Embed Size (px)
Citation preview

248
Biochimica et Biophysica Acta, 561 (1979) 248--260 © Elsevier/North-Holland Biomedical Press
BBA 99366
COMPARATIVE STUDIES ON HIGHLY METABOLICALLY ACTIVE HISTONE ACETYLATION
MICHAEL MOORE, VAUGHN JACKSON, LINDA SEALY and ROGER CHALKLEY *
Department of Biochemistry, School of Medicine, The University of Iowa, Iowa City, IA 52242 (U.S.A.)
(Received May 22nd, 1978)
Key words: Histone modification; Acetylation
Summary
Histone acetate is hydrolyzed rapidly in logarithmically dividing hepatoma tissue culture cells (Jackson, V., Shires, A., Chalkley, R. and Granner, D.K. (1975) J. Biol. Chem. 250, 4856--4863). The phenomenon has been analyzed further in hepatoma tissue culture cells at various stages of the cell cycle, in stationary phase, and in the presence of actinomycin D. We also investigated the phenomenon in Tetrahymena pyriformis macronuclei, bovine thymocytes, and human foreskin fibroblasts. The data suggest that this highly metabolically active histone acetylation while altered in mitotic cells, is independent of the overall rate of cell division, and is only slightly sensitive to actinomycin D. Finally, we conclude that the same general phenomenon is found in both cancerous and normal cells and is apparently common to cells from various stages of the evolutionary scale.
Introduction
Histones can be modified by methylation, phosphorylation and acetylation. The latter two modifications produce a change in the overall charge on the histone molecule and as such have been of great interest as possible devices for modulating histone-DNA interactions. Phosphorylation of histones is par- ticularly active during cell multiplication [1--4]. On the other hand acetylation seems to be continuously active except in the quiescent germ line cells of certain organisms which contain sperm histones [5].
While all histone fraction are modified by acetylation [6--9] only histones H1, H2a and H4 possess a metabolically stable amino-terminal acetate group
* To whom correspondence should b e s e n t at t h e p r e s e n t address.

249
[10]. The four non-H1 histones are all modified at internal lysine residues [10], a modification which is metabolically very active [11]. Recent evidence in exponentially growing hepatoma tissue culture cells has indicated that more than 50% of this acetate is hydrolyzed from the histone with a tl/2 ~ 3 min, while most of the remaining acetate is removed with a t in ~ 30 min [11]. There appears to be a steady-state occurring between the acetylation reaction and its subsequent hydrolysis so that a significant amount of material exists in the modified form at any given time [12]. For example in hepatoma tissue cul- ture cells modification of histone H4 occurs primarily (30%) as the mono- acetylated form, 5--10% as the diacetylated and somewhat less than 2% in the form of higher levels of acetylation. Histone H3 is acetylated to a degree com- parable to histone H4, bu t histones H2a and H2b show only low levels of the modified species, presumably reflecting a lower overall rate of acetylation [13]. The extent to which a given histone is acetylated appears to depend upon the cell type and the stage of the cell cycle. D'Anna et al. [14] have shown that histone H4 acetylation levels are decreased in mitotic as opposed to interphase Chinese hamster CHO cells. Whether variation in the degree of modification is due to changes in rate of acetylation or rate of hydrolysis has not been defined.
Although the function of histone acetylation is not known, several groups have surmised that it may play a role in gene expression. Thus AUfrey and his colleagues [15] have proposed that diffuse chromatin contains more exten- sively acetylated histone and that an increase in histone acetylation follows hormonal or environmental stimuli which result in major phenotypic changes in the cell [16.17]. More recently it has been argued that DNAase I selectively releases acetylated histone from the nucleosome, at the same time selectively digesting active genetic material [18]. If histone acetylation does play a role in gene activation there is no indication whether is is a causative effect or if it is simply playing a permissive role.
In order to understand more about the function of histone acetylation we have investigated both acetylation and deacetylation, asking particularly whether the exceedingly rapid rate of deacetylat ion found in logarithmic hepatoma tissue culture cells is a characteristic of both dividing and non- dividing cells, whether it is found at all stages of the cell cycle, and whether other mammalian and non-mammalian cell types possess this type of rapid deacetylation. Our results indicate that it may be a general property of essenti- ally all cells.
Materials and Methods
Labeling o f hepatoma tissue culture cells in G1, S, and Mitosis. Hepatoma tissue culture cells at various stages of the cell cycle were prepared as described previously [1,19]. The cells were grown to stationary phase in Swim's $77 medium, and 5-ml aliquots were added to each of 51 prescription bott les of 100 cm 2 surface area. 40 ml of fresh medium containing 2.41 mM CaC12 was added and the cells were incubated at 37°C for 36 h. At this time, the medium was replaced with fresh medium containing 0.1 mM colcemid and 2.41 mM CaC12. 8 h later the mitotic cells were shaken off the surface of the flasks, pelleted, and resuspended at 300 000 cells/ml in 1200 ml. The cells were

250
greater than 95% mitotic. 3 h after resuspension, 600 ml of the cells were pelleted (300 Xg for 10 min) and at 3.5 h after resuspension added to 60 ml of fresh medium with [3H]acetate (25 mCi, 100 Ci/mol). At 10, 30, and 60 min, 20-ml samples were taken and the cells pelleted and frozen rapidly in solid CO2 acetone. The G1 histones were later isolated and analzyed as' previously described [ 11 ].
To prepare S phase cells, 10 h after resuspension the remaining 600 ml of cells were pelleted and resuspended in medium containing [3H]acetate as described above. This process takes approx. 30 min. At 10, 30, and 60 min after resuspension 20-ml samples were taken, cooled, pelleted, frozen and the S phase histones analyzed as above.
