Embed Size (px)
Citation preview

doi:10.1182/blood-2006-08-040139Prepublished online November 21, 2006;
Anna Krop-Watorek, Gregory R Halverson, Marcin Czerwinski and Steven L SpitalnikDavid Austin Schirmer, Shuh-Chyung Song, Jeffrey P Baliff, Stephanie O Harbers, Raphael A Clynes, Mouse models of IgG- and IgM-mediated hemolysis
(238 articles)Transfusion Medicine �Articles on similar topics can be found in the following Blood collections
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
digital object identifier (DOIs) and date of initial publication. theindexed by PubMed from initial publication. Citations to Advance online articles must include
final publication). Advance online articles are citable and establish publication priority; they areappeared in the paper journal (edited, typeset versions may be posted when available prior to Advance online articles have been peer reviewed and accepted for publication but have not yet
Copyright 2011 by The American Society of Hematology; all rights reserved.20036.the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 1 of 31
Mouse models of IgG- and IgM-mediated hemolysis
David A. Schirmer*, Shuh-Chyung Song*, Jeffrey P. Baliff**, Stephanie O. Harbers¶, Raphael
A. Clynes¶, Anna Krop-Watorek¶¶, Gregory R. Halverson†, Marcin Czerwinski¶¶, and Steven L.
Spitalnik*
Affiliations note: From the Departments of Pathology* and Microbiology¶, Columbia
University, New York, NY; Department of Pathology and Laboratory Medicine, University of
Rochester, Rochester, NY**; Ludwik Hirszfeld Institute of Immunology and Experimental
Therapy, Wroclaw, Poland¶¶; and the Department of Immunochemistry, New York Blood
Center, New York, NY†
Corresponding Author and Reprints: Steven L. Spitalnik, Department of Pathology, 630 West
168th St, College of Physicians & Surgeons of Columbia University, New York, NY 10023
Phone: 212-305-2204; FAX: 212-305-3693; email: [email protected]
Grant support: Supported, in part, by grants from the National Blood Foundation (to S.L.S.)
and the MetLife Foundation (to G.R.H.).
Statement of authorship: All authors participated in designing and performing the research;
D.A.S and S.L.S controlled and analyzed the data; D.A.S and S.L.S. wrote the paper; and all
authors critiqued various drafts and checked the final version of the manuscript.
Left running head: SCHIRMER et al
Right running head: MOUSE MODELS OF ALLOIMMUNE HEMOLYSIS
Journal section designation: TRANSFUSION MEDICINE
Abbreviations:
HTR: hemolytic transfusion reaction; IgM/G-HTR: IgM/G-mediated HTR; RBC: red blood
cell(s); AIHA: autoimmune hemolytic anemia; Tg: transgenic or transgene; KO: knockout;
mRBC: mouse RBC; hGPA: human glycophorin A; MAb: monoclonal antibody; CWB: column
Blood First Edition Paper, prepublished online November 21, 2006; DOI 10.1182/blood-2006-08-040139
Copyright © 2006 American Society of Hematology
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 2 of 31
wash buffer; RT: room temperature; SN: supernatant; NS: normal saline; GVB: gelatin veronal
buffer; mPCR: multiplex polymerase chain reaction; FISH: fluorescence in situ hybridization;
EDTA: ethylenediaminetetraacetic acid; pRBC: packed RBC
Abstract: 179 words
Text: Introduction (286) + Materials and Methods (1175) + Results (1505) +
Discusssion (1617) = 4573 words
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 3 of 31
ABSTRACT
Well-characterized mouse models of alloimmune antibody-mediated hemolysis would provide a
valuable approach for gaining greater insight into the pathophysiology of hemolytic transfusion
reactions. To this end, mouse red blood cells (mRBC) from human glycophorin A transgenic
(hGPA-Tg) donor mice were transfused into non-Tg recipients that had been passively
immunized with IgG or IgM hGPA-specific monoclonal antibodies (MAb). In this novel murine
“blood group system,” mRBC from hGPA-Tg mice are “antigen positive” and mRBC from non-
Tg mice are “antigen negative.” Passive immunization of non-Tg mice with the IgG1 10F7 and
IgG3 NaM10-2H12 anti-hGPA Mab each induced rapid clearance of incompatible transfused
hGPA-Tg-mRBC in a dose-response manner. Using various knockout (KO) mice as transfusion
recipients, both the complement system and activating Fcγ receptors were found to be important
in the clearance of incompatible mRBC by each of these IgG Mab. In addition, the IgM E4 anti-
hGPA Mab induced complement-dependent intravascular hemolysis of transfused incompatible
hGPA-Tg-mRBC accompanied by gross hemoglobinuria. These initial studies validate the
relevance of these new mouse models for addressing important questions in the field of
transfusion medicine.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 4 of 31
INTRODUCTION
Hemolytic transfusion reactions (HTRs) are dangerous complications of blood transfusion. IgM-
mediated HTRs (IgM-HTRs) occur acutely, are usually due to ABO incompatibility, typically
cause intravascular hemolysis, and can lead to shock, renal failure, coagulopathy, and death1.
Although infrequent, IgM-HTRs have a high mortality rate. IgG-mediated HTRs (IgG-HTRs),
are more common, but usually less severe, than IgM-HTRs; they can occur acutely or be
delayed, typically cause extravascular hemolysis, and occasionally result in renal failure,
coagulopathy, and death2. Symptomatic immune-mediated red blood cell (RBC) destruction also
occurs in autoimmune hemolytic anemia (AIHA)3, blood-group incompatible transplantation4,
and immune thrombocytopenic purpura treated with Rh-immune globulin5. Although various
different modalities are used to treat HTRs, there is no definitive evidence regarding efficacy.
Given the sporadic nature of HTRs, designing human trials to evaluate treatment options is
difficult. Therefore, to study the mechanisms underlying HTRs, to evaluate the efficacy of
existing treatments, and to develop new treatments, a relevant, inexpensive, and tractable animal
model is required6. A mouse model would be ideal for this purpose7, given the abundance of
reagents and relevant knockout (KO) and transgenic (Tg) animals. Although mouse blood group
polymorphisms exist8, and transfused incompatible mouse RBC (mRBC) are rapidly cleared9,10,
no one has exploited these findings to develop a model of HTRs. The availability of a Tg mouse
expressing human glycophorin A (hGPA) on mRBC11 offers an approach using a well-described
glycoprotein target and well-characterized reagents.
We describe initial studies validating mouse models of IgG- and IgM-mediated
alloimmune hemolysis. Thus, mRBC from hGPA-Tg donors were transfused into non-Tg
recipients that were passively immunized with IgG or IgM hGPA-specific monoclonal antibodies
(MAb). The clearance of the transfused mRBC was quantified and the roles of complement and
Fcγ receptors were evaluated.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 5 of 31
MATERIALS AND METHODS
Antibodies
Purified MAb NaM26-2H12 (IgG3), NaM26-3F4 (IgG1), and NaM10-6G4 (IgG2a), which
recognize peptide epitopes on the M and N forms of hGPA12, were purchased from EFS (Nantes,
France). Hybridomas producing MAb 6A7, an IgG1 specific for a sialic acid-dependent epitope
on M-type hGPA13-15, 10F7, an IgG1 recognizing a non-polymorphic epitope on hGPA13,14, and
J11d, a rat IgM recognizing CD24 on mRBC16,17, were purchased from the ATCC (Manassas,
VA). The hybridoma producing MAb N92, an IgG2a anti-N Mab18, was provided by Elwira
Lisowska (Ludwik Hirszfeld Institute, Wroclaw, Poland). The hybridoma producing MAb E4, an
IgM that recognizes a non-polymorphic epitope present on hGPA19, was provided by Marilyn
Telen (Duke University, Durham, NC).
Throughout culture and MAb purification, endotoxin contamination was minimized. IgG
MAbs were purified by immunoaffinity chromatography. Thus, 5 ml of goat anti-mouse IgG
agarose (Sigma, St. Louis, MO) in a disposable 10 ml chromatography column (Pierce,
Rockford, IL) were equilibrated with 50 ml of column wash buffer (CWB; 50 mM Tris, pH 8.0,
prepared using endotoxin-free water and 1.0 M Tris-HCl, pH 8.0 (Mediatech, Herndon, VA)). A
peristaltic pump and pyrogen-free tubing (VWR, West Chester, PA) were used to perfuse the
column with 100 ml of conditioned medium or ascites at 1 ml/min overnight at 4°C. The column
was then washed with 50 ml of CWB and eluted with Immunopure IgG elution buffer (Pierce,
Rockford, IL) in 4 ml fractions into tubes containing 1 ml of 1.0 M Tris-HCl in endotoxin-free
water, pH. 8.0. Peak fractions (i.e. A280nm > 0.050) were pooled, changed into endotoxin-free
Tris-buffered saline (20 mM Tris-HCl, pH 7.4, 0.15 M NaCl (Teknova, Hollister, CA)), and
concentrated to 1 mg/ml (determined by the BCA Assay (Pierce)) using Centriprep YM-10
centrifugal filters (Millipore, Billerica, MA). Similarly, the IgM J11d and E4 MAbs were
purified using a protein L column (Pierce) and the mannose binding protein-based Immunopure
IgM Purification Kit (Pierce), respectively.
