Embed Size (px)
DESCRIPTION
PMN
Citation preview

SKELETAL MUSCLE
Djoko PrakosaDepartment of Anatomy, Embryology & Anthropology
Faculty of Medicine Gadjah Mada University
One characteristic of animal including man is its ability to move. Movement is carried out by specialized cells called muscle fibers, the latent energy of which is, or can be, controlled by the nervous system. Muscle fibers are classified as skeletal or striated, cardiac, and smooth. They are all derived from myocyte (myo- and sarco- are frequently used in naming structures associated with muscle).
Skeletal Muscle Skeletal muscle fibers are long, multinucleated cells having a characteristic cross-striated appearance under the microscope. These cells are supplied by motor fibers from cells in the central nervous system. The skeletal muscle fibers size tend to be consistent within a given muscle, but in different muscles may range from 10 – 100 in diameter and from milimeters to many centimeters in length. Skeletal muscles are sometimes called voluntary muscles, owing to the fact that they can usually be controlled voluntarily. However, many of the actions of skeletal muscle are automatic, and the actions of some of them are reflex and only to a limited extent under voluntary control.
General Characteristics Most muscles are discrete structures that cross one or more joints and, by contracting, can cause movements at these joints. Exceptions are certain subcutaneous muscles (e.g. facial muscles) that move or wrinkle the skin or close orifices, the muscles that move the eyes, and other muscles associated with the respiratory and digestive systems. Each muscle fiber is surrounded by a connective tissue sheath, the endomysium. Muscle fibers are grouped into fasciculi, each of which is enclosed by a connective tissue sheath termed perimysium. A muscle as a whole is composed of many fasciculi and is surrounded by epimysium, which is closely associated with fascia.
Origin and Insertion Most muscles are attached either directly or by means of their tendons or aponeuroses to bones, cartilages, ligaments, or fasciae. Other muscles are attached to organs, such as the eyeball, and still other are attachjed to skin. When a muscle contracts and shortens, one of its attachments usually remains fixed and the other one moves. The fixed attachments is called the origin, the mobile one the insertion. However, the terms origin and insertion are convenient merely for purposes of description. Very often the anatomical insertion remains fixed and the origin moves. Sometimes both ends remain fixed; the muscle then stabilizeds a joint.
Shape and Fiber Architecture It is possible to attempt a classification of muscles based on their general shape and the predominant orientation of their fibers relative to the direction of pull. Muscles with
1

fibers that are largely parallel to the line of pull vary in form from flat, short and quadrilateral (e.g. thyrohyoid) to long and strap-like (e.g. sternohyoid, sartorius). In a fusiform muscle, the fibers may be close to parallel in the “belly”, but converge to a tendon at one or both ends. Where fibers are oblique to the line of pull, muscles may be triangular (e.g. temporalis, adductor longus) or pennate (=”feather-like”) in construction. The latter vary in complexity from unipennate (e.g. flexor pollicis longus), and bipennate (e.g. rectus femoris, dorsal interossei) to multipennate (e.g. deltoid). In some muscles the fibers pass obliquely between deep and superficial aponeuroses, in a kind of ‘unipennate’ form (e.g. soleus). In other sites muscle fibers start from the walls of osteofascial compartment, and converge obliquely on a central tendon in circumpennate fashion (e.g. tibialis anterior). Some muscles have a spiral or twisted arrangement. Some muscles spiral around a bone (e.g. supinator). Another type of spiral arrangement sometimes reffered to as cruciate, that have two or more planes of fibers arranged in differing directions. Sternocleidomastoid, masseter and adductor magnus are partially spiral and cruciate.
Naming of Muscles The names of muscles usually indicate some structural or functional feature. A name may indicate shape, e.g., trapezius, rhomboid, or gracilis. A name may refer to location, e.g., tibialis posterior. The number of heads of origin is indicated by the terms biceps, triceps, and quadriceps. Action is reflected in terms such as levator scapulae and extensor digitorum. Action and shape are combined in the term flexor digitorum profundus. In anatomical text, individual muscles are usually described according their origin, insertion, nerve supply, and action.
Blood Supply to Skeletal Muscles Muscles are supplied by adjacent vessels. The pattern of supply varies. Some muscles are supplied by vessels that arise from a single stem and enter at one end of the muscle or in the middle of the belly. Other muscles are supplied by a succession of anastomosing vessels. The major source of artery enters on the deep surface, frequently in close association with the principal vein and nerve, which together form a neurovascular hilum. The arteries that enter a muscle branch repeatedly within the connective tissue framework of the muscle, with the smaller arteries and arterioles ramifying in the perimysial septa and giving off capillaries that run in the endomysium. Fibers that are involved in sustained activities –such as posture- are serve by a denser capillary network than fibers which are recruited only infrequently. Veins branch in a similar way, forming venous territories that correspond closely to the arterial territories. The veins is equiped by valves. The pressure exerted on valved intramuscular veins during contraction enables them to function as a ‘muscle pump’, promoting venous return to the heart. Various experiments, provide evidence for the existence in muscle of arteriovenous anastomoses, through which blood can returned directly to the venous system without having traversed the capillaries. The extent to which the muscle capillary bed is perfused can thus be varied in accordance with functional demand.
2