Mitotic cells were prepared as described above [1]. Histone acetylation turnover studies with mitotic cells were done following a 20 min pulse with [3H]acetate.
Pulse-chase studies with [3H]acetate in rapidly dividing and stationary hepatoma tissue culture cells were done as previously described [11] utilizing a 10 min pulse period. When act inomycin D was used, cells were preincubated in normal medium including 1 pg/ml act inomycin D for 1 h prior to the pulse and also during the chase period.
Human foreskin fibroblasts. Human foreskin fibroblasts were grown in roller bott les at 37°C in Eagle's minimal essential medium supplemented with 15% fetal calf serum and aureomycin (50 pg/ml). Pulse-chase experiments were done with actively-dividing rather than stationary cells. Each roller bott le of cells (about 4 • 107 cells) was pulsed with 3.6 mCi [3H]acetate in 10 ml of the same medium for 10 min at 37°C and at various time points the chase was s topped by adding ice-cold physiological saline, pH 7. Two roller bottles of cells were used for each time point. Cells were then harvested, centrifuged, frozen, and histones isolated and analyzed as described above for hepatoma tissue culture cell histones.
Bovine thymocytes. Bovine thymus tissue was obtained immediately after slaughter from a local slaughterhouse. It was transported on ice to the labora- tory within 45 min. The thymus (25 g) was minced with scissors and shaken with tweezers to release thymocytes which were incubated with [3H]acetate (25 mCi, 100 Ci/mol) for 10 min at 37°C in 12.5 ml of Swims $77 medium at a cell concentrat ion of 2 . 1 0 8 cells/ml. The reaction was s topped by adding 10 volumes of cold medium. One-seventh of the total volume was taken as the zero time sample and the cells were pelleted and frozen. The remaining cells were pelleted at 300 X g for 10 min and resuspended at a cell density of 5 • 106/ml in fresh medium lacking [3H]acetate; 70-ml samples were taken at each time point during the chase, pelleted and frozen. Histones were isolated and analyzed as above.
Tetrahymena pyriformis. T. pyriformis, syngen I, strain B, were grown in medium containing 2% proteose peptone (Difco), 0.2% dextrose, 0.1% yeast extract (Difco), and 0.002~o Fe2+-EDTA complex [20]. Cultures were aerated and maintained at 29°C. Cells were harvested in log phase (5 • 10s--6 • l 0 s cells/ml) by centrifugation at 370 X g for 5 min. Cells were resuspended in a defined, inorganic medium (47 mM NaC1, 1 mM MgSO4"7H20 in 5 m M potassium phosphate buffer, pH.7 [21]) at 1/20 the previous culture volume.
2 5 1
[3H]Acetate (40 mCi, 100 Ci/mol, New England Nuclear) was added. After a 10 min pulse period, the cells were pelleted by centrifugation at 370 × g for 2 min and placed on ice. They were washed once with ice-cold inorganic medium before resuspension in fresh inorganic medium at 29°C and normal cell densities (5 . l 0 s cells/ml) for the chase period. Cells were harvested at the desired times and frozen quickly. A pellet highly enriched in macronuclei was prepared by homogenizing the cells gently in a solution 1% Triton X-100, 0.25 M sucrose, 0.1 M MgC12, 0.01 M Tris, and 0.05 M NaHSO3, pH 6.5, as previously described [11]. Macronuclei were washed once and histones extracted in 0.2 M H2SO4, 2.5 M urea for 4 h. Insoluble material was removed by centrifugation at 1000 × g for 10 rain, and the supernatant was dialyzed against 95% ethanol at 4°C. Precipitated histone was collected, dried, and dissolved in 20% sucrose, 0.9 M acetic acid, 0.5 M ~-mercaptoethanol. Electro- phoresis was performed on 25-cm gels by the procedure of Panyim and Chalk- ley [22] except that 15% acrylamide, 8 M urea was used. Gels were cut, digested and counted as previously described [11].
Results
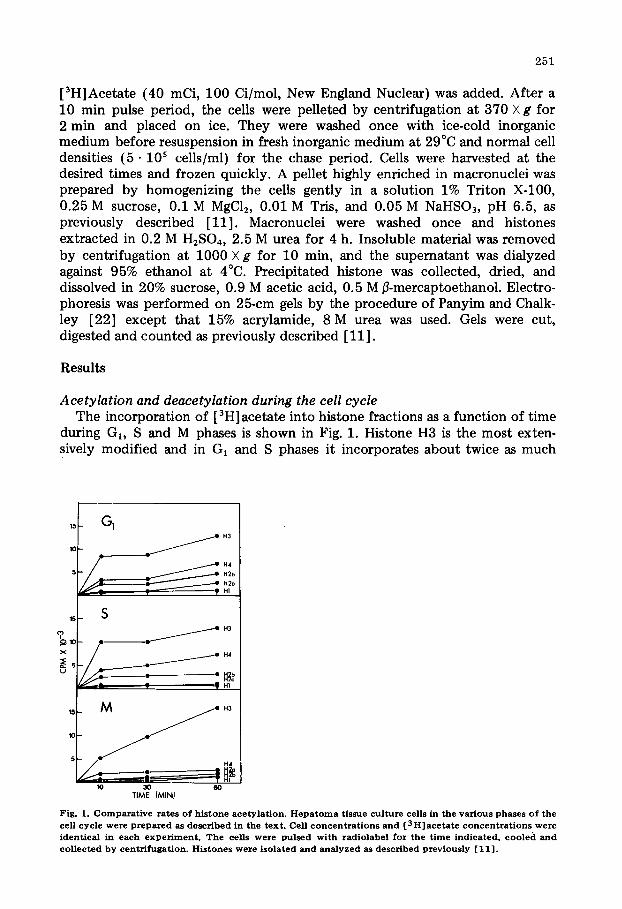
Acetylation and deacetylation during the cell cycle The incorporation of [ 3H] acetate into histone fractions as a function of t ime
during GI, S and M phases is shown in Fig. 1. Histone H3 is the most exten- sively modified and in Gi and S phases it incorporates about twice as much
@ /
H2b
H2a
= ? HI _ _ - T
u ~ - ; 9 .1
TIME (MIN)
Fig. 1. Compazat ive rates of histone acetylation. Hepatoma tissue culture cells in the various phases of the cel l cyc le were prepared as descr ibed in the t ex t . Cel l concentrations and [3H]acetate concentrations were ident ica l in each e x p e r i m e n t . Th e cells were pu lsed wi th radio label for the t ime ind ica ted , coo led an d c o l l e c t e d by centr i fugat ion . H i s t o n e s w e r e i so la ted and ana lyzed as descr ibed prev ious ly [ 1 1 ] .
