Embed Size (px)
Citation preview

CLINICAL IMMUNOLOGY AND IMMUNOPATHOLOGY 53, 113-122 (1989)
Human Polyclonal and Monoclonal IgG and IgM Complement 3 Nephritic Factors: Evidence for ldiotypic Commonality
GEORGE C. TSOKOS, ANN E. STITZEL, ANEETA D. PATEL, MISAKO HIRAMATSU, JAMES E. BALOW, AND ROGER E. SPITZER
Kidney Diseases Section, National Institute of Diabetes and Digestive und Kidney Diseases, Department of Medicine. Uniformed Services University of the Health Sciences,
Bethesda, Maryland 20892; Department of Pediatrics, State University of New York, Svrucuse. Neti3 York 13210; and Department of Pediatrics, Kumamoto University,
Kumamoto, Japan
Complement 3 nephritic factors (C3NeF) were isolated from the sera of patients with membranoproliferative glomerulonephritis (MPGN) and the supematants of pokeweed mitogen-stimulated mononuclear cells from patients with MPGN. Three human mono- clonal C3NeF antibodies (two IgGs, CK and PH. and one IgM, K3C4) were established. Using an exhaustive series of affinity columns, we isolated antiC3NeF idiotypic anti- bodies (anti-IdNeF l (three from normal and two from patient sera). Anti-IdNeF prepara- tions bound to F(ab’),-NeF and prevented its ability to stabilize C3bBb convertase. We have used the above reagents to address questions on the genesis and the diversity of C3NeF antibodies. The following results were obtained: All anti-IdN” preparations bound to C3NeF isolated from patient sera. ceil culture supernatants. and IgG and IgM monoclonal C3NeF. None of the monoclonal C3NeF bound to an extensive battery of common antigens, including Fc portion of IgG, TNP. P-galactosidase, DNA, and bacterial products. These data indicate that C3NeF express one common idiotype and that these antibodies are not raised in response to an obvious antigen. B 1989 Academic
PET\. Inc.
INTRODUCTION
Complement 3 nephritic factors (C3NeF) are unique autoantibodies character- ized by their ability to stabilize the C3bBb convertase of the alternative pathway of complement (I-5); they were initially detected in the sera of patients with membranoproliferative glomerulonephritis (MPGN) and partial lipodystrophy and later in the supernatants of polyclonally stimulated normal and MPGN patient mononuclear cells (MNC) (6). Monoclonal C3NeF antibodies have been success- fully produced from Epstein-Barr virus (EBV)-transformed MNC of MPGN pa- tients (7).
Patients with systemic autoimmune disorders have circulating autoantibodies and anti-idiotypic antibodies which are thought to have an immunoregulatory role (g-13). Questions pertinent to the genesis of autoantibodies in general are also applicable to the origin of C3NeF autoantibodies. C3NeF antibodies bind specif- ically to a neoantigen arising from the formation of the C3bBb complex (5, 15). The structure of the C3NeF target neoantigen is not known. It is possible that more than one neoantigen is formed, recognizable by different idiotypic C3NeF antibodies. Hence, the first question refers to the number of C3NeF antibodies (utilizing different variable and/or other immunoglobulin regions) in one patient or different patients. Various autoantibodies have been found to cross-react with
II3 0090-1229189 $1.50 Copyright 0 1989 hy Academic Press, Inc. All rights of reproduction in any form reserved.

113 I SOKOS hl \ I
substances contained by infectious agents, suggesting a pathogenic role for the latter (12-16). The second question, in reference to the origin of C3NeF antibod- ies, relates to the possibility that these antibodies may cross-react with environ- mental pathogens.
The availability of (a) C3NeF preparations from various patients. (b) human monoclonal C3NeF of both IgG and IgM class, and (c) C3NeF anti-idiotypic antibodies has enabled us to address the above two questions. Our data indicate that the various ClNeF preparations share one idiotype determined by one or more specific VDJ immunoglobulin gene rearrangement which recognizes one specific epitope expressed as neoantigen following the formation of the C3bBb convertase complex.
MATERIALS AND METHODS
Reagents. Supplies for column chromatography and protein separations includ- ing CM Sephadex C-50, Sephadex G-200, Sephacryl 200 Superfine, and CNBr- activated Sepharose 4B were purchased from Pharmacia (Piscataway, NJ). DEAE-cellulose was obtained from Bio-Rad (Richmond. CA) and DEAE- cellulose from Whatman (Hillsboro, OR).
