subclass are just around6 [34 36]
IgG3 is the IgG-subclass exerting the most effectiveactivation of the complement cascade through the classicalpathway [34] Since complement activation is associated withthe pathogenesis of APS [21 32 37] the high proportionof complement-activating anti-1205732GP1-IgG3 in the childrenseems to contradict a possible protective effect against APS ofthese kinds of antibodies However the role of complement inthe etiopathogenesis of autoimmune diseases is manifold asthe complement components C1q C2 and C4 contribute tothe clearance of postapoptotic secondary necrotic cells andtheir deficiency is therefore a risk factor for autoimmunityand autoinflammation [38] In a similar manner studiesin mouse in vivo models of various autoimmune diseasesdemonstrated that IgG mediated tissue inflammation wasblocked in mice deficient in activating Fc120574Rs although thecomplement component C3 was still abundantly depositedin the tissue [39ndash41] Thus the contribution of complementdeposits in tissue with respect to tissue inflammation remainsto be established
To obtain new insights into the involvement of thecomplement system in the pathogenesis of APS we analysedthe C1q- and C3c-binding to anti-1205732GP1 We observed thatanti-1205732GP1 in the sera of the healthy children and in patientswith APS similarly bound C1q the first molecule of theclassical pathway of complement activation [42] Performinganti-1205732GP1-IgM-ELISAs we detected significantly higherIgM values in the patients with APS One would expecta higher C1q-binding of the IgM positive sera since thisimmunoglobulin binds and activates C1q more strongly thanIgG However the children mainly harbour IgG3 autoanti-bodies the most potent subclass for the activation of theclassical complement pathway [34]This may compensate forthe lower IgM level in their sera and thus be responsiblefor the similar C1q-binding of both cohorts This suggeststhat preconditions for complement activation via the classicalpathway are comparable in both cohorts We conclude thatclassical complement activation has a minor impact on thedifferential pathogenesis of aPL in healthy children andpatients with APS
What then is the culprit that induces their pathogenicpotential
Nextwe quantified the contribution ofC3c to the immunecomplexes bound to 1205732GP1 C3c is a domain in the C3bmolecule and one of its cleavage products created aftercleavage by factor I of C3b [42 43] C3c not just representsa marker of complement activation but also serves as anopsonin improving the clearance of bound targets Weobserved significantly more C3c bound to anti-1205732GP1 inthe sera of the healthy children than in those of patientswith APS This reflects a better opsonization by C3c ofthe potentially harmful anti-1205732GP1 immune complexes
anti-1205732GP1-opsonized cells or cellular fragments in thesera of the healthy children Consequently the anti-1205732GP1complexes can be cleared more efficiently by the powerfulcomplement system of the healthy children In patients withchronic inflammatory autoimmune diseases complement isusually low which may result from null alleles [44] or fromcomplement consumption [38]
But why do these processes not cause inflammation andaberrant coagulation in the childrenApart fromanti-1205732GP1-IgG subclass distribution and distinct ability to activate thecomplement system it is likely that other properties of theanti-1205732GP1 in the sera of children contribute to their lowpathogenicity We hypothesized that an increased sialylationof the anti-1205732GP1-IgG in healthy individuals renders themharmless for their hosts IgG is usually N-glycosylated at theCH2-domain of its Fc-part This N-glycan crucially modifiesthe effector functions the IgG molecule is endowed with[45ndash47] This biantennary core-fucosylated oligosaccharidecarries a variable amount of sugar monomers attached toboth arms [28 48] IgG antibodies terminating with sialicacids on one or both antennae have been shown to behaveanti-inflammatorily after infusion of IVIG [29 31]The signalexerted by the terminal sialic acid is dominant since thefraction of sialylated IgG in healthy plasma is just 4 to 14[28 46 49]
In mice these effects are mainly ascribed to decreasedbinding of sialylated IgG to activating Fc120574-receptors andan upregulation of the inhibitory Fc120574RIIB [29 31] Weobserved that anti-1205732GP1-IgG isolated from sera of healthychildren was significantly more sialylated than that fromsymptomatic patients with APS One may predict that theseautoantibodies in complex with 1205732GP1 show low affinity foractivating Fc120574Rs and therefore do not provoke proinflam-matory and procoagulant effector functions Fc120574RIIA is anexample for an activating receptor It is located on a plethoraof cells On the surfaces of platelets it mediates activationby immune complexes The activation is accompanied byplatelet aggregation and the release of mediators such asthe prothromboticThromboxaneA2 [50] Consecutively thiscascade can result in thrombus formation [51 52] Sialylatedanti-1205732GP1-IgG is unable to bind to and to activate plateletsvia Fc120574RIIA
Further mechanisms involved in thrombus formation inAPS comprise the interaction of anti-1205732GP1 with variouspattern recognition receptors [53] and its interference withregulatory functions ofmolecules involved in the coagulationprocess (protein C prothrombin or tissue factor [52 54])Direct binding to 1205732GP1 on endothelial cells (EC) shiftstheir phenotype towards coagulation and inflammation [54]Considering that Fc120574R-binding is involved in many of thesemechanisms it seems likely that a high sialylation of anti-1205732GP1-IgG leads to a low pathogenicity of these autoan-tibodies and does not cause APS Therefore we concludethat the higher sialylation of anti-1205732GP1-IgG in the seraof the healthy children plays a major role in protectingthem from typical complications of APS This hypothesis isfurther supported by the observation that antiphospholipidantibodies from sera of asymptomatic adult carriers aremore sialylated than those from patients with APS A high
10 Journal of Immunology Research
sialylation status of antiphospholipid antibodies seems toprevent the development of symptoms of APS also in theseindividuals
We conclude that a plethora of factors contributes tothe low pathogenicity of anti-1205732GP1-IgG in the sera ofhealthy children and asymptomatic adults We observed adistinct spectrum of IgG-subclasses compared to patientswith APS and a better binding of C3c indicating a moreefficient clearance of potentially harmful anti-1205732GP1 Thehypersialylation of the anti-1205732GP1-IgG is prone to amelioratetheir inflammatory and procoagulatory effects in healthycarriers whereas hyposialylation and inflammationmutuallyprecipitate the antiphospholipid syndrome
Disclosure
Thepresent workwas performed in fulfillment of the require-ments for obtaining the degree ldquoDr medrdquo for ChristophFickentscher
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Laura Andreoli andMartin Herrmann contributed equally tosenior authorship
Acknowledgments
This project was supported by the German Research Foun-dation (training research programme of the collaborativeresearch center 643) by the Emerging Fields Initiative (EFI)of the FAU Erlangen-Nuremberg and by the K und RWucherpfennig-Stiftung
References
[1] S Miyakis M D Lockshin T Atsumi et al ldquoInternationalconsensus statement on an update of the classification criteriafor definite antiphospholipid syndrome (APS)rdquo Journal ofThrombosis and Haemostasis vol 4 no 2 pp 295ndash306 2006
[2] W A Wilson A E Gharavi T Koike et al ldquoInternationalconsensus statement on preliminary classification criteria fordefinite antiphospholipid syndrome report of an internationalworkshoprdquo Arthritis and Rheumatism vol 42 no 7 pp 1309ndash1311 1999
[3] P G de Groot and R T Urbanus ldquoThe significance of autoan-tibodies against beta2-glycoprotein Irdquo Blood vol 120 no 2 pp266ndash274 2012
[4] C Nalli S Piantoni L Andreoli M Motta and A TincanildquoAntiphospholipid syndrome and antiphospholipid antibodiesin children the two sides of the coinrdquo Israel Medical AssociationJournal vol 14 no 5 pp 310ndash312 2012
[5] I Schousboe ldquo1205732-glycoprotein I a plasma inhibitor of thecontact activation of the intrinsic blood coagulation pathwayrdquoBlood vol 66 no 5 pp 1086ndash1091 1985
[6] J George M Blank Y Levy et al ldquoDifferential effects of anti-1205732-glycoprotein I antibodies on endothelial cells and on themanifestations of experimental antiphospholipid syndromerdquoCirculation vol 97 no 9 pp 900ndash906 1998
[7] R Willis and S S Pierangeli ldquoAnti-1205732-glycoprotein I antibod-iesrdquo Annals of the New York Academy of Sciences vol 1285 no1 pp 44ndash58 2013
[8] M J Fischer J Rauch and J S Levine ldquoThe antiphospholipidsyndromerdquo Seminars in Nephrology vol 27 no 1 pp 35ndash462007
[9] H Amital M Govoni R Maya et al ldquoRole of infectious agentsin systemic rheumatic diseasesrdquo Clinical and ExperimentalRheumatology vol 26 no 1 supplement 48 pp S27ndashS32 2008
[10] T Avcin ldquoAntiphospholipid syndrome in childrenrdquo CurrentOpinion in Rheumatology vol 20 no 5 pp 595ndash600 2008
[11] M Garcıa-Carrasco C Galarza-Maldonado C Mendoza-Pinto R O Escarcega and R Cervera ldquoInfections and theantiphospholipid syndromerdquo Clinical Reviews in Allergy ampImmunology vol 36 no 2-3 pp 104ndash108 2009
[12] P Cruz-Tapias M Blank J-M Anaya and Y ShoenfeldldquoInfections and vaccines in the etiology of antiphospholipidsyndromerdquo Current Opinion in Rheumatology vol 24 no 4 pp389ndash393 2012
[13] P von Landenberg Y Doring S Modrow and K J LacknerldquoAre antiphospholipid antibodies an essential requirement foran effective immune response to infectionsrdquoAnnals of the NewYork Academy of Sciences vol 1108 pp 578ndash583 2007
[14] M Blank I Krause M Fridkin et al ldquoBacterial induction ofautoantibodies to 1205732-glycoprotein-I accounts for the infectiousetiology of antiphospholipid syndromerdquo Journal of ClinicalInvestigation vol 109 no 6 pp 797ndash804 2002
[15] Y Shoenfeld M Blank R Cervera J Font E Raschi and P-LMeroni ldquoInfectious origin of the antiphospholipid syndromerdquoAnnals of the Rheumatic Diseases vol 65 no 1 pp 2ndash6 2006
[16] A E Gharavi S S Pierangeli R G Espinola X Liu MColden-Stanfield and E N Harris ldquoAntiphospholipid antibod-ies induced in mice by immunization with a cytomegalovirus-derived peptide cause thrombosis and activation of endothelialcells in vivordquo Arthritis and Rheumatism vol 46 no 2 pp 545ndash552 2002
[17] M Motta G Chirico C B Rebaioli et al ldquoAnticardiolipin andanti-beta2 glycoprotein I antibodies in infants born to motherswith antiphospholipid antibody-positive autoimmune diseasea follow-up studyrdquoAmerican Journal of Perinatology vol 23 no4 pp 247ndash251 2006
[18] T Avcin A Ambrozic M Kuhar T Kveder and B RozmanldquoAnticardiolipin and anti-1205732 glycoprotein I antibodies in seraof 61 apparently healthy children at regular preventive visitsrdquoRheumatology vol 40 no 5 pp 565ndash573 2001
[19] A Ambrozic ldquoAnti-beta2-glycoprotein I antibodies in children
with atopic dermatitisrdquo International Immunology vol 14 no 7pp 823ndash830
[20] J A Giron-Gonzalez E Garcıa Del Rıo C Rodrıguez JRodrıguez-Martorell and A Serrano ldquoAntiphospholipid syn-drome and asymptomatic carriers of antiphospholipid anti-body prospective analysis of 404 individualsrdquo Journal ofRheumatology vol 31 no 8 pp 1560ndash1567 2004
[21] K A Breen P Seed K Parmar GWMoore S E Stuart-Smithand B J Hunt ldquoComplement activation in patients with isolatedantiphospholipid antibodies or primary antiphospholipid syn-dromerdquo Thrombosis and Haemostasis vol 107 no 3 pp 423ndash429 2012
Journal of Immunology Research 11
[22] F Fischetti P Durigutto V Pellis et al ldquoThrombus formationinduced by antibodies to 1205732-glycoprotein I is complementdependent and requires a priming factorrdquo Blood vol 106 no7 pp 2340ndash2346 2005
[23] J Guerin E Casey C Feighery and J Jackson ldquoAnti-Beta 2-glycoprotein I antibody isotype and IgG subclass in antiphos-pholipid syndrome patientsrdquo Autoimmunity vol 31 no 2 pp109ndash116 1999
[24] L Andreoli C Nalli M Motta et al ldquoAnti-1205732-glycoprotein I
IgG antibodies from 1-year-old healthy children born to moth-ers with systemic autoimmune diseases preferentially targetdomain 45 might it be the reason for their lsquoinnocentrsquo profilerdquoAnnals of the Rheumatic Diseases vol 70 no 2 pp 380ndash3832011
[25] B de Laat R H W M Derksen M van Lummel M T T Pen-nings and P G deGroot ldquoPathogenic anti-beta2-glycoprotein Iantibodies recognize domain I of beta2-glycoprotein I only aftera conformational changerdquo Blood vol 107 no 5 pp 1916ndash19242006
[26] M Samarkos K A Davies C Gordon M J Walport and SLoizou ldquoIgG subclass distribution of antibodies against 1205732-GP1and cardiolipin in patients with systemic lupus erythematosusand primary antiphospholipid syndrome and their clinicalassociationsrdquo Rheumatology vol 40 no 9 pp 1026ndash1032 2001
[27] J Arvieux B Roussel D Ponard and M G Colomb ldquoIgG2subclass restriction of anti-1205732 glycoprotein 1 antibodies inautoimmune patientsrdquo Clinical and Experimental Immunologyvol 95 no 2 pp 310ndash315 1994
[28] J N Arnold M R Wormald R B Sim P M Rudd and R ADwek ldquoThe impact of glycosylation on the biological functionand structure of human immunoglobulinsrdquo Annual Review ofImmunology vol 25 pp 21ndash50 2007
[29] Y Kaneko F Nimmerjahn and J V Ravetch ldquoAnti-inflamma-tory activity of immunoglobulin G resulting from Fc sialyla-tionrdquo Science vol 313 no 5787 pp 670ndash673 2006
[30] RM Anthony and F Nimmerjahn ldquoThe role of differential IgGglycosylation in the interaction of antibodies with FcgammaRsin vivordquo Current Opinion in Organ Transplantation vol 16 no1 pp 7ndash14 2011
[31] R M Anthony and J V Ravetch ldquoA novel role for the IgGFc glycan the anti-inflammatory activity of sialylated IgG FcsrdquoJournal of Clinical Immunology vol 30 no 1 pp S9ndashS14 2010
[32] K Oku T Atsumi M Bohgaki et al ldquoComplement activationin patients with primary antiphospholipid syndromerdquoAnnals ofthe Rheumatic Diseases vol 68 no 6 pp 1030ndash1035 2009
[33] H P McNeil R J Simpson C N Chesterman and S AKrilis ldquoAnti-phospholipid antibodies are directed against acomplex antigen that includes a lipid-binding inhibitor of coag-ulation 1205732-Glycoprotein I (apolipoprotein H)rdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 87 no 11 pp 4120ndash4124 1990