All MAbs used in vivo were purified to electrophoretic homogeneity. Endotoxin levels
were quantified using the LAL QCL-1000 kit (Cambrex, East Rutherford, NJ). The purified
MAbs were clear solutions with no visible precipitates; however, they were not further evaluated
to exclude the presence of higher oligomers or aggregates. To measure Mab activity,
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 6 of 31
hemagglutination titers were determined (see below), using commercially-available reagent-
quality human RBC (Ortho-Clinical Diagnostics, Raritan, NJ). To evaluate IgM Mab function in
vitro, hemolytic complement assays were performed. Thus, RBC were incubated for 1h at room
temperature (RT) in MAb E4 or J11d diluted in normal saline (NS; 0.154 M NaCl, pH 7.4). Ten
microliters of washed RBC (see below) in NS (2 x 108 cells/ml) were added to 190 µl of an
18.75% (v/v) mixture of guinea pig serum (Sigma) in gelatin veronal buffer +2 (GVB+2)
(Sigma) in a microtiter plate and incubated at 37°C for 30 min. Following centrifugation for 5
min at 4°C, SN absorbance at 414 nm was determined. Non-opsonized human RBC and mRBC
were negative controls. As positive controls, RBC were completely hemolyzed using distilled
water instead of GVB+2 buffer; after hemolysis, 10 µl of guinea pig serum were added.
Mice
Wild type C57BL/6 and BALB/C mice, and FcγRIIb knockout (KO) mice on the BALB/C
background, were from Taconic Labs (Hudson, NY). FVB/NJ mice were purchased from
Jackson Laboratories (Bar Harbor, ME). FcRγ KO20, complement C3 KO21, and FcγR/C3
double-KO mice (all on the C57BL/6 background) were maintained by Dr. Clynes; C3 KO
breeding pairs were provided by Michael Carroll (Harvard University, Boston, MA). Animal
protocols were approved by the Institutional Animal Care and Use Committee at the Columbia
University. Narla Mohandas (New York Blood Center) provided breeding pairs of hGPA-Tg
mice11, which were constructed on the FVB/NJ background using human BAC clone 159F4
(Catalog #982132; Invitrogen), which contains hGPA. FVB/NJ mice do not express functional
complement C5 protein22.
A multiplex polymerase chain reaction (mPCR) assay identified hGPA-Tg mice, using
genomic DNA extracted from tail biopsies to amplify a 291-bp hGPA sequence using forward
(5’-TATTAGCTCAGAGCCTCACACATT-3’) and reverse (5’-
GAAATTTCTGAAACTTCATGAGCTC-3’) primers; as a positive control, an 88 bp segment of
the mouse glycophorin gene Gypa23 was amplified (forward: 5’-CTTTGTCATTCGGCTGATA-
3’; reverse: 5’-TGATGATACTCATAATTTTGGG-3’). Fluorescence in situ hybridization
(FISH) was performed by standard methods using the 159F4 BAC clone to probe bone marrow
cells from femurs of wild type and hGPA-Tg mice.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 7 of 31
In humans, hGPA encodes the MN blood group antigens. The MN phenotype of hGPA-
Tg-mRBC was determined by standard microtiter plate hemagglutination assays using the 6A7
anti-M13,14 and N92 anti-N18 MAbs. Briefly, 25 µl aliquots of serial two-fold dilutions of the
MAbs were prepared in NS. Washed mRBC, obtained from retro-orbital plexus blood, were re-
suspended to a 3% hematocrit in NS and 25 µl aliquots were added to each well. Following a 1h
incubation at 37°C, agglutinates were identified macroscopically. Controls included reagent-
quality M+N-, M+N+, and M-N+ human RBC and wild type FVB/NJ mRBC.
Passive immunization
Prior to injection, MAbs were dialyzed against endotoxin-free NS (Phoenix Scientific, St.
Joseph, MO). Recipients received IgG MAbs by intraperitoneal injection 16h before transfusion;
alternatively, the IgM MAb was infused into the right jugular vein 2 min before transfusion.
Preparation of 51-Cr labeled mRBC
Donors were anesthetized, exsanguinated by cardiac puncture using 25-gauge needles and
heparin-coated syringes, and euthanized. For each transfusion, a 1 ml aliquot of a 3% suspension
of washed mRBC in a pyrogen-free 1.5 ml microfuge tube was centrifuged at 10,000 x g for 3s,
SN was removed, and packed RBC (pRBC) were re-suspended in 100 µl of endotoxin-free NS
and 20 µl of 51-Cr (1 mCi/ml; GE Healthcare, Piscataway, NJ). Following 30 min at RT, the
tube was centrifuged, SN removed, and the pRBC re-suspended in 100 µl of NS.
Transfusion of 51-Cr labeled mRBC
The recipients were anesthetized and, using sterile technique, a 1 cm vertical midline incision
was made and the left jugular vein exposed by blunt dissection. The 51-Cr labeled mRBC (~30
µl of pRBC) in a 25-gauge butterfly was transfused over 5s into the jugular vein. Following
transfusion, hemostasis was achieved, and the wound was closed. This procedure was completed
in less than 5 min. A zero timepoint (T=0) sample of ~25 µl of blood was then obtained from the
retro-orbital plexus.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 8 of 31
Survival of transfused RBC
At designated times post-transfusion, mice were anesthetized and an endpoint sample of ~25 µl
of retro-orbital plexus blood was obtained. Microhematocrit tubes containing the T=0 and
terminal blood samples were centrifuged for 1 min; the height of the pRBC column was
measured and the tubes analyzed for counts per minute (CPM) in a gamma counter (Perkin-
Elmer, Wellesley, MA). The percentage of radiolabeled mRBC remaining at time “n” following
transfusion (T=n) was calculated as:
{(CPM/mm of pRBC in the T=n sample)/(CPM/mm of pRBC in the T=0 sample)} x 100
Following the terminal time point sample, recipients were exsanguinated and euthanized.
Determination of hemoglobinuria
In some experiments, recipients were maintained in metabolic cages. Urine was collected from
metabolic cages while mice were alive; bladder urine was also collected at autopsy. Urine
absorbance was measured by wavelength scanning spectrophotometry at 300-700 nm using a
spectrophotometer (Nanodrop Technologies, Wilmington, DE).
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 9 of 31
RESULTS
Further characterization of hGPA-Tg mice
Animal husbandry
There were no behavioral differences between hGPA-Tg and wild type FVB/NJ mice. hGPA-Tg
mice were healthy, bred well, bore litters of expected size (averaging ~8 neonates/litter), and
nurtured their offspring attentively.
Serological characterization of hGPA-Tg-mRBC
In the initial study of hGPA-Tg mice, hGPA expression on mRBC was characterized using only
two anti-hGPA MAbs, which had no blood group MN specificity11. We extended these studies
using well-characterized anti-hGPA MAbs12-15,18,19. Hemagglutination with anti-M and -N MAbs
(6A7 and N92, respectively) revealed that hGPA-Tg-mRBC expressed the M allele (Table 1).
These results are particularly interesting because 6A7 recognizes a sialic acid-dependent
epitope15, and mouse and human RBC sialic acids differ24. In addition, the NaM16-1B10,
NaM10-2H12, NaM26-3F4, and 10F7 MAbs, which recognize non-allelic hGPA peptide
epitopes12-14, agglutinated human RBC and hGPA-Tg-mRBC, but not wild type mRBC (Table
1). Similar results were obtained with the hGPA peptide-specific NaM10-6G4 and E4 MAbs (not
shown). The variation in agglutination titer seen with 10F7 and NaM26-3F4 with different RBC
isolates is intriguing, but is not yet understood. Taken together, these results demonstrate that a
broad array of anti-hGPA MAbs recognize a glycosylated recombinant form of hGPA on hGPA-
Tg-mRBC.
Genotyping and phenotyping hGPA-Tg mice
We developed mPCR and hemagglutination assays to provide a definitive means of identifying
hGPA-Tg mice. Thus, mPCR of genomic DNA yielded a 218 bp band when the hGPA Tg was
present, and an 88 bp band corresponding to mouse glycophorin23, as an internal standard (Figure
1). In contrast, mPCR results from mice lacking the hGPA Tg yielded only the 88 bp mouse
glycophorin band (Figure 1). For older mice, hemagglutination assays using the 6A7 anti-hGPA
MAb allowed phenotypic confirmation of the presence or absence of the Tg. Thus, MAb 6A7
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 10 of 31
agglutinated both M+N- human RBC (positive control) and hGPA-Tg-mRBC, but not wild type
mRBC or M-N+ human RBC (negative controls; not shown). These assays always produced
concordant results.