Goat antibody specific for Fc fragments of human IgG was obtained from Coo- per Biomedical Inc. (Malvern. PA). Rabbit antibody to goat IgG, alkaline phos- phatase-labeled goat antibody to human IgG, and affinity-purified pooled human IgG were purchased from Sigma (St. Louis, MO). Monospecific antibodies to IgG. C7, C8, and albumin were purchased from Miles Laboratories (Naperville, IL). A human IgM monoclonal antibody specific for TNP and cross-reacting with P-galactosidase has been prepared in our laboratory (17). P-galactosidase, single- stranded DNA, and Fc portion of IgG were purchased from Sigma. Tetanus toxoid was purchased from Lederle Laboratories (Pearl River, NY).
Trypsin and soybean trypsin inhibitor came from Worthington Diagnostic Sys- tems, Inc. (Freehold, NJ); protein A-Sepharose from Pharmacia (Piscataway): Linbro PVC microtiter plates from Flow Laboratories, Inc. (McLean, VA); p- nitrophenyl phosphate (Sigma-104), pepsin, Tween 20, and BSA from Sigma: octanoic acid from Fisher Scientific (Rochester, NY); and diethanolamine from Eastman Kodak (Rochester, NY). Rat serum came from our own laboratory animals.
Preparution of puri$ed wrnplerne~zt components and cellular intermedicrtes. Preparation of C3, factor B, factor D, and factor H are standard procedures in our laboratory and were done by established and published techniques (15).
EC3bBb cells were prepared by incubating sheep erythrocytes (E) with purified C3 in the presence of trypsin so that the amount of bound C3b was equivalent to approximately one C3b hemolytic site/cell (18). Following incubation with trypsin, soybean trypsin inhibitor was added and the cells were centrifuged and washed. Purified factors B and D (18) were then added in amounts necessary to provide one hemolytic site/cell.
Sera. Sera were obtained from patients with MPGN; the histologic types of

HUMAN C3 NEPHRITIC FACTORS 115
MPGN were determined by light, electron, and immunofluorescence microscopy of the renal tissue. Normal human serum was obtained from normal, healthy donors.
MNC isolation. MNC from three patients with MPGN were obtained from heparinized peripheral blood by standard Ficoll-Paque gradient centrifugation. Purified B cells were obtained by depleting T cells which were rosetted with sheep erythrocytes and separated on Ficoll-Paque gradients.
Establishment of lines. B cell-enriched MNC in RPM1 1640 culture medium, supplemented with (10%) fetal calf serum, were infected with 1 ml of EBV- containing supernatant of a lymphoblastoid B cell line (B95-8) and were incubated at 106/ml in 10 ml of medium in tissue culture flasks (Costar 3050; Costar, Cam- bridge, MA) for 20 days at 37°C in a humidified atmosphere containing 5% CO?. On Day 20, the cells were subcultured in larger flasks with 100 ml of media. Subsequently the cells were cloned by the limiting dilution method. Culture wells were screened for growth and were tested for Ig secretion by ELISA. Clones have been developed which were stable and which secreted C3NeF; these have been maintained in culture. Two of them secrete IgG (CK and PH) and one IgM (K3C4) (7, 17).
Cell culture techniques. Usually I x 10’ mononuclear cells in RPM1 containing Hepes and 20% fetal calf serum were added to plastic Falcon tubes. Pokeweed mitogen at a concentration of 5 pgiculture was then added and the cells were cultured at 37°C for 14 days (culture conditions found to be optimal) in 5% CO,.
Isolation and characterization of C3NeF. C3NeF has been isolated and purified from the sera of patients with MPGN. Serum was first separated by DEAE- cellulose column chromatography and was then applied to a column of CM Seph- adex C-50. C3NeF was eluted and separated from properdin with a linear NaCl gradient. Further purification was achieved using molecular sieve chromatogra- phy on Sephadex G-200.
After precipitation and dialysis, the IgG fraction was further purified on DEAE- cellulose. All fractions prepared in this way contained no properdin and were identified as a single IgG protein. Purity of the preparations was tested in all cases by SDS-polyacrylamide gel electrophoresis. A similar procedure was followed to isolate IgG from cell culture supernatants and monoclonal cell line supernatants.