[34] R Jefferis and D S Kumararatne ldquoSelective IgG subclassdeficiency quantification and clinical relevancerdquo Clinical andExperimental Immunology vol 81 no 3 pp 357ndash367 1990
[35] P H Schur F Rosen and M E Norman ldquoImmunoglobulinsubclasses in normal childrenrdquo Pediatric Research vol 13 no3 pp 181ndash183 1979
[36] M A H French and G Harrison ldquoSerum IgG subclass concen-trations in healthy adults a study using monoclonal antiserardquoClinical and Experimental Immunology vol 56 no 2 pp 473ndash475 1984
[37] M Odorczuk L B Keil and V A DeBari ldquoBinding of com-plement component C1q to anti-beta2 glycoprotein I antibodiesfrom patients with antiphospholipid syndromerdquo InflammationResearch vol 48 no 10 pp 524ndash526 1999
[38] L Truedsson A A Bengtsson and G Sturfelt ldquoComplementdeficiencies and systemic lupus erythematosusrdquoAutoimmunityvol 40 no 8 pp 560ndash566 2007
[39] S Aschermann C H K Lehmann S Mihai G Schett DDudziak and F Nimmerjahn ldquoB cells are critical for autoim-mune pathology in Scurfy micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 110 no47 pp 19042ndash19047 2013
[40] Y Kaneko F Nimmerjahn M P Madaio and J V RavetchldquoPathology and protection in nephrotoxic nephritis is deter-mined by selective engagement of specific Fc receptorsrdquo TheJournal of Experimental Medicine vol 203 no 3 pp 789ndash7972006
[41] D Sylvestre R Clynes M Ma H Warren M C Carrolland J V Ravetch ldquoImmunoglobulin G-mediated inflammatoryresponses develop normally in complement-deficient micerdquoJournal of ExperimentalMedicine vol 184 no 6 pp 2385ndash23921996
[42] G E Ritchie B E Moffatt R B Sim B P Morgan R A Dwekand P M Rudd ldquoGlycosylation and the complement systemrdquoChemical Reviews vol 102 no 2 pp 305ndash320 2002
[43] Y Palarasah K Skjodt J Brandt et al ldquoGeneration of aC3c specific monoclonal antibody and assessment of C3c as aputative inflammatorymarker derived from complement factorC3rdquo Journal of ImmunologicalMethods vol 362 no 1-2 pp 142ndash150 2010
[44] D G Doherty J A Underhill P T Donaldson et al ldquoPoly-morphism in the human complement C4 genes and geneticsusceptibility to autoimmune hepatitisrdquo Autoimmunity vol 18no 4 pp 243ndash249 1994
[45] M-H Tao and S L Morrison ldquoStudies of aglycosylatedchimeric mouse-human IgG Role of carbohydrate in thestructure and effector functions mediated by the human IgGconstant regionrdquoThe Journal of Immunology vol 143 no 8 pp2595ndash2601 1989
[46] R Jefferis ldquoGlycosylation of recombinant antibody therapeu-ticsrdquo Biotechnology Progress vol 21 no 1 pp 11ndash16 2005
[47] Y Mimura S Church R Ghirlando et al ldquoThe influence ofglycosylation on the thermal stability and effector functionexpression of human IgG1-Fc properties of a series of truncatedglycoformsrdquoMolecular Immunology vol 37 no 12-13 pp 697ndash706 2001
[48] C Huhn M H J Selman L R Ruhaak A M Deelder and MWuhrer ldquoIgG glycosylation analysisrdquo Proteomics vol 9 no 4pp 882ndash913 2009
[49] J Lund N Takahashi J D Pound M Goodall and R JefferisldquoMultiple interactions of IgG with its core oligosaccharide canmodulate recognition by complement and human Fc gammareceptor I and influence the synthesis of its oligosaccharidechainsrdquo The Journal of Immunology vol 157 no 11 pp 4963ndash4969 1996
[50] R Forastiero M Martinuzzo L O Carreras and J MacloufldquoAnti-1205732 glycoprotein I antibodies and platelet activation inpatients with antiphospholipid antibodies association withincreased excretion of platelet-derived thromboxane urinarymetabolitesrdquoThrombosis and Haemostasis vol 79 no 1 pp 42ndash45 1998
12 Journal of Immunology Research
[51] J-C Reverter and D Tassies ldquoMechanism of thrombosis inantiphospholipid syndrome binding to plateletsrdquo in HughesSyndrome M A Khamashta Ed pp 403ndash414 Springer Lon-don UK 2006
[52] G Espinosa R Cervera J Font and Y Shoenfeld ldquoAntiphos-pholipid syndrome pathogenic mechanismsrdquo AutoimmunityReviews vol 2 no 2 pp 86ndash93 2003
[53] T Shi B Giannakopoulos X Yan et al ldquoAnti-1205732-glycoprotein
I antibodies in complex with 1205732-glycoprotein I can activate
platelets in a dysregulated manner via glycoprotein Ib-IX-VrdquoArthritis amp Rheumatism vol 54 no 8 pp 2558ndash2567 2006
[54] P LMeroni E RaschiMCamera et al ldquoEndothelial activationby aPL a potential pathogenetic mechanism for the clinicalmanifestations of the syndromerdquo Journal of Autoimmunity vol15 no 2 pp 237ndash240 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Immunology Research 7
Table 4 Number (frequency) of seropositive samples of the anti-1205732GP1-IgG subclasses
Number of seropositive samples (gtmOD[NHD] + 2 times SD[NHD])IgG1
(gt29mOD)IgG2
(gt79mOD)IgG3
(gt85mOD)IgG4
(gt319mOD)NHD (119899 = 17) 0 (0) 0 (0) 1 (6) 1 (6)Children (119899 = 13) 13 (100) 2 (15) 8 (62) 2 (15)PAPS (119899 = 12) 6 (50) 5 (42) 0 (0) 1 (8)SAPS (119899 = 6) 1 (17) 2 (33) 0 (0) 1 (17)APS (119899 = 18) 7 (39) 7 (39) 0 (0) 2 (11)OD = optical density 450 nm PAPS = primary antiphospholipid syndrome SAPS = secondary antiphospholipid syndrome APS = PAPS and SAPS NHD =normal healthy donorsThe cut-off for seropositivity was calculated from the mean OD of the corresponding anti-1205732GP1-IgG subclass of the NHD cohort andits standard deviation (SD) according to the formula mean OD[NHD] + 2 times SD[NHD] The respective cut-off OD values are indicated in parentheses below theanti-1205732GP1-IgG subclasses
Table 5 Cohorts studied with the lectin ELISA
Cohort Abbreviation Number of samplesPatients with primary APS PAPS 12Patients with APS and SLE as underlying disease SAPS 10Patients with SLE without symptoms of APS harbouring circulating aPL SLE + aPL 15Asymptomatic carriers of aPL aaPL 6Healthy children with circulating anti-1205732GP1 mdash 16APS = antiphospholipid syndrome aPL = antiphospholipid antibodies SLE = systemic lupus erythematosus
34 Analysis of the Sialylation of Anti-1205732GP1 We analysedthe glycosylation of anti-1205732GP1-IgG focusing on its sialy-lation since highly sialylated Fc-parts of IgG are known tocause rather anti-inflammatory responses [30] Wemeasuredthe binding of the sialic acid specific lectin SNA to the anti-1205732GP1-IgG As shown in Table 5 we employed five cohorts ofindividuals 12 patientswith PAPS 10 patientswith SAPS (SLEas primary disease) 15 patients with SLE who did not showclinical sign of APS but with aPL in their sera (SLE + aPL)6 asymptomatic adult carriers of aPL (aaPL) and 16 healthychildren harbouring aPL
Analysing the SNAIgG-ratios (Figure 4) we identifiedthe highest sialylation level in the sera of the healthy childrenThe significances were rather high and decreased from PAPS(119901 lt 00001) to SAPS (119901 lt 00001) to SLE + aPL (119901 lt00001) and to aaPL (119901 lt 0001)
4 Discussion
We analysed two distinct sets of sera One was used toinvestigate anti-1205732GP1-IgG its subclasses and its associatedproteins C1q and C3c For the analysis of the sialylation ofanti-1205732GP1-IgG we had to collect fresh sera from patientswith APS and the healthy children since the stability of theantibody-glycosylation is unknown In order to avoid itspossible decay by processes like enzymatic degradation thisperiod has been kept as short as possible and was followed byimmediate investigation of the glycosylation
The antiphospholipid syndrome is characterized by clin-ical manifestations such as arterial and venous thrombosisor pregnancy-related complications and the detection of spe-cific autoantibodies (lupus anticoagulant anti-1205732GP1- and
anticardiolipin-autoantibodies) [1] In particular anti-1205732GP1sare considered responsible for provoking the pathologicalcomplications [3]We firstmeasured the total amount of anti-1205732GP1-IgG in the sera of 16 healthy children without clinicalmanifestations of APS and 46 adult patients with APS toidentify individual cohorts seropositive for anti-1205732GP1 17adult normal healthy donors served as negative control
The samples of the patientswithAPS showedhigher levelsof anti-1205732GP1-IgG than those of NHD We also observeda higher concentration of anti-1205732GP1-IgG in the sera ofhealthy children compared to NHD This is due to the factthat the detection of these antibodies has been the inclusioncriterion for the healthy children examined in this studyInterestingly the children showed even higher OD values forthe putatively pathogenic anti-1205732GP1-IgG than the patientswith APS This finding was surprising since the childrenwere clinically healthy and did not show any signs of APSsuch as thrombosis or thrombocytopenia complications inwhich the existence of aPL especially anti-1205732GP1 is maderesponsible for [7 33]
High levels of circulating aPL in 61 healthy children whencompared to healthy adults have already been described byAvcin and colleagues [18] The anti-1205732GP1-IgG was report-edly higher in preschool children with a mean age of 5years than in adolescents with a mean age of 135 years[18] The strongly elevated circulating anti-1205732GP1-IgG in ourcohort of healthy children may be explained by the fact thatthey had an even lower age (mean age was nine months)One possible explanation for this result is the inductionof anti-1205732GP1 by viral and bacterial infections [10 11] ornutritional exposure to 1205732GP1 [17] As the immune systemin children is still to evolve they are more susceptible to
8 Journal of Immunology Research
0
05
10
15
20
25SN
AIg
G
ChildrenPAPS SAPS aaPLSLE + aPL
lowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowast
Figure 4 SNAanti-1205732GP1-IgG ratio in PAPS SAPS SLE + aPLaaPL and children Healthy children show a significantly higherSNAanti-1205732GP1-IgG ratio than PAPS SAPS SLE + aPL and aaPL(lowastlowast119901 lt 00001 lowastlowastlowast119901 lt 000001) This suggests higher sialylation ofthe oligosaccharides attached to Fc fragments of the childrenrsquos anti-1205732GP1No significant differenceswere foundbetween the remainingcohorts Horizontal lines indicate the mean SNAIgG ratio of thecohort SNA = Sambucus nigra agglutinin APS = antiphospholipidsyndrome PAPS = patients with primary APS SAPS = patients withAPS and SLE as underlying disease SLE + aaPL = patients withSLE without symptoms of APS harbouring circulating aPL aaPL =asymptomatic carriers of aPL aPL = antiphospholipid antibodiesSLE = systemic lupus erythematosus
infections than adults and their immune system has to copewith a plethora of viral and bacterial pathogens for the firsttime in their life This scenario is prone to result in thestimulation of autoantibody-production Molecular mimicryof certain pathogens can lead to the production of antibodieswhich then cross-react with 1205732GP1 [7 12ndash15] When infantsdiversify their dietary habits and start to consume productslike milk and meat they can get into contact with nutritional1205732GP1 [19] This is a further condition capable triggeringthe production of anti-1205732GP1 Exposure to pathogen-derivedand dietary 1205732GP1 could explain the high level of circulatinganti-1205732GP1-IgG in healthy childrenThis low affine reactivityis consecutively lost during the maturation of the immunesystem and replaced by the emergence in adults of high affineantibodies that more specifically recognize pathogens Apartfrom possible origins of these autoantibodies it is obviousthat they do not provoke any of the hallmark symptoms ofAPS In contrast to their adult counterparts anti-1205732GP1-IgGscirculating in the plasma of healthy children are obviouslynot pathogenic So it seems decent to hypothesize thatanti-1205732GP1 autoantibodies in healthy children biochemicallydiffer from those in adults
To test this hypothesis we examined the anti-1205732GP1-IgG subclass distribution in the seropositive sera of the anti-1205732GP1-IgG ELISA The healthy children showed higher ODvalues for the IgG1- and IgG3-subclasses than patients withAPS and NHD This was consistent with the number ofseropositive samples In the anti-1205732GP1-IgG1-subclass every
tested serum of the healthy children surpassed the thresholdset for seropositivity (gtmean OD
[NHD] + 2 times SD[NHD]) and
8 samples (62) did so in the IgG3-subclass In the seraof patients with APS just 7 (39) and no samples wereseropositive for IgG1 or IgG3 In the sera of patients with APScompared to NHD we observed higher OD levels of anti-1205732GP1 in all subclasses except for IgG4This result is consis-tent with the findings of Guerin et al who reported elevatedlevels in all anti-1205732GP1-IgG subclasses even including IgG4in patients with APS compared to a control group of healthyadults [23]
It might be that the elevated absolute OD values of theanti-1205732GP1-IgG subclasses are the logical consequence ofthe generally higher level of anti-1205732GP1-IgG in the patientswith APS and healthy children who were selected for furthertesting Therefore we also analysed the subclass distributionof this antibody We first analysed the anti-1205732GP1-IgG sub-class distribution within the individual cohorts and observedthat in the healthy children the IgG3 antibody was mostabundant with a contribution of 40 to the total reactivityThis is consistent with the finding that all sera of the healthychildren were seropositive in this subclass and that theiranti-1205732GP1-IgG3 OD values were the highest of all cohortsThis indicates a predominance of this subclass in the healthychildren The remaining anti-1205732GP1-IgG subclasses in thesera of the children ranged from 15 (IgG1) over 20 (IgG2)to 25 (IgG4) but the differences between their proportionswere not statistically significant In contrast in the sera ofthe patients with APS the anti-1205732GP1-IgG2 subclass had thehighest contribution with 38
