Embed Size (px)
Citation preview

Acetylation represses the binding of CheY to itstarget proteinsmmi_7148 932..943
Orna Liarzi,1 Rina Barak,1 Vered Bronner,2
Monica Dines,2† Yael Sagi,1‡ Alla Shainskaya3 andMichael Eisenbach1*1Department of Biological Chemistry, and 3BiologicalMass Spectrometry Facility of the Department ofBiological Services, The Weizmann Institute of Science,76100 Rehovot, Israel.2Bio-Rad Haifa, 32000 Haifa, Israel.
Summary
The ability of CheY, the response regulator of bacte-rial chemotaxis, to generate clockwise rotation isregulated by two covalent modifications – phospho-rylation and acetylation. While the function and signalpropagation of the former are widely understood, themechanism and role of the latter are still obscure. Toobtain information on the function of this acetylation,we non-enzymatically acetylated CheY to a levelsimilar to that found in vivo, and examined its bindingto its kinase CheA, its phosphatase CheZ and theswitch protein FliM – its target at the flagellar switchcomplex. Acetylation repressed the binding to allthree proteins. These results suggest that both phos-phorylation and acetylation determine CheY’s abilityto bind to its target proteins, thus providing two levelsof regulation, fast and slow respectively. The fastlevel is modulated by environmental signals (e.g.chemotactic and thermotactic stimuli). The slow oneis regulated by the metabolic state of the cell and itdetermines, at each metabolic state, the fraction ofCheY molecules that can participate in signalling.
Introduction
CheY, the response regulator of bacterial chemotaxis,transduces the chemotactic signal from the receptorsupramolecular complex, located at the cell pole, to theswitch elements in the flagellar–motor supramolecular
complexes, which are randomly distributed around thecell. Phosphorylation of CheY by its cognate receptor-bound histidine kinase, CheA, reduces their mutualbinding affinity and elevates the binding affinity of CheYfor the switch protein FliM and for the phosphatase CheZ.The binding of phosphorylated CheY (CheY~P) to theswitch promotes clockwise rotation. Downregulation ofthe phosphorylation level of CheY is achieved by thephosphatase CheZ (for reviews, see Eisenbach, 2004;Wadhams and Armitage, 2004; Baker et al., 2006).
Acetylation is another covalent modification that acti-vates CheY to generate clockwise rotation (Wolfe et al.,1988; Barak et al., 1992; 1998; Ramakrishnan et al.,1998). Two mechanisms of CheY acetylation have beenrevealed: AcCoA synthetase (Acs)-catalysed acetylationwith acetate or acetyl coenzyme A (AcCoA) as an acetyldonor (Barak et al., 1992; 2004), and autoacetylation(Barak et al., 2006) with AcCoA as the acetyl donor. Bothacetylation mechanisms were shown to be physiologicallyimportant for chemotaxis, as defects in any of them lead todefective chemotaxis (Barak and Eisenbach, 2001; Baraket al., 2006). Furthermore, the level of CheY acetylation invivo is remarkably high: on average, every CheY moleculeis acetylated to a level of one acetyl group per molecule;however, it is essentially a mixture of CheY molecules withone (primarily), zero and two acetyl groups per molecule(Yan et al., 2008). Both covalent modifications of CheY ap-pear to be linked. Thus, CheY’s phosphorylating and de-phosphorylating agents inhibit and enhance, respectively,CheY autoacetylation (Barak et al., 2006) and Acs-mediated acetylation (Barak and Eisenbach, 2004). Fur-thermore, the acetylating enzyme Acs elevates the phos-phorylation level of CheY (Barak and Eisenbach, 2004).
CheY is acetylated at multiple sites, all clustered at theC-terminus of the protein (Barak et al., 2004). These sites(lysine residues 91, 92, 109, 119, 122 and 126)(Ramakrishnan et al., 1998; Barak et al., 2004) are local-ized to the surface of CheY that binds its target proteins:CheA, CheZ and FliM (Shukla and Matsumura, 1995; Zhuet al., 1997; Shukla et al., 1998; Welch et al., 1998;McEvoy et al., 1999). Furthermore, structural studiesdemonstrated that several of these acetylation sites aredirectly involved in CheY binding to FliM (Lee et al., 2001;Dyer et al., 2004; Dyer and Dahlquist, 2006), CheZ (Zhaoet al., 2002; Guhaniyogi et al., 2006) and CheA (Welch
Accepted 18 March, 2010. *For correspondence. [email protected]; Tel. (+972) 8 9343923; Fax (+972) 89472722. †Department of Neurobiology and Ethology, Faculty ofScience and Science Education, University of Haifa, Mt. Carmel31905, Israel; ‡Department of Medicine, Division of Oncology, Stan-ford University School of Medicine, Stanford, CA 94305, USA.
Molecular Microbiology (2010) 76(4), 932–943 � doi:10.1111/j.1365-2958.2010.07148.xFirst published online 14 April 2010
© 2010 Blackwell Publishing Ltd
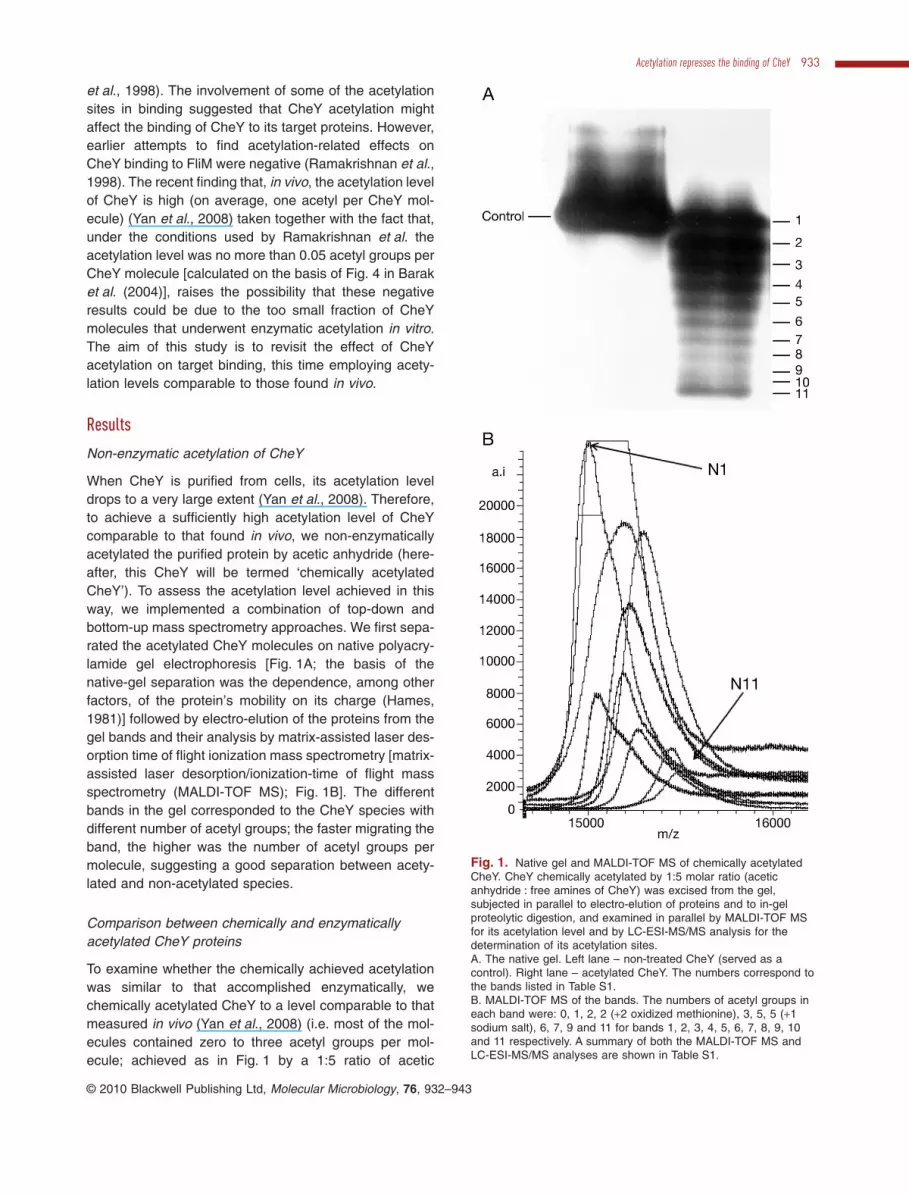
et al., 1998). The involvement of some of the acetylationsites in binding suggested that CheY acetylation mightaffect the binding of CheY to its target proteins. However,earlier attempts to find acetylation-related effects onCheY binding to FliM were negative (Ramakrishnan et al.,1998). The recent finding that, in vivo, the acetylation levelof CheY is high (on average, one acetyl per CheY mol-ecule) (Yan et al., 2008) taken together with the fact that,under the conditions used by Ramakrishnan et al. theacetylation level was no more than 0.05 acetyl groups perCheY molecule [calculated on the basis of Fig. 4 in Baraket al. (2004)], raises the possibility that these negativeresults could be due to the too small fraction of CheYmolecules that underwent enzymatic acetylation in vitro.The aim of this study is to revisit the effect of CheYacetylation on target binding, this time employing acety-lation levels comparable to those found in vivo.
