Embed Size (px)
Citation preview

Pa
KJa
b
c
d
e
f
a
ARRAA
KPAMNMH
1
o
s(cora
DT
0h
Peptides 48 (2013) 124– 133
Contents lists available at ScienceDirect
Peptides
jo ur nal home p age: www. elsev ier .com/ locate /pept ides
roline rich-oligopeptides: Diverse mechanisms for antihypertensivection
atia L.P. Moraisa,b,1, Danielle Ianzera,c,1, José Rodolfo R. Mirandaa,b, Robson L. Meloa,uliano R. Guerreirod, Robson A.S. Santose, Henning Ulrichf, Claudiana Lameuf,∗
Laboratório Especial de Toxinologia Aplicada (LETA/CAT-Cepid), Instituto Butantan, SP, BrazilDepartamento de Bioquímica, Universidade Federal de São Paulo, SP, BrazilDepartamento de Ciências Biológicas, Instituto de Ciências Exatas e Biológicas, Universidade Federal de Ouro Preto, Ouro Preto, MG, BrazilLaboratório de Bioquímica de Proteínas, Centro de Biotecnologia Agrícola – CEBTEC, Departamento de Ciências Biológicas, ESALQ/USP, SP, BrazilLaboratório de Hipertensão, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, MG, BrazilDepartamento de Bioquímica, Instituto de Química, Universidade de São Paulo, SP, Brazil
r t i c l e i n f o
rticle history:eceived 16 May 2013eceived in revised form 12 July 2013ccepted 12 July 2013vailable online xxx
eywords:roline-rich oligopeptidesrgininosuccinate synthetaseuscarinic acetylcholine receptoritric oxideean arterial pressureeart rate
a b s t r a c t
Bradykinin-potentiating peptides from Bothrops jararaca (Bj) discovered in the early 1960s, were the firstnatural inhibitors of the angiotensin-converting enzyme (ACE). These peptides belong to a large familyof snake venom proline-rich oligopeptides (PROs). One of these peptides, Bj-PRO-9a, was essential fordefining ACE as effective drug target and development of captopril, an active site-directed inhibitor ofACE used worldwide for the treatment of human arterial hypertension. Recent experimental evidencesdemonstrated that cardiovascular effects exerted by different Bj-PROs are due to distinct mechanismsbesides of ACE inhibition. In the present work, we have investigated the cardiovascular actions of four Bj-PROs, namely Bj-PRO-9a, -11e, -12b and -13a. Bj-PRO-9a acts upon ACE and BK activities to promote bloodpressure reduction. Although the others Bj-PROs are also able to inhibit the ACE activity and to potentiatethe BK effects, our results indicate that antihypertensive effect evoked by them involve new mechanisms.Bj-PRO-11e and Bj-PRO-12b involves induction of [Ca2+]i transients by so far unknown receptor proteins.Moreover, we have suggested argininosuccinate synthetase and M3 muscarinic receptor as targets for
cardiovascular effects elicited by Bj-PRO-13a. In summary, the herein reported results provide evidencethat Bj-PRO-mediated effects are not restricted to ACE inhibition or potentiation of BK-induced effects andsuggest different actions for each peptide for promoting arterial pressure reduction. The present studyreveals the complexity of the effects exerted by Bj-PROs for cardiovascular control, opening avenuesfor the better understanding of blood pressure regulation and for the development of novel therapeutic approaches.. Introduction
Bradykinin-potentiating peptides are a family of proline-richligopeptides (PROs) containing 5–14 amino acid residues [6,14]
Abbreviations: ACE, angiotensin I-converting enzyme; ASS, argininosuccinateynthetase; Bj, Bothrops jararaca; BK, bradykinin; PRO, proline-rich oligopeptide;[Ca2+]i), intracellular calcium concentration; CNP, C-type natriuretic peptide; CNS,entral nervous system; HR, heart rate; mAChRs, muscarinic receptors; NO, nitricxide; NOS, nitric oxide synthase; MAP, mean arterial pressure; PAP, pulsatile arte-ial pressure; SHR, spontaneously hypertensive rat; MDLA, �-methyl-dl-asparticcid; 4-DAMP, 4-[(diphenylacetyl)oxy]-1,1-dimethyl-piperidinium iodide.∗ Corresponding author at: Universidade de São Paulo, Instituto de Química,epartamento de Bioquímica, Av. Lineu Prestes 748, São Paulo, SP 05508-900, Brazil.el.: +55 11 3091 9745; fax: +55 11 3815 5579.
E-mail addresses: [email protected], [email protected] (C. Lameu).1 These authors should be considered co-first authors.
196-9781/$ – see front matter © 2013 Elsevier Inc. All rights reserved.ttp://dx.doi.org/10.1016/j.peptides.2013.07.016
© 2013 Elsevier Inc. All rights reserved.
found in the venom gland and in the neuroendocrine tissue ofBothrops jararaca brain as part of the C-type natriuretic peptideprecursor [13,30]. These small molecules are known as naturalinhibitors of the angiotensin-converting enzyme (ACE). Specifi-cally, the Bj-PRO-9a led to the development of captopril, one of themost successful drugs for the treatment of human hypertension[7,32]. However, systematic studies performed in conscious spon-taneously hypertensive rats (SHRs) revealed that the most potentin vitro inhibitor of ACE (Bj-PRO-10c) was able to elicit a strongantihypertensive effect independent on in vivo ACE inhibition [16].Recently, we found that the same oligopeptide specifically inter-acts with kidney argininosuccinate synthetase (ASS) promoting itsactivation. Once activated, ASS substantially increases the produc-
tion of l-arginine and nitric oxide (NO), accounting for almost 50%of the antihypertensive activity of Bj-PRO-10c in SHR [10].Another important evidence regarding the action mechanismof Bj-PROs arose recently, as Bj-PRO-10c mobilized intracellular

eptide
crptr
lhBbrobb
cmglrfmi
cibBiHni
2
2
b11pw(UwmtspaufleMSdFue
2
b
K.L.P. Morais et al. / P
alcium ([Ca2+]i) fluxes in neurons and induced release of the neu-otransmitters GABA and glutamate, which are involved in bloodressure control [22]. These results allowed the comprehension ofhe action mechanism of Bj-PRO-10c in the CNS, where the peptideegulates the arterial baroreflex of SHRs [23].
Recently, we have demonstrated that Bj-PRO-5a also causesong-lasting antihypertensive effect [17]. Despite of this peptideas always been regarded as an efficient potentiating agent ofK induced-effects, it mediates NO production by activating ofradykinin-B2 receptor as well as of M1 muscarinic acetylcholineeceptors (mAChRs) and inducing vasodilatation in vivo [28]. More-ver, the characterization of Bj-PRO-7a as M1 mAChR agonist [31]rings new insights for explaining antihypertensive action exertedy this peptide [16].
