Embed Size (px)
Citation preview

Tbx2 Represses Expression of Connexin43 in Osteoblastic-like Cells
J.-R. Chen,1,2,3 B. Chatterjee,4 R. Meyer,4 J. C. Yu,5 J. L. Borke,6 C. M. Isales,1,7,8 M. L. Kirby,1,2
C. W. Lo,4 R. J. Bollag1,2
1Department of Cell Biology and Anatomy, Medical College of Georgia, Augusta, GA 30912, USA2Developmental Biology Program, Institute of Molecular Medicine and Genetics, Medical College of Georgia, Augusta, GA 30912, USA3Department of Biological Science and Technology, I-Shou University, Kaohsiung, Taiwan R.O.C.4Laboratory of Developmental Biology, NHLBI/NIH, Bethesda, MD 20892, USA5Department of Surgery, Section of Plastic Surgery, Medical College of Georgia, Augusta, GA 30912, USA6Department of Oral Biology and Maxillofacial Pathology, Medical College of Georgia, School of Dentistry, Augusta, GA 30912, USA7Program in Synapses and Cell Signaling, Institute of Molecular Medicine and Genetics, Medical College of Georgia, Augusta, GA 30912, USA8Augusta VA Medical Center, Augusta, GA 30910, USA
Received: 8 May 2003 / Accepted: 14 October 2003 / Online publication: 17 February 2004
Abstract. Tbx2 belongs to a family of developmentaltranscription regulatory factors. We evaluated whetherthe gap junction protein Connexin43 (Cx43), an im-portant regulator of osteoblast function and bone de-velopment, may be a downstream target gene regulatedby Tbx2. The Cx43 promoter contains direct repeats ofthe consensus T-box binding motif, TCACAC, andmoreover, Tbx2 and Cx43 show overlapping expressiondomains in precursors to bone and in osteoblasts. Invitro analysis showed that the Cx43 promoter containstwo Tbx2 binding sites, and this binding was dependenton the TCACAC consensus sequence. Transient trans-fection analysis with a Cx43 promoter-driven lacZ re-porter construct revealed negative regulation mediatedby these two Tbx2 binding sites in osteoblast-like cells.Thus, downregulation of Tbx2 led to de-repression ofwild-type Cx43 promoter activity, whereas a promoterconstruct with mutated binding sites showed no de-re-pression. In stably transfected osteosarcoma cells inwhich expression of the endogenous Tbx2 gene wasdownregulated with a Tbx2 antisense construct, amarked de-repression of the endogenous Cx43 gene wasobserved. This was accompanied by a marked increasein the abundance of Cx43 gap junctions and increasedfunctional gap junction-mediated cell–cell communica-tion. Analysis of lacZ expression in transgenic micecontaining the mutated Cx43 promoter-driven lacZconstruct further suggested de-repression of the Cx43promoter in limb buds, a region destined to give rise tolong bones of the limbs. Taken together, these findingsindicate that the promoter of Cx43 is repressible byTbx2, both in cultured osteoblast-like cells in vitro andlikely in the developing embryo.
Key words: Bone — Development — Gap junctions —T-box
The mouse T-box gene family was identified on the basisof a conserved DNA binding domain, the T-box motif[1]. The T-box gene family has attracted considerableattention because of the ability of individual members toaffect a wide variety of developmental processes. T-boxgenes have been found to influence identity of axialmesoderm (T, spadetail) [2–4], development of pharyn-geal arch arteries (Tbx1) [5–7], determination of paraxialmesoderm (Tbx6) [8, 9], heart and limb specification(Tbx5) [10–13], development of mammary glands andlimbs (Tbx3) [14–16], limb position and identity (Tbx4and Tbx5) [17–20], trophoblast development (eomeso-dermin) [21], identity of pituicytes (Tpit) [22], palato-genesis (TBX22) [23], and differentiation of thymocytes(T-bet) [24]. Indeed, the importance of the T-box genefamily in controlling development is manifested by thediscoveries that haploinsufficiencies for certain T-boxgenes play roles in three human developmental syn-dromes: ulnar-mammary syndrome, Holt-Oram syn-drome, and DiGeorge syndrome [5–7, 10, 12, 14, 15].One common theme in these disorders is a pathologiccondition of bone, including limb deformities in theformer two and craniofacial deformities in the latter.
Based on sequences generated by the Caenorhabditiselegans genome-sequencing project, there are at least fiveT-box genes in nematodes. To date, the only gene incommon between mice and nematodes is Tbx2 [25].Orthologs (direct descendants) of a primordial Tbx2gene have survived in the mouse genes, Tbx2 and Tbx3,as well as in the Drosophila gene, omb. This suggests thata Tbx2 ortholog will be common to all species with thiscommon ancestor. The mouse and human Tbx2 genesshare 100% predicted amino acid identity in the T-box
Present address of M.L.K: Neonatal/Perinatal Research Insti-tute, Department of Pediatrics/Neonatology, Duke UniversityMedical Center, Durham, NC 27710, USA
Present address of R.M: Department of Biomedical Sciences,Creighton University Medical School, Omaha, NE 68178,USA
Correspondence to: R. J. Bollag; E-mail: [email protected]
Calcif Tissue Int (2004) 74:561–573
DOI: 10.1007/s00223-003-0106-5

region [26]. This conservation implies a vital role forTbx2 and underscores its importance.
Although the phenotypic ramifications of geneticmodulation of these genes implies a complex function,limited information is available concerning the targetgenes directly controlled by T-box proteins. Studieswith mouse Brachyury (T), the archetype of the T-boxgene family, and Tbx2 showed that these T-box proteinsare localized in the nucleus, and are able to modulatethe activity of a reporter gene by binding to a responseelement, TCACAC [16, 27–34]. Thus far few studieshave examined downstream target genes regulated byT-box genes. One approach to identifying potentialcandidate genes that may be regulated by Tbx2 is tostudy genes that have similar spatial-temporal embry-onic expression patterns and that also have the puta-tive consensus, palindromic T-box binding site or halfsite in the promoter region. On the basis of commonexpression in osteogenic progenitors and osteoblasts,Cx43 is one such candidate gene, and is the focus of thepresent study examining transcriptional regulation byTbx2.
Cx43, encoded by the gene Gja1, is a 43-kDa conn-exin protein that constitutes gap junctions found inneural crest cells, osteoblasts, myocytes, and many othercells and tissues. Cx43 plays an essential role in medi-ating cell–cell coupling in osteoblasts and in develop-ment by modulating the migratory behavior of neuralcrest cells [35, 36]. It has been shown that the regulatedtemporal pattern of Cx43 expression plays a critical rolein normal ossification and osteoblast function [37], andthat gap junctional communication modulates differen-tiation and proliferation in osteoblasts [38]. As deter-mined by nucleotide analysis, the promoter of Cx43 [39]contains direct repeats of T half sites (Fig. 1). In addi-tion, the expression levels of Cx43.4, an ortholog of themammalian connexin45 gene, and mouse connexin40are depressed in the zebrafish T locus mutant, no tail[40], and in mouse Tbx5 mutants [13], respectively.Moreover, previously published studies suggest thatCx43 and Tbx2 are both expressed abundantly in thepharyngeal arches [41, 42]. Together these findingssuggest that connexin genes could be subject to regula-tion by T-box genes.
In light of these observations, we hypothesized thatTbx2 may regulate Cx43 expression. In accordance withthis hypothesis, we have recently shown that there is areciprocal staining pattern for Tbx2 and Cx43 indeveloping rat coronal sutures and that Tbx2 can neg-atively regulate Cx43 promoter activity in Tbx2-modu-lated osteoblasts [43], implying an antitheticalrelationship between the two proteins. In the presentwork, we seek to establish direct physical binding of theTbx2 binding domain to Cx43 promoter elements, andto demonstrate repression of Cx43 promoter and func-tion in cell lines as well as in transgenic mice. Our results
strongly suggest that Cx43 is a direct transcriptionaltarget of Tbx2 in osteoblasts.
