Embed Size (px)
Citation preview


Temelji neuroznanosti Vodič kroz vježbe
1. Stanična građa, izgled i raspodjela sive i bijele tvari kralježnične moždine i mozga
Cilj vježbe je prikazati studentima građu središnjega živčanog sustava
studiranjem mikroskopskih i makroskopskih preparata mozga i kralježnične moždine.
1.1 Uvod
Različitim neurohistološkim tehnikama moguće je na mikroskopskim
preparatima prikazati raznolikost građe središnjega živčanog sustava. Golgijevom
metodom prikazuje se izgled neurona, Nisslovom metodom oboje se tijela neurona, a
Weigertovom metodom bojenja vizualiziraju se mijelinizirani aksoni.
Makropreparatima mozga fiksiranog u formaldehidu prikazujemo sivu i bijelu
tvar velikog mozga, malog mozga i moždanog debla. Osim prikaza vanjskog izgleda,
moguće je različitim presjecima u nekoliko ravnina istražiti i unutrašnju građu tih
struktura.
Tri temeljne neurohistološke metode uveli su krajem prošlog stoljeća Camillo
Golgi, Franz Nissl i Carl Weigert.
Golgijevom metodom možemo prikazati stvarni izgled neurona
Srebrnim solima možemo obojiti samo neke neurone u malom komadiću
moždanog tkiva, no tada se oboje svi dijelovi neurona (soma, dendriti i akson). Ta je
(Golgijeva) metoda po prvi put omogućila spoznaju iznimne raznolikosti oblika
neurona i njihovih međusobnih odnosa. Golgijeva metoda pokazuje stvarni izgled
neurona, ali ne i citološke pojedinosti (cijeli je neuron zacrnjen talogom srebrnih soli,
pa se niti jezgra ne vidi) ili neurotransmitersku narav.
Nisslovom metodom prikazujemo citoarhitektoniku, tj. opći plan staničnog
ustrojstva sive tvari središnjega živčanog sustava.
Tom se metodom oboji Nisslova tvar, tj. nukleinske kiseline jezgre i
ribosoma, pa se tako prikažu tijela svih stanica središnjega živčanog sustava. Pritom
se prikažu cijela tijela neurona te samo početni dijelovi dendrita i aksona. Stoga se
tom metodom ne može prikazati stvarni izgled neurona (kao Golgijevom metodom),
ali se može prikazati opći plan stanične građe sive tvari središnjega živčanog sustava
3
Temelji neuroznanosti Vodič kroz vježbe
1. Stanična građa, izgled i raspodjela sive i bijele tvari kralježnične moždine i mozga
Cilj vježbe je prikazati studentima građu središnjega živčanog sustava
studiranjem mikroskopskih i makroskopskih preparata mozga i kralježnične moždine.
1.1 Uvod
Različitim neurohistološkim tehnikama moguće je na mikroskopskim
preparatima prikazati raznolikost građe središnjega živčanog sustava. Golgijevom
metodom prikazuje se izgled neurona, Nisslovom metodom oboje se tijela neurona, a
Weigertovom metodom bojenja vizualiziraju se mijelinizirani aksoni.
Makropreparatima mozga fiksiranog u formaldehidu prikazujemo sivu i bijelu
tvar velikog mozga, malog mozga i moždanog debla. Osim prikaza vanjskog izgleda,
moguće je različitim presjecima u nekoliko ravnina istražiti i unutrašnju građu tih
struktura.
Tri temeljne neurohistološke metode uveli su krajem prošlog stoljeća Camillo
Golgi, Franz Nissl i Carl Weigert.
Golgijevom metodom možemo prikazati stvarni izgled neurona
Srebrnim solima možemo obojiti samo neke neurone u malom komadiću
moždanog tkiva, no tada se oboje svi dijelovi neurona (soma, dendriti i akson). Ta je
(Golgijeva) metoda po prvi put omogućila spoznaju iznimne raznolikosti oblika
neurona i njihovih međusobnih odnosa. Golgijeva metoda pokazuje stvarni izgled
neurona, ali ne i citološke pojedinosti (cijeli je neuron zacrnjen talogom srebrnih soli,
pa se niti jezgra ne vidi) ili neurotransmitersku narav.
Nisslovom metodom prikazujemo citoarhitektoniku, tj. opći plan staničnog
ustrojstva sive tvari središnjega živčanog sustava.
Tom se metodom oboji Nisslova tvar, tj. nukleinske kiseline jezgre i
ribosoma, pa se tako prikažu tijela svih stanica središnjega živčanog sustava. Pritom
se prikažu cijela tijela neurona te samo početni dijelovi dendrita i aksona. Stoga se
tom metodom ne može prikazati stvarni izgled neurona (kao Golgijevom metodom),
ali se može prikazati opći plan stanične građe sive tvari središnjega živčanog sustava
3

Temelji neuroznanosti Vodič kroz vježbe
(broj, veličina, oblik i međusobni raspored tijela neurona i glije), tj. citoarhitektonika
različitih moždanih područja.
Mijeloarhitektonika temelji se na Weigertovoj metodi bojanja mijeliniziranih
aksona solima nekih teških metala. Mijelinske ovojnice mijeliniziranih aksona mogu
se obojiti tamnosmeđe ili crno, kad rezove moždanog tkiva uronimo u vodene otopine
soli nekih teških metala. Takvo (Weigertovo) bojanje otkriva nam opći plan građe
bijele tvari, odnosno međusobni raspored i usmjerenost već ih i manjih snopova
mijeliniziranih aksona (ali ne i nemijeliniziranih aksona!). Stoga govorimo o
mijeloarhitektonici pojedinih moždanih područja. Temeljna vrijednost te metode je u
tome što nam pokazuje topografski položaj i usmjerenost glavnih moždanih putova,
tj. većih snopova aksona što polaze od neurona smještenih u jednom području sive
tvari (jezgri, sloju), imaju istu putanju (i obično istu funkciju), a završe
(uspostavljajući sinapse) na neuronima u nekom drugom području sive tvari (jezgri,
sloju).
Neuronske putove može se istražiti metodama što se temelje na prirodnoj
pojavi aksonskog prenošenja. Putanju aksona može se prikazati autoradiografijom
radioaktivno označenih aminokiselina što aksonskim prenošenjem putuju
anterogradno (od some neurona do presinaptičkog završetka aksona). Ta se metoda
temelji na pojavi anterogradnog aksonskog prenošenja, tj. polaganog premještanja
raznovrsnih molekula i organela od tijela neurona do presinaptičkog završetka aksona
(za opis pojave vidi 6. poglavlje udžbenika!). U neko područje sive tvari (jezgru, sloj)
uštrca se finom iglom ili mikropipetom mala količina aminokiseline (npr. leucin ili
prolin) označene tricijem. Ta aminokiselina anterogradno se prenosi duž aksona te se
(nakon određenog vremena) može autoradiografskim postupkom (na histološkom rezu
tkiva) prikazati i dotični akson i njegovo završno razgranjenje, tj. ciljno područje u
kojemu završava. Tako su primjerice David Hubel i Thorsten Wiesel (Nobelova
nagrada za fiziologiju i medicinu 1981. godine!) uspješno prikazali kolumnarni
završetak talamokortikalnih aksona vidnog puta u primarnoj vidnoj moždanoj kori
(vidi 28. poglavlje). Putanju aksona može se prikazati histokemijski, na temelju
retrogradnog (od presinaptičkog aksonskog završetka prema somi) aksonskog
prenošenja peroksidaze hrena (HRP).
Ta se metoda temelji na pojavi retrogradnog aksonskog prenošenja, tj.
putovanja raznovrsnih molekula i organela od presinaptičkog završetka aksona do
some neurona (vidi također 6. poglavlje). U lokalizirano područje sive tvari uštrcamo
4
Temelji neuroznanosti Vodič kroz vježbe
(broj, veličina, oblik i međusobni raspored tijela neurona i glije), tj. citoarhitektonika
različitih moždanih područja.
Mijeloarhitektonika temelji se na Weigertovoj metodi bojanja mijeliniziranih
aksona solima nekih teških metala. Mijelinske ovojnice mijeliniziranih aksona mogu
se obojiti tamnosmeđe ili crno, kad rezove moždanog tkiva uronimo u vodene otopine
soli nekih teških metala. Takvo (Weigertovo) bojanje otkriva nam opći plan građe
bijele tvari, odnosno međusobni raspored i usmjerenost već ih i manjih snopova
mijeliniziranih aksona (ali ne i nemijeliniziranih aksona!). Stoga govorimo o
mijeloarhitektonici pojedinih moždanih područja. Temeljna vrijednost te metode je u
tome što nam pokazuje topografski položaj i usmjerenost glavnih moždanih putova,
tj. većih snopova aksona što polaze od neurona smještenih u jednom području sive
tvari (jezgri, sloju), imaju istu putanju (i obično istu funkciju), a završe
(uspostavljajući sinapse) na neuronima u nekom drugom području sive tvari (jezgri,
sloju).
Neuronske putove može se istražiti metodama što se temelje na prirodnoj
pojavi aksonskog prenošenja. Putanju aksona može se prikazati autoradiografijom
radioaktivno označenih aminokiselina što aksonskim prenošenjem putuju
anterogradno (od some neurona do presinaptičkog završetka aksona). Ta se metoda
temelji na pojavi anterogradnog aksonskog prenošenja, tj. polaganog premještanja
raznovrsnih molekula i organela od tijela neurona do presinaptičkog završetka aksona
(za opis pojave vidi 6. poglavlje udžbenika!). U neko područje sive tvari (jezgru, sloj)
uštrca se finom iglom ili mikropipetom mala količina aminokiseline (npr. leucin ili
prolin) označene tricijem. Ta aminokiselina anterogradno se prenosi duž aksona te se
(nakon određenog vremena) može autoradiografskim postupkom (na histološkom rezu
tkiva) prikazati i dotični akson i njegovo završno razgranjenje, tj. ciljno područje u
kojemu završava. Tako su primjerice David Hubel i Thorsten Wiesel (Nobelova
nagrada za fiziologiju i medicinu 1981. godine!) uspješno prikazali kolumnarni
završetak talamokortikalnih aksona vidnog puta u primarnoj vidnoj moždanoj kori
(vidi 28. poglavlje). Putanju aksona može se prikazati histokemijski, na temelju
retrogradnog (od presinaptičkog aksonskog završetka prema somi) aksonskog
prenošenja peroksidaze hrena (HRP).
Ta se metoda temelji na pojavi retrogradnog aksonskog prenošenja, tj.
putovanja raznovrsnih molekula i organela od presinaptičkog završetka aksona do
some neurona (vidi također 6. poglavlje). U lokalizirano područje sive tvari uštrcamo
4

Temelji neuroznanosti Vodič kroz vježbe
malu količinu jednog enzima, peroksidaze hrena (engl., horseradish- peroxidase,
HRP). Enzim prodire u okolne presinaptičke aksonske završetke (endocitoza
posredstvom receptora - 6. poglavlje!) i potom retrogradnim aksonskim prenošenjem
dospije u somu neurona. Kako je na enzim prije uštrcavanja vezana neka
fluorescentna boja, tijela dotičnih neurona možemo prikazati histokemijskom
obradom histoloških rezova (histofluorescencija). Tako dobijemo odgovor na sljedeće
pitanje: odakle (i od koje vrste neurona) u dotično područje mozga dolaze aksoni?
Slika 1.1 Praćenje neuronskih putova metodom retrogradnog (lijevo) i anterogradnog (desno) aksonskog prenošenja. Za pojedinosti vidi tekst.
Dakle, metoda je posebno pogodna za istraživanje aferentnih projekcijskih
veza (npr. aferentnih putova moždane kore ili talamusa). Metodom dvostrukog
označavanja (retrogradnim prenošenjem dviju različitih fluorescentnih boja) može se
pokazati da se jedan akson svojim ograncima projicira u dva različita moždana
područja. Uvođenje te metode dovelo je do pravog prevrata u istraživanju nekih
moždanih putova. Jednu fluorescentnu boju uštrcamo u jedno, a drugu u drugo
područje sive tvari (npr. u dva različita kortikalna polja iste ili suprotne moždane
polutke). Obje boje retrogradnim aksonskim prenošenjem putuju do tijela neurona.
Ukoliko posebnom metodom histofluorescencije uočimo da tijelo jednog neurona
fluoresceira u dvije različite boje, doznali smo da je jedan ogranak aksona tog neurona
sinaptički završio u jednom, a drugi ogranak u drugom području sive tvari. Tako je
primjerice Patricia Goldman-Rakić u rezus majmuna pokazala da neki piramidni
neuroni III. sloja asocijacijske moždane kore čeonog režnja istodobno inerviraju
(preko ogranaka svojih aksona) i homolognu (čeonu) moždanu koru suprotne
5
Temelji neuroznanosti Vodič kroz vježbe
malu količinu jednog enzima, peroksidaze hrena (engl., horseradish- peroxidase,
HRP). Enzim prodire u okolne presinaptičke aksonske završetke (endocitoza
posredstvom receptora - 6. poglavlje!) i potom retrogradnim aksonskim prenošenjem
dospije u somu neurona. Kako je na enzim prije uštrcavanja vezana neka
fluorescentna boja, tijela dotičnih neurona možemo prikazati histokemijskom
obradom histoloških rezova (histofluorescencija). Tako dobijemo odgovor na sljedeće
pitanje: odakle (i od koje vrste neurona) u dotično područje mozga dolaze aksoni?
Slika 1.1 Praćenje neuronskih putova metodom retrogradnog (lijevo) i anterogradnog (desno) aksonskog prenošenja. Za pojedinosti vidi tekst.
Dakle, metoda je posebno pogodna za istraživanje aferentnih projekcijskih
veza (npr. aferentnih putova moždane kore ili talamusa). Metodom dvostrukog
označavanja (retrogradnim prenošenjem dviju različitih fluorescentnih boja) može se
pokazati da se jedan akson svojim ograncima projicira u dva različita moždana
područja. Uvođenje te metode dovelo je do pravog prevrata u istraživanju nekih
moždanih putova. Jednu fluorescentnu boju uštrcamo u jedno, a drugu u drugo
područje sive tvari (npr. u dva različita kortikalna polja iste ili suprotne moždane
polutke). Obje boje retrogradnim aksonskim prenošenjem putuju do tijela neurona.
Ukoliko posebnom metodom histofluorescencije uočimo da tijelo jednog neurona
fluoresceira u dvije različite boje, doznali smo da je jedan ogranak aksona tog neurona
sinaptički završio u jednom, a drugi ogranak u drugom području sive tvari. Tako je
primjerice Patricia Goldman-Rakić u rezus majmuna pokazala da neki piramidni
neuroni III. sloja asocijacijske moždane kore čeonog režnja istodobno inerviraju
(preko ogranaka svojih aksona) i homolognu (čeonu) moždanu koru suprotne
5

Temelji neuroznanosti Vodič kroz vježbe
moždane polutke (komisurna vlakna!) i tjemenu moždanu koru iste moždane
hemisfere (asocijacijska vlakna!) Upravo te veze, značajne za neurobiologiju
pozornosti i pamćenja, opisane su u 42. poglavlju udžbenika. Na sličan način moglo
se otkriti zapanjujuću činjenicu da jedan noradrenalinski neuron smješten u
moždanom deblu (locus coeruleus) istodobno inervira različita područja međumozga i
velikog mozga. Naime, aksoni takvih neurona imaju tisuće kolaterala, pri čemu jedna
kolaterala može uspostaviti sinapsu u moždanom deblu, druga u talamusu ili
hipotalamusu, a treća u moždanoj kori!
Sve tri opisane metode praćenja neuronskih veza mogu se primijeniti jedino na
pokusnim životinjama. No, odnedavno postoje karbocijaninske fluorescentne boje
(DiI, DiO) što se u istu svrhu mogu vrlo uspješno primijeniti (i za anterogradno, i za
retrogradno praćenje, i za dvostruko označavanje!) na debelim rezovima mrtvog tkiva
ljudskog mozga fiksiranog u formalinu. Time su otvorene neslućene mogućnosti za
istraživanje neuronskih veza izravno u ljudskom mozgu, kako odraslom, tako i
tijekom prenatalnog i postnatalnog razvoja. Dotad se o postojanju takvih veza
zaključivalo gotovo isključivo neizravno, uglavnom na temelju nalaza u mozgu
majmuna. Ne treba posebno isticati da čak ni mozak čimpanze ne može služiti kao
potpun model za eksperimentalno proučavanje ljudskog mozga. Dovoljno je
spomenuti sposobnost govora ili duševne bolesti. Stoga vam je zasigurno posve jasno
koliko je golemo značenje uvođenja tih metoda, ali isto tako i dostupnosti za izravno
proučavanje tkiva fetalnog i odraslog ljudskog mozga, naravno, uz poštivanje svih
etičkih načela. Imunocitokemijskim metodama možemo prikazati raznovrsne signalne
molekule i neurotransmitersku narav neurona. Moguće je načiniti monoklonska i
poliklonska protutijela na neki neurotransmiter ili njegov sintetski enzim i tako obojiti
samo one neurone što sintetiziraju taj neurotransmiter. Naime, na protutijelo se još
doda fluorescentna boja, ili se rabi neki drugi postupak vizualizacije, a takav neuron
lako promatramo svjetlosnim mikroskopom. Riječ je o imunocitokemijskim ili
imunohistokemijskim metodama prikaza odabranih tkivnih elemenata u histološkim
rezovima moždanog tkiva. Tako se mogu prikazati ključni enzimi za sintezu klasičnih
neurotransmitera: kolin-acetiltransferaza (acetilkolinski neuroni), tirozin-hidroksilaza
(dopaminski neuroni), dopamin-hidroksilaza (noradrenalinski neuroni), a u novije
doba mogu se prikazati i sami neurotransmiteri (npr. acetilkolin, glutamat, GABA,
serotonin). No, ta je metoda posebno pogodna za prikaz neuroaktivnih peptida (npr.
neuropeptida Y). Ukratko, ključna prednost imunocitokemijske metode je u tome što
6
Temelji neuroznanosti Vodič kroz vježbe
moždane polutke (komisurna vlakna!) i tjemenu moždanu koru iste moždane
hemisfere (asocijacijska vlakna!) Upravo te veze, značajne za neurobiologiju
pozornosti i pamćenja, opisane su u 42. poglavlju udžbenika. Na sličan način moglo
se otkriti zapanjujuću činjenicu da jedan noradrenalinski neuron smješten u
moždanom deblu (locus coeruleus) istodobno inervira različita područja međumozga i
velikog mozga. Naime, aksoni takvih neurona imaju tisuće kolaterala, pri čemu jedna
kolaterala može uspostaviti sinapsu u moždanom deblu, druga u talamusu ili
hipotalamusu, a treća u moždanoj kori!
Sve tri opisane metode praćenja neuronskih veza mogu se primijeniti jedino na
pokusnim životinjama. No, odnedavno postoje karbocijaninske fluorescentne boje
(DiI, DiO) što se u istu svrhu mogu vrlo uspješno primijeniti (i za anterogradno, i za
retrogradno praćenje, i za dvostruko označavanje!) na debelim rezovima mrtvog tkiva
ljudskog mozga fiksiranog u formalinu. Time su otvorene neslućene mogućnosti za
istraživanje neuronskih veza izravno u ljudskom mozgu, kako odraslom, tako i
tijekom prenatalnog i postnatalnog razvoja. Dotad se o postojanju takvih veza
zaključivalo gotovo isključivo neizravno, uglavnom na temelju nalaza u mozgu
majmuna. Ne treba posebno isticati da čak ni mozak čimpanze ne može služiti kao
potpun model za eksperimentalno proučavanje ljudskog mozga. Dovoljno je
spomenuti sposobnost govora ili duševne bolesti. Stoga vam je zasigurno posve jasno
koliko je golemo značenje uvođenja tih metoda, ali isto tako i dostupnosti za izravno
proučavanje tkiva fetalnog i odraslog ljudskog mozga, naravno, uz poštivanje svih
etičkih načela. Imunocitokemijskim metodama možemo prikazati raznovrsne signalne
molekule i neurotransmitersku narav neurona. Moguće je načiniti monoklonska i
poliklonska protutijela na neki neurotransmiter ili njegov sintetski enzim i tako obojiti
samo one neurone što sintetiziraju taj neurotransmiter. Naime, na protutijelo se još
doda fluorescentna boja, ili se rabi neki drugi postupak vizualizacije, a takav neuron
lako promatramo svjetlosnim mikroskopom. Riječ je o imunocitokemijskim ili
imunohistokemijskim metodama prikaza odabranih tkivnih elemenata u histološkim
rezovima moždanog tkiva. Tako se mogu prikazati ključni enzimi za sintezu klasičnih
neurotransmitera: kolin-acetiltransferaza (acetilkolinski neuroni), tirozin-hidroksilaza
(dopaminski neuroni), dopamin-hidroksilaza (noradrenalinski neuroni), a u novije
doba mogu se prikazati i sami neurotransmiteri (npr. acetilkolin, glutamat, GABA,
serotonin). No, ta je metoda posebno pogodna za prikaz neuroaktivnih peptida (npr.
neuropeptida Y). Ukratko, ključna prednost imunocitokemijske metode je u tome što
6

Temelji neuroznanosti Vodič kroz vježbe
prikaže cijeli neuron kao i Golgijeva metoda, ali pritom još i doznamo koji neuroni
sintetiziraju koji neurotransmiter ili neuropeptid, što je prvi korak u upoznavanju
njihove funkcije u sklopu neuronskih krugova. Osim neurotransmitera i njihovih
enzima, tom metodom mogu se prikazati i drugi sastojci neurona, npr. sastojci
citoskeleta, podjedinice receptornih molekula ili ionskih kanala, te druge vrste
zanimljivih molekula.
Ključna prednost imunocitokemijskih metoda je u tome što se mogu izravno
primijeniti na histološke rezove fiksiranog ljudskog mozga.
1.2 Mikroskopiranje
Nastavno štivo: Temelji neuroznanosti, Poglavlje 2.
Svaki će student za vježbu mikroskopiranja dobiti tri različita mikroskopska
preparata. Molimo Vas da preparate proučite i opišete koje ste stanice vidjeli, te
kojom je histološkom tehnikom preparat priređen:
Mikroskopski preparat 1:
Mikroskopski preparat 2:
Mikroskopski preparat 3:
7
Temelji neuroznanosti Vodič kroz vježbe
prikaže cijeli neuron kao i Golgijeva metoda, ali pritom još i doznamo koji neuroni
sintetiziraju koji neurotransmiter ili neuropeptid, što je prvi korak u upoznavanju
njihove funkcije u sklopu neuronskih krugova. Osim neurotransmitera i njihovih
enzima, tom metodom mogu se prikazati i drugi sastojci neurona, npr. sastojci
citoskeleta, podjedinice receptornih molekula ili ionskih kanala, te druge vrste
zanimljivih molekula.
Ključna prednost imunocitokemijskih metoda je u tome što se mogu izravno
primijeniti na histološke rezove fiksiranog ljudskog mozga.
1.2 Mikroskopiranje
Nastavno štivo: Temelji neuroznanosti, Poglavlje 2.
Svaki će student za vježbu mikroskopiranja dobiti tri različita mikroskopska
preparata. Molimo Vas da preparate proučite i opišete koje ste stanice vidjeli, te
kojom je histološkom tehnikom preparat priređen:
Mikroskopski preparat 1:
Mikroskopski preparat 2:
Mikroskopski preparat 3:
7

Temelji neuroznanosti Vodič kroz vježbe
1.3 Sekcija mozga i kralježnične moždine
Nastavno štivo: Temelji neuroznanosti, Poglavlje 4 i 5.
Za proučavanje izgleda i raspodjele sive i bijele tvari kralježnične moždine i
mozga potrebno je napraviti presjeke anatomskih preparata ovih organa, a potom
prepoznati svaku pojedinu strukturu na horizontalnom, sagitalnom i frontalnom
presjeku. Postupite prema naputku za svaki pojedini presjek.
1.3.1 Veliki mozak
1.3.1.1 Veliki mozak – mediosagitalni presjek
Naputak: Velikim nožem presijecite mozak u mediosagitalnoj ravnini i
odvojite hemisfere. Označite one strukture koje ste na presjeku vidjeli:
Comissura anterior ٱ
Lamina terminalis ٱ
Chiasma opticum ٱ
Hypothalamus ٱ
Infundibulum ٱ
Corpus mamillare ٱ
Thalamus ٱ
Corpus callosum ٱ
Septum pellucidum ٱ
Fornix ٱ
Foramen interventriculare ٱ
Comisura epithalamica ٱ
Gyrus cinguli ٱ
Sulcus cinguli ٱ
Gyrus hippocampi ٱ
Sulcus collateralis ٱ
Sulcus parietooccipitalis ٱ
8
Temelji neuroznanosti Vodič kroz vježbe
1.3 Sekcija mozga i kralježnične moždine
Nastavno štivo: Temelji neuroznanosti, Poglavlje 4 i 5.
Za proučavanje izgleda i raspodjele sive i bijele tvari kralježnične moždine i
mozga potrebno je napraviti presjeke anatomskih preparata ovih organa, a potom
prepoznati svaku pojedinu strukturu na horizontalnom, sagitalnom i frontalnom
presjeku. Postupite prema naputku za svaki pojedini presjek.
1.3.1 Veliki mozak
1.3.1.1 Veliki mozak – mediosagitalni presjek
Naputak: Velikim nožem presijecite mozak u mediosagitalnoj ravnini i
odvojite hemisfere. Označite one strukture koje ste na presjeku vidjeli:
Comissura anterior ٱ
Lamina terminalis ٱ
Chiasma opticum ٱ
Hypothalamus ٱ
Infundibulum ٱ
Corpus mamillare ٱ
Thalamus ٱ
Corpus callosum ٱ
Septum pellucidum ٱ
Fornix ٱ
Foramen interventriculare ٱ
Comisura epithalamica ٱ
Gyrus cinguli ٱ
Sulcus cinguli ٱ
Gyrus hippocampi ٱ
Sulcus collateralis ٱ
Sulcus parietooccipitalis ٱ
8

Temelji neuroznanosti Vodič kroz vježbe
1.3.1.2 Veliki mozak-frontalni presjek
Naputak: Velikim nožem napravite frontalni presjek kroz corpora mamillaria.
Označite strukture koje ste na presjeku vidjeli:
Corpus callosum ٱ
Septum pellucidum ٱ
Ventriculus lateralis, pars centralis ٱ
Fornix ٱ
Nucleus caudatus, corpus ٱ
Thalamus ٱ
Capsula interna ٱ
Globus pallidus ٱ
Putamen ٱ
Capsula externa ٱ
Claustrum ٱ
Capsula exstrema ٱ
Ventriculus tertius ٱ
Hypothalamus ٱ
Hippocampus ٱ
Corpus amigdaloideum ٱ
Nucleus caudatus, cauda ٱ
Ventriculus lateralis, cornu temporale ٱ
1.3.1.3 Veliki mozak-horizontalni presjeci
Naputak: Velikim nožem napravite seriju horizontalnih presjeka režući od
vrha hemisfera prema bazi mozga. Na horizontalnom presjeku kroz središnji dio treće
moždane komore označite strukture koje ste vidjeli:
Corpus callosum ٱ
Ventriculus lateralis, cornu frontale ٱ
Fornix ٱ
Nucleus caudatus, caput ٱ
Thalamus ٱ
Capsula interna ٱ
9
Temelji neuroznanosti Vodič kroz vježbe
1.3.1.2 Veliki mozak-frontalni presjek
Naputak: Velikim nožem napravite frontalni presjek kroz corpora mamillaria.
Označite strukture koje ste na presjeku vidjeli:
Corpus callosum ٱ
Septum pellucidum ٱ
Ventriculus lateralis, pars centralis ٱ
Fornix ٱ
Nucleus caudatus, corpus ٱ
Thalamus ٱ
Capsula interna ٱ
Globus pallidus ٱ
Putamen ٱ
Capsula externa ٱ
Claustrum ٱ
Capsula exstrema ٱ
Ventriculus tertius ٱ
Hypothalamus ٱ
Hippocampus ٱ
Corpus amigdaloideum ٱ
Nucleus caudatus, cauda ٱ
Ventriculus lateralis, cornu temporale ٱ
1.3.1.3 Veliki mozak-horizontalni presjeci
Naputak: Velikim nožem napravite seriju horizontalnih presjeka režući od
vrha hemisfera prema bazi mozga. Na horizontalnom presjeku kroz središnji dio treće
moždane komore označite strukture koje ste vidjeli:
Corpus callosum ٱ
Ventriculus lateralis, cornu frontale ٱ
Fornix ٱ
Nucleus caudatus, caput ٱ
Thalamus ٱ
Capsula interna ٱ
9

Temelji neuroznanosti Vodič kroz vježbe
Globus pallidus ٱ
Putamen ٱ
Capsula externa ٱ
Claustrum ٱ
Capsula exstrema ٱ
Ventriculus tertius ٱ
Nucleus caudatus, cauda ٱ
Ventriculus lateralis, cornu occipitale ٱ
Hippocampus ٱ
Gyrus hippocampi ٱ
1.3.2 Mali mozak
Naputak: Odvojite mali mozak od velikog mozga i moždanog debla rezom
kroz korijenove malog mozga. Označite strukture koje ste vidjeli na površini malog
mozga:
Vermis ٱ
Nodulus ٱ
Flocculus ٱ
Hemispherium cerebelli ٱ
1.3.3 Moždano deblo
1.3.3.1 Moždano deblo – baza
Naputak: Moždano deblo odvojite od velikog mozga rezom kroz srednji
mozak. Na bazi moždanog debla označite strukture koje ste vidjeli:
Pons ٱ
Fissura mediana anterior medullae oblongatae ٱ
Pyramis ٱ
Oliva ٱ
10
Temelji neuroznanosti Vodič kroz vježbe
Globus pallidus ٱ
Putamen ٱ
Capsula externa ٱ
Claustrum ٱ
Capsula exstrema ٱ
Ventriculus tertius ٱ
Nucleus caudatus, cauda ٱ
Ventriculus lateralis, cornu occipitale ٱ
Hippocampus ٱ
Gyrus hippocampi ٱ
1.3.2 Mali mozak
Naputak: Odvojite mali mozak od velikog mozga i moždanog debla rezom
kroz korijenove malog mozga. Označite strukture koje ste vidjeli na površini malog
mozga:
Vermis ٱ
Nodulus ٱ
Flocculus ٱ
Hemispherium cerebelli ٱ
1.3.3 Moždano deblo
1.3.3.1 Moždano deblo – baza
Naputak: Moždano deblo odvojite od velikog mozga rezom kroz srednji
mozak. Na bazi moždanog debla označite strukture koje ste vidjeli:
Pons ٱ
Fissura mediana anterior medullae oblongatae ٱ
Pyramis ٱ
Oliva ٱ
10

Temelji neuroznanosti Vodič kroz vježbe
1.3.3.2 Moždano deblo – dorzalni dio
Naputak: na dorzalnom dijelu moždanog debla označite strukture koje ste
vidjeli:
Colliculus superior ٱ
Colliculus inferior ٱ
Pedunculus cerebri ٱ
Pedunculus cerebellaris medius ٱ
Pedunculus cerebellaris superior ٱ
Velum medullare superius ٱ
Fossa rhomboidea ٱ
Sulcus medianus posterior medullae oblongatae ٱ
1.3.4 Kralježnična moždina – poprečni presjek
Naputak: Na poprečnom presjeku označite strukture koje ste vidjeli:
Cornu ventrale ٱ
Cornu laterale ٱ
Cornu dorsale ٱ
Funiculus ventarlis ٱ
Funiculus lateralis ٱ
Funiculus dorsalis ٱ
Sulcus dorsolateralis ٱ
Sulcus ventrolateralis ٱ
Fissura mediana ventralis ٱ
Sulcus medianus dorsalis ٱ
Radix ventralis ٱ
Radix dorsalis ٱ
11
Temelji neuroznanosti Vodič kroz vježbe
1.3.3.2 Moždano deblo – dorzalni dio
Naputak: na dorzalnom dijelu moždanog debla označite strukture koje ste
vidjeli:
Colliculus superior ٱ
Colliculus inferior ٱ
Pedunculus cerebri ٱ
Pedunculus cerebellaris medius ٱ
Pedunculus cerebellaris superior ٱ
Velum medullare superius ٱ
Fossa rhomboidea ٱ
Sulcus medianus posterior medullae oblongatae ٱ
1.3.4 Kralježnična moždina – poprečni presjek
Naputak: Na poprečnom presjeku označite strukture koje ste vidjeli:
Cornu ventrale ٱ
Cornu laterale ٱ
Cornu dorsale ٱ
Funiculus ventarlis ٱ
Funiculus lateralis ٱ
Funiculus dorsalis ٱ
Sulcus dorsolateralis ٱ
Sulcus ventrolateralis ٱ
Fissura mediana ventralis ٱ
Sulcus medianus dorsalis ٱ
Radix ventralis ٱ
Radix dorsalis ٱ
11

Temelji neuroznanosti Vodič kroz vježbe
2. Kliničko-anatomski sindromi kralježnične moždine
Cilj vježbe je osposobiti studenta da primjenom dosadašnjih saznanja o građi i
unutrašnjem ustrojstvu kralježnične moždine riješi zadani klinički problem.
2.1 Uvod
Osim usvajanja činjenica o građi središnjega živčanog sustava osobito je
važno razvijati sposobnost primjene tog znanja u kliničkoj praksi. To se najbolje
postiže rješavanjem kliničkih problema. U središtu pozornosti svakog kliničkog
problema nije struktura, već pacijent s određenim simptomima. Na Vama je da
primjenom znanja o anatomskim strukturama pretpostavite koja je od struktura
zahvaćena bolešću ili ozlijeđena.
Za vježbu je potreban program Lamperti A, Sodicoff M., Oxbery B,
Neuroanatomy Laboratory Assistant. Program sadrži atlas s opisom struktura i
kliničke primjere. Kao uvod u vježbu pregledat ćemo atlas kralježnične moždine, a
potom riješiti zadane kliničke slučajeve.
2.2 1. Vježba
Nastavno štivo: Temelji neuroznanosti, Poglavlje 14.
Naputak: Pokrenite program Neuroanatomy. Otvorite atlas kliknuvši na ikonu
atlas i odaberite kralježničnu moždinu. Obnovite znanje o putovima i jezgrama
kralježnične moždine studirajući presjeke različitih segmenata.
12
Temelji neuroznanosti Vodič kroz vježbe
2. Kliničko-anatomski sindromi kralježnične moždine
Cilj vježbe je osposobiti studenta da primjenom dosadašnjih saznanja o građi i
unutrašnjem ustrojstvu kralježnične moždine riješi zadani klinički problem.
2.1 Uvod
Osim usvajanja činjenica o građi središnjega živčanog sustava osobito je
važno razvijati sposobnost primjene tog znanja u kliničkoj praksi. To se najbolje
postiže rješavanjem kliničkih problema. U središtu pozornosti svakog kliničkog
problema nije struktura, već pacijent s određenim simptomima. Na Vama je da
primjenom znanja o anatomskim strukturama pretpostavite koja je od struktura
zahvaćena bolešću ili ozlijeđena.
Za vježbu je potreban program Lamperti A, Sodicoff M., Oxbery B,
Neuroanatomy Laboratory Assistant. Program sadrži atlas s opisom struktura i
kliničke primjere. Kao uvod u vježbu pregledat ćemo atlas kralježnične moždine, a
potom riješiti zadane kliničke slučajeve.
2.2 1. Vježba
Nastavno štivo: Temelji neuroznanosti, Poglavlje 14.
Naputak: Pokrenite program Neuroanatomy. Otvorite atlas kliknuvši na ikonu
atlas i odaberite kralježničnu moždinu. Obnovite znanje o putovima i jezgrama
kralježnične moždine studirajući presjeke različitih segmenata.
12
Temelji neuroznanosti Vodič kroz vježbe
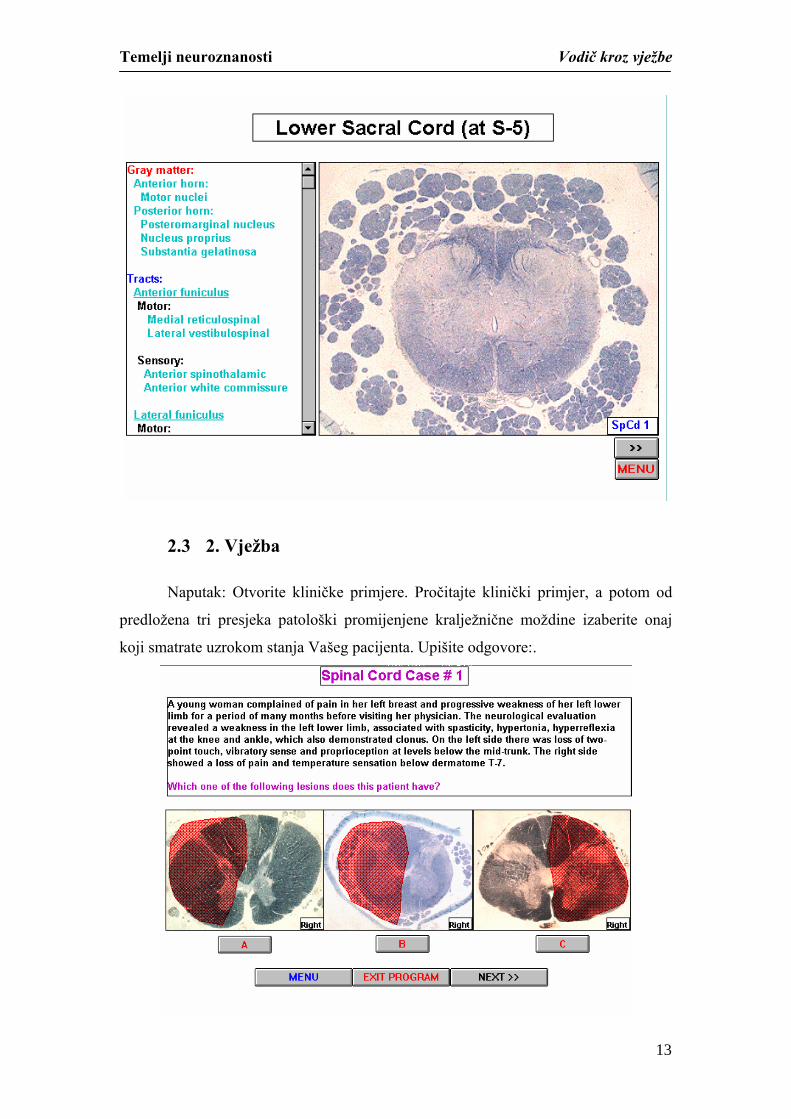
2.3 2. Vježba
Naputak: Otvorite kliničke primjere. Pročitajte klinički primjer, a potom od
predložena tri presjeka patološki promijenjene kralježnične moždine izaberite onaj
koji smatrate uzrokom stanja Vašeg pacijenta. Upišite odgovore:.
13
Temelji neuroznanosti Vodič kroz vježbe
2.3 2. Vježba
Naputak: Otvorite kliničke primjere. Pročitajte klinički primjer, a potom od
predložena tri presjeka patološki promijenjene kralježnične moždine izaberite onaj
koji smatrate uzrokom stanja Vašeg pacijenta. Upišite odgovore:.
13

Temelji neuroznanosti Vodič kroz vježbe
Primjer 1:______________________________________________________
Primjer 2:______________________________________________________
Primjer 3:______________________________________________________
Primjer 4:______________________________________________________
Primjer 5:______________________________________________________
Primjer 6:______________________________________________________
Primjer 7:______________________________________________________
Primjer 8:______________________________________________________
Primjer 9:______________________________________________________
Primjer 10:_____________________________________________________
14
Temelji neuroznanosti Vodič kroz vježbe
Primjer 1:______________________________________________________
Primjer 2:______________________________________________________
Primjer 3:______________________________________________________
Primjer 4:______________________________________________________
Primjer 5:______________________________________________________
Primjer 6:______________________________________________________
Primjer 7:______________________________________________________
Primjer 8:______________________________________________________
Primjer 9:______________________________________________________
Primjer 10:_____________________________________________________
14

Temelji neuroznanosti Vodič kroz vježbe
2A. Računalni pregled neuroanatomskih struktura
Cilj vježbe je omogućiti studentu da pregledom brojnih presjeka mozga i
kralježnične moždine ostvari uvid u povezanost struktura središnjega živčanog
sustava.
2.4 Uvod
Studiranje morfologije središnjega živčanog sustava podijeljeno je iz
edukativnih razloga u nekoliko cjelina. Ipak, sve su strukture, bez obzira da li
pripadaju malom mozgu, velikom mozgu, moždanom deblu ili kralježničnoj moždini,
međusobno povezane ili se neprekinute protežu iz jednog dijela središnjega živčanog
sustava u drugi. Stoga, osim znanja o postojanju i smještaju neke jezgre ili puta,
osobito je važno dobro naučiti međusobne odnose i povezanost te strukture sa
susjednim ili udaljenim strukturama.
Naputak: Pokrenite program Lamperti A, Sodicoff M., Oxbery B,
Neuroanatomy Laboratory Assistant. Svaki student dobit će pet zadataka. Na prvi
zadatak morate uz pomoć atlasa odgovoriti kući.
Zadatak 1. Nađite u atlasu strukture središnjega živčanog sustava koje prenose
osjet boli i stavite ih u odgovarajući redoslijed počevši od periferije (kože).
Osjet boli prenose:
Zadatak 2:
15
Temelji neuroznanosti Vodič kroz vježbe
2A. Računalni pregled neuroanatomskih struktura
Cilj vježbe je omogućiti studentu da pregledom brojnih presjeka mozga i
kralježnične moždine ostvari uvid u povezanost struktura središnjega živčanog
sustava.
2.4 Uvod
Studiranje morfologije središnjega živčanog sustava podijeljeno je iz
edukativnih razloga u nekoliko cjelina. Ipak, sve su strukture, bez obzira da li
pripadaju malom mozgu, velikom mozgu, moždanom deblu ili kralježničnoj moždini,
međusobno povezane ili se neprekinute protežu iz jednog dijela središnjega živčanog
sustava u drugi. Stoga, osim znanja o postojanju i smještaju neke jezgre ili puta,
osobito je važno dobro naučiti međusobne odnose i povezanost te strukture sa
susjednim ili udaljenim strukturama.
Naputak: Pokrenite program Lamperti A, Sodicoff M., Oxbery B,
Neuroanatomy Laboratory Assistant. Svaki student dobit će pet zadataka. Na prvi
zadatak morate uz pomoć atlasa odgovoriti kući.
Zadatak 1. Nađite u atlasu strukture središnjega živčanog sustava koje prenose
osjet boli i stavite ih u odgovarajući redoslijed počevši od periferije (kože).
Osjet boli prenose:
Zadatak 2:
15

Temelji neuroznanosti Vodič kroz vježbe
Odgovor
Zadatak 3:
Odgovor
Zadatak 4:
Odgovor
Zadatak 5:
Odgovor
16
Temelji neuroznanosti Vodič kroz vježbe
Odgovor
Zadatak 3:
Odgovor
Zadatak 4:
Odgovor
Zadatak 5:
Odgovor
16

Temelji neuroznanosti Vodič kroz vježbe
4. Potencijal u stanju mirovanja
4.1 Uvod
Razumijevanje mehanizma nastanka potencijala u stanju mirovanja temelj je
razumijevanja elektrofiziologije podražljivosti neurona i aksona.
Zajedničko svojstvo svih stanica jest postojanje razlike električnog potencijala
s dvije strane stanične membrane. Tu razliku nazivamo membranski ili
transmembranski potencijal. Membranski potencijal jako je važno stanično svojstvo
koje se može izjednačiti sa životom kao takvim. Stanice za održavanje života moraju
imati membranske potencijale. Stanice koje su izgubile sposobnost održavanja
električnog potencijala kroz svoju membranu su mrtve stanice.
Membranski potencijal neophodan je za izvođenje različitih staničnih funkcija
putem njenih organela. U živčanim stanicama, te mišićnim i žljezdanim stanicama
koje su s njima u vezi, membranski potencijal izvor je posebnih funkcija. Brze
promjene membranskog potencijala osnova su električnog signaliziranja. Iz
navedenoga očito je kako se studiranje električkih zbivanja u stanicama i izvan njih
mora započeti detaljnim razumijevanjem osnova membranskog potencijala.
U razmatranju sila koje djeluju na ione s jedne i druge strane membrane
moramo istaknuti jednadžbu koja opisuje stanje ravnoteže tj. ono stanje u kojemu su
dvije sile, električna i kemijska, jednake. To je jednadžba koju je izveo njemački
kemičar Walther Nernst 1988., a koja definira potencijal pri kojemu nema neto
protoka dotičnog iona kroz membranu. Ukoliko u vašem studiranju elektrofiziologije
želite upamtiti makar jednu jednadžbu, neka to bude upravo Nernstova jednadžba koja
opisuje temeljnu prirodu električnog potencijala u svim stanicama. Promjene
membranskog potencijala povezane su s različitim vidovima električne aktivnosti
stanica, kao što je akcijski potencijal, signaliziranje pomoću sinaptičkih veza i
stvaranje postsinaptičkih potencijala.
4.2 Ionske ravnoteže i membranski potencijal mirovanja
Kroz plazmatske membrane životinjskih stanica postoji razlika u električnom
potencijalu (napon). Citoplazma je obično električki negativna za razliku od
izvanstanične tekućine. Budući da razlika u električnom potencijalu postoji i u
17
Temelji neuroznanosti Vodič kroz vježbe
4. Potencijal u stanju mirovanja
4.1 Uvod
Razumijevanje mehanizma nastanka potencijala u stanju mirovanja temelj je
razumijevanja elektrofiziologije podražljivosti neurona i aksona.
Zajedničko svojstvo svih stanica jest postojanje razlike električnog potencijala
s dvije strane stanične membrane. Tu razliku nazivamo membranski ili
transmembranski potencijal. Membranski potencijal jako je važno stanično svojstvo
koje se može izjednačiti sa životom kao takvim. Stanice za održavanje života moraju
imati membranske potencijale. Stanice koje su izgubile sposobnost održavanja
električnog potencijala kroz svoju membranu su mrtve stanice.
Membranski potencijal neophodan je za izvođenje različitih staničnih funkcija
putem njenih organela. U živčanim stanicama, te mišićnim i žljezdanim stanicama
koje su s njima u vezi, membranski potencijal izvor je posebnih funkcija. Brze
promjene membranskog potencijala osnova su električnog signaliziranja. Iz
navedenoga očito je kako se studiranje električkih zbivanja u stanicama i izvan njih
mora započeti detaljnim razumijevanjem osnova membranskog potencijala.
U razmatranju sila koje djeluju na ione s jedne i druge strane membrane
moramo istaknuti jednadžbu koja opisuje stanje ravnoteže tj. ono stanje u kojemu su
dvije sile, električna i kemijska, jednake. To je jednadžba koju je izveo njemački
kemičar Walther Nernst 1988., a koja definira potencijal pri kojemu nema neto
protoka dotičnog iona kroz membranu. Ukoliko u vašem studiranju elektrofiziologije
želite upamtiti makar jednu jednadžbu, neka to bude upravo Nernstova jednadžba koja
opisuje temeljnu prirodu električnog potencijala u svim stanicama. Promjene
membranskog potencijala povezane su s različitim vidovima električne aktivnosti
stanica, kao što je akcijski potencijal, signaliziranje pomoću sinaptičkih veza i
stvaranje postsinaptičkih potencijala.
4.2 Ionske ravnoteže i membranski potencijal mirovanja
Kroz plazmatske membrane životinjskih stanica postoji razlika u električnom
potencijalu (napon). Citoplazma je obično električki negativna za razliku od
izvanstanične tekućine. Budući da razlika u električnom potencijalu postoji i u
17

Temelji neuroznanosti Vodič kroz vježbe
stanicama koje miruju, taj se potencijal obično naziva membranski potencijal
mirovanja. Membranski potencijal mirovanja vrlo je važan za podražljivost živčanih
i mišićnih stanica te za neke druge stanične reakcije.
Glavna je zadaća ovog prikaza razmotriti način kako gradijenti
elektrokemijskog potencijala pojedinih iona kroz plazmatsku membranu proizvode
membranski potencijal mirovanja.
U prvom se dijelu obrađuju neki osnovni pojmovi i definicije kojima se
opisuje protok iona kroz membrane.
4.3 Ionske ravnoteže
4.3.1 Elektrokemijski potencijali iona
Zamislimo jednostavan model u kojemu membrana odjeljuje vodenu otopinu u
dva odjeljka (A i B). Pretpostavimo da je koncentracija Na+ veća u odjeljku A nego u
odjeljku B. Ako između odjeljaka A i B nema razlike u električnom potencijalu, Na+
će difundirati iz odjeljka A u odjeljak B, baš kao da je riječ o molekuli bez naboja.
No, ako je odjeljak A električki negativan u odnosu prema odjeljku B, prilike postaju
mnogo složenije. Difuzija Na+ iz odjeljka A u odjeljak B zbog razlike u koncentraciji
postoji i dalje, ali se sada Na+ kreće i u suprotnome smjeru (iz B u A) zbog razlike u
električnom potencijalu kroz membranu. Smjer neto-putovanja Na+ ovisi o tome koje
djelovanje prevladava - učinak razlike u koncentraciji ili učinak razlike u električnom
potencijalu. Uspoređujući ta dva učinka, koncentracijski i električni, može se
predvidjeti smjer neto-protoka Na+.
Veličina pomoću koje možemo usporediti relativni udio koncentracije nekog
iona i električnog potencijala zove se elektrokemijski potencijal (µ) tog iona.
Elektrokemijski potencijal definira se kao
µ = µ0 + RTlnC + zFE (1)
gdje su: µ0 = elektrokemijski potencijal iona u nekom određenom stanju (npr. koncentracija 1 M, temperatura 0 ºC, električni potencijal jednak nuli), R = opća plinska konstanta, T = apsolutna temperatura, InC = prirodni logaritam koncentracije, z = broj naboja iona (+2 za Ca++, -1 za Cl-, itd.), F = Faradayev broj, E = električni potencijal.
18
Temelji neuroznanosti Vodič kroz vježbe
stanicama koje miruju, taj se potencijal obično naziva membranski potencijal
mirovanja. Membranski potencijal mirovanja vrlo je važan za podražljivost živčanih
i mišićnih stanica te za neke druge stanične reakcije.
Glavna je zadaća ovog prikaza razmotriti način kako gradijenti
elektrokemijskog potencijala pojedinih iona kroz plazmatsku membranu proizvode
membranski potencijal mirovanja.
U prvom se dijelu obrađuju neki osnovni pojmovi i definicije kojima se
opisuje protok iona kroz membrane.
4.3 Ionske ravnoteže
4.3.1 Elektrokemijski potencijali iona
Zamislimo jednostavan model u kojemu membrana odjeljuje vodenu otopinu u
dva odjeljka (A i B). Pretpostavimo da je koncentracija Na+ veća u odjeljku A nego u
odjeljku B. Ako između odjeljaka A i B nema razlike u električnom potencijalu, Na+
će difundirati iz odjeljka A u odjeljak B, baš kao da je riječ o molekuli bez naboja.
No, ako je odjeljak A električki negativan u odnosu prema odjeljku B, prilike postaju
mnogo složenije. Difuzija Na+ iz odjeljka A u odjeljak B zbog razlike u koncentraciji
postoji i dalje, ali se sada Na+ kreće i u suprotnome smjeru (iz B u A) zbog razlike u
električnom potencijalu kroz membranu. Smjer neto-putovanja Na+ ovisi o tome koje
djelovanje prevladava - učinak razlike u koncentraciji ili učinak razlike u električnom
potencijalu. Uspoređujući ta dva učinka, koncentracijski i električni, može se
predvidjeti smjer neto-protoka Na+.
Veličina pomoću koje možemo usporediti relativni udio koncentracije nekog
iona i električnog potencijala zove se elektrokemijski potencijal (µ) tog iona.
Elektrokemijski potencijal definira se kao
µ = µ0 + RTlnC + zFE (1)
gdje su: µ0 = elektrokemijski potencijal iona u nekom određenom stanju (npr. koncentracija 1 M, temperatura 0 ºC, električni potencijal jednak nuli), R = opća plinska konstanta, T = apsolutna temperatura, InC = prirodni logaritam koncentracije, z = broj naboja iona (+2 za Ca++, -1 za Cl-, itd.), F = Faradayev broj, E = električni potencijal.
18

Temelji neuroznanosti Vodič kroz vježbe
Jedinica µ svakoga člana jednadžbe (1) jest energija po molu. Elektrokemijski
je potencijal potencijalna kemijska energija koju ima jedan mol nekog iona zbog
svoje koncentracije i zbog električnog potencijala.
Neto-protok nekog iona zbivat će se iz mjesta gdje mu je elektrokemijski
potencijal veći prema mjestu gdje mu je elektrokemijski potencijal manji. Zamislimo
da membrana odjeljuje dva odjeljka (A i B), od kojih svaki sadrži otopljene ione x.
Sklonost iona kretanju iz A u B razmjerna je µ(x) na strani A, a sklonost iona kretanju
iz B u A razmjerna je µ(x) na strani B. Neto-sklonost iona x kretanju iz A u B je
µA(x) - µB(x). Ta razlika jest razlika u elektrokemijskom potencijalu iona x kroz
membranu (∆µ).
∆µ(x) = µΑ(x) - µΒ(x) (2)
Ako se u jednadžbu (2) uvrste vrijednosti µA(x) i µB(x) iz jednadžbe (1), dobit
će se:
µA(x) = µ0(x) + RTln[x]A + zFEA
µB(x) = µ0(x) + RTln[x]B + zFEB
pa je:
∆µ(x) = µΑ(x) - µΒ(x) = RTln([x]A/[x]B) + zF(EA-EB) (3)
Prvi član s desne strane jednadžbe (3) RTln([x]A/[x]B) predočuje težnju iona x
da se kreće iz A u B zbog razlike u koncentraciji, a drugi član zF(EA - EB) nastojanje
iona da se kreće iz A u B zbog razlike u električnom potencijalu. Prvi član označava
razliku u kemijskom potencijalu između jednog mola iona x u odjeljku A i jednog
mola iona x u odjeljku B koja nastaje zbog razlike u koncentraciji. Drugi pak član
označava razliku u električnom potencijalu između jednog mola iona x u odjeljku A i
jednog mola iona x u odjeljku B koja nastaje zbog razlike u električnom potencijalu
između A i B. Prema tome, ∆µ opisuje razliku u kemijskome potencijalu između
jednog mola iona x u odjeljku A i jednog mola iona x u odjeljku B koja nastaje i zbog
razlike u koncentraciji i zbog razlike u električnom potencijalu.
Ioni x putovat će iz mjesta gdje je elektrokemijski potencijal veći prema
mjestu gdje je manji. ∆µ definira se kao elektrokemijski potencijal iona u odjeljku A
minus elektrokemijski potencijal tog iona u odjeljku B. Ako je ∆µ pozitivan, ioni se
kreću iz A u B; ako je ∆µ nula, neto-putovanja iona uopće nema, a ako je ∆µ
negativan, ioni se kreću iz B u A.
19
Temelji neuroznanosti Vodič kroz vježbe
Jedinica µ svakoga člana jednadžbe (1) jest energija po molu. Elektrokemijski
je potencijal potencijalna kemijska energija koju ima jedan mol nekog iona zbog
svoje koncentracije i zbog električnog potencijala.
Neto-protok nekog iona zbivat će se iz mjesta gdje mu je elektrokemijski
potencijal veći prema mjestu gdje mu je elektrokemijski potencijal manji. Zamislimo
da membrana odjeljuje dva odjeljka (A i B), od kojih svaki sadrži otopljene ione x.
Sklonost iona kretanju iz A u B razmjerna je µ(x) na strani A, a sklonost iona kretanju
iz B u A razmjerna je µ(x) na strani B. Neto-sklonost iona x kretanju iz A u B je
µA(x) - µB(x). Ta razlika jest razlika u elektrokemijskom potencijalu iona x kroz
membranu (∆µ).
∆µ(x) = µΑ(x) - µΒ(x) (2)
Ako se u jednadžbu (2) uvrste vrijednosti µA(x) i µB(x) iz jednadžbe (1), dobit
će se:
µA(x) = µ0(x) + RTln[x]A + zFEA
µB(x) = µ0(x) + RTln[x]B + zFEB
pa je:
∆µ(x) = µΑ(x) - µΒ(x) = RTln([x]A/[x]B) + zF(EA-EB) (3)
Prvi član s desne strane jednadžbe (3) RTln([x]A/[x]B) predočuje težnju iona x
da se kreće iz A u B zbog razlike u koncentraciji, a drugi član zF(EA - EB) nastojanje
iona da se kreće iz A u B zbog razlike u električnom potencijalu. Prvi član označava
razliku u kemijskom potencijalu između jednog mola iona x u odjeljku A i jednog
mola iona x u odjeljku B koja nastaje zbog razlike u koncentraciji. Drugi pak član
označava razliku u električnom potencijalu između jednog mola iona x u odjeljku A i
jednog mola iona x u odjeljku B koja nastaje zbog razlike u električnom potencijalu
između A i B. Prema tome, ∆µ opisuje razliku u kemijskome potencijalu između
jednog mola iona x u odjeljku A i jednog mola iona x u odjeljku B koja nastaje i zbog
razlike u koncentraciji i zbog razlike u električnom potencijalu.
Ioni x putovat će iz mjesta gdje je elektrokemijski potencijal veći prema
mjestu gdje je manji. ∆µ definira se kao elektrokemijski potencijal iona u odjeljku A
minus elektrokemijski potencijal tog iona u odjeljku B. Ako je ∆µ pozitivan, ioni se
kreću iz A u B; ako je ∆µ nula, neto-putovanja iona uopće nema, a ako je ∆µ
negativan, ioni se kreću iz B u A.
19

Temelji neuroznanosti Vodič kroz vježbe
4.3.2 Elektrokemijska ravnoteža i Nernstova jednadžba
∆µ možemo zamisliti kao neto-silu na ion, dok je RTln([x]A/[x]B) sila
uzrokovana razlikom u koncentraciji, a zF(EA - EB) je sila uzrokovana razlikom u
električnom potencijalu. Kad su te dvije sile jednake i suprotnoga smjera, ∆µ = 0, tada
nema neto-sile na ion. Kad nema neto-sile na ion, neće biti ni neto-putovanja iona, pa
se kaže da kroz membranu za taj ion postoji elektrokemijska ravnoteža. U ravnoteži
∆µ = 0, pa iz jednadžbe (3) slijedi:
RTln([x]A/[x]B) + zF(EA - EB) = 0 (4) Potraživši rješenje za EA - EB, dobit ćemo:
EA - EB = -(RT/zF) . In([x]A/[x]B) = (RT/zF) . In([x]B/[x]A) (5)
Jednadžba (5) zove se Nernstova jednadžba. U njezinu se izvođenju
pretpostavlja da postoji stanje ravnoteže i zato Nernstova jednadžba vrijedi samo za
one ione koji su u ravnoteži. Pomoću nje može se izračunati potrebna razlika u
električnom potencijalu, EA - EB, da bi se proizvela električna sila zF(EA - EB), koja bi
bila jednaka i suprotna koncentracijskoj sili, RTln([x]A/[x]B) što pokreće ion iz A u B.
4.3.3 Primjena Nernstove jednadžbe.
Obično je Nernstovu jednadžbu korisno prevesti u oblik u kojemu se
primjenjuju dekadski (log10x) umjesto prirodnih logaritama (lnx = 2,303 log10x).
Budući da se biološki potencijali obično izražavaju u milivoltima (mV), jedinice za R
odabiru se tako da RT daje rezultat u milivoltima. Na 29,2 °C vrijednost 2,303RT/F
jednaka je 60 mV. Ta je vrijednost razmjerna apsolutnoj temperaturi, a približno se
mijenja za samo 1/273 za svaki stupanj. Prema tome, vrijednost od 60 mV za
2,303RT/F približno će vrijediti za većinu eksperimentalnih uvjeta, pa će koristan
oblik Nernstove jednadžbe biti:
EA - EB = -60 mV/z . Iog([x]A/[x]B) = 60 mV/z . Iog([x]B/[x]A) (6)
20
Temelji neuroznanosti Vodič kroz vježbe
4.3.2 Elektrokemijska ravnoteža i Nernstova jednadžba
∆µ možemo zamisliti kao neto-silu na ion, dok je RTln([x]A/[x]B) sila
uzrokovana razlikom u koncentraciji, a zF(EA - EB) je sila uzrokovana razlikom u
električnom potencijalu. Kad su te dvije sile jednake i suprotnoga smjera, ∆µ = 0, tada
nema neto-sile na ion. Kad nema neto-sile na ion, neće biti ni neto-putovanja iona, pa
se kaže da kroz membranu za taj ion postoji elektrokemijska ravnoteža. U ravnoteži
∆µ = 0, pa iz jednadžbe (3) slijedi:
RTln([x]A/[x]B) + zF(EA - EB) = 0 (4) Potraživši rješenje za EA - EB, dobit ćemo:
EA - EB = -(RT/zF) . In([x]A/[x]B) = (RT/zF) . In([x]B/[x]A) (5)
Jednadžba (5) zove se Nernstova jednadžba. U njezinu se izvođenju
pretpostavlja da postoji stanje ravnoteže i zato Nernstova jednadžba vrijedi samo za
one ione koji su u ravnoteži. Pomoću nje može se izračunati potrebna razlika u
električnom potencijalu, EA - EB, da bi se proizvela električna sila zF(EA - EB), koja bi
bila jednaka i suprotna koncentracijskoj sili, RTln([x]A/[x]B) što pokreće ion iz A u B.
4.3.3 Primjena Nernstove jednadžbe.
Obično je Nernstovu jednadžbu korisno prevesti u oblik u kojemu se
primjenjuju dekadski (log10x) umjesto prirodnih logaritama (lnx = 2,303 log10x).
Budući da se biološki potencijali obično izražavaju u milivoltima (mV), jedinice za R
odabiru se tako da RT daje rezultat u milivoltima. Na 29,2 °C vrijednost 2,303RT/F
jednaka je 60 mV. Ta je vrijednost razmjerna apsolutnoj temperaturi, a približno se
mijenja za samo 1/273 za svaki stupanj. Prema tome, vrijednost od 60 mV za
2,303RT/F približno će vrijediti za većinu eksperimentalnih uvjeta, pa će koristan
oblik Nernstove jednadžbe biti:
EA - EB = -60 mV/z . Iog([x]A/[x]B) = 60 mV/z . Iog([x]B/[x]A) (6)
20

Temelji neuroznanosti Vodič kroz vježbe
Primjeri za uporabu Nernstove jednadžbe: Primjer 1. Ako je K+ deset puta koncentriraniji u odjeljku A (0,1 M) nego u
odjeljku B (0,01 M), izračunat ćemo kolika razlika u električnom potencijalu mora
postojati između odjeljaka kako bi K+ bio u ravnoteži s jedne i s druge strane
membrane.
Budući da smo postavili zahtjev da K+ bude u ravnoteži, vrijedit će Nernstova
jednadžba.
EA - EB = -60 mV/+1 x log([K+]A/[K+]B)
= -60 mV log(0,1/0,01)(7) (7) = -60 mV log(10) = -60 mV
Nernstova nam jednadžba kaže da u stanju ravnoteže odjeljak A mora biti 60
mV negativan u odnosu prema odjeljku B. Možemo se uvjeriti da je takva polarnost
ispravna. Naime, zbog te električne sile K+ će se kretati iz B u A i time poništiti
njegovo kretanje iz A u B zbog razlike u koncentraciji.
Taj primjer pokazuje da razlika u električnom potencijalu od oko 60 mV može
održati ravnotežu desetorostrukoj razlici u koncentraciji nekog jednovalentnog iona.
To je korisno nepisano pravilo.
Primjer 2. Pomoću Nernstove jednadžbe možemo odlučiti primjerice je li
HCO3- u ravnoteži ili nije. Na primjer, ako je HCO3
- deset puta koncentriraniji u
odjeljku A (1 M) u kojem je pozitivan naboj, nego u odjeljku B (0,1 M) u kojem je
negativan naboj te EA - EB = +100 mV. Ukoliko HCO3-, nije u ravnoteži, Nernstova će
jednadžba odrediti smjer neto-protoka HCO3-.
Nernstova jednadžba kaže nam kolika je razlika u električnom potencijalu, EA
- EB, koja će upravo poništiti učinak razlike u koncentraciji HCO3- kroz membranu.
EA - EB = -60 mV/-1 x log([HCO3
-]A/[ HCO3-]B)
= 60 mV log(1/0,1) (8) = 60 mV log(10) = 60 mV
Dakle, razlika u potencijalu od +60 mV između A i B upravo bi održala
ravnotežu kretanju HCO3- iz A u B zbog razlike u koncentraciji. No, stvarni EA - EB
jest +100 mV, što znači da je predznak električnog potencijala takav da se može
suprotstaviti koncentracijskoj sili, ali da je istodobno za 40 mV veći od onog koji bi
bio potreban da upravo održi ravnotežu koncentracijskoj sili. Dakle, električna sila u
21
Temelji neuroznanosti Vodič kroz vježbe
Primjeri za uporabu Nernstove jednadžbe: Primjer 1. Ako je K+ deset puta koncentriraniji u odjeljku A (0,1 M) nego u
odjeljku B (0,01 M), izračunat ćemo kolika razlika u električnom potencijalu mora
postojati između odjeljaka kako bi K+ bio u ravnoteži s jedne i s druge strane
membrane.
Budući da smo postavili zahtjev da K+ bude u ravnoteži, vrijedit će Nernstova
jednadžba.
EA - EB = -60 mV/+1 x log([K+]A/[K+]B)
= -60 mV log(0,1/0,01)(7) (7) = -60 mV log(10) = -60 mV
Nernstova nam jednadžba kaže da u stanju ravnoteže odjeljak A mora biti 60
mV negativan u odnosu prema odjeljku B. Možemo se uvjeriti da je takva polarnost
ispravna. Naime, zbog te električne sile K+ će se kretati iz B u A i time poništiti
njegovo kretanje iz A u B zbog razlike u koncentraciji.
Taj primjer pokazuje da razlika u električnom potencijalu od oko 60 mV može
održati ravnotežu desetorostrukoj razlici u koncentraciji nekog jednovalentnog iona.
To je korisno nepisano pravilo.
Primjer 2. Pomoću Nernstove jednadžbe možemo odlučiti primjerice je li
HCO3- u ravnoteži ili nije. Na primjer, ako je HCO3
- deset puta koncentriraniji u
odjeljku A (1 M) u kojem je pozitivan naboj, nego u odjeljku B (0,1 M) u kojem je
negativan naboj te EA - EB = +100 mV. Ukoliko HCO3-, nije u ravnoteži, Nernstova će
jednadžba odrediti smjer neto-protoka HCO3-.
Nernstova jednadžba kaže nam kolika je razlika u električnom potencijalu, EA
- EB, koja će upravo poništiti učinak razlike u koncentraciji HCO3- kroz membranu.
EA - EB = -60 mV/-1 x log([HCO3
-]A/[ HCO3-]B)
= 60 mV log(1/0,1) (8) = 60 mV log(10) = 60 mV
Dakle, razlika u potencijalu od +60 mV između A i B upravo bi održala
ravnotežu kretanju HCO3- iz A u B zbog razlike u koncentraciji. No, stvarni EA - EB
jest +100 mV, što znači da je predznak električnog potencijala takav da se može
suprotstaviti koncentracijskoj sili, ali da je istodobno za 40 mV veći od onog koji bi
bio potreban da upravo održi ravnotežu koncentracijskoj sili. Dakle, električna sila u
21