Results
Non-enzymatic acetylation of CheY
When CheY is purified from cells, its acetylation leveldrops to a very large extent (Yan et al., 2008). Therefore,to achieve a sufficiently high acetylation level of CheYcomparable to that found in vivo, we non-enzymaticallyacetylated the purified protein by acetic anhydride (here-after, this CheY will be termed ‘chemically acetylatedCheY’). To assess the acetylation level achieved in thisway, we implemented a combination of top-down andbottom-up mass spectrometry approaches. We first sepa-rated the acetylated CheY molecules on native polyacry-lamide gel electrophoresis [Fig. 1A; the basis of thenative-gel separation was the dependence, among otherfactors, of the protein’s mobility on its charge (Hames,1981)] followed by electro-elution of the proteins from thegel bands and their analysis by matrix-assisted laser des-orption time of flight ionization mass spectrometry [matrix-assisted laser desorption/ionization-time of flight massspectrometry (MALDI-TOF MS); Fig. 1B]. The differentbands in the gel corresponded to the CheY species withdifferent number of acetyl groups; the faster migrating theband, the higher was the number of acetyl groups permolecule, suggesting a good separation between acety-lated and non-acetylated species.
Comparison between chemically and enzymaticallyacetylated CheY proteins
To examine whether the chemically achieved acetylationwas similar to that accomplished enzymatically, wechemically acetylated CheY to a level comparable to thatmeasured in vivo (Yan et al., 2008) (i.e. most of the mol-ecules contained zero to three acetyl groups per mol-ecule; achieved as in Fig. 1 by a 1:5 ratio of acetic
Fig. 1. Native gel and MALDI-TOF MS of chemically acetylatedCheY. CheY chemically acetylated by 1:5 molar ratio (aceticanhydride : free amines of CheY) was excised from the gel,subjected in parallel to electro-elution of proteins and to in-gelproteolytic digestion, and examined in parallel by MALDI-TOF MSfor its acetylation level and by LC-ESI-MS/MS analysis for thedetermination of its acetylation sites.A. The native gel. Left lane – non-treated CheY (served as acontrol). Right lane – acetylated CheY. The numbers correspond tothe bands listed in Table S1.B. MALDI-TOF MS of the bands. The numbers of acetyl groups ineach band were: 0, 1, 2, 2 (+2 oxidized methionine), 3, 5, 5 (+1sodium salt), 6, 7, 9 and 11 for bands 1, 2, 3, 4, 5, 6, 7, 8, 9, 10and 11 respectively. A summary of both the MALDI-TOF MS andLC-ESI-MS/MS analyses are shown in Table S1.
Acetylation represses the binding of CheY 933
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 76, 932–943

anhydride : free amines of CheY), and then identified theacetylation sites by proteolytic digestion of the proteinfollowed by peptide fingerprinting and sequencing by liquidchromatography electrospray ionization tandem massspectrometry (LC-ESI-MS/MS). The chemical acetylationresulted in acetylation at multiple sites (lysine residues 4,7, 26, 70, 91, 92, 109, 119 and 122; Table S1). To evaluatethe relative abundances of the chemical acetylation sites,we performed precursor ion scan on the band correspond-ing to singly acetylated CheY species. We found thatLys-122 was the site most acetylated, followed by Lys-92(Fig. 2 and Fig. S1). To examine whether these are alsothe main acetylation sites in vivo, we compared the levelsof CheY acetylation in vivo (employing Western blots withanti-acetyl-lysine antibody) between strains overproduc-ing CheY-K92R/K122T, CheY-K92R/K109R and wild-typeCheY (all in a DcheY background). The basal acetylation
levels of CheY in the mutant CheY-K92R/K122T was sig-nificantly lower than the other two strains (Fig. 3), suggest-ing that lysine residues 122 and 92 are the mainacetylation sites in vivo and that there was no, or little,compensation of the mutation K92R/K122T by otheracetylation sites. This pattern is somewhat different fromthe precursor ion scan obtained for Acs-mediated acety-lation and autoacetylation, where the highly abundantacetylation sites were all located at the C-terminus of theprotein (lysine residues 91, 92, 109, 119, 122 and 126)(Barak et al., 2004; 2006).
Effect of acetylation on the binding of CheY to itstarget proteins
To determine the effect of CheY acetylation on its bindingto the kinase CheA, the phosphatase CheZ, and the
Fig. 2. Acetylated lysine-specific precursor ion scan of singly acetylated CheY at m/z 126.1. CheY acetylated by 1:5 molar ratio (aceticanhydride : free amines of CheY) was separated on native gel. Each band was excised from the gel and examined by MALDI-TOF MS for thedetermination of the number of acetyl groups per molecule. The band corresponding to one acetyl group per molecule was subjected toprecursor ion scan analysis to determine the relative abundances of each of the acetylation sites. The peaks of peptides, which appeared inthe ion scan and were later confirmed by tandem mass spectrometric analysis, are designated on the spectrum. Inset, acetylated lysineresidues identified by Mascot search. Mascot scores higher than 35 indicate high reliability of identification. The relative abundance of theacetylation sites was determined on the basis of both the precursor ion scan and the Mascot score.
934 O. Liarzi et al. �
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 76, 932–943

switch protein FliM, we chemically acetylated CheY tovarious levels, ranging from below the level found in vivoto higher levels (Fig. 4; the acetylation levels defined as 1,2, 3 etc.; non-acetylated CheY defined as acetylation level0; the acetylation level in vivo was roughly comparable tolevel 1 or to between levels 1 and 2, depending on theacetylation batch). For the binding measurement, weemployed surface plasmon resonance (SPR), carried outwith ProteOn (6 ¥ 6 channels). We carried out the bindingassays to FliM and CheZ with phosphorylated CheY (i.e.in the presence of the phosphodonor acetyl phosphate),and to CheA with non-phosphorylated CheY (i.e. in theabsence of acetyl phosphate) – conditions that yield thestrongest binding of CheY to its target proteins (Schusteret al., 1993; Welch et al., 1993; Blat and Eisenbach,1994). We had to carry out two different sets of experi-ments, one set for CheA and CheZ, and the other for FliM.In the former, we immobilized His-CheY (at various acety-lation levels) onto the sensor chip, and CheA and CheZ(one at a time) were the analytes. In the other set,because non-tagged FliM was not sufficiently stable as ananalyte, we immobilized His-tagged FliM to the chip andused non-tagged CheY with various acetylation levels (allinjected simultaneously at a constant concentration) asanalytes.
