Embed Size (px)
Citation preview

Cardiovascular Research 53 (2002) 921–935www.elsevier.com/ locate /cardiores
Redistribution of connexin45 in gap junctions of connexin43-deficienthearts
a a b c ,1Carolyn M. Johnson , Evelyn M. Kanter , Karen G. Green , James G. Laing ,a c ,2 a a,b,dTetsuo Betsuyaku , Eric C. Beyer , Thomas H. Steinberg , Jeffrey E. Saffitz ,
a,d ,*Kathryn A. YamadaaDepartment of Medicine, Washington University School of Medicine, St. Louis, MO, USA
bDepartment of Pathology, Washington University School of Medicine, St. Louis, MO, USAcDepartment of Pediatrics, Washington University School of Medicine, St. Louis, MO, USA
dCenter for Cardiovascular Research, Washington University School of Medicine, St. Louis, MO, USA
Received 22 May 2001; accepted 26 October 2001
Abstract
Objective: Adult ventricular myocytes express two gap junction channel proteins: connexin43 (Cx43) and connexin45 (Cx45).Cx43-deficient mice exhibit slow ventricular epicardial conduction, suggesting that Cx43 plays an important role in intercellular couplingin the ventricle. Cx45 is much less abundant than Cx43 in working ventricular myocytes. Its role in ventricular conduction has not beendefined, nor is it known whether expression or distribution of Cx45 is altered in Cx43-deficient mice. The present study was undertaken todetermine (1) whether expression of Cx45 is upregulated and (2) whether gap junction structure and distribution are altered in
1 / 1 1 / 2 2 / 2 1 / 1 1 / 2Cx43-deficient mice. Methods: Ventricular tissue from neonatal Cx43 , Cx43 and Cx43 and adult Cx43 and Cx43mice was analyzed by immunoblotting and confocal immunofluorescence microscopy. Results: Total Cx45 protein abundance measuredby immunoblotting was not different in Cx43-deficient or null hearts compared to wild-type control hearts. However, the amount and
1 / 2distribution of Cx45 immunoreactive signal measured by quantitative confocal analysis were markedly reduced in both Cx43 and2 / 2Cx43 hearts. Conclusion: Although the total content of Cx45 is not upregulated in Cx43-deficient hearts, the localization of Cx45 to
cardiac gap junctions depends on the expression level of Cx43 and is dramatically altered in mice that express no Cx43. 2002Elsevier Science B.V. All rights reserved.
Keywords: Cell communication; Developmental biology; Gap junctions; Histo(patho)logy; Myocytes; Remodeling
1. Introduction cardiac myocytes. Ventricular myocytes in the adult heartexpress connexin43 (Cx43) and connexin45 (Cx45)
Gap junction channels comprised of connexin subunits whereas atrial myocytes of most species including human,are responsible for cell-to-cell communication and inter- dog and mouse express Cx43, Cx45 and Cx40 [1–5]. Cx43cellular propagation of electrical signals throughout the is the predominant cardiac connexin [1,4,6–8]. Studies inheart. At least three different connexins are expressed in genetically engineered mice indicate that Cx43 is respon-
sible for cell-to-cell communication in the ventricles[9,10], whereas Cx40 is a major determinant of thatfunction in the atria [10–12]. The functional role of Cx43*Corresponding author. Cardiovascular Division, Box 8086, Washing-
ton University School of Medicine, 660 South Euclid Avenue, St. Louis, in development, normal conduction and arrhythmogenesisMO 63110, USA. Tel.: 11-314-3628-909; fax: 11-314-3628-957. is, however, complex. Conflicting reports describe modest
E-mail address: [email protected] (K.A. Yamada). conduction slowing [9,10,13] or lack thereof [14] in Cx43-1Present address: Washington University School of Medicine, Infecti-deficient hearts. Cx43 null myocytes survive well inous Diseases Division, Box 8051, St. Louis, MO 63110, USA.
2Present address: The University of Chicago, Department of Pediatrics,Section of Hematology/Oncology, Chicago, IL 60637, USA. Time for primary review 24 days.
0008-6363/02/$ – see front matter 2002 Elsevier Science B.V. All rights reserved.PI I : S0008-6363( 01 )00522-3

922 C.M. Johnson et al. / Cardiovascular Research 53 (2002) 921 –935
culture [15,16], are well coupled based on measurements control hearts, the area occupied by Cx45 immuno-of junctional conductance [15], and exhibit synchronized, fluorescent signal in confocal images was markedly re-rhythmic contractions [15,16]. Nevertheless, absence of duced in Cx43-deficient hearts, suggesting that distributioncardiac Cx43 results in ventricular arrhythmias and sudden of Cx45 in ventricular myocyte gap junctions is alteredcardiac death [17], consistent with demonstrated enhance- dramatically in cells lacking Cx43. Thus, Cx43 expressionment of ischemia-induced ventricular tachycardia in Cx43- level is a major determinant of the distribution of connexindeficient hearts [18]. proteins within ventricular myocyte gap junctions.