The finding of an elevated proportion of the IgG2 subclassin the patients with APS confirms former published datawhich also describe a skewing towards anti-1205732GP1-IgG2 inAPS [19 23 26 27] The authors detected an even highercontribution of anti-1205732GP1-IgG2 (702 [26] and 87 [27]resp) In contrast to our results the IgG1 subclass wasmore prominent in the published studies described aboveImportantly in the cohort of the healthy infants IgG2 wasrelatively lowTherefore onemay argue that anti-1205732GP1-IgG2is not just a marker of the disease but is also involved inthe pathogenic action of the aPL autoantibodies When wecompared the anti-1205732GP1-IgG subclass distribution betweenthe three cohorts we noticed a significant difference in thedistribution concerning the children on one hand and NHDand patients with APS on the other Healthy children showeda higher proportion of anti-1205732GP1-IgG3 and a lower one ofanti-1205732GP1-IgG2 In NHD and patients with APS it was theother way round IgG1 and IgG4 did not significantly differbetween healthy children and patients with APS orNHDThepredominance of anti-1205732GP1-IgG3 in the healthy children isa hint to an immune response elicited by a proteinaceousantigen [26 27 34] This supports the hypothesis thatthe harmless anti-1205732GP1 in children are infection-inducedwith pathogen-derived proteins serving as trigger for theirproduction However it is still elusive if the IgG3-dominatedanti-1205732GP1 response found in the sera of healthy childrencontributes to its low pathogenic potential Alternativelyit can just be ascribed to the proceeding evolution of theinfantrsquos immune system typically associated with an elevated
Journal of Immunology Research 9
proportion of IgG3 antibodies [34 35] During childhoodthe proportion of IgG3 continuously decreases to adult rangesusually reached at the age of 10ndash12 years However the aboveaverage elevated proportion of the anti-1205732GP1-IgG3 (40) inthe sera of the healthy childrenmay thus not just be due to theinfant immune system In particular when bearing in mindthat normal adult levels of the IgG3 subclass are just around6 [34 36]
IgG3 is the IgG-subclass exerting the most effectiveactivation of the complement cascade through the classicalpathway [34] Since complement activation is associated withthe pathogenesis of APS [21 32 37] the high proportionof complement-activating anti-1205732GP1-IgG3 in the childrenseems to contradict a possible protective effect against APS ofthese kinds of antibodies However the role of complement inthe etiopathogenesis of autoimmune diseases is manifold asthe complement components C1q C2 and C4 contribute tothe clearance of postapoptotic secondary necrotic cells andtheir deficiency is therefore a risk factor for autoimmunityand autoinflammation [38] In a similar manner studiesin mouse in vivo models of various autoimmune diseasesdemonstrated that IgG mediated tissue inflammation wasblocked in mice deficient in activating Fc120574Rs although thecomplement component C3 was still abundantly depositedin the tissue [39ndash41] Thus the contribution of complementdeposits in tissue with respect to tissue inflammation remainsto be established
To obtain new insights into the involvement of thecomplement system in the pathogenesis of APS we analysedthe C1q- and C3c-binding to anti-1205732GP1 We observed thatanti-1205732GP1 in the sera of the healthy children and in patientswith APS similarly bound C1q the first molecule of theclassical pathway of complement activation [42] Performinganti-1205732GP1-IgM-ELISAs we detected significantly higherIgM values in the patients with APS One would expecta higher C1q-binding of the IgM positive sera since thisimmunoglobulin binds and activates C1q more strongly thanIgG However the children mainly harbour IgG3 autoanti-bodies the most potent subclass for the activation of theclassical complement pathway [34]This may compensate forthe lower IgM level in their sera and thus be responsiblefor the similar C1q-binding of both cohorts This suggeststhat preconditions for complement activation via the classicalpathway are comparable in both cohorts We conclude thatclassical complement activation has a minor impact on thedifferential pathogenesis of aPL in healthy children andpatients with APS
What then is the culprit that induces their pathogenicpotential
Nextwe quantified the contribution ofC3c to the immunecomplexes bound to 1205732GP1 C3c is a domain in the C3bmolecule and one of its cleavage products created aftercleavage by factor I of C3b [42 43] C3c not just representsa marker of complement activation but also serves as anopsonin improving the clearance of bound targets Weobserved significantly more C3c bound to anti-1205732GP1 inthe sera of the healthy children than in those of patientswith APS This reflects a better opsonization by C3c ofthe potentially harmful anti-1205732GP1 immune complexes
anti-1205732GP1-opsonized cells or cellular fragments in thesera of the healthy children Consequently the anti-1205732GP1complexes can be cleared more efficiently by the powerfulcomplement system of the healthy children In patients withchronic inflammatory autoimmune diseases complement isusually low which may result from null alleles [44] or fromcomplement consumption [38]
But why do these processes not cause inflammation andaberrant coagulation in the childrenApart fromanti-1205732GP1-IgG subclass distribution and distinct ability to activate thecomplement system it is likely that other properties of theanti-1205732GP1 in the sera of children contribute to their lowpathogenicity We hypothesized that an increased sialylationof the anti-1205732GP1-IgG in healthy individuals renders themharmless for their hosts IgG is usually N-glycosylated at theCH2-domain of its Fc-part This N-glycan crucially modifiesthe effector functions the IgG molecule is endowed with[45ndash47] This biantennary core-fucosylated oligosaccharidecarries a variable amount of sugar monomers attached toboth arms [28 48] IgG antibodies terminating with sialicacids on one or both antennae have been shown to behaveanti-inflammatorily after infusion of IVIG [29 31]The signalexerted by the terminal sialic acid is dominant since thefraction of sialylated IgG in healthy plasma is just 4 to 14[28 46 49]
In mice these effects are mainly ascribed to decreasedbinding of sialylated IgG to activating Fc120574-receptors andan upregulation of the inhibitory Fc120574RIIB [29 31] Weobserved that anti-1205732GP1-IgG isolated from sera of healthychildren was significantly more sialylated than that fromsymptomatic patients with APS One may predict that theseautoantibodies in complex with 1205732GP1 show low affinity foractivating Fc120574Rs and therefore do not provoke proinflam-matory and procoagulant effector functions Fc120574RIIA is anexample for an activating receptor It is located on a plethoraof cells On the surfaces of platelets it mediates activationby immune complexes The activation is accompanied byplatelet aggregation and the release of mediators such asthe prothromboticThromboxaneA2 [50] Consecutively thiscascade can result in thrombus formation [51 52] Sialylatedanti-1205732GP1-IgG is unable to bind to and to activate plateletsvia Fc120574RIIA
Further mechanisms involved in thrombus formation inAPS comprise the interaction of anti-1205732GP1 with variouspattern recognition receptors [53] and its interference withregulatory functions ofmolecules involved in the coagulationprocess (protein C prothrombin or tissue factor [52 54])Direct binding to 1205732GP1 on endothelial cells (EC) shiftstheir phenotype towards coagulation and inflammation [54]Considering that Fc120574R-binding is involved in many of thesemechanisms it seems likely that a high sialylation of anti-1205732GP1-IgG leads to a low pathogenicity of these autoan-tibodies and does not cause APS Therefore we concludethat the higher sialylation of anti-1205732GP1-IgG in the seraof the healthy children plays a major role in protectingthem from typical complications of APS This hypothesis isfurther supported by the observation that antiphospholipidantibodies from sera of asymptomatic adult carriers aremore sialylated than those from patients with APS A high
10 Journal of Immunology Research
sialylation status of antiphospholipid antibodies seems toprevent the development of symptoms of APS also in theseindividuals
We conclude that a plethora of factors contributes tothe low pathogenicity of anti-1205732GP1-IgG in the sera ofhealthy children and asymptomatic adults We observed adistinct spectrum of IgG-subclasses compared to patientswith APS and a better binding of C3c indicating a moreefficient clearance of potentially harmful anti-1205732GP1 Thehypersialylation of the anti-1205732GP1-IgG is prone to amelioratetheir inflammatory and procoagulatory effects in healthycarriers whereas hyposialylation and inflammationmutuallyprecipitate the antiphospholipid syndrome
Disclosure
Thepresent workwas performed in fulfillment of the require-ments for obtaining the degree ldquoDr medrdquo for ChristophFickentscher
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Laura Andreoli andMartin Herrmann contributed equally tosenior authorship
Acknowledgments
This project was supported by the German Research Foun-dation (training research programme of the collaborativeresearch center 643) by the Emerging Fields Initiative (EFI)of the FAU Erlangen-Nuremberg and by the K und RWucherpfennig-Stiftung
References
[1] S Miyakis M D Lockshin T Atsumi et al ldquoInternationalconsensus statement on an update of the classification criteriafor definite antiphospholipid syndrome (APS)rdquo Journal ofThrombosis and Haemostasis vol 4 no 2 pp 295ndash306 2006
[2] W A Wilson A E Gharavi T Koike et al ldquoInternationalconsensus statement on preliminary classification criteria fordefinite antiphospholipid syndrome report of an internationalworkshoprdquo Arthritis and Rheumatism vol 42 no 7 pp 1309ndash1311 1999
[3] P G de Groot and R T Urbanus ldquoThe significance of autoan-tibodies against beta2-glycoprotein Irdquo Blood vol 120 no 2 pp266ndash274 2012
[4] C Nalli S Piantoni L Andreoli M Motta and A TincanildquoAntiphospholipid syndrome and antiphospholipid antibodiesin children the two sides of the coinrdquo Israel Medical AssociationJournal vol 14 no 5 pp 310ndash312 2012
[5] I Schousboe ldquo1205732-glycoprotein I a plasma inhibitor of thecontact activation of the intrinsic blood coagulation pathwayrdquoBlood vol 66 no 5 pp 1086ndash1091 1985
[6] J George M Blank Y Levy et al ldquoDifferential effects of anti-1205732-glycoprotein I antibodies on endothelial cells and on themanifestations of experimental antiphospholipid syndromerdquoCirculation vol 97 no 9 pp 900ndash906 1998
[7] R Willis and S S Pierangeli ldquoAnti-1205732-glycoprotein I antibod-iesrdquo Annals of the New York Academy of Sciences vol 1285 no1 pp 44ndash58 2013
[8] M J Fischer J Rauch and J S Levine ldquoThe antiphospholipidsyndromerdquo Seminars in Nephrology vol 27 no 1 pp 35ndash462007
[9] H Amital M Govoni R Maya et al ldquoRole of infectious agentsin systemic rheumatic diseasesrdquo Clinical and ExperimentalRheumatology vol 26 no 1 supplement 48 pp S27ndashS32 2008
[10] T Avcin ldquoAntiphospholipid syndrome in childrenrdquo CurrentOpinion in Rheumatology vol 20 no 5 pp 595ndash600 2008
[11] M Garcıa-Carrasco C Galarza-Maldonado C Mendoza-Pinto R O Escarcega and R Cervera ldquoInfections and theantiphospholipid syndromerdquo Clinical Reviews in Allergy ampImmunology vol 36 no 2-3 pp 104ndash108 2009
[12] P Cruz-Tapias M Blank J-M Anaya and Y ShoenfeldldquoInfections and vaccines in the etiology of antiphospholipidsyndromerdquo Current Opinion in Rheumatology vol 24 no 4 pp389ndash393 2012
[13] P von Landenberg Y Doring S Modrow and K J LacknerldquoAre antiphospholipid antibodies an essential requirement foran effective immune response to infectionsrdquoAnnals of the NewYork Academy of Sciences vol 1108 pp 578ndash583 2007
[14] M Blank I Krause M Fridkin et al ldquoBacterial induction ofautoantibodies to 1205732-glycoprotein-I accounts for the infectiousetiology of antiphospholipid syndromerdquo Journal of ClinicalInvestigation vol 109 no 6 pp 797ndash804 2002
[15] Y Shoenfeld M Blank R Cervera J Font E Raschi and P-LMeroni ldquoInfectious origin of the antiphospholipid syndromerdquoAnnals of the Rheumatic Diseases vol 65 no 1 pp 2ndash6 2006
[16] A E Gharavi S S Pierangeli R G Espinola X Liu MColden-Stanfield and E N Harris ldquoAntiphospholipid antibod-ies induced in mice by immunization with a cytomegalovirus-derived peptide cause thrombosis and activation of endothelialcells in vivordquo Arthritis and Rheumatism vol 46 no 2 pp 545ndash552 2002
[17] M Motta G Chirico C B Rebaioli et al ldquoAnticardiolipin andanti-beta2 glycoprotein I antibodies in infants born to motherswith antiphospholipid antibody-positive autoimmune diseasea follow-up studyrdquoAmerican Journal of Perinatology vol 23 no4 pp 247ndash251 2006
[18] T Avcin A Ambrozic M Kuhar T Kveder and B RozmanldquoAnticardiolipin and anti-1205732 glycoprotein I antibodies in seraof 61 apparently healthy children at regular preventive visitsrdquoRheumatology vol 40 no 5 pp 565ndash573 2001
[19] A Ambrozic ldquoAnti-beta2-glycoprotein I antibodies in children
with atopic dermatitisrdquo International Immunology vol 14 no 7pp 823ndash830
[20] J A Giron-Gonzalez E Garcıa Del Rıo C Rodrıguez JRodrıguez-Martorell and A Serrano ldquoAntiphospholipid syn-drome and asymptomatic carriers of antiphospholipid anti-body prospective analysis of 404 individualsrdquo Journal ofRheumatology vol 31 no 8 pp 1560ndash1567 2004
[21] K A Breen P Seed K Parmar GWMoore S E Stuart-Smithand B J Hunt ldquoComplement activation in patients with isolatedantiphospholipid antibodies or primary antiphospholipid syn-dromerdquo Thrombosis and Haemostasis vol 107 no 3 pp 423ndash429 2012
Journal of Immunology Research 11
[22] F Fischetti P Durigutto V Pellis et al ldquoThrombus formationinduced by antibodies to 1205732-glycoprotein I is complementdependent and requires a priming factorrdquo Blood vol 106 no7 pp 2340ndash2346 2005
[23] J Guerin E Casey C Feighery and J Jackson ldquoAnti-Beta 2-glycoprotein I antibody isotype and IgG subclass in antiphos-pholipid syndrome patientsrdquo Autoimmunity vol 31 no 2 pp109ndash116 1999
[24] L Andreoli C Nalli M Motta et al ldquoAnti-1205732-glycoprotein I
IgG antibodies from 1-year-old healthy children born to moth-ers with systemic autoimmune diseases preferentially targetdomain 45 might it be the reason for their lsquoinnocentrsquo profilerdquoAnnals of the Rheumatic Diseases vol 70 no 2 pp 380ndash3832011
[25] B de Laat R H W M Derksen M van Lummel M T T Pen-nings and P G deGroot ldquoPathogenic anti-beta2-glycoprotein Iantibodies recognize domain I of beta2-glycoprotein I only aftera conformational changerdquo Blood vol 107 no 5 pp 1916ndash19242006
[26] M Samarkos K A Davies C Gordon M J Walport and SLoizou ldquoIgG subclass distribution of antibodies against 1205732-GP1and cardiolipin in patients with systemic lupus erythematosusand primary antiphospholipid syndrome and their clinicalassociationsrdquo Rheumatology vol 40 no 9 pp 1026ndash1032 2001