Materials and Methods
Bacterial Expression and Purification of Tbx2 Protein
Bacterial expression of full length Tbx2 with a decahistidinetag was accomplished by cloning the entire mouse Tbx2 codingsequence [1] into pET19b after incorporation of an NdeI site atthe second ATG, followed by transformation into BL21 (DE3)pLysS. Optimal expression was achieved by adding 1 mMisopropyl-thio-ß-D-galactopyranoside (IPTG) to the bacterialculture at 30�C for 3 hours. The protein was solubilized in 6 Murea and purified by passing over a nickel resin column, withsubsequent elution by 80 to 100 mM imidazole. Urea was thenremoved via dialysis through successive changes of buffer to afinal buffer concentration of 100 mM HEPES, 150 mM NaCl.Truncated proteins containing the N-terminus (AA 1–550), orC-terminus (AA 548–701), were generated via pET19b usingan internal NdeI site to generate upstream or downstreamdeletions. Bacterial expression of the conserved DNA bindingbox region of Tbx2 protein, box2, (AA 86–286) was accom-plished by polymerase chain reaction (PCR) amplification theT-box region of Tbx2 and cloning into pET19b followed bytransformation into BL21 (DE3) pLysS. The expression wasinduced by 0.3 mM IPTG at 30�C overnight. The purificationwas similar to that for the full length Tbx2 protein, except thatbox2 protein remained in the soluble fraction so that urea wasomitted from the solubilization buffer.
Southwestern Blot Analysis
Protein samples were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and trans-ferred to a nitrocellulose membrane in 25 mM Tris-HCl,200 mM glycine, and 25% methanol at 4�C for 1.5 hours at45 V. The membrane was incubated in renaturation buffer (5%dry milk; 10 mM Tris-HCl, pH 7.5, 150 mM NaCl; 10% glyc-erol; 2.5% NP)40; 1 mM DTT; and 0.1 mM ZnCl) overnight.The membrane was probed with 106 cpm 32P end-labeled frag-ments corresponding to double-stranded oligonucleotides BS-T[27–29] (5¢–GGGAATTTCACACCTAGGTGTGAAATTCCC–3¢) or BS-neg (5¢–GGGAATTTGT GTGGATCCACACAAATTCCC-3¢) in hybridization solution (0.25% dry milk;10 mM Tris-HCl, pH 7.5, 50 mM NaCl; and 1 mM DTT) atroom temperature for 2 hours. After washing in 10 mM Tris-HCl, pH 7.5, 150 mM NaCl, and 1 mM DTT, the blot wasexposed to film for autoradiography.
Electrophoretic Mobility Shift Analysis
Electrophoretic mobility shift assays were carried out usingpurified Tbx2 DNA-binding domain (box2) protein, as de-scribed previously. Full-length Tbx2 protein was insolubleunder standard growth conditions in bacteria, and this neces-sitated the use of the DNA binding domain exclusively. Thebox2 region is a protein of approximately 30 kDa. The fol-lowing synthesized oligonucleotides were used as probes orcompetitors in subsequent electrophoretic mobility shift as-says. WtCx43: 5¢GATCACTCACACCCA CCTCAAGCTT-CACACACCTTGG3¢ (37 bp) contains two Tbx2 bindingsites; whereas Cx43L (19 bp): 5¢GATCACTCACACCCACCTC3¢ or Cx43R (19 bp): 5¢AAGCTTCACACACCTTGG3¢contains only individual binding sites. Mut3: 5¢-GAT-CACTCAACACCACCTCAAGCTTCAACAACCTTGG-3¢(37 bp) oligonucleotide contains CAC to ACA mutations atboth binding sites. The complementary oligonucleotides wereannealed prior to subsequent experiments.
Probes (10,000 cpm) were incubated with purified box2protein in the presence of nonspecific poly (dI-dC) competitor
562 J.-R. Chen et al.: Tbx2 Represses Cx43 Expression

and unlabeled WtCx43 or mut3 oligonucleotides as competi-tors, at concentrations ranging from 0.01 to 1 lM, in a totalvolume of 10 lL binding buffer containing 10% glycerol,25 mM HEPES (pH 8.7), 84 mM KCl, and 5 mM DTT for15 minutes at 30�C. Reactions were separated by electropho-resis through 4% acrylamide (29:1) in TBE buffer at 270 V for60 minutes, Gels were dried and visualized by exposure eitherto PhosphorImager screens or to film.
Mammalian Expression Constructs
Full-length mouse Tbx2 cDNA in the sense or antisenseorientation was cloned into pcDNA3 (Invitrogen), with theCMV promoter driving high-level gene expression. Tbx2cDNA was first cut with BamHI and BglII and cloned intothe pcDNA3 BamHI site. The plasmid pHXPL is a reporter
gene construct containing 6.7 kb of the upstream sequencefrom the mouse Cx43 promoter driving lacZ expression [39].The plasmid pHXPLmut3 is the same as pHXPL describedpreviously except that the two binding sites were changed fromTCACAC to TCAACA. A PCR fragment containing themutations was first cut with XbaI and PacI, and then ligatedinto pHXPL that had been cut by PacI and partially digestedwith XbaI to delete 700 bp. Mutations were verified by directsequencing.
Cell Culture and Transfection
NIH3T3 cells were purchased from the ATCC (Rockville,MD, USA) and maintained in DMEM supplemented with10% fetal bovine serum, 4 mM glutamine and penicillin/streptomycin at 37�C in a 5% CO2 humidified incubator.
Fig. 1. Cx43 promoter sequence. Panel A: Alignment ofproximal promoter regions of the Cx43 gene from mouse,human, and rat [53–55]. Transcriptional regulatory sequencesidentified previously are indicated [53–55, 60, 61], as are theputative T-box binding sites characterized in this work. The37-mer oligonucleotide encompassing the TCACAC sequences(Tbx2 binding sites) used in electrophoretic mobility shift as-says is boxed, and the pair of CAC nucleotides that arechanged to ACA in the mut3 oligonucleotide and also in the
pHXPLmut3 constructs are marked with diamonds. Theproposed transcription start site is indicated. Panel B: Depic-tion of oligonucleotides used in the mobility shift analyses ofFigure 3. WtCx43 consists of the murine promoter regionbetween )466 and )430 of panel A, whereas Cx43mut3 is theidentical probe with three nucleotides in each TCACAC half-site mutated as indicated. Cx43L ()466 to )449) and Cx43R()448 to)430), used in panel B of Figure 3, comprise the leftand right half-sites as indicated.
J.-R. Chen et al.: Tbx2 Represses Cx43 Expression 563

ROS17/2.8 cells were maintained in F12/DMEM supple-mented with 10% fetal bovine serum, 4 mM glutamine andpenicillin/streptomycin at 37�C in a 5% CO2 humidified incu-bator as described previously [44].
Reporter constructs were introduced into cells by liposome-mediated gene transfer (Gibco BRL). Cells were plated onto a35-mm 6-well plate at a seeding density of 2.5 · 105 cells perwell. A Lipofectamine/DNA ratio of 4 lL/lg was experimen-tally determined to be optimal for transfecting pcDNA3 con-structs into NIH3T3. A Lipofectin/DNA ratio of 6lL/lg wasexperimentally determined to be optimal for transfect-ing pcDNA3 constructs into ROS17/2.8. Forty-eight hoursafter transfection, cells were analyzed for beta-galactosidaseand luciferase activity for normalization of transfection effi-ciency. Reporter gene activities were measured using theDual-Light� luminescent reporter gene assay kit from Tropix(Bedford, MA, USA) incorporating the luminescent sub-strates luciferin and Galacton-Plus� for the detection of lucif-erase and ß-galactosidase respectively [45]. Briefly, cell lysateswere mixed with buffer A and light signal from the luciferaseenzyme present in the extract was measured immediately afterthe addition of buffer B containing luciferin and Galacton-Plus�. After a 60-minute incubation, an accelerator with al-kaline Sapphire-II� macromolecular enhancer was added andß-galactosidase light signal activity was recorded.