Cx45 appears to be the first cardiac connexin expressedduring the initial stage of cardiac contractions in the mouseand then is developmentally downregulated [19]. Original 2. Methodsstudies in adult canine hearts showed that Cx45 wasexpressed throughout the ventricular myocardium [1,4,20]. The investigation conforms with the Guide for the CareSubsequently, Coppen and co-workers [21,22] observed and Use of Laboratory Animals published by the USthat Cx45 immunofluorescence was concentrated at the National Institutes of Health (NIH Publication No. 85-23,endocardial surface with a distribution pattern overlapping revised 1996).that of Cx40-expressing myocytes of the ventricular con-duction system in the adult heart. Ventricular Cx45 protein 2.1. Cx43 micelevels were below the limit of detection on immunoblots
1 / 1 1 / 2[21]. Other investigators have since observed Cx45 in low We maintain a colony of Cx43 and Cx43 adultamounts in foci dispersed in the ventricular free walls of mice for breeding in a standard barrier facility. Founder
tm1Kdrthe adult heart [19], and faint b-gal staining representing mice (B6,129-Gja1 ) were obtained from Jackson2 / 2Cx45 expression in myocytes of adult working myocar- Laboratory (Bar Harbor, ME, USA). Because Cx43
dium [23]. mice die soon after birth, pregnancies were timed byThe functional role of Cx45 in cardiac conduction in housing breeding pairs overnight only. No changes in litter
adult working myocardium is entirely unknown. Compared size, genotype distribution, or Cx43 expression have beenwith channels made by Cx43 and Cx40, Cx45 channels observed over the past 4 years. The genotypes of all micehave low unitary conductances and are more voltage were determined by polymerase chain reaction (PCR) asdependent [24–27]. Although the amount of Cx45 in described previously [16].ventricular myocytes may be considerably less than Cx43,substantial evidence indicates that Cx43 and Cx45 can 2.2. Characterization of anti-Cx45 antibodyform hybrid gap junction channels with unique biophysicalproperties [28–30]. Thus, the functional consequences of The anti-Cx45 antibody used in the present study wasCx45 expression in the ventricles may be greater than its produced at Washington University, St. Louis, MO, USA,low level of expression would suggest, especially in mice against a mouse 6 his-carboxy terminus (amino acidsdeficient in Cx43. Previously, we have used genetically 259–396) fusion protein as described by Lecanda et al.altered mice deficient in Cx43 protein [31] for detailed [33]. Detailed experiments demonstrating specificity of theinvestigations into the expression, distribution and func- anti-Cx45 antibody for Cx45 and absence of crossreactivi-tional role of Cx43 in the heart [9,10,32]. In a recent study ty with Cx43 have been performed by us (Lin andinvestigating the relationship between the amount of Steinberg, unpublished observations) and in the laboratoryconnexin expressed and the structure of gap junctions, we of Professor Habo Jongsma [34]. To further validate thefound that the number, but not the size, of gap junctions is specificity of the anti-Cx45 antibody, we performed im-
1 / 2reduced in Cx43 hearts in which 50% of the wild-type munostaining experiments on control HeLa cells and HeLalevel of Cx43 was expressed [32]. In the present study, we cells transfected to overexpress either mouse Cx45 (HeLa-undertook a systematic approach to determine whether 45) or rat Cx43 (HeLa-43). Anti-Cx45 antibody staining ofcompensatory upregulation of Cx45 occurs in Cx43-de- control HeLa cells demonstrated minimal to no back-ficient hearts, and whether the structure or distribution of ground expression of Cx45 in control HeLa cells (Fig. 1A).gap junctions containing Cx45 is altered in Cx43-deficient The level of anti-Cx45 staining of HeLa-43 cells was nothearts. We used a new anti-Cx45 antibody produced in one significantly different from control HeLa cells demon-of our laboratories (T.H.S.), which has been extensively strating that the anti-Cx45 antibody did not recognizecharacterized and found to be monospecific for Cx45. Cx43 protein (Fig. 1A). In contrast, HeLa-45 cells ex-
We report two new findings. First, Cx45 immuno- hibited intense staining with the anti-Cx45 antibody (Fig.fluorescent signal was observed in both adult and neonatal 1A). In addition, immunoblot analysis of control HeLa,mouse ventricles albeit at lower levels than described HeLa-45 and HeLa-43 cells (Fig. 1B) confirmed thepreviously [1,4,20]. Second, although total Cx45 protein immunofluorescence data, demonstrating that the anti-levels measured in immunoblots were unchanged in Cx45 antibody did not recognize Cx43 protein in the
1 / 2 2 / 2 1 / 1Cx43 and Cx43 hearts compared to Cx43 HeLa-43 cell lane while demonstrating an intense band in

C.M. Johnson et al. / Cardiovascular Research 53 (2002) 921 –935 923
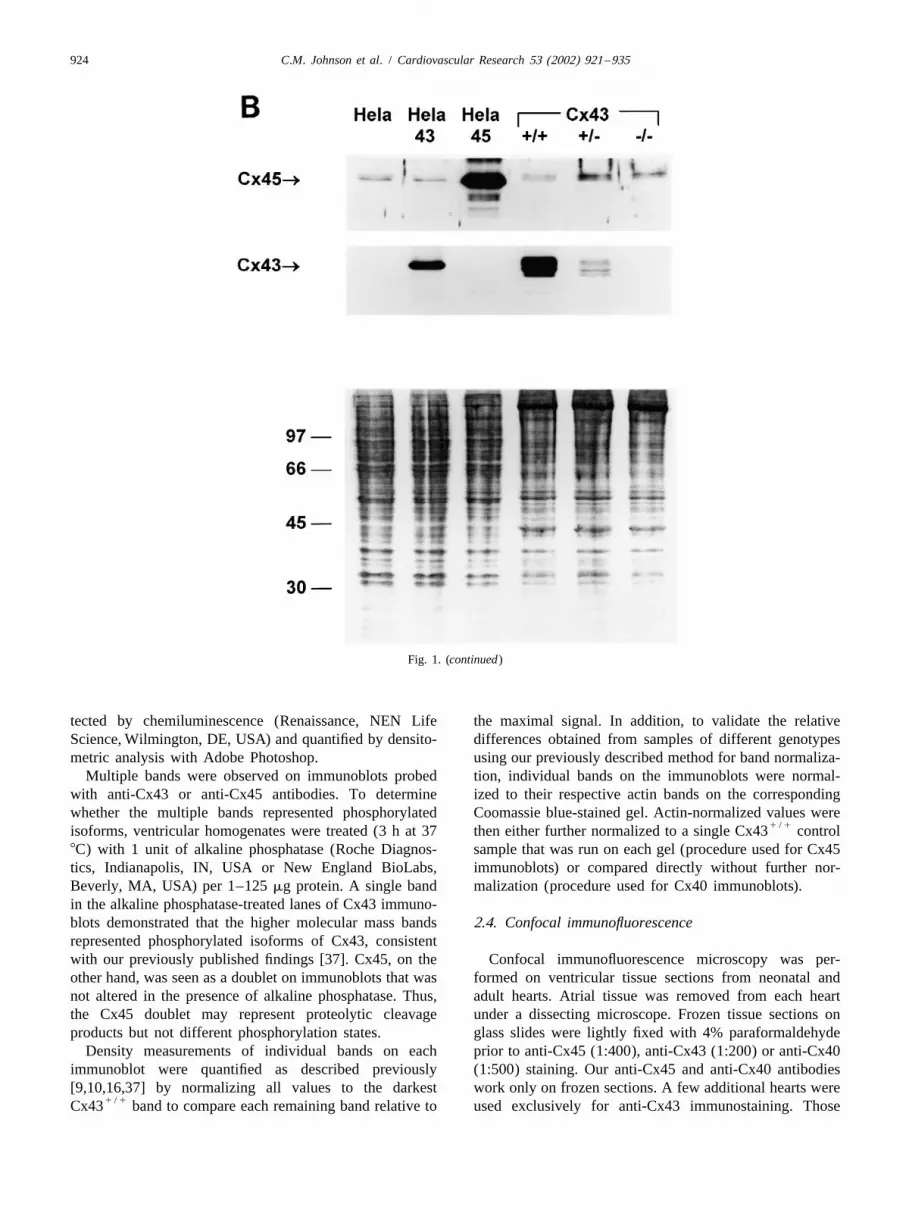
Fig. 1. (A) Cx45 (panels on the top) and Cx43 (panels on the bottom) immunostaining in HeLa cells showing monospecificity and absence ofcrossreactivity of anti-Cx45 antisera used in the present study. Left: Control HeLa cells exhibit low background levels of Cx45 expression. Center: HeLacells overexpressing Cx45 (HeLa-Cx45) exhibit intense Cx45 staining reflecting abundant Cx45 expression and background levels of Cx43. Right: HeLacells overexpressing Cx43 (HeLa-Cx43) exhibit low levels of Cx45 expression comparable to control HeLa cells and marked overexpression of Cx43.