Identification of the hGPA Tg integration site
Initially, it was not clear whether the hGPA Tg integrated at one or multiple loci11. FISH
provides a definitive means of examining chromosomal integration sites. As a positive control,
hGPA is a single copy gene in human leukocytes and maps to chromosome 4q28-q31 (Figure
2A). Similarly, in hGPA-Tg mouse bone marrow cells, the hGPA Tg integrated at a single locus
(Figure 2C-D). No hybridization was observed with wild type FVB/NJ mouse bone marrow cells
(Figure 2B).
RBC survival studies
IgG-mediated hemolysis in vivo of transfused incompatible mRBC
When transfused into wild type FVB/NJ and C57BL/6 mice that had been passively immunized
with varying amounts of the 10F7, NaM10-2H12, and NaM10-6G4 IgG MAbs, incompatible
hGPA-Tg-mRBC were promptly cleared from the circulation in amounts proportional to the
amount of infused anti-hGPA MAb. For example, when immunized with increasing amounts of
MAb 10F7, 24-hour RBC survival progressively decreased (Figure 3). Because of these results,
500 µg of MAb 10F7 were used subsequently. Similar dose-response experiments with NaM10-
2H12 and NaM10-6G4 (not shown), revealed that 50 µg of each of these MAbs would suffice for
inducing dramatic RBC clearance (Table 2). As negative controls, no destruction of transfused,
compatible, wild type mRBC was observed in mice immunized with any IgG anti-hGPA MAbs
(not shown). In addition, when an isotype-matched irrelevant IgG1 MAb was used as a control
for 10F7, the survival of hGPA-Tg-mRBC was normal at 24 hr (D.A.S, S.L.S., and James C.
Zimring, unpublished data). Similarly, no destruction of transfused hGPA-Tg-mRBC was
detected in unimmunized recipients (not shown and see Table 3, below). Therefore, despite the
H-2 difference between donors (FVB/NJ background) and recipients (i.e. C57BL/6 and
BALB/C), no “naturally-occurring” anti-H-2 antibodies caused accelerated mRBC
destruction8,10.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 11 of 31
Transfused incompatible hGPA-Tg-mRBC were rapidly cleared from passively
immunized mice. For example, in mice immunized with MAb 10F7 or NaM10-2H12,
incompatible hGPA-Tg-mRBC clearance was complete by 4 hours post-transfusion (Figure 4).
To evaluate the roles of activating Fcγ receptors and the complement system in IgG-
mediated clearance of incompatible mRBC, FcRγ KO, C3 KO, and FcγR/C3 double-KO mice,
all on the C57BL/6 background, were passively immunized with IgG anti-hGPA MAbs and
transfused with incompatible mRBC. When coated by IgG MAbs of different subclasses,
incompatible mRBC were cleared by activating different pathways. For example, as compared
with wild type C57BL/6 mice, IgG1-mediated clearance of hGPA-Tg-mRBC by MAb 10F7 was
markedly inhibited in FcRγ KO mice and only moderately inhibited in C3 KO mice (Table 3B).
Thus, although complement participates in IgG1-mediated clearance of hGPA-Tg-mRBC,
activating Fcγ receptors play a dominant role. In contrast, when immunized with the IgG3
NaM10-2H12 anti-hGPA MAb, hGPA-Tg-mRBC destruction was markedly inhibited in C3 KO
mice and only moderately inhibited in FcRγ KO mice (Table 3A); thus, complement is dominant,
with some contribution by activating Fcγ receptors. With both of these Mabs, clearance of
hGPA-Tg-mRBC was markedly, but not completely, inhibited in FcγR/C3 double-KO mice,
suggesting that other mechanisms may also be involved in mRBC clearance.
Inhibitory Fcγ receptors were also investigated using FcγRIIb KO mice (BALB/C
background). No difference in IgG3-mediated clearance of incompatible mRBC by Mab NaM10-
2H12 was observed when comparing FcγRIIb KO and wild type BALB/C mice (not shown).
This was not unexpected given the low affinity of FcγRII for IgG325; in addition, Kupffer cells,
which appear to play a major role in mRBC destruction (D.A.S. and S.L.S., unpublished data;
and Ref. 26-29), either lack FcγRIIb30 or have defective regulation of phagocytosis by FcγRIIb29.
To examine the role of terminal complement components in IgG-mediated mRBC
clearance, we compared wild type FVB/NJ mice, which lack complement component C522, with
wild type C57BL/6 mice. Clearance of hGPA-Tg-mRBC in MAb 10F7-immunized FVB/NJ
mice was markedly reduced compared with C5-replete C57BL/6 mice (Tables 2-3). This
suggests that complement C5 plays a significant role, perhaps because C5a enhances activating
Fcγ receptor function31. In contrast, there was little difference between FVB/NJ and C57BL/6
mice in the clearance of incompatible mRBC by the IgG3 NaM10-2H12 MAb (Tables 2-3).
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 12 of 31
These differences may relate to the individual epitope specificities, affinities, or IgG subclasses
of these two MAbs. In addition, it will be interesting to determine whether C5 is completely
responsible for the findings with 10F7 by using congenic mouse strains that differ only at the C5
locus32.
Complement mediated hemolysis in vitro of IgM-coated hGPA-Tg-mRBC
hGPA-Tg-mRBC are susceptible to complement-mediated hemolysis in vitro in the presence of
guinea pig serum. Thus, hGPA-Tg-mRBC coated in vitro with either the IgM E4 anti-GPA MAb
or the IgM J11d anti-CD24 MAb were hemolyzed following incubation in complement-
containing serum, as were wild type mRBC coated with J11d (Table 4). As negative controls,
wild type mRBC incubated with the E4 MAb, and uncoated hGPA-Tg-mRBC, were not
hemolyzed. Thus, the hGPA epitope density on hGPA-Tg-mRBC and the avidity of MAb E4
were sufficient to activate complement, up to and including formation of functional C5b-9
membrane attack complexes.
IgM mediated hemolysis in vivo of transfused incompatible hGPA-Tg-mRBC
Dose-response and time course studies demonstrated that passive immunization intravenously of
wild type C57BL/6 mice with 100 µg of the IgM anti-hGPA E4 MAb caused dramatic and very
rapid clearance of transfused incompatible hGPA-Tg-mRBC (Table 5). For example, by
comparing the CPM/mm of pRBC at T=0 of MAb E4-immunized recipients transfused with
either wild type or hGPA-Tg-mRBC, approximately 50% of the hGPA-Tg-mRBC were cleared
in the short time between transfusion and drawing the T=0 sample (i.e. 2-5 min). In addition, by
2h post-transfusion, hGPA-Tg-mRBC clearance in E4-immunized mice was virtually complete,
with little change at the 24hr time point (not shown). Finally, no significant RBC clearance
occurred in mice immunized with MAb E4 and transfused with wild type mRBC (Table 5).
By 2h post-transfusion, abundant, soluble, red-brown pigment was seen in the urine of all
MAb E4-immunized wild type C57BL/6 mice transfused with hGPA-Tg-mRBC (Figure 5A).
This pigment was not pelleted by centrifugation (not shown); thus, this color change was not due
to gross hematuria. However, spectrophotometry confirmed that the pigment was free
hemoglobin with a peak absorbance at 414 nm (Figure 5B); thus, the rapid clearance of
transfused hGPA-Tg-mRBC produced gross hemoglobinuria (Figure 5; Table 5). In contrast, no
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 13 of 31
abnormal pigment was seen with MAb E4-immunized mice transfused with wild type mRBC.
These results strongly suggest that the IgM E4 anti-hGPA MAb rapidly produced dramatic,
intravascular, complement-mediated hemolysis of transfused incompatible hGPA-Tg-mRBC.
To confirm the role of the membrane attack complex, FVB/NJ mice were used, which are
C5 deficient22. Interestingly, the survival of transfused hGPA-Tg-mRBC in MAb E4-immunized
FVB/NJ mice was only slightly less than that observed in complement-replete wild type
C57BL/6 mice (not shown). However, no hemoglobinuria was observed in FVB/NJ recipients.