[27] J Arvieux B Roussel D Ponard and M G Colomb ldquoIgG2subclass restriction of anti-1205732 glycoprotein 1 antibodies inautoimmune patientsrdquo Clinical and Experimental Immunologyvol 95 no 2 pp 310ndash315 1994
[28] J N Arnold M R Wormald R B Sim P M Rudd and R ADwek ldquoThe impact of glycosylation on the biological functionand structure of human immunoglobulinsrdquo Annual Review ofImmunology vol 25 pp 21ndash50 2007
[29] Y Kaneko F Nimmerjahn and J V Ravetch ldquoAnti-inflamma-tory activity of immunoglobulin G resulting from Fc sialyla-tionrdquo Science vol 313 no 5787 pp 670ndash673 2006
[30] RM Anthony and F Nimmerjahn ldquoThe role of differential IgGglycosylation in the interaction of antibodies with FcgammaRsin vivordquo Current Opinion in Organ Transplantation vol 16 no1 pp 7ndash14 2011
[31] R M Anthony and J V Ravetch ldquoA novel role for the IgGFc glycan the anti-inflammatory activity of sialylated IgG FcsrdquoJournal of Clinical Immunology vol 30 no 1 pp S9ndashS14 2010
[32] K Oku T Atsumi M Bohgaki et al ldquoComplement activationin patients with primary antiphospholipid syndromerdquoAnnals ofthe Rheumatic Diseases vol 68 no 6 pp 1030ndash1035 2009
[33] H P McNeil R J Simpson C N Chesterman and S AKrilis ldquoAnti-phospholipid antibodies are directed against acomplex antigen that includes a lipid-binding inhibitor of coag-ulation 1205732-Glycoprotein I (apolipoprotein H)rdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 87 no 11 pp 4120ndash4124 1990
[34] R Jefferis and D S Kumararatne ldquoSelective IgG subclassdeficiency quantification and clinical relevancerdquo Clinical andExperimental Immunology vol 81 no 3 pp 357ndash367 1990
[35] P H Schur F Rosen and M E Norman ldquoImmunoglobulinsubclasses in normal childrenrdquo Pediatric Research vol 13 no3 pp 181ndash183 1979
[36] M A H French and G Harrison ldquoSerum IgG subclass concen-trations in healthy adults a study using monoclonal antiserardquoClinical and Experimental Immunology vol 56 no 2 pp 473ndash475 1984
[37] M Odorczuk L B Keil and V A DeBari ldquoBinding of com-plement component C1q to anti-beta2 glycoprotein I antibodiesfrom patients with antiphospholipid syndromerdquo InflammationResearch vol 48 no 10 pp 524ndash526 1999
[38] L Truedsson A A Bengtsson and G Sturfelt ldquoComplementdeficiencies and systemic lupus erythematosusrdquoAutoimmunityvol 40 no 8 pp 560ndash566 2007
[39] S Aschermann C H K Lehmann S Mihai G Schett DDudziak and F Nimmerjahn ldquoB cells are critical for autoim-mune pathology in Scurfy micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 110 no47 pp 19042ndash19047 2013
[40] Y Kaneko F Nimmerjahn M P Madaio and J V RavetchldquoPathology and protection in nephrotoxic nephritis is deter-mined by selective engagement of specific Fc receptorsrdquo TheJournal of Experimental Medicine vol 203 no 3 pp 789ndash7972006
[41] D Sylvestre R Clynes M Ma H Warren M C Carrolland J V Ravetch ldquoImmunoglobulin G-mediated inflammatoryresponses develop normally in complement-deficient micerdquoJournal of ExperimentalMedicine vol 184 no 6 pp 2385ndash23921996
[42] G E Ritchie B E Moffatt R B Sim B P Morgan R A Dwekand P M Rudd ldquoGlycosylation and the complement systemrdquoChemical Reviews vol 102 no 2 pp 305ndash320 2002
[43] Y Palarasah K Skjodt J Brandt et al ldquoGeneration of aC3c specific monoclonal antibody and assessment of C3c as aputative inflammatorymarker derived from complement factorC3rdquo Journal of ImmunologicalMethods vol 362 no 1-2 pp 142ndash150 2010
[44] D G Doherty J A Underhill P T Donaldson et al ldquoPoly-morphism in the human complement C4 genes and geneticsusceptibility to autoimmune hepatitisrdquo Autoimmunity vol 18no 4 pp 243ndash249 1994
[45] M-H Tao and S L Morrison ldquoStudies of aglycosylatedchimeric mouse-human IgG Role of carbohydrate in thestructure and effector functions mediated by the human IgGconstant regionrdquoThe Journal of Immunology vol 143 no 8 pp2595ndash2601 1989
[46] R Jefferis ldquoGlycosylation of recombinant antibody therapeu-ticsrdquo Biotechnology Progress vol 21 no 1 pp 11ndash16 2005
[47] Y Mimura S Church R Ghirlando et al ldquoThe influence ofglycosylation on the thermal stability and effector functionexpression of human IgG1-Fc properties of a series of truncatedglycoformsrdquoMolecular Immunology vol 37 no 12-13 pp 697ndash706 2001
[48] C Huhn M H J Selman L R Ruhaak A M Deelder and MWuhrer ldquoIgG glycosylation analysisrdquo Proteomics vol 9 no 4pp 882ndash913 2009
[49] J Lund N Takahashi J D Pound M Goodall and R JefferisldquoMultiple interactions of IgG with its core oligosaccharide canmodulate recognition by complement and human Fc gammareceptor I and influence the synthesis of its oligosaccharidechainsrdquo The Journal of Immunology vol 157 no 11 pp 4963ndash4969 1996
[50] R Forastiero M Martinuzzo L O Carreras and J MacloufldquoAnti-1205732 glycoprotein I antibodies and platelet activation inpatients with antiphospholipid antibodies association withincreased excretion of platelet-derived thromboxane urinarymetabolitesrdquoThrombosis and Haemostasis vol 79 no 1 pp 42ndash45 1998
12 Journal of Immunology Research
[51] J-C Reverter and D Tassies ldquoMechanism of thrombosis inantiphospholipid syndrome binding to plateletsrdquo in HughesSyndrome M A Khamashta Ed pp 403ndash414 Springer Lon-don UK 2006
[52] G Espinosa R Cervera J Font and Y Shoenfeld ldquoAntiphos-pholipid syndrome pathogenic mechanismsrdquo AutoimmunityReviews vol 2 no 2 pp 86ndash93 2003
[53] T Shi B Giannakopoulos X Yan et al ldquoAnti-1205732-glycoprotein
I antibodies in complex with 1205732-glycoprotein I can activate
platelets in a dysregulated manner via glycoprotein Ib-IX-VrdquoArthritis amp Rheumatism vol 54 no 8 pp 2558ndash2567 2006
[54] P LMeroni E RaschiMCamera et al ldquoEndothelial activationby aPL a potential pathogenetic mechanism for the clinicalmanifestations of the syndromerdquo Journal of Autoimmunity vol15 no 2 pp 237ndash240 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 Journal of Immunology Research
0
05
10
15
20
25SN
AIg
G
ChildrenPAPS SAPS aaPLSLE + aPL
lowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowast
Figure 4 SNAanti-1205732GP1-IgG ratio in PAPS SAPS SLE + aPLaaPL and children Healthy children show a significantly higherSNAanti-1205732GP1-IgG ratio than PAPS SAPS SLE + aPL and aaPL(lowastlowast119901 lt 00001 lowastlowastlowast119901 lt 000001) This suggests higher sialylation ofthe oligosaccharides attached to Fc fragments of the childrenrsquos anti-1205732GP1No significant differenceswere foundbetween the remainingcohorts Horizontal lines indicate the mean SNAIgG ratio of thecohort SNA = Sambucus nigra agglutinin APS = antiphospholipidsyndrome PAPS = patients with primary APS SAPS = patients withAPS and SLE as underlying disease SLE + aaPL = patients withSLE without symptoms of APS harbouring circulating aPL aaPL =asymptomatic carriers of aPL aPL = antiphospholipid antibodiesSLE = systemic lupus erythematosus
infections than adults and their immune system has to copewith a plethora of viral and bacterial pathogens for the firsttime in their life This scenario is prone to result in thestimulation of autoantibody-production Molecular mimicryof certain pathogens can lead to the production of antibodieswhich then cross-react with 1205732GP1 [7 12ndash15] When infantsdiversify their dietary habits and start to consume productslike milk and meat they can get into contact with nutritional1205732GP1 [19] This is a further condition capable triggeringthe production of anti-1205732GP1 Exposure to pathogen-derivedand dietary 1205732GP1 could explain the high level of circulatinganti-1205732GP1-IgG in healthy childrenThis low affine reactivityis consecutively lost during the maturation of the immunesystem and replaced by the emergence in adults of high affineantibodies that more specifically recognize pathogens Apartfrom possible origins of these autoantibodies it is obviousthat they do not provoke any of the hallmark symptoms ofAPS In contrast to their adult counterparts anti-1205732GP1-IgGscirculating in the plasma of healthy children are obviouslynot pathogenic So it seems decent to hypothesize thatanti-1205732GP1 autoantibodies in healthy children biochemicallydiffer from those in adults
To test this hypothesis we examined the anti-1205732GP1-IgG subclass distribution in the seropositive sera of the anti-1205732GP1-IgG ELISA The healthy children showed higher ODvalues for the IgG1- and IgG3-subclasses than patients withAPS and NHD This was consistent with the number ofseropositive samples In the anti-1205732GP1-IgG1-subclass every
tested serum of the healthy children surpassed the thresholdset for seropositivity (gtmean OD
[NHD] + 2 times SD[NHD]) and
8 samples (62) did so in the IgG3-subclass In the seraof patients with APS just 7 (39) and no samples wereseropositive for IgG1 or IgG3 In the sera of patients with APScompared to NHD we observed higher OD levels of anti-1205732GP1 in all subclasses except for IgG4This result is consis-tent with the findings of Guerin et al who reported elevatedlevels in all anti-1205732GP1-IgG subclasses even including IgG4in patients with APS compared to a control group of healthyadults [23]
It might be that the elevated absolute OD values of theanti-1205732GP1-IgG subclasses are the logical consequence ofthe generally higher level of anti-1205732GP1-IgG in the patientswith APS and healthy children who were selected for furthertesting Therefore we also analysed the subclass distributionof this antibody We first analysed the anti-1205732GP1-IgG sub-class distribution within the individual cohorts and observedthat in the healthy children the IgG3 antibody was mostabundant with a contribution of 40 to the total reactivityThis is consistent with the finding that all sera of the healthychildren were seropositive in this subclass and that theiranti-1205732GP1-IgG3 OD values were the highest of all cohortsThis indicates a predominance of this subclass in the healthychildren The remaining anti-1205732GP1-IgG subclasses in thesera of the children ranged from 15 (IgG1) over 20 (IgG2)to 25 (IgG4) but the differences between their proportionswere not statistically significant In contrast in the sera ofthe patients with APS the anti-1205732GP1-IgG2 subclass had thehighest contribution with 38
The finding of an elevated proportion of the IgG2 subclassin the patients with APS confirms former published datawhich also describe a skewing towards anti-1205732GP1-IgG2 inAPS [19 23 26 27] The authors detected an even highercontribution of anti-1205732GP1-IgG2 (702 [26] and 87 [27]resp) In contrast to our results the IgG1 subclass wasmore prominent in the published studies described aboveImportantly in the cohort of the healthy infants IgG2 wasrelatively lowTherefore onemay argue that anti-1205732GP1-IgG2is not just a marker of the disease but is also involved inthe pathogenic action of the aPL autoantibodies When wecompared the anti-1205732GP1-IgG subclass distribution betweenthe three cohorts we noticed a significant difference in thedistribution concerning the children on one hand and NHDand patients with APS on the other Healthy children showeda higher proportion of anti-1205732GP1-IgG3 and a lower one ofanti-1205732GP1-IgG2 In NHD and patients with APS it was theother way round IgG1 and IgG4 did not significantly differbetween healthy children and patients with APS orNHDThepredominance of anti-1205732GP1-IgG3 in the healthy children isa hint to an immune response elicited by a proteinaceousantigen [26 27 34] This supports the hypothesis thatthe harmless anti-1205732GP1 in children are infection-inducedwith pathogen-derived proteins serving as trigger for theirproduction However it is still elusive if the IgG3-dominatedanti-1205732GP1 response found in the sera of healthy childrencontributes to its low pathogenic potential Alternativelyit can just be ascribed to the proceeding evolution of theinfantrsquos immune system typically associated with an elevated
Journal of Immunology Research 9
proportion of IgG3 antibodies [34 35] During childhoodthe proportion of IgG3 continuously decreases to adult rangesusually reached at the age of 10ndash12 years However the aboveaverage elevated proportion of the anti-1205732GP1-IgG3 (40) inthe sera of the healthy childrenmay thus not just be due to theinfant immune system In particular when bearing in mindthat normal adult levels of the IgG3 subclass are just around6 [34 36]
IgG3 is the IgG-subclass exerting the most effectiveactivation of the complement cascade through the classicalpathway [34] Since complement activation is associated withthe pathogenesis of APS [21 32 37] the high proportionof complement-activating anti-1205732GP1-IgG3 in the childrenseems to contradict a possible protective effect against APS ofthese kinds of antibodies However the role of complement inthe etiopathogenesis of autoimmune diseases is manifold asthe complement components C1q C2 and C4 contribute tothe clearance of postapoptotic secondary necrotic cells andtheir deficiency is therefore a risk factor for autoimmunityand autoinflammation [38] In a similar manner studiesin mouse in vivo models of various autoimmune diseasesdemonstrated that IgG mediated tissue inflammation wasblocked in mice deficient in activating Fc120574Rs although thecomplement component C3 was still abundantly depositedin the tissue [39ndash41] Thus the contribution of complementdeposits in tissue with respect to tissue inflammation remainsto be established
To obtain new insights into the involvement of thecomplement system in the pathogenesis of APS we analysedthe C1q- and C3c-binding to anti-1205732GP1 We observed thatanti-1205732GP1 in the sera of the healthy children and in patientswith APS similarly bound C1q the first molecule of theclassical pathway of complement activation [42] Performinganti-1205732GP1-IgM-ELISAs we detected significantly higherIgM values in the patients with APS One would expecta higher C1q-binding of the IgM positive sera since thisimmunoglobulin binds and activates C1q more strongly thanIgG However the children mainly harbour IgG3 autoanti-bodies the most potent subclass for the activation of theclassical complement pathway [34]This may compensate forthe lower IgM level in their sera and thus be responsiblefor the similar C1q-binding of both cohorts This suggeststhat preconditions for complement activation via the classicalpathway are comparable in both cohorts We conclude thatclassical complement activation has a minor impact on thedifferential pathogenesis of aPL in healthy children andpatients with APS