1 / 1 1 / 2 2 / 2Bar510 mm and applies to all panels. (B) Immunoblots of HeLa, HeLa-43, and HeLa-45 cells and Cx43 , Cx43 and Cx43 ventricular tissuesamples from left to right probed with anti-Cx45 antisera (top) and anti-Cx43 antibody (middle). Corresponding Coomassie blue-stained gel is shown
1 / 1 1 / 2below. (C) Cx45 (panels on the left) and X-gal (panels on the right) staining of frozen tissue sections of whole Cx45 (top), Cx45 (middle) and2 / 2 1 / 1 1 / 2 2 / 2Cx45 (bottom) embryos (E8.5). Specific Cx45 signal is visible as punctate bright red spots in the Cx45 and Cx45 embryos. Cx45 embryo is
1 / 2 2 / 2devoid of Cx45 signal. Positive X-gal staining (dark purple) identifies tissue in Cx45 and Cx45 embryos in which nls-lacZ has replaced Cx45. Barscorrespond to 50 and 100 mm as indicated.
the HeLa-45 cell lane. Finally, we tested the specificity of EDTA, 1 EGTA, pH 8.0, containing the following proteasethe anti-Cx45 antibody on Cx45-deficient E8.5 embryos inhibitors: 1 mM pepstatin, 100 nM aprotinin, 1 mM[35] obtained from Professor Yosaburo Shibata and co- benzamidine, 1 mM iodoacetamide, 1 mM leupeptin, 1
1 / 1workers. Our anti-Cx45 antibody clearly stained Cx45 mM phenylmethylsulfonyl fluoride (PMSF), and sonicated1 / 2and Cx45 embryos hidentified both by PCR [35] and by (3315 s after each homogenization cycle).
positive X-gal (Invitrogen Life Technologies, Carlsbad, Aliquots of protein (7 mg for anti-Cx43 blots; 20 or 302 / 2CA, USA) staining [36]j but did not stain Cx45 mg for anti-Cx40 and anti-Cx45 blots) were added to 43
embryos (Fig. 1C), further demonstrating specificity of the sample buffer [0.0625 M Tris, 11.25% glycerol, 0.5%antibody used in the present study. sodium dodecyl sulfate (SDS), 0.001% bromphenol blue,
1% b-mercaptoethanol) and resolved by SDS–poly-2.3. Immunoblotting acrylamide (10%) gel electrophoresis and transferred
(semi-dry) to nitrocellulose membranes (Schleicher andImmunoblot analysis was performed on ventricular Schuell, Keene, NH, USA). Membranes were blocked in
tissue from neonatal (1-day-old pups) and adult hearts. 5% nonfat, dry milk in Tris-buffered saline–Tween 20,Although Cx40 expression is not normally detectable in then incubated with rabbit anti-rat Cx43 (1:5000, Zymedworking adult ventricular myocytes, we measured Cx40 Laboratories, South San Francisco, CA, USA), proteinlevels in addition to Cx45 and Cx43 in neonatal and adult A-purified rabbit anti-mouse Cx45 (1:500, Washingtonhearts to determine whether compensatory changes in its University), or rabbit anti-rat Cx40 (1:500, derived in ourexpression occurred in Cx43-deficient hearts. Hearts were laboratories against rat Cx40 carboxy terminus amino acidsrapidly removed from anesthetized mice and frozen in 235–355) antibodies as described previously [16]. Goatliquid nitrogen. The frozen tissue was pulverized and anti-rabbit antibody (1:5000, Jackson ImmunoResearchhomogenized (20 strokes32 in a Duall homogenizer) in a Laboratories, West Grove, PA, USA) was used as thehydrogencarbonate buffer solution (mM): 1 NaHCO , 5 secondary antibody. Immunoreactivity of blots was de-3
924 C.M. Johnson et al. / Cardiovascular Research 53 (2002) 921 –935
Fig. 1. (continued)
tected by chemiluminescence (Renaissance, NEN Life the maximal signal. In addition, to validate the relativeScience, Wilmington, DE, USA) and quantified by densito- differences obtained from samples of different genotypesmetric analysis with Adobe Photoshop. using our previously described method for band normaliza-
Multiple bands were observed on immunoblots probed tion, individual bands on the immunoblots were normal-with anti-Cx43 or anti-Cx45 antibodies. To determine ized to their respective actin bands on the correspondingwhether the multiple bands represented phosphorylated Coomassie blue-stained gel. Actin-normalized values were
1 / 1isoforms, ventricular homogenates were treated (3 h at 37 then either further normalized to a single Cx43 control8C) with 1 unit of alkaline phosphatase (Roche Diagnos- sample that was run on each gel (procedure used for Cx45tics, Indianapolis, IN, USA or New England BioLabs, immunoblots) or compared directly without further nor-Beverly, MA, USA) per 1–125 mg protein. A single band malization (procedure used for Cx40 immunoblots).in the alkaline phosphatase-treated lanes of Cx43 immuno-blots demonstrated that the higher molecular mass bands 2.4. Confocal immunofluorescencerepresented phosphorylated isoforms of Cx43, consistentwith our previously published findings [37]. Cx45, on the Confocal immunofluorescence microscopy was per-other hand, was seen as a doublet on immunoblots that was formed on ventricular tissue sections from neonatal andnot altered in the presence of alkaline phosphatase. Thus, adult hearts. Atrial tissue was removed from each heartthe Cx45 doublet may represent proteolytic cleavage under a dissecting microscope. Frozen tissue sections onproducts but not different phosphorylation states. glass slides were lightly fixed with 4% paraformaldehyde
Density measurements of individual bands on each prior to anti-Cx45 (1:400), anti-Cx43 (1:200) or anti-Cx40immunoblot were quantified as described previously (1:500) staining. Our anti-Cx45 and anti-Cx40 antibodies[9,10,16,37] by normalizing all values to the darkest work only on frozen sections. A few additional hearts were
1 / 1Cx43 band to compare each remaining band relative to used exclusively for anti-Cx43 immunostaining. Those

C.M. Johnson et al. / Cardiovascular Research 53 (2002) 921 –935 925
Fig. 1. (continued)
hearts were fixed in 10% formalin, embedded in paraffin, rabbit secondary antibody (1:400, Jackson Immuno-sectioned (6 mm thick), and subjected to microwave Research).antigen recovery for anti-Cx43 (1:200) staining [38]. All hearts were sliced in transverse section approximate-Sections were then incubated in Cy3-conjugated goat anti- ly midway between the apex and the base. Histologic

926 C.M. Johnson et al. / Cardiovascular Research 53 (2002) 921 –935
sections were subsequently prepared in the same plane field). An arbitrary signal intensity threshold was identifiedfrom the cut surface. Immunofluorescence images were for each set of sections stained with a given primarycollected from sections that exhibited intact tissue structure antibody that clearly distinguished high intensity signalwith minimal tears, folds, gaps or artefacts. All confocal concentrated in discrete spots from all other areas in theimages were acquired from the left ventricular midmyocar- test field. Signal intensity that exceeded this threshold wasdial layer where the myocytes were longitudinally (circum- assumed to represent gap junctions. A much lower thres-ferentially) oriented. Subendocardial and subepicardial hold value was also identified below which areas of thelayers were excluded from analysis, as were myocytes slide not covered by cells were defined. The proportion oforiented in an oblique plane of sectioning. Within the total tissue area occupied by gap junctions (% cell area)midmyocardium, fields containing the most robust staining was defined as the number of pixels with signal intensitywere selected for imaging so we could compare the exceeding the high threshold divided by the total numbermaximal signal across genotypes. Confocal images were of pixels exceeding the low threshold.acquired through a Zeiss microscope using an ArKr laser Total number and mean size of gap junctions were(568 nm, Molecular Dynamics) as described previously determined by defining an individual gap junction as 5 or[32]. Five separate images (fields) were scanned from more contiguous pixels exceeding the high threshold valuesections of each heart. Five to 11 hearts of each genotype [32]. The quantitative procedures developed previously onwere analyzed. adult ventricular tissue sections [32] were verified in the
Quantitative confocal analysis of connexin immuno- present study on neonatal ventricular tissue sections. Fig. 2fluorescent signal has been validated and described in demonstrates representative confocal images of neonataldetail in a previous study [32]. Briefly, test fields were gap junctions before and after subtraction of pixels that did
6digitized into a 102431024 matrix (|1.05310 pixels / test not achieve the signal threshold that distinguished gap
Fig. 2. Quantitative analysis of gap junction number and size by confocal microscopy and digital image processing in neonatal ventricular tissue. (A)Representative confocal image of neonatal ventricular myocardium cut in longitudinal section. Myocytes appear grey; gap junctions are bright white spots.(B) Same image as in A after subtraction of pixels that did not achieve the signal threshold that distinguished gap junctions (clusters of pixels exhibitingspecific immunoreactive signal at cell–cell junctions) from other structures. (C) High-resolution image of a cluster of Cx43 immunoreactive signalidentified between the arrows in A and B. (D) Same image as in C following subtraction of pixels that did not achieve the signal threshold thatdistinguished gap junctions from other structures. Image processing software identified eight individual gap junctions (clusters $5 contiguous pixels). Barscorrespond to 10 and 1 mm as indicated.