The lymphatic drainage of muscles commences as capillaries in epimysial and perimysial, but not endomysial sheath. These converge to form large lymphatic vessels which accompany the veins, draining to the regional lymh nodes.
Innervation of Skeletal Muscle Each muscle is supplied by one or more nerves. In the limbs, face and neck there is usually a single nerve, although its axons may correspond to several spinal cord segment. Muscles such as those of the abdominal wall, which originate from several embryonic segments, are supplied by more than one nerve. In most cases the nerve travels with principal blood vessels as a neurovascular bundle, approaches the muscle near to its least mobile attachment and enters the deep surface, at a position wich is more or less constant for each muscle. Muscle nerves contain both motor and sensory components. The major motor component , i.e., the -motor axons (-motoneurons) consist of the large, myelinated axons that supply the muscle fibers. In addition, the nerve carries small -motoneuron, which innervate the intrafusal muscle fibers of neuromuscular spindle, and fine, non-myelinated autonomic efferents (C fibers), which innervate vascular smooth muscle. The sensory component consists of the large, myelinated 2A afferents from neuromuscular spindles, the slightly smaller myelinated 2B afferens from the Golgi tendon organs and fine myelineted and non-myelinatedfibers conveying pain and other sensations from free terminals in the connective tissue sheaths of the muscle. -motor axons branch repeatedly before they lose their myelinated sheaths and terminate near the middle of muscle fibers. These terminals tend to cluster in a narrow zone towards the center of the muscle belly known as the motor point. Clinically, this is the place on the muscle from which it is easiest to elicit a contraction with stimulating electrode. A specialized synapse, the neuromuscular junction, is formed where the terminal branch of an -motor axons contacts the muscle fiber. The axon terminal gives off several short, curling branches over an elliptical area, the motor end plate. The arrival of an action potential at the motor end plate causes acetylcholine (ACh) to be released from storage vesicles into the 30 – 50 nm synaptic cleft that separates nerve ending from the sarcolemma. The Ach is rapidly bound by receptor molecules located in the junctional folds, triggering an almost instataneous increase in the permeability, and hence conductance, of the postsynaptic membrane. This generates a local depolarization (the end plate-potential) whose duration is self limited by voltage dependent conformational changes in the membrane. The activity of the neurotransmetter is rapidly terminated by the enzyme acetylcholinesterase (AchE), which is bound to the basement membrane in the junctional folds. A muscle action potential is generated for each nervous impulse. The action potential propagates along the length of the muscle fiber at about 5 meters per second and spreads into its interior along the T-tubules, initiating contraction.
Motor Units and Motor ControlMotor units and their recruitment The terminal branches of -motor axons are normally in a ‘one-to-one’ relationship with their muscle fibers. When a motor neuron is excited, an action potential is propagated along the axon and its branches to all of the muscle fibers that it supplies. The
3

motor neuron and the muscle fibers that it innervates can therefore be regarded as a functional unit, the motor unit, which accounts for the more or less simultaneous contraction of a number of fibers within the muscle. The actual size of the motor units varies considerably; in muscles that are employed for precision tasks each motor neuron innervates about 10 muscle fibers, whereas in a large limb muscle the ratio may exceed 1000. Within a muscle, the fibers belonging to one motor unit are distributed over a wide territory, without regard to fascicular boundaries, and intermingled with the fibers of other motor units. The passage of a single potential through a motor unit elecits a twitch contraction that lasts 25 – 75 ms. However, the motor neuron can deliver a second nervous impulse in less time than it takes for the muscle fibers to relax. When this happens, the muscle fibers contract again, building the tension to a higher level. Through this mechanism, a sequence of impulses can evoke a large force than a single impulse, and –within certain limits- the higher the the impulse frequency, the more force is produced. This force-frequency relationship is one of the strategies used by the nervous system to gradate the contraction it elicits from the muscle. The other strategy is to recruit more motor units. In practice, the two mechanisms appear to operate in parallel. Recruitment is not a random process; neither is there an alteration of active motor units. Low-level contractions are associated with low levels of excitation within the spinal cord, and these conditions always produce regular firing of the same population of small motor neurons. As the force increases, motor neurons of larger size become involved, and the largest motoneuron are activated only when the highest forces have to be generated. Since larger motor neurons have axons of larger diameter, which divide into more numerous branches, the order of recruitment of motor units in every sense one of increasing size. Fiber types of adult skeletal muscle There are two types of skeletal muscle fibers: tonic and twitch. Tonic muscle fibers are slow contracting and cannot propagate action potentials over their membranes. They are found in birds and reptiles. In man this type of fibers only found in the extrinsic ocular muscles and the stapedius muscle of the middle ear. Almost all mammalian skeletal muscles are composed entirely of fibers of twitch type. These fibers can all conduct action potentials but they are not the same in other respect. Some fibers obtain their energy very efficiently by aerobic oxidations of substrates.They have high content of mitochondria and contain myoglobin. They are supported by a well-developped network of capillaries. Such fibers are well suited to functions such as postural maintenance. At the other extreme are fibers that have few mitochondria, little myoglobin, and a sparse capillary network. Their immediate energy requirements are met largely through anaerobic glycolysis, a route that provides prompt access to energy stores but is less efficient and less sustainable than oxidative metabolism. Such fibers are capable of brief bursts of intense activity. In man, all muscles are of the mixed variety, in which fibers that are specialized for aerobic working conditions intermingle with fibers of a more anaerobic or intermediate metabolic in character. These different types of fiber can be seen clearly in sections stained by histochemical techniques. On the basis of the metabolic differences, it is possible to classify the individual fibers as oxidative, glycolytic and intermediate (or oxidative-glycolytic).
4