What then is the culprit that induces their pathogenicpotential
Nextwe quantified the contribution ofC3c to the immunecomplexes bound to 1205732GP1 C3c is a domain in the C3bmolecule and one of its cleavage products created aftercleavage by factor I of C3b [42 43] C3c not just representsa marker of complement activation but also serves as anopsonin improving the clearance of bound targets Weobserved significantly more C3c bound to anti-1205732GP1 inthe sera of the healthy children than in those of patientswith APS This reflects a better opsonization by C3c ofthe potentially harmful anti-1205732GP1 immune complexes
anti-1205732GP1-opsonized cells or cellular fragments in thesera of the healthy children Consequently the anti-1205732GP1complexes can be cleared more efficiently by the powerfulcomplement system of the healthy children In patients withchronic inflammatory autoimmune diseases complement isusually low which may result from null alleles [44] or fromcomplement consumption [38]
But why do these processes not cause inflammation andaberrant coagulation in the childrenApart fromanti-1205732GP1-IgG subclass distribution and distinct ability to activate thecomplement system it is likely that other properties of theanti-1205732GP1 in the sera of children contribute to their lowpathogenicity We hypothesized that an increased sialylationof the anti-1205732GP1-IgG in healthy individuals renders themharmless for their hosts IgG is usually N-glycosylated at theCH2-domain of its Fc-part This N-glycan crucially modifiesthe effector functions the IgG molecule is endowed with[45ndash47] This biantennary core-fucosylated oligosaccharidecarries a variable amount of sugar monomers attached toboth arms [28 48] IgG antibodies terminating with sialicacids on one or both antennae have been shown to behaveanti-inflammatorily after infusion of IVIG [29 31]The signalexerted by the terminal sialic acid is dominant since thefraction of sialylated IgG in healthy plasma is just 4 to 14[28 46 49]
In mice these effects are mainly ascribed to decreasedbinding of sialylated IgG to activating Fc120574-receptors andan upregulation of the inhibitory Fc120574RIIB [29 31] Weobserved that anti-1205732GP1-IgG isolated from sera of healthychildren was significantly more sialylated than that fromsymptomatic patients with APS One may predict that theseautoantibodies in complex with 1205732GP1 show low affinity foractivating Fc120574Rs and therefore do not provoke proinflam-matory and procoagulant effector functions Fc120574RIIA is anexample for an activating receptor It is located on a plethoraof cells On the surfaces of platelets it mediates activationby immune complexes The activation is accompanied byplatelet aggregation and the release of mediators such asthe prothromboticThromboxaneA2 [50] Consecutively thiscascade can result in thrombus formation [51 52] Sialylatedanti-1205732GP1-IgG is unable to bind to and to activate plateletsvia Fc120574RIIA
Further mechanisms involved in thrombus formation inAPS comprise the interaction of anti-1205732GP1 with variouspattern recognition receptors [53] and its interference withregulatory functions ofmolecules involved in the coagulationprocess (protein C prothrombin or tissue factor [52 54])Direct binding to 1205732GP1 on endothelial cells (EC) shiftstheir phenotype towards coagulation and inflammation [54]Considering that Fc120574R-binding is involved in many of thesemechanisms it seems likely that a high sialylation of anti-1205732GP1-IgG leads to a low pathogenicity of these autoan-tibodies and does not cause APS Therefore we concludethat the higher sialylation of anti-1205732GP1-IgG in the seraof the healthy children plays a major role in protectingthem from typical complications of APS This hypothesis isfurther supported by the observation that antiphospholipidantibodies from sera of asymptomatic adult carriers aremore sialylated than those from patients with APS A high
10 Journal of Immunology Research
sialylation status of antiphospholipid antibodies seems toprevent the development of symptoms of APS also in theseindividuals
We conclude that a plethora of factors contributes tothe low pathogenicity of anti-1205732GP1-IgG in the sera ofhealthy children and asymptomatic adults We observed adistinct spectrum of IgG-subclasses compared to patientswith APS and a better binding of C3c indicating a moreefficient clearance of potentially harmful anti-1205732GP1 Thehypersialylation of the anti-1205732GP1-IgG is prone to amelioratetheir inflammatory and procoagulatory effects in healthycarriers whereas hyposialylation and inflammationmutuallyprecipitate the antiphospholipid syndrome
Disclosure
Thepresent workwas performed in fulfillment of the require-ments for obtaining the degree ldquoDr medrdquo for ChristophFickentscher
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Laura Andreoli andMartin Herrmann contributed equally tosenior authorship
Acknowledgments
This project was supported by the German Research Foun-dation (training research programme of the collaborativeresearch center 643) by the Emerging Fields Initiative (EFI)of the FAU Erlangen-Nuremberg and by the K und RWucherpfennig-Stiftung
References
[1] S Miyakis M D Lockshin T Atsumi et al ldquoInternationalconsensus statement on an update of the classification criteriafor definite antiphospholipid syndrome (APS)rdquo Journal ofThrombosis and Haemostasis vol 4 no 2 pp 295ndash306 2006
[2] W A Wilson A E Gharavi T Koike et al ldquoInternationalconsensus statement on preliminary classification criteria fordefinite antiphospholipid syndrome report of an internationalworkshoprdquo Arthritis and Rheumatism vol 42 no 7 pp 1309ndash1311 1999
[3] P G de Groot and R T Urbanus ldquoThe significance of autoan-tibodies against beta2-glycoprotein Irdquo Blood vol 120 no 2 pp266ndash274 2012
[4] C Nalli S Piantoni L Andreoli M Motta and A TincanildquoAntiphospholipid syndrome and antiphospholipid antibodiesin children the two sides of the coinrdquo Israel Medical AssociationJournal vol 14 no 5 pp 310ndash312 2012
[5] I Schousboe ldquo1205732-glycoprotein I a plasma inhibitor of thecontact activation of the intrinsic blood coagulation pathwayrdquoBlood vol 66 no 5 pp 1086ndash1091 1985
[6] J George M Blank Y Levy et al ldquoDifferential effects of anti-1205732-glycoprotein I antibodies on endothelial cells and on themanifestations of experimental antiphospholipid syndromerdquoCirculation vol 97 no 9 pp 900ndash906 1998
[7] R Willis and S S Pierangeli ldquoAnti-1205732-glycoprotein I antibod-iesrdquo Annals of the New York Academy of Sciences vol 1285 no1 pp 44ndash58 2013
[8] M J Fischer J Rauch and J S Levine ldquoThe antiphospholipidsyndromerdquo Seminars in Nephrology vol 27 no 1 pp 35ndash462007
[9] H Amital M Govoni R Maya et al ldquoRole of infectious agentsin systemic rheumatic diseasesrdquo Clinical and ExperimentalRheumatology vol 26 no 1 supplement 48 pp S27ndashS32 2008
[10] T Avcin ldquoAntiphospholipid syndrome in childrenrdquo CurrentOpinion in Rheumatology vol 20 no 5 pp 595ndash600 2008
[11] M Garcıa-Carrasco C Galarza-Maldonado C Mendoza-Pinto R O Escarcega and R Cervera ldquoInfections and theantiphospholipid syndromerdquo Clinical Reviews in Allergy ampImmunology vol 36 no 2-3 pp 104ndash108 2009
[12] P Cruz-Tapias M Blank J-M Anaya and Y ShoenfeldldquoInfections and vaccines in the etiology of antiphospholipidsyndromerdquo Current Opinion in Rheumatology vol 24 no 4 pp389ndash393 2012
[13] P von Landenberg Y Doring S Modrow and K J LacknerldquoAre antiphospholipid antibodies an essential requirement foran effective immune response to infectionsrdquoAnnals of the NewYork Academy of Sciences vol 1108 pp 578ndash583 2007
[14] M Blank I Krause M Fridkin et al ldquoBacterial induction ofautoantibodies to 1205732-glycoprotein-I accounts for the infectiousetiology of antiphospholipid syndromerdquo Journal of ClinicalInvestigation vol 109 no 6 pp 797ndash804 2002
[15] Y Shoenfeld M Blank R Cervera J Font E Raschi and P-LMeroni ldquoInfectious origin of the antiphospholipid syndromerdquoAnnals of the Rheumatic Diseases vol 65 no 1 pp 2ndash6 2006
[16] A E Gharavi S S Pierangeli R G Espinola X Liu MColden-Stanfield and E N Harris ldquoAntiphospholipid antibod-ies induced in mice by immunization with a cytomegalovirus-derived peptide cause thrombosis and activation of endothelialcells in vivordquo Arthritis and Rheumatism vol 46 no 2 pp 545ndash552 2002
[17] M Motta G Chirico C B Rebaioli et al ldquoAnticardiolipin andanti-beta2 glycoprotein I antibodies in infants born to motherswith antiphospholipid antibody-positive autoimmune diseasea follow-up studyrdquoAmerican Journal of Perinatology vol 23 no4 pp 247ndash251 2006
[18] T Avcin A Ambrozic M Kuhar T Kveder and B RozmanldquoAnticardiolipin and anti-1205732 glycoprotein I antibodies in seraof 61 apparently healthy children at regular preventive visitsrdquoRheumatology vol 40 no 5 pp 565ndash573 2001
[19] A Ambrozic ldquoAnti-beta2-glycoprotein I antibodies in children
with atopic dermatitisrdquo International Immunology vol 14 no 7pp 823ndash830
[20] J A Giron-Gonzalez E Garcıa Del Rıo C Rodrıguez JRodrıguez-Martorell and A Serrano ldquoAntiphospholipid syn-drome and asymptomatic carriers of antiphospholipid anti-body prospective analysis of 404 individualsrdquo Journal ofRheumatology vol 31 no 8 pp 1560ndash1567 2004
[21] K A Breen P Seed K Parmar GWMoore S E Stuart-Smithand B J Hunt ldquoComplement activation in patients with isolatedantiphospholipid antibodies or primary antiphospholipid syn-dromerdquo Thrombosis and Haemostasis vol 107 no 3 pp 423ndash429 2012
Journal of Immunology Research 11
[22] F Fischetti P Durigutto V Pellis et al ldquoThrombus formationinduced by antibodies to 1205732-glycoprotein I is complementdependent and requires a priming factorrdquo Blood vol 106 no7 pp 2340ndash2346 2005
[23] J Guerin E Casey C Feighery and J Jackson ldquoAnti-Beta 2-glycoprotein I antibody isotype and IgG subclass in antiphos-pholipid syndrome patientsrdquo Autoimmunity vol 31 no 2 pp109ndash116 1999
[24] L Andreoli C Nalli M Motta et al ldquoAnti-1205732-glycoprotein I
IgG antibodies from 1-year-old healthy children born to moth-ers with systemic autoimmune diseases preferentially targetdomain 45 might it be the reason for their lsquoinnocentrsquo profilerdquoAnnals of the Rheumatic Diseases vol 70 no 2 pp 380ndash3832011
[25] B de Laat R H W M Derksen M van Lummel M T T Pen-nings and P G deGroot ldquoPathogenic anti-beta2-glycoprotein Iantibodies recognize domain I of beta2-glycoprotein I only aftera conformational changerdquo Blood vol 107 no 5 pp 1916ndash19242006
[26] M Samarkos K A Davies C Gordon M J Walport and SLoizou ldquoIgG subclass distribution of antibodies against 1205732-GP1and cardiolipin in patients with systemic lupus erythematosusand primary antiphospholipid syndrome and their clinicalassociationsrdquo Rheumatology vol 40 no 9 pp 1026ndash1032 2001
[27] J Arvieux B Roussel D Ponard and M G Colomb ldquoIgG2subclass restriction of anti-1205732 glycoprotein 1 antibodies inautoimmune patientsrdquo Clinical and Experimental Immunologyvol 95 no 2 pp 310ndash315 1994
[28] J N Arnold M R Wormald R B Sim P M Rudd and R ADwek ldquoThe impact of glycosylation on the biological functionand structure of human immunoglobulinsrdquo Annual Review ofImmunology vol 25 pp 21ndash50 2007
[29] Y Kaneko F Nimmerjahn and J V Ravetch ldquoAnti-inflamma-tory activity of immunoglobulin G resulting from Fc sialyla-tionrdquo Science vol 313 no 5787 pp 670ndash673 2006
[30] RM Anthony and F Nimmerjahn ldquoThe role of differential IgGglycosylation in the interaction of antibodies with FcgammaRsin vivordquo Current Opinion in Organ Transplantation vol 16 no1 pp 7ndash14 2011
[31] R M Anthony and J V Ravetch ldquoA novel role for the IgGFc glycan the anti-inflammatory activity of sialylated IgG FcsrdquoJournal of Clinical Immunology vol 30 no 1 pp S9ndashS14 2010
[32] K Oku T Atsumi M Bohgaki et al ldquoComplement activationin patients with primary antiphospholipid syndromerdquoAnnals ofthe Rheumatic Diseases vol 68 no 6 pp 1030ndash1035 2009
[33] H P McNeil R J Simpson C N Chesterman and S AKrilis ldquoAnti-phospholipid antibodies are directed against acomplex antigen that includes a lipid-binding inhibitor of coag-ulation 1205732-Glycoprotein I (apolipoprotein H)rdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 87 no 11 pp 4120ndash4124 1990
[34] R Jefferis and D S Kumararatne ldquoSelective IgG subclassdeficiency quantification and clinical relevancerdquo Clinical andExperimental Immunology vol 81 no 3 pp 357ndash367 1990
[35] P H Schur F Rosen and M E Norman ldquoImmunoglobulinsubclasses in normal childrenrdquo Pediatric Research vol 13 no3 pp 181ndash183 1979
[36] M A H French and G Harrison ldquoSerum IgG subclass concen-trations in healthy adults a study using monoclonal antiserardquoClinical and Experimental Immunology vol 56 no 2 pp 473ndash475 1984
[37] M Odorczuk L B Keil and V A DeBari ldquoBinding of com-plement component C1q to anti-beta2 glycoprotein I antibodiesfrom patients with antiphospholipid syndromerdquo InflammationResearch vol 48 no 10 pp 524ndash526 1999
[38] L Truedsson A A Bengtsson and G Sturfelt ldquoComplementdeficiencies and systemic lupus erythematosusrdquoAutoimmunityvol 40 no 8 pp 560ndash566 2007
[39] S Aschermann C H K Lehmann S Mihai G Schett DDudziak and F Nimmerjahn ldquoB cells are critical for autoim-mune pathology in Scurfy micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 110 no47 pp 19042ndash19047 2013
[40] Y Kaneko F Nimmerjahn M P Madaio and J V RavetchldquoPathology and protection in nephrotoxic nephritis is deter-mined by selective engagement of specific Fc receptorsrdquo TheJournal of Experimental Medicine vol 203 no 3 pp 789ndash7972006
[41] D Sylvestre R Clynes M Ma H Warren M C Carrolland J V Ravetch ldquoImmunoglobulin G-mediated inflammatoryresponses develop normally in complement-deficient micerdquoJournal of ExperimentalMedicine vol 184 no 6 pp 2385ndash23921996
[42] G E Ritchie B E Moffatt R B Sim B P Morgan R A Dwekand P M Rudd ldquoGlycosylation and the complement systemrdquoChemical Reviews vol 102 no 2 pp 305ndash320 2002
[43] Y Palarasah K Skjodt J Brandt et al ldquoGeneration of aC3c specific monoclonal antibody and assessment of C3c as aputative inflammatorymarker derived from complement factorC3rdquo Journal of ImmunologicalMethods vol 362 no 1-2 pp 142ndash150 2010
