Embed Size (px)
Citation preview

EAP1/Daxx interacts with ETS1 and represses transcriptional activationof ETS1 target genes
Runzhao Li*,1, Huiping Pei1, Dennis K Watson1 and Takis S Papas1
1Center for Molecular and Structural Biology, Department of Medicine, and Hollings Cancer Center, Medical University of SouthCarolina, Charleston, South Carolina, SC 29425, USA
ETS1 is a member of the evolutionarily conserved familyof ets genes, which are transcription factors that bind tounique DNA sequences, either alone or by associationwith other proteins. In this study, we have used the yeasttwo-hybrid system to identify an ETS1 interactingprotein. The ETS1 N-terminal amino acid region wasused as bait and an interaction was identi®ed with theDaxx protein, referred to as EAP1 (ETS1 AssociatedProtein 1)/Daxx. This interactin has been shown to existin yeast and in vitro. EAP1/Daxx and ETS1 are co-localized in the nucleus of mammalian cells. The regionin EAP1/Daxx which speci®cally binds to ETS1 islocated within its carboxy terminal 173 amino acidregion. The ETS1 interaction region is located within itsN-terminal 139 amino acids and is referred as the DaxxInteraction Domain (DID). The DID appears to beconserved in several other ets family members, as well asin other proteins known to interact with Daxx. TheEAP1/Daxx interacts with both isoforms of ETS1, p51-ETS1 and p42-ETS1. Interaction of EAP1/Daxx withETS1 causes the repression of transcriptional activationof the MMP1 and BCL2 genes. The interaction domainsof both ETS1 and EAP1/Daxx are required for thisrepression and deletion of either domain abolishes thisactivity. Oncogene (2000) 19, 745 ± 753.
Keywords: ETS1; EAP1/Daxx; protein interaction;transcriptional repression; DID motif
Introduction
Ets proteins constitute a family of sequence speci®cDNA binding proteins, which share a highly conserved85 amino acid residue DNA-binding domain (ETSdomain) and function as transcription factors (Watsonet al., 1988; Ho et al., 1990; Bhat and Papas, 1994;Bassuk and Leiden, 1997). ETS proteins are foundfrom C. elegans to human. Amino acid sequencealignment of 49 ETS genes has been used to constructa phylogenetic tree, allowing sub-classi®cation intonine groups, including PEA3, ETS, ERG, ERF, ELG,ELK, ELF, YAN and SPI (Lautenberger et al., 1992;Graves and Petersen, 1998). ETS proteins carry outcritical roles in many biological processes, involvingdevelopment and function of the mammalian immunesystem (Bhat et al., 1989, 1990; Bassuk and Leiden,
1997), di�erentiation (Athanasiou et al., 1996), onco-genesis (Seth et al., 1989; Seth and Papas, 1990;Watson et al., 1997; Ghysdael and Boureux, 1997)and embryonic development (Maroulakou et al., 1994;Barton et al., 1998).
ETS1, the founding member of the ETS family, isone of the cellular genes forming the transformingprotein of the E26 virus. ETS1 plays important roles incell proliferation, di�erentiation, lymphoid cells devel-opment, transformation, invasiveness, angiogenesis,and apoptosis (Bhat et al., 1996; Huang et al., 1997;Li et al., 1999). ETS1 controls the expression of criticalgenes involved in these processes by binding to etsbinding sites (EBS) present in the transcriptionalregulatory regions. The mechanisms whereby indivi-dual ETS proteins regulate the expression of a uniqueset of target genes remains to be determined, but islikely to be controlled at multiple levels. Protein ±protein interactions provide one important mechanismto control the speci®city of target gene selection byETS proteins. Transcriptional regulation is dependentupon the combinatorial interactions between multiplenuclear proteins. Unlike many other well-knownfamilies of transcription factors that can associate ashomo- or heterodimers, Ets proteins are able to formcomplexes with other transcription factors. Suchinteractions may strengthen the transactivating activityand/or de®ne target gene speci®city. Many transcrip-tion factors have their DNA binding sites adjacent toets binding sites (EBS). Depending on the precisesequence context, binding of Ets protein near othertranscription factors results in higher a�nity interac-tions, synergistic repression or activation of speci®ctarget genes. For example, the presence of both ETS1and CBF proteins enhance the binding a�nity of bothproteins for beta E2 (Halle et al., 1997; Wotton et al.,1994). The basic and zipper region of MafB, an AP-1like protein, interacts with the ets domain of ETS1 in aDNA-dependent manner and inhibits ETS1-mediatedtransactivation of the transferrin receptor, which isknown to be essential for erythroid di�erentiation(Sieweke et al., 1996). Physical interaction of ETS1with GHF-1 is required for establishing lactotroph-speci®c PRL gene expression (Bradford et al., 1997).ETS1, as well as other family members, binds to theearly B-cell-speci®c mb-1 promoter by using Pax-5 as acell type-speci®c docking protein (Fitzsimmons et al.,1996). AP-1 tethers ETS1 to the TIMP-1 promoter viaprotein ± protein interaction to achieve ETS-dependenttranscriptional activation (Logan et al., 1996).
To identify additional proteins that may modulateETS1 function, we initiated a study to isolate proteinsbased upon their interaction with ETS1. The N-terminal 139 amino acids of human ETS1 was chosen
Oncogene (2000) 19, 745 ± 753ã 2000 Macmillan Publishers Ltd All rights reserved 0950 ± 9232/00 $15.00
www.nature.com/onc
*Correspondence: R Li, CMSB, Room #334, Hollings CancerCenter, Medical University of South Carolina, 86 Jonathan LucasStreet, Charleston, South Carolina, SC 29425, USAReceived 2 September 1999; revised 18 November 1999; accepted 2December 1999

as bait for a yeast two-hybrid interactive screen. In thisreport, we describe the identi®cation and characteriza-tion of a protein interacting with human ETS1, termedEAP1 (ETS1 associated protein-1)/Daxx. The interac-tion between ETS1 and EAP1/Daxx identi®ed in yeastoccurs in vitro in the absence of DNA and EAP1/Daxxis co-localized with ETS1 in nucleus of mammaliancells. Signi®cantly, we ®nd that EAP1/Daxx is able torepress ETS1-dependent transactivation of MMP1 andBCL2 promoters. Collectively, the data suggest thatEAP1/Daxx is able to bind to ETS1 in vivo andsubsequently function as a transrepressor. We havealso identi®ed two putative motifs located in the DIDdomain of ETS1 and other EAP1/Daxx interactingproteins required for protein ± protein interaction.