C.M. Johnson et al. / Cardiovascular Research 53 (2002) 921 –935 927
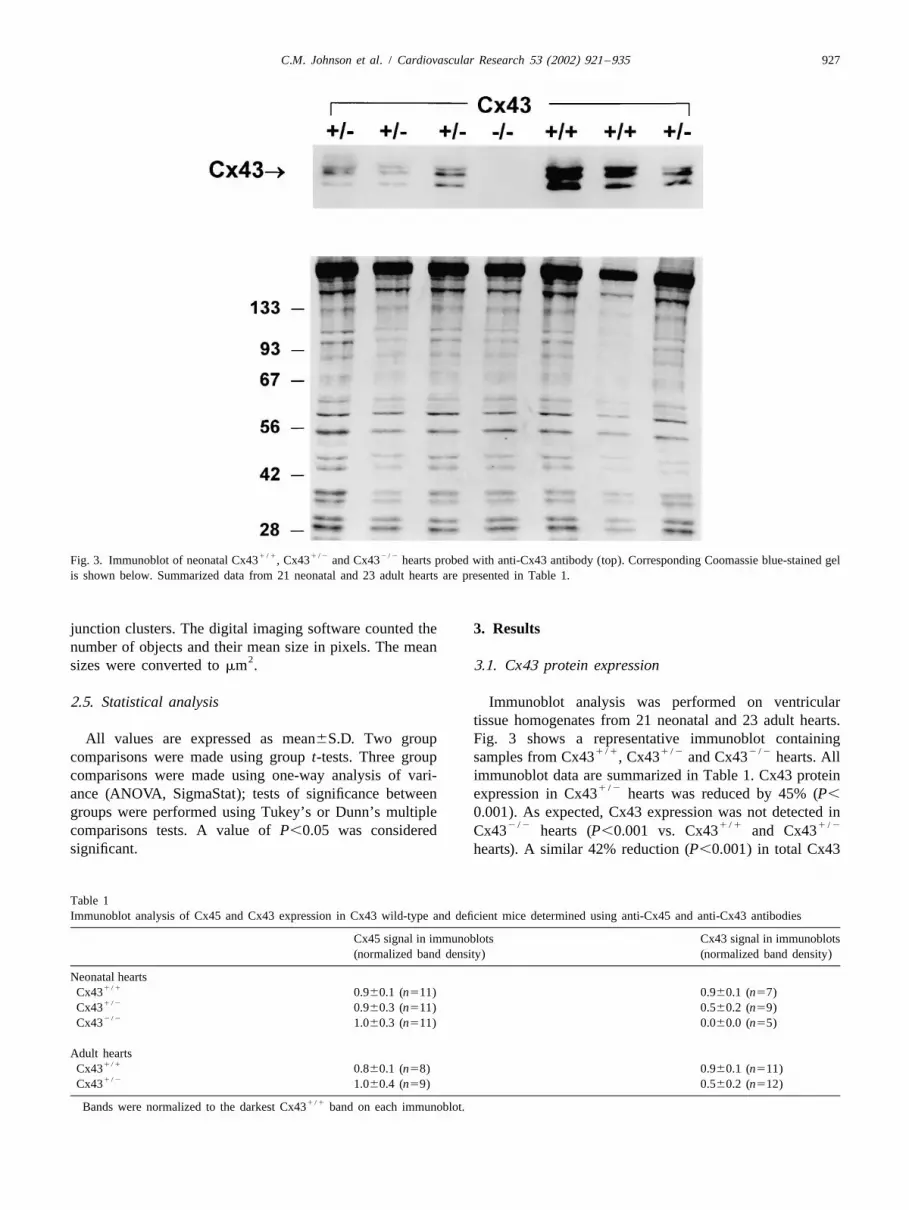
1 / 1 1 / 2 2 / 2Fig. 3. Immunoblot of neonatal Cx43 , Cx43 and Cx43 hearts probed with anti-Cx43 antibody (top). Corresponding Coomassie blue-stained gelis shown below. Summarized data from 21 neonatal and 23 adult hearts are presented in Table 1.
junction clusters. The digital imaging software counted the 3. Resultsnumber of objects and their mean size in pixels. The mean
2sizes were converted to mm . 3.1. Cx43 protein expression
2.5. Statistical analysis Immunoblot analysis was performed on ventriculartissue homogenates from 21 neonatal and 23 adult hearts.
All values are expressed as mean6S.D. Two group Fig. 3 shows a representative immunoblot containing1 / 1 1 / 2 2 / 2comparisons were made using group t-tests. Three group samples from Cx43 , Cx43 and Cx43 hearts. All
comparisons were made using one-way analysis of vari- immunoblot data are summarized in Table 1. Cx43 protein1 / 2ance (ANOVA, SigmaStat); tests of significance between expression in Cx43 hearts was reduced by 45% (P,
groups were performed using Tukey’s or Dunn’s multiple 0.001). As expected, Cx43 expression was not detected in2 / 2 1 / 1 1 / 2comparisons tests. A value of P,0.05 was considered Cx43 hearts (P,0.001 vs. Cx43 and Cx43
significant. hearts). A similar 42% reduction (P,0.001) in total Cx43
Table 1Immunoblot analysis of Cx45 and Cx43 expression in Cx43 wild-type and deficient mice determined using anti-Cx45 and anti-Cx43 antibodies
Cx45 signal in immunoblots Cx43 signal in immunoblots(normalized band density) (normalized band density)
Neonatal hearts1 / 1Cx43 0.960.1 (n511) 0.960.1 (n57)1 / 2Cx43 0.960.3 (n511) 0.560.2 (n59)2 / 2Cx43 1.060.3 (n511) 0.060.0 (n55)
Adult hearts1 / 1Cx43 0.860.1 (n58) 0.960.1 (n511)1 / 2Cx43 1.060.4 (n59) 0.560.2 (n512)
1 / 1Bands were normalized to the darkest Cx43 band on each immunoblot.

928 C.M. Johnson et al. / Cardiovascular Research 53 (2002) 921 –935
1 / 1 1 / 2 2 / 2Fig. 4. Representative confocal images of Cx43 immunofluorescence in neonatal Cx43 (left), Cx43 (center) and Cx43 (right) ventricular tissuesections stained with anti-Cx43 antibody. Bar510 mm and applies to all panels.