[44] D G Doherty J A Underhill P T Donaldson et al ldquoPoly-morphism in the human complement C4 genes and geneticsusceptibility to autoimmune hepatitisrdquo Autoimmunity vol 18no 4 pp 243ndash249 1994
[45] M-H Tao and S L Morrison ldquoStudies of aglycosylatedchimeric mouse-human IgG Role of carbohydrate in thestructure and effector functions mediated by the human IgGconstant regionrdquoThe Journal of Immunology vol 143 no 8 pp2595ndash2601 1989
[46] R Jefferis ldquoGlycosylation of recombinant antibody therapeu-ticsrdquo Biotechnology Progress vol 21 no 1 pp 11ndash16 2005
[47] Y Mimura S Church R Ghirlando et al ldquoThe influence ofglycosylation on the thermal stability and effector functionexpression of human IgG1-Fc properties of a series of truncatedglycoformsrdquoMolecular Immunology vol 37 no 12-13 pp 697ndash706 2001
[48] C Huhn M H J Selman L R Ruhaak A M Deelder and MWuhrer ldquoIgG glycosylation analysisrdquo Proteomics vol 9 no 4pp 882ndash913 2009
[49] J Lund N Takahashi J D Pound M Goodall and R JefferisldquoMultiple interactions of IgG with its core oligosaccharide canmodulate recognition by complement and human Fc gammareceptor I and influence the synthesis of its oligosaccharidechainsrdquo The Journal of Immunology vol 157 no 11 pp 4963ndash4969 1996
[50] R Forastiero M Martinuzzo L O Carreras and J MacloufldquoAnti-1205732 glycoprotein I antibodies and platelet activation inpatients with antiphospholipid antibodies association withincreased excretion of platelet-derived thromboxane urinarymetabolitesrdquoThrombosis and Haemostasis vol 79 no 1 pp 42ndash45 1998
12 Journal of Immunology Research
[51] J-C Reverter and D Tassies ldquoMechanism of thrombosis inantiphospholipid syndrome binding to plateletsrdquo in HughesSyndrome M A Khamashta Ed pp 403ndash414 Springer Lon-don UK 2006
[52] G Espinosa R Cervera J Font and Y Shoenfeld ldquoAntiphos-pholipid syndrome pathogenic mechanismsrdquo AutoimmunityReviews vol 2 no 2 pp 86ndash93 2003
[53] T Shi B Giannakopoulos X Yan et al ldquoAnti-1205732-glycoprotein
I antibodies in complex with 1205732-glycoprotein I can activate
platelets in a dysregulated manner via glycoprotein Ib-IX-VrdquoArthritis amp Rheumatism vol 54 no 8 pp 2558ndash2567 2006
[54] P LMeroni E RaschiMCamera et al ldquoEndothelial activationby aPL a potential pathogenetic mechanism for the clinicalmanifestations of the syndromerdquo Journal of Autoimmunity vol15 no 2 pp 237ndash240 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Immunology Research 9
proportion of IgG3 antibodies [34 35] During childhoodthe proportion of IgG3 continuously decreases to adult rangesusually reached at the age of 10ndash12 years However the aboveaverage elevated proportion of the anti-1205732GP1-IgG3 (40) inthe sera of the healthy childrenmay thus not just be due to theinfant immune system In particular when bearing in mindthat normal adult levels of the IgG3 subclass are just around6 [34 36]
IgG3 is the IgG-subclass exerting the most effectiveactivation of the complement cascade through the classicalpathway [34] Since complement activation is associated withthe pathogenesis of APS [21 32 37] the high proportionof complement-activating anti-1205732GP1-IgG3 in the childrenseems to contradict a possible protective effect against APS ofthese kinds of antibodies However the role of complement inthe etiopathogenesis of autoimmune diseases is manifold asthe complement components C1q C2 and C4 contribute tothe clearance of postapoptotic secondary necrotic cells andtheir deficiency is therefore a risk factor for autoimmunityand autoinflammation [38] In a similar manner studiesin mouse in vivo models of various autoimmune diseasesdemonstrated that IgG mediated tissue inflammation wasblocked in mice deficient in activating Fc120574Rs although thecomplement component C3 was still abundantly depositedin the tissue [39ndash41] Thus the contribution of complementdeposits in tissue with respect to tissue inflammation remainsto be established
To obtain new insights into the involvement of thecomplement system in the pathogenesis of APS we analysedthe C1q- and C3c-binding to anti-1205732GP1 We observed thatanti-1205732GP1 in the sera of the healthy children and in patientswith APS similarly bound C1q the first molecule of theclassical pathway of complement activation [42] Performinganti-1205732GP1-IgM-ELISAs we detected significantly higherIgM values in the patients with APS One would expecta higher C1q-binding of the IgM positive sera since thisimmunoglobulin binds and activates C1q more strongly thanIgG However the children mainly harbour IgG3 autoanti-bodies the most potent subclass for the activation of theclassical complement pathway [34]This may compensate forthe lower IgM level in their sera and thus be responsiblefor the similar C1q-binding of both cohorts This suggeststhat preconditions for complement activation via the classicalpathway are comparable in both cohorts We conclude thatclassical complement activation has a minor impact on thedifferential pathogenesis of aPL in healthy children andpatients with APS
What then is the culprit that induces their pathogenicpotential
Nextwe quantified the contribution ofC3c to the immunecomplexes bound to 1205732GP1 C3c is a domain in the C3bmolecule and one of its cleavage products created aftercleavage by factor I of C3b [42 43] C3c not just representsa marker of complement activation but also serves as anopsonin improving the clearance of bound targets Weobserved significantly more C3c bound to anti-1205732GP1 inthe sera of the healthy children than in those of patientswith APS This reflects a better opsonization by C3c ofthe potentially harmful anti-1205732GP1 immune complexes
anti-1205732GP1-opsonized cells or cellular fragments in thesera of the healthy children Consequently the anti-1205732GP1complexes can be cleared more efficiently by the powerfulcomplement system of the healthy children In patients withchronic inflammatory autoimmune diseases complement isusually low which may result from null alleles [44] or fromcomplement consumption [38]
But why do these processes not cause inflammation andaberrant coagulation in the childrenApart fromanti-1205732GP1-IgG subclass distribution and distinct ability to activate thecomplement system it is likely that other properties of theanti-1205732GP1 in the sera of children contribute to their lowpathogenicity We hypothesized that an increased sialylationof the anti-1205732GP1-IgG in healthy individuals renders themharmless for their hosts IgG is usually N-glycosylated at theCH2-domain of its Fc-part This N-glycan crucially modifiesthe effector functions the IgG molecule is endowed with[45ndash47] This biantennary core-fucosylated oligosaccharidecarries a variable amount of sugar monomers attached toboth arms [28 48] IgG antibodies terminating with sialicacids on one or both antennae have been shown to behaveanti-inflammatorily after infusion of IVIG [29 31]The signalexerted by the terminal sialic acid is dominant since thefraction of sialylated IgG in healthy plasma is just 4 to 14[28 46 49]
In mice these effects are mainly ascribed to decreasedbinding of sialylated IgG to activating Fc120574-receptors andan upregulation of the inhibitory Fc120574RIIB [29 31] Weobserved that anti-1205732GP1-IgG isolated from sera of healthychildren was significantly more sialylated than that fromsymptomatic patients with APS One may predict that theseautoantibodies in complex with 1205732GP1 show low affinity foractivating Fc120574Rs and therefore do not provoke proinflam-matory and procoagulant effector functions Fc120574RIIA is anexample for an activating receptor It is located on a plethoraof cells On the surfaces of platelets it mediates activationby immune complexes The activation is accompanied byplatelet aggregation and the release of mediators such asthe prothromboticThromboxaneA2 [50] Consecutively thiscascade can result in thrombus formation [51 52] Sialylatedanti-1205732GP1-IgG is unable to bind to and to activate plateletsvia Fc120574RIIA
Further mechanisms involved in thrombus formation inAPS comprise the interaction of anti-1205732GP1 with variouspattern recognition receptors [53] and its interference withregulatory functions ofmolecules involved in the coagulationprocess (protein C prothrombin or tissue factor [52 54])Direct binding to 1205732GP1 on endothelial cells (EC) shiftstheir phenotype towards coagulation and inflammation [54]Considering that Fc120574R-binding is involved in many of thesemechanisms it seems likely that a high sialylation of anti-1205732GP1-IgG leads to a low pathogenicity of these autoan-tibodies and does not cause APS Therefore we concludethat the higher sialylation of anti-1205732GP1-IgG in the seraof the healthy children plays a major role in protectingthem from typical complications of APS This hypothesis isfurther supported by the observation that antiphospholipidantibodies from sera of asymptomatic adult carriers aremore sialylated than those from patients with APS A high
10 Journal of Immunology Research
sialylation status of antiphospholipid antibodies seems toprevent the development of symptoms of APS also in theseindividuals
We conclude that a plethora of factors contributes tothe low pathogenicity of anti-1205732GP1-IgG in the sera ofhealthy children and asymptomatic adults We observed adistinct spectrum of IgG-subclasses compared to patientswith APS and a better binding of C3c indicating a moreefficient clearance of potentially harmful anti-1205732GP1 Thehypersialylation of the anti-1205732GP1-IgG is prone to amelioratetheir inflammatory and procoagulatory effects in healthycarriers whereas hyposialylation and inflammationmutuallyprecipitate the antiphospholipid syndrome
Disclosure
Thepresent workwas performed in fulfillment of the require-ments for obtaining the degree ldquoDr medrdquo for ChristophFickentscher
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Laura Andreoli andMartin Herrmann contributed equally tosenior authorship
Acknowledgments
This project was supported by the German Research Foun-dation (training research programme of the collaborativeresearch center 643) by the Emerging Fields Initiative (EFI)of the FAU Erlangen-Nuremberg and by the K und RWucherpfennig-Stiftung
References
[1] S Miyakis M D Lockshin T Atsumi et al ldquoInternationalconsensus statement on an update of the classification criteriafor definite antiphospholipid syndrome (APS)rdquo Journal ofThrombosis and Haemostasis vol 4 no 2 pp 295ndash306 2006
[2] W A Wilson A E Gharavi T Koike et al ldquoInternationalconsensus statement on preliminary classification criteria fordefinite antiphospholipid syndrome report of an internationalworkshoprdquo Arthritis and Rheumatism vol 42 no 7 pp 1309ndash1311 1999
[3] P G de Groot and R T Urbanus ldquoThe significance of autoan-tibodies against beta2-glycoprotein Irdquo Blood vol 120 no 2 pp266ndash274 2012
[4] C Nalli S Piantoni L Andreoli M Motta and A TincanildquoAntiphospholipid syndrome and antiphospholipid antibodiesin children the two sides of the coinrdquo Israel Medical AssociationJournal vol 14 no 5 pp 310ndash312 2012
[5] I Schousboe ldquo1205732-glycoprotein I a plasma inhibitor of thecontact activation of the intrinsic blood coagulation pathwayrdquoBlood vol 66 no 5 pp 1086ndash1091 1985
[6] J George M Blank Y Levy et al ldquoDifferential effects of anti-1205732-glycoprotein I antibodies on endothelial cells and on themanifestations of experimental antiphospholipid syndromerdquoCirculation vol 97 no 9 pp 900ndash906 1998
[7] R Willis and S S Pierangeli ldquoAnti-1205732-glycoprotein I antibod-iesrdquo Annals of the New York Academy of Sciences vol 1285 no1 pp 44ndash58 2013
[8] M J Fischer J Rauch and J S Levine ldquoThe antiphospholipidsyndromerdquo Seminars in Nephrology vol 27 no 1 pp 35ndash462007
[9] H Amital M Govoni R Maya et al ldquoRole of infectious agentsin systemic rheumatic diseasesrdquo Clinical and ExperimentalRheumatology vol 26 no 1 supplement 48 pp S27ndashS32 2008
[10] T Avcin ldquoAntiphospholipid syndrome in childrenrdquo CurrentOpinion in Rheumatology vol 20 no 5 pp 595ndash600 2008
[11] M Garcıa-Carrasco C Galarza-Maldonado C Mendoza-Pinto R O Escarcega and R Cervera ldquoInfections and theantiphospholipid syndromerdquo Clinical Reviews in Allergy ampImmunology vol 36 no 2-3 pp 104ndash108 2009
[12] P Cruz-Tapias M Blank J-M Anaya and Y ShoenfeldldquoInfections and vaccines in the etiology of antiphospholipidsyndromerdquo Current Opinion in Rheumatology vol 24 no 4 pp389ndash393 2012
[13] P von Landenberg Y Doring S Modrow and K J LacknerldquoAre antiphospholipid antibodies an essential requirement foran effective immune response to infectionsrdquoAnnals of the NewYork Academy of Sciences vol 1108 pp 578ndash583 2007
[14] M Blank I Krause M Fridkin et al ldquoBacterial induction ofautoantibodies to 1205732-glycoprotein-I accounts for the infectiousetiology of antiphospholipid syndromerdquo Journal of ClinicalInvestigation vol 109 no 6 pp 797ndash804 2002
[15] Y Shoenfeld M Blank R Cervera J Font E Raschi and P-LMeroni ldquoInfectious origin of the antiphospholipid syndromerdquoAnnals of the Rheumatic Diseases vol 65 no 1 pp 2ndash6 2006
[16] A E Gharavi S S Pierangeli R G Espinola X Liu MColden-Stanfield and E N Harris ldquoAntiphospholipid antibod-ies induced in mice by immunization with a cytomegalovirus-derived peptide cause thrombosis and activation of endothelialcells in vivordquo Arthritis and Rheumatism vol 46 no 2 pp 545ndash552 2002
[17] M Motta G Chirico C B Rebaioli et al ldquoAnticardiolipin andanti-beta2 glycoprotein I antibodies in infants born to motherswith antiphospholipid antibody-positive autoimmune diseasea follow-up studyrdquoAmerican Journal of Perinatology vol 23 no4 pp 247ndash251 2006
[18] T Avcin A Ambrozic M Kuhar T Kveder and B RozmanldquoAnticardiolipin and anti-1205732 glycoprotein I antibodies in seraof 61 apparently healthy children at regular preventive visitsrdquoRheumatology vol 40 no 5 pp 565ndash573 2001
[19] A Ambrozic ldquoAnti-beta2-glycoprotein I antibodies in children
with atopic dermatitisrdquo International Immunology vol 14 no 7pp 823ndash830
[20] J A Giron-Gonzalez E Garcıa Del Rıo C Rodrıguez JRodrıguez-Martorell and A Serrano ldquoAntiphospholipid syn-drome and asymptomatic carriers of antiphospholipid anti-body prospective analysis of 404 individualsrdquo Journal ofRheumatology vol 31 no 8 pp 1560ndash1567 2004