Results
Two hybrid system screen
We have used the yeast two-hybrid interaction trapmethod (Gyuris et al., 1993) to screen for proteins thatinteract with the ETS1 protein. Full-length ETS1 hasbeen shown to possess transcriptional activity. In orderto select an appropriate ETS1 protein bait, a series oflexA202-fused truncated ETS1 protein constructs wereprepared. Protein expression was con®rmed by Westernblot analysis utilizing a LexA antibody and therepression assay was used to verify the transport ofthe baits into the nucleus. As shown in Figure 1, only
the N139-ETS1 fragment derived from the N-terminaldomain of ETS1 and the C362-ETS1 fragment derivedfrom the C-terminal domain met the requirement as abait. To screen for binding proteins, an EGY48 yeaststrain that expresses the LexA-N139-ETS1 fusionprotein was transformed with a fetal brain cDNAlibrary. The cDNAs were fused to the B42 transactiva-tion domain (Gyuris et al., 1993). Of the approximately16106 transformants, 35 colonies were capable ofgalactose-dependent growth on media lacking leucineand production of b-galactosidase. Plasmid DNA wasisolated from positive yeast colonies and 23 individualclones were identi®ed by restriction mapping of PCRproducts. All clones were sequenced and eight of the 23clones were found to be novel. Three overlappingclones (17, 27 and 30) were found to encode di�erentparts of an identical protein. We have named thisETS1 interacting protein as EAP1 (ETS-1 AssociatedProtein-1). The protein encoded by these clones wassubsequently identi®ed as the Daxx gene (Yang et al.,1997). A schematic representation of the Daxx proteinalong with the clones interacting with the N139-ETS1bait is shown in Figure 2A. Figure 2B shows therelative b-galactosidase activity generated by theinteractions of the EAP1 clones and the bait N139-ETS1, with clone 27 exhibiting the highest activity. Thespeci®city of binding of Daxx clone 27 with ETS1 wascon®rmed in yeast by measuring the relative b-galactosidase activity utilizing several baits (Table 1).The data show that only the N139 bait bindsspeci®cally to Daxx. It is also interesting to note that
Figure 1 ETS1 bait constructs and their selection for the two hybrid system. The left panel includes schematic representations ofthe ETS1 protein [The Pointed (PNT), Activation (ACT) and DNA binding (ETS) domains are indicated.] and protein fragmentsfused to the LexA protein 1 ± 202 amino acids. Intrinsic transcription activity was measured in yeast cells co-transfected with thepSH-18-34 reporter gene and the LexA-ETS1 fusion constructs. The relative galactosidase activity of the Lac-Z reporter wasmeasured. This activity was controlled by the Lex-A operator. Plasmids pSH-17-4 and pRHMI were used as positive and negativecontrols, respectively. Yeast cells transformed with the indicated LexA-ETS1 fusion protein were grown in the presence of 2%galactose and 2% ra� medium. Leucine auxotrophy was performed in the above yeast cells by plating them on Gal/CM-Ura, -His,-Leu dropout plates. Yeast clones were checked on day 3 (++) and up to day 6 (+). Repression was measured by using thereporter gene pJK101 and the LexA-ETS1 fusion constructs. Expression levels of fusion proteins were monitored by Westernblotting analysis using the LexA antibody
EAP1/Daxx represses ETS1 dependent transcriptionR Li et al
746
Oncogene

only MafB, which is known to bind to ETS1 (Siewekeet al., 1996), was also capable of interacting with clone27.
Characterization of full-length EAP1/Daxx
The full-length EAP1/Daxx was cloned by PCR fromhuman T lymphocyte Jurkat cell cDNA (Materials andmethods). The sequence of EAP1/Daxx was depositedin the GenBank (Accession number: AF097742).Amino acid alignment between the mouse and humanEAP1/Daxx showed 70% homology. In vitro translatedEAP1/Daxx was found to migrate at a molecularweight of 120 kD, similar to what was previouslyreported (Kiriakidou et al., 1997). We have found thatEAP1/Daxx has nearly ubiquitous expression (data notshown) as previously described (Herberg et al., 1998;Kiriakidou et al., 1997; Yang et al., 1997).
