Embed Size (px)
Citation preview

693Research Article
IntroductionConnexin (Cx) proteins are subunits of gap junction channelsthat permit diffusional exchange of ions, secondary messengermolecules and metabolites up to 1000 Da and can serve fortransmission of electrical signals between two contacting cells.Each gap junction channel consists of two hemichannels, theconnexons, which are contributed by two neighboring cells.Each connexon is composed of six connexin protein subunits(Nicholson, 2003). In the mouse genome, 20 connexin genes(Cx) have been identified (Söhl and Willecke, 2003). Theyshow cell type- and differentiation-dependent expressionpatterns, whereby most cell types express more than oneconnexin isoform. Connexins can assemble into homomeric orheteromeric hemichannels. The hemichannels can dock tohemichannels of the same or different connexin compositionin the plasma membrane of apposed cells to form homotypicor heterotypic channels, respectively (Willecke et al., 2002).Gap junction channels composed of different connexinisoforms differ from each other in unitary conductance andpermeability to small molecules (Harris, 2001). This raised thequestion ‘to what extent do the different connexin isoforms
limit or support functional specialization of different celltypes?’.
Of the 20 known mouse connexin genes, connexin43 (Cx43)is most abundantly expressed in different cell types. In mouseheart, Connexin43 protein (Cx43) is found in the workingmyocardium and Purkinje fibers (van Veen et al., 2001). Cx43-deficient mice died shortly after birth due to obstruction of theright ventricular outflow tract of the heart (Reaume et al.,1995). It has been described that the lethality of Cx43-deficientmice could be rescued by Cx32 or Cx40, indicating that Cx43,Cx40 and Cx32 share at least some common functions.However, Cx43 knock-in Cx32 (Cx4332/32) and Cx43 knock-inCx40 (Cx4340/40) mice differed functionally andmorphologically from each other and from wild-type mice(Plum et al., 2000). Cx31 is not expressed in the heart and Cx31deficiency in mice caused transient placentaldysmorphogenesis (Plum et al., 2001). In transfected HeLacells or in RNA-injected Xenopus oocytes, Cx31 hemichannelsdid not form functional heterotypic channels with any otherconnexin, including Cx43 hemichannels, when analyzed bytransfer of microinjected tracer molecules (Elfgang et al.,
In the gastrulating mouse embryo, the gap junction proteinconnexin43 is expressed exclusively in cells derived fromthe inner cell mass, whereas connexin31 is expressed in cellsof the trophoblast lineage. Since connexin43 andconnexin31 do not form heterotypic gap junction channelsin exogenous expression systems, such as HeLa cells andXenopus oocytes, previous studies have suggested that theincompatibility of these two connexins could contribute tothe separation of connexin43-expressing and connexin31-expressing compartments between embryo andextraembryonic tissues at gastrulation, respectively. Thus,we have generated connexin43 knock-in connexin31 mice,in which the coding region of the connexin43 gene wasreplaced by that of connexin31. Interbreeding ofheterozygous connexin43 knock-in connexin31 miceresulted in homozygous connexin43 knock-in connexin31mice, but none of them survived to adulthood. As thesemice were born at the expected Mendelian frequency, we
conclude that the reported incompatibility of connexin43and connexin31 to form heterotypic gap junction channelsdoes not interfere with normal embryonic development.Neonatal homozygous connexin43 knock-in connexin31hearts showed malformation in the subpulmonary outlet ofthe right ventricle, similar to general connexin43-deficientmice. Electrocardiograms of neonatal hearts inhomozygous connexin43 knock-in connexin31 micerevealed significantly low voltage of the QRS complex. Thisis in contrast to previous results from our laboratory whichshowed that replacement of connexin43 by connexin40resulted in morphologically and functionally normalhearts. We conclude that connexin31 cannot functionallyreplace connexin43 during cardiac morphogenesis.
Key words: Gap junction, Connexin, Knock-in mice, Couplingcompartment, Subpulmonary outlet, Low voltage QRS complex
Summary
Connexin31 cannot functionally replace connexin43during cardiac morphogenesis in miceQingyi Zheng-Fischhöfer1, Alexander Ghanem2, Jung-Sun Kim3, Mark Kibschull4, Gaby Schwarz1,Jörg O. Schwab2, James Nagy5, Elke Winterhager4, Klaus Tiemann2 and Klaus Willecke1,*1Institut für Genetik, Universität Bonn, 53117 Bonn, Germany2Medizinische Klinik und Poliklinik II, Universitätsklinikum Bonn, 53105 Bonn, Germany3University of Ulsan, College of Medicine, Seoul, 138-736 Republic of Korea4Institut für Anatomie, Universität Duisburg-Essen, 45122 Essen, Germany5Department of Physiology, University of Manitoba, Canada*Corresponding author (e-mail: [email protected])
Accepted 18 November 2005Journal of Cell Science 119, 693-701 Published by The Company of Biologists 2006doi:10.1242/jcs.02800
Jour
nal o
f Cel
l Sci
ence

694
1995) or by electrical conductance (Yeager and Nicholson,2000). We refer to this as the incompatibility of Cx43 and Cx31hemichannels. During embryonic development, both Cx43 andCx31 are present in the compacted morula and throughout theblastocyst. After implantation, Cx43 protein is detectedexclusively in cells derived from the inner cell mass, whereasCx31 protein is localized in cells of the trophectoderm lineage(Dahl et al., 1996; Reuss et al., 1996). It has been hypothesizedthat the incompatibility of these two connexins mightcontribute to the separation of Cx43-expressing and Cx31-expressing compartments between embryo and extraembryonictissues at gastrulation (Dahl et al., 1996). Using Cx43 knock-in Cx31 mice (Cx4331/31), in which the coding region of theCx43 gene was replaced by that of Cx31, we have nowinvestigated whether the incompatibility between these twoisoforms is necessary for embryonic development and whetherthe loss of Cx43 can be compensated for by Cx31.
ResultsGeneration of floxCx43KI31neo miceTo circumvent possible embryonic lethality, we first generateda mouse line that carries a ‘floxed’ Cx43 coding region, i.e.flanked by loxP recognition sites for the Cre recombinase(Nagy, 2000). The Cx31 coding region was inserted behind thesecond loxP site (Fig. 1A). After crossing the floxed mice withPGK-Cre mice that expressed the Cre recombinase undercontrol of the phosphoglycerate kinase (PGK) promoter, theCx43 coding region was removed by Cre-mediated deletionand Cx31 was expressed under control of the Cx43 promoter.This approach allowed us not only to study the generalreplacement of Cx43 by Cx31, but will also be useful in thefuture for a cell-type-specific replacement by crossing withother mouse lines that express Cre under control of cell-type-specific promoters.