In the first set of experiments, acetylated His-CheY,having the compositions shown in Fig. 4A (CheA andCheZ columns), bound CheA and CheZ to a lower extentthan did non-acetylated His-CheY, the higher the acetyla-tion level the lower the binding (Fig. 4B and C). The dis-sociation constant (Kd), however, did not significantly
change with the acetylation level and remained similar atall acetylation levels in which the Kd could be calculated(Figs S2–S4). Furthermore, at high acetylation levels His-CheY did not bind at all to CheA and very little to CheZ(Fig. 4B). These results suggest that the main, if not sole,species that bind CheA and CheZ is non-acetylated His-CheY, with Kd values (averaged for all measurable acety-lation levels) of 5.8 � 1.6 nM (� SEM, n = 8) for CheAand 0.15 � 0.04 mM (n = 5) for CheZ. These Kd values arecomparable or even lower than those reported in theliterature: 30 nM for binding of CheA to CheY immobilizedonto a different kind of SPR chip (Schuster et al., 1993)and ~2 mM for the binding of the P2 domain of CheA thatcontains the CheY binding site (Swanson et al., 1995);and 26 mM for the CheZ peptide that contains the CheY~Pbinding site (McEvoy et al., 1999). [The differencebetween the two SPR results of CheY-CheA binding maybe the consequence of the different techniques employedto immobilize CheY to the chip: an interaction between theHis-tag on CheY and Tris-nitrilotriacetic acid that coats thechip (this study), and a disulphide bond to an engineeredcysteine at residue 51 of CheY (Schuster et al., 1993), aprocedure that may have affected the binding propertiesof CheY.]
In the other set of experiments, a similar phenomenonwas observed: acetylated CheY, having the compositionsshown in Fig. 4A (His-FliM column), bound His-FliM to alower extent than did non-acetylated CheY, the higher theacetylation level the lower the binding (Fig. 4B and C). Inthis case, however, the association and dissociation(Fig. 4B, His-FliM column) were too fast to be fitted into akinetic model for calculating the kinetic and dissociationconstants. On the basis of the similarity of the His-FliMbinding results to those of CheA and CheZ (Fig. 4), andbecause the interface of CheY that binds FliM overlapsthe interface that binds CheA and CheZ (Zhu et al., 1997),it is reasonable that, here too, the binding of His-FliM ispredominantly to non-acetylated CheY.
We also measured by SPR the binding of acetylated butnon-phosphorylated CheY to FliM and CheZ, yieldingsimilar binding trends (Fig. S4 for CheZ) and, in the caseof CheZ, the Kd was two orders of magnitude higher thanthe Kd value obtained under phosphorylating conditions.Binding of phosphorylated CheY to CheA was not mea-sured, as this form does not bind CheA (Schuster et al.,1993).
To address the possibility that the reduced binding wasdue to the way by which the acetylation had beenachieved, i.e. due to the chemical rather than enzymaticacetylation, we measured the binding of purified, enzy-matically (by Acs) acetylated His-CheY to a preparation ofisolated, intact switch complexes of the flagellar motor(Sagi et al., 2003) by employing a highly sensitive double-labelling assay, described in Supporting information.
Fig. 3. In vivo basal acetylation of wild-type and double-mutantCheY proteins. Cell-free extracts were loaded on two equal gelsand subjected to SDS-PAGE. One gel was used for Western blotswith an anti-acetyl-lysine antibody and the other for Coomassieblue staining. The values shown are the normalized ratios betweenthe intensity (measured with a densitometer) of the CheY band inthe Western blot and the relative intensity of the equivalent band inthe Coomassie blue gel (mean � SEM of three independentexperiments). An asterisk indicates a statistically significantdifference from the wild type (P = 0.018 according to two-wayANOVA, followed by Dunnett’s post-test).
Acetylation represses the binding of CheY 935
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 76, 932–943

Fig. 4. Binding of CheY to its target proteins.A. Native gels of the CheY samples used for each experiment. For the binding of CheY to His-FliM, acetylation levels 1, 2, 3, 4 and 5 wereachieved by using 1:5, 1:3, 1:2, 1:1.2 and 1:1.0 molar ratios of acetic anhydride : free amines of CheY respectively. For the binding of CheAand CheZ to His-CheY, acetylation levels 1, 2 and 3 were achieved by using 1:5, 1:2 (different acetylation batch) and 1:2 molar ratio of aceticanhydride : free amines of CheY respectively. (Note that acetylation level 2 was obtained from an acetylation batch different from the one usedin all other lanes, resulting in a different acetylation level; this level was absent in the binding to CheZ.) In all experiments, each band fromacetylation level 1 was excised from the gel and examined by MALDI-TOF MS for the number of acetyl groups per molecule. Non-treatedCheY (i.e. acetylation level 0) served as a control.B. A representative SPR sensorgram of the interaction between the CheY samples shown in (A) and the target protein. The immobilizationbinding signals were 3900 RU for the binding His-FliM to CheY (70 mM); 520, 650, 560 and 500 RU for the binding of His-CheY withacetylation levels 0, 1, 2 and 3, respectively, to CheA (460 nM); and 1000, 800 and 850 RU for the binding of His-CheY with acetylation levels0, 1 and 3, respectively, to CheZ (3.6 mM). In the case of FliM and CheZ, phosphorylation of CheY was achieved by the presence of AcP bothin the running buffer and in the injected protein sample. The apparently ‘negative’ CheA binding at acetylation levels 2 and 3 was due to thefact that the binding to CheY acetylated to these levels was lower than the negative control [FliM, which does not interact with CheA (Brenand Eisenbach, 2000), immobilized onto the very same chip in a separate channel], subtracted from it. The negative control for the bindingbetween CheY~P and FliM or CheZ was the immobilization to the same chip of the irrelevant protein Barstar fused to cellulose. The arrowindicates analyte removal.C. Average binding signal just prior to analyte removal for each of the acetylation levels (mean � SEM). Acetylation levels 2 and 3 are notshown in the CheA panel because of the ‘negative’ binding (for explanation, see B above). One asterisk, a statistically significant differencefrom non-acetylated CheY, P < 0.05 according to repeated measures ANOVA with Tukey–Kramer post-test (n = 12 runs) for His-FliM, andaccording to paired t-test (n = 4) for CheA. Two asterisks, P < 0.01 according to repeated measures ANOVA with Tukey–Kramer post-test(n = 3). Three asterisks, P < 0.001 according to repeated measures ANOVA with Tukey–Kramer post-test (n = 12).
936 O. Liarzi et al. �
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 76, 932–943

Consistent with the results described above, the moreacetylated CheY became, the lower was the binding ofenzymatically acetylated CheY to the switch complex(Fig. S5).