Interaction of ETS1 with EAP1/Daxx
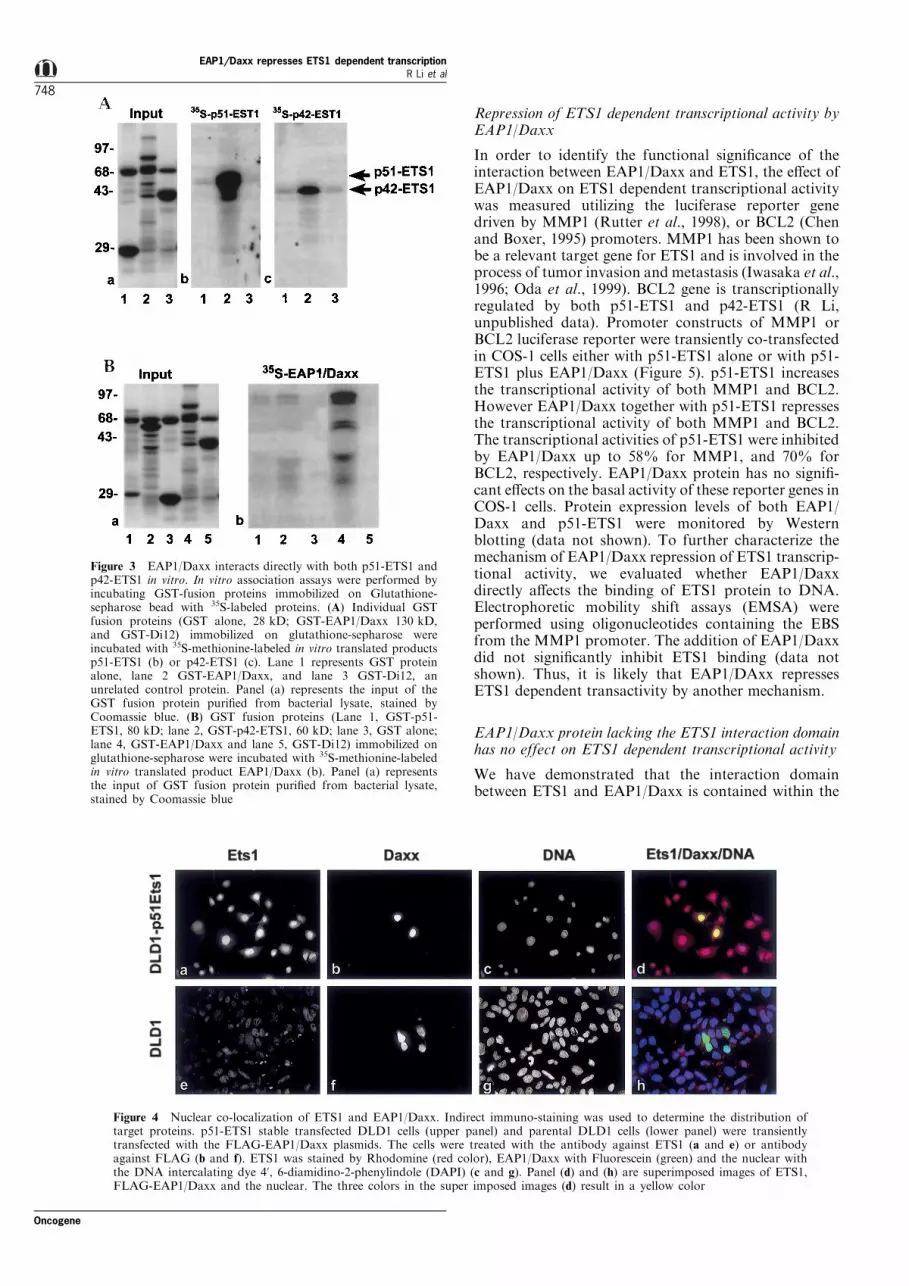
We showed above that the 139 amino acid regionfrom the amino terminus of the ETS1 proteininteracts with the 173 amino acid region from thecarboxyl terminus of EAP1/Daxx protein. To furtherverify this interaction, a GST-EAP1/Daxx fusionprotein was incubated with a 35S-methionine-labeledtranslation product p51-ETS1 (Figure 3A,b) or p42-ETS1 (Figure 3A,c). It is clear that both p51-ETS1(Figure 3A,b, lane 2) and p42-ETS1 (Figure 3A,c, lane2) interact speci®cally with EAP1/Daxx. No suchinteraction occurs between ETS1 and an unrelatedprotein Di12 (Figure 3A,b,c, lane 3). The reverseexperiment was also performed. In this case, either aGST-p51-ETS1 or a GST-p42-ETS1 fusion proteinwas immobilized and was incubated with a 35S-methionine-labeled translation product of EAP1/Daxx(Figure 3B,b). Again, no interaction occurs betweenDaxx and the unrelated protein Di12. It is noteworthythat EAP1/Daxx binds very strongly to itself (Figure3B,b, lane 4).
Subcellular localization of the EAP1/Daxx protein
It is known that ETS1 is a transcription factor bindingto a unique DNA sequence (-GGAA/T-), and islocalized in the nucleus. To determine whetherinteractions exist between ETS1 and EAP1/Daxx invivo, we determined the sub-cellular localization ofEAP1/Daxx. We utilized the stably transfected DLD1-p51-ETS1 cell line expressing high levels of p51-ETS1.This cell line was transiently transfected with a FLAG-EAP1/Daxx construct. Detection of EAP1/Daxx wasperformed utilizing an antibody directed againstFLAG. Thus, only the exogenously expressed EAP1/Daxx protein was detected. The upper panel of Figure4 shows that both ETS1 and EAP1/Daxx are localizedin the nucleus (Figure 4a,b) as de®ned by DNA nuclearstaining (Figure 4c). Shown in the lower panel ofFigure 4 (e ± h) is the parental DLD1 cell line, whichdoes not express endogenous ETS1. The exogenouslyexpressed FLAG-EAP1/Daxx is localized in thenucleus. We have identi®ed two putative nuclearlocalization signals from the predicted amino acidsequence, at position 397 ± 402 and 639 ± 646 aminoacid, respectively. A polyclonal antibody against full-length EAP1/Daxx protein was used to localize theendogenous EAP1/Daxx and the exogenously ex-pressed Daxx protein. Both forms of EAP1/Daxx weredetected in nucleus (data not shown).