To check whether expression of Cx43 and Cx31 in the
Journal of Cell Science 119 (4)
Fig. 1. Generation and characterization ofCx4331/31 mice. (A) Vector pCx43KI31DNA was transfected into HM-1embryonic stem cells. Homologousrecombination was tested by PCR andSouthern blot analysis. Correctlyrecombined clones were injected intoC57BL/6 blastocysts to generate firstchimeras and in the next generation micecarrying the floxCx43KI31neo allele. Bymeans of Cre activity, the coding region ofconnexin43 and the frt-flanked selectioncassette were deleted, resulting in theCx43KI31 allele. Thus, in mice carryingthis allele, Cx31 is expressed under controlof Cx43 regulatory elements. (B) Southernblot analysis of PstI digested Cx43+/+,Cx43+/31 and Cx4331/31 DNA using theexternal Cx43 probe. A 8 kb fragment isindicative of the Cx43 wild-type allele,whereas a 2.9 kb fragment indicates theCx43KI31 allele. (C) Southern blotanalysis of PstI digested Cx43+/+, Cx43+/31
and Cx4331/31 DNA using the internal Cx43probe. Both Cx43+/+ and Cx43+/31 DNAshowed a wild-type fragment of 8 kb, butCx4331/31 did not. (D) Southern blotanalysis of HindIII digested Cx43+/+,Cx43+/31 and Cx4331/31 DNA using theinternal Cx31 probe. All three genotypesyielded a 12 kb wild-type fragment,whereas Cx43+/31 and Cx4331/31 showed anadditional 5.3 kb recombinant fragment.(E) PCR analysis of different genotypes.Lane 1: 100 bp marker; lane 2: a 2.1 kbamplicon indicates that homologousrecombination has occurred in thefloxCx43KI31neo allele using primer 1(Cx31Ki2) and 2 (Cx43-3�-RO3); lane 3:using primer 3 (Cx43/31fw) and primer 4(Cx43/31rev), the wild-type allele yielded a326 bp DNA fragment, whereasfloxCx43KI31neo allele gave rise to a 375bp fragment due to an additional loxP site;lane 4: after Cre activity, a 736 bp amplicon was generated using primer 3 (Cx43/31fw) and primer 5 (Cx31Ki3rev); lane 5-7: a multiplex PCRwas established using primer 6 (Cx43-3�-HO2), primer 7 (Cx31Ki3) and primer 8 (Cx43-3�-RO4). A 381 bp fragment indicates the wild-typeallele and a 615 bp fragment the knock-in allele. Lane 6: the Cx43+/31 genotype.
Jour
nal o
f Cel
l Sci
ence

695Cx43 knock-in Cx31 mice
floxCx43KI31neo mice (here abbreviated as Cx43flox/flox) wasinfluenced by the loxP sites, the PGK-neomycin cassette or thesilent Cx31 gene, Cx43+/flox mice were crossed with each other.Cx43flox/flox animals were born at the expected Mendelianfrequency. We performed western blot analysis of heart andbrain lysates using anti-Cx43 and anti-Cx31 antibodies.Compared to wild-type animals, the protein level of Cx43 inthe heart was slightly increased in Cx43+/flox or Cx43flox/flox mice(data not shown). By contrast, no significant change of Cx43expression was found in the brain. As in wild-type mice, Cx31was not expressed in brain or heart of Cx43+/flox and Cx43flox/flox
mice (data not shown). Therefore, we concluded thatfloxCx43KI31neo mice resemble wild-type mice with regardto Cx43 or Cx31 expression.
Generation of Cx4331/31 miceTo generate Cx4331/31 mice, in which the coding region of Cx43was replaced by the coding region of Cx31, heterozygous malefloxCx43KI31neo mice (Cx43+/flox) were mated with femalePGK-Cre mice. Under these conditions, Cre activitycommences in the diploid phase of oogenesis (Lallemand etal., 1998). The Cre-mediated recombination led to excision ofthe coding region of Cx43 and the PGK-neomycin cassette.The Cx31 gene was then expressed under control ofendogenous Cx43 gene regulatory elements. Cx43+/31 micewere viable and fertile (Fig. 5). The offspring Cx43+/31 micewere intercrossed. Occurrence of three genotypes Cx43+/+,Cx43+/31 and Cx4331/31 of mice was proved by PCR (Fig. 1E),Southern blot (Fig. 1B-D), western blot (Fig. 2) andimmunofluorescence (Figs 3, 4) analyses.
A multiplex PCR (Fig. 1A) was established for genotypingusing one common reverse primer from the 3� untranslatedregion of Cx43 (primer 8), one forward primer from the codingregion of Cx43 (primer 6) and the other from the coding regionof Cx31 (primer 7). A 381 bp amplicon was derived from thewild-type allele and a 615 bp amplicon was delivered from theknock-in allele (Fig. 1E). In the Southern blot hybridization, theCx43 external probe recognized a 8 kb wild-type fragment anda 2.9 kb knock-in fragment (Fig. 1B). The Cx43 internal probedetected the 8 kb wild-type fragment only in wild-type but notin homozygous mutant DNA (Fig. 1C). The Cx31 internal probelabeled the 12 kb wild-type fragment in all three genotypes andthe additional 5.3 kb knock-in fragment in heterozygous andhomozygous mutants (Fig. 1D). Western blot analyses ofpostnatal day 7 (P7) heart and brain lysates demonstrated thatCx43 protein was absent in the Cx4331/31 mice and Cx31 proteinwas translated in Cx43+/31 and Cx4331/31 mice in a gene dosage-dependent manner (Fig. 2). Immunofluorescence analysis of P0ventricular myocardium confirmed the results of the westernblot analysis: no gap junctional plaques due to Cx43 proteinwere detected in the Cx4331/31 mice, whereas signals of theCx31 protein were found in Cx43+/31 and Cx4331/31 mutants
(Fig. 3). In addition, we performeddouble immunofluorescenceanalysis of Cx43 and Cx31 on theCx43+/31 heart tissue (Fig. 4). Theworking cardiomyocytes of theadult ventricle were extensivelyinterconnected at the intercalateddisk (Fig. 4D,E). Cx43- andCx31-containing plaques were
Fig. 2. Western blot analyses of Cx43 and Cx31 in heart and brainlysates. Lane 1: Cx43+/+; lane 2: Cx43+/31; lane 3: Cx4331/31. Cx43protein was not detected in Cx4331/31 animals (lane 3). Cx31 was notfound in the Cx43+/+ heart and brain (lane 1), whereas it wasexpressed in Cx43+/31 mice and there was nearly double the amountin Cx4331/31 animals (lane 2 and lane 3). Loading of equal amountsof homogenates was verified by re-immunostaining with monoclonalanti-tubulin.
Fig. 3. Immunofluorescence analysisof Cx43 and Cx31 in heart.Cryosections of Cx43+/+ (A,D),Cx43+/31 (B,E) and Cx4331/31 (C,F)hearts at P0 were immunolabeledwith anti-Cx43 (A-C, red), anti-Cx31(D-F, green) and counterstained withpropidium iodide (shown as bluenuclear staining). Cx43 wasexpressed in Cx43+/+ and Cx43+/31
ventricles, but not in Cx4331/31
ventricles. Cx31 was not detected inCx43+/+ hearts, but in Cx43+/31 andCx4331/31 ventricles. Bars, 20 �m.
Jour
nal o
f Cel
l Sci
ence

696
predominantly co-localized (Fig. 4F). By contrast, P0ventricular cardiomyocytes did not show intercalated disks (Fig.4A,B) and the co-localization of Cx43 and Cx31 was low (Fig.4C). Immunofluorescence analysis of Cx4331/31 heart showedregular expression of Cx40 in the atria and vessels.Upregulation of Cx40 was not found in Cx4331/31 heart (datanot shown). Western blot analysis showed that the total amountof Cx40 was not changed in the P0 heart lysate of Cx4331/31
mice (data not shown). By immunofluorescence, Cx45 wasdetected in the conduction system, but not in the workingcardiomyocytes of either Cx43+/+ or Cx4331/31 mice (data notshown). In the western blot analyses, Cx45 was not found to beupregulated in the Cx4331/31 heart lysate (data not shown).
