Embed Size (px)
Citation preview

J. Anat. (1995) 187, pp. 279-292, with 14 figures Printed in Great Britain
Glycobiology: 'The function of sugar in the IgG molecule'
R. A. DWEK, A. C. LELLOUCH AND M. R. WORMALD
Department of Biochemistry, University of Oxford, UK
(Accepted 7 February 1995)
ABSTRACT
Immunoglobulin G (IgG) is glycosylated in both the Fc and the Fab regions of the protein with aheterogeneous ensemble of structures (glycoforms) that is both highly reproducible (i.e. nonrandom) and sitespecific. In normal IgG, the 2 highly conserved oligosaccharides of the Fc region are found buried betweenthe CH2 domains, forming specific protein-saccharide interactions with the Fc protein surface. One of thefunctions attributed to the Fc oligosaccharides of normal IgG is to maintain the conformationalarrangements of the Fc domains as well as the hinge regions. These structural features are necessary for Fceffector functions such as Clq and monocyte binding. A hallmark of rheumatoid arthritis (RA) patients is adramatic increase in the presence of serum IgG containing Fc oligosaccharides lacking an outer armgalactose residue (termed 'GO' glycoforms). The increased level of GO has been shown to be directly relatedto the pathogenesis of RA. Nuclear magnetic resonance relaxation studies of the Fc region from normal andRA IgG, as well as examination of x-ray structures, show that the GO oligosaccharides have an increasedmobility resulting from the loss of binding between the GO oligosaccharide and the Fc protein surface. Fromthese observations it follows that regions of the protein surface that are normally covered by theoligosaccharide are revealed. The newly accessible protein surface could have lectin-like activity and also beinherently antigenic. In addition, the more mobile GO oligosaccharide can be recognised by mannose bindingprotein. As the mannose binding protein can activate complement, and the Fc oligosaccharide would notnormally be accessible to protein recognition, this finding might suggest a specific role for the GO glycoformin inflammation when the appropriate IgG glycoforms are clustered.
Key words: Glycosylation; antibodies; rheumatoid arthritis; nuclear magnetic resonance; lectins.
INTRODUCTION
There is no single unifying function for oligo-saccharides. They can modify the intrinsic propertiesof proteins to which they are attached by altering thestability, dynamic properties, protease resistance orquaternary structure. The large size of oligosac-charides (each monosaccharide residue is 0.6 nmacross, so a hexasaccharide could extend to about3.6 nm), may allow them to cover functionallyimportant areas of proteins, to modulate the inter-actions ofglycoconjugates with other molecules and toaffect the rate of processes which involve conforma-tional changes. Glycosylation is highly sensitive toalterations in cellular function, and abnormal glyco-sylation is diagnostic of a number of disease statesincluding rheumatoid arthritis and cancer. In additionto their roles as modulators of protein structure and
function, a major function of oligosaccharides is toserve as recognition markers. The control of glyco-sylation by the cell affords, in principle, a means ofputting the same recognition markers on quitedifferent proteins without having to code the in-formation into the DNA of that protein. Temporalcontrol of the levels and activities ofenzymes involvedin the glycosylation process may also be important.This can result in proteins produced at different timesin a cell having different glycosylation patterns.
In this article we will illustrate many of the functionsresulting from the presence of an oligosaccharideattached to a protein, as exemplified by the immuno-globulin G (IgG) molecule. Of particular significanceare the conserved oligosaccharides found in the Fcregion of the molecule. The Fc region is involved inthe elimination of antigen antibody complexes follow-ing the initial recognition of antigen. Among the
Correspondence to Professor R. A. Dwek, The Glycobiology Institute, Department of Biochemistry, University of Oxford, South ParksRoad, Oxford OXI 3QU, UK.
279
280 R. A. Dwek, A. C. Lellouch and M. R. Wormald
notable features we will discuss are: (1) the structuralorganisation of the protein and oligosaccharidecomponents of the IgG molecule; (2) the hetero-geneous yet nonrandom nature of the IgG glycosyl-ation; (3) the possible structural and functionalroles for the oligosaccharides of normal IgG; and (4)the change in oligosaccharide populations (glyco-forms) with rheumatoid arthritis (RA) and thepossible consequences including pathogenic glyco-forms, generation of new antigenic determinants andrecognition of RA associated IgG oligosaccharides bythe mannose binding protein (MBP).
THE DOMAIN STRUCTURE OF IgG
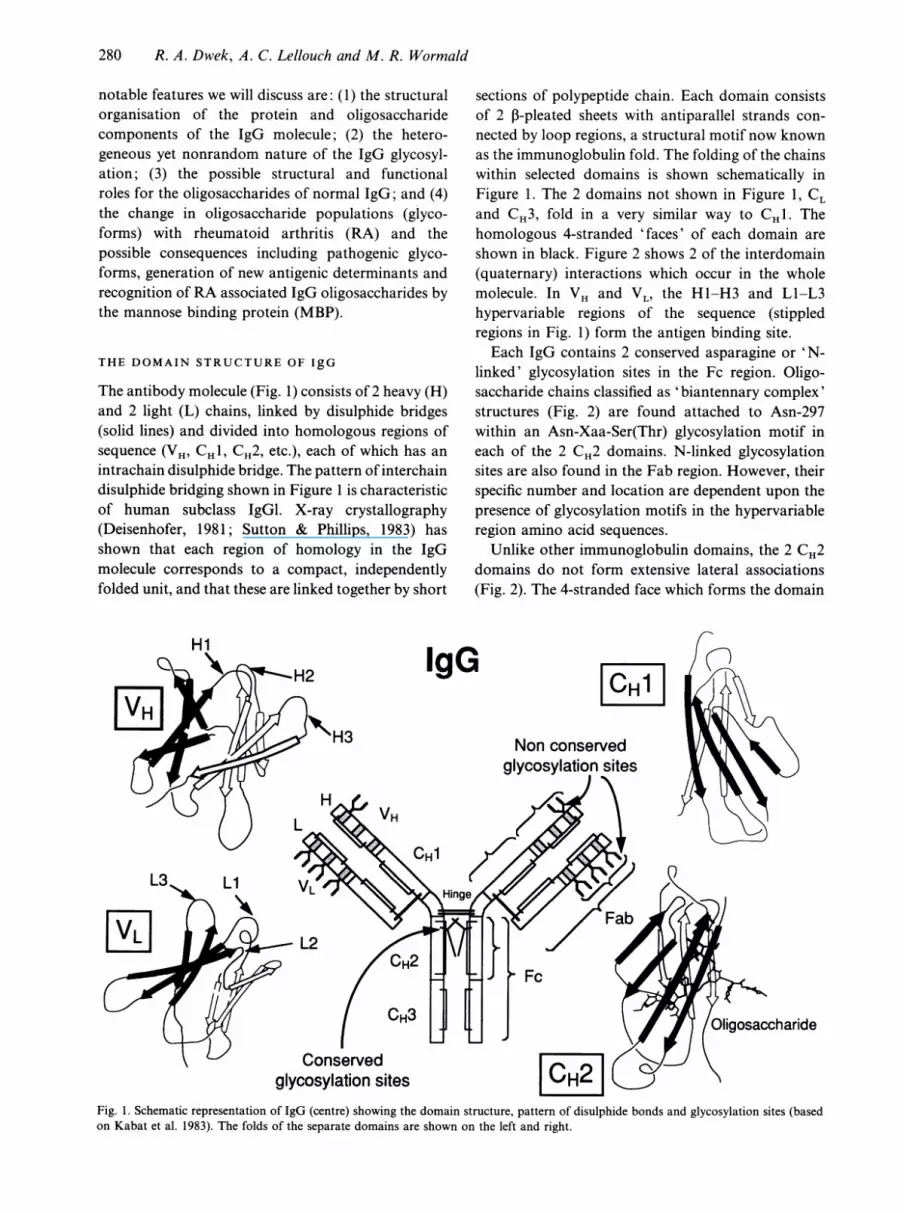