Thus, although complement-mediated intravascular hemolysis was strikingly evident in C57BL/6
mice, the predominant clearance mechanism was probably CR3-mediated phagocytosis of
IgM/C3b-coated mRBC. This will be evaluated further in future studies using C3 KO and/or
CVF-treated, complement deficient mice. The specific role of CR3, as opposed to CRIg33, can
also be evaluated using CR3 KO mice34 and the M1/70 CR3-blocking MAb35,36. Finally, the role
of C5 can be explored further using congenic strains that only differ at this locus32.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 14 of 31
DISCUSSION
The need for mouse models to study immune-mediated hemolysis
Various animal models were used to study immune-mediated RBC destruction and its
sequelae7,37-40. However, these are not optimal for studying human HTRs due to their expense
and the limited flexibility. Although mRBC blood group systems were described8, and although
antibody-mediated clearance of transfused, incompatible mRBC occurs9,10, these polymorphisms
were not exploited to study HTRs. However, murine AIHA models, which clear mRBC
following infusion of monoclonal mRBC-specific autoantibodies26-28,41-44, allowed evaluation of
the roles of antibody isotype, IgG subclass, complement, and Fcγ receptors. Nonetheless,
incomplete knowledge of target epitopes, the inability to evaluate early kinetics of mRBC
destruction, and in vivo agglutination by IgM41,45, make these less than ideal models for human
HTRs. For example, autoantibody-induced agglutination in vivo almost never occurs in
humans46. Finally, transfusion of hen egg lysozyme (HEL) Tg mRBC into wild-type mice
actively immunized to HEL surprisingly caused removal of HEL from the mRBC, with no
mRBC destruction47. Therefore, the current study describes novel mouse models of alloantibody-
mediated intravascular and extravascular hemolysis, potentially suitable for studying HTR
pathogenesis.
The relevance of Tg hGPA for immune-mediated hemolysis
Using hGPA-Tg-mRBC to study antibody-induced mRBC clearance is highly relevant for the
human situation. Thus, hGPA is the most abundant human RBC glycoprotein48,49. Its primary
importance results from carrying the MN antigens48, glycopeptide antigens encoded by amino
acid polymorphisms at positions 1 and 550,51 that also depend on O-glycosylation at positions 2-
415,18,48,49,52,53. In addition, hGPA carries other types of blood group antigens54,55. Antibodies to
hGPA antigens cause HTRs56, hemolytic disease of the newborn57, and AIHA55,58,59.
The hGPA-Tg mouse provides a genetically inheritable, murine “blood group system.”
Thus, wild-type mRBC are “antigen-negative” and hGPA-Tg-mRBC are “antigen positive.”
Wild-type mice can be actively immunized with hGPA53 and many mouse MAbs are available12-
15,18,19,53,60-63. This type of blood group polymorphism (i.e. the presence or absence of an entire
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 15 of 31
protein) also occurs in humans: En(a-) RBC lack hGPA, S-s-U- RBC lack human glycophorin B,
and “Rh-negative” RBC lack the Rh(D) protein.
IgG-mediated hemolysis in passively immunized mice transfused with hGPA-Tg-mRBC
This study provides useful insights into the pathogenesis of antibody-mediated hemolysis. For
example, the kinetics of IgG1- and IgG3-mediated clearance of hGPA-Tg-mRBC (Figure 4;
Table 2) correlated well with that seen in human IgG-HTRs64. In addition, although complement
participates in IgG1-mediated clearance of hGPA-Tg-mRBC, Fcγ receptors play a dominant role
(Table 3). Interestingly, in a recent study, the IgG1 subclass caused minimal platelet destruction
due to its strong binding to the inhibitory FcγRII25; the efficacy of the IgG1 10F7 MAb in hGPA-
Tg-mRBC clearance is consistent with these findings due to the liver being the likely locus of
RBC clearance in mice (Ref. 26-29; D.A.S. and S.L.S., unpublished data) and the absence of, or
defective regulation by, FcγRII on mouse Kupffer cells29,30. Indeed, when using IgG class-switch
variants of a monoclonal mRBC-specific autoantibody to induce AIHA, the IgG1 variant was
moderately pathogenic27,43; furthermore, this variant’s pathogenicity resulted from its interaction
with activating Fcγ receptors. Therefore, the efficacy 10F7 (Table 2; Figures 3-4) correlates well
with what is known of IgG1’s in murine immune-mediated hematological disorders.
The difference between FVB/NJ and C57BL/6 mice in IgG1-mediated mRBC clearance
(Tables 2-3) was particularly intriguing because FVB/NJ mice are deficient in C5. Therefore,
C5a and/or the C5b-C9 membrane attack complex may be important in mRBC clearance by
MAb 10F7. In contrast, in a mouse AIHA model (i.e. comparing mRBC clearance in DBA/2 (C5
deficient) and BALB/C mice (C5 replete)), C5 was not involved in the anemia induced by
several mRBC-specific MAbs, including an IgG141. Thus, our alloimmune IgG-HTR model
provides novel information as compared to existing AIHA mouse models. Additional studies
using congenic mice that only differ at the C5 locus32 will further clarify these findings.
Both complement and activating Fcγ receptors are important in IgG3-mediated clearance
by MAb NaM10-2H12, with complement being dominant (Table 3). Although in the initial
description murine IgG3 did not activate complement65, later studies clearly demonstrated
complement activation26,66,67. Our results that activating Fcγ receptors contribute to IgG3-
mediated clearance are particularly interesting in light of recent papers concerning murine IgG3
antibodies. Thus, it was originally proposed that a unique IgG3-specific Fcγ receptor facilitates
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 16 of 31
phagocytosis of IgG3-coated mRBC by murine macrophages68. In addition, recent results
suggest that no known murine Fcγ receptors function as IgG3 receptors because they do not bind
IgG3 in a quantitative binding assay25. Our results in vivo with FcRγ KO mice (Table 3) are
consistent with this hypothesis; that is, there may be an additional, as yet undescribed, activating
FcγR requiring the FcRγ chain. Finally, when human RBC or hGPA-Tg-mRBC are coated in
vitro with NaM10-2H12 in serum-free medium, they are phagocytosed by RAW 264.769 mouse
macrophages (D.A.S., S.L.S., and Sunny Seo, unpublished data), also suggesting the presence of
an activating Fcγ receptor.
In contrast, others suggested that FcγRI is the only IgG3 receptor, in that IgG3 binds to
transfected cells expressing recombinant FcγRI and macrophages from FcγRI KO mice do not
phagocytose IgG3-coated RBC70,71. Interestingly, although FcRγ KO mice express ~20% of
normal levels of FcγRI, they cannot phagocytose IgG3-coated RBC because phagocytosis
presumably requires the ITAM function of FcRγ70.
Finally, in mouse ITP25 and AIHA models26,27 IgG3 antibodies are either non-pathogenic
or slightly pathogenic, respectively. In addition, in the latter, only complement was involved for
this mild effect. However, in our in vivo studies NaM10-2H12 led to rapid and pronounced
clearance of incompatible mRBC involving both complement and Fcγ receptors. These results
are similar to those found with an IgG3 monoclonal anti-mRBC autoantibody72, although the
roles of complement and Fcγ receptors were not assessed. Thus, the mechanisms of mRBC
clearance by IgG3 Mabs may depend on additional factors, such as antigen specificity and
antigen affinity. Studies directly comparing these systems (e.g. infusing anti-hGPA Mabs into
hGPA-Tg mice as an AIHA model, and constructing Tg mice expressing hGPA on platelets as an
ITP model) would provide additional insights.
Thus, this new model has characteristics that will deepen and extend the mechanistic
understanding of IgG-mediated RBC destruction beyond that provided by existing models.
IgM-mediated hemolysis in passively immunized mice transfused with hGPA-Tg-mRBC
The IgM E4 anti-hGPA MAb produced dramatic, intravascular, complement-mediated hemolysis
of transfused hGPA-Tg-mRBC (Table 5; Figure 5). To our knowledge, this represents the first
time that an IgM has been effectively used to cause intravascular hemolysis in mice. Although
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 17 of 31
previous studies used monoclonal IgM anti-RBC MAbs to induce AIHA, the anemia was due to
intravascular agglutination41. In contrast, given the small quantities of incompatible mRBC
transfused in the current study, their clearance could only be caused by complement-mediated
hemolysis and/or phagocytosis. Furthermore, the rapid kinetics of this IgM-mediated clearance
agrees well with that in human IgM-HTRs64. Interestingly, despite the presence of the CD59
complement regulatory protein on hGPA-Tg-mRBC, and despite the impression that mouse
complement is “weak”73, E4 caused complement-mediated intravascular hemolysis. Thus, this
model will prove useful in unraveling and clarifying the pathogenic mechanisms of IgM-HTRs.
IgM-mediated intravascular hemolysis could also be enhanced by constructing hGPA-Tg x
CD59 KO mice74 and using these mRBC for transfusion.
When IgM-opsonized RBC were previously transfused into guinea pigs37,75, they were
rapidly cleared by the liver, and then released back into the circulation within 24h. The
mechanism involved complement activation and adherence of IgM/C3b-coated RBC to
complement receptors (presumably CR3), but not phagocytosis. In contrast, passively infused
IgM E4 MAb not only rapidly cleared hGPA-Tg-mRBC, but the cleared mRBC were also not
released back into the circulation (Table 5). One explanation is that the CR3 integrins in our
mice were already activated (even though the mice are maintained in a barrier facility and less
than 0.5 µg of endotoxin contaminated the infused E4 MAb). Alternatively, the newly described
CRIg complement receptor33, which does not require prior activation, may be responsible.