Cx4331/31 mice die postnatally because of adysmorphogenesis of the right ventricular outflow tractTo obtain Cx4331/31 mice, Cx43+/31 mice were intercrossed. Atbirth, we found the expected ratio of homozygous,heterozygous and wild-type mice (Fig. 5). This indicated thatthe embryos developed to term, although Cx31 waspresumably expressed both in the extraembryonic tissue and
the embryo proper (instead of Cx43). However, no Cx4331/31
mice survived to adulthood. 50% of the Cx4331/31 animalssurvived at P0, 30% at P1, none of them lived longer than 10days (Fig. 5). At P0, the average weight of the wild-type micewas 1.4±0.15 g and the weight of Cx4331/31 mice was1.2±0.14 g. Several days later, the living Cx4331/31 animalscould be identified by their lower weight. They did not growas fast as their Cx43+/+ and Cx43+/31 littermates. There was noobvious difference in the phenotype between Cx43+/+ andCx43+/31 mice.
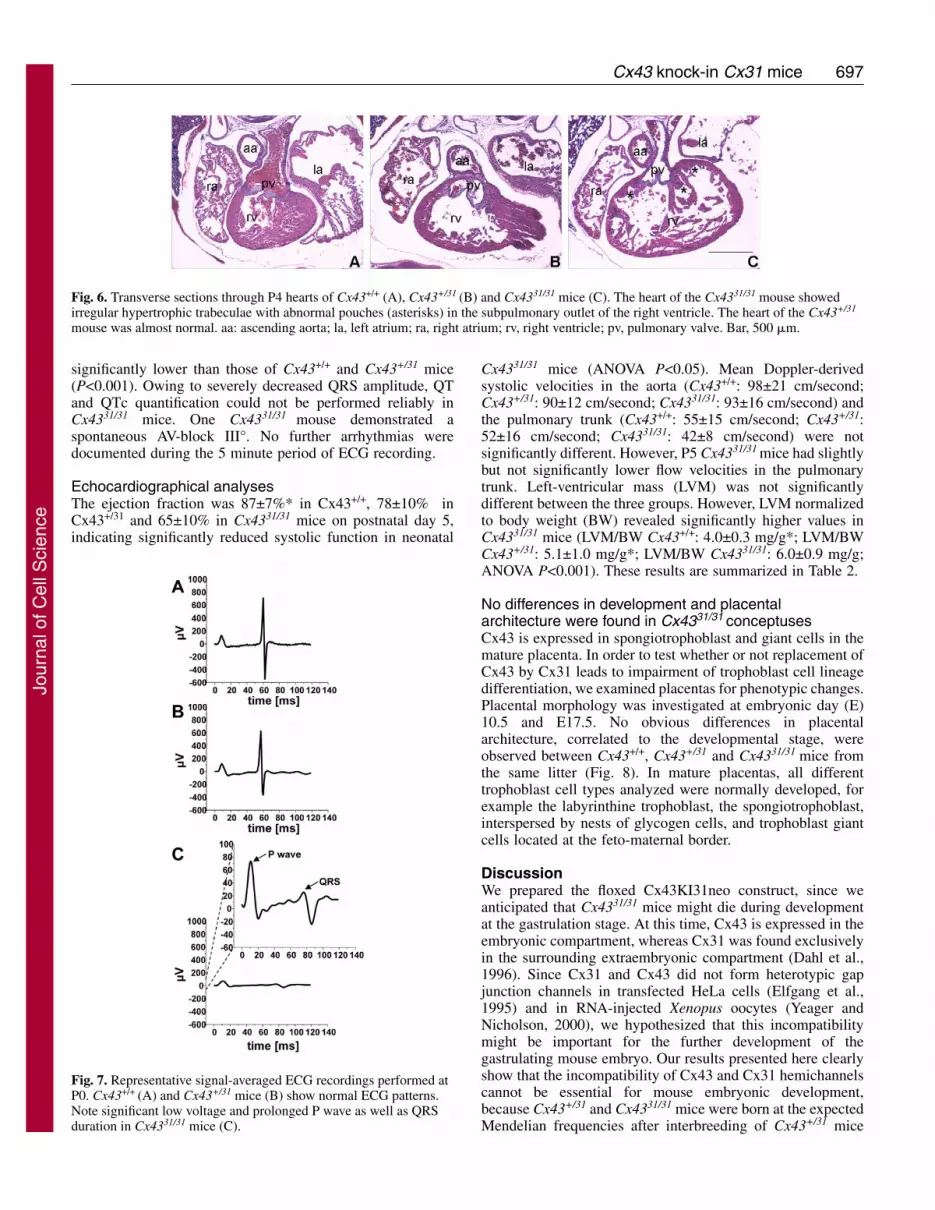
Since Cx43-deficient mice died shortly after birth due to adysmorphogenesis of the right ventricular outflow tract(Reaume et al., 1995), we investigated neonatal Cx4331/31
hearts histologically (Fig. 6). We examined six Cx43+/+ (threeP0, two P4 and one P5), six Cx43+/31 (two P0, two P4 and twoP5) and ten Cx4331/31 (eight P0, one P4 and one P5) hearts. Thepulmonary outlet of all ten Cx4331/31 heart was prominent withirregular hypertrophic trabeculation forming abnormal pouch-like spaces, surrounding the narrow but still open substantiallumen. The morphology of Cx43+/31 hearts was basicallynormal. We conclude that the morphological defect ofCx4331/31 hearts was similar to Cx43-deficient hearts and Cx31could not replace the functions of Cx43 during cardiacmorphogenesis.
Neonatal Cx4331/31 mice show low voltage of the QRScomplex in the electrocardiogramA total of 81 ECG recordings of P0 mice were performed (Fig.7), 23 Cx43+/+, 43 Cx43+/31 and 15 Cx4331/31 mice. Mean heartrate was 362±65 beats per minute* (bpm; asterisk denotesstatistical significance, see Materials and Methods), 325±35bpm and 295±25 bpm, respectively. Thus, it was significantlylower in Cx4331/31 mice than in Cx43+/+ mice (ANOVA,P<0.05). Results of signal-averaged ECG analyses aresummarized in Table 1. Significant prolongation of P wave(P<0.01) and QRS duration (P<0.001) were detected inCx4331/31 mice. The QRS amplitudes of Cx4331/31 mice were
Journal of Cell Science 119 (4)
Fig. 4. Double immunofluorescenceanalysis using monoclonal anti-Cx43 and polyclonal anti-Cx31antibodies. (A-C) Cx43+/31 P0 and(D-F) adult ventricles. Cx43 stainingis shown in red (A,D), Cx31staining in green (B,E). C is anoverlay of A and B; F is an overlayof D and E. Bars, 20 �m.
Fig. 5. Cx4331/31 mice die postnatally. Litters of 10 heterozygousmatings (86 pups) were used in the analysis. Cx4331/31 animals wereborn at the expected Mendelian ratio of 24.4% (21 animals), but diedbefore P10.
Jour
nal o
f Cel
l Sci
ence
697Cx43 knock-in Cx31 mice
significantly lower than those of Cx43+/+ and Cx43+/31 mice(P<0.001). Owing to severely decreased QRS amplitude, QTand QTc quantification could not be performed reliably inCx4331/31 mice. One Cx4331/31 mouse demonstrated aspontaneous AV-block III°. No further arrhythmias weredocumented during the 5 minute period of ECG recording.
Echocardiographical analysesThe ejection fraction was 87±7%* in Cx43+/+, 78±10% inCx43+/31 and 65±10% in Cx4331/31 mice on postnatal day 5,indicating significantly reduced systolic function in neonatal
Cx4331/31 mice (ANOVA P<0.05). Mean Doppler-derivedsystolic velocities in the aorta (Cx43+/+: 98±21 cm/second;Cx43+/31: 90±12 cm/second; Cx4331/31: 93±16 cm/second) andthe pulmonary trunk (Cx43+/+: 55±15 cm/second; Cx43+/31:52±16 cm/second; Cx4331/31: 42±8 cm/second) were notsignificantly different. However, P5 Cx4331/31 mice had slightlybut not significantly lower flow velocities in the pulmonarytrunk. Left-ventricular mass (LVM) was not significantlydifferent between the three groups. However, LVM normalizedto body weight (BW) revealed significantly higher values inCx4331/31 mice (LVM/BW Cx43+/+: 4.0±0.3 mg/g*; LVM/BWCx43+/31: 5.1±1.0 mg/g*; LVM/BW Cx4331/31: 6.0±0.9 mg/g;ANOVA P<0.001). These results are summarized in Table 2.