Isolation oj’speci’c C3NeF. The above procedures yielded a purified IgG prep- aration containing C3NeF. To further isolate specific C3NeF. packed cells bear- ing EC3bBb were mixed with the IgG preparation at 30°C for 15 min. The IgG fraction was adsorbed with E and EC3b as well as with purified factor B prior to the reaction with EC3bBb in an effort to remove other antibodies which might bind to the cells in addition to C3NeF. Adsorptions were done at 0°C for 10 min and were usually repeated three times. The reacted EC3bBb cells were then centrifuged, washed, and eluted in 0.02 M EDTA at 37°C for 2 hr. The elution was repeated three times and the eluates were combined and concentrated. No further C3NeF remained on the cells as determined by ELISA. The resultant mixture was reseparated by protein A-Sepharose adsorption and elution in 0.1 M glycine buffer, pH 2.5, except for the K3C4 monoclonal NeF.

II6 I SOKOS El ,\I
Prcpuxtiou (?/‘~(LI~‘)~ C3Nrb’. Specific C3NeF. isolated as described above. was used as starting material. Pepsin digestion was carried out at low pH and the F(ab’), fragments of C3NeF were further purified by gel filtration on Sephacryl- 200 Superfine and the protein was measured by the Lowry method using purified IgG as the standard. Specificity of the preparation, i.e., the absence of undigested IgG or Fc fragments. was assured by the lack of reactivity with antibody to Fc fragments in an ELISA assay.
Assay .fi)r C3NeF ,fitnctiomd activity. The functional activity of C3NeF was measured by its effect on the rate of decay of preformed EC3bBb cells t IX). In this method, 0.5 ml of the sample was added to a button of I x 10’ cells at 30°C. At varying times from 0 to 30 min, aliquots of 0.1 ml were removed and added to tubes containing 0. I ml of 0.01 M EDTA and 0.3 ml of a I: 15 dilution of whole rat serum in 0.04 M EDTA. This mixture was incubated for 60 min at 37°C and the amount of oxyhemoglobin is then determined spectrophotometrically. Control cells are incubated with buffer. Residual hemolytic activity was then plotted as a function of time.
Isolution of’ unti-idiotypic antibody to C3NrF. Affinity columns t I .6 x 15-20 cm) with goat anti-human IgG (Column I), pooled human IgG (Column II), and F(ab’)? C3NeF (Column III) were made by coupling the respective proteins to CNBr-activated Sepharose 4B as recommended by the manufacturer. One milli- liter of serum was diluted fivefold with phosphate-buffered saline (PBS) and ap- plied to the anti-IgG column (Column I) at 4°C. After I5 min. a j-ml aliquot was collected and reapplied to the column. This procedure was again repeated fol- lowed by extensive washing with PBS and stripping with 0.1 M glycine-HCI, pH 2.5. The protein-containing fractions were pooled, immediately neutralized. dia- lyzed against PBS, and concentrated to 5 ml. This S-ml fraction was then placed on the human IgG affinity column (Column II), equilibrated as above, and washed through with PBS. Approximately 30 ml was collected, concentrated to 2.5 ml. and placed on the F(ab’), C3NeF affinity column (Column III). After extensive washing with PBS to remove nonadherent protein, the column was eluted with 0. I M glycine-HCI, pH 2.5. A 60-ml fraction was collected, neutralized. concen- trated, and dialyzed against PBS. Quantitation for total IgG and anti-idiotypic antibody to C3NeF (see below) was then done by ELISA. If the total IgG ex- ceeded the amount of anti-idiotypic antibody by more than 5%. the specimen was reapplied to Columns II and III. The final preparations failed to bind to nonspe- cific IgG and contained greater than 95% anti-idiotypic antibody. In addition. no C3NeF activity (inadvertently eluted from Column III) could be detected, as evidenced by lack of stabilization of C3bBb activity.
Inhibition oj’NeF,firnctionul clctillity. Samples containing the anti-idiotypic an- tibody were mixed for 30 min at 30°C with the NeF-containing samples and were then tested for the ability of the mixture to stabilize EC3bBb as described above. Percentage inhibition was calculated using the number of residual sites per cell at IO min.
Determination oj binding of‘anti-idiotypic antibodies to C3NeF preprrrations. Purified anti-idiotypic antibody (18.6 &ml) was incubated with several concen- trations of purified C3NeF. After 60 min at 30°C the mixtures were centrifuged at
HUMAN C3 NEPHRITIC FACTORS 117
high speed to remove any complexes and the supernatants were added to Immu- Ion plates coated with F(ab’), fragments of C3NeF. Residual anti-idiotypic anti- body binding to the F(ab’), fragments of NeF was quantitated using alkaline phosphatase-labeled goat antibody to human Fc fragment followed by proper development. The amount of the bound anti-idiotypic antibody was then calcu- lated as a percentage of the original amount present and plotted against the amount of C3NeF. The best fit line was plotted and the ability of the anti-idiotypic antibody to bind to C3NeF is expressed as the nanograms of C3NeF which bound 50% of the anti-idiotypic antibody.