1 / 2 1 / 2 1 / 1protein was observed in tissue from adult Cx43 (n512) neonatal Cx43 compared to Cx43 hearts was the1 / 1vs. Cx43 (n511) hearts. These data corroborate previ- result of reductions in both the number and size of gap
ous reports that Cx43 protein expression is reduced by junctions. No Cx43 immunostaining was detected in1 / 2 2 / 2|50% in Cx43 ventricles [9,10,16] and define Cx43 Cx43 tissue as expected.
protein levels in hearts used for analysis of Cx45 expres-sion and distribution. 3.3. Cx45 protein expression
Immunoblot analysis was performed to determine ex-3.2. Confocal microscopy of Cx43 immunostaining pression levels of Cx45 in 33 neonatal and 17 adult
ventricles. Fig. 5 shows a representative immunoblotCx43 immunostaining was performed on neonatal ven- demonstrating specific bands for Cx45 protein in neonatal
1 / 1 1 / 2 2 / 2tricular tissue sections from 11 hearts of each genotype. Cx43 , Cx43 and Cx43 ventricular tissue1 / 1Fig. 4 shows representative images from neonatal homogenates. Quantitative data from 11 Cx43 , 11
1 / 1 1 / 2 2 / 2 1 / 2 2 / 2Cx43 , Cx43 , and Cx43 mice. Neonatal ven- Cx43 and 11 Cx43 hearts are given in Table 1 andtricular tissue showed the characteristic pattern [39–41] of demonstrate that the level of total Cx45 protein contentpunctate spots of immunofluorescent signal distributed was similar for all three genotypes. Table 1 includesaround the cell borders. As expected, based on immuno- summarized Cx45 expression data from adult hearts asfluorescence data reported in adult hearts [32], Cx43 signal well; no significant difference in total Cx45 protein content
1 / 2 1 / 1 1 / 2was significantly reduced in Cx43 compared to between Cx43 and Cx43 hearts was observed.1 / 1 2 / 2Cx43 hearts and completely absent in Cx43 hearts. Cx45 immunoblots were subjected to additional analysis.
As shown in Table 2, the % cell area occupied by Cx43 Individual bands on each immunoblot were normalized to1 / 2signal in Cx43 tissue was reduced to 41% of the their respective actin bands on the corresponding Coomas-
1 / 1Cx43 value. The significant reduction in % cell area in sie blue-stained gel. All bands were compared to a single
Table 2Confocal analysis of Cx43 immunostaining in Cx43 wild-type and deficient mice
Cx43 signal2% Cell area Gap junction number Gap junction size (mm )
Neonatal hearts1 / 1Cx43 (n511) 0.8560.30 254696 0.2660.051 / 2Cx43 (n511) 0.3560.12* 147636* 0.1860.02*2 / 2Cx43 (n511) – – –
1 / 1*P,0.05 versus Cx43 ; – no signal detected.

C.M. Johnson et al. / Cardiovascular Research 53 (2002) 921 –935 929
munoreactive signal in the midmyocardium from the left1 / 1 1 / 2 2 / 2ventricles of Cx43 , Cx43 and Cx43 mice was
analyzed. Although Cx45 protein expression was shown byimmunoblot analysis to be unchanged, inspection of theconfocal images showed an obvious decrease in the areaover which specific Cx45 signal was distributed in ven-
1 / 2 2 / 2tricular myocardium from both Cx43 and Cx43mice.
Quantitative analysis by digital image processing of theimmunofluorescence signals confirmed this unexpectedfinding. As shown in Table 3, the area occupied by Cx45signal expressed as a % of cell area was reduced by 22%
1 / 2in neonatal Cx43 hearts (P50.42) and by 68% in2 / 2 1 / 1Cx43 hearts (P,0.001) compared to Cx43 hearts.
The number of gap junctions containing Cx45 was reduced1 / 2significantly in both neonatal Cx43 (P,0.001) and
2 / 2 1 / 1Cx43 (P,0.001) hearts compared to Cx43 controls(Table 3). The size of gap junctions containing Cx45 was
1 / 2not reduced in neonatal Cx43 hearts (P50.69), but was2 / 2reduced significantly in neonatal Cx43 hearts (P,
2 / 20.001). Thus, in Cx43 hearts with complete absence ofCx43, both number and size of Cx45 gap junctions aremarkedly reduced. Dramatic alterations in Cx45 gap
1 / 2junctions were also observed in adult Cx43 hearts.Cx45 immunostaining expressed as a % of cell area was
1 / 2reduced in adult Cx43 hearts by 69% (P50.005), dueto decreases in both number (P50.009) and size (P,
1 / 20.001) of Cx45 gap junctions in Cx43 compared to1 / 1 1 / 2 2 / 2Fig. 5. Immunoblot of Cx43 , Cx43 and Cx43 hearts probed 1 / 1Cx43 hearts.
with anti-Cx45 antisera (top). Corresponding Coomassie blue-stained gel 1 / 1 1 / 2Sections from Cx43 and Cx43 hearts wereis shown below. Summarized data from 33 neonatal and 17 adult heartsdouble labeled with both anti-Cx45 and anti-Cx43 anti-are presented in Table 1.
bodies. Fig. 7 demonstrates extensive colocalization ofCx45 and Cx43 in gap junctions in the ventricular myocar-dium of the mouse.
1 / 1 1 / 1adult Cx43 control or a single neonatal Cx43control, which were given a value of 1. There was nosignificant difference in Cx45 protein content in adult
1 / 1 1 / 2Cx43 (1.0) vs. Cx43 (1.460.7, n59; P50.728) or 3.5. Cx40 protein expression1 / 1 1 / 2neonatal Cx43 (1.0) vs. Cx43 (1.160.3, n510) vs.
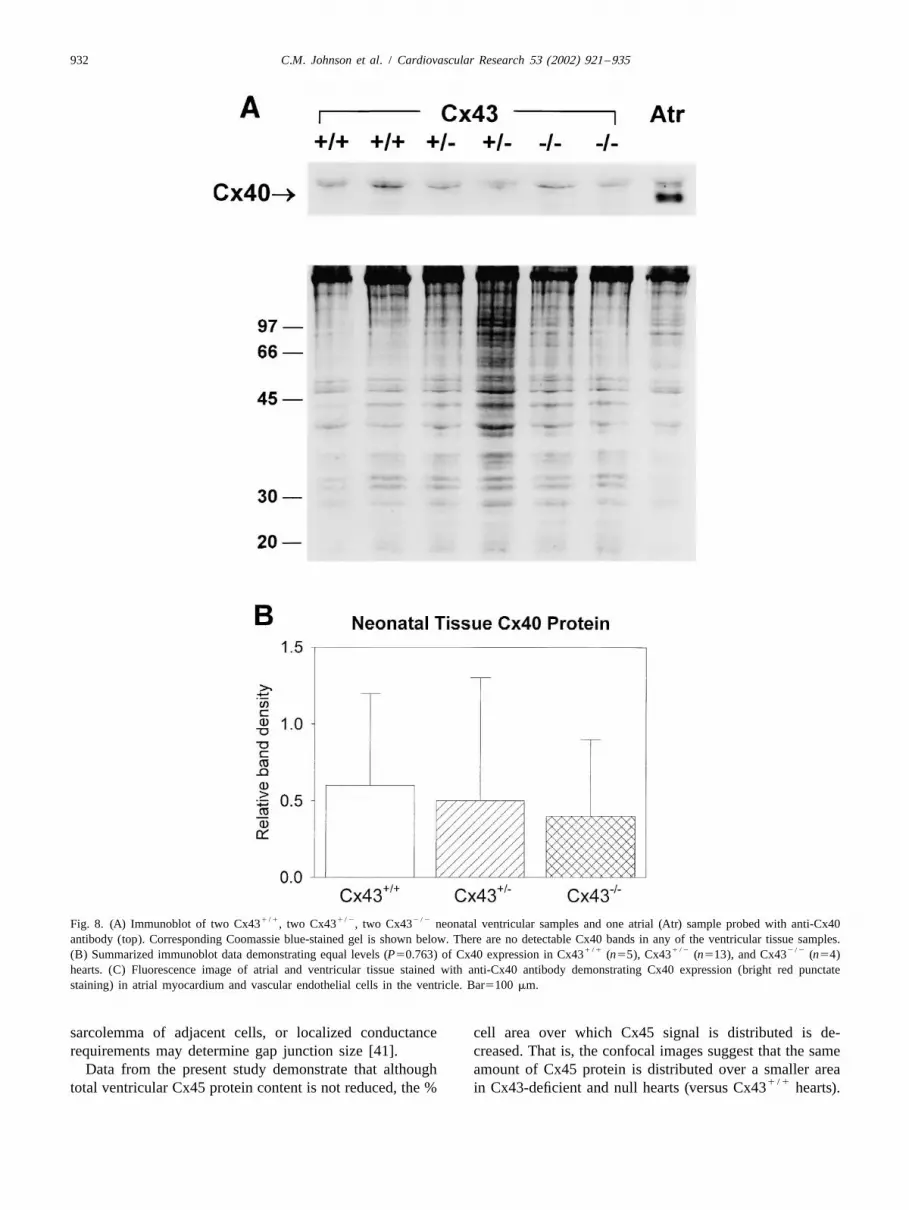
2 / 2 Cx40 protein expression was assessed in ventricularCx43 (2.161.1, n511; P50.413) ventricles. Thesetissue from 22 hearts. When homogenized atria weredata demonstrate that the total amount of ventricular Cx45included on the gels as a positive control, it was obviousdid not change in Cx43-deficient or null hearts.that the level of Cx40 expression was extremely low orabsent in ventricular tissue (Fig. 8A). Expression of Cx40
3.4. Confocal microscopy of Cx45 immunostaining has been reported to be restricted to the Purkinje systemand vascular endothelium [2–5,20,42] and would not be
Fig. 6 shows representative images from neonatal expected to yield a strong signal, if any, on immunoblots1 / 1 1 / 2 2 / 2Cx43 , Cx43 and Cx43 hearts and adult of whole ventricular homogenates. Because atrial samples1 / 1 1 / 2Cx43 and Cx43 hearts stained with anti-Cx45 were not run on every gel, we compared all Cx40 band
antisera. Cx45 expression was not confined exclusively to densities normalized to each respective actin band onthe endocardium and interventricular septum in the present Coomassie blue-stained gels. Fig. 8B summarizes Cx40study. Although specific Cx45 signal was observed in the protein expression and shows that total ventricular Cx40endocardium consistent with expression in the conduction protein levels were virtually identical, albeit minimal, in allsystem, we also observed Cx45 signal at intercellular three genotypes. These data demonstrate that ventricularjunctions between working ventricular myocytes through- Cx40 protein expression is not upregulated in Cx43-de-out the left ventricular myocardial free wall. Cx45 im- ficient or null hearts.