The antibody molecule (Fig. 1) consists of 2 heavy (H)and 2 light (L) chains, linked by disulphide bridges(solid lines) and divided into homologous regions ofsequence (VH, CHI, CH2, etc.), each of which has anintrachain disulphide bridge. The pattern of interchaindisulphide bridging shown in Figure 1 is characteristicof human subclass IgGl. X-ray crystallography(Deisenhofer, 1981; Sutton & Phillips, 1983) hasshown that each region of homology in the IgGmolecule corresponds to a compact, independentlyfolded unit, and that these are linked together by short
IgG
sections of polypeptide chain. Each domain consistsof 2 n-pleated sheets with antiparallel strands con-nected by loop regions, a structural motif now knownas the immunoglobulin fold. The folding of the chainswithin selected domains is shown schematically inFigure 1. The 2 domains not shown in Figure 1, CLand CH3, fold in a very similar way to CH1. Thehomologous 4-stranded 'faces' of each domain areshown in black. Figure 2 shows 2 of the interdomain(quaternary) interactions which occur in the wholemolecule. In VH and VL, the H1-H3 and L1-L3hypervariable regions of the sequence (stippledregions in Fig. 1) form the antigen binding site.Each IgG contains 2 conserved asparagine or 'N-
linked' glycosylation sites in the Fc region. Oligo-saccharide chains classified as 'biantennary complex'structures (Fig. 2) are found attached to Asn-297within an Asn-Xaa-Ser(Thr) glycosylation motif ineach of the 2 CH2 domains. N-linked glycosylationsites are also found in the Fab region. However, theirspecific number and location are dependent upon thepresence of glycosylation motifs in the hypervariableregion amino acid sequences.
Unlike other immunoglobulin domains, the 2 CH2domains do not form extensive lateral associations(Fig. 2). The 4-stranded face which forms the domain
ICH
Non conservedglycosylation sites
\ L'/ Conservedglycosylation sites CHA
Fig. 1. Schematic representation of IgG (centre) showing the domain structure, pattern of disulphide bonds and glycosylation sites (basedon Kabat et al. 1983). The folds of the separate domains are shown on the left and right.

Glycobiology
* - 4 nm - >
a(1-3) arma Neu5Ac
b Gal
c GlcNW
d Man
a(1-6) armNeu5Ac
I Gal
kc GIcNAcI +
I GIcNAc Man
Man
GIcNAc
GIcNAc*- Fuc
v\^ Asn/x/Ser(Thr)vxA
a'
b'
co
d'
h
Fig. 2. The size of the oligosaccharide, typical dimensions shown on the left, is approximately the size of an Ig domain. Details of 2 of thedomain-pair interactions from the rabbit IgG Fc crystal structure (Sutton & Philips, 1983) are shown on the right. The association between5- and 4-stranded twisted 1-sheets, of VH and VL respectively, forms a 'barrel' of 9 strands and brings the hypervariable loop regions (H1,Ll, etc.) together to form the antigen binding site. The arrows represent the strands of 3-sheet, and the solid bars are the intrachain disulphidebridges which link the 2 1-sheets of each domain. Potential oligosaccharide attachment sites are illustrated in the loops (right, top). The 2CH2 domains are uniquely isolated. The branched oligosaccharide chains cover part of their 4-stranded faces and occupy the space betweenthem (right, bottom).
pair interface in the CH1:CL and the CH3:CH3interactions is partly covered by the branched oligo-saccharide chain in the CH2 domain. The resultinginterstitial region accommodates the complex oligo-saccharides (Fig. 3). The quaternary orientation ofthe CH2 domains is subject to the following twoconstraints: (1) the hinge disulphide limits the N-terminal freedom of the CH2 domains, and (2) theCH3 domain limits the C-terminal freedom of theCH2 domains (Fig. 3). Using computer modelling(Leatherbarrow, 1983), it is possible to construct an Fcfragment in which the 4-stranded faces of the CH2domains interact exactly as in the CH3 region.However, in this arrangement the N-termini of the 2CH2 domains are too far apart to allow the formationof the hinge disulphide bonds. The disulphide bondconstraint in the hinge therefore provides a rationale
for the large interstitial space between the 2 CH2domains and for the fact that there are no extensivelateral interactions between them. The presence of theoligosaccharides may limit further the possible orient-ations of the CH2 domains due to oligosaccharide-oligosaccharide steric interactions.Each of the 2 biantennary complex oligosaccharide
chains linked to Asn 297 range in size from 8 to 10saccharide residues and are found packed within thespace between the CH2 domains (Fig. 3). One shouldnote that the relative sizes of a single immunoglobulindomain and a fully extended complex oligosaccharide,such as those found on CH2, are similar (Fig. 2). In thestructure of the rabbit IgG Fc (Sutton & Phillips,1983), the a(1-3) oligosaccharide arm of one oligo-saccharide interacts with the trimannose core of theopposing oligosaccharide in the interstitial space. In
e
f
3nm
!I
1 nm
/l
281
9

282 R. A. Dwek, A. C. Lellouch and M. R. Wormald
.*. ~ . ........