To investigate the possibility that acetylation of CheYmay interfere with the ability of the protein to autophos-phorylate with acetyl phosphate as the donor, we incu-bated acetyl [32P]phosphate with CheY molecules, pre-acetylated to a level that did not contain non-acetylatedmolecules (Fig. 5A). Clearly, CheY became radiolabelled(i.e. phosphorylated) irrespective of whether or not it waspre-acetylated (Fig. 5B). Since all the CheY molecules inthis sample were pre-acetylated, we conclude that acety-lation does not interfere with the ability of CheY toundergo autophosphorylation with acetyl phosphate asthe donor (but it does interfere with its ability to undergophosphorylation by CheA because of the apparent inabil-ity of acetylated CheY to bind to CheA).
Discussion
The results of this study demonstrate that acetylation ofCheY interferes with the binding of this protein to all itstarget proteins – CheA, CheZ and FliM. These resultstaken together with the observation that CheY is highlyacetylated in vivo (Yan et al., 2008) have far-reachingimplications for the mechanism by which the chemotactic
signal is regulated and, as discussed below, they chal-lenge our current view that only phosphorylation regulatesthe signal.
Are the results obtained with chemically acetylatedCheY physiologically meaningful?
The relatively high acetylation level of CheY in vivo [onaverage, one acetyl per CheY molecule (Yan et al., 2008)]cannot be achieved in vitro by the currently known mecha-nisms – autoacetylation (Barak et al., 2006) and Acs-mediated acetylation (Barak et al., 2004). Thesemechanisms can acetylate CheY (after many hours) to amaximal level of 0.15 acetyl groups per CheY moleculeonly (Barak et al., 2004; 2006), suggesting that, in addi-tion, an acetyltransferase is involved in acetylating CheYin vivo (Yan et al., 2008). Because the identity of thisacetylating agent is not yet known, the only way to cur-rently achieve in vitro acetylation levels of CheY compa-rable to those measured in vivo is to chemically (i.e.non-enzymatically) acetylate CheY. The similaritybetween the lysine residue positions chemically acety-lated and those acetylated in vivo (Figs 2 and 3 andrelated text) suggests that the results obtained withchemically acetylated CheY are physiologicallymeaningful. This is endorsed by the observed reducedbinding of enzymatically acetylated CheY to the isolatedswitch complex (Fig. S5).
How does acetylation block CheY binding to itstarget proteins?
One of the first questions that comes to mind is howacetylation of lysine residues results in blocking CheYbinding to its target proteins. Co-crystallization of CheYwith its targets revealed that some lysine residues directlyparticipate in the binding of CheY to FliM (Lys-92, -119and -122) (Lee et al., 2001; Dyer et al., 2004; Dyer andDahlquist, 2006), to CheZ (Lys-109 and -119) (Zhao et al.,2002; Guhaniyogi et al., 2006; 2008), and to CheA (Lys-92, -122 and -126) (Swanson et al., 1995; Welch et al.,1998) (Protein Data Bank codes 3CHY, 1U8T, 1JBE,1KM1, 2FMI, 2PL9 and 1A0O). Therefore, the moststraightforward reason for the repression of binding byacetylation may be the covalent modification of residuesthat are directly involved in the binding. Neutralization ofthe positive charge of each acetylated lysine residue mayalso change the electrostatic interactions between CheYand its target proteins (McEvoy et al., 1999; Guhaniyogiet al., 2006; 2008). In addition, Lys-109 is known to forma salt bridge with the phosphorylation site, Asp-57 (Lukatet al., 1991). Even though it is not the main acetylationsite, its acetylation may lead to a conformational changeof the protein due to the missing salt bridge, and this
Fig. 5. Autophosphorylation of acetylated His-CheY with acetylphosphate as the donor. Acetylation was achieved by a 1:1 molarratio of acetic anhydride : free amines of CheY. Non-acetylatedHis-CheY served as a control.A. Native gel of non-acetylated and chemically acetylatedHis-tagged CheY used for this experiment.B. The same His-CheY samples on 12% SDS gel, following 30 sincubation with acetyl [32P]phosphate and stopping the reaction withSDS-sample buffer. Top: the SDS gel stained with Coomassie blue.Bottom: A phosphoimager scan of the SDS gel. NA stands fornon-acetylated.
Acetylation represses the binding of CheY 937
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 76, 932–943

change may interfere with the binding of CheY to its targetproteins. Acetylation of all other target sites is notexpected to affect the conformation of the proteinbecause they are all on the surface of the protein,exposed to the medium, and do not participate in otherintramolecular interactions.
What is the function of N-Lys acetylation of CheY inchemotaxis? – A model
By employing a genetic approach that impaired CheYacetylation by either Acs deletion (Barak and Eisenbach,2001) or lowering the AcCoA pool in the cell (Barak et al.,2006), CheY acetylation was demonstrated to beinvolved in bacterial chemotaxis. A number of functionswere proposed for this involvement, including a tuningmechanism that compensates for cell-to-cell variations inthe intracellular concentrations of CheA and CheZ (Barakand Eisenbach, 2004), linking the chemotaxis system tothe metabolic state of the cell (Barak and Eisenbach,2001; 2004), and direct involvement in the chemotacticresponse to repellents (Barak and Eisenbach, 2001;Barak et al., 2006). While these suggested functions maywell be correct, none of them considers reduced bindingby acetylation. What is the function of the acetylation-dependent reduction in binding? A conceivable possibility(shown schematically in the model in Fig. 6 for the inter-action with FliM only) is that the binding of CheY to itstarget proteins is controlled at two levels: fast, positiveregulation by phosphorylation according to environmen-tal signals (e.g. chemotactic and thermotactic stimuli),and slow, negative regulation by acetylation according to
the metabolic state of the cell. According to this hypoth-esis, acetylated CheY is essentially sequestered fromsignalling: it cannot bind to CheA at the receptorsupramolecular complex (and, consequently, it is prob-ably free in the cytoplasm), and it can therefore neitherbe phosphorylated by CheA nor bind to FliM and CheZ.According to this hypothesis, the magnitude of the frac-tion of CheY that is not acetylated and therefore free forsignalling depends on the metabolic state of the cell. Theobservations (i) that the intracellular concentrations ofthe precursors of the acetylation process (acetate andAcCoA) and of the inducible enzyme (Acs), which is con-sidered to be one of the acetylating enzymes of CheY(Barak et al., 1992; 2004; Ramakrishnan et al., 1998),are all dependent on the metabolic state of the cell(Prüss and Wolfe, 1994; Shin et al., 1997; Wolfe, 2005),(ii) that the level of this acetylation in vivo depends onthe growth phase (Yan et al., 2008; Yu et al., 2008), and(iii) that lysine acetylation is a dynamic and regulatedprocess in Escherichia coli (Zhang et al., 2009) – are allconsistent with the hypothesis. In other words, acetyla-tion determines at each metabolic state the fraction ofCheY molecules that can participate in signalling, andphosphorylation regulates the binding of acetyl-freeCheY molecules to FliM (and CheZ). This model demon-strates for CheY the advantage of a dual covalent modi-fication over a single modification, taking advantage ofthe difference in the stability of these modifications –stable acetylation (Barak et al., 2004; 2006) versusunstable phosphorylation (Hess et al., 1988a,b). Therecent observations that many proteins in E. coli arelysine acetylated in vivo (Yan et al., 2008; Yu et al., 2008;Zhang et al., 2009) make the question of regulation byacetylation of general interest.
