Embed Size (px)
Citation preview

Autoantibodies to alpha-synuclein in inherited Parkinson’sdisease
Katerina K. Papachroni*, Natalia Ninkina†, Angeliki Papapanagiotou*, Georgios M.Hadjigeorgiou‡, Georgia Xiromerisiou‡, Alexandros Papadimitriou‡, AnastasiosKalofoutis*, and Vladimir L. Buchman†
*Laboratory of Biological Chemistry, University of Athens Medical School, Athens, Greece†School of Biosciences, Cardiff University, Cardiff, UK‡Department of Neurology, Neurogenetics Unit, Medical School, University of Thessaly, Larissa,Greece
AbstractNeurodegeneration in Parkinson’s disease (PD) is accompanied by a local immune reaction in theaffected brain regions. It is well established that α-synuclein is directly implicated in thepathogenesis of PD. Development of the disease is often associated with changes of expressionand cellular compartmentalisation of this protein; moreover, its oligomers or protofibrils are oftenreleased to the CSF and plasma of patients. Aggregated α-synuclein can trigger the activation ofmicroglia; however, its capacity to induce production of specific autoantibodies (AAb) has notbeen assessed. In this study, we examined the presence of AAb against synuclein family membersin the peripheral blood serum of PD patients and control individuals. Presence of AAb against β-synuclein or γ-synuclein showed no association with PD. Multi-epitopic AAb against α-synucleinwere detected in 65% of all patients tested and their presence strongly correlated with an inheritedmode of the disease but not with other disease-related factors. The frequency of the presence ofAAb in the studied group of patients with sporadic form of PD was not significantly different fromthe frequency in the control group but very high proportion (90%) of patients with familial form ofthe disease were positive for AAb against α-synuclein. We hypothesise that these AAb could beinvolved in pathogenesis of the inherited form of PD.
Keywordsalpha-synuclein; autoantibody; Parkinson’s disease
The principal pathological feature of Parkinson’s disease (PD) is a massive death ofdopaminergic neurons. With the exception of a few hereditary and toxin-induced forms, theaetiology and mechanism of the development of this pathology are not known. Recently,immunological components of the disease came to the limelight, because local inflammatoryand immune reactions in PD patients were identified as factors that exacerbate theneurodegenerative processes in the affected regions of the brain. Pro-inflammatory eventsthat might trigger these reactions, like head trauma and intrauterine foetal exposure tocertain viruses, are considered to be pre-disposing factors to the disease (Ringheim and
© 2007 International Society for Neurochemistry
Address correspondence and reprint requests to V. L. Buchman, School of Biosciences, Cardiff University, Museum Avenue, CardiffCF10 3US, UK. [email protected].
Europe PMC Funders GroupAuthor ManuscriptJ Neurochem. Author manuscript; available in PMC 2011 August 11.
Published in final edited form as:J Neurochem. 2007 May ; 101(3): 749–756. doi:10.1111/j.1471-4159.2006.04365.x.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Conant 2004), and epidemiological studies revealed that chronic use of non-steroidal anti-inflammatory drugs may reduce the risk of PD by about 45% (Chen et al. 2003).
Local inflammation and immune response in PD involve both CNS resident cells (mainlyactivated microglia) and infiltrating peripheral monocytes and leukocytes (T-cells and B-cells). A profound microglia proliferation is found in the substantia nigra (SN) and striatumof PD patients as well as in the basal ganglia of the experimental animal models of PD(Hunot and Hirsch 2003). Activated microglia could develop into functional antigenpresenting cells, activate the classical complement pathway, induce opsonisation ofdamaged neurons and promote chemotaxis (Wucherpfennig 1994). Possible consequences ofthis initial immune activation in the affected region of PD brains are the localpermeabilisation of blood–brain barrier. These could lead to infiltration of the affectedregions by B- and/or T-lymphocytes, which is believed to be a critical step for thedevelopment of autoimmune reactions (Racke et al. 2000).
Recent studies linked the PD-associated immune reaction with the dysfunction of α-synuclein, an abundant neuronal protein with developmentally regulated pattern ofexpression in many neuronal populations, including the dopaminergic neurons of the SN.The normal function(s) of α-synuclein is not known but the ability of over-expressed α-synuclein to functionally compensate for the loss of certain chaperone complexes involvedin the regulation of neurotransmitter release in neuronal synapses (Chandra et al. 2005) isconsistent with the hypothesis that synucleins are scaffold/adaptor proteins that enhanceefficiency of various presynaptic processes (Papachroni et al. 2005). The implication of α-synuclein in PD is well documented and supported by observations that (i) aggregates offibrillated α-synuclein are the main constituents of Lewy bodies, (ii) certain mis-sensemutations, as well as duplication or triplication of the α-synuclein gene, cause autosomaldominant PD and (iii) the principal molecular, cellular and pathophysiological aspects of PDcan be modelled by expression of α-synuclein in neuronal cell lines or transgenic organisms(reviewed in Maries et al. 2003; Forman et al. 2005). Certain intermediates of α-synucleinaggregation are believed to be toxic to neurons, particularly to dopaminergic neurons(Lundvig et al. 2005). Death of neurons or damage of axons and synaptic terminals couldresult in the release of various soluble or aggregated forms of α-synuclein into theextracellular space and, in accord with this scenario, monomeric and oligomeric forms of α-synuclein have been found in the CSF and plasma of PD patients (El-Agnaf et al. 2006).Experiments in primary mesencephalic neuron–glia cultures have demonstrated that theextracellular aggregated form of human α-synuclein induces microglia activation, whichaggravates the degeneration of neurons (Zhang et al. 2005). One of the effects of activatedmicroglia on co-cultured dopaminergic neurons is nitration of α-synuclein, which isbelieved to be a crucial step that triggers death of these neurons (Shavali et al. 2006). Thesedata suggest that α-synuclein is involved in the initial steps of the local immune responseassociated with PD. However, its involvement in further steps, particularly in the inductionof autoimmune reactions, has never been studied. In this study, we assessed the presence ofautoantibodies (AAb) against all three synucleins in the peripheral blood serum of PDpatients and control healthy individuals.