.v~~~~~~~~~~N
\ ~~~~~~~~~~~wi
L. H'vlrj e , X d, 0 . 0 ,, 0 00 \t:; W j ! t!: .; : :' i, 0 50~~~~~~~~~~~~~VU
:<St;WAD0;, , ,00 f4' t 4 4Xif ;,-;.\'S000. . t ,i, .- 0 i 'd0S t000;:0:00tV~~~~~~~~~~~~~~~~'
Fig. 3. Human IgG Fc crystal structure (Deisenhofer, 1981), showing the peptide backbone and the conserved oligosaccharides. The pseudo-C2 axis runs vertically through the centre of the molecule.
addition, it is seen that the a(l-6) antenna of eacholigosaccharide interacts with both hydrophobic andpolar residues Phe 243 (Man 5 and GlcNAc 6), Pro244/245 (Gal 7) and Thr 260 (GlcNAc 6 and Gal 7) onthe Fc domain surface. This specific protein-carbohydrate interaction involves the presence ofsugar binding subsites on the protein surface involvingboth aromatic residues and hydrogen bond formationbetween the protein and water molecules. To date thisis the only X-ray crystal structure in which extensiveinteractions between an oligosaccharide and theprotein to which the oligosaccharide is covalentlybound, are observed. This interaction will be discussedin more detail later in this article as it relates to themechanism of rheumatoid arthritis.-pathogenesis.
GLYCOSYLATED IgG GIVES RISE TO MULTIPLEGLYCOFORMS AND IS SITE SPECIFIC
Post-translational modification of a polypeptide byN-glycosylation leads to the formation of discretesubsets or glycoforms which may have differentphysical and biochemical properties. Glycoformsfrom the same cell arise from the modification of a
single polypeptide by the action of glycosyl trans-ferases and glycosidases. As a result of this, manystructures may be found at the same glycosylationsite. Greater complexity emerges if the polypeptidecontains more than one glycosylation site. The patternof oligosaccharide heterogeneity during a constantphysiological condition is reproducible, not random.
Analysis of the N-linked oligosaccharides chemic-ally released from serum IgG, Fab and Fc hasshown that, on average, 2.8 N-linked oligosaccharidesare attached to each IgG molecule of which 2.0 arelocated in the Fc (Parekh et al. 1985). The additionaloligosaccharides are found in the hypervariableregions in the Fab.
Exoglycosidase sequencing of oligosaccharideschemically released from human serum IgG (Parekhet al. 1985) has shown that IgG carries at least 30different N-linked oligosaccharides all of which are ofthe complex biantennary type (Fig. 4). There areglycosylation site specific differences between the Faband Fc. Characteristics of Fc N-glycosylation includea low incidence (10%) of monosialylated structures,the absence of disialylated structures, a low incidenceof cores carrying a 'bisecting' GIcNAc, a high

Glycobiology
Neu tralss _ ____\__
. __ _
<_:_ _ _,{
_:^ _ ____\
_=_ _{ Monosialyated^ . _a__-\__ _ -.__/___
_:ta___Ns
a__r__/___
<_:, _ _
a
_:{o___
__. _ _ ^
_5______
, _ _
___\_\_ _ _
. _ _ __ _r___\
_r _
i . _ _, _ _ _s\___e___
i___\___
i * _ _
v __
_So___
-__1__{r-__
DisiatytatedA &
V
Fig. 4. Primary sequences of the N-linked oligosaccharides associated with IgG.
incidence of cores carrying fucose and heterogeneityin the galactose residues. In general, the Fc oligo-saccharides are mainly restricted to biantennaryoligosaccharides terminating in 2, 1 and 0 galactose
residues (termed G2, GI and GO respectively). FabN-glycosylation is characterised by a high incidence ofdi- and monosialylated structures and of cores withthe 'bisecting' GlcNAc residue (Rademacher et al.
NeuSAcx2-m 66Ipl-s. 4cGLcNAcl2Manal Fuctlva 6 * 6
GlcNAcpl .'4Manp1-4GIcNAc,l1-o 4(3cNAcM03 * n
NeuSAca 2- 6Gdpl - 4GLcNAc 1-a2M- no7# 6A s A 40 3 2 1
283
284 R. A. Dwek, A. C. Lellouch and M. R. Wormald
1986). Similar heterogeneity is found on analysis ofmyeloma and hybridoma-derived, as well as poly-clonal, IgG. The observed differences in Fab andFc glycosylation may reflect differences in the3-dimensional environment which could limit process-ing enzyme accessibility during glycosylation.
A ROLE FOR IgG GLYCOSYLATION
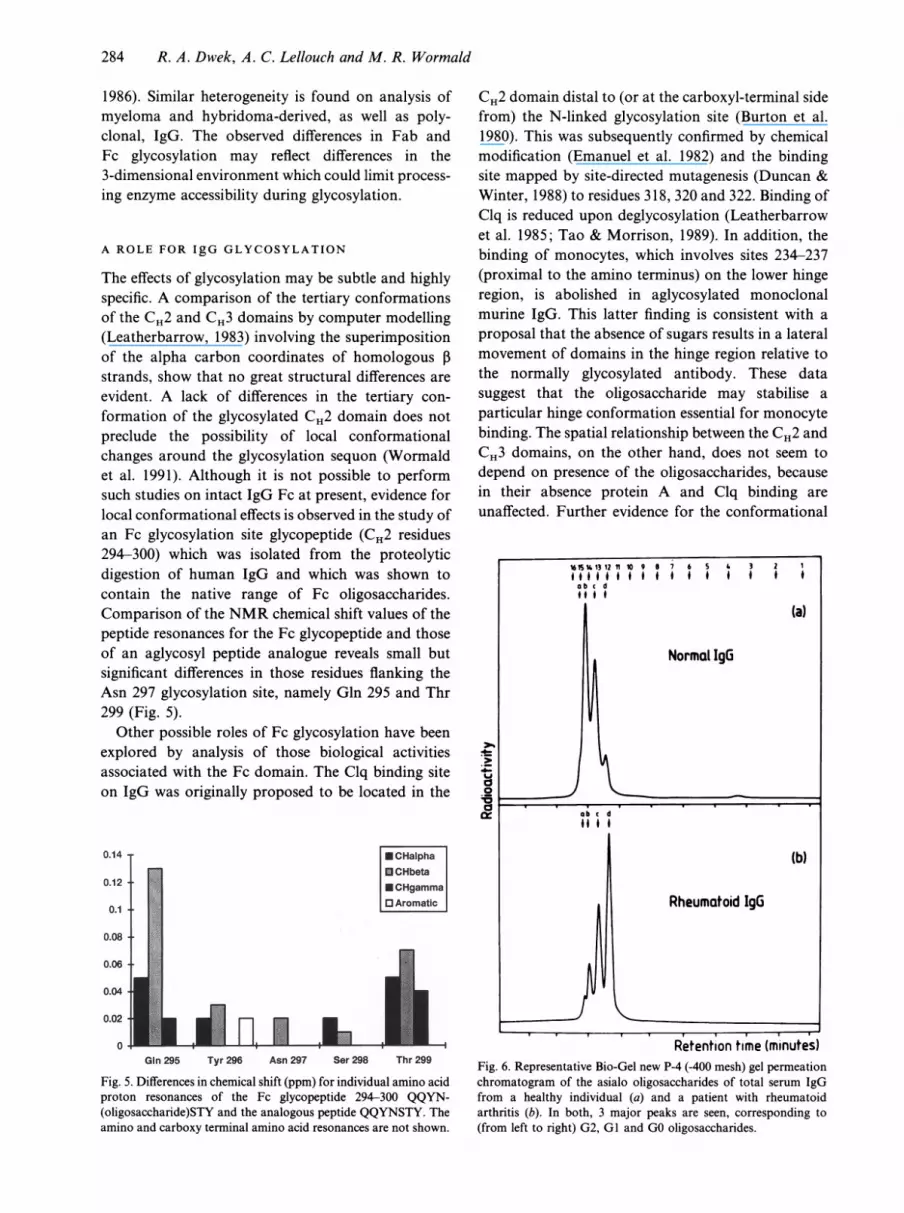