A covalent modification involved in signal transductionshould be regulatable, meaning that there must be amechanism to deacetylate CheY, not only acetylate it. Theidentity of the protein(s), if any, that deacetylate CheY isnot yet known. One candidate is the acetylating enzymeitself (Acs), which mediates reversible acetylation of CheY(Barak et al., 2004). However, it appears that the deacety-lating power of this enzyme alone cannot account for thepresumed deacetylating activity in the cell becausedeacetylation of CheY by Acs is incomplete and very slow(Barak et al., 2004; 2006). Another candidate is thedeacetylase CobB – a bacterial nicotinamide adeninedinucleotide (NAD+)-dependent deacetylase that, at leastin Salmonella, contributes to the activation of Acs bydeacetylating this enzyme (Starai et al., 2002; 2003;2004). Interestingly, Lys-609, which undergoes acetyla-tion by protein acetyltransferase in Salmonella (Starai andEscalante-Semerena, 2004), is conserved in E. coli Acs.Two scenarios seem possible: (i) CobB regulates theactivity of Acs in E. coli and thus indirectly regulates the
Fig. 6. Suggested role for the dual covalent modification of CheY.According to the model, the metabolic state of the cell determinesthe fraction of CheY molecules that are acetylated. Onlynon-acetylated CheY molecules can bind to CheA and bephosphorylated by it, and only they can bind to FliM with aresultant change in the direction of flagellar rotation. Thus, whileacetylation, which is the slower process of these two covalentmodifications, determines the fraction of CheY molecules that canparticipate in chemotactic signalling, phosphorylation, whose levelis regulated by chemotactic and thermotactic stimuli, determinesthe extent of CheY binding to FliM. Although a CheY molecule canbe both phosphorylated by AcP and acetylated at the same time(Fig. 5), this situation is not presented in the model because themajor phosphorylating agent in vivo is CheA, and not AcP(Mayover et al., 1999).
938 O. Liarzi et al. �
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 76, 932–943

acetylation level of CheY, and (ii) CobB deacetylatesCheY directly. Future studies should identify the enzymethat deacetylates CheY and the signal for deacetylation.They should also resolve the apparent inconsistencybetween the observed acetylation-dependent reduction ofCheY binding to FliM, on the one hand, and the evidence,obtained with cytoplasm-free envelopes and intact guttedcells expressing CheY (Wolfe et al., 1988; Barak et al.,1992; 1998; Ramakrishnan et al., 1998), suggesting thatCheY acetylation promotes clockwise rotation, on theother hand.
Experimental procedures
Chemicals
[32P]Orthophosphate was obtained from Amersham (Bucking-hamshire, UK); ampicillin, chloramphenicol, carbenicillin,CelLytic-B™, and acetyl phosphate from Sigma (Rehovot,Israel); acetic anhydride from Frutarom (Haifa, Israel); nickel-nitrilotriacetic acid (Ni-NTA) agarose from Qiagen (Hilden,Germany); and GelCode® blue stain reagent from ThermoScientific (Rockford, IL, USA). All the compounds were of thehighest purity available.
Bacterial strains and growth conditions
The strains and plasmids used in this study are listed inTable 1. All bacterial cultures were grown aerobically withshaking (250 rotations per min) at 35°C (or 30°C for FliM
purification) either in Luria broth (LB, for the purification ofHis-CheY and His-FliM) or in Casamino medium [M-9minimal medium (Miller, 1992), 0.5% (w/v) Casamino acids,and 5 mg ml-1 thiamine, for the purification of non-taggedCheY, CheZ and CheA].
Protein purification
The procedures for the purification of non-tagged CheY(Barak and Eisenbach, 1992) and CheZ (Blat and Eisenbach,1994) were described earlier as indicated. His-tagged CheYwas purified basically as described (Sagi et al., 2003), exceptthat the protein was purified twice on the Ni2+ column. CheAwas purified as described (Hess et al., 1988b), with somemodifications. The CheA-containing fractions from the Ciba-cron column were pooled, concentrated by ultrafiltrationthrough a 10 kDa cut-off membrane, and then loaded onto aSephadex G-100 column. The eluted protein was concen-trated by ultrafiltration, and dialysed against PBS containingsurfactant P20 (0.005%). The Barstar protein was a gift fromG. Schreiber, Israel. The coding sequence for Barstar fromBacillus amyloliquefaciens, fused to the coding sequence ofcel5A cellulase from Thermobifida fusca, was cloned into thegeneral expression vector pET-28a (Novagen) N-terminal toa 10¥His tag. The protein was expressed in BL21 (DE3)(Novagen) bacterial cells grown at 35°C to OD590 = 0.7,induced with 1 mM IPTG for 3 h, and harvested. The proteinwas purified from the supernatant by Ni-NTA affinity chroma-tography (Qiagen) as described for the production of 6¥His-CheY, with some modifications. The protein was loaded onceonto the column and the elution was carried out by a stepgradient of increasing imidazole (up to 500 mM). Peak frac-
Table 1. List of strains and plasmids used in this study.
Strain/plasmid Relevant genotype/phenotype Reference and/or source
StrainsRP437 Wild type for chemotaxis; his thr leu metE thi eda rpsL Parkinson and Houts (1982)BL21 (DE3) HsdS gal ompT rB
-mB- [DC1857(Ts)indl Sam7 min 5 lacUV5-T7 gene 1] Studier et al. (1990); Novagen
BL21 (DE3) + pET-28a HsdS gal ompT rB-mB
- [DC1857(Ts)indl Sam7 min 5 lacUV5-T7 gene 1] KanR G. SchreiberEW126 RP437 + pRL22DPvuII; AmpR Barak et al. (2004)M15 [pREP4] LacZ StrRKanR Zamenhof and Villarejo (1972)M15 [pREP4] + pQE12-
His-tag-CheYLacZ StrRKanR Sagi et al. (2003)
RP5232 RP437D(CheY)6021 J.S. ParkinsonEW237 RP5232 + pBtac/wild-type CheY This studyEW238 RP5232 + pBtac/CheY-K92R/K122T This studyEW239 RP5232 + pBtac/CheY-K92R/K109R This studyEW86 BL21(DE3) + pEWM1; AmpRCapR Bren and Eisenbach (1998)ST1 + pDV4 PenRAmpR R.M. MacnabRP437 + pRL22 StrepRPenR Blat and Eisenbach (1994)
PlasmidspRL22DPvuII CheY overproduction P. MatsumurapQE12-CheY-His-tag 6¥His-CheY overproduction A. WolfepEWM1 6¥His-FliM overproduction Bren and Eisenbach (1998)pDV4 CheA and CheW overproduction Hess et al. (1987)pRL22 CheY and CheZ overproduction Matsumura et al. (1984)pET-28a 10¥His-BarStar-cel5A overproduction G. SchreiberpBtac/wild-type CheY CheY (wild type) overproduction This studypBtac/CheY-K92R/K122T CheY-K92R/K122T overproduction This studypBtac/CheY-K92R/K109R CheY-K92R/K109R overproduction Barak et al. (2004)
Acetylation represses the binding of CheY 939
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 76, 932–943

tions were pooled, extensively dialysed against PBS buffer,and kept at -80°C.
Protein modifications
Chemically acetylated CheY was prepared as describedearlier (Yan et al., 2008). Briefly, CheY, chemically acetylatedto various levels, was prepared by 30 min incubation of puri-fied CheY at room temperature with acetic anhydride(0.2–1.0:1 ratio of acetic anhydride : free amines of CheY).The reaction was carried out under basic pH conditions(pH 9.0) using 2.5% Na2CO3, conditions that allowN-acetylation but not O-acetylation (Riordan and Vallee,1967). The reaction was terminated by dialysis. The level ofacetylation of CheY was estimated by loading CheY aliquotson a native gel (12.5% or 15% polyacrylamide), staining thegel by Coomassie blue, destaining, cutting the multiple bandsof acetylated CheY from the gel, and determining theirdegrees of acetylation by MALDI-TOF MS.