Figure 2 Interaction between ETS1 and the EAP1/Daxx cDNAclones. (A) Schematic representation of EAP1/Daxx clones thatinteract with N139 ETS1 bait. The solid box in Daxx representsthe region interacting with Fas death domain (628 ± 739 aminoacid) and the striped box indicates the JNK activation domain.(B) EGY48 yeast strain containing the pSH18-34 reporter and theLexA-ETS1-N139 bait was transformed with EAP1/Daxx clones#17, #27 and #30. Transfectants were grown in Gal/CM-Ura,-His, -Leu, -Trp media. The galactosidase activity was measuredand normalized for the same amount of protein. pSH-17-4 andpRFHM1 were the positive and negative controls, respectively.Assays were performed in triplicate and repeated at least twotimes from di�erent individual clones
Table 1 EAP1/Daxx clone 27 interacts speci®cally with N139 ETS1protein
Baits Blue clones in glucose Blue clones in Gal/Raf
peg202 7 7C145 7 +C282 7 +N139 7 +++249-418 + +C362 7 +N418 +++ +++ETS2 +++ +++MafB + +++
EGY 48 Yeast Strain containing the pSH18-34 reporter and Clone 27was transformed with di�erent ETS1 as well as ETS2 and MafB.Transfectants were grown on X-gal plates (-His, -Trp, -Leu, -Ura)containing either Glucose or Gal/Raf. After 3 days the color changeswere observed
EAP1/Daxx represses ETS1 dependent transcriptionR Li et al
747
Oncogene
Repression of ETS1 dependent transcriptional activity byEAP1/Daxx
In order to identify the functional signi®cance of theinteraction between EAP1/Daxx and ETS1, the e�ect ofEAP1/Daxx on ETS1 dependent transcriptional activitywas measured utilizing the luciferase reporter genedriven by MMP1 (Rutter et al., 1998), or BCL2 (Chenand Boxer, 1995) promoters. MMP1 has been shown tobe a relevant target gene for ETS1 and is involved in theprocess of tumor invasion and metastasis (Iwasaka et al.,1996; Oda et al., 1999). BCL2 gene is transcriptionallyregulated by both p51-ETS1 and p42-ETS1 (R Li,unpublished data). Promoter constructs of MMP1 orBCL2 luciferase reporter were transiently co-transfectedin COS-1 cells either with p51-ETS1 alone or with p51-ETS1 plus EAP1/Daxx (Figure 5). p51-ETS1 increasesthe transcriptional activity of both MMP1 and BCL2.However EAP1/Daxx together with p51-ETS1 repressesthe transcriptional activity of both MMP1 and BCL2.The transcriptional activities of p51-ETS1 were inhibitedby EAP1/Daxx up to 58% for MMP1, and 70% forBCL2, respectively. EAP1/Daxx protein has no signi®-cant e�ects on the basal activity of these reporter genes inCOS-1 cells. Protein expression levels of both EAP1/Daxx and p51-ETS1 were monitored by Westernblotting (data not shown). To further characterize themechanism of EAP1/Daxx repression of ETS1 transcrip-tional activity, we evaluated whether EAP1/Daxxdirectly a�ects the binding of ETS1 protein to DNA.Electrophoretic mobility shift assays (EMSA) wereperformed using oligonucleotides containing the EBSfrom the MMP1 promoter. The addition of EAP1/Daxxdid not signi®cantly inhibit ETS1 binding (data notshown). Thus, it is likely that EAP1/DAxx repressesETS1 dependent transactivity by another mechanism.
EAP1/Daxx protein lacking the ETS1 interaction domainhas no effect on ETS1 dependent transcriptional activity
We have demonstrated that the interaction domainbetween ETS1 and EAP1/Daxx is contained within the
Figure 3 EAP1/Daxx interacts directly with both p51-ETS1 andp42-ETS1 in vitro. In vitro association assays were performed byincubating GST-fusion proteins immobilized on Glutathione-sepharose bead with 35S-labeled proteins. (A) Individual GSTfusion proteins (GST alone, 28 kD; GST-EAP1/Daxx 130 kD,and GST-Di12) immobilized on glutathione-sepharose wereincubated with 35S-methionine-labeled in vitro translated productsp51-ETS1 (b) or p42-ETS1 (c). Lane 1 represents GST proteinalone, lane 2 GST-EAP1/Daxx, and lane 3 GST-Di12, anunrelated control protein. Panel (a) represents the input of theGST fusion protein puri®ed from bacterial lysate, stained byCoomassie blue. (B) GST fusion proteins (Lane 1, GST-p51-ETS1, 80 kD; lane 2, GST-p42-ETS1, 60 kD; lane 3, GST alone;lane 4, GST-EAP1/Daxx and lane 5, GST-Di12) immobilized onglutathione-sepharose were incubated with 35S-methionine-labeledin vitro translated product EAP1/Daxx (b). Panel (a) representsthe input of GST fusion protein puri®ed from bacterial lysate,stained by Coomassie blue
Figure 4 Nuclear co-localization of ETS1 and EAP1/Daxx. Indirect immuno-staining was used to determine the distribution oftarget proteins. p51-ETS1 stable transfected DLD1 cells (upper panel) and parental DLD1 cells (lower panel) were transientlytransfected with the FLAG-EAP1/Daxx plasmids. The cells were treated with the antibody against ETS1 (a and e) or antibodyagainst FLAG (b and f). ETS1 was stained by Rhodomine (red color), EAP1/Daxx with Fluorescein (green) and the nuclear withthe DNA intercalating dye 4', 6-diamidino-2-phenylindole (DAPI) (c and g). Panel (d) and (h) are superimposed images of ETS1,FLAG-EAP1/Daxx and the nuclear. The three colors in the super imposed images (d) result in a yellow color
EAP1/Daxx represses ETS1 dependent transcriptionR Li et al
748
Oncogene

carboxyl terminal end of EAP1/Daxx. To determinewhether the interaction domain is required for
repression of ETS1 transcriptional activity, we madetwo deletion mutations lacking the interaction domain.The two deletion constructs are: (1) N567-EAP1/Daxxlacking 173 amino acids of the carboxyl terminalregion, and (2) C312-EAP1/Daxx, lacking 311 aminoacids of the amino terminal region. C312-EAP1/Daxxrepresses the activity of ETS1 by 50% in the BCL2system (Figure 6A). However, N567-EAP1/Daxx,which lacks the interaction domain does not repressthis activity. These results indicate that the interactiondomain is functionally important in repressing tran-scriptional activity. Essentially identical results wereobtained utilizing the promoter of another functionaltarget of ETS1, MMP1 (Figure 6B). These datastrongly support the conclusion that the interactiondomain of EAP1/Daxx plays a role in the transcrip-tional repression of at least two genes regulated byETS1. It will be of interest to determine if EAP1/Daxxis also involved in repressing the activity of other ETS1target genes.