The effects of glycosylation may be subtle and highlyspecific. A comparison of the tertiary conformationsof the CH2 and CH3 domains by computer modelling(Leatherbarrow, 1983) involving the superimpositionof the alpha carbon coordinates of homologous 1Bstrands, show that no great structural differences areevident. A lack of differences in the tertiary con-formation of the glycosylated CH2 domain does notpreclude the possibility of local conformationalchanges around the glycosylation sequon (Wormaldet al. 1991). Although it is not possible to performsuch studies on intact IgG Fc at present, evidence forlocal conformational effects is observed in the study ofan Fc glycosylation site glycopeptide (CH2 residues294-300) which was isolated from the proteolyticdigestion of human IgG and which was shown tocontain the native range of Fc oligosaccharides.Comparison of the NMR chemical shift values of thepeptide resonances for the Fc glycopeptide and thoseof an aglycosyl peptide analogue reveals small butsignificant differences in those residues flanking theAsn 297 glycosylation site, namely Gln 295 and Thr299 (Fig. 5).
Other possible roles of Fc glycosylation have beenexplored by analysis of those biological activitiesassociated with the Fc domain. The Clq binding siteon IgG was originally proposed to be located in the
0.14 T
0.12
0.1
0.08
0.06
0.04
* CHalphaOCHbeta
* CHgammaAromatic
0.02 -_
0 n I h ,
Gin 295 Tyr296 Asn 297 Ser 298 Thr 299
Fig. 5. Differences in chemical shift (ppm) for individual amino acidproton resonances of the Fc glycopeptide 294-300 QQYN-(oligosaccharide)STY and the analogous peptide QQYNSTY. Theamino and carboxy terminal amino acid resonances are not shown.
CH2 domain distal to (or at the carboxyl-terminal sidefrom) the N-linked glycosylation site (Burton et al.1980). This was subsequently confirmed by chemicalmodification (Emanuel et al. 1982) and the bindingsite mapped by site-directed mutagenesis (Duncan &Winter, 1988) to residues 318, 320 and 322. Binding ofClq is reduced upon deglycosylation (Leatherbarrowet al. 1985; Tao & Morrison, 1989). In addition, thebinding of monocytes, which involves sites 234-237(proximal to the amino terminus) on the lower hingeregion, is abolished in aglycosylated monoclonalmurine IgG. This latter finding is consistent with aproposal that the absence of sugars results in a lateralmovement of domains in the hinge region relative tothe normally glycosylated antibody. These datasuggest that the oligosaccharide may stabilise aparticular hinge conformation essential for monocytebinding. The spatial relationship between the CH2 andCH3 domains, on the other hand, does not seem todepend on presence of the oligosaccharides, becausein their absence protein A and Clq binding areunaffected. Further evidence for the conformational
Retention time (minutes)Fig. 6. Representative Bio-Gel new P-4 (-400 mesh) gel permeationchromatogram of the asialo oligosaccharides of total serum IgGfrom a healthy individual (a) and a patient with rheumatoidarthritis (b). In both, 3 major peaks are seen, corresponding to(from left to right) G2, GI and GO oligosaccharides.

Glycobiology
Pregnancy
-300 -100 0 100Conception Birth
80
60
000-
40
20
300 500Conception
Remission ofJuvenile R.A.
Time (days) Age (years)Fig. 7. Variations in the galactosylation of IgG. There are changes with age and disease activity as illustrated for juvenile arthritis (right).A comparison between the changes in percentage GO during the course of pregnancy for a normal and rheumatoid arthritis patient (left),shows that both patients tend to increase their galactose levels during pregnancy. The dashed line indicates the aged matched expected valueof a normal healthy individual. It is seen that the pregnant patient with rheumatoid arthritis achieves this level and this correlates with a
remission of arthritis in the patient.
influence of the Fc oligosaccharides comes from thefact that removal of the oligosaccharides results in a
reduced stability for the CH2 domain as suggested bya 60-fold increase in its rate of cleavage by pepsin.From Figure 1 we can see that the presence ofoligosaccharides in the Fab hypervariable regioncomprising the antigen binding site could well affectantigen recognition.
IgG GLYCOSYLATION ALTERS IN RHEUMATOID
ARTHRITIS AND ALSO WITH AGE AND WITH
PREGNANCY
Serum IgG from patients with rheumatoid arthritis(RA) contains the same set of biantennary oligo-saccharides found in normal individuals, but in very
different proportions (Fig. 6). The incidence ofstructures with outer-arm galactose (GI or G2) isdramatically decreased, and the incidence of thosestructures terminating in outer-arm N-acetylglucos-amine (GO) correspondingly increased (Parekh et al.1985). Clinically, the GO value can be used both as an
indicator of disease severity and in the early diagnosisof rheumatoid arthritis (Young et al. 1991).