It is well known that the regulation of arterial pressure involvesomplex, time-dependent interactions among multiple neural, hor-onal and intrinsic regulatory systems [20,34,35]. Nowadays a
reat progress has been achieved to understand the pathophysio-ogy of human hypertension, although a large number of questionsemain to be answered [19]. The cardiovascular properties of theamily of proline-rich oligopeptides seem to provide interesting
olecular tools to explore the yet unrevealed metabolic and phys-ological pathways involving cardiovascular homeostasis.
This study was undertaken in order to evaluate the enzymatic,ellular and pharmacological properties of four Bj-PROs, previouslydentified in both crude venom and neuroendrocrine tissue. Despiteeing similar in their size and amino acid sequences, Bj-PRO-9a,j-PRO-11e, Bj-PRO-12b and Bj-PRO-13a, present large differences
n their biological activities when compared among themselves.erein, we demonstrate that complex, but independent mecha-isms are involved in the cardiovascular effects evoked by Bj-PROs
n SHRs.
. Materials and methods
.1. Synthesis and purification of peptides
Peptides were synthesized essentially as previously describedy [8]. Briefly, the synthesis of Bj-PRO-9a (<EWPRPQIPP), Bj-PRO-1e (<EARPPHPPIPP), Bj-PRO-12b (<EWGRPPGPPIPP) and Bj-PRO-3a (<EGGWPRPGPEIPP) were performed on an automated PSSM-8eptide synthesizer (Shimadzu Corp., Kyoto, Japan) by a step-ise solid-phase method using N-9-fluorenylmethoxycarbonyl
Fmoc) chemistry (Novabiochem-EMD Chemicals, San Diego, CA,SA). Peptide cleavage from the resin was achieved by treatmentith a mixture of trifluoroacetic acid (TFA)/1,2-ethanedithiol/ethylethyl sulfide for 2 h at room temperature. After removal of
he resin by filtration and washing twice with TFA, the crudeynthetic peptide was purified by preparative reversed-phase high-erformance liquid chromatography (HPLC) (Shimadzu Corp.) on
YMC-Pack ODS column 20 mm × 150 mm (YMC, Kyoto, Japan),sing a linear gradient from 3 to 20% CH3CN in 0.1% TFA, at aow rate of 7 ml/min. Both, the purity and primary structure ofach synthetic peptide were confirmed by analytical HPLC andALDI-TOF mass spectrometry (Amersham Biosciences, Uppsala,
weden). Samples were frozen in liquid nitrogen and then freezeried (Edwards Freeze Dryer Super Modulyo Pirani 1001, Thermoisher Scientific, Waltham, MA, USA) for 48 h at −50 ◦C under vac-um. Following this procedure, the counter ion TFA was completelyxchanged as determined by mass spectrometry analysis.
.2. Animals
Experiments were performed using male Wistar rats (250–280 gody weight) from COBIO, Instituto Butantan and male SHRs
s 48 (2013) 124– 133 125
(280–350 g) from CEBIO, Universidade Federal de Minas Gerais andBiotério Central, ICB, Universidade de São Paulo. The animals werebred at the animal facility of the Laboratório Especial de Toxinolo-gia Aplicada, Instituto Butantan and at the animal facility of theLaboratório de Hipertensão, UFMG. The animals had free access tofood and water and were submitted to a light/dark cycle (12 h each)before the preparation for the experiments. All animals were han-dled under ethical conditions according to international rules ofanimal care, stated by the International Animal Welfare Recom-mendations, and in accordance with the guidelines established bythe local institutional animal welfare committee (CEUAIB/ButantanInstitute, protocol 520/08).
2.3. Cardiovascular parameters recording in rats
The cardiovascular parameters, pulsatile arterial pressure (PAP),mean arterial pressure (MAP) and heart rate (HR) were monitoredby a solid-state strain gauge transducer connected to a computerthrough a data acquisition system (MP 100; BIOPAC Systems Inc.,USA). The PAP, MAP and HR were monitored jointly during experi-ments in different monitor channels and recorded in the computerhard disk for later analysis.
2.4. Evaluation of cardiovascular parameters in conscious SHRs
This assay was performed according to Ianzer and colleagues[16]. The choice of dose (71 nmol/kg) was based on the effectsof Bj-PROs 5a, 7a and 10c found in a dose-response curve(0.47–710 nmol/kg) in SHRs [16,17]. Under anesthesia with tribro-moethanol (2.5 g/kg, intraperitoneally), 20 h before the experimentpolyethylene catheters (PE-10 connected to PE-50) were placedinto the abdominal aorta through the femoral artery and vein tomeasure cardiovascular parameters and to inject drugs, respec-tively. After recovery from anesthesia, rats were kept in individualcages with free access to water and chow until the end of the exper-iments.
Cardiovascular parameters were initially followed for 1 h (base-line period). Subsequently, intravenous (iv) bolus injection of theBj-PROs or vehicle (NaCl 0.9%) in a total volume of 0.5 ml wasperformed. A Bj-PRO dose of 71 nmol/kg was used (n = 5–6). Thecardiovascular parameters were monitored continuously for 6 hafter injection. The time-course changes were calculated from thedifference between basal average value (10 min average sampledimmediately before iv injection) and value of every 10 min postinjection time. The peak changes were observed in each rat. For thepeak change evaluation the time was not considered.
2.5. Hypotensive effect of bradykinin in anesthetized Wistar rats
The evaluation of the capacity of Bj-PROs to potentiate thehypotensive effect of bradykinin (BK) in rats was performed aspreviously described [16]. The choice of dose (60 nmol) was basedon previously published data [8,15,37]. Briefly, Wistar rats wereanesthetized with sodium urethane (14 g/kg, intraperitoneally),polyethylene catheters (PE-10 connected to a PE-50) was insertedinto the abdominal aorta through the femoral artery for bloodpressure measurements. For iv bolus injections, a polyethylenecatheter was implanted into the femoral vein. Rats were main-tained anesthetized during the experiments. After blood pressurestabilization, standard hypotensive responses evoked by iv injec-tions of BK (0.5 �g and 1.0 �g) were recorded. All oligopeptides
were dissolved in sterile isotonic saline immediately before use.Following, the injection of the Bj-PROs (60 nmol) was performed.Subsequently, injections of BK (0.5 �g) were repeated at 5, 10, 15,20, 25, and 30 min after peptide administration.