No differences in development and placentalarchitecture were found in Cx4331/31 conceptusesCx43 is expressed in spongiotrophoblast and giant cells in themature placenta. In order to test whether or not replacement ofCx43 by Cx31 leads to impairment of trophoblast cell lineagedifferentiation, we examined placentas for phenotypic changes.Placental morphology was investigated at embryonic day (E)10.5 and E17.5. No obvious differences in placentalarchitecture, correlated to the developmental stage, wereobserved between Cx43+/+, Cx43+/31 and Cx4331/31 mice fromthe same litter (Fig. 8). In mature placentas, all differenttrophoblast cell types analyzed were normally developed, forexample the labyrinthine trophoblast, the spongiotrophoblast,interspersed by nests of glycogen cells, and trophoblast giantcells located at the feto-maternal border.
DiscussionWe prepared the floxed Cx43KI31neo construct, since weanticipated that Cx4331/31 mice might die during developmentat the gastrulation stage. At this time, Cx43 is expressed in theembryonic compartment, whereas Cx31 was found exclusivelyin the surrounding extraembryonic compartment (Dahl et al.,1996). Since Cx31 and Cx43 did not form heterotypic gapjunction channels in transfected HeLa cells (Elfgang et al.,1995) and in RNA-injected Xenopus oocytes (Yeager andNicholson, 2000), we hypothesized that this incompatibilitymight be important for the further development of thegastrulating mouse embryo. Our results presented here clearlyshow that the incompatibility of Cx43 and Cx31 hemichannelscannot be essential for mouse embryonic development,because Cx43+/31 and Cx4331/31 mice were born at the expectedMendelian frequencies after interbreeding of Cx43+/31 mice
Fig. 6. Transverse sections through P4 hearts of Cx43+/+ (A), Cx43+/31 (B) and Cx4331/31 mice (C). The heart of the Cx4331/31 mouse showedirregular hypertrophic trabeculae with abnormal pouches (asterisks) in the subpulmonary outlet of the right ventricle. The heart of the Cx43+/31
mouse was almost normal. aa: ascending aorta; la, left atrium; ra, right atrium; rv, right ventricle; pv, pulmonary valve. Bar, 500 �m.
Fig. 7. Representative signal-averaged ECG recordings performed atP0. Cx43+/+ (A) and Cx43+/31 mice (B) show normal ECG patterns.Note significant low voltage and prolonged P wave as well as QRSduration in Cx4331/31 mice (C).
Jour
nal o
f Cel
l Sci
ence

698
(Fig. 5). There might be several reasons why the separationbetween embryonic and placental lineage might not have anyeffect in our experimental system. For example, the separationbetween the embryo and the surrounding extraembryonictissues at gastrulation might have other explanations, such asdiscrete tissue masses that do not establish gap-junctionalcommunication across an extracellular matrix barrier or,alternatively, the abolition of the separation does not impair thedevelopment of the two cell lineages.
Cx4331/31 newborns showed a significantly reduced weightcompared to their Cx43+/+ and Cx43+/31 littermates. The weightof embryos or newborns has not been analyzed so far for Cx43–/–
mice (Reaume et al., 1995), but recently Kibschull et al.(Kibschull et al., 2005) showed for the Cx31/Cx43 doubleknockout mouse that at 17.5 dpc the Cx31+/–/Cx43–/– embryoswere also significantly smaller compared to theirCx31+/–/Cx43+/– and Cx31+/–/Cx43+/+ littermates. Neither theweight nor the morphology of the Cx31/Cx43 double knockoutplacentas appeared to be affected by Cx43 expression. Inaccordance to this data, the Cx4331/31 placentas also did not showany morphological changes compared to wild-type littermates.Therefore, we assume that the weight of the Cx4331/31 embryosis unlikely to have been influenced by placental development orfunction. The reduced weight of the Cx4331/31 newborns isprobably the result of the loss of Cx43 in the embryo itself, ashas been discussed for the Cx31/Cx43 double knockout mouse(Kibschull et al., 2005). Since the replacement of Cx43 by Cx31did not lead to any obvious impairment in embryogenesis andplacental development, we focused our characterization ofCx4331/31 mice on the cardiac phenotype.
Journal of Cell Science 119 (4)
Table 1. ECG parameters of 23 Cx43+/+, 43 Cx43+/31 and 15 Cx4331/31 mice at P0Heart rate
Genotype (1/min) P (ms) PQ (ms) QRS (ms) QRS (�V) QT (ms) QTc (ms)
Cx43+/+ 362±65 11±2 54±7 11±2 1300±418 25±5 18±3Cx43+/31 325±35 12±2 59±11 12±2 1096±390 29±6 21±4Cx4331/31 295±25* 19±2** 63±10 28±7*** 188±77*** n.d. n.d.
Durations and intervals are given as group means ± s.e.m.; *P<0.05; **P<0.01; ***P<0.001; n.d., evaluation not possible because voltage was too low.
Table 2. Echocardiographic data from five Cx43+/+, sixteenCx43+/31 and five Cx4331/31 mice at P5
Cx43+/+ Cx43+/31 Cx4331/31 ANOVA
Ejection fraction (%) 87±7* 78±10 65±10 P<0.05Vmean Aorta (cm/second) 98±21 90±12 93±16 n.s.Vmean Pulmonary artery 55±15 52±16 42±8 n.s.
(cm/second)LVM/BW (mg/g) 4.0±0.3*** 5.1±1.0*** 6.0±0.9 P<0.001
Vmean, mean systolic velocity; LVM, left ventricular mass; BW, bodyweight; n.s., not significant (P>0.05).
Fig. 8. Placental development in Cx4331/31 conceptuses. HE-stained paraffin sections of Cx43+/+ (A,C) and Cx4331/31 (B,D) placentas at E17.5.No differences in development and placental architecture were found. All trophoblast cell types [labyrinth (L); spongiotrophoblast (S);glycogen cells (G)] are normal in Cx4331/31 placenta compared with Cx43+/+ controls. Bars, 600 �m (A,B); 150 �m (C,D).
Jour
nal o
f Cel
l Sci
ence

699Cx43 knock-in Cx31 mice
Our results show that Cx31 cannot replace the essential roleof Cx43 during cardiac morphogenesis. Mice lacking Cx43 dieshortly after birth from an obstruction of the right ventricularoutflow tract (Reaume et al., 1995). In Cx4331/31 mice, the rightventricular outflow tract is still open, but its lumen is so narrowthat the supply of blood is likely to be limited. As aconsequence, all of them died within ten days (Fig. 5). Bycontrast, Cx4340/40 and Cx4332/32 mice can survive to adults,although 40% of them died during the first 3 weeks after birthand most of them were infertile (Plum et al., 2000). Thus, Cx40and Cx32 can replace Cx43 during cardiac development, butCx31 cannot. Our results fit into the general scheme, that thedifferent connexin isoforms can replace each other in certaintissues but fulfill unique essential functions in other tissues(Plum et al., 2000; White et al., 2002; Alcolea et al., 2004).