Temelji neuroznanosti Vodič kroz vježbe
ovom primjeru na HCO3- veća je od koncentracijske sile, pa će to odrediti smjer neto-
kretanja iona HCO3-.
Neto-protok HCO3- bit će iz B u A.
Ukratko, pomoću Nernstove jednadžbe može se predvidjeti u kojem će smjeru
teći ioni i to:
1. Ako je razlika u potencijalu koja je izmjerena kroz membranu jednaka
razlici u potencijalu koja je za neki ion izračunata Nernstovom jednadžbom, onda je
taj ion u elektrokemijskoj ravnoteži pa kroz membranu neće biti neto-protoka tog
iona.
2. Ako izmjereni električni potencijal ima isti predznak kao potencijal
izračunat Nernstovom jednadžbom za neki određeni ion, ali je brojčano veći, onda je
električna sila veća od koncentracijske pa će neto-kretanje tog iona biti u smjeru
određenom električnom silom.
3. Kad je razlika u električnome potencijalu istoga predznaka, ali je brojčano
manja od vrijednosti izračunate Nernstovom jednadžbom za neki određeni ion, onda
je koncentracijska sila veća od električne sile, pa će neto-kretanje tog iona teći u
smjeru koji određuje razlika u koncentraciji.
4. Ako je izmjerena razlika u električnome potencijalu kroz membranu
suprotna predznaka od onog koji za određeni ion predviđa Nernstova jednadžba, onda
i električna sila i koncentracijska sila djeluju u istome smjeru. Prema tome, taj ion ne
može biti u ravnoteži i kretat će se u smjeru koji je određen i električnom i
koncentracijskom silom.
4.3.4 Izvođenje vježbe
U ovoj ćemo vježbi izvesti pokuse na mehanizmima kojima se ostvaruje
potencijal u stanju mirovanja.
Alan Hodgkin, Andrew Huxley i Bernard Katz radeći u Plymouth Biological
Laboratory u Engleskoj iskoristili su prednost rada na velikom, neki vole reći
divovskom (približno 1 mm u promjeru), aksonu lignje za intracelularno snimanje i
ispitivanje uloge iona u nastanaku akcijskog potencijala (Hodgkin & Katz, 1949).
Za simulaciju pokusa potrebno je pokrenuti aplikaciju CCWin. Preko
padajućeg izbornika File izaberite naredbu Open i pokrenite datoteku REST.CCS.
Simulaciju možete započeti tako da pomoću padajućeg izbornika Run izaberete
22
Temelji neuroznanosti Vodič kroz vježbe
ovom primjeru na HCO3- veća je od koncentracijske sile, pa će to odrediti smjer neto-
kretanja iona HCO3-.
Neto-protok HCO3- bit će iz B u A.
Ukratko, pomoću Nernstove jednadžbe može se predvidjeti u kojem će smjeru
teći ioni i to:
1. Ako je razlika u potencijalu koja je izmjerena kroz membranu jednaka
razlici u potencijalu koja je za neki ion izračunata Nernstovom jednadžbom, onda je
taj ion u elektrokemijskoj ravnoteži pa kroz membranu neće biti neto-protoka tog
iona.
2. Ako izmjereni električni potencijal ima isti predznak kao potencijal
izračunat Nernstovom jednadžbom za neki određeni ion, ali je brojčano veći, onda je
električna sila veća od koncentracijske pa će neto-kretanje tog iona biti u smjeru
određenom električnom silom.
3. Kad je razlika u električnome potencijalu istoga predznaka, ali je brojčano
manja od vrijednosti izračunate Nernstovom jednadžbom za neki određeni ion, onda
je koncentracijska sila veća od električne sile, pa će neto-kretanje tog iona teći u
smjeru koji određuje razlika u koncentraciji.
4. Ako je izmjerena razlika u električnome potencijalu kroz membranu
suprotna predznaka od onog koji za određeni ion predviđa Nernstova jednadžba, onda
i električna sila i koncentracijska sila djeluju u istome smjeru. Prema tome, taj ion ne
može biti u ravnoteži i kretat će se u smjeru koji je određen i električnom i
koncentracijskom silom.
4.3.4 Izvođenje vježbe
U ovoj ćemo vježbi izvesti pokuse na mehanizmima kojima se ostvaruje
potencijal u stanju mirovanja.
Alan Hodgkin, Andrew Huxley i Bernard Katz radeći u Plymouth Biological
Laboratory u Engleskoj iskoristili su prednost rada na velikom, neki vole reći
divovskom (približno 1 mm u promjeru), aksonu lignje za intracelularno snimanje i
ispitivanje uloge iona u nastanaku akcijskog potencijala (Hodgkin & Katz, 1949).
Za simulaciju pokusa potrebno je pokrenuti aplikaciju CCWin. Preko
padajućeg izbornika File izaberite naredbu Open i pokrenite datoteku REST.CCS.
Simulaciju možete započeti tako da pomoću padajućeg izbornika Run izaberete
22
Temelji neuroznanosti Vodič kroz vježbe
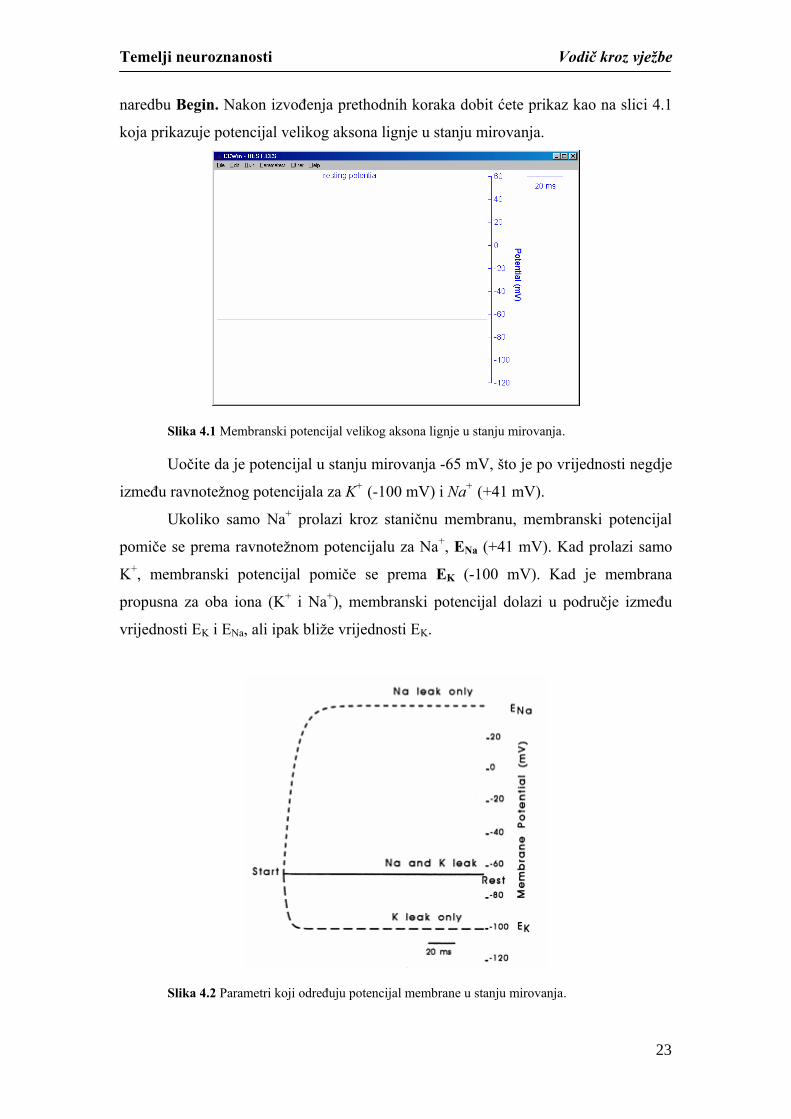
naredbu Begin. Nakon izvođenja prethodnih koraka dobit ćete prikaz kao na slici 4.1
koja prikazuje potencijal velikog aksona lignje u stanju mirovanja.
Slika 4.1 Membranski potencijal velikog aksona lignje u stanju mirovanja.
Uočite da je potencijal u stanju mirovanja -65 mV, što je po vrijednosti negdje
između ravnotežnog potencijala za K+ (-100 mV) i Na+ (+41 mV).
Ukoliko samo Na+ prolazi kroz staničnu membranu, membranski potencijal
pomiče se prema ravnotežnom potencijalu za Na+, ENa (+41 mV). Kad prolazi samo
K+, membranski potencijal pomiče se prema EK (-100 mV). Kad je membrana
propusna za oba iona (K+ i Na+), membranski potencijal dolazi u područje između
vrijednosti EK i ENa, ali ipak bliže vrijednosti EK.
Slika 4.2 Parametri koji određuju potencijal membrane u stanju mirovanja.
23
Temelji neuroznanosti Vodič kroz vježbe
naredbu Begin. Nakon izvođenja prethodnih koraka dobit ćete prikaz kao na slici 4.1
koja prikazuje potencijal velikog aksona lignje u stanju mirovanja.
Slika 4.1 Membranski potencijal velikog aksona lignje u stanju mirovanja.
Uočite da je potencijal u stanju mirovanja -65 mV, što je po vrijednosti negdje
između ravnotežnog potencijala za K+ (-100 mV) i Na+ (+41 mV).
Ukoliko samo Na+ prolazi kroz staničnu membranu, membranski potencijal
pomiče se prema ravnotežnom potencijalu za Na+, ENa (+41 mV). Kad prolazi samo
K+, membranski potencijal pomiče se prema EK (-100 mV). Kad je membrana
propusna za oba iona (K+ i Na+), membranski potencijal dolazi u područje između
vrijednosti EK i ENa, ali ipak bliže vrijednosti EK.
Slika 4.2 Parametri koji određuju potencijal membrane u stanju mirovanja.
23

Temelji neuroznanosti Vodič kroz vježbe
Prolazak iona kroz staničnu membranu u stanju mirovanja važan je za
nastanak potencijala mirovanja. Ioni prolaze niz koncentracijski gradijent (iz područja
više u područje niže koncentracije) ukoliko se nalaze u različitim koncentracijama s
vanjske i unutarnje strane membrane, a da pri tome postoji mogućnost prolaza kroz
membranu. Posljedica takvog kretanja iona je promjena membranskog potencijala kao
posljedica električkog naboja iona. Tako nastala promjena membranskog potencijala
ima učinak na distribuciju iona. Prisjetite se na trenutak srednje škole gdje ste učili da
se istoimeni naboji, primjerice pozitivni i pozitivni odbijaju, dok se raznoimeni
(pozitivni i negativni) privlače.
Kad se utjecaji koncentracijskog gradijenta i membranskog potencijala
izjednače, kažemo da se ioni nalaze "u stanju ravnoteže". Vrijednost potencijala
membrane koja u ravnoteži drži razliku u koncentracijskom gradijentu (primjerice -
100 mV potrebno je za sprječavanje izlaska K+ iz stanice zbog veće koncentracije u
stanici) naziva se ravnotežni potencijal. Ravnotežni potencijal može se izračunati
korištenjem Nernstove jednadžbe.
Goldman, Hodgkin i Katz napravili su modifikaciju Nernstove jednadžbe tj.
Goldman-Hodgkin-Katzovu jednadžbu konstantnog polja (GHK).
eClClP
iNaNaP
iKKP
iClClP
eNaNaP
eKKP
FRT
mV
−+
++
+
−+
++
+
= ln
GHK omogućuje izračunavanje potencijala s unutarnje strane membrane u
čijem nastanku sudjeluju dva jednovalentna kationa, natrij (Na+) i kalij (K+) i jedan
jednovalentan anion, klorid (Cl-). Detaljnije objašnjenje značenja te jednadžbe može
se vidjeti u udžbeniku.
4.4 Zadaci za vježbu
Vježba 4.1: Ravnotežni potencijal
Cilj vježbe je simulacija mijenjanja ravnotežnoga membranskog potencijala
ovisno o propusnosti membrane za ione Na+ i K+. U aplikaciji CCWIN propusnost za
K+ označena je s pKleak, dok je propusnost za Na+ označena s pNaleak.
24
Temelji neuroznanosti Vodič kroz vježbe
Prolazak iona kroz staničnu membranu u stanju mirovanja važan je za
nastanak potencijala mirovanja. Ioni prolaze niz koncentracijski gradijent (iz područja
više u područje niže koncentracije) ukoliko se nalaze u različitim koncentracijama s
vanjske i unutarnje strane membrane, a da pri tome postoji mogućnost prolaza kroz
membranu. Posljedica takvog kretanja iona je promjena membranskog potencijala kao
posljedica električkog naboja iona. Tako nastala promjena membranskog potencijala
ima učinak na distribuciju iona. Prisjetite se na trenutak srednje škole gdje ste učili da
se istoimeni naboji, primjerice pozitivni i pozitivni odbijaju, dok se raznoimeni
(pozitivni i negativni) privlače.
Kad se utjecaji koncentracijskog gradijenta i membranskog potencijala
izjednače, kažemo da se ioni nalaze "u stanju ravnoteže". Vrijednost potencijala
membrane koja u ravnoteži drži razliku u koncentracijskom gradijentu (primjerice -
100 mV potrebno je za sprječavanje izlaska K+ iz stanice zbog veće koncentracije u
stanici) naziva se ravnotežni potencijal. Ravnotežni potencijal može se izračunati
korištenjem Nernstove jednadžbe.
Goldman, Hodgkin i Katz napravili su modifikaciju Nernstove jednadžbe tj.
Goldman-Hodgkin-Katzovu jednadžbu konstantnog polja (GHK).
eClClP
iNaNaP
iKKP
iClClP
eNaNaP
eKKP
FRT
mV
−+
++
+
−+
++
+
= ln
GHK omogućuje izračunavanje potencijala s unutarnje strane membrane u
čijem nastanku sudjeluju dva jednovalentna kationa, natrij (Na+) i kalij (K+) i jedan
jednovalentan anion, klorid (Cl-). Detaljnije objašnjenje značenja te jednadžbe može
se vidjeti u udžbeniku.
4.4 Zadaci za vježbu
Vježba 4.1: Ravnotežni potencijal
Cilj vježbe je simulacija mijenjanja ravnotežnoga membranskog potencijala
ovisno o propusnosti membrane za ione Na+ i K+. U aplikaciji CCWIN propusnost za
K+ označena je s pKleak, dok je propusnost za Na+ označena s pNaleak.
24

Temelji neuroznanosti Vodič kroz vježbe
Stanični potencijal u stanju mirovanja može se izračunati pomoću GHK
jednadžbe. Pojava curenja (engl., leak) odnosi se na činjenicu da se propusnost
stanične membrane za ione Na+ i K+ ne mijenja s promjenama membranskog
potencijala. To dodatno objašnjava i činjenicu da i u stanju mirovanja postoji protok
iona kroz staničnu membranu.
Za ispitivanje utjecaja protoka iona potrebno je napraviti slijedeće:
• Ukoliko ste pokrenuli aplikaciju CCWIN i otvorili datoteku REST.CCS te
izabrali naredbu Begin iz padajućeg izbornika Run, na zaslonu je prikazan
početni dijagram koji pokazuje membranski potencijal u stanju mirovanja od
-65 mV (slika 4.1). Da bi simulirali promjenu membranskog potencijala
ovisno o propusnosti za ione Na+ i K+ potrebno je izabrati naredbu
Conductance iz padajućeg izbornika Parameters. Promijenite vrijednost za
natrij (pNaleak) u tablici s 0.06 u 0. Nakon što ste promijenili vrijednost
morate pritisnuti OK, a da bi vidjeli rezultat promjene trebate odabrati
naredbu Overlay iz padajućeg izbornika Run.
• Uočite kako se početna vrijednost membranskog potencijala od -65 mV brzo
promijenila na vrijednost približno -100 mV, koja odgovara ravnotežnom
potencijalu za K+ (vidi slike 4.2 i 4.3).
• Koristeći aplikaciju CCWIN u tablicu 4.1 upišite parametre koji nedostaju
kako bi se tablica u cijelosti popunila.
Tablica 4.1 Vrijednosti membranskog potencijala ovisno o parametrima propusnosti membrane za ione Na+ i K+
pNaleak pKleak Potencijal0.5 1 0 2
0.06 -65mV
• Proizvoljno smanjujte i povećavajte vrijednost propusnosti za ione Na+ i K+.
Zapišite rezultate barem deset vaših pokusa.
25
Temelji neuroznanosti Vodič kroz vježbe
Stanični potencijal u stanju mirovanja može se izračunati pomoću GHK
jednadžbe. Pojava curenja (engl., leak) odnosi se na činjenicu da se propusnost
stanične membrane za ione Na+ i K+ ne mijenja s promjenama membranskog
potencijala. To dodatno objašnjava i činjenicu da i u stanju mirovanja postoji protok
iona kroz staničnu membranu.
Za ispitivanje utjecaja protoka iona potrebno je napraviti slijedeće:
• Ukoliko ste pokrenuli aplikaciju CCWIN i otvorili datoteku REST.CCS te
izabrali naredbu Begin iz padajućeg izbornika Run, na zaslonu je prikazan
početni dijagram koji pokazuje membranski potencijal u stanju mirovanja od
-65 mV (slika 4.1). Da bi simulirali promjenu membranskog potencijala
ovisno o propusnosti za ione Na+ i K+ potrebno je izabrati naredbu
Conductance iz padajućeg izbornika Parameters. Promijenite vrijednost za
natrij (pNaleak) u tablici s 0.06 u 0. Nakon što ste promijenili vrijednost
morate pritisnuti OK, a da bi vidjeli rezultat promjene trebate odabrati
naredbu Overlay iz padajućeg izbornika Run.
• Uočite kako se početna vrijednost membranskog potencijala od -65 mV brzo
promijenila na vrijednost približno -100 mV, koja odgovara ravnotežnom
potencijalu za K+ (vidi slike 4.2 i 4.3).
• Koristeći aplikaciju CCWIN u tablicu 4.1 upišite parametre koji nedostaju
kako bi se tablica u cijelosti popunila.
Tablica 4.1 Vrijednosti membranskog potencijala ovisno o parametrima propusnosti membrane za ione Na+ i K+
pNaleak pKleak Potencijal0.5 1 0 2
0.06 -65mV
• Proizvoljno smanjujte i povećavajte vrijednost propusnosti za ione Na+ i K+.
Zapišite rezultate barem deset vaših pokusa.
25

Temelji neuroznanosti Vodič kroz vježbe
Slika 4.3 Promjena membranskog potencijala nakon promjene vrijednosti protoka Na+ iona (pNaleak = 0).
Poznato je da je koncentracija kalijevih iona značajno veća u unutarstaničnom
prostoru nego u izvanstaničnom. Kad smo započeli simulaciju na -65 mV, kalijevi
ioni prolazili su kroz staničnu membranu niz koncentracijski gradijent (smjerom iz
stanice prema van). Prilikom takvog kretanja kalijevih iona membranski potencijal
postaje više negativan što stvara silu (engl., 'pull') na K+ ione koja im ne dozvoljava
da napuste unutarstanični prostor. Nakon što težnja K+ iona za kretanjem niz
koncentracijski gradijent, tj. za izlaskom u izvanstanični prostor, bude izjednačena s
težnjom K+ da uđe u stanicu poradi negativnog naboja unutar stanice, ioni dolaze u
stanje ravnoteže. Kao posljedica tih procesa membranski potencijal ostaje jednak
ravnotežnom potencijalu.
Uočite kako, premda postoji neto protok iona kad je membranski potencijal po
vrijednosti daleko od ravnotežnog potencijala za odgovarajući ion, broj iona koji
prolaze kroz staničnu membranu u odnosu na ukupan broj iona unutar stanice i u
tekućem mediju između stanica izuzetno je malen. Stoga se koncentracija s obje
strane membrane mijenja u zanemarivo malim vrijednostima.
VJEŽBA 4.2: Učinci promjena koncentracije iona.
Cilj vježbe je ispitivanje učinka promjene unutarstanične i izvanstanične
koncentracije na vrijednost ravnotežnog potencijala.
Za izvođenje simulacije potrebno je napraviti nekoliko koraka:
• Ukoliko imate otvorenu datoteku REST.CCS odaberite naredbu Ions iz
padajućeg izbornika Parameters. Propusnost za natrij (pNaleak) ostavite na
26
Temelji neuroznanosti Vodič kroz vježbe
Slika 4.3 Promjena membranskog potencijala nakon promjene vrijednosti protoka Na+ iona (pNaleak = 0).
Poznato je da je koncentracija kalijevih iona značajno veća u unutarstaničnom
prostoru nego u izvanstaničnom. Kad smo započeli simulaciju na -65 mV, kalijevi
ioni prolazili su kroz staničnu membranu niz koncentracijski gradijent (smjerom iz
stanice prema van). Prilikom takvog kretanja kalijevih iona membranski potencijal
postaje više negativan što stvara silu (engl., 'pull') na K+ ione koja im ne dozvoljava
da napuste unutarstanični prostor. Nakon što težnja K+ iona za kretanjem niz
koncentracijski gradijent, tj. za izlaskom u izvanstanični prostor, bude izjednačena s
težnjom K+ da uđe u stanicu poradi negativnog naboja unutar stanice, ioni dolaze u
stanje ravnoteže. Kao posljedica tih procesa membranski potencijal ostaje jednak
ravnotežnom potencijalu.
Uočite kako, premda postoji neto protok iona kad je membranski potencijal po
vrijednosti daleko od ravnotežnog potencijala za odgovarajući ion, broj iona koji
prolaze kroz staničnu membranu u odnosu na ukupan broj iona unutar stanice i u
tekućem mediju između stanica izuzetno je malen. Stoga se koncentracija s obje
strane membrane mijenja u zanemarivo malim vrijednostima.
VJEŽBA 4.2: Učinci promjena koncentracije iona.
Cilj vježbe je ispitivanje učinka promjene unutarstanične i izvanstanične
koncentracije na vrijednost ravnotežnog potencijala.
Za izvođenje simulacije potrebno je napraviti nekoliko koraka:
• Ukoliko imate otvorenu datoteku REST.CCS odaberite naredbu Ions iz
padajućeg izbornika Parameters. Propusnost za natrij (pNaleak) ostavite na
26
Temelji neuroznanosti Vodič kroz vježbe
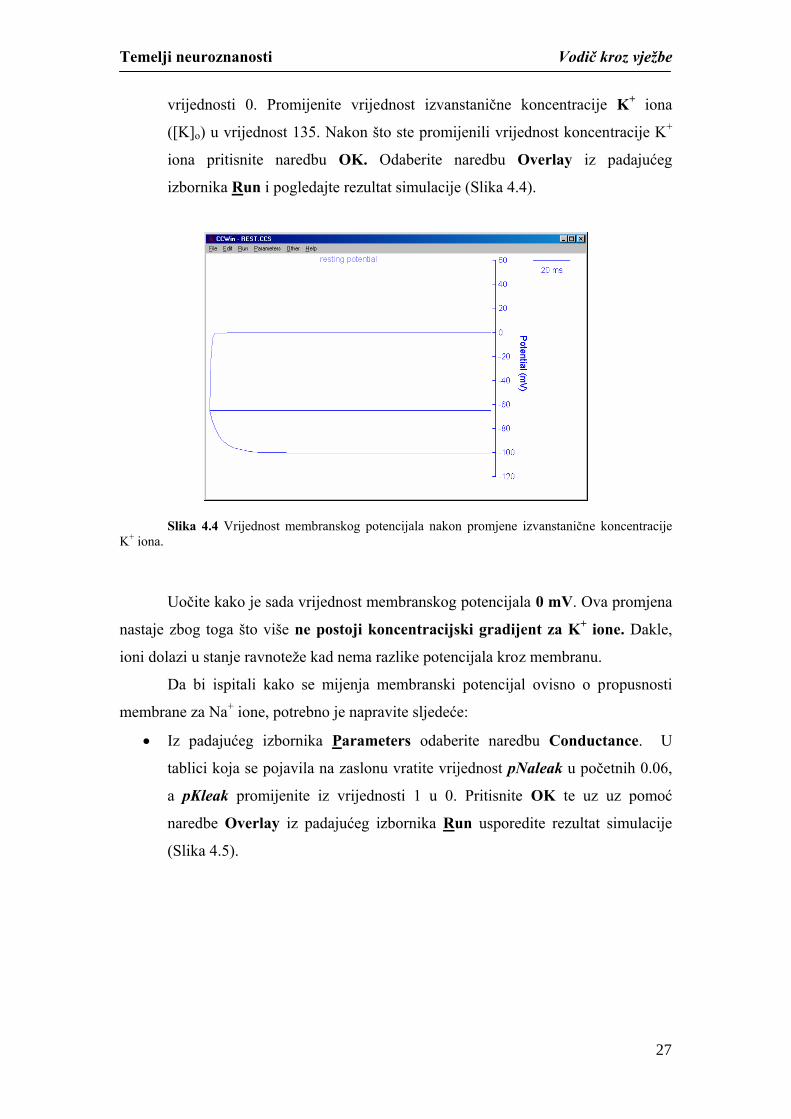
vrijednosti 0. Promijenite vrijednost izvanstanične koncentracije K+ iona
([K]o) u vrijednost 135. Nakon što ste promijenili vrijednost koncentracije K+
iona pritisnite naredbu OK. Odaberite naredbu Overlay iz padajućeg
izbornika Run i pogledajte rezultat simulacije (Slika 4.4).
Slika 4.4 Vrijednost membranskog potencijala nakon promjene izvanstanične koncentracije K+ iona.
Uočite kako je sada vrijednost membranskog potencijala 0 mV. Ova promjena
nastaje zbog toga što više ne postoji koncentracijski gradijent za K+ ione. Dakle,
ioni dolazi u stanje ravnoteže kad nema razlike potencijala kroz membranu.
Da bi ispitali kako se mijenja membranski potencijal ovisno o propusnosti
membrane za Na+ ione, potrebno je napravite sljedeće:
• Iz padajućeg izbornika Parameters odaberite naredbu Conductance. U
tablici koja se pojavila na zaslonu vratite vrijednost pNaleak u početnih 0.06,
a pKleak promijenite iz vrijednosti 1 u 0. Pritisnite OK te uz uz pomoć
naredbe Overlay iz padajućeg izbornika Run usporedite rezultat simulacije
(Slika 4.5).
27
Temelji neuroznanosti Vodič kroz vježbe
vrijednosti 0. Promijenite vrijednost izvanstanične koncentracije K+ iona
([K]o) u vrijednost 135. Nakon što ste promijenili vrijednost koncentracije K+
iona pritisnite naredbu OK. Odaberite naredbu Overlay iz padajućeg
izbornika Run i pogledajte rezultat simulacije (Slika 4.4).
Slika 4.4 Vrijednost membranskog potencijala nakon promjene izvanstanične koncentracije K+ iona.
Uočite kako je sada vrijednost membranskog potencijala 0 mV. Ova promjena
nastaje zbog toga što više ne postoji koncentracijski gradijent za K+ ione. Dakle,
ioni dolazi u stanje ravnoteže kad nema razlike potencijala kroz membranu.
Da bi ispitali kako se mijenja membranski potencijal ovisno o propusnosti
membrane za Na+ ione, potrebno je napravite sljedeće:
• Iz padajućeg izbornika Parameters odaberite naredbu Conductance. U
tablici koja se pojavila na zaslonu vratite vrijednost pNaleak u početnih 0.06,
a pKleak promijenite iz vrijednosti 1 u 0. Pritisnite OK te uz uz pomoć
naredbe Overlay iz padajućeg izbornika Run usporedite rezultat simulacije
(Slika 4.5).
27
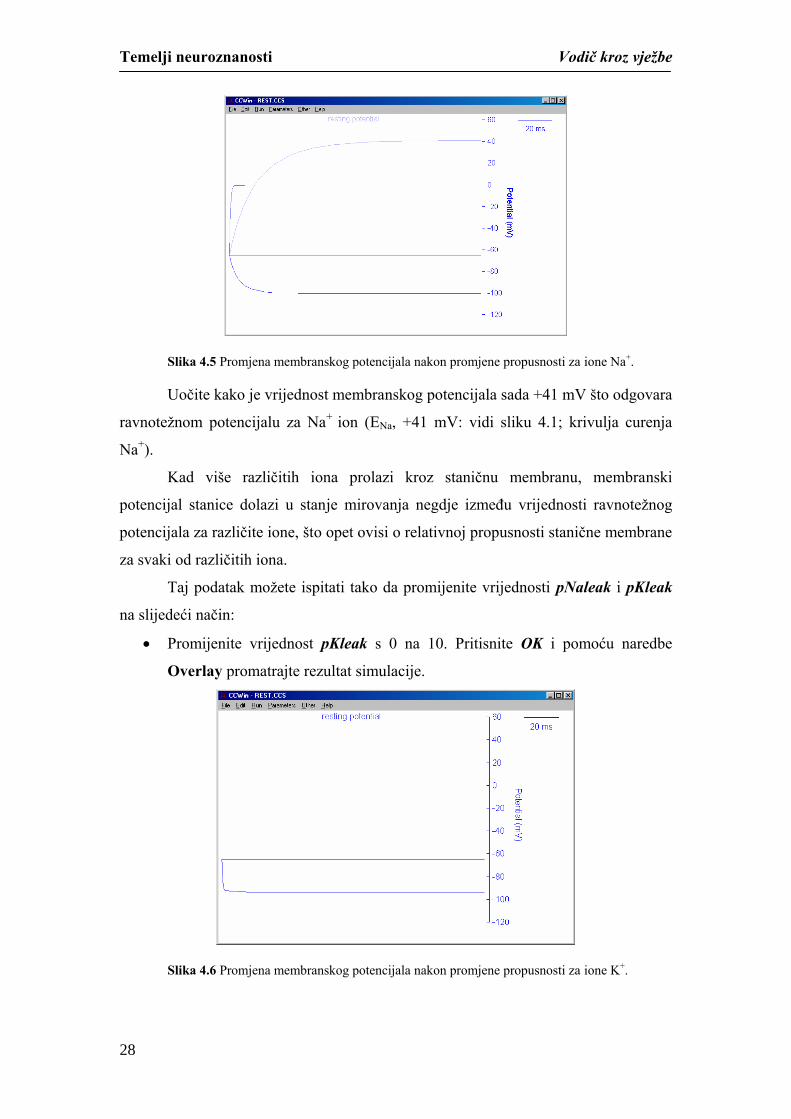
Temelji neuroznanosti Vodič kroz vježbe
Slika 4.5 Promjena membranskog potencijala nakon promjene propusnosti za ione Na+.
Uočite kako je vrijednost membranskog potencijala sada +41 mV što odgovara
ravnotežnom potencijalu za Na+ ion (ENa, +41 mV: vidi sliku 4.1; krivulja curenja
Na+).
Kad više različitih iona prolazi kroz staničnu membranu, membranski
potencijal stanice dolazi u stanje mirovanja negdje između vrijednosti ravnotežnog
potencijala za različite ione, što opet ovisi o relativnoj propusnosti stanične membrane
za svaki od različitih iona.
Taj podatak možete ispitati tako da promijenite vrijednosti pNaleak i pKleak
na slijedeći način:
• Promijenite vrijednost pKleak s 0 na 10. Pritisnite OK i pomoću naredbe
Overlay promatrajte rezultat simulacije.
Slika 4.6 Promjena membranskog potencijala nakon promjene propusnosti za ione K+.
28
Temelji neuroznanosti Vodič kroz vježbe
Slika 4.5 Promjena membranskog potencijala nakon promjene propusnosti za ione Na+.
Uočite kako je vrijednost membranskog potencijala sada +41 mV što odgovara
ravnotežnom potencijalu za Na+ ion (ENa, +41 mV: vidi sliku 4.1; krivulja curenja
Na+).
Kad više različitih iona prolazi kroz staničnu membranu, membranski
potencijal stanice dolazi u stanje mirovanja negdje između vrijednosti ravnotežnog
potencijala za različite ione, što opet ovisi o relativnoj propusnosti stanične membrane
za svaki od različitih iona.
Taj podatak možete ispitati tako da promijenite vrijednosti pNaleak i pKleak
na slijedeći način:
• Promijenite vrijednost pKleak s 0 na 10. Pritisnite OK i pomoću naredbe
Overlay promatrajte rezultat simulacije.
Slika 4.6 Promjena membranskog potencijala nakon promjene propusnosti za ione K+.
28

Temelji neuroznanosti Vodič kroz vježbe
Uočite kako je membranski potencijal bliže vrijednosti EK (-100 mV) nego
kada je vrijednost pKleak jednaka 1.
Poznato je da na veličinu potencijala u stanju mirovanja uglavnom utječe
distribucija K+, što je izravna posljedica veće propusnosti stanične membrane za K+ u
odnosu na propusnost za Na+. Pojednostavnili smo prilike u simulaciji tako da smo
namjestili propusnost za Na+ samo na dio propusnosti za K+ u stanju mirovanja.
Ispitajte što će se dogoditi ukoliko povećate izvanstaničnu koncentraciju Ca2+
iona s 2 na 10? Upišite zapažanja.
Provjerite kolike moraju biti unutarstanične i vanstanične koncentracije iona
da bi potencijal mirovanja bio -100mV? Koji ioni imaju najviše utjecaja na potencijal
membrane u mirovanju?
Vježba 4.3: Pasivna električna svojstva membrane
Osim snimanja membranskog potencijala aksona i neurona, uporabom
posebnih elektroničkih pojačala može se membranski potencijal izravno mijenjati s
promjenama veličine injicirane struje tj. podražaja. Dobiveni rezultati izravna su
posljedica aktivnih svojstava ionskih kanala smještenih u samoj membrani, a ne samo
pasivnih električnih svojstava plazmatske membrane.
Za izvođenje simulacije potrebno je napraviti slijedeće korake:
• Pokrenite program CCWIN te pomoću padajućeg izbornika File i naredbe
Open otvorite datoteku PASSIVE.CSS. Nakon što ste otvorili datoteku
izaberite naredbu Begin iz padajućeg izbornika Run. Dakle, na grafičkom
prikazu vidljiv je pasivni odgovor membrane.
29
Temelji neuroznanosti Vodič kroz vježbe
Uočite kako je membranski potencijal bliže vrijednosti EK (-100 mV) nego
kada je vrijednost pKleak jednaka 1.
Poznato je da na veličinu potencijala u stanju mirovanja uglavnom utječe
distribucija K+, što je izravna posljedica veće propusnosti stanične membrane za K+ u
odnosu na propusnost za Na+. Pojednostavnili smo prilike u simulaciji tako da smo
namjestili propusnost za Na+ samo na dio propusnosti za K+ u stanju mirovanja.
Ispitajte što će se dogoditi ukoliko povećate izvanstaničnu koncentraciju Ca2+
iona s 2 na 10? Upišite zapažanja.
Provjerite kolike moraju biti unutarstanične i vanstanične koncentracije iona
da bi potencijal mirovanja bio -100mV? Koji ioni imaju najviše utjecaja na potencijal
membrane u mirovanju?
Vježba 4.3: Pasivna električna svojstva membrane
Osim snimanja membranskog potencijala aksona i neurona, uporabom
posebnih elektroničkih pojačala može se membranski potencijal izravno mijenjati s
promjenama veličine injicirane struje tj. podražaja. Dobiveni rezultati izravna su
posljedica aktivnih svojstava ionskih kanala smještenih u samoj membrani, a ne samo
pasivnih električnih svojstava plazmatske membrane.
Za izvođenje simulacije potrebno je napraviti slijedeće korake:
• Pokrenite program CCWIN te pomoću padajućeg izbornika File i naredbe
Open otvorite datoteku PASSIVE.CSS. Nakon što ste otvorili datoteku
izaberite naredbu Begin iz padajućeg izbornika Run. Dakle, na grafičkom
prikazu vidljiv je pasivni odgovor membrane.
29

Temelji neuroznanosti Vodič kroz vježbe
• Putem padajućeg izbornika Parameters izaberite naredbu Protocol.
Promijenite vrijednost Injected current s 1.5 u 2.0. Nakon promjene pritisnite
OK, te pomoću naredbe Overlay iz izbornika Run pogledajte krajnji rezultat
(slika 4.7 i 4.8).
Slika 4.7 Pasivni i aktivni odgovor membrane na modelu aksona. (A) injiciranje strujnog udara od 1.5 nA ima za posljedicu pasivni odgovor koji je posljedica značajki membrane u vidu otpora i kapaciteta. (B) injiciranje struje od 2.0 nA rezultira serijom akcijskih potencijala.
Slika 4.8 Promjena membranskog potencijala nakon promjene veličine podražaja.
30
Temelji neuroznanosti Vodič kroz vježbe
• Putem padajućeg izbornika Parameters izaberite naredbu Protocol.
Promijenite vrijednost Injected current s 1.5 u 2.0. Nakon promjene pritisnite
OK, te pomoću naredbe Overlay iz izbornika Run pogledajte krajnji rezultat
(slika 4.7 i 4.8).
Slika 4.7 Pasivni i aktivni odgovor membrane na modelu aksona. (A) injiciranje strujnog udara od 1.5 nA ima za posljedicu pasivni odgovor koji je posljedica značajki membrane u vidu otpora i kapaciteta. (B) injiciranje struje od 2.0 nA rezultira serijom akcijskih potencijala.
Slika 4.8 Promjena membranskog potencijala nakon promjene veličine podražaja.
30

Temelji neuroznanosti Vodič kroz vježbe
4.5 Pitanja: Potencijal u stanju mirovanja
1. Koja su dva glavna čimbenika koja određuju prolaz iona kroz staničnu
membranu?
a) ___________________________
b) ___________________________
2. Što podrazumijeva pojam "ravnotežni potencijal" iona?
3. Zašto je ravnotežni potencijal K+ iona -100 mV, a Na+ iona +41 mV, premda
su oba pozitivni ioni?
4. Ako je potencijal stanice u stanju mirovanja -65mV, što bi se dogodilo ukoliko
mijenjate koncentraciju iona K+ i Na+ u izvanstaničnom i unutarstaničnom
prostoru? Provjerite ispravnost tvrdnje mijenjajući koncentracije iona pomoću
simulacije koristeći program CCWIN i datoteku REST.CCS.
5. Otvorite PASSIVE.CCS. Što očekujete da će se dogoditi s veličinom
membranskog potencijala i odgovorom na depolarizacijsku struju ukoliko
udvostručite i pKleak i pNaleak? Pokušajte izvesti taj eksperiment i provjerite
jeste li u pravu. Ukratko objasnite rezultat.
31
Temelji neuroznanosti Vodič kroz vježbe
4.5 Pitanja: Potencijal u stanju mirovanja
1. Koja su dva glavna čimbenika koja određuju prolaz iona kroz staničnu
membranu?
a) ___________________________
b) ___________________________
2. Što podrazumijeva pojam "ravnotežni potencijal" iona?
3. Zašto je ravnotežni potencijal K+ iona -100 mV, a Na+ iona +41 mV, premda
su oba pozitivni ioni?
4. Ako je potencijal stanice u stanju mirovanja -65mV, što bi se dogodilo ukoliko
mijenjate koncentraciju iona K+ i Na+ u izvanstaničnom i unutarstaničnom
prostoru? Provjerite ispravnost tvrdnje mijenjajući koncentracije iona pomoću
simulacije koristeći program CCWIN i datoteku REST.CCS.
5. Otvorite PASSIVE.CCS. Što očekujete da će se dogoditi s veličinom
membranskog potencijala i odgovorom na depolarizacijsku struju ukoliko
udvostručite i pKleak i pNaleak? Pokušajte izvesti taj eksperiment i provjerite
jeste li u pravu. Ukratko objasnite rezultat.
31

Temelji neuroznanosti Vodič kroz vježbe
6. Koja je razlika između pojmova vodljivost i propusnost membrane?
7. Napišite Nernstovu jednadžbu i provjerite značenje svakog simbola. Što znači
kad kažemo da neki ion zadovoljava Nernstovu jednadžbu?
8. Odjeljci A i B odvojeni su membranom. U odjeljku A je 1 mol KNO3, dok
odjeljak B sadrži 0,1 mol KNO3-. K+ ioni nalaze se u ravnoteži. Odgovorite
kolika je razlika potencijala EA-EB, u standardnim uvjetima pri temperaturi
29,2oC? Koja je strana električki pozitivna?
9. U slijedećoj tablici nalaze se podaci koji se odnose na m. sartorius u žabe
("South American spotted tree frog").
IONI: unutarst. konc. izvanst. konc. vodljivost Na+ 12 mM 120 mM 0,05 K+ 120 mM 4 mM 0,5 Cl- 4 mM 120 mM 0,45
Izračunajte potencijal mirovanja za tu mišićnu stanicu. Je li unutarstanični
prostor električki negativan ili pozitivan u odnosu na izvanstanični prostor?
32
Temelji neuroznanosti Vodič kroz vježbe
6. Koja je razlika između pojmova vodljivost i propusnost membrane?
7. Napišite Nernstovu jednadžbu i provjerite značenje svakog simbola. Što znači
kad kažemo da neki ion zadovoljava Nernstovu jednadžbu?
8. Odjeljci A i B odvojeni su membranom. U odjeljku A je 1 mol KNO3, dok
odjeljak B sadrži 0,1 mol KNO3-. K+ ioni nalaze se u ravnoteži. Odgovorite
kolika je razlika potencijala EA-EB, u standardnim uvjetima pri temperaturi
29,2oC? Koja je strana električki pozitivna?
9. U slijedećoj tablici nalaze se podaci koji se odnose na m. sartorius u žabe
("South American spotted tree frog").
IONI: unutarst. konc. izvanst. konc. vodljivost Na+ 12 mM 120 mM 0,05 K+ 120 mM 4 mM 0,5 Cl- 4 mM 120 mM 0,45
Izračunajte potencijal mirovanja za tu mišićnu stanicu. Je li unutarstanični
prostor električki negativan ili pozitivan u odnosu na izvanstanični prostor?
32

Temelji neuroznanosti Vodič kroz vježbe
5. Akcijski potencijal
5.1 Uvod
Mehanizam nastanka akcijskog potencijala, kao i potencijala u stanju
mirovanja, neobično je važan za razumijevanje elektrofizioloških obilježja
podražljivih stanica. Neuroni i njihovi aksoni "rade" mnogo više od jednostavnog
postizanja potencijala mirovanja. Oni aktivno stvaraju, i svojom membranom provode
akcijske potencijale s jednog dijela stanice na drugi, te uz pomoć oslobađanja
neurotransmitora, omogućuju i komunikaciju s drugim stanicama.
Akcijski potencijal brza je promjena membranskoga potencijala, nakon koje
slijedi povratak na membranski potencijal mirovanja (slika 5.1). Veličina i oblik
akcijskoga potencijala znatno se razlikuju od jednoga podražljivog tkiva do drugoga.
Akcijski se potencijal, ne mijenjajući oblik i veličinu, širi cijelom dužinom aksona
živca ili mišićne stanice. Sposobnost živčane stanice da prenosi signale zasniva se na
pojavi akcijskoga potencijala. Akcijski potencijal omogućuje da se svi dijelovi neke
izdužene mišićne stanice kontrahiraju gotovo istodobno. U ovom ćemo poglavlju
raspravljati o ionskim strujama koje proizvode akcijski potencijal, o ionskim kanalima
koji omogućuju tok struje i o načinu kako se akcijski potencijal rasprostire i provodi.
Primjenjuju li se sve jači i jači depolarizacijski udari strujom, napokon se
dosegne točka u kojoj se pojavljuje drukčija vrsta reakcije, akcijski potencijal.
Akcijski potencijal izbija kad je depolarizacija tolika da membranski potencijal
dosegne vrijednost praga, a taj za m. sartorius žabe iznosi oko -60 mV.
Akcijski se potencijal razlikuje od lokalne depolarizacijske reakcije prema
ovim dvjema važnim pojavama:
akcijski je potencijal mnogo jača reakcija, tako da se polarnost
membranskog potencijala čak obrće (unutrašnjost stanice postaje pozitivna
u odnosu prema okolici),
akcijski se potencijal širi bez slabljenja cijelom duljinom mišićnoga vlakna.
Dok putuje duž mišićnoga vlakna, veličina i oblik akcijskoga potencijala
ostaju jednaki, tj. veličina se ne smanjuje razmjerno s udaljenošću, kao što
je to u lokalnoj reakciji. Primijeni li se podražaj koji je jači od praga,
veličina se i oblik akcijskoga potencijala ne mijenjaju, tj. veličina se
33
Temelji neuroznanosti Vodič kroz vježbe
5. Akcijski potencijal
5.1 Uvod
Mehanizam nastanka akcijskog potencijala, kao i potencijala u stanju
mirovanja, neobično je važan za razumijevanje elektrofizioloških obilježja
podražljivih stanica. Neuroni i njihovi aksoni "rade" mnogo više od jednostavnog
postizanja potencijala mirovanja. Oni aktivno stvaraju, i svojom membranom provode
akcijske potencijale s jednog dijela stanice na drugi, te uz pomoć oslobađanja
neurotransmitora, omogućuju i komunikaciju s drugim stanicama.
Akcijski potencijal brza je promjena membranskoga potencijala, nakon koje
slijedi povratak na membranski potencijal mirovanja (slika 5.1). Veličina i oblik
akcijskoga potencijala znatno se razlikuju od jednoga podražljivog tkiva do drugoga.
Akcijski se potencijal, ne mijenjajući oblik i veličinu, širi cijelom dužinom aksona
živca ili mišićne stanice. Sposobnost živčane stanice da prenosi signale zasniva se na
pojavi akcijskoga potencijala. Akcijski potencijal omogućuje da se svi dijelovi neke
izdužene mišićne stanice kontrahiraju gotovo istodobno. U ovom ćemo poglavlju
raspravljati o ionskim strujama koje proizvode akcijski potencijal, o ionskim kanalima
koji omogućuju tok struje i o načinu kako se akcijski potencijal rasprostire i provodi.
Primjenjuju li se sve jači i jači depolarizacijski udari strujom, napokon se
dosegne točka u kojoj se pojavljuje drukčija vrsta reakcije, akcijski potencijal.
Akcijski potencijal izbija kad je depolarizacija tolika da membranski potencijal
dosegne vrijednost praga, a taj za m. sartorius žabe iznosi oko -60 mV.
Akcijski se potencijal razlikuje od lokalne depolarizacijske reakcije prema
ovim dvjema važnim pojavama:
akcijski je potencijal mnogo jača reakcija, tako da se polarnost
membranskog potencijala čak obrće (unutrašnjost stanice postaje pozitivna
u odnosu prema okolici),
akcijski se potencijal širi bez slabljenja cijelom duljinom mišićnoga vlakna.
Dok putuje duž mišićnoga vlakna, veličina i oblik akcijskoga potencijala
ostaju jednaki, tj. veličina se ne smanjuje razmjerno s udaljenošću, kao što
je to u lokalnoj reakciji. Primijeni li se podražaj koji je jači od praga,
veličina se i oblik akcijskoga potencijala ne mijenjaju, tj. veličina se
33

Temelji neuroznanosti Vodič kroz vježbe
akcijskoga potencijala ne povećava pojačanjem podražaja. Nekim
podražajem akcijski se potencijal ili neće izazvati (subliminalni podražaj),
ili će se proizvesti akcijski potencijal potpune veličine i oblika. Prema
tome, akcijski je potencijal zapravo reakcija po načelu “sve ili ništa”.
Oblik akcijskog potencijala Na slici 5.3 prikazan je oblik akcijskoga potencijala u divovskom aksonu
lignje. Kad se membrana depolarizira do razine praga, nastane eksplozivna
depolarizacija kojom se membrana, ne samo potpuno depolarizira, već se pojavi i
prebačaj, tj. membrana se polarizira u suprotnome smjeru. Vršak akcijskoga
potencijala doseže vrijednost od oko +50 mV. Potom se membranski potencijal vraća
prema vrijednosti membranskoga potencijala mirovanja, i to gotovo jednakom
brzinom kao što se depolarizirao. Nakon repolarizacije uslijedi prolazna
hiperpolarizacija koja se naziva hiperpolarizacijski naknadni potencijal, a traje
otprilike 4 ms. U idućem odsječku razmotrit ćemo ionske struje koje uzrokuju
pojedine faze akcijskoga potencijala.
Slika 5.1 Reakcija membranskog potencijala na sve jače udare depolarizacijske struje. Kad se stanica depolarizira do razine praga, izbija akcijski potencijal.
Ionski mehanizmi akcijskog potencijala
Ranije ste pomoću jednadžbe za vodljivost kabela mogli vidjeti da je
membranski potencijal rezultat međusobno suprotnih djelovanja gradijenta za K+, koji
nastoji električni potencijal membrane (Em) približiti ravnotežnome potencijalu za K+,
te gradijenta za Na+, koji nastoji Em približiti ravnotežnome potencijalu za Na+.
Povećanje vodljivosti za bilo koji od tih iona povećat će silu kojom taj ion "vuče" Em
34
Temelji neuroznanosti Vodič kroz vježbe
akcijskoga potencijala ne povećava pojačanjem podražaja. Nekim
podražajem akcijski se potencijal ili neće izazvati (subliminalni podražaj),
ili će se proizvesti akcijski potencijal potpune veličine i oblika. Prema
tome, akcijski je potencijal zapravo reakcija po načelu “sve ili ništa”.
Oblik akcijskog potencijala Na slici 5.3 prikazan je oblik akcijskoga potencijala u divovskom aksonu
lignje. Kad se membrana depolarizira do razine praga, nastane eksplozivna
depolarizacija kojom se membrana, ne samo potpuno depolarizira, već se pojavi i
prebačaj, tj. membrana se polarizira u suprotnome smjeru. Vršak akcijskoga
potencijala doseže vrijednost od oko +50 mV. Potom se membranski potencijal vraća
prema vrijednosti membranskoga potencijala mirovanja, i to gotovo jednakom
brzinom kao što se depolarizirao. Nakon repolarizacije uslijedi prolazna
hiperpolarizacija koja se naziva hiperpolarizacijski naknadni potencijal, a traje
otprilike 4 ms. U idućem odsječku razmotrit ćemo ionske struje koje uzrokuju
pojedine faze akcijskoga potencijala.
Slika 5.1 Reakcija membranskog potencijala na sve jače udare depolarizacijske struje. Kad se stanica depolarizira do razine praga, izbija akcijski potencijal.
Ionski mehanizmi akcijskog potencijala
Ranije ste pomoću jednadžbe za vodljivost kabela mogli vidjeti da je
membranski potencijal rezultat međusobno suprotnih djelovanja gradijenta za K+, koji
nastoji električni potencijal membrane (Em) približiti ravnotežnome potencijalu za K+,
te gradijenta za Na+, koji nastoji Em približiti ravnotežnome potencijalu za Na+.
Povećanje vodljivosti za bilo koji od tih iona povećat će silu kojom taj ion "vuče" Em
34
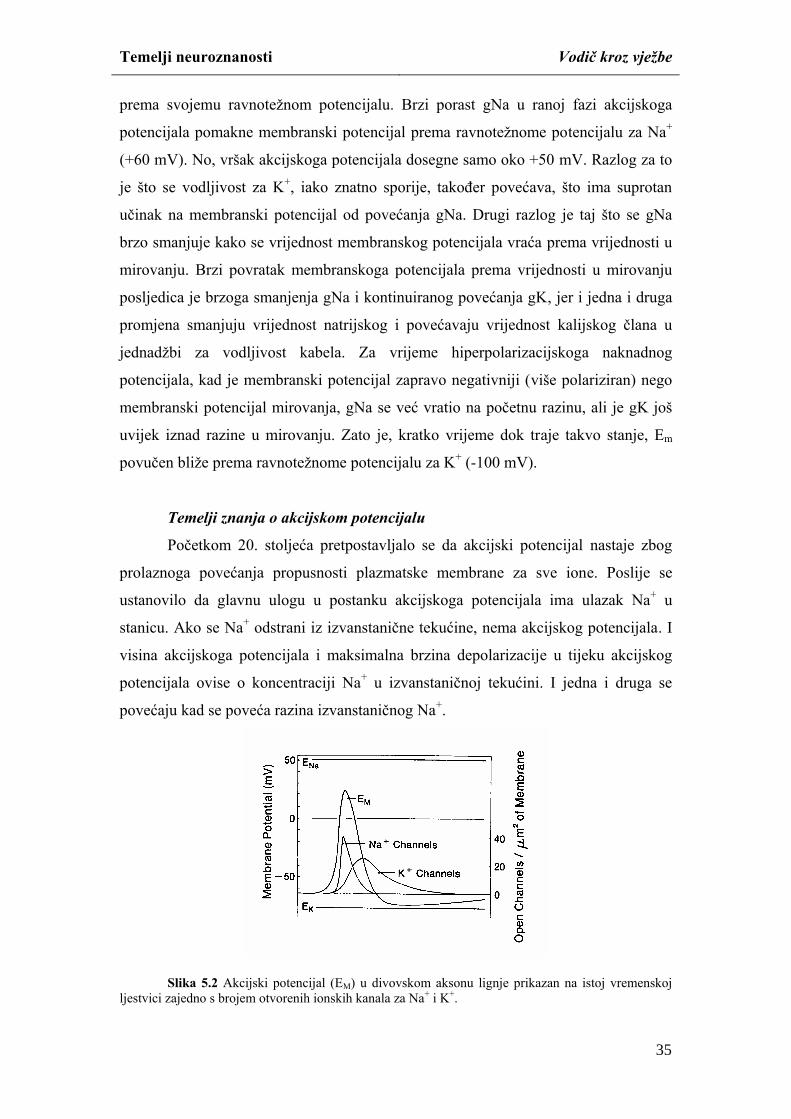
Temelji neuroznanosti Vodič kroz vježbe
prema svojemu ravnotežnom potencijalu. Brzi porast gNa u ranoj fazi akcijskoga
potencijala pomakne membranski potencijal prema ravnotežnome potencijalu za Na+
(+60 mV). No, vršak akcijskoga potencijala dosegne samo oko +50 mV. Razlog za to
je što se vodljivost za K+, iako znatno sporije, također povećava, što ima suprotan
učinak na membranski potencijal od povećanja gNa. Drugi razlog je taj što se gNa
brzo smanjuje kako se vrijednost membranskog potencijala vraća prema vrijednosti u
mirovanju. Brzi povratak membranskoga potencijala prema vrijednosti u mirovanju
posljedica je brzoga smanjenja gNa i kontinuiranog povećanja gK, jer i jedna i druga
promjena smanjuju vrijednost natrijskog i povećavaju vrijednost kalijskog člana u
jednadžbi za vodljivost kabela. Za vrijeme hiperpolarizacijskoga naknadnog
potencijala, kad je membranski potencijal zapravo negativniji (više polariziran) nego
membranski potencijal mirovanja, gNa se već vratio na početnu razinu, ali je gK još
uvijek iznad razine u mirovanju. Zato je, kratko vrijeme dok traje takvo stanje, Em
povučen bliže prema ravnotežnome potencijalu za K+ (-100 mV).
Temelji znanja o akcijskom potencijalu
Početkom 20. stoljeća pretpostavljalo se da akcijski potencijal nastaje zbog
prolaznoga povećanja propusnosti plazmatske membrane za sve ione. Poslije se
ustanovilo da glavnu ulogu u postanku akcijskoga potencijala ima ulazak Na+ u
stanicu. Ako se Na+ odstrani iz izvanstanične tekućine, nema akcijskog potencijala. I
visina akcijskoga potencijala i maksimalna brzina depolarizacije u tijeku akcijskog
potencijala ovise o koncentraciji Na+ u izvanstaničnoj tekućini. I jedna i druga se
povećaju kad se poveća razina izvanstaničnog Na+.
Slika 5.2 Akcijski potencijal (EM) u divovskom aksonu lignje prikazan na istoj vremenskoj ljestvici zajedno s brojem otvorenih ionskih kanala za Na+ i K+.
35
Temelji neuroznanosti Vodič kroz vježbe
prema svojemu ravnotežnom potencijalu. Brzi porast gNa u ranoj fazi akcijskoga
potencijala pomakne membranski potencijal prema ravnotežnome potencijalu za Na+
(+60 mV). No, vršak akcijskoga potencijala dosegne samo oko +50 mV. Razlog za to
je što se vodljivost za K+, iako znatno sporije, također povećava, što ima suprotan
učinak na membranski potencijal od povećanja gNa. Drugi razlog je taj što se gNa
brzo smanjuje kako se vrijednost membranskog potencijala vraća prema vrijednosti u
mirovanju. Brzi povratak membranskoga potencijala prema vrijednosti u mirovanju
posljedica je brzoga smanjenja gNa i kontinuiranog povećanja gK, jer i jedna i druga
promjena smanjuju vrijednost natrijskog i povećavaju vrijednost kalijskog člana u
jednadžbi za vodljivost kabela. Za vrijeme hiperpolarizacijskoga naknadnog
potencijala, kad je membranski potencijal zapravo negativniji (više polariziran) nego
membranski potencijal mirovanja, gNa se već vratio na početnu razinu, ali je gK još
uvijek iznad razine u mirovanju. Zato je, kratko vrijeme dok traje takvo stanje, Em
povučen bliže prema ravnotežnome potencijalu za K+ (-100 mV).
Temelji znanja o akcijskom potencijalu
Početkom 20. stoljeća pretpostavljalo se da akcijski potencijal nastaje zbog
prolaznoga povećanja propusnosti plazmatske membrane za sve ione. Poslije se
ustanovilo da glavnu ulogu u postanku akcijskoga potencijala ima ulazak Na+ u
stanicu. Ako se Na+ odstrani iz izvanstanične tekućine, nema akcijskog potencijala. I
visina akcijskoga potencijala i maksimalna brzina depolarizacije u tijeku akcijskog
potencijala ovise o koncentraciji Na+ u izvanstaničnoj tekućini. I jedna i druga se
povećaju kad se poveća razina izvanstaničnog Na+.
Slika 5.2 Akcijski potencijal (EM) u divovskom aksonu lignje prikazan na istoj vremenskoj ljestvici zajedno s brojem otvorenih ionskih kanala za Na+ i K+.
35

Temelji neuroznanosti Vodič kroz vježbe
Opetovanim podraživanjem divovskog aksona lignje u mediju koji sadrži
radioaktivni 22Na+ ustanovljeno je da u tijeku jednoga akcijskog potencijala u akson
uđe 3 do 4 x 10-12 mola Na+ kroz svaki četvorni centimetar površine membrane. Ta se
količina Na+ dobro slaže s vrijednošću izračunatom na osnovi električnih razmatranja
o tome kolika količina Na+ mora ući u akson. Kapacitet membrane (C) definira se kao
dQ/dV, tj. kao količina naboja koja mora proteći da bi se razlika u potencijalu
promijenila za 1 V. Zna se da je kapacitet 1 cm2 aksonske membrane lignje otprilike 1
mikrofarad (10-6 farada). Količina naboja koja mora proteći da bi se svaki cm2
membranskoga kondenzatora izbio za 100 mV (tolika je približna vrijednost
akcijskoga potencijala) računa se kao umnožak kapaciteta (C) i potencijala (V): 10-6
farada x 10-1 V = 10-7 kulona. Ako tu vrijednost podijelimo s količinom naboja koju
nosi 1 mol iona Na+ (96 500 kulona/molu), dobit ćemo da približno 10-12 mola iona
Na+ mora proći kroz svaki četvorni centimetar aksonske membrane da bi se
membrana depolarizirala za 100 mV.
Ako su u proces uključeni samo Na+ i K+, onda količina K+ koja napušta
stanicu u tijeku repolarizacije mora biti jednaka količini Na+ koja je tijekom
depolarizacije ušla u stanicu. Na+/K+-ATPaza izbacuje Na+ koji je ušao u stanicu i
ponovno nagomilava izgubljeni K+. Budući da je broj iona koji prolaze sa svakim
impulsom mali, Na+/K+-ATPaza, gledano na kraći rok, nije nužna. Akson lignje koji
je otrovan cijanidom ili uabainom tako da je aktivno prebacivanje iona obustavljeno,
još će uvijek odaslati približno 100 000 akcijskih potencijala prije no što u tome
zakaže. Manji aksoni, kod kojih je omjer volumena prema površini mnogo manji,
zakazat će nakon odašiljanja manjega broja impulsa, u slučaju da je Na+/K+ crpka
otrovana.
36
Temelji neuroznanosti Vodič kroz vježbe
Opetovanim podraživanjem divovskog aksona lignje u mediju koji sadrži
radioaktivni 22Na+ ustanovljeno je da u tijeku jednoga akcijskog potencijala u akson
uđe 3 do 4 x 10-12 mola Na+ kroz svaki četvorni centimetar površine membrane. Ta se
količina Na+ dobro slaže s vrijednošću izračunatom na osnovi električnih razmatranja
o tome kolika količina Na+ mora ući u akson. Kapacitet membrane (C) definira se kao
dQ/dV, tj. kao količina naboja koja mora proteći da bi se razlika u potencijalu
promijenila za 1 V. Zna se da je kapacitet 1 cm2 aksonske membrane lignje otprilike 1
mikrofarad (10-6 farada). Količina naboja koja mora proteći da bi se svaki cm2
membranskoga kondenzatora izbio za 100 mV (tolika je približna vrijednost
akcijskoga potencijala) računa se kao umnožak kapaciteta (C) i potencijala (V): 10-6
farada x 10-1 V = 10-7 kulona. Ako tu vrijednost podijelimo s količinom naboja koju
nosi 1 mol iona Na+ (96 500 kulona/molu), dobit ćemo da približno 10-12 mola iona
Na+ mora proći kroz svaki četvorni centimetar aksonske membrane da bi se
membrana depolarizirala za 100 mV.
Ako su u proces uključeni samo Na+ i K+, onda količina K+ koja napušta
stanicu u tijeku repolarizacije mora biti jednaka količini Na+ koja je tijekom
depolarizacije ušla u stanicu. Na+/K+-ATPaza izbacuje Na+ koji je ušao u stanicu i
ponovno nagomilava izgubljeni K+. Budući da je broj iona koji prolaze sa svakim
impulsom mali, Na+/K+-ATPaza, gledano na kraći rok, nije nužna. Akson lignje koji
je otrovan cijanidom ili uabainom tako da je aktivno prebacivanje iona obustavljeno,
još će uvijek odaslati približno 100 000 akcijskih potencijala prije no što u tome
zakaže. Manji aksoni, kod kojih je omjer volumena prema površini mnogo manji,
zakazat će nakon odašiljanja manjega broja impulsa, u slučaju da je Na+/K+ crpka
otrovana.
36
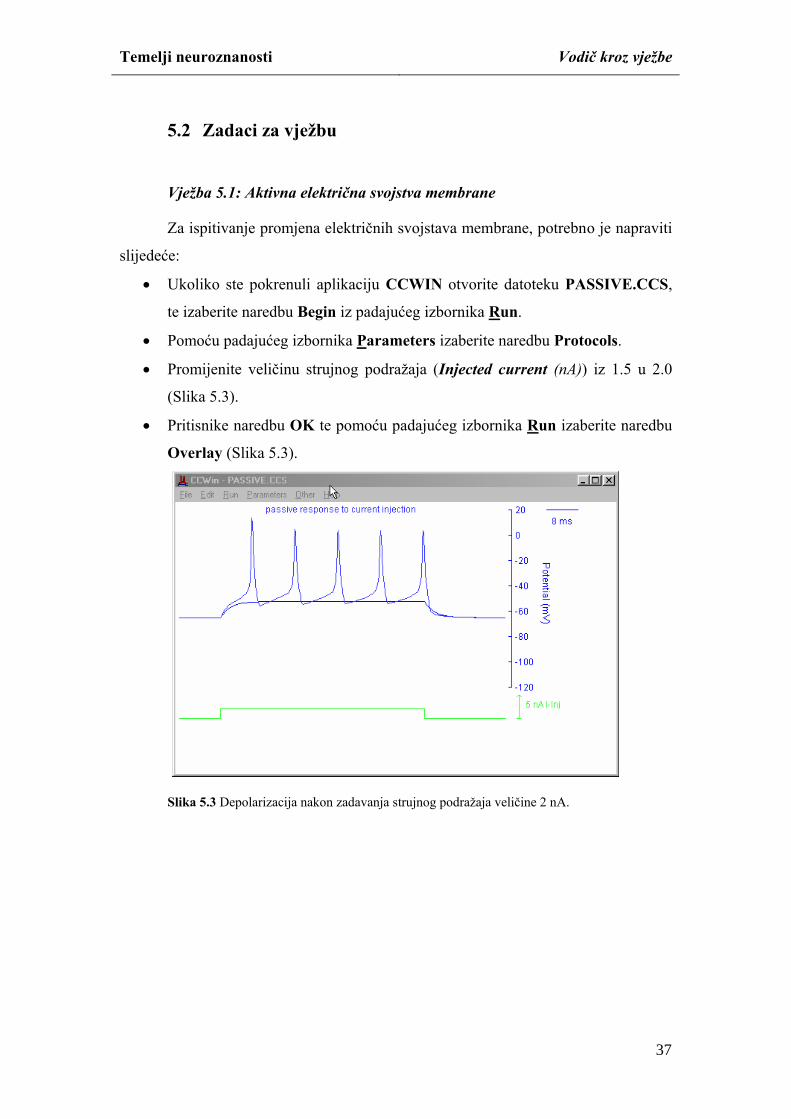
Temelji neuroznanosti Vodič kroz vježbe
5.2 Zadaci za vježbu
Vježba 5.1: Aktivna električna svojstva membrane
Za ispitivanje promjena električnih svojstava membrane, potrebno je napraviti
slijedeće:
• Ukoliko ste pokrenuli aplikaciju CCWIN otvorite datoteku PASSIVE.CCS,
te izaberite naredbu Begin iz padajućeg izbornika Run.
• Pomoću padajućeg izbornika Parameters izaberite naredbu Protocols.
• Promijenite veličinu strujnog podražaja (Injected current (nA)) iz 1.5 u 2.0
(Slika 5.3).
• Pritisnike naredbu OK te pomoću padajućeg izbornika Run izaberite naredbu
Overlay (Slika 5.3).
Slika 5.3 Depolarizacija nakon zadavanja strujnog podražaja veličine 2 nA.
37
Temelji neuroznanosti Vodič kroz vježbe
5.2 Zadaci za vježbu
Vježba 5.1: Aktivna električna svojstva membrane
Za ispitivanje promjena električnih svojstava membrane, potrebno je napraviti
slijedeće:
• Ukoliko ste pokrenuli aplikaciju CCWIN otvorite datoteku PASSIVE.CCS,
te izaberite naredbu Begin iz padajućeg izbornika Run.
• Pomoću padajućeg izbornika Parameters izaberite naredbu Protocols.
• Promijenite veličinu strujnog podražaja (Injected current (nA)) iz 1.5 u 2.0
(Slika 5.3).
• Pritisnike naredbu OK te pomoću padajućeg izbornika Run izaberite naredbu
Overlay (Slika 5.3).
Slika 5.3 Depolarizacija nakon zadavanja strujnog podražaja veličine 2 nA.
37
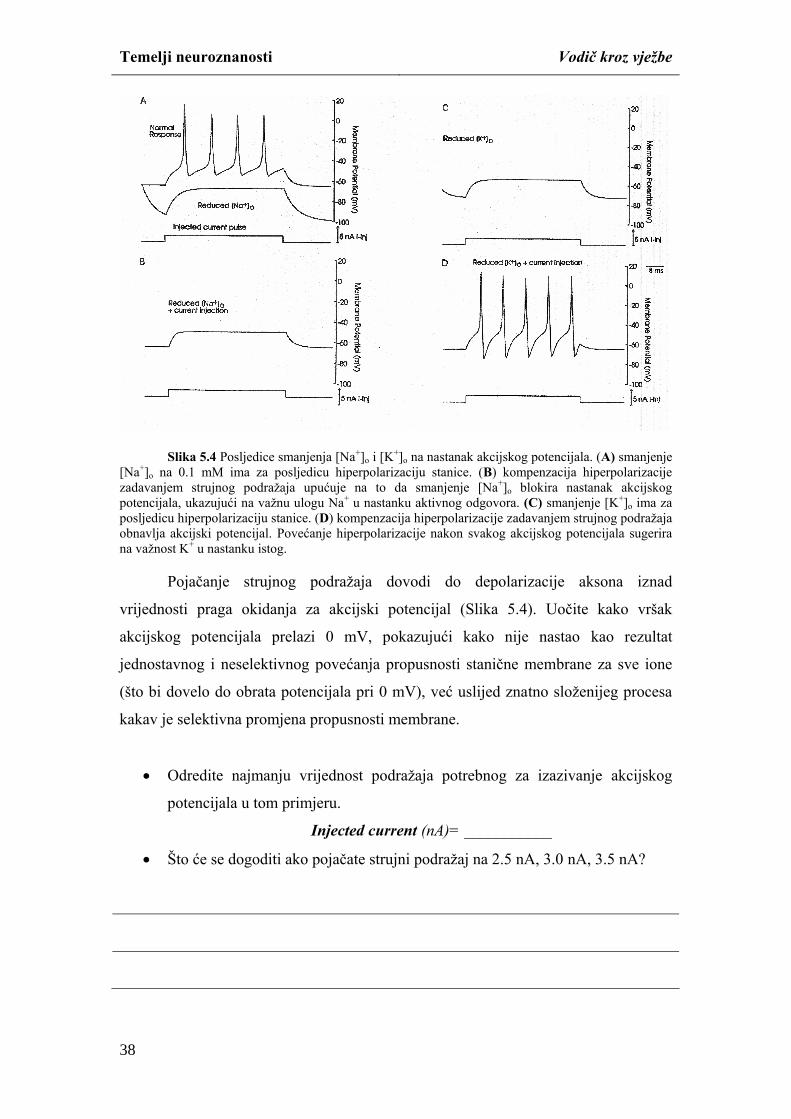
Temelji neuroznanosti Vodič kroz vježbe
Slika 5.4 Posljedice smanjenja [Na+]o i [K+]o na nastanak akcijskog potencijala. (A) smanjenje [Na+]o na 0.1 mM ima za posljedicu hiperpolarizaciju stanice. (B) kompenzacija hiperpolarizacije zadavanjem strujnog podražaja upućuje na to da smanjenje [Na+]o blokira nastanak akcijskog potencijala, ukazujući na važnu ulogu Na+ u nastanku aktivnog odgovora. (C) smanjenje [K+]o ima za posljedicu hiperpolarizaciju stanice. (D) kompenzacija hiperpolarizacije zadavanjem strujnog podražaja obnavlja akcijski potencijal. Povećanje hiperpolarizacije nakon svakog akcijskog potencijala sugerira na važnost K+ u nastanku istog.
Pojačanje strujnog podražaja dovodi do depolarizacije aksona iznad
vrijednosti praga okidanja za akcijski potencijal (Slika 5.4). Uočite kako vršak
akcijskog potencijala prelazi 0 mV, pokazujući kako nije nastao kao rezultat
jednostavnog i neselektivnog povećanja propusnosti stanične membrane za sve ione
(što bi dovelo do obrata potencijala pri 0 mV), već uslijed znatno složenijeg procesa
kakav je selektivna promjena propusnosti membrane.
• Odredite najmanju vrijednost podražaja potrebnog za izazivanje akcijskog
potencijala u tom primjeru.
Injected current (nA)= ___________
• Što će se dogoditi ako pojačate strujni podražaj na 2.5 nA, 3.0 nA, 3.5 nA?
38
Temelji neuroznanosti Vodič kroz vježbe
Slika 5.4 Posljedice smanjenja [Na+]o i [K+]o na nastanak akcijskog potencijala. (A) smanjenje [Na+]o na 0.1 mM ima za posljedicu hiperpolarizaciju stanice. (B) kompenzacija hiperpolarizacije zadavanjem strujnog podražaja upućuje na to da smanjenje [Na+]o blokira nastanak akcijskog potencijala, ukazujući na važnu ulogu Na+ u nastanku aktivnog odgovora. (C) smanjenje [K+]o ima za posljedicu hiperpolarizaciju stanice. (D) kompenzacija hiperpolarizacije zadavanjem strujnog podražaja obnavlja akcijski potencijal. Povećanje hiperpolarizacije nakon svakog akcijskog potencijala sugerira na važnost K+ u nastanku istog.
Pojačanje strujnog podražaja dovodi do depolarizacije aksona iznad
vrijednosti praga okidanja za akcijski potencijal (Slika 5.4). Uočite kako vršak
akcijskog potencijala prelazi 0 mV, pokazujući kako nije nastao kao rezultat
jednostavnog i neselektivnog povećanja propusnosti stanične membrane za sve ione
(što bi dovelo do obrata potencijala pri 0 mV), već uslijed znatno složenijeg procesa
kakav je selektivna promjena propusnosti membrane.
• Odredite najmanju vrijednost podražaja potrebnog za izazivanje akcijskog
potencijala u tom primjeru.
Injected current (nA)= ___________
• Što će se dogoditi ako pojačate strujni podražaj na 2.5 nA, 3.0 nA, 3.5 nA?
38