252
I0~
6.(~
4.C
•'--°x I.C
u 0.(
0.,~
0.;
O]
TIME (MIN)
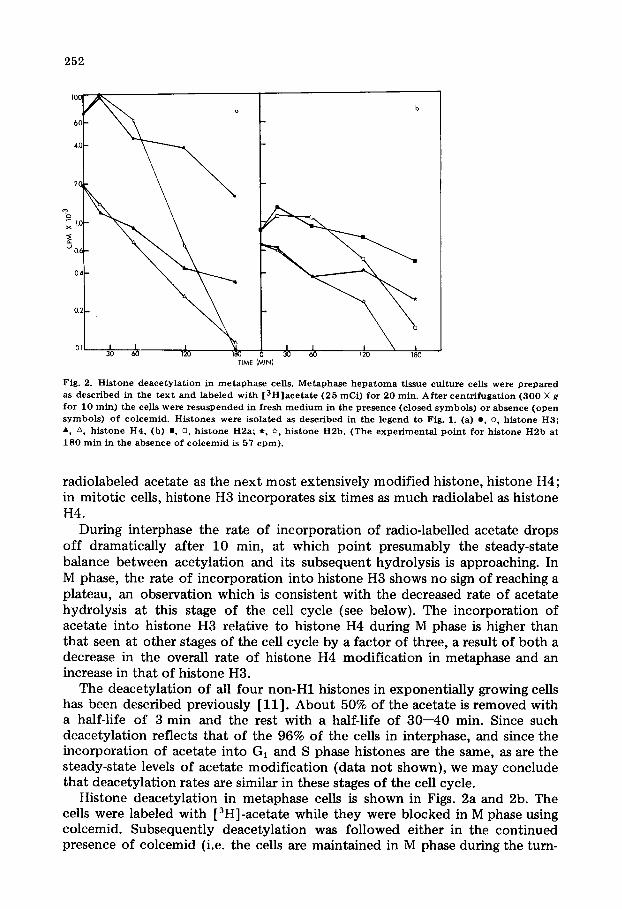
Fig. 2. H i s t o n e d e a c e t y l a t i o n in m e t a p h a s e cells. M e t a p h a s e h e p a t o m a t issue c u l t u r e cells were p r e p a r e d as d e s c r i b e d in t he t e x t a n d l abe l ed w i th [ 3 H ] a c e t a t e ( 2 5 mCi ) f o r 2 0 min . A f t e r c e n t r i f u g a t i o n ( 3 0 0 × g f o r 1 0 ra in ) t he cells we re r e s u s p e n d e d in f resh m e d i u m in t h e p r e s e n c e ( c l o sed s y m b o l s ) o r a b s e n c e ( o p e n s y m b o l s ) o f c o l c e m i d . H i s t o n e s were i s o l a t e d as d e s c r i b e d in the l e g e n d to Fig . 1. (a) e, o, h i s t o n e H3 ; A z~ h i s t o n e H 4 , (b) m • h i s t o n e H 2 a ; *, ~, h i s t o n e H 2 b . (The e x p e r i m e n t a l p o i n t fo r h i s t o n e H 2 b at 1 8 0 m i n in the a b s e n c e o f c o l c e m i d is 57 c p m ) .
radiolabeled acetate as the next most extensively modified histone, histone H4; in mitotic cells, histone H3 incorporates six times as much radiolabel as histone H4.
During interphase the rate of incorporation of radio-labelled acetate drops off dramatically after 10 min, at which point presumably the steady-state balance between acetylation and its subsequent hydrolysis is approaching. In M phase, the rate of incorporation into histone H3 shows no sign of reaching a plateau, an observation which is consistent with the decreased rate of acetate hydrolysis at this stage of the cell cycle (see below). The incorporation of acetate into histone H3 relative to histone H4 during M phase is higher than that seen at other stages of the cell cycle by a factor of three, a result of both a decrease in the overall rate of histone H4 modification in metaphase and an increase in that of histone H3.
The deacetylation of all four non-H1 histones in exponentially growing cells has been described previously [11]. About 50% of the acetate is removed with a half-life of 3 min and the rest with a half-life of 30--40 min. Since such deacetylation reflects that of the 96% of the cells in interphase, and since the incorporation of acetate into G1 and S phase histones are the same, as are the steady-state levels of acetate modification (data not shown}, we may conclude that deacetylation rates are similar in these stages of the cell cycle.