1 eptide
2p
TasOBwwpaTtHeapidm
2
up(ttwtommw
2
1s1w
2c
tfcsmiKfi(aSro
26 K.L.P. Morais et al. / P
.6. Human argininosuccinate synthetase (ASS) expression andurification
The cDNA coding for human ASS was obtained from Lifeechonologies (San Diego, CA, USA) and was cloned together with
N-terminal His-tag fusion coding sequence into pDEST17 expres-ion vector. Then this vector was transformed into the E. coli BL21rigami (DE3) pLysS strain and liquid-culture growth in Terrificroth using standard methods, expression of the recombinant ASSas induced at 18 ◦C by addition of 0.2% l-arabinose. Bacterial cellsere harvested, resuspended in incubation buffer (20 mM Tris–HClH 7.5, 100 mM KCl, 20 mM imidazole, 10 mM �-mercaptoethanolnd 10% glycerol) and lysed for 20 s by two rounds of sonication.he soluble fraction of the lysate was collected after further cen-rifugation (250 × g, 4 ◦C, 45 min) and loaded directly onto a 1 mlis-Bind Ni2+ affinity resin (Qiagen, Hilden, Germany), previouslyquilibrated with incubation buffer supplemented 80 mM imid-zole. Following ten washing steps, the recombinant expressedrotein was then eluted with incubation buffer containing 500 mM
midazole in a single 2 ml fraction. Eluted fractions were thenialyzed against 20 mM Tris–HCl pH 7.5, containing 10 mM �-ercaptoethanol and 10% glycerol.
.7. ASS activity assay
ASS activity was determined based on accumulation of the prod-ct pyrophosphate as inorganic phosphate, following cleavage withyrophosphatase [11]. ASS (1 �g) was added to the reaction buffer20 mM Tris–HCl, pH 7.8, 2 mM ATP, 2 mM citrulline, 2 mM aspar-ate, 6 mM MgCl2, 20 mM KCl, and 0.2 units of pyrophosphatase)o a final volume of 0.2 ml. Bj-PROs (concentrations: 0.01–100 �M)ere incubated at 37 ◦C in 96-well microtiter plates, and the reac-
ions were stopped after 30 min by the addition of an equal volumef molybdate buffer (10 mM ascorbic acid, 2.5 mM ammoniumolybdate, 2% sulfuric acid). Accumulation of phosphate was deter-ined spectrophotometrically at 650 nm, and its concentrationas extrapolated from a standard curve of inorganic phosphate.
.8. HEK 293 cells culture
Human Embryonic Kidney 293 (HEK293) cells [ATCC no. CRL-573] were cultured in Dulbecco’s modified Eagle’s mediumupplemented with 10% fetal bovine serum, 2 mM l-glutamine,00 �g/ml streptomycin sulfate and 100 U/ml penicillin G. Culturesere kept at 37 ◦C in 5% CO2 atmosphere.
.9. Measurements of changes in free intracellular calciumoncentration ([Ca2+]i) measurements by microfluorimetry
Changes in [Ca2+]i were determined by microfluorimetry usinghe FlexStation III (Molecular Devices Corp., Sunny Valley, CA, USA),ollowing previous published protocols [22,28]. Briefly, HEK293ells were seeded a night before starting the experiment at a den-ity of 5 × 104 cells/well in 96-well black walled and clear bottomedicroplates, with 100 �l of cell culture medium per well. Cells were
ncubated for 60 min at 37 ◦C with the FlexStation Calcium Assay
it (Molecular Devices Corp.) containing 2.5 mM probenecid in anal volume of 200 �l per well. Before and after addition of Bj-PROsconcentrations: 0.003–1 �M), fluorescence of samples was excitedt 485 nm, and fluorescence emission was detected at 525 nm.amples were read for 120 s at 1.52 s intervals with a total of 79ead-outs per well. Responses were measured at the peak intensityf fluorescence compared to baseline background values.s 48 (2013) 124– 133
2.10. Chemiluminescence assay for measurement of nitric-oxideproducts
For NO products quantification, 1 × 106 HEK293 cells wereplaced in 6 wells plates containing culture medium supplementedwith 10% fetal bovine serum. After the period of cell adhesion, thecultures were kept for 24 h in medium without serum togetherwith final concentrations of 0.03–3 �M Bj-PROs. Cells and culturemedium were processed as follows. The culture medium was col-lected and centrifuged for 5 min at 90 × g and 4 ◦C to eliminatepossible debris. The resulting supernatant was analyzed for nitratecontent. Cells were resuspended in RIPA buffer (50 mM Tris–HCl,pH 7.5, 150 mM NaCl, Nonidet 1% P-40 (NP-40), 0.5% sodium deoxy-cholate, 0.1% SDS, 1 mM DTPA and 10 mM N-ethylmaleimide) andincubated for 20 min on ice for cell lysis. The resulting cell extractwas centrifuged for 15 min at 21,000 × g and 4 ◦C. The methodol-ogy and instrumentation for the measurements were performedas described by Feelisch and colleagues [5]. Purified intracellularmedium and extracellular medium were directly injected into avessel containing a saturated solution of vanadium (III) chloridein 1 N HCl maintained at 90 ◦C. Under these conditions, all nitricoxide-derived products (nitrate, nitrite, nitrosothiol, nitrosamines,and iron-nitrosyl complexes) were reduced and compared withthose of standard solutions of nitrate under the same experimentalconditions.
Other set of assay was performed in order to clarify themechanism involved in NO production in HEK293 cells by Bj-PROs. Before to incubate with oligopeptides, the cells werepre-treated with inhibitors or antagonists: ASS inhibitor: 10 mM�-methyl-dl-aspartic acid (MDLA); bradykinin-B2 receptor antag-onist: 1 �M HOE-140; �2-adrenergic receptor antagonist: 10 �Myohimbine; muscarinic receptors (mAChR) M1, M2 or M3 subtypesantagonists: 1 mM pirenzepine, 100 �M gallamine or 10 �M 4-[(diphenylacetyl)oxy]-1,1-dimethyl-piperidinium iodide4-DAMP,respectively. MDLA was used at 10 mM, an essential concentrationto inhibit allosteric activation of ASS evoked by Bj-PRO-10c [10].The concentrations of antagonists used correspond to that requiredto inhibit the receptors when submitted to stimulation with theirrespective agonists [26,28].
2.11. Statistical analysis
Data are expressed as mean ± standard error (S.E.). The criteriafor statistical significance were set at P < 0.05. Statistical analyseswere made using GraphPad Prism 5.0 software (GraphPad SoftwareInc., San Diego, CA, USA). For in vivo assays: Statistical signifi-cances were determined by the T-test; one-way analysis of variance(ANOVA) followed by the Dunnett post-test or by two-way ANOVAwith Bonferroni post-test when appropriate. Statistical analyses forBK-potentiation effects were performed using the effect of 0.5 �g ofBK before Bj-PRO injection as reference. For in vitro assays: Statisti-cal significances were determined by one-way analysis of variance(ANOVA) followed by the Dunnett post-test.