NIH 3T3 cells were transfected stably with expressionconstruct pcDNA-3 or with pcDNA-3-Tbx2sense (Table 1).Transfectants were selected in 400 lg/mL G418, and colo-nies were subcloned and selected based on positive immu-nohistochemical staining for Tbx2 protein (in the case ofpcDNA-3-Tbx2sense transfectants). Among selected trans-fectants, two representative subcloned cell lines were chosenfor subsequent analysis based on immunohistochemistry:K2 contained the pcDNA-3-Tbx2sense construct, whereasP1 contained pcDNA-3 alone (described in [30]). For ROS17/2.8 cells, pcDNA-3 vectors with sense or antisense orien-tations of the Tbx2 coding sequence were used for transfec-tion. Positive transfectants were selected in 600 lg/mLG418, and cell lines were screened for Tbx2 immunoreactivity.Tbx2 antisense transfectants were chosen by their minimalTbx2 immunoreactivity, whereas Tbx2 sense transfectantswere selected for enhanced immunoreactivity. Cell linesAS8 (antisense) and S14 (sense) were chosen for subsequentanalysis.
Immunohistochemistry and Western Blot Analysis
Antibodies were raised in rabbits against bacterially purifiedmouse Tbx2 protein by conventional protocols [30, 46]. Toeliminate cross-reactivity with other TBX proteins, the poly-clonal serum was subtracted by using chelated histidine-taggedmouse-TBX3 protein (epitope-tagged in pET19b incorporatinga murine sequence comparable to amino acids 99–295 in ac-cession number NP_005987), since among T-boxes this proteinhas the greatest sequence homology with Tbx2.
For immunofluorescence staining of tissue culture cells,cells were grown on coverslips, washed with phosphate buff-ered saline (PBS), fixed in 4% formaldehyde, and permeabi-
lized by incubation with cold methanol ()20�C). Cells werefirst stained with rabbit polyclonal antibodies raised againstfull length Tbx2 or rabbit polyclonal antibody to Cx43 (18-A-8; kindly provided by Dr. E. Hertzberg). Secondary antibodiesused were either FITC- or Cy3-conjugated goat-anti-rabbitIgG (Jackson ImmunoResearch Laboratories, Inc., WestGrove, PA, USA). Cells were examined and photographedunder epifluorescent illumination on a Nikon BX-40 or LeicaDMRE upright microscope.
For Western blotting, confluent cells (500,000/pair of lanes)were scraped into ice-cold PBS (pH 7.4) and disrupted by so-nication for 60 seconds in ice-cold homogenization buffer[60 mM Tris buffer, pH 7.4, 0 25 M sucrose, 10 mM EGTA,2 mM EDTA, 10 mM b-mercaptoethanol and protease in-hibitor cocktail (Sigma, St. Louis, MO, USA)]. Lysates withequalized protein sample were placed in sample buffer (0.5 MTris, pH 6.8, 4% SDS, 20% glycerol, and 0.1% bromophenolblue) and boiled. The denatured protein was separated bySDS-PAGE, transferred to NitroBind nitrocellulose mem-branes (Micron Separations Inc, Westborough, MA, USA)and incubated with rabbit anti-Tbx2 antibody at a 1:250 di-lution. Immunoreactive bands were visualized with a horse-radish peroxidase–conjugated secondary goat anti-rabbitserum and developed with enhanced chemiluminescence (ECL)(Pierce, Rockford, IL, USA).
Dye Coupling Studies
To evaluate gap junctional communication, cells were plated at200,000 cells per 35-mm dish. Two days after plating, gapjunctional communication was analyzed by using dye injec-tion. Microelectrode impalements were performed in L15medium containing 10% serum on a heated stage. Single cellswere impaled with glass microelectrodes containing 5% 6-carboxyfluorescein. Dye was iontophoretically ejected into theimpaled cell using 2 nA hyperpolarizing current pulses of 0.5-second duration at a frequency of once per second. Impale-ments were held for either 1 or 3 minutes. After the designatedtime, the microelectrode was removed and the preparation wasviewed under darkfield epifluorescent illumination. The num-ber of dye-filled cells (excluding the impaled cell) in the pri-mary, secondary, and tertiary order of cell contacts werecounted immediately after the impalement was terminated.
Assaying Promoter Activity in Transgenic Embryos
Transgenic mice containing the pHXPLmut3 construct weregenerated by the University of Pennsylvania TransgenicMouse Facility. This entailed pronuclear injection of mouseeggs with the purified promoter-driven lacZ construct, fol-lowed by transfer of the injected eggs into foster mothers. Theembryos that developed were harvested at E10.5, E11.5, andE13.5, and lightly fixed in 2% paraformaldehyde, followed bystaining with X-gal to detect the expression of the lacZ re-porter construct. After staining overnight, the embryos werepostfixed in 10% formalin and examined and photographedusing a light stereomicroscope.
Table 1. Plasmid list
Plasmid Description
pcDNA-3-Tbx2sense Full-length murine Tbx2 driven by CMV promoter in sense orientationpcDNA-3-Tbx2antisense Full-length murine Tbx2 driven by CMV promoter in antisense orientationpET19b-FLTbx2 Full-length Tbx2 cDNA in frame with His-Tag of pET-19b (Novagen)pET19b-NtermTbx2 cDNA constituting Tbx2 amino acids 1–550 in frame with His-TagpET19b-CtermTbx2 cDNA constituting Tbx2 amino acids 548–701 in frame with His-TagpET19b-box2 cDNA constituting Tbx2 amino acids 86–286 in frame with His-TagpHXPL 6.7-kb promoter of murine Gjo1 driving lacZ in pPD46.21pHXPLmut3 pHXPL with six nucleotides modified within T-box binding half-site
564 J.-R. Chen et al.: Tbx2 Represses Cx43 Expression
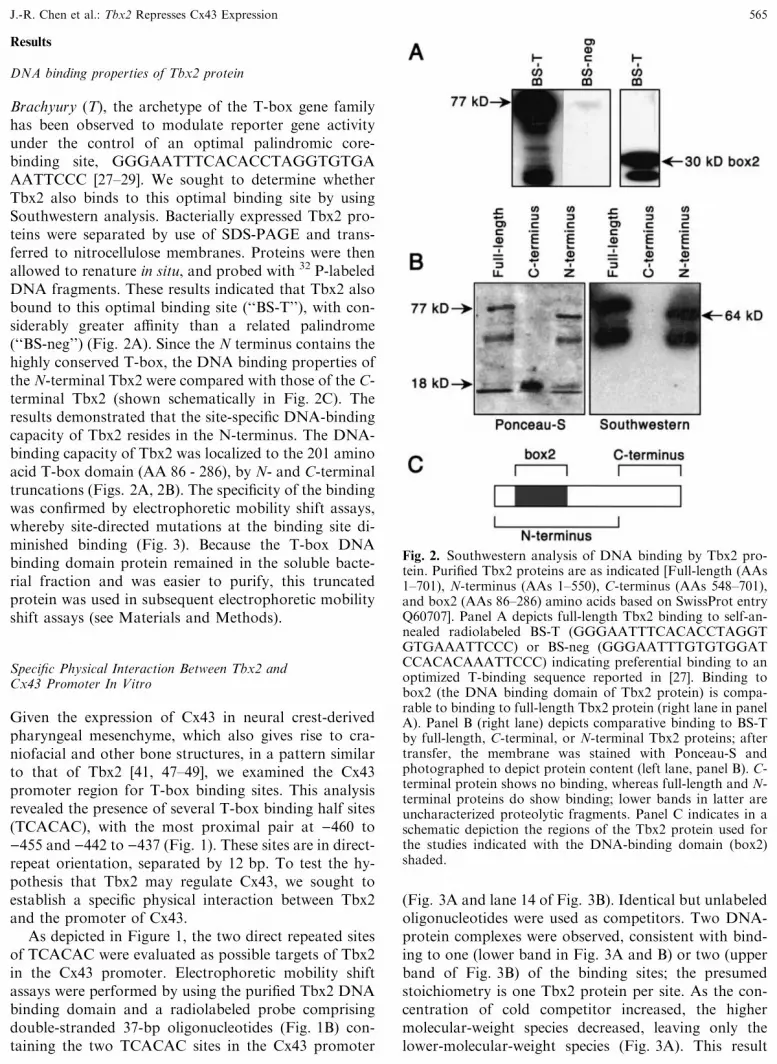
Results
DNA binding properties of Tbx2 protein
Brachyury (T), the archetype of the T-box gene familyhas been observed to modulate reporter gene activityunder the control of an optimal palindromic core-binding site, GGGAATTTCACACCTAGGTGTGAAATTCCC [27–29]. We sought to determine whetherTbx2 also binds to this optimal binding site by usingSouthwestern analysis. Bacterially expressed Tbx2 pro-teins were separated by use of SDS-PAGE and trans-ferred to nitrocellulose membranes. Proteins were thenallowed to renature in situ, and probed with 32 P-labeledDNA fragments. These results indicated that Tbx2 alsobound to this optimal binding site (‘‘BS-T’’), with con-siderably greater affinity than a related palindrome(‘‘BS-neg’’) (Fig. 2A). Since the N terminus contains thehighly conserved T-box, the DNA binding properties ofthe N-terminal Tbx2 were compared with those of the C-terminal Tbx2 (shown schematically in Fig. 2C). Theresults demonstrated that the site-specific DNA-bindingcapacity of Tbx2 resides in the N-terminus. The DNA-binding capacity of Tbx2 was localized to the 201 aminoacid T-box domain (AA 86 - 286), by N- and C-terminaltruncations (Figs. 2A, 2B). The specificity of the bindingwas confirmed by electrophoretic mobility shift assays,whereby site-directed mutations at the binding site di-minished binding (Fig. 3). Because the T-box DNAbinding domain protein remained in the soluble bacte-rial fraction and was easier to purify, this truncatedprotein was used in subsequent electrophoretic mobilityshift assays (see Materials and Methods).