It is also noted that the ‘red’ muscles (muscles with predominant oxidative fibers) contracted and relaxed more slowly than the ‘white’ muscles (predominant glycolytic fibers). These difference in contractile speed is now known to be in large part to molecular differences between myosin heavy chains in these types of muscles. The different molecular isoforms of myosin may be demonstrated by histochemical techniques. In many muscles, a classification of fibers into Types 1, 2A, and 2B is reasonably robust, based as it is on qualitative differences in myosin. Correspondense between this classification and more quantitative schemes based on metabolism is considerably less secure, because of the amount of metabolic variety within fibers of given type. In broad sense, however, Type 1 and type 2A fibers tend to be predominantly oxidative in character, where as Type 2B fibers rely more heavily on anaerobic glycolysis. Physiologically: - Type 1 fibers are slow contracting and fatigue resistant, Type 2A fibers are fast contracting and fatigue resistant, Type 2B fibers are fast contracting and susceptible to fatigue.
Fatigue and the functional significance of motor unit organization Diversity of muscle fibers properties is an important element in the organization of the motor system. Motor units that are active for much of the time are composed of slow, type 1 fibers. Motor units that are active only infrequently are composed of fast type 2B fibers. Between these extremes, the type 2A motor units cope with the routine activity against a background of more continuous postural tension provided by the type 1 motor units. This organization minimizes the possibility of muscle fatigue. Fatigue can also be deferred if the muscle is activated in an economical way. First, motor units discharge at a rate that correlate closely with the contractile speed of the muscle fbers. Second, motor units discharge asynchronously. Third, individual motor units do not fire at constant frequency.
Functional Implications of Shape and Fiber ArchitectureDirection of action The resultant force generated by a muscle is directed along the line of the tendon; any forces tranverse to this direction must therefore be in balance. In strap-like forms the transverse component is negligible. In the fusiform, bipennate and multipennate forms, symmetry in the arrangement of fibers produces a balanced opposition between transverse component. In asymmetrical muscles, the fibers generate an unopposed lateral component of force which is balanced by intramuscular pressure. Muscles that incorporate a twist in their geometry unwind it as they contract, so that they tend not only to approximate their attachments but also to bring them into the same plane.
Force and range of contraction The force developed by an active muscle is the summation of the tractive forces exerted by millions of cross-bridges as they work asynchronously in repeated cycles of attachment and detachment. This force depends on the amount of contractile machinary that is assambled in parallel, and therefore on the cross-sectional area of the muscle.
5

The range of contraction generated by an active muscle depends on the relative motion that can take place between the overlapping arrays of thick and thin filament in each sarcomere. In vertebrate muscle, the amount of shortening that can take place: from the minimum overlap to the maximum overlap of the thick and thin filaments represents a shortening of about 30%. Since the sarcomeres are arranged in series, the muscle fibers shorten by the same percentage. The actual movement that take place at the ends of the fibers will depend on the number of sarcomeres in series, i.e, it will be proportional to fiber length. Muscles in which the fibers are predominantly parallel to the line of pull are often long and thin. Such muscles develop rather low forces, but are capable of a longe range of contraction. Where greater force is required the cross sectional area must be increased, and the pennate type of construction is a way of achieving this in a compact way. Although the terms power and strength are often used interchangeably with force, they are not synonymous. Power is the rate at which a muscle can perform external work and is equal to force * velocity. Since force depends on the total cross-sectional area of fibers, and velocity depends on their length, power is related to the total mass of a muscle. Strength is usually measured on intact subjects in tasks that require the participation of several muscles; it is then as much an expression of the skilful activation and co-ordination of these muscles as it is a measure of the forces that they contribute individually. Thus it is possible for strength to increase without a concomitant increase in the true force-generating capacities of the muscles iunvolved, espicially during the early stages of training.
Muscle and MovementThe study of muscle action
1. By gross observation2. By localized electrical stimulation3. Manual palpation4. Recording the electrical activity that accompanies mechanical contraction (EMG)5. EMG + goniometer and force plates + recording the movements simultaneously
on videotype or with a full three-dimensional motion analysis system.6. Clinical method
Actions of muscles Classically, the action of a muscle is defined as movement that takes place when it contracts. This should be treated as an operational definition, because a view that equates ‘contraction’ with shortening and ‘relaxation with lengthening, is simplistic in the context of whole muscles and real movements. Contraction can be considered as an attempt on the part of the muscle to approximate its attachments. Whether it actually does so depends on the degree to which it is activated and the forces against which it has to act. These opposing forces are: gravitational and inertial forces; forces generated by any external contact or impact; forces generated by opposing muscles; and forces generated passively by elastic and viscous resistance of all the structures that undergo extension and deformation, some within the muscle it self, others in joints, inactive muscles and soft tissues. Depending on the conditions, an active muscle may maintain its original length, shorten, or lengthen, and during this time its tension may increase, decrease or stay the
6

same. Movements that involve shortening of the active muscle are termed concentric; movements in which the active muscles undergoes lengthening are termed eccentric. Natural movements are accomplished by groups of muscles. Each muscle may be classified, according to its role in the movement, as a: prime mover, antagonist, fixator, and synergist. Prime mover muscles that are consistently active in initiating and maintaining a movement. Muscles that wholy oppose the movement, or initiate and maintain the opposite movement, are the antagonists. When prime movers and antagonists contract together they behave as fixators, stabilizing the corresponding joint by increased transarticular compression, and creating an immobile base on which other prime mover acts. Prime movers that act at multiaxial joints or across more than one joint may produce complex movements, containing elements that have to be eliminated by contraction of other muscles. Those muscles, which assist in accomplishing the movement, are considered to be synergists, although they may act as fixators, or even partial antagonists of the prime mover. In the context of different movements, a given muscle may act as a prime mover, antagonist, fixator, or synergist.
Attachments and levers: the biomechanics of movement Skeletal muscles have a limited range of contraction. Where it is necessary to produce a greater range of movement, this is achieved through the action of the muscles on the bony levers of the skeleton. To produce a large range of movement, force has to be applied close to the joint axis. Example: the action of the triceps on the olecranon, which produces extension at the elbow. However, the motion could be prevented by applying to the hand a force very much less than that developed by the muscle. In fact, the force available at any point along the forearm, wrist and hand is reduced by the same factor as the range of motion there is increased. This type of lever is known as a lever of the first class. In levers of the second class, the load appears between the force and joint axis. Example: plantar flexion of weight-bearing foot raises the body. Here, the range of motion of the load is actually less than that produced by the muscle, and the force is multiplied by the same factor. In levers of the third class, the force is applied between the load and the joint axis. Example: flexion at the elbow produced by the action of bices brachii. The effect is similar to that of the first class lever. Most of the levers of the body are the first and third class.
ReferencesGardner E, Gray DJ & O’Rahilly R. 1975: Anatomy. A regional study of human
structure. 4th ed. WB Saunders Co. Philadelphia.Williams PL, Bannister LH, Berry MM, Collins P, Dyson M, Dussek JE & Fergusson
MWJ. 1995: Gray’s Anatomy. The anatomical basis of medicine and surgery. ELBS with Churchill Livingstone, Edinburgh.
NEURON MOTOR PERIFER
7