Materials and methodsPatients and healthy controls
The study sample consisted of three groups of subjects – patients with sporadic (n = 31) orfamilial (n = 20) PD (fPD) (total 32 men, 19 women; mean age 65.22 ± 12.08 years) and acontrol group included 26 healthy individuals (16 men, 10 women; mean age 64.9 ± 10.9years) with matching gender, age and ethnic characteristics and with no history ofneurological illness. Demographics data of patients and control individuals are shown in
Papachroni et al. Page 2
J Neurochem. Author manuscript; available in PMC 2011 August 11.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Table 1. All patients were residents of Thessaly (Central Greece) and were recruited fromthe outpatient clinic for movement disorders in the University Hospital of Larissa andfollowed up for at least 1 year. Mutations in autosomal dominant PD genes (α-synuclein,ubiquitin carboxylterminal hydrolase L1 (UCHL1) and leucine rich repeat kinase 2(LRRK2)) were excluded by direct sequencing in all fPD patients. Dosage alterations in α-synuclein gene were excluded by quantitative duplex PCR. The LRRK2 G2019S mutationthat may cause sporadic PD was excluded in all sporadic PD patients. Skilled neurologists(G.M.H and A.P.) performed all clinical assessments including PD diagnosis, stagingaccording to Hoehn and Yahr scale, age-at-onset, etc. Fifteen probands with fPD had fiveaffected family members and five probands with fPD had six affected family members,apparently on autosomal dominant mode of inheritance based on family history and pedigreeanalysis (see Fig. 1 and Xiromerisiou et al. 2006).
In the absence of knowing the causative gene in these families, or haplotype data around aknown locus, it is premature to conclude whether or not these families are unrelated. In twofamilies, two affected members were examined in each family, while diagnosis of othermembers was based on medical records or self-report of the proband. In the remainingfamilies (n = 18), at least three affected members were examined by movement disorders’specialists (G.M.H or A.P). Moreover, PD was excluded in most unaffected family membersafter clinical examination.
All PD patients were under pharmaceutical treatment. Controls were subjects living in thesame geographical area, who visited other, non-neurological outpatient clinics and were freeof disease (PD included). The samples of peripheral blood serum of all subjects werealiquoted and stored at −80°C. This study was approved by the institutional ethical reviewcommittees. All subjects or their families were informed of the investigational nature of thestudy and informed consent was obtained for their participation.
Preparation of bacterially expressed recombinant proteinsThe cDNA fragments encoding full-length α-synuclein, β-synuclein and γ-synuclein, andoverlapping peptides of α-synuclein were PCR amplified from corresponding plasmidtemplates (α-syn/pRK172 and β-syn/pRK172 and γ-syn/H1) using Pfu polymerase(Stratagene, La Jolla, CA, USA) and cloned into pGEX-4T-1 (Amersham PharmaciaBiotech, St. Albans, UK) vector in frame with glutathione-S-transferase (GST). Nucleotidesequences of all plasmid constructs were verified by DNA sequencing. Expression ofrecombinant proteins was induced with 1 mmol/L isopropyl-b-D-thiogalactosidase inexponentially growing Escherichia coli BL21 cells transformed with correspondingrecombinant plasmids. GST-fusion proteins were purified using standard affinitypurification protocol described by the manufacturer of Glutathione Sepharose 4B(Amersham Pharmacia). When required, eluted GST fusion proteins were treated with 10units of human thrombin (Sigma, St. Louis, MO, USA) per 0.5 mg of protein at 18–21°C for1.5 h and the GST fragment was removed by re-absorption on the fresh GlutathioneSepharose 4B beads.
Immunoblot analysisSodium dodecyl sulfate – polyacrylamide gel electrophoresis (SDS–PAGE) of recombinantsynucleins was performed as described (Buchman et al. 1998), using 4–12% polyacrylamidegels. Separated recombinant proteins (1–2 μg per lane) were transferred to nitrocellulosemembrane (ECL Hybond, Amersham Pharmacia). Following blocking with 5% non-fat drymilk in Tris-buffered saline–0.1% Tween 20 (TBST), the membranes were incubated withhuman serum (1 : 100 dilution in 5% milk/TBST) for 14–16 h at 4°C. The presence of AAbwas revealed using secondary horseradish peroxidase-conjugated anti-human antibody
Papachroni et al. Page 3
J Neurochem. Author manuscript; available in PMC 2011 August 11.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

(Sigma) and ECL detection system (Amersham Pharmacia). In some experiments, filterswere re-probed with specific anti-synuclein antibodies. Monoclonal anti-α-synuclein(sc-32280, Santa Cruz Biotechnology), polyclonal anti-β-synuclein (sc-9565, Santa CruzBiotechnology) and anti-γ-synuclein SK109 (Ninkina et al. 1998) antibodies were used in1 : 1000 dilution.