The reasons, at the molecular level, for the malformation ofhearts during development of general Cx43-deficient mice arestill a matter of debate. The hearts of mice with cardiac-restricted inactivation of Cx43 by use of the Cre/loxP systemare structurally normal (Gutstein et al., 2001). Studies oftransgenic mice exhibiting gain or loss of Cx43 suggest thatthe migration rate of cardiac neural crest cells is increased withoverexpression of Cx43 and decreased with deficiency of Cx43(Lo et al., 1999). Additional studies have shown involvementof another extracardiac cell population, the proepicardial cellsin these cardiac anomalies (Li et al., 2002; Wei et al., 2004).Cx4331/31 mice show similar cardiac malformations as thegeneral Cx43-deficient mice. The hearts of Cx4332/32 mice alsosuggest a similar morphologic defect, but to a much lesserextent. This underlines that the special properties of Cx43 leadto normal morphogenesis of the heart.
Functionally, replacement of Cx43 by Cx31 in the heartleads to low voltage of the QRS complex, and prolonged P andQRS duration in the electrocardiogram. Recently, Danik et al.(Danik et al., 2004) generated a heart-specific Cx43-deficientsubline (O-CKO mice) in which cardiac Cx43 expressiondecreased progressively throughout the ventricularmyocardium. From 25 to 45 days, the number ofimmunofluorescent clusters in the O-CKO hearts decreasedfrom 67% of control to 34% and the percentage area from 59%to 30%. The QRS amplitude of O-CKO mice showed an age-dependent decrease, which closely paralleled the loss of Cx43expression. In order to determine the mechanism of diminishedQRS amplitude, the authors later used voltage-sensitive dyesand high resolution optical mapping techniques and found thatreduced intercellular coupling leads to paradoxical propagationacross Purkinje-ventricular junctions and aberrant myocardialactivation (Morley et al., 2005). The diminution of the QRSamplitude seems to be related to the pattern of ventricularexcitation. Compared with O-CKO mice, in Cx43+/31 andCx4331/31 hearts Cx43 is replaced by Cx31. Doubleimmunofluorescence using anti-Cx43 and anti-Cx31 showed inthe ventricle of Cx43+/31 mice at P0 that Cx43- and Cx31-containing plaques were not co-localized, whereas in the adultCx+/31 ventricle, Cx43 and Cx31 were predominantly co-localized in intercalated disks (Fig. 4). Since the ECG ofCx43+/31 mice was normal, we assume that the presence ofCx31 plaques does not lead to an increase in impulsepropagation across the Purkinje-ventricular junction. Thereduction of the QRS amplitude in the Cx4331/31 heart ispossibly due to deletion of Cx43 and the inability of Cx31 to
function like Cx43 in the myocardium. Although Cx43 is thepredominant connexin expressed by ventricular and atrialcardiomyocytes in wild-type mice, Cx45 is also expressed atvery low levels (Severs et al., 2004). The total amount of Cx45in the Cx4331/31 heart was not upregulated, as determined bywestern blot analysis (data not shown).
The relationship between QRS duration and uncoupling dueto Cx43-deletion is controversial in the literature. A modestincrease in QRS duration was detected only at 25 days in theO-CKO mice. At 35 and 45 days of age, however, there wasno significant difference in the QRS duration between the CKOand control mice (Danik et al., 2004). Our data are consistentwith the results of Eckardt et al. (Eckardt et al., 2004) whoshowed in inducible Cx43Cre-Er(T)/fl mice that ventricularactivation was significantly delayed after deletion of Cx43, asindicated by significant prolongation of the QRS complex.Prolongation of the P wave was found in Cx40-deficient mice(Kirschhoff et al., 1998; Simon et al., 1998). Gap junctionchannels among atrial myocytes contain abundant Cx40, co-localized with Cx43 (Vozzi et al., 1999; Severs et al., 2001).An increase in P wave has not been shown in conditional Cx43-deficient mice (Danik et al., 2004; Eckardt et al., 2004). TheCx4331/31 mice exhibited significant prolongation of the Pwave. This might be due to the different level of atrial Cx43 indifferent mouse lines. Atrial expression of Cx43 in the O-CKOhearts at 45 days of age was not changed in immunoblotanalyses (Danik et al., 2004). By contrast, no Cx43 protein wasdetected in the Cx4331/31 heart, either by western blot (Fig. 2)or by immunofluorescence (Fig. 3) analyses. Directcompensation of Cx43 deletion by upregulation of Cx40 wasnot found in Cx4331/31 heart, similar to O-CKO mice (Danik etal., 2004) and inducible Cx43Cre-Er(T)/fl mice (Eckhardt et al.,2004).
Quantitative echocardiography of neonatal mice on P0 couldnot be performed in this study because of the small size of thehearts. On P5, echocardiography revealed an impaired systolicfunction in surviving Cx4331/31. In these animals, meanDoppler-derived systolic velocities in aorta and pulmonarytrunk showed no significant differences between the genotypes.Assessment of Doppler-derived flow velocities within thestenotic outflow tract was not possible at this age. Slightlylower velocities measured in the putative post-stenoticpulmonary trunk of Cx4331/31 mice could be the consequenceof right-ventricular outflow tract stenosis.
In recent years, several heart-specific conditional Cx43-deficient mice have been generated (Gutstein et al., 2001;Eckardt et al., 2004; van Rijen et al., 2004; Danik et al., 2004;Gros et al., 2004). In summary, 80-95% decrease of Cx43expression resulted in delayed ventricular activation,arrhythmia and sudden death. Cx4331/31 mice died before P10.At this young age, telemetric ECGs could not be measured.Thus, we do not know the ECG shortly before death. Since wecan induce cardiac-specific replacement of Cx43 with Cx31 inadult mice by breeding floxCx43KI31neo mice withCx43Cre-ER(T)/+ mice, we can study the consequence of thisreplacement also in adult hearts in the future. Furthermore, thefloxCx43KI31 mice that were generated in this work could bebred with mice that express the Cre recombinase cell typespecifically, in order to study the functional consequence ofreplacement of Cx43 by Cx31 in other cell types. We assumethat the functional diversity of different connexin isoforms in
Jour
nal o
f Cel
l Sci
ence

700
different cell types is probably due to specific regulatoryfeatures of these proteins, such as permeability characteristics,posttranslational modifications, specific binding proteins or acombination of these differences.
Materials and MethodsGeneration of miceA fragment of genomic 129/Sv mouse DNA spanning about 12 kb of the Cx43 locuswas isolated as previously described (Plum et al., 2000) from a recombinant lamdaphage library (Stratagene, La Jolla, CA). The Cx43KI31 construct (Fig. 1A) wasembedded in a pBluescriptII SK+ vector backbone (Stratagene). A 5.3 kb SacI-HincII fragment and a 1.6 kb ClaI-AvaI fragment were used as 5� and 3� homologousregions, respectively. The first loxP site was inserted into the HincII site, 188 bpupstream of the Cx43 start codon. A 160 bp DNA fragment harboring a SV40poly(A) signal was inserted into the ClaI site 26 bp downstream of the Cx43 stopcodon, followed by a 1.95 kb phosphoglycerate kinase (PGK) promoter-neomycinselection marker flanked by two frt sites and the second loxP site (Meyers et al.,1998). Between the second loxP site and the 3� homologous region of Cx43, severalDNA fragments were inserted: a 188 bp fragment upstream of the Cx43 start codonincluding the endogenous splice acceptor site, the 813 bp coding region of Cx31and the following 80 bp downstream of the Cx31 stop codon. In order to generatea NcoI site, a cytosine residue was inserted two nucleotides upstream of the Cx43start codon. The final targeting vector pCx43KI31 was analyzed by restrictionmapping and partial sequencing. The functions of loxP sites and frt sites wereverified by transformation of Cre or Flp recombinase-expressing E. coli bacteria(Buchholz et al., 1996).