Finally, unique aspects of the specificity and/or affinity of MAb E4 may be relevant.
These mouse studies provided a surprising result that may be relevant to the pathogenesis
of human IgM-HTRs. Thus, complement-mediated intravascular hemolysis has long been
thought to be primarily responsible for destroying transfused incompatible RBC, as well as for
the associated morbidity and mortality of human IgM-HTRs. Indeed, the signs of human IgM-
HTRs, including hemoglobinemia and hemoglobiunuria, provide solid evidence of intravascular
hemolysis. However, without studying IgM-HTRs in a controlled setting, it has not been possible
to determine the extent to which intravascular hemolysis clears transfused, incompatible RBC.
For example, the dramatic hemoglobinuria in MAb E4-immunized C57BL/6 recipients
transfused with hGPA-Tg-mRBC provides clear evidence of intravascular hemolysis (Figure 5).
Coupled with its rapid kinetics, it initially seemed that most, if not all, of the incompatible
mRBC were destroyed intravascularly. Thus, the results of MAb E4-mediated mRBC clearance
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 18 of 31
in C5-deficient FVB/NJ recipients were particularly interesting. As expected, no hemoglobinuria
occurred in any FVB/NJ mice; however, these recipients cleared incompatible mRBC with the
same kinetics and almost the same efficiency as the complement-replete C57BL/6 recipients.
This suggests that, in contrast to what is believed with human IgM-HTRs, although complement
does cause intravascular hemolysis of transfused, IgM-coated RBC, the IgM-mediated clearance
of the incompatible RBC may be predominantly due to complement-mediated extravascular
hemolysis resulting from phagocytosis by hepatic Kupffer cells and/or splenic macrophages.
Studies evaluating complement receptors (e.g. CR3 KO mice), early components in the
complement cascade (e.g. C3 KO mice), and macrophage phagocytosis (e.g. using liposomal
chlodronate) will provide more insight.
Future directions
Future studies will evaluate the relevance of this mouse model for human HTRs. In the
current study, small amounts of radiolabeled incompatible mRBC were transfused into
anesthetized mice. Despite rapid clearance of transfused mRBC, recipients did not display the
clinical sequelae seen in human HTRs. Therefore, future studies will transfuse large quantities of
incompatible mRBC and will monitor various physiological parameters76.
Acknowledgements: This work was initially inspired by conversations with Dr. Louis Fink. The
authors wish to thank Drs. Elwira Lisowska, Marilyn Telen, and Narla Mohandas for providing
valuable reagents, Dr. Murty Vundavalli for performing the FISH analysis, Drs. Raymond Baggs
and Carl Pinkert for advice regarding the design of mouse experiments, Mary Bolognino and
Sunny Seo for technical assistance during the initial studies, Drs. Robert Pierce and James
Zimring for helpful discussions, and Dr. Michael Shelanski for support and encouragement.
All authors participated in designing and performing the research; D.A.S and S.L.S
controlled and analyzed the data; D.A.S and S.L.S. wrote the paper; and all authors critiqued
various drafts and checked the final version of the manuscript.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 19 of 31
FIGURE LEGENDS
Figure 1. Identification of hGPA-Tg mice by mPCR. The presence of the hGPA Tg was
detected using mPCR of genomic DNA extracted from biopsied tail tissue with the primers
described in Materials and Methods. This yielded a 219 bp band if the hGPA Tg was present.
Amplification of the endogenous mouse glycophorin gene was used as an internal standard and
yielded an 88 bp band in each case.
Figure 2. Localizing the hGPA gene by FISH. FISH was performed using the 159F4 BAC
probe to examine the chromosomal location of the hGPA gene, which encodes the hGPA protein.
As a positive control, the probe hybridized to human chromosome 4 (A), and, as a negative
control, the probe did not hybridize to any wild type FVB/NJ mouse chromosome (B). However,
the probe did hybridize to one chromosome from hemizygous hGPA-Tg mice (C-D). The
locations of hybridization are indicated by white arrowheads.
Figure 3. IgG1-mediated clearance of incompatible mRBC. Wild type FVB/NJ mice were
passively immunized intraperitoneally with the indicated amounts of purified 10F7, an anti-
hGPA IgG1 MAb; 16h later they were transfused with 51Cr-labeled wild type (i.e. (-) control) or
hGPA-Tg-mRBC, and 24h RBC survivals were quantified. Results are recorded as percentage of
transfused RBC remaining in the vasculature after 24 hours for groups of 2-5 mice (mean + 1
SD). These results show that incompatible hGPA-Tg-mRBC were cleared from the circulation of
mice in proportion to the dose of purified 10F7 MAb that they received.
Figure 4. IgG-mediated clearance of incompatible hGPA-Tg-mRBC: Time course studies.
Wild type FVB/NJ mice were passively immunized intraperitoneally with 500 µg of the purified
10F7 anti-hGPA IgG1 MAb (A) or with 50 µg of the purified NaM10-2H12 anti-hGPA IgG3
MAb (B). Sixteen hours later they were transfused with 51Cr-labeled hGPA-Tg-mRBC, and then
RBC survival was quantified at the indicated time points. The number of mice evaluated at each
time point is indicated in parentheses; the mean + 1 SD is provided for each time point.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 20 of 31
Figure 5. Gross hemoglobinuria in wild type C57BL/6 mice following passive immunization
with the IgM E4 anti-hGPA MAb and transfusion with incompatible hGPA-Tg-mRBC.
Mice were passively immunized intravenously with 100 µg of the purified IgM E4 MAb. (A)
Representative post-transfusion urine samples were obtained at autopsy from mice receiving
incompatible hGPA-Tg-mRBC (left sample) or receiving compatible wild type mRBC (right
sample). (B) Urine samples from mice transfused with incompatible hGPA-Tg-mRBC displayed
absorbance peaks at 414 nm (for example, curve A); no absorbance peak at 414 nm was
detectable in any urine sample obtained from mice transfused with compatible wild type mRBC
(for example, curve B).
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 21 of 31
Table 1. Serological characterization of hGPA expressed on the surface of hGPA-Tg-mRBC.
MAb 6A7 N92 NaM16-1B10 10F7 NaM10-2H12 NaM26-3F4 Specificity M N hGPA hGPA hGPA hGPA Isotype IgG1 IgG2a IgM IgG1 IgG3 IgG1 Source of RBC RBC type Human #1 M+N- >128 0 64 16 1600 64
Human #2 M-N+ 0 + 64 16 1600 32
Mouse #1 hGPA-Tg 128 0 64 64 1600 8
Mouse #2 hGPA-Tg 128 0 64 128 1600 8
Mouse #3 hGPA-Tg >128 0 64 128 1600 8
Mouse #4 wild type 0 0 0 0 0 0
A panel of mouse MAbs recognizing either a sialic acid-dependent M epitope on hGPA (i.e. 6A7), a sialic-independent N epitope on hGPA (i.e. N92), or peptide epitopes on both M- and N-type hGPA (i.e. NaM16-1B10, 10F7, NaM10-2H12, and NaM26-3F4) were used to evaluate hGPA expression on hGPA-Tg-mRBC. Human RBC of defined MN type were positive controls. RBC from a wild type FVB/NJ mouse were used as a negative control. The inverse agglutination titers are provided in each case, except for the results of the N92 MAb with the RBC obtained from Human #2, in which strong agglutination was seen, but titers were not performed. As additional controls, wild type and hGPA-Tg-mRBC were agglutinated by a rabbit polyclonal anti-mRBC IgG antibody (not shown). Table 2. IgG anti-hGPA MAbs rapid cleared hGPA-Tg-mRBC in vivo.
Mab IgG subclass # of mice % 4h RBC survival (mean + 1 SD) 10F7 IgG1 5 59.9 + 3.4
NaM10-6G4 IgG2a 4 29.6 + 9.2 NaM10-2H12 IgG3 10 17.0 + 7.2
Wild type FVB/NJ recipient mice that had been passively immunized IP with MAb 10F7 (500 µg), NaM10-6G4 (50 µg), or NaM10-2H12 (50 µg), were then transfused 16 hours later with 51Cr-labeled hGPA-Tg-mRBC and the 4 hour RBC survivals were determined as described in Materials and Methods.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 22 of 31
Table 3. Evaluation of the role of activating Fcγ receptors and complement component C3 in IgG-mediated clearance of incompatible hGPA-Tg-mRBC.