3. Results
3.1. Effect of Bj-PROs on cardiovascular parameters
3.1.1. Potentiation of the hypotensive effect of BK by Bj-PROsIn order to investigate the cardiovascular effects evoked by iv
bolus injection of Bj-PROs (71 nmol/kg), we monitored during 6 h
the arterial pressure (MAP) and heart rate (HR) of conscious SHRs.As observed in similar experimental conditions [16,17], injection ofvehicle provoked slight changes in MAP and HR, likely attributed tothe experimental procedure in free-behaving animals (Fig. 1A and
K.L.P. Morais et al. / Peptides 48 (2013) 124– 133 127
Fig. 1. Time-course changes on cardiovascular parameters of SHRs produced by Bj-PROs. Cardiovascular parameters were sampled each 10 min during 6 h after Bj-PROs(71 nmol/kg, iv injection) in conscious SHRs. The control group received 0.9% saline (vehicle). Fill bars represents mean arterial pressure (MAP) and open bars representsheart rate (HR). (A and B) Vehicle; (C and D) Bj-PRO-9a; (E and F) Bj-PRO-11e; (G and H) Bj-PRO-12b; (I and J) Bj-PRO-13a. Data are presented as mean values ± S.E. *P < 0.05compared to vehicle.

1 eptides 48 (2013) 124– 133
Bed
wobi
aowegt(
i3Tr
it−Bbt
p
3
tFtte(birPaio
3
bBAcP(
3
dhcBa
ivit
ies
of
syn
thet
ic
Bj-P
RO
s.
Act
ion
s
of
vari
ous
Bj-P
RO
s
on
card
iova
scu
lar
par
amet
ers
of
con
scio
us
SHR
as
wel
l as
on
the
BK
-in
du
ced
hyp
oten
sion
, act
ivat
ion
of
AsS
, NO
pro
du
ctio
n
and
ind
uct
ion
of
[Ca2+
] itr
ansi
ents
.ex
pre
ssed
as
mea
n
valu
es
±
stan
dar
d
erro
rs. N
D:
not
det
erm
ined
;
NS:
not
sign
ifica
nt.
Am
ino
acid
sequ
ence
Ori
gin
AC
E
Ki (
nM
)
Max
imal
red
uct
ion
in
PAM
(mm
Hg)
Max
imu
mre
du
ctio
n
in
FC(b
pm
)
Max
imal
BK
pot
enti
atio
n
in
vivo
(%)
Max
imal
acti
vati
onof
ASS
(%)
Max
imal
ind
uct
ion
of
NO
pro
du
ctio
n(n
mol
/106
cell
s)
Ind
uct
ion
of
[Ca2+
] itr
ansi
ents
N-s
ite
C-s
ite
<EK
WA
P
Ven
oma
Bra
inb
400b
1280
b−3
0
±
5e−6
6
±
9e48
±
5g17
e8.
08
±
3.6h
7.93
±
0.1
h
<EW
PRPQ
IPP
Ven
oma
100b
1b−2
2
±
3*,j
−40
± 15
j13
8
±
17j
0j0j
0j
<EN
WPH
PQIP
P
Ven
oma
Bra
inb
200b
0.5b
−53
±
6*,d
−67
± 10
*,d
120
±
23i
85f
3.1
±
0.4
f7.
23
±
0.0
g
<EA
RPP
HPP
IPP
Bra
inb
100b
300b
−26
±
3*,j
−61
±
9*,j
48
±
10j
0j0j
7.15
±
0.11
j
<EW
GR
PPG
PPIP
P
Bra
inb
5b15
0b−2
5
±
2*,j
−54
±
9*,j
102
±
31j
50j
0j8.
63
±
0.15
j
<EG
GW
PRPG
PEIP
P
Ven
omc
Bra
inc
50b
50b
−27
±
2*,j
−67
±
10*,
jN
Sj41
j9.
09
±
2.08
j7.
82
±
0.17
j
t
al. (
1970
)
[6].
t
al. (
2003
)
[13]
.
al. (
2004
)
[14]
.
al. (
2007
)
[16]
.
al. (
2011
)
[17]
.
et
al. (
2009
)
[10]
.
al. (
2010
)
[22]
.
al. (
2011
)
[28]
.et
al. (
2012
)
[37]
.ep
ort.
omp
ared
wit
h
con
trol
(−11
±
2
mm
Hg
and
−31
± 9
bpm
).
28 K.L.P. Morais et al. / P
). All tested Bj-PROs produce antihypertensive and bradycardicffects in SHRs (Fig. 1C–J). However, the magnitude of these car-iovascular responses differed among proline-rich oligopeptides.
As showed in Fig. 1C, the antihypertensive effect of Bj-PRO-9aas oscillatory and reached about −16 mmHg, close to the end
f experimental period. The Bj-PRO-9a caused slight and transientidirectional cardiac chronotropic effects during the entire exper-
mental period (Fig. 1D).The Bj-PRO-11e caused gradual reduction in MAP, reaching
bout −18 mmHg at 330 min, which was sustained until the endf the recording (Fig. 1E). The time-course of HR of SHRs treatedith Bj-PRO-11e presented similar profile of MAP. The bradycardic
ffect evoked by Bj-PRO-11e was evident after 60 min and increasedradually up to the end experimental period. The average HR reduc-ion reached −40 bpm, likely close to end of experimental periodFig. 1F).
The Bj-PRO-12b caused a gradual decrease in MAP, reach-ng approximately −20 mmHg at 240 min, which was kept until60 min (Fig. 1G). This peptide also promoted a bradycardic effect.he HR reductions were evident immediately after injection andeached −40 bpm at 280 min (Fig. 1H).
The antihypertensive effect observed following Bj-PRO-13anjection displayed a gradual and stable reduction in MAP untilhe end of experimental period, reaching maximal average of25 mmHg at 290 min (Fig. 1I). The marked bradycardia caused byj-PRO-13a was observed at 40 min (about −42 bpm), but a smallradycardic effect was predominant and persisted until the end ofhe experimental period (Fig. 1J).
Table 1 presents the maximal peak changes in cardiovasculararameters (MAP and HR) of SHR caused by Bj-PROs.
.2. Potentiation of the hypotensive effect of BK by Bj-PROs
Effects of Bj-PROs upon hypotensive activation caused by injec-ion 0.5 �g of BK in anesthetized normotensive rats are shown inig. 2. Bj-PROs 11e and 13a (60 nmol) were not able to changehe amplitude of the hypotensive effect evoked by iv bolus injec-ion of BK (Fig. 2, panels C and E). 60 nmol of Bj-PRO-9a (asxpected) and 12b potentiated the hypotensive effect of BK (0.5 �g)Fig. 2B), but the amplitude of this potentiation was different foroth oligopeptides. Different from the other oligopeptides studied
n the present work, Bj-PRO-9a evoked slight and transient MAPeduction, although it did not differ from vehicle (Fig. 2A and B). Bj-RO-9a and Bj-PRO-12b elicited BK potentiation at 5–20 min; and 5nd 10 min after iv injection, respectively (Fig. 2B and D). The max-mal potentiation of BK-induced hypotensive effects after injectionf the Bj-PROs is presented in Table 1.