A. Brodal: Neurological Anatomy. In relation to clinical Medicine. 3ed New York. Oxford University Press 1981
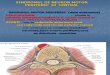
Sel motor di cornu anterius medulla spinalis. Neuron motor perifer atau motoneuron mempunyai badan sel di cornu anterius med. spin. dan beberapa nuclei nn. craniales di truncus cerebri. Dengan ditemukannya inervasi eferen ke fusus neuromuscularis (muscle spindle), motoneuron dibagi menjadi dan . Alpha motoneuron bersifat multipolar. Dendritnya bercabang-cabang di substansia grisea cornu anterius. Axonnya meninggalkan med. spin. melalui radix anterior. Setelah ikut membentuk saraf perifer, axon berakhir di otot. Badan sel motoneuron terletak di lamina IX Rexed, bercampur dengan berbagai neuron yang lebih kecil. Sel-sel kecil tadi terdiri atas motoneuron , interneuron dan sel Renshaw. Motoneuron di cornu anterius dan di nuclei craniales adalah “lintasan akhir bersama” untuk semua impuls yang di transmisikan ke otot skelet. Mereka dipengaruhi oleh berbagai impuls dari berbagai sumber, antara lain dari serabut saraf ordo I. Terhadap motoneuron juga beraksi, terutama melalui interneuron, berbagai serabut desenden yang berasal dari cortex cerebri dan beberapa nuclei di truncus cerebri. Gerak, yang terjadi adalah sebagai pengaruh akhir aksi motoneuron, ditentukan oleh berbagai impuls yang datang di motoneuron. Yang terakhir ini dipengaruhi oleh berbagai sistem. Oleh karena itu, berbagai macam gerak yang terjadi baik pada keadaan normal atau oleh adanya penyakit sistem saraf, tidak seluruhnya ditentukan oleh motoneuron itu sendiri (kecuali apabila motoneuron tersebut secara langsung terlibat dalam proses patologi).
Susunan motoneuron Motoneuron di lamina IX Rexed dan di nuclei nn. craniales tersusun dalam pola yang khas. Mereka tersusun dalam berbagai kelompok berbentuk pilar. Pilar-pilar tadi terdapat di segmen-segmen tertentu. Pada pokoknya, kelompok medial menginervasi otot truncus dan leher. Kelompok ini terdapat di seluruh panjang med. spin. Sebaliknya, kelompok lateral terutama terdapat di intumescentia cervicalis dan lumbalis, dan mereka menginervasi otot anggota badan. Di kelompok lateral, neuron yang menginervasi otot distal terletak di dorsal neuron yang menginervasi otot proksimal. Dengan demikian motoneuron tersusun secara somatotopis.
Unit motor Unit motor adalah motoneuron bersama dengan axon dan seluruh serabut otot yang diinervasinya. Pada saat sebuah motoneuron beraksi, seluruh serabut otot yang diinervasinya berkontraksi. Karena satu motoneuron mungkin menginervasi dari sangat sedikit sampai seribu atau lebih serabut otot, maka ukuran unit motor sangat bervariasi,. Unit motor yang kecil terdapat pada otot-otot yang kecil, misalnya otot ekstraokular dan otot tangan. Demikian juga, unit motor yang kecil terdapat pada otot-otot yang melakukan berbagai gerak yang halus, misalnya otot-otot kecil tangan, otot larynx dan otot ekstraokular. Unit motor yang besar misalnya terdapat pada m. tibialis anterior, m. gastrocnemius. Serabut saraf unit yang kecil umumnya juga berdiameter lebih kecil dibandingkan unit yang besar. Satu serabut saraf dapat menginervasi banyak serabut otot
8