Histone deacetylation in metaphase cells is shown in Figs. 2a and 2b. The cells were labeled with [3H]-acetate while they were blocked in M phase using colcemid. Subsequently deacetylation was followed either in the continued presence of colcemid (i.e. the cells are maintained in M phase during the turn-

253
over study) or in the absence of colcemid in which case the metaphase~labeled cells are experiencing deacetylation as they enter G, phase. Cells labeled in metaphase and undergoing deacetylation in metaphase show a much reduced deacetylation rate compared to that seen in interphase. No indication was seen of the exceedingly rapid deacetylation (t1~2 ~ 3 min) observed in interphase cells. If acetate-labeled M phase cells are allowed to progress into G,, a remark- able difference in removal of acetate is seen. Acetate associated with histone H3 is now removed quite rapidly. The label associated with histone H4 and the other non-H1 histones is likewise released at a somewhat faster rate than that seen in metaphase.
Recently D'Anna et al. [14] reported that in metaphase cells the steady-state level of histone H4 acetate decreases markedly. We have made a similar observa- tion. Analysis of the decreased rate of histone H4 deacetylation and of the decreased acetylation do not provide an immediately obvious explanation for this behavior. Perhaps the most likely explanation lies in a decreased number of histone H4 molecules which can partake in the acetylation reaction, possibly due to the increased degree of packing of the nucleoprotein within the meta- phase chromosomes.
Acetylation as a function of cell growth The pattern of histone acetylation and deacetylation following a short pulse
of [3H]acetate had been established previously in this laboratory for logarithmic hepatoma tissue culture cells. It seemed appropriate to next inquire into the activity of these parameters in quiescent cells. Therefore we have compared the rates of acetylation and deacetylation in logarithmically dividing and in stationary phase cells, showing the deacetylation results in Fig. 3.
Although stationary cells took up somewhat more [3H]acetate into their cytoplasm during a 10 min pulse than did the logarithmic cells, the stationary cells incorporated slightly less (90%) [3H]acetate into each of the histones. Whether these observations reflect variation in permeability, pool size, or small acetylation rate changes is not known. As shown in Fig. 3, the deacetylation of histones H3 and H4 in the non-dividing cells is comparable to that observed previously in log phase cells, indicating that DNA synthesis is not an absolute requirement for the vigorous acetylation and deacetylation. In addition, in logarithmic and stationary cells all the various subfractions of histones H3 and H4 are deacetylated at the same rates. These data do not preclude more vigorous acetylation of histones at the t ime of histone and DNA synthesis as part of the depositional process and in fact there is good evidence that this occurs; however, the fraction of histone which is 'newly synthesized' is only a small component of the whole and one does not detect its absence in non- dividing cells.
Deacetylation in the presence of actinomycin D It has been postulated that regions of the genome involved in transcriptional
activity are more vigorously acetylated [17,18]. We wondered if the acetyla- tion reaction was in some way physically coupled to RNA synthesis, for example, as migh be found if the modification enzymes were directly
100
80
60
40
2O
u
~o
IOC
8C
4C
254
I I I I I I I I I 10 30 60 120 0 10 30 60 120
CHASE DURATION [MIN)
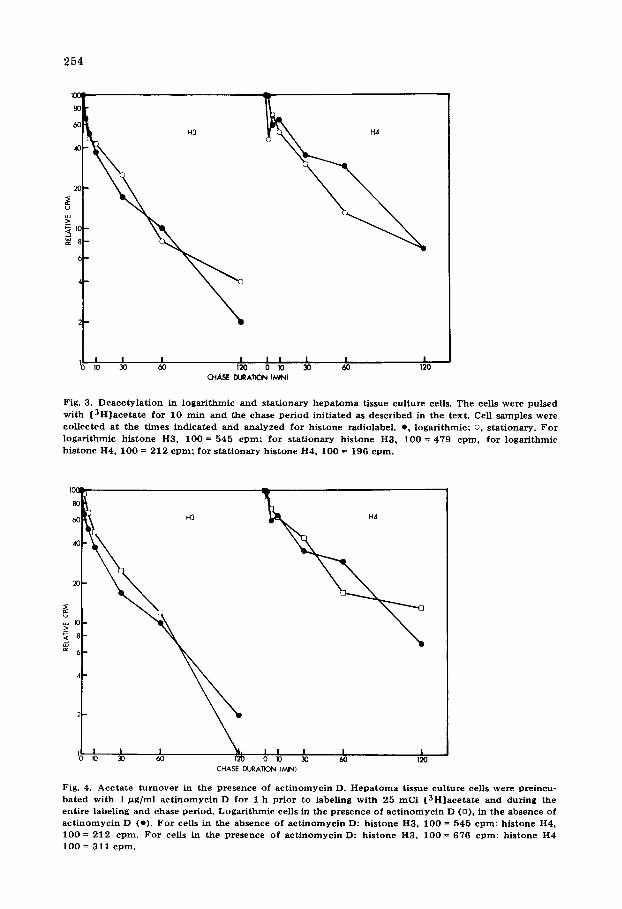
Fig. 3. D e a c e t y l a t i o n in l o g a r i t h m i c and s t a t i o n a r y h e p a t o m a t issue cu l tu re cells. The cells were pu l sed w i t h [ 3 H ] a c e t a t e for 10 ra in and the chase p e r i o d in i t i a t ed as desc r ibed in the tex t . Cell s a mp le s were co l lec ted a t the t i m e s i n d i c a t e d and a n a l y z e d fo r h i s t one radiolabel , e , l oga r i t hmic ; o, s t a t iona ry . F o r
l o g a r i t h m i c h i s t o n e H3, 100 = 545 c p m ; fo r s t a t i o n a r y h i s t one H3, 100 = 479 e p m , fo r l o g a r i t h m i c h i s t on e H4, 100 = 212 c p m ; for s t a t i o n a r y h i s t one H4, 100 = 196 c p m .