[21] K A Breen P Seed K Parmar GWMoore S E Stuart-Smithand B J Hunt ldquoComplement activation in patients with isolatedantiphospholipid antibodies or primary antiphospholipid syn-dromerdquo Thrombosis and Haemostasis vol 107 no 3 pp 423ndash429 2012
Journal of Immunology Research 11
[22] F Fischetti P Durigutto V Pellis et al ldquoThrombus formationinduced by antibodies to 1205732-glycoprotein I is complementdependent and requires a priming factorrdquo Blood vol 106 no7 pp 2340ndash2346 2005
[23] J Guerin E Casey C Feighery and J Jackson ldquoAnti-Beta 2-glycoprotein I antibody isotype and IgG subclass in antiphos-pholipid syndrome patientsrdquo Autoimmunity vol 31 no 2 pp109ndash116 1999
[24] L Andreoli C Nalli M Motta et al ldquoAnti-1205732-glycoprotein I
IgG antibodies from 1-year-old healthy children born to moth-ers with systemic autoimmune diseases preferentially targetdomain 45 might it be the reason for their lsquoinnocentrsquo profilerdquoAnnals of the Rheumatic Diseases vol 70 no 2 pp 380ndash3832011
[25] B de Laat R H W M Derksen M van Lummel M T T Pen-nings and P G deGroot ldquoPathogenic anti-beta2-glycoprotein Iantibodies recognize domain I of beta2-glycoprotein I only aftera conformational changerdquo Blood vol 107 no 5 pp 1916ndash19242006
[26] M Samarkos K A Davies C Gordon M J Walport and SLoizou ldquoIgG subclass distribution of antibodies against 1205732-GP1and cardiolipin in patients with systemic lupus erythematosusand primary antiphospholipid syndrome and their clinicalassociationsrdquo Rheumatology vol 40 no 9 pp 1026ndash1032 2001
[27] J Arvieux B Roussel D Ponard and M G Colomb ldquoIgG2subclass restriction of anti-1205732 glycoprotein 1 antibodies inautoimmune patientsrdquo Clinical and Experimental Immunologyvol 95 no 2 pp 310ndash315 1994
[28] J N Arnold M R Wormald R B Sim P M Rudd and R ADwek ldquoThe impact of glycosylation on the biological functionand structure of human immunoglobulinsrdquo Annual Review ofImmunology vol 25 pp 21ndash50 2007
[29] Y Kaneko F Nimmerjahn and J V Ravetch ldquoAnti-inflamma-tory activity of immunoglobulin G resulting from Fc sialyla-tionrdquo Science vol 313 no 5787 pp 670ndash673 2006
[30] RM Anthony and F Nimmerjahn ldquoThe role of differential IgGglycosylation in the interaction of antibodies with FcgammaRsin vivordquo Current Opinion in Organ Transplantation vol 16 no1 pp 7ndash14 2011
[31] R M Anthony and J V Ravetch ldquoA novel role for the IgGFc glycan the anti-inflammatory activity of sialylated IgG FcsrdquoJournal of Clinical Immunology vol 30 no 1 pp S9ndashS14 2010
[32] K Oku T Atsumi M Bohgaki et al ldquoComplement activationin patients with primary antiphospholipid syndromerdquoAnnals ofthe Rheumatic Diseases vol 68 no 6 pp 1030ndash1035 2009
[33] H P McNeil R J Simpson C N Chesterman and S AKrilis ldquoAnti-phospholipid antibodies are directed against acomplex antigen that includes a lipid-binding inhibitor of coag-ulation 1205732-Glycoprotein I (apolipoprotein H)rdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 87 no 11 pp 4120ndash4124 1990
[34] R Jefferis and D S Kumararatne ldquoSelective IgG subclassdeficiency quantification and clinical relevancerdquo Clinical andExperimental Immunology vol 81 no 3 pp 357ndash367 1990
[35] P H Schur F Rosen and M E Norman ldquoImmunoglobulinsubclasses in normal childrenrdquo Pediatric Research vol 13 no3 pp 181ndash183 1979
[36] M A H French and G Harrison ldquoSerum IgG subclass concen-trations in healthy adults a study using monoclonal antiserardquoClinical and Experimental Immunology vol 56 no 2 pp 473ndash475 1984
[37] M Odorczuk L B Keil and V A DeBari ldquoBinding of com-plement component C1q to anti-beta2 glycoprotein I antibodiesfrom patients with antiphospholipid syndromerdquo InflammationResearch vol 48 no 10 pp 524ndash526 1999
[38] L Truedsson A A Bengtsson and G Sturfelt ldquoComplementdeficiencies and systemic lupus erythematosusrdquoAutoimmunityvol 40 no 8 pp 560ndash566 2007
[39] S Aschermann C H K Lehmann S Mihai G Schett DDudziak and F Nimmerjahn ldquoB cells are critical for autoim-mune pathology in Scurfy micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 110 no47 pp 19042ndash19047 2013
[40] Y Kaneko F Nimmerjahn M P Madaio and J V RavetchldquoPathology and protection in nephrotoxic nephritis is deter-mined by selective engagement of specific Fc receptorsrdquo TheJournal of Experimental Medicine vol 203 no 3 pp 789ndash7972006
[41] D Sylvestre R Clynes M Ma H Warren M C Carrolland J V Ravetch ldquoImmunoglobulin G-mediated inflammatoryresponses develop normally in complement-deficient micerdquoJournal of ExperimentalMedicine vol 184 no 6 pp 2385ndash23921996
[42] G E Ritchie B E Moffatt R B Sim B P Morgan R A Dwekand P M Rudd ldquoGlycosylation and the complement systemrdquoChemical Reviews vol 102 no 2 pp 305ndash320 2002
[43] Y Palarasah K Skjodt J Brandt et al ldquoGeneration of aC3c specific monoclonal antibody and assessment of C3c as aputative inflammatorymarker derived from complement factorC3rdquo Journal of ImmunologicalMethods vol 362 no 1-2 pp 142ndash150 2010
[44] D G Doherty J A Underhill P T Donaldson et al ldquoPoly-morphism in the human complement C4 genes and geneticsusceptibility to autoimmune hepatitisrdquo Autoimmunity vol 18no 4 pp 243ndash249 1994
[45] M-H Tao and S L Morrison ldquoStudies of aglycosylatedchimeric mouse-human IgG Role of carbohydrate in thestructure and effector functions mediated by the human IgGconstant regionrdquoThe Journal of Immunology vol 143 no 8 pp2595ndash2601 1989
[46] R Jefferis ldquoGlycosylation of recombinant antibody therapeu-ticsrdquo Biotechnology Progress vol 21 no 1 pp 11ndash16 2005
[47] Y Mimura S Church R Ghirlando et al ldquoThe influence ofglycosylation on the thermal stability and effector functionexpression of human IgG1-Fc properties of a series of truncatedglycoformsrdquoMolecular Immunology vol 37 no 12-13 pp 697ndash706 2001
[48] C Huhn M H J Selman L R Ruhaak A M Deelder and MWuhrer ldquoIgG glycosylation analysisrdquo Proteomics vol 9 no 4pp 882ndash913 2009
[49] J Lund N Takahashi J D Pound M Goodall and R JefferisldquoMultiple interactions of IgG with its core oligosaccharide canmodulate recognition by complement and human Fc gammareceptor I and influence the synthesis of its oligosaccharidechainsrdquo The Journal of Immunology vol 157 no 11 pp 4963ndash4969 1996
[50] R Forastiero M Martinuzzo L O Carreras and J MacloufldquoAnti-1205732 glycoprotein I antibodies and platelet activation inpatients with antiphospholipid antibodies association withincreased excretion of platelet-derived thromboxane urinarymetabolitesrdquoThrombosis and Haemostasis vol 79 no 1 pp 42ndash45 1998
12 Journal of Immunology Research
[51] J-C Reverter and D Tassies ldquoMechanism of thrombosis inantiphospholipid syndrome binding to plateletsrdquo in HughesSyndrome M A Khamashta Ed pp 403ndash414 Springer Lon-don UK 2006
[52] G Espinosa R Cervera J Font and Y Shoenfeld ldquoAntiphos-pholipid syndrome pathogenic mechanismsrdquo AutoimmunityReviews vol 2 no 2 pp 86ndash93 2003
[53] T Shi B Giannakopoulos X Yan et al ldquoAnti-1205732-glycoprotein
I antibodies in complex with 1205732-glycoprotein I can activate
platelets in a dysregulated manner via glycoprotein Ib-IX-VrdquoArthritis amp Rheumatism vol 54 no 8 pp 2558ndash2567 2006
[54] P LMeroni E RaschiMCamera et al ldquoEndothelial activationby aPL a potential pathogenetic mechanism for the clinicalmanifestations of the syndromerdquo Journal of Autoimmunity vol15 no 2 pp 237ndash240 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 Journal of Immunology Research
sialylation status of antiphospholipid antibodies seems toprevent the development of symptoms of APS also in theseindividuals
We conclude that a plethora of factors contributes tothe low pathogenicity of anti-1205732GP1-IgG in the sera ofhealthy children and asymptomatic adults We observed adistinct spectrum of IgG-subclasses compared to patientswith APS and a better binding of C3c indicating a moreefficient clearance of potentially harmful anti-1205732GP1 Thehypersialylation of the anti-1205732GP1-IgG is prone to amelioratetheir inflammatory and procoagulatory effects in healthycarriers whereas hyposialylation and inflammationmutuallyprecipitate the antiphospholipid syndrome
Disclosure
Thepresent workwas performed in fulfillment of the require-ments for obtaining the degree ldquoDr medrdquo for ChristophFickentscher
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Laura Andreoli andMartin Herrmann contributed equally tosenior authorship
Acknowledgments
This project was supported by the German Research Foun-dation (training research programme of the collaborativeresearch center 643) by the Emerging Fields Initiative (EFI)of the FAU Erlangen-Nuremberg and by the K und RWucherpfennig-Stiftung
References
[1] S Miyakis M D Lockshin T Atsumi et al ldquoInternationalconsensus statement on an update of the classification criteriafor definite antiphospholipid syndrome (APS)rdquo Journal ofThrombosis and Haemostasis vol 4 no 2 pp 295ndash306 2006
[2] W A Wilson A E Gharavi T Koike et al ldquoInternationalconsensus statement on preliminary classification criteria fordefinite antiphospholipid syndrome report of an internationalworkshoprdquo Arthritis and Rheumatism vol 42 no 7 pp 1309ndash1311 1999
[3] P G de Groot and R T Urbanus ldquoThe significance of autoan-tibodies against beta2-glycoprotein Irdquo Blood vol 120 no 2 pp266ndash274 2012
[4] C Nalli S Piantoni L Andreoli M Motta and A TincanildquoAntiphospholipid syndrome and antiphospholipid antibodiesin children the two sides of the coinrdquo Israel Medical AssociationJournal vol 14 no 5 pp 310ndash312 2012
[5] I Schousboe ldquo1205732-glycoprotein I a plasma inhibitor of thecontact activation of the intrinsic blood coagulation pathwayrdquoBlood vol 66 no 5 pp 1086ndash1091 1985
[6] J George M Blank Y Levy et al ldquoDifferential effects of anti-1205732-glycoprotein I antibodies on endothelial cells and on themanifestations of experimental antiphospholipid syndromerdquoCirculation vol 97 no 9 pp 900ndash906 1998
[7] R Willis and S S Pierangeli ldquoAnti-1205732-glycoprotein I antibod-iesrdquo Annals of the New York Academy of Sciences vol 1285 no1 pp 44ndash58 2013
[8] M J Fischer J Rauch and J S Levine ldquoThe antiphospholipidsyndromerdquo Seminars in Nephrology vol 27 no 1 pp 35ndash462007
[9] H Amital M Govoni R Maya et al ldquoRole of infectious agentsin systemic rheumatic diseasesrdquo Clinical and ExperimentalRheumatology vol 26 no 1 supplement 48 pp S27ndashS32 2008
[10] T Avcin ldquoAntiphospholipid syndrome in childrenrdquo CurrentOpinion in Rheumatology vol 20 no 5 pp 595ndash600 2008
[11] M Garcıa-Carrasco C Galarza-Maldonado C Mendoza-Pinto R O Escarcega and R Cervera ldquoInfections and theantiphospholipid syndromerdquo Clinical Reviews in Allergy ampImmunology vol 36 no 2-3 pp 104ndash108 2009
[12] P Cruz-Tapias M Blank J-M Anaya and Y ShoenfeldldquoInfections and vaccines in the etiology of antiphospholipidsyndromerdquo Current Opinion in Rheumatology vol 24 no 4 pp389ndash393 2012
[13] P von Landenberg Y Doring S Modrow and K J LacknerldquoAre antiphospholipid antibodies an essential requirement foran effective immune response to infectionsrdquoAnnals of the NewYork Academy of Sciences vol 1108 pp 578ndash583 2007
[14] M Blank I Krause M Fridkin et al ldquoBacterial induction ofautoantibodies to 1205732-glycoprotein-I accounts for the infectiousetiology of antiphospholipid syndromerdquo Journal of ClinicalInvestigation vol 109 no 6 pp 797ndash804 2002
[15] Y Shoenfeld M Blank R Cervera J Font E Raschi and P-LMeroni ldquoInfectious origin of the antiphospholipid syndromerdquoAnnals of the Rheumatic Diseases vol 65 no 1 pp 2ndash6 2006
[16] A E Gharavi S S Pierangeli R G Espinola X Liu MColden-Stanfield and E N Harris ldquoAntiphospholipid antibod-ies induced in mice by immunization with a cytomegalovirus-derived peptide cause thrombosis and activation of endothelialcells in vivordquo Arthritis and Rheumatism vol 46 no 2 pp 545ndash552 2002
[17] M Motta G Chirico C B Rebaioli et al ldquoAnticardiolipin andanti-beta2 glycoprotein I antibodies in infants born to motherswith antiphospholipid antibody-positive autoimmune diseasea follow-up studyrdquoAmerican Journal of Perinatology vol 23 no4 pp 247ndash251 2006
[18] T Avcin A Ambrozic M Kuhar T Kveder and B RozmanldquoAnticardiolipin and anti-1205732 glycoprotein I antibodies in seraof 61 apparently healthy children at regular preventive visitsrdquoRheumatology vol 40 no 5 pp 565ndash573 2001
[19] A Ambrozic ldquoAnti-beta2-glycoprotein I antibodies in children
with atopic dermatitisrdquo International Immunology vol 14 no 7pp 823ndash830
[20] J A Giron-Gonzalez E Garcıa Del Rıo C Rodrıguez JRodrıguez-Martorell and A Serrano ldquoAntiphospholipid syn-drome and asymptomatic carriers of antiphospholipid anti-body prospective analysis of 404 individualsrdquo Journal ofRheumatology vol 31 no 8 pp 1560ndash1567 2004
[21] K A Breen P Seed K Parmar GWMoore S E Stuart-Smithand B J Hunt ldquoComplement activation in patients with isolatedantiphospholipid antibodies or primary antiphospholipid syn-dromerdquo Thrombosis and Haemostasis vol 107 no 3 pp 423ndash429 2012
Journal of Immunology Research 11
[22] F Fischetti P Durigutto V Pellis et al ldquoThrombus formationinduced by antibodies to 1205732-glycoprotein I is complementdependent and requires a priming factorrdquo Blood vol 106 no7 pp 2340ndash2346 2005
[23] J Guerin E Casey C Feighery and J Jackson ldquoAnti-Beta 2-glycoprotein I antibody isotype and IgG subclass in antiphos-pholipid syndrome patientsrdquo Autoimmunity vol 31 no 2 pp109ndash116 1999
[24] L Andreoli C Nalli M Motta et al ldquoAnti-1205732-glycoprotein I
IgG antibodies from 1-year-old healthy children born to moth-ers with systemic autoimmune diseases preferentially targetdomain 45 might it be the reason for their lsquoinnocentrsquo profilerdquoAnnals of the Rheumatic Diseases vol 70 no 2 pp 380ndash3832011
[25] B de Laat R H W M Derksen M van Lummel M T T Pen-nings and P G deGroot ldquoPathogenic anti-beta2-glycoprotein Iantibodies recognize domain I of beta2-glycoprotein I only aftera conformational changerdquo Blood vol 107 no 5 pp 1916ndash19242006
[26] M Samarkos K A Davies C Gordon M J Walport and SLoizou ldquoIgG subclass distribution of antibodies against 1205732-GP1and cardiolipin in patients with systemic lupus erythematosusand primary antiphospholipid syndrome and their clinicalassociationsrdquo Rheumatology vol 40 no 9 pp 1026ndash1032 2001