RESULTS
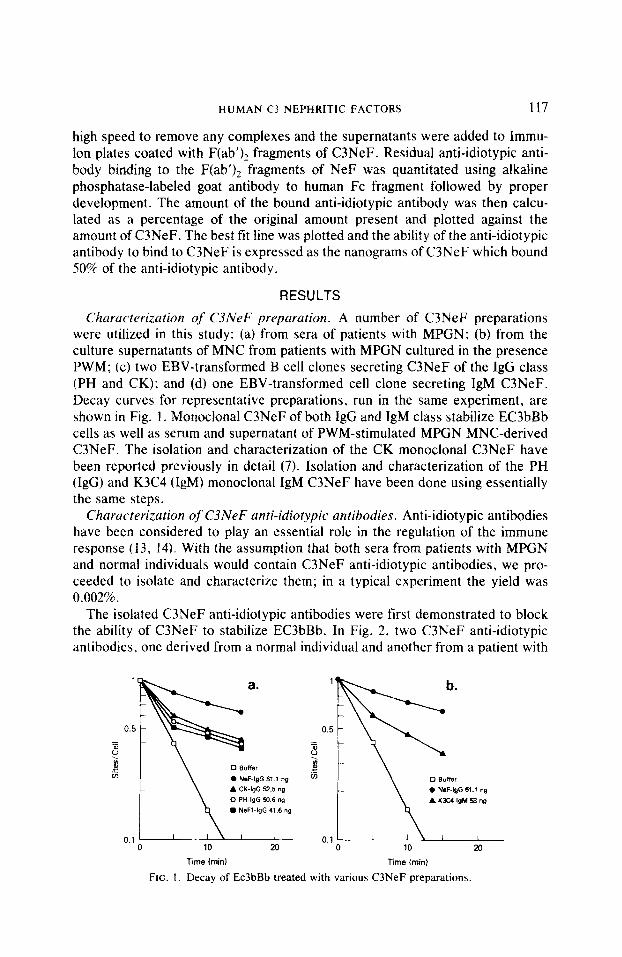
Churucterizution qf C3NeF preparation. A number of C3NeF preparations were utilized in this study: (a) from sera of patients with MPGN; (b) from the culture supernatants of MNC from patients with MPGN cultured in the presence PWM; (c) two EBV-transformed B cell clones secreting C3NeF of the IgG class (PH and CK); and (d) one EBV-transformed cell clone secreting IgM C3NeF. Decay curves for representative preparations, run in the same experiment, are shown in Fig. 1. Monoclonal C3NeF of both IgG and IgM class stabilize EC3bBb cells as well as serum and supernatant of PWM-stimulated MPGN MNC-derived C3NeF. The isolation and characterization of the CK monoclonal C3NeF have been reported previously in detail (7). Isolation and characterization of the PH (IgG) and K3C4 (IgM) monoclonal IgM C3NeF have been done using essentially the same steps.
Characterization of C3NeF unti-idiotypic antibodies. Anti-idiotypic antibodies have been considered to play an essential role in the regulation of the immune response (13, 141. With the assumption that both sera from patients with MPGN and normal individuals would contain C3NeF anti-idiotypic antibodies, we pro- ceeded to isolate and characterize them; in a typical experiment the yield was 0.002%.