m
1 H3
I I 0 I0 30 60 0 10
CHASE DURATION (MIN)
I I I 30 60 120
Fig. 4. A c e t a t e t u r n o v e r in the p resence o f a c t i n o m y c i n D. H e p a t o m a t issue cu l tu re cells were pre incu- b a t e d w i t h 1 /~g/ml a c t i n o m y c i n D fo r 1 h p r io r to label ing w i t h 25 m C i [ 3 H ] a c e t a t e and du r ing the en t i r e label ing and chase pe r iod . L o g a r i t h m i c cells in the p r e sence of a c t i n o m y c i n D (o) , in the absence of
a c t i n o m y c i n D (o) . F o r cells in the absence of a c t i n o m y c i n D: h i s tone H3, 100 = 545 c p m ; h i s t one H4, I 0 0 = 212 c p m . F o r cells in the p r e sence of a c t i n o m y c i n D: h i s t one H3, I 0 0 = 676 c p m ; h i s tone H4 100 = 311 c p m .
255
associated with RNA polymerase. Accordingly we have analyzed acetylation and deacetylat ion as well as acetate uptake into the cytoplasm in the presence of a level of act inomycin D which inhibits the rate of RNA synthesis in logarithmic hepatoma tissue culture cells by about 75%. As shown in Fig. 4, inhibition of RNA synthesis in this way evidently has no discernible effect on the total deacetylation rate of all histone subfractions combined for histones H3 and H4. We found that the cytoplasm of the actinomycin D-treated cells contained about 30% more [3H]acetate after a 10 min pulse than the untreated logarithmic cells. We further found that the histones were about 50% more heavily labeled than in untreated cells, possibly reflecting either a change in the pool size of acetate, a nuclear membrane permeability change, or a change in the chromatin structure, allowing more rapid acetylation.
Hydrolysis of histone-acetate on other cell types Thus far, the exceedingly rapid modification and hydrolysis has been
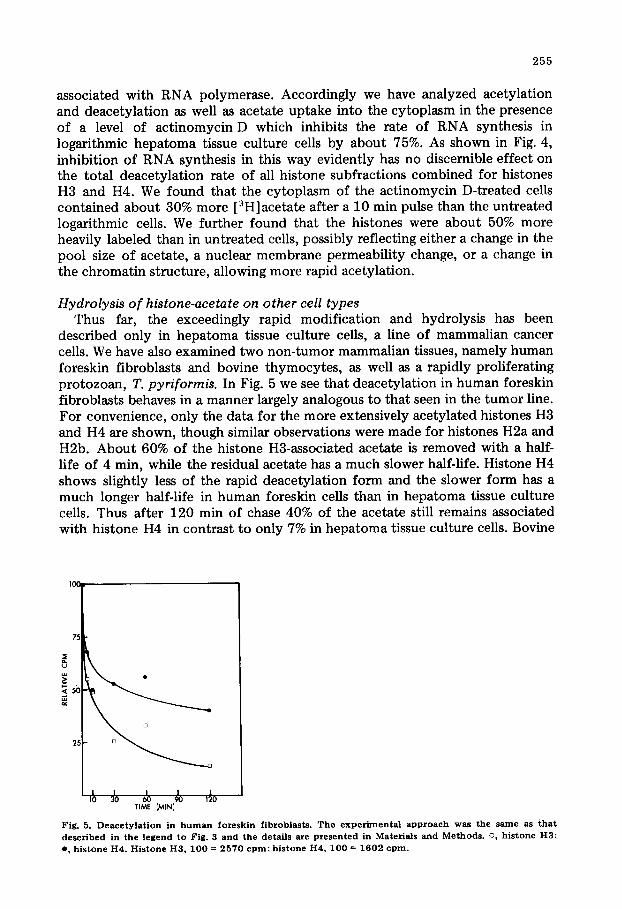
described only in hepatoma tissue culture cells, a line of mammalian cancer cells. We have also examined two non-tumor mammalian tissues, namely human foreskin fibroblasts and bovine thymocytes , as well as a rapidly proliferating protozoan, T. pyriformis. In Fig. 5 we see that deacetylation in human foreskin fibroblasts behaves in a manner largely analogous to that seen in the tumor line. For convenience, only the data for the more extensively acetylated histones H3 and H4 are shown, though similar observations were made for histones H2a and H2b. About 60% of the histone H3-associated acetate is removed with a half- life of 4 min, while the residual acetate has a much slower half-life. Histone H4 shows slightly less of the rapid deacetylat ion form and the slower form has a much longer half-life in human foreskin cells than in hepatoma tissue culture cells. Thus after 120 min of chase 40% of the acetate still remains associated with histone H4 in contrast to only 7% in hepatoma tissue culture cells. Bovine
t j
~b ~o ~ ~ ,~o TIME (MIN}
Fig. 5. Deaeety]ation in h u m a n foreskin f ibroblasts. The e x p e d m e n t a l approach was the same as tha t d e s c r i b e d in the l e g e n d to Fig. 3 and the detai l s are p z e s e n t e d i n M a t e r i a l s and M e t h o d s . o, h i s t o n e H 3 ;
e , h i s t o n e H4 . H i s t o n e H 3 , 1 0 0 = 2 5 7 0 c p m ; h i s t o n e H4 , 1 0 0 = 1 6 0 2 c p m .