EAP1/Daxx represses ETS1 dependent transcriptionalactivity in stable transfected DLD1 cell lines
To further study the role of EAP1/Daxx in repressingtranscriptional activity, we have utilized stable trans-fectants, expressing EAP1/Daxx, p42-ETS1 and p51-ETS1 alone or combination. The results are similar tothose previously shown utilizing transient transfection.The EAP1/Daxx expressing cell line did not activate
Figure 5 EAP1/Daxx represses p51-ETS1 dependent transcrip-tional activity. COS-1 cells were transfected with constructs ofMMP1 or BCL2 promoters coupled with the luciferase reportergene. Each construct was co-transfected with human p51-ETS1alone or with increasing concentrations of EAP1/Daxx (0.25, 0.5and 1 mg). Adding appropriate amount of vector pcDNA3.1normalized the total amounts of DNA. The results werenormalized by b-galactosidase activity from a co-transfectedpCH110-Lac-Z vector. Assays were performed in triplicate andrepresent at least two experiments
Figure 6 E�ect of EAP1/Daxx protein lacking the interacting domain with ETS1 on the ETS1 dependent transcriptional activity.(A) The BCL2 promoter construct with the luciferase reporter was co-transfected in COS-1 cells with p51-ETS1 alone, p51-ETS1and N567-EAP1/Daxx (Del 567 ± 740aa), or p51-ETS1 N312-EAP1/Daxx (Del 1 ± 312aa). Similarly, the construct of BCL2 was alsoco-transfected with p42-ETS1 alone, p42-ETS1 and N567-EAP1/Daxx (Del 567 ± 740aa), or p42-ETS1 and N312-EAP1/Daxx (Del1 ± 312aa). Total amounts of DNA were normalized by the addition of carrier vector pcDNA3.1. Luciferase activities were measuredafter 24 h of transfection. The values were normalized by b-galactosidase activity from a co-transfected CH110-Lac-Z vector. Assayswere performed in triplicate and represented two independent experiments. (B) The MMP1 promoter construct with the luciferasereporter was co-transfected in COS-1 cells with p42-ETS1 alone, p42-ETS1 plus an increased amount of either N567-EAP1/Daxx(Del 567 ± 740aa), or with N312-EAP1/Daxx (Del 1 ± 312aa). Total amounts of DNA were normalized by the addition of carriervector pcDNA3.1. Luciferase activities were measured after 24 h of transfection. The values were normalized by b-galactosidaseactivity from a co-transfected CH110-Lac-Z vector. Assays were performed in triplicate and represented two independentexperiments
EAP1/Daxx represses ETS1 dependent transcriptionR Li et al
749
Oncogene

the MMP1 promoter, while this promoter was active inboth the p51-ETS1 and p42-ETS1 expressing cell lines.A dramatic decrease in the transcriptional activity ofthe MMP1 gene was obtained in cell lines expressingboth p51-ETS1 and EAP1/Daxx. (Figure 7B, lanes 5and 6).
Discussion
The ETS family of transcription factors is conservedthroughout evolution. Family members are present inDrosophila and humans, indicating that the family hasexisted for more than 600 million years (Pribyl et al.,1998, 1991; Lautenberger et al., 1992). They bind tounique DNA sequences, either by themselves or byforming complexes with partner proteins. ETS1controls the expression of target genes that play criticalroles in cell proliferation, di�erentiation, lymphoid celldevelopment, transformation, angiogenesis, and apop-
tosis (Bhat et al., 1996). Sequence recognition can bein¯uenced by other factors in the vicinity of the EBS.Therefore, the geometry of the binding site and theadjacent factors will in¯uence the selection of targetgenes.
In this paper, we have utilized the yeast two-hybridsystem to identify ETS1 interacting proteins. A 139amino acid region of the amino terminus of the ETS1protein was used as bait. This region includes thePointed (PNT) domain and an adjacent mitogen-activated protein (MAP) kinase phosphorylation site.PNT is conserved in a subset of the ets familymembers. Nuclear magnetic resonance (NMR) spectro-scopy analysis of this domain indicates that it forms anindependent structure with unique architecture of amonomeric ®ve-helix bundle (Slupsky et al., 1998).With this bait, we have identi®ed several putativeinteracting proteins. One of the clones was furthercharacterized and found to be Daxx, which wasoriginally reported to be involved in the Fas deathpathway (Yang et al., 1997).
The interaction region of EAP1/Daxx is located inthe c-terminal 173 amino acids. This region was testedin the yeast system with several ETS1 baits (Table 1).The only region that interacts with EAP1/Daxx is 139amino acid from the amino terminus of ETS1. Thus,we have identi®ed the interaction domains of bothETS1 and EAP1/Daxx. EAP1 clone 30, which containsall the amino acids present in clone 27, is not as activeas clone 27. This may be due to conformationaldi�erences between the clone 27 and 30 proteins, suchthat the additional amino terminal amino acids ofclone 30 protein interfere with its ability to bind ETS1.It became important to examine whether the nativeETS1 and EAP1/Daxx protein interact. Our datademonstrates that EAP1/Daxx interacts with bothp51-ETS1 and its spliced variant, p42-ETS1 insolution. The p42-ETS1 lacks exon VII, which encodes87 amino acids directly adjacent to the DNA bindingdomain (Jorcyk et al., 1991). Since the interactiondomain is present in both ETS1 isoforms and both ofthese proteins interact with EAP1/Daxx, amino acidsderived from exon VII do not contribute to theconformation of the Daxx interaction domain ofETS1 protein. This is consistent with the conclusionfrom structural studies that the N-terminal 139 aminoacids of ETS1 protein forms an independent structurewith unique architecture (Slupsky et al., 1998). Wehave also observed that EAP1/Daxx binds to itself(Figure 3B,a, lane 4) and the signi®cance of this self-interaction remains to be further evaluated.
That ETS1 and EAP1/Daxx are predicted to belocalized in the nucleus, strongly suggests the possibi-lity for a physiological interaction. We have tested thishypothesis by examining the subcellular localization ofboth ETS1 and EAP1/Daxx. Our results clearlydemonstrate that ETS1 and EAP1/Daxx are co-localized in the nucleus supporting the suggestion ofa possible interaction (Figure 4). The functionalimportance of the interaction between ETS1 andEAP1/Daxx is clearly demonstrated. We have shownthat this interaction leads to the transcriptionalrepression of two relevant ETS1 target genes MMP1and BCL2. Removal of the ETS1 interacting domainfrom EAP1/Daxx results in the loss of transcriptionalrepression.