HM-1 embryonic stem cells were transfected with 300 �g DNA of the targetingvector pCx43KI31, linearized upstream of the 5� homologous region by NotI andselected by 350 �g G418 (Sigma, St Louis, MO) per ml of medium as describedpreviously (Magin et al., 1992; Degen et al., 2004). Resistant clones were analyzedfor homologous recombination by PCR and Southern blot hybridization. Correctlyrecombined ES clones were injected into C57BL/6 blastocysts to generate chimeras,as described by Nagy et al. (Nagy et al., 2003). For germline transmission of themutated allele, the chimeras were crossed to C57BL/6 mice. Male heterozygousoffspring, designated as floxCx43KI31neo, were further crossed to female PGK-Cre mice (Lallemand et al., 1998) to generate the final allele Cx43KI31. All analyseswere carried out on a mixed 129/Ola/C57BL/6 genetic background using littermatesas controls.
Mice were kept under standard housing conditions with a fixed 12 hours light:12hours dark cycle, and food as well as water ad libitum. All experiments were carriedout in accordance with the German law for animal welfare and with permission fromthe local state authorities.
Genotyping of ES cells and mouse tissuesG418 resistant ES clones were prepared and processed for two PCR analyses (Fig.1A,E). In the first PCR, recombination at the 3� homologous site was examinedusing a Cx31-coding-region-specific upstream primer (primer 1: 5�-GTG AGA TCTGCT ACC TCA TCT TCC-3�) and a 3� downstream primer external to the targetingvector (primer 2: 5�-GCA GCA ACA TAC CAT TGC ACA GAG-3�). A 2.1 kbamplicon was indicative of homologous recombination. In the second PCR, theexistence of the 5� located loxP site was verified using the primers: primer 3 (5�-GCA CTT GGT AGG TAG AGC CTG TCA GGT C-3�) and primer 4 (5�´-CTCCAG TCA CCC ATG GTC TGG GCA CCT C-3�). The wild-type allele yielded a326 bp DNA fragment, whereas the mutated allele gave a 375 bp fragment becauseof the first loxP site.
For genomic Southern blot hybridization, DNA from PCR-positive clones wasdigested by XbaI. A 550 bp AvaI/AvaI fragment outside the 3� homologous regionof Cx43 served as external probe. The Cx43 internal probe was a 762 bp HincII/ClaIfragment of the coding region. In addition, a Cx31 internal probe (544 bp, PstI/PstI)was used. After XbaI digestion and hybridization to the Cx43 external probe, theCx43 wild-type allele yielded a 10 kb fragment compared to a 6.4 kb fragment ofmutated allele. The Cx43 internal probe detected a 10 kb fragment of the wild-typeallele and a 4 kb fragment of the mutated allele. The Cx31 internal probe hybridizedto a 5.3 kb fragment of wild-type and a 6.4 kb fragment of the recombined allele.
After action of the Cre recombinase, the deletion of the Cx43 coding region andPGK-neo cassette was analyzed by using primer 3 (5�-GCA CTT GGT AGG TAGAGC CTG TCA GGT C-3�) and primer 5 (5�-CTA AAC AAG TAG GTC CACCAC AGG-3�). The PCR yielded a 736 bp amplicon covering the junction betweenthe intron of Cx43 and the coding region of Cx31. For detection of the PGK-Cretransgene, the primers up-pgk1 (5�-GCT GTT CTC CTC TTC CTC ATC TCC-3�)and IntCre_rev (5�-TCC ATG AGT GAA CGA ACC TGG TCG-3�) were used,generating a 500 bp amplicon.
For routine genotype analysis, genomic DNAs from tail tips were used for PCR,including three primers: primer 6 (coding region of Cx43; 5�-CGC AAT TAC AACAAG CAA GCC AGC- 3�), primer 8 (3� untranslated region of Cx43; 5�-CGC CTCATT ACT GAG GTT GTT GAG-3�) and primer 7 (coding region of Cx31; 5�-CCT
GTG GTG GAC CTA CTT GTT TAG-3�) that generated a 381 bp amplicon of wild-type and a 615 bp amplicon of the knock-in allele.
Southern blot analysisFor Southern blot hybridization of wild-type (Cx43+/+), heterozygous (Cx43+/31) andhomozygous (Cx4331/31) Cx43 knock-in Cx31 mice, genomic DNA was preparedfrom neonatal kidneys and digested with PstI for the Cx43 external and internalprobe, and HindIII for the Cx31 internal probe. After digestion, genomic DNA wasfractionated on 0.7% agarose gels and transferred onto nylon membrane(HybondTM-N+; Amersham Biosciences, UK). The Cx43 external probe (487 bpoutside the 3� homologous region), Cx43 internal probe (654 bp of Cx43 codingregion) and Cx31 internal probe (508 bp of Cx31 coding region) were synthesizedusing a PCR DIG Synthesis Kit (Roche diagnostics GmbH, Penzberg, Germany).Hybridization, washing and detection were performed using DIG Prime DNALabeling and Detection Starter Kit II (Roche Diagnostics GmbH, Penzberg,Germany) according to instructions provided by the manufacturer.
Western blot analysisMouse hearts and brains were dissected, immediately frozen on dry ice and storedat –70°C until further use. Homogenized tissue was taken up in protein lysis buffer(60 mM Tris HCl pH 7.4 and 3% sodium dodecyl sulphate), supplemented withprotease inhibitor ‘Complete’ (Roche, Mannheim, Germany) and sonicated threetimes for 10 seconds on ice. Protein concentration was determined using thebicinchoninic acid protein assay (Sigma, Taufkirchen, Germany). 50 �g proteinwere separated on 12.5% SDS-polyacrylamide gels and electroblotted onnitrocellulose membranes (Hybond ECL Biosciences, Bucks, UK) for 45 minutesat 100 V. Membranes were blocked for 1 hour with WBWP buffer (20 mM Tris-HCl pH 7.4, 150 mM NaCl, 0.1% Tween 20) and 5% milk powder (w/v) for anti-Cx43 and with Roti-Block (Roth, Karlsruhe, Germany) for anti-Cx31. Afterwardsthe membranes were incubated with rabbit anti-Cx43 (1:4000) (Traub et al., 1994)and rabbit anti-Cx31 (1:1000; Zymed, San Francisco, CA) overnight at 4°C. Anti-Cx43 was diluted in WBWP buffer and anti-Cx31 was diluted in Roti-Block. Afterincubation, the membranes were washed for 30 minutes in WBWP buffer (anti-Cx43) or in phosphate-buffered saline (PBS) plus 0.1% Tween 20 (anti-Cx31),followed by incubation with anti-rabbit horseradish peroxidase-conjugatedsecondary antibodies (1: 30000; Dianova, Hamburg, Germany) for 1 hour at roomtemperature. Membranes were washed as described before and incubated with anECL chemiluminescence detection system (Amersham Biosciences, Freiburg,Germany). In order to prove that equal amounts of protein were loaded in all lanes,the immunoblots were stripped using RestoreTM Western Blot Stripping (Pierce,Rockford, IL) and incubated with monoclonal anti-�-tubulin (1:10.000; Sigma,Taufkirchen, Germany) for 1 hour at room temperature. After washing, themembranes were incubated with anti-mouse horseradish peroxidase-conjugatedsecondary antibodies (1: 30.000; Dianova, Hamburg, Germany), followed by ECLdetection. Monoclonal anti-Cx45 (Chemicon, Hampshire, UK) and polyclonal anti-Cx40 (Alpha Diagnostic, San Antonio, TX) were used as described for anti-Cx43.