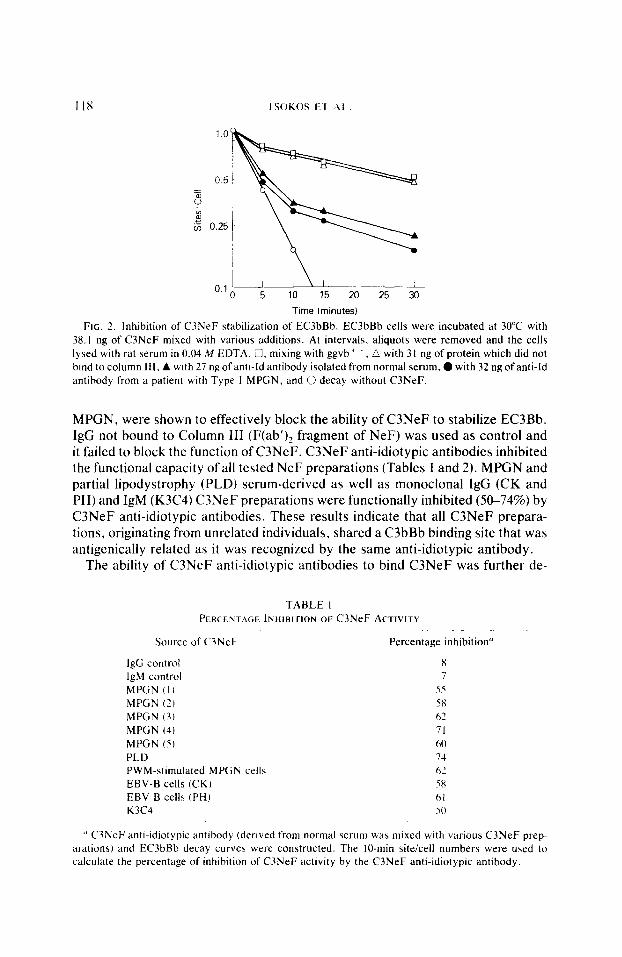
The isolated C3NeF anti-idiotypic antibodies were first demonstrated to block the ability of C3NeF to stabilize EC3bBb. In Fig. 2. two C3NeF anti-idiotypic antibodies, one derived from a normal individual and another from a patient with
a.
0
?
E 0 *utter i7 . N-F-,gG B,. I “g
. CK-lgG 52.5 ng . NSWJG 61.1 ng
0 PWgG cc.6 ng * K3CClgM 53 ng
. NeFl-IgG 41.6 ng
I ,
0 10 m 0 10 m Time (mid Time (mid
FIG. I. Decay of Ec3bBb treated with various C3NeF preparations.
0.1 1 0 5 10 15 20 25 30 Time (minutes)
FIG. 2. Inhibition of C3NeF stabilization of EC3bBb. EC3bBb cells were incubated at 30°C with 38.1 ng of C3NeF mixed with various additions. At intervals, aliquots were removed and the cells lysed with rat serum in 0.04 M EDTA. i?, mixing with ggvb ’ .. a with 3 I ng of protein which did not bind to column Ill. A with 27 ng of anti-Id antibody isolated from normal serum, 0 with 32 ng of anti-Id antibody from a patient with Type I MPGN, and 0 decay without C3NeF.
MPGN, were shown to effectively block the ability of C3NeF to stabilize EC3Bb. IgG not bound to Column III (F(ab’)? fragment of NeF) was used as control and it failed to block the function of C3NeF. C3NeF anti-idiotypic antibodies inhibited the functional capacity of all tested NeF preparations (Tables 1 and 2). MPGN and partial lipodystrophy (PLD) serum-derived as well as monoclonal IgG (CK and PH) and IgM (K3C4) C3NeF preparations were functionally inhibited (50-74%) by C3NeF anti-idiotypic antibodies. These results indicate that all C3NeF prepara- tions, originating from unrelated individuals, shared a C3bBb binding site that was antigenically related as it was recognized by the same anti-idiotypic antibody.
The ability of C3NeF anti-idiotypic antibodies to bind C3NeF was further de-
TABLE I PERIXNTACE INHIBITION OF C3NeF ACTIVITY
Source of C3NeF Percentage inhibition”
IgG control x IgM control 7 MPGN (1) 55 MPGN (2) 5x MPGN (3) 62 MPGN (4) 71 MPGN (5) 60 PLD 74 PWM-stimulated MPGN cells 62 EBV-B cells (CK) 5x EBV-B cells (PH) hl K3C4 50
” C3NeF anti-idiotypic antibody (derived from normal serum was mixed with various C3NeF prep- arations) and EC3bBb decay curves were constructed. The IO-min site/cell numbers were used to calculate the percentage of inhibition of C3NeF activity by the C3NeF anti-idiotypic antibody.
HUMAN C3 NEPHRITIC FACTORS 119
TABLE 2 INHIBITION OF C3NeF ACTIVITY
Anti-idiotypic antibody Percentage inhibition of C3NeF preparations
..__- ~~ to C3NeF 1 2 3
Normal 51 66 58 Normal 66 69 71 Normal 49 55 51 MPGN 52 58 61 MPGN 56 61 31 IgG control 8 5 2
Nare. Percentage of inhibition was calculated as in legend for Table 1.