Figure 7 EAP1/Daxx represses ETS1 dependent transcriptionalactivity in stably transfected DLD1 cells. Parental DLD1 cells(lane 1) and stably transfected DLD1 transfectants expressingEAP1/Daxx (lane 2), p42-ETS1 (lane 3), p42-ETS1 plus EAP1/Daxx (lane 4), p51-ETS1 (lane 5) and p51-ETS1 plus EAP1/Daxx(lane 6) were utilized. Detection of these proteins was performedby Western analysis utilizing the appropriate antibodies for ETS1and EAP1/Daxx. The results are shown in (A). Each of the abovestable cell lines were transfected with a luciferase reporter drivenby MMP1 promoter (B). The luciferase activity was measured andnormalized by b-galactosidase activity from a co-transfectedCH110-Lac-Z vector and the experiments were performed intriplicate
EAP1/Daxx represses ETS1 dependent transcriptionR Li et al
750
Oncogene

The data in this paper are summarized diagramati-cally in Figure 8, which shows the structure of theEAP1/Daxx gene. The domain that we have found tointeract with ETS1 is designated the ETS1 BindingDomain. We also have identi®ed the region of ETS1that binds to EAP1/Daxx, which we refer to as theDaxx Interaction Domain (DID). Ets family membersare known to share many sequence homologies. Themost widely conserved region within the gene family isthe DNA binding domain of 85 amino acids, and itsstructure has been determined (Werner et al., 1995).The DID overlaps with the previously described HLH(Seth and Papas, 1990) and PNT (Pointed) domains,which is the second most highly conserved domain. Itbecame, therefore, of interest to determine if other etsfamily members have homologous sequences. Align-ment of several ets family proteins demonstratessequence homology in the putative DID of two otherfamily members, ETS2 and FLI1. Greatest homologyis found in two separate regions within the DID (DIDmotif I and II). This homology predicts that EAP1/Daxx will bind to ETS2 and FLI1 and regulate theiractivity. This may lead to the coordinate regulation ofthe ets family which would also regulate the expressionof their common target genes. We are presentlyinvestigating the interaction of EAP1/Daxx with otherfamily members.
A search for the existence of DID motifs in otherproteins known to bind to Daxx, including PAX3(Hollenbach et al., 1999) and CENP-C (Pluta et al.,1998) was carried out. It is signi®cant that these motifs
are conserved in both of these proteins. We, therefore,propose that the EAP1/Daxx protein may be a generalregulatory molecule involved in the control of theactivities of many nuclear proteins by binding to theDID motifs of those proteins. It has been shown thatseveral ets proteins are phosphorylated in response toextracellular signals. The threonine 38 (T38) in ETS1or threonine 72 in ETS2 are the only phosphorylationsites responding to these signals (Wasylyk et al., 1997).It is interesting to note that the putative motif of DIDin both ETS1 and ETS2 contain these phosphorylationsites. So that it is possible that the extracellular signalfrom ERK pathway a�ects the interaction of ETS1with its partner such as EAP1/Daxx.
As mentioned above, H-Daxx has been previouslyidenti®ed as a PAX3 interacting protein and repressesPAX3 dependent transactivity (Hollenback et al.,1999). PAX3 is a member of an evolutionarilyconserved family of Pax genes. Similar to the etsfamily, Pax family members function as transcriptionalregulators that control gene expression (Dahl et al.,1997). It is likely that the biological signi®cance ofEAP1/Daxx may be determined by its predominateinteracting protein, which may be controlled during thecell cycle, developmental stage and the speci®c cell line.
The recent report on the Drosophila Groucho genefamily describes the function of these proteins as non-DNA binding co-repressors for speci®c subsets ofDNA binding transcription factors (Fisher and Caudy,1998). These proteins repress transcription via aconserved intrinsic repression domain. We have shown
Figure 8 Diagram of Daxx ETS1 interaction regions. The organization of the Daxx gene is shown, and the ETS1 binding region isindicated by the full black section in the N-terminus, from amino acid 567 ± 740. The Daxx Interaction Domain (DID) regions inETS1, PAX3 and CENP-C are shown in full black. Speci®c sequence motifs within the DIDs of the three known (ETS1, PAX3 andCENP-C) and two putative (ETS2 and FLI1) Daxx binding proteins are also included
EAP1/Daxx represses ETS1 dependent transcriptionR Li et al
751
Oncogene

in this paper that the Daxx protein has manysimilarities to Groucho. For example, it also repressestranscription of a subset of genes by interaction with aunique domain of DNA binding transcription factors.In addition, it has been shown previously that Daxxalso binds to other transcription factors, repressingtheir activity (Hollenbach et al., 1999) and to anintrinsic centromeric protein which may have aregulatory role in mitosis (Pluta et al., 1998).
Materials and methods
Two hybrid system
The interaction trap assay system and a human fetal braincDNA library were kindly provided by Dr Roger Brent(Harvard Medical School, Boston, MA, USA). All experi-ments were performed in the yeast reporter strain EGY48yeast cells [MATa leu27 his7 trp7 upa7LEU2 : : plexao-p6LEU2(DUASLEU2)]. The yeast vector pEG202 is a LexADNA-binding domain (1 ± 202aa) encoding vector that allowsfusion of desired protein into its C-terminal part. A series ofETS1 bait constructs were inserted into pEG202 vector inframe of LexA. DNA fragments for these constructs weregenerated by PCR ampli®cation using Pfu polymerase(Stratagene, La Jolla, CA, USA) and primers designed withappropriate restriction sites. The human fetal brain cDNAlibrary was constructed using the yeast vector pJG-45, whichcontains a B42 activation domain that allows fusion ofdesired protein to its C-terminus. Plasmid constructs weresequenced by the Taq Dideoxy Terminator Cycle sequencingmethod (Applied Biosystems, Foster City, CA, USA) usingan Applied Biosystems model 373 automated sequencer. Thelibrary screen, b-galactosidase assays and general yeastmanipulate were performed as described previously (Ausubelet al., 1995).