Temelji neuroznanosti Vodič kroz vježbe
• Što će se dogoditi ako zadate podražaj negativnog predznaka npr. -1.5 nA?
Kako to objašnjavate?
Vježba 5.2 Učinci različitih iona na nastanak akcijskog potencijala
Kako bi dokazali koji su ioni značajni u nastanku akcijskog potencijala,
možete mijenjati pojedinačnu koncentraciju različitih iona u umjetnoj otopini
("morskoj vodi" koja okružuje vanstaničnu površinu aksona) te promatrati rezultate.
Poznato je da među različitim ionima u morskoj vodi značajnu količinu čine ioni Na+,
K+, Ca++, Mg++ i Cl-.
• Pomoću padajućeg izbornika Parameters izaberite naredbu Ions i promijenite
veličinu izvanstanične koncentracije Mg++ ([Mg]o) u 0.1.
• Ponovno zadajte strujni podražaj u stanicu te pomoću padajućeg izbornika
Run izaberite naredbu Overlay i proučite dobiveni rezultat.
Zamijetite kako promjena koncentracije izvanstaničnoga magnezija ne utječe
na potencijal u stanju mirovanja i na nastanak akcijskog potencijala.
Ponovite isti eksperiment koristeći aplikaciju CCWIN tako da mijenjate
koncentracije drugih iona [Na+]o, [K+]o, [Ca++]o, [Cl-]o na niže vrijednosti (0.1 mM).
Izmjene uvijek radite zasebno za svaki pojedini ion. To znači da prije svake slijedeće
promjene vratite sve vrijednosti na početne. Zapamtite da uvijek možete vratiti
početne vrijednosti pomoću datoteke ACTIVE.CCS nakon svake pojedine
simulacije.
• Upišite rezultate vaših zapažanja.
39
Temelji neuroznanosti Vodič kroz vježbe
• Što će se dogoditi ako zadate podražaj negativnog predznaka npr. -1.5 nA?
Kako to objašnjavate?
Vježba 5.2 Učinci različitih iona na nastanak akcijskog potencijala
Kako bi dokazali koji su ioni značajni u nastanku akcijskog potencijala,
možete mijenjati pojedinačnu koncentraciju različitih iona u umjetnoj otopini
("morskoj vodi" koja okružuje vanstaničnu površinu aksona) te promatrati rezultate.
Poznato je da među različitim ionima u morskoj vodi značajnu količinu čine ioni Na+,
K+, Ca++, Mg++ i Cl-.
• Pomoću padajućeg izbornika Parameters izaberite naredbu Ions i promijenite
veličinu izvanstanične koncentracije Mg++ ([Mg]o) u 0.1.
• Ponovno zadajte strujni podražaj u stanicu te pomoću padajućeg izbornika
Run izaberite naredbu Overlay i proučite dobiveni rezultat.
Zamijetite kako promjena koncentracije izvanstaničnoga magnezija ne utječe
na potencijal u stanju mirovanja i na nastanak akcijskog potencijala.
Ponovite isti eksperiment koristeći aplikaciju CCWIN tako da mijenjate
koncentracije drugih iona [Na+]o, [K+]o, [Ca++]o, [Cl-]o na niže vrijednosti (0.1 mM).
Izmjene uvijek radite zasebno za svaki pojedini ion. To znači da prije svake slijedeće
promjene vratite sve vrijednosti na početne. Zapamtite da uvijek možete vratiti
početne vrijednosti pomoću datoteke ACTIVE.CCS nakon svake pojedine
simulacije.
• Upišite rezultate vaših zapažanja.
39

Temelji neuroznanosti Vodič kroz vježbe
Uočite kako pojedinačno najveći utjecaj imaju ioni Na+ i K+. Nakon što ste
smanjili veličinu koncentracije Na+ u izvanstaničnom prostoru na 0.1, membranski
potencijal stanice postaje značajno negativniji. To ukazuje na činjenicu da pasivni
utok Na+ u stanicu utječe na membranski potencijal u stanju mirovanja tako da dovodi
do depolarizacije od -65 mV. Onog trenutka kad onemogućite ulazak, ili obrnete
smjer ulaska iona Na+, membranski potencijal postaje značajno hiperpolariziran.
"Nadoknadu" nastale hiperpolarizacije možete postići tako da zadate strujni podražaj
pomoću mikroelektrode i vratite membranski potencijal u stanici na -65 mV.
Simulaciju možete izvesti na slijedeći način:
• Pomoću padajućeg izbornika Parameters i naredbe Protocol promijenite
veličinu Base current (nA) u 2.25. Pritisnite OK te pomoću padajućeg
izbornika Run i naredbe Begin promatrajte rezultat.
Uočite kako je onemogućen nastanak akcijskog potencijala što ukazuje na
značenje izvanstaničnoga Na+ u nastanku istog. Čak i kad povećate jačinu strujnog
podražaja na 4 nA, u stanici neće nastati akcijski potencijal.
• Pronađite kolika mora biti izvanstanična koncentracija natrijevih iona pri kojoj
dolazi do nastanka akcijskog potencijala, ako pritom zadate strujni podražaj od
2.25 nA.
[Na]o=___________
Sad ćemo ispitati utjecaj K+ iona:
• Otvorite datoteku ACTIVE.CCS i pomoću naredbe Begin iz padajućeg
izbornika Run započnite simulaciju.
• Preko izbornika Parameters pronađite naredbu Ions i u tablici promijenite
koncentraciju izvanstaničnog K+ na 0.1 mM.
• Pritisnite OK i pomoću naredbe Overlay iz izbornika Run promatrajte
rezultat (Slika 5.5).
40
Temelji neuroznanosti Vodič kroz vježbe
Uočite kako pojedinačno najveći utjecaj imaju ioni Na+ i K+. Nakon što ste
smanjili veličinu koncentracije Na+ u izvanstaničnom prostoru na 0.1, membranski
potencijal stanice postaje značajno negativniji. To ukazuje na činjenicu da pasivni
utok Na+ u stanicu utječe na membranski potencijal u stanju mirovanja tako da dovodi
do depolarizacije od -65 mV. Onog trenutka kad onemogućite ulazak, ili obrnete
smjer ulaska iona Na+, membranski potencijal postaje značajno hiperpolariziran.
"Nadoknadu" nastale hiperpolarizacije možete postići tako da zadate strujni podražaj
pomoću mikroelektrode i vratite membranski potencijal u stanici na -65 mV.
Simulaciju možete izvesti na slijedeći način:
• Pomoću padajućeg izbornika Parameters i naredbe Protocol promijenite
veličinu Base current (nA) u 2.25. Pritisnite OK te pomoću padajućeg
izbornika Run i naredbe Begin promatrajte rezultat.
Uočite kako je onemogućen nastanak akcijskog potencijala što ukazuje na
značenje izvanstaničnoga Na+ u nastanku istog. Čak i kad povećate jačinu strujnog
podražaja na 4 nA, u stanici neće nastati akcijski potencijal.
• Pronađite kolika mora biti izvanstanična koncentracija natrijevih iona pri kojoj
dolazi do nastanka akcijskog potencijala, ako pritom zadate strujni podražaj od
2.25 nA.
[Na]o=___________
Sad ćemo ispitati utjecaj K+ iona:
• Otvorite datoteku ACTIVE.CCS i pomoću naredbe Begin iz padajućeg
izbornika Run započnite simulaciju.
• Preko izbornika Parameters pronađite naredbu Ions i u tablici promijenite
koncentraciju izvanstaničnog K+ na 0.1 mM.
• Pritisnite OK i pomoću naredbe Overlay iz izbornika Run promatrajte
rezultat (Slika 5.5).
40
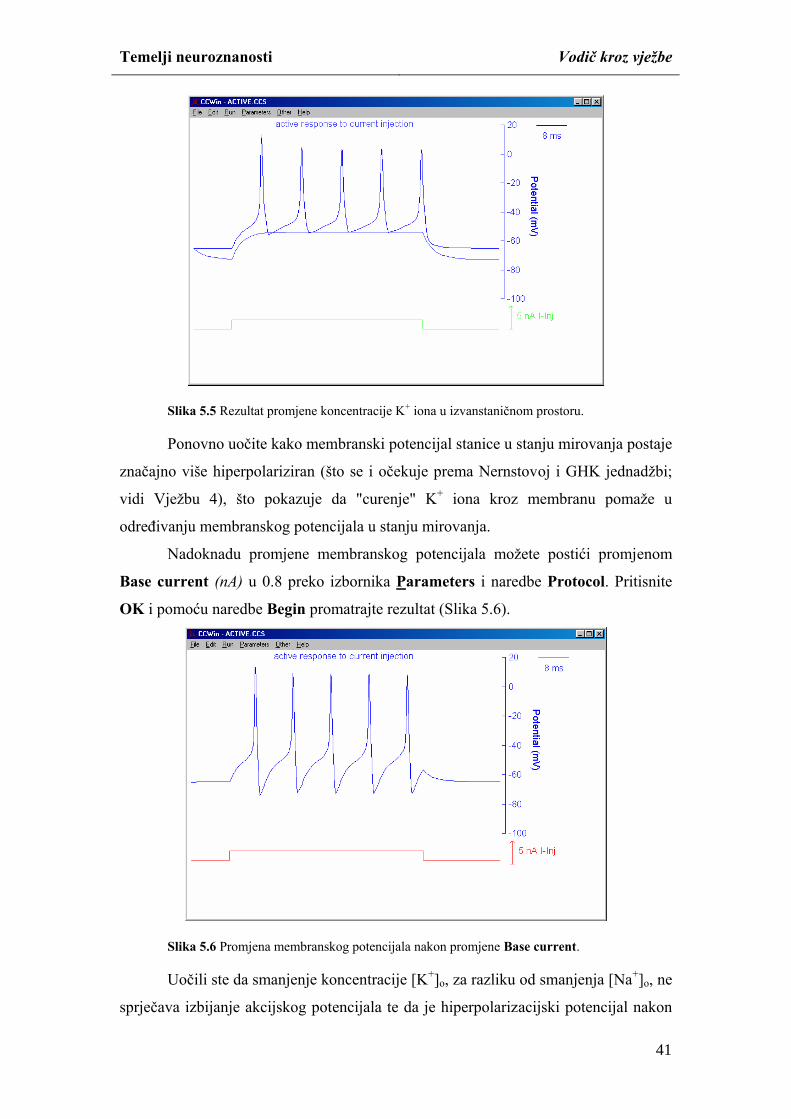
Temelji neuroznanosti Vodič kroz vježbe
Slika 5.5 Rezultat promjene koncentracije K+ iona u izvanstaničnom prostoru.
Ponovno uočite kako membranski potencijal stanice u stanju mirovanja postaje
značajno više hiperpolariziran (što se i očekuje prema Nernstovoj i GHK jednadžbi;
vidi Vježbu 4), što pokazuje da "curenje" K+ iona kroz membranu pomaže u
određivanju membranskog potencijala u stanju mirovanja.
Nadoknadu promjene membranskog potencijala možete postići promjenom
Base current (nA) u 0.8 preko izbornika Parameters i naredbe Protocol. Pritisnite
OK i pomoću naredbe Begin promatrajte rezultat (Slika 5.6).
Slika 5.6 Promjena membranskog potencijala nakon promjene Base current.
Uočili ste da smanjenje koncentracije [K+]o, za razliku od smanjenja [Na+]o, ne
sprječava izbijanje akcijskog potencijala te da je hiperpolarizacijski potencijal nakon
41
Temelji neuroznanosti Vodič kroz vježbe
Slika 5.5 Rezultat promjene koncentracije K+ iona u izvanstaničnom prostoru.
Ponovno uočite kako membranski potencijal stanice u stanju mirovanja postaje
značajno više hiperpolariziran (što se i očekuje prema Nernstovoj i GHK jednadžbi;
vidi Vježbu 4), što pokazuje da "curenje" K+ iona kroz membranu pomaže u
određivanju membranskog potencijala u stanju mirovanja.
Nadoknadu promjene membranskog potencijala možete postići promjenom
Base current (nA) u 0.8 preko izbornika Parameters i naredbe Protocol. Pritisnite
OK i pomoću naredbe Begin promatrajte rezultat (Slika 5.6).
Slika 5.6 Promjena membranskog potencijala nakon promjene Base current.
Uočili ste da smanjenje koncentracije [K+]o, za razliku od smanjenja [Na+]o, ne
sprječava izbijanje akcijskog potencijala te da je hiperpolarizacijski potencijal nakon
41

Temelji neuroznanosti Vodič kroz vježbe
svakog akcijskog potencijala veći nego ranije (usporedite sliku 5.4A i 5.4D). Te
promjene možete izravno promatrati pomoću datoteke ACTIVE.CCS i naredbe
Overlay iz padajućeg izbornika Run. Porast hiperpolarizacije nakon svakog
akcijskog potencijala uz smanjenje [K+]o ukazuje na značaj pomaka K+ kroz
membranu.
Dakle, uočili smo da smanjenje [K+]o dovodi do porasta hiperpolarizacije
nakon akcijskog potencijala, kao i da ta hiperpolarizacija ide ispod početne razine
membranskog potencijala ("undershoot"). Iz toga slijedi da je koncentracija K+ veća
unutar stanice nego u izvanstaničnom prostoru te da se kalij kreće niz koncentracijski
gradijent tijekom faze repolarizacije akcijskog potencijala!
• Kako bi ispitali tu činjenicu u eksperimentu promijenite i intracelularnu
koncentraciju K+ iona ([K+]i). Obrazložite dobiveni rezultat.
Vježba 5.3 "Toothpaste" eksperiment
Baker, Hodgkin i Shaw (1962) u izvornom su eksperimentu iskoristili veličinu
divovskog aksona lignje i iz njega istisnuli aksoplazmu (kao da je akson nalik na
zubnu pastu, odakle i potječe naziv ovog eksperimenta). Zamijenili su aksoplazmu
umjetnim nadomjestkom različitih ionskih koncentracija.
Za simulaciju tog pokusa u aplikaciji CCWIN potrebno je napraviti slijedeće
korake:
• Otvorite datoteku K_INTRA.CCS i izaberite naredbu Begin iz padajućeg
izbornika Run.
• Preko naredbe Ions Smanjite koncentraciju [K+]i na 0.6 mM.
• Pritisnike OK te odaberite naredbu Begin i uočite kako se depolarizira
membranski potencijal (Slika 5.7A).
• Za poništenje te promjene odaberite naredbu Protocol pomoću izbornika
Parameters, pa promijenite Base current (nA) u -3 nA i odaberite naredbu
Begin. U ovom trenutku akson stvara akcijski potencijal, ali ne dolazi do
42
Temelji neuroznanosti Vodič kroz vježbe
svakog akcijskog potencijala veći nego ranije (usporedite sliku 5.4A i 5.4D). Te
promjene možete izravno promatrati pomoću datoteke ACTIVE.CCS i naredbe
Overlay iz padajućeg izbornika Run. Porast hiperpolarizacije nakon svakog
akcijskog potencijala uz smanjenje [K+]o ukazuje na značaj pomaka K+ kroz
membranu.
Dakle, uočili smo da smanjenje [K+]o dovodi do porasta hiperpolarizacije
nakon akcijskog potencijala, kao i da ta hiperpolarizacija ide ispod početne razine
membranskog potencijala ("undershoot"). Iz toga slijedi da je koncentracija K+ veća
unutar stanice nego u izvanstaničnom prostoru te da se kalij kreće niz koncentracijski
gradijent tijekom faze repolarizacije akcijskog potencijala!
• Kako bi ispitali tu činjenicu u eksperimentu promijenite i intracelularnu
koncentraciju K+ iona ([K+]i). Obrazložite dobiveni rezultat.
Vježba 5.3 "Toothpaste" eksperiment
Baker, Hodgkin i Shaw (1962) u izvornom su eksperimentu iskoristili veličinu
divovskog aksona lignje i iz njega istisnuli aksoplazmu (kao da je akson nalik na
zubnu pastu, odakle i potječe naziv ovog eksperimenta). Zamijenili su aksoplazmu
umjetnim nadomjestkom različitih ionskih koncentracija.
Za simulaciju tog pokusa u aplikaciji CCWIN potrebno je napraviti slijedeće
korake:
• Otvorite datoteku K_INTRA.CCS i izaberite naredbu Begin iz padajućeg
izbornika Run.
• Preko naredbe Ions Smanjite koncentraciju [K+]i na 0.6 mM.
• Pritisnike OK te odaberite naredbu Begin i uočite kako se depolarizira
membranski potencijal (Slika 5.7A).
• Za poništenje te promjene odaberite naredbu Protocol pomoću izbornika
Parameters, pa promijenite Base current (nA) u -3 nA i odaberite naredbu
Begin. U ovom trenutku akson stvara akcijski potencijal, ali ne dolazi do
42
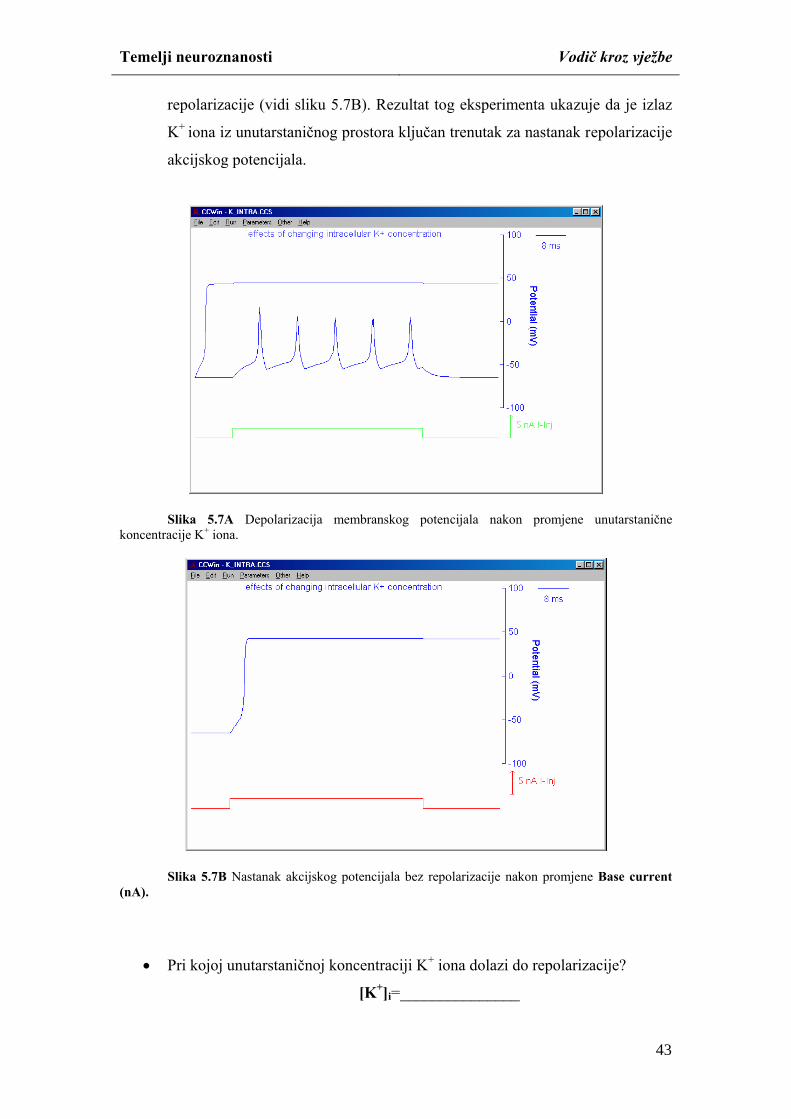
Temelji neuroznanosti Vodič kroz vježbe
repolarizacije (vidi sliku 5.7B). Rezultat tog eksperimenta ukazuje da je izlaz
K+ iona iz unutarstaničnog prostora ključan trenutak za nastanak repolarizacije
akcijskog potencijala.
Slika 5.7A Depolarizacija membranskog potencijala nakon promjene unutarstanične koncentracije K+ iona.
Slika 5.7B Nastanak akcijskog potencijala bez repolarizacije nakon promjene Base current (nA).
• Pri kojoj unutarstaničnoj koncentraciji K+ iona dolazi do repolarizacije?
[K+]i=_______________
43
Temelji neuroznanosti Vodič kroz vježbe
repolarizacije (vidi sliku 5.7B). Rezultat tog eksperimenta ukazuje da je izlaz
K+ iona iz unutarstaničnog prostora ključan trenutak za nastanak repolarizacije
akcijskog potencijala.
Slika 5.7A Depolarizacija membranskog potencijala nakon promjene unutarstanične koncentracije K+ iona.
Slika 5.7B Nastanak akcijskog potencijala bez repolarizacije nakon promjene Base current (nA).
• Pri kojoj unutarstaničnoj koncentraciji K+ iona dolazi do repolarizacije?
[K+]i=_______________
43

Temelji neuroznanosti Vodič kroz vježbe
• Što će se dogoditi ako povećate unutarstaničnu koncentraciju K+ iona i zašto?
5.3 Pitanja: Akcijski potencijal
• Što je akcijski potencijal i zašto nastaje?
• Nacrtajte krivulju akcijskog potencijala divovskog aksona lignje (membranski
potencijal u mV, vrijeme u ms). Na krivulji označite: potencijal mirovanja,
prag, vršak, prebačaj, razdoblje depolarizacije, razdoblje hiperpolarizacije.
Nacrtajte krivulje ionskih struja i označite koja krivulja odgovara struji Na+, a
koja struji K+.
a) akcijski potencijal
-80
-60
-40
-20
0
20
40
60
80
t (ms)
U (m
V)
44
Temelji neuroznanosti Vodič kroz vježbe
• Što će se dogoditi ako povećate unutarstaničnu koncentraciju K+ iona i zašto?
5.3 Pitanja: Akcijski potencijal
• Što je akcijski potencijal i zašto nastaje?
• Nacrtajte krivulju akcijskog potencijala divovskog aksona lignje (membranski
potencijal u mV, vrijeme u ms). Na krivulji označite: potencijal mirovanja,
prag, vršak, prebačaj, razdoblje depolarizacije, razdoblje hiperpolarizacije.
Nacrtajte krivulje ionskih struja i označite koja krivulja odgovara struji Na+, a
koja struji K+.
a) akcijski potencijal
-80
-60
-40
-20
0
20
40
60
80
t (ms)
U (m
V)
44

Temelji neuroznanosti Vodič kroz vježbe
b) ionske struje
• U kojem smjeru prolaze ioni za vrijeme uzlaznog dijela krivulje akcijskog
potencijala, a u kojem za vrijeme silaznog dijela krivulje akcijskog
potencijala?
• Zbog čega vršak akcijskog potencijala ne doseže ravnotežni potencijal za
natrij? Odgovor možete pronaći ako pobliže ispitate ionske struje pomoću
pokretanja datoteke actn_pot.ccs.
• Što uzrokuje apsolutnu refrakternost (nepodražljivost)? Što uzrokuje relativnu
refrakternost? Zašto akcijski potencijal ima prag?
45
Temelji neuroznanosti Vodič kroz vježbe
b) ionske struje
• U kojem smjeru prolaze ioni za vrijeme uzlaznog dijela krivulje akcijskog
potencijala, a u kojem za vrijeme silaznog dijela krivulje akcijskog
potencijala?
• Zbog čega vršak akcijskog potencijala ne doseže ravnotežni potencijal za
natrij? Odgovor možete pronaći ako pobliže ispitate ionske struje pomoću
pokretanja datoteke actn_pot.ccs.
• Što uzrokuje apsolutnu refrakternost (nepodražljivost)? Što uzrokuje relativnu
refrakternost? Zašto akcijski potencijal ima prag?
45

Temelji neuroznanosti Vodič kroz vježbe
• Koja je uloga Na+/K+ ATP-aze u nastajanju akcijskog potencijala?
46
Temelji neuroznanosti Vodič kroz vježbe
• Koja je uloga Na+/K+ ATP-aze u nastajanju akcijskog potencijala?
46

Temelji neuroznanosti Vodič kroz vježbe
6. Sinaptički potencijali
6.1 Uvod
Nekad je postojalo mišljenje kako su sve sinaptičke veze u živčanom sustavu
ekscitacijskoga tipa. Charles Sherrington (1906.) je u svojoj studiji o spinalnim
refleksima pokazao postojanje inhibicijskih i ekscitacijskih veza među neuronima.
Četrdesetih i pedestih godina Sir John Eccles vršio je intracelularno snimanje
motornih neurona leđne moždine i došao do otkrića tzv. inhibicijskih
postsinaptičkih potencijala (IPSPs), koji su jednako važni kao i ekscitacijski
postsinaptički potencijali (EPSPs) u prenošenju informacija u neuronskim
krugovima.
Danas znamo da u zamjetno brzom prijenosu informacija pomoću
neurotransmitera (milisekunde) postoje dva osnovna mehanizma: ekscitacija i
inhibicija. Osnovna razlika je u tome što ekscitacijska neurotransmisija povećava
mogućnost nastanka akcijskog potencijala, dok inhibicijska neurotransmisija
smanjuje mogućnost nastanka akcijskog potencijala. Vjeruje se kako je najčešći
ekscitacijski neurotransmiter u mozgu glutamat ili neke druge slične aminokiseline
koje mogu aktivirati oba postsinaptička receptora (AMPA/kainatni i NMDA
receptor). Najvažniji inhibicijski neurotransmiter je GABA koji aktivira GABAA i
GABAB receptore.
Vježba 6.1: Ekscitacijski postsinaptički potencijali
Zamislite na trenutak kako snimate hipokampalne ili kortikalne piramidne
stanice (ili neurone iz bilo kojeg dijela mozga čovjeka ili životinje) i aktivirate
aferentni ekscitacijski put pomoću kratkotrajnog električnog podražaja. Zapis koji
dobijete snimanjem predstavlja brzi EPSP praćen bifazičnom hiperpolarizacijom zbog
IPSP-a (slika 6.1; desni trag zapisa). Pretpostavite da IPSP nastaje zbog aktivacije
lokalnih GABA-ergičkih neurona. I odista, kad snimate te stanice vidjet ćete kako se
one kratkotrajno aktiviraju, kao odgovor na kratkotrajnu aferentnu stimulaciju (slika
6.1.; lijevi trag zapisa). Ekscitacija nastaje zbog otpuštanja glutamata nakon aferentne
stimulacije, dok posljedični inhibicijski potencijal nastaje zbog otpuštanja GABA-e iz
lokalnih GABA-ergičkih interneurona. I te inhibicijske stanice također se podraže
zbog djelovanja glutamata nakon aferentne stimulacije (slika 6.2.)
47
Temelji neuroznanosti Vodič kroz vježbe
6. Sinaptički potencijali
6.1 Uvod
Nekad je postojalo mišljenje kako su sve sinaptičke veze u živčanom sustavu
ekscitacijskoga tipa. Charles Sherrington (1906.) je u svojoj studiji o spinalnim
refleksima pokazao postojanje inhibicijskih i ekscitacijskih veza među neuronima.
Četrdesetih i pedestih godina Sir John Eccles vršio je intracelularno snimanje
motornih neurona leđne moždine i došao do otkrića tzv. inhibicijskih
postsinaptičkih potencijala (IPSPs), koji su jednako važni kao i ekscitacijski
postsinaptički potencijali (EPSPs) u prenošenju informacija u neuronskim
krugovima.
Danas znamo da u zamjetno brzom prijenosu informacija pomoću
neurotransmitera (milisekunde) postoje dva osnovna mehanizma: ekscitacija i
inhibicija. Osnovna razlika je u tome što ekscitacijska neurotransmisija povećava
mogućnost nastanka akcijskog potencijala, dok inhibicijska neurotransmisija
smanjuje mogućnost nastanka akcijskog potencijala. Vjeruje se kako je najčešći
ekscitacijski neurotransmiter u mozgu glutamat ili neke druge slične aminokiseline
koje mogu aktivirati oba postsinaptička receptora (AMPA/kainatni i NMDA
receptor). Najvažniji inhibicijski neurotransmiter je GABA koji aktivira GABAA i
GABAB receptore.
Vježba 6.1: Ekscitacijski postsinaptički potencijali
Zamislite na trenutak kako snimate hipokampalne ili kortikalne piramidne
stanice (ili neurone iz bilo kojeg dijela mozga čovjeka ili životinje) i aktivirate
aferentni ekscitacijski put pomoću kratkotrajnog električnog podražaja. Zapis koji
dobijete snimanjem predstavlja brzi EPSP praćen bifazičnom hiperpolarizacijom zbog
IPSP-a (slika 6.1; desni trag zapisa). Pretpostavite da IPSP nastaje zbog aktivacije
lokalnih GABA-ergičkih neurona. I odista, kad snimate te stanice vidjet ćete kako se
one kratkotrajno aktiviraju, kao odgovor na kratkotrajnu aferentnu stimulaciju (slika
6.1.; lijevi trag zapisa). Ekscitacija nastaje zbog otpuštanja glutamata nakon aferentne
stimulacije, dok posljedični inhibicijski potencijal nastaje zbog otpuštanja GABA-e iz
lokalnih GABA-ergičkih interneurona. I te inhibicijske stanice također se podraže
zbog djelovanja glutamata nakon aferentne stimulacije (slika 6.2.)
47

Temelji neuroznanosti Vodič kroz vježbe
• Pokrenite aplikaciju CCWIN te otvorite datoteku EP_IPSP.CCS. Sad možete
izolirati EPSP od bifazičkog IPSP-a primjenom antagonista GABA receptora.
• Pomoću padajućeg izbornika Parameters i naredbe Synaptic Currents
pronađite gIPSP i promijenite vrijednost u 0. Pritisnite OK i pomoću naredbe
Overlay iz podajućeg izbornika Run usporedite EPSP prije i nakon blokiranja
inhibicije.
Možete vidjeti da EPSP postaje značajno veći nakon blokiranja inhibicije
(slika 6.2.). Kod stvarnog neuronskog kruga, jaki porast amplitude EPSP-a nakon
blokiranja inhibicije može imati za posljedicu "bježanje" ekscitacije među spojenim
ekscitacijskim neuronima (obrazac karakterističan za epileptiformne poremećaje).
Farmakolozi su iskoristili tu spoznaju u liječenju epilepsije uspostavljanjem ravnoteže
između ekscitacijskih i inhibicijskih mehanizama.
Ispitivanjem izoliranog EPSP-a otkrivate postojanje dva tipa receptora koje
selektivno aktiviraju agonisti AMPA i NMDA. Ispitivanjem naponske ovisnosti i
ionske osjetljivosti odgovora na podražaj posredovanog AMPA i NMDA receptorima,
provjerite kako oba receptor/kanal sustava propuštaju Na+ i K+ ione i stoga odgovor
nastaje na oko 0 mV.
• Pomoću padajućeg izbornika Parameters i naredbe Protocol promijenite
Base current(nA) u 1.78 i vrijednost Starting Vm(mV) u 20. Pritisnite OK i
pomoću naredbe Overlay iz padajućeg izbornika Run promatrajte rezultat
simulacije.
EPSP se okreće prema hiperpolarizaciji obzirom da je membranski potencijal
pozitivniji prema potencijalu obrata (slika 6.2.; obrnuti EPSP). NMDA komponenta
EPSP-a ima neobično veliku naponsku ovisnost, postaje veća uz depolarizaciju iako
ste bliži potencijalu obrata! Kako biste ispitali tu činjenicu izolirajte NMDA
komponentu blokiranjem AMPA komponente pomoću selektivnog antagonista.
• Pomoću aplikacije CCWIN otvorite datoteku NMDA.CCS (slika 6.3.). Sad će
aferentna stimulacija imati za posljedicu nastanak EPSP-a posredovanog
NMDA receptorom.
48
Temelji neuroznanosti Vodič kroz vježbe
• Pokrenite aplikaciju CCWIN te otvorite datoteku EP_IPSP.CCS. Sad možete
izolirati EPSP od bifazičkog IPSP-a primjenom antagonista GABA receptora.
• Pomoću padajućeg izbornika Parameters i naredbe Synaptic Currents
pronađite gIPSP i promijenite vrijednost u 0. Pritisnite OK i pomoću naredbe
Overlay iz podajućeg izbornika Run usporedite EPSP prije i nakon blokiranja
inhibicije.
Možete vidjeti da EPSP postaje značajno veći nakon blokiranja inhibicije
(slika 6.2.). Kod stvarnog neuronskog kruga, jaki porast amplitude EPSP-a nakon
blokiranja inhibicije može imati za posljedicu "bježanje" ekscitacije među spojenim
ekscitacijskim neuronima (obrazac karakterističan za epileptiformne poremećaje).
Farmakolozi su iskoristili tu spoznaju u liječenju epilepsije uspostavljanjem ravnoteže
između ekscitacijskih i inhibicijskih mehanizama.
Ispitivanjem izoliranog EPSP-a otkrivate postojanje dva tipa receptora koje
selektivno aktiviraju agonisti AMPA i NMDA. Ispitivanjem naponske ovisnosti i
ionske osjetljivosti odgovora na podražaj posredovanog AMPA i NMDA receptorima,
provjerite kako oba receptor/kanal sustava propuštaju Na+ i K+ ione i stoga odgovor
nastaje na oko 0 mV.
• Pomoću padajućeg izbornika Parameters i naredbe Protocol promijenite
Base current(nA) u 1.78 i vrijednost Starting Vm(mV) u 20. Pritisnite OK i
pomoću naredbe Overlay iz padajućeg izbornika Run promatrajte rezultat
simulacije.
EPSP se okreće prema hiperpolarizaciji obzirom da je membranski potencijal
pozitivniji prema potencijalu obrata (slika 6.2.; obrnuti EPSP). NMDA komponenta
EPSP-a ima neobično veliku naponsku ovisnost, postaje veća uz depolarizaciju iako
ste bliži potencijalu obrata! Kako biste ispitali tu činjenicu izolirajte NMDA
komponentu blokiranjem AMPA komponente pomoću selektivnog antagonista.
• Pomoću aplikacije CCWIN otvorite datoteku NMDA.CCS (slika 6.3.). Sad će
aferentna stimulacija imati za posljedicu nastanak EPSP-a posredovanog
NMDA receptorom.
48

Temelji neuroznanosti Vodič kroz vježbe
• Izazovite hiperpolarizaciju neurona tako da iz izbornika Parameters izaberete
Protocol te promijenite vrijednosti Base current(nA) sa 0 u -0.52 nA i Starting
Vm u -90. Pritisnite OK i pomoću Overlay promatrajte rezultat simulacije.
Uočite kako je EPSP posredovan NMDA-receptorom vrlo malen kod
hiperpolariziranog membranskog potencijala (slika 6.3).
• Promijenite vrijednost Base current (nA) u 0.525, a vrijednost Starting Vm u –
30. Pritisnite OK i pomoću naredbe Overlay promatrajte rezultat simulacije.
Uočite kako je EPSP veći kod depolariziranog membranskog potencijala iako
ste bliži potencijalu obrata.
Slika 6.1. Aktivacija ekscitacijske aferentne veze humanih kortikalnih piramidnih stanica ima za posljedicu ekscitaciju praćenu inhibicijskim postsinaptičkim potencijalom. Električno podraživanje aferentnog ekscitacijskog puta ima za posljedicu jaku aktivnost akcijskih potencijala u lokalnim intrakortikalnim inhibicijskim (GABAergičkim) interneuronima (lijevo) kao i izravne depolarizacije piramidnih neurona (desno, EPSP). Snažna aktivnost GABAergičkih neurona ima za posljedicu hiperpolarizaciju (inhibiciju) piramidnih stanica povećanjem vodljivosti za Cl- putem GABAA receptora i povećanjem vodljivosti za K+ putem GABAB receptora.
49
Temelji neuroznanosti Vodič kroz vježbe
• Izazovite hiperpolarizaciju neurona tako da iz izbornika Parameters izaberete
Protocol te promijenite vrijednosti Base current(nA) sa 0 u -0.52 nA i Starting
Vm u -90. Pritisnite OK i pomoću Overlay promatrajte rezultat simulacije.
Uočite kako je EPSP posredovan NMDA-receptorom vrlo malen kod
hiperpolariziranog membranskog potencijala (slika 6.3).
• Promijenite vrijednost Base current (nA) u 0.525, a vrijednost Starting Vm u –
30. Pritisnite OK i pomoću naredbe Overlay promatrajte rezultat simulacije.
Uočite kako je EPSP veći kod depolariziranog membranskog potencijala iako
ste bliži potencijalu obrata.
Slika 6.1. Aktivacija ekscitacijske aferentne veze humanih kortikalnih piramidnih stanica ima za posljedicu ekscitaciju praćenu inhibicijskim postsinaptičkim potencijalom. Električno podraživanje aferentnog ekscitacijskog puta ima za posljedicu jaku aktivnost akcijskih potencijala u lokalnim intrakortikalnim inhibicijskim (GABAergičkim) interneuronima (lijevo) kao i izravne depolarizacije piramidnih neurona (desno, EPSP). Snažna aktivnost GABAergičkih neurona ima za posljedicu hiperpolarizaciju (inhibiciju) piramidnih stanica povećanjem vodljivosti za Cl- putem GABAA receptora i povećanjem vodljivosti za K+ putem GABAB receptora.
49

Temelji neuroznanosti Vodič kroz vježbe
Slika 6.2. Ekscitacijski i inhibicijski postsinaptički potencijal. Ekscitacijski postsinaptički potencijal posredovan je AMPA i NMDA receptorima, dok je inhibicijski postsinaptički potencijal posredovan GABAA i GABAB receptorima staničnog modela. Blokiranje inhibicijskog potencijala ima za posljedicu povećanje amplitude EPSP-a. EPSP dovodi do obrata polarnosti na 0 mV.
Izmjenom izvanstanične koncentracije različitih iona, Linda Nowak, Philip
Ascher i suradnici, došli su do spoznaje o značenju iona Mg++ u izvanstaničnoj
tekućini u naponskoj ovisnosti EPSP-a posredovanog NMDA receptorom (Nowak i
sur., 1984.).
• Vratite vrijednosti Base current na 0 i Starting Vm na -55mV.
• Pomoću padajućeg izbornika Parameters i naredbe Ions promijenite
izvanstaničnu koncentraciju [Mg++]o iz vrijednosti 1.2 u 0.01 mM. Pritisnite
OK i pomoću naredbe Overlay promatrajte rezultat simulacije. Uočite kako je
EPSP znatno veći nakon što ste uklonili Mg++ iz tekućeg medija (slika 6.3.).
• Ispitajte je li moguće dobiti isti učinak ako je membrana depolarizirana tako
da pomoću padajućeg izbornika Parameters i naredbe Protocol promijenite
vrijednosti Base current (nA)= u 0.525 i Starting Vm u -30. Pritisnite OK i
pomoću naredbe Overlay promatrajte rezultat simulacije.
• Provjerite kakav je učinak ako je membrana hiperpolarizirana. Promijenite
vrijednost Base current u –0.52, a Starting Vm u –90mV. Pritisnite OK i
Overlay i pokušajte objasniti promjene koje vidite.
Ukoliko na zaslonu vidite samo promjene sa smanjenim vrijednostima
magnezija, učinite slijedeće:
50
Temelji neuroznanosti Vodič kroz vježbe
Slika 6.2. Ekscitacijski i inhibicijski postsinaptički potencijal. Ekscitacijski postsinaptički potencijal posredovan je AMPA i NMDA receptorima, dok je inhibicijski postsinaptički potencijal posredovan GABAA i GABAB receptorima staničnog modela. Blokiranje inhibicijskog potencijala ima za posljedicu povećanje amplitude EPSP-a. EPSP dovodi do obrata polarnosti na 0 mV.
Izmjenom izvanstanične koncentracije različitih iona, Linda Nowak, Philip
Ascher i suradnici, došli su do spoznaje o značenju iona Mg++ u izvanstaničnoj
tekućini u naponskoj ovisnosti EPSP-a posredovanog NMDA receptorom (Nowak i
sur., 1984.).
• Vratite vrijednosti Base current na 0 i Starting Vm na -55mV.
• Pomoću padajućeg izbornika Parameters i naredbe Ions promijenite
izvanstaničnu koncentraciju [Mg++]o iz vrijednosti 1.2 u 0.01 mM. Pritisnite
OK i pomoću naredbe Overlay promatrajte rezultat simulacije. Uočite kako je
EPSP znatno veći nakon što ste uklonili Mg++ iz tekućeg medija (slika 6.3.).
• Ispitajte je li moguće dobiti isti učinak ako je membrana depolarizirana tako
da pomoću padajućeg izbornika Parameters i naredbe Protocol promijenite
vrijednosti Base current (nA)= u 0.525 i Starting Vm u -30. Pritisnite OK i
pomoću naredbe Overlay promatrajte rezultat simulacije.
• Provjerite kakav je učinak ako je membrana hiperpolarizirana. Promijenite
vrijednost Base current u –0.52, a Starting Vm u –90mV. Pritisnite OK i
Overlay i pokušajte objasniti promjene koje vidite.
Ukoliko na zaslonu vidite samo promjene sa smanjenim vrijednostima
magnezija, učinite slijedeće:
50

Temelji neuroznanosti Vodič kroz vježbe
• Promijenite [Mg++]o s 0 na 1.2 te pritisnite OK i pomoću naredbe Overlay
promatrajte rezultat. Uočite kako ioni magnezija imaju malen učinak na EPSP
posredovan NMDA receptorom kad je membrana depolarizirana, nasuprot
hiperpolariziranom membranskom potencijalu (slika 6.3.).
Slika 6.3. Naponska ovisnost NMDA receptora posredovana je blokirajućim učinkom iona magnezija. Prilikom smanjenja koncentracije magnezija u izvanstaničnom prostoru, amplituda EPSP-a posredovanog NMDA receptorom veća je pri hiperpolariziranom membranskom potencijalu (-90 mV) nego pri depolariziranom membranskom potencijalu (-30 mV) zahvaljujući približavanju ravnotežnom potencijalu za NMDA struju (0 mV). Nasuprot tome, uz normalne vrijednosti Mg++ iona u izvanstaničnom prostoru, ovaj ion blokira NMDA struju pri hiperpolariziranom membranskom potencijalu (-55 i -90 mV) i stoga rezultira neobičnom naponskom ovisnošću.
51
Temelji neuroznanosti Vodič kroz vježbe
• Promijenite [Mg++]o s 0 na 1.2 te pritisnite OK i pomoću naredbe Overlay
promatrajte rezultat. Uočite kako ioni magnezija imaju malen učinak na EPSP
posredovan NMDA receptorom kad je membrana depolarizirana, nasuprot
hiperpolariziranom membranskom potencijalu (slika 6.3.).
Slika 6.3. Naponska ovisnost NMDA receptora posredovana je blokirajućim učinkom iona magnezija. Prilikom smanjenja koncentracije magnezija u izvanstaničnom prostoru, amplituda EPSP-a posredovanog NMDA receptorom veća je pri hiperpolariziranom membranskom potencijalu (-90 mV) nego pri depolariziranom membranskom potencijalu (-30 mV) zahvaljujući približavanju ravnotežnom potencijalu za NMDA struju (0 mV). Nasuprot tome, uz normalne vrijednosti Mg++ iona u izvanstaničnom prostoru, ovaj ion blokira NMDA struju pri hiperpolariziranom membranskom potencijalu (-55 i -90 mV) i stoga rezultira neobičnom naponskom ovisnošću.
51
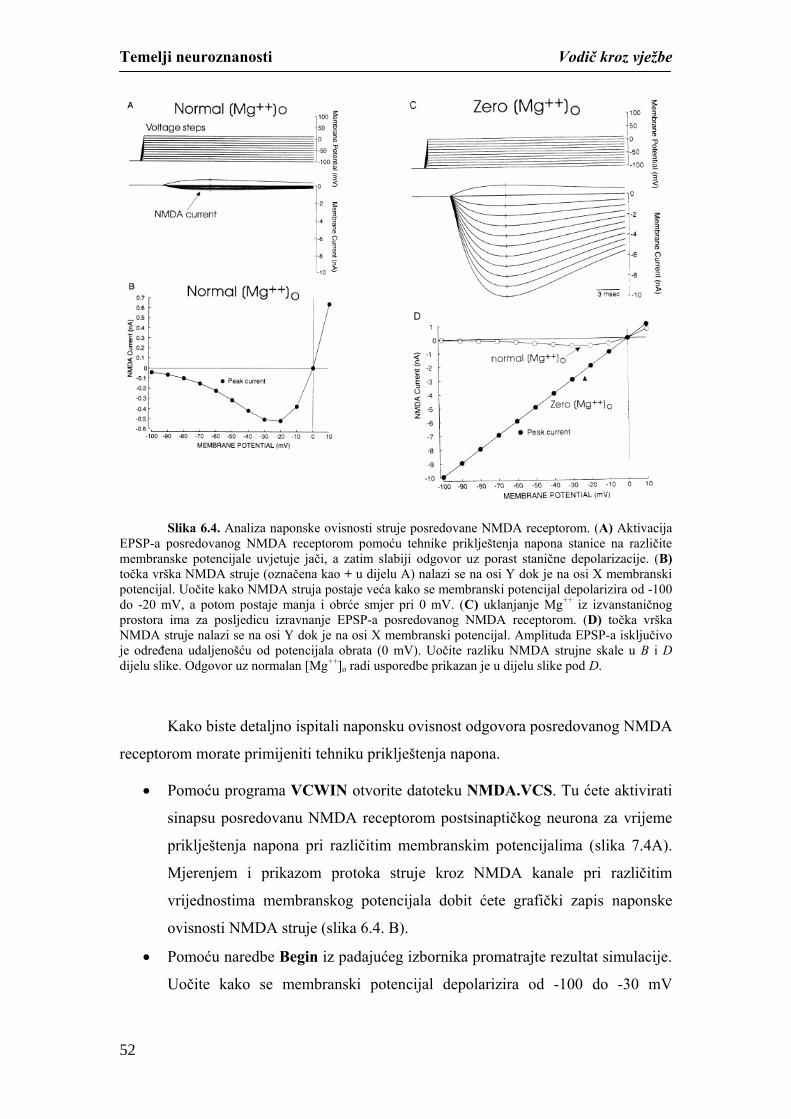
Temelji neuroznanosti Vodič kroz vježbe
Slika 6.4. Analiza naponske ovisnosti struje posredovane NMDA receptorom. (A) Aktivacija EPSP-a posredovanog NMDA receptorom pomoću tehnike priklještenja napona stanice na različite membranske potencijale uvjetuje jači, a zatim slabiji odgovor uz porast stanične depolarizacije. (B) točka vrška NMDA struje (označena kao + u dijelu A) nalazi se na osi Y dok je na osi X membranski potencijal. Uočite kako NMDA struja postaje veća kako se membranski potencijal depolarizira od -100 do -20 mV, a potom postaje manja i obrće smjer pri 0 mV. (C) uklanjanje Mg++ iz izvanstaničnog prostora ima za posljedicu izravnanje EPSP-a posredovanog NMDA receptorom. (D) točka vrška NMDA struje nalazi se na osi Y dok je na osi X membranski potencijal. Amplituda EPSP-a isključivo je određena udaljenošću od potencijala obrata (0 mV). Uočite razliku NMDA strujne skale u B i D dijelu slike. Odgovor uz normalan [Mg++]o radi usporedbe prikazan je u dijelu slike pod D.
Kako biste detaljno ispitali naponsku ovisnost odgovora posredovanog NMDA
receptorom morate primijeniti tehniku priklještenja napona.
• Pomoću programa VCWIN otvorite datoteku NMDA.VCS. Tu ćete aktivirati
sinapsu posredovanu NMDA receptorom postsinaptičkog neurona za vrijeme
priklještenja napona pri različitim membranskim potencijalima (slika 7.4A).
Mjerenjem i prikazom protoka struje kroz NMDA kanale pri različitim
vrijednostima membranskog potencijala dobit ćete grafički zapis naponske
ovisnosti NMDA struje (slika 6.4. B).
• Pomoću naredbe Begin iz padajućeg izbornika promatrajte rezultat simulacije.
Uočite kako se membranski potencijal depolarizira od -100 do -30 mV
52
Temelji neuroznanosti Vodič kroz vježbe
Slika 6.4. Analiza naponske ovisnosti struje posredovane NMDA receptorom. (A) Aktivacija EPSP-a posredovanog NMDA receptorom pomoću tehnike priklještenja napona stanice na različite membranske potencijale uvjetuje jači, a zatim slabiji odgovor uz porast stanične depolarizacije. (B) točka vrška NMDA struje (označena kao + u dijelu A) nalazi se na osi Y dok je na osi X membranski potencijal. Uočite kako NMDA struja postaje veća kako se membranski potencijal depolarizira od -100 do -20 mV, a potom postaje manja i obrće smjer pri 0 mV. (C) uklanjanje Mg++ iz izvanstaničnog prostora ima za posljedicu izravnanje EPSP-a posredovanog NMDA receptorom. (D) točka vrška NMDA struje nalazi se na osi Y dok je na osi X membranski potencijal. Amplituda EPSP-a isključivo je određena udaljenošću od potencijala obrata (0 mV). Uočite razliku NMDA strujne skale u B i D dijelu slike. Odgovor uz normalan [Mg++]o radi usporedbe prikazan je u dijelu slike pod D.
Kako biste detaljno ispitali naponsku ovisnost odgovora posredovanog NMDA
receptorom morate primijeniti tehniku priklještenja napona.
• Pomoću programa VCWIN otvorite datoteku NMDA.VCS. Tu ćete aktivirati
sinapsu posredovanu NMDA receptorom postsinaptičkog neurona za vrijeme
priklještenja napona pri različitim membranskim potencijalima (slika 7.4A).
Mjerenjem i prikazom protoka struje kroz NMDA kanale pri različitim
vrijednostima membranskog potencijala dobit ćete grafički zapis naponske
ovisnosti NMDA struje (slika 6.4. B).
• Pomoću naredbe Begin iz padajućeg izbornika promatrajte rezultat simulacije.
Uočite kako se membranski potencijal depolarizira od -100 do -30 mV
52

Temelji neuroznanosti Vodič kroz vježbe
(NMDA struja postaje veća iako pri većim vrijednostima depolarizacije
ponovno postaje manja i čak dolazi do obrata smjera pri 0 mV, slika 7.4.B).
• Smanjite koncentraciju izvanstaničnog magnezija [Mg++]o na 0.001 mM.
Pritisnite OK i pomoću naredbe Overlay promatrajte rezultat simulacije.
Uočite kako EPSP posredovan NMDA receptorom više nema naponsku
ovisnost (slika 6.4. C i D)
Rezultat te simulacije navodi Vas na postavljanje hipoteze prema kojoj
NMDA receptori imaju naponsku ovisnost zato što kod hiperpolariziranog
membranskog potencijala, negativni potencijal unutar stanice privlači Mg++ koji će
pokušati ući u stanicu. Onog trenutka kada ioni magnezija uđu u NMDA kanale, isti
postaju "začepljeni" tako da nema prolaza za druge ione.
Kad je membranski potencijal depolariziran, privlačenje Mg++ za ulazak u
stanicu znatno je smanjeno. Stoga, kad su NMDA kanali otvoreni, svega ih nekoliko
ostaje začepljeno ionima Mg++.
Možete izvesti sličan pokus s izoliranim EPSP-om posredovanim AMPA
receptorom. Zaključak bi trebao biti da AMPA receptori nisu ovisni o naponu i na njih
ne utječe [Mg++]o.
Vježba 6.2: Inhibicijski postsinaptički potencijali
Blokiranjem ekscitacijskih postsinaptičkih receptora posredno pomoću
farmakoloških pripravaka ili izravno stimulacijom inhibicijskih interneurona, ispitat
ćete inhibicijske postsinaptičke potencijale.
• Pomoću aplikacije CCWIN otvorite datoteku IPSPS.CCS te preko naredbe
Begin iz padajućeg izbornika Run promatrajte rezultat. Uočite bifazični ili
dvodijelni, inhibicijski potencijal. Provjerite kako je prvi dio posredovan
aktivacijom GABAA receptora, dok je drugi dio posredovan aktivacijom
GABAB receptora (slika 6.5.).
• Sad ispitajte ionski mehanizam nastanka različitih IPSP-a mijenjanjem
membranskog potencijala i koncentracije iona u tekućem izvanstaničnom
mediju. Pomoću padajućeg izbornika Parameters i naredbe Protocol
promijenite vrijednost Base current (nA) od 0 u -0.38 nA te Starting Vm u -85
mV. Pritisnite OK i pomoću naredbe Overlay promotrite rezultat simulacije.
53
Temelji neuroznanosti Vodič kroz vježbe
(NMDA struja postaje veća iako pri većim vrijednostima depolarizacije
ponovno postaje manja i čak dolazi do obrata smjera pri 0 mV, slika 7.4.B).
• Smanjite koncentraciju izvanstaničnog magnezija [Mg++]o na 0.001 mM.
Pritisnite OK i pomoću naredbe Overlay promatrajte rezultat simulacije.
Uočite kako EPSP posredovan NMDA receptorom više nema naponsku
ovisnost (slika 6.4. C i D)
Rezultat te simulacije navodi Vas na postavljanje hipoteze prema kojoj
NMDA receptori imaju naponsku ovisnost zato što kod hiperpolariziranog
membranskog potencijala, negativni potencijal unutar stanice privlači Mg++ koji će
pokušati ući u stanicu. Onog trenutka kada ioni magnezija uđu u NMDA kanale, isti
postaju "začepljeni" tako da nema prolaza za druge ione.
Kad je membranski potencijal depolariziran, privlačenje Mg++ za ulazak u
stanicu znatno je smanjeno. Stoga, kad su NMDA kanali otvoreni, svega ih nekoliko
ostaje začepljeno ionima Mg++.
Možete izvesti sličan pokus s izoliranim EPSP-om posredovanim AMPA
receptorom. Zaključak bi trebao biti da AMPA receptori nisu ovisni o naponu i na njih
ne utječe [Mg++]o.
Vježba 6.2: Inhibicijski postsinaptički potencijali
Blokiranjem ekscitacijskih postsinaptičkih receptora posredno pomoću
farmakoloških pripravaka ili izravno stimulacijom inhibicijskih interneurona, ispitat
ćete inhibicijske postsinaptičke potencijale.
• Pomoću aplikacije CCWIN otvorite datoteku IPSPS.CCS te preko naredbe
Begin iz padajućeg izbornika Run promatrajte rezultat. Uočite bifazični ili
dvodijelni, inhibicijski potencijal. Provjerite kako je prvi dio posredovan
aktivacijom GABAA receptora, dok je drugi dio posredovan aktivacijom
GABAB receptora (slika 6.5.).
• Sad ispitajte ionski mehanizam nastanka različitih IPSP-a mijenjanjem
membranskog potencijala i koncentracije iona u tekućem izvanstaničnom
mediju. Pomoću padajućeg izbornika Parameters i naredbe Protocol
promijenite vrijednost Base current (nA) od 0 u -0.38 nA te Starting Vm u -85
mV. Pritisnite OK i pomoću naredbe Overlay promotrite rezultat simulacije.
53

Temelji neuroznanosti Vodič kroz vježbe
Uočite prvi IPSP koji je obrnut (depolarizacijski), dok kasniji IPSP nije (još
uvijek je hiperpolarizacijski) (slika 6.5.). Vjerojatno je razlog utjecaj različitih iona
na njihov nastanak.
Ispitajte sada ovisnost različitih IPSP-a o koncentracijama iona u
izvanstaničnoj tekućini.
• Ponovno otvorite datoteku IPSPS.CCS. Pretpostavite sljedeće: rani GABAA
IPSP posredovan je ionima Cl-, što proizlazi iz rezultata ispitivanja ovisnosti o
naponu koji su pokazali obrat prema ravnotežnom potencijalu za Cl- (ECl; -75
mV).
• Smanjite [Cl-]o sa 120 na 7 mM. Pritisnite OK i pomoću naredbe Overlay
promotrite rezultat simulacije.
Uočite kako se GABAA IPSP sad depolarizira, što ukazuje na promjenu
ravnotežnog potencijala za Cl- na način da membranski potencijal stanice postaje
pozitivan (što je u skladu s promjenom smjera protoka kloridnih iona). U stanju
mirovanja, ioni Cl- prolaze iz izvanstaničnog prostora u unutarstanični, a potom se
pomiču u obrnutom smjeru tj. iz unutarstaničnog prostora u izvanstanični (slika 6.5.;
[Cl-]i=[Cl-]o).
• Ponovno otvorite datoteku IPSPS.CSS (vraćamo se na početne vrijednosti).
• Promijenite izvanstaničnu koncentraciju K+ s 3.1 na 25 mM te vrijednost Base
current(nA) u -0.5. Uočite da se kasni GABAB IPSP depolarizira, što je
rezultat povećanja K+ vodljivosti (slika 6.5.; [K+]o=25).
Često zbunjuje podatak da se sinaptička transmisija matematički prikazuje kao
jednakost između depolarizacijskog potencijala i ekscitacijske transmisije, te
hiperpolarizacijskog sinaptičkog potencijala i inhibicijske sinaptičke transmisije.
Vidjeli ste kako hiperpolarizacijom stanice iznad ECl, hiperpolarizacijski IPSP
može postati depolarizacijski. Dovodi li to do promjene inhibicijskog sinaptičkog
potencijala u ekscitacijski? Odgovor je NE, jer je činjenica da iako se IPSP
depolarizira, njegov ravnotežni potencijal još je uvijek -75 mV, dakle 20 mV iznad
praga okidanja za nastanak akcijskog potencijala (obično -55 mV).
54
Temelji neuroznanosti Vodič kroz vježbe
Uočite prvi IPSP koji je obrnut (depolarizacijski), dok kasniji IPSP nije (još
uvijek je hiperpolarizacijski) (slika 6.5.). Vjerojatno je razlog utjecaj različitih iona
na njihov nastanak.
Ispitajte sada ovisnost različitih IPSP-a o koncentracijama iona u
izvanstaničnoj tekućini.
• Ponovno otvorite datoteku IPSPS.CCS. Pretpostavite sljedeće: rani GABAA
IPSP posredovan je ionima Cl-, što proizlazi iz rezultata ispitivanja ovisnosti o
naponu koji su pokazali obrat prema ravnotežnom potencijalu za Cl- (ECl; -75
mV).
• Smanjite [Cl-]o sa 120 na 7 mM. Pritisnite OK i pomoću naredbe Overlay
promotrite rezultat simulacije.
Uočite kako se GABAA IPSP sad depolarizira, što ukazuje na promjenu
ravnotežnog potencijala za Cl- na način da membranski potencijal stanice postaje
pozitivan (što je u skladu s promjenom smjera protoka kloridnih iona). U stanju
mirovanja, ioni Cl- prolaze iz izvanstaničnog prostora u unutarstanični, a potom se
pomiču u obrnutom smjeru tj. iz unutarstaničnog prostora u izvanstanični (slika 6.5.;
[Cl-]i=[Cl-]o).
• Ponovno otvorite datoteku IPSPS.CSS (vraćamo se na početne vrijednosti).
• Promijenite izvanstaničnu koncentraciju K+ s 3.1 na 25 mM te vrijednost Base
current(nA) u -0.5. Uočite da se kasni GABAB IPSP depolarizira, što je
rezultat povećanja K+ vodljivosti (slika 6.5.; [K+]o=25).
Često zbunjuje podatak da se sinaptička transmisija matematički prikazuje kao
jednakost između depolarizacijskog potencijala i ekscitacijske transmisije, te
hiperpolarizacijskog sinaptičkog potencijala i inhibicijske sinaptičke transmisije.
Vidjeli ste kako hiperpolarizacijom stanice iznad ECl, hiperpolarizacijski IPSP
može postati depolarizacijski. Dovodi li to do promjene inhibicijskog sinaptičkog
potencijala u ekscitacijski? Odgovor je NE, jer je činjenica da iako se IPSP
depolarizira, njegov ravnotežni potencijal još je uvijek -75 mV, dakle 20 mV iznad
praga okidanja za nastanak akcijskog potencijala (obično -55 mV).
54
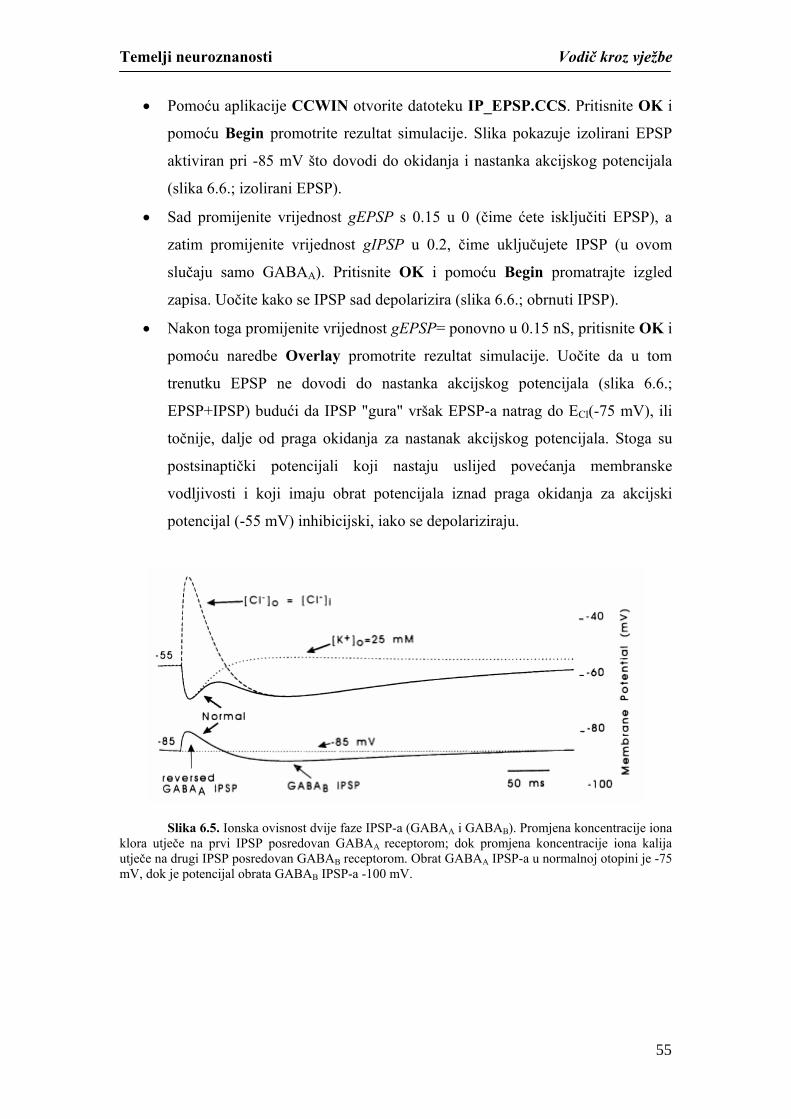
Temelji neuroznanosti Vodič kroz vježbe
• Pomoću aplikacije CCWIN otvorite datoteku IP_EPSP.CCS. Pritisnite OK i
pomoću Begin promotrite rezultat simulacije. Slika pokazuje izolirani EPSP
aktiviran pri -85 mV što dovodi do okidanja i nastanka akcijskog potencijala
(slika 6.6.; izolirani EPSP).
• Sad promijenite vrijednost gEPSP s 0.15 u 0 (čime ćete isključiti EPSP), a
zatim promijenite vrijednost gIPSP u 0.2, čime uključujete IPSP (u ovom
slučaju samo GABAA). Pritisnite OK i pomoću Begin promatrajte izgled
zapisa. Uočite kako se IPSP sad depolarizira (slika 6.6.; obrnuti IPSP).
• Nakon toga promijenite vrijednost gEPSP= ponovno u 0.15 nS, pritisnite OK i
pomoću naredbe Overlay promotrite rezultat simulacije. Uočite da u tom
trenutku EPSP ne dovodi do nastanka akcijskog potencijala (slika 6.6.;
EPSP+IPSP) budući da IPSP "gura" vršak EPSP-a natrag do ECl(-75 mV), ili
točnije, dalje od praga okidanja za nastanak akcijskog potencijala. Stoga su
postsinaptički potencijali koji nastaju uslijed povećanja membranske
vodljivosti i koji imaju obrat potencijala iznad praga okidanja za akcijski
potencijal (-55 mV) inhibicijski, iako se depolariziraju.
Slika 6.5. Ionska ovisnost dvije faze IPSP-a (GABAA i GABAB). Promjena koncentracije iona klora utječe na prvi IPSP posredovan GABAA receptorom; dok promjena koncentracije iona kalija utječe na drugi IPSP posredovan GABAB receptorom. Obrat GABAA IPSP-a u normalnoj otopini je -75 mV, dok je potencijal obrata GABAB IPSP-a -100 mV.
55
Temelji neuroznanosti Vodič kroz vježbe
• Pomoću aplikacije CCWIN otvorite datoteku IP_EPSP.CCS. Pritisnite OK i
pomoću Begin promotrite rezultat simulacije. Slika pokazuje izolirani EPSP
aktiviran pri -85 mV što dovodi do okidanja i nastanka akcijskog potencijala
(slika 6.6.; izolirani EPSP).
• Sad promijenite vrijednost gEPSP s 0.15 u 0 (čime ćete isključiti EPSP), a
zatim promijenite vrijednost gIPSP u 0.2, čime uključujete IPSP (u ovom
slučaju samo GABAA). Pritisnite OK i pomoću Begin promatrajte izgled
zapisa. Uočite kako se IPSP sad depolarizira (slika 6.6.; obrnuti IPSP).
• Nakon toga promijenite vrijednost gEPSP= ponovno u 0.15 nS, pritisnite OK i
pomoću naredbe Overlay promotrite rezultat simulacije. Uočite da u tom
trenutku EPSP ne dovodi do nastanka akcijskog potencijala (slika 6.6.;
EPSP+IPSP) budući da IPSP "gura" vršak EPSP-a natrag do ECl(-75 mV), ili
točnije, dalje od praga okidanja za nastanak akcijskog potencijala. Stoga su
postsinaptički potencijali koji nastaju uslijed povećanja membranske
vodljivosti i koji imaju obrat potencijala iznad praga okidanja za akcijski
potencijal (-55 mV) inhibicijski, iako se depolariziraju.
Slika 6.5. Ionska ovisnost dvije faze IPSP-a (GABAA i GABAB). Promjena koncentracije iona klora utječe na prvi IPSP posredovan GABAA receptorom; dok promjena koncentracije iona kalija utječe na drugi IPSP posredovan GABAB receptorom. Obrat GABAA IPSP-a u normalnoj otopini je -75 mV, dok je potencijal obrata GABAB IPSP-a -100 mV.
55
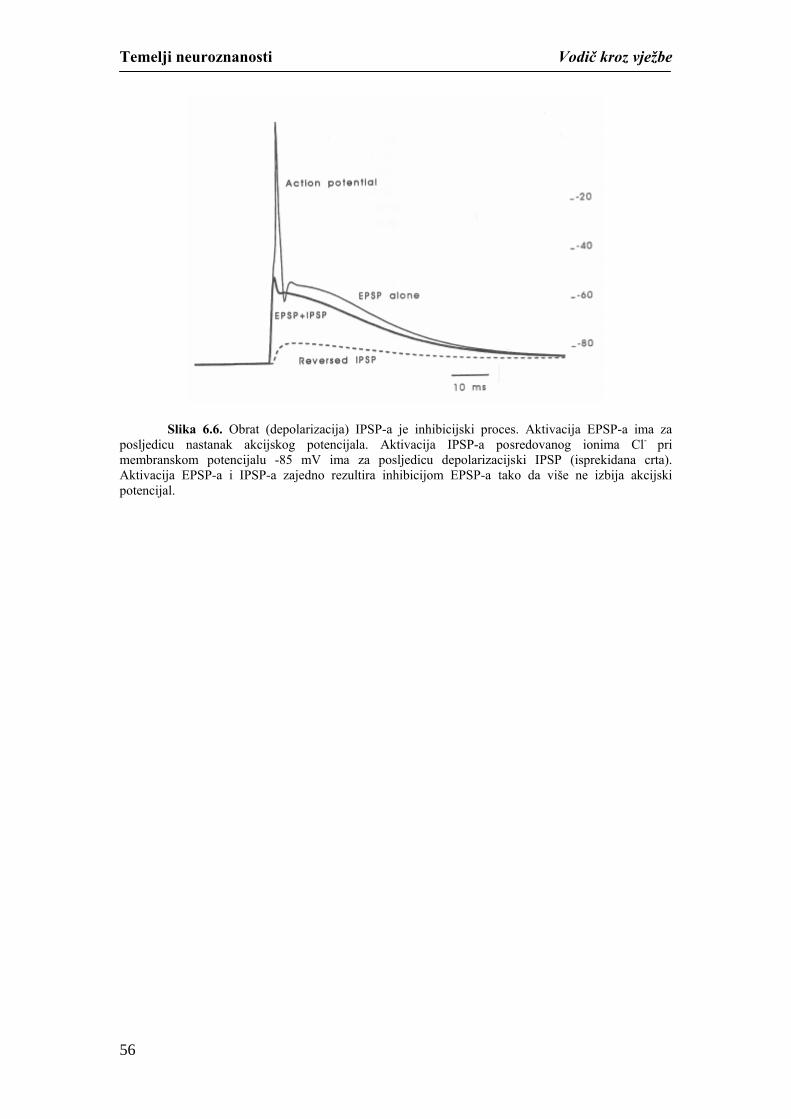
Temelji neuroznanosti Vodič kroz vježbe
Slika 6.6. Obrat (depolarizacija) IPSP-a je inhibicijski proces. Aktivacija EPSP-a ima za posljedicu nastanak akcijskog potencijala. Aktivacija IPSP-a posredovanog ionima Cl- pri membranskom potencijalu -85 mV ima za posljedicu depolarizacijski IPSP (isprekidana crta). Aktivacija EPSP-a i IPSP-a zajedno rezultira inhibicijom EPSP-a tako da više ne izbija akcijski potencijal.
56
Temelji neuroznanosti Vodič kroz vježbe
Slika 6.6. Obrat (depolarizacija) IPSP-a je inhibicijski proces. Aktivacija EPSP-a ima za posljedicu nastanak akcijskog potencijala. Aktivacija IPSP-a posredovanog ionima Cl- pri membranskom potencijalu -85 mV ima za posljedicu depolarizacijski IPSP (isprekidana crta). Aktivacija EPSP-a i IPSP-a zajedno rezultira inhibicijom EPSP-a tako da više ne izbija akcijski potencijal.
56