Statistical analysisData were analysed and power analysis was performed with the statistical software SPSSv12 (SPSS Inc., Chicago, IL, USA). Two by two associations were tested using the Fisher’sexact test and the odds ratio with the corresponding 95% confidence interval. The differencein continuous variables was tested using the Mann–Whitney U-test. The concordance inpositive immunosignal between any two constructs was tested using the McNemar’s test andthe concordance for positive immunosignal for more than two constructs was tested usingthe Cochran’s Q-test. p-values >0.05 for Conhran’s Q-test and McNemar’s test areindicative of concordance. In all cases, p < 0.05 was considered statistically significant.
ResultsTo assess the presence of AAb against synucleins in the peripheral blood serum of PDpatients and healthy control individuals, we used each diluted sera as a source of primaryantibody for immunoblot detection of denatured recombinant human α-, β- and γ-synucleins. A positive signal for at least one synuclein was detected with 69% of all serumsamples when they were used in a 1 : 100 dilution. The use of less diluted sera produced thesame results but on significantly higher background over the whole surface of the filter.Typical examples of immunoblots probed with human peripheral blood sera are shown inFigs 2 and 3. The AAb against α-synuclein was present with significantly (p = 0.007) higherfrequency in the tested PD patient samples (65%) than in the control group samples (31%).This association was due to high frequency of AAb in fPD patients (90%; p < 0.001), whilethere was no statistically different distribution of AAb between sporadic PD patients (48%)and controls (p = 0.17) (Table 2). The observed frequency of α-synuclein AAb presence inthe control population is in accordance with other reports referring to the frequency ofnatural antibodies against various autoantigens in healthy subjects (Terryberry et al. 1998).Control population showed higher frequency of a positive immune reaction for β-synuclein;however, the difference from the PD group did not bear statistical significance (Table 2).Only six serum samples (two from the PD and four from the control group) reacted with γ-synuclein. The analysis of correlations between the presence of AAb against either α- or β-synuclein in the PD subjects and the age or sex of the patients, the stage of the disease, theinheritance mode and other pre-disposing factors (pesticide exposure, traumatic brain injury(TBI), smoking) demonstrated that only inheritance mode of the disease (which isconsidered the primary investigated parameter) significantly correlates with the presence ofthe autoantibody – the unadjusted odds ratio was 9.60 with 95% confidence interval (1.90–48.60) and the power to detect the observed association at 5% significance level was 91%.No association was established with the other factors (adjusted ORs are shown in Table 3).The age of onset or the disease duration also did not show correlation (two-tailed Mann–Whitney U-test) with the presence of α-synuclein AAb (p = 0.89 for the age of onset, p =0.98 for the duration) or β-synuclein AAb (p = 0.33 for age-at-onset, p = 0.50 for duration).
To reveal whether α-synuclein AAb in PD patients display increased specificity to particularepitopes of the protein, overlapping α-synuclein fragments fused to a GST-tag, weredesigned (Fig. 4a), expressed in E. coli and purified as described in Materials and methodssection. For larger fragments, fusion proteins were digested with thrombin to remove theGST-tag, while fragments 3, 317 and 99 were used in GST-fusion form, because their sizes(<5 kDa) were too small for efficient SDS–PAGE analysis. The fragments were separated
Papachroni et al. Page 4
J Neurochem. Author manuscript; available in PMC 2011 August 11.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

by SDS–PAGE, transferred to nitrocellulose membrane and incubated with diluted serum ofcontrol or PD subjects. Serum samples that were previously found positive for the presenceof AAb against the full-length α-synuclein reacted with one or several α-synuclein peptides.A sample immunoblot is shown in Fig. 4b.
The association between antigenicity of each of the fragments and disease-related factors(sex, mode of inheritance, age, disease stage, smoking, TBI, pesticide exposure) wasassessed and no statistically significant correlations were found (Table 4). However,combined immunoreactivity to all three peptides (N-terminal peptide 3, C-terminal peptide99 and middle protein region peptide 317) or combined immunoreactivity for any twopeptides of those three is concordant, as was shown by statistical analysis (Cochran’s Q-testanalysis of antigenicity of the three peptides, p = 0.78; McNemar’s two-sided test forimmunopositivity between any two peptides of the three: 3 versus 317, p = 0.75; 99 versus3, p = 1.0; 317 versus 99, p = 0.73).
DiscussionA growing body of evidence from clinical studies and experiments with animal modelsemphasise a role for an immune component in the development of PD. Neurodegenerationin the brain of PD patients is accompanied by inflammatory-like response, primarilycharacterised by activation of microglia. Direct inhibition of microglial activation (Delgadoand Ganea 2003) or therapeutic immunisation using adoptive transfer of immune cells(Benner et al. 2004) is neuroprotective in model systems. Further development of the localimmune response is characterised by infiltration of the damaged regions with various typesof immune cells, including B- and T-lymphocytes. Particularly interesting is the highrepresentation in this lymphocyte population of activated (CD25+) γδ-T cells that play akey role in autoimmunity. The increased number of γδ-T cells is often seen in peripheralblood of PD patients but the elevated levels of activated (CD25+) γδ-T cells are foundsolely in the CSF and not peripherally, suggesting that activation of these cells is associatedwith degenerative processes in the CNS (Fiszer et al. 1994). Therefore, degeneration ofdopaminergic neurons and associated local inflammation in the SN of PD patients sets upconditions for the development of an autoimmune reaction. Indeed, AAb against variousantigens that recognise dopaminergic neurons have been found in the CSF and peripheralblood serum of patients with PD (Defazio et al. 1994; Chen et al. 1998; Rowe et al. 1998;Bas et al. 2001; Orr et al. 2005).