Phosphorylation of CheY by acetyl phosphate was per-formed according to Lukat et al. (1992) by incubating CheY(3–70 mM) in PBS buffer containing surfactant P20 (0.005%)for 2 min at room temperature with 9 mM acetyl phosphate inthe presence of 10 mM MgCl2.
Phosphorylation of CheY (67 mM) by acetyl [32P]phosphate[200–400 cpm pmole-1; synthesized as described byMcCleary and Stock (1994)] was performed by 30 s incuba-tion at room temperature. Both non-acetylated CheY andCheY pre-acetylated to a level that does not contain non-acetylated molecules (achieved by a 1:1 ratio of acetic anhy-dride : free amines of CheY) were used (one at a time). Thereaction was stopped by the addition of SDS sample buffer.The proteins were subjected to SDS-polyacrylamide gel elec-trophoresis (12.5% polyacrylamide) and stained with Coo-massie blue, destained, dried, and the relative radioactivity ofCheY was measured with a phosphoimager (Fujix).
Measurement of CheY acetylation level in vivo
The measurement of the acetylation level of CheY in vivowas carried out essentially as described by Yan et al. (2008),except that the overnight LB cultures of strains EW237,EW238, EW239 and RP5232 were diluted 20-fold with LBand continued to grow until OD590 = 2.0, harvested, washedtwice with Tris-HCl (50 mM, pH 7.0) and rapidly frozen inliquid nitrogen. Cell-free extracts were loaded on two equalgels (12% polyacrylamide) and were exposed to SDS-gelelectrophoresis. The proteins of one gel were transferred toa nitrocellulose membrane as previously described (Brenand Eisenbach, 2001) and the acetylation level on CheY wasdetected by Western blotting with Immunoglobulin G (IgG)-enriched serum containing anti acetyl-lysine antibody(1:1500, v/v) (Yan et al., 2008), followed by peroxidase-conjugated goat-anti rabbit (1:40 000, v/v) antibody. Theresults were analysed by a densitometer (Bio-Rad). Theother gel was stained with GelCode® blue stain reagent, anddried.
Real-time analysis interactions
All real-time binding interactions were measured by SPR,using the ProteOn XPR36™ Protein Interaction Array System
(Bio-Rad, Haifa, Israel). Since the ProteOn chip contains sixhorizontal and six vertical channels, it allowed us to simulta-neously measure different acetylation levels of immobilizedHis-CheY with six different analyte concentrations. Prior toeach experiment, we assessed the composition of eachacetylated sample by native polyacrylamide gel electrophore-sis and then we immobilized the differently acetylatedsamples onto a His-tag-binding chip – a ProteOn NLC (Bio-Rad) Sensor Chip (a modified alginate matrix chip pre-coatedwith NeutrAvidin) that was pre-coated with 200 nM biotin-conjugated tris-nitrilotriacetic acid (Reichel et al., 2007)(200 RU; a gift from G. Schreiber, Israel).
For the first set of binding, i.e. the binding of CheA andCheZ to His-CheY, the latter, acetylated to the indicatedlevels, was immobilized onto a sensor chip surface. Immobi-lization was achieved by injecting (at a rate of 30 ml min-1)5 mM His-CheY for its binding to CheA, 30–35 mM for bindingto CheZ under phosphorylating conditions, and 3–7 mM forbinding to CheZ under non-phosphorylating conditions. About500–650 RU (CheA), 800–1000 RU (CheZ phosphorylatingconditions) and 800–970 RU (CheZ non-phosphorylatingconditions) of His-CheY were typically immobilized underthese conditions. The analytes were diluted in PBS buffercontaining surfactant P20 (0.005%) and 10 mM MgCl2, andflowed at a rate of 50 ml min-1. For phosphorylating conditionsacetyl phosphate (9 mM) was supplemented. Regenerationof the surface was carried out with NaOH (50 mM) at50 ml min-1 for 1 min. The analyte concentrations ranged from29 to 460 nM for CheA, and 0.2 to 3.6 mM and 1 to 30 mM forCheZ under phosphorylating and non-phosphorylating condi-tions respectively. The effect of acetylation was examinedusing a fixed concentration of the analyte as indicated in thelegend to Fig. 4. In the experiments with CheA and CheZunder non-phosphorylating conditions, the negative controlfor non-specific binding was His-tagged FliM, which does notinteract with CheA or CheZ (Bren and Eisenbach, 2000). Inthe experiments with CheZ under phosphorylating conditions,the negative control was the irrelevant ligand His-taggedBarstar (an RNase inhibitor) fused to cel5A (a cellulase). Thekinetic parameters were calculated by ProteOn Manager(Bio-Rad) software package.
For the second set of binding, i.e. the binding of CheY toHis-FliM, the latter (5 mM) was immobilized to the chip(binding signal 3900 RU), as described above. CheY at theindicated acetylation levels was diluted to a constant concen-tration (70 mM) in the same buffer as above. The negativecontrol for non-specific binding was an irrelevant ligand(Barstar fused to cel5A). No regeneration step was neededbetween the injections. All other conditions were as describedabove.
Mass spectrometry
Electroelution was performed in GeBAflex tube (Gene BioApplication, Israel) at 150 V for 2 h as described (Montignyet al., 2004). Protein bands were excised from the SDS gel,stained with GelCode, and in-gel digested with bovine trypsin(sequencing grade, Roche Diagnostics, Germany) andEndoproteinase GluC (V8), as described (Shevchenko et al.,1996). The resulting peptide mixtures were reconstituted in80% formic acid and immediately diluted 1:10 with Milli-Q
940 O. Liarzi et al. �
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 76, 932–943

water prior to the analysis by online reversed-phase nano-LC-ESI-MS/MS.
Intact molecular mass measurements were performed on aBruker ReflexIII™ MALDI-TOF mass spectrometer (Bruker,Bremen, Germany) equipped with delayed ion extraction,reflector and a 337 nm nitrogen laser. Each mass spectrumwas generated from accumulated data of 200 laser shots.External calibration for proteins was achieved by using BSAand myoglobin proteins, obtained from Sigma.
Precursor ion scan at m/z 126.1 and 143.1 for the detectionof peptides incorporated acetylated Lysine amino acidresidue specific was performed on Q-STAR PulsariElectrospray-Quadrupole TOF (ABI, Sciex, Toronto,Canada), as described (Barak et al., 2004).
For the identification of acetylated lysine residues peptidemixtures were separated by online reversed-phase nanos-cale capillary LC online with LTQ-OrbitrapXL hybrid ion trapand FT mass spectrometer (Thermo Fisher Scientific,Bremen, Germany) operated in the positive ion mode.