A comparison of the N-glycosylation of Fab and Fcfragments derived from total serum IgG of patientswith rheumatoid arthritis and from a control group
shows that the decreased galactosylation found on
analysis of total serum IgG is largely due to changesin the N-linked oligosaccharides of the Fc. There are
also quantitatively minor but potentially significantdifferences in Fab glycosylation (Chang, 1993). It hasrecently been shown in this laboratory by Dr A.Youings that the changes in Fc glycoforms inrheumatoid arthritis occur in all 4 IgG subclasses.The change in galactosylation of the serum IgG of
patients with rheumatoid arthritis is not common toall other autoimmune or inflammatory disorders.However, agalactosyl IgG has been consistently foundin patients with juvenile rheumatoid arthritis, Crohn'sdisease, and tuberculosis (Parekh et al. 1988 a, 1989).The Fc glycoform distribution also changes with
age (Parekh et al. 1988b) and with pregnancy (Rooket al. 1991) (Fig. 7). These changes presumably reflectthe control of galactosyl transferase activity underdifferent physiological conditions. In an arthriticwoman with pathologically elevated levels of GOglycoforms, pregnancy induced changes in GO corre-
53
46
36
0(5o0o
26
16
9
285
286 R. A. Dwek, A. C. Lellouch and M. R. Wormald
IgG(O) Glycoforms are Involved in CIA
Heat DenaturedType II Collagen
+ FCA
Native Type II
Collagen +FCA(Day 15 vs 35)
ArthritisIgG +IgG(O)++
I
IgGP-galactosidase
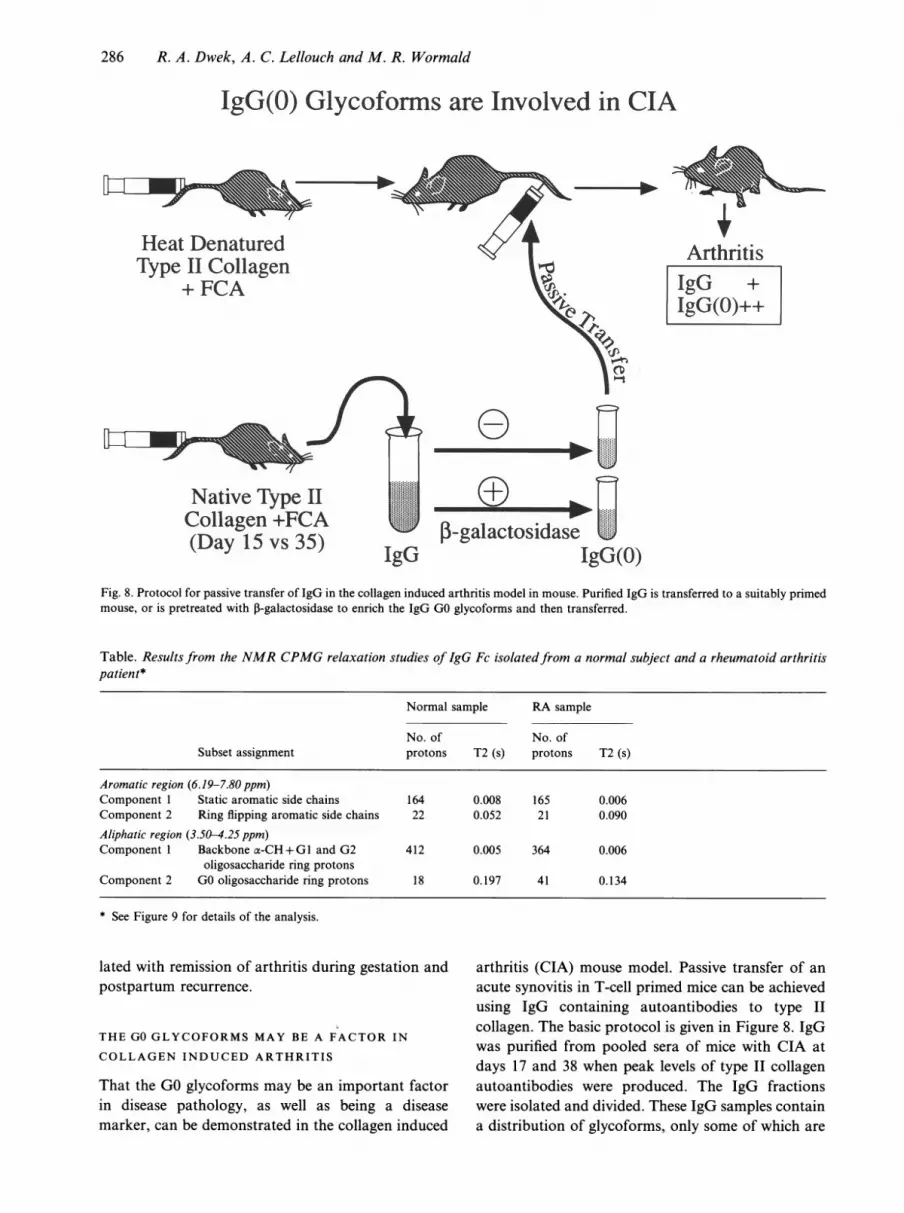
IgG(O)Fig. 8. Protocol for passive transfer of IgG in the collagen induced arthritis model in mouse. Purified IgG is transferred to a suitably primedmouse, or is pretreated with 3-galactosidase to enrich the IgG GO glycoforms and then transferred.
Table. Results from the NMR CPMG relaxation studies of IgG Fc isolatedfrom a normal subject and a rheumatoid arthritispatient*
Normal sample RA sample
No. of No. ofSubset assignment protons T2 (s) protons T2 (s)
Aromatic region (6.19-7.80 ppm)Component 1 Static aromatic side chains 164 0.008 165 0.006Component 2 Ring flipping aromatic side chains 22 0.052 21 0.090Aliphatic region (3.50-4.25 ppm)Component 1 Backbone a-CH + GI and G2 412 0.005 364 0.006
oligosaccharide ring protonsComponent 2 GO oligosaccharide ring protons 18 0.197 41 0.134
* See Figure 9 for details of the analysis.
lated with remission of arthritis during gestation andpostpartum recurrence.