A
Mice Passive Immunization # of mice % 4h RBC survival (mean + 1 SD) Wild type None 6 98.2 + 1.6 Wild type NaM10-2H12 6 26.8 + 8.1 FcRγ KO NaM10-2H12 2 56.5 + 0.4 C3 KO NaM10-2H12 6 81.7 + 7.2
FcγR/C3 double-KO NaM10-2H12 6 80.6 + 9.1
B
Mice Passive Immunization # of mice % 4h RBC survival (mean + 1 SD) Wild type None 6 98.2 + 1.6 Wild type 10F7 6 27.2 + 14.5 FcRγ KO 10F7 6 85.7 + 19.9 C3 KO 10F7 6 58.6 + 4.5
FcγR/C3 double-KO 10F7 4 80.2 + 9.2 All mice were on the C57BL/6 background. Mice were passively immunized IP with either 50 µg of the IgG3 NaM10-2H12 anti-hGPA MAb (Panel A) or 500 µg of the IgG1 10F7 anti-hGPA MAb (Panel B) 16h prior to transfusion. All mice were then transfused with 51Cr-labeled hGPA-Tg-mRBC. In mice immunized with 10F7, clearance of incompatible mRBC was highly dependent on the presence of activating Fcγ receptors, and moderately dependent on C3. In contrast, clearance of incompatible RBC in mice immunized with NaM10-2H12 was moderately dependent on activating Fcγ receptors and highly dependent on C3. Table 4. Complement mediated lysis in vitro of IgM-opsonized mRBC.
MAb Test RBC J11d (2 µg/ml) E4 (0.25 µg/mL) E4 (0.1 µg/mL) E4 (0.05 µg/ml) Wild type mRBC 91.8 0.0 0.0 1.9 hGPA-Tg-mRBC 118.0 67.9 38.9 31.9
Wild type and hGPA-Tg-mRBC were incubated with distilled water as a positive control (defined as 100% hemolysis) or with guinea pig serum alone as a negative control (defined as background hemolysis). Test samples contained guinea pig serum as a source of complement and defined concentrations of purified MAbs. Hemolysis was determined by spectrophotometric evaluation of absorbance at 414 nm of the supernatant of each reaction and the results are presented as percent hemolysis compared to the distilled water control. All samples were tested in duplicate and the average results are presented. Binding of the mRBC-specific J11d anti-CD24 IgM MAb led to hemolysis of both types of mRBC; the E4 anti-hGPA IgM MAb only hemolyzed hGPA-Tg-mRBC.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom
Page 23 of 31
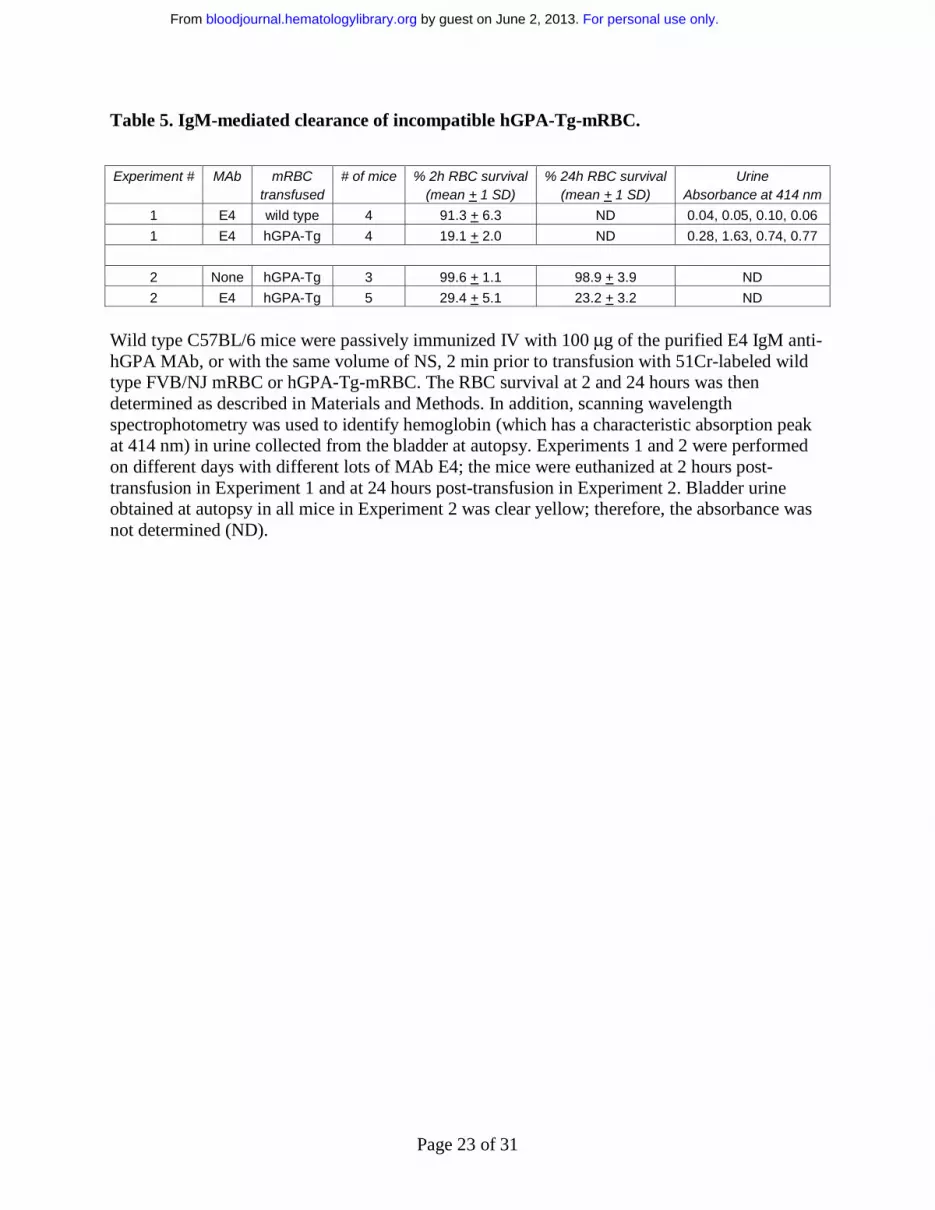
Table 5. IgM-mediated clearance of incompatible hGPA-Tg-mRBC. Experiment # MAb mRBC
transfused # of mice % 2h RBC survival
(mean + 1 SD) % 24h RBC survival
(mean + 1 SD) Urine
Absorbance at 414 nm
1 E4 wild type 4 91.3 + 6.3 ND 0.04, 0.05, 0.10, 0.06
1 E4 hGPA-Tg 4 19.1 + 2.0 ND 0.28, 1.63, 0.74, 0.77
2 None hGPA-Tg 3 99.6 + 1.1 98.9 + 3.9 ND
2 E4 hGPA-Tg 5 29.4 + 5.1 23.2 + 3.2 ND
Wild type C57BL/6 mice were passively immunized IV with 100 µg of the purified E4 IgM anti-hGPA MAb, or with the same volume of NS, 2 min prior to transfusion with 51Cr-labeled wild type FVB/NJ mRBC or hGPA-Tg-mRBC. The RBC survival at 2 and 24 hours was then determined as described in Materials and Methods. In addition, scanning wavelength spectrophotometry was used to identify hemoglobin (which has a characteristic absorption peak at 414 nm) in urine collected from the bladder at autopsy. Experiments 1 and 2 were performed on different days with different lots of MAb E4; the mice were euthanized at 2 hours post-transfusion in Experiment 1 and at 24 hours post-transfusion in Experiment 2. Bladder urine obtained at autopsy in all mice in Experiment 2 was clear yellow; therefore, the absorbance was not determined (ND).
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 24 of 31
100
101
102
103
105
104
107
106
108
Agglutination - - -+ + + + + +
Mouse #
219 bp
88 bp
100
101
102
103
105
104
107
106
108
Agglutination - - -+ + + + + +
Mouse #
219 bp219 bp
88 bp88 bp
Human 4q28-q31 Wild-type FVB mouse
hGPA-Tg mouse hGPA-Tg mouse
BA
C D
Human 4q28-q31 Wild-type FVB mouse
hGPA-Tg mouse hGPA-Tg mouse
Human 4q28-q31 Wild-type FVB mouseHuman 4q28-q31 Wild-type FVB mouse
hGPA-Tg mouse hGPA-Tg mousehGPA-Tg mouse hGPA-Tg mouse
BA
C D
Figure 1
Figure 2
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 25 of 31
0
20
40
60
80
100
(-) Control; 500 (2) 50 (2) 200 (3) 500 (5)
Passive Immunization (10F7: ug per mouse (# mice))
% 2
4 h
r su
rviv
al o
f h
GP
A-T
g m
RB
C
A B
0
20
40
60
80
100
0 4 8 12 16 20 24
Time (hrs)
% S
urv
ival
of
hG
PA
-Tg
mR
BC
(4)
(5)(2)
(5)
(5)
0
20
40
60
80
100
0 4 8 12 16 20 24
Time (hrs)
% S
urv
ival
of
hG
PA
-Tg
mR
BC
(4)
(5)(2)
(5)
(5)
0
20
40
60
80
100
0 1 4 8 16 24
Time (hrs)
%S
urv
ival
of
hG
PA
-Tg
mR
BC
(4) (10)
(4)
(2)(7)
0
20
40
60
80
100
0 1 4 8 16 24
Time (hrs)
%S
urv
ival
of
hG
PA
-Tg
mR
BC
(4) (10)
(4)
(2)(7)
Figure 3
Figure 4
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 26 of 31
Figure 5
A B
A B
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 27 of 31
REFERENCES
1. Capon SM, Goldfinger D. Acute hemolytic transfusion reaction, a paradigm of the systemic inflammatory response: new insights into pathophysiology and treatment. Transfusion. 1995;35:513-520.