Temelji neuroznanosti Vodič kroz vježbe
6.2 Pitanja: Sinaptički potencijali
• Kakav je učinak glutamata na stanicu? Kako djeluje GABA?
• Ako se koncentracija izvanstaničnog i unutarstaničnog Cl- izjednači, hoće li
aktivacija GABAA receptora biti inhibicijska ili aktivacijska? Zašto? (To
možete ispitati ako otvorite datoteku IPSP.CCS koristeći aplikaciju CCWin,
izjednačenjem [Cl-]o=[Cl-]i, uključite Na+ i kasnu odgođenu struju K+ na 10
odnosno 2 µS).
• Koja su tri glavna čimbenika kontrole amplitude EPSP-a posredovanog
NMDA receptorom?
• Zašto EPSP posredovan NMDA i AMPA receptorom obrće polarnost pri 0
mV?
57
Temelji neuroznanosti Vodič kroz vježbe
6.2 Pitanja: Sinaptički potencijali
• Kakav je učinak glutamata na stanicu? Kako djeluje GABA?
• Ako se koncentracija izvanstaničnog i unutarstaničnog Cl- izjednači, hoće li
aktivacija GABAA receptora biti inhibicijska ili aktivacijska? Zašto? (To
možete ispitati ako otvorite datoteku IPSP.CCS koristeći aplikaciju CCWin,
izjednačenjem [Cl-]o=[Cl-]i, uključite Na+ i kasnu odgođenu struju K+ na 10
odnosno 2 µS).
• Koja su tri glavna čimbenika kontrole amplitude EPSP-a posredovanog
NMDA receptorom?
• Zašto EPSP posredovan NMDA i AMPA receptorom obrće polarnost pri 0
mV?
57

Temelji neuroznanosti Vodič kroz vježbe
7. Prostorna i vremenska konstanta
7.1 Uvod
Aplikacija Neurosim simulira pasivna svojstva neurona koji ne stvara akcijske
potencijale. Pretpostavimo izgled tog neurona. Neuron ima beskonačno dug akson u
kojemu su postavljene četiri elektrode. Jedna elektroda služi za injiciranje pozitivne
struje, dok preostale elektrode služe za mjerenje napona. Program daje mogućnost
promjene amplitude i trajanja injicirane struje te promjene položaja elektroda za
snimanje napona u odnosu na položaj elektrode za injiciranje struje. Također, postoji
mogućnost izmjene građe aksona prema vlastitoj želji mijenjanjem membranskih
svojstava i promjera aksona izborom odgovarajućih opcija u izborniku.
Cilj vježbe je ispitati kako promjene napona nakon injiciranja struje variraju
vremenom i udaljenošću u ovisnosti o temeljnim svojstvima aksona.
Detaljno proučite dobivene rezultate budući da je razumijevanje osnovnih
obilježja građe aksona neobično važno za razumijevanje funkcioniranja neurona u
SŽS-u.
• Pronađite folder naziva "Neurosim" i u njemu "Cable", a zatim ikonu naziva
"Cable". S obzirom da je taj program pisan u DOS verziji nećete moći koristiti
miša u daljnjem dijelu vježbe. Umjesto toga koristite tipkovnicu kao i ranije.
• Otvorite "experiment" i uočite četiri zapisa na zaslonu ekrana. Krivulje
predstavljaju napone snimljene pomoću tri elektrode u aksonu. Najveći otklon
daje elektroda za snimanje postavljena najbliže mjestu injiciranja struje.
Ljestvica u lijevom dijelu zapisa pokazuje vrijednost napona. Četvrti zapis ima
kvadratasti otklon i predstavlja podražaj injiciranom strujom. Ljestvica u
donjem desnom dijelu zapisa predstavlja jakost injicirane struje.
• GRAĐA AKSONA ponuđeni su niže navedeni parametri:
Otpor aksoplazme (Ri): To je unutarnji otpor citoplazme aksona. Ne pokazuje
značajne razlike među različitim aksonima. Prihvatljiv raspon veličine je 0,02
– 0,5 kOhm cm2
Otpor membrane (Rm): To je otpor dijela membrane po centimetru površine.
Pokazuje značajne varijacije među različitim tipovima aksona. Prihvatljiv
raspon veličine je 0,1 - 50 kOhm cm2.
58
Temelji neuroznanosti Vodič kroz vježbe
7. Prostorna i vremenska konstanta
7.1 Uvod
Aplikacija Neurosim simulira pasivna svojstva neurona koji ne stvara akcijske
potencijale. Pretpostavimo izgled tog neurona. Neuron ima beskonačno dug akson u
kojemu su postavljene četiri elektrode. Jedna elektroda služi za injiciranje pozitivne
struje, dok preostale elektrode služe za mjerenje napona. Program daje mogućnost
promjene amplitude i trajanja injicirane struje te promjene položaja elektroda za
snimanje napona u odnosu na položaj elektrode za injiciranje struje. Također, postoji
mogućnost izmjene građe aksona prema vlastitoj želji mijenjanjem membranskih
svojstava i promjera aksona izborom odgovarajućih opcija u izborniku.
Cilj vježbe je ispitati kako promjene napona nakon injiciranja struje variraju
vremenom i udaljenošću u ovisnosti o temeljnim svojstvima aksona.
Detaljno proučite dobivene rezultate budući da je razumijevanje osnovnih
obilježja građe aksona neobično važno za razumijevanje funkcioniranja neurona u
SŽS-u.
• Pronađite folder naziva "Neurosim" i u njemu "Cable", a zatim ikonu naziva
"Cable". S obzirom da je taj program pisan u DOS verziji nećete moći koristiti
miša u daljnjem dijelu vježbe. Umjesto toga koristite tipkovnicu kao i ranije.
• Otvorite "experiment" i uočite četiri zapisa na zaslonu ekrana. Krivulje
predstavljaju napone snimljene pomoću tri elektrode u aksonu. Najveći otklon
daje elektroda za snimanje postavljena najbliže mjestu injiciranja struje.
Ljestvica u lijevom dijelu zapisa pokazuje vrijednost napona. Četvrti zapis ima
kvadratasti otklon i predstavlja podražaj injiciranom strujom. Ljestvica u
donjem desnom dijelu zapisa predstavlja jakost injicirane struje.
• GRAĐA AKSONA ponuđeni su niže navedeni parametri:
Otpor aksoplazme (Ri): To je unutarnji otpor citoplazme aksona. Ne pokazuje
značajne razlike među različitim aksonima. Prihvatljiv raspon veličine je 0,02
– 0,5 kOhm cm2
Otpor membrane (Rm): To je otpor dijela membrane po centimetru površine.
Pokazuje značajne varijacije među različitim tipovima aksona. Prihvatljiv
raspon veličine je 0,1 - 50 kOhm cm2.
58

Temelji neuroznanosti Vodič kroz vježbe
Kapacitet membrane (Cm): To je kapacitet 1 cm2 površine membrane.
Obzirom da je najvećim dijelom određen debljinom i sastavom tj. građom
lipidnog dvosloja (sličan je kod većine stanica) ne mijenja se značajno među
aksonima. Prihvatljiv raspon vrijednosti je 0,1 - 10 µF/cm2.
Radijus aksona: Prihvatljiv raspon veličine je 0,1 - 500 µm.
Vremenska i prostorna konstanta. Razmatrajući gore navedene značajke
aksona koje određuju vremensku i prostornu konstantu i otpor injiciranom
podražaju proizlazi sljedeće:
VREMENSKA KONSTANTA= CmRm*
PROSTORNA KONSTANTA= rarm /
slijedi:
rm= radius
Rm**2 π
ra= radiusRi
2*π
ulazni otpor= 2* rarm
Izvedene parametre ne možete izravno mijenjati, dok osnovne značajke
neurona možete odrediti i mijenjati u glavnom izborniku.
• Određivanje podražaja ("SET STIMULUS")
Ulaskom u taj podizbornik glavnog izbornika možete primijeniti dva podražaja
struje u akson. Više od dva podražaja možete dodati pritiskom na tipkovnicu
"space bar". Pogledajte u "Options and operations-Change Parameters" kako
unijeti novi parametar i kako se vratiti u glavni izbornik. Određeni su sljedeći
parametri:
Amplituda: Određuje amplitudu injicirane struje. Prihvatljiv raspon vrijednosti
je 0 - 999 nA.
Trajanje podražaja: Određuje trajanje injicirane struje. Prihvatljiv raspon
vrijednosti je 0 - 999 ms.
59
Temelji neuroznanosti Vodič kroz vježbe
Kapacitet membrane (Cm): To je kapacitet 1 cm2 površine membrane.
Obzirom da je najvećim dijelom određen debljinom i sastavom tj. građom
lipidnog dvosloja (sličan je kod većine stanica) ne mijenja se značajno među
aksonima. Prihvatljiv raspon vrijednosti je 0,1 - 10 µF/cm2.
Radijus aksona: Prihvatljiv raspon veličine je 0,1 - 500 µm.
Vremenska i prostorna konstanta. Razmatrajući gore navedene značajke
aksona koje određuju vremensku i prostornu konstantu i otpor injiciranom
podražaju proizlazi sljedeće:
VREMENSKA KONSTANTA= CmRm*
PROSTORNA KONSTANTA= rarm /
slijedi:
rm= radius
Rm**2 π
ra= radiusRi
2*π
ulazni otpor= 2* rarm
Izvedene parametre ne možete izravno mijenjati, dok osnovne značajke
neurona možete odrediti i mijenjati u glavnom izborniku.
• Određivanje podražaja ("SET STIMULUS")
Ulaskom u taj podizbornik glavnog izbornika možete primijeniti dva podražaja
struje u akson. Više od dva podražaja možete dodati pritiskom na tipkovnicu
"space bar". Pogledajte u "Options and operations-Change Parameters" kako
unijeti novi parametar i kako se vratiti u glavni izbornik. Određeni su sljedeći
parametri:
Amplituda: Određuje amplitudu injicirane struje. Prihvatljiv raspon vrijednosti
je 0 - 999 nA.
Trajanje podražaja: Određuje trajanje injicirane struje. Prihvatljiv raspon
vrijednosti je 0 - 999 ms.
59

Temelji neuroznanosti Vodič kroz vježbe
Interval između dva podražaja: Dva podražaja okidaju automatski u toj
simulaciji. Prvi nastaje na početku krivulje potencijala, dok je razmak do
početka drugog automatski određen. Podražaj može nastati i automatski
pritiskom na "space bar" tijekom simulacije. Prihvatljiv raspon veličine je 0,5
- 999 ms.
• Postavljanje elektroda ("PLACE ELECTRODES")
Na raspolaganju Vam stoje tri mikroelektrode za snimanje napona označene
kao E1-E3 koje mogu biti postavljene u aksonu na različitim udaljenostima
(mjereno u milimetrima) od mjesta strujnog podražaja. Bilo koja elektroda
postavljena dalje od otprilike 4 prostorne konstante od mjesta injiciranja struje
snima izuzetno male ili gotovo nezamjetne promjene napona. Najveću i
najbržu promjenu napona zabilježit ćete ako elektrodu postavite točno na
mjesto injiciranja. Možete postaviti više od jedne elektrode na to mjesto, pa će
u tom slučaju doći do pojačanja odgovora. Prihvatljiv raspon veličina je 0 -
999 mm.
• Prikazivanje krivulja "TOGGLE TRACE"
Možete mijenjati svaku pojedinu krivulju zapisa elektroda za snimanje, zatim
brisati pomoću ("n") ili ponovno upisati pomoću ("y").
• Prikazivanje ljestvica mjerenja "DISPLAY SCALES"
Time se određuje horizontalni i vertikalni trag zapisa. Za izbor imate:
Napon: Određuje ljestvicu mjerenja za napon. Odgovara okomitoj crti na
lijevom dijelu zaslona. Prihvatljiv raspon vrijednosti je 1 - 999 mV.
Struja: Određuje ljestvicu mjerenja za struju. Odgovara okomitoj crti na
desnom dijelu zaslona. Prihvatljiv raspon vrijednosti je 1 - 999 nA.
Vrijeme: Odgovara vodoravnoj crti na zaslonu (apscisa). Ukoliko je vrijeme
kraće od desetine vremenske konstante, izgled zapisa na zaslonu neće biti
pregledan. Prihvatljiv raspon vrijednosti je 1 - 999 ms.
• "ZAPIS"
• Pohranjivanje parametara "SAVE PARAMETERS"
Automatski izbor imena u slučaju pohranjivanja podataka je ".cab".
• Prikazivanje pohranjenih parametara "LOAD PARAMETERS"
60
Temelji neuroznanosti Vodič kroz vježbe
Interval između dva podražaja: Dva podražaja okidaju automatski u toj
simulaciji. Prvi nastaje na početku krivulje potencijala, dok je razmak do
početka drugog automatski određen. Podražaj može nastati i automatski
pritiskom na "space bar" tijekom simulacije. Prihvatljiv raspon veličine je 0,5
- 999 ms.
• Postavljanje elektroda ("PLACE ELECTRODES")
Na raspolaganju Vam stoje tri mikroelektrode za snimanje napona označene
kao E1-E3 koje mogu biti postavljene u aksonu na različitim udaljenostima
(mjereno u milimetrima) od mjesta strujnog podražaja. Bilo koja elektroda
postavljena dalje od otprilike 4 prostorne konstante od mjesta injiciranja struje
snima izuzetno male ili gotovo nezamjetne promjene napona. Najveću i
najbržu promjenu napona zabilježit ćete ako elektrodu postavite točno na
mjesto injiciranja. Možete postaviti više od jedne elektrode na to mjesto, pa će
u tom slučaju doći do pojačanja odgovora. Prihvatljiv raspon veličina je 0 -
999 mm.
• Prikazivanje krivulja "TOGGLE TRACE"
Možete mijenjati svaku pojedinu krivulju zapisa elektroda za snimanje, zatim
brisati pomoću ("n") ili ponovno upisati pomoću ("y").
• Prikazivanje ljestvica mjerenja "DISPLAY SCALES"
Time se određuje horizontalni i vertikalni trag zapisa. Za izbor imate:
Napon: Određuje ljestvicu mjerenja za napon. Odgovara okomitoj crti na
lijevom dijelu zaslona. Prihvatljiv raspon vrijednosti je 1 - 999 mV.
Struja: Određuje ljestvicu mjerenja za struju. Odgovara okomitoj crti na
desnom dijelu zaslona. Prihvatljiv raspon vrijednosti je 1 - 999 nA.
Vrijeme: Odgovara vodoravnoj crti na zaslonu (apscisa). Ukoliko je vrijeme
kraće od desetine vremenske konstante, izgled zapisa na zaslonu neće biti
pregledan. Prihvatljiv raspon vrijednosti je 1 - 999 ms.
• "ZAPIS"
• Pohranjivanje parametara "SAVE PARAMETERS"
Automatski izbor imena u slučaju pohranjivanja podataka je ".cab".
• Prikazivanje pohranjenih parametara "LOAD PARAMETERS"
60

Temelji neuroznanosti Vodič kroz vježbe
7.2 Zadaci – prostorna i vremenska konstanta
1. Prema prethodno navedenim formulama izračunajte:
Vremensku konstantu_______________________________________
Prostornu konstantu________________________________________
Ulazni otpor______________________________________________
2. Odredite pomoću podataka dobivenih iz mjerenja vršnog potencijala iz
krivulja dobivenih snimanjem elektrodama za snimanje kolika je vremenska konstanta
u datim eksperimentalnim uvjetima. Podudara li se izmjereni rezultat s rezultatom
dobivenim matematički u prethodnom postupku?
Vremenska konstanta=___________ms
3. U glavnom izborniku nalazi se podizbornik Build axon s temeljnim
pasivnim svojstvima neurona koje možete mijenjati prema vlastitim željama. Ispitajte
i odgovorite kako ovisi prostorna konstanta o promjeni radijusa aksona?
Ukoliko ste u pokusu dokazali tu ovisnost, potvrdite rezultat računanjem
pomoću prethodno navedenih formula.
4. Ispitajte i odgovorite kako ovisi prostorna konstanta o promjeni otpora
membrane? Što se događa ako otpor raste, a što ako otpor pada?
61
Temelji neuroznanosti Vodič kroz vježbe
7.2 Zadaci – prostorna i vremenska konstanta
1. Prema prethodno navedenim formulama izračunajte:
Vremensku konstantu_______________________________________
Prostornu konstantu________________________________________
Ulazni otpor______________________________________________
2. Odredite pomoću podataka dobivenih iz mjerenja vršnog potencijala iz
krivulja dobivenih snimanjem elektrodama za snimanje kolika je vremenska konstanta
u datim eksperimentalnim uvjetima. Podudara li se izmjereni rezultat s rezultatom
dobivenim matematički u prethodnom postupku?
Vremenska konstanta=___________ms
3. U glavnom izborniku nalazi se podizbornik Build axon s temeljnim
pasivnim svojstvima neurona koje možete mijenjati prema vlastitim željama. Ispitajte
i odgovorite kako ovisi prostorna konstanta o promjeni radijusa aksona?
Ukoliko ste u pokusu dokazali tu ovisnost, potvrdite rezultat računanjem
pomoću prethodno navedenih formula.
4. Ispitajte i odgovorite kako ovisi prostorna konstanta o promjeni otpora
membrane? Što se događa ako otpor raste, a što ako otpor pada?
61

Temelji neuroznanosti Vodič kroz vježbe
5. Ispitajte i odgovorite kako ovisi prostorna konstanta o promjeni otpora
aksoplazme? Što se događa ako otpor raste, a što ako otpor pada?
6. Ukoliko ste u eksperimentu dokazali kako prostorna konstanta ovisi o
promjeni otpora, potvrdite rezultat računanjem pomoću prethodno navedenih formula.
7. Što je vremenska konstanta? Što je prostorna konstanta? Je li sumacija
lakša (ili teža) uz veću (ili manju) prostornu i vremensku konstantu i zašto?
7.3 Vremenska konstanta i otpor ulaznog signala
Otvorite datoteku "c1". Uočite na zaslonu zapis jedne elektrode, E1 koja je
postavljena na mjesto injiciranja struje. U glavnom izborniku otvorite "experiment".
Pomoću pokazivača (u gornjem desnom dijelu zaslona nalazi se ikona u znaku
križića) izmjerite konačnu vrijednost potencijala izmjerenu elektrodom E1 i potvrdite
da je otpor ulaznog signala istovjetan zapisu u glavnom izborniku (ako se ne možete
prisjetiti te vrijednosti, vratite se u glavni izbornik pomoću <ESC>, provjerite
vrijednost i pritisnite "view").
Konačna vrijednost napona=______mV
62
Temelji neuroznanosti Vodič kroz vježbe
5. Ispitajte i odgovorite kako ovisi prostorna konstanta o promjeni otpora
aksoplazme? Što se događa ako otpor raste, a što ako otpor pada?
6. Ukoliko ste u eksperimentu dokazali kako prostorna konstanta ovisi o
promjeni otpora, potvrdite rezultat računanjem pomoću prethodno navedenih formula.
7. Što je vremenska konstanta? Što je prostorna konstanta? Je li sumacija
lakša (ili teža) uz veću (ili manju) prostornu i vremensku konstantu i zašto?
7.3 Vremenska konstanta i otpor ulaznog signala
Otvorite datoteku "c1". Uočite na zaslonu zapis jedne elektrode, E1 koja je
postavljena na mjesto injiciranja struje. U glavnom izborniku otvorite "experiment".
Pomoću pokazivača (u gornjem desnom dijelu zaslona nalazi se ikona u znaku
križića) izmjerite konačnu vrijednost potencijala izmjerenu elektrodom E1 i potvrdite
da je otpor ulaznog signala istovjetan zapisu u glavnom izborniku (ako se ne možete
prisjetiti te vrijednosti, vratite se u glavni izbornik pomoću <ESC>, provjerite
vrijednost i pritisnite "view").
Konačna vrijednost napona=______mV
62

Temelji neuroznanosti Vodič kroz vježbe
Izmjerite napon nakon latencije jedne vremenske konstante (npr. 10ms) nakon
početka injiciranja struje.
Napon=_________mV
Izračunajte koliki je to postotak konačne vrijednosti potencijala?
=________%
Napomena: U većini udžbenika možete pročitati da je vremenska konstanta
vrijeme potrebno da injicirani strujni podražaj nabije membranski kapacitet na 63%
njegove konačne vrijednosti. Pa ipak, takva se definicija može koristiti samo za
stanicu sferičkog oblika, dok na beskonačno dugom aksonu kakav ovdje promatramo
nije primjenjiva.
Pokušajte zapamtiti i usporediti taj rezultat sa sličnim pokusom kojim se
koristi program "psp.exe" (vidi ispod).
Vratite se u glavni izbornik bez brisanja zapisa (pritisnite <ESC>).
Otvorite datoteku "c2". Uočite da je izgled zapisa istovjetan prethodnom, osim
što je membranski kapacitet dvostruko veći. Otvorite "experiment" i ponovite ista
mjerenja kao u "c1".
Konačna vrijednost napona=______mV
Izmjerite napon nakon latencije jedne vremenske konstante (npr. 10ms) nakon
početka injiciranja struje.
Napon=_________mV
Izračunajte koliki je to postotak konačne vrijednosti potencijala?
=________%
Vratite se u glavni izbornik bez brisanja zapisa (pritisnite <ESC>).
63
Temelji neuroznanosti Vodič kroz vježbe
Izmjerite napon nakon latencije jedne vremenske konstante (npr. 10ms) nakon
početka injiciranja struje.
Napon=_________mV
Izračunajte koliki je to postotak konačne vrijednosti potencijala?
=________%
Napomena: U većini udžbenika možete pročitati da je vremenska konstanta
vrijeme potrebno da injicirani strujni podražaj nabije membranski kapacitet na 63%
njegove konačne vrijednosti. Pa ipak, takva se definicija može koristiti samo za
stanicu sferičkog oblika, dok na beskonačno dugom aksonu kakav ovdje promatramo
nije primjenjiva.
Pokušajte zapamtiti i usporediti taj rezultat sa sličnim pokusom kojim se
koristi program "psp.exe" (vidi ispod).
Vratite se u glavni izbornik bez brisanja zapisa (pritisnite <ESC>).
Otvorite datoteku "c2". Uočite da je izgled zapisa istovjetan prethodnom, osim
što je membranski kapacitet dvostruko veći. Otvorite "experiment" i ponovite ista
mjerenja kao u "c1".
Konačna vrijednost napona=______mV
Izmjerite napon nakon latencije jedne vremenske konstante (npr. 10ms) nakon
početka injiciranja struje.
Napon=_________mV
Izračunajte koliki je to postotak konačne vrijednosti potencijala?
=________%
Vratite se u glavni izbornik bez brisanja zapisa (pritisnite <ESC>).
63

Temelji neuroznanosti Vodič kroz vježbe
Otvorite datoteku "c3". Uočite da je membranski kapacitet ponovno vraćen na
početne vrijednosti, dok je otpor membrane dvostruko veći. Otvorite "experiment" i
ponovite ista mjerenja kao u "c2".
Konačna vrijednost napona=______mV
Izmjerite napon nakon latencije jedne vremenske konstante (npr. 10ms) nakon
početka injiciranja struje.
Napon=_________mV
Izračunajte koliki je to postotak konačne vrijednosti potencijala?
=________%
Zašto je vrijednost konačnog napona različita?
Što se događa s vremenskom konstantom?
Vratite se u glavni izbornik bez brisanja zapisa (pritisnite <ESC>).
Pronađite i otvorite datoteku "c4". Membranski kapacitet i membranski otpor
ponovno su vraćeni na početne vrijednosti, dok se promijenio promjer aksona.
64
Temelji neuroznanosti Vodič kroz vježbe
Otvorite datoteku "c3". Uočite da je membranski kapacitet ponovno vraćen na
početne vrijednosti, dok je otpor membrane dvostruko veći. Otvorite "experiment" i
ponovite ista mjerenja kao u "c2".
Konačna vrijednost napona=______mV
Izmjerite napon nakon latencije jedne vremenske konstante (npr. 10ms) nakon
početka injiciranja struje.
Napon=_________mV
Izračunajte koliki je to postotak konačne vrijednosti potencijala?
=________%
Zašto je vrijednost konačnog napona različita?
Što se događa s vremenskom konstantom?
Vratite se u glavni izbornik bez brisanja zapisa (pritisnite <ESC>).
Pronađite i otvorite datoteku "c4". Membranski kapacitet i membranski otpor
ponovno su vraćeni na početne vrijednosti, dok se promijenio promjer aksona.
64

Temelji neuroznanosti Vodič kroz vježbe
Kakve posljedice ima promjena promjera aksona u tom pokusu?
Vratite se u glavni izbornik bez brisanja zapisa (pritisnite <ESC>).
Otvorite datoteku "c5". Svi parametri osim otpora aksoplazme istovjetni su
prethodnim.
Kakve posljedice ima promjena otpora aksoplazme na ovaj pokus?
Pritisnite "c" za brisanje svih zapisa.
7.4 Prostorna konstanta
Otvorite datoteku "c6". U prikazanom pokusu tri su elektrode postavljene na
različitim udaljenostima od mjesta na kojemu je položena elektroda za injiciranje
struje. Otvorite "experiment".
Izmjerite konačne vrijednosti potencijala kod tri elektrode
E1=______mV
E2=______mV
E3=______mV Mijenjajte položaj elektroda za snimanje i odaberite barem još pet različitih udaljenosti od
elektrode za snimanje i nacrtajte grafikon odnosa konačnih vrijednosti potencijala (os y, mV) i
udaljenosti (os x, mm).
65
Temelji neuroznanosti Vodič kroz vježbe
Kakve posljedice ima promjena promjera aksona u tom pokusu?
Vratite se u glavni izbornik bez brisanja zapisa (pritisnite <ESC>).
Otvorite datoteku "c5". Svi parametri osim otpora aksoplazme istovjetni su
prethodnim.
Kakve posljedice ima promjena otpora aksoplazme na ovaj pokus?
Pritisnite "c" za brisanje svih zapisa.
7.4 Prostorna konstanta
Otvorite datoteku "c6". U prikazanom pokusu tri su elektrode postavljene na
različitim udaljenostima od mjesta na kojemu je položena elektroda za injiciranje
struje. Otvorite "experiment".
Izmjerite konačne vrijednosti potencijala kod tri elektrode
E1=______mV
E2=______mV
E3=______mV Mijenjajte položaj elektroda za snimanje i odaberite barem još pet različitih udaljenosti od
elektrode za snimanje i nacrtajte grafikon odnosa konačnih vrijednosti potencijala (os y, mV) i
udaljenosti (os x, mm).
65

Temelji neuroznanosti Vodič kroz vježbe
Na kojoj udaljenosti dolazi do smanjenja elektronički vođene promjene
membranskog potencijala na 37% početne vrijednosti?
Usporedite dobiveni rezultat s prostornom konstantom ponuđenom u glavnom
izborniku.
Promijenite vrijednosti pojedinačnih parametara građe aksona kao u
prethodnoj vježbi i proučite kakve učinke to ima na prostornu konstantu:
dvostruko veći kapacitet membrane:
dvostruko veći otpor membrane:
66
Temelji neuroznanosti Vodič kroz vježbe
Na kojoj udaljenosti dolazi do smanjenja elektronički vođene promjene
membranskog potencijala na 37% početne vrijednosti?
Usporedite dobiveni rezultat s prostornom konstantom ponuđenom u glavnom
izborniku.
Promijenite vrijednosti pojedinačnih parametara građe aksona kao u
prethodnoj vježbi i proučite kakve učinke to ima na prostornu konstantu:
dvostruko veći kapacitet membrane:
dvostruko veći otpor membrane:
66

Temelji neuroznanosti Vodič kroz vježbe
promjena promjera aksona:
promjena otpora aksoplazme:
Otvorite datoteku "c7". Razlika između tog i prethodnog pokusa je u tome što
su kod ovog ljestvice proširene, a vremenski zapis brži. Uočite da pomicanjem
elektroda duž aksona, tj. dalje od mjesta injiciranja strujnog podražaja, dolazi do sve
većeg kašnjenja u promjeni potencijala. Iako teorijski dolazi do promjene potencijala
kod svih elektroda postavljenih dalje od mjesta injiciranja struje, oblik krivulje
mijenja se tako da ona postaje gotovo izravnata sigmoidna krivulja, to više što je
elektroda postavljena dalje od mjesta injiciranja struje.
67
Temelji neuroznanosti Vodič kroz vježbe
promjena promjera aksona:
promjena otpora aksoplazme:
Otvorite datoteku "c7". Razlika između tog i prethodnog pokusa je u tome što
su kod ovog ljestvice proširene, a vremenski zapis brži. Uočite da pomicanjem
elektroda duž aksona, tj. dalje od mjesta injiciranja strujnog podražaja, dolazi do sve
većeg kašnjenja u promjeni potencijala. Iako teorijski dolazi do promjene potencijala
kod svih elektroda postavljenih dalje od mjesta injiciranja struje, oblik krivulje
mijenja se tako da ona postaje gotovo izravnata sigmoidna krivulja, to više što je
elektroda postavljena dalje od mjesta injiciranja struje.
67

Temelji neuroznanosti Vodič kroz vježbe
8. Sinaptička signalizacija
8.1 Uvod
U ovoj vježbi putem Interneta učit ćete o sinaptičkoj signalizaciji, o sintezi
neurotransmitera, njihovom otpuštanju u sinaptičku pukotinu, o ulozi i načinu rada
postsinaptičkih receptora te o inaktivaciji neurotransmitera. Multimedia
Neuroscience Education Project financiran je od strane National Science
Foundation (NSF) da bi se unaprijedila primjena tehnologije u neuroznanstvenoj
edukaciji. Dvodimenzionalni prikazi postaju nedostatni u prikazivanju kompleksnih
fenomena kao što je sinaptička signalizacija. U vježbi ćete sažeti i utvrditi već
postojeća znanja uz pomoć trodimenzijskih animacija na adresi:
http://www.williams.edu/imput/synapse/pages/index.html.
Kliknite na enter the site i nakon što pročitate uvodni tekst odaberite prvo od
četiri ponuđena poglavlja (I Synthesis and Storage of Neurotransmitters). Proučite
sintezu malih molekula neurotranmitera (A) odnosno sintezu neuropeptida (B)
pomažući se ponuđenim trodimenzijskim animacijama i odgovorite na pitanja
postavljena na kraju vježbe.
Vratite se na uvodnu stranicu odabirom Introduction ili Return to
Introduction, pa potom odaberite drugo poglavlje (II Neurotransmitter Release).
Proučite ciklus sinaptičkih mjehurića (A Vesicles) te odgovorite na pitanja
postavljena na kraju vježbe.
Opet se vratite na Introduction i odaberite treće poglavlje (III
Neurotransmitter Postsynaptic Receptors) te proučite karakteristike ionotropnih (A) i
metabotropnih (B) receptora koristeći ponuđene animacijama. Na kraju vježbe
odgovorite na postavljena pitanja.
Odaberite i četvrto poglavlje (IV Inactivation of Neurotransmitters), proučite
enzimatsku inaktivaciju (A) te “reuptake” inaktivaciju (B) uz pomoć animacija, pa
odgovorite na postavljena pitanja.
68
Temelji neuroznanosti Vodič kroz vježbe
8. Sinaptička signalizacija
8.1 Uvod
U ovoj vježbi putem Interneta učit ćete o sinaptičkoj signalizaciji, o sintezi
neurotransmitera, njihovom otpuštanju u sinaptičku pukotinu, o ulozi i načinu rada
postsinaptičkih receptora te o inaktivaciji neurotransmitera. Multimedia
Neuroscience Education Project financiran je od strane National Science
Foundation (NSF) da bi se unaprijedila primjena tehnologije u neuroznanstvenoj
edukaciji. Dvodimenzionalni prikazi postaju nedostatni u prikazivanju kompleksnih
fenomena kao što je sinaptička signalizacija. U vježbi ćete sažeti i utvrditi već
postojeća znanja uz pomoć trodimenzijskih animacija na adresi:
http://www.williams.edu/imput/synapse/pages/index.html.
Kliknite na enter the site i nakon što pročitate uvodni tekst odaberite prvo od
četiri ponuđena poglavlja (I Synthesis and Storage of Neurotransmitters). Proučite
sintezu malih molekula neurotranmitera (A) odnosno sintezu neuropeptida (B)
pomažući se ponuđenim trodimenzijskim animacijama i odgovorite na pitanja
postavljena na kraju vježbe.
Vratite se na uvodnu stranicu odabirom Introduction ili Return to
Introduction, pa potom odaberite drugo poglavlje (II Neurotransmitter Release).
Proučite ciklus sinaptičkih mjehurića (A Vesicles) te odgovorite na pitanja
postavljena na kraju vježbe.
Opet se vratite na Introduction i odaberite treće poglavlje (III
Neurotransmitter Postsynaptic Receptors) te proučite karakteristike ionotropnih (A) i
metabotropnih (B) receptora koristeći ponuđene animacijama. Na kraju vježbe
odgovorite na postavljena pitanja.
Odaberite i četvrto poglavlje (IV Inactivation of Neurotransmitters), proučite
enzimatsku inaktivaciju (A) te “reuptake” inaktivaciju (B) uz pomoć animacija, pa
odgovorite na postavljena pitanja.
68

Temelji neuroznanosti Vodič kroz vježbe
8.2 Multimedia Neuroscience Education Project1
Synaptic Transmission: A Four Step Proces
I.Synthesis and Storage of Neurotransmitters
A. Synthesis of small molecule neurotransmitters
Acetylcholine: a representative small molecule neurotransmitter
Uptake of choline, a precursor of acetylcholine
Choline acetyltransferase: the binding of choline and acetate
Storage of acetylcholine in vesicles
Clinical Application: Acetylcholine and Alzheimer's Disease
B. Synthesis of neuropeptide neurotransmitters
Endorphins: a representative family of neuropeptide neurotransmitters
Translation of genomic message to endorphin prepropeptide
Packaging, transport and final processing of the endorphin propeptides
Arrival of the synthesized peptide at the terminal
Clinical Application: Endorphins and Autism
II. Neurotransmitter Release
A. Vesicles
Calcium influx: initation of neurotransmitter release
Mobilization of synaptic vesicles
Role of specialized proteins
Vesicle docking, fusion, and exocytosis
Recycling of synaptic vesicles
III. Neurotransmitter Postsynaptic Receptors
A. Ionotropic (ligand-gated) receptors
1 © Williams College Neuroscience, 1998
69
Temelji neuroznanosti Vodič kroz vježbe
8.2 Multimedia Neuroscience Education Project1
Synaptic Transmission: A Four Step Proces
I.Synthesis and Storage of Neurotransmitters
A. Synthesis of small molecule neurotransmitters
Acetylcholine: a representative small molecule neurotransmitter
Uptake of choline, a precursor of acetylcholine
Choline acetyltransferase: the binding of choline and acetate
Storage of acetylcholine in vesicles
Clinical Application: Acetylcholine and Alzheimer's Disease
B. Synthesis of neuropeptide neurotransmitters
Endorphins: a representative family of neuropeptide neurotransmitters
Translation of genomic message to endorphin prepropeptide
Packaging, transport and final processing of the endorphin propeptides
Arrival of the synthesized peptide at the terminal
Clinical Application: Endorphins and Autism
II. Neurotransmitter Release
A. Vesicles
Calcium influx: initation of neurotransmitter release
Mobilization of synaptic vesicles
Role of specialized proteins
Vesicle docking, fusion, and exocytosis
Recycling of synaptic vesicles
III. Neurotransmitter Postsynaptic Receptors
A. Ionotropic (ligand-gated) receptors
1 © Williams College Neuroscience, 1998
69

Temelji neuroznanosti Vodič kroz vježbe
GABAa receptors: a representative family of ligand-gated receptors
GABAa receptor function: inhibitory postsynaptic potentials
Postsynaptic GABA recognition by its receptor
Binding of GABA to receptor
Structure of the GABAa receptor
Ion channel opening: mechanism of action
Positive neuromodulation of GABAa receptors: tranquilizers
Negative neuromodualtion of GABAa receptors: anxiogenics
Clinical Application: GABA and Anxiety
B. Metabotropic (G-protein coupled) receptors
Dopamine Receptors: a representative family of metabotropic
receptors
Dopamine D1 receptor
Function of the G-protein
G-proteins, cAMP, and ion channel opening
Dopamine receptor blockade: antipsychotic drugs
Clinical Application: Dopamine and Schizophrenia
IV. Inactivation of Neurotransmitters
A. Enzymatic inactivation of neurotransmitters
Acetylcholinesterase: a representative inactivation enzyme
Location, structure and function of acetylcholinesterase
Enzyme inhibition: anticholinesterases
Clinical Application: Acetylcholine, Nerve Gas and Myasthenia Gravis
B. Presynaptic transporters: inactivation of neurotransmitters by reuptake
Serotonin transporter: a representative reuptake pump
Location and structure of the serotonin transporter
Mechanism of action of the serotonin transporter
Transporter inhibition: selective serotonin reuptake inhibitors
Clinical Application: Serotonin and Depression
70
Temelji neuroznanosti Vodič kroz vježbe
GABAa receptors: a representative family of ligand-gated receptors
GABAa receptor function: inhibitory postsynaptic potentials
Postsynaptic GABA recognition by its receptor
Binding of GABA to receptor
Structure of the GABAa receptor
Ion channel opening: mechanism of action
Positive neuromodulation of GABAa receptors: tranquilizers
Negative neuromodualtion of GABAa receptors: anxiogenics
Clinical Application: GABA and Anxiety
B. Metabotropic (G-protein coupled) receptors
Dopamine Receptors: a representative family of metabotropic
receptors
Dopamine D1 receptor
Function of the G-protein
G-proteins, cAMP, and ion channel opening
Dopamine receptor blockade: antipsychotic drugs
Clinical Application: Dopamine and Schizophrenia
IV. Inactivation of Neurotransmitters
A. Enzymatic inactivation of neurotransmitters
Acetylcholinesterase: a representative inactivation enzyme
Location, structure and function of acetylcholinesterase
Enzyme inhibition: anticholinesterases
Clinical Application: Acetylcholine, Nerve Gas and Myasthenia Gravis
B. Presynaptic transporters: inactivation of neurotransmitters by reuptake
Serotonin transporter: a representative reuptake pump
Location and structure of the serotonin transporter
Mechanism of action of the serotonin transporter
Transporter inhibition: selective serotonin reuptake inhibitors
Clinical Application: Serotonin and Depression
70

Temelji neuroznanosti Vodič kroz vježbe
INTRODUCTION
The nervous system is comprised of billions of specialized cells called
neurons. Efficient communication between these cells is crucial to the normal
functioning of the central and peripheral nervous systems. In this section we will
investigate the way in which the unique morphology and biochemistry of neurons
makes such communication possible.
The cell body, or soma, of a neuron is like that of any other cell, containing
mitochondria, ribosomes, a nucleus, and other essential organelles. Extending from
the cell membrane, however, is a system of dendritic branches which serve as receptor
sites for information sent from other neurons. If the dendrites receive a strong enough
signal from a neighboring nerve cell, or from several neighboring nerve cells, the
resting electrical potential of the receptor cell's membrane becomes depolarized.
Regenerating itself, this electrical signal travels down the cell's axon, a specialized
extension from the cell body which ranges from a few hundred micrometers in some
nerve cells, to over a meter in length in others. This wave of depolarization along the
axon is called an action potential. Most axons are covered by myelin, a fatty substance
that serves as an insulator and thus greatly enhances the speed of an action potential.
In between each sheath of myelin is an exposed portion of the axon called a node of
Ranvier. It is in these uninsulated areas that the actual flow of ions along the axon
takes place.
The end of the axon branches off into several terminals. Each axon terminal is
highly specialized to pass along action potentials to adjacent neurons, or target tissue,
in the neural pathway. Some cells communicate this information via electrical
synapses. In such cases, the action potential simply travels from one cell to the next
through specialized channels, called gap junctions, which connect the two cells.
Most cells, however, communicate via chemical synapses. Such cells are
separated by a space called a synaptic cleft and thus cannot transmit action potentials
directly. Instead, chemicals called neurotransmitters are used to communicate the
signal from one cell to the next. Some neurotransmitters are excitatory and depolarize
the next cell, increasing the probability that an action potential will be fired. Others
are inhibitory, causing the membrane of the next cell to hyperpolarize, thus decreasing
the probability of that the next neuron will fire an action potential.
71
Temelji neuroznanosti Vodič kroz vježbe
INTRODUCTION
The nervous system is comprised of billions of specialized cells called
neurons. Efficient communication between these cells is crucial to the normal
functioning of the central and peripheral nervous systems. In this section we will
investigate the way in which the unique morphology and biochemistry of neurons
makes such communication possible.
The cell body, or soma, of a neuron is like that of any other cell, containing
mitochondria, ribosomes, a nucleus, and other essential organelles. Extending from
the cell membrane, however, is a system of dendritic branches which serve as receptor
sites for information sent from other neurons. If the dendrites receive a strong enough
signal from a neighboring nerve cell, or from several neighboring nerve cells, the
resting electrical potential of the receptor cell's membrane becomes depolarized.
Regenerating itself, this electrical signal travels down the cell's axon, a specialized
extension from the cell body which ranges from a few hundred micrometers in some
nerve cells, to over a meter in length in others. This wave of depolarization along the
axon is called an action potential. Most axons are covered by myelin, a fatty substance
that serves as an insulator and thus greatly enhances the speed of an action potential.
In between each sheath of myelin is an exposed portion of the axon called a node of
Ranvier. It is in these uninsulated areas that the actual flow of ions along the axon
takes place.
The end of the axon branches off into several terminals. Each axon terminal is
highly specialized to pass along action potentials to adjacent neurons, or target tissue,
in the neural pathway. Some cells communicate this information via electrical
synapses. In such cases, the action potential simply travels from one cell to the next
through specialized channels, called gap junctions, which connect the two cells.
Most cells, however, communicate via chemical synapses. Such cells are
separated by a space called a synaptic cleft and thus cannot transmit action potentials
directly. Instead, chemicals called neurotransmitters are used to communicate the
signal from one cell to the next. Some neurotransmitters are excitatory and depolarize
the next cell, increasing the probability that an action potential will be fired. Others
are inhibitory, causing the membrane of the next cell to hyperpolarize, thus decreasing
the probability of that the next neuron will fire an action potential.
71

Temelji neuroznanosti Vodič kroz vježbe
The process by which this information is communicated is called synaptic
transmission and can be broken down into four steps. First, the neurotransmitter must
be synthesized and stored in vesicles so that when an action potential arrives at the
nerve ending, the cell is ready to pass it along to the next neuron. Next, when an
action potential does arrive at the terminal, the neurotransmitter must be quickly and
efficiently released from the terminal and into the synaptic cleft. The neurotransmitter
must then be recognized by selective receptors on the postsynaptic cell so that it can
pass along the signal and initiate another action potential. Or, in some cases, the
receptors act to block the signals of other neurons also connecting to that postsynaptic
neuron. After its recognition by the receptor, the neurotransmitter must be inactivated
so that it does not continually occupy the receptor sites of the postsynaptic cell.
Inactivation of the neurotransmitter avoids constant stimulation of the postsynaptic
cell, while at the same time freeing up the receptor sites so that they can receive
additional neurotransmitter molecules, should another action potential arrive.
Most neurotransmitters are specific for the kind of information that they are
used to convey. As a result, a certain neurotransmitter may be more highly
concentrated in one area of the brain than it is in another. In addition, the same
neurotransmitter may elicit a variety of different responses based on the type of tissue
being targeted and which other neurotransmitters, if any, are co-released. The integral
role of neurotransmitters on the normal functioning of the brain makes it clear to see
how an imbalance in any one of these chemicals could very possibly have serious
clinical implications for an individual. Whether due to genetics, drug use, the aging
process, or other various causes, biological disfunction at any of the four steps of
synaptic transmission often leads to such imbalances and is the ultimately source of
conditions such as schizophrenia, Parkinson's disease, and Alzheimer's disease. The
causes and characteristics of these conditions and others will be studied more closely
are as we focus specifically on the four steps of synaptic transmission, and trace the
actions of several important neurotransmitters.
72
Temelji neuroznanosti Vodič kroz vježbe
The process by which this information is communicated is called synaptic
transmission and can be broken down into four steps. First, the neurotransmitter must
be synthesized and stored in vesicles so that when an action potential arrives at the
nerve ending, the cell is ready to pass it along to the next neuron. Next, when an
action potential does arrive at the terminal, the neurotransmitter must be quickly and
efficiently released from the terminal and into the synaptic cleft. The neurotransmitter
must then be recognized by selective receptors on the postsynaptic cell so that it can
pass along the signal and initiate another action potential. Or, in some cases, the
receptors act to block the signals of other neurons also connecting to that postsynaptic
neuron. After its recognition by the receptor, the neurotransmitter must be inactivated
so that it does not continually occupy the receptor sites of the postsynaptic cell.
Inactivation of the neurotransmitter avoids constant stimulation of the postsynaptic
cell, while at the same time freeing up the receptor sites so that they can receive
additional neurotransmitter molecules, should another action potential arrive.
Most neurotransmitters are specific for the kind of information that they are
used to convey. As a result, a certain neurotransmitter may be more highly
concentrated in one area of the brain than it is in another. In addition, the same
neurotransmitter may elicit a variety of different responses based on the type of tissue
being targeted and which other neurotransmitters, if any, are co-released. The integral
role of neurotransmitters on the normal functioning of the brain makes it clear to see
how an imbalance in any one of these chemicals could very possibly have serious
clinical implications for an individual. Whether due to genetics, drug use, the aging
process, or other various causes, biological disfunction at any of the four steps of
synaptic transmission often leads to such imbalances and is the ultimately source of
conditions such as schizophrenia, Parkinson's disease, and Alzheimer's disease. The
causes and characteristics of these conditions and others will be studied more closely
are as we focus specifically on the four steps of synaptic transmission, and trace the
actions of several important neurotransmitters.
72

Temelji neuroznanosti Vodič kroz vježbe
I. Synthesis and Storage of Neurotransmitters
The first step in synaptic transmission is the synthesis and storage of
neurotransmitters. There are two broad categories of neurotransmitters. Small-
molecule neurotransmitters are synthesized locally within the axon terminal. Some of
the precursors necessary for the synthesis of these molecules are taken up by selective
transporters on the membrane of the terminal. Others are byproducts of cellular
processes that take place within the neuron itself and are thus readily available. The
enzymes necessary to catalyze an interaction among these precursors are usually
produced in the cell body and transported to the terminal by slow axonal transport.
Acetylcholine (ACh), is an example of an excitatory small-molecule
neurotransmitter. This important, well-studied neurotransmitter, made up of choline
and acetate, is found at various locations throughout the central and peripheral
nervous systems and at all neuromuscular junctions. The synthesis of ACh requires
the enzyme choline actyltransferase and, like all small-molecule neurotransmitters,
takes place within the nerve terminal.
Neuropeptides are the second category of neurotransmitters. These messengers
differ from small-molecule neurotransmitters in both size and in the way that they are
synthesized. Neuropeptides generally range from 3 to 36 amino acids in length, and
are thus larger than small-molecule neurotransmitters. Also, neuropeptides must made
in the cell body because their synthesis requires peptide bond formation. This process
is a great deal more involved than the simple enzymatic reactions involved in making
smaller neurotransmitters.
The synthesis of a neuropeptide is very much like the synthesis of any
secretory protein made by the cell. First, within the cell nucleus, gene transcription
takes place, during which a specific peptide-coding sequence of DNA is used as a
template to construct a corresponding strand of messenger RNA. The mRNA then
travels to a ribosome, where the process of translation begins. During translation, the
sequence of nucleotides that make up the mRNA act as a code to string together a
corresponding sequence of amino acids that will eventually become the neuropeptide
needed at the terminal. Before this molecule can be transported to the terminal for
release into the synaptic cleft, it must be processed in the endoplasmic reticulum (ER),
73
Temelji neuroznanosti Vodič kroz vježbe
I. Synthesis and Storage of Neurotransmitters
The first step in synaptic transmission is the synthesis and storage of
neurotransmitters. There are two broad categories of neurotransmitters. Small-
molecule neurotransmitters are synthesized locally within the axon terminal. Some of
the precursors necessary for the synthesis of these molecules are taken up by selective
transporters on the membrane of the terminal. Others are byproducts of cellular
processes that take place within the neuron itself and are thus readily available. The
enzymes necessary to catalyze an interaction among these precursors are usually
produced in the cell body and transported to the terminal by slow axonal transport.
Acetylcholine (ACh), is an example of an excitatory small-molecule
neurotransmitter. This important, well-studied neurotransmitter, made up of choline
and acetate, is found at various locations throughout the central and peripheral
nervous systems and at all neuromuscular junctions. The synthesis of ACh requires
the enzyme choline actyltransferase and, like all small-molecule neurotransmitters,
takes place within the nerve terminal.
Neuropeptides are the second category of neurotransmitters. These messengers
differ from small-molecule neurotransmitters in both size and in the way that they are
synthesized. Neuropeptides generally range from 3 to 36 amino acids in length, and
are thus larger than small-molecule neurotransmitters. Also, neuropeptides must made
in the cell body because their synthesis requires peptide bond formation. This process
is a great deal more involved than the simple enzymatic reactions involved in making
smaller neurotransmitters.
The synthesis of a neuropeptide is very much like the synthesis of any
secretory protein made by the cell. First, within the cell nucleus, gene transcription
takes place, during which a specific peptide-coding sequence of DNA is used as a
template to construct a corresponding strand of messenger RNA. The mRNA then
travels to a ribosome, where the process of translation begins. During translation, the
sequence of nucleotides that make up the mRNA act as a code to string together a
corresponding sequence of amino acids that will eventually become the neuropeptide
needed at the terminal. Before this molecule can be transported to the terminal for
release into the synaptic cleft, it must be processed in the endoplasmic reticulum (ER),
73

Temelji neuroznanosti Vodič kroz vježbe
packaged in the golgi apparatus, and transported in storage vesicles down the axon to
the terminal.
The endogenous opioids, a large family of neuropeptides that act as natural
analgesics, provide a good example of how post-translational processing of just one
precursor molecule can result in a whole spectrum of different, but related,
neurotransmitters. Selective cleaving and splicing of each just three precursor
molecules results in the production of the various opioids included in this family of
neurotransmitters.
Once they are synthesized, neurotransmitters, both small molecules and
neuropeptides, are stored in vesicles within the axon terminal until an action potential
arrives and they are released. Most small-molecule neurotransmitters are stored in
small vesicles that range from 40 to 60 nm in diameter and, in electron micrographs,
appear to have clear centers. The vesicles that store neuropeptides are larger, ranging
from 90 to 250 nm in diameter. These vesicles appear dark and electron-dense in
electron micrographs. For a table of small molecule neurotransmitters and their site of
synthesis, go to http://web.indstate.edu/thcme/mwking/nerves.html#table
A. Synthesis of small molecule neurotransmitters
IA1. Acetylcholine: A Representative Small Molecule
Neurotransmitter This is a side view of a human brain, facing left. The lights are showing you a
few of the areas that use the neurotransmitter acetylcholine. Let's look at one neuron
close up. Here's a neuron that starts in the septal nucleus of the hypothalamus and
sends its axon to the hippocampus. Let's follow the action potential down the axon to
the terminal to see how acetylcholine is synthesized.
Acetylcholine (ACh), the first neurotransmitter ever to be identified, is a
small- molecule excitatory neurotransmitter with a wide variety of known functions.
In the sympathetic and parasympathetic nervous systems and at all neuromuscular
junctions, ACh is used to signal muscle movement. Within the central nervous
system, cholinergic cells (neurons that use ACh as a neurotransmitter) are found in
several different locations of the brain, including the striatal complex, the basal
forebrain, the diencephalon, pontomesencephalic cell groups, and the medulla.
Depending on which area of the brain it is found in, ACh may be involved in any one
74
Temelji neuroznanosti Vodič kroz vježbe
packaged in the golgi apparatus, and transported in storage vesicles down the axon to
the terminal.
The endogenous opioids, a large family of neuropeptides that act as natural
analgesics, provide a good example of how post-translational processing of just one
precursor molecule can result in a whole spectrum of different, but related,
neurotransmitters. Selective cleaving and splicing of each just three precursor
molecules results in the production of the various opioids included in this family of
neurotransmitters.
Once they are synthesized, neurotransmitters, both small molecules and
neuropeptides, are stored in vesicles within the axon terminal until an action potential
arrives and they are released. Most small-molecule neurotransmitters are stored in
small vesicles that range from 40 to 60 nm in diameter and, in electron micrographs,
appear to have clear centers. The vesicles that store neuropeptides are larger, ranging
from 90 to 250 nm in diameter. These vesicles appear dark and electron-dense in
electron micrographs. For a table of small molecule neurotransmitters and their site of
synthesis, go to http://web.indstate.edu/thcme/mwking/nerves.html#table
A. Synthesis of small molecule neurotransmitters
IA1. Acetylcholine: A Representative Small Molecule
Neurotransmitter This is a side view of a human brain, facing left. The lights are showing you a
few of the areas that use the neurotransmitter acetylcholine. Let's look at one neuron
close up. Here's a neuron that starts in the septal nucleus of the hypothalamus and
sends its axon to the hippocampus. Let's follow the action potential down the axon to
the terminal to see how acetylcholine is synthesized.
Acetylcholine (ACh), the first neurotransmitter ever to be identified, is a
small- molecule excitatory neurotransmitter with a wide variety of known functions.
In the sympathetic and parasympathetic nervous systems and at all neuromuscular
junctions, ACh is used to signal muscle movement. Within the central nervous
system, cholinergic cells (neurons that use ACh as a neurotransmitter) are found in
several different locations of the brain, including the striatal complex, the basal
forebrain, the diencephalon, pontomesencephalic cell groups, and the medulla.
Depending on which area of the brain it is found in, ACh may be involved in any one
74

Temelji neuroznanosti Vodič kroz vježbe
of several different functions. Some of these functions include the conduction of pain,
the regulation of neuroendocrine function, the regulation of REM sleep cycles, and
the process of learning and memory formation.
Within the basal forebrain, it is the cholinergic cells of the septal nuclei that
play a large role in learning and memory. These neurons send major projections to the
hippocampus, a structure particularly important for the normal formation of
declarative memories. Several noteworthy clinical cases have shown that lesions in
this area of the brain alone can greatly impair an individual's ability to form new
declarative memories. It is this cholinergic system, among others, that has been shown
to suffer serious neurodegeneration in Alzheimer's disease, a condition characterized
by a significant failure of memory and other cognitive functions.
IA2. Uptake of Choline, a Precursor of Acetylcholine Here we see a section of the terminal bouton. Choline is transported into the
cell by a transporter protein in the membrane of the presynaptic neuron where it will
eventually combine with acetyl CoA to synthesize the neurotransmitter acetylcholine.
IA3. Choline Acetyltransferase: The Binding of Choline and
Acetate This animation shows the synthesis of the neurotransmitter acetylcholine.
Choline approaches from the left, while Cofactor A approaches from the top right
carrying acetate. Both bind to the much larger enzyme choline acetyltransferase. The
enzyme facilitates the transfer of acetate to the choline, producing the
neurotransmitter acetylcholine. After completing the reaction, the enzyme releases the
neurotransmitter and the Cofactor A carrier molecule.
The acetate required for the synthesis of ACh comes from Acetyl CoenzymeA.
Coenzyme A (CoA) forms a covalent bond with the acyl group in acetate and thus
serves as a carrier of the acetate. Acetyl CoA is synthesized in the mitochondria of all
cells as a by-product of the pyruvate dehydrogenase complex. Much of the Acetyl
CoA produced by this process is used to generate ATP in the Krebs cycle. However,
in cholinergic cells, some of this acetyl CoA is transported across the mitochondrial
membrane into the cytoplasm for use in the synthesis of ACh.
75
Temelji neuroznanosti Vodič kroz vježbe
of several different functions. Some of these functions include the conduction of pain,
the regulation of neuroendocrine function, the regulation of REM sleep cycles, and
the process of learning and memory formation.
Within the basal forebrain, it is the cholinergic cells of the septal nuclei that
play a large role in learning and memory. These neurons send major projections to the
hippocampus, a structure particularly important for the normal formation of
declarative memories. Several noteworthy clinical cases have shown that lesions in
this area of the brain alone can greatly impair an individual's ability to form new
declarative memories. It is this cholinergic system, among others, that has been shown
to suffer serious neurodegeneration in Alzheimer's disease, a condition characterized
by a significant failure of memory and other cognitive functions.
IA2. Uptake of Choline, a Precursor of Acetylcholine Here we see a section of the terminal bouton. Choline is transported into the
cell by a transporter protein in the membrane of the presynaptic neuron where it will
eventually combine with acetyl CoA to synthesize the neurotransmitter acetylcholine.
IA3. Choline Acetyltransferase: The Binding of Choline and
Acetate This animation shows the synthesis of the neurotransmitter acetylcholine.
Choline approaches from the left, while Cofactor A approaches from the top right
carrying acetate. Both bind to the much larger enzyme choline acetyltransferase. The
enzyme facilitates the transfer of acetate to the choline, producing the
neurotransmitter acetylcholine. After completing the reaction, the enzyme releases the
neurotransmitter and the Cofactor A carrier molecule.
The acetate required for the synthesis of ACh comes from Acetyl CoenzymeA.
Coenzyme A (CoA) forms a covalent bond with the acyl group in acetate and thus
serves as a carrier of the acetate. Acetyl CoA is synthesized in the mitochondria of all
cells as a by-product of the pyruvate dehydrogenase complex. Much of the Acetyl
CoA produced by this process is used to generate ATP in the Krebs cycle. However,
in cholinergic cells, some of this acetyl CoA is transported across the mitochondrial
membrane into the cytoplasm for use in the synthesis of ACh.
75

Temelji neuroznanosti Vodič kroz vježbe
Coenzyme A remains bonded to the acyl group of acetate until it reacts with
choline to form ACh. This simple, one-step reaction requires the enzyme choline
acetyltransferase (CAT). CAT is synthesized in the rough endoplasmic reticulum of
all cholinergic cells and then transported by slow axoplasmic transport to the axon
terminal, where it is readily available for the synthesis of ACh.
It is thought that acetylcholine synthesis begins with CAT first forming a
complex with acetyl CoA. Next, choline binds to an adjacent site on the enzyme.
After both precursors are bound, choline is transferred to acetyl CoA and the two
molecules form a complex. CAT and coenzyme A are then released, and what remains
is acetylcholine.
IA4. Storage of Acetylcholine in Vesicles Following its synthesis, the neurotransmitter acetylcholine travels to a vesicle,
where a transporter protein actively pumps the choline into the vesicle. Acetylcholine
is stored until the neurotransmitter needs to be released.
After its synthesis, acetylcholine (ACh) is packaged into vesicles. ACh is
transported from the cytoplasm into individual vesicles by means of a carrier protein
on the vesicle membrane called the vesicular ACh transporter (VAChT). VAChT is an
antiporter that couples an influx of ACh with an efflux of protons (H+). Once
packaged in a vesicle, acetylcholine is stored at the nerve ending until an action
potential arrives and allows for its release into the synaptic cleft.
IA5. Clinical Application: Acetylcholine and Alzheimer's Disease
The acetylcholine has been synthesized and stored in a vesicle. We zoom out
to see other vesicles inside the terminal bouton, one of the many endings of an axon.
These neurons form the septal-hippocampal pathway, a pathway that is important for
learning and memory.
During the course of the normal aging process, concentrations of ACh tend to
decrease, resulting in the sporadic lapses of short-term memory that many elderly
individuals tend to experience from time to time. This normal, non-debilitating
decline in memory, referred to as Age-Associated Memory Impairment, or benign
senescent forgetfulness, should not be confused with Alzheimer's Disease, a serious
brain disorder, in which levels of ACh can drop by up to 90 percent. The gradual
76
Temelji neuroznanosti Vodič kroz vježbe
Coenzyme A remains bonded to the acyl group of acetate until it reacts with
choline to form ACh. This simple, one-step reaction requires the enzyme choline
acetyltransferase (CAT). CAT is synthesized in the rough endoplasmic reticulum of
all cholinergic cells and then transported by slow axoplasmic transport to the axon
terminal, where it is readily available for the synthesis of ACh.
It is thought that acetylcholine synthesis begins with CAT first forming a
complex with acetyl CoA. Next, choline binds to an adjacent site on the enzyme.
After both precursors are bound, choline is transferred to acetyl CoA and the two
molecules form a complex. CAT and coenzyme A are then released, and what remains
is acetylcholine.
IA4. Storage of Acetylcholine in Vesicles Following its synthesis, the neurotransmitter acetylcholine travels to a vesicle,
where a transporter protein actively pumps the choline into the vesicle. Acetylcholine
is stored until the neurotransmitter needs to be released.
After its synthesis, acetylcholine (ACh) is packaged into vesicles. ACh is
transported from the cytoplasm into individual vesicles by means of a carrier protein
on the vesicle membrane called the vesicular ACh transporter (VAChT). VAChT is an
antiporter that couples an influx of ACh with an efflux of protons (H+). Once
packaged in a vesicle, acetylcholine is stored at the nerve ending until an action
potential arrives and allows for its release into the synaptic cleft.
IA5. Clinical Application: Acetylcholine and Alzheimer's Disease
The acetylcholine has been synthesized and stored in a vesicle. We zoom out
to see other vesicles inside the terminal bouton, one of the many endings of an axon.
These neurons form the septal-hippocampal pathway, a pathway that is important for
learning and memory.
During the course of the normal aging process, concentrations of ACh tend to
decrease, resulting in the sporadic lapses of short-term memory that many elderly
individuals tend to experience from time to time. This normal, non-debilitating
decline in memory, referred to as Age-Associated Memory Impairment, or benign
senescent forgetfulness, should not be confused with Alzheimer's Disease, a serious
brain disorder, in which levels of ACh can drop by up to 90 percent. The gradual
76