4 5
61 62 78 72 51 59 60 62 66 62
2 4
termined by incubating standard amounts of them with variable concentrations of C3NeF preparations. The amount of each C3NeF preparation which bound 50% of the anti-idiotypic antibody was used as an indicator of the avidity of the idio- type anti-idiotypic interaction. As shown in Table 3, MPGN serum-derived, MPGN MNC PWM-stimulated culture supernatant-derived, and IgG and IgM monocional NeF preparations bound to a C3NeF anti-idiotypic antibody with comparable avidities.
Monoclonal C3NeF preparations fail to bind to common antigens. The avail- ability of both IgG and IgM NeF and the finding that they, along with serum- derived C3NeF preparations, share a common idiotype, the assumption that anti- idiotypic antibodies may resemble antigenically to environmental antigens, and the fact that many autoantibodies have been found to cross-react with environ- mental antigens (19) tested the ability of all three monoclonal C3NeF to bind to a battery of antigens (Fig. 3). Among the tested antigens were TNP, DNA, P-galactosidase, Fc portion of human IgG, C3b, C3d, factor H, lipopolysaccharide
TABLE 3 BINDING OF ANTI-IDIOTYPIC ANTIBODY TO C3NeF
~~~~- Origin of C3NeF ng of C3NeF”
Serum MPGN IgG MPGN IgG MPGN IgG
Culture K3C4 CK PH PWM-stimulated MPGN cells
Control IgG Control IgM
0.19 0.29 0.19
0.23 0.42 0.54 0.45 ,>20 ‘230
LI Increasing quantities of C3NeF were incubated with a standard (18 ng) of anti-idiotypic antibody to C3NeF. Free anti-idiotypic antibody was estimated by ELISA using plates coated with Ftab’), fragments of C3NeF. The numbers represent the amount of C3NeF which bound half of the used anti-idiotypic antibody.
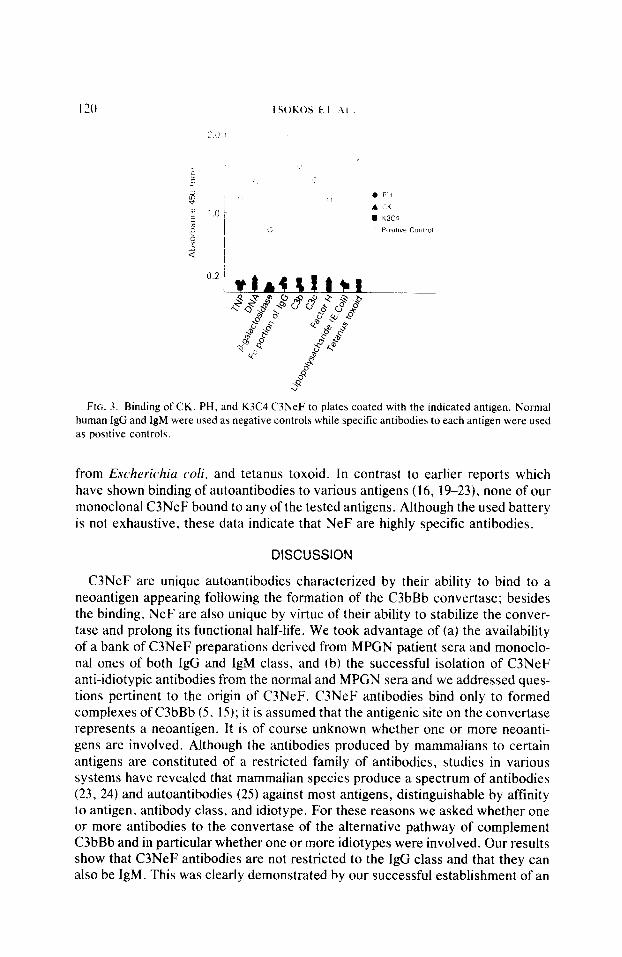
FIG. 3. Binding of CK, PH, and K3C4 C3NeF to plates coated with the indicated antigen. Normal human IgG and IgM were used as negative controls while specific antibodies to each antigen were used as positive controls.
from Escherichia cd, and tetanus toxoid. In contrast to earlier reports which have shown binding of autoantibodies to various antigens (16, 1%23), none of our monoclonal C3NeF bound to any of the tested antigens. Although the used battery is not exhaustive, these data indicate that NeF are highly specific antibodies.