Cloning and plasmid construct
For cloning of full-length EAP1/Daxx: proof reading PCRwas used to amplify a 2158 bp fragment from human Tlymphocyte Jurkat cell cDNA by the forward primer, 5'-ATCCCCTATGGCCACCGCTAACA-3' (Kiriakidou et al.,1997) and reverse primer, 5'-CACACTTGTCTTGCAAG-TACCAG-3' (derived from the sequence of our initial clone).The product was digested by EcoRI and NcoI and insertedinto p2tk2/2700, which was constructed by inserting the718 bp EcoRI/XhoI fragment of clone 27 into the p2tk2vector. The plasmid generated above yields a GST fused full-length EAP1/Daxx protein. pcDNA3.1-EAP1/Daxx weremade by inserting a BamHI/XhoI fragment into pcDNA3.1(Invitrogen, Carlsbad, CA, USA). Deletion constructs ofN567-EAP1/Daxx (del 567 ± 740) and C312-Daxx (del 1 ± 312)were generated by inserting the PCR fragment containing thenative start codon into appropriated cloning sites ofpcDNA3.1 vector. The XhoI/HindIII fragment ofpcDNA3.1-EAP1/Daxx was inserted into the SalI andHindIII sites of pFlag-cmv-2 vector to generate FLAG-EAP1/Daxx.
Indirect immune-staining
Cells were grown on glass 35 mm dish and ®xed in PBScontaining 4% paraformaldehyde and 0.5% Triton X-100 for30 min at room temperature. The cells were then washedtwice with PBS, and nonspeci®c binding was blocked for25 min with 3% bovine serum albumin and 10% fetal bovineserum in PBS. The cells were then incubated with a�nity-puri®ed Anti-¯ag M5 Mab (Eastman Kodak, New Haven,CT, USA) or anti-ETS1 polyclone (C-20 Santa Cruz, CA,
USA) for 1 h at room temperature, and washed ®ve timeswith PBS, followed by incubation with Fluorescein con-jugated anti mouse IgG (Pierce, Rockford, IL, USA) forFLAG and Rhodamine conjugated anti rabbit IgG (Pierce,Rockford, IL, USA) for recognized Ets1 C-20 polyclone(Santa Cruz, CA, USA) for 1 h, respectively. The cells wereadditionally washed four times with PbS containing 0.02% 4',6-diamidino-2-phenylindole (DAPI) for staining of nucleus,and the coverslips were mounted with Gel/Mount aqueousmounting medium (Fisher, Pittsburgh, PA, USA) andobserved using a ¯uorescent microscope.
GST pull-down assays
Plasmid constructs of GST-EAP1/Daxx, GST-p51-ETS1 andGST-p42-ETS1 were generated in pGEX-2TK2 derivedpGEX-2TK (Pharmacia-Biotech, Piscataway, NJ, USA) byinserting multiple-cloning sites. The fusion proteins werepuri®ed as described in the manufacturer's protocol. 35S-labeled protein were obtained by in vitro translation reactions(TNT coupled reticulocyte lysate system, Promega). Twomicrograms of DNA were used in the presence of completeamino acid mix with T7 RNA Polymerase as per themanufacturer's protocol. GST fusion protein (2 ± 5 mg) wasbound to glutathione-agarose beads and incubated with 35S-labeled protein (5 ml), in 0.2 ml binding bu�er (20 mM
HEPES pH 7.6, 3 mM MgCL2, 0.1 mM EDTA, 0.1%Tween-20, 10% Glycerol, 1 mM DTT, 0.2 M KCL, 2%BSA). The mixture was incubated by rocking for 2 h at 48C.The glutathione-agarose beads were washed four times inbinding bu�er (without BSA), resuspended in 20 ml of SDSsample bu�er, and the proteins were released from the beadby boiling for 5 min. The proteins were analysed by 10%SDS±PAGE and autoradiography.
Cell line and transfection
COS-1 and the human colon cancer DLD1 (human coloncarcinoma cell, ATCC-CCL221) and DLD1 stable transfec-tants were used in these experiments. Transient transfectionswere carried out using Superfect (Qiagen, Valencia, CA,USA) as per the manufacturer's directions. For reporterassays, cells were seeded 24 h prior to transfection in 24-wellplates. Equal amount of DNA was maintained in eachtransfection by addition of pcDNA3.1 vector. Cells wereharvested 24 h after transfection and luciferase activity wasmeasured by a luciferase assay kit (Promega). Preparation ofcell lysates and reporter assays were done as per themanufacturer's protocols. Assay results were corrected forequal amount of protein. All transfections were performed atleast three times in duplicate or triplicate. In addition, stableDLD1 transfectants expressing EAP1/Daxx and EAP1/Daxxplus either p51-ETS1 or p42-ETS1 were established and theclones were maintained in RPMI with 10% fetal bovineserum (FBS) in the presence of G418 (400 mg/ml). Westernblots were developed by enhanced chemiluminescence (ECL).Anti-EAP1/Daxx polyclonal antibody was generated againstfull-length EAP1/Daxx generated from GST-EAP1/Daxxafter thrombin cleavage. Antibody against ETS1 (c-20,sc350) was purchased from Santa Cruz Biotechnology, Inc.(Santa Cruz, CA, USA).
AcknowledgmentsWe thank Dr R Brent (Harvard Medical School, MA,USA) for generously providing the yeast two hybridreagents; Dr L Boxer (Stanford University, CA, USA)for providing BCL-2 luciferase reporter plasmid; Drs JRutter and C Brinckerho� (Dartmouth Medical School,NH, USA) for the MMP1-luciferase reporter plasmids andDr M Sieweke (EMBL, Germany) for MafB plasmid. We
EAP1/Daxx represses ETS1 dependent transcriptionR Li et al
752
Oncogene

thank Drs J Vournakis, T Hsu and C Schweinfest forcritical review of the manuscript. The GeneBank accessionnumber for the sequence of EAP1/Daxx reported in thispaper is AF097742. This work was supported in part by
National Institutes of Health Grants P01CA78582,RO1CA76231 and American Cancer Society Grant (GN-164) to TS Papas. This study is dedicated to the memory ofDr Takis S Papas, a mentor, scienti®c colleague and friend.
References
Athanasiou M, Clausen PA, Mavrothalassitis GJ, ZhangXK, Watson DK and Blair DG. (1996). Cell GrowthDi�er., 7, 1525 ± 1534.
Ausubel F, Brent R and Kingston R (eds). (1995). CurrentProtocols in Molecular Biology. Wiley Interscience, NewYork.