For the nano-LC-ESI-MS/MS, the samples were injectedonto an in-house-made 15 cm reversed-phase sprayingfused-silica capillary column (inner diameter 75 mm, packedwith 3 mm ReproSil-Pur C18A18 media (Dr Maisch GmbH,Ammerbuch-Entringen, Germany), using an UltiMate 3000Capillary/Nano LC System, consisting of Famos™ MicroAutosampler, Switchos™ Micro Column Switching Module(LC Packings, Dionex). The LC set-up was connected to theLTQ Orbitrap mass spectrometer (Thermo Fisher Scientific,Bremen, Germany) equipped with a nano-electrospray ionsource (Thermo Fisher Scientific, Bremen, Germany). Theflow rate through the column was 250 nl min-1. An acetonitrilegradient was employed with a mobile phase containing 0.1%and 0.2% formic acid in Milli-Q water in buffers A and Brespectively. The injection volume was 5 ml. The peptideswere separated with 50 min gradients from 5% to 65%CH3CN. In the nano-electrospray ionization source, the endof the capillary from the nano-LC column was connected tothe emitter with pico-tip silica tubing, i.d. 20 mm (New Objec-tive) by stainless steel union, with a PEEK sleeve for cou-pling the nano-spray with the online nano-LC. The voltageapplied to the union in order to produce an electrospray was2.4 kV. Helium was introduced as a collision gas at a pres-sure of 3 psi. The mass spectrometer was operated in thedata-dependent mode. Survey MS scans were acquired inthe Orbitrap with the resolution set to a value of 60 000. Upto the seven most intense ions per scan were fragmentedand analysed in the linear trap. For the analysis of trypticpeptides, survey scans were recorded in the FT mode fol-lowed by data-dependent collision-induced dissociation(CID) of the seven most intense ions in the linear ion trap(LTQ). Raw spectra were processed using open-source soft-ware DTASuperCharge (http://msquant.sourceforge.net).The data were searched with MASCOT (Matrix Science,London, UK) and Proteome Discoverer against a Swissprotor NCBI database. Search parameters included variablemodifications of 57.02146 Da (carboxyamidomethylation) onCys, 15.99491 Da (oxidation) on Met and 0.984016 Da (dea-midation) on Asn and Gln and 42.0367 (acetylation) on Lys.The search parameters were as follows: maximum twomissed cleavages, initial precursor ion mass tolerance10 p.p.m., fragment ion mass tolerance 0.6 Da. The identity
of the peptides and the assignment of acetylated lysine resi-dues were concluded from the detected collision-induceddissociation products by Mascot and Proteome Discoverersoftware followed by manual inspection of the fragmentationseries.
Acknowledgements
We thank M. Wilchek for advice and assistance in chemicallyacetylating CheY, L. Armon for statistical analyses and G.Schreiber for the Barstar protein. M.E. is an incumbent of theJack and Simon Djanogly Professorial Chair in Biochemistry.This study was supported by Grant No. 290/06 from the IsraelScience Foundation.
References
Baker, M.D., Wolanin, P.M., and Stock, J.B. (2006) Signaltransduction in bacterial chemotaxis. Bioessays 28: 9–22.
Barak, R., and Eisenbach, M. (1992) Correlation betweenphosphorylation of the chemotaxis protein CheY and itsactivity at the flagellar motor. Biochemistry 31: 1821–1826.
Barak, R., and Eisenbach, M. (2001) Acetylation of theresponse regulator, CheY, is involved in bacterialchemotaxis. Mol Microbiol 40: 731–743.
Barak, R., and Eisenbach, M. (2004) Co-regulation of acety-lation and phosphorylation of CheY, a response regulator inchemotaxis of Escherichia coli. J Mol Biol 342: 375–381.
Barak, R., Welch, M., Yanovsky, A., Oosawa, K., and Eisen-bach, M. (1992) Acetyladenylate or its derivative acetylatesthe chemotaxis protein CheY in vitro and increases itsactivity at the flagellar switch. Biochemistry 31: 10099–10107.
Barak, R., Abouhamad, W.N., and Eisenbach, M. (1998) Bothacetate kinase and acetyl Coenzyme A synthetase areinvolved in acetate-stimulated change in the direction offlagellar rotation in Escherichia coli. J Bacteriol 180: 985–988.
Barak, R., Prasad, K., Shainskaya, A., Wolfe, A.J., and Eisen-bach, M. (2004) Acetylation of the chemotaxis responseregulator CheY by acetyl-CoA synthetase from Escherichiacoli. J Mol Biol 342: 383–401.
Barak, R., Yan, J., Shainskaya, A., and Eisenbach, M. (2006)The chemotaxis response regulator CheY can catalyze itsown acetylation. J Mol Biol 359: 251–265.
Blat, Y., and Eisenbach, M. (1994) Phosphorylation-dependent binding of the chemotaxis signal moleculeCheY to its phosphatase, CheZ. Biochemistry 33: 902–906.
Bren, A., and Eisenbach, M. (1998) The N terminus of theflagellar switch protein, FliM, is the binding domain for thechemotactic response regulator, CheY. J Mol Biol 278:507–514.
Bren, A., and Eisenbach, M. (2000) How signals are heardduring bacterial chemotaxis: protein–protein interactions insensory signal propagation. J Bacteriol 182: 6865–6873.
Bren, A., and Eisenbach, M. (2001) Changing the direction offlagellar rotation in bacteria by modulating the ratiobetween the rotational states of the switch protein FliM.J Mol Biol 312: 699–709.
Dyer, C.M., and Dahlquist, F.W. (2006) Switched or not?: the
Acetylation represses the binding of CheY 941
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 76, 932–943

structure of unphosphorylated CheY bound to the N termi-nus of FliM. J Bacteriol 188: 7354–7363.
Dyer, C.M., Quillin, M.L., Campos, A., Lu, J., McEvoy, M.M.,Hausrath, A.C., et al. (2004) Structure of the constitutivelyactive double mutant CheYD13K Y106W alone and in complexwith a FliM peptide. J Mol Biol 342: 1325–1335.
Eisenbach, M. (2004) Bacterial chemotaxis. In Chemotaxis.Eisenbach, M. (ed.). London: Imperial College Press, pp.53–215.
Guhaniyogi, J., Robinson, V.L., and Stock, A.M. (2006)Crystal structures of beryllium fluoride-free and berylliumfluoride-bound CheY in complex with the conservedC-terminal peptide of CheZ reveal dual binding modesspecific to CheY conformation. J Mol Biol 359: 624–645.
Guhaniyogi, J., Wu, T., Patel, S.S., and Stock, A.M. (2008)Interaction of CheY with the C-terminal peptide of CheZ.J Bacteriol 190: 1419–1428.
Hames, B.D. (1981) An introduction to polyacrilamide gelelectrophoresis. In Gel Electrophoresis of Proteins: A Prac-tical Approach. Hames, B.D., and Rickwood, D. (eds).Washington, DC: IRL Press Oxford, pp. 23–64.
Hess, J.F., Oosawa, K., Matsumura, P., and Simon, M.I.(1987) Protein phosphorylation is involved in bacterialchemotaxis. Proc Natl Acad Sci USA 84: 7609–7613.
Hess, J.F., Bourret, R.B., Oosawa, K., Matsumura, P., andSimon, M.I. (1988a) Protein phosphorylation and bacterialchemotaxis. Cold Spring Harb Symp Quant Biol 53:41–48.
Hess, J.F., Oosawa, K., Kaplan, N., and Simon, M.I. (1988b)Phosphorylation of three proteins in the signaling pathwayof bacterial chemotaxis. Cell 53: 79–87.
Lee, S.-Y., Cho, H.S., Pelton, J.G., Yan, D.L., Henderson,R.K., King, D.S., et al. (2001) Crystal structure of an acti-vated response regulator bound to its target. Nat Struct Biol8: 52–56.
Lukat, G.S., Lee, B.H., Mottonen, J.M., Stock, A., and Stock,J.B. (1991) Roles of the highly conserved aspartate andlysine residues in the response regulator of bacterialchemotaxis. J Biol Chem 266: 8348–8354.
Lukat, G.S., McCleary, W.R., Stock, A.M., and Stock, J.B.(1992) Phosphorylation of bacterial response regulatorproteins by low molecular weight phospho-donors. ProcNatl Acad Sci USA 89: 718–722.
McCleary, W.R., and Stock, J.B. (1994) Acetyl phosphateand the activation of two component response regulators.J Biol Chem 269: 31567–31572.
McEvoy, M., Bren, A., Eisenbach, M., and Dahlquist, F.W.(1999) Identification of the binding interfaces on CheY fortwo of its targets, the phosphatase CheZ and the flagellarswitch protein FliM. J Mol Biol 289: 1423–1433.