DISCUSSION
C3NeF are unique autoantibodies characterized by their ability to bind to a neoantigen appearing following the formation of the C3bBb convertase; besides the binding, NeF are also unique by virtue of their ability to stabilize the conver- tase and prolong its functional half-life. We took advantage of (a) the availability of a bank of C3NeF preparations derived from MPGN patient sera and monoclo- nal ones of both IgG and IgM class, and Cb) the successful isolation of C3NeF anti-idiotypic antibodies from the normal and MPGN sera and we addressed ques- tions pertinent to the origin of C3NeF. C3NeF antibodies bind only to formed complexes of C3bBb (5, 15); it is assumed that the antigenic site on the convertase represents a neoantigen. It is of course unknown whether one or more neoanti- gens are involved. Although the antibodies produced by mammalians to certain antigens are constituted of a restricted family of antibodies, studies in various systems have revealed that mammalian species produce a spectrum of antibodies (23, 24) and autoantibodies (25) against most antigens, distinguishable by affinity to antigen, antibody class, and idiotype. For these reasons we asked whether one or more antibodies to the convertase of the alternative pathway of complement C3bBb and in particular whether one or more idiotypes were involved. Our results show that C3NeF antibodies are not restricted to the IgG class and that they can also be IgM. This was clearly demonstrated by our successful establishment of an

HUMAN C3 NEPHRITIC FACTORS 121
IgM C3NeF (K3C4) by transforming peripheral B cells with EBV from a patient with MPGN. Previously, murine monoclonal C3NeF antibodies of IgG lk and IgM classes have been produced by immunizing mice with factor B (26, 27). In reference to the number of idiotypes expressed by the various C3NeF, including both the IgG and IgM classes, our results show that all of them express only one idiotype binding to the C3bB convertase. This conclusion arose from the following observations: (a) each C3NeF anti-idiotypic antibody inhibited the binding and the functional stabilization of the C3bBb convertase induced by all C3NeF antibodies. and (b) all C3NeF antibodies bound avidly to C3NeF anti-Id antibodies. Although the affinity of each NeF for both the C3bBb convertase and the NeF anti-idiotypic antibodies was not formally calculated, we can assume that it is approximately the same for all C3NeF antibodies. We reached this conclusion because the percent- ages of the functional inhibition of the C3NeF antibodies induced by the C3NeF anti-idiotypic antibodies (Fig. 3) and the amount of anti-idiotypic antibody bound by the various C3NeF (Table 3) were similar.
The similarity in antibody reactivity among the various C3NeF preparations was not an unexpected observation. Various autoantibodies originating in mice (20, 28, 29) or in humans [to autoantigens (16, 28) or to foreign antigens (1911 express extensive similarities in terms of class. affinity, and idiotypy.
Anti-idiotypic antibodies are generated during the course of a normal immune response to extrinsic antigens (14). Antibodies to foreign antigens and autoanti- bodies may share idiotypes (19); shared idiotypy implies that anti-idiotypic anti- bodies could provide the means for expansion of the autoantibody response as envisioned by Plotz (30).
C3NeF could theoretically arise in response to a foreign antigen, although, as it is the case with most autoimmune responses. the inciting agent remains at large. Our effort to detect cross-reactivity between the monoclonal C3NeF antibodies and a limited battery of antigens was fruitless.
In this communication we have provided evidence that human C3NeF antibod- ies of either IgG or IgM class, derived from different individuals, share a common idiotype and have similar binding characteristics. These findings suggest but do not prove that germline genes may be used in the formation of C3NeF antibodies. Finally, C3NeF autoantibodies do not bind to a battery of antigens to which other autoantibodies have been reported to cross-react (3 I ).
REFERENCES I. Spitzer. R. E.. Vallota. E. H., Forristal. J.. Sudora. E.. Stitzel. A., Davis. N. C., and West.
C. D.. Science 164, 436, 1969. 2. Sissons. J. G. P.. West, R. J., Fallows. J.. Williams, D. Ci.. Boucher, B. J., Amos. N.. and
Peters. D. K.. N. Engl. J. Med. 294, 461, 1976. 3. Vallota. E. J.. Forristal. J.. Spitzer. R. E.. Davis, N. C.. and West, C. D.. J. Exp. Med. 131, 1306.
1970. 4. Schreiber, R. D.. Gotze, 0.. and Muller-Eberhard. H. J., Sc,und. J. fmmunol. 5, 705. 1976. 5. Daha, M. R. ~ Austen, K. R., and Fearon. D. T.. J. Immunol. 119, 812, 1977. 6. Spitzer, R. E.. and Stitzel. A.. Manuscript submitted. 7. Hirdmatsu. M.. Balow. J. E., and Tsokos, G. C.. J. Immrcnol. 136, 4451. 1986. 8. Abdou. N. I.. Wall. H.. Lindsley. H. B.. Halsey, J. F.. and Suzuki. T.. J. C/in. Invrs/. 67, 1297.