Here, we have demonstrated for the first time the presence of AAb against synucleins in thehuman peripheral blood serum. A positive correlation with PD has been found only for AAbagainst α-synuclein, which is consistent with the role of this protein but not two othersynuclein family members in the aetiology and pathogenesis of PD. This correlation wasevident only for PD patients with familial but not sporadic form of the disease, suggestingthat production of AAb against α-synuclein is associated with this commonly moreaggressive form of PD. We have shown that the majority of studied antisera were multi-epitopic and the epitopes were spread across the length of α-synuclein molecule. Thisobservation confirms that not a single peptide but the whole α-synuclein molecule isautoimmunogenic and dismisses a possibility that the observed immune reaction was theresult of an accidental cross-reactivity with a α-synuclein epitope of AAb produced againstan antigenically similar epitope of another protein. Although the protocol used in this study(immunoblot analysis of denatured recombinant synucleins) was designed to detect onlyAAb against linear antigens, it is feasible that using different detection methods AAbspecific to covalently modified α-synuclein species or aggregation-dependent epitopes couldalso be found in PD patients.
Papachroni et al. Page 5
J Neurochem. Author manuscript; available in PMC 2011 August 11.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

The question about functional importance of antibodies against disease-associated neuronalproteins remains wide open. It has been demonstrated that an immunoglobulin G fractionpurified from serum of PD patients causes death of dopaminergic neurons in vivo followingstereotaxic injection in the SN of experimental animals (Chen et al. 1998), and the presenceof immunoglobulins in PD brain tissue could lead to the targeting of dopaminergic nigralneurons for destruction (Orr et al. 2005). Thus, it is feasible to suggest that AAb against α-synuclein could also be neurotoxic. However, results of other experiments suggest thatantibodies against certain proteins implicated in neurodegenerative disorders could bebeneficial for slowing down diseases progression. Immunisation-mediated neuroprotectionfollowing central or peripheral nervous trauma has been demonstrated (Moalem et al. 1999)and vaccination approaches have been effective in the removal of huntingtin (Luthi-Carter2003) and β-amyloid1–42 peptide (Janus et al. 2000) deposits in animal models. Recently, aneuroprotective role of anti-α-synuclein antibodies has been shown in a human α-synucleintransgenic mouse model of PD. Mice that produced high affinity antibodies followingvaccination with human α-synuclein displayed reduced accumulation of the aggregatedhuman α-synuclein in the neuronal cell bodies and the synapses, and decreased degenerationof neurons, probably because the antibodies recognised the aggregated human α-synucleinand promoted its degradation via lysosomal pathways (Masliah et al. 2005). It is possiblethat α-synuclein AAb in the serum of fPD patients are also part of a protective reactionaimed at elimination of extracellular toxic α-synuclein species or even cells bearing thesespecies. Such cleansing could not eradicate the cause of the disease, which in these cases isobviously genetic, but could at least break the vicious cycle of events and slow the diseaseprogression. To clarify a role for α-synuclein AAb, further longitudinal studies of largercohorts of PD patients and healthy individuals, including healthy members of the affectedfamilies, as well as animal models of the disease are necessary. These studies should revealwhether the monitoring of the AAb titres could have a diagnostic value and whetherimmunisation with α-synuclein could be used as a novel approach in PD therapy.
AcknowledgmentsWe are grateful to Michel Goedert for α- and β-synuclein plasmids, Julia Wanless for help with protein expression,the DNA Sequencing Unit, University of Dundee for excellent sequencing service, Elias Zintzaras for his usefulcomments on statistical analysis and Anne Rosser for critical reading of the article. This study was partiallysupported by grants from the Universities of Athens and Thessaly (Greece) and by grants from The Wellcome Trustto VLB. KP visit to the UK laboratory was sponsored by Eurogendis.