Specific Physical Interaction Between Tbx2 andCx43 Promoter In Vitro
Given the expression of Cx43 in neural crest-derivedpharyngeal mesenchyme, which also gives rise to cra-niofacial and other bone structures, in a pattern similarto that of Tbx2 [41, 47–49], we examined the Cx43promoter region for T-box binding sites. This analysisrevealed the presence of several T-box binding half sites(TCACAC), with the most proximal pair at )460 to)455 and )442 to )437 (Fig. 1). These sites are in direct-repeat orientation, separated by 12 bp. To test the hy-pothesis that Tbx2 may regulate Cx43, we sought toestablish a specific physical interaction between Tbx2and the promoter of Cx43.
As depicted in Figure 1, the two direct repeated sitesof TCACAC were evaluated as possible targets of Tbx2in the Cx43 promoter. Electrophoretic mobility shiftassays were performed by using the purified Tbx2 DNAbinding domain and a radiolabeled probe comprisingdouble-stranded 37-bp oligonucleotides (Fig. 1B) con-taining the two TCACAC sites in the Cx43 promoter
(Fig. 3A and lane 14 of Fig. 3B). Identical but unlabeledoligonucleotides were used as competitors. Two DNA-protein complexes were observed, consistent with bind-ing to one (lower band in Fig. 3A and B) or two (upperband of Fig. 3B) of the binding sites; the presumedstoichiometry is one Tbx2 protein per site. As the con-centration of cold competitor increased, the highermolecular-weight species decreased, leaving only thelower-molecular-weight species (Fig. 3A). This result
Fig. 2. Southwestern analysis of DNA binding by Tbx2 pro-tein. Purified Tbx2 proteins are as indicated [Full-length (AAs1–701), N-terminus (AAs 1–550), C-terminus (AAs 548–701),and box2 (AAs 86–286) amino acids based on SwissProt entryQ60707]. Panel A depicts full-length Tbx2 binding to self-an-nealed radiolabeled BS-T (GGGAATTTCACACCTAGGTGTGAAATTCCC) or BS-neg (GGGAATTTGTGTGGATCCACACAAATTCCC) indicating preferential binding to anoptimized T-binding sequence reported in [27]. Binding tobox2 (the DNA binding domain of Tbx2 protein) is compa-rable to binding to full-length Tbx2 protein (right lane in panelA). Panel B (right lane) depicts comparative binding to BS-Tby full-length, C-terminal, or N-terminal Tbx2 proteins; aftertransfer, the membrane was stained with Ponceau-S andphotographed to depict protein content (left lane, panel B). C-terminal protein shows no binding, whereas full-length and N-terminal proteins do show binding; lower bands in latter areuncharacterized proteolytic fragments. Panel C indicates in aschematic depiction the regions of the Tbx2 protein used forthe studies indicated with the DNA-binding domain (box2)shaded.
J.-R. Chen et al.: Tbx2 Represses Cx43 Expression 565

suggests that Tbx2 binds to both TCACAC sites in theCx43 promoter (as shown in the schematic to Fig. 3A).To evaluate whether Tbx2 protein is indeed able to bindto both TCACAC sites, increasing amounts of Tbx2binding domain were co-incubated with either a radio-labeled 19-bp double-stranded oligonucleotide encom-passing the 5¢ binding site or a comparable radiolabeled19-bp double-stranded oligonucleotide encompassingthe 3¢ binding site. The relative levels of bound oligo-nucleotide imply that the 3¢ binding site has higherbinding specificity for box2, suggesting that flankingsequences may influence the binding affinity (Fig. 3,lanes 1–13).
Tbx2 Negatively Regulates Cx43 Promoter-lacZ Constructs
To determine if Tbx2 can regulate Cx43 expression invivo, we transfected into cultured cells a pHXPL-lacZreporter construct driven by 6.7 kb of the upstream se-quence from the mouse Cx43 promoter [39]. This con-
struct was cotransfected with a Tbx2 expression vectorinto NIH3T3 cells, a cell line that expresses Cx43 butnot Tbx2. As shown in Figure 4A, Tbx2 expressionleads to a decrease in lacZ expression, thereby indicatingthat Tbx2 represses Cx43 promoter activity in NIH3T3cells.
We repeated similar studies in ROS 17/2.8 cells, anosteoblastic cell line that constitutively expresses boththe endogenous Cx43 and Tbx2 genes [30, 43]. For theseexperiments, ROS cells were transfected with eitherpHXPL or the construct pHXPLmut3 in which the twoT-box binding sites in the Cx43 promoter were mutatedfrom TCACAC to TCAACA (Mut3, Fig. 1B) pHXPLshowed a twofold decrease in reporter gene activity, ascompared to pHXPLmut3, suggesting that endogenousTbx2 represses the Cx43 promoter more efficiently thanthe mutant promoter in ROS 17/2.8 cells.
Next, we sought to evaluate the effect of attenuatingendogenous Tbx2 expression on Cx43 expression inROS 17/2.8 cells. Cx43 promoter reporter constructs
Fig. 3. Binding to both TCACAC bindingsites in Cx43 promoter. Panel A:Electrophoretic mobility shift analysisusing radiolabeled WtCx43 37mer, whichcontains two TCACAC binding sites, asprobe (see text and Fig. 1 for sequence ofWtCx43 probe). Two DNA proteincomplexes form in the absence ofcompetitor (lane 2), both of which aredisplaced with increasing concentrations ofunlabeled WtCx43 competitor. Asindicated in the schematic, the upper bandis consistent with two proteins bound, oneto each half-site. Panel B: In order tocompare the binding affinity of theindividual binding sites, gel-shift assayswere performed using radiolabeled 19mers(depicted in Figure 1B) containing eachindividual TCACAC site (lanes 1–13).Whereas only a single DNA-proteinspecies is observed with each probecontaining only a single half-site, twobands are observed as in panel A withradiolabeled WtCx43 37mer, whichcontains two TCACAC binding sites (farright, lane 14); several lanes intercedingbetween lanes 13 and 14 were spliced outto simplify this figure. Cx43L contains theleft TCACAC site, and Cx43R containsthe right TCACAC site, as indicated inpanel B of Figure 1. Increasingconcentrations of box2 protein (0.1 to10 lM) were included as indicatedshowing a dose-dependent increase inmobility shift. Based on Phosporlmagerquantitation analysis, the 3¢ site binds tobox2 with approximately threefold higheraffinity than does the 5¢ site.