Temelji neuroznanosti Vodič kroz vježbe
death of cholinergic brain cells results in a progressive and significant loss of
cognitive and behavioral function.
Research in the last two decades has revealed that Alzheimer's Disease begins
in the entorhinal cortex and proceeds to the hippocampus. As hippocampal cells lose
their connection to other neurons and die, short-term memory falters and, as a result,
individuals may become easily confused. Once competent individuals may lose the
ability to remember simple facts like where they live, what year it is, or what they had
for breakfast. Next, neurons in the cerebral cortex begin to degenerate, leading to a
greater difficulty with language and judgement. Appetite may decline and individuals
may also begin to lose control over bladder and bowel function. As the disease
progresses even further, many patients cannot recognize even close members of their
family and become unable to perform simple tasks such as bathing or eating. Many
suffer from anxiety, suspiciousness, and may wander or pace. The disease finally
leads to a complete loss of mental function and an inability to communicate in any
way.
Significant loss of cholinergic neurons is not the only biological explanation
for this devastating condition. Autopsies of those who have died from Alzheimer's
reveal an abnormal presence of intracellular neurofibrillary tangles and extracellular
beta amyloid patches, called neuritic plaques, in the cholinergic pathways of the brain.
It is thought that neurofibrillary tangles are the result of a mutated protein normally
involved in the healthy functioning of microtubules. In affected cells, these tangles
interfere with axonal transport of enzymes, nutrients, and other materials essential to
the cell. The exact role of neuritic plaques in the progression of Alzheimer's disease
is, however, somewhat less clear. Excess concentrations of beta amyloid are
associated with neuron death, yet the mechanism by which this occurs is still not
known.
Though there is no cure for Alzheimer's, there are two drugs presently on the
market which can help to ease some of the symptoms of the disease. Both of these
drugs work to inhibit acetylcholinesterase, the enzyme which inactivates ACh at the
synapse. Inhibiting this enzyme prevents the normal breakdown of ACh and is a way
of compensating for the lowered concentrations of ACh characteristic of Alzheimer's
disease. Tetrohydroaminoacridine (THA), marketed in 1993 under the name Tacrine
or Cognex, was the first Alzheimer's drug to be approved by the U.S. FDA and has
77
Temelji neuroznanosti Vodič kroz vježbe
death of cholinergic brain cells results in a progressive and significant loss of
cognitive and behavioral function.
Research in the last two decades has revealed that Alzheimer's Disease begins
in the entorhinal cortex and proceeds to the hippocampus. As hippocampal cells lose
their connection to other neurons and die, short-term memory falters and, as a result,
individuals may become easily confused. Once competent individuals may lose the
ability to remember simple facts like where they live, what year it is, or what they had
for breakfast. Next, neurons in the cerebral cortex begin to degenerate, leading to a
greater difficulty with language and judgement. Appetite may decline and individuals
may also begin to lose control over bladder and bowel function. As the disease
progresses even further, many patients cannot recognize even close members of their
family and become unable to perform simple tasks such as bathing or eating. Many
suffer from anxiety, suspiciousness, and may wander or pace. The disease finally
leads to a complete loss of mental function and an inability to communicate in any
way.
Significant loss of cholinergic neurons is not the only biological explanation
for this devastating condition. Autopsies of those who have died from Alzheimer's
reveal an abnormal presence of intracellular neurofibrillary tangles and extracellular
beta amyloid patches, called neuritic plaques, in the cholinergic pathways of the brain.
It is thought that neurofibrillary tangles are the result of a mutated protein normally
involved in the healthy functioning of microtubules. In affected cells, these tangles
interfere with axonal transport of enzymes, nutrients, and other materials essential to
the cell. The exact role of neuritic plaques in the progression of Alzheimer's disease
is, however, somewhat less clear. Excess concentrations of beta amyloid are
associated with neuron death, yet the mechanism by which this occurs is still not
known.
Though there is no cure for Alzheimer's, there are two drugs presently on the
market which can help to ease some of the symptoms of the disease. Both of these
drugs work to inhibit acetylcholinesterase, the enzyme which inactivates ACh at the
synapse. Inhibiting this enzyme prevents the normal breakdown of ACh and is a way
of compensating for the lowered concentrations of ACh characteristic of Alzheimer's
disease. Tetrohydroaminoacridine (THA), marketed in 1993 under the name Tacrine
or Cognex, was the first Alzheimer's drug to be approved by the U.S. FDA and has
77

Temelji neuroznanosti Vodič kroz vježbe
been shown to improve memory and language deficits in early stages of the disease.
The drug is less effective in later stages when the total number of cholinergic neurons
has been reduced even further. THA is not helpful in alleviating the paranoia,
hallucinations, and aggressive behavior that many patients exhibit and has many side
effects including nausea, liver dysfunction, vomiting, diarrhea, anorexia, and
dizziness. The wide range of side effects produced by THA are due to a lack of the
drug's substrate specificity. Though it can successfully target and prevent the
breakdown of ACh in cholinergic pathways, it also reacts with additional, unrelated
chemicals in the body, thus producing the undesired symptoms. A second drug, one
with more substrate specificity, was approved by the FDA in 1996. Donepezil,
marketed under the name Aricept, targets only ACh in the brain and allows for the
same improvements in cognitive function as THA, yet without as many side effects.
Neither of these drugs, however, is a cure and neither can slow the progression
of the disease. The difficulty in finding a cure for Alzheimer's Disease reflects the
immense complexity of the condition. A deficit in ACh is only one of the many
factors contributing to this devastating brain disorder. To learn more about
Alzheimer's a helpful list of informative links can be reached at
http://www.alzweb.org.
B. Synthesis of neuropeptide neurotransmitters
IB1. Endorphins: A Representative Family of Neuropeptide
Neurotransmitter Many areas of the brain use endorphin neuropeptides for neurrotansmitters.
The endogenous opioids are a large family of neuropeptides widely distributed
throughout the central and peripheral nervous systems. These neurotransmitters are
crucial to the normal functioning of important processes such as motor coordination,
learning and memory, gastrointestinal function, the control of seizures, and the
hormonal regulation of the reproductive system, yet they are most often recognized
for their central role in the modulation of pain. Opiate pain-killers such as Morphine
and Demorol, as well as narcotic street drugs like Heroin, act at the receptors for these
neuropeptide neurotransmitters. In fact, opium derivatives were used for centuries in
the treatment of pain before the mechanism behind their analgesic effect was ever
understood. Once it was discovered that these chemicals work at specific receptor
78
Temelji neuroznanosti Vodič kroz vježbe
been shown to improve memory and language deficits in early stages of the disease.
The drug is less effective in later stages when the total number of cholinergic neurons
has been reduced even further. THA is not helpful in alleviating the paranoia,
hallucinations, and aggressive behavior that many patients exhibit and has many side
effects including nausea, liver dysfunction, vomiting, diarrhea, anorexia, and
dizziness. The wide range of side effects produced by THA are due to a lack of the
drug's substrate specificity. Though it can successfully target and prevent the
breakdown of ACh in cholinergic pathways, it also reacts with additional, unrelated
chemicals in the body, thus producing the undesired symptoms. A second drug, one
with more substrate specificity, was approved by the FDA in 1996. Donepezil,
marketed under the name Aricept, targets only ACh in the brain and allows for the
same improvements in cognitive function as THA, yet without as many side effects.
Neither of these drugs, however, is a cure and neither can slow the progression
of the disease. The difficulty in finding a cure for Alzheimer's Disease reflects the
immense complexity of the condition. A deficit in ACh is only one of the many
factors contributing to this devastating brain disorder. To learn more about
Alzheimer's a helpful list of informative links can be reached at
http://www.alzweb.org.
B. Synthesis of neuropeptide neurotransmitters
IB1. Endorphins: A Representative Family of Neuropeptide
Neurotransmitter Many areas of the brain use endorphin neuropeptides for neurrotansmitters.
The endogenous opioids are a large family of neuropeptides widely distributed
throughout the central and peripheral nervous systems. These neurotransmitters are
crucial to the normal functioning of important processes such as motor coordination,
learning and memory, gastrointestinal function, the control of seizures, and the
hormonal regulation of the reproductive system, yet they are most often recognized
for their central role in the modulation of pain. Opiate pain-killers such as Morphine
and Demorol, as well as narcotic street drugs like Heroin, act at the receptors for these
neuropeptide neurotransmitters. In fact, opium derivatives were used for centuries in
the treatment of pain before the mechanism behind their analgesic effect was ever
understood. Once it was discovered that these chemicals work at specific receptor
78

Temelji neuroznanosti Vodič kroz vježbe
sites in the body, it was proposed that our bodies must naturally produce chemicals
that are similar in nature and action. The endogenous opioids were subsequently
isolated and named to reflect the amazing ability of our bodies to produce its own
painkillers. These neuropeptides are often referred to as endorphins, endo- derived
from "endogenous," and -orphin derived from the suffix common in the names of
many opiates.
The endorphins consist of three subfamilies, each derived from a separate
precursor molecule. Each precursor molecule, called a "pre-propeptide," is coded for
by a separate gene and thus a separate molecule of mRNA. Each of these precursors is
made up of a chain of amino acids that can be selectively cleaved and spliced by
specific enzymes to synthesize any number of several related neuropeptides.
Proenkephalin (PENK), one of the three endogenous opioid precursors, consists of
267 amino acids and is cleaved at several loci to produce Leu-enkephalin, Met-
enkephalin, Peptides E, F, and B, in addition to several other peptides. The most
abundant of the three families of endorphins, these neuropeptides are found in many
different areas of both the central and peripheral nervous systems. They are most
concentrated, however, in the medium spiny neurons of the striatum, the ventromedial
nucleus of the hypothalamus, and the granule cells of the dentate gyrus, within the
hippocampus.
Prodynorphin (PDYN), another one of the three endogenous opioid precursors,
is a chain of 254 amino acids and can be cleaved to produce Leu-enkephalin, and
several Leu-enkephalin-containing peptides, including dynorphinA, dynorphin B, and
alpha- and beta-neoendorphin. For the most part, the distribution of these
neuropeptides closely parallels that of the neuropeptides derived from PENK. Levels
of PDYN mRNA are, however, particularly concentrated in the magnocellular cells of
the hypothalamus and in the granule cell layer of the dentate gyrus, within the
hippocampal formation.
Proopiomelanocortin (POMC) is made up of 267 amino acids which can be
cleaved to produce beta-lipoprotein and beta-endorphin, in addition to several non-
opioid neuropeptides, including ACTH, alpha-MSH, beta-MSH, and gamma-MSH.
POMC neuropeptides are found in the basal ganglia, cortex, amygdala, anterior and
intermediate lobes of the pituitary, the arcuate nucleus of the hypothalamus, and the
nucleus tractus solitarius in the medulla.
79
Temelji neuroznanosti Vodič kroz vježbe
sites in the body, it was proposed that our bodies must naturally produce chemicals
that are similar in nature and action. The endogenous opioids were subsequently
isolated and named to reflect the amazing ability of our bodies to produce its own
painkillers. These neuropeptides are often referred to as endorphins, endo- derived
from "endogenous," and -orphin derived from the suffix common in the names of
many opiates.
The endorphins consist of three subfamilies, each derived from a separate
precursor molecule. Each precursor molecule, called a "pre-propeptide," is coded for
by a separate gene and thus a separate molecule of mRNA. Each of these precursors is
made up of a chain of amino acids that can be selectively cleaved and spliced by
specific enzymes to synthesize any number of several related neuropeptides.
Proenkephalin (PENK), one of the three endogenous opioid precursors, consists of
267 amino acids and is cleaved at several loci to produce Leu-enkephalin, Met-
enkephalin, Peptides E, F, and B, in addition to several other peptides. The most
abundant of the three families of endorphins, these neuropeptides are found in many
different areas of both the central and peripheral nervous systems. They are most
concentrated, however, in the medium spiny neurons of the striatum, the ventromedial
nucleus of the hypothalamus, and the granule cells of the dentate gyrus, within the
hippocampus.
Prodynorphin (PDYN), another one of the three endogenous opioid precursors,
is a chain of 254 amino acids and can be cleaved to produce Leu-enkephalin, and
several Leu-enkephalin-containing peptides, including dynorphinA, dynorphin B, and
alpha- and beta-neoendorphin. For the most part, the distribution of these
neuropeptides closely parallels that of the neuropeptides derived from PENK. Levels
of PDYN mRNA are, however, particularly concentrated in the magnocellular cells of
the hypothalamus and in the granule cell layer of the dentate gyrus, within the
hippocampal formation.
Proopiomelanocortin (POMC) is made up of 267 amino acids which can be
cleaved to produce beta-lipoprotein and beta-endorphin, in addition to several non-
opioid neuropeptides, including ACTH, alpha-MSH, beta-MSH, and gamma-MSH.
POMC neuropeptides are found in the basal ganglia, cortex, amygdala, anterior and
intermediate lobes of the pituitary, the arcuate nucleus of the hypothalamus, and the
nucleus tractus solitarius in the medulla.
79

Temelji neuroznanosti Vodič kroz vježbe
IB2. Translation of Genomic Message to Endorphin
Prepropeptide Synthesis of neuropeptides starts with the transcription of the DNA code by
messenger RNA. The messenger RNA then binds with ribosomes to create a long
chain of amino acids called a propetide. The propeptide, with the enzyme protease,
enters the Golgi apparatus where they are both packaged into a vesicle to be sent
down the axon. The processing of neuropeptide precursors, which takes place within
the endoplasmic reticulum, Golgi apparatus, and transporting vesicles, makes it
possible for neurons to synthesize a large number of endogenous opioids from just
three different pe ptide chains. Such post-translational processing also makes it
possible for cells to efficiently react to changing environmental conditions. Because
much of the post-translational processing is regulated by feedback mechanisms, the
concentration of a pa rticular neurotransmitter can be altered simply, by splicing one
of the three precursor peptides at different signal sites instead of altering the level of
mRNA that is constructed during transcription in the nucleus.
Gene transcription in the nucleus results in a newly formed protein-coding
strand of mRNA. This mRNA molecule travels to the cytoplasm, where translation
begins on a free-floating ribosome. In the case of neuropeptides, translation starts with
th e construction of a "signal sequence" at the N-terminus of the peptide. This short
sequence of hydrophobic amino acids directs the mRNA, the attached ribosome, and
the "nascent" peptide chain to the membrane of the endoplasmic reticulum (ER).
The translational complex binds to the endoplasmic reticulum (ER)
membrane, translation continues, and the growing chain of polypeptides is "fed" into
the lumen of the ER. When transcription is complete, the result is a string of amino
acids contained completely within the ER lumen and anchored to the membrane by
the signal sequence.
Processing of the pre-propeptide in the endoplasmic reticulum (ER) begins
with the cleavage of the signal sequence by an enzyme called a signal peptidase.
Peptidases are enzymes which cleave designated sections of a propeptide by
hydrolyzing the peptide bonds that hold amino acids together. Specific sequences
along the chain of amino acids serve as signals to direct the enzymes where to cleave.
The ER contains numerous peptidases, each type recognizing different signals and
cleaving at different locations, resulting in the specificity necessary to synthesize the
80
Temelji neuroznanosti Vodič kroz vježbe
IB2. Translation of Genomic Message to Endorphin
Prepropeptide Synthesis of neuropeptides starts with the transcription of the DNA code by
messenger RNA. The messenger RNA then binds with ribosomes to create a long
chain of amino acids called a propetide. The propeptide, with the enzyme protease,
enters the Golgi apparatus where they are both packaged into a vesicle to be sent
down the axon. The processing of neuropeptide precursors, which takes place within
the endoplasmic reticulum, Golgi apparatus, and transporting vesicles, makes it
possible for neurons to synthesize a large number of endogenous opioids from just
three different pe ptide chains. Such post-translational processing also makes it
possible for cells to efficiently react to changing environmental conditions. Because
much of the post-translational processing is regulated by feedback mechanisms, the
concentration of a pa rticular neurotransmitter can be altered simply, by splicing one
of the three precursor peptides at different signal sites instead of altering the level of
mRNA that is constructed during transcription in the nucleus.
Gene transcription in the nucleus results in a newly formed protein-coding
strand of mRNA. This mRNA molecule travels to the cytoplasm, where translation
begins on a free-floating ribosome. In the case of neuropeptides, translation starts with
th e construction of a "signal sequence" at the N-terminus of the peptide. This short
sequence of hydrophobic amino acids directs the mRNA, the attached ribosome, and
the "nascent" peptide chain to the membrane of the endoplasmic reticulum (ER).
The translational complex binds to the endoplasmic reticulum (ER)
membrane, translation continues, and the growing chain of polypeptides is "fed" into
the lumen of the ER. When transcription is complete, the result is a string of amino
acids contained completely within the ER lumen and anchored to the membrane by
the signal sequence.
Processing of the pre-propeptide in the endoplasmic reticulum (ER) begins
with the cleavage of the signal sequence by an enzyme called a signal peptidase.
Peptidases are enzymes which cleave designated sections of a propeptide by
hydrolyzing the peptide bonds that hold amino acids together. Specific sequences
along the chain of amino acids serve as signals to direct the enzymes where to cleave.
The ER contains numerous peptidases, each type recognizing different signals and
cleaving at different locations, resulting in the specificity necessary to synthesize the
80

Temelji neuroznanosti Vodič kroz vježbe
wide range of neurotransmitters that may be derived from just one propeptide (in this
case, the synthesis of one of several possible endogenous opioids from one of three
precursor peptides). Which enzymes are present in a particular neuron dictates which
neuropeptides will be synthesized and ultimately depends upon the genes that are
expressed by the cell, the changing demands of the cell, and which area of the brain
the cell is located in.
After processing in the Endoplasmic Reticulum (ER), a sequence of amino
acids on the propeptide directs the molecule to the Golgi apparatus, where it is further
processed by peptidases, packaged into a vesicle along with a pool of peptidases, and
released for transport down the axon.
IB3. Packaging, Transport and Final Processing of the Endorphin
Propeptides The vesicle is actively transported down the axon along microtubules, seen in
yellow, getting passed from one kinesin protein to the next. Meanwhile, inside the
vesicle, enzymes called proteases, which were packaged into the vesicle with the
propeptide, cleave the longer propeptide chain into several smaller pieces. These
small chains, shown here in dark green, are the active neurotransmitters. The extra
yellow pieces are inactive. Leaving the cell body, the vesicle contains both chains of
amino acids and proteases which cleave the peptide chain, creating a vesicle filled
with both active neurotransmitter and excess amino acid chains. Fast anterograde
transport is the means by which materials produced in the cell body are carried,
enclosed by vesicles, down the axon to the terminals of the nerve cell. This process is
dependent upon a system of microtubules running lengthwise along the interior of the
axon. These microtubules provide a track by which the direction of vesicle movement
is guided. The actual movement of vesicles down the axon is facilitated by a second
protein called kinesin. Kinesin is a rod-shaped ATP-ase which forms cross-bridges
between individual vesicles and microtubules. At one end of the kinesin molecule are
two globular heads which bind to the microtubule. The other end is characterized by a
fan-shaped tail which binds to proteins on the vesicular membrane. A hinge along the
middle of the rod allows the kinesin molecule to bend and pass the vesicle on to other
kinesin molecules further down the axon. The vesicle thus slides along the length of
81
Temelji neuroznanosti Vodič kroz vježbe
wide range of neurotransmitters that may be derived from just one propeptide (in this
case, the synthesis of one of several possible endogenous opioids from one of three
precursor peptides). Which enzymes are present in a particular neuron dictates which
neuropeptides will be synthesized and ultimately depends upon the genes that are
expressed by the cell, the changing demands of the cell, and which area of the brain
the cell is located in.
After processing in the Endoplasmic Reticulum (ER), a sequence of amino
acids on the propeptide directs the molecule to the Golgi apparatus, where it is further
processed by peptidases, packaged into a vesicle along with a pool of peptidases, and
released for transport down the axon.
IB3. Packaging, Transport and Final Processing of the Endorphin
Propeptides The vesicle is actively transported down the axon along microtubules, seen in
yellow, getting passed from one kinesin protein to the next. Meanwhile, inside the
vesicle, enzymes called proteases, which were packaged into the vesicle with the
propeptide, cleave the longer propeptide chain into several smaller pieces. These
small chains, shown here in dark green, are the active neurotransmitters. The extra
yellow pieces are inactive. Leaving the cell body, the vesicle contains both chains of
amino acids and proteases which cleave the peptide chain, creating a vesicle filled
with both active neurotransmitter and excess amino acid chains. Fast anterograde
transport is the means by which materials produced in the cell body are carried,
enclosed by vesicles, down the axon to the terminals of the nerve cell. This process is
dependent upon a system of microtubules running lengthwise along the interior of the
axon. These microtubules provide a track by which the direction of vesicle movement
is guided. The actual movement of vesicles down the axon is facilitated by a second
protein called kinesin. Kinesin is a rod-shaped ATP-ase which forms cross-bridges
between individual vesicles and microtubules. At one end of the kinesin molecule are
two globular heads which bind to the microtubule. The other end is characterized by a
fan-shaped tail which binds to proteins on the vesicular membrane. A hinge along the
middle of the rod allows the kinesin molecule to bend and pass the vesicle on to other
kinesin molecules further down the axon. The vesicle thus slides along the length of
81

Temelji neuroznanosti Vodič kroz vježbe
the axon in a stop-and-go (saltatory) fashion, from kinesin to kinesin, until it reaches
the terminal.
The majority of neuropeptide-processing takes place within the vesicle as it
travels down the axon. This is where PENK, for example, may be cleaved to produce
four copies of Met-enkephalin (ME), a single copy of Leu-enkephalin (LE), and
single copies of the extended MEs, ME-Arg-Gly-Leu and ME-Arg-Phe. PENK may
also be cleaved into larger ME- and LE-containing fragments to produce Peptides E,
F, and B. ME and LE share similar structures. Both are relatively simple peptides
made up of five amino acids (pentapeptides). The chemical structure of LE is H-
tyrosine-glycine-glycine-phenylalanine-leucine-OH, while the structure for ME is the
same, except for the last amino acid: H-tyrosine-glycine-glycine-phenylalanine-
methionine-OH. The larger LE- or ME-containing peptides derived from PENK
contain within their structure one of these two base chains in addition to any number
of adjacent amino acids that make up the propeptide chain.
IB4. Arrival of the Synthesized Peptide at the Terminal The vesicle, with the small active neuropeptides inside, comes to the end of
the axon and binds to the vesicle cage in the terminal.
Vesicles containing neuropeptide neurotransmitters are referred to as large
dense-core vesicles, as compared to the small clear-core vesicles containing small
molecule neurotransmitters. This refers to the larger amount of material inside the
dense-core vesicles, which contain not only neurotransmitters, but also proteases and
other peptide chains that have been cleaved from the active neurotransmitter. Large
dense core vesicles also tend to be stored, in cages in the bouton, farther from the
synapse than the small clear-core vesicles; this means that the release of neuropeptide
neurotransmitters generally has a longer delay than the release of small molecule
neurotransmitters. When it arrives at the terminal, the necessary neurotransmitter has
been fully synthesized and processed and is ready for release.
The vesicles are held in place by the interaction of synapsin and actin
molecules. Synapsin I, found on the membrane of the pre-synaptic vesicles, binds to
actin filaments, microtubules, spectrin, and other elements of the cytoskeleton to hold
the vesicle in a "cage," awaiting an action potential. For a more in depth report on the
82
Temelji neuroznanosti Vodič kroz vježbe
the axon in a stop-and-go (saltatory) fashion, from kinesin to kinesin, until it reaches
the terminal.
The majority of neuropeptide-processing takes place within the vesicle as it
travels down the axon. This is where PENK, for example, may be cleaved to produce
four copies of Met-enkephalin (ME), a single copy of Leu-enkephalin (LE), and
single copies of the extended MEs, ME-Arg-Gly-Leu and ME-Arg-Phe. PENK may
also be cleaved into larger ME- and LE-containing fragments to produce Peptides E,
F, and B. ME and LE share similar structures. Both are relatively simple peptides
made up of five amino acids (pentapeptides). The chemical structure of LE is H-
tyrosine-glycine-glycine-phenylalanine-leucine-OH, while the structure for ME is the
same, except for the last amino acid: H-tyrosine-glycine-glycine-phenylalanine-
methionine-OH. The larger LE- or ME-containing peptides derived from PENK
contain within their structure one of these two base chains in addition to any number
of adjacent amino acids that make up the propeptide chain.
IB4. Arrival of the Synthesized Peptide at the Terminal The vesicle, with the small active neuropeptides inside, comes to the end of
the axon and binds to the vesicle cage in the terminal.
Vesicles containing neuropeptide neurotransmitters are referred to as large
dense-core vesicles, as compared to the small clear-core vesicles containing small
molecule neurotransmitters. This refers to the larger amount of material inside the
dense-core vesicles, which contain not only neurotransmitters, but also proteases and
other peptide chains that have been cleaved from the active neurotransmitter. Large
dense core vesicles also tend to be stored, in cages in the bouton, farther from the
synapse than the small clear-core vesicles; this means that the release of neuropeptide
neurotransmitters generally has a longer delay than the release of small molecule
neurotransmitters. When it arrives at the terminal, the necessary neurotransmitter has
been fully synthesized and processed and is ready for release.
The vesicles are held in place by the interaction of synapsin and actin
molecules. Synapsin I, found on the membrane of the pre-synaptic vesicles, binds to
actin filaments, microtubules, spectrin, and other elements of the cytoskeleton to hold
the vesicle in a "cage," awaiting an action potential. For a more in depth report on the
82

Temelji neuroznanosti Vodič kroz vježbe
energy requirements and kinetics of synapsin's binding to synaptic vesicles, go to
http://physiology.cup.cam.ac.uk/JPhysiol/1997/504p3/7005/7005
IB5. Clinical Applications: Endorphins and Autism The vesicles, filled with active neurotransmitter and other material, remain in
the bouton awaiting an action potential. In times of stress and injury, endorphin levels
increase and their release allows us, and other animals, to ignore pain. Such a
mechanism is adaptive, allowing us to focus our energy on productive behaviors that
will help us to escape potentially harmful situations. Thus, if you are bitten by a snake
in the woods, you may not feel the pain as you run away at top speed from the
dangerous animal.
Endogenous opioids, through their analgesic qualities, are also thought to
mediate the motivation that many animals exhibit for forming social attachments. In
the absence of pain, the release of endorphins can result in feelings of well-being and
sensations of pleasure. Accepting the fact that social attachments tend to be adaptive
(i.e. lions are more likely to find food if they hunt as a pride, children are more likely
to survive if they are taken care of by their mother, etc.), it is not unreasonable to
hypothesize that a reward mechanism, in the form of endorphin release, exists to
promote such adaptive behaviors. Thus, we seek social attachments and interactions to
increase the natural release of endorphins, which, in turn motivates us to continue to
seek such attachments.
The idea that the formation of social attachments is mediated by endogenous
opiates is supported by the fact that, in most cases, social isolation results in a state of
distress. When juvenile guinea pigs are removed from their mothers and isolated from
all social interaction, their level of distress can be quantified by measuring distress
vocalizations (i.e. crying). If separation distress is indeed mediated by a lack (or
lowered level) of endogenous opioids, then administering an opiate agonist, a
substance that mimics the action of the endogenous opioids, should alleviate such
distress. This is just what has been shown. When juvenile guinea pigs are separated
from their mothers and isolated from any social contact, those treated with Morphine,
an opiate agonist, show a markedly lower level of distress vocalizations than those in
a control condition. (Herman and Panksepp, 1977).
83
Temelji neuroznanosti Vodič kroz vježbe
energy requirements and kinetics of synapsin's binding to synaptic vesicles, go to
http://physiology.cup.cam.ac.uk/JPhysiol/1997/504p3/7005/7005
IB5. Clinical Applications: Endorphins and Autism The vesicles, filled with active neurotransmitter and other material, remain in
the bouton awaiting an action potential. In times of stress and injury, endorphin levels
increase and their release allows us, and other animals, to ignore pain. Such a
mechanism is adaptive, allowing us to focus our energy on productive behaviors that
will help us to escape potentially harmful situations. Thus, if you are bitten by a snake
in the woods, you may not feel the pain as you run away at top speed from the
dangerous animal.
Endogenous opioids, through their analgesic qualities, are also thought to
mediate the motivation that many animals exhibit for forming social attachments. In
the absence of pain, the release of endorphins can result in feelings of well-being and
sensations of pleasure. Accepting the fact that social attachments tend to be adaptive
(i.e. lions are more likely to find food if they hunt as a pride, children are more likely
to survive if they are taken care of by their mother, etc.), it is not unreasonable to
hypothesize that a reward mechanism, in the form of endorphin release, exists to
promote such adaptive behaviors. Thus, we seek social attachments and interactions to
increase the natural release of endorphins, which, in turn motivates us to continue to
seek such attachments.
The idea that the formation of social attachments is mediated by endogenous
opiates is supported by the fact that, in most cases, social isolation results in a state of
distress. When juvenile guinea pigs are removed from their mothers and isolated from
all social interaction, their level of distress can be quantified by measuring distress
vocalizations (i.e. crying). If separation distress is indeed mediated by a lack (or
lowered level) of endogenous opioids, then administering an opiate agonist, a
substance that mimics the action of the endogenous opioids, should alleviate such
distress. This is just what has been shown. When juvenile guinea pigs are separated
from their mothers and isolated from any social contact, those treated with Morphine,
an opiate agonist, show a markedly lower level of distress vocalizations than those in
a control condition. (Herman and Panksepp, 1977).
83

Temelji neuroznanosti Vodič kroz vježbe
Administering opiate agonists not only alleviates the distress that results from
social isolation but has also been shown to reduce the need to seek out social
attachments. Measures of infant/maternal proximity-maintenance time provide an
animal model to which we can compare approach attachment behavior normally
exhibited by humans. When guinea pigs are treated with Morphine, the amount of
time they choose to spend in close proximity to their mothers is significantly lower
than juveniles not given the opiate agonist. (Herman and Panksepp, 1977). Such
findings support the idea that direct administration of opiate agonists replaces the
need to form social bonds, a process which naturally raises endorphin levels. If
artificial opiates are administered to an individual, that individual then has no
motivation to take the actions (i.e. seeking out social attachments) that would
naturally raise levels of endogenous opioids. Such individuals thus avoid these
actions.
This finding can be observed in humans who are addicted to narcotic
analgesics such as Morphine and Heroin. Individuals dependent on such drugs are
activating their opioid receptors on a regular basis, thus substituting the need for
normal social behavior. As a result, these individuals tend to be emotionally and
socially withdrawn. They also tend to exhibit a decreased interest in other endorphin-
releasing stimuli, such as food and sex. Interestingly, many of the withdrawal
symptoms exhibited by recovering heroin addicts are similar to the expressions of
grief exhibited by those who have recently suffered the loss a loved social one: crying,
depression, insomnia, irritability, and anorexia.
If individuals who artificially introduce opiates into their bodies exhibit
abnormal social and emotional behaviors, it seems possible that similar avoidance
behaviors in non-dependent individuals could signal a clinical condition in which
naturally-occurring endogenous opioids are present in abnormally high
concentrations. In fact, overactive opioid neurons are thought to be one of the
physiological explanations behind Autism, a pervasive developmental disorder
characterized by mildly to severely impaired social interaction. Autism affects two to
ten out of every 10,000 people, with males four times as likely as females to develop
the condition. It is thought that autistic children lack normal concentrations of the
enzymes necessary to break down endorphins. As a result, the brain of an autistic
individual remains saturated with opioids.
84
Temelji neuroznanosti Vodič kroz vježbe
Administering opiate agonists not only alleviates the distress that results from
social isolation but has also been shown to reduce the need to seek out social
attachments. Measures of infant/maternal proximity-maintenance time provide an
animal model to which we can compare approach attachment behavior normally
exhibited by humans. When guinea pigs are treated with Morphine, the amount of
time they choose to spend in close proximity to their mothers is significantly lower
than juveniles not given the opiate agonist. (Herman and Panksepp, 1977). Such
findings support the idea that direct administration of opiate agonists replaces the
need to form social bonds, a process which naturally raises endorphin levels. If
artificial opiates are administered to an individual, that individual then has no
motivation to take the actions (i.e. seeking out social attachments) that would
naturally raise levels of endogenous opioids. Such individuals thus avoid these
actions.
This finding can be observed in humans who are addicted to narcotic
analgesics such as Morphine and Heroin. Individuals dependent on such drugs are
activating their opioid receptors on a regular basis, thus substituting the need for
normal social behavior. As a result, these individuals tend to be emotionally and
socially withdrawn. They also tend to exhibit a decreased interest in other endorphin-
releasing stimuli, such as food and sex. Interestingly, many of the withdrawal
symptoms exhibited by recovering heroin addicts are similar to the expressions of
grief exhibited by those who have recently suffered the loss a loved social one: crying,
depression, insomnia, irritability, and anorexia.
If individuals who artificially introduce opiates into their bodies exhibit
abnormal social and emotional behaviors, it seems possible that similar avoidance
behaviors in non-dependent individuals could signal a clinical condition in which
naturally-occurring endogenous opioids are present in abnormally high
concentrations. In fact, overactive opioid neurons are thought to be one of the
physiological explanations behind Autism, a pervasive developmental disorder
characterized by mildly to severely impaired social interaction. Autism affects two to
ten out of every 10,000 people, with males four times as likely as females to develop
the condition. It is thought that autistic children lack normal concentrations of the
enzymes necessary to break down endorphins. As a result, the brain of an autistic
individual remains saturated with opioids.
84

Temelji neuroznanosti Vodič kroz vježbe
It is thought that excessive opioid activity in autistic infants may interfere with
many of the various early childhood socialization experiences. The first symptoms
usually appear during the first three years of childhood and may include a failure of
the child to respond to his or her name, a failure to respond to other people's emotions,
difficulty in interpreting tone of voice, or difficulty in interpreting the facial
expressions of others. Autistic children tend to develop speech at a later age and may
refer to themselves by name, instead of "I" or "me." Later, they seem uninterested in
developing friendships and seem unable to initiate or sustain logical conversations
with others. Many use a sing-song voice to speak on a narrow range of seemingly odd
topics, with little regard for the interest level of the other person.
In addition to demonstrating an abnormal disregard for seeking social
attachments and interactions, autism is often marked by a stubborn adherence to
specific routines and rituals. Individuals often become upset by any change in their
normal routine. Many autistics also exhibit abnormal responses to auditory, tactile,
and other sensory stimulation. Perhaps because of increased opioid activity, these
individuals often exhibit a reduced sensitivity to pain, yet a negative hyper-sensitivity
to other sensations ordinarily thought of as benign, or even pleasurable. This might
explain why certain autistic children avoid hugging or cuddling with parents.
(www.autism.org) Such individuals often engage in repetitive, "self-stimulatory," and
sometimes self-injurious behavior, such as rocking, hair-twirling, biting, and head-
banging. One theory suggests that autistics engage in this seemingly self-injurious
behavior to stimulate the pain-induced release of endogenous opiates.
If an excess of opoid activity is, in fact, responsible for causing the
characteristic behaviors associated with autism, then blocking the receptors for these
neurotransmitters should prove to be an effective treatment for this disorder.
Naltrexone is an opiate antagonist which is thought to aid in the treatment of autistic
behaviors. An antagonist is a substance which blocks the receptors for a specific
neurotransmitter. In several trials, naltrexone helped to increase socialization, eye
contact, and general happiness in autistic individuals. In addition, pain sensitivity was
normalized, and a decrease in self-injurious and other self stimulatory behaviors was
observed. (www.autism.org/naltrex.html) Not all trials, however, offer such promising
results. In some cases the administration of Naltrexone helped to significantly reduce
85
Temelji neuroznanosti Vodič kroz vježbe
It is thought that excessive opioid activity in autistic infants may interfere with
many of the various early childhood socialization experiences. The first symptoms
usually appear during the first three years of childhood and may include a failure of
the child to respond to his or her name, a failure to respond to other people's emotions,
difficulty in interpreting tone of voice, or difficulty in interpreting the facial
expressions of others. Autistic children tend to develop speech at a later age and may
refer to themselves by name, instead of "I" or "me." Later, they seem uninterested in
developing friendships and seem unable to initiate or sustain logical conversations
with others. Many use a sing-song voice to speak on a narrow range of seemingly odd
topics, with little regard for the interest level of the other person.
In addition to demonstrating an abnormal disregard for seeking social
attachments and interactions, autism is often marked by a stubborn adherence to
specific routines and rituals. Individuals often become upset by any change in their
normal routine. Many autistics also exhibit abnormal responses to auditory, tactile,
and other sensory stimulation. Perhaps because of increased opioid activity, these
individuals often exhibit a reduced sensitivity to pain, yet a negative hyper-sensitivity
to other sensations ordinarily thought of as benign, or even pleasurable. This might
explain why certain autistic children avoid hugging or cuddling with parents.
(www.autism.org) Such individuals often engage in repetitive, "self-stimulatory," and
sometimes self-injurious behavior, such as rocking, hair-twirling, biting, and head-
banging. One theory suggests that autistics engage in this seemingly self-injurious
behavior to stimulate the pain-induced release of endogenous opiates.
If an excess of opoid activity is, in fact, responsible for causing the
characteristic behaviors associated with autism, then blocking the receptors for these
neurotransmitters should prove to be an effective treatment for this disorder.
Naltrexone is an opiate antagonist which is thought to aid in the treatment of autistic
behaviors. An antagonist is a substance which blocks the receptors for a specific
neurotransmitter. In several trials, naltrexone helped to increase socialization, eye
contact, and general happiness in autistic individuals. In addition, pain sensitivity was
normalized, and a decrease in self-injurious and other self stimulatory behaviors was
observed. (www.autism.org/naltrex.html) Not all trials, however, offer such promising
results. In some cases the administration of Naltrexone helped to significantly reduce
85

Temelji neuroznanosti Vodič kroz vježbe
only some characteristic behaviors, while other behaviors remained at baseline. In
other trials there was no observable change in behavior at all.
Because autism is a disorder defined by a wide range of symptoms and
behaviors, each individual presents a unique range of characteristics and differs in his
or her level of severity. In addition, many autistics are plagued by a number of
associated neurological disorders ranging from attention deficit hyperactivity disorder
to profound mental retardation. The efficacy of Naltrexone varies from patient to
patient and seems to depend on the specific condition of the individual. As a result,
individual differences must be taken into consideration when treatment methods are
selected.
Further complicating the matter of treatment is the fact that endorphins are not
the only neurotransmitters thought to be involved in autism. Abnormal concentrations
of serotonin and dopamine are also thought contribute to the condition. As a result,
while Naltrexone may help to alleviate some of the symptoms modulated by the
endogenous opioids, this powerful antagonist does not present an ultimate answer in
the treatment of autism.
II. Neurotransmitter Release
At rest, neurotransmitter-containing vesicles are stored at the terminal of the
neuron in one of two places. A small number of vesicles are positioned along the pre-
synaptic membrane in places called "active zones." This is where neurotransmitter
release occurs. Most vesicles, however, are held close to these zones, yet further from
the membrane itself until they are needed. These vesicles are held in place by Ca2+-
sensitive vesicle membrane proteins (VAMPs), which bind to actin filaments,
microtubules, and various other elements of the cytoskeleton. When an action
potential reaches the terminal of a presynaptic neuron, voltage-dependent calcium
(Ca2+) channels embedded in the pre-synaptic membrane open and Ca2+ rushes in.
This influx of calcium ions triggers a series of events, which ultimately results in the
release of the neurotransmitter from a storage vesicle into the synaptic cleft.
The first step in this process involves freeing the neurotransmitter-containing
vesicles from the bonds that hold them to the cytoskeleton. The vesicles are then free
86
Temelji neuroznanosti Vodič kroz vježbe
only some characteristic behaviors, while other behaviors remained at baseline. In
other trials there was no observable change in behavior at all.
Because autism is a disorder defined by a wide range of symptoms and
behaviors, each individual presents a unique range of characteristics and differs in his
or her level of severity. In addition, many autistics are plagued by a number of
associated neurological disorders ranging from attention deficit hyperactivity disorder
to profound mental retardation. The efficacy of Naltrexone varies from patient to
patient and seems to depend on the specific condition of the individual. As a result,
individual differences must be taken into consideration when treatment methods are
selected.
Further complicating the matter of treatment is the fact that endorphins are not
the only neurotransmitters thought to be involved in autism. Abnormal concentrations
of serotonin and dopamine are also thought contribute to the condition. As a result,
while Naltrexone may help to alleviate some of the symptoms modulated by the
endogenous opioids, this powerful antagonist does not present an ultimate answer in
the treatment of autism.
II. Neurotransmitter Release
At rest, neurotransmitter-containing vesicles are stored at the terminal of the
neuron in one of two places. A small number of vesicles are positioned along the pre-
synaptic membrane in places called "active zones." This is where neurotransmitter
release occurs. Most vesicles, however, are held close to these zones, yet further from
the membrane itself until they are needed. These vesicles are held in place by Ca2+-
sensitive vesicle membrane proteins (VAMPs), which bind to actin filaments,
microtubules, and various other elements of the cytoskeleton. When an action
potential reaches the terminal of a presynaptic neuron, voltage-dependent calcium
(Ca2+) channels embedded in the pre-synaptic membrane open and Ca2+ rushes in.
This influx of calcium ions triggers a series of events, which ultimately results in the
release of the neurotransmitter from a storage vesicle into the synaptic cleft.
The first step in this process involves freeing the neurotransmitter-containing
vesicles from the bonds that hold them to the cytoskeleton. The vesicles are then free
86

Temelji neuroznanosti Vodič kroz vježbe
to travel to the active zones, where docking takes place. Docking is the process by
which the vesicle and the pre-synaptic membrane line up in a position that will allow
them to fuse easily. Fusion then takes place, in which the vesicle membrane and the
pre-synaptic membrane connect to form a small opening, a pore, connecting the
lumen of the vesicle with the synaptic cleft. This pore grows larger and larger until the
vesicle membrane collapses into the pre-synaptic membrane and releases its contents
into the synaptic cleft, a process called exocytosis.
Following exocytosis, the vesicular membrane, presently a continuous
extension of the pre-synaptic membrane, forms a pit and pinches off into the terminal
to form a new, vacant vesicle. This vesicle is then either recycled and refilled with
more of the neurotransmitter, or sent to the cell body, where it is broken down,
processed into a new vesicle, and transported to the terminal where it can then be
filled with the neurotransmitter.
A. Vesicles: two types, one process
IIA1. Calcium Influx: Initiation of Neurotransmitter Release
This is a side view of a human brain, facing left, showing an action potential
travelling down the axon. This action potential is initiated when the cell body has
received enough excitatory signals from other neurons. When the action potential
reaches the terminal, it activates voltage-dependent calcium channels, allowing
calcium ions to flow into theterminal.
In the resting state, neurotransmitters are stored in vesicles at the pre-synaptic
terminal. Most neurons contain at least two types of synaptic vesicles, small (about 50
nm diameter) and large (70-200 nm diameter). Neurotransmitters are released in
quantal units as each vesicle contains a given amount of transmitter. The vesicles are
concentrated near "active zones," dense bodies along the pre-synaptic membrane
where neurotransmitter release occurs. At rest, only a small number of vesicles are
actually positioned at the active zones. Rather, most are held near the active zones by
the proteins synapsin I and actin. Synapsin I, a protein associated with the vesicle
membrane, binds to actin, a cytoskeletal filament holding the vesicles in place.
Calcium (Ca2+) is a vital element in the process of neurotransmitter release;
when Ca2+ channels are blocked, neurotransmitter release is inhibited. When the
87
Temelji neuroznanosti Vodič kroz vježbe
to travel to the active zones, where docking takes place. Docking is the process by
which the vesicle and the pre-synaptic membrane line up in a position that will allow
them to fuse easily. Fusion then takes place, in which the vesicle membrane and the
pre-synaptic membrane connect to form a small opening, a pore, connecting the
lumen of the vesicle with the synaptic cleft. This pore grows larger and larger until the
vesicle membrane collapses into the pre-synaptic membrane and releases its contents
into the synaptic cleft, a process called exocytosis.
Following exocytosis, the vesicular membrane, presently a continuous
extension of the pre-synaptic membrane, forms a pit and pinches off into the terminal
to form a new, vacant vesicle. This vesicle is then either recycled and refilled with
more of the neurotransmitter, or sent to the cell body, where it is broken down,
processed into a new vesicle, and transported to the terminal where it can then be
filled with the neurotransmitter.
A. Vesicles: two types, one process
IIA1. Calcium Influx: Initiation of Neurotransmitter Release
This is a side view of a human brain, facing left, showing an action potential
travelling down the axon. This action potential is initiated when the cell body has
received enough excitatory signals from other neurons. When the action potential
reaches the terminal, it activates voltage-dependent calcium channels, allowing
calcium ions to flow into theterminal.
In the resting state, neurotransmitters are stored in vesicles at the pre-synaptic
terminal. Most neurons contain at least two types of synaptic vesicles, small (about 50
nm diameter) and large (70-200 nm diameter). Neurotransmitters are released in
quantal units as each vesicle contains a given amount of transmitter. The vesicles are
concentrated near "active zones," dense bodies along the pre-synaptic membrane
where neurotransmitter release occurs. At rest, only a small number of vesicles are
actually positioned at the active zones. Rather, most are held near the active zones by
the proteins synapsin I and actin. Synapsin I, a protein associated with the vesicle
membrane, binds to actin, a cytoskeletal filament holding the vesicles in place.
Calcium (Ca2+) is a vital element in the process of neurotransmitter release;
when Ca2+ channels are blocked, neurotransmitter release is inhibited. When the
87

Temelji neuroznanosti Vodič kroz vježbe
action potential reaches the nerve terminal, voltage-dependent Ca2+ channels open and
Ca2+ rushes into the neuron terminal due to a greater extracellular concentration.
Ca2+ channels appear to be localized near the active zones of the vesicular
membrane. In the giant squid neuron, it has been found that Ca2+ influx is ten times
greater in the area of the active zone than elsewhere in the neuron. Furthermore,
during an action potential Ca2+ concentration at the active zone can rise one-
thousandfold, from 100 nM (nano-molar concentration) to 100 micro-M within a few
hundred microseconds. The observation of intramembranous proteins thought to be
Ca2+ channels near active zones is consistent with rapid neurotransmitter release
following Ca2+ influx.
IIA2. Mobilization of synaptic vesicles
When the calcium ions enter the pre-synaptic terminal, they activate the
protein calmodulin, shown in blue, which then activates protein kinase II, seen in
aqua-white. The protein kinase then phosphoylates Synapsin, releasing the vesicle
from its actin cage. The vesicle falls to the active zone at the presynaptic terminal.
Once in the pre-synaptic terminal, calcium triggers a series of events that lead
to neurotransmitter release. The entire process, from the opening of Ca2+ channels to
neurotransmitter release, may occur quickly within 200 micro-seconds or take up to
50 milli-seconds. Time variations are most likely attributed to the location of pre-
synaptic vesicles when Ca2+ enters the neuron. For rapid release it is assumed that the
synaptic vesicles involved and the site of exocytosis are located at or very close to the
Ca2+ channels. In delayed reactions, synaptic vesicles must be recruited and guided to
the active zones before neurotransmitter release can occur.
The first step in the Ca2+ dependent process of neurotransmitter release is to
free synaptic vesicles from actin and other cytoskeletal elements. At rest, the synaptic
vesicle protein synapsin I is found in a dephosphorylated state where it binds to
elements of the cytoskeleton including actin filaments, microtubules, and spectrin as
well as exhibiting a high affinity toward synaptic vesicles. Synapsin I, by binding to
both the cytoskeleton and the vesicle, immobilizes vesicles.
Synapsin I, however, when phosphorylated, loses its binding affinity for
synaptic vesicles. Phosphorylation, a process where phosphate (PO4-) is transferred
88
Temelji neuroznanosti Vodič kroz vježbe
action potential reaches the nerve terminal, voltage-dependent Ca2+ channels open and
Ca2+ rushes into the neuron terminal due to a greater extracellular concentration.
Ca2+ channels appear to be localized near the active zones of the vesicular
membrane. In the giant squid neuron, it has been found that Ca2+ influx is ten times
greater in the area of the active zone than elsewhere in the neuron. Furthermore,
during an action potential Ca2+ concentration at the active zone can rise one-
thousandfold, from 100 nM (nano-molar concentration) to 100 micro-M within a few
hundred microseconds. The observation of intramembranous proteins thought to be
Ca2+ channels near active zones is consistent with rapid neurotransmitter release
following Ca2+ influx.
IIA2. Mobilization of synaptic vesicles
When the calcium ions enter the pre-synaptic terminal, they activate the
protein calmodulin, shown in blue, which then activates protein kinase II, seen in
aqua-white. The protein kinase then phosphoylates Synapsin, releasing the vesicle
from its actin cage. The vesicle falls to the active zone at the presynaptic terminal.
Once in the pre-synaptic terminal, calcium triggers a series of events that lead
to neurotransmitter release. The entire process, from the opening of Ca2+ channels to
neurotransmitter release, may occur quickly within 200 micro-seconds or take up to
50 milli-seconds. Time variations are most likely attributed to the location of pre-
synaptic vesicles when Ca2+ enters the neuron. For rapid release it is assumed that the
synaptic vesicles involved and the site of exocytosis are located at or very close to the
Ca2+ channels. In delayed reactions, synaptic vesicles must be recruited and guided to
the active zones before neurotransmitter release can occur.
The first step in the Ca2+ dependent process of neurotransmitter release is to
free synaptic vesicles from actin and other cytoskeletal elements. At rest, the synaptic
vesicle protein synapsin I is found in a dephosphorylated state where it binds to
elements of the cytoskeleton including actin filaments, microtubules, and spectrin as
well as exhibiting a high affinity toward synaptic vesicles. Synapsin I, by binding to
both the cytoskeleton and the vesicle, immobilizes vesicles.
Synapsin I, however, when phosphorylated, loses its binding affinity for
synaptic vesicles. Phosphorylation, a process where phosphate (PO4-) is transferred
88

Temelji neuroznanosti Vodič kroz vježbe
from adenosine triphosphate (ATP) to a protein, is a mechanism for changing the state
or shape of a protein, and thus affecting its function.
Synapsin I is phosphorylated by the Ca2+ dependent protein, Ca2+ /calmodulin-
dependent protein kinase II (CaM kinase II). With Ca2+ influx, Ca2+ /calmodulin is
formed, which then binds to CaM kinase II. In its inactive state, CaM kinase II has an
auto-inhibitory sequence which overlaps with the Ca2+ /calmodulin binding domain.
Binding of Ca2+ /calmodulin changes the conformation of the kinase, relieves the
inhibitory block, and activates the kinase. When activated, CaM kinase II
phosphorylates synapsin I. Consequently, synapsin I loses its affinity for synaptic
vesicles and the vesicles are released from the cytoskeleton. It has been found that
most synaptic vesicles within 30 nm of the terminal membrane do not have synapsin I
(Hirokawa et al 1989). Thus, it seems dephosphorylated synapsin I holds vesicles in
place at a distance from the synaptic terminal. Vesicles released by phosphorylated
synapsin I, however, move to the active zone where they are ready for exocytosis.
IIA3. Role of Specialized Proteins The vesicle, shown in green, arrives at the active zone, shown in purple.
Docking the vesicle to the presynaptic membrane involves the cooperation of many
specialized proteins. Some of these proteins are found in the wall of the vesicle, some
in the presynaptic membrane, and some in the cytoplasm between the two
membranes. Interactions between these proteins bring the vesicle very close to the
presynaptic membrane.
Specialized proteins which are involved in neurotransmitter release have been
identified on the vesicle membrane and the pre-synaptic membrane, as well as within
the cytoplasm. The many proteins are thought to be involved in keeping the vesicles
in place, sensing calcium influx, guiding the vesicles to active zones, docking vesicles
at the pre-synaptic membrane, and in allowing fusion and exocytosis to occur. The
exact role of the identified proteins continues to be a major topic in neuroscience.
VAMP's (vesicle membrane proteins) are involved in holding vesicles in
place, sensing Ca++ influx, and in the initial phases of exocytosis. The synapsin
group, a family of four proteins of which synapsin I and synapsin II are the best
studied, have been identified on the vesicle membrane and are thought to play a role
in holding vesicles in place. Rab3a and Rab3b, two members of a class of proteins
89
Temelji neuroznanosti Vodič kroz vježbe
from adenosine triphosphate (ATP) to a protein, is a mechanism for changing the state
or shape of a protein, and thus affecting its function.
Synapsin I is phosphorylated by the Ca2+ dependent protein, Ca2+ /calmodulin-
dependent protein kinase II (CaM kinase II). With Ca2+ influx, Ca2+ /calmodulin is
formed, which then binds to CaM kinase II. In its inactive state, CaM kinase II has an
auto-inhibitory sequence which overlaps with the Ca2+ /calmodulin binding domain.
Binding of Ca2+ /calmodulin changes the conformation of the kinase, relieves the
inhibitory block, and activates the kinase. When activated, CaM kinase II
phosphorylates synapsin I. Consequently, synapsin I loses its affinity for synaptic
vesicles and the vesicles are released from the cytoskeleton. It has been found that
most synaptic vesicles within 30 nm of the terminal membrane do not have synapsin I
(Hirokawa et al 1989). Thus, it seems dephosphorylated synapsin I holds vesicles in
place at a distance from the synaptic terminal. Vesicles released by phosphorylated
synapsin I, however, move to the active zone where they are ready for exocytosis.
IIA3. Role of Specialized Proteins The vesicle, shown in green, arrives at the active zone, shown in purple.
Docking the vesicle to the presynaptic membrane involves the cooperation of many
specialized proteins. Some of these proteins are found in the wall of the vesicle, some
in the presynaptic membrane, and some in the cytoplasm between the two
membranes. Interactions between these proteins bring the vesicle very close to the
presynaptic membrane.
Specialized proteins which are involved in neurotransmitter release have been
identified on the vesicle membrane and the pre-synaptic membrane, as well as within
the cytoplasm. The many proteins are thought to be involved in keeping the vesicles
in place, sensing calcium influx, guiding the vesicles to active zones, docking vesicles
at the pre-synaptic membrane, and in allowing fusion and exocytosis to occur. The
exact role of the identified proteins continues to be a major topic in neuroscience.
VAMP's (vesicle membrane proteins) are involved in holding vesicles in
place, sensing Ca++ influx, and in the initial phases of exocytosis. The synapsin
group, a family of four proteins of which synapsin I and synapsin II are the best
studied, have been identified on the vesicle membrane and are thought to play a role
in holding vesicles in place. Rab3a and Rab3b, two members of a class of proteins
89

Temelji neuroznanosti Vodič kroz vježbe
that bind GTP, are thought to be involved in guiding mobile vesicles to active zones
where docking occurs. Synaptobrevin, another VAMP, is a protein that may be
involved in recognition of the plasma membrane.
Two other well studied proteins associated with the vesicle membrane are
synaptotagmin and synaptophysin. Synaptotagmin, with a membrane spanning region,
has a Ca++ binding domain and, in the presence of Ca++, binds phospholipids. It is
thought to play a role in sensing Ca++ influx and starting the process of vesicle
fusion. Synaptophysin, of which there are about 10-20 copies per vesicle, may be
involved in forming a fusion pore with the pre-synaptic membrane.
Proteins have also been identified on the presynaptic membrane which are
necessary for neurotransmitter release. Syntaxin, a membrane spanning protein, may
be involved in recognizing the vesicle membrane. SNAP-25 (no relation to
cytoplasmic SNAP's) attaches to the plasma membrane through fatty acid groups and
acts as a SNARE (SNAP receptor protein) in vesicle docking. Another protein on the
presynaptic membrane, physophilin, may be involved in forming a fusion pore. In
vitro, physophilin and synaptophysin join to form a pore.
Finally, there are two sets of cytoplasmic proteins involved in vesicle docking.
The N-ethylmaleimide sensitive fusion protein (NSF) is one group and the second is
called the soluble NSF attachment proteins (SNAP). In the fusion of synaptic vesicles
to the pre-synaptic membrane, these general cytoplasmic proteins are thought to
intervene between two specific proteins, one on the vesicle and one of the pre-
synaptic membrane, to create a fusion pore.
IIA4. Vesicle Docking, Fusion, and Exocytosis The arrival of the action potential causes the first influx of calcium ions,
allowing the vesicle to dock at the active zone. But the fusion proteins are only
activated by a second influx of calcium ions through additional channels at the zone.
When the proteins fuse the vesicle to the presynaptic membrane, the vesicle opens and
empties its neurotransmitters into the synaptic cleft.
Docking is the process during which the vesicle and pre-synaptic membrane
line up in a fusion-ready state. Following docking, the membranes fuse to create a
small opening which grows larger until the vesicle membrane collapses into the pre-
synaptic membrane and exocytosis occurs. While the exact mechanisms behind
90
Temelji neuroznanosti Vodič kroz vježbe
that bind GTP, are thought to be involved in guiding mobile vesicles to active zones
where docking occurs. Synaptobrevin, another VAMP, is a protein that may be
involved in recognition of the plasma membrane.
Two other well studied proteins associated with the vesicle membrane are
synaptotagmin and synaptophysin. Synaptotagmin, with a membrane spanning region,
has a Ca++ binding domain and, in the presence of Ca++, binds phospholipids. It is
thought to play a role in sensing Ca++ influx and starting the process of vesicle
fusion. Synaptophysin, of which there are about 10-20 copies per vesicle, may be
involved in forming a fusion pore with the pre-synaptic membrane.
Proteins have also been identified on the presynaptic membrane which are
necessary for neurotransmitter release. Syntaxin, a membrane spanning protein, may
be involved in recognizing the vesicle membrane. SNAP-25 (no relation to
cytoplasmic SNAP's) attaches to the plasma membrane through fatty acid groups and
acts as a SNARE (SNAP receptor protein) in vesicle docking. Another protein on the
presynaptic membrane, physophilin, may be involved in forming a fusion pore. In
vitro, physophilin and synaptophysin join to form a pore.
Finally, there are two sets of cytoplasmic proteins involved in vesicle docking.
The N-ethylmaleimide sensitive fusion protein (NSF) is one group and the second is
called the soluble NSF attachment proteins (SNAP). In the fusion of synaptic vesicles
to the pre-synaptic membrane, these general cytoplasmic proteins are thought to
intervene between two specific proteins, one on the vesicle and one of the pre-
synaptic membrane, to create a fusion pore.
IIA4. Vesicle Docking, Fusion, and Exocytosis The arrival of the action potential causes the first influx of calcium ions,
allowing the vesicle to dock at the active zone. But the fusion proteins are only
activated by a second influx of calcium ions through additional channels at the zone.
When the proteins fuse the vesicle to the presynaptic membrane, the vesicle opens and
empties its neurotransmitters into the synaptic cleft.
Docking is the process during which the vesicle and pre-synaptic membrane
line up in a fusion-ready state. Following docking, the membranes fuse to create a
small opening which grows larger until the vesicle membrane collapses into the pre-
synaptic membrane and exocytosis occurs. While the exact mechanisms behind
90

Temelji neuroznanosti Vodič kroz vježbe
synaptic fusion have not been determined in full, the process is known to be calcium
dependent.
There is evidence that rab 3A and rab 3B, two G-proteins, guide synaptic
vesicles to active zones. Once at the active zone, vesicle and plasma membrane
proteins, with the assistance of cytoplasmic proteins, recognize each other and
docking occurs. The cytoplasmic proteins, NSF and SNAPs, are involved in assuring
proper vesicle targeting. NSF and SNAPs along with SNAP receptors (SNAREs) may
form an initial complex between the vesicle and presynaptic membrane. Possible
proteins on the vesicle membrane and pre-synaptic membrane that act as SNAREs
are, respectively, synaptobrevin and syntaxin. These two proteins may allow the
vesicle and presynaptic membrane to recognize each other.
Following docking, there is a second influx of calcium at the active zone,
which causes the vesicle membrane to fuse to the presynaptic membrane, forming a
temporary ion channel. This early fusion pore, similar to a gap junction, is a
cytoplasmic bridge that connects the lumen of the vesicle with the space outside the
neuronal terminal. The fusion pore is composed of two halves, one in the vesicle
membrane and one in the pre-synaptic membrane. Proteins potentially involved in
forming the fusion pore include synaptophysin on the vesicle membrane and
physophilin on the pre-synaptic membrane. When aligned, these proteins may fuse the
vesicle membrane to the pre-synaptic membrane. The formation of this pore is an
energetically unfavorable event and, thus requires energy provided when NSF
hydrolyzes ATP.
Without calcium influx, there is no fusion pore formed. There is evidence that
the vesicle protein synaptotagmin is the key calcium sensor and plays a significant
role in activating fusion. Synaptotagmin has membrane binding abilities, binds
calcium, and has been observed in close proximity to the site of calcium influx. In the
absence of calcium, synaptotagmin acts as a clamp and inhibits vesicle fusion; the
vesicle remains in a fusion-ready state but does not form a fusion pore.
Synaptotagmin acts as a calcium sensor and, with calcium influx, activates fusion.
Synaptotagmin does not appear to participate in the actual fusion event but may pull
the vesicle and pre-synaptic membrane into closer contact with each other or induce
conformational changes in fusion proteins.
91
Temelji neuroznanosti Vodič kroz vježbe
synaptic fusion have not been determined in full, the process is known to be calcium
dependent.
There is evidence that rab 3A and rab 3B, two G-proteins, guide synaptic
vesicles to active zones. Once at the active zone, vesicle and plasma membrane
proteins, with the assistance of cytoplasmic proteins, recognize each other and
docking occurs. The cytoplasmic proteins, NSF and SNAPs, are involved in assuring
proper vesicle targeting. NSF and SNAPs along with SNAP receptors (SNAREs) may
form an initial complex between the vesicle and presynaptic membrane. Possible
proteins on the vesicle membrane and pre-synaptic membrane that act as SNAREs
are, respectively, synaptobrevin and syntaxin. These two proteins may allow the
vesicle and presynaptic membrane to recognize each other.
Following docking, there is a second influx of calcium at the active zone,
which causes the vesicle membrane to fuse to the presynaptic membrane, forming a
temporary ion channel. This early fusion pore, similar to a gap junction, is a
cytoplasmic bridge that connects the lumen of the vesicle with the space outside the
neuronal terminal. The fusion pore is composed of two halves, one in the vesicle
membrane and one in the pre-synaptic membrane. Proteins potentially involved in
forming the fusion pore include synaptophysin on the vesicle membrane and
physophilin on the pre-synaptic membrane. When aligned, these proteins may fuse the
vesicle membrane to the pre-synaptic membrane. The formation of this pore is an
energetically unfavorable event and, thus requires energy provided when NSF
hydrolyzes ATP.
Without calcium influx, there is no fusion pore formed. There is evidence that
the vesicle protein synaptotagmin is the key calcium sensor and plays a significant
role in activating fusion. Synaptotagmin has membrane binding abilities, binds
calcium, and has been observed in close proximity to the site of calcium influx. In the
absence of calcium, synaptotagmin acts as a clamp and inhibits vesicle fusion; the
vesicle remains in a fusion-ready state but does not form a fusion pore.
Synaptotagmin acts as a calcium sensor and, with calcium influx, activates fusion.
Synaptotagmin does not appear to participate in the actual fusion event but may pull
the vesicle and pre-synaptic membrane into closer contact with each other or induce
conformational changes in fusion proteins.
91

Temelji neuroznanosti Vodič kroz vježbe
Once an initial pore is formed, it quickly dilates leading to exocytosis and
neurotransmitter release. The vesicle membrane joins the plasma membrane and the
contents of the vesicle are released into the synaptic cleft.
IIA5. Recycling of Synaptic Vesicles After the vesicle has fused with the cell membrane, the vesicular membrane
moves away from the active zone. It then pinches off back into the presynaptic
terminal, and travels away from the presynaptic membrane. The vesicle is eventually
absorbed into a cistern, shown in purple, to begin the process again.
In exocytosis, the membrane of the vesicle fuses with the membrane of the
synaptic terminal. Without a compensatory mechanism, the pre-synaptic membrane
would enlarge as vesicular membrane would continuously be added to the plasma
membrane. This enlargement, however, does not occur as the vesicle membrane is
retrieved from the pre-synaptic membrane and is recycled to form another synaptic
vesicle. Following exocytosis, excess membrane on the synaptic terminal forms a pit
coated with the protein clathrin. This clathrin-coated pit pinches off from the pre-
synaptic membrane and, after the clathrin is removed, re-forms a synaptic vesicle.
Not all of the retrieved membrane is directly recycled into vesicles through the
cistern. Some are returned to the cell body where it is degraded and processed into
new vesicles which are then transported to the synaptic terminal.
III. Neurotransmitter Postsynaptic Receptors After release into the synaptic cleft, neurotransmitters interact with receptor
proteins on the membrane of the postsynaptic cell, causing ionic channels on the
membrane to either open or close. When these channels open, depolarization occurs,
res ulting in the initiation of another action potential.
There are two types of postsynaptic receptors that recognize neurotransmitters.
Ionotropic receptors, also referred to as ligand-gated ion channels , act quickly to
depol arize the neuron and pass on the action potential (or hyperpolarize the neuron
and inhibit additional action potentials). These receptors are made up of five
individual protein subunits embedded in the cell membrane, and arranged to form a
single pore th at spans this membrane. When a neurotransmitter associates with the
92
Temelji neuroznanosti Vodič kroz vježbe
Once an initial pore is formed, it quickly dilates leading to exocytosis and
neurotransmitter release. The vesicle membrane joins the plasma membrane and the
contents of the vesicle are released into the synaptic cleft.
IIA5. Recycling of Synaptic Vesicles After the vesicle has fused with the cell membrane, the vesicular membrane
moves away from the active zone. It then pinches off back into the presynaptic
terminal, and travels away from the presynaptic membrane. The vesicle is eventually
absorbed into a cistern, shown in purple, to begin the process again.
In exocytosis, the membrane of the vesicle fuses with the membrane of the
synaptic terminal. Without a compensatory mechanism, the pre-synaptic membrane
would enlarge as vesicular membrane would continuously be added to the plasma
membrane. This enlargement, however, does not occur as the vesicle membrane is
retrieved from the pre-synaptic membrane and is recycled to form another synaptic
vesicle. Following exocytosis, excess membrane on the synaptic terminal forms a pit
coated with the protein clathrin. This clathrin-coated pit pinches off from the pre-
synaptic membrane and, after the clathrin is removed, re-forms a synaptic vesicle.
Not all of the retrieved membrane is directly recycled into vesicles through the
cistern. Some are returned to the cell body where it is degraded and processed into
new vesicles which are then transported to the synaptic terminal.
III. Neurotransmitter Postsynaptic Receptors After release into the synaptic cleft, neurotransmitters interact with receptor
proteins on the membrane of the postsynaptic cell, causing ionic channels on the
membrane to either open or close. When these channels open, depolarization occurs,
res ulting in the initiation of another action potential.
There are two types of postsynaptic receptors that recognize neurotransmitters.
Ionotropic receptors, also referred to as ligand-gated ion channels , act quickly to
depol arize the neuron and pass on the action potential (or hyperpolarize the neuron
and inhibit additional action potentials). These receptors are made up of five
individual protein subunits embedded in the cell membrane, and arranged to form a
single pore th at spans this membrane. When a neurotransmitter associates with the
92