930 C.M. Johnson et al. / Cardiovascular Research 53 (2002) 921 –935
1 / 1 1 / 2 2 / 2Fig. 6. Representative confocal images of Cx45 immunofluorescence in Cx43 (left), Cx43 (center) and Cx43 (right) ventricular tissue sectionsstained with anti-Cx45 antisera. Neonatal tissue images are shown on top. Adult tissue images are shown below. Bar510 mm and applies to all panels.
3.6. Confocal microscopy of Cx40 immunostaining expression in ventricular tissue as shown by immunoblotanalysis (Fig. 8A and B).
As expected, immunoreactive signal in ventricular tissuestained with anti-Cx40 antibody was confined to suben-docardial foci (presumably His-Purkinje staining) and 4. Discussionintramural coronary vascular endothelial cells. No Cx40signal was observed in ventricular myocytes. Fig. 8C These data demonstrate that Cx45 is expressed inshows a representative fluorescence image demonstrating neonatal and adult ventricular myocytes of the mouse.expected atrial staining of Cx40 and absence of Cx40 in Cx45 and Cx40 are developmentally downregulatedventricular myocardium except in vascular endothelial [19,43–45] and exhibit limited expression in adult ventri-cells. These results confirm the low level of Cx40 protein cles. There has been uncertainty as to the level of Cx45
Table 3Confocal analysis of Cx45 immunostaining in Cx43 wild-type and deficient mice
Cx45 Signal2% Cell area Gap junction number Gap junction size (mm )
Neonatal hearts1 / 1Cx43 (n511) 0.4160.12 173630 0.1760.021 / 2Cx43 (n510) 0.3260.22 117626* 0.1660.022 / 2Cx43 (n510) 0.1360.11** 48634** 0.1260.02**
Adult hearts1 / 1Cx43 (n55) 0.3260.12 137643 0.5660.051 / 2Cx43 (n55) 0.1060.04* 63623* 0.3960.02*
1 / 1 1 / 1 1 / 2*P,0.05 versus Cx43 ; **P,0.05 versus Cx43 and Cx43 .

C.M. Johnson et al. / Cardiovascular Research 53 (2002) 921 –935 931
1 / 1Fig. 7. Double labeling of Cx43 ventricular myocardium with both anti-Cx43 (left) and anti-Cx45 (right) antibodies. Antibodies used in the doublelabel experiments were: primary for Cx43, mouse monoclonal (1:400, Chemicon International, Temecula, CA, USA); secondary for Cx43, Cy3-conjugatedgoat anti-mouse (1:400, Jackson ImmunoResearch Laboratories); primary for Cx45, rabbit polyclonal (1:500, Washington University); secondary for Cx45,Alexa Fluor 488 goat anti-rabbit (1:400, Molecular Probes, Eugene, OR, USA). Extensive colocalization of Cx45 and Cx43 immunoreactive signal isclearly apparent at gap junctions. Bar510 mm and applies to both panels.
expression in adult mammalian ventricular myocardium that exhibit synchronized contractions [15,16]. Our databased on the finding that a commercial anti-Cx45 antibody suggest that Cx45 expression is responsible, at least in
2 / 2crossreacts with Cx43 [21]. The present study was per- part, for intercellular coupling in Cx43 hearts. Cx45formed using new, monospecific anti-Cx45 antisera that do protein expression is not, however, upregulated in Cx43-not crossreact with Cx43 in cells overexpressing Cx43 deficient hearts to compensate for diminished or absent[34]. Using this antibody, we observed that Cx45 is present Cx43 protein. Our data also suggest that Cx40 is notin both neonatal and adult mouse ventricles as assessed by upregulated in Cx43-deficient hearts, although Spray et al.immunoblot analysis and quantitative confocal immuno- have reported that Cx40 and possibly Cx45 channels may
2 / 2fluorescence microscopy, but not at the levels reported be present in Cx43 hearts based on characteristicpreviously in the dog [1,4,20]. properties of single channel activity recorded from those
The physiologic role of Cx45 in developing and mature hearts [15].1 / 2hearts has not been fully elucidated. Cx45 expression does, The pattern of Cx43 immunostaining in Cx43 hearts
however, appear to be critical during early cardiogenesis as highlights an interesting difference in the way a limited2 / 2Cx45 embryos do not survive [23,35]. In the adult amount of Cx43 protein is packaged into gap junctions in
1 / 2heart, because Cx45 is present in the conduction system neonatal versus adult Cx43 hearts. In the latter, gap[21], it may play a role in propagation from the AV node to junction number is reduced, yet size remains the same
1 / 2the ventricular muscle. In working ventricular myocardium [32]. In neonatal Cx43 hearts, however, both numberCx45 may play a modulatory role in influencing Cx43 gap and size are reduced. Our data suggest that during matura-junction channel conductances. Based on a large body of tion of Cx43-deficient hearts, cardiac Cx43 gap junctionsevidence in cell lines [28–30] and our new data demon- attain a relatively normal, consistent size in the face ofstrating colocalization of Cx45 and Cx43 in the mouse reduced Cx43 protein expression, whereas immatureheart, Cx43/Cx45 hybrid channels are likely to exist and neonatal gap junctions are smaller in both number and size.function in vivo. Thus, even if present in small amounts Regulatory control of connexin synthesis, packaging,relative to Cx43, Cx45 could influence cell-to-cell com- degradation and turnover in the adult heart may maintainmunication in normal and diseased hearts. an optimal size of gap junctions for efficient coordination
2 / 2Cx43 myocytes are coupled throughout develop- of structure and functional activity. Specifically, the2 / 2ment. Although Cx43 hearts exhibit a conotruncal amount of Cx43 expressed does not determine the size of
2 / 2malformation [31,46–48] and Cx43 mice do not sur- cardiac gap junctions as long as some Cx43 is produced2 / 2 1 / 2vive long after birth, neonatal Cx43 ventricular (i.e., as in the Cx43 heart). Other factors such as the
myocytes grow well in culture and form clusters of cells structural stability imposed by mechanical junctions on the
932 C.M. Johnson et al. / Cardiovascular Research 53 (2002) 921 –935
1 / 1 1 / 2 2 / 2Fig. 8. (A) Immunoblot of two Cx43 , two Cx43 , two Cx43 neonatal ventricular samples and one atrial (Atr) sample probed with anti-Cx40antibody (top). Corresponding Coomassie blue-stained gel is shown below. There are no detectable Cx40 bands in any of the ventricular tissue samples.