.3. Effect of Bj-PROs on catalytic activity of ASS
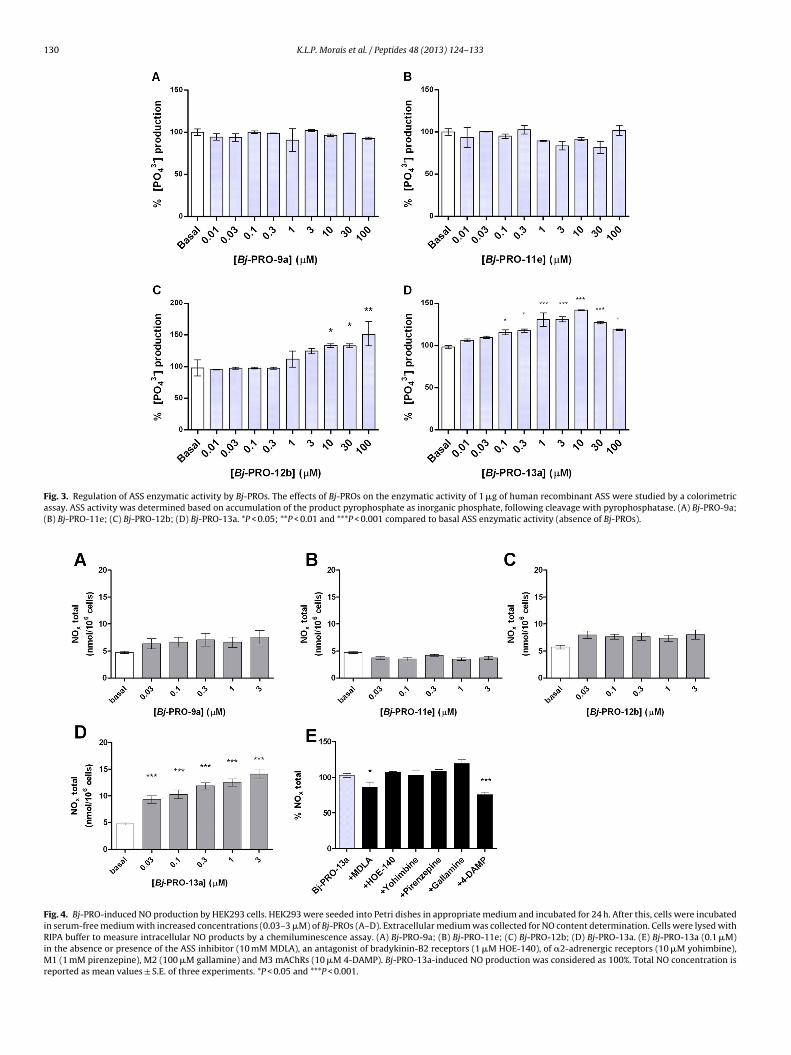
Increasing concentrations of Bj-PROs 12b and 13a on recom-inant ASS modulated its positively catalytic activity. 100 �Mj-PRO-12b and 10 �M Bj-PRO-13a reached a maximal increase ofSS activity at about 50% and 42%, respectively (Fig. 3C and D). Inontrast, at least in the concentrations studied (0.01–100 �M), Bj-RO-9a and Bj-PRO-11e did not exert any effects on ASS activityFig. 3A, B and E).
.4. Effect of Bj-PROs in the NO production by cells HEK293
Previous studies have shown that Bj-PRO-10c induced NO pro-uction in various cell lines [4,10]. Within this framework, we
ave studied the effects of Bj-PROs on NO production in HEK293ells. After treatment of the cells with increasing amounts of eachj-PRO, we observed that Bj-PRO-9a, Bj-PRO-11e and Bj-PRO-12bt concentrations of 0.03–3 �M did not promote any significant Table
1B
iolo
gica
l act
The
dat
a
are
Bj-P
RO
Bj-P
RO
-5a
Bj-P
RO
-9a
Bj-P
RO
-10c
Bj-P
RO
-11e
Bj-P
RO
-12b
Bj-P
RO
-13a
aFe
rrei
ra
eb
Hay
ash
i ec
Ian
zer
etd
Ian
zer
ete
Ian
zer
etf
Gu
erre
iro
gLa
meu
eth
Mor
ais
eti
Tash
ima
jPr
esen
t
r*
P
<
0.05
c

K.L.P. Morais et al. / Peptides 48 (2013) 124– 133 129
Fig. 2. BK-mediated hypotensive effects in anesthetized Wistar rats before and after administration of Bj-PROs. Intravenous bolus injection of each Bj-PRO was made at thed j-PROo
cBat1
i1a
ose of 60 nmol. (A) Vehicle; (B) Bj-PRO-9a; (C) Bj-PRO-11e; (D) Bj-PRO-12b; (E) Bbtained with 0.5 �g of BK before peptide injection; #P < 0.05 compared to vehicle.
hanges of NO production in HEK293 cells (Fig. 4A–C). However,j-PRO-13a dose-dependently increased NO production, startingt concentration of 0.03 �M. Maximal response was observed inhe presence of 3 �M Bj-PRO-13a from basal levels of 4.75 ± 0.68 to4.09 ± 1.4 nmol/106 cells (Fig. 4D).
The effects of specific inhibition of ASS and different receptorsnvolved in regulation of NO production were studied for Bj-PRO-3a (0.1 �M). Pretreatment of cells with the ASS inhibitor, MDLA;nd antagonist of the subtype M3 mAChR, 4-DAMP, inhibited
-13a. Data are expressed as mean values ± S.E. *P < 0.05 compared with the values
Bj-PRO-13a-induced NO production by 10 and 25%, respectively(Fig. 4E).
3.5. Bj-PROs-induced [Ca2+]i transients in HEK293 cells
Bj-PROs-induced [Ca2+]i transients after stimulation with theoligopeptides were monitored by microfluorimetry [22,28]. Thedose-response curves of peak values of [Ca2+]i transients evokedby Bj-PRO-11e, Bj-PRO-12b and Bj-PRO-13a revealed potencies
130 K.L.P. Morais et al. / Peptides 48 (2013) 124– 133
Fig. 3. Regulation of ASS enzymatic activity by Bj-PROs. The effects of Bj-PROs on the enzymatic activity of 1 �g of human recombinant ASS were studied by a colorimetricassay. ASS activity was determined based on accumulation of the product pyrophosphate as inorganic phosphate, following cleavage with pyrophosphatase. (A) Bj-PRO-9a;(B) Bj-PRO-11e; (C) Bj-PRO-12b; (D) Bj-PRO-13a. *P < 0.05; **P < 0.01 and ***P < 0.001 compared to basal ASS enzymatic activity (absence of Bj-PROs).