Barton K, Muthusamy N, Fischer C, Ting CN, Walunas TL,Lanier LL and Leiden JM. (1998). Immunity, 9, 555 ± 563.
Bassuk A and Leiden J. (1997). Advances in Immunology,Vol. 64. Academic Press: San Diego, pp. 65 ± 104.
Bhat NK, Fischinger PJ, Seth A, Watson DK and Papas T.(1996). Inter. J. Oncol., 8, 841 ± 846.
Bhat NK, Komschlies KL, Fujiwara S, Fisher RJ, MathiesonBJ, Gregorio TA, Young HA, Kasik JW, Ozato K andPapas TS. (1989). J. Immunol., 142, 672 ± 678.
Bhat NK and Papas TS. (1994). Challenges of ModernMedicine, Vol. 5: Ares-Serono Symposia 5. Verna R andShamoo A (eds). pp. 63 ± 68.
Bhat NK, Thompson CB, Lindsten T, June CH, Fujiwara S,Koizumi S, Fisher RJ and Papas TS. (1990). Proc. Natl.Acad. Sci. USA, 87, 3723 ± 3727.
Bradford AP, Wasylyk C, Wasylyk B and Gutierrezhart-mann A. (1997). Mol. Cell Biol., 17, 1065 ± 1074.
Chen HM and Boxer LM. (1995). Mol. Cell. Biol., 15, 3840 ±3847.
Dahl E, Koseki H and Balling R. (1997). Bioessays, 19, 755 ±765.
Fisher AL and Caudy M. (1998). Genes Dev., 12, 1931 ± 1940.Fitzsimmons D, Hodsdon W, Wheat W, Maira SM, Wasylyk
B and Hagman J. (1996). Genes Dev., 10, 2198 ± 2211.Ghysdael J and Boureux A. (1997). The ETS family of
Transcriptional Regulators. Birkhauser Verlag, Basel,Switzerland.
Graves B and Petersen J. (1998). Advances in CancerResearch, Vol. 75. Academic Press: San Diego, California,pp. 1 ± 55.
Gyuris J, Golemis E, Chertkov H and Brent R. (1993). Cell,75, 791 ± 803.
Halle JP, Haus-Seu�ert P, Woltering C, Stelzer G andMeisterernst M. (1997). Mol. Cell. Biol., 17, 4220 ± 4229.
Herberg JA, Beck S and Trowsdale J. (1998). J. Mol. Biol.,277, 839 ± 857.
Ho IC, Bhat NK, Gottschalk LR, Lindsten T, ThompsonCB, Papas TS and Leiden JM. (1990). Science, 250, 814 ±818.
Hollenbach AD, Sublett JE, McPherson CJ and Grosveld G.(1999). EMBO J., 18, 3702 ± 3211.
Huang CC, Papas TS and Bhat NK. (1997). Oncogene, 15,851 ± 856.
Iwasaka C, Tanaka K, Abe M and Sato Y. (1996). J. CellPhysiol., 169, 522 ± 531.
Jorcyk CL, Watson DK, Mavrothalassitis GJ and Papas TS.(1991). Oncogene, 6, 523 ± 532.
Kiriakidou M, Driscoll DA, Lopez-Guisa JM and StraussJF, III. (1997). DNA Cell Biol., 16, 1289 ± 1298.
Lautenberger JA, Burdett LA, Gunnell MA, Qi S, WatsonDK, O'Brien SJ and Papas TS. (1992).Oncogene, 7, 1713 ±1719.
Li R, Pei H and Papas TS. (1999). Proc. Natl. Acad. Sci.USA, 96, 3876 ± 3881.
Logan SK, Garabedian MJ, Campbell CE and Werb Z.(1996). J. Biol. Chem., 271, 774 ± 782.
Maroulakou IG, Papas TS and Green JE. (1994). Oncogene,9, 1551 ± 1565.
Oda N, Abe M and Sato Y. (1999). J. Cell Physiol., 178,121 ± 132.
Pluta AF, Earnshaw WC and Goldberg IG. (1998). J. CellSci., 111, 2029 ± 2041.
Pribyl LJ, Watson DK, McWilliams MJ, Ascione R andPapas TS. (1988). Dev. Biol., 127, 45 ± 53.
Pribyl LJ, Watson DK, Schulz RA and Papas TS. (1991).Oncogene, 6, 1175 ± 1183.
Rutter JL, Mitchell TI, Buttice G, Meyers J, Gusella JF,Ozelius LJ and Brinckerho� CE. (1998). Cancer Res., 58,5321 ± 5325.
Seth A and Papas TS. (1990). Oncogene, 5, 1761 ± 1767.Seth A, Watson DK, Blair DG and Papas TS. (1989). Proc.
Natl. Acad. Sci. USA, 86, 7833 ± 7877.Sieweke MH, Tekotte H, Frampton J and Graf T. (1996).
Cell, 85, 49 ± 60.Slupsky CM, Gentile LN, Donaldson LW, Mackereth CD,
Seidel JJ, Graves BJ and McIntosh LP. (1998). Proc. Natl.Acad. Sci. USA, 95, 12129 ± 12134.
Wasylyk C, Bradford AP, Gutierrez-Hartmann A andWasylyk B. (1997). Oncogene, 14, 899 ± 913.
Watson DK, McWilliams MJ, Lapis P, Lautenberger JA,Schweinfest CW and Papas TS. (1998). Proc. Natl. Acad.Sci. USA, 85, 7862 ± 7866.
Watson DK, Robinson L, Hodge DR, Kola I, Papas TS andSeth A. (1997). Oncogene, 14, 213 ± 221.
Werner MH, Clore M, Fisher CL, Fisher RJ, Trinh L,Shiloach J and Gronenborn AM. (1995). Cell, 83, 761 ±771.
Wotton D, Ghysdael J, Wang S, Speck NA and Owen MJ.(1994). Mol. Cell. Biol., 14, 840 ± 850.
Yang X, Khosravi-Far R, Chang HY and Baltimore D.(1997). Cell, 89, 1067 ± 1076.
EAP1/Daxx represses ETS1 dependent transcriptionR Li et al
753
Oncogene





























