Embed Size (px)
Citation preview

Dental microwear in grazing and browsing Gotland sheep(Ovis aries) and its implications for dietary reconstruction
Ingrid L. Mainland*
Department of Archaeological Sciences, University of Bradford, Bradford BD7 1DP, UK
Received 10 July 2002; received in revised form 8 April 2003; accepted 10 April 2003
Abstract
Microwear patterns and formation processes are examined in modern browsing and grazing Gotland sheep from Denmark. Cleardifferences in microwear patterning are identified between Gotland sheep, which included a high proportion of leaves, bark andtwigs of deciduous trees and shrubs in their diet, and those which primarily had access to graze species, with the browsing groupexhibiting enamel surfaces dominated by pitted microwear features, the grazers a predominance of striations. Analysis of abrasivesin dung suggests that these trends can be attributed to higher levels of soil ingestion in the grazing population and are a consequenceboth of grassland management practices and height of feeding within the vegetation canopy. These findings question previousinterpretations of microwear patterning in browsing and grazing ungulates, which have equated striated enamel features with theingestion of high levels of opal phytoliths, and thus have significant implications for palaeodietary reconstruction in such species,both within archaeological and palaeontological contexts. To address further the implications of these results, comparison is madebetween the Gotland sheep and modern grazing and fodder-fed sheep and goats from elsewhere in Europe. It is concluded thatdental microwear analysis has the potential to address questions relating to grassland management practices and the use of certainfodders, such as leafy-hay, but that it may ultimately prove more difficult to distinguish ‘fresh’ browsing sheep from some grazingpopulations, specifically those in which levels of soil ingestion are reduced.� 2003 Elsevier Ltd. All rights reserved.
Keywords: Dental microwear; Diet; Sheep; Browsing; Grazing; Archaeology
1. Introduction
Dental microwear analysis is now widely used inpalaeodietary reconstruction [27,37]. Although largelyfocused on the diet of hominids and primates, micro-wear analysis has also been applied to other mammalianspecies within both palaeontological [1,31,33] and, morerecently, archaeological contexts [4,16,21]. Interpret-ation of microwear patterning in ancient specimens isbased on analogy with modern individuals, and, as such,one of the primary concerns of microwear research is toestablish the variation evident in modern species withdiets relevant to particular archaeological or palaeonto-logical questions [18,37]. This article presents furtherresearch on microwear patterning