[27] J Arvieux B Roussel D Ponard and M G Colomb ldquoIgG2subclass restriction of anti-1205732 glycoprotein 1 antibodies inautoimmune patientsrdquo Clinical and Experimental Immunologyvol 95 no 2 pp 310ndash315 1994
[28] J N Arnold M R Wormald R B Sim P M Rudd and R ADwek ldquoThe impact of glycosylation on the biological functionand structure of human immunoglobulinsrdquo Annual Review ofImmunology vol 25 pp 21ndash50 2007
[29] Y Kaneko F Nimmerjahn and J V Ravetch ldquoAnti-inflamma-tory activity of immunoglobulin G resulting from Fc sialyla-tionrdquo Science vol 313 no 5787 pp 670ndash673 2006
[30] RM Anthony and F Nimmerjahn ldquoThe role of differential IgGglycosylation in the interaction of antibodies with FcgammaRsin vivordquo Current Opinion in Organ Transplantation vol 16 no1 pp 7ndash14 2011
[31] R M Anthony and J V Ravetch ldquoA novel role for the IgGFc glycan the anti-inflammatory activity of sialylated IgG FcsrdquoJournal of Clinical Immunology vol 30 no 1 pp S9ndashS14 2010
[32] K Oku T Atsumi M Bohgaki et al ldquoComplement activationin patients with primary antiphospholipid syndromerdquoAnnals ofthe Rheumatic Diseases vol 68 no 6 pp 1030ndash1035 2009
[33] H P McNeil R J Simpson C N Chesterman and S AKrilis ldquoAnti-phospholipid antibodies are directed against acomplex antigen that includes a lipid-binding inhibitor of coag-ulation 1205732-Glycoprotein I (apolipoprotein H)rdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 87 no 11 pp 4120ndash4124 1990
[34] R Jefferis and D S Kumararatne ldquoSelective IgG subclassdeficiency quantification and clinical relevancerdquo Clinical andExperimental Immunology vol 81 no 3 pp 357ndash367 1990
[35] P H Schur F Rosen and M E Norman ldquoImmunoglobulinsubclasses in normal childrenrdquo Pediatric Research vol 13 no3 pp 181ndash183 1979
[36] M A H French and G Harrison ldquoSerum IgG subclass concen-trations in healthy adults a study using monoclonal antiserardquoClinical and Experimental Immunology vol 56 no 2 pp 473ndash475 1984
[37] M Odorczuk L B Keil and V A DeBari ldquoBinding of com-plement component C1q to anti-beta2 glycoprotein I antibodiesfrom patients with antiphospholipid syndromerdquo InflammationResearch vol 48 no 10 pp 524ndash526 1999
[38] L Truedsson A A Bengtsson and G Sturfelt ldquoComplementdeficiencies and systemic lupus erythematosusrdquoAutoimmunityvol 40 no 8 pp 560ndash566 2007
[39] S Aschermann C H K Lehmann S Mihai G Schett DDudziak and F Nimmerjahn ldquoB cells are critical for autoim-mune pathology in Scurfy micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 110 no47 pp 19042ndash19047 2013
[40] Y Kaneko F Nimmerjahn M P Madaio and J V RavetchldquoPathology and protection in nephrotoxic nephritis is deter-mined by selective engagement of specific Fc receptorsrdquo TheJournal of Experimental Medicine vol 203 no 3 pp 789ndash7972006
[41] D Sylvestre R Clynes M Ma H Warren M C Carrolland J V Ravetch ldquoImmunoglobulin G-mediated inflammatoryresponses develop normally in complement-deficient micerdquoJournal of ExperimentalMedicine vol 184 no 6 pp 2385ndash23921996
[42] G E Ritchie B E Moffatt R B Sim B P Morgan R A Dwekand P M Rudd ldquoGlycosylation and the complement systemrdquoChemical Reviews vol 102 no 2 pp 305ndash320 2002
[43] Y Palarasah K Skjodt J Brandt et al ldquoGeneration of aC3c specific monoclonal antibody and assessment of C3c as aputative inflammatorymarker derived from complement factorC3rdquo Journal of ImmunologicalMethods vol 362 no 1-2 pp 142ndash150 2010
[44] D G Doherty J A Underhill P T Donaldson et al ldquoPoly-morphism in the human complement C4 genes and geneticsusceptibility to autoimmune hepatitisrdquo Autoimmunity vol 18no 4 pp 243ndash249 1994
[45] M-H Tao and S L Morrison ldquoStudies of aglycosylatedchimeric mouse-human IgG Role of carbohydrate in thestructure and effector functions mediated by the human IgGconstant regionrdquoThe Journal of Immunology vol 143 no 8 pp2595ndash2601 1989
[46] R Jefferis ldquoGlycosylation of recombinant antibody therapeu-ticsrdquo Biotechnology Progress vol 21 no 1 pp 11ndash16 2005
[47] Y Mimura S Church R Ghirlando et al ldquoThe influence ofglycosylation on the thermal stability and effector functionexpression of human IgG1-Fc properties of a series of truncatedglycoformsrdquoMolecular Immunology vol 37 no 12-13 pp 697ndash706 2001
[48] C Huhn M H J Selman L R Ruhaak A M Deelder and MWuhrer ldquoIgG glycosylation analysisrdquo Proteomics vol 9 no 4pp 882ndash913 2009
[49] J Lund N Takahashi J D Pound M Goodall and R JefferisldquoMultiple interactions of IgG with its core oligosaccharide canmodulate recognition by complement and human Fc gammareceptor I and influence the synthesis of its oligosaccharidechainsrdquo The Journal of Immunology vol 157 no 11 pp 4963ndash4969 1996
[50] R Forastiero M Martinuzzo L O Carreras and J MacloufldquoAnti-1205732 glycoprotein I antibodies and platelet activation inpatients with antiphospholipid antibodies association withincreased excretion of platelet-derived thromboxane urinarymetabolitesrdquoThrombosis and Haemostasis vol 79 no 1 pp 42ndash45 1998
12 Journal of Immunology Research
[51] J-C Reverter and D Tassies ldquoMechanism of thrombosis inantiphospholipid syndrome binding to plateletsrdquo in HughesSyndrome M A Khamashta Ed pp 403ndash414 Springer Lon-don UK 2006
[52] G Espinosa R Cervera J Font and Y Shoenfeld ldquoAntiphos-pholipid syndrome pathogenic mechanismsrdquo AutoimmunityReviews vol 2 no 2 pp 86ndash93 2003
[53] T Shi B Giannakopoulos X Yan et al ldquoAnti-1205732-glycoprotein
I antibodies in complex with 1205732-glycoprotein I can activate
platelets in a dysregulated manner via glycoprotein Ib-IX-VrdquoArthritis amp Rheumatism vol 54 no 8 pp 2558ndash2567 2006
[54] P LMeroni E RaschiMCamera et al ldquoEndothelial activationby aPL a potential pathogenetic mechanism for the clinicalmanifestations of the syndromerdquo Journal of Autoimmunity vol15 no 2 pp 237ndash240 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Immunology Research 11
[22] F Fischetti P Durigutto V Pellis et al ldquoThrombus formationinduced by antibodies to 1205732-glycoprotein I is complementdependent and requires a priming factorrdquo Blood vol 106 no7 pp 2340ndash2346 2005
[23] J Guerin E Casey C Feighery and J Jackson ldquoAnti-Beta 2-glycoprotein I antibody isotype and IgG subclass in antiphos-pholipid syndrome patientsrdquo Autoimmunity vol 31 no 2 pp109ndash116 1999
[24] L Andreoli C Nalli M Motta et al ldquoAnti-1205732-glycoprotein I
IgG antibodies from 1-year-old healthy children born to moth-ers with systemic autoimmune diseases preferentially targetdomain 45 might it be the reason for their lsquoinnocentrsquo profilerdquoAnnals of the Rheumatic Diseases vol 70 no 2 pp 380ndash3832011
[25] B de Laat R H W M Derksen M van Lummel M T T Pen-nings and P G deGroot ldquoPathogenic anti-beta2-glycoprotein Iantibodies recognize domain I of beta2-glycoprotein I only aftera conformational changerdquo Blood vol 107 no 5 pp 1916ndash19242006
[26] M Samarkos K A Davies C Gordon M J Walport and SLoizou ldquoIgG subclass distribution of antibodies against 1205732-GP1and cardiolipin in patients with systemic lupus erythematosusand primary antiphospholipid syndrome and their clinicalassociationsrdquo Rheumatology vol 40 no 9 pp 1026ndash1032 2001
[27] J Arvieux B Roussel D Ponard and M G Colomb ldquoIgG2subclass restriction of anti-1205732 glycoprotein 1 antibodies inautoimmune patientsrdquo Clinical and Experimental Immunologyvol 95 no 2 pp 310ndash315 1994
[28] J N Arnold M R Wormald R B Sim P M Rudd and R ADwek ldquoThe impact of glycosylation on the biological functionand structure of human immunoglobulinsrdquo Annual Review ofImmunology vol 25 pp 21ndash50 2007
[29] Y Kaneko F Nimmerjahn and J V Ravetch ldquoAnti-inflamma-tory activity of immunoglobulin G resulting from Fc sialyla-tionrdquo Science vol 313 no 5787 pp 670ndash673 2006
[30] RM Anthony and F Nimmerjahn ldquoThe role of differential IgGglycosylation in the interaction of antibodies with FcgammaRsin vivordquo Current Opinion in Organ Transplantation vol 16 no1 pp 7ndash14 2011
[31] R M Anthony and J V Ravetch ldquoA novel role for the IgGFc glycan the anti-inflammatory activity of sialylated IgG FcsrdquoJournal of Clinical Immunology vol 30 no 1 pp S9ndashS14 2010
[32] K Oku T Atsumi M Bohgaki et al ldquoComplement activationin patients with primary antiphospholipid syndromerdquoAnnals ofthe Rheumatic Diseases vol 68 no 6 pp 1030ndash1035 2009
[33] H P McNeil R J Simpson C N Chesterman and S AKrilis ldquoAnti-phospholipid antibodies are directed against acomplex antigen that includes a lipid-binding inhibitor of coag-ulation 1205732-Glycoprotein I (apolipoprotein H)rdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 87 no 11 pp 4120ndash4124 1990
[34] R Jefferis and D S Kumararatne ldquoSelective IgG subclassdeficiency quantification and clinical relevancerdquo Clinical andExperimental Immunology vol 81 no 3 pp 357ndash367 1990
[35] P H Schur F Rosen and M E Norman ldquoImmunoglobulinsubclasses in normal childrenrdquo Pediatric Research vol 13 no3 pp 181ndash183 1979
[36] M A H French and G Harrison ldquoSerum IgG subclass concen-trations in healthy adults a study using monoclonal antiserardquoClinical and Experimental Immunology vol 56 no 2 pp 473ndash475 1984
[37] M Odorczuk L B Keil and V A DeBari ldquoBinding of com-plement component C1q to anti-beta2 glycoprotein I antibodiesfrom patients with antiphospholipid syndromerdquo InflammationResearch vol 48 no 10 pp 524ndash526 1999
[38] L Truedsson A A Bengtsson and G Sturfelt ldquoComplementdeficiencies and systemic lupus erythematosusrdquoAutoimmunityvol 40 no 8 pp 560ndash566 2007
[39] S Aschermann C H K Lehmann S Mihai G Schett DDudziak and F Nimmerjahn ldquoB cells are critical for autoim-mune pathology in Scurfy micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 110 no47 pp 19042ndash19047 2013
[40] Y Kaneko F Nimmerjahn M P Madaio and J V RavetchldquoPathology and protection in nephrotoxic nephritis is deter-mined by selective engagement of specific Fc receptorsrdquo TheJournal of Experimental Medicine vol 203 no 3 pp 789ndash7972006
[41] D Sylvestre R Clynes M Ma H Warren M C Carrolland J V Ravetch ldquoImmunoglobulin G-mediated inflammatoryresponses develop normally in complement-deficient micerdquoJournal of ExperimentalMedicine vol 184 no 6 pp 2385ndash23921996
[42] G E Ritchie B E Moffatt R B Sim B P Morgan R A Dwekand P M Rudd ldquoGlycosylation and the complement systemrdquoChemical Reviews vol 102 no 2 pp 305ndash320 2002
[43] Y Palarasah K Skjodt J Brandt et al ldquoGeneration of aC3c specific monoclonal antibody and assessment of C3c as aputative inflammatorymarker derived from complement factorC3rdquo Journal of ImmunologicalMethods vol 362 no 1-2 pp 142ndash150 2010
[44] D G Doherty J A Underhill P T Donaldson et al ldquoPoly-morphism in the human complement C4 genes and geneticsusceptibility to autoimmune hepatitisrdquo Autoimmunity vol 18no 4 pp 243ndash249 1994
[45] M-H Tao and S L Morrison ldquoStudies of aglycosylatedchimeric mouse-human IgG Role of carbohydrate in thestructure and effector functions mediated by the human IgGconstant regionrdquoThe Journal of Immunology vol 143 no 8 pp2595ndash2601 1989
[46] R Jefferis ldquoGlycosylation of recombinant antibody therapeu-ticsrdquo Biotechnology Progress vol 21 no 1 pp 11ndash16 2005
[47] Y Mimura S Church R Ghirlando et al ldquoThe influence ofglycosylation on the thermal stability and effector functionexpression of human IgG1-Fc properties of a series of truncatedglycoformsrdquoMolecular Immunology vol 37 no 12-13 pp 697ndash706 2001
[48] C Huhn M H J Selman L R Ruhaak A M Deelder and MWuhrer ldquoIgG glycosylation analysisrdquo Proteomics vol 9 no 4pp 882ndash913 2009
[49] J Lund N Takahashi J D Pound M Goodall and R JefferisldquoMultiple interactions of IgG with its core oligosaccharide canmodulate recognition by complement and human Fc gammareceptor I and influence the synthesis of its oligosaccharidechainsrdquo The Journal of Immunology vol 157 no 11 pp 4963ndash4969 1996
[50] R Forastiero M Martinuzzo L O Carreras and J MacloufldquoAnti-1205732 glycoprotein I antibodies and platelet activation inpatients with antiphospholipid antibodies association withincreased excretion of platelet-derived thromboxane urinarymetabolitesrdquoThrombosis and Haemostasis vol 79 no 1 pp 42ndash45 1998
12 Journal of Immunology Research
[51] J-C Reverter and D Tassies ldquoMechanism of thrombosis inantiphospholipid syndrome binding to plateletsrdquo in HughesSyndrome M A Khamashta Ed pp 403ndash414 Springer Lon-don UK 2006
[52] G Espinosa R Cervera J Font and Y Shoenfeld ldquoAntiphos-pholipid syndrome pathogenic mechanismsrdquo AutoimmunityReviews vol 2 no 2 pp 86ndash93 2003
[53] T Shi B Giannakopoulos X Yan et al ldquoAnti-1205732-glycoprotein
I antibodies in complex with 1205732-glycoprotein I can activate
platelets in a dysregulated manner via glycoprotein Ib-IX-VrdquoArthritis amp Rheumatism vol 54 no 8 pp 2558ndash2567 2006
[54] P LMeroni E RaschiMCamera et al ldquoEndothelial activationby aPL a potential pathogenetic mechanism for the clinicalmanifestations of the syndromerdquo Journal of Autoimmunity vol15 no 2 pp 237ndash240 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

12 Journal of Immunology Research
[51] J-C Reverter and D Tassies ldquoMechanism of thrombosis inantiphospholipid syndrome binding to plateletsrdquo in HughesSyndrome M A Khamashta Ed pp 403ndash414 Springer Lon-don UK 2006
[52] G Espinosa R Cervera J Font and Y Shoenfeld ldquoAntiphos-pholipid syndrome pathogenic mechanismsrdquo AutoimmunityReviews vol 2 no 2 pp 86ndash93 2003
[53] T Shi B Giannakopoulos X Yan et al ldquoAnti-1205732-glycoprotein
I antibodies in complex with 1205732-glycoprotein I can activate
platelets in a dysregulated manner via glycoprotein Ib-IX-VrdquoArthritis amp Rheumatism vol 54 no 8 pp 2558ndash2567 2006
[54] P LMeroni E RaschiMCamera et al ldquoEndothelial activationby aPL a potential pathogenetic mechanism for the clinicalmanifestations of the syndromerdquo Journal of Autoimmunity vol15 no 2 pp 237ndash240 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom























![[The incidence of the pituitary autoantibodies in Graves' disease]](https://img.dokumen.tips/doc/110x75/635915fc68c5ffb42104606f/the-incidence-of-the-pituitary-autoantibodies-in-graves-disease.jpg)