Temelji neuroznanosti Vodič kroz vježbe
extracellular recognition site, the membrane-spanning subunits of the receptor quickly
open to form a pore through which the necessary ions can pass. Depolarization
usually occurs a m illisecond or two after the action potential has been received and
lasts only up to ten milliseconds.
?-aminobutyric acid(GABA) is one example of a neurotransmitter recognized
by an ionotropic receptor. GABA is an inhibitory neurotransmitter used at roughly
one-third of the synapses in the brain. The binding of GABA at the GABA
recognition site c auses the membrane-spanning channel of the receptor protein to
open and allow an influx of negatively charged chloride ions. This influx of negative
ions serves to hyperpolarize the cell thus inhibiting the firing of an action potential.
Though in the c ase of GABA, the ionotropic receptor is used to inhibit the firing of
an action potential, there are other ionotropic receptors which recognize excitatory
neurotransmitters and thus stimulate the firing of action potentials in post-synaptic
cells. < /P>
Metabotropic receptors, or G-protein linked receptors , do not work as simply
as ligand-gated ion channels do. Like ionotropic receptors, metabotropic receptors
also have an extracellular neurotransmitter recognition site, yet these receptors do not
form a membrane-spanning pore that can allow the direct passage of ions. Instead,
when a neurotransmitter associates with the extracellular recognition site, an
intermediate molecule within the postsynaptic cell, called a G-protein, is activated
and, either directly or through a series of enzymatic reactions, opens or closes ion
channels located at other places on the cell membrane. Because the action of
metabotropic recepto rs is not as direct, their action is slower. Depolarization takes
longer, typically lasting up to hundreds of milliseconds, and in some cases, going on
for several minutes, hours, or even days.
Metabotropic receptors are used in the recognition of all neuropeptides and
several small-molecule neurotransmitters. Dopamine (DA), for example, is a small-
molecule neurotransmitter recognized by a G-protein coupled receptor. The binding of
DA a t the recognition site of a post-synaptic DA receptor sets off a chain of reactions
which ultimately cause ion pores along the post-synaptic membrane to open and an
action potential to be stimulated. Found in four major tracts of the brain (the nigrostri
al tract, the tuberoinfundibular tract, the mesolimbic tract, and the mesocortical tract),
DA plays an important role in the control of both motor and emotional behavior.
93
Temelji neuroznanosti Vodič kroz vježbe
extracellular recognition site, the membrane-spanning subunits of the receptor quickly
open to form a pore through which the necessary ions can pass. Depolarization
usually occurs a m illisecond or two after the action potential has been received and
lasts only up to ten milliseconds.
?-aminobutyric acid(GABA) is one example of a neurotransmitter recognized
by an ionotropic receptor. GABA is an inhibitory neurotransmitter used at roughly
one-third of the synapses in the brain. The binding of GABA at the GABA
recognition site c auses the membrane-spanning channel of the receptor protein to
open and allow an influx of negatively charged chloride ions. This influx of negative
ions serves to hyperpolarize the cell thus inhibiting the firing of an action potential.
Though in the c ase of GABA, the ionotropic receptor is used to inhibit the firing of
an action potential, there are other ionotropic receptors which recognize excitatory
neurotransmitters and thus stimulate the firing of action potentials in post-synaptic
cells. < /P>
Metabotropic receptors, or G-protein linked receptors , do not work as simply
as ligand-gated ion channels do. Like ionotropic receptors, metabotropic receptors
also have an extracellular neurotransmitter recognition site, yet these receptors do not
form a membrane-spanning pore that can allow the direct passage of ions. Instead,
when a neurotransmitter associates with the extracellular recognition site, an
intermediate molecule within the postsynaptic cell, called a G-protein, is activated
and, either directly or through a series of enzymatic reactions, opens or closes ion
channels located at other places on the cell membrane. Because the action of
metabotropic recepto rs is not as direct, their action is slower. Depolarization takes
longer, typically lasting up to hundreds of milliseconds, and in some cases, going on
for several minutes, hours, or even days.
Metabotropic receptors are used in the recognition of all neuropeptides and
several small-molecule neurotransmitters. Dopamine (DA), for example, is a small-
molecule neurotransmitter recognized by a G-protein coupled receptor. The binding of
DA a t the recognition site of a post-synaptic DA receptor sets off a chain of reactions
which ultimately cause ion pores along the post-synaptic membrane to open and an
action potential to be stimulated. Found in four major tracts of the brain (the nigrostri
al tract, the tuberoinfundibular tract, the mesolimbic tract, and the mesocortical tract),
DA plays an important role in the control of both motor and emotional behavior.
93

Temelji neuroznanosti Vodič kroz vježbe
A. Ionotropic (ligand-gated) receptors
1. GABAa Receptors: a representative family of ligand-gated receptors
2. GABAa receptor function: inhibitory postsynaptic potentials
3. Postsynaptic GABA recognition by its receptor
4. Binding of GABA to receptor
5. Structure of the GABAa receptor
6. Ion channel opening: mechanism of action
7. Positive neuromodulation of GABAa receptors: tranquilizers
8. Negative neuromodualtion of GABAa receptors: anxiogenics
9. Clinical Application: GABA and Anxiety
B. Metabotropic (G-protein coupled) receptors
1. Dopamine Receptors: a representative family of metabotropic receptors
2. Dopamine D1 Receptor
3. Function of the G-protein
4. G-proteins, cAMP, and Ion Channel Opening
5. Dopamine Receptor Blockade: Antipsychotic Drugs
6. Clinical Application: Dopamine and Schizophrenia
A. Ionotropic (ligand-gated) receptors
IIIA1. GABAa Receptors: A Representative Family of Ligand-
Gated Receptors This is a cross-section of a human brain, facing left. Inside lies the Limbic
System. One part of the limbic system is the amygdala, shown in pink. Inside the
amygdala are many neurons that use the neurotransmitter GABA to control feelings of
fear and anxiety.
GABA (gamma-aminobutyric acid) is the primary inhibitory neurotransmitter
in the brain. Unlike excitatory neurons, which "pass along" a message to the next
neuron by causing an Excitatory Post-Synaptic Potential (EPSP), inhibitory neurons
make the next neuron less likely to fire by causing an Inhibitory Post-Synaptic
Potential (IPSP). So when the arrival of an action potential at the axon terminal of a
gabaminergic neuron causes the release of GABA into the synaptic cleft to interact
94
Temelji neuroznanosti Vodič kroz vježbe
A. Ionotropic (ligand-gated) receptors
1. GABAa Receptors: a representative family of ligand-gated receptors
2. GABAa receptor function: inhibitory postsynaptic potentials
3. Postsynaptic GABA recognition by its receptor
4. Binding of GABA to receptor
5. Structure of the GABAa receptor
6. Ion channel opening: mechanism of action
7. Positive neuromodulation of GABAa receptors: tranquilizers
8. Negative neuromodualtion of GABAa receptors: anxiogenics
9. Clinical Application: GABA and Anxiety
B. Metabotropic (G-protein coupled) receptors
1. Dopamine Receptors: a representative family of metabotropic receptors
2. Dopamine D1 Receptor
3. Function of the G-protein
4. G-proteins, cAMP, and Ion Channel Opening
5. Dopamine Receptor Blockade: Antipsychotic Drugs
6. Clinical Application: Dopamine and Schizophrenia
A. Ionotropic (ligand-gated) receptors
IIIA1. GABAa Receptors: A Representative Family of Ligand-
Gated Receptors This is a cross-section of a human brain, facing left. Inside lies the Limbic
System. One part of the limbic system is the amygdala, shown in pink. Inside the
amygdala are many neurons that use the neurotransmitter GABA to control feelings of
fear and anxiety.
GABA (gamma-aminobutyric acid) is the primary inhibitory neurotransmitter
in the brain. Unlike excitatory neurons, which "pass along" a message to the next
neuron by causing an Excitatory Post-Synaptic Potential (EPSP), inhibitory neurons
make the next neuron less likely to fire by causing an Inhibitory Post-Synaptic
Potential (IPSP). So when the arrival of an action potential at the axon terminal of a
gabaminergic neuron causes the release of GABA into the synaptic cleft to interact
94

Temelji neuroznanosti Vodič kroz vježbe
with its post-synaptic receptors, these receptors open channels for chloride ions. The
rush of these negatively-charged chloride ions into the receiving neuron makes it
more negative inside, and thus less likely to fire.
GABA is recognized by both ligand-gated and G-protein-linked receptors. The
ligand-gated GABA receptor is called GABAa and represents a typical ligand-gated
receptor molecule. High concentrations of GABA and GABAa receptors are found
in the limbic system, an area of the brain where personal feelings and emotional
memories are generated and stored. When an individual experiences a strong emotion,
or is faced with a threatening challenge, it is the limbic system that interprets the
ambiguous physiological responses of the autonomic nervous system that accompany
these situations (i.e. change in heart rate, blood pressure, breathing, perspiration,
etc.) to determine exactly which emotion is being experienced. Is it fear? Excitement?
Anxiety? Anger? By analyzing the particular situation and comparing it to any
memories associated with that or similar experiences, the limbic system helps us to
identify and name the emotions we feel.
GABA neurons are present throughout the nervous system, from the eye (see
http://retina.anatomy.upenn.edu/~lance/Neurochem/gaba.html ), to the limbic system.
The amygdala (or amygdaloid complex), located in the medial temporal lobe, is one
of the most important elements of the limbic system, playing a crucial role in emotion
and memory. More specifically, it is thought that the amygdala plays a large role in
associating certain stimuli with rewards and in thus moderating some of our
behavioral responses. Several studies have illustrated the crucial role that
the amygdala plays in modulating the connection between our emotions and
our behavior, most specifically in the areas of fear and aggression. In one experiment,
rats were conditioned to experience a fear response to a neutral stimulus. This
response, characterized by increased blood pressure and periods of "freezing"
(immobility), was abolished after lesioning of the amygdala. In a second lesioning
experiment, usually aggressive rhesus monkeys became placid and fearless
around their caretakers after the removal of their amygdalas. In humans, electrical
stimulation of the amygdala has been shown to lead to uncontrolled rage.
The amygdala connects various sensory processing regions with
the hypothalamus and brain stem. Its inputs include projections from the olfactory
cortex, somatic sensory pathways, and the auditory complex. This diverse sensory
95
Temelji neuroznanosti Vodič kroz vježbe
with its post-synaptic receptors, these receptors open channels for chloride ions. The
rush of these negatively-charged chloride ions into the receiving neuron makes it
more negative inside, and thus less likely to fire.
GABA is recognized by both ligand-gated and G-protein-linked receptors. The
ligand-gated GABA receptor is called GABAa and represents a typical ligand-gated
receptor molecule. High concentrations of GABA and GABAa receptors are found
in the limbic system, an area of the brain where personal feelings and emotional
memories are generated and stored. When an individual experiences a strong emotion,
or is faced with a threatening challenge, it is the limbic system that interprets the
ambiguous physiological responses of the autonomic nervous system that accompany
these situations (i.e. change in heart rate, blood pressure, breathing, perspiration,
etc.) to determine exactly which emotion is being experienced. Is it fear? Excitement?
Anxiety? Anger? By analyzing the particular situation and comparing it to any
memories associated with that or similar experiences, the limbic system helps us to
identify and name the emotions we feel.
GABA neurons are present throughout the nervous system, from the eye (see
http://retina.anatomy.upenn.edu/~lance/Neurochem/gaba.html ), to the limbic system.
The amygdala (or amygdaloid complex), located in the medial temporal lobe, is one
of the most important elements of the limbic system, playing a crucial role in emotion
and memory. More specifically, it is thought that the amygdala plays a large role in
associating certain stimuli with rewards and in thus moderating some of our
behavioral responses. Several studies have illustrated the crucial role that
the amygdala plays in modulating the connection between our emotions and
our behavior, most specifically in the areas of fear and aggression. In one experiment,
rats were conditioned to experience a fear response to a neutral stimulus. This
response, characterized by increased blood pressure and periods of "freezing"
(immobility), was abolished after lesioning of the amygdala. In a second lesioning
experiment, usually aggressive rhesus monkeys became placid and fearless
around their caretakers after the removal of their amygdalas. In humans, electrical
stimulation of the amygdala has been shown to lead to uncontrolled rage.
The amygdala connects various sensory processing regions with
the hypothalamus and brain stem. Its inputs include projections from the olfactory
cortex, somatic sensory pathways, and the auditory complex. This diverse sensory
95

Temelji neuroznanosti Vodič kroz vježbe
information is integrated and processed in the amygdala, where evidence of long term
potentiation (LTP), and thus associative learning, has been shown. The processed
signals are then sent through projections to the hypothalamus and brain stem. In this
way, the amygdala has the ability to control both autonomic and somatic systems.
IIIA2. GABAa Receptor Function: Inhibitory Postsynaptic
Potentials First, the output neuron on the left receives excitatory signals from two
neurons on the right, generating an action potential. Then the third neuron on the right
fires. This time, the output neuron on the left does not generate an action potential,
and so no feelings of anxiety leave the amygdala. That's because the third neuron
produced an inhibitory postsynaptic potential. Let's follow the action potential down
the axon and find out how the GABA receptor works to produce this inhibition.
The various inputs received by the amygdala often transmit conflicting signals.
It is not uncommon for excitatory and inhibitory neurons to synapse on the same
neuron. The arrival of an excitatory signal triggers a wave of depolarization along the
membrane of a post-synaptic neuron known as an excitatory post-synaptic potential
(EPSP). Inhibitory signals have an opposite effect. Such signals cause a wave
of hyperpolarization along the membrane of a post-synaptic cell known as
an inhibitory post-synaptic potential (IPSP). When they arrive at the same neuron,
these opposing signals travel along the post-synaptic membrane until they reach the
axon terminal, where they are then integrated. If enough EPSPs arrive at the axon
terminal simultaneously, the membrane potential will rise above threshold, and an
action potential will fire. The presence of IPSPs, however, will lower the membrane
potential. If enough IPSPs have fired, these inhibitory signals will stop the
neuron from firing.
In times of stress, excitatory neurons in the amygdala fire rapidly, sending
excitatory signals to other areas of the brain. It is this type of firing that leads to a
feeling of panic or fear. The inhibitory interneurons in the amygdala modulate these
emotions by releasing GABA. The release of GABA, and its subsequent recognition
by post-synaptic receptors like the GABAa receptor, inhibits the excitatory signals
that result in feelings of anxiety and fear. GABA thus has a calming, tranquilizing
effect on our emotions and prevents us from becoming overwhelmed in stressful
96
Temelji neuroznanosti Vodič kroz vježbe
information is integrated and processed in the amygdala, where evidence of long term
potentiation (LTP), and thus associative learning, has been shown. The processed
signals are then sent through projections to the hypothalamus and brain stem. In this
way, the amygdala has the ability to control both autonomic and somatic systems.
IIIA2. GABAa Receptor Function: Inhibitory Postsynaptic
Potentials First, the output neuron on the left receives excitatory signals from two
neurons on the right, generating an action potential. Then the third neuron on the right
fires. This time, the output neuron on the left does not generate an action potential,
and so no feelings of anxiety leave the amygdala. That's because the third neuron
produced an inhibitory postsynaptic potential. Let's follow the action potential down
the axon and find out how the GABA receptor works to produce this inhibition.
The various inputs received by the amygdala often transmit conflicting signals.
It is not uncommon for excitatory and inhibitory neurons to synapse on the same
neuron. The arrival of an excitatory signal triggers a wave of depolarization along the
membrane of a post-synaptic neuron known as an excitatory post-synaptic potential
(EPSP). Inhibitory signals have an opposite effect. Such signals cause a wave
of hyperpolarization along the membrane of a post-synaptic cell known as
an inhibitory post-synaptic potential (IPSP). When they arrive at the same neuron,
these opposing signals travel along the post-synaptic membrane until they reach the
axon terminal, where they are then integrated. If enough EPSPs arrive at the axon
terminal simultaneously, the membrane potential will rise above threshold, and an
action potential will fire. The presence of IPSPs, however, will lower the membrane
potential. If enough IPSPs have fired, these inhibitory signals will stop the
neuron from firing.
In times of stress, excitatory neurons in the amygdala fire rapidly, sending
excitatory signals to other areas of the brain. It is this type of firing that leads to a
feeling of panic or fear. The inhibitory interneurons in the amygdala modulate these
emotions by releasing GABA. The release of GABA, and its subsequent recognition
by post-synaptic receptors like the GABAa receptor, inhibits the excitatory signals
that result in feelings of anxiety and fear. GABA thus has a calming, tranquilizing
effect on our emotions and prevents us from becoming overwhelmed in stressful
96

Temelji neuroznanosti Vodič kroz vježbe
situations. GABA is important in maintaining a normal level of firing for all kinds of
different systems; for information about GABA and epilepsy, a disease characterized
by seizures due to uncontrollable firing of certain neurons, go to
http://www.sfn.org/briefings/epilepsy.html.
IIIA3. GABA Release into the Synapse We now zoom in to the synapse, with the presynaptic membrane on top and
the postsynaptic membrane on bottom. Notice the input neuron releasing the
neurotransmitter GABA into the synaptic cleft. Now you see the GABA receptor on
the postsynaptic side.
GABA is a small-molecule neurotransmitter synthesized within
the presynaptic terminal of GABA-containing neurons. Once synthesized, GABA is
packaged in vesicles and stored at the post-synaptic terminal until the arrival of an
action potential causes these vesicles to release their contents into the synaptic cleft.
Once released, GABA molecules travel across the synapse to the post-synaptic
membrane, where they are recognized by GABA receptors.
97
Temelji neuroznanosti Vodič kroz vježbe
situations. GABA is important in maintaining a normal level of firing for all kinds of
different systems; for information about GABA and epilepsy, a disease characterized
by seizures due to uncontrollable firing of certain neurons, go to
http://www.sfn.org/briefings/epilepsy.html.
IIIA3. GABA Release into the Synapse We now zoom in to the synapse, with the presynaptic membrane on top and
the postsynaptic membrane on bottom. Notice the input neuron releasing the
neurotransmitter GABA into the synaptic cleft. Now you see the GABA receptor on
the postsynaptic side.
GABA is a small-molecule neurotransmitter synthesized within
the presynaptic terminal of GABA-containing neurons. Once synthesized, GABA is
packaged in vesicles and stored at the post-synaptic terminal until the arrival of an
action potential causes these vesicles to release their contents into the synaptic cleft.
Once released, GABA molecules travel across the synapse to the post-synaptic
membrane, where they are recognized by GABA receptors.
97

Temelji neuroznanosti Vodič kroz vježbe
IIIA4. Binding of GABA to Receptor Receptors for GABA are embedded in the post-synaptic membrane. When two
molecules of GABA bind to its receptor, the receptor channel opens, and chloride ions
rush into the neuron.
The GABAa receptor is made up of five subunit proteins. Each subunit
transverses the postsynaptic membrane in a generally symmetrical manner. The
five subunits are arranged to form a central pore, which, when opened, allows for the
passage of chloride ions into the postsynaptic neuron. The GABAa receptor molecule
has a specific site at which molecules of GABA bind. The interactions between
neurotransmitter and receptor at this site do not involve the formation of chemical
bonds, but do result in changingthe conformation of the receptor to consequently open
up the central pore. Opening this pore allows for the influx of chloride ions and the
initiation of an IPSP. After recognition, GABA is released from the receptor and taken
up by surrounding glial cells, which recycle the neurotransmitter for future use.
IIIA5. Structure of the GABA Receptor Let's look closely at the GABA receptor structure. Like all ligand-gated
receptors, it is composed of five subunit proteins, each a chain of amino acids that
runs in and out of the cell membrane four times. Notice a small kink in one of the
subunit proteins.
The GABAa receptor's structure is typical of most ligand-gated (ionotropic)
receptors. It is made up of five protein subunits arranged in a circle to form a pore, or
channel, that remains closed until its specific ligand (in this case, GABA) binds to the
recognition site. Each protein subunit is actually a string of amino acids which passes
in and out of the cell membrane four times. At the extracellular end of this string is a
large N-terminal; this end-chain is thought to mediate GABA-channel interactions. In
the middle of the string is a large intracellular loop of amino acids with four sites
where phosphorylation occurs.
Each of the five subunits that make up the receptor is about 50,000 daltons in
size. These subunits have been labeled with Greek letters, such as alpha, beta, gamma
and delta. It appears as though GABA requires both alpha and beta components in
order to bind. GABAa receptors are typically made up of two alpha and two beta
subunits among the five subunits, though the particular subunit composition often
98
Temelji neuroznanosti Vodič kroz vježbe
IIIA4. Binding of GABA to Receptor Receptors for GABA are embedded in the post-synaptic membrane. When two
molecules of GABA bind to its receptor, the receptor channel opens, and chloride ions
rush into the neuron.
The GABAa receptor is made up of five subunit proteins. Each subunit
transverses the postsynaptic membrane in a generally symmetrical manner. The
five subunits are arranged to form a central pore, which, when opened, allows for the
passage of chloride ions into the postsynaptic neuron. The GABAa receptor molecule
has a specific site at which molecules of GABA bind. The interactions between
neurotransmitter and receptor at this site do not involve the formation of chemical
bonds, but do result in changingthe conformation of the receptor to consequently open
up the central pore. Opening this pore allows for the influx of chloride ions and the
initiation of an IPSP. After recognition, GABA is released from the receptor and taken
up by surrounding glial cells, which recycle the neurotransmitter for future use.
IIIA5. Structure of the GABA Receptor Let's look closely at the GABA receptor structure. Like all ligand-gated
receptors, it is composed of five subunit proteins, each a chain of amino acids that
runs in and out of the cell membrane four times. Notice a small kink in one of the
subunit proteins.
The GABAa receptor's structure is typical of most ligand-gated (ionotropic)
receptors. It is made up of five protein subunits arranged in a circle to form a pore, or
channel, that remains closed until its specific ligand (in this case, GABA) binds to the
recognition site. Each protein subunit is actually a string of amino acids which passes
in and out of the cell membrane four times. At the extracellular end of this string is a
large N-terminal; this end-chain is thought to mediate GABA-channel interactions. In
the middle of the string is a large intracellular loop of amino acids with four sites
where phosphorylation occurs.
Each of the five subunits that make up the receptor is about 50,000 daltons in
size. These subunits have been labeled with Greek letters, such as alpha, beta, gamma
and delta. It appears as though GABA requires both alpha and beta components in
order to bind. GABAa receptors are typically made up of two alpha and two beta
subunits among the five subunits, though the particular subunit composition often
98

Temelji neuroznanosti Vodič kroz vježbe
varies widely among brain regions and species. In fact, among GABAa receptors, 17
different combinations of subunits have been identified. Having a family of receptors
that all share the same basic structure, but which differ in specific subunit
composition, allows for a greater level of functional diversity. Each of the receptor
varieties vary in binding affinity, channel activity, and the degree to which GABA
binding is affected by different endogenous modulators. As a result, inhibitory signals
can be more finely controlled. Such receptor heterogeneity also allows for more
control at the genomic level, allowing post-synaptic cells to respond to changing
developmental needs or variable activity at the synapse.
IIIA6. Ion Channel Opening: Mechanism of Action When two molecules of GABA bind to the outside of the receptor, chloride
ions rush into the cell. The channel opens because the subunit proteins rotate sideways
so that "kinks" keeping the channel closed swivel and open the channel.
The five protein subunits of the GABAa receptor are arranged in
circular formation to form a pore which transverses the membrane of the post-
synaptic neuron. When GABA is not present, this pore is shaped somewhat like an
hourglass. The openings at either end are relatively large, reaching a diameter of about
3 nm, yet the inside of the channel narrows to about .5 nm. The narrowing of the
channel is due to a kink in the three dimensional structure of the five protein units
which form the receptor. It is this narrow region which prevents chloride ions
from entering the neuron when GABA is not present. However, when GABA binds
to the recognition site, it induces several conformational changes in the receptor
molecule. One of these changes involves the rotation of the five protein subunits such
that the diameter of the kink is widened. A wider channel makes it possible for
choride ions to pass freely into the neuron.
IIIA7. Positive Neuromodulation of GABAa Receptors:
Tranquilizers Here we see how a drug can improve the binding of GABA. When GABA
binds to the receptor, negative chloride ions rush into the cell. This increases the
negative charge inside the postsynaptic cell. Now a tranquilizer, coming in from the
99
Temelji neuroznanosti Vodič kroz vježbe
varies widely among brain regions and species. In fact, among GABAa receptors, 17
different combinations of subunits have been identified. Having a family of receptors
that all share the same basic structure, but which differ in specific subunit
composition, allows for a greater level of functional diversity. Each of the receptor
varieties vary in binding affinity, channel activity, and the degree to which GABA
binding is affected by different endogenous modulators. As a result, inhibitory signals
can be more finely controlled. Such receptor heterogeneity also allows for more
control at the genomic level, allowing post-synaptic cells to respond to changing
developmental needs or variable activity at the synapse.
IIIA6. Ion Channel Opening: Mechanism of Action When two molecules of GABA bind to the outside of the receptor, chloride
ions rush into the cell. The channel opens because the subunit proteins rotate sideways
so that "kinks" keeping the channel closed swivel and open the channel.
The five protein subunits of the GABAa receptor are arranged in
circular formation to form a pore which transverses the membrane of the post-
synaptic neuron. When GABA is not present, this pore is shaped somewhat like an
hourglass. The openings at either end are relatively large, reaching a diameter of about
3 nm, yet the inside of the channel narrows to about .5 nm. The narrowing of the
channel is due to a kink in the three dimensional structure of the five protein units
which form the receptor. It is this narrow region which prevents chloride ions
from entering the neuron when GABA is not present. However, when GABA binds
to the recognition site, it induces several conformational changes in the receptor
molecule. One of these changes involves the rotation of the five protein subunits such
that the diameter of the kink is widened. A wider channel makes it possible for
choride ions to pass freely into the neuron.
IIIA7. Positive Neuromodulation of GABAa Receptors:
Tranquilizers Here we see how a drug can improve the binding of GABA. When GABA
binds to the receptor, negative chloride ions rush into the cell. This increases the
negative charge inside the postsynaptic cell. Now a tranquilizer, coming in from the
99

Temelji neuroznanosti Vodič kroz vježbe
left, binds to the receptor. This changes the receptor's conformation so GABA binds
more tightly. The channel is open for longer, letting in more chloride ions.
In addition to a distinct binding site for the neurotransmitter GABA, the
GABAa receptor has a second recognition site at which certain drugs can bind. In
1957, quite by accident, scientists at a drug company discovered that an unknown
compound reduced fear in animals. When tested on humans with anxiety disorders,
this drug, called Librium, was found to be a powerful tranquilizer. Since then, many
other drug companies have made minor chemical changes in the structure of Librium,
producing a large number of similarly-shaped drugs called Benzodiazepines. Two
well known Benzodiazepines are Valium and Xanax, tranquilizers which work to
reduce fear and anxiety.
Because they do not bind to the GABA recognition site, the Benzodiazepines
do not work as directly as GABA in allowing the free passage of chloride ions
through the pore of the receptor. Instead, when a drug like Valium of Xanax binds to
its specific recognition site, the 3-dimensional shape of the GABAa receptor changes
in such a way as to allow GABA to bind more easily to its own binding site.
Increased GABA-binding leads to an increased influx of choride ions at the post-
synaptic cell. Whether this increase in chloride influx is due to an increase in the
amount of time that the channel stays open, or to a greater affinity for the GABA
molecule at its binding site is not currently known. It is, however, clear that increased
chloride influx causes an increased number of inhibitory inputs (IPSPs) at the post-
synaptic neuron. This increase in IPSPs makes it difficult for any excitatory inputs
(EPSPs) to cause an action potential in the post-synaptic cell. When the excitatory
signals caused by fear or stress are inhibited in this way, the result is a marked
decrease in anxiety and a greater feeling of calm.
IIIA8. Negative Neuromodulation of GABAa Receptors:
Anxiogenics Some drugs, and even some endogenously produced proteins, such as DBI,
can impair the binding of GABA by changing the receptor's conformation and causing
GABA to bind less tightly. Now the protein DBI, coming in from the right, binds to
the receptor. Fewer chloride ions will now enter the cell.
100
Temelji neuroznanosti Vodič kroz vježbe
left, binds to the receptor. This changes the receptor's conformation so GABA binds
more tightly. The channel is open for longer, letting in more chloride ions.
In addition to a distinct binding site for the neurotransmitter GABA, the
GABAa receptor has a second recognition site at which certain drugs can bind. In
1957, quite by accident, scientists at a drug company discovered that an unknown
compound reduced fear in animals. When tested on humans with anxiety disorders,
this drug, called Librium, was found to be a powerful tranquilizer. Since then, many
other drug companies have made minor chemical changes in the structure of Librium,
producing a large number of similarly-shaped drugs called Benzodiazepines. Two
well known Benzodiazepines are Valium and Xanax, tranquilizers which work to
reduce fear and anxiety.
Because they do not bind to the GABA recognition site, the Benzodiazepines
do not work as directly as GABA in allowing the free passage of chloride ions
through the pore of the receptor. Instead, when a drug like Valium of Xanax binds to
its specific recognition site, the 3-dimensional shape of the GABAa receptor changes
in such a way as to allow GABA to bind more easily to its own binding site.
Increased GABA-binding leads to an increased influx of choride ions at the post-
synaptic cell. Whether this increase in chloride influx is due to an increase in the
amount of time that the channel stays open, or to a greater affinity for the GABA
molecule at its binding site is not currently known. It is, however, clear that increased
chloride influx causes an increased number of inhibitory inputs (IPSPs) at the post-
synaptic neuron. This increase in IPSPs makes it difficult for any excitatory inputs
(EPSPs) to cause an action potential in the post-synaptic cell. When the excitatory
signals caused by fear or stress are inhibited in this way, the result is a marked
decrease in anxiety and a greater feeling of calm.
IIIA8. Negative Neuromodulation of GABAa Receptors:
Anxiogenics Some drugs, and even some endogenously produced proteins, such as DBI,
can impair the binding of GABA by changing the receptor's conformation and causing
GABA to bind less tightly. Now the protein DBI, coming in from the right, binds to
the receptor. Fewer chloride ions will now enter the cell.
100

Temelji neuroznanosti Vodič kroz vježbe
The finding that Benzodiazepines bind to the GABAa receptor raises a very
interesting question: Why is that our brains have a naturally-occurring recognition site
for these synthetic chemicals? This question led scientists to the conclusion that there
had to be an endogenous neurochemical shaped like the benzodiazepines, which could
bind to the same spot on the GABAa receptor. Such a compound was
subsequently identified in the brains of rats and in the cerebral spinal fluid of humans.
This chemical, a peptide named Diazepam Binding Inhibitor (DBI), was indeed,
found to bind to the same site as the benzodiazepines, yet caused an effect that was
opposite to these tranquilizers. Upon binding, DBI changes the conformation of the
GABAa receptor such that the ability of GABA to bind to its receptor site is
decreased. This leads to a decrease in the influx of chloride ions into the postsynaptic
cell and, as a result, fewer inhibitory post-synaptic potentials. DBI is thus a negative
modulator of the GABAa receptor.
In order to determine the behavioral effect of this endogenous peptide,
neuroscientists worked on developing synthetic compounds which would bind to the
same recognition site as DBI and which, like DBI, would act as negative modulators
of the GABAa receptor. The production of these compounds, known as Inverse
Agonists, have helped to confirm the role of DBI as an endogenous anxiogenic. When
injected into human volunteers, one of these synthetic inverse agonists, FG7142,
caused sweating, nausea, palpitations, tightness in the chest, restlessness, tremors, and
feelings of worry and impending doom- all markers of increased anxiety. Some
people are naturally more anxious than others. Such behavioral variation most likely
results from individual differences in the steady state production of DBI, and thus the
amount of this peptide normally found in the brain. The steady state production of
DBI is thought to be a result of genetic background, as well as early conditioning
experiences.
It has recently been discovered that certain steroids also have a binding site on
the GABAa receptor. This binding site is again distinct -- it is not the place where
GABA binds, nor is it the place where DBI and the benzodiazepines bind. The
steroids that bind at these sites are unique in that they are synthesized from circulating
cholesterol locally in the glial cells of the brain. In times of stress, cholesterol is
converted to progesterone, which is then converted to allopregnanolone. Because of
its direct effect on neurotransmission, allopregnanolone is called a neuroactive
101
Temelji neuroznanosti Vodič kroz vježbe
The finding that Benzodiazepines bind to the GABAa receptor raises a very
interesting question: Why is that our brains have a naturally-occurring recognition site
for these synthetic chemicals? This question led scientists to the conclusion that there
had to be an endogenous neurochemical shaped like the benzodiazepines, which could
bind to the same spot on the GABAa receptor. Such a compound was
subsequently identified in the brains of rats and in the cerebral spinal fluid of humans.
This chemical, a peptide named Diazepam Binding Inhibitor (DBI), was indeed,
found to bind to the same site as the benzodiazepines, yet caused an effect that was
opposite to these tranquilizers. Upon binding, DBI changes the conformation of the
GABAa receptor such that the ability of GABA to bind to its receptor site is
decreased. This leads to a decrease in the influx of chloride ions into the postsynaptic
cell and, as a result, fewer inhibitory post-synaptic potentials. DBI is thus a negative
modulator of the GABAa receptor.
In order to determine the behavioral effect of this endogenous peptide,
neuroscientists worked on developing synthetic compounds which would bind to the
same recognition site as DBI and which, like DBI, would act as negative modulators
of the GABAa receptor. The production of these compounds, known as Inverse
Agonists, have helped to confirm the role of DBI as an endogenous anxiogenic. When
injected into human volunteers, one of these synthetic inverse agonists, FG7142,
caused sweating, nausea, palpitations, tightness in the chest, restlessness, tremors, and
feelings of worry and impending doom- all markers of increased anxiety. Some
people are naturally more anxious than others. Such behavioral variation most likely
results from individual differences in the steady state production of DBI, and thus the
amount of this peptide normally found in the brain. The steady state production of
DBI is thought to be a result of genetic background, as well as early conditioning
experiences.
It has recently been discovered that certain steroids also have a binding site on
the GABAa receptor. This binding site is again distinct -- it is not the place where
GABA binds, nor is it the place where DBI and the benzodiazepines bind. The
steroids that bind at these sites are unique in that they are synthesized from circulating
cholesterol locally in the glial cells of the brain. In times of stress, cholesterol is
converted to progesterone, which is then converted to allopregnanolone. Because of
its direct effect on neurotransmission, allopregnanolone is called a neuroactive
101

Temelji neuroznanosti Vodič kroz vježbe
steroid. Like the benzodiazepines, this hormone acts as a positive modulator of the
GABAa receptor, increasing the ability of GABA to bind to the receptor, and thus
increasing the influx of chloride ions at the post-synaptic membrane. A second
neuroactive steroid is a progesterone precursor known as pregnenolone. This
neuroactive steroid differs from allopregnanolone in that it is a negative modulator of
the GABAa receptor, thus producing the same effects as DBI. For a list of some
agonists/facilitators and antagonists of GABA, go to
http://www.pharmcentral.com/GABA.htm.
IIIA9. Clinical Application: GABA and Anxiety To sum up, GABA receptors open channels for negative chloride ions, making
it less likely that action potentials can be generated in output neurons in the amygdala.
These output neurons will then stop sending signals from the amygdala in the limbic
system to the frontal cortex.
In the inhibitory neurons of the amygdala, the arrival of an action potential at
the axon terminal causes the release of GABA into the synaptic cleft. Recognition by
GABAa post-synaptic receptors causes an influx of chloride through the post-synaptic
cell membrane, decreasing the effectiveness of EPSPs and thus decreasing the ability
of that neuron to fire action potentials. If no action potential is fired in this cell,
one excitatory circuit in the amygdala has thus been quieted. When this inhibitory
action occurs in many circuits at the same time, the activity of the amygdala as a
whole decreases, and it fires less excitatory signals to other areas of the brain. This in
turn leads to a reduction in the physiological and psychological markers of stress and
anxiety. To learn more about anxiety disorders, go to
http://www.nlm.nih.gov/medlineplus/anxiety.html.
B. Metabotropic (G-protein coupled) receptors
IIIB1. Dopamine Receptors: A Representative Family of
Metabotropic Receptors This is a cross-section of a human brain, facing left. The lights are showing
you some of the areas in the brain that use the neurotransmitter dopamine. One of
these areas is the mesolimbic pathway. Mesolimbic system neurons have their cell
bodies in the midbrain and send their axons forward to the limbic system and cortex.
102
Temelji neuroznanosti Vodič kroz vježbe
steroid. Like the benzodiazepines, this hormone acts as a positive modulator of the
GABAa receptor, increasing the ability of GABA to bind to the receptor, and thus
increasing the influx of chloride ions at the post-synaptic membrane. A second
neuroactive steroid is a progesterone precursor known as pregnenolone. This
neuroactive steroid differs from allopregnanolone in that it is a negative modulator of
the GABAa receptor, thus producing the same effects as DBI. For a list of some
agonists/facilitators and antagonists of GABA, go to
http://www.pharmcentral.com/GABA.htm.
IIIA9. Clinical Application: GABA and Anxiety To sum up, GABA receptors open channels for negative chloride ions, making
it less likely that action potentials can be generated in output neurons in the amygdala.
These output neurons will then stop sending signals from the amygdala in the limbic
system to the frontal cortex.
In the inhibitory neurons of the amygdala, the arrival of an action potential at
the axon terminal causes the release of GABA into the synaptic cleft. Recognition by
GABAa post-synaptic receptors causes an influx of chloride through the post-synaptic
cell membrane, decreasing the effectiveness of EPSPs and thus decreasing the ability
of that neuron to fire action potentials. If no action potential is fired in this cell,
one excitatory circuit in the amygdala has thus been quieted. When this inhibitory
action occurs in many circuits at the same time, the activity of the amygdala as a
whole decreases, and it fires less excitatory signals to other areas of the brain. This in
turn leads to a reduction in the physiological and psychological markers of stress and
anxiety. To learn more about anxiety disorders, go to
http://www.nlm.nih.gov/medlineplus/anxiety.html.
B. Metabotropic (G-protein coupled) receptors
IIIB1. Dopamine Receptors: A Representative Family of
Metabotropic Receptors This is a cross-section of a human brain, facing left. The lights are showing
you some of the areas in the brain that use the neurotransmitter dopamine. One of
these areas is the mesolimbic pathway. Mesolimbic system neurons have their cell
bodies in the midbrain and send their axons forward to the limbic system and cortex.
102

Temelji neuroznanosti Vodič kroz vježbe
At the axon terminal, vesicles containing dopamine fuse with the presynaptic cell
membrane, releasing the neurotransmitter into the synaptic cleft.
Dopamine (DA) is a small-molecule neurotransmitter involved in the control
of both motor and emotional behavior. Despite the large number of crucial functions it
performs, this chemical messenger is found in a relatively small number of brain cells.
In fact, while there are a total of 10 billion total cells in the cerebral cortex alone,
there are only one million dopaminergic cells in the entire brain.
Though small in number, dopaminergic cells are well dispersed among four
major tracts in the brain. The nigrostriatal tract begins in the substantia nigra. These
neurons extend to the putamen and caudate nucleus and are involved in the fine tuning
of movement. The breakdown of neurons in this pathway is associated with the
tremors, rigidity, bent posture, and slowness of movement characteristic of
Parkinson's Disease.
The neurons in the tuberoinfundibular tract have cell bodies in the arcurate
nucleus of the hypothalamus and extend to the pituitary stalk. These cells are involved
in the control of hormones.
There are two dopaminergic tracts within the mesolimbocortical system. Both
are involved in motivated and emotional behavior, and in organized thought
processes. The mesolimbic tract begins in the ventral tegmental area and projects to
many components of the limbic system. The mesocortical system also stems from the
ventral tegmental area, but then projects to the neocortex, mostly to the prefrontal
areas of the neocortex. Excess dopaminergic transmission within the two tracts of the
mesolimbocortical system is associated with the disordered thought and inappropriate
emotional behavior characteristic of schizophrenia.
Like all small-molecule neurotransmitters, dopamine (DA) is synthesized in
the nerve terminal and stored there until an action potential arrives. Once released into
the synaptic cleft, receptors on the postsynaptic cell recognize and interact with the
DA molecules to cause depolarization of the postsynaptic cell and the initiation of a
new action potential.
Six dopaminergic receptors have been well-characterized. All of these
receptors, known as D1, D2a, D2b, D3, D4, and D5, are metabotropic, or G-linked,
named so because they are linked to heterotrimeric GTP-binding (G) proteins. G-
proteins relay signals received by receptors at the cell surface to effector molecules
103
Temelji neuroznanosti Vodič kroz vježbe
At the axon terminal, vesicles containing dopamine fuse with the presynaptic cell
membrane, releasing the neurotransmitter into the synaptic cleft.
Dopamine (DA) is a small-molecule neurotransmitter involved in the control
of both motor and emotional behavior. Despite the large number of crucial functions it
performs, this chemical messenger is found in a relatively small number of brain cells.
In fact, while there are a total of 10 billion total cells in the cerebral cortex alone,
there are only one million dopaminergic cells in the entire brain.
Though small in number, dopaminergic cells are well dispersed among four
major tracts in the brain. The nigrostriatal tract begins in the substantia nigra. These
neurons extend to the putamen and caudate nucleus and are involved in the fine tuning
of movement. The breakdown of neurons in this pathway is associated with the
tremors, rigidity, bent posture, and slowness of movement characteristic of
Parkinson's Disease.
The neurons in the tuberoinfundibular tract have cell bodies in the arcurate
nucleus of the hypothalamus and extend to the pituitary stalk. These cells are involved
in the control of hormones.
There are two dopaminergic tracts within the mesolimbocortical system. Both
are involved in motivated and emotional behavior, and in organized thought
processes. The mesolimbic tract begins in the ventral tegmental area and projects to
many components of the limbic system. The mesocortical system also stems from the
ventral tegmental area, but then projects to the neocortex, mostly to the prefrontal
areas of the neocortex. Excess dopaminergic transmission within the two tracts of the
mesolimbocortical system is associated with the disordered thought and inappropriate
emotional behavior characteristic of schizophrenia.
Like all small-molecule neurotransmitters, dopamine (DA) is synthesized in
the nerve terminal and stored there until an action potential arrives. Once released into
the synaptic cleft, receptors on the postsynaptic cell recognize and interact with the
DA molecules to cause depolarization of the postsynaptic cell and the initiation of a
new action potential.
Six dopaminergic receptors have been well-characterized. All of these
receptors, known as D1, D2a, D2b, D3, D4, and D5, are metabotropic, or G-linked,
named so because they are linked to heterotrimeric GTP-binding (G) proteins. G-
proteins relay signals received by receptors at the cell surface to effector molecules
103

Temelji neuroznanosti Vodič kroz vježbe
within the cell, often setting off a cascade of intracellular reactions. In this section, we
will use the D1 receptor to illustrate one way that metabotropic receptors, and the
proteins they are linked to, can indirectly open the ion channels necessary to cause a
wave of depolarization.
IIIB2. Dopamine D1 Receptor When dopamine is released into the synaptic cleft, this neurotransmitter binds
to a G-protein linked receptor embedded in the post-synaptic membrane The
dopamine receptor protein is a string of amino acids that weaves in and out of the
postsynaptic cell membrane seven times.
The D1 receptor is made up of a string of 446 amino acids folded to form a
three dimensional protein embedded in the post-synaptic cell membrane. This string
of amino acids weaves in and out of the post-synaptic membrane to form seven
membrane-spanning domains. These transmembrane regions form a pocket which
functions as the recognition site for dopamine molecules. More specifically, it is the
third and fifth regions that are thought to be involved in transiently binding the
neurotransmitter to the receptor. The extracellular loops of the structure are also
thought to be involved in this ligand-binding action. It is the third intracellular loop of
the protein, however, which serves as the binding-site for the G-protein associated
with the receptor.
IIIB3. Function of the G-protein The dopamine receptor, shown in violet, is adjacent to a G-protein shown here
in yellow. In the inactive state, the G protein binds the nucleotide GDP. The G-protein
has three subunits, alpha, beta and gamma. Activation of the receptor by the
neurotransmitter dopamine causes the alpha subunit to exchange its GDP for a GTP.
The G protein then disassociates. The alpha subunit, with GTP, pulls away leaving
behind the beta and gamma subunits.
Heterotrimeric G-proteins are made up of alpha, beta, and gamma subunits.
The chemical qualities of the alpha subunit allow it to bind easily to one of two
guanine subunits, GDP or GTP. The protein thus has two functional formations. When
GDP is bound to the alpha subunit, the alpha subunit remains bound to the beta-
gamma subunit to form an inactive trimeric protein. However, when GTP binds to the
104
Temelji neuroznanosti Vodič kroz vježbe
within the cell, often setting off a cascade of intracellular reactions. In this section, we
will use the D1 receptor to illustrate one way that metabotropic receptors, and the
proteins they are linked to, can indirectly open the ion channels necessary to cause a
wave of depolarization.
IIIB2. Dopamine D1 Receptor When dopamine is released into the synaptic cleft, this neurotransmitter binds
to a G-protein linked receptor embedded in the post-synaptic membrane The
dopamine receptor protein is a string of amino acids that weaves in and out of the
postsynaptic cell membrane seven times.
The D1 receptor is made up of a string of 446 amino acids folded to form a
three dimensional protein embedded in the post-synaptic cell membrane. This string
of amino acids weaves in and out of the post-synaptic membrane to form seven
membrane-spanning domains. These transmembrane regions form a pocket which
functions as the recognition site for dopamine molecules. More specifically, it is the
third and fifth regions that are thought to be involved in transiently binding the
neurotransmitter to the receptor. The extracellular loops of the structure are also
thought to be involved in this ligand-binding action. It is the third intracellular loop of
the protein, however, which serves as the binding-site for the G-protein associated
with the receptor.
IIIB3. Function of the G-protein The dopamine receptor, shown in violet, is adjacent to a G-protein shown here
in yellow. In the inactive state, the G protein binds the nucleotide GDP. The G-protein
has three subunits, alpha, beta and gamma. Activation of the receptor by the
neurotransmitter dopamine causes the alpha subunit to exchange its GDP for a GTP.
The G protein then disassociates. The alpha subunit, with GTP, pulls away leaving
behind the beta and gamma subunits.
Heterotrimeric G-proteins are made up of alpha, beta, and gamma subunits.
The chemical qualities of the alpha subunit allow it to bind easily to one of two
guanine subunits, GDP or GTP. The protein thus has two functional formations. When
GDP is bound to the alpha subunit, the alpha subunit remains bound to the beta-
gamma subunit to form an inactive trimeric protein. However, when GTP binds to the
104

Temelji neuroznanosti Vodič kroz vježbe
alpha subunit, the alpha subunit dissociates from the beta-gamma complex and is then
free to interact with other effector molecules.
The unique chemical qualities of G-proteins are what allow metabotropic
receptors to indirectly control the opening and closing of ion channels located at other
places on the postsynaptic membrane. When there is no dopamine in the synaptic
cleft, GDP remains bound to the alpha subunit and the G-protein is inactive. However,
the binding of dopamine at the extracellular recognition site of the receptor causes the
receptor to change shape. This conformational change causes the alpha subunit to give
up its GDP in exchange for GTP. Once bound to GTP, the alpha subunit dissociates
from the beta-gamma complex. The free alpha-GTP complex then binds with
adenylate cyclase, a transmembrane protein which converts adenosine triphosphate
(ATP) to adenosine 3,5 monophosphate (cyclic AMP, or cAMP).
IIIB4. G-proteins, cAMP, and Ion Channel Opening The alpha subunit activates adenylate cyclase, in purple, and loses GTP.
Adenylate cyclase converts ATP to cyclic AMP, which then activates Protein Kinase,
shown in blue. Protein Kinase phosphorylates an ion channel, letting sodium ions rush
into the cell. Notice that the alpha subunit, now carrying GDP, has reattached to the
beta and gamma G protein subunits by the dopamine receptor.
Cyclic AMP (cAMP) is a protein messenger present in many different types of
cells which activates cAMP-dependent protein kinases, causing them to transfer
phosphate groups from free molecules of ATP to various proteins in the cell. In this
case, protein kinase A (PKA) transfers these phosphate groups to proteins on ion
channels on the cell membrane. Phosphorylating these channel proteins increases the
chances of the channels opening, thus increasing the chances of depolarizing the
postsynaptic neuron at the axon hillock and setting off an action potential. Such
events are called excitatory postsynaptic potentials (EPSPs). In this case, when
dopamine receptors are activated by dopamine, the linked G-protein dissociates and
stimulates the production of cAMP, which then activates the protein kinase, which in
turn phosphorylates the ion channel, allowing sodium (Na+) ions to rush into the
postsynaptic call and depolarize it. If enough depolarization occurs, an action
potential will be generated and a signal sent down the axon of the postsynaptic cell.
105
Temelji neuroznanosti Vodič kroz vježbe
alpha subunit, the alpha subunit dissociates from the beta-gamma complex and is then
free to interact with other effector molecules.
The unique chemical qualities of G-proteins are what allow metabotropic
receptors to indirectly control the opening and closing of ion channels located at other
places on the postsynaptic membrane. When there is no dopamine in the synaptic
cleft, GDP remains bound to the alpha subunit and the G-protein is inactive. However,
the binding of dopamine at the extracellular recognition site of the receptor causes the
receptor to change shape. This conformational change causes the alpha subunit to give
up its GDP in exchange for GTP. Once bound to GTP, the alpha subunit dissociates
from the beta-gamma complex. The free alpha-GTP complex then binds with
adenylate cyclase, a transmembrane protein which converts adenosine triphosphate
(ATP) to adenosine 3,5 monophosphate (cyclic AMP, or cAMP).
IIIB4. G-proteins, cAMP, and Ion Channel Opening The alpha subunit activates adenylate cyclase, in purple, and loses GTP.
Adenylate cyclase converts ATP to cyclic AMP, which then activates Protein Kinase,
shown in blue. Protein Kinase phosphorylates an ion channel, letting sodium ions rush
into the cell. Notice that the alpha subunit, now carrying GDP, has reattached to the
beta and gamma G protein subunits by the dopamine receptor.
Cyclic AMP (cAMP) is a protein messenger present in many different types of
cells which activates cAMP-dependent protein kinases, causing them to transfer
phosphate groups from free molecules of ATP to various proteins in the cell. In this
case, protein kinase A (PKA) transfers these phosphate groups to proteins on ion
channels on the cell membrane. Phosphorylating these channel proteins increases the
chances of the channels opening, thus increasing the chances of depolarizing the
postsynaptic neuron at the axon hillock and setting off an action potential. Such
events are called excitatory postsynaptic potentials (EPSPs). In this case, when
dopamine receptors are activated by dopamine, the linked G-protein dissociates and
stimulates the production of cAMP, which then activates the protein kinase, which in
turn phosphorylates the ion channel, allowing sodium (Na+) ions to rush into the
postsynaptic call and depolarize it. If enough depolarization occurs, an action
potential will be generated and a signal sent down the axon of the postsynaptic cell.
105

Temelji neuroznanosti Vodič kroz vježbe
IIIB5. Dopamine Receptor Blockade: Antipsychotic Drugs Here you can see the entire G-protein receptor process, from the activation of
the receptor by the neurotransmitter dopamine to the opening of the sodium ion
channel. Now an anti-psychotic drug comes in from the left and one binds to the
receptor. It binds better than dopamine, but can't activate the G-protein. Dopamine
tries to bind, but its binding site is occupied by the drug. This is how antipsychotic
drugs prevent sodium ions from entering the postsynaptic cell.
Excessive neurotransmission of dopamine is associated with schizophrenia , a
clinical condition marked by seriously disordered thought. Antipsychotics, also called
neuroleptics, are a class of compounds with a high affinity for several subtypes of
dopamine receptors. The chemical structure of the various antipsychotics allows them
to bind to dopamine receptors without triggering the postsynaptic response that the
binding of dopamine normally would. Because of their ability to block dopamine
receptors without causing the opening of ion channels and setting off an action
potential, neuroleptics can be administered to schizophrenic patients to help reduce
excess levels of dopamine, and to thus help alleviate the positive symptoms of the
disorder.
There are approximately thirty antipsychotic drugs presently available in
North America. Developed in the 1950's, the "typical antipsychotic drugs" include the
phenotiazines (e.g. chloropromazine), the thioanthenes (e.g. chlorprothixene), the
butyrophenones (e.g. haloperidol), the diphenypbutylpiperidines (e.g. pimozide), and
the dihydroindolones (e.g. molindone). These drugs illustrate a high affinity for the
D2 family of dopamine receptors and it is at these sites that they are thought to exert
their therapeutic action. Because D2 dopamine receptors are present not only on the
post-synaptic membrane, but on the cell bodies, dendrites and nerve terminals of
presynaptic cells as well, antipsychotic compounds can interfere with dopaminergic
neurotransmission at various sites in both the pre- and postsynaptic cell. Some
neuroleptics interfere with the release of DA at the presynaptic terminal, while others
block postsynaptic dopamine receptors so that postsynaptic cells cannot recognize
dopamine.
Using typical antipsychotic drugs to interfere with dopaminergic
neurotransmission in the limbic system and in the cerebral cortex (areas involved in
the control of motivated and emotional behavior and in facilitating organized thought)
106
Temelji neuroznanosti Vodič kroz vježbe
IIIB5. Dopamine Receptor Blockade: Antipsychotic Drugs Here you can see the entire G-protein receptor process, from the activation of
the receptor by the neurotransmitter dopamine to the opening of the sodium ion
channel. Now an anti-psychotic drug comes in from the left and one binds to the
receptor. It binds better than dopamine, but can't activate the G-protein. Dopamine
tries to bind, but its binding site is occupied by the drug. This is how antipsychotic
drugs prevent sodium ions from entering the postsynaptic cell.
Excessive neurotransmission of dopamine is associated with schizophrenia , a
clinical condition marked by seriously disordered thought. Antipsychotics, also called
neuroleptics, are a class of compounds with a high affinity for several subtypes of
dopamine receptors. The chemical structure of the various antipsychotics allows them
to bind to dopamine receptors without triggering the postsynaptic response that the
binding of dopamine normally would. Because of their ability to block dopamine
receptors without causing the opening of ion channels and setting off an action
potential, neuroleptics can be administered to schizophrenic patients to help reduce
excess levels of dopamine, and to thus help alleviate the positive symptoms of the
disorder.
There are approximately thirty antipsychotic drugs presently available in
North America. Developed in the 1950's, the "typical antipsychotic drugs" include the
phenotiazines (e.g. chloropromazine), the thioanthenes (e.g. chlorprothixene), the
butyrophenones (e.g. haloperidol), the diphenypbutylpiperidines (e.g. pimozide), and
the dihydroindolones (e.g. molindone). These drugs illustrate a high affinity for the
D2 family of dopamine receptors and it is at these sites that they are thought to exert
their therapeutic action. Because D2 dopamine receptors are present not only on the
post-synaptic membrane, but on the cell bodies, dendrites and nerve terminals of
presynaptic cells as well, antipsychotic compounds can interfere with dopaminergic
neurotransmission at various sites in both the pre- and postsynaptic cell. Some
neuroleptics interfere with the release of DA at the presynaptic terminal, while others
block postsynaptic dopamine receptors so that postsynaptic cells cannot recognize
dopamine.
Using typical antipsychotic drugs to interfere with dopaminergic
neurotransmission in the limbic system and in the cerebral cortex (areas involved in
the control of motivated and emotional behavior and in facilitating organized thought)
106

Temelji neuroznanosti Vodič kroz vježbe
can be helpful in alleviating the positive symptoms of schizophrenia. The inhibition of
dopamine transmission in other pathways, however, can result in a wide range of
highly undesirable side effects. Unfortunately, when typical antipsychotics are
administered, all D2 receptors are blocked, including those in areas of the brain
involved in the fine tuning of motor movement (namely, the basal ganglia and
cerebellum). As a result, when these drugs are administered to schizophrenic patients,
many experience the motor dysfunction characteristic of Parkinson's disease: tremors,
akinesia (a slowing of voluntary movements), spasticity and rigidity, and a kathesia
(discomfort and a feeling of restlessness in the legs). The use of typical antipsychotics
may also interfere with normal endocrine function, and can exert anticholinergic,
antiadrenergic, antihistaminic and antiserotonergic actions.
A second class of neuroleptics, known as "atypical antipsychotics," exert the
same therapeutic effects of the traditional antipsychotics without producing the
undesirable side effects of the older, typical antipsychotics. The atypical
antipsychotics illustrate a lower affinity for D2 receptors, yet readily bind with D3
and D4 dopamine receptors. Since the expression of D3 and D4 is restricted to the
neurons of the limbic system and cerebral cortex only, the action of these newer
antipsychotics is confined to areas involved in the pathology of schizophrenia.
IIIB6. Clinical Application: Dopamine and Schizophrenia Learning about the function of the dopamine g-protein linked receptor and the
role of neurons in the mesolimbic system which use dopamine as their
neurotransmitter will help you to understand about behaviors such as motivation and
disorders such as schizophrenia.
Schizophrenia is a class of mental disorders marked by psychotic episodes
during which affected individuals exhibit gross distortions of thought and an inability
to test reality. The word schizophrenia , translated from Greek, means "split brain"
and refers to the splitting of cognitive function from emotional and motor behavior. In
patients with schizophrenia, the integrative centers of the brain, namely the limbic
system and the cortex of the prefrontal and temporal lobes, are not functioning
normally. The currently accepted hypothesis for this disturbance in function postulates
that, in patients with the condition, there is an overproduction of dopamine in the
mesolimbic neurons that project to the limbic system and to the prefrontal cortex. This
107
Temelji neuroznanosti Vodič kroz vježbe
can be helpful in alleviating the positive symptoms of schizophrenia. The inhibition of
dopamine transmission in other pathways, however, can result in a wide range of
highly undesirable side effects. Unfortunately, when typical antipsychotics are
administered, all D2 receptors are blocked, including those in areas of the brain
involved in the fine tuning of motor movement (namely, the basal ganglia and
cerebellum). As a result, when these drugs are administered to schizophrenic patients,
many experience the motor dysfunction characteristic of Parkinson's disease: tremors,
akinesia (a slowing of voluntary movements), spasticity and rigidity, and a kathesia
(discomfort and a feeling of restlessness in the legs). The use of typical antipsychotics
may also interfere with normal endocrine function, and can exert anticholinergic,
antiadrenergic, antihistaminic and antiserotonergic actions.
A second class of neuroleptics, known as "atypical antipsychotics," exert the
same therapeutic effects of the traditional antipsychotics without producing the
undesirable side effects of the older, typical antipsychotics. The atypical
antipsychotics illustrate a lower affinity for D2 receptors, yet readily bind with D3
and D4 dopamine receptors. Since the expression of D3 and D4 is restricted to the
neurons of the limbic system and cerebral cortex only, the action of these newer
antipsychotics is confined to areas involved in the pathology of schizophrenia.
IIIB6. Clinical Application: Dopamine and Schizophrenia Learning about the function of the dopamine g-protein linked receptor and the
role of neurons in the mesolimbic system which use dopamine as their
neurotransmitter will help you to understand about behaviors such as motivation and
disorders such as schizophrenia.
Schizophrenia is a class of mental disorders marked by psychotic episodes
during which affected individuals exhibit gross distortions of thought and an inability
to test reality. The word schizophrenia , translated from Greek, means "split brain"
and refers to the splitting of cognitive function from emotional and motor behavior. In
patients with schizophrenia, the integrative centers of the brain, namely the limbic
system and the cortex of the prefrontal and temporal lobes, are not functioning
normally. The currently accepted hypothesis for this disturbance in function postulates
that, in patients with the condition, there is an overproduction of dopamine in the
mesolimbic neurons that project to the limbic system and to the prefrontal cortex. This
107

Temelji neuroznanosti Vodič kroz vježbe
hypothesis is, of course, only one of many possible explanations behind this highly
complex brain disorder
The symptoms that result from the hypothesized dopminergic dysfunction can
be broken into two categories. Positive symptoms include cognitive, emotional, and
behavioral excesses. Hallucinations, delusions, incoherent speech, inappropriate
emotional responses, and an overly animated affect are all examples of positive
symptoms. Negative symptoms include cognitive, emotional, and behavioral deficits.
Blank stares, blunted affect, slowed movement and speech, and social withdrawal are
all examples of negative symptoms. Absorbed by distorted beliefs and out of touch
with the world around them, many schizophrenics exhibit bizarre behaviors such as
talking to themselves or sitting for several hours at a time in the same awkward
position.
Psychiatrists distinguish between five different types of schizophrenia, each
characterized by a different combination of stereotypic behaviors. Disorganized
schizophrenia is used to describe individuals who exhibit illogical thinking,
incoherent speech, childish mannerisms, overly animated emotions, and who often
neglect personal hygiene. Paranoid schizophrenics are characterized by a
preoccupation with delusions or hallucinations, often becoming irrationally suspicious
or hostile. Catatonic schizophrenia is a more rare form of the condition marked by
extremes in motor behavior. Individuals with catatonic schizophrenia may spend
hours in the same, motionless pose or, at the other extreme, may become overcome
with energetic bursts of hyperactivity. Undifferentiated schizophrenia is a term used
to describe those individuals who exhibit a wide range of symptoms and cannot be
classified under one specific category. Residual schizophrenics are individuals who
have had previous schizophrenic episodes but who are currently not experiencing any
major symptoms of the disorder.
It is estimated that close to 1% of the American population is afflicted with a
schizophrenic disorder. For men, the onset of symptoms tends to occur between the
ages of eighteen to twenty-five. For women, the onset is usually later, occurring
between the ages of twenty-six to forty-five. In some cases, after striking suddenly
and unexpectedly, the disorder disappears and may be followed by a full recovery. In
other cases, however, symptoms develop more gradually and remain problematic, if
left untreated, throughout the individual's lifetime. To learn more about this disturbing
108
Temelji neuroznanosti Vodič kroz vježbe
hypothesis is, of course, only one of many possible explanations behind this highly
complex brain disorder
The symptoms that result from the hypothesized dopminergic dysfunction can
be broken into two categories. Positive symptoms include cognitive, emotional, and
behavioral excesses. Hallucinations, delusions, incoherent speech, inappropriate
emotional responses, and an overly animated affect are all examples of positive
symptoms. Negative symptoms include cognitive, emotional, and behavioral deficits.
Blank stares, blunted affect, slowed movement and speech, and social withdrawal are
all examples of negative symptoms. Absorbed by distorted beliefs and out of touch
with the world around them, many schizophrenics exhibit bizarre behaviors such as
talking to themselves or sitting for several hours at a time in the same awkward
position.
Psychiatrists distinguish between five different types of schizophrenia, each
characterized by a different combination of stereotypic behaviors. Disorganized
schizophrenia is used to describe individuals who exhibit illogical thinking,
incoherent speech, childish mannerisms, overly animated emotions, and who often
neglect personal hygiene. Paranoid schizophrenics are characterized by a
preoccupation with delusions or hallucinations, often becoming irrationally suspicious
or hostile. Catatonic schizophrenia is a more rare form of the condition marked by
extremes in motor behavior. Individuals with catatonic schizophrenia may spend
hours in the same, motionless pose or, at the other extreme, may become overcome
with energetic bursts of hyperactivity. Undifferentiated schizophrenia is a term used
to describe those individuals who exhibit a wide range of symptoms and cannot be
classified under one specific category. Residual schizophrenics are individuals who
have had previous schizophrenic episodes but who are currently not experiencing any
major symptoms of the disorder.
It is estimated that close to 1% of the American population is afflicted with a
schizophrenic disorder. For men, the onset of symptoms tends to occur between the
ages of eighteen to twenty-five. For women, the onset is usually later, occurring
between the ages of twenty-six to forty-five. In some cases, after striking suddenly
and unexpectedly, the disorder disappears and may be followed by a full recovery. In
other cases, however, symptoms develop more gradually and remain problematic, if
left untreated, throughout the individual's lifetime. To learn more about this disturbing
108

Temelji neuroznanosti Vodič kroz vježbe
disorder, visit the schizophrenia homepage, an informative site with many helpful
links.
IV. Inactivation of Neurotransmitters After a neurotransmitter molecule has been recognized by a post-synaptic
receptor, it is released back into the synaptic cleft. Once in the synapse, it must be
quickly removed or chemically inactivated in order to prevent constant stimulation of
the post-synaptic cell and an excessive firing of action potentials.
Some neurotransmitters are removed from the synaptic cleft by special
transporter proteins on the pre-synaptic membrane. These transporter proteins carry
the neurotransmitter back into the pre-synaptic cell, where it is either re-packaged into
a vesicle and stored until it is once again needed to transmit a chemical message, or
broken down by enzymes. Serotonin is one neurotransmitter that gets recycled in this
way. Serotonin, a small-molecule neurotransmitter found in many areas throughout
the brain, is involved in a wide range of behaviors, including sleep, appetite, memory,
sexual behavior, neuroendocrine function, and mood.
Not all neurotransmitters are recycled by the presynaptic cell. Neuropeptide
neurotransmitters merely quickly diffuse away from the receptors into the surrounding
medium. One important neurotransmitter, acetylcholine, has a specialized enzyme for
inactivation right in the synaptic cleft called acetylcholinesterase (AChE. AChE is an
enzyme present at all cholinergic synapses which serves to inactivate acetylcholine by
hydrolysis. Acetylcholine (ACh), an excitatory small-molecule neurotransmitter found
at various locations throughout the central and peripheral nervous systems and at all
neuromuscular junctions, is composed of acetate and choline. AChE breaks ACh into
its component parts; acetate and choline. After hydrolysis, acetate quickly diffuses
into the surrounding medium, while choline gets taken back into the presynaptic cell
by a high affinity choline uptake (HACU) system. Choline is then recycled by the pre-
synaptic cell for use in the synthesis of more ACh.
109
Temelji neuroznanosti Vodič kroz vježbe
disorder, visit the schizophrenia homepage, an informative site with many helpful
links.
IV. Inactivation of Neurotransmitters After a neurotransmitter molecule has been recognized by a post-synaptic
receptor, it is released back into the synaptic cleft. Once in the synapse, it must be
quickly removed or chemically inactivated in order to prevent constant stimulation of
the post-synaptic cell and an excessive firing of action potentials.
Some neurotransmitters are removed from the synaptic cleft by special
transporter proteins on the pre-synaptic membrane. These transporter proteins carry
the neurotransmitter back into the pre-synaptic cell, where it is either re-packaged into
a vesicle and stored until it is once again needed to transmit a chemical message, or
broken down by enzymes. Serotonin is one neurotransmitter that gets recycled in this
way. Serotonin, a small-molecule neurotransmitter found in many areas throughout
the brain, is involved in a wide range of behaviors, including sleep, appetite, memory,
sexual behavior, neuroendocrine function, and mood.
Not all neurotransmitters are recycled by the presynaptic cell. Neuropeptide
neurotransmitters merely quickly diffuse away from the receptors into the surrounding
medium. One important neurotransmitter, acetylcholine, has a specialized enzyme for
inactivation right in the synaptic cleft called acetylcholinesterase (AChE. AChE is an
enzyme present at all cholinergic synapses which serves to inactivate acetylcholine by
hydrolysis. Acetylcholine (ACh), an excitatory small-molecule neurotransmitter found
at various locations throughout the central and peripheral nervous systems and at all
neuromuscular junctions, is composed of acetate and choline. AChE breaks ACh into
its component parts; acetate and choline. After hydrolysis, acetate quickly diffuses
into the surrounding medium, while choline gets taken back into the presynaptic cell
by a high affinity choline uptake (HACU) system. Choline is then recycled by the pre-
synaptic cell for use in the synthesis of more ACh.
109

Temelji neuroznanosti Vodič kroz vježbe
A. Enzymatic inactivation of neurotransmitters 1. Acetylcholinesterase: a representative inactivation enzyme
2. Location, structure and function of acetylcholinesterase
3. Enzyme inhibition: anticholinesterases
4. Clinical Application: Acetylcholine, Nerve Gas and Myasthenia Gravis
B. Presynaptic transporters: inactivation of neurotransmitters by
reuptake
1. Serotonin Transporter: a representative reuptake pump
2. Location and structure of the serotonin transporter
3. Mechanism of action of the serotonin transporter
4. Transporter inhibition: selective serotonin reuptake inhibitors
5. Clinical Application: Serotonin and Depression
A. Enzymatic inactivation of neurotransmitters
IVA1. Acetylcholinesterase: A Representative Inactivation
Enzyme
This is the side view of the human brain, facing left. The lights are showing
you some of the places where neurons use acetylcholine as their neurotransmitter. We
now zoom in on one of these neurons which uses acetylcholine. The cell body, located
in the septal nucleus of the hypothalamus, sends its axon to the hippocampus. At the
end of the axon is the terminal bouton. After the arrival of an action potential, vesicles
containing acetylcholine fuse with the presynaptic membrane and release
acetylcholine into the synaptic cleft.
Acetylcholine (ACh), the first neurotransmitter ever to be identified, is a
small- molecule excitatory neurotransmitter with a wide variety of known functions.
Within the central nervous system, cholinergic cells (neurons that use ACh as a
neurotransmitter) are found in several different locations of the brain, including the
striatal complex, the basal forebrain, the diencephalon, pontomesencephalic cell
groups, and the medulla. Depending on which area of the brain it is found in, ACh
may be involved in any one of several different functions. Some of these functions
include the conduction of pain, the regulation of neuroendocrine function, the
110
Temelji neuroznanosti Vodič kroz vježbe
A. Enzymatic inactivation of neurotransmitters 1. Acetylcholinesterase: a representative inactivation enzyme
2. Location, structure and function of acetylcholinesterase
3. Enzyme inhibition: anticholinesterases
4. Clinical Application: Acetylcholine, Nerve Gas and Myasthenia Gravis
B. Presynaptic transporters: inactivation of neurotransmitters by
reuptake
1. Serotonin Transporter: a representative reuptake pump
2. Location and structure of the serotonin transporter
3. Mechanism of action of the serotonin transporter
4. Transporter inhibition: selective serotonin reuptake inhibitors
5. Clinical Application: Serotonin and Depression
A. Enzymatic inactivation of neurotransmitters
IVA1. Acetylcholinesterase: A Representative Inactivation
Enzyme
This is the side view of the human brain, facing left. The lights are showing
you some of the places where neurons use acetylcholine as their neurotransmitter. We
now zoom in on one of these neurons which uses acetylcholine. The cell body, located
in the septal nucleus of the hypothalamus, sends its axon to the hippocampus. At the
end of the axon is the terminal bouton. After the arrival of an action potential, vesicles
containing acetylcholine fuse with the presynaptic membrane and release
acetylcholine into the synaptic cleft.
Acetylcholine (ACh), the first neurotransmitter ever to be identified, is a
small- molecule excitatory neurotransmitter with a wide variety of known functions.
Within the central nervous system, cholinergic cells (neurons that use ACh as a
neurotransmitter) are found in several different locations of the brain, including the
striatal complex, the basal forebrain, the diencephalon, pontomesencephalic cell
groups, and the medulla. Depending on which area of the brain it is found in, ACh
may be involved in any one of several different functions. Some of these functions
include the conduction of pain, the regulation of neuroendocrine function, the
110