karena axon mempunyai banyak cabang. Serabut-serabut otot yang berasal dari satu unit motor tersebar merata di otot. Ujung cabang-cabang motoneuron bersama dengan membran otot yang diinervasinya membentuk motor-end plate (junctio neuromuscularis). Gambaran pokok dari sebuah motor end plate adalah sbb. Motor end plate terdiri atas dua bagian, yaitu saraf dan otot yang saling dipisahkan oleh celah. Jadi motor end plate ini dalam beberapa hal mirip sinapsis di sistem saraf sentral. Bagian otot mengandung beberapa nuklei dan banyak mitokhondria serta miofibril. Bagian otot dilengkapi dengan sejumlah benjolan seperti buah anggur, sangat mirip benik terminal. Setiap benjolan “melesak” ke dalam serabut otot dan mengandung vesikel sinapsis dan mitokhondria. Telah diketahui bahwa substansi transmiter di end plate adalah asetilkholin. Ia masuk ke dalam celah, berikatan dengan membran otot, dan mengakibatkan perubahan permiabilitas membran tersebut. Satu impuls saraf menghasilkan suatu potensial end plate, dan apabila potensial ini mecapai ambang maka terjadilah potensial aksi yang disebarkan ke sepanjang serabut otot dan menimbulkan kontraksi. Asetilkholin yang dilepaskan pada saat datangnya aksi potensial saraf akan segara dipecah oleh asetilkholinesterase. Transmisi impuls di junctio neuromuscularis dapat dipengaruhi melalui beberapa cara. Curare, misalnya, mengurangi potensial end plate, dengan demikian mencegah timbulnya potensial aksi. Akbiatnya terjadi paralisis otot. (Bandingkan dengan penggunaan substansi seperti curare untuk memperoleh relaksasi pada anestesi). Kerusakan yang terjadi pada miastenia gravis adalah adalah kerusakan pada transmisi di end plate. Potensial yang direkam pada EMG adalah aksi potensial serabut otot tersebut di atas. Apabila serabut saraf dipotong, maka motor end plate dan serabut saraf mengalami degenerasi. Pada umumnya satu serabut otot diinervasi oleh satu axon dan mempunyai satu motor end plate. Setelah lahir ukuran motor unit mengecil, mungkin karena pada mulanya satu serabut otot diinervasi oleh lebih dari satu motoneuron. Setelah tercapai bentuk dewasa yaitu satu serabut otot diinervasi oleh satu motoneuron, maka ukuran unit motor menjadi konstan.
Fungsi unit motor. Elektromiografi Unit motor merupakan unit fungsional terkecil dari alat lokomotor. Berbagai variasi gerak – dalam rentang, daya, dan jenis – pada akhirnya ditentukan oleh perbedaan interaksi dan kolaborasi dari unit motor. Informasi tentang fungsi unit elementer ini terutama diperoleh dari EMG. Apabila perubahan potensial yang terjadi pada otot yang berkontraksi secara maksimal direkam, maka akan tampak banyak sekali gelombang negatif dan positif muncul di rekaman, mengakibatkan sulit dan kompleknya interpretasinya. Sebaliknya, apabila otot dikontraksikan lemah saja, maka aksi potensial tunggal akan muncul dengan interval tertentu. Setiap potensial merefleksikan aktivitas satu unit motor. Oleh karena potensial dari berbagai unit mungkin amplitudonya berbeda, beberapa unit potensial dapat dikenali dengan syarat unit itu hanya sedikit dan kontraksinya lemah. Kekuatan suatu kontraksi otot sebagian tergantung kepada jumlah unit motor yang diaktivasi (direkrut) dan sebagian dari frekuensi impuls yang dikrim motoneuron ke serabut otot yang diinervasinya. Tampaknya rekrutmen merupakan mekanisme yang diinginkan untuk gradasi pada saat gerakan berdaya rendah, dan pada gerak yang sangat singkat yang waktunya hanya cukup untuk satu impuls. Bagaimanapun juga, melihat
9

luasnya rentang fisiologis daya otot, peningkatan frekuensi tampaknya merupakan faktor yang paling penting. Pengungkapan unit motor, dan cara merekam fungsinya, membuka perspektif baru dan meungkinkan pemahaman terhadap berbagai masalah penting. EMG memberi informasi yang berharga untuk membantu diagnosis terutama pada kasus atrofi dan distrofi otot . Pada lesi saraf perifer, EMG dapat dipakai untuk menetukan, pada stadium yang lebih awal dibandingkan dengan cara lainnya, apakah regenerasi terjadi dengan memuaskan. Pemunculan unit motor pada rekaman otot yang paralisis dapat dipakai sebagai bukti bahwa beberapa saraf masih berfungsi dan intak (lolos dari kerusakan atau mengalami regenerasi), meskipun kontraksi tidak tampak. EMG juga terbukti merupakan alat yang bernilai dalam riset fungsi otot. Meskipun EMG menggambarkan aktivitas motoneuron, ia hanya memberi gambaran yang relatif kasar terhadap fungsinya.
Reseptor di otot Otot skelet dilengkapi dengan beberapa jenis reseptor, yang dapat dikelompokkan sbb: fusus neuromuskularis (muscle spindle), organ tendon (Golgi), korpuskulum Vater-Pacini, berbagai jenis akhiran di kapsul sendi, dan akhiran saraf bebas. Dari berbagai reseptor di atas muscle spindle secara morfologis dan fungsional paling banyak diketahui dan organisasinya paling kompleks. Muscle spindle terdapat di semua otot alat lokomotor, dan pada beberapa otot jang diinervasi saraf kranial, misalnya otot larynx, otot mastikasi, lidah, dan otot ekstraokular. Jumlah muscle spindle sangat bervariasi pada berbagai otot. Otot yang dipakai untuk gerak yang halus mempunyai lebih banyak muscle spindle dibandingkan dengan otot yang digunakan untuk gerak kasar. Pokok-pokok penting di dalam organisasi muscle spindle adalah sebagai berikut. Muscle spidle Tersusun atas berupa 3 – 8 (pada manusia rata-rata 10) buah serabut otot yang dibungkus oleh kapsul jaringan ikat berbentuk gelendong. Serabut otot yang disebut serabut otot intrafusal ini kedua ujungnya melekat di jaringan ikat kapsul dan dipisahkan darinya oleh ruang yang berisi cairan. Dengan demikian serabut intrafusal tersusun secara paralel dengan serabut ekstrafusal, dan keduanya akhirnya secara langsung atau tidak langsung, melalui serabut kolagen, dilekatkan ke tendo. Pada saat serabut ekstrafusal berkontraksi, serabut intrafusal akan mengalami penurunan regangan, sebaliknya regangan otot secara keseluruhan akan menambah panjang dan regangan serabut intrafusal. Bagian sentral serabut intrafusal tidak kontraktil, dan di situ banyak nuklei. Mereka dikelilingi oleh serabut saraf yang berjalan spiral, yaitu akhiran annulospiral atau sensor primer, yang merupakan cabang-cabang serabut aferen yang relatif besar. Akhiran sensris kedua, atau akhiran flower spray berasal dari serabut yang relatif lebih kecil mungkin terdapat di kedua sisi akhiran anulospiral. Sebagai tambahan, muscle spindle mendapat inervasi motor. Serabut ini relatif kecil ( motoneuron) dan berasal dari cornu anterius. Akhiran serabut ini terletak di bagian yang kontraktil dari serabut intrafusal. Berdasar ini kontraksi bagian tepi dari serabut intrafusal sebagai respon dari impuls yang datang melalui motoneuron akan menyebabkan regangan bagian sentral serabut intrafusal yang bersifat “sensoris”, dengan konsekuensi terpacunya akhiran sensoris, seperti halnya pada saat seluruh otot diregangkan.
10