Fig. 4. Bj-PRO-induced NO production by HEK293 cells. HEK293 were seeded into Petri dishes in appropriate medium and incubated for 24 h. After this, cells were incubatedin serum-free medium with increased concentrations (0.03–3 �M) of Bj-PROs (A–D). Extracellular medium was collected for NO content determination. Cells were lysed withRIPA buffer to measure intracellular NO products by a chemiluminescence assay. (A) Bj-PRO-9a; (B) Bj-PRO-11e; (C) Bj-PRO-12b; (D) Bj-PRO-13a. (E) Bj-PRO-13a (0.1 �M)in the absence or presence of the ASS inhibitor (10 mM MDLA), an antagonist of bradykinin-B2 receptors (1 �M HOE-140), of �2-adrenergic receptors (10 �M yohimbine),M1 (1 mM pirenzepine), M2 (100 �M gallamine) and M3 mAChRs (10 �M 4-DAMP). Bj-PRO-13a-induced NO production was considered as 100%. Total NO concentration isreported as mean values ± S.E. of three experiments. *P < 0.05 and ***P < 0.001.

K.L.P. Morais et al. / Peptides 48 (2013) 124– 133 131
F k heigw nd Fle( epend
((utmtv
4
4i
e9aoctcaet
ig. 5. Bj-PROs-induced [Ca2+]i elevations in HEK293 cells. Changes of maximal peaere measured in HEK293 cells by microfluorimetry using the FlexStation III device a
C) Bj-PRO-12b; (D) Bj-PRO-13a. The data shown are mean values ± S.E. of three ind
pD2 values) of 7.15 ± 0.11, 8.63 ± 0.15 and 7.82 ± 0.17, respectivelyFig. 5), whereas no calcium response was detected following stim-lation of cells with Bj-PRO-9a (Fig. 5). Moreover, we observed thathe increases of [Ca2+]i were instantaneous and transient, i.e., the
aximal peak response of [Ca2+]i was reached immediately afterhe stimulation of HEK293 cells by Bj-PROs, returning to baselinealues within 120 s (data not shown).
. Discussion
.1. The cardiovascular action of Bj-PROs is unrelated tonhibition of ACE activity or BK potentiation
In this study, we used SHRs to evaluate the cardiovascular effectsvoked by intravenous injection of 71 nmol/kg of Bj-PROs. Bj-PRO-a is already known to affect the blood pressure of hypertensivenimal models [36,38]. Strikingly, it was unexpected that the dosef 71 nmol/kg, which is approximately 15-fold lower than thatompletely blocking ACE activity [21], would cause this substan-ial activity on arterial pressure. The Bj-PROs 11e, 12b and 13a
aused potent and long-lasting reduction of arterial blood pressurend heart rates of hypertensive animals, being similar to responseslicited by Bj-PROs 5a and 10c [16,17] (Fig. 1; Table 1). Sincehe cardiac output contributes to pressoric levels, it is plausiblehts of [Ca2+]i responses induced by several concentrations of Bj-PROs (0.003–1 �M)xStation Calcium Assay Kit (Molecular Devices Corp.). (A) Bj-PRO-9a; (B) Bj-PRO-11e;ent experiments.
that bradycardia may have contributed to the observed antihyper-tensive effect. This is congruent with below-listed experimentalevidence showing that cardiovascular activity of Bj-PROs is notrelated to ACE inhibition or BK potentiation.
I. The bradycardia that happened concomitant to the antihy-pertensive effect of these peptides is unrelated to BK-effectpotentiation, since BK-mediated hypotension is associated withtachycardia by the unloading of baroreceptors [1]. Accordingly,following identification of ASS as molecular target for Bj-PRO-10c actions, resulting in increases of NO production [10], it wassuggested that this oligopeptide also regulates baroreflex sen-sitivity in SHR. Its central action on baroreceptors results froma neurochemical effect, such as the releasing of GABA [23], themain inhibitor of neuronal firing that plays a key role in thecentral control of cardiovascular homeostasis [9].
II. Cardiovascular actions may also be related to a central effect,as recently suggested [2]. Biodistribution assays revealed thatBj-PRO-10c was able to cross the blood-brain barrier [33].
Furthermore, arterial pressure of SHR was reduced followingcentral injection of Bj-PRO-10c [22]. Probably, the burst of NOrelease after ASS activation by Bj-PRO-10c exerts its functionon regions/nuclei in the central nervous system involved in
1 eptide
I
sPat
4
nHCtan
aAbmacIpbcpp
r1o
[
[
[
32 K.L.P. Morais et al. / P
cardiovascular regulation, such as the paraventricular nucleus(PVN) [27].
III. There is no apparent relationship between the pharmacolog-ical properties of Bj-PROs. Recent research indicated that theeffects of Bj-PROs 5a and 10c on arterial pressure occurredwithout changing the standard responses to the administrationof angiotensin I (Ang I) and BK [16,17]. Moreover, no furtherchanges were observed during antihypertensive effects elicitedby Bj-PRO-9a (unpublished data). According to the current view,the results presented in Fig. 1 and Fig. 2 and in Table 1 are rathersurprising. Analysis of the results obtained in the presence of Bj-PROs provide additional support to a concept of being unrelatedto ACE inhibition. Herein, we demonstrate that only Bj-PRO-9aand Bj-PRO-12b significantly potentiated the hypotensive effectof BK (Fig. 2 and Table 1). In agreement, a synthetic compoundsimilar to Bj-PRO-9a, presented high BK-potentiating activityand weak ACE inhibition [29]. Accordingly, we have shownthat some of the investigated peptides, such as i.e., Bj-PRO-7a, markedly potentiate the hypotensive effect of BK, withoutchanging ACE activity in vitro or in vivo [16]. Moreover, in linewith our study, the capacities of several peptides from snakevenom in potentiating BK effects are not directly proportionalto their efficacies in inhibiting ACE in vitro [3,13,16,29] (Table 1).This is especially true when we compare the inhibitory speci-ficities of these compounds for the C-active site of ACE. Exceptfor Bj-PROs 9a and 10c, specificities of the remaining Bj-PROsare about 50–1000-fold lower than that of captopril [12].
V. Preliminary results revealed that Bj-PROs 5a and 10c improvethe renal function of SHR by increasing the urinary flow rate andsodium excretion [18]. This is in turn an indicative for Bj-PROeffects through multiple targets, acting on renal arterioles as avasodilator [17] and on renal tubules by augmenting sodiumand water excretion [18].