2. Pineda AA, Vamvakas EC, Gorden LD, Winters JL, Moore SB. Trends in the incidence of delayed hemolytic and delayed serologic transfusion reactions. Transfusion. 1999;39:1097-1103.
3. Ulvestad E, Berentsen S, Bo K, Shammas FV. Clinical immunology of chronic cold agglutinin disease. European Journal of Haematology. 1999;63:259-266.
4. Salmon JP, Michaux S, Hermanne JP, et al. Delayed massive immune hemolysis mediated by minor ABO incompatibility after allogeneic peripheral blood progenitor cell transplantation. Transfusion. 1999;39:824-827.
5. Gaines AR. Disseminated intravascular coagulation associated with acute hemoglobinemia or hemoglobinuria following Rh(0)(D) immune globulin intravenous administration for immune thrombocytopenic purpura. Blood. 2005;106:1532-1537.
6. Davenport RD. Pathophysiology of hemolytic transfusion reactions. Seminars in Hematology. 2005;42:165-168.
7. Ness PM, Shirey RS, Weinstein MH, King KE. An animal model for delayed hemolytic transfusion reactions. Transfusion Medicine Reviews. 2001;15:305-317.
8. Stimpfling JH, Reichert AE, Blanchard S. Genetic control of the immune response to the Ea-2 alloantigen system of the mouse. I. The humoral antibody response. Journal of Immunology. 1976;116:1096-1098.
9. Erickson RP, Herzenberg LA, Goor R. Partial elimination of homologous red blood cells in mice. Transplantation. 1964;2:175-182.
10. Moller G. Biologic properties of 19S and 7S mouse isoantibodies directed against isoantigens of the H-2 system. Journal of Immunology. 1966;96:430-439.
11. Auffray I, Marfatia S, de Jong K, et al. Glycophorin A dimerization and band 3 interaction during erythroid membrane biogenesis: in vivo studies in human glycophorin A transgenic mice. Blood. 2001;97:2872-2878.
12. Rasamoelisolo M, Czerwinski M, Bruneau V, Lisowska E, Blanchard D. Fine characterization of a series of new monoclonal antibodies directed against glycophorin A. Vox Sanguinis. 1997;72:185-191.
13. Bigbee WL, Vanderlaan M, Fong SS, Jensen RH. Monoclonal antibodies specific for the M- and N-forms of human glycophorin A. Molecular Immunology. 1983;20:1353-1362.
14. Bigbee WL, Langlois RG, Vanderlaan M, Jensen RH. Binding specificities of eight monoclonal antibodies to human glycophorin A--studies with McM, and MkEn(UK) variant human erythrocytes and M- and MNV-type chimpanzee erythrocytes. Journal of Immunology. 1984;133:3149-3155.
15. Blackall DP, Ugorski M, Pahlsson P, Shakin-Eshleman SH, Spitalnik SL. A molecular biologic approach to study the fine specificity of antibodies directed to the MN human blood group antigens. Journal of Immunology. 1994;152:2241-2247.
16. Bruce J, Symington FW, McKearn TJ, Sprent J. A monoclonal antibody discriminating between subsets of T and B cells. Journal of Immunology. 1981;127:2496-2501.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 28 of 31
17. Nielsen PJ, Lorenz B, Muller AM, et al. Altered erythrocytes and a leaky block in B-cell development in CD24/HSA-deficient mice. Blood. 1997;89:1058-1067.
18. Wasniowska K, Duk M, Steuden I, Czerwinski M, Wiedlocha A, Lisowska E. Two monoclonal antibodies recognizing different epitopes of blood group N antigen. Archivum Immunologiae et Therapiae Experimentalis. 1988;36:623-632.
19. Telen MJ, Scearce RM, Haynes BF. Human erythrocyte antigens. III. Characterization of a panel of murine monoclonal antibodies that react with human erythrocyte and erythroid precursor membranes. Vox Sanguinis. 1987;52:236-243.
20. Clynes R, Ravetch JV. Cytotoxic antibodies trigger inflammation through Fc receptors. Immunity. 1995;3:21-26.
21. Wessels MR, Butko P, Ma M, Warren HB, Lage AL, Carroll MC. Studies of group B streptococcal infection in mice deficient in complement component C3 or C4 demonstrate an essential role for complement in both innate and acquired immunity. Proceedings of the National Academy of Sciences of the United States of America. 1995;92:11490-11494.
22. Taketo M, Schroeder AC, Mobraaten LE, et al. FVB/N: an inbred mouse strain preferable for transgenic analyses. Proceedings of the National Academy of Sciences of the United States of America. 1991;88:2065-2069.
23. Matsui Y, Natori S, Obinata M. Isolation of the cDNA clone for mouse glycophorin, erythroid-specific membrane protein. Gene. 1989;77:325-332.
24. Varki A, Kornfeld S. An autosomal dominant gene regulates the extent of 9-O-acetylation of murine erythrocyte sialic acids. A probable explanation for the variation in capacity to activate the human alternate complement pathway. Journal of Experimental Medicine. 1980;152:532-544.
25. Nimmerjahn F, Ravetch JV. Divergent immunoglobulin G subclass activity through selective Fc receptor binding. Science. 2005;310:1510-1512.
26. Azeredo da Silveira S, Kikuchi S, Fossati-Jimack L, et al. Complement activation selectively potentiates the pathogenicity of the IgG2b and IgG3 isotypes of a high affinity anti-erythrocyte autoantibody. Journal of Experimental Medicine. 2002;195:665-672.
27. Fossati-Jimack L, Ioan-Facsinay A, Reininger L, et al. Markedly different pathogenicity of four immunoglobulin G isotype-switch variants of an antierythrocyte autoantibody is based on their capacity to interact in vivo with the low-affinity Fcgamma receptor III. Journal of Experimental Medicine. 2000;191:1293-1302.
28. Fossati-Jimack L, Reininger L, Chicheportiche Y, et al. High pathogenic potential of low-affinity autoantibodies in experimental autoimmune hemolytic anemia. Journal of Experimental Medicine. 1999;190:1689-1696.
29. Schiller C, Janssen-Graalfs I, Baumann U, et al. Mouse FcgammaRII is a negative regulator of FcgammaRIII in IgG immune complex-triggered inflammation but not in autoantibody-induced hemolysis. European Journal of Immunology. 2000;30:481-490.
30. Kumar V, Ali SR, Konrad S, et al. Cell-derived anaphylatoxins as key mediators of antibody-dependent type II autoimmunity in mice. Journal of Clinical Investigation. 2006;116:512-520.
31. Shushakova N, Skokowa J, Schulman J, et al. C5a anaphylatoxin is a major regulator of activating versus inhibitory FcgammaRs in immune complex-induced lung disease. Journal of Clinical Investigation. 2002;110:1823-1830.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 29 of 31
32. Kyriakides C, Austen W, Jr., Wang Y, et al. Skeletal muscle reperfusion injury is mediated by neutrophils and the complement membrane attack complex. American Journal of Physiology - Heart & Circulatory Physiology. 1999;277:C1263-1268.
33. Helmy KY, Katschke KJ, Jr., Gorgani NN, et al. CRIg: a macrophage complement receptor required for phagocytosis of circulating pathogens. Cell. 2006;124:915-927.
34. Rosenkranz AR, Coxon A, Maurer M, et al. Impaired mast cell development and innate immunity in Mac-1 (CD11b/CD18, CR3)-deficient mice. Journal of Immunology. 1998;161:6463-6467.
35. Skeel A, Leonard EJ. Action and target cell specificity of human macrophage-stimulating protein (MSP). Journal of Immunology. 1994;152:4618-4623.
36. Thornton BP, Vetvicka V, Pitman M, Goldman RC, Ross GD. Analysis of the sugar specificity and molecular location of the beta-glucan-binding lectin site of complement receptor type 3 (CD11b/CD18). Journal of Immunology. 1996;156:1235-1246.
37. Schreiber AD, Frank MM. Role of antibody and complement in the immune clearance and destruction of erythrocytes. II. Molecular nature of IgG and IgM complement-fixing sites and effects of their interaction with serum. Journal of Clinical Investigation. 1972;51:583-589.
38. Giger U, Akol KG. Acute hemolytic transfusion reaction in an Abyssinian cat with blood type B. Journal of Veterinary Internal Medicine. 1990;4:315-316.
39. Giger U, Gelens CJ, Callan MB, Oakley DA. An acute hemolytic transfusion reaction caused by dog erythrocyte antigen 1.1 incompatibility in a previously sensitized dog. Journal of the American Veterinary Medical Association. 1995;206:1358-1362.