1 / 1 1 / 2 2 / 2(B) Summarized immunoblot data demonstrating equal levels (P50.763) of Cx40 expression in Cx43 (n55), Cx43 (n513), and Cx43 (n54)hearts. (C) Fluorescence image of atrial and ventricular tissue stained with anti-Cx40 antibody demonstrating Cx40 expression (bright red punctatestaining) in atrial myocardium and vascular endothelial cells in the ventricle. Bar5100 mm.
sarcolemma of adjacent cells, or localized conductance cell area over which Cx45 signal is distributed is de-requirements may determine gap junction size [41]. creased. That is, the confocal images suggest that the same
Data from the present study demonstrate that although amount of Cx45 protein is distributed over a smaller area1 / 1total ventricular Cx45 protein content is not reduced, the % in Cx43-deficient and null hearts (versus Cx43 hearts).

C.M. Johnson et al. / Cardiovascular Research 53 (2002) 921 –935 933
Fig. 8. (continued)
An alternative interpretation is that there may be a pool of coordinate expression of cardiac connexins undercellular Cx45 protein that is not detected in the confocal pathophysiologic conditions such as heart failure in whichimages from Cx43-deficient and null hearts. Our data Cx43 is downregulated.cannot distinguish between either of these two possibilities
1 / 2or a combination of both. In adult Cx43 hearts, both %cell area occupied by Cx45 immunoreactive signal and AcknowledgementsCx45 gap junction number were reduced significantly(Table 3). This result is not surprising given the fact that We are grateful to Drs. Kiyomasa Nishii and YosaburoCx45 colocalizes with Cx43 in gap junctions, and it Shibata for helpful discussions, sharing protocols and for
1 / 21 / 1 1 / 2 2 / 2suggests that the Cx45 protein expressed in Cx43 and sending us Cx45 , Cx45 and Cx45 embryos
1 / 1Cx43 hearts is distributed to gap junctions that contain used in characterization of the anti-Cx45 antibody used inCx43. That Cx45 gap junction size was also reduced in the present study. This work was supported by NIH/
1 / 2Cx43 hearts raises the interesting possibility of altered NHLBI grants HL50598 and HL58507 (J.E.S.) and1 / 2Cx43/Cx45 stoichiometry in Cx43 hearts. For exam- HL66350 (K.A.Y.).
ple, the pathophysiologic effects of reduced expression of1 / 2Cx43 in Cx43 hearts [9,10,13,17,18] may result not
only from a reduction in the number of homotypic orReferenceshomomeric Cx43 channels, but also from an increase in the
number or composition of Cx45/Cx43 hybrid channels.[1] Kanter HL, Saffitz JE, Beyer EC. Cardiac myocytes express multipleThe latter would be expected to have lower conductances
gap junction proteins. Circ Res 1992;70:438–444.[28–30] and altered gating [49]. [2] Gourdie RG, Severs NH, Green CR et al. The spatial distribution
In summary, Cx43 expression has a marked influence on and relative abundance of gap-junctional connexin40 andthe packaging (number and size) of Cx45 gap junctions. connexin43 correlate to functional properties of components of the
cardiac atrioventricular conduction system. J Cell SciVentricular Cx45 protein redistributes markedly in Cx43-1993;105:985–991.deficient and null hearts. It is not known what regulates
[3] Bastide B, Neyses L, Ganten D et al. Gap junction proteinCx45 and Cx40 expression. Although there is no evidence connexin40 is preferentially expressed in vascular endothelium andfor coordinate upregulation of Cx45 or Cx40 in Cx43- conductive bundles of rat myocardium and is increased underdeficient hearts, this does not exclude the possibility of hypertensive conditions. Circ Res 1993;73:1138–1149.

934 C.M. Johnson et al. / Cardiovascular Research 53 (2002) 921 –935
[4] Kanter HL, Laing JG, Beyer EC, Green KG, Saffitz JE. Multiple Circ Res 1994;75:483–490.connexins colocalize in canine cardiac myocyte gap junctions. Circ [25] Moreno AP, Laing JG, Beyer EC, Spray DC. Properties of gapRes 1993;73:344–350. junction channels formed of connexin45 endogenously expressed in
[5] Gros D, Jarry-Guichard T, Ten Velde I et al. Restricted distribution human hepatoma (SKHep1) cells. Am J Physiol 1995;268:C356–of connexin40, a gap junctional protein, in mammalian heart. Circ C365.
´Res 1994;74:839–851. [26] Elfgang C, Eckert R, Lichtenberg-Frate H et al. Specific permeabili-[6] Van Kempen MJ, Fromaget C, Gros D, Moorman AF, Lamers WH. ty and selective formation of gap junction channels in connexin-
Spatial distribution of connexin43, the major cardiac gap junction transfected HeLa cells. J Cell Biol 1995;129:805–817.protein, in the developing and adult rat heart. Circ Res [27] Barrio LC, Capel J, Jarillo JA, Castro C, Revilla A. Species-specific1991;68:1638–1651. voltage-gating properties of connexin-45 junctions expressed in
[7] Oosthoek PW, Viragh S, Lamers WH, Moorman AFM. Immuno- Xenopus oocytes. Biophys J 1997;73:757–769.histochemical delineation of the conduction system. II. The atrioven- [28] Moreno AP, Fishman GI, Beyer EC, Spray DC. Voltage dependenttricular node and the Purkinje fibers. Circ Res 1993;73:482–491. gating and single channel analysis of heterotypic channels formed
[8] Oosthoek PW, Viragh S, Mayen AEM et al. Immunohistochemical by Cx45 and Cx43. Prog Cell Res 1995;4:405–408.delineation of the conduction system. I. The sinoatrial node. Circ [29] Koval M, Geist ST, Westphale EM. et al. Transfected connexin45Res 1993;73:473–481. alters gap junction permeability in cells expressing endogenous
[9] Guerrero PA, Schuessler RB, Davis LM et al. Slow ventricular connexin43. J Cell Biol 1995;130:987–995.conduction in mice heterozygous for a Cx43 null mutation. J Clin [30] Steiner E, Ebihara L. Functional characterization of canineInvest 1997;99:1991–1998. connexin45. J Membr Biol 1996;150:153–161.