Abbreviations used
AAb autoantibodies
fPD familial PD
GST glutathione-S-transferase
PD Parkinson’s disease
SDS–PAGE sodium dodecyl sulfate – polyacrylamide gel electrophoresis
SN substantia nigra
TBI traumatic brain injury
TBST Tris-buffered saline with 0.1% Tween 20
Papachroni et al. Page 6
J Neurochem. Author manuscript; available in PMC 2011 August 11.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

ReferencesBas J, Calopa M, Mestre M, Mollevi DG, Cutillas B, Ambrosio S, Buendia E. Lymphocyte population
in Parkinson’s disease and rat models of parkinsonism. J. Neuroimmunol. 2001; 113:146–152.[PubMed: 11137586]
Benner EJ, Mosley RL, Destache CJ, Lewis TB, Jackson-Lewis V, Gorantla S, Nemachek C, GreenSR, Przedborski S, Gendelman HE. Therapeutic immunization protects dopaminergic neurons in amouse model of Parkinson’s disease. Proc. Natl Acad. Sci. USA. 2004; 101:9435–9440. [PubMed:15197276]
Buchman VL, Hunter HJA, Pinon LGP, Thompson J, Privalova EM, Ninkina NN, Davies AM. Persyn,a member of the synuclein family, has a distinct pattern of expression in the developing nervoussystem. J. Neurosci. 1998; 18:9335–9341. [PubMed: 9801372]
Chandra S, Gallardo G, Fernández-Chacón R, Schlüter O, Südhof T. Alpha-synuclein cooperates withCSPalpha in preventing neurodegeneration. Cell. 2005; 123:383–396. [PubMed: 16269331]
Chen S, Weidong DL, Wenjie JX, Alexianu ME, Engelhardt JI, Siklos L, Appel SH. Experimentaldestruction of subtantia nigra initiated by Parkinson’s disease immunoglobulins. Arch. Neurol.1998; 55:1075–1080. [PubMed: 9708957]
Chen A, Zhang B, Hernan MA, Schwarzschild MA, Willett WC, Colditz GA, Speizer FE, Ascherio A.Nonsteroidal anti-inflammatory drugs and the risk of Parkinson’s disease. Arch. Neurol. 2003;60:1059–1064. [PubMed: 12925360]
Defazio G, Dal Toso R, Benvegnu D, Minozzi MC, Cananzi AR, Leon A. Parkinsonian serum carriescomplement-dependent toxicity for rat mesencephalic dopaminergic neurons in culture. Brain Res.1994; 633:206–212. [PubMed: 7907931]
Delgado M, Ganea D. Neuroprotective effect of vasoactive intestinal peptide (VIP) in a mouse modelof Parkinson’s disease by blocking microglial activation. FASEB J. 2003; 17:944–946. [PubMed:12626429]
El-Agnaf OMA, Salem SA, Paleologou KE, et al. Detection of oligomeric forms of alpha-synucleinprotein in human plasma as a potential biomarker for Parkinson’s disease. FASEB J. 2006; 20:419–425. [PubMed: 16507759]
Fiszer U, Mix E, Fredrikson S, Kostulas V, Olsson T, Link H. Gamma delta+ T cells are increased inpatients with Parkinson’s disease. J. Neurol. Sci. 1994; 121:39–45. [PubMed: 8133310]
Forman MS, Lee VM, Trojanowski JQ. Nosology of Parkinson’s disease, looking for the way out of aquackmire. Neuron. 2005; 47:479–482. [PubMed: 16102530]
Hunot S, Hirsch EC. Neuroinflammatory processes in Parkinson’s disease. Ann. Neurol. 2003;53:S49–S60. [PubMed: 12666098]
Janus C, Pearson J, McLaurin J, et al. A beta peptide immunization reduces behavioural impairmentand plaques in a model of Alzheimer’s disease. Nature. 2000; 408:979–982. [PubMed: 11140685]
Lundvig D, Lindersson E, Jensen PH. Pathogenic effects of alpha-synuclein aggregation. Brain Res.Mol. Brain Res. 2005; 134:3–17. [PubMed: 15790525]
Luthi-Carter R. Progress towards a vaccine for Huntington’s disease. Mol. Ther. 2003; 7:569–570.[PubMed: 12718897]
Maries E, Dass B, Collier TJ, Kordower JH, Steece-Collier K. The role of alpha-synuclein inParkinson’s disease, insights from animal models. Nat. Rev. Neurosci. 2003; 4:727–738.[PubMed: 12951565]
Masliah E, Rockenstein E, Adame A, et al. Effects of alpha-synuclein immunisation in a mouse modelof Parkinson’s disease. Neuron. 2005; 46:857–868. [PubMed: 15953415]
Moalem G, Leibowitz-Amit R, Yoles E, Mor F, Cohen IR, Schwartz M. Autoimmune T cells protectneurons from secondary degeneration after central nervous system axotomy. Nat. Med. 1999;5:49–55. [PubMed: 9883839]
Ninkina NN, Alimova-Kost MV, Paterson JW, Delaney L, Cohen BB, Imreh S, Gnuchev NV, DaviesAM, Buchman VL. Organization, expression and polymorphism of the human persyn gene. Hum.Mol. Genet. 1998; 7:1417–1424. [PubMed: 9700196]
Orr CF, Rowe DB, Mizuno Y, Mori H, Halliday GM. A possible role for humoral immunity in thepathogenesis of Parkinson’s disease. Brain. 2005; 128:2665–2674. [PubMed: 16219675]
Papachroni et al. Page 7
J Neurochem. Author manuscript; available in PMC 2011 August 11.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Papachroni K, Ninkina N, Wanless J, Kalofoutis AT, Gnuchev NV, Buchman VL. Peripheral sensoryneurons survive in the absence of alpha and gamma synucleins. J. Mol. Neurosci. 2005; 25:157–164. [PubMed: 15784963]
Racke MK, Ratts RB, Arredondo L, Perrin PJ, Lovett-Racke A. The role of costimulation inautoimmune demyelination. J. Neuroimmunol. 2000; 107:205–215. [PubMed: 10854658]
Ringheim GE, Conant K. Neurodegenerative disease and the neuroimmune axis (Alzheimer’s andParkinson’s disease and viral infections). J. Neuroimmunol. 2004; 147:43–49. [PubMed:14741426]
Rowe DB, Le W, Smith RG, Appel SH. Antibodies from patients with Parkinson’s disease react withprotein modified by dopamine oxidation. J. Neurosci. Res. 1998; 53:551–558. [PubMed: 9726426]
Shavali S, Combs CK, Ebadi M. Reactive macrophages increase oxidative stress and alpha-synucleinnitration during death of dopaminergic neuronal cells in co-culture: relevance to Parkinson’sdisease. Neurochem. Res. 2006; 31:85–94. [PubMed: 16475001]
Terryberry JW, Thor G, Peter JB. Autoantibodies in neurodegenerative diseases: antigen-specificfrequencies and intrathecal analysis. Neurobiol. Aging. 1998; 19:205–216. [PubMed: 9661995]
Wucherpfennig KW. Autoimmunity in the central nervous system: mechanisms of antigen presentationand recognition. Clin. Immunol. Immunopathol. 1994; 72:293–306. [PubMed: 8062444]
Xiromerisiou G, Hadjigeorgiou GM, Gourbali V, Johnson J, Papakonstantinou I, Papadimitriou A,Singleton AB. Screening for SNCA and LRRK2 mutations in Greek sporadic and autosomaldominant Parkinson’s disease: identification of two novel LRRK2 variants. Eur. J. Neurol. 2006published online September 21 2006, doi: 10.1111/j.1468-1331.2006.01551.x.