40. Lopas H, Birndorf NI, Bell CE, Jr., Robboy SJ, Fortwengler HP, Biddison WE. Experimental transfusion reactions in monkeys: haemolytic, coagulant and renal effects of transfused isoimmune IgG and IgM. British Journal of Haematology. 1972;23:765-776.
41. Shibata T, Berney T, Reininger L, et al. Monoclonal anti-erythrocyte autoantibodies derived from NZB mice cause autoimmune hemolytic anemia by two distinct pathogenic mechanisms. International Immunology. 1990;2:1133-1141.
42. Berney T, Shibata T, Merino R, et al. Murine autoimmune hemolytic anemia resulting from Fc gamma receptor- mediated erythrophagocytosis: protection by erythropoietin but not by interleukin-3, and aggravation by granulocyte-macrophage colony- stimulating factor. Blood. 1992;79:2960-2964.
43. Izui S, Fossati-Jimack L, da Silveira SA, Moll T. Isotype-dependent pathogenicity of autoantibodies: analysis in experimental autoimmune hemolytic anemia. Springer Seminars in Immunopathology. 2001;23:433-445.
44. Jordan MB, van Rooijen N, Izui S, Kappler J, Marrack P. Liposomal clodronate as a novel agent for treating autoimmune hemolytic anemia in a mouse model. Blood. 2003;101:594-601.
45. Shibata T, Kindler V, Chicheportiche Y, Vassalli P, Izui S. Interleukin 3 perfusion prevents death due to acute anemia induced by monoclonal antierythrocyte autoantibody. Journal of Experimental Medicine. 1990;171:1809-1814.
46. Nowak-Wegrzyn A, King KE, Shirey RS, Chen AR, McDonough C, Lederman HM. Fatal warm autoimmune hemolytic anemia resulting from IgM autoagglutinins in an infant with severe combined immunodeficiency. Journal of Pediatric Hematology/Oncology. 2001;23:250-252.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 30 of 31
47. Zimring JC, Hair GA, Chadwick TE, et al. Nonhemolytic antibody-induced loss of erythrocyte surface antigen. Blood. 2005;106:1105-1112.
48. Tomita M, Marchesi VT. Amino-acid sequence and oligosaccharide attachment sites of human erythrocyte glycophorin. Proceedings of the National Academy of Sciences of the United States of America. 1975;72:2964-2968.
49. Pisano A, Redmond JW, Williams KL, Gooley AA. Glycosylation sites identified by solid-phase Edman degradation: O-linked glycosylation motifs on human glycophorin A. Glycobiology. 1993;3:429-435.
50. Blumenfeld OO, Adamany AM. Structural polymorphism within the amino-terminal region of MM, NN, and MN glycoproteins (glycophorins) of the human erythrocyte membrane. Proceedings of the National Academy of Sciences of the United States of America. 1978;75:2727-2731.
51. Wasniowska K, Drzeniek Z, Lisowska E. The amino acids of M and N blood group glycopeptides are different. Biochemical & Biophysical Research Communications. 1976;76:385-390.
52. Judd WJ, Issitt PD, Pavone BG, Anderson J, Aminoff D. Antibodies that define NANA-independent MN-system antigens. Transfusion. 1979;19:12-18.
53. Czerwinski M, Siemaszko D, Siegel DL, Spitalnik SL. Only selected light chains combine with a given heavy chain to confer specificity for a model glycopeptide antigen. Journal of Immunology. 1998;160:4406-4417.
54. Ugorski M, Blackall DP, Pahlsson P, Shakin-Eshleman SH, Moore J, Spitalnik SL. Recombinant Miltenberger I and II human blood group antigens: the role of glycosylation in cell surface expression and antigenicity of glycophorin A. Blood. 1993;82:1913-1920.
55. Silberstein LE, Goldman J, Kant JA, Spitalnik SL. Comparative biochemical and genetic characterization of clonally related human B-cell lines secreting pathogenic anti-Pr2 cold agglutinins. Archives of Biochemistry & Biophysics. 1988;264:244-252.
56. Parry-Jones N, Gore ME, Taylor J, Treleaven JG. Delayed haemolytic transfusion reaction caused by anti-M antibody in a patient receiving interleukin-2 and interferon for metastatic renal cell cancer. Clinical Laboratory Haematology. 1999;21:407-408.
57. Rearden A, Frandson S, Carry JB. Severe hemolytic disease of the newborn due to anti-Vw and detection of glycophorin A antigens on the Miltenberger I sialoglycoprotein by Western blotting. Vox Sanguinis. 1987;52:318-321.
58. Garratty G, Arndt P, Domen R, et al. Severe autoimmune hemolytic anemia associated with IgM warm autoantibodies directed against determinants on or associated with glycophorin A. Vox Sanguinis. 1997;72:124-130.
59. Brain MC, Prevost JM, Pihl CE, Brown CB. Glycophorin A-mediated haemolysis of normal human erythrocytes: evidence for antigen aggregation in the pathogenesis of immune haemolysis. British Journal of Haematology. 2002;118:899-908.
60. Czerwinski M, Blackall DP, Abrams WR, Rubocki RJ, Spitalnik SL. Restricted VH gene usage by murine hybridomas directed against the human N, but not M, blood group antigen. Molecular Immunology. 1994;31:279-288.
61. Czerwinski M, Krop-Watorek A, Siegel DL, Spitalnik SL. A molecular approach for isolating high-affinity Fab fragments that are useful in blood group serology. Transfusion. 1999;39:364-371.
62. Anstee DJ, Lisowska E. Monoclonal antibodies against glycophorins and other glycoproteins. Journal of Immunogenetics. 1990;17:301-308.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

Page 31 of 31
63. Lisowska E, Messeter L, Duk M, Czerwinski M, Lundblad A. A monoclonal anti-glycophorin A antibody recognizing the blood group M determinant: studies on the subspecificity. Molecular Immunology. 1987;24:605-613.
64. Mollison PL. Survival curves of incompatible red cells: An analytical review. Transfusion. 1986;26:43-50.
65. Grey HM, Hirst JW, Cohn M. A new mouse immunoglobulin: IgG3. Journal of Experimental Medicine. 1971;133:289-304.
66. Fulpius T, Lemoine R, Berney T, Pastore Y, Moll S, Izui S. Polymorphonuclear leukocytes play a key role in the generation of "wire- loop" lesions induced by a murine IgG3 rheumatoid factor. Kidney International. 1996;49:647-655.
67. Hjelm F, Carlsson F, Verbeek S, Heyman B. IgG3-mediated enhancement of the antibody response is normal in FcgammaRI-deficient mice. Scandinavian Journal of Immunology. 2005;62:453-461.
68. Diamond B, Yelton DE. A new Fc receptor on mouse macrophages binding IgG3. Journal of Experimental Medicine. 1981;153:514-519.
69. Raschke WC, Baird S, Ralph P, Nakoinz I. Functional macrophage cell lines transformed by Abelson leukemia virus. Cell. 1978;15:261-267.
70. Barnes N, Gavin AL, Tan PS, Mottram P, Koentgen F, Hogarth PM. FcgammaRI-deficient mice show multiple alterations to inflammatory and immune responses. Immunity. 2002;16:379-389.
71. Gavin AL, Barnes N, Dijstelbloem HM, Hogarth PM. Identification of the mouse IgG3 receptor: implications for antibody effector function at the interface between innate and adaptive immunity. Journal of Immunology. 1998;160:20-23.
72. Meryhew NL, Handwerger BS, Messner RP. Monoclonal antibody-induced murine hemolytic anemia. Journal of Laboratory and Clinical Medicine. 1984;104:591-601.
73. Ish C, Ong GL, Desai N, Mattes MJ. The specificity of alternative complement pathway-mediated lysis of erythrocytes: a survey of complement and target cells from 25 species. Scandinavian Journal of Immunology. 1993;38:113-122.
74. Qin X, Krumrei N, Grubissich L, et al. Deficiency of the mouse complement regulatory protein mCd59b results in spontaneous hemolytic anemia with platelet activation and progressive male infertility. Immunity. 2003;18:217-227.
75. Schreiber AD, Frank MM. Role of antibody and complement in the immune clearance and destruction of erythrocytes. I. In vivo effects of IgG and IgM complement-fixing sites. Journal of Clinical Investigation. 1972;51:575-582.
76. Nieswandt B, Echtenacher B, Wachs FP, et al. Acute systemic reaction and lung alterations induced by an antiplatelet integrin gpIIb/IIIa antibody in mice. Blood. 1999;94:684-693.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom




















![IGM janfeb11-Ipatov[1]](https://img.dokumen.tips/doc/110x75/63457d2b596bdb97a908f42a/igm-janfeb11-ipatov1.jpg)