Temelji neuroznanosti Vodič kroz vježbe
regulation of REM sleep cycles, and the process of learning and memory formation.
In the sympathetic and parasympathetic nervous systems and at all neuromuscular
junctions, ACh is used to signal muscle movement.
Whether it is within the brain, or at the neuromuscular junction, when ACh is
released into the synaptic cleft, it transiently binds with a ligand-gated receptor
molecule on the post-synaptic neuron or muscle cell. This binding causes the receptor
molecule to open up, allowing for an exchange of ions which subsequently results in a
depolarization of the post-synaptic neuron. If this depolarization is strong enough, an
action potential is stimulated.
After depolarization, the neurotransmitter must be swiftly removed from the
receptor molecule and inactivated in order to prevent constant stimulation of the post-
synaptic membrane and an excessive firing of action potentials. ACh is one of several
neurotransmitters whose action at the synaptic cleft is terminated by the hydrolyizing
action of an enzyme. Once ACh has been recognized and the post-synaptic membrane
has been depolarized, ACh quickly dissociates from the receptor and binds to
acetylcholinesterase (AChE), an enzyme named for its ability to break the ester bond
that holds acetate and choline together. Removal of ACh from the recognition site
causes the membrane-spanning pore of the receptor to close. The electric potential of
the membrane then normalizes and the action potential ceases to fire. The breakdown
of ACh by AChE inactivates the neurotransmitter and prevents it from reassociating
with the receptor molecule.
111
Temelji neuroznanosti Vodič kroz vježbe
regulation of REM sleep cycles, and the process of learning and memory formation.
In the sympathetic and parasympathetic nervous systems and at all neuromuscular
junctions, ACh is used to signal muscle movement.
Whether it is within the brain, or at the neuromuscular junction, when ACh is
released into the synaptic cleft, it transiently binds with a ligand-gated receptor
molecule on the post-synaptic neuron or muscle cell. This binding causes the receptor
molecule to open up, allowing for an exchange of ions which subsequently results in a
depolarization of the post-synaptic neuron. If this depolarization is strong enough, an
action potential is stimulated.
After depolarization, the neurotransmitter must be swiftly removed from the
receptor molecule and inactivated in order to prevent constant stimulation of the post-
synaptic membrane and an excessive firing of action potentials. ACh is one of several
neurotransmitters whose action at the synaptic cleft is terminated by the hydrolyizing
action of an enzyme. Once ACh has been recognized and the post-synaptic membrane
has been depolarized, ACh quickly dissociates from the receptor and binds to
acetylcholinesterase (AChE), an enzyme named for its ability to break the ester bond
that holds acetate and choline together. Removal of ACh from the recognition site
causes the membrane-spanning pore of the receptor to close. The electric potential of
the membrane then normalizes and the action potential ceases to fire. The breakdown
of ACh by AChE inactivates the neurotransmitter and prevents it from reassociating
with the receptor molecule.
111

Temelji neuroznanosti Vodič kroz vježbe
IVA2. Location, Structure and Function of Acetylcholinesterase Presynaptic vesicles release acetylcholine into the synaptic cleft where it binds
to its receptor. Right next to the receptor is acetylcholinesterase, the enzyme which
breaks up acetylcholine into acetate and choline.
AChE is a glycoprotein that exists is several forms. For images of
acetylcholinesterase's structure displayed in ribbon form, go to
http://jsdnt.claremont.edu/biochem98/acetylcholinesterase/AChEcontents.html. As a
result of the variety in chemical structure, some forms of AChE are hydrophobic,
while others are hydrophilic. The hydrophilic species generally work within the cell to
break down excess concentrations of intracellular ACh. The lipid-linked (hydophobic)
varieties, however, are the primary agents of ACh inactivation, working at the
synaptic cleft or the neuromuscular junction to break ACh into its component parts:
acetate and choline. Lipid-linked species of AChE are embedded within the post-
synaptic membrane and placed strategically close to the post-synaptic receptor
molecules in order to ensure quick inactivation of ACh.
Though the individual chemical qualities and anatomical arrangements vary
widely between the various forms of AChE, the mechanism of catalysis for all the
species remains strikingly similar. In all cases, the active site of AChE is made up of
two subsites, both of which are critical to the breakdown of ACh. The anionic site
serves to bind a molecule of ACh to the enzyme. When first discovered, this site was
thought to be made up of one or more negatively charged groups which
electrostatically interacted with the positively charged quaternary nitrogen of the ACh
molecule. More recently, however, it is thought that hydrophobic interactions are
equally, if not more important in binding this region of the substrate to the enzyme.
Once the ACh is bound, the hydrolytic reaction occurs at a second region of
the active site called the esteratic subsite. Here, the ester bond of ACh is broken,
releasing acetate and choline. Choline is then either temporarily trapped within the
junctional folds of the muscular endplate or immediately taken up again by the high
affinity choline uptake (HACU) system on the presynaptic membrane. Acetate,
however, becomes covalently bonded to serine residues within the esteratic subsite,
forming a temporary acetylated form of AChE. A molecule of water then reacts with
this intermediate, liberating the acetate group, which diffuses into the surrounding
112
Temelji neuroznanosti Vodič kroz vježbe
IVA2. Location, Structure and Function of Acetylcholinesterase Presynaptic vesicles release acetylcholine into the synaptic cleft where it binds
to its receptor. Right next to the receptor is acetylcholinesterase, the enzyme which
breaks up acetylcholine into acetate and choline.
AChE is a glycoprotein that exists is several forms. For images of
acetylcholinesterase's structure displayed in ribbon form, go to
http://jsdnt.claremont.edu/biochem98/acetylcholinesterase/AChEcontents.html. As a
result of the variety in chemical structure, some forms of AChE are hydrophobic,
while others are hydrophilic. The hydrophilic species generally work within the cell to
break down excess concentrations of intracellular ACh. The lipid-linked (hydophobic)
varieties, however, are the primary agents of ACh inactivation, working at the
synaptic cleft or the neuromuscular junction to break ACh into its component parts:
acetate and choline. Lipid-linked species of AChE are embedded within the post-
synaptic membrane and placed strategically close to the post-synaptic receptor
molecules in order to ensure quick inactivation of ACh.
Though the individual chemical qualities and anatomical arrangements vary
widely between the various forms of AChE, the mechanism of catalysis for all the
species remains strikingly similar. In all cases, the active site of AChE is made up of
two subsites, both of which are critical to the breakdown of ACh. The anionic site
serves to bind a molecule of ACh to the enzyme. When first discovered, this site was
thought to be made up of one or more negatively charged groups which
electrostatically interacted with the positively charged quaternary nitrogen of the ACh
molecule. More recently, however, it is thought that hydrophobic interactions are
equally, if not more important in binding this region of the substrate to the enzyme.
Once the ACh is bound, the hydrolytic reaction occurs at a second region of
the active site called the esteratic subsite. Here, the ester bond of ACh is broken,
releasing acetate and choline. Choline is then either temporarily trapped within the
junctional folds of the muscular endplate or immediately taken up again by the high
affinity choline uptake (HACU) system on the presynaptic membrane. Acetate,
however, becomes covalently bonded to serine residues within the esteratic subsite,
forming a temporary acetylated form of AChE. A molecule of water then reacts with
this intermediate, liberating the acetate group, which diffuses into the surrounding
112

Temelji neuroznanosti Vodič kroz vježbe
medium. What remains is an active form of the enzyme, ready to react with a newly
released molecule of ACh.
IVA3. Enzyme Inhibition: Anticholinesterases After the release of acetylcholine from vesicles, it binds to post-synaptic
receptors and is then broken down by the enzyme acetylcholinesterase. However,
when anticholinesterases bind to the enzyme, they prevent the neurotransmitter from
being broken down. Acetylcholine continues to activate its receptor.
Anticholinesterases (anti-AChE) are a class of chemicals which can take the
place of ACh at the active site of AChE, significantly reducing the neurotransmitter's
ability to bind with AChE and essentially limiting the rate at which ACh can be
broken down. Since there is no pre-synaptic re-uptake mechanism to transport ACh
back into the presynaptic cell and since ACh cannot readily dissolve into the
surrounding medium, the presence of AChE inhibitors result in an excess build-up of
ACh at the synapse. This build-up leads to an increased level of ACh activity at post-
synaptic receptor molecules and, subsequently, a continuous firing of action
potentials.
IVA4. Clinical Application: Acetylcholine, Nerve Gas and
Myasthenia Gravis Once acetylcholine is released into the synaptic cleft, it can bind to a post-
synaptic receptor and then be split into acetate and choline. Choline can then be
pumped back into the pre-synaptic terminal by its transporter to be reused for new
synthesis. Keeping a steady cycle of acetylcholine in the hippocampus helps maintain
memories.
The effect that a specific AChE inhibitor can have on the body depends largely
on the chemical properties of the molecule and the strength of the bond it forms with
AChE. Irreversible, or competitive AChE inhibitors are highly toxic. These
chemicals, also known as organophosphorus compounds or nerve gases, form
incredibly stable phosphorus bonds with AChE. These bonds can resist hydrolytic
cleavage for up to several hours, meanwhile inhibiting all normal AChE activity and
leading to an excessive, and possibly toxic, build-up of ACh at cholinergic synapses.
113
Temelji neuroznanosti Vodič kroz vježbe
medium. What remains is an active form of the enzyme, ready to react with a newly
released molecule of ACh.
IVA3. Enzyme Inhibition: Anticholinesterases After the release of acetylcholine from vesicles, it binds to post-synaptic
receptors and is then broken down by the enzyme acetylcholinesterase. However,
when anticholinesterases bind to the enzyme, they prevent the neurotransmitter from
being broken down. Acetylcholine continues to activate its receptor.
Anticholinesterases (anti-AChE) are a class of chemicals which can take the
place of ACh at the active site of AChE, significantly reducing the neurotransmitter's
ability to bind with AChE and essentially limiting the rate at which ACh can be
broken down. Since there is no pre-synaptic re-uptake mechanism to transport ACh
back into the presynaptic cell and since ACh cannot readily dissolve into the
surrounding medium, the presence of AChE inhibitors result in an excess build-up of
ACh at the synapse. This build-up leads to an increased level of ACh activity at post-
synaptic receptor molecules and, subsequently, a continuous firing of action
potentials.
IVA4. Clinical Application: Acetylcholine, Nerve Gas and
Myasthenia Gravis Once acetylcholine is released into the synaptic cleft, it can bind to a post-
synaptic receptor and then be split into acetate and choline. Choline can then be
pumped back into the pre-synaptic terminal by its transporter to be reused for new
synthesis. Keeping a steady cycle of acetylcholine in the hippocampus helps maintain
memories.
The effect that a specific AChE inhibitor can have on the body depends largely
on the chemical properties of the molecule and the strength of the bond it forms with
AChE. Irreversible, or competitive AChE inhibitors are highly toxic. These
chemicals, also known as organophosphorus compounds or nerve gases, form
incredibly stable phosphorus bonds with AChE. These bonds can resist hydrolytic
cleavage for up to several hours, meanwhile inhibiting all normal AChE activity and
leading to an excessive, and possibly toxic, build-up of ACh at cholinergic synapses.
113

Temelji neuroznanosti Vodič kroz vježbe
In the early 1930's, parathion, an irreversible AChE inhibitor, was
manufactured for use as an insecticide. However, when it was discovered that this
compound was toxic not only to insects, but to humans and other animals as well, i's
use was discontinued. Its potential as a lethal weapon, however, did not go unnoticed.
Several irreversible AChE inhibitors, including Sarin, Soman, and Tabun were
secretly manufactured in Germany prior to and during World War II for use as
chemical weapons. Though chemical warfare never actually broke out during this
time, the Allied forces, in an effort to respond to the German threat, also
manufactured their own nerve gas, called diisopropylfluoro-phosphate (DFP).
Organophosphorus compounds are stored as liquids but may be released as a
vapor cloud or a spray. Whether they come into direct contact with the skin or they
are inhaled, these deadly chemicals penetrate quickly into the bloodstream due to their
high liquid solubility. They readily pass across the blood-brain barrier, inactivating
AChE throughout the central and peripheral nervous systems and at all neuromuscular
junctions. As little as one milligram of these noxious compounds causes profuse
sweating, dimmed vision, uncontrollable vomiting and defecation, convulsions, filling
of the bronchial passages with mucus, bronchial constriction, and at last, paralysis and
asphyxiation from respiratory failure. (Feldman, Meyer, and Quenzer, 1997)
Though our bodies cannot naturally break the phosphate bond holding
organophosphorus compounds to AChE, several antidotes, including pralidoxim and
trimedoxime, can. These chemicals successfully break up the toxin-enzyme complex,
freeing the enzyme and allowing for the breakdown of accumulated ACh.
Not all ACh inhibitors are as toxic as the irreversible/competitive compounds.
Reversible AChE inhibitors are considered non-competitive because they can only
bind to AChE for a limited amount of time before the unstable complex dissociates
and ACh can once again bind to the enzyme. Because of the non-permanent nature of
their action, reversible anti-AChEs can be safely administered as therapeutic agents in
the treatment of disorders characterized by a decrease in cholinergic function. By
transiently inhibiting the enzyme, these drugs allow for the multiple use of one
molecule of ACh at the same receptor, thus compensating for the decrease in function
caused by the condition. The bond formed with AChE is, however, broken before the
accumulation of ACh in the synapse can become toxic.
114
Temelji neuroznanosti Vodič kroz vježbe
In the early 1930's, parathion, an irreversible AChE inhibitor, was
manufactured for use as an insecticide. However, when it was discovered that this
compound was toxic not only to insects, but to humans and other animals as well, i's
use was discontinued. Its potential as a lethal weapon, however, did not go unnoticed.
Several irreversible AChE inhibitors, including Sarin, Soman, and Tabun were
secretly manufactured in Germany prior to and during World War II for use as
chemical weapons. Though chemical warfare never actually broke out during this
time, the Allied forces, in an effort to respond to the German threat, also
manufactured their own nerve gas, called diisopropylfluoro-phosphate (DFP).
Organophosphorus compounds are stored as liquids but may be released as a
vapor cloud or a spray. Whether they come into direct contact with the skin or they
are inhaled, these deadly chemicals penetrate quickly into the bloodstream due to their
high liquid solubility. They readily pass across the blood-brain barrier, inactivating
AChE throughout the central and peripheral nervous systems and at all neuromuscular
junctions. As little as one milligram of these noxious compounds causes profuse
sweating, dimmed vision, uncontrollable vomiting and defecation, convulsions, filling
of the bronchial passages with mucus, bronchial constriction, and at last, paralysis and
asphyxiation from respiratory failure. (Feldman, Meyer, and Quenzer, 1997)
Though our bodies cannot naturally break the phosphate bond holding
organophosphorus compounds to AChE, several antidotes, including pralidoxim and
trimedoxime, can. These chemicals successfully break up the toxin-enzyme complex,
freeing the enzyme and allowing for the breakdown of accumulated ACh.
Not all ACh inhibitors are as toxic as the irreversible/competitive compounds.
Reversible AChE inhibitors are considered non-competitive because they can only
bind to AChE for a limited amount of time before the unstable complex dissociates
and ACh can once again bind to the enzyme. Because of the non-permanent nature of
their action, reversible anti-AChEs can be safely administered as therapeutic agents in
the treatment of disorders characterized by a decrease in cholinergic function. By
transiently inhibiting the enzyme, these drugs allow for the multiple use of one
molecule of ACh at the same receptor, thus compensating for the decrease in function
caused by the condition. The bond formed with AChE is, however, broken before the
accumulation of ACh in the synapse can become toxic.
114

Temelji neuroznanosti Vodič kroz vježbe
Reversible AChE inhibitors are used to treat myasthenia gravis, an
autoimmune disorder characterized by debilitating muscle weakness. This weakness is
caused by a progressive breakdown of ACh receptor sites on the muscular endplate.
Though ACh is present in normal concentrations, a lowered number of receptors on
post-synaptic muscle cells limits the transmission of nerve signals at neuromuscular
junctions, making muscle movement difficult. By causing a temporary excess of ACh
at the synapse, non-competitive anti-AChEs are helpful in utilizing each remaining
receptor molecule to its fullest capacity.
Pyridostigmine bromide, marketed under the name Mestinon or Regonol, is
the most widely used anti-AChE in the treatment of Myasthenia gravis. Others include
ambenonium chloride (Mytelase), and neostigmine methyl sulfate (Prostigmin). These
drugs cannot pass the blood-brain barrier, so their action is limited to the inactivation
of AChE at the neuromuscular junction. Because of this, they can be administered to
treat muscle weakness, without causing the psychotropic side-effects they might if
they could penetrate the cholinergic pathways of the central nervous system.
Reversible AChE inhibitors are also used in the treatment of Alzheimer's
disease , a serious brain disorder in which the gradual death of cholinergic brain cells
results in a progressive and significant loss of cognitive and behavioral function.
Because Alzheimer's affects brain cells, drugs used to treat this condition must readily
cross the blood-brain barrier. Tetrohydroaminoacridine (THA), known as Tacrine or
Cognex, and Donepezil, known as Aricept, are both reversible anti-AChEs with the
ability to penetrate the central nervous system. Though neither drug works as a cure or
can be used to slow the progression of the condition, administering these compounds
to Alzheimer's patients can help to ease some of the memory and language deficits
characteristic of this devastating disease.
B. Presynaptic transporters: inactivation of neurotransmitters by
reuptake
IVB1. Serotonin Transporter: A Representative Reuptake Pump This is the side view of the human brain, facing left. We see a major pathway
in the brain that uses serotonin for its neurotransmitter. The cell bodies for these
neurons are found in the raphe nucleus of the midbrain and the axons terminate in the
115
Temelji neuroznanosti Vodič kroz vježbe
Reversible AChE inhibitors are used to treat myasthenia gravis, an
autoimmune disorder characterized by debilitating muscle weakness. This weakness is
caused by a progressive breakdown of ACh receptor sites on the muscular endplate.
Though ACh is present in normal concentrations, a lowered number of receptors on
post-synaptic muscle cells limits the transmission of nerve signals at neuromuscular
junctions, making muscle movement difficult. By causing a temporary excess of ACh
at the synapse, non-competitive anti-AChEs are helpful in utilizing each remaining
receptor molecule to its fullest capacity.
Pyridostigmine bromide, marketed under the name Mestinon or Regonol, is
the most widely used anti-AChE in the treatment of Myasthenia gravis. Others include
ambenonium chloride (Mytelase), and neostigmine methyl sulfate (Prostigmin). These
drugs cannot pass the blood-brain barrier, so their action is limited to the inactivation
of AChE at the neuromuscular junction. Because of this, they can be administered to
treat muscle weakness, without causing the psychotropic side-effects they might if
they could penetrate the cholinergic pathways of the central nervous system.
Reversible AChE inhibitors are also used in the treatment of Alzheimer's
disease , a serious brain disorder in which the gradual death of cholinergic brain cells
results in a progressive and significant loss of cognitive and behavioral function.
Because Alzheimer's affects brain cells, drugs used to treat this condition must readily
cross the blood-brain barrier. Tetrohydroaminoacridine (THA), known as Tacrine or
Cognex, and Donepezil, known as Aricept, are both reversible anti-AChEs with the
ability to penetrate the central nervous system. Though neither drug works as a cure or
can be used to slow the progression of the condition, administering these compounds
to Alzheimer's patients can help to ease some of the memory and language deficits
characteristic of this devastating disease.
B. Presynaptic transporters: inactivation of neurotransmitters by
reuptake
IVB1. Serotonin Transporter: A Representative Reuptake Pump This is the side view of the human brain, facing left. We see a major pathway
in the brain that uses serotonin for its neurotransmitter. The cell bodies for these
neurons are found in the raphe nucleus of the midbrain and the axons terminate in the
115

Temelji neuroznanosti Vodič kroz vježbe
cortex. After the arrival of an action potential, vesicles fuse with the presynaptic
membrane and release serotonin into the synaptic cleft.
Serotonin (5-hydroxytryptamine, 5HT) , a monoamine neurotransmitter, plays
an important role in many behaviors including sleep, appetite, memory, sexual
behavior, neuroendocrine function, and mood. In the brain, the highest level of
serotonin is found in the dorsal and median raphe nuclear complex. Neurons in the
raphe system project beyond the raphe nucleus, most often in the lateral direction.
There are also many serotonergic neurons found in the reticular region of the lower
brain stem. For convenience, neurons in the brain stem are often divided into a caudal
system and a rostral system. Those in the caudal system descend to the spinal chord
along many pathways and are largely involved in sensory, motor, and autonomic
functioning. Serotonergic cells in the rostral system of the brainstem largely terminate
in the dorsal and median raphe nuclei.
Another major serotonergic pathway in the brain is one going from the
cerebellum that terminates in the cerebellar cortex and the cerebellar nuclei. Many
serotonergic neurons contribute to this pathway. The presence of serotonin has also
been observed in the pons, medulla, thalamus, hypothalamus, substantia nigra, and
locus coeruleus.
Serotonin is synthesized from the amino acid precursor tryptophan, packaged
into vesicles, and released into the synapse following an action potential. Once in the
synapse, serotonin can interact with both the pre- and postsynaptic receptors.
However, immediately after reacting with the pre- and postsynaptic receptors, it is
critically important that serotonin be removed from the synapse.
Re-uptake, the process of removing transmitters after release, determines the
extent, duration, and spatial domain of receptor activation. Any transmitter not
removed from the cleft prevents further signals from getting through. Active removal
reduces the level of transmitter in the cleft faster than diffusion, constrains the effects
of released transmitter to smaller areas, and allows at least part of the released
chemical to be recycled for further use. Re-uptake is carried out by transporter
proteins which bind to the released transmitter and carry it across the plasma
membrane into the pre-synaptic neuron.
116
Temelji neuroznanosti Vodič kroz vježbe
cortex. After the arrival of an action potential, vesicles fuse with the presynaptic
membrane and release serotonin into the synaptic cleft.
Serotonin (5-hydroxytryptamine, 5HT) , a monoamine neurotransmitter, plays
an important role in many behaviors including sleep, appetite, memory, sexual
behavior, neuroendocrine function, and mood. In the brain, the highest level of
serotonin is found in the dorsal and median raphe nuclear complex. Neurons in the
raphe system project beyond the raphe nucleus, most often in the lateral direction.
There are also many serotonergic neurons found in the reticular region of the lower
brain stem. For convenience, neurons in the brain stem are often divided into a caudal
system and a rostral system. Those in the caudal system descend to the spinal chord
along many pathways and are largely involved in sensory, motor, and autonomic
functioning. Serotonergic cells in the rostral system of the brainstem largely terminate
in the dorsal and median raphe nuclei.
Another major serotonergic pathway in the brain is one going from the
cerebellum that terminates in the cerebellar cortex and the cerebellar nuclei. Many
serotonergic neurons contribute to this pathway. The presence of serotonin has also
been observed in the pons, medulla, thalamus, hypothalamus, substantia nigra, and
locus coeruleus.
Serotonin is synthesized from the amino acid precursor tryptophan, packaged
into vesicles, and released into the synapse following an action potential. Once in the
synapse, serotonin can interact with both the pre- and postsynaptic receptors.
However, immediately after reacting with the pre- and postsynaptic receptors, it is
critically important that serotonin be removed from the synapse.
Re-uptake, the process of removing transmitters after release, determines the
extent, duration, and spatial domain of receptor activation. Any transmitter not
removed from the cleft prevents further signals from getting through. Active removal
reduces the level of transmitter in the cleft faster than diffusion, constrains the effects
of released transmitter to smaller areas, and allows at least part of the released
chemical to be recycled for further use. Re-uptake is carried out by transporter
proteins which bind to the released transmitter and carry it across the plasma
membrane into the pre-synaptic neuron.
116

Temelji neuroznanosti Vodič kroz vježbe
IVB2. Location and Structure of the Serotonin Transporter After the serotonin binds to its post-synaptic receptor, an ion dependent
serotonin transporter pumps the serotonin back into the pre-synaptic cell. The
transporter protein is a string of amino acids that passes in and out of the membrane
twelve times.
Transporter proteins are specific to their respective transmitter. In the case of
serotonin, a transporter protein found in the plasma membrane of serotonergic
neurons is responsible for re-uptake of the transmitter. The transporter protein acts as
a carrier of serotonin molecules across the membrane. Unlike channels which stay
open or closed, transporters undergo conformational changes (changes in their three
dimensional shape) and move one molecule in each cycle.
The serotonin transporter (SERT) is similar to other biogenic amine
transporters (e.g. norepinephrine and dopamine transporters) and is part of a family of
sodium (Na+) and chloride (Cl-) dependent transporters. The transporter gene has
been cloned from rat brain, human brain, and human placenta. While there have been
different species of mRNA found for the gene, all SERTs seem to have come from a
common gene. SERTs, with molecular weights of 60-80 kDA, have twelve
transmembrane domains (TM) with a large extracellular loop between TMs 3 and 4.
Both the N- and C-termini are located within the cytoplasm. There are six potential
sites of phosphorylation by protein kinase A and protein kinase C, and regulation of
SERTs by protein kinases has been reported. The large extracellular loop and the
intracellular parts of the N- and C-termini do not appear to be the significant sites that
determine interactions with 5-HT or transporter inhibitors. Rather, the areas important
for selective 5-HT affinity appear to be localized within TMs 1-3 and TMs 8-12. It is
believed that SERTs have a common binding site for 5HT and many of its inhibitors.
SERT activity, like serotonin, is seen most often in the raphe nuclear complex.
SERTs have also been seen in the amygdala, thalamus, hypothalamus, substantia
nigra, and locus coeruleus. In addition to being found on neurons, SERTs are seen in
the placenta, lungs, and blood platelets. Blood platelets utilize SERTs to obtain
serotonin from the environment because they cannot synthesize it themselves. In the
placenta, SERTs may protect heavily vascularized embryonic tissue from constricting
too early due to maternal serotonin.
117
Temelji neuroznanosti Vodič kroz vježbe
IVB2. Location and Structure of the Serotonin Transporter After the serotonin binds to its post-synaptic receptor, an ion dependent
serotonin transporter pumps the serotonin back into the pre-synaptic cell. The
transporter protein is a string of amino acids that passes in and out of the membrane
twelve times.
Transporter proteins are specific to their respective transmitter. In the case of
serotonin, a transporter protein found in the plasma membrane of serotonergic
neurons is responsible for re-uptake of the transmitter. The transporter protein acts as
a carrier of serotonin molecules across the membrane. Unlike channels which stay
open or closed, transporters undergo conformational changes (changes in their three
dimensional shape) and move one molecule in each cycle.
The serotonin transporter (SERT) is similar to other biogenic amine
transporters (e.g. norepinephrine and dopamine transporters) and is part of a family of
sodium (Na+) and chloride (Cl-) dependent transporters. The transporter gene has
been cloned from rat brain, human brain, and human placenta. While there have been
different species of mRNA found for the gene, all SERTs seem to have come from a
common gene. SERTs, with molecular weights of 60-80 kDA, have twelve
transmembrane domains (TM) with a large extracellular loop between TMs 3 and 4.
Both the N- and C-termini are located within the cytoplasm. There are six potential
sites of phosphorylation by protein kinase A and protein kinase C, and regulation of
SERTs by protein kinases has been reported. The large extracellular loop and the
intracellular parts of the N- and C-termini do not appear to be the significant sites that
determine interactions with 5-HT or transporter inhibitors. Rather, the areas important
for selective 5-HT affinity appear to be localized within TMs 1-3 and TMs 8-12. It is
believed that SERTs have a common binding site for 5HT and many of its inhibitors.
SERT activity, like serotonin, is seen most often in the raphe nuclear complex.
SERTs have also been seen in the amygdala, thalamus, hypothalamus, substantia
nigra, and locus coeruleus. In addition to being found on neurons, SERTs are seen in
the placenta, lungs, and blood platelets. Blood platelets utilize SERTs to obtain
serotonin from the environment because they cannot synthesize it themselves. In the
placenta, SERTs may protect heavily vascularized embryonic tissue from constricting
too early due to maternal serotonin.
117

Temelji neuroznanosti Vodič kroz vježbe
IVB3. Mechanism of Action of the Serotonin Transporter The serotonin transporter first binds a sodium ion, followed by serotonin, and
then a chloride ion. The transporter then flips inside the cell, releasing serotonin. A
potassium ion binds, and the transporter flips back out, ready to receive another
serotonin molecule.
Serotonin reuptake transporters (SERTs) are dependent on extracellular Na+
and extracellular Cl-. Unlike Na+, Cl- can be at least partly substituted for by NO2-, Br-
, and other anions. Intracellular potassium (K+) is also used in the process but can be
replaced by other ions, most notably hydrogen (H+). The driving force for the
energetically unfavorable transport of serotonin is the Na+ influx down its
concentration gradient. The Na+/K+ pump (Na+/K+ ATPase) maintains the
extracellular Na+ concentration as well as the intracellular K+ concentration. Na+/K+
ATPase pumps three Na+ ions our for each two K+ ions pumped into the cell. The
electrical potential produced, in addition to creating the Na+ concentration used by the
transporter protein, also leads to the loss of Cl- ions from the cell which are also used
in transport.
According to the present model of SERT function, the first step occurs when
Na+ binds to the carrier protein. Serotonin, in its protonated form (5HT+), then binds
to the transporter followed by Cl-. Chloride ions are not required for 5HT+ binding to
occur but are necessary for net transport to take place. The initial complex of
serotonin, Na+, and Cl- creates a conformational change in the transporter protein. The
protein, which began by facing the outside of the neuron, moves to an inward position
where the neurotransmitter and ions are released into the cytoplasm of the neuron.
Intracellular K+ then binds to the SERT to promote reorientation of the carrier for
another transport cycle. The unoccupied binding site becomes, once again, exposed to
the outside of the cell and the K+ is released outside the cell.
IVB4. Transporter Inhibition: Selective Serotonin Reuptake
Inhibitors Antidepressant drugs also bind to the serotonin transporter. These drugs have a
higher affinity for the transporter, so they block serotonin from binding and the
transporter from flipping inside the presynaptic cell. Excess serotonin in the synaptic
cleft means over-activation of the post-synaptic receptors.
118
Temelji neuroznanosti Vodič kroz vježbe
IVB3. Mechanism of Action of the Serotonin Transporter The serotonin transporter first binds a sodium ion, followed by serotonin, and
then a chloride ion. The transporter then flips inside the cell, releasing serotonin. A
potassium ion binds, and the transporter flips back out, ready to receive another
serotonin molecule.
Serotonin reuptake transporters (SERTs) are dependent on extracellular Na+
and extracellular Cl-. Unlike Na+, Cl- can be at least partly substituted for by NO2-, Br-
, and other anions. Intracellular potassium (K+) is also used in the process but can be
replaced by other ions, most notably hydrogen (H+). The driving force for the
energetically unfavorable transport of serotonin is the Na+ influx down its
concentration gradient. The Na+/K+ pump (Na+/K+ ATPase) maintains the
extracellular Na+ concentration as well as the intracellular K+ concentration. Na+/K+
ATPase pumps three Na+ ions our for each two K+ ions pumped into the cell. The
electrical potential produced, in addition to creating the Na+ concentration used by the
transporter protein, also leads to the loss of Cl- ions from the cell which are also used
in transport.
According to the present model of SERT function, the first step occurs when
Na+ binds to the carrier protein. Serotonin, in its protonated form (5HT+), then binds
to the transporter followed by Cl-. Chloride ions are not required for 5HT+ binding to
occur but are necessary for net transport to take place. The initial complex of
serotonin, Na+, and Cl- creates a conformational change in the transporter protein. The
protein, which began by facing the outside of the neuron, moves to an inward position
where the neurotransmitter and ions are released into the cytoplasm of the neuron.
Intracellular K+ then binds to the SERT to promote reorientation of the carrier for
another transport cycle. The unoccupied binding site becomes, once again, exposed to
the outside of the cell and the K+ is released outside the cell.
IVB4. Transporter Inhibition: Selective Serotonin Reuptake
Inhibitors Antidepressant drugs also bind to the serotonin transporter. These drugs have a
higher affinity for the transporter, so they block serotonin from binding and the
transporter from flipping inside the presynaptic cell. Excess serotonin in the synaptic
cleft means over-activation of the post-synaptic receptors.
118

Temelji neuroznanosti Vodič kroz vježbe
The reuptake process is susceptible to drug manipulation. By blocking the
action of serotonin reuptake inhibitors (SERTs), the amount of serotonin in the
synaptic cleft increases. Selective serotonin reuptake inhibitors (SSRIs) act primarily
at the 5HT transporter protein and have limited, if any, reaction with other
neurotransmitter systems. SSRIs bind to the transporter protein directly and block the
reuptake process. Consequently, more serotonin remains in the cleft where it is free to
travel further to more distant receptors as well as continue to react with nearby
receptors. Like the binding of substrates, antagonist binding to SERTs is also
dependent on extracellular Na+ although ion dependency is different for each SERT
antagonist. It is unclear whether SSRIs bind to the same SERT domain as serotonin or
operate through more indirect mechanisms. Recent evidence suggests that binding of
SSRIs to SERTs occurs at the same site as 5HT binding, but it has not been
determined conclusively.
There are many SSRIs either in the market or in development. SSRIs currently
available include citalopram, fluoxetine (Prozac), fluvoxamine, paroxetine (Paxil),
and sertraline (Zoloft). In addition, while not as selective as the above mentioned,
drugs of abuse such as cocaine, fenfluramine, and (3,4-methylenedioxy)
methamphetamine (MDMA or ecstasy) are inhibitors of serotonin uptake.
IVB5. Clinical Application: Serotonin and Depression The recycling of serotonin is controlled by the serotoin transporter. The
serotonin pathway from the midbrain to the cortex may be an important component in
the regulation of affect.
Selective serotonin reuptake inhibitors (SSRIs) are the focus of extensive
research into finding beneficial pharmacological therapies. There is evidence that
serotonergic pathways are the most closely related systems to mood disorders,
especially depression, and thus SSRIs may lead to significant therapy. Clinical
depression is one of the most common psychiatric disorders, with an incidence of
about 4% and a life-time prevalence of 15-20%. Despite significant research, the
neurobiological dysfunctions of major depression remain elusive. Findings implicate
multiple abnormalities in serotonergic pathways in the cause of depression. Findings
include 1) low concentrations of the major serotonin metabolite; 2) a low density of
brain and platelet serotonin transporters in depressed individuals; 3) a high density of
119
Temelji neuroznanosti Vodič kroz vježbe
The reuptake process is susceptible to drug manipulation. By blocking the
action of serotonin reuptake inhibitors (SERTs), the amount of serotonin in the
synaptic cleft increases. Selective serotonin reuptake inhibitors (SSRIs) act primarily
at the 5HT transporter protein and have limited, if any, reaction with other
neurotransmitter systems. SSRIs bind to the transporter protein directly and block the
reuptake process. Consequently, more serotonin remains in the cleft where it is free to
travel further to more distant receptors as well as continue to react with nearby
receptors. Like the binding of substrates, antagonist binding to SERTs is also
dependent on extracellular Na+ although ion dependency is different for each SERT
antagonist. It is unclear whether SSRIs bind to the same SERT domain as serotonin or
operate through more indirect mechanisms. Recent evidence suggests that binding of
SSRIs to SERTs occurs at the same site as 5HT binding, but it has not been
determined conclusively.
There are many SSRIs either in the market or in development. SSRIs currently
available include citalopram, fluoxetine (Prozac), fluvoxamine, paroxetine (Paxil),
and sertraline (Zoloft). In addition, while not as selective as the above mentioned,
drugs of abuse such as cocaine, fenfluramine, and (3,4-methylenedioxy)
methamphetamine (MDMA or ecstasy) are inhibitors of serotonin uptake.
IVB5. Clinical Application: Serotonin and Depression The recycling of serotonin is controlled by the serotoin transporter. The
serotonin pathway from the midbrain to the cortex may be an important component in
the regulation of affect.
Selective serotonin reuptake inhibitors (SSRIs) are the focus of extensive
research into finding beneficial pharmacological therapies. There is evidence that
serotonergic pathways are the most closely related systems to mood disorders,
especially depression, and thus SSRIs may lead to significant therapy. Clinical
depression is one of the most common psychiatric disorders, with an incidence of
about 4% and a life-time prevalence of 15-20%. Despite significant research, the
neurobiological dysfunctions of major depression remain elusive. Findings implicate
multiple abnormalities in serotonergic pathways in the cause of depression. Findings
include 1) low concentrations of the major serotonin metabolite; 2) a low density of
brain and platelet serotonin transporters in depressed individuals; 3) a high density of
119

Temelji neuroznanosti Vodič kroz vježbe
brain and platelet serotonin binding sites; and 4) a low concentration of tryptophan
which is used in serotonin synthesis. Of these, the low level of SERTs in depressed
patients has received the most attention. Still, while the precise cause of depression
eludes neuroscientists, SSRIs have been shown to alleviate the mood disorder and are
a common therapy for depression.
Fluoxetine, more commonly Prozac, is one antidepressant SSRI that has been
a popular topic in medical as well as social and psychological discussions. Because of
increased selectivity as compared to previous drugs (such as tricyclics and non-
selective MAO inhibitors which have affinity for amines, acetylcholine, and
norepinephrine transporters), SSRIs produce significantly fewer unwelcome side
effects because they work exclusively on serotonin pathways.
Fluoxetine blocks SERT activity immediately, however, therapeutic effects are
seen only after 2-3 weeks of treatment. A patient given Prozac may experience greater
depression before feeling better. Initially, a SERT antagonist blocks the reuptake
process, extracellular serotonin remains elevated and can activate receptors at greater
distances from the presynaptic neuron. High levels of serotonin over an enduring time
period create changes in the amount of pre- and postsynaptic receptors, the amount of
serotonin produced, as well as in the number of SERTs expressed. Neurons may set
the level of SERT expression based on the level of clearance needed. An increase in
serotonin creates a change in SERT expression and consequently effects mood and
other serotonin affected behaviors. It is these compensatory responses at receptors and
transporters that is thought to produce therapeutic effects.
To learn more about depression and the various treatments available, visit
these informative websites: http://www.cmhc.com/guide/depress.htm and
http://depression.cmhc.com.
120
Temelji neuroznanosti Vodič kroz vježbe
brain and platelet serotonin binding sites; and 4) a low concentration of tryptophan
which is used in serotonin synthesis. Of these, the low level of SERTs in depressed
patients has received the most attention. Still, while the precise cause of depression
eludes neuroscientists, SSRIs have been shown to alleviate the mood disorder and are
a common therapy for depression.
Fluoxetine, more commonly Prozac, is one antidepressant SSRI that has been
a popular topic in medical as well as social and psychological discussions. Because of
increased selectivity as compared to previous drugs (such as tricyclics and non-
selective MAO inhibitors which have affinity for amines, acetylcholine, and
norepinephrine transporters), SSRIs produce significantly fewer unwelcome side
effects because they work exclusively on serotonin pathways.
Fluoxetine blocks SERT activity immediately, however, therapeutic effects are
seen only after 2-3 weeks of treatment. A patient given Prozac may experience greater
depression before feeling better. Initially, a SERT antagonist blocks the reuptake
process, extracellular serotonin remains elevated and can activate receptors at greater
distances from the presynaptic neuron. High levels of serotonin over an enduring time
period create changes in the amount of pre- and postsynaptic receptors, the amount of
serotonin produced, as well as in the number of SERTs expressed. Neurons may set
the level of SERT expression based on the level of clearance needed. An increase in
serotonin creates a change in SERT expression and consequently effects mood and
other serotonin affected behaviors. It is these compensatory responses at receptors and
transporters that is thought to produce therapeutic effects.
To learn more about depression and the various treatments available, visit
these informative websites: http://www.cmhc.com/guide/depress.htm and
http://depression.cmhc.com.
120

Temelji neuroznanosti Vodič kroz vježbe
8.3 Pitanja – sinaptička signalizacija
I. Sinteza i pohrana neurotransmitera:
• Objasnite razliku između “gap junctions” i kemijskih sinapsi.
• Ukratko opišite sintezu acetilkolina iz kolina i acetata uz pomoć enzima
• Endorfini…?
II. Otpuštanje neurotransmitera:
• Objasnite ciklus sinaptičkih mjehurića
• Koja je uloga Ca2+ iona?
• Koji proteini sudjeluju u ciklusu sinaptičkih mjehurića i što im je zadaća?
• Objasnite proces egzocitoze i endocitoze sinaptičkih mjehurića.
III. Postinaptički receptori
• U koju skupinu receptora spadaju GABAa receptori?
• Jeli njihova uloga ekscitacijska ili inhibicijska?
• Objasnite strukturu i mehanizam djelovanja GABAa receptora.
• Što znate o kliničkoj aplikaciji agonista GABAa receptora.
• Koji je važan predstavnik metabotropnih receptora?
• Objasnite funkciju G-proteina.
IV. Inaktivacija neurotransmitera
• Koje sve načine inaktivacije neurotransmitera poznajete?
• Objasnite ulogu acetilkolintransferaze.
• Objasnite mehanizam “reuptake” tj ponovnog preuzimanja na primjeru
serotonina.
121
Temelji neuroznanosti Vodič kroz vježbe
8.3 Pitanja – sinaptička signalizacija
I. Sinteza i pohrana neurotransmitera:
• Objasnite razliku između “gap junctions” i kemijskih sinapsi.
• Ukratko opišite sintezu acetilkolina iz kolina i acetata uz pomoć enzima
• Endorfini…?
II. Otpuštanje neurotransmitera:
• Objasnite ciklus sinaptičkih mjehurića
• Koja je uloga Ca2+ iona?
• Koji proteini sudjeluju u ciklusu sinaptičkih mjehurića i što im je zadaća?
• Objasnite proces egzocitoze i endocitoze sinaptičkih mjehurića.
III. Postinaptički receptori
• U koju skupinu receptora spadaju GABAa receptori?
• Jeli njihova uloga ekscitacijska ili inhibicijska?
• Objasnite strukturu i mehanizam djelovanja GABAa receptora.
• Što znate o kliničkoj aplikaciji agonista GABAa receptora.
• Koji je važan predstavnik metabotropnih receptora?
• Objasnite funkciju G-proteina.
IV. Inaktivacija neurotransmitera
• Koje sve načine inaktivacije neurotransmitera poznajete?
• Objasnite ulogu acetilkolintransferaze.
• Objasnite mehanizam “reuptake” tj ponovnog preuzimanja na primjeru
serotonina.
121

Temelji neuroznanosti Vodič kroz vježbe
9. Fiziologija osjeta
9.1 Uvod
Cilj vježbe je približiti studentima pojam osjetila, osjetnih i njima sličnih
fenomena te osjetnih iluzija.
Postoji pet osnovnih osjetila: za vid, sluh, okus, miris i dodir (te stoga i osjeti
vida, sluha, okusa, mirisa i dodira). To je pojednostavljeni model. Dodatni osjetni
modaliteti uključuju i osjet temperature, bola, vibracije, položaja zglobova i
propriocepcije.
Mnogi kratki, ali informativni pokusi iz fiziologije osjeta, poput onih
prikazanih u ovoj vježbi, mogu se izvesti vrlo jednostavnom opremom i predmetima
iz svakodnevne uporabe.
Oprema potrebna za vježbu uključuje:
o igle i spajalice za papir
o bijeli i crni papir te olovku
o malu baterijsku lampu
o veliku laboratorijsku posudu
o hladnu, vruću i mlaku vodu
o štipalicu za nos
o jabuku, sirovi krumpir i sirovi crveni luk
o šećer, kuhinjsku sol i limunsku kiselinu
o 3 čaše
o štapiće s vrhom od pamuka
o rotirajuću stolicu
Uporaba navedene opreme kao i sve daljnje pripreme bit će objašnjene tijekom
vježbe.
122
Temelji neuroznanosti Vodič kroz vježbe
9. Fiziologija osjeta
9.1 Uvod
Cilj vježbe je približiti studentima pojam osjetila, osjetnih i njima sličnih
fenomena te osjetnih iluzija.
Postoji pet osnovnih osjetila: za vid, sluh, okus, miris i dodir (te stoga i osjeti
vida, sluha, okusa, mirisa i dodira). To je pojednostavljeni model. Dodatni osjetni
modaliteti uključuju i osjet temperature, bola, vibracije, položaja zglobova i
propriocepcije.
Mnogi kratki, ali informativni pokusi iz fiziologije osjeta, poput onih
prikazanih u ovoj vježbi, mogu se izvesti vrlo jednostavnom opremom i predmetima
iz svakodnevne uporabe.
Oprema potrebna za vježbu uključuje:
o igle i spajalice za papir
o bijeli i crni papir te olovku
o malu baterijsku lampu
o veliku laboratorijsku posudu
o hladnu, vruću i mlaku vodu
o štipalicu za nos
o jabuku, sirovi krumpir i sirovi crveni luk
o šećer, kuhinjsku sol i limunsku kiselinu
o 3 čaše
o štapiće s vrhom od pamuka
o rotirajuću stolicu
Uporaba navedene opreme kao i sve daljnje pripreme bit će objašnjene tijekom
vježbe.
122

Temelji neuroznanosti Vodič kroz vježbe
9.2 Binokularni vid
Binokularni vid zahtijeva da se odvojene slike lijevog i desnog oka "fuziraju" i
daju jedinstven pogled. Fuzija, tj. spajanje slika nekog objekta, moguća je jedino
ukoliko slike padaju na odgovarajući dio lijeve i desne retine. Ukoliko se to ipak ne
dogodi, rezultat će biti dvoslika promatranog objekta.
Postupak:
o Držite jednu ruku ispruženu i podignite uspravni kažiprst u razinu s
nekim udaljenim predmetom (npr. sat na zidu u blizini). Pogledajte
svoj prst (zadržite prst u fokusu), ali se pokušajte koncentrirati na
udaljeni predmet. Zamijetite da se udaljeni predmet vidi dvostruko -
postoje dvije slike jedna uz drugu.
o Pokrijte desno oko. Zamijetite kako slika udaljenog predmeta s desne
strane nestaje.
o Pogledajte s oba otvorena oka udaljeni predmet. Zamijetite kako sada
vidite dvostruku sliku prsta.
o Pokrijte desno oko i zamijetite kako nestaje lijeva slika vašega prsta.
o Zamolite dragovoljca da prvo pogleda udaljeni predmet, a potom drugi
predmet na neposrednoj udaljenosti od lica, otprilike 15 cm. Zamijetite
kako se prilikom gledanja drugog predmeta oči ispitanika okreću
prema unutra.
9.3 Akomodacija (fokusiranje)
Oko se može akomodirati (promijeniti fokus) tj. prilagoditi za gledanje bliskih
i udaljenih predmeta, promjenom oblika očne leće.
o Pokrijte ili zatvorite jedno oko i držite iglu 15 cm ispred drugog oka u
razini s nekim udaljenim predmetom.
o Pogledajte u udaljeni predmet i zamijetite kako se pojavljuje slika igle,
mutna i nejasna (nalazi se izvan fokusa).
o Sada pogledajte iglu. Zamijetite kako je sada slika udaljenog predmeta
postala mutna i nejasna. Zamijetite isto tako da akomodacija oka za
123
Temelji neuroznanosti Vodič kroz vježbe
9.2 Binokularni vid
Binokularni vid zahtijeva da se odvojene slike lijevog i desnog oka "fuziraju" i
daju jedinstven pogled. Fuzija, tj. spajanje slika nekog objekta, moguća je jedino
ukoliko slike padaju na odgovarajući dio lijeve i desne retine. Ukoliko se to ipak ne
dogodi, rezultat će biti dvoslika promatranog objekta.
Postupak:
o Držite jednu ruku ispruženu i podignite uspravni kažiprst u razinu s
nekim udaljenim predmetom (npr. sat na zidu u blizini). Pogledajte
svoj prst (zadržite prst u fokusu), ali se pokušajte koncentrirati na
udaljeni predmet. Zamijetite da se udaljeni predmet vidi dvostruko -
postoje dvije slike jedna uz drugu.
o Pokrijte desno oko. Zamijetite kako slika udaljenog predmeta s desne
strane nestaje.
o Pogledajte s oba otvorena oka udaljeni predmet. Zamijetite kako sada
vidite dvostruku sliku prsta.
o Pokrijte desno oko i zamijetite kako nestaje lijeva slika vašega prsta.
o Zamolite dragovoljca da prvo pogleda udaljeni predmet, a potom drugi
predmet na neposrednoj udaljenosti od lica, otprilike 15 cm. Zamijetite
kako se prilikom gledanja drugog predmeta oči ispitanika okreću
prema unutra.
9.3 Akomodacija (fokusiranje)
Oko se može akomodirati (promijeniti fokus) tj. prilagoditi za gledanje bliskih
i udaljenih predmeta, promjenom oblika očne leće.
o Pokrijte ili zatvorite jedno oko i držite iglu 15 cm ispred drugog oka u
razini s nekim udaljenim predmetom.
o Pogledajte u udaljeni predmet i zamijetite kako se pojavljuje slika igle,
mutna i nejasna (nalazi se izvan fokusa).
o Sada pogledajte iglu. Zamijetite kako je sada slika udaljenog predmeta
postala mutna i nejasna. Zamijetite isto tako da akomodacija oka za
123

Temelji neuroznanosti Vodič kroz vježbe
gledanje bliskih predmeta (u ovom primjeru igle) biva praćena
osjećajem napora.
o Pokrijte jedno oko i držite iglu u razini ruke. Dok gledate u vršak igle,
polako primičite iglu licu sve dok slika postane nejasna. Najkraća
udaljenost na kojoj maksimalno akomodirano oko još uvijek oštro
razabire ono što gleda (u ovom primjeru igla) naziva se bliska točka.
9.4 Slijepa pjega
Sastavni dio vidnog polja oba oka je slijepa pjega koja predstavlja optički
disk - dio mrežnice bez fotoreceptora.
o Promatrajte olovku koja piše crnom tintom, a ima bijeli držač.
Omotajte bijelim papirom držač olovke te ostavite nepokriven samo
crni dio za pisanje.
o Zabilježite križić na bijelom papiru. Zatvorite lijevo oko i mirno
gledajte križić na udaljenosti od 25cm. U nastavku te vježbe nastojte
glavu držati mirno i nastavite gledati u križić.
o Pomičite olovku udesno od križića. Kad dosegnete stanovitu udaljenost
vrh olovke postat će nevidljiv. Označite tu točku na papiru.
o Pomičite olovku i dalje udesno dok ponovno ne postane nevidljiva.
Obilježite i tu točku.
o Obilježite jednako tako gornju i donju granicu slijepe pjege.
Pažljivim i nadasve strpljivim radom moguće je odrediti granice slijepe pjege
u cijelosti. Slijepa pjega ima ovalan oblik i nepravilne rubove. Nepravilnosti
predstavljaju mjesta gdje mrežnične krvne žile izlaze iz optičkog diska.
124
Temelji neuroznanosti Vodič kroz vježbe
gledanje bliskih predmeta (u ovom primjeru igle) biva praćena
osjećajem napora.
o Pokrijte jedno oko i držite iglu u razini ruke. Dok gledate u vršak igle,
polako primičite iglu licu sve dok slika postane nejasna. Najkraća
udaljenost na kojoj maksimalno akomodirano oko još uvijek oštro
razabire ono što gleda (u ovom primjeru igla) naziva se bliska točka.
9.4 Slijepa pjega
Sastavni dio vidnog polja oba oka je slijepa pjega koja predstavlja optički
disk - dio mrežnice bez fotoreceptora.
o Promatrajte olovku koja piše crnom tintom, a ima bijeli držač.
Omotajte bijelim papirom držač olovke te ostavite nepokriven samo
crni dio za pisanje.
o Zabilježite križić na bijelom papiru. Zatvorite lijevo oko i mirno
gledajte križić na udaljenosti od 25cm. U nastavku te vježbe nastojte
glavu držati mirno i nastavite gledati u križić.
o Pomičite olovku udesno od križića. Kad dosegnete stanovitu udaljenost
vrh olovke postat će nevidljiv. Označite tu točku na papiru.
o Pomičite olovku i dalje udesno dok ponovno ne postane nevidljiva.
Obilježite i tu točku.
o Obilježite jednako tako gornju i donju granicu slijepe pjege.
Pažljivim i nadasve strpljivim radom moguće je odrediti granice slijepe pjege
u cijelosti. Slijepa pjega ima ovalan oblik i nepravilne rubove. Nepravilnosti
predstavljaju mjesta gdje mrežnične krvne žile izlaze iz optičkog diska.
124

Temelji neuroznanosti Vodič kroz vježbe
9.5 Mehanička stimulacija mrežnice
Oko je sastavljeno od elemenata sličnih onima kod kamere te je slika koja
nastaje na mrežnici obrnuta. Svjetlost koja dolazi na mrežnicu s jedne strane oka daje
vizualni odgovor na suprotnoj strani vidnog polja. Mehanička stimulacija mrežnice,
pritiskom na očnu jabučicu, jednako tako daje obrnuti vizualni odgovor.
o Usmjerite pogled ulijevo i zatvorite oba oka. Nastavite gledati ulijevo.
o Vrškom prsta nježno pritisnite desnu stranu desne očne jabučice.
Zamijetite vidni učinak što pri tome nastaje.
o Klizite lagano prstom prema gore i dolje te zamijetite smjer pomaka
vizualnog odgovora.
o Usmjerite pogled udesno i slično pritisnite lijevu stranu lijeve očne
jabučice, u lijevom kutu oka. Zamijetite nastali vidni učinak.
Trebali biste zamijetiti da je glavni odgovor na podražaj svijetli krug ili disk,
što nastaje na suprotnoj strani vidnog polja od one na kojoj je učinjen podražaj.
Podraživanje mrežnice na desnoj strani oka daje odgovor na lijevoj strani i obrnuto.
9.6 Pozitivna naknadna slika
Mrežnični fotoreceptori imaju iznenađujuće dugačak i spor odgovor na
svjetlosni podražaj. Kratkotrajan vidni podražaj uzrokuje odgovor koji traje duže od
same stimulacije dovoljno za nastanak pozitivne naknadne slike.
o Pogledajte u svijetlu i kontrastnu površinu kao što je prozor okrenut
suncu ili u jako osvijetljenu površinu radnog stola.
o Zatvorite oba oka i pokrijte ih rukama. Sačekajte 30 sekundi.
o Odmaknite ruke, otvorite oči za najkraće moguće vrijeme te ih
ponovno zatvorite.
o Zamijetite naknadnu sliku. Svijetli izgled površine koju ste prethodno
gledali ostaje vidljiv kroz kratko, ali mjerljivo vrijeme (značajan dio
sekunde).
125
Temelji neuroznanosti Vodič kroz vježbe
9.5 Mehanička stimulacija mrežnice
Oko je sastavljeno od elemenata sličnih onima kod kamere te je slika koja
nastaje na mrežnici obrnuta. Svjetlost koja dolazi na mrežnicu s jedne strane oka daje
vizualni odgovor na suprotnoj strani vidnog polja. Mehanička stimulacija mrežnice,
pritiskom na očnu jabučicu, jednako tako daje obrnuti vizualni odgovor.
o Usmjerite pogled ulijevo i zatvorite oba oka. Nastavite gledati ulijevo.
o Vrškom prsta nježno pritisnite desnu stranu desne očne jabučice.
Zamijetite vidni učinak što pri tome nastaje.
o Klizite lagano prstom prema gore i dolje te zamijetite smjer pomaka
vizualnog odgovora.
o Usmjerite pogled udesno i slično pritisnite lijevu stranu lijeve očne
jabučice, u lijevom kutu oka. Zamijetite nastali vidni učinak.
Trebali biste zamijetiti da je glavni odgovor na podražaj svijetli krug ili disk,
što nastaje na suprotnoj strani vidnog polja od one na kojoj je učinjen podražaj.
Podraživanje mrežnice na desnoj strani oka daje odgovor na lijevoj strani i obrnuto.
9.6 Pozitivna naknadna slika
Mrežnični fotoreceptori imaju iznenađujuće dugačak i spor odgovor na
svjetlosni podražaj. Kratkotrajan vidni podražaj uzrokuje odgovor koji traje duže od
same stimulacije dovoljno za nastanak pozitivne naknadne slike.
o Pogledajte u svijetlu i kontrastnu površinu kao što je prozor okrenut
suncu ili u jako osvijetljenu površinu radnog stola.
o Zatvorite oba oka i pokrijte ih rukama. Sačekajte 30 sekundi.
o Odmaknite ruke, otvorite oči za najkraće moguće vrijeme te ih
ponovno zatvorite.
o Zamijetite naknadnu sliku. Svijetli izgled površine koju ste prethodno
gledali ostaje vidljiv kroz kratko, ali mjerljivo vrijeme (značajan dio
sekunde).
125

Temelji neuroznanosti Vodič kroz vježbe
9.7 Negativna naknadna slika
Osjetljivost mrežničnih fotoreceptora raste postupno dok su podraženi
svjetlošću, a smanjuje se dok nisu podraženi. Adaptacija na svjetlo i tamu dopušta
funkciju vida u velikom rasponu svjetlosnih intenziteta. To ima i sporedni učinak koji
se ogleda u nastanku negativnih naknadnih slika.
o Postavite crni predmet na komad bijelog papira ili nacrtajte crni
kvadrat na papiru.
o Fiksirajte pogled na crni predmet 30 sekundi. Možete trepnuti, ali
nastojte što duže zadržati pogled fiksiranim.
o Premjestite pogled na komad ravnog bijelog papira i zamijetite
naknadnu sliku crnog predmeta. Slika traje nekoliko sekundi. Slika je
obrnuti kontrast (crni predmet daje svijetlu naknadnu sliku), odatle i
ime "negativna naknadna slika".
o Ponovite isto korištenjem obojanog predmeta i zamijetite promjenu
boje naknadne slike. Navodimo primjer: crveni predmet daje zelenu
naknadnu sliku.
9.8 Mrežnične krvne žile
Mrežnične krvne žile nalaze se u prednjem dijelu neuralnog i fotosenzitivnog
sloja. Apsorpcija i rasprostranjenost svjetla u krvnim žilama trebala bi prouzročiti
nastajanje slike. Slika je normalno suprimirana, ali mehanizmom koji nije dovoljno
razjašnjen. Mijenjanje smjera osvjetljenja krvnih žila privremeno nadvlada supresiju i
stvara sliku vidljivom. Dostatno osvjetljenje može ući u oko kroz bjeloočnicu, ukoliko
se malen izvor svjetla drži blizu, i to tako da malo svjetla ulazi kroz zjenicu.
o Zatvorite oba oka i usmjerite pogled ulijevo. Osvijetlite malom
baterijskom svjetiljkom kapak na desnoj strani desnog oka. Zadržite
svjetiljku u blizini oka tako da stvori malu svijetlu točku.
o Obrazac poput čipkaste mreže morao bi postati vidljiv za kratko
vrijeme, nakon čega počinje blijediti. Polaganim pomicanjem svjetla
126
Temelji neuroznanosti Vodič kroz vježbe
9.7 Negativna naknadna slika
Osjetljivost mrežničnih fotoreceptora raste postupno dok su podraženi
svjetlošću, a smanjuje se dok nisu podraženi. Adaptacija na svjetlo i tamu dopušta
funkciju vida u velikom rasponu svjetlosnih intenziteta. To ima i sporedni učinak koji
se ogleda u nastanku negativnih naknadnih slika.
o Postavite crni predmet na komad bijelog papira ili nacrtajte crni
kvadrat na papiru.
o Fiksirajte pogled na crni predmet 30 sekundi. Možete trepnuti, ali
nastojte što duže zadržati pogled fiksiranim.
o Premjestite pogled na komad ravnog bijelog papira i zamijetite
naknadnu sliku crnog predmeta. Slika traje nekoliko sekundi. Slika je
obrnuti kontrast (crni predmet daje svijetlu naknadnu sliku), odatle i
ime "negativna naknadna slika".
o Ponovite isto korištenjem obojanog predmeta i zamijetite promjenu
boje naknadne slike. Navodimo primjer: crveni predmet daje zelenu
naknadnu sliku.
9.8 Mrežnične krvne žile
Mrežnične krvne žile nalaze se u prednjem dijelu neuralnog i fotosenzitivnog
sloja. Apsorpcija i rasprostranjenost svjetla u krvnim žilama trebala bi prouzročiti
nastajanje slike. Slika je normalno suprimirana, ali mehanizmom koji nije dovoljno
razjašnjen. Mijenjanje smjera osvjetljenja krvnih žila privremeno nadvlada supresiju i
stvara sliku vidljivom. Dostatno osvjetljenje može ući u oko kroz bjeloočnicu, ukoliko
se malen izvor svjetla drži blizu, i to tako da malo svjetla ulazi kroz zjenicu.
o Zatvorite oba oka i usmjerite pogled ulijevo. Osvijetlite malom
baterijskom svjetiljkom kapak na desnoj strani desnog oka. Zadržite
svjetiljku u blizini oka tako da stvori malu svijetlu točku.
o Obrazac poput čipkaste mreže morao bi postati vidljiv za kratko
vrijeme, nakon čega počinje blijediti. Polaganim pomicanjem svjetla
126

Temelji neuroznanosti Vodič kroz vježbe
slika ponovno nastaje. Ukoliko nastavite ritmično pomicati izvor
svjetla, slika će ostati vidljiva.
o Krvne žile jasnije se mogu vidjeti ukoliko oči držite otvorenima, a
svjetlost usmjerite izravno prema bjeloočnici. Takav pokus zahtijeva
izvođenje u tamnoj sobi, ili ispitanik mora gledati nekakav bezličan
prizor poput ravne tamno obojane podloge ili zida.
o Strpljivim promatranjem postaje jasno vidljivo grananje i vijugav tok
krvnih žila. Središte vidnog polja ne sadrži krvne žile. Krvne žile
približavaju se radijalnim smjerom iz vanjskih dijelova polja.
o Većina ljudi može vidjeti izvorno grananje krvnih žila u području
desno od središta. To područje nazivamo optički disk ili pločica vidnog
živca (vidite vježbu 10.3).
9.9 Razlikovanje dviju točaka
Gustoća taktilnih receptora kože uvelike se razlikuje u različitim dijelovima
tijela.
o Uzmite metalnu spajalicu za papir i raširite je. Savijte je u oblik slova
U tako da razmak raširenih točaka bude oko 1 cm.
o Dotaknite lagano dvije točke na dlanu ispružene ispitanikove ruke.
Zamolite ga da Vam kaže je li osjetio jednu ili dvije točke dodira. Uz
razmak od 1 cm, lako se mogu razlikovati dva podražaja kao takva.
o Zamolite ispitanika da zatvori oči. Savijte spajalicu tako da vrhovi
budu bliže jedan drugom. Ponavljanjem pokušaja s različitim
razmakom vrhova spajalice pronađite najmanju udaljenost na kojoj
ispitanik može razlikovati dvije točke. Možete ispitati istinitost
ispitanikovih odgovora s vremena na vrijeme, ukošenjem spajalice i
pritiskanjem samo jedne točke.
o Izmjerite udaljenost dviju točaka ravnalom.
o Ponavljte korake 3 i 4 na različitim dijelovima tijela (npr. vršak prsta,
stražnji dio šake, stražnji dio podlaktice, na leđima i sl.).
127
Temelji neuroznanosti Vodič kroz vježbe
slika ponovno nastaje. Ukoliko nastavite ritmično pomicati izvor
svjetla, slika će ostati vidljiva.
o Krvne žile jasnije se mogu vidjeti ukoliko oči držite otvorenima, a
svjetlost usmjerite izravno prema bjeloočnici. Takav pokus zahtijeva
izvođenje u tamnoj sobi, ili ispitanik mora gledati nekakav bezličan
prizor poput ravne tamno obojane podloge ili zida.
o Strpljivim promatranjem postaje jasno vidljivo grananje i vijugav tok
krvnih žila. Središte vidnog polja ne sadrži krvne žile. Krvne žile
približavaju se radijalnim smjerom iz vanjskih dijelova polja.
o Većina ljudi može vidjeti izvorno grananje krvnih žila u području
desno od središta. To područje nazivamo optički disk ili pločica vidnog
živca (vidite vježbu 10.3).
9.9 Razlikovanje dviju točaka
Gustoća taktilnih receptora kože uvelike se razlikuje u različitim dijelovima
tijela.
o Uzmite metalnu spajalicu za papir i raširite je. Savijte je u oblik slova
U tako da razmak raširenih točaka bude oko 1 cm.
o Dotaknite lagano dvije točke na dlanu ispružene ispitanikove ruke.
Zamolite ga da Vam kaže je li osjetio jednu ili dvije točke dodira. Uz
razmak od 1 cm, lako se mogu razlikovati dva podražaja kao takva.
o Zamolite ispitanika da zatvori oči. Savijte spajalicu tako da vrhovi
budu bliže jedan drugom. Ponavljanjem pokušaja s različitim
razmakom vrhova spajalice pronađite najmanju udaljenost na kojoj
ispitanik može razlikovati dvije točke. Možete ispitati istinitost
ispitanikovih odgovora s vremena na vrijeme, ukošenjem spajalice i
pritiskanjem samo jedne točke.
o Izmjerite udaljenost dviju točaka ravnalom.
o Ponavljte korake 3 i 4 na različitim dijelovima tijela (npr. vršak prsta,
stražnji dio šake, stražnji dio podlaktice, na leđima i sl.).
127