THE GO GLYCOFORMS MAY BE A FACTOR IN
COLLAGEN INDUCED ARTHRITIS
That the GO glycoforms may be an important factorin disease pathology, as well as being a diseasemarker, can be demonstrated in the collagen induced
arthritis (CIA) mouse model. Passive transfer of anacute synovitis in T-cell primed mice can be achievedusing IgG containing autoantibodies to type IIcollagen. The basic protocol is given in Figure 8. IgGwas purified from pooled sera of mice with CIA atdays 17 and 38 when peak levels of type II collagenautoantibodies were produced. The IgG fractionswere isolated and divided. These IgG samples containa distribution of glycoforms, only some of which are

Glycobiology
VerticalScale
x 5b
x 1
7.6 7.2 6.8 6.4 4.2 3.9 3.6
Chemical Shift (ppm)Fig. 9. 1H NMR relaxation weighted spectra of IgG Fc from a rheumatoid arthritis patient. The top insert shows the 1D spectrum recordedon a Varian Unity 500 with a probe temperature of 30 'C. Selected regions of the CPMG (Meiboom & Gill, 1958) spectra at (a) 0.001 s and(b) 0.128 s total relaxation time are shown. By measuring the total intensity in each region as a function of relaxation time and fitting to a
multi-exponential decay, the relaxation rates of the different proton subsets present in each region and the number of protons in each subsetcan be extracted. For each region of the spectrum shown, the data can be fitted well to a bi-exponential decay (2-component system). Theproton subsets can be assigned on the basis of their chemical shift dispersion and their relative relaxation rates. These results are given inthe Table. The backbone a-CH and static aromatic ring protons have very similar T2 times, dependent on the bulk tumbling rate of theprotein. Larger T2 values then indicate more rapid motion (extra internal mobility) of the protons concerned. The values for theoligosaccharide protons compared with typical T2 times of 0.2-0.5 s for the ring protons of released oligosaccharides.
the GO glycoform. One fraction was treated with3-galactosidase from Streptococcus strain 6646K to
generate the IgG (GO) glycoforms exclusively. Byincreasing the level of agalactosyl IgG glycoforms ofthe anti-type II collagen antibodies present in thepreparation, the IgG becomes more effective atcausing arthritis. This indicates that the glycosylationstatus of these autoantibodies is one factor indetermining if they are pathogenic (Rademacher et al.1994).
STRUCTURAL IMPLICATIONS OF CHANGES IN
IgG GLYCOFORMS
NMR studies and x-ray data on the intact Fcfragment from normal and RA samples provide astructural basis for describing the changes resultingfrom the loss of galactose residues. The x-ray data(Deisenhofer, 1981; Sutton & Phillips, 1983) show the
Fc oligosaccharides held rigidly in place relative tothe protein with the ct(I-6) arms interacting withthe protein surface. None of the oligosaccharide resi-dues is accessible to recognition by other proteins. Inthis model, loss of the galactose residues simplyexposes the regions of the protein surface previouslycovered by the galactoses. These vacant sites couldcreate a lectin-like activity by binding galactoseresidues from other IgG molecules, resulting in theformation of complexes or autoaggregates typical ofRA (Rademacher et al. 1988) without an actualautoimmune response. Alternatively, the sites couldreveal antigenic determinants, creating a direct auto-immune response. The contribution of such factors tothe pathology of the disease has still to be assessed.NMR relaxation studies (Fig. 9) from normal (26%
GO) and RA (55% GO) samples show that there is avery slowly relaxing subset of proton resonances (seenas residual sharp peaks in the relaxation weighted
'&L-A,- -AL,- --A L. -.k.. !Poo%1-1 -1-T I O"WT
-.1 ....I....I ....I.... I----I.... I.Frn
a
.... I---- I....I.... I.... I.... I----I---- ....I ....I---- I.... .... I....I
287

288 R. A. Dwek, A. C. Lellouch and M. R. Wormald
residues in each GO oligosaccharide must be under-going rapid motion relative to the protein.These observed changes in relative mobility lead to
a proposed model of GI and G2 oligosaccharidesbeing bound to, or having restricted motion relativeto, the peptide surface (as seen in the crystal structure)whilst the GO oligosaccharides are not bound to theprotein surface and have extra mobility relative to thepeptide (Fig. 10). This model has various structuralimplications. (1) Removal of galactoses not onlyexposes the protein surface previously covered by thegalactose residues, but also other regions previouslycovered by other oligosaccharide residues (Fig. 11).(2) The extra mobility of the GO oligosaccharides,together with inspection of the crystal structure,suggests that the oligosaccharides could now bedisplaced far enough to make the terminal oligo-saccharide residues accessible for direct recognitionby other proteins. (3) The ability to displace the Fcoligosaccharides would allow for greater changes inthe relative positions (quaternary organisation) ofthe 2 CH2 domains (given the constraints discussedearlier in the section on the domain structure ofIgG) without unfavourable oligosaccharide-oligo-saccharide steric interactions occurring. This in turnwould affect the structure of the hinge region.
Fig. 10. Schematic representation of the oligosaccharides of IgG Fclooking along the pseudo-C2 axis. The squares show the positionsof the GlcNAc 1 residues, the circles the positions of the othermonosaccharide residues, G indicates a nonreducing terminalgalactose residue on the 6-arm of the oligosaccharide. (a) Two GIoligosaccharides (as seen in the crystal structure). Binding of the 6-arm to the protein surface holds the rest of the oligosaccharide rigidwith respect to the protein. (b) A GO (left) and GI (right)oligosaccharide. Loss ofinteraction between the GO oligosaccharide6-arm and the protein surface leaves the whole of the oligo-saccharide free to move.
spectra) between 3.50 and 4.25 ppm which can beassigned to the oligosaccharides (Malhotra et al.1995). Slow NMR relaxation corresponds to more
rapid motion of the protons in this subset. There are
approximately 18 and 41 protons in this subset in thenormal and rheumatoid samples respectively. Theseare not enough to account for all the oligosaccharideprotons in either sample, the remaining havingrelaxation properties very similar to the peptidebackbone. The 2-fold increase in number of protonsin the slowly relaxing set mirrors the 2-fold increase inGO value between the 2 samples allowing assignmentof these protons to GO oligosaccharides only. In orderto account for the number of rapidly moving protonsin this subset, at least 6 of the 8 monosaccharide
OLIGOSACCHARIDE RECOGNITION
That oligosaccharides act as recognition elementsis now widely accepted. There is evidence fortheir involvement in many different processes includ-ing the specific recognition of one cell by another,cell-cell adhesion and the clearance of serum glyco-proteins. Recognition of oligosaccharides impliesthat there must be other molecules (lectins) that can
selectively interact with them. Many animal lectinshave now been isolated from different tissues and celltypes and have been classified mainly on the basis ofsequence homology. The largest group is the C-typelectins (Drickamer, 1988) which bind carbohydratesin a Ca2"-dependent manner. These lectins con-
tain common sequence motifs, called carbohydrate-recognition domains (CRDs), which consist ofapproximately 120 amino acid residues and are
characterised by 31 invariant or highly conservedresidues (Weis et al. 1991). C-type CRDs have now
been reported in a large number of proteins inassociation with a variety of effector domains.One important C-type lectin is the mannose-binding
protein (MBP) which binds to pathogens containing a
high concentration of mannose or N-acetylglucos-
(a)
(b)

Glycobiology
I I
J~~~~~~~~~~~~~~~~~~~~~1
Fig. 11. The change in solvent accessible protein surface of the CH2 domain when the terminal galactose residues are removed and theoligosaccharide allowed to move from its crystallographic position. Left, crystallographic structure (Deisenhofer, 1981). Right, modelledstructure based on the NMR results with a GO glycoform. Molecular modelling was performed on a Silicon Graphics Personal Irisworkstation using the programs Insight II and Discover from Biosym Technologies Ltd.