Organ tendon.jauh lebih sederhana dibandingkan muscle spindle. Pada intinya, organ tendon terdiri atas sekelompok cabang serabut saraf bermielin yang berakhir di antara serabut-serabut kolagen tendo, biasanya di dekat junctio musculotendinosa. Satu serabut mungkin menginervasi lebih dari satu tendo. Biasanya mereka dibungkus dengan jaringan ikat yang halus. Untuk memahami fungsi organ tendon, perlu untuk mengingat kenyataan bahwa mereka tersusun secara seri dengan serabut ekstrafusal. Apakah otot berkontraksi atau teregang, organ tendon akan terangsang, karena pada kedua keadaan regangan di tendo akan meningkat. Penelitain fisiologis menunjukkan bahwa apabila otot diregangkan frekuensi aksi potensial serabut aferen dari muscle spindle dan organ tendon meningkat, dan apabila regangan dilepaskan, setelah istirahat sejenak, frekuensi kembali ke keadaan istirahat. Apabila otot berkontraksi, organ tendon kembali merespon dengan menaikkan frekuensinya karena tertariknya tendo. Bagaimanapun juga pada saat otot berkontraksi muscle spindle akan kehilangan beban dan akan istirahat. Apabila kontraksinya berhenti, spindle yang kendor kembali meregang dan sebagai akibatnya akan timbul aksi potensial, sedangkan organ tendon tidak memberi respon karena regangan mengendor. Berdasar temuan ini, organ tenton diduga merupakan perekam regangan, sedangkan muscle spindle memberi informasi mengenai panjang otot.
Stretch reflex dan pengendalian otot Bahkan bila dipisahkan dari bagian otak lainnya, med spin mampu memediasi sejumlah refleks, somatik dan autonomik. Dasar morfologis refleks saraf umumnya disebut arkus refleks, yang dalam bentuknya yang paling sederhana tersusun atas (1) reseptor, yang bereaksi terhadap stimulus; (2) penghantar eferen, yang membawa impuls ke “pusat refleks” (Penghantar aferen adalah serabut sensorik aferen, yang kebanyakan mempunyai badan sel di ganglion spinal atau kranial); (3) “Pusat refleks”, tempat pesan aferen dari reseptor berkumpul dengan impuls aferen dari reseptor lainnya, atau dengan aferen dari sumber lain, yang mungkin mengubah pengaruh impuls aferen dari reseptor; (4) penghantar eferen, yaitu serabut saraf yang menuju ke efektor; (5) efektor, yang menghasilkan reaksi, yang mungkin adalah otot, kelenjar atau vasa darah, atau mungkin melibatkan beberapa komponen itu. Refleks sangat bervariasi, dari yang sangat kompleks, misalnya refleks menelan, yang melibatkan berbagai efektor; sampai yang paling sederhana. Di sini kita uraikan mengenai refleks spinal, dan hanya yang efektornya adalah otot skelet, yaitu refleks-refleks somatik. Beberapa jenis refleks dapat disebutkan. Refleks fleksor adalah yang responnya adalah fleksi anggota badan. Stimulus yang paling poten adalah noksiseptif, dan hasilnya adalah tarikan anggota badan (withdrawal reflex). Pada refleks lain ada ekstensi anggota badan, misalnya pada crossed extensor reflex yang mungkin menyertai refleks fleksor. Masih ada lagi refleks yang lebih kompleks, misalnya scratch reflex. Semua refleks tersebut biasanya melibatkan beberapa otot, dan respon refleksnya mungkin berbagai macam tergantung pada keadaan (jenis dan tempat pengenaan stimulus, intensitas stimulus, pengenaan stimulus lain secara bersamaan, dll). Arkus refleks semacam ini sangat kompleks. Refleks lain adalah stretch reflex, yaitu kontraksi satu otot karena diregangkan. Ini merupakan refleks elementer yang mungkin terjadi di
11