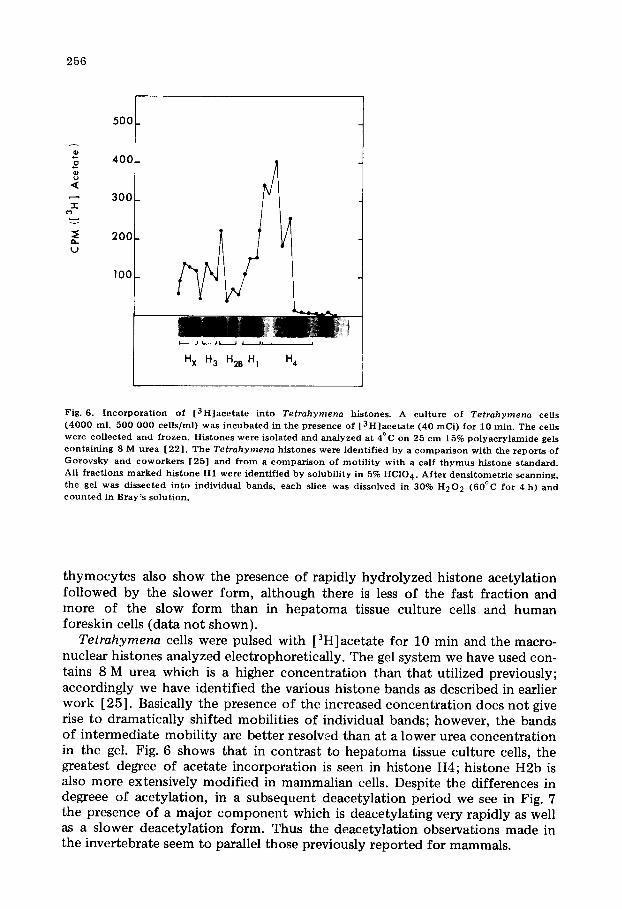
256
500
.,.? 4 0 0
.<
300 -1-
200 U
100
L I L ~ - - J ~ J I I ,
H x H 3 H2B H I H4
Fig. 6. I n c o r p o r a t i o n of [ 3 H ] a c e t a t e in to Tetrahymena histones. A cul ture of Tetrahymena cells ( 4 0 0 0 ml, 500 000 cel ls /ml) was i n c u b a t e d in the presence of [ 3 H ] a c e t a t e (40 mCi) for 10 min . The cells were co l lec ted and f rozen. His tones were isolated and ana lyzed at 4°C on 25 cm 15% p o l y a c r y l a m i d e gels con ta in ing 8 M u r e a [22 ] . The Tetrahymena his tones were ident i f ied by a c o m p a r i s o n wi th the r epor t s of G o r o v s k y and c o w o r k e r s [25] and f r o m a c o m p a r i s o n of mot i l i t y wi th a calf t h y m u s h i s tone s tandard . All f rac t ions m a r k e d h i s tone H1 were ident i f ied by solubi l i ty in 5% HC104. A f t e r dens i tome t r i c scanning, the gel was dissected in to individual bands , each slice was dissolved in 30% H 2 0 2 (60°C for 4 h) and c o u n t e d in Bray ' s solut ion.
thymocytes also show the presence of rapidly hydrolyzed histone acetylation followed by the slower form, although there is less of the fast fraction and more of the slow form than in hepatoma tissue culture cells and human foreskin cells {data not shown).
Tetrahymena cells were pulsed with [3H]acetate for 10 min and the macro- nuclear histones analyzed electrophoretically. The gel system we have used con- tains 8 M urea which is a higher concentration than that utilized previously; accordingly we have identified the various histone bands as described in earlier work [25]. Basically the presence of the increased concentration does not give rise to dramatically shifted mobilities of individual bands; however, the bands of intermediate mobility are better resolved than at a lower urea concentration in the gel. Fig. 6 shows that in contrast to hepatoma tissue culture cells, the greatest degree of acetate incorporation is seen in histone H4; histone H2b is also more extensively modified in mammalian cells. Despite the differences in degreee of acetylation, in a subsequent deacetylation period we see in Fig. 7 the presence of a major component which is deacetylating very rapidly as well as a slower deacetylation form. Thus the deacetylation observations made in the invertebrate seem to parallel those previously reported for mammals.
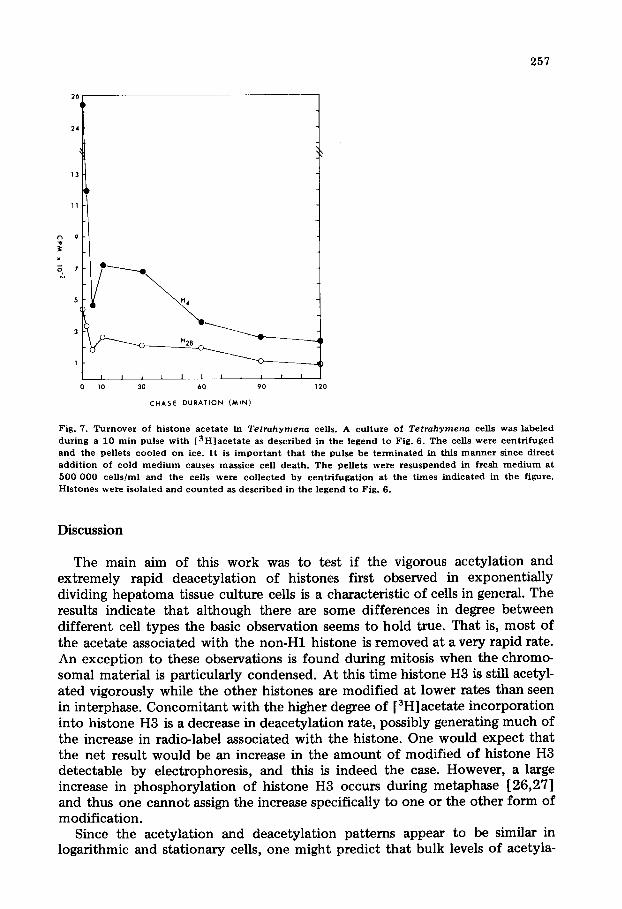
257
o
5 H 4
1 I I I l I 1 I 1 l I I I
0 10 30 6 0 9 0 120
C H A S E D U R A T I O N ( M I N )
Fig. 7. T u r n o v e r of h i s t one ace t a t e in Tetrahymena cells. A cu l tu re of Tetrahymena cells was labe led
d u r i n g a 10 ra in pulse w i t h [ 3 H ] a c e t a t e as desc r ibed in the l egend to Fig. 6. T h e cells were c e n t r i f u g e d and the pe l le ts c o o l e d on ice. I t is i m p o r t a n t that the pulse be t e r m i n a t e d in th is m a n n e r s ince d i r ec t
a d d i t i o n of cold m e d i u m causes mas s i ce cell dea th . T h e pel le ts were r e s u s p e n d e d in f resh m e d i u m a t
500 000 cells/m1 and the cells were co l l ec t ed by c e n t r i f u g a t i o n a t the t imes i n d i c a t e d in the f igure .