566 J.-R. Chen et al.: Tbx2 Represses Cx43 Expression

(pHXPL) were transiently cotransfected with an anti-sense or sense Tbx2 expression construct under controlof the CMV promoter or with an empty pcDNA3 vector
into ROS 17/2.8 cells. A third vector encoding lucif-erase, pGL3-luc, was included to control for transfec-tion efficiency. In order to show that the regulation isspecific, the regulatory relationship between Tbx2 andCx43 was also examined by using the reporter constructpHXPLmut3. The effect of decreasing Tbx2 expressionon Cx43 promoter activity was evaluated 48 hours aftertransfection by measuring ß-galactosidase activity, andß-galactosidase activity was normalized to luciferaseactivity and expressed as fold over control. The transientexpression of antisense Tbx2 led to an increase in ß-galactosidase activity in cells transfected with pHXPL,suggesting that attenuation of Tbx2 transcription byantisense Tbx2 led to de-repression of the reporter geneactivity. In contrast, no increase in ß-galactosidase ac-tivity was observed when a reporter construct withmutated Tbx2 binding sites (pHXPLmut3) was used(Fig. 4B). Together these results indicate that the de-repression lacZ reporter expression by antisense-Tbx2 ismediated via the two Tbx2 binding sites in the Cx43promoter.
Endogenous Tbx2 Expression Negatively Regulatesthe Endogenous Cx43 Gene
The experiments described earlier showed that Tbx2could repress the Cx43 promoter contained within thelacZ expression vector. To determine if Tbx2 also canmodulate the endogenous Cx43 promoter, ROS17/2.8cells were transfected with either sense or antisense Tbx2constructs to overexpress or attenuate endogenous Tbx2expression, respectively. Transfections of pcDNA3 vec-tor alone served as controls. Western blot analysis wascarried out to examine Tbx2 and Cx43 protein expres-sion in the transfected cells (Fig. 5). With the antisenseTbx2 construct, Tbx2 expression was markedly reduced.However, this was associated with only a small increasein the level of Cx43 expression. In contrast, transfectedcells expressing the sense Tbx2 construct exhibited amarked reduction in Cx43 expression (Fig. 5).
In stably transfected ROS cells, immunostaining us-ing a Cx43 polyclonal antibody showed an abundance ofCx43 gap junctions in ROS cells transfected with theempty vector (Fig. 6C), whereas cells expressing thesense Tbx2 transcript exhibited a marked decrease inCx43 gap junction plaques (Fig. 6B). In contrast, cellsexpressing antisense Tbx2 showed no obvious change inCx43 gap junction abundance (Fig. 6A), indicating apossible threshold for detection by immunohistochem-istry.
Analysis of these same stably transfected cells toquantitate the level of gap junctional communication bydye coupling analysis showed consistent changes in thelevel of dye coupling [48]. For these studies, individualcells were iontophoretically microinjected with carb-oxyfluorescein, and the spread of dye to the surrounding
Fig. 4. Tbx2 represses Cx43 promoter activity in culturedcells. (A) Transcriptional repression of Cx43 promoter-lacZconstructs in NIH3T3 cells. NIH3T3 cells were cotransfectedwith reporter constructs pHXPL and normalization controlpGL3-luc, together with either pcDNA3-Tbx2, pcDNA3alone, or with no expression vector. Luciferase and ß-gal ac-tivities were determined as presented in ‘‘Materials andMethods’’. Results are presented as the ratio of the ß-galac-tosidase activity normalized to luciferase compared with noexpression vector control; values represent means from threeseparate transfections. When the Tbx2 overexpression con-structs and the Cx43 promoter lacZ reporter genes were co-transfected into NIH3T3 cells, a cell line that does not haveendogenous Tbx2 expression, the expression of Tbx2 in 3T3cells represses Cx43 promoter activity (P < 0.05, Tbx2 com-pared to pcDNA3 or control based on unpaired t test). (B)Transfection of ROS 17/2.8 cells with antisense to Tbx2 de-represses the Cx43 promoter, ROS 17/2.8 cells were cotrans-fected with reporter constructs pHXPL or pHXPLmut3 withnormalization control pGL3-luc together with eitherpcDNA3-Tbx2antisense or with pcDNA3 vector alone as acontrol. Results are presented as the ratio of normalized ß-galactosidase activity compared with pcDNA3 control valuefor the respective reporter construct (pHXPL or pHXPL-mut3), set as 1 (dashed line); values represent means from threeseparate transfections. With the wild-type Cx43 promoterpresent in pHXPL, antisense transcripts to Tbx2 elevate ß-galactosidase expression, whereas with the pHXPLmut3 re-porter construct this derepression is abrogated (P < 0.0001,pHXPL compared to pHXPLmut3 or control; P > 0.25,pHXPLmut3 compared to control based on unpaired t test).
J.-R. Chen et al.: Tbx2 Represses Cx43 Expression 567

cells was used to quantitate the extent of gap junction–mediated dye transfer. As shown in Table 2, controlNIH3T3 or ROS cells exhibited a high level of dyecoupling, reflecting high levels of gap-junction expres-sion. With expression of Tbx2 in NIH3T3 cells, therewas a significant (>2.5-fold) decrease in functionalcoupling between cells. Likewise ROS 17/2.8 cellsoverexpressing Tbx2 through transfection with a senseTbx2 construct also exhibited a reduction in cell–cellcoupling. In contrast, no change in dye coupling wasobserved in ROS 17/2.8 cells expressing antisense Tbx2,a result consistent with no obvious change in theabundance of Cx43 gap junction plaques in thesetransfected cells. These findings suggest that Tbx2 canexert repression on the endogenous Cx43 gene in bothROS and NIH3T3 cells, but in ROS cells, the consti-tutive Tbx2 expression level may be insufficient tocompletely repress the endogenous Cx43 gene.
Analysis of Tbx2 Regulation of Cx43 PromoterConstructs In Transgenic Mice
The pHXPLmut3 construct was incorporated intotransgenic mice to examine the possible developmentaleffects of mutating the two Tbx2 binding sites on lacZreporter expression in the embryo. For this analysis,transgenic founder embryos were harvested at E10.5,E11.5, and E13.5 and stained with X-gal to detect lacZreporter gene expression. From this analysis, four X-gal-stained embryos were obtained, two at E10.5, one atE11.5, and one at E13.5. On a gross level, all embryosexhibited a pattern of LacZ expression similar to thatpreviously described for the pHXPL construct [39]: ex-pression was observed in the dorsal neural tube and inthe dorsal root ganglia, the pharyngeal arches, fronto-nasal mass, and other regions where neural crest deriv-atives are expected. However, one striking differencewas the observation in all four transgenic embryos car-
rying the mutated Cx43 promoter-lacZ construct of ß-galactosidase expression in the apical ectodermal ridge(Figs. 7A, B, C). This is a region where endogenousCx43 expression is expected [42, 50], but where ß-ga-lactosidase expression has never been observed in over adozen independent transgenic mouse lines possessing thepHXPL vector (LacZ expression driven by the wildtypeCx43 promoter sequence) (Fig. 7D) [39]. It is interestingto note that like Cx43, Tbx2 is also expressed in theapical ectodermal ridge during limb development [41].These observations suggest the interesting possibilitythat in the developing embryo, Tbx2 may play a rolein modulating Cx43 expression during limb morpho-genesis.