Immunofluorescence analysisCryostat sections (10 �m) of hearts were stained with the following antibodies:mouse anti-Cx43 (1:1000, Tytomed, Berlin, Germany) and rabbit anti-Cx31(1:1000, Zymed). Cryostat sections were fixed in 4% paraformaldehyde for 5minutes, blocked with 4% goat serum in TBST buffer (50 mM Tris pH 7.6, 0.257M NaCl, 0.3% Triton-X-100) and incubated with antibodies diluted in blockingreagent overnight at 4°C. Afterwards, sections were washed 3 times in TBST bufferand incubated with Alexa Fluor 488-conjugated goat anti-rabbit immunoglobulin(1:1000; MoBiTech, Goettingen, Germany) and Cy5-conjugated goat anti-mouseimmunoglobulin (1:1000; Dianova, Hamburg) for 1 hour at room temperature.Nuclei were stained with propidium iodide. Slices were mounted with the aqueousmounting medium Permafluor (Beckman Coulter, Marseille, France) and analyzedusing an Zeiss confocal microscope (LSM 510). Polyclonal anti-Cx45 was fromUlrike Janssen-Bienhold (Oldenburg, Germany). Cx40 was detected using apolyclonal antibody (Alpha Diagnostic).
Histological analysisFor morphological analyses, hearts from postnatal days (P)0, P4 and P5 mice andplacentas from E10.5 and E17.5 conceptuses were fixed in 4% paraformaldehydein PBS for at least 2 days at 4°C, dehydrated and embedded in paraffin. Sectionswere stained with Hematoxylin and Eosin (HE) and mounted in Entellan (Merck,Darmstadt, Germany).
ECG recording of neonatal miceECG recordings were obtained on P0 from 81 neonates of both genders weighingbetween 1 and 1.4 g (1.3±0.2 g). Mice were fixed on a heating pad warmed tomaintain body temperature. A surface 6-lead ECG was acquired digitally by meansof a multichannel amplifier (PowerLabTM System, ADInstruments, Milford, MA).ECG channels were amplified, filtered between 10 and 100 Hz and sampled at arate of 1 kHz per channel. Heart rate, P-wave duration, PR interval, QRS duration,QRS amplitude and QT interval were determined off-line as described recently
Journal of Cell Science 119 (4)
Jour
nal o
f Cel
l Sci
ence

701Cx43 knock-in Cx31 mice
(Maass et al., 2004). Sums of QRS voltage were calculated as described previously(Kishimoto et al., 1984). The QT interval was rate-corrected (QTc) according toMitchell et al. (Mitchell et al., 1998).
Echocardiography of neonatal miceHigh resolution mouse echocardiography was performed in 26 mice (five Cx43+/+,sixteen Cx43+/31 and five Cx4331/31) on P5 using a commercially available ultrasoundsystem (HDI 5000, Philips-Ultrasound, Bothell, WA) equipped with a 15 MHzlinear array transducer. Volumetric analysis of left ventricular (LV) structures wasperformed to assess mass and function (Tiemann et al., 2003; Ghanem et al., 2005).Structures that are known to be prone to congenital defects in Cx43-deficient mice,in particular, the right-ventricular outflow tract, were investigated carefully by B-mode and pw-Doppler. Additionally, indirect signs of congestive heart failure orpulmonary hypertension (Doppler-derived velocities of the aorta and pulmonarytrunk) were assessed.
Statistical analysisParameters were compared between the three genotypes by means of one-wayANOVA along with post-hoc Tuckey-Kramer Multiple Comparisons Test. P values<0.05 for ANOVA were considered as significant. Statistical significance of post-hoc analyses of Cx43+/31 or Cx43+/+ versus Cx4331/31 mice are defined as values ofP<0.05 (*), P<0.01 (**), and P<0.001 (***). For comparisons of two groups, 2-sided unpaired Student’s t-test was performed. P values <0.05 were considered assignificant.
We gratefully acknowledge the technical assistance of Ina Fiedler.We thank Joachim Degen and Dominik Eckardt as well as MariaKreuzberg for helpful discussions. Work in the Bonn laboratory wassupported by the German Research Association (Wi270/25-1,2 andthrough the research group on keratinocytes at Bonn University) toK.W.
ReferencesAlcolea, S., Jarry-Guichard, T., de Bakker, J., Gonzalez, D., Lamers, W., Coppen,
S., Barrio, L., Jongsma, H., Gros, D. and van Rijen, H. (2004). Replacement ofconnexin40 by connexin45 in the mouse: impact on cardiac electrical conduction. Circ.Res. 94, 100-109.
Buchholz, F., Angrand, P. O. and Stewart, A. F. (1996). A simple assay to determinethe functionality of Cre or FLP recombination targets in genomic manipulationconstructs. Nucleic Acids Res. 24, 3118-3119.
Dahl, E., Winterhager, E., Reuss, B., Traub, O., Butterweck, A. and Willecke, K.(1996). Expression of the gap junction proteins connexin31 and connexin43 correlateswith communication compartments in extraembryonic tissues and in the gastrulatingmouse embryo, respectively. J. Cell Sci. 109, 191-197.
Danik, S. B., Liu, F., Zhang, J., Suk, H. J., Morley, G. E., Fishman, G. I. and Gutstein,D. E. (2004). Modulation of cardiac gap junction expression and arrhythmicsusceptibility. Circ. Res. 95, 1035-1041.
Degen, J., Meier, C., Van Der Giessen, R. S., Söhl, G., Petrasch-Parwez, E., Urschel,S., Dermietzel, R., Schilling, K., De Zeeuw, C. I. and Willecke, K. (2004).Expression pattern of lacZ reporter gene representing connexin36 in transgenic mice.J. Comp. Neurol. 473, 511-525.
Eckardt, D., Theis, M., Degen, J., Ott, T., van Rijen, H. V., Kirchhoff, S., Kim, J. S.,de Bakker, J. M. and Willecke, K. (2004). Functional role of connexin43 gap junctionchannels in adult mouse heart assessed by inducible gene deletion. J. Mol. Cell Cardiol.36, 101-110.
Elfgang, C., Eckert, R., Lichtenberg-Frate, H., Butterweck, A., Traub, O., Klein, R.A., Hülser, D. F. and Willecke, K. (1995). Specific permeability and selectiveformation of gap junction channels in connexin-transfected HeLa cells. J. Cell Biol.129, 805-817.
Ghanem, A., Röll, W., Hashemi, T., Dewald, O., Djoufack, P. C., Fink, K. B.,Schrickel, J., Lewalter, T., Lüderitz, B. and Tiemann, K. (2005). Echocardiographicassessment of left ventricular mass in neonatal and adult mice–accuracy of differentechocardiographic methods. Echocardiography (in press).
Gros, D., Dupays, L., Alcolea, S., Meysenm, S., Miquerol, L. and Theveniau-Ruissy,M. (2004). Genetically modified mice: tools to decode the functions of connexins inthe heart-new models for cardiovascular research. Cardiovasc. Res. 62, 299-308.
Gutstein, D. E., Morley, G. E., Tamaddon, H., Vaidya, D., Schneider, M. D., Chen,J., Chien, K. R., Stuhlmann, H. and Fishman, G. I. (2001). Conduction slowing andsudden arrhythmic death in mice with cardiac-restricted inactivation of connexin43.Circ. Res. 88, 333-339.
Harris, A. (2001). Emerging issues of connexin channels: biophysics fills the gap. Q. Rev.Biophys. 34, 325-472.
Kibschull, M., Magin, T. M., Traub, O. and Winterhager, E. (2005). Cx31 and Cx43double-deficient mice reveal independent functions in murine placental and skindevelopment. Dev. Dyn. 233, 853-863.