Zhang W, Wang T, Pei Z, et al. Aggregated alpha-synuclein activates microglia: a process leading todisease progression in Parkinson’s disease. FASEB J. 2005; 19:533–542. [PubMed: 15791003]
Papachroni et al. Page 8
J Neurochem. Author manuscript; available in PMC 2011 August 11.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Fig. 1.Pedigrees for the patients with familiar form of PD participated in the study. Affectedmembers of the family are shown as filled circles or squares. Arrows show studied patients.Of 20 patients with familiar form of the disease, only patients GRFPD14 and GRFPD17 didnot have detectable levels of AAb against α-synuclein.
Papachroni et al. Page 9
J Neurochem. Author manuscript; available in PMC 2011 August 11.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Fig. 2.Examples of immunoreactivity of sera obtained from PD patients with recombinant full-length synucleins. To confirm that protein bands detected by the PD serum GRFPD16 (a,top panel) represent α-synuclein, the membrane was re-probed with an anti-α-synucleinantibody (a, bottom panel, see also Materials and methods section). The right panels showthe same membrane stained with Ponceau-S to visualize full-length recombinant synucleins(b, top panel) and probed with another PD serum GRFPD6, which has AAb against α- andβ- but not γ-synuclein (b, bottom panel). Panel c shows examples of immunoblots probedwith a control serum, sera from patients with sporadic form of PD (GRSPD) or sera frompatients from PD families (GRFDP, designated by arrows in Fig. 1).
Papachroni et al. Page 10
J Neurochem. Author manuscript; available in PMC 2011 August 11.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
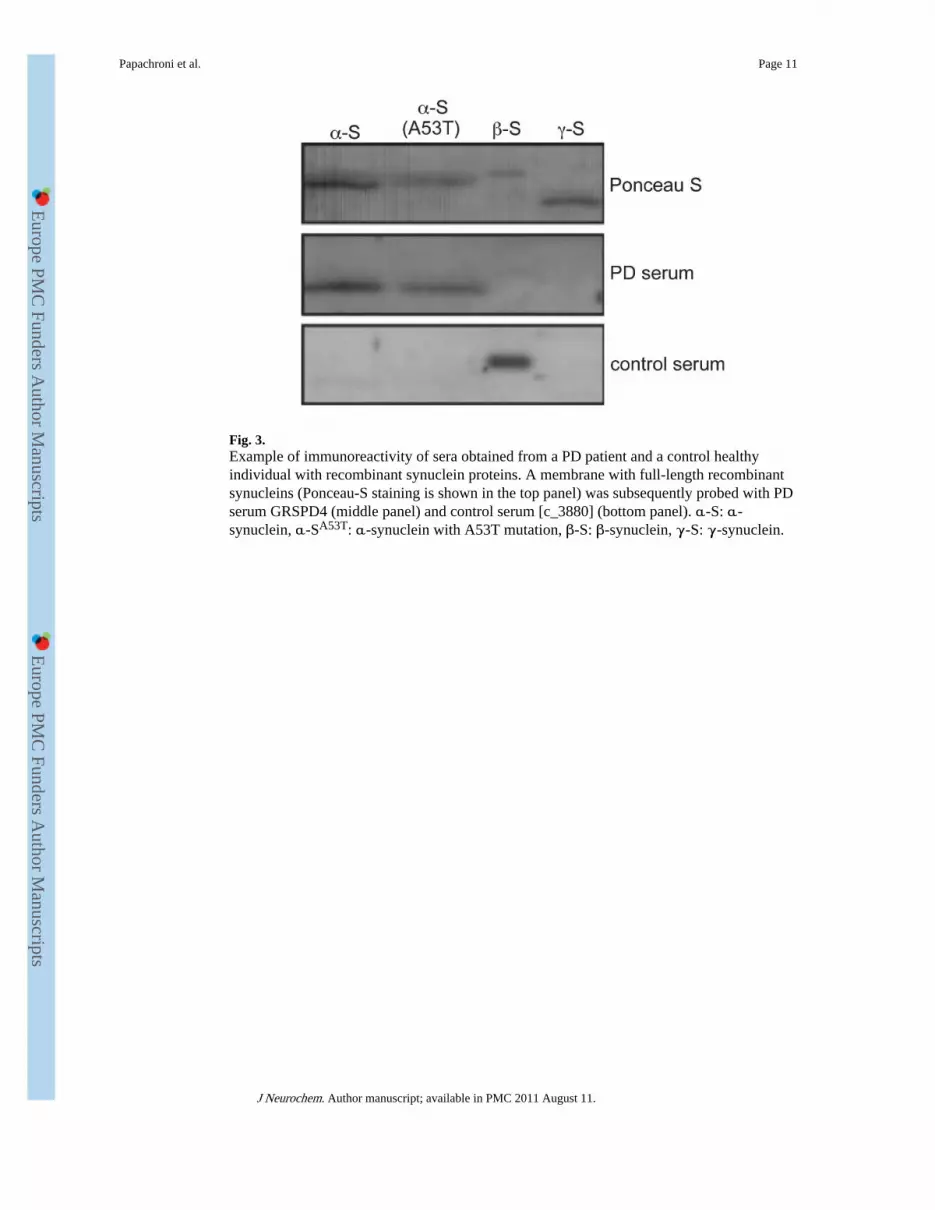
Fig. 3.Example of immunoreactivity of sera obtained from a PD patient and a control healthyindividual with recombinant synuclein proteins. A membrane with full-length recombinantsynucleins (Ponceau-S staining is shown in the top panel) was subsequently probed with PDserum GRSPD4 (middle panel) and control serum [c_3880] (bottom panel). α-S: α-synuclein, α-SA53T: α-synuclein with A53T mutation, β-S: β-synuclein, γ-S: γ-synuclein.
Papachroni et al. Page 11