Discussion
The similar expression patterns of Tbx2 and Cx43 in theneural crest–derived mesenchyme in the pharyngealarches and other regions of the embryo [41, 42], togetherwith the presence of T-box consensus motifs in the Cx43promoter sequence led us to investigate the role of Tbx2in modulating Cx43 expression in osteoblast lineage–derived cells. By electrophoretic mobility shift assays, weconfirmed that there are at least two specific Tbx2binding sites in the 6.7-kb Cx43 promoter region. Weshowed that Tbx2 negatively regulates Cx43 promoter-lacZ reporter constructs and this negative regulation isdependent on the T-box sites, and is attenuated by ex-pression of an antisense Tbx2 construct. In addition, weshowed that the endogenous Cx43 gene can be repressedby de novo expression or overexpression of Tbx2 inNIH3T3 and ROS 17/2.8 cells, respectively. This re-pression can be observed as a decrease in Cx43 proteinexpression, a reduction in Cx43 gap-junction plaques,and decreased functional coupling between cells. How-ever, downregulating Tbx2 expression in ROS cells with
Fig. 5. Transcriptional de-repression ofendogenous Cx43 by antisense-Tbx2 inROS 17/2.8 cells. ROS cells weretransfected with either sense (S), antisenseTbx2 (AS), or vector alone (pcDNA3, C)respectively. The expression of Tbx2 (toppanel) and Cx43 (middle panel) proteinswere analyzed in cultures by use ofWestern blot analysis. The expression ofantisense to Tbx2 led to a markedreduction in Tbx2 protein levels and anincrease in the level of Cx43. The levelsof expression of Tbx2 and Cx43 werenormalized to actin expression (bottompanel).
568 J.-R. Chen et al.: Tbx2 Represses Cx43 Expression

Fig. 6. Reduction of Cx43 gap junctions in ROS17/2.8 cellsexpressing an antisense Tbx2 construct. Stably transfectedROS 17/2.8 cells containing antisense Tbx2 (panel A, D), senseTbx2 (panel B, E), or vector control (panel C, F) plasmids wereexamined for expression of Cx43 gap-junction contacts byusing immunofluorescence staining with a rabbit polyclonal
antibody to Cx43. Control cells (A, D) and cells transfectedwith antisense Tbx2 (C, F) show an abundance of the typicalpunctate Cx43 immunostaining at areas of cell–cell contact. Incontrast, cells transfected with the antisense construct (B, E)show a marked reduction in Cx43 immunostaining (scale bar is50 lm).
J.-R. Chen et al.: Tbx2 Represses Cx43 Expression 569

antisense Tbx2 only moderately increased Cx43 proteinexpression and did not significantly alter the level offunctional coupling or the abundance of Cx43 gap-junction plaques. The latter result suggests that theconstitutive level of Tbx2 expression in ROS cells isinsufficient to repress expression from the endogenousCx43 gene. This is perhaps not surprising given theconstitutively high level of Cx43 expression in ROS cells.On the other hand, when a Tbx2 sense transcript wasexpressed in ROS cells, Tbx2 protein expression waselevated, and Cx43 expression was reduced to barelydetectable levels. Taken together, these results suggestthat Cx43 is a bona fide target gene of Tbx2 and thatTbx2 exerts negative regulation on Cx43 gene expres-sion.
The crystal structure of the Brachyury (T) T-box-DNA co-crystal suggests that DNA binding by T(Brachyury) requires two half-sites [29]. Recently, it wasfound that Tbx2 could bind to a T half-site and repressthe melanocyte-specific tyrosinase-related protein-1(TRP-1) gene [32]. Moreover, a repression domain hasbeen localized to the C terminal domain of the Tbx2protein [34]. Of the four potential Tbx2 sites located inthe 6.7-kb Cx43 promoter, the most proximal two sitesare located )676 and )656 bp upstream of the start site()217). The results from the electrophoretic mobilityshift assays presented here suggest a model in whichTbx2 protein binds to each TCACAC site as a mono-mer. The realization that each T-box gene familymember has similar DNA binding requirements suggeststhe importance of the binding partners in mediating T-box gene function. Our results are consistent with anemerging picture that T-box–containing proteins (i.e.,TBX2, TBX3, and Brachyury) bind to TCACAC, whichis similar to half of the canonical consensus palindromicT-binding sequence determined by Herrmann et al. [6,27, 31–33]. Several transcription factor families (e.g.,homeobox and myoD) exhibit similarity in in vitro DNAbinding specificity, although functional heterogeneity isevident among different family members [51, 52]. Therepression of Cx43 by Tbx2 was more prominent inROS 17/2.8 cells than in NIH3T3 cells, suggesting thatthe differences in DNA-binding specificity may be
modulated in vivo by additional cellular factors. It is ofinterest that the T-box binding sites upstream of theCx43 promoter that mediate the observed repression areembedded in a region with multiple elements that me-diate activation in response to steroid and peptide hor-mones, which are known to have prominent effects onbone remodeling [53–57]. Future studies directed towardidentification of the molecular interactions between T-box genes and cofactors that bind to them are likely tocontribute to a better understanding of the functions ofthe T-box genes. While the present study demonstratedthat Tbx2 represses Cx43 in the mouse, there is prece-dence for T-box protein regulation of connexins, albeitin a transcriptional activation role: the expression levelof an ortholog of the mammalian connexin45 gene isdepressed in the zebrafish no tail (zebrafish T gene)mutant [40], and mouse Cx40 is depressed in Tbx5mutants [13].
Previous studies using transgenic and knockout micehave shown that the precise level of Cx43 expression inneural crest cells is of critical importance in regulatingcardiac outflow tract development and function. Thepresent study suggests that Tbx2 may play a rolein downregulating Cx43 expression. The in vivo co-expression patterns of Tbx2 and Cx43 and in vitrorepressor activity of Tbx2 for Cx43 constitutes an os-tensible paradox, yet the two proteins must coexist ifthey comprise a regulatory system; for Tbx2 to exert aneffect on the Cx43 promoter, whether positive or nega-tive, it must be present in the nucleus of cells expressingCx43. However, given that neural crest cells, like ROScells, express both Cx43 and Tbx2, the constitutive levelof Tbx2 expression in neural crest cells may not providesignificant repression of Cx43 on its own. Moreover,recent studies have demonstrated a reciprocal expressionpattern at developing rat suture blastemas, with Tbx2expression predominating centrally and Cx43 at theperiphery [43]. Such a reciprocal relationship would beanticipated in an antagonistic pairing, with one partnerrepressing its antagonist. The concept of a repressor andits target being co-expressed is difficult to reconcilewithout invoking an argument that Tbx2 is a componentin a regulatory network for Cx43, responsible for fine-tuning expression, rather than complete ablation. Re-pression by Tbx2 at the TRP1 promoter in melanocytesis subject to counterbalance by the activator Pax3,shown to bind to an overlapping site in the TRP1 pro-moter [58]. The thesis that fine-tune regulation by Tbx2may be exquisitely sensitive to dosage is supported bythe observation that, at minimum, four T-box genes(Tbx1, Tbx3, Tbx5, and Tbx22) manifest striking pa-thology in haploinsufficient individuals [5–7, 10, 12–15,23], and in each syndrome there is an associated elementof bone pathology.