1981.

I’? k... I-SOKOS El Al
Y. Zouali. M.. and Eyquem. A.. .4,1r,. //l.cr. Ptrsrrrrr /n~~nrtn~~/. 1346. 377. IY83.
IO. Abe. T.. Takeuchi. ‘I... Kyotaki. M.. Koide. J.. Ho\ono. 0.. Homma. M.. Otake. I and Kane.
S., J. Imtn/rno/. 132. 2381. 1984.
I 1. Cunningham-RundIe\. C., J. /Lip. Mctl. 155, 71 I, 1%:.
I?. Dwyer. D.. Vakil. M.. and Kearney. J. F.. J. E.r/>. Mrtl. 164, 1340. 19%
I?. Zanetti. M.. <‘Kc‘ (‘rir. /?(,I.. /~,?,nro~o/. 6, 151. 19x6.
14. Bona, C. A.. Heber Kat/. E.. and Paul. W. E.. J. E.rp. Mtd 153, Y.Sl. IYXI.
1. Daha. M. R.. Fearon, D. ‘I‘., and Austen. K. F.. J. fn~mrrrrrd. 116. 1. lY76.
16. Mackworth-Young. C.. Sabbaga. J.. and Schwartz. R. S.. J. C/in. Int~esr. 79, 572. 1987.
17. Golding, B.. Inphirami. G.. Peter\, E.. Hoffman T.. Balow. J. E.. and Tsokos. G. C.. J. Irnmzcno/. 139, 4061. 1987.
IX. Medicus. R. G.. Gotre. 0.. and Miiller-Eberhard. H. J.. J. Erp. Med. 144. 1076. 1976.
19. Monestier, M., Bonin. B.. Migliorini. I’.. Dang, H.. Datta, S., Kuppers. R.. Rose. N., Maurer. P..
Talal. N.. and Bona, C.. J. Exp. Med. 166, 1109, 1987.
20. Migliorini. P.. Ardman. B.. Kahuraki. J.. and Schwartz. R.. J. Exp. Med. 164. 483, 1987.
71. Lymperi. P.. Dighiero. G.. Ternynck. T.. and Arrameas. S.. Eftu. J. fmmunol. 15, 702. 1985.
22. Kofler. R.. Noonan. 0. J.. Lev). D. E.. Wilson. M. C.. Moller. N. P. H.. Dixon, F. J., and
Theofilopoulo\. A. N.. J. Exp. ,Mcd. 161, 805. 19X5.
‘3. Perlmuter. R. M.. Cl-ewh, S. T.. Douglas. R.. Sorensen, G.. Johnson. N.. Nivera. N.. Gearhart.
P., and Hood. L.. ild~.. /nrf?~~rroi. 35. I, 19X5.
24. Berek. C.. Griffitha. G. M.. and Milstein. C.. Narrtrc, U.ondon) 316. 412. 1985.
35. Kotler. R.. Noonan. I). J.. Strohal. R.. Balderas. R. S.. Moller, N. P. H.. Dixon. F. J.. and
Theotilopoulos. A. N.. EIU. J. Itnmrrnd. 17, Yl. 1987.
26. Daha. M. R.. Deedler. A. M.. and Van Es. L. A.. ./. Immrtnol. 132, 2538. 1984.
77. Veda. A., Kearney. J. F., Rout. K., and Vollanakis. J. E.. J. Immrnol. 138, 1143. 1987.
78. Bergen III. H. R.. Losman. M. J.. O’Connor. T.. Zacharias. W.. Larson. J. E.. Accavitti. M. A.,
Wells. R. I>.. and Koopman. M’. J.. J. Immrcnd. 139. 743. 1987.
29. Rauch, J.. Murphy. I-.. Roth\. J. B.. Stellar. B. D.. and Schwartz. R. S.. ./. Imnrrmd. 129, 136.
lY81.
30. Plotz. P. H.. L<inc,c,/ 2. X24, 19x3.
?I. Shoenfeld. Y.. Rauch. J.. Masicotte. H.. Dalton, S. K.. Andre-Schwart/. J.. Stellar. B. 0.. and
Schwartz. R. S.. !L’. Eqc(. J. Mvd. 308. 414. lY83.
Received March 15. IYXY: accented with revision June 6. IYXY





