semua otot. Stretch refleks menjadi dasar banyak sekali postural reflex, yang secara garis besar bertujuan untuk menjaga sikap tubuh yang benar, dan menyesuaikan diri dengan berbagai kebutuhan, baik itu karena daya dari luar atau disebabkan karena gerak yang dilakukan oleh organisme. Salah satu contoh dari stretch reflex adalah refleks lutut. Ini merupakan refleks monosinapsis, satu-satunya sinapsis terdapat di med spin. Refleks ini memerlukan stimulus melebihi keadaan fisologis. Tidak semua otot mempunyai stretch refleks, dan kadang-kadang refleks ini absen pada subyek yang sehat. Respon refleks terhadap regangan otot, yang terjadi dengan masa laten yang lebih lama dibanding stretch reflex yang monosinaptik, tampaknya mempunyai arti fungsional yang lebih signifikan. Lintasan stretch refleks polisinaptik ini kurang begitu diketahui. Di dalam istilah klinik refleks-refleks yang disebutkan di atas disebut tendon reflexes atau deep reflexes. Kadang-kadang ini juga disebut refleks miotatik.. Gambaran umum stretch reflex adalah sebagai berikut. Pukulan pada ligamentum patellae akan meregangkan baik otot ekstrafusal maupun muscle spindle m. quadriceps. Muscle spindle menjawab dengan mengirimkan serentetan impuls ke med spin. Serabut aferen di radix dorsale yang membawa impuls membentuk sinapsis dengan motoneuron yang kemudian tereksitasi dan menyebabkan kontraksi m. quadriceps. Agar hal ini terjadi, maka harus disertai pengurangan tensi di otot antagonis m. quadriceps, yaitu otot fleksor lutut. Hal ini terjadi sbb. Aferen dari muscle spindle memberi cabang kolateral ke sekelompok neuron intermedier di substasia grisea. Neuron ini ini bersifat inhibitor dan mengirimkan axonnya ke motoneuron yang menginervasi otot antagonis. Contoh di atas menggambarkan refleks fasik, di situ muscle spindle dengan melalui impuls aferen, secara monosinaptik mengaktipkan motoneuron yang menginervasi otot ekstrafusalnya sehingga menyebabkan kontraksi otot dan menyebabkan relaksasi antagonis. Pada saat kontraksi, karena panjangnya berkurang muscle spindle akan berhenti mengirimkan impuls, motoneuron tidak lagi tidak tereksitasi, dan otot akan kembali relaksasi. Meskipun demikian, stretch reflex juga dapat bersifat tonik. Reflex tonik ini mungkin dimediasi oleh lintasan polisinaptik. Contoh refleksnya adalah refleks biceps dan refleks tendo flexor pollicis longus. Jawaban reflek ini dua macam. Satu mempunyai letensi pendek dan yang lain lebih panjang. Secara tidak langsung terdapat bukti bahwa pada refleks ini cortex cerebri ikut terlibat. Reseptornya adalah muscle spindle, meskipun demikian dapat juga dari reseptor kulit dan sendi. Telah dibuktikan bahwa informasi baik akhiran primer maupun sekunder dari muscle spindle mencapai cortex cerebri. Hilangnya masa latensi panjang dari stretch refleks akibat adanya lesi pada capsula interna, columna dorsalis, atau cortex sensorimotor memberi dukungan terhadap dugaan bahwa suatu jirat transcortical ikut terlibat.
Tonus otot Apabila otot normal yang istirahat dipalpasi, dapat dirasa bahwa otot tersebut sama sekali tidak flaksid tetapi mempunyai regangan tertentu. Ini juga impresi yang diperoleh saat otot tersebut secara pasif digerakkan. Keadaan ini disebut tonus otot atau tonus istirahat. Pda berbagai keadaan patologis tonus yang normal ini berubah. Kadang-kadang meningkat (hipertonus), dan pada keadaan lain berkurang (hipotonus). Klinis, tonus otot diperiksa dengan palpasi dan gerak pasif. Bagaimanapun juga, pada kedua metode pemeriksaan tersebut terdapat bukti ada dua komponen tonus otot. Sehingga tidak jarang
12

terlihat, misalnya pada hemiplegia kapsular, tonus otot pada palpasi menurun (konsistensi otot berkurang), sedangkan pada gerak pasif tampak meningkat, karena resistensi terhadap gerakan lebih besar daripada normal. Resistensi otot karena digerakkan secara pasif secara prinsip dapat disebabkan oleh dua faktor: yaitu sifat viskoelastik otot itu sendiri dan tegangan yang diakibatkan oleh kontraksi. Kedua fkator tentu saja sama pentingnya, tetapi kontribusi mereka masing-masing jelas berbeda pada berbagai macam keadaan. Dari penelitian binatang yang dideserebrasi terbukti bahwa tonus otot terutama disebabkan oleh refleks, yang disebabkan oleh aliran impuls yang berkesinambungan dari muscle spindle, yang mengaktivasi motoneuron. Meskipun demikian, hasil ini sulit diterapkan pada manusia pada keadaan sadar. Mereka mungkin paling relevan untuk otot yang menjaga posisi tegak, yang memperlihatkan aktivitas berkesinambungan atau intermiten pada orang yang berdiri dengan relaks (misalnya, beberapa ekstensor punggung, m. psoas major, dan m. soleus; kebanyakan otot lain tidak menunjukkan aktivitas EMG pada posisi ini). Dengan demikian, konsistensi otot mungkin tergantung tidak hanya kepada aktivitas otot yang sedang berlangsung, tetapi juga oleh berbagai sifat ototnya itu sendiri. Oleh karena motoneuron dapat mengubah sensitivitas spindle, mereka penting untuk tonus otot. Aktivitas yang meningkat mengatur spindle pada tingkat yang lebih aktif, sehingga dapat meningkatkan tonus otot ekstrafusal. Pada saat gerakan dilaksanakan, inervasi penting dalam menentukan keadaan otot yang akan dipakai dan ia aktif dalam mengendalikannya, sedangkan motoneuron bertanggung jawab terhadap kontraksinya sendiri. Kerjasama dan neuron pada pelaksanaan gerak banyak diteliti. Tampaknya, sistem saraf pusat mampu mengendalikan statik dan dinamik dan neuron secara sendiri-sendiri. Sampai seberapa jauh kendali sentral ini dimediasi oleh lintasan terpisah tidak diketahui, tetapi berbagai struktur tertentu terlihat terutama mempengaruhi satu jenis neuron fusimotor. Dengan demikian, stimulasi bagian medial formatio reticularis menyebabkan meningkatnya sensitivitas statik spindle, sedangkan stimulasi bagian lateral menyebabkan peningkatan aktivitas dinamik. Faktor lain yang mempengaruhi stretch reflex dan tonus otot adalah organ tendon. Pada peregangan dan terutama kontraksi otot, reseptor ini terstimulasi. Aferennya tidak langsung ke motoneuron, tetapi melalui interneuron, yang mengirimkan impulsnya ke motoneuron. Fungsi organ tendon diduga mencegah kontraksi berlebihan dan untuk pengerem dan pada saat yang sama memfasilitasi antagonis. Selain itu juga ada yang untuk eksitasi motoneuron sinergis dan inhibisi antagonis. Jadi fungsinya lebih kompleks. Sebagai tambahan dari organ tendon, ada mekanisme lain yang cenderung untuk menghambat aktivitas motoneuron yang tereksitasi, yaitu inhibisi Renshaw. Hal ini berdasar pengamatan fisiologis. Impuls motoneuron melalui kolateral di cornu anterius merangsang sel Renshaw yang pada gilirannya akan menghambat motoneuron.
Beberapa gambaran organisasi motoneuron Kinerja gerakan sangat kompleks. Gerak ini memerlukan kerjasama yang harmonis dari sejumlah unit motor, yang terdistribusi di berbagai tempat di med spin dan menginervasi berbagai otot. Aktivitas ini dikoordinasi dalam waktu dan intensitas; waktu fasilitasi dan inhibisi unit harus secara tepat dilakukan. Kekomplekan itu dapat digambarkan dengan baik dengan kenyataan bahwa bagian-bagian dari suatu otot dapat
13