H i s t o n e s were i so la t ed and c o u n t e d as d e s c r i b e d in the l egend to Fig. 6.
Discussion
The main aim of this work was to test if the vigorous acetylation and extremely rapid deacetylation of histones first observed in exponentially dividing hepatoma tissue culture cells is a characteristic of cells in general. The results indicate that although there are some differences in degree between different cell types the basic observation seems to hold true. That is, most of the acetate associated with the non-H1 histone is removed at a very rapid rate. An exception to these observations is found during mitosis when the chromo- somal material is particularly condensed. At this time histone H3 is still acetyl- ated vigorously while the other histones are modified at lower rates than seen in interphase. Concomitant with the higher degree of [3H]acetate incorporation into histone H3 is a decrease in deacetylat ion rate, possibly generating much of the increase in radio-label associated with the histone. One would expect that the net result would be an increase in the amount of modified of histone H3 detectable by electrophoresis, and this is indeed the case. However, a large increase in phosphorylat ion of histone H3 occurs during metaphase [26,27] and thus one cannot assign the increase specifically to one or the other form of modification.
Since the acetylation and deacetylat ion patterns appear to be similar in logarithmic and stationary cells, one might predict that bulk levels of acetyla-

258
tion of the histones would also be similar in these two stages of cell growth. However, analysis of the amount of acetylation of histone H4 from logarithmic and stationary cells shows a statistically significant (p = 0.001) increase in bulk level histone H4 acetylation from 35.8% + 3.7% (logarithmic) to 53% + 8.1% (stationary). Since there is no obvious change in the apparent rates of acetyla- tion and deacetylat ion this phenomenon may result from an increase in pool size of acetyl-coenzyme A, as the stationary cells show a decreased require- ment for continued lipid synthesis.
An interesting finding was the fact that actinomycin D treatment of the logarithmic hepatoma tissue culture cells caused an increase in the apparent rate of acetylation of all non-H1 histones, although the deacetylation pattern was the same as in control cells. A possible explanation is that the actinomy- cin D, in binding to the DNA, alters the chromatin structure sufficiently to allow the acetylating enzymes greater access to the histones. Ruiz-Carillo et al. [28] showed that rifamycin AF/013, actinomycin D, and a-amanitin, other RNA synthesis inhibitors, faild to affect acetylation or deacetylation of his- tones in peripheral erythrocytes from anemic ducks. The above discrepancy in effect of act inomycin D on histone acetylation may be due to the fact that we were using widely different cell types [29]. Our results on deacetylation are in agreement with theirs.
The fraction of histone molecules which is modified is quite substantial. Thus in a 10 min pulse 95% or more of the label which becomes associated with histone H4 is not at the metabolically stable amino-terminus site [ 11 ]. The bulk of the acetate is incorporated onto pre-existing histone and we have previ- ously calculated that at least 30% of the total histone H4 complement is so modified in a 10 min period [11]. A point of great interest is whether all histones are available for acetylation or if only a specific population can be so modified. Since we know from densitometric scans of polyacrylamide gels that in the steady state abou t 35% of the histone H4 complement is modified and that a substantial fraction of this turns over very rapidly it seems unavoidable that a significant fraction of the total H4 must be available for such modification. It is difficult to estimate the fraction of histone H4 that might be unavailable (if there is any histone H4 unavailable) wi thout a detailed knowledge of the rate constant for the acetylation reaction in vivo, and such a determination is obviously subject to the model describing the acetylation reaction and the interconversions between fast and slow deacetylation forms. However, unless the rate constant for the acetylation reactions were very high (and the data of Fig. 1 indicate that while this is a vigorous reaction it is not excessively so) one must conclude that the bulk of the histone H4 is available for acetylation.
Clearly a much larger proport ion of histones in the cell are acetylated and deacetylated than would appear to be necessary if acetylation were involved only in activating specific genes or even specific regions of the genome. A likely explanation for these results might lie in a 2-fold function f o r acetylation. On the one hand a possible total lack of modification of a small fraction of the histone H4 might reflect its occurrence in heterochromatin. This would be con- sistent with the recent results of D'Anna et al. [14] who observed that the extent of histone H4 acetylation decreases in mitotic cells. Allfrey and his colleagues [30,15] have also argued that acetylation might play a role in the

259
distinction between euchromatin and heterochromatin, though one clearly cannot say whether this is a causitive role or simply a consequence of failure to acetylate in the heterochromatin environment. On the other hand, although all the H4 histone in the euchromatin might undergo a single level of modifica- tion, acetylation to levels of 3 and 4 acetate groups might be a more effective way of temporarily breaking histone-DNA bonds. Certainly the level of such higher modifications is quite small. Recent estimates of the number of histone H4 molecules which can be so modified indicates that they are of the order of 10--20% [31,32] a number which is a much more likely figure if acetylation is involved in a permissive effect in gene activation.