amine on their surface. This recognition event canlead to antibody-independent complement fixation oropsonisation (Ikeda et al. 1987). Recent x-ray data onthe CRD from this protein (Weis et al. 1992) showthat the major interaction between the CRD and thesugar involves binding of the Ca2" ion to saccharidecis-hydroxyl groups (Fig. 12). Site-specific muta-genesis studies (Iobst & Drickamer, 1994) have shownthat additional nonbonded interactions between thesugar and the protein amino acid side chains areresponsible for the observed monosaccharide bindingspecificity.The CRD of the MBP is connected to a collagen-
like domain. The individual polypeptides of theprotein associate into groups of 3, forming a collagentriple helix attached to a cluster of 3 CRDs in aglobular head. These trimeric building blocks as-sociate, via the collagenous region, to form largerstructures with up to 6 globular heads (i.e. 18 CRDs).Each CRD binds a terminal monosaccharide residueweakly. Physiologically significant binding resultsfrom multiple CRDs interacting with a multivalenttarget. MBP resembles Clq in structure (Fig. 13). The
mechanism by which MBP activates complement isless well documented than is the case with Clq, butboth require a multivalent target.
RECOGNITION OF GO GLYCOFORMS BY THE
MANNOSE BINDING PROTEIN
Removal of the terminal galactose residues in the GOglycoforms ofIgG results in terminal GlcNAc residueswhich have the potential to interact with the CRDfrom MBP. Molecular modelling shows that thisinteraction cannot occur without displacement ofthe oligosaccharide from the position observed in theFc x-ray structure. The NMR data show thatsuch displacements occur spontaneously on loss ofgalactose. Molecular modelling indicates that thesedisplacements could make either the 3-arm or 6-armterminal GlcNAc residues available for binding to theCRD. One possible model is shown in Figure 14.
Recent observations (Malhotra et al. 1995) indicatethat MBP does bind to agalactosyl IgG in vitrovia the Fc terminal GlcNAc residues and is thencapable of activating complement. These experiments
289

290 R. A. Dwek, A. C. Lellouch and M. R. Wormald
Asn 187
Glu 185
His 189-F W
Man
lie 207
.I
Glu 193Asn 205
Fig. 12. Structure of the C-type CRD from mannose binding protein (Weis et al. 1992). The boxed area is enlarged to illustrate the detailsof the interaction with mannose.
CIq
Nr
IgG MBP
~Y
1II1 Onm
Fig. 13. Schematic representations of Clq, MBP and IgG based on the references (Dwek et al. 1984; Drickamer et al. 1986; Malhotra et al.1990). Clq has 6 globular regions each consisting of 3 similar, but nonidentical independently folding domains. MBP occurs not only as thehexameric structure drawn here, but also as trimer, tetramers and pentamers (Lu et al. 1990) of the triple helical subunit and has head regionscomposed of identical protein domains each containing a CRD.
I

Glycobiology 291
'; *) " \ s
.,~~~~~9 24, ..* t>\ > w1 ..~~Fig. 14. Molecular model of a complex formed between the Fc (Deisenhofer, 1981) and the mannose binding protein (Weis et al. 1992) basedon the mannose binding protein interacting with the 6-arm terminal N-acetylglucosamine residue of the IgG GO glycoform. Molecularmodelling was performed on a Silicon Graphics Personal Iris workstation using the programs Insight II and Discover from BiosymTechnologies Ltd.
involve binding IgG or its fragments to microtitreplates. IgG fixed on these plates is aggregated so thatevidence produced is that aggregates of agalactosylIgG bind MBP and activate complement. In RApatients, autoantibodies carrying GO structures maybe localised in the joints bound to the collagenmatrix and therefore essentially aggregated on animmobilised matrix presenting a multivalent surfaceto MBP. The levels ofGO are elevated in the synoviumcompared with serum (Rademacher et al. 1988;Tsuchiya et al. 1993). Within the synovium, it is apossibility that activation of the complement pathwayby MBP could contribute to the chronic inflammationof the synovial membrane of affected joints.The capacity of IgG carrying GO structures to
activate complement via MBP may be of considerablesignificance in vivo. This additional mechanismleading to complement activation is only available tothose IgG molecules with appropriate glycosylation.It provides a possible link between the elevated levelsofGO structures on IgG found in rheumatoid arthritisand the onset of tissue damage, and therefore a
possible role for GO structures in the pathogenesis ofthe disease.
ACKNOWLEDGEMENTS
We wish to acknowledge the contributions from ourcolleagues, both past and present, in the GlycobiologyInstitute to the work described in this paper. Inparticular, we acknowledge the work of ProfessorT. W. Rademacher and Dr P. M. Rudd. We thankthe Biochemical Society for permission to reproduceFigures 20, 22 and 24-28 from the 7th Wellcome TrustAward for Research in Biochemistry Related toMedicine, 1994, given by R. A. Dwek and publishedin the Biochemical Society Transactions (in press).
REFERENCES
BURTON DR, BOYD J, BRAMPTON AD, EASTERBROOK-SMITH SB,EMANUEL EJ, NOVoTNY J et al. (1980) The Clq receptor site onimmunoglobulin G. Nature 288, 338-344.
CHANG S-C (1993) Altered glycosylation in immunoglobulin G and itsfragments in rheumatoid arthritis. D.Phil. thesis, University ofOxford.
DEISENHOFER J (1981) Crystallographic refinement and atomicmodels of human Fc fragment and its complex with Fragment

292 R. A. Dwek, A. C. Lellouch and M. R. Wormald
B of Protein A from Staphylococcus aureus at 2.9 and 2.8 Aresolution. Biochemistry 20, 2361-2370.
DRICKAMER K (1988) Two distinct classes of carbohydrate-recognition domains in animal lectins. Journal of BiologicalChemistry 263, 9557-9560.