Matsumura, P., Rydel, J.J., Linzmeier, R., and Vacante, D.(1984) Overexpression and sequence of the Escherichiacoli cheY gene and biochemical activities of the CheYprotein. J Bacteriol 160: 36–41.
Mayover, T.L., Halkides, C.J., and Stewart, R.C. (1999)Kinetic characterization of CheY phosphorylation reac-tions: comparison of P-CheA and small-moleculephosphodonors. Biochemistry 38: 2259–2271.
Miller, J.H. (1992) A Short Course in Bacterial Genetics. ColdSpring Harbor, NY: Cold Spring Harbor Laboratory.
Montigny, C., Jaxel, C., Shainskaya, A., Vinh, J., Labas, V.,Moller, J.V., et al. (2004) Fe2+-catalyzed oxidative cleav-ages of Ca2+-ATPase reveal novel features of its pumpingmechanism. J Biol Chem 279: 43971–43981.
Parkinson, J.S., and Houts, S.E. (1982) Isolation and behav-ior of Escherichia coli deletion mutants lacking chemotaxisfunctions. J Bacteriol 151: 106–113.
Prüss, B.M., and Wolfe, A.J. (1994) Regulation of acetylphosphate synthesis and degradation, and the control offlagellar expression in Escherichia coli. Mol Microbiol 12:973–984.
Ramakrishnan, R., Schuster, M., and Bourret, R.B. (1998)Acetylation at Lys-92 enhances signaling by the chemot-axis response regulator protein CheY. Proc Natl Acad SciUSA 95: 4918–4923.
Reichel, A., Schaible, D., Al Furoukh, N., Cohen, M.,Schreiber, G., and Piehler, J. (2007) Noncovalent, site-specific biotinylation of histidine-tagged proteins. AnalChem 79: 8590–8600.
Riordan, J.F., and Vallee, B.L. (1967) Acetylation. MethodsEnzymol 11: 565–570.
Sagi, Y., Khan, S., and Eisenbach, M. (2003) Binding of thechemotaxis response regulator CheY to the isolated, intactswitch complex of the bacterial flagellar motor: lack ofcooperativity. J Biol Chem 278: 25867–25871.
Schuster, S.C., Swanson, R.V., Alex, L.A., Bourret, R.B., andSimon, M.I. (1993) Assembly and function of a quaternarysignal transduction complex monitored by surface plasmonresonance. Nature 365: 343–347.
Shevchenko, A., Wilm, M., Vorm, O., and Mann, M. (1996)MALDI Quadrupole time-of-flight mass spectrometry: apowerful tool for proteomic research. Anal Chem 68: 850–858.
Shin, S., Song, S.G., Lee, D.S., Pan, J.G., and Park, C.(1997) Involvement of iclR and rpoS in the induction of acs,the gene for acetyl coenzyme A synthetase of Escherichiacoli K-12. FEMS Microbiol Lett 146: 103–108.
Shukla, D., and Matsumura, P. (1995) Mutations leading toaltered CheA binding cluster on a face of CheY. J BiolChem 270: 24414–24419.
Shukla, D., Zhu, X.Y., and Matsumura, P. (1998) Flagellarmotor-switch binding face of CheY and the biochemicalbasis of suppression by CheY mutants that compensate formotor-switch defects in Escherichia coli. J Biol Chem 273:23993–23999.
Starai, V.J., and Escalante-Semerena, J.C. (2004) Identifica-tion of the protein acetyltransferase (Pat) enzyme thatacetylates acetyl-CoA synthetase in Salmonella enterica.J Mol Biol 340: 1005–1012.
Starai, V.J., Celic, I., Cole, R.N., Boeke, J.D., and Escalante-Semerena, J.C. (2002) Sir2-dependent activation of acetyl-CoA synthetase by deacetylation of active lysine. Science298: 2390–2392.
Starai, V.J., Takahashi, H., Boeke, J.D., and Escalante-Semerena, J.C. (2003) Short chain fatty acid activation byacyl-coenzyme A synthetases requires SIR2 protein func-tion in Salmonella enterica and Saccharomyces cerevisiae.Genetics 163: 545–555.
Starai, V.J., Takahashi, H., Boeke, J.D., and Escalante-Semerena, J.C. (2004) A link between transcription andintermediary metabolism: a role for Sir2 in the control of
942 O. Liarzi et al. �
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 76, 932–943

acetyl-coenzyme A synthetase. Curr Opin Microbiol 7: 115–119.
Studier, F.W., Rosenberg, A.H., Dunn, J.J., and Dubendorff,J.W. (1990) Use of T7 RNA polymerase to direct expres-sion of cloned genes. Methods Enzymol 185: 60–89.
Swanson, R.V., Lowry, D.F., Matsumura, P., McEvoy, M.M.,Simon, M.I., and Dahlquist, F.W. (1995) Localized pertur-bations in CheY structure monitored by NMR identify aCheA binding interface. Nat Struct Biol 2: 906–910.
Wadhams, G.H., and Armitage, J.P. (2004) Making sense of itall: bacterial chemotaxis. Nat Rev Mol Cell Biol 5: 1024–1037.
Welch, M., Oosawa, K., Aizawa, S.-I., and Eisenbach, M.(1993) Phosphorylation dependent binding of a signal mol-ecule to the flagellar switch of bacteria. Proc Natl Acad SciUSA 90: 8787–8791.
Welch, M., Chinardet, N., Mourey, L., Birck, C., and Samama,J.-P. (1998) Structure of the CheY-binding domain of histi-dine kinase CheA in complex with CheY. Nat Struct Biol 5:25–29.
Wolfe, A.J. (2005) The acetate switch. Microbiol Mol Biol Rev69: 12–50.
Wolfe, A.J., Conley, M.P., and Berg, H.C. (1988) Acetylad-enylate plays a role in controlling the direction of flagellarrotation. Proc Natl Acad Sci USA 85: 6711–6715.
Yan, J., Barak, R., Liarzi, O., Shainskaya, A., and Eisenbach,M. (2008) In vivo acetylation of CheY, a response regulatorin chemotaxis of Escherichia coli. J Mol Biol 376: 1260–1271.
Yu, B.J., Kim, J.A., Moon, J.H., Ryu, S.E., and Pan, J.G.(2008) The diversity of lysine acetylated proteins inEscherichia coli. J Microbiol Biotechnol 18: 1529–1536.
Zamenhof, P.J., and Villarejo, M. (1972) Constructionand properties of Escherichia coli strains exhibitingcomplementation of galactosidase fragments in vivo.J Bacteriol 110: 171–178.
Zhang, J., Sprung, R., Pei, J., Tan, X., Kim, S., Zhu, H., et al.(2009) Lysine acetylation is a highly abundant and evolu-tionarily conserved modification in Escherichia coli. MolCell Proteomics 8: 215–225.
Zhao, R., Collins, E.J., Bourret, R.B., and Silversmith, R.E.(2002) Structure and catalytic mechanism of the E. colichemotaxis phosphatase CheZ. Nat Struct Biol 9: 570–575.
Zhu, X.Y., Volz, K., and Matsumura, P. (1997) The CheZ-binding surface of CheY overlaps the CheA- and FliM-binding surfaces. J Biol Chem 272: 23758–23764.
Supporting information
Additional supporting information may be found in the onlineversion of this article.
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting materials suppliedby the authors. Any queries (other than missing material)should be directed to the corresponding author for thearticle.
Acetylation represses the binding of CheY 943
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 76, 932–943





