Temelji neuroznanosti Vodič kroz vježbe
9.10 Dodirna iluzija
Prekrižite dva susjedna prsta, tako da nokti budu jedan uz drugoga, ali u
položaju suprotnom od normalnog. Većini ljudi najlakše je prekrižiti srednjak i
kažiprst.
Stavite malen predmet poput olovke u prostor između noktiju u obliku slova V
i lagano ga pomičite naprijed - natrag.
Pojavit će se očita iluzija u kojoj osjećate kako u ruci imate dva predmeta.
9.11 Temperaturna iluzija
Mnogi osjetni sustavi pokazuju adaptaciju: opadanje odgovora kod
kontinuiranog jednakomjernog podražaja. Temperaturni receptori u koži pokazuju
adaptaciju tako da osjećaj topline za toplo i hladno više ovisi o promjenama
temperature nego o samoj temperaturi.
o Napunite tri posude vodom. Jednu napunite vrućom vodom, ali ne tako
vrućom vodom da biste se mogli opeći. Drugu napunite hladnom, a
treću mlakom vodom.
o Stavite jednu ruku u hladnu vodu, drugu u vruću vodu. Zadržite ih tako
30 sekundi.
o Potom stavite obje ruke u mlaku vodu. Vodu ćete osjetiti kao toplu
jednom, a kao hladnu drugom rukom.
9.12 Okus i miris
Veliki dio osjeta koji se uobičajeno naziva "okusom", u stvari je miris.
o Zamolite dobrovoljca da zatvori oči i drži obje nosnice zatvorenima
tako da zrak kroz njih ne može prolaziti.
o Prethodno pripremljeni komad jabuke dajte ispitaniku u usta te ga
zamolite da identificira što mu je u ustima prema okusu.
o Ponovite to isto sa svježim krumpirom, a zatim sa svježim lukom.
Razlikovanje je izuzetno teško.
128
Temelji neuroznanosti Vodič kroz vježbe
9.10 Dodirna iluzija
Prekrižite dva susjedna prsta, tako da nokti budu jedan uz drugoga, ali u
položaju suprotnom od normalnog. Većini ljudi najlakše je prekrižiti srednjak i
kažiprst.
Stavite malen predmet poput olovke u prostor između noktiju u obliku slova V
i lagano ga pomičite naprijed - natrag.
Pojavit će se očita iluzija u kojoj osjećate kako u ruci imate dva predmeta.
9.11 Temperaturna iluzija
Mnogi osjetni sustavi pokazuju adaptaciju: opadanje odgovora kod
kontinuiranog jednakomjernog podražaja. Temperaturni receptori u koži pokazuju
adaptaciju tako da osjećaj topline za toplo i hladno više ovisi o promjenama
temperature nego o samoj temperaturi.
o Napunite tri posude vodom. Jednu napunite vrućom vodom, ali ne tako
vrućom vodom da biste se mogli opeći. Drugu napunite hladnom, a
treću mlakom vodom.
o Stavite jednu ruku u hladnu vodu, drugu u vruću vodu. Zadržite ih tako
30 sekundi.
o Potom stavite obje ruke u mlaku vodu. Vodu ćete osjetiti kao toplu
jednom, a kao hladnu drugom rukom.
9.12 Okus i miris
Veliki dio osjeta koji se uobičajeno naziva "okusom", u stvari je miris.
o Zamolite dobrovoljca da zatvori oči i drži obje nosnice zatvorenima
tako da zrak kroz njih ne može prolaziti.
o Prethodno pripremljeni komad jabuke dajte ispitaniku u usta te ga
zamolite da identificira što mu je u ustima prema okusu.
o Ponovite to isto sa svježim krumpirom, a zatim sa svježim lukom.
Razlikovanje je izuzetno teško.
128

Temelji neuroznanosti Vodič kroz vježbe
o Ponovite korake 2 i 3, ali ovoga puta dozvolite ispitaniku disanje kroz
nos. Sad je razlikovanje vrlo lagano.
9.13 Okus i raspodjela
Okusni receptori ("okusni pupoljci") uglavnom su pronađeni na jeziku, no
jednako tako i na nepcu te u farinksu. Razlikuju se četiri vrste okusa: slatko, kiselo,
slano i gorko. Svaka vrsta okusnih pupoljaka ima karakterističnu prostornu
rasprostranjenost na jeziku.
o Pripremite u plastične čaše sljedeće otopine.
• šećer (saharozu): 15 grama u 50 ml vode
• kuhinjsku sol (natrijev klorid): 5g u 50 ml vode
• limunsku kiselinu: 2 g u 50 ml vode
o Navlažite mali komadić čistog pamuka prvom otopinom, odstranite
suvišak tekućine i dotaknite pamukom stražnji dio ispitanikova jezika.
Zamolite ispitanika da Vam kaže što osjeća. Ponovite isto samo sada
dotaknite pamukom jednu stranu jezika, te potom vršak jezika.
Zamijetit ćete kako upravo vršak jezika daje najintenzivniji osjećaj
slatkog.
o Ponovite korak 2, ali ovaj put sa slanom otopinom. Zamijetite različitu
distribuciju osjeta za slano.
o Ponovite korak 3, ali ovaj put s otopinom limunske kiseline. Zamijetite
različitu distribuciju osjeta za kiselo.
129
Temelji neuroznanosti Vodič kroz vježbe
o Ponovite korake 2 i 3, ali ovoga puta dozvolite ispitaniku disanje kroz
nos. Sad je razlikovanje vrlo lagano.
9.13 Okus i raspodjela
Okusni receptori ("okusni pupoljci") uglavnom su pronađeni na jeziku, no
jednako tako i na nepcu te u farinksu. Razlikuju se četiri vrste okusa: slatko, kiselo,
slano i gorko. Svaka vrsta okusnih pupoljaka ima karakterističnu prostornu
rasprostranjenost na jeziku.
o Pripremite u plastične čaše sljedeće otopine.
• šećer (saharozu): 15 grama u 50 ml vode
• kuhinjsku sol (natrijev klorid): 5g u 50 ml vode
• limunsku kiselinu: 2 g u 50 ml vode
o Navlažite mali komadić čistog pamuka prvom otopinom, odstranite
suvišak tekućine i dotaknite pamukom stražnji dio ispitanikova jezika.
Zamolite ispitanika da Vam kaže što osjeća. Ponovite isto samo sada
dotaknite pamukom jednu stranu jezika, te potom vršak jezika.
Zamijetit ćete kako upravo vršak jezika daje najintenzivniji osjećaj
slatkog.
o Ponovite korak 2, ali ovaj put sa slanom otopinom. Zamijetite različitu
distribuciju osjeta za slano.
o Ponovite korak 3, ali ovaj put s otopinom limunske kiseline. Zamijetite
različitu distribuciju osjeta za kiselo.
129

Temelji neuroznanosti Vodič kroz vježbe
9.14 Osjećaj pritiska
Kapsule i ligamenti zglobova zaprimaju osjetnu inervaciju čime imaju
mogućnost zamjećivanja promjene položaja zglobova. Učinkovitost toga manje
poznatog osjeta kroz vježbu ćemo jednostavno demonstrirati.
o Zamolite dobrovoljca da ispruži ruku dlanom okrenutim prema gore i
ispruženim prstima.
o Držite ispitanikov kažiprst svojim palcem s jedne, a kažiprstom s druge
strane. Nemojte držati ispitanikov kažiprst s dorzalne i palmarne
strane, s obzirom da bi to moglo dati interpretaciju pokreta izazvanu
silama povlačenja prema dolje ili podizanja.
o Savijte ispitanikov prst prema gore i istovremeno recite: "Ovo je gore".
Sada potegnite prst prema dolje u početni položaj i istovremeno recite:
"Ovo je dolje".
o Zamolite ispitanika da zatvori oči. Sada testirajte njegovu/njezinu
sposobnost zamjećivanja promjene položaja prsta. Pokušajte izvesti
veće i manje pomake prsta.
9.15 Polukružni kanali
Polukružni kanali ispunjeni su tekućinom te su smješteni u temporalnoj kosti
lubanje i čine dio unutarnjeg uha. Imaju mogućnost zamjećivanja rotacijskih pokreta
glave u tri osovine.
Za izvođenje vježbe potrebna je rotacijska stolica (mora se moći rotirati lagano
u svojoj vertikalnoj osovini) npr. uredska stolica.
o Zamolite dobrovoljca da sjedne s objema nogama u zraku te da zatvori
oči.
o Zamolite ispitanika da Vam kaže kad počinje osjećati rotaciju i u
kojemu smjeru. Ispitajte njegovu mogućnost zamjećivanja rotacijskih
pokreta tako da rotirate stolicu različitim brzinama i u različitim
smjerovima. Trebali biste zamijetiti kako je moguće osjetiti i vrlo
lagane pomake.
130
Temelji neuroznanosti Vodič kroz vježbe
9.14 Osjećaj pritiska
Kapsule i ligamenti zglobova zaprimaju osjetnu inervaciju čime imaju
mogućnost zamjećivanja promjene položaja zglobova. Učinkovitost toga manje
poznatog osjeta kroz vježbu ćemo jednostavno demonstrirati.
o Zamolite dobrovoljca da ispruži ruku dlanom okrenutim prema gore i
ispruženim prstima.
o Držite ispitanikov kažiprst svojim palcem s jedne, a kažiprstom s druge
strane. Nemojte držati ispitanikov kažiprst s dorzalne i palmarne
strane, s obzirom da bi to moglo dati interpretaciju pokreta izazvanu
silama povlačenja prema dolje ili podizanja.
o Savijte ispitanikov prst prema gore i istovremeno recite: "Ovo je gore".
Sada potegnite prst prema dolje u početni položaj i istovremeno recite:
"Ovo je dolje".
o Zamolite ispitanika da zatvori oči. Sada testirajte njegovu/njezinu
sposobnost zamjećivanja promjene položaja prsta. Pokušajte izvesti
veće i manje pomake prsta.
9.15 Polukružni kanali
Polukružni kanali ispunjeni su tekućinom te su smješteni u temporalnoj kosti
lubanje i čine dio unutarnjeg uha. Imaju mogućnost zamjećivanja rotacijskih pokreta
glave u tri osovine.
Za izvođenje vježbe potrebna je rotacijska stolica (mora se moći rotirati lagano
u svojoj vertikalnoj osovini) npr. uredska stolica.
o Zamolite dobrovoljca da sjedne s objema nogama u zraku te da zatvori
oči.
o Zamolite ispitanika da Vam kaže kad počinje osjećati rotaciju i u
kojemu smjeru. Ispitajte njegovu mogućnost zamjećivanja rotacijskih
pokreta tako da rotirate stolicu različitim brzinama i u različitim
smjerovima. Trebali biste zamijetiti kako je moguće osjetiti i vrlo
lagane pomake.
130

Temelji neuroznanosti Vodič kroz vježbe
Polukružni kanalići zamjećuju rotaciju, ali nisu u mogućnosti signalizirati
položaj tijela. To možete ispitati tako da pokažete kako ispitanikov osjećaj u kojem se
smjeru okreće postaje nepouzdan, primjerice prilikom okretanja za jednu četvrtinu
kruga u jednom smjeru, a zatim za pola kruga u drugom smjeru. Međutim test nije
jednostavno izvesti jer se ispitanik može služiti drugim informacijama o smjeru
kretanja. Intenzitet osvjetljenja u prostoru može se mijenjati s rotacijama što se može
zamijetiti i kroz zatvorene kapke. Jednako tako i akustično okruženje nije
ravnomjerno. Trebalo bi obratiti pozornost na navedene čimbenike i pokušati umanjiti
njihov utjecaj.
131
Temelji neuroznanosti Vodič kroz vježbe
Polukružni kanalići zamjećuju rotaciju, ali nisu u mogućnosti signalizirati
položaj tijela. To možete ispitati tako da pokažete kako ispitanikov osjećaj u kojem se
smjeru okreće postaje nepouzdan, primjerice prilikom okretanja za jednu četvrtinu
kruga u jednom smjeru, a zatim za pola kruga u drugom smjeru. Međutim test nije
jednostavno izvesti jer se ispitanik može služiti drugim informacijama o smjeru
kretanja. Intenzitet osvjetljenja u prostoru može se mijenjati s rotacijama što se može
zamijetiti i kroz zatvorene kapke. Jednako tako i akustično okruženje nije
ravnomjerno. Trebalo bi obratiti pozornost na navedene čimbenike i pokušati umanjiti
njihov utjecaj.
131

Temelji neuroznanosti Vodič kroz vježbe
10. VRIJEME REAKCIJE
10.1 Uvod
Cilj vježbe je upoznavanje studenata sa značajkama vremena reakcije različitih
psihomotoričkih i kognitivnih procesa. Za ispitivanje vremena reakcije
psihomotoričkih i kognitivnih procesa koristit će se CRD serija psihodijagnostičkih
testova.
CRD (Complex Reactionmeter Drenovac) primjer je elektroničke izvedbe
psihologijskog mjernog instrumenta. Sastoji se od 38 standardiziranih testova koji se
izvode na četiri elektroničke ploče CRD 1-4.
Slika 10.1 CRD sustav
Elektronički instrumenti zajedno s računalom tvore cjelinu tog
psihodijagnostičkog sustava (slika 10.1). Korisno je napomenuti da CRD pripada
instrumentima koji koriste računalo isključivo u svrhu automatizacije mjerenja,
prikupljanja i obrade dobivenih podataka. Dakle, ne pripada instrumentima koji
koriste specifične elektroničke sklopove za simulaciju određenih kognitivnih ili
psihomotoričkih procesa.
Teorijska i metodološka osnova CRD psihodijagnostičkog instrumenta nije
novijeg datuma. Pojava prvog instrumenta datira još od sredine šezdesetih godina.
132
Temelji neuroznanosti Vodič kroz vježbe
10. VRIJEME REAKCIJE
10.1 Uvod
Cilj vježbe je upoznavanje studenata sa značajkama vremena reakcije različitih
psihomotoričkih i kognitivnih procesa. Za ispitivanje vremena reakcije
psihomotoričkih i kognitivnih procesa koristit će se CRD serija psihodijagnostičkih
testova.
CRD (Complex Reactionmeter Drenovac) primjer je elektroničke izvedbe
psihologijskog mjernog instrumenta. Sastoji se od 38 standardiziranih testova koji se
izvode na četiri elektroničke ploče CRD 1-4.
Slika 10.1 CRD sustav
Elektronički instrumenti zajedno s računalom tvore cjelinu tog
psihodijagnostičkog sustava (slika 10.1). Korisno je napomenuti da CRD pripada
instrumentima koji koriste računalo isključivo u svrhu automatizacije mjerenja,
prikupljanja i obrade dobivenih podataka. Dakle, ne pripada instrumentima koji
koriste specifične elektroničke sklopove za simulaciju određenih kognitivnih ili
psihomotoričkih procesa.
Teorijska i metodološka osnova CRD psihodijagnostičkog instrumenta nije
novijeg datuma. Pojava prvog instrumenta datira još od sredine šezdesetih godina.
132

Temelji neuroznanosti Vodič kroz vježbe
Nastao je integracijom različitih metodologija istraživanja koje su bile prisutne i prije
same izrade instrumenta. Pokazatelji učinka i njihovo tumačenje počiva na
kronometriji i pretpostavkama funkcioniranja neurofizioloških mehanizama koji
potječu od I.P. Pavlova. Postavke kronometrije polaze od hipoteze da se iz vremena
ispitanikovih reakcija mogu dobiti informacije o ustroju i funkcijama psihomotoričkih
i kognitivnih procesa.
Kako se razvijala tehnologija i postupci mjerenja, tako su napredovale i nove
izvedbe CRD serije. Počele su se uvoditi različite ideje, teorije i terminologija iz
područja kognitivne psihologije i računalne znanosti (engl., computer science). U
današnjoj izvedbi iskorištene su prednosti primjene računala, programskih jezika
novije generacije, relacijskih modela baza podataka i statističkih programskih paketa.
No, u području neuroznanosti postoji još dosta prostora za istraživanje i razvoj, kako
CRD serije, tako i konstrukcije novih instrumenata.
Uočljivo je da su i na današnjem stupnju informatičke i računalne razvijenosti
nedovoljno istraženi potencijali računalne primjene neuropsihologijskih testova. U
prvom redu to se odnosi na prikupljanje dodatnih podataka o ispitaniku i njihovu
dijagnostičku primjenu. Važno je napomenuti da tehnološke inovacije nisu same po
sebi dovoljne bez dobro osmišljene teorijske osnove, metodologije mjerenja i
znanstvene evaluacije i validacije psihodijagnostičkih instrumenata.
CRD testovi posjeduju formu složenog reakcionmetra. Namijenjeni su
iniciranju kognitivnih procesa, od uočavanja i diskriminacije promjena svjetlosnih i
zvučnih signala, pa do ispitivanja funkcija pamćenja i mišljenja.
CRD serija primjenjuje se u diferencijalnoj dijagnostici, profesionalnoj
orijentaciji i selekciji, te u dijagnostici i praćenju psihomotoričkih i kognitivnih
funkcija u znanstvenim istraživanjima.
Testovi CRD serije prilagođeni su ispitivanju:
perceptivnih sposobnosti (opažanje, diskriminacija, vizualna orijentacija),
pamćenja,
mišljenja (operativno mišljenje, zaključivanje, rješavanje problema),
različitih oblika jednostavnih i izbornih psihomotoričkih reakcija.
Primarni cilj primjene testova CRD serije je dijagnosticiranje i praćenje
vremena i točnosti izvođenja kognitivnih i psihomotoričkih procesa. Testovi su
133
Temelji neuroznanosti Vodič kroz vježbe
Nastao je integracijom različitih metodologija istraživanja koje su bile prisutne i prije
same izrade instrumenta. Pokazatelji učinka i njihovo tumačenje počiva na
kronometriji i pretpostavkama funkcioniranja neurofizioloških mehanizama koji
potječu od I.P. Pavlova. Postavke kronometrije polaze od hipoteze da se iz vremena
ispitanikovih reakcija mogu dobiti informacije o ustroju i funkcijama psihomotoričkih
i kognitivnih procesa.
Kako se razvijala tehnologija i postupci mjerenja, tako su napredovale i nove
izvedbe CRD serije. Počele su se uvoditi različite ideje, teorije i terminologija iz
područja kognitivne psihologije i računalne znanosti (engl., computer science). U
današnjoj izvedbi iskorištene su prednosti primjene računala, programskih jezika
novije generacije, relacijskih modela baza podataka i statističkih programskih paketa.
No, u području neuroznanosti postoji još dosta prostora za istraživanje i razvoj, kako
CRD serije, tako i konstrukcije novih instrumenata.
Uočljivo je da su i na današnjem stupnju informatičke i računalne razvijenosti
nedovoljno istraženi potencijali računalne primjene neuropsihologijskih testova. U
prvom redu to se odnosi na prikupljanje dodatnih podataka o ispitaniku i njihovu
dijagnostičku primjenu. Važno je napomenuti da tehnološke inovacije nisu same po
sebi dovoljne bez dobro osmišljene teorijske osnove, metodologije mjerenja i
znanstvene evaluacije i validacije psihodijagnostičkih instrumenata.
CRD testovi posjeduju formu složenog reakcionmetra. Namijenjeni su
iniciranju kognitivnih procesa, od uočavanja i diskriminacije promjena svjetlosnih i
zvučnih signala, pa do ispitivanja funkcija pamćenja i mišljenja.
CRD serija primjenjuje se u diferencijalnoj dijagnostici, profesionalnoj
orijentaciji i selekciji, te u dijagnostici i praćenju psihomotoričkih i kognitivnih
funkcija u znanstvenim istraživanjima.
Testovi CRD serije prilagođeni su ispitivanju:
perceptivnih sposobnosti (opažanje, diskriminacija, vizualna orijentacija),
pamćenja,
mišljenja (operativno mišljenje, zaključivanje, rješavanje problema),
različitih oblika jednostavnih i izbornih psihomotoričkih reakcija.
Primarni cilj primjene testova CRD serije je dijagnosticiranje i praćenje
vremena i točnosti izvođenja kognitivnih i psihomotoričkih procesa. Testovi su
133

Temelji neuroznanosti Vodič kroz vježbe
posebno korisni u praćenju i dijagnostici unutar različitih zanimanja koja zahtijevaju
aktualizaciju specifičnih znanja, vještina i sposobnosti u kratkom vremenu.
Konstrukcija zadataka u pojedinim testovima izvedena je u skladu s
pretpostavljenim manifestacijama i hijerarhijskim ustrojem kognitivnih procesa.
Prema navedenom, individualni rezultat na testu ili testovima CRD serije predstavlja
skup pokazatelja koji ukazuju na:
specifične sposobnosti (percepcija, pamćenje, mišljenje, i sl.),
funkcionalne karakteristike SŽS-a (brzina, stabilnost, i sl.),
funkcionalne smetnje (perseveracije, regresije).
Unutar perioda latencije uklopljena su vremena različitog trajanja određenih
neurofizioloških procesa. Period latencije predstavlja vrijeme od početka zadavanja
podražaja do pojave odgovora ispitanika na taj podražaj. U toku perioda latencije
odvijaju se različiti procesi na neurofiziološkoj razini.
U periodu latencije sadržana su vremena različitog trajanja potrebna za:
1. odvijanje pojednih neurofizioloških mehanizama pri primanju podražaja,
2. obradu podražajnog sadržaja na kortikalnoj razini,
3. izvođenje odgovora na podražaj,
4. djelovanje individualnih posebnih čimbenika,
5. djelovanje situacijskih čimbenika.
Mjerenjem vremena koje je potrebno za rješavanje zadataka u testovima CRD
serije nastoji se dobiti slika o organizaciji, varijacijama i čimbenicima koji određuju
vremenske odnose unutar elemenata različito složenih kognitivnih i psihomotoričkih
reakcija. Utvrđivanje strukture oslanja se na primjeni postupka frakcioniranja
ukupnog vremena reakcije tj. izdvajanju parcijalnih vremena potrebnih za odvijanje
pojedinog procesa. No, postoje i drugi pristupi koji nastoje proniknuti u strukturu
vremena koje je potrebno za odvijanje kognitivnih i psihomotoričkih procesa. Takvi
pristupi zahtijevaju multidisciplinarni i eksperimentalni rad i koriste primjenu
različitih tehnologija i fizioloških parametara koji čine osnovu vremenskih varijacija.
Dva su temeljna problema kronometrijskih mjerenja:
a) Koliko je vrijeme kognitivnih i psihomotoričkih procesa koji su rezultat
brzine širenja ekscitacije ovisno o karakteristikama neuralnih putova
(debljina i duljina vlakana)?
134
Temelji neuroznanosti Vodič kroz vježbe
posebno korisni u praćenju i dijagnostici unutar različitih zanimanja koja zahtijevaju
aktualizaciju specifičnih znanja, vještina i sposobnosti u kratkom vremenu.
Konstrukcija zadataka u pojedinim testovima izvedena je u skladu s
pretpostavljenim manifestacijama i hijerarhijskim ustrojem kognitivnih procesa.
Prema navedenom, individualni rezultat na testu ili testovima CRD serije predstavlja
skup pokazatelja koji ukazuju na:
specifične sposobnosti (percepcija, pamćenje, mišljenje, i sl.),
funkcionalne karakteristike SŽS-a (brzina, stabilnost, i sl.),
funkcionalne smetnje (perseveracije, regresije).
Unutar perioda latencije uklopljena su vremena različitog trajanja određenih
neurofizioloških procesa. Period latencije predstavlja vrijeme od početka zadavanja
podražaja do pojave odgovora ispitanika na taj podražaj. U toku perioda latencije
odvijaju se različiti procesi na neurofiziološkoj razini.
U periodu latencije sadržana su vremena različitog trajanja potrebna za:
1. odvijanje pojednih neurofizioloških mehanizama pri primanju podražaja,
2. obradu podražajnog sadržaja na kortikalnoj razini,
3. izvođenje odgovora na podražaj,
4. djelovanje individualnih posebnih čimbenika,
5. djelovanje situacijskih čimbenika.
Mjerenjem vremena koje je potrebno za rješavanje zadataka u testovima CRD
serije nastoji se dobiti slika o organizaciji, varijacijama i čimbenicima koji određuju
vremenske odnose unutar elemenata različito složenih kognitivnih i psihomotoričkih
reakcija. Utvrđivanje strukture oslanja se na primjeni postupka frakcioniranja
ukupnog vremena reakcije tj. izdvajanju parcijalnih vremena potrebnih za odvijanje
pojedinog procesa. No, postoje i drugi pristupi koji nastoje proniknuti u strukturu
vremena koje je potrebno za odvijanje kognitivnih i psihomotoričkih procesa. Takvi
pristupi zahtijevaju multidisciplinarni i eksperimentalni rad i koriste primjenu
različitih tehnologija i fizioloških parametara koji čine osnovu vremenskih varijacija.
Dva su temeljna problema kronometrijskih mjerenja:
a) Koliko je vrijeme kognitivnih i psihomotoričkih procesa koji su rezultat
brzine širenja ekscitacije ovisno o karakteristikama neuralnih putova
(debljina i duljina vlakana)?
134

Temelji neuroznanosti Vodič kroz vježbe
b) Koji su čimbenici i koliko vremena otpada na pojedine čimbenike koji
uvjetuju varijacije vremena latencije?
Prema kronometrijskom pristupu, cjelokupno vrijeme rješavanja pojedinog
zadatka odgovarajućeg testa je određeno. Po tom modelu ne postoji slučajni faktor
pogreške. Svako produženo vrijeme rješavanja zadatka ili pogreška uvjetovana je
jednim ili većim brojem čimbenika koji se nastoje tumačiti u terminima afektivne
inkontinencije, afektivnog stupora, mentalnim blokadama i perseveracijama.
Jedini pravi pokazatelj “potencijala” i sposobnosti svakog pojedinca za proces
koji mjeri odgovarajući test CRD serije je najkraće vrijeme rješavanja zadatka u testu
(TMIN). Ponavljanjem većeg broja zadataka u testu postoji i veća vjerojatnost da
ćemo “uhvatiti” ili se asimptotski u funkciji odvijanja testa približiti pravoj veličini
koju test mjeri. To znači da je ta mjera (TMIN) bolja mjera (prave veličine mjerenja)
od ukupnog vremena rješavanja testa u cjelini (UT). S druge strane, MT (srednje
vrijeme rješavanja testa CRD serije) je u slučaju uspoređivanja rezultata među
ispitanicima najprikladnija mjera. Naime, pri obavljanju različitih svakodnevnih
aktivnosti najveća je vjerojatnost da će se vremena obavljanja psihomotoričkih i
kognitivnih procesa kretati oko srednje vrijednosti.
Pokazatelji učinka na CRD testovima
U većini CRD testova učinak i uspješnost izražava se različitim vremenskim i
količinskim parametrima. Svi pokazatelji dostupni su putem računala. Za upravljanje
uređajima CRD serije koristi se aplikacija CRD-Psychodiagnostic System koja
omogućuje automatsko izračunavanje, prikaz i spremanje podataka u bazu.
Pokazatelji vremena rješavanja
1. Ukupno vrijeme rješavanja testa (TT) – ukupno vrijeme rješavanja testa,
predstavlja zbroj pojedinačnih vremena rješavanja zadataka; UT=∑(Ti),
gdje je Ti vrijeme potrebno za rješavanje pojedinog zadatka u testu.
Varijacije ukupnog vremena rješavanja cijelog testa rezultat su intereakcije
različitih čimbenika kao što su brzina, stabilnost, točnost i izdržljivost pri
rješavanju zadataka.
2. Prosječno vrijeme rješavanja zadataka u testu (MT) – za razliku od
ukupnog vremena rješavanja cjelokupnog testa, pogodnija je mjera i
pokazatelj interindividualnih razlika.
135
Temelji neuroznanosti Vodič kroz vježbe
b) Koji su čimbenici i koliko vremena otpada na pojedine čimbenike koji
uvjetuju varijacije vremena latencije?
Prema kronometrijskom pristupu, cjelokupno vrijeme rješavanja pojedinog
zadatka odgovarajućeg testa je određeno. Po tom modelu ne postoji slučajni faktor
pogreške. Svako produženo vrijeme rješavanja zadatka ili pogreška uvjetovana je
jednim ili većim brojem čimbenika koji se nastoje tumačiti u terminima afektivne
inkontinencije, afektivnog stupora, mentalnim blokadama i perseveracijama.
Jedini pravi pokazatelj “potencijala” i sposobnosti svakog pojedinca za proces
koji mjeri odgovarajući test CRD serije je najkraće vrijeme rješavanja zadatka u testu
(TMIN). Ponavljanjem većeg broja zadataka u testu postoji i veća vjerojatnost da
ćemo “uhvatiti” ili se asimptotski u funkciji odvijanja testa približiti pravoj veličini
koju test mjeri. To znači da je ta mjera (TMIN) bolja mjera (prave veličine mjerenja)
od ukupnog vremena rješavanja testa u cjelini (UT). S druge strane, MT (srednje
vrijeme rješavanja testa CRD serije) je u slučaju uspoređivanja rezultata među
ispitanicima najprikladnija mjera. Naime, pri obavljanju različitih svakodnevnih
aktivnosti najveća je vjerojatnost da će se vremena obavljanja psihomotoričkih i
kognitivnih procesa kretati oko srednje vrijednosti.
Pokazatelji učinka na CRD testovima
U većini CRD testova učinak i uspješnost izražava se različitim vremenskim i
količinskim parametrima. Svi pokazatelji dostupni su putem računala. Za upravljanje
uređajima CRD serije koristi se aplikacija CRD-Psychodiagnostic System koja
omogućuje automatsko izračunavanje, prikaz i spremanje podataka u bazu.
Pokazatelji vremena rješavanja
1. Ukupno vrijeme rješavanja testa (TT) – ukupno vrijeme rješavanja testa,
predstavlja zbroj pojedinačnih vremena rješavanja zadataka; UT=∑(Ti),
gdje je Ti vrijeme potrebno za rješavanje pojedinog zadatka u testu.
Varijacije ukupnog vremena rješavanja cijelog testa rezultat su intereakcije
različitih čimbenika kao što su brzina, stabilnost, točnost i izdržljivost pri
rješavanju zadataka.
2. Prosječno vrijeme rješavanja zadataka u testu (MT) – za razliku od
ukupnog vremena rješavanja cjelokupnog testa, pogodnija je mjera i
pokazatelj interindividualnih razlika.
135

Temelji neuroznanosti Vodič kroz vježbe
3. Najkraće vrijeme rješavanja testa (TMIN) – najkraće je vrijeme rješavanja
zadatka u pojedinom testu. Najkraće vrijeme predstavlja “potencijalnu
mentalnu brzinu” izvođenja kognitivne ili psihomotoričke zadaće.
4. Najduže vrijeme rješavanja zadatka (TMAX) – predstavlja maksimalan
učinak nepovoljnog djelovanja većeg broja čimbenika na brzinu izvođenja
određene psihomotoričke ili kognitivne aktivnosti.
Pokazatelji točnosti rješavanja zadataka
1. Ukupni broj pogrešaka u testu (UP) – pokazatelj pouzdanosti
neuropsihologijskih funkcija i točnosti rješavanja zadataka u testu. Broj
pogrešaka kod rješavanja jednog zadatka spada u pokazatelje
funkcionalnih smetnji (perseveracije, rigidnosti ili mentalne
inkontinencije), dok je UP pokazatelj koncentracije pozornosti.
2. Broj pokušaja ulaza u labirint (NU) – pokazatelj uspješnosti učenja i
pokazatelj koncentracije pozornosti.
3. Ukupni broj “lažnih uzbuna” (ULU) – pokazatelj slabe kontroliranosti
ponašanja uslijed povećane ekscitacije.
Pokazatelji fluktuacije tijekom testa:
1. Ukupni balast (ukupno izgubljeno vrijeme - UB) – pokazatelj stabilnosti
odvijanja određene psihomotoričke ili kognitivne funkcije. Ukupno
izgubljeno vrijeme, kao mjera suviška (“balasta”), dobiva se po slijedećem
algoritmu UB=∑(Ti – TMIN); gdje je Ti – vrijeme rješavanja pojedinačnog
zadatka u testu, UB – ukupni “balast”, TMIN – najkraće vrijeme rješavanja
zadatka u određenom testu.
2. Izgubljeno vrijeme u pojedinim sekvencijama (D1...Dn1) – predstavlja
parcijalni balast. To je suma razlika između vremena rješavanja pojedinih
zadatka u sekvenciji i najkraćeg vremena rješavanja zadatka u cijelom
testu.
3. Startna disocijacija (D1/UB) – daje informaciju o izgubljenom vremenu u
rješavanju prvih pet zadataka u testu. Startna disocijacija uvjetovana je
emocionalnim smetnjama u prvom susretu ispitanika s novim sadržajem.
1 D1…Dn predstavlja balast za pojedini niz zadataka. Tako npr. za zadatke od 5 do 10
parcijalni balast možemo označiti kao D5 i on predstavlja zbroj razlika između vremena rješavanja zadataka 5,6,7,8,9,10 i TMIN
136
Temelji neuroznanosti Vodič kroz vježbe
3. Najkraće vrijeme rješavanja testa (TMIN) – najkraće je vrijeme rješavanja
zadatka u pojedinom testu. Najkraće vrijeme predstavlja “potencijalnu
mentalnu brzinu” izvođenja kognitivne ili psihomotoričke zadaće.
4. Najduže vrijeme rješavanja zadatka (TMAX) – predstavlja maksimalan
učinak nepovoljnog djelovanja većeg broja čimbenika na brzinu izvođenja
određene psihomotoričke ili kognitivne aktivnosti.
Pokazatelji točnosti rješavanja zadataka
1. Ukupni broj pogrešaka u testu (UP) – pokazatelj pouzdanosti
neuropsihologijskih funkcija i točnosti rješavanja zadataka u testu. Broj
pogrešaka kod rješavanja jednog zadatka spada u pokazatelje
funkcionalnih smetnji (perseveracije, rigidnosti ili mentalne
inkontinencije), dok je UP pokazatelj koncentracije pozornosti.
2. Broj pokušaja ulaza u labirint (NU) – pokazatelj uspješnosti učenja i
pokazatelj koncentracije pozornosti.
3. Ukupni broj “lažnih uzbuna” (ULU) – pokazatelj slabe kontroliranosti
ponašanja uslijed povećane ekscitacije.
Pokazatelji fluktuacije tijekom testa:
1. Ukupni balast (ukupno izgubljeno vrijeme - UB) – pokazatelj stabilnosti
odvijanja određene psihomotoričke ili kognitivne funkcije. Ukupno
izgubljeno vrijeme, kao mjera suviška (“balasta”), dobiva se po slijedećem
algoritmu UB=∑(Ti – TMIN); gdje je Ti – vrijeme rješavanja pojedinačnog
zadatka u testu, UB – ukupni “balast”, TMIN – najkraće vrijeme rješavanja
zadatka u određenom testu.
2. Izgubljeno vrijeme u pojedinim sekvencijama (D1...Dn1) – predstavlja
parcijalni balast. To je suma razlika između vremena rješavanja pojedinih
zadatka u sekvenciji i najkraćeg vremena rješavanja zadatka u cijelom
testu.
3. Startna disocijacija (D1/UB) – daje informaciju o izgubljenom vremenu u
rješavanju prvih pet zadataka u testu. Startna disocijacija uvjetovana je
emocionalnim smetnjama u prvom susretu ispitanika s novim sadržajem.
1 D1…Dn predstavlja balast za pojedini niz zadataka. Tako npr. za zadatke od 5 do 10
parcijalni balast možemo označiti kao D5 i on predstavlja zbroj razlika između vremena rješavanja zadataka 5,6,7,8,9,10 i TMIN
136

Temelji neuroznanosti Vodič kroz vježbe
4. Startni balast (SB) – pokazatelj je izgubljenog vremena kod rješavanja
zadataka u prvoj polovini testa.
5. Završni balast (ZB) – pokazatelj je izgubljenog vremena u drugoj polovini
rješavanja testa.
Navedeni pokazatelji učinka dostupni su za sve testove koji se koriste u CRD-
seriji.
Označavanje varijabli unutar pojedinog testa vrlo je jednostavno. Tako za test
CRD-11 imamo slijedeće varijable:
1) CRD11_1 do CRD11_35 - vremena rješavanja pojedinačnih zadataka u
testu
2) TT_CRD11 - ukupno vrijeme rješavanja cjelokupnog testa od 35 zadataka
3) MT_CRD11 - prosječno vrijeme rješavanja zadataka u testu
4) TMIN_CRD11 - najkraće vrijeme rješavanja zadataka u testu
5) TMAX_CRD11 - najduže vrijeme rješavanja zadataka u testu
6) UP_CRD11 - ukupan broj pogreški u testu CRD11
7) UKTP_CRD11 - ukupno vrijeme provedeno u pogreškama u testu CRD11
8) TB_CRD11 - ukupni balast ili potrošeno vrijeme u testu
9) D1_UB_CRD11 - startna disocijacija
10) SB_CRD11 - startni balast
11) ZB_CRD11 - završni balast
12) SB_ZB_CRD11 - transfer iskustva tj. omjer startnog balasta i završnog
balasta
13) ZB_SB_CRD11 - indeks izdržljivosti tj. omjer završnog i startnog balasta
10.2 Upute za primjenu testova CRD serije:
Za izvedbu vježbe s CRD uređajima potrebno je:
1) uključiti sva četiri CRD uređaja,
2) pokrenuti aplikaciju CRD - Psychodiagnostic System,
137
Temelji neuroznanosti Vodič kroz vježbe
4. Startni balast (SB) – pokazatelj je izgubljenog vremena kod rješavanja
zadataka u prvoj polovini testa.
5. Završni balast (ZB) – pokazatelj je izgubljenog vremena u drugoj polovini
rješavanja testa.
Navedeni pokazatelji učinka dostupni su za sve testove koji se koriste u CRD-
seriji.
Označavanje varijabli unutar pojedinog testa vrlo je jednostavno. Tako za test
CRD-11 imamo slijedeće varijable:
1) CRD11_1 do CRD11_35 - vremena rješavanja pojedinačnih zadataka u
testu
2) TT_CRD11 - ukupno vrijeme rješavanja cjelokupnog testa od 35 zadataka
3) MT_CRD11 - prosječno vrijeme rješavanja zadataka u testu
4) TMIN_CRD11 - najkraće vrijeme rješavanja zadataka u testu
5) TMAX_CRD11 - najduže vrijeme rješavanja zadataka u testu
6) UP_CRD11 - ukupan broj pogreški u testu CRD11
7) UKTP_CRD11 - ukupno vrijeme provedeno u pogreškama u testu CRD11
8) TB_CRD11 - ukupni balast ili potrošeno vrijeme u testu
9) D1_UB_CRD11 - startna disocijacija
10) SB_CRD11 - startni balast
11) ZB_CRD11 - završni balast
12) SB_ZB_CRD11 - transfer iskustva tj. omjer startnog balasta i završnog
balasta
13) ZB_SB_CRD11 - indeks izdržljivosti tj. omjer završnog i startnog balasta
10.2 Upute za primjenu testova CRD serije:
Za izvedbu vježbe s CRD uređajima potrebno je:
1) uključiti sva četiri CRD uređaja,
2) pokrenuti aplikaciju CRD - Psychodiagnostic System,
137
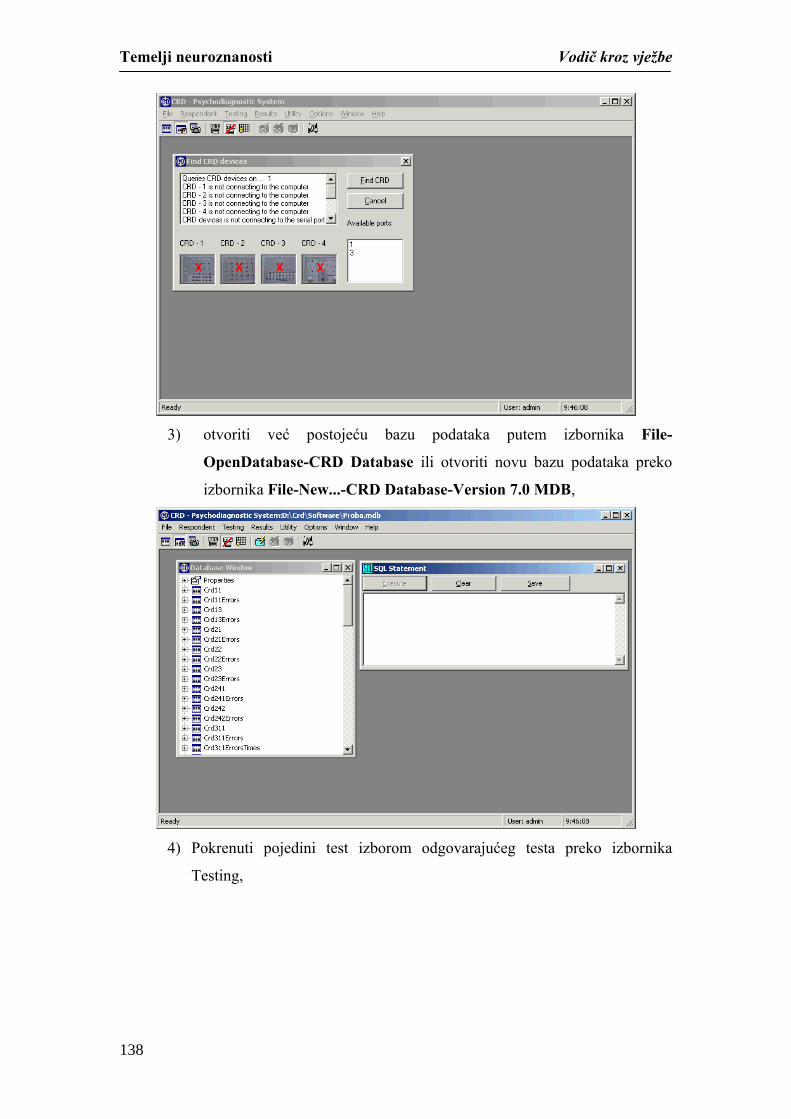
Temelji neuroznanosti Vodič kroz vježbe
3) otvoriti već postojeću bazu podataka putem izbornika File-
OpenDatabase-CRD Database ili otvoriti novu bazu podataka preko
izbornika File-New...-CRD Database-Version 7.0 MDB,
4) Pokrenuti pojedini test izborom odgovarajućeg testa preko izbornika
Testing,
138
Temelji neuroznanosti Vodič kroz vježbe
3) otvoriti već postojeću bazu podataka putem izbornika File-
OpenDatabase-CRD Database ili otvoriti novu bazu podataka preko
izbornika File-New...-CRD Database-Version 7.0 MDB,
4) Pokrenuti pojedini test izborom odgovarajućeg testa preko izbornika
Testing,
138
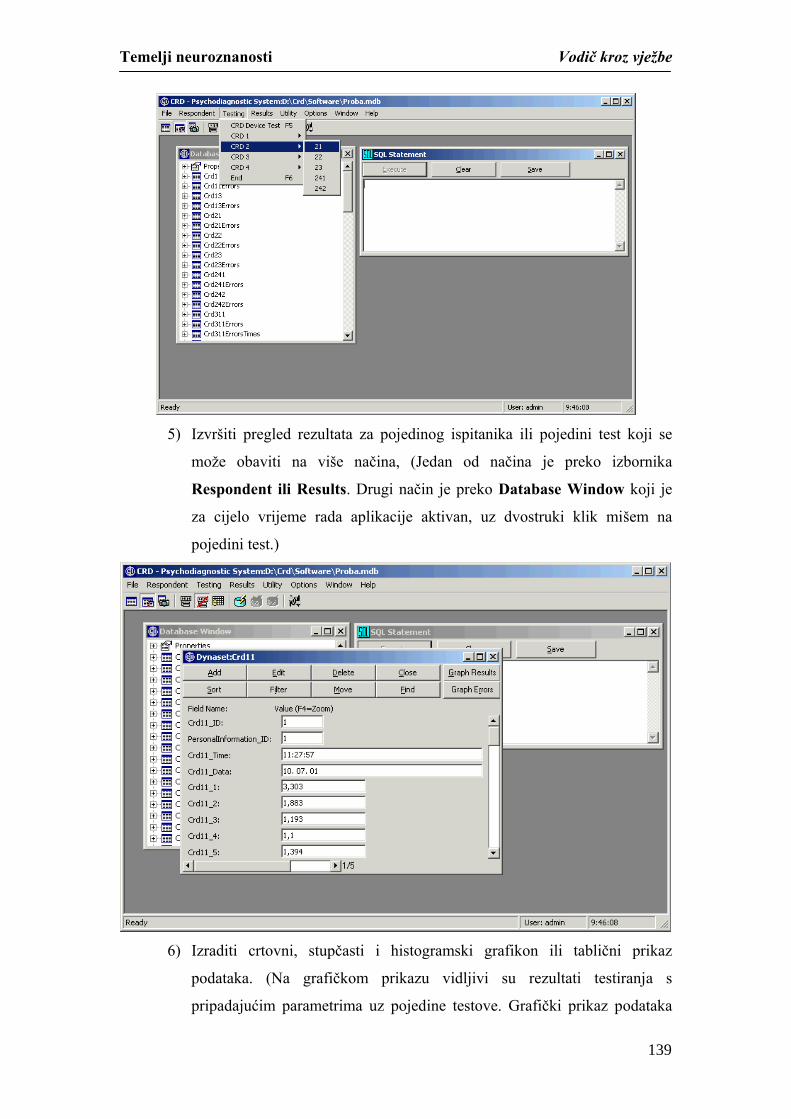
Temelji neuroznanosti Vodič kroz vježbe
5) Izvršiti pregled rezultata za pojedinog ispitanika ili pojedini test koji se
može obaviti na više načina, (Jedan od načina je preko izbornika
Respondent ili Results. Drugi način je preko Database Window koji je
za cijelo vrijeme rada aplikacije aktivan, uz dvostruki klik mišem na
pojedini test.)
6) Izraditi crtovni, stupčasti i histogramski grafikon ili tablični prikaz
podataka. (Na grafičkom prikazu vidljivi su rezultati testiranja s
pripadajućim parametrima uz pojedine testove. Grafički prikaz podataka
139
Temelji neuroznanosti Vodič kroz vježbe
5) Izvršiti pregled rezultata za pojedinog ispitanika ili pojedini test koji se
može obaviti na više načina, (Jedan od načina je preko izbornika
Respondent ili Results. Drugi način je preko Database Window koji je
za cijelo vrijeme rada aplikacije aktivan, uz dvostruki klik mišem na
pojedini test.)
6) Izraditi crtovni, stupčasti i histogramski grafikon ili tablični prikaz
podataka. (Na grafičkom prikazu vidljivi su rezultati testiranja s
pripadajućim parametrima uz pojedine testove. Grafički prikaz podataka
139
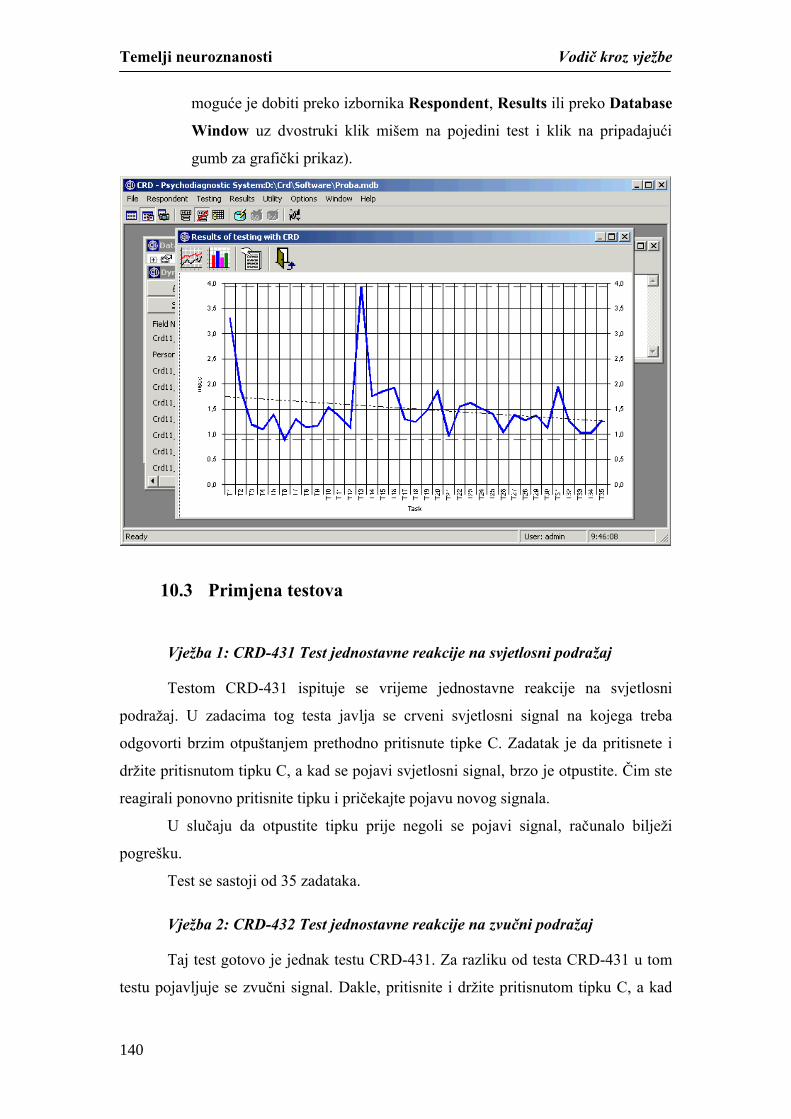
Temelji neuroznanosti Vodič kroz vježbe
moguće je dobiti preko izbornika Respondent, Results ili preko Database
Window uz dvostruki klik mišem na pojedini test i klik na pripadajući
gumb za grafički prikaz).
10.3 Primjena testova
Vježba 1: CRD-431 Test jednostavne reakcije na svjetlosni podražaj
Testom CRD-431 ispituje se vrijeme jednostavne reakcije na svjetlosni
podražaj. U zadacima tog testa javlja se crveni svjetlosni signal na kojega treba
odgovorti brzim otpuštanjem prethodno pritisnute tipke C. Zadatak je da pritisnete i
držite pritisnutom tipku C, a kad se pojavi svjetlosni signal, brzo je otpustite. Čim ste
reagirali ponovno pritisnite tipku i pričekajte pojavu novog signala.
U slučaju da otpustite tipku prije negoli se pojavi signal, računalo bilježi
pogrešku.
Test se sastoji od 35 zadataka.
Vježba 2: CRD-432 Test jednostavne reakcije na zvučni podražaj
Taj test gotovo je jednak testu CRD-431. Za razliku od testa CRD-431 u tom
testu pojavljuje se zvučni signal. Dakle, pritisnite i držite pritisnutom tipku C, a kad
140
Temelji neuroznanosti Vodič kroz vježbe
moguće je dobiti preko izbornika Respondent, Results ili preko Database
Window uz dvostruki klik mišem na pojedini test i klik na pripadajući
gumb za grafički prikaz).
10.3 Primjena testova
Vježba 1: CRD-431 Test jednostavne reakcije na svjetlosni podražaj
Testom CRD-431 ispituje se vrijeme jednostavne reakcije na svjetlosni
podražaj. U zadacima tog testa javlja se crveni svjetlosni signal na kojega treba
odgovorti brzim otpuštanjem prethodno pritisnute tipke C. Zadatak je da pritisnete i
držite pritisnutom tipku C, a kad se pojavi svjetlosni signal, brzo je otpustite. Čim ste
reagirali ponovno pritisnite tipku i pričekajte pojavu novog signala.
U slučaju da otpustite tipku prije negoli se pojavi signal, računalo bilježi
pogrešku.
Test se sastoji od 35 zadataka.
Vježba 2: CRD-432 Test jednostavne reakcije na zvučni podražaj
Taj test gotovo je jednak testu CRD-431. Za razliku od testa CRD-431 u tom
testu pojavljuje se zvučni signal. Dakle, pritisnite i držite pritisnutom tipku C, a kad
140

Temelji neuroznanosti Vodič kroz vježbe
začujete zvučni signal, brzo otpustite tipku. U slučaju da otpustite tipku prije pojave
zvučnog signala, računalo bilježi pogrešku.
Test se sastoji od 35 zadataka i važno je što brže i točnije rješavati zadatke.
Vježba 3: CRD-443 Brzina i točnost razlikovanja svjetlosnog signala
(otpuštanje)
Testom CRD-443 ispituje se brzina i točnost razlikovanja svjetlosnih signala i
izbora odgovora. Za razliku od prethodnih testova, u tom testu na signalnom polju C
može se pojaviti crveni ili zeleni signal.
Prije negoli test započne, morate objema rukama pritisnuti velike tipke na
komandnoj ploči. Kad se pojavi zeleni signal, otpustit ćete lijevu tipku, a kad se
pojavi crveni signal, otpustit ćete desnu tipku.
U slučaju da otpustite bilo koju tipku prije pojave signala bilježi se pogreška.
Isto tako, ako otpustite krivu tipku prilikom pojave signala, računalo bilježi pogrešku.
Čim ste reagirali, ponovno pritisnite tipke i pričekajte pojavu novog signala.
Vježba 4: CRD-21 Jednostavna konvergentna vizualna orijentacija
Kod instrumenta CRD-2 signalni i komandni sklop čine jednu cjelinu. Uz
rubove instrumenta CRD-2 smještene su signalne lampice, a u središnjem dijelu tipke
za odgovor.
Zadatak počinje paljenjem dvije signalne lampice koje određuju stupac i red
položaja tipke za odgovor (Slika 10.2). Dakle, tipka za odgovor nalazi se na mjestu
križanja stupca i retka koje označavaju signalne lampice. U slučaju paljenja pete
lampice uz gornji rub ploče, ona označava da se tipka za odgovor nalazi u petom
stupcu. S druge strane, ako se upali lampica u četvrtom bočnom lijevom nizu, tipka za
odgovor nalazi se u četvrtom redu. Točan odgovor na taj zadatak je pritisak na tipku
koja se nalazi na križanju petog stupca i četvrtog retka.
141
Temelji neuroznanosti Vodič kroz vježbe
začujete zvučni signal, brzo otpustite tipku. U slučaju da otpustite tipku prije pojave
zvučnog signala, računalo bilježi pogrešku.
Test se sastoji od 35 zadataka i važno je što brže i točnije rješavati zadatke.
Vježba 3: CRD-443 Brzina i točnost razlikovanja svjetlosnog signala
(otpuštanje)
Testom CRD-443 ispituje se brzina i točnost razlikovanja svjetlosnih signala i
izbora odgovora. Za razliku od prethodnih testova, u tom testu na signalnom polju C
može se pojaviti crveni ili zeleni signal.
Prije negoli test započne, morate objema rukama pritisnuti velike tipke na
komandnoj ploči. Kad se pojavi zeleni signal, otpustit ćete lijevu tipku, a kad se
pojavi crveni signal, otpustit ćete desnu tipku.
U slučaju da otpustite bilo koju tipku prije pojave signala bilježi se pogreška.
Isto tako, ako otpustite krivu tipku prilikom pojave signala, računalo bilježi pogrešku.
Čim ste reagirali, ponovno pritisnite tipke i pričekajte pojavu novog signala.
Vježba 4: CRD-21 Jednostavna konvergentna vizualna orijentacija
Kod instrumenta CRD-2 signalni i komandni sklop čine jednu cjelinu. Uz
rubove instrumenta CRD-2 smještene su signalne lampice, a u središnjem dijelu tipke
za odgovor.
Zadatak počinje paljenjem dvije signalne lampice koje određuju stupac i red
položaja tipke za odgovor (Slika 10.2). Dakle, tipka za odgovor nalazi se na mjestu
križanja stupca i retka koje označavaju signalne lampice. U slučaju paljenja pete
lampice uz gornji rub ploče, ona označava da se tipka za odgovor nalazi u petom
stupcu. S druge strane, ako se upali lampica u četvrtom bočnom lijevom nizu, tipka za
odgovor nalazi se u četvrtom redu. Točan odgovor na taj zadatak je pritisak na tipku
koja se nalazi na križanju petog stupca i četvrtog retka.
141

Temelji neuroznanosti Vodič kroz vježbe
Slika 10.2 Elektronička ploča CRD-1
Vježba 5: CRD-11 Test konvergentnog mišljenja
Testom CRD-11 ispituje se brzina i točnost rješavanja jednostavnih računskih
zadataka, tj. zbrajanje ili oduzimanje brojeva. Na gornjem dijelu instrumenta CRD-1
nalazi se signalna ploča u čijem je središtu 12 signalnih lampica.
Uz rubove signalne ploče nalaze se prozorčići s brojevima. Uz gornji rub
nalaze se brojevi 13, 11, 12, 10, a uz bočne 4, 1 i 3. Osim brojeva, u lijevom i desnom
gornjem kutu signalne ploče smještena je po jedna signalna lampica. Te dvije lampice
označavaju računsku operaciju koju treba obaviti. Uz lampicu, u lijevom gornjem
kutu je oznaka zbrajanja, a u desnom oduzimanja.
Ispod signalne ploče nalazi se komandna ploča na kojoj je u dva reda
raspoređeno dvanaest tipki za odgovore ispod kojih se nalaze brojevi od 6 do 17.
U svakom zadatku istovremeno se pale dvije signalne lampice. Jedna lampica
u signalnom dijelu ploče označava stupac i red na čijim su krajevima prozorčići s
brojevima koje treba povezati u zadatak. Druga lampica upaljena je u gornjem lijevom
ili desnom kutu pokazujući koju računsku operaciju treba izvršiti sa zadanim
brojevima.
Odgovor se daje pritiskom tipke smještene ispod broja koji predstavlja rezultat
računanja.
Test se sastoji od 35 zadataka i pri rješavanju zadataka u testu bitni su točnost i
brzina. Nakon točno riješenog zadatka odmah se pojavljuje novi zadatak. Ukoliko se
ne pojavi novi zadatak, znači da niste točno odgovorili na tekući.
142
Temelji neuroznanosti Vodič kroz vježbe
Slika 10.2 Elektronička ploča CRD-1
Vježba 5: CRD-11 Test konvergentnog mišljenja
Testom CRD-11 ispituje se brzina i točnost rješavanja jednostavnih računskih
zadataka, tj. zbrajanje ili oduzimanje brojeva. Na gornjem dijelu instrumenta CRD-1
nalazi se signalna ploča u čijem je središtu 12 signalnih lampica.
Uz rubove signalne ploče nalaze se prozorčići s brojevima. Uz gornji rub
nalaze se brojevi 13, 11, 12, 10, a uz bočne 4, 1 i 3. Osim brojeva, u lijevom i desnom
gornjem kutu signalne ploče smještena je po jedna signalna lampica. Te dvije lampice
označavaju računsku operaciju koju treba obaviti. Uz lampicu, u lijevom gornjem
kutu je oznaka zbrajanja, a u desnom oduzimanja.
Ispod signalne ploče nalazi se komandna ploča na kojoj je u dva reda
raspoređeno dvanaest tipki za odgovore ispod kojih se nalaze brojevi od 6 do 17.
U svakom zadatku istovremeno se pale dvije signalne lampice. Jedna lampica
u signalnom dijelu ploče označava stupac i red na čijim su krajevima prozorčići s
brojevima koje treba povezati u zadatak. Druga lampica upaljena je u gornjem lijevom
ili desnom kutu pokazujući koju računsku operaciju treba izvršiti sa zadanim
brojevima.
Odgovor se daje pritiskom tipke smještene ispod broja koji predstavlja rezultat
računanja.
Test se sastoji od 35 zadataka i pri rješavanju zadataka u testu bitni su točnost i
brzina. Nakon točno riješenog zadatka odmah se pojavljuje novi zadatak. Ukoliko se
ne pojavi novi zadatak, znači da niste točno odgovorili na tekući.
142

Temelji neuroznanosti Vodič kroz vježbe
Vježba 6: CRD-13 Test prostorne vizualizacije
Testom prostorne vizualizacije ispituje se brzina i točnost prepoznavanja
likova. Likovi se javljaju u promjenjivim veličinama i različitim položajima u
prostoru.
U središnjem dijelu ploče CRD-1 nalazi se signalni dio s 12 lampica
raspoređenih u tri reda i četiri stupca. U svakom zadatku pali se određeni broj lampica
koje ocrtavaju nekakav lik. Lik može biti crta, rašlje, trokut, četverokut i sl. Važno je
napomenuti da isti likovi u različitim zadacima mogu biti različite veličine, ali i
različito zaokrenuti u prostoru.
Na donjem dijelu ploče CRD-1 nalazi se komandni dio. Na komandnoj su
ploči crteži likova, i to na gornjem dijelu ili u prvom retku komandne ploče likovi
otvorenih linija, a na donjem dijelu ili u drugom retku geometrijski likovi.
Zadatak je ispitanika da pronađe na komandnom dijelu ploče lik koji odgovara
liku ocrtanom lampicama na signalnom dijelu ploče.
Pri rješavanju zadatka nastojte što brže prepoznati lik kojeg tvori skupina
upaljenih signalnih lampica te među tipkama za odgovor (na komandnom dijelu
ploče) brzo pronaći i pritisnuti tipku iznad crteža zadanog lika.
Test se sastoji od 35 zadataka. Nakon točno riješenog zadatka odmah se
pojavljuje novi zadatak. Ukoliko se ne pojavi novi zadatak, znači da niste točno
odgovorili na tekući zadatak.
Pri rješavanju zadatka bitni su točnost i brzina.
Vježba 7: CRD-343
Testovi CRD-34x različite su varijante testova učenja labirinta. Testom CRD-
343 ispituje se brzina i točnost učenja labirinta. Na ploči CRD-3 nalaze se dva niza
tipki, i to gornji i donji niz, te središnji niz signalnih lampica. Svaka pojedina signalna
lampica ima svoj par tipki za odgovor. Dakle, jedna tipka u gornjem nizu i druga tipka
u donjem nizu.
Zadatak je ispitanika 'gašenje' signalnih lampica redom od A do I. Dakle, od
početka do kraja niza pritiskanjem 'ispravnih' tipki za odgovor u pripadajućem paru. U
svakom pripadajućem paru samo je jedna od dvije tipke 'ispravna' za odgovor i
pokretanje signalne lampice na slijedeći korak.
143
Temelji neuroznanosti Vodič kroz vježbe
Vježba 6: CRD-13 Test prostorne vizualizacije
Testom prostorne vizualizacije ispituje se brzina i točnost prepoznavanja
likova. Likovi se javljaju u promjenjivim veličinama i različitim položajima u
prostoru.
U središnjem dijelu ploče CRD-1 nalazi se signalni dio s 12 lampica
raspoređenih u tri reda i četiri stupca. U svakom zadatku pali se određeni broj lampica
koje ocrtavaju nekakav lik. Lik može biti crta, rašlje, trokut, četverokut i sl. Važno je
napomenuti da isti likovi u različitim zadacima mogu biti različite veličine, ali i
različito zaokrenuti u prostoru.
Na donjem dijelu ploče CRD-1 nalazi se komandni dio. Na komandnoj su
ploči crteži likova, i to na gornjem dijelu ili u prvom retku komandne ploče likovi
otvorenih linija, a na donjem dijelu ili u drugom retku geometrijski likovi.
Zadatak je ispitanika da pronađe na komandnom dijelu ploče lik koji odgovara
liku ocrtanom lampicama na signalnom dijelu ploče.
Pri rješavanju zadatka nastojte što brže prepoznati lik kojeg tvori skupina
upaljenih signalnih lampica te među tipkama za odgovor (na komandnom dijelu
ploče) brzo pronaći i pritisnuti tipku iznad crteža zadanog lika.
Test se sastoji od 35 zadataka. Nakon točno riješenog zadatka odmah se
pojavljuje novi zadatak. Ukoliko se ne pojavi novi zadatak, znači da niste točno
odgovorili na tekući zadatak.
Pri rješavanju zadatka bitni su točnost i brzina.
Vježba 7: CRD-343
Testovi CRD-34x različite su varijante testova učenja labirinta. Testom CRD-
343 ispituje se brzina i točnost učenja labirinta. Na ploči CRD-3 nalaze se dva niza
tipki, i to gornji i donji niz, te središnji niz signalnih lampica. Svaka pojedina signalna
lampica ima svoj par tipki za odgovor. Dakle, jedna tipka u gornjem nizu i druga tipka
u donjem nizu.
Zadatak je ispitanika 'gašenje' signalnih lampica redom od A do I. Dakle, od
početka do kraja niza pritiskanjem 'ispravnih' tipki za odgovor u pripadajućem paru. U
svakom pripadajućem paru samo je jedna od dvije tipke 'ispravna' za odgovor i
pokretanje signalne lampice na slijedeći korak.
143

Temelji neuroznanosti Vodič kroz vježbe
Na početku testa pali se prva lampica u nizu s lijeva na desno označena
slovom A. Ispitanik treba pogoditi koja od dvije tipke, gornja ili donja u stupcu A gasi
tu signalnu lampicu i pali slijedeću signalnu lampicu u nizu. U slučaju da ispitanik
pritisne krivu tipku ili bilo koju tipku osim one koja gasi upaljenu signalnu lampicu,
vraća se na početak niza tj. ponovno se pali signalna lampica A. Kad ispitanik dođe do
kraja labirinta, tj. kad bez greške ugasi svih devet lampica jednu za drugom, završen
je jedan cjelokupan prolaz kroz labirint bez pogreške. Na kraju uspješnog prvog
prolaza kroz labirint pojavljuje se zvučni signal i lampica se vraća na prvu lampicu A.
Vrlo je važno napomenuti, ukoliko izostane zvučni signal na zadnjoj lampici, došlo je
do pogreške.
Test je završen kad se tri puta zaredom uspješno bez pogreške prođe kroz
labirint.
144
Temelji neuroznanosti Vodič kroz vježbe
Na početku testa pali se prva lampica u nizu s lijeva na desno označena
slovom A. Ispitanik treba pogoditi koja od dvije tipke, gornja ili donja u stupcu A gasi
tu signalnu lampicu i pali slijedeću signalnu lampicu u nizu. U slučaju da ispitanik
pritisne krivu tipku ili bilo koju tipku osim one koja gasi upaljenu signalnu lampicu,
vraća se na početak niza tj. ponovno se pali signalna lampica A. Kad ispitanik dođe do
kraja labirinta, tj. kad bez greške ugasi svih devet lampica jednu za drugom, završen
je jedan cjelokupan prolaz kroz labirint bez pogreške. Na kraju uspješnog prvog
prolaza kroz labirint pojavljuje se zvučni signal i lampica se vraća na prvu lampicu A.
Vrlo je važno napomenuti, ukoliko izostane zvučni signal na zadnjoj lampici, došlo je
do pogreške.
Test je završen kad se tri puta zaredom uspješno bez pogreške prođe kroz
labirint.
144

Temelji neuroznanosti Vodič kroz vježbe
10.4 PITANJA:
1) Koji sve čimbenici stoje u osnovi razlika između rezultata na testovima
različite psihomotoričke i kognitivne složenosti?
2) Koji sve čimbenici mogu uvjetovati pogreške i fluktuacije vremena
reakcije na pojedine testove?
3) Objasnite aferentne i eferentne putove koji sudjeluju pri rješavanju
zadataka kod testova CRD-431 i CRD-432? Objasnite razliku u
prosječnom vremenu reakcije na svjetlo i zvuk? Opišite neurofiziološku
osnovu.
4) Je li najkraće vrijeme rješavanja zadatka u pojedinom testu (TMIN)
fiziološka granica pojedinog ispitanika?
5) U testu CRD-343 ispituju se funkcije kratkoročne memorije. Objasnite
neurofiziološku osnovu kratkoročne memorije.
6) Koji sve lijekovi mogu utjecati na vrijeme reakcije? Pogađaju li jednako
jednostavne psihomotoričke testove i testove koji ispituju brzinu
procesiranja viših kognitivnih funkcija?
7) Zamislite da imate pacijenta s jednostranim difuznim oštećenjem desne
hemisfere. Na kojim testovima CRD-serije možete očekivati produženje
vremena reakcije i veći broj pogrešaka?
8) Na kojim CRD testovima možete očekivati razliku u vremenu rješavanja
zadataka između muških i ženskih ispitanika?
145
Temelji neuroznanosti Vodič kroz vježbe
10.4 PITANJA:
1) Koji sve čimbenici stoje u osnovi razlika između rezultata na testovima
različite psihomotoričke i kognitivne složenosti?
2) Koji sve čimbenici mogu uvjetovati pogreške i fluktuacije vremena
reakcije na pojedine testove?
3) Objasnite aferentne i eferentne putove koji sudjeluju pri rješavanju
zadataka kod testova CRD-431 i CRD-432? Objasnite razliku u
prosječnom vremenu reakcije na svjetlo i zvuk? Opišite neurofiziološku
osnovu.
4) Je li najkraće vrijeme rješavanja zadatka u pojedinom testu (TMIN)
fiziološka granica pojedinog ispitanika?
5) U testu CRD-343 ispituju se funkcije kratkoročne memorije. Objasnite
neurofiziološku osnovu kratkoročne memorije.
6) Koji sve lijekovi mogu utjecati na vrijeme reakcije? Pogađaju li jednako
jednostavne psihomotoričke testove i testove koji ispituju brzinu
procesiranja viših kognitivnih funkcija?
7) Zamislite da imate pacijenta s jednostranim difuznim oštećenjem desne
hemisfere. Na kojim testovima CRD-serije možete očekivati produženje
vremena reakcije i veći broj pogrešaka?
8) Na kojim CRD testovima možete očekivati razliku u vremenu rješavanja
zadataka između muških i ženskih ispitanika?
145