DRICKAMER K, DORDAL MS, REYNOLDS L (1986) Mannose bindingprotein isolated from rat liver contains carbohydrate-recognitiondomains linked to collagenous tails. Complete primary structureand homology with pulmonary surfactant apoprotein. Journal ofBiological Chemistry 261, 6878-6887.
DUNCAN AR, WINTER G (1988) The binding site for Clq on IgG.Nature 332, 738-740.
DWEK RA, SUTTON BJ, PERKINS SJ, RADEMACHER TW (1984)Structure-function relationships in immunoglobulins. Bio-chemistry Society Symposium 49, 123-136.
EMANUEL EJ, BRAMPTON AD, GAGNON J, DWEK RA (1982)Chemical verification of the Clq receptor site on IgG. FEBSLetters 137, 298-302.
IKEDA K, SANNOH T, KAWASAKI N, KAWASAKI T, YAMASHINA I(1987) Serum lectin with known structure activates complementthrough the classical pathway. Journal of Biological Chemistry262,451-454.
IOBST ST, DRICKAMER K (1994) Binding of sugar ligands to Ca2+-dependent animal lectins. II. Generation of high-affinity galactosebinding by site-directed mutagenesis. Journal of BiologicalChemistry 269, 15512-15519.
KABAT EA, WU TT, BILOFSKY H, REID-MILLER M, PERRY H (1983)Sequences of proteins of immunological interest. Publications ofUS Department of Health and Human Services.
LEATHERBARROW RJ (1983) Structural studies on DNP bindingantibodies. D.Phil. thesis, University of Oxford.
LEATHERBARROW RJ, RADEMACHER TW, DWEK RA, WOOF JM,CLARK A, BURTON DR et al. (1985) Effector functions ofmonoclonal aglycosylated mouse IgG2a; binding and activationof complement component Cl and interaction with human Fcreceptor. Molecular Immunology 22, 407-415.
LU J, THIEL S, WIEDEMANN H, TIMPL R, REID KBM (1990) Bindingof the pentamer/hexamer forms of mannan-binding protein tozymosan activates the proenzyme Clr2 Cls2 complex, of theclassical pathway of complement, without involvement of Clq.Journal of Immunology 144, 2287-2294.
MALHOTRA R, THIEL S, REID KBM, SIM RB (1990) Humanleukocyte Clq receptor binds other soluble proteins with collagendomains. Journal of Experimental Medicine 172, 955-959.
MALHOTRA R, WORMALD MR, RUDD PM, FISCHER PB, DWEK RA,SIM RB (1995) Glycosylation changes of IgG associated withrheumatoid arthritis can activate complement via the mannose-
binding protein. Nature Medicine 3, 237-243.MEIBOOM S, GILL D (1958) Modified spin-echo method for
measuring nuclear relaxation times. Review of Scientific Instru-mentation 29, 688-691.
PAREKH RB, DWEK RA, SUTTON BJ, FERNANDES DL, LEUNG A,STANWORTH D et al. (1985) Association of rheumatoid arthritis
and primary osteoarthritis with changes in the glycosylationpattern of total serum IgG. Nature 316, 452-457.
PAREKH RB, ISENBERG DA, ANSELL BM, ROITT IM, DWEK RA,RADEMACHER TW (1988 a) Galactosylation of IgG associatedoligosaccharides: reduction in patients with adult and juvenileonset rheumatoid arthritis and relation to disease activity. Lancet1(8592), 966-969.
PAREKH RB, ISENBERG DA, RoITT IM, DWEK RA, RADEMACHERTW (1988b) Age-related galactosylation of the N-linked oligo-saccharides of human serum IgG. Journal of ExperimentalMedicine 167, 1713-1736.
PAREKH RB, ISENBERG DA, ROOK G, RoITT IM, DWEK RA,RADEMACHER TW (1989) A comparative analysis of disease-associated changes in the galactosylation of serum IgG. Journalof Autoimmunity 2, 101-1 14.
RADEMACHER TW, HOMANS SW, PAREKH RB, DWEK RA (1986)Immunoglobulin G as a glycoprotein. Biochemistry SocietySymposium 51, 131-148.
RADEMACHER TW, PAREKH RB, DWEK RA, ISENBERG D, ROOK G,AXFORD J et al. (1988) The role of IgG glycoforms in thepathogenesis of rheumatoid arthritis. Springer Seminars onImmunopathology 10, 231-249.
RADEMACHER TW, WILLIAMS P, DWEK RA (1994) Agalactosylglycoforms of IgG autoantibodies are pathogenic. Proceedings ofthe National Academy of Sciences of the USA 91, 6123--6127.
ROOK GAW, STEELE J, BREALEY R, WHYTE A, ISENBERG D, SUMARN et al. (1991) Changes in IgG glycoform levels are associatedwith remission of arthritis during pregnancy. Journal of Auto-immunity 4, 779-794.
SUTTON BJ, PHILLIPS DC (1983) The three-dimensional structure ofthe carbohydrate within the Fc fragment of immunoglobulin G.Biochemistry Society Transactions 11, 130-132.
TAO M-H, MORRISON SL (1989) Studies of aglycosylated chimericmouse-human IgG. Journal of Immunology 143, 2595-2601.
TSUCHIYA N, ENDO T, MATSUTA K, YOSHINOYA S, TAKENCHI F,NAGANO Y et al. (1993) Detection of glycosylation abnormalityin rheumatoid IgG using N-acetylglucosamine-specificPsathyrella velutina lectin. Journal of Immunology 151,1137-1146.
WEIS W, KAHN R, FOuRME R, DRICKAMER K, HENDRICKSON WA(1991) Structure of the calcium-dependent lectin domain from a
rat mannose-binding protein determined by MAD phasing.Science 254, 1608-1615.
WEIS WI, DRICKAMER K, HENDRICKSON WA (1992) Structure ofa C-type mannose-binding protein complexed with an oligo-saccharide. Nature 360, 127-134.
WORMALD MR, WOOTEN EW, BAZZO R, EDGE CJ, FEINSTEIN A,RADEMACHER TW et al. (1991) The conformational effects of N-glycosylation on the tailpiece from serum IgM. European Journalof Biochemistry 198, 131-139.
YOUNG A, SUMAR N, BODMAN K, GOYAL S, SINCLAIR H, ROITT I etal. (1991) Agalactosyl IgG: an aid to differential diagnosis inearly synovitis. Arthritis and Rheumatism 34, 1425-1429.





