Taken together, evidences obtained in the present and previoustudies clearly confirm that the cardiovascular activity of all Bj-ROs can be dissociated regarding their inhibitory activities on ACEnd potentiation of BK-mediated effects, suggesting an update inhis classical concept.
.2. Mechanisms involved in Bj-PROs cardiovascular effects
Bj-PRO-9a – This peptide did not interfere with ASS activity,or it stimulated NO production or elicited [Ca2+]i responses inEK293 cells. Although Bj-PRO-9a is an efficient inhibitor of the-terminal site of the ACE [3], we cannot rule out the possibilityhat this peptide could induce its antihypertensive effects by inter-cting with tissue-specific receptors, which so far are unknown andot expressed in HEK293 cells.
Bj-PRO-11e and Bj-PRO-12b – While Bj-PRO-11e poorly potenti-tes BK-induced hypotensive effects and is inefficient in inhibitingCE activity, Bj-PRO-12b potentiates BK-exerted effects. However,oth oligopeptides induce [Ca2+]i mobilization. This inotropic effectight be induced by these two Bj-PROs in a plenty of tissues. Inter-
ctions of these peptides with central or systematic regulators ofardiovascular system, such as Ca2+/calmodulin-dependent kinaseI [24,25], could explain the effects observed in vivo. ASS is sup-osedly not a physiological target of Bj-PRO-12b, since interactionsetween Bj-PRO-12b and ASS are only observed at high peptideoncentrations. Moreover, there was no significant increase in NOroduction in HEK293 cells treated with Bj-PRO-12b when com-ared to untreated cells.
Bj-PRO-13a – Our in vitro results showed that Bj-PRO-13aesembles similar activity to that previous described for Bj-PRO-0c [10,22]. Bj-PRO-13a-mediated effects include thus increasingf ASS activity, NO production and [Ca2+]i mobilization in order to
[
s 48 (2013) 124– 133
achieve the described cardiovascular effects. Furthermore, Bj-PRO-13a stimulates the M3 muscarinic receptor (mAChR). The maineffects mediated by Bj-PRO-13a agonism on peripheral mAChRs,specifically of M3 mAChRs, might involve the control of vascu-lar tonus, induction of negative cardiac chronotropy and smoothmuscle relaxation [39].
5. Conclusion
This study shows that the cardiovascular action evoked by someBj-PROs, characterized by on-going in vivo, ex vivo and in vitrostudies, result from synergic interactions of each oligopeptide witheither ACE and/or with ASS, and certainly with other unknowntargets. There is no direct correlation between BK potentiation,cardiovascular activity and inhibition of the ACE, thus suggestingthat Bj-PROs act through distinct pathways, which do not nec-essarily include ACE inhibition. Therefore, Bj-PROs are useful aspharmacological tools, possibly revealing novel therapeutic targetsfor treatment of hypertension and other related diseases as well ascontributing to the better understanding of the pathophysiologyunderlying cardiovascular diseases.
Acknowledgements
This work was supported by grants from Fundac ão de Amparoà Pesquisa do Estado de São Paulo (FAPESP), through the Cen-ter for Applied Toxinology (CAT-CEPID) and Conselho Nacionalde Pesquisa (CNPq), Brazil. We are grateful to Dr. Carlos Hen-rique Xavier (ICB-UFG) for critically reviewing the manuscript. Wethank Maria José da Silva and Isaías Franc a da Silva for secretarialassistance and Maria Aparecida Siqueira and Roberto Siqueira fortechnical support during experiments.
References
[1] Bunag RD, Walaszek EJ, Mueting N. Sex differences in reflex tachycar-dia induced by hypotensive drugs in unanesthetized rats. Am J Physiol1975;229:652–6.
[2] Camargo AC, Ianzer D, Guerreiro JR, Serrano SM. Bradykinin-potentiating pep-tides: beyond captopril. Toxicon 2012;59:516–23.
[3] Cotton J, Hayashi MA, Cuniasse P, Vazeux G, Ianzer D, De Camargo AC, et al.Selective inhibition of the C-domain of angiotensin I converting enzyme bybradykinin potentiating peptides. Biochemistry 2002;41:6065–71.
[4] de Oliveira EF, Guerreiro JR, Silva CA, Benedetti GF, Lebrun I, Ulrich H,et al. Enhancement of the citrulline-nitric oxide cycle in astroglioma cellsby the proline-rich peptide-10c from Bothrops jararaca venom. Brain Res2010;1363:11–9.
[5] Feelisch M, Rassaf T, Mnaimneh S, Singh N, Bryan NS, Jourd’Heuil D, et al.Concomitant S-, N-, and heme-nitros(yl)ation in biological tissues and fluids:implications for the fate of NO in vivo. Faseb J 2002;16:1775–85.
[6] Ferreira SH, Bartelt DC, Greene LJ. Isolation of bradykinin-potentiating peptidesfrom Bothrops jararaca venom. Biochemistry 1970;9:2583–93.
[7] Georgopapadakou NH, Liu FY, Ryono DE, Neubeck R, Ondetti MA. Chemicalmodifications of the active site of Streptomyces R61 DD-carboxypeptidase. EurJ Biochem 1981;115:53–7.
[8] Gomes CL, Konno K, Conceicao IM, Ianzer D, Yamanouye N, Prezoto BC, et al.Identification of novel bradykinin-potentiating peptides (BPPs) in the venomgland of a rattlesnake allowed the evaluation of the structure-function rela-tionship of BPPs. Biochem Pharmacol 2007;74:1350–60.
[9] Gordon FJ, Sved AF. Neurotransmitters in central cardiovascular regulation:glutamate and GABA. Clin Exp Pharmacol Physiol 2002;29:522–4.
10] Guerreiro JR, Lameu C, Oliveira EF, Klitzke CF, Melo RL, Linares E, et al.Argininosuccinate synthetase is a functional target for a snake venom anti-hypertensive peptide: role in arginine and nitric oxide production. J Biol Chem2009;284:20022–33.
11] Hao G, Xie L, Gross SS. Argininosuccinate synthetase is reversibly inactivatedby S-nitrosylation in vitro and in vivo. J Biol Chem 2004;279:36192–200.
12] Hayashi MA, Camargo AC. The Bradykinin-potentiating peptides from venomgland and brain of Bothrops jararaca contain highly site specific inhibitors of
the somatic angiotensin-converting enzyme. Toxicon 2005;45:1163–70.13] Hayashi MA, Murbach AF, Ianzer D, Portaro FC, Prezoto BC, Fernandes BL,et al. The C-type natriuretic peptide precursor of snake brain containshighly specific inhibitors of the angiotensin-converting enzyme. J Neurochem2003;85:969–77.