Kirchhoff, S., Nelles, E., Hagendorff, A., Kruger, O., Traub, O. and Willecke, K.(1998). Reduced cardiac conduction velocity and predisposition to arrhythmias inconnexin40-deficient mice. Curr. Biol. 8, 299-302.
Kishimoto, C., Matsumori, A., Ohmae, M., Tomioka, N. and Kawai, C. (1984).Electrocardiographic findings in experimental myocarditis in DBA/2 mice: complete
atrioventricular block in the acute stage, low voltage of the QRS complex in the subacutestage and arrhythmias in the chronic stage. J. Am. Coll. Cardiol. 3, 1461-1468.
Lallemand, Y., Luria, V., Haffner-Krausz, R. and Lonai, P. (1998). Maternallyexpressed PGK-Cre transgene as a tool for early and uniform activation of the Cre site-specific recombinase. Transgenic Res. 7, 105-112.
Li, W. E., Waldo, K., Linask, K. L., Chen, T., Wessels, A., Parmacek, M. S., Kirby,M. L. and Lo, C. W. (2002). An essential role for connexin43 gap junctions in mousecoronary artery development. Development 129, 2031-2042.
Lo, C. W., Waldo, K. L. and Kirby, M. L. (1999). Gap junction communication and themodulation of cardiac neural crest cells. Trends Cardiovasc. Med. 9, 63-69.
Maass, K., Ghanem, A., Kim, J. S., Saathoff, M., Urschel, S., Kirfel, G., Grummer,R., Kretz, M., Lewalter, T., Tiemann, K. et al. (2004). Defective epidermal barrierin neonatal mice lacking the C-terminal region of connexin43. Mol. Biol. Cell 15, 4597-4608.
Magin, T. M., McWhir, J. and Melton, D. W. (1992). A new mouse embryonic stemcell line with good germ line contribution and gene targeting frequency. Nucleic AcidsRes. 20, 795-796.
Meyers, E. N., Lewandoski, M. and Martin, G. R. (1998). An Fgf8 mutant allelic seriesgenerated by Cre- and Flp-mediated recombination. Nat. Genet. 18, 136-141.
Mitchell, G. F., Jeron, A. and Koren, G. (1998). Measurement of heart rate and Q-Tinterval in the conscious mouse. Am. J. Physiol. 274, 747-751.
Morley, G. E., Danik, S. B., Bernstein, S., Sun, Y., Rosner, G., Gutstein, D. E. andFishman, G. I. (2005). Reduced intercellular coupling leads to paradoxicalpropagation across the Purkinje-ventricular junction and aberrant myocardialactivation. Proc. Natl. Acad. Sci. USA 102, 4126-4129.
Nagy, A. (2000). Cre recombinase: the universal reagent for genome tailoring. Genesis26, 99-109.
Nagy, A., Gerstenstein, M., Vintersten, K. and Behringer, R. (2003). Manipulating theMouse Embryo: A Laboratory Manual. New York: Cold Spring Habor LaboratoryPress.
Nicholson, B. J. (2003). Gap junctions – from cell to molecule. J. Cell Sci. 116, 4479-4481.
Plum, A., Hallas, G., Magin, T., Dombrowski, F., Hagendorff, A., Schumacher, B.,Wolpert, C., Kim, J., Lamers, W. H., Evert, M. et al. (2000). Unique and sharedfunctions of different connexins in mice. Curr. Biol. 10, 1083-1091.
Plum, A., Winterhager, E., Pesch, J., Lautermann, J., Hallas, G., Rosentreter, B.,Traub, O., Herberhold, C. and Willecke, K. (2001). Connexin31-deficiency in micecauses transient placental dysmorphogenesis but does not impair hearing and skindifferentiation. Dev. Biol. 231, 334-347.
Reaume, A. G., de Sousa, P. A., Kulkarni, S., Langille, B. L., Zhu, D., Davies, T. C.,Juneja, S. C., Kidder, G. M. and Rossant, J. (1995). Cardiac malformation inneonatal mice lacking connexin43. Science 267, 1831-1834.
Reuss, B., Hellmann, P., Dahl, E., Traub, O., Butterweck, A., Grummer, R. andWinterhager, E. (1996). Connexins and E-cadherin are differentially expressedduring trophoblast invasion and placenta differentiation in the rat. Dev. Dyn. 205, 172-182.
Severs, N. J., Rothery, S., Dupont, E., Coppen, S. R., Yeh, H. I., Ko, Y. S., Matsushita,T., Kaba, R. and Halliday, D. (2001). Immunocytochemical analysis of connexinexpression in the healthy and diseased cardiovascular system. Microsc. Res. Tech. 52,301-322.
Severs, N. J., Coppen, S. R., Dupont, E., Yeh, H., Ko, Y. and Matsushita, T. (2004).Gap junction alterations in human cardiac disease. Cardiovasc. Res. 62, 368-377.
Simon, A. M., Goodenough, D. A. and Paul, D. L. (1998). Mice lacking connexin40have cardiac conduction abnormalities characteristic of atrioventricular block andbundle branch block. Curr. Biol. 8, 295-298.
Söhl, G. and Willecke, K. (2003). An update on connexin genes and their nomenclaturein mouse and man. Cell Commun. Adhes. 10, 173-180.
Tiemann, K., Weyer, D., Djoufack, P. C., Ghanem, A., Lewalter, T., Dreiner, U.,Meyer, R., Grohe, C. and Fink, K. B. (2003). Increasing myocardial contraction andblood pressure in C57BL/6 mice during early postnatal development. Am. J. Physiol.Heart Circ. Physiol. 282, 464-474.
Traub, O., Eckert, R., Lichtenberg-Frate, H., Elfgang, C., Bastide, B., Scheidtmann,K. H., Hülser, D. F. and Willecke, K. (1994). Immunochemical andelectrophysiological characterization of murine connexin40 and -43 in mouse tissuesand transfected human cells. Eur. J. Cell Biol. 64, 101-112.
van Rijen, H. V., Eckardt, D., Degen, J., Theis, M., Ott, T., Willecke, K., Jongsma,H., Opthof, T. and de Bakker, J, M. (2004). Slow conduction and enhancedanisotropy increase the propensity for ventricular tachyarrhythmias in adult mice withinduced deletion of connexin43. Circulation 109, 1048-1055.
van Veen, A. A., van Rijen, H. V. and Opthof, T. (2001). Cardiac gap junction channels:modulation of expression and channel properties. Cardiovasc. Res. 51, 217-229.
Vozzi, C., Dupont, E., Coppen, S. R., Yeh, H. I. and Severs, N. J. (1999). Chamber-related differences in connexin expression in the human heart. J. Mol. Cell Cardiol.31, 991-1003.
Wei, C. J., Xu, X. and Lo, C. W. (2004). Connexins and cell signaling in developmentand disease. Annu. Rev. Cell Dev. Biol. 20, 811-838.
White, T. W. (2002). Unique and redundant connexin contributions to lens development.Science 295, 319-320.
Willecke, K., Eiberger, J., Degen, J., Eckardt, D., Romualdi, A., Güldenagel, M.,Deutsch, U. and Sohl, G. (2002). Structural and functional diversity of connexin genesin the mouse and human genome. Biol. Chem. 383, 725-737.
Yeager, M. and Nicholson, B. J. (2000). Structure and biochemistry of gap junctions. InGap Junctions: Advances in Molecular and Cell Biology (ed. E. L. Hertzberg and E.E. Bittar), pp. 31-98. Stamford: Jai Press.
Jour
nal o
f Cel
l Sci
ence





