Consistent with the thesis that Tbx2 plays an ancil-lary but important role in regulation of Cx43 in vivo,
Table 2. Effect of Tbx2 expression on dye couplinga
Cell Lines Dye Transfer
Control/3T3 (P1) 4.11 ± 0.190Tbx2sense/3T3 (K2) 1.48 ± 0.095
P < 0.0001Control/ROS 7.27 ± 0.400Tbx2antisense/ROS (AS8) 7.27 ± 0.470Tbx2sense/ROS (S14) 3.02 ± 0.258
P < 0.0001AS8 or control compared to S14
a Statistics based on paired t test
570 J.-R. Chen et al.: Tbx2 Represses Cx43 Expression

Fig. 7. Ectopic expression of ß-galactosidase localized toapical ectodermal ridge in transgenic mice–injected Cx43promoter-LacZ constructs lacking Tbx binding sites in a site-directed mutant Cx43 promoter-LacZ construct (pHXPL-mut3). Panels A, B, and C show forelimb buds from threeindependent transgenic embryos at E10.5–11.5 days postcoi-tus, derived from eggs injected with the pHXPLmut3 plasmid.Cells in the apical ectodermal ridge exhibit ß-galactosidase
expression (see white arrows). The expression levels are vari-able, likely due to insertion site effects on transcriptional ac-tivity of the LacZ insert. Shown in panel D is an E11.5 controlembryo limb bud derived from injection of the wild-type Cx43promoter-lacZ construct (pHXPL). Although there is exten-sive labeling on the dorsal surface of the limb bud, there is nolabeling in the apical ectodermal ridge (see white arrow) (scalebar is 200 lm).
J.-R. Chen et al.: Tbx2 Represses Cx43 Expression 571

transgenic mice containing this mutated pHXPL con-struct exhibited the same pattern of LacZ expression asseen in mice containing the wildtype Cx43 promoterdriven pHXPL construct: expression is localized to thedorsal neural tube and neural crest derivatives. Giventhese results, we hypothesize that Tbx2 requires the co-expression of other developmentally regulated tran-scription factors to modulate Cx43 expression in mi-grating neural crest cells. Perhaps Tbx2 helps poise theneural crest cells for rapid Cx43 downregulation—suchas may occur when subpopulations of neural crest cellsreach their target sites [59]. It is interesting to note thatinjection of the mutant pHXPLmut3 construct yieldedß-galactosidase expression in the apical ectodermalridge, which has never been observed in transgenic micecarrying the wildtype pHXPL construct (Fig. 7 [39]).The apical ectodermal ridge, like cardiac neural crestcells, is another tissue where endogenous Cx43 and Tbx2genes are co-expressed. These results further support arole for Tbx2 in repressing the Cx43 promoter in oste-oblastic progenitor cells. The fact that the endogenousCx43 gene but not the pHXPL construct is expressed inthe apical ectodermal ridge would suggest that the roleof Tbx2 in modulating transcription from the Cx43promoter may be dependent on other parameters. Forexample, the chromatin context may serve to limit ac-cessibility of the promoter to binding by Tbx2 and/orother regulatory factors, a subject of investigation forfuture studies.
Acknowledgments. We thank Drs. Mark G. Anderson, andJuren Huang for assistance in interpreting Cx43 promoteranalyses, as well as Kimberly Crawford and Jamie Cranfordfor excellent technical support. This study was supported by aGrant-in-Aid from the American Heart Association/South-eastern Affiliate, the National Space Biomedical Research In-stitute (NSBRI) through a NASA Cooperative AgreementNCC-9-58, and by grants HD34149, HL36059, HD17069,DE12812, and HL51533 from the National Institute of Health.
References
1. Bollag RJ, Siegfried Z, Cebra-Thomas JA, Garvey N,Davison EM, et al. (1994) An ancient family of embryo-nically expressed mouse genes sharing a conserved proteinmotif with the T locus. Nat Genet 7:383–389
2. Herrmann BG, Labeit S, Poustka A, King TR, Lehrach H(1990) Cloning of the T gene required in mesoderm for-mation in the mouse. Nature 343:617–622
3. Herrmann BG, Kispert A (1994) The T genes in embryo-genesis. Trends Genet 10:280–286
4. Griffin KJP, Amacher SL, Kimmel CB, Kimelman D(1998) Molecular identification of spadetail: regulation ofzebrafish trunk and tail mesoderm formation by T-boxgenes. Development 125:379–388
5. Jerome LA, Papaioannou VE (2001) DiGeorge syndromephenotype in mice mutant for the T-box gene, Tbx1. NatGenet 27:286–291
6. Lindsay EA, Vitelli F, Su H, Morishima M, Huynh T,et al. (2001) Tbx1 haploinsufficieny in the DiGeorge syn-drome region causes aortic arch defects in mice. Nature410:97–101
7. Merscher S, Funke B, Epstein JA, Heyer J, Puech A, et al.(2001) TBX1 is responsible for cardiovascular defects invelo-cardio-facial/DiGeorge syndrome. Cell 104:619–629
8. Hug B, Walter V, Grunwald DJ (1997) Tbx6, a Brachy-ury-related gene expressed by ventral mesendodermalprecursors in the zebrafish embryo. Dev Biol 183:61–73
9. Chapman DL, Papaioannou VE (1998) Three neural tubesin mouse embryos with mutations in T-box gene Tbx6.Nature 391:695–697
10. Li QY, Newbury-Ecob RA, Terrett JA, Wilson DI, CurtisARJ, et al. (1997) Holt-Oram syndrome is caused bymutations in TBX5, a member of the Brachyury (T) genefamily. Nat Genet 15:21–29
11. Horb ME, Thomsen GH (1999) Tbx5 is essential for heartdevelopment. Development 126:1739–1751
12. Basson CT, Bachinski DR, Lin RC, Levi T, Elkins JA,et al. (1997) Mutations in humans cause limb and cardiacmalformation in Holt-Oram syndrome. Nat Genet 15:30–35
13. Bruneau BG, Nemer G, Schmitt JP, Charron F, RobitailleL, et al. (2001) A murine model of Holt-Oram syndromedefines roles of T-box transcription factor Tbx5 in cardi-ogenesis and disease. Cell 106:709–721
14. Bamshad M, Le T, Watkins WS, Dixon ME, Kramer BE,et al. (1997) The spectrum of mutations in TBX3: geno-type/phenotype relationship in ulnar-mammary syndrome.Am J Hum Genet 64:1550–1562
15. Bamshad M, Lin RC, Law DJ, Watkins WS, KrakowiakPA, et al. (1997) Mutations in human TBX3 alter limb,apocrine and genital development in ulnar-mammarysyndrome. Nat Genet 16:311–315
16. He M-L, Wen L, Campbell CE, Wu JY, Rao Y (1999)Transcription repression by Xenopus ET and its humanortholog TBX3, a gene involved in ulnar-mammary syn-drome. Proc Natl Acad Sci U S A 96:10212–10217
17. Gibson-Brown JJ, Agulnik SI, Silver LM, Niswander L,Papaioannou VE (1998) Involvement of T-box genesTbx2-Tbx5 in vertebrate limb specification and develop-ment. Development 125:2499–2509
18. Logan M, Simon H-G, Tabin C (1998) Differential regu-lation of T-box and homeobox transcription factors sug-gests roles in controlling chick limb-type identity.Development 125:2825–2835
19. Ohuchi H, Takeuchi J, Yoshioka H, Ishimaru Y, OguraK, et al. (1998) Correlation of wing-leg identity in ectopicFGF-induced chimeric limbs with the differential expres-sion of chick Tbx5 and Tbx4. Development 125:51–60
20. Rodriguez-Esteban C, Tsukui T, Yonei S, Magallon J,Tamura K, et al. (1999) The T-box genes Tbx4 andTbx5 regulate limb outgrowth and identity. Nature398:814–818
21. Russ AP, Wattler S, Colledge WH, Aparicio SAJR,Carlton MBL, et al. (2000) Eomesodermin is required formouse trophoblast development and mesoderm forma-tion. Nature 404:95–99