digunakan pada berbagai gerak yang kadang-kadang saling antagonistis. Dengan demikian, bagian tertentu dari m. deltoideus pada satu posisi dapat berfungsi sebagai abduktor sendi bahu, dan sebagai adduktor pada posisi yang lain. Jelas, serabut pada satu radix dorsalis bersinapsis dengan interneuron, dan melalui ini dapat mempengaruhi beberapa segmen med spin. Secara tradisional interneuron termasuk tipe Golgi II, yang mempunyai ciri khas aksonnya bercabang-cabang di dekat badan sel. Konsep ini dipakai untuk menjelaskan berbagai macam observasi fisiologis. Alpha motoneuron dapat dibagi menjadi dua tipe: tonik dan fasik. Motoneuron tonik antara lain mempunyai frekuensi impuls lebih rendah, kecepatan konduksi lebih rendah, berdiameter lebih kecil, ukuran unit motor lebih kecil daripada motoneuron fasik. Pada saat otot berkontraksi, ini bukan merupakan aktivasi satu atau beberapa unit motor, tetapi melibatkan “pool” motoneuron. Apabila diasumsikan bahwa input sinaptik pada pool ini pada berbagai keadaan secara relatif terdistribusi ke semua selnya, yang mempunyai berbagai ukuran, urutan aktivasi yang biasanya diobservasi ( neuron - neuron tonik - neuron fasik) dapat diterangkan berdasarkan morfologinya.
Simtom setelah lesi neuron motor perifer Berdasarkan uraian di atas maka apabila semua sel di cornu anterius yang menginerrvasi otot atau bagian otot rusak, maka otot atau bagian otot tersebut akan tidak mampu berkontraksi. Contoh keadaan ini adalah poliomielitis yang menyerang motoneuron baik med spin maupun truncus cerebri. Pada paralisis akibat destruksi semua atau hampir semua motoneuron yang menginervasi otot, tidak hanya gerak volunter saja yang hilang, tetapi juga kontraksi refleksnya. Lebih jauh lagi, karena terputusnya lengkung refleks miotatik maka otot menjadi flaksid. Konsistensinya turun dan tidak ada resistensi pada gerak pasif. Ciri lain adalah munculnya atrofi otot, yang semakin jelas pada paralisis komplet dan waktu yang lama. Apabila neuron motor perifer yang menginervasi otot tidak seluruhnya rusak, maka hanya timbul paralisis partial atau paresis. Simptom lain sama dengan pada paralisis tetapi dengan derajad yang lebih ringan. Termasuk didalamnya adalah atrofi, penurunan tonus dan refleks miotatik yang melemah. Gerak yang dilakukan lebih lemah dan biasanya juga lebih lambat. Simtom di atas, paresis, atrofi, hipotonus, refleks yang hilang atau melemah, dan retardasi tentu saja tidak hanya terjadi pada lesi badan sel saja, tetapi juga pada lesi aksonnya yang ada di radix anterior, plexus atau saraf perifer. Gangguan tersebut pada beberapa keadaan hanya bersifat fungsional, tanpa kerusakan morfologis serabut saraf, misalnya tumor intraspinal yang menekan radix motor, atau saraf perifer yang menyebabkan anoksi saraf karena kompresi sirkulasi. Kadang-kadang, pada pasien dengan kerusakan neuron motor perifer timbul fibrilasi, yaitu kontraksi sekelompok kecil otot yang bersifat halus, ritmik atau sering ireguler yang dapat terlihat di bawah kulit atau teraba pada palpasi. Keadaan lain adalah fasikulasi. Keduanya dapat direkam dengan EMG. Fasikulasi diduga disebabkan oleh kerusakan di badan sel (misalnya yang terjadi pada amyotropik lateral sclerosis atau progressive spinal muscular atrophy), sehingga simtom tersebut dapat untuk diferensial diagnosis apakah lesi bersifat radikular atau nulear.m
14

Pada otot yang mengalami denervasiada fenomena lain yang dapat dilihat pada EMG, yaitu timbulnya aksi potensial kecil berfrekuensi tinggi. Potensial fibrilasi ini tampaknya berasal dari satu serabut otot yang kehilangan inervasi motoris. Penyebab fibrilasi diduga adalah sensitisasi membran otot setelah denervasi. Tiadanya transmitter, asetilkholin, menyebabkan serabut otot merespon terhadap sejumlah kecil asetilkholin yang beredar di sirkulasi. Prostigmin, yang menghambat pemecahan asetilkholin, cenderung meningkatkan fibrilasi pada otot yang denervasi.
15