J Neurochem. Author manuscript; available in PMC 2011 August 11.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Fig. 4.(a). Schematic representation of the GST-fused peptides of α-synuclein used in this study.The domain organisation of the α-synuclein molecule with the positions of the three knownpoint mutations associated with hereditary forms of PD is shown above the scheme of theoverlapping GST-fusion peptides 3, 6, 99, 305 and 317. (b). Immunoreactivity of PD serumGRSPD5 against the full-length synuclein proteins and the GST-peptides of α-synuclein.The numbers on top of the lanes correspond to the peptides shown on panel a. This serumshows immune reaction with full-length α-synuclein and α-synuclein peptide 99 but notwith other α-synuclein peptides.
Papachroni et al. Page 12
J Neurochem. Author manuscript; available in PMC 2011 August 11.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
Papachroni et al. Page 13
Tabl
e 1
Dem
ogra
phic
dat
a of
PD
pat
ient
s
Spor
adic
(n =
31)
Fam
ilial
(n =
20)
Con
trol
s(n
= 2
6)
Age
at t
ime
of s
tudy
, ave
rage
, yea
rs (
SD)
65.1
(11
.6)
66.1
(12
.7)
64.9
(10
.9)
Age
at o
nset
, ave
rage
, yea
rs (
SD)
57.9
(13
.7)
56.7
(12
.5)
NA
Fem
ale,
(%
)12
(38
.7)
7 (3
5.0)
10 (
38.5
)
His
tory
of
smok
ing
(%)
12 (
38.7
)3
(15.
0)7
(26.
9)
His
tory
of
trau
mat
ic b
rain
inju
ry, T
BI
(%)
5 (1
6.1)
3 (1
5.0)
5 (1
9.2)
His
tory
of
expo
sure
in p
estic
ides
(%
)14
(45
.2)
7 (3
5.0)
8 (3
0.8)
Clin
ical
cha
ract
eris
tics
and
find
ings
, num
ber
Ons
et w
ith tr
emor
(%
)12
(38
.7)
11 (
55.0
)N
A
Bra
dyki
nesi
a (%
)16
(51
.6)
14 (
70.0
)N
A
Mus
cula
r ri
gidi
ty (
%)
15 (
48.4
)9
(45.
0)N
A
Post
ural
inst
abili
ty (
%)
17 (
54.8
)10
(50
.0)
NA
Pers
iste
nt a
sym
met
ry o
f si
gns
(%)
2 (6
.5)
1 (0
.05)
NA
Res
t tre
mor
(%
)23
(74
.2)
17 (
85.0
)N
A
Hoe
hn a
nd Y
ahn
scor
e (s
tagi
ng 0
–5)
mea
n (S
D)
2.5
(1.3
)2.
4 (1
.7)
NA
J Neurochem. Author manuscript; available in PMC 2011 August 11.

Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
Papachroni et al. Page 14
Tabl
e 2
Inci
denc
e of
α-
and β-
synu
clei
n A
Ab
in th
e pe
riph
eral
blo
od s
erum
of
PD a
nd c
ontr
ol g
roup
. The
pre
senc
e of
AA
b w
as r
evea
led
by im
mun
oblo
t ass
ayw
ith f
ull-
leng
th r
ecom
bina
nt p
rote
ins
as a
ntig
ens.