22. Lamolet B, Pulichino A-M, Lamonerie T, Gauthier Y,Brue T, et al. (2001) A pituitary cell-restricted T box fac-tor, Tpit, activates POMC transcription in cooperationwith Pitx homeoproteins. Cell 104:849–859
23. Braybrook C, Doudney K, Mancano ACB, Arnason A,Bjornsson A, et al. (2001) The T-box transcription factorgene TBX22 is mutated in X-linked cleft palate and an-kyloglossia. Nat Genet 29:179–183
24. Finotto S, Neurath MF, Glickman JN, Qin S, Lehr HA,et al. (2002) Development of spontaneous airway changesconsistent with human asthma in mice lacking T-bet.Science 295:336–338
25. Agulnik SI, Bollag RJ, Silver LM (1995) Conservation ofthe T-box gene family from M. musculus to C. elegans.Genomics 25:214–219
26. Campbell C, Goodrich K, Casey G, Beatty B (1995)Cloning and mapping of a human gene (TBX2) sharing ahighly conserved protein motif with the Drosophila ombgene. Genomics 28:255–260
572 J.-R. Chen et al.: Tbx2 Represses Cx43 Expression

27. Kispert A, Herrmann BG (1993) The Brachyury gene en-codes a novel DNA binding protein. EMBO J 12:3211–3220
28. Kispert A, Koschorz B, Hermann BG (1995) The T pro-tein encoded by Brachyury is a tissue-specific transcriptionfactor. EMBO J 14:4763–4772
29. Muller CW, Herrmann BG (1997) Crystallographicstructure of the T domain-DNA complex of the Brachyurytranscription factor. Nature 389:884–888
30. Chen J-R, Zhong Q, Wang J, Cameron RS, Borke JL,et al. (2001) Microarray analysis of Tbx2-directed geneexpression: a possible role in osteogenesis. Mol Cell End-ocrinol 177:43–54
31. Casey ES, O’Reilly M-AJ, Conlon FL, Smith JC (1998)The T-box transcription factor Brachyury regulates ex-pression of eFGF through binding to a non-palindromicresponse element. Development 125:3887–3894
32. Carreira S, Dexter TJ, Yavuzer , Goding CR (1998)Brachyury-related transcription factor Tbx2 and repres-sion of the melanocyte-specific TRP-1 promoter. Mol CellBiol 18:5099–5108
33. Sinha S, Abraham S, Gronostajski RM, Campbell CE(2000) Differential DNA binding and transcription mod-ulation by three T-box proteins, T,TBX1 and TBX2. Gene258:15–29
34. Paxton C, Zhao H, Chin Y, Langner K, Reecy J (2002)Murine Tbx2 contains domains that activate and repressgene transcription. Gene 283:117–124
35. Ewart JL, Cohen MF, Meyer RA, Huang GY, Wessels A,et al. (1997) Heart and neural tube defects in transgenicmice overexpressing the Cx43 gap junction gene. Devel-opment 124:1281–1292
36. Huang GY, Wessels A, Smith BR, Linask KK, Ewart JL,et al. (1998) Alteration in connexin 43 gap junction genedosage impairs conotruncal heart development. Dev Biol198:32–44
37. Lecanda F, Warlow PM, Sheikh S, Furlan F, SteinbergTH, et al. (2000) Connexin43 deficiency causes delayedossification, craniofacial abnormalities, and osteoblastdysfunction. J Cell Biol 151:931–943
38. Li Z, Zhou Z, Yellowley CE, Donahue HJ (1999) Inhib-iting gap junctional intercelular communication alters ex-pression of differentiation markers in osteoblastic cells.Bone 25:661–666
39. Lo CW, Cohen MF, Huang G-Y, Lazatin BO, Patel N,et al. (1997) Cx43 gap junction gene expression and gapjunctional communication in mouse neural crest cells. DevGenet 20:119–132
40. Essner JJ, Laing JG, Beyer EC, Johnson RG, Hackett PB(1996) Expression of zebrafish connexin43.4 in the noto-chord and tail bud of wild-type and mutant no tail em-bryos. Dev Biol 177:449–462
41. Chapman DL, Garvey N, Hancock S, Alexiou M, AgulnikSI, et al. (1996) Expression of the T-box family genes,Tbx1-Tbx5, during early mouse development. Dev Dyn206:379–390
42. Ruangvoravat CP, Lo CW (1992) Connexin43 expressionin the mouse embryo: localization of transcripts withindevelopmentally significant domains. Dev Dyn 194:261–281
43. Borke JL, Chen JR, Yu JC, Bollag RJ, Orellana MF, et al.(2003) Negative transcriptional regulation of connexin 43
by Tbx2 in rat immature coronal sutures and ROS 17/2.8cells in culture. Cleft Palate Craniofac. J 40 (in press)
44. Bollag RJ, Zhong Q, Phillips P, Min L, Qin F, et al. (2000)Osteoblast-derived cells express functional glucose-dependent insulinotropic receptors. Endocrinology 141:1228–1235
45. Martin CS, Wight PA, Dobretsova A, Bronstein I (1996)Dual luminescence-based reporter gene assay for luciferaseand ß-galactosidase. Bio techniques 21:520–524
46. Harlow E, Lane D (1988) Antibodies: A LaboratoryManual. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, NY
47. Waldo KL, Lo CW, Kirby ML (1999) Connexin 43 ex-pression reflects neural crest patterns during cardiovascu-lar development. Dev Biol 208:307–323
48. Sullivan R, Lo CW (1995) Expression of a connexin 43/ß-galactosidase fusion protein inhibits gap junctional com-munication in NIH3T3 cells. J Cell Biol 130:419–429
49. Yamada M, Rivelli J-P, Eichele G, Barron M, SchwartzRJ (2000) Expression of chick Tbx-2, Tbx-3, and Tbx-5genes during early heart development: evidence for BMP2induction of Tbx-2. Dev Biol 228:95–105
50. Meyer RA, Cohen MF, Recalde S, Zakany J, Bell SM,et al. (1997) Developmental regulation and asymmetricexpression of the gene encoding Cx43 gap junctions in themouse limb bud. Dev Genet 21:290–300
51. McGinnis W, Krumlauf R (1992) Homeobox genes andaxial patterning. Cell 68:283–302
52. Olson EN, Klein WH (1994) bHLH factors in muscledevelopment: dead lines and commitments, what to leavein what to leave out. Genes Dev 8:1–8
53. Yu W, Dahl G, Werner R (1994) The connexin43 gene isresponsive to oestrogen. Proc R Soc Lond B Biol Sci255:125–132
54. Chen Z-Q, Lefebvre D, Bai X-H, Reaume A, Rossant J,et al. (1995) Identification of two regulatory elementswithin the promoter region of the mouse connexin43 gene.J Biol Chem 270:3863–3868
55. Geimonen E, Wei J, Ali M, Fishman GI, Garfield RE,et al. (1996) Activation of protein kinase C in humanuterine smooth muscle induces connexin43 gene tran-scription through AP-1 site in the promoter sequence. JBiol Chem 271:23667–23674
56. Civitelli R, Ziambaras K, Warlow PM, Lecanda F, NelsonT, et al. (1998) Regulation of connexin43 expression andfunction by prostaglandin E2 (PGE2) and parathyroidhormone (PTH) in osteoblastic cells. J Cell Biochem 68
57. Stock A, Sies H (2000) Thyroid hormone receptors bind toan element in the connexin43 promoter. Biol Chem381:973–979
58. Galibert M-D, Yavuzer U, Dexter TJ, Coding CR (1999)Pax3 and regulation of the melanocyte-specific tyrosinase-related protein-1 promoter. J Biol Chem 274:26894–26900
59. Lo CW (1996) The role of gap junction membrane chan-nels in development. J Bioenerg Biomembr 28:379–385
60. Echetebu CO, Ali M, Izban MG, MacKay L, Garfield RE(1999) Localization of regulatory protein binding sites inthe proximal region of human myometrial connexin 43gene. Mol Hum Rep 5:757–766
61. Fernandez-Cobo M, Stewart D, Drujan D, De Maio A(2001) Promoter activity of the rat connexin 43 gene inNRK cells. J Cell Biochem 81:514–522
J.-R. Chen et al.: Tbx2 Represses Cx43 Expression 573





