If highly modified forms of histone H4 (and histone H3) play a role in the overall transcriptional process, this poses a very real problem in a consideration of the fast and slow hydrolyzing forms of acetylation, in view of the observa- tion that the sites in the histones at which these modifications occur are identi- cal (Dixon, G. and Chalkley, R., unpublished data). It seems likely that the different deacetylat ion rates must occur in histone H3 and H4 molecules in different environments so that they have different susceptibilities to deacetyla- tion, but the nature of the difference in environment remains obscure.
Acknowledgements
The authors would like to thank Dr. Mark F. Stinsky of the Department of Microbiology, University of Iowa, for the use of foreskin cells in tissue culture. We also thank Dr. Normal Williams of the Department of Zoology, University of Iowa for his starter culture of Tetrahymena pyriformis. Finally, we wish to acknowledge the excellent technical assistance of Shin Liao. M.R.M. was a post- doctoral trainee of the National Institutes of Health. This work was supported by U.S.P.H.S. grants CA 10871 and Ca 17224.
References
1 Balhorn, R., BordweU, J., Sellers, L., Granner, D. and Chalkley, R. (19#2) Biochem. Biophys. Res. Commun. 46, 1326--1333
2 Balhorn, R., Balhom, M., Morris, H.P. and Chalkley, R. (1972) Cancer Res. 32, 1775--1784 3 Gurley, L., Walters, R. and Tobey, R. (1974) J. Cell Biol. 60, 356--364 4 Marks, D.B., Paik, W.K. and Borun, T.W. (1973) J. Biol. Chem. 248, 5660--5667 5 Easton, D. and Chalkley, R. (1972) Exp. Cell Res. 72, 502--508 6 Gershey, E.L., Vidali, G. and Allfrey, V.G. (1968) J. Biol. Chem. 243, 5018--5022 7 Vidali, G., Gershey, E.L. and Anfrey, V.G. (1968) J. Biol. Chem. 243, 6361---6366 8 Sanders, L.A., Schechter, N.M. and McCarthy, K.S. (1973) Biochemistry 12, 783--791 9 Philips, D.M.P. (1963) Biochem. J. 87, 258--263
10 Hnilica, L.S. (1972) in The Structure and Biological Funct ion of Histones, pp. 79--92, CRC Press, Cleveland
11 Jackson, V., Shires, A., Chalkley, R. and Granner, D.K. (1975) J. Biol. Chem. 250, 4856--4863 12 Jackson, V., Shires, A., Tanphaichitr , N. and Chalkley, R. (1976) J. Mol. Biol. 104, 471--483 13 Panyim, S. and Chalkley, R. (1969) Biochemistry 8, 3972--3979 14 D'Anna, J.A., Tobey, R.A., Barham, S.S. and Gurley, L.R. (1977) Biochem. Biophys. Res. Commun.
77, 187--194 15 Allfrey, V.G. (1971) in Histones and Nucleohis tones (Phillips, D.M.P., ed.), pp. 241--294, Plenum
Press, London 16 Libby, P.R. (1973) Biochem. J. 134, 90#--912 17 Pogo, B.G.T., Allfrey, V.G. and Mirsky, A.E. (1966) Proc. Natl. Acad. Sci. U.S. 55, 805--812 18 Wong, Lee-Jun C. and Alberts, B.M. (197#) Fed. Proc. 36, 784

2 6 0
19 Balhorn, R., Jackson, V., Granner, D. and Cha]kley, R. (1975) Biochemistry 14, 2504--2511 20 Thompson, Jr., G.A. (1967) Biochemistry 6, 2015--2022 21 Plesner, P., Rasmussen, L. and Zeuthen, E. (1964) in Synchrony in Cell Division and Growth
(Zeuthen, E., ed.), pp. 544--563, John Wiley and Sons, New York 22 Panyim, S. and Chalkley, R. (1969) Arch. Biochem. Biophys. 130, 337--346 23 Sung, M.T. and Dixon, G.H. (1970) Proc. Natl. Acad. Sci. U.S. 67, 1616--1623 24 Ruiz-Carillo, A., Wangh, L. and Allfrey, V.G. (1975) Science 190, 117--128 25 Johmann, C.A. and Gorovsky, M.A. (1976) Arch. Bioehem. Biophys. 175, 694--699 26 Lake, R.S. and Sa]zman, N.P. (1972) Biochemistry 11, 4817--4826 27 Lake, R.S. (1973) J. Cell Biol. 58, 317--331 28 Ruiz-Carillo, A., Wangh, L.J. and Allfrey, V.G. (1976) Arch. Biochem. Biophys. 174, 273--290 29 Ruiz-Carillo, A., Wangh, L.J., Lit tau, V.C. and Allfrey, V.G. (1974) J. Biol. Chem. 249, 7358--7368 30 Allfrey, V.G. (1965) Proc. Can. Cancer Conf. 6, 313--335 31 Sea]y, L. and Chalkley, R. (1978) Cell 14, 115--121 32 Riggs, M.G., Whittaker, R.G., Newmann, J.R. and Ingrarn, V.N. (1977) Nature 268, 462--464





