The
num
ber
of s
erum
sam
ples
in e
ach
grou
p po
sitiv
e fo
r α
-syn
ucle
in a
utoa
ntib
ody
(α-s
yn+),
neg
ativ
efo
r α
-syn
ucle
in a
utoa
ntib
ody
(α-s
yn−),
pos
itive
for
β-s
ynuc
lein
aut
oant
ibod
y (β
-syn
+)
and
nega
tive
for β-
synu
clei
n au
toan
tibod
y (β
-syn
−)
is s
how
n
α-S
yn−
α-S
yn+
Tot
alO
R (
95%
CI)
pβ-
Syn−
β-Sy
n+T
otal
OR
(95
% C
I)p
Con
trol
188
26R
efer
ence
NA
1313
26R
efer
ence
NA
Fam
ilial
PD
218
2020
.25
(3.7
7–10
8.83
)<
0.00
115
520
0.33
(0.
09–1
.19)
0.12
9
Spor
adic
PD
1615
312.
11 (
0.71
–6.2
8)0.
278
229
310.
41 (
0.14
–1.2
2)0.
172
Tot
al P
D18
3351
4.13
(1.
50–1
1.34
)0.
007
3714
510.
38 (
0.14
–1.0
1)0.
076
J Neurochem. Author manuscript; available in PMC 2011 August 11.

Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
Papachroni et al. Page 15
Tabl
e 3
Cor
rela
tion
for
the
pres
ence
of α
- an
d β-
synu
clei
n A
Ab
in th
e pe
riph
eral
blo
od s
erum
of
PD p
atie
nts
with
pre
disp
osin
g (t
raum
atic
bra
in in
jury
, pes
ticid
eex
posu
re)
or o
ther
fac
tors
(in
heri
tanc
e, d
isea
se s
tage
, sex
, age
and
sm
okin
g) r
elat
ed to
the
dise
ase
α-S
ynuc
lein
β-Sy
nucl
ein
Fac
tor
p (F
ishe
r’s
exac
t te
st)
OR
(95
% C
I)p
(Fis
her’
s ex
act
test
)O
R (
95%
CI)
Sex
0.07
3.33
(0.
99–1
1.12
)0.
991.
10 (
0.31
–3.9
4)
Age
0.99
0.85
(0.
27–2
.69)
0.99
0.85
(0.
25–2
.91)
Smok
e0.
202.
86 (
0.68
–11.
92)
0.73
1.50
(0.
40–5
.57)
TB
I0.
200.
48 (
0.11
–2.2
2)0.
193.
3 (0
.70–
15.6
4)
Pest
icid
es0.
202.
45 (
0.71
–8.4
3)0.
531.
64 (
0.48
–5.6
8)
PD s
tage
0.20
2.13
(0.
64-7
.02)
0.99
0.88
0.2
6–3.
10)
Inhe
rita
nce
0.02
5.67
(1.
37-2
3.46
)0.
520.
58 (
0.15
–2.2
4)
J Neurochem. Author manuscript; available in PMC 2011 August 11.

Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
Papachroni et al. Page 16
Tabl
e 4
Ass
ocia
tive
anal
ysis
of
the
antig
enic
ity o
f α
-syn
ucle
in f
ragm
ents
aga
inst
dis
ease
-rel
ated
fac
tors
Con
stru
ctSe
xIn
heri
tanc
eA
geSm
oke
TB
IP
esti
cide
sP
D s
tage
3p
1.00
0.29
0.30
1.00
0.61
0.73
1.00
OR
0.95
(0.
20–4
.41)
2.75
(0.
66–1
1.54
)2.
36 (
0.58
–9.5
8)1.
27 (
0.30
–5.3
3)2.
80 (
0.26
–30.
18)
0.70
(0.
18–2
.77)
0.88
(0.
22–3
.45)
99p
1.00
1.00
0.73
0.47
1.00
0.49
0.49
OR
0.95
(0.
20–4
.41)
0.98
(0.
25–3
.96)
1.43
(0.
36–5
.66)
2.2
(0.5
0–9.
61)
0.81
(0.
10–6
.58)
1.88
(0.
47–7
.53)
0.53
(0.
13–2
.14)
317
p1.
000.
280.
080.
471.
000.
730.
49
OR
0.70
(0.
14–3
.49)
2.74
(0.
64–1
1.75
)4.
18 (
0.94
–18.
61)
0.50
(0.
12–2
.13)
2.11
(0.
19–2
2.90
)0.
70 (
0.17
–2.8
5)0.
51 (
0.12
–2.1
2)
6p
0.46
0.50
0.49
0.30
0.11
0.30
0.49
OR
2.00
(0.
40–9
.91)
1.69
(0.
41–6
.88)
1.88
(0.
47–7
.53)
2.28
(0.
54–9
.67)
0.64
(0.
13–3
.17)
2.36
(0.
58–9
.58)
1.88
(0.
47–7
.53)
305
p0.
710.
070.
730.
070.
340.
080.
17
OR
1.48
(0.
32–6
.90)
4.80
(1.
07–2
1.45
)0.
69 (
0.18
–2.7
3)4.
88 (
1.01
–23.
57)
0.27
(0.
03–2
.92)
4.03
(0.
95–1
7.22
)3.
06 (
0.74
–12.
63)
J Neurochem. Author manuscript; available in PMC 2011 August 11.











![[The incidence of the pituitary autoantibodies in Graves' disease]](https://img.dokumen.tips/doc/110x75/635915fc68c5ffb42104606f/the-incidence-of-the-pituitary-autoantibodies-in-graves-disease.jpg)

