[10] Thomas SA, Schuessler RB, Berul CI et al. Disparate effects of [31] Reaume AG, de Sousa PA, Kulkarni S et al. Cardiac malformationdeficient expression of connexin43 on atrial and ventricular conduc- in neonatal mice lacking connexin43. Science 1995;267:1831–1834.tion: Evidence for chamber-specific molecular determinants of [32] Saffitz JE, Green KG, Kraft WJ, Schechtman KB, Yamada KA.conduction. Circulation 1998;97:686–691. Effects of diminished expression of connexin43 on gap junction
[11] Simon AM, Goodenough DA, Paul DL. Mice lacking connexin40 number and size in ventricular myocardium. Am J Physiolhave cardiac conduction abnormalities characteristic of atrioven- 2000;278:H1662–H1670.tricular block and bundle branch block. Curr Biol 1998;8:295–298. [33] Lecanda F, Towler DA, Ziambaras K et al. Gap junctional com-
¨[12] Hagendorff A, Schumacher B, Kirchhoff S, Luderitz B, Willecke K. munication modulates gene expression in osteoblastic cells. MolConduction disturbances and increased atrial vulnerability in Biol Cell 1998;9:2249–2258.connexin40-deficient mice analyzed by transesophageal stimulation. [34] Van Veen TAB, van Rijen HVM, Jongsma HJ. Electrical conduct-Circulation 1999;99:1508–1515. ance of mouse connexin45 gap junction channels is modulated by
[13] Eloff BC, Lerner DL, Yamada KA et al. High resolution optical phosphorylation. Cardiovasc Res 2000;46:496–510.mapping reveals conduction slowing in connexin43 deficient mice. [35] Kumai M, Nishii K, Nakamura K et al. Loss of connexin45 causes aCardiovasc Res 2001;51:681–690. cushion defect in early cardiogenesis. Development 2000;127:3501–
[14] Morley GE, Vaidya D, Samie FH et al. Characterization of conduc- 3512.tion in the ventricles of normal and heterozygous Cx43 knockout [36] Gossler A, Zachgo J. Gene and enhancer trap screens in ES cellmice using optical mapping. J Cardiovasc Electrophysiol chimeras. In: Joyner AL, editor, Gene targeting. A practical ap-1999;10:1361–1375. proach, New York: Oxford University Press, 1993, pp. 181–227.
[15] Spray DC, Vink MJ, Scemes E et al. Characteristics of coupling in [37] Beardslee MA, Lerner DL, Tadros PN et al. Dephosphorylation andcardiac myocytes and CNS astrocytes cultured from wildtype and intracellular redistribution of ventricular connexin43 during electri-Cx43-null mice. In: Werner R, editor, Gap junctions, Amsterdam: cal uncoupling induced by ischemia. Circ Res 2000;87:656–662.IOS Press, 1998, pp. 281–285. [38] Kwong KF, Schuessler RB, Green KG, Boineau JP, Saffitz JE.
1[16] Johnson CM, Green KG, Kanter EM et al. Voltage-gated Na Differential expression of gap junction proteins in the canine sinuschannel activity and connexin expression in Cx43-deficient node. Circ Res 1998;82:604–612.myocytes. J Cardiovasc Electrophysiol 1999;10:1390–1401. [39] Gourdie RG, Green CR, Severs NJ, Thompson RP. Immunolabelling
[17] Gutstein DE, Morley GE, Tamaddon H et al. Conduction slowing patterns of gap junction connexins in the developing and mature ratand sudden arrhythmic death in mice with cardiac-restricted in- heart. Anat Embryol 1992;185:363–378.activation of connexin43. Circ Res 2001;88:333–339. [40] Peters NS, Severs NJ, Rothery S et al. Spatiotemporal relations
[18] Lerner DL, Yamada KA, Schuessler RB, Saffitz JE. Accelerated between gap junctions and fascia adherens junctions during postnat-onset and increased incidence of ventricular arrhythmias induced by al development of human ventricular myocardium. Circulationischemia in Cx43-deficient mice. Circulation 2000;101:547–552. 1994;90:713–725.
´ ´[19] Alcolea S, Theveniau-Ruissy M, Jarry-Guichard T et al. Downregu- [41] Angst BD, Khan LUR, Severs NJ et al. Dissociated spatial pattern-lation of connexin 45 gene products during mouse heart develop- ing of gap junctions and cell adhesion junctions during postnatalment. Circ Res 1999;84:1365–1379. differentiation of ventricular myocardium. Circ Res 1997;80:88–94.
[20] Davis LM, Kanter HL, Beyer EC, Saffitz JE. Distinct gap junction [42] Saffitz JE, Kanter HL, Green KG, Tolley TK, Beyer EC. Tissue-protein phenotypes in cardiac tissues with disparate conduction specific determinants of anisotropic conduction velocity in canineproperties. J Am Coll Cardiol 1994;24:1124–1132. atrial and ventricular myocardium. Circ Res 1994;74:1065–1070.
[21] Coppen SR, Dupont E, Rothery S, Severs NJ. Connexin45 expres- [43] Beyer EC. Molecular cloning and developmental expression of twosion is preferentially associated with the ventricular conduction chick embryo gap junction proteins. J Biol Chem 1990;265:14439–system in mouse and rat heart. Circ Res 1998;82:232–243. 14443.
[22] Coppen SR, Severs NJ, Gourdie RG. Connexin 45 (a6) expression [44] Delorme B, Dahl E, Jarry-Guichard T et al. Expression pattern ofdelineates an extended conduction system in the embryonic and connexin genes products at the early developmental stages of themature rodent heart. Dev Genet 1999;24:82–90. mouse cardiovascular system. Circ Res 1997;81:423–437.
¨[23] Kruger O, Plum A, Kim J-S et al. Defective vascular development in [45] Delorme B, Dahl E, Jarry-Guichard T et al. Developmental regula-connexin 45-deficient mice. Development 2000;127:4179–4193. tion of connexin 40 gene expression in mouse heart correlates with
[24] Veenstra RD, Wang H-Z, Beyer EC, Brink PR. Selective dye and the differentiation of the conduction system. Dev Dyn 1995;20:358–ionic permeability of gap junction channels formed by connexin45. 371.

C.M. Johnson et al. / Cardiovascular Research 53 (2002) 921 –935 935
[46] Ewart JL, Cohen MF, Meyer RA et al. Heart and neural tube defects [48] Ya J, Erdtsieck-Ernste EBHW, de Boer PAJ et al. Heart defects inin transgenic mice overexpressing the Cx43 gap junction gene. connexin43-deficient mice. Circ Res 1998;82:360–366.Development 1997;124:1281–1292. [49] Elenes S, Martinez AD, Delmar M, Beyer EC, Moreno AP.
[47] Huang GY, Wessels A, Smith BR et al. Alteration in connexin 43 Heterotypic docking of Cx43 and Cx45 connexons blocks fastgap junction gene dosage impairs conotruncal heart development. voltage gating of Cx43. Biophys J 2001;81:1406–1418.Dev Biol 1998;193:32–44.





