eptide
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
K.L.P. Morais et al. / P
14] Ianzer D, Konno K, Marques-Porto R, Vieira Portaro FC, Stocklin R, Martins deCamargo AC, et al. Identification of five new bradykinin potentiating peptides(BPPs) from Bothrops jararaca crude venom by using electrospray ionizationtandem mass spectrometry after a two-step liquid chromatography. Peptides2004;25:1085–92.
15] Ianzer D, Konno K, Xavier CH, Stocklin R, Santos RA, de Camargo AC, et al.Hemorphin and hemorphin-like peptides isolated from dog pancreas andsheep brain are able to potentiate bradykinin activity in vivo. Peptides2006;27:2957–66.
16] Ianzer D, Santos RA, Etelvino GM, Xavier CH, de Almeida Santos J, Mendes EP,et al. Do the cardiovascular effects of angiotensin-converting enzyme (ACE) Iinvolve ACE-independent mechanisms? new insights from proline-rich pep-tides of Bothrops jararaca. J Pharmacol Exp Ther 2007;322:795–805.
17] Ianzer D, Xavier CH, Fraga FC, Lautner RQ, Guerreiro JR, Machado LT, et al. BPP-5a produces a potent and long-lasting NO-dependent antihypertensive effect.Ther Adv Cardiovasc Dis 2011;5:281–95.
18] Ianzer DA, Silveira KD, Teixeira MM, Simões e Silva AC, Camargo ACM. BPPs fromBothrops jararaca venom change renal function of SHR. FASEB J 2011;25:643.9.
19] Kakar P, Lip GY. Towards improving the clinical assessment and managementof human hypertension: An overview from this Journal. J Hum Hypertens2006;20:913–6.
20] Kanagy NL. Alpha(2)-adrenergic receptor signalling in hypertension. Clin Sci(Lond) 2005;109:431–7.
21] Keim Jr GR, Kirpan J, Peterson AE, Murphy BF, Hassert Jr GL, Poutsiaka JW. Inhi-bition of angiotensin I-initiated hemodynamic changes in anesthetized dogsby a synthetic nonapeptide. Proc Soc Exp Biol Med 1972;140:149–52.
22] Lameu C, Hayashi MA, Guerreiro JR, Oliveira EF, Lebrun I, Pontieri V, et al. Thecentral nervous system as target for antihypertensive actions of a proline-richpeptide from Bothrops jararaca venom. Cytometry A 2010;77:220–30.
23] Lameu C, Pontieri V, Guerreiro JR, Oliveira EF, da Silva CA, Giglio JM, et al. Brainnitric oxide production by a proline-rich decapeptide from Bothrops jararacavenom improves baroreflex sensitivity of spontaneously hypertensive rats.Hypertens Res 2010;33:1283–8.
24] Leenders AG, Sheng ZH. Modulation of neurotransmitter release by the secondmessenger-activated protein kinases: implications for presynaptic plasticity.Pharmacol Ther 2005;105:69–84.
25] Maier LS. Ca(2+)/calmodulin-dependent protein kinase II (CaMKII) in the heart.Adv Exp Med Biol 2012;740:685–702.
26] Majumder P, Trujillo CA, Lopes CG, Resende RR, Gomes KN, Yuahasi KK, et al.New insights into purinergic receptor signaling in neuronal differentiation,neuroprotection, and brain disorders. Purinergic Signal 2007;3:317–31.
[
s 48 (2013) 124– 133 133
27] Martins-Pinge MC, Mueller PJ, Foley CM, Heesch CM, Hasser EM. Regulation ofarterial pressure by the paraventricular nucleus in conscious rats: interactionsamong glutamate, GABA, and nitric oxide. Front Physiol 2012;3:490.
28] Morais KL, Hayashi MA, Bruni FM, Lopes-Ferreira M, Camargo AC, Ulrich H,et al. Bj-PRO-5a, a natural angiotensin-converting enzyme inhibitor, promotesvasodilatation mediated by both bradykinin B(2)and M1 muscarinic acetyl-choline receptors. Biochem Pharmacol 2011;81:736–42.
29] Mueller S, Gothe R, Siems WD, Vietinghoff G, Paegelow I, Reissmann S. Poten-tiation of bradykinin actions by analogues of the bradykinin potentiatingnonapeptide BPP9alpha. Peptides 2005;26:1235–47.
30] Murayama N, Hayashi MA, Ohi H, Ferreira LA, Hermann VV, Saito H, et al.Cloning and sequence analysis of a Bothrops jararaca cDNA encoding a precur-sor of seven bradykinin-potentiating peptides and a C-type natriuretic peptide.Proc Natl Acad Sci U S A 1997;94:1189–93.
31] Negraes PD, Lameu C, Hayashi MA, Melo RL, Camargo AC, Ulrich H. The snakevenom peptide Bj-PRO-7a is a M1 muscarinic acetylcholine receptor agonist.Cytometry A 2011;79:77–83.
32] Ondetti MA, Williams NJ, Sabo EF, Pluscec J, Weaver ER, Kocy O. Angiotensin-converting enzyme inhibitors from the venom of Bothrops jararaca. Isolation,elucidation of structure, and synthesis. Biochemistry 1971;10:4033–9.
33] Silva CA, Portaro FC, Fernandes BL, Ianzer DA, Guerreiro JR, Gomes CL, et al.Tissue distribution in mice of BPP 10c, a potent proline-rich anti-hypertensivepeptide of Bothrops jararaca. Toxicon 2008;51:515–23.
34] Solomonson LP, Flam BR, Pendleton LC, Goodwin BL, Eichler DC. The caveo-lar nitric oxide synthase/arginine regeneration system for NO production inendothelial cells. J Exp Biol 2003;206:2083–7.
35] Somsen RJ, Jennings JR, Van der Molen MW. The cardiac cycle time effect revis-ited: temporal dynamics of the central-vagal modulation of heart rate in humanreaction time tasks. Psychophysiology 2004;41:941–53.
36] Svendsen UG. The importance of thymus for hypertension and hypertensivevascular disease in rats and mice. Acta Pathol Microbiol Scand Suppl 1978:1–15.
37] Tashima AK, Zelanis A, Kitano ES, Ianzer D, Melo RL, Rioli V, et al. Peptidomicsof three Bothrops snake venoms: insights into the molecular diversification ofproteomes and peptidomes. Mol Cell Proteomics 2012;11:1245–62.
38] Vollmer RR, Boccagno JA, Harris DN, Murthy VS. Hypotension induced by inhi-
bition of angiotensin-converting enzyme in pentobarbital-anesthetized dogs.Eur J Pharmacol 1978;51:39–45.39] Wess J. Muscarinic acetylcholine receptor knockout mice: novel pheno-types and clinical implications. Annu Rev Pharmacool Toxicol 2004;44:423–50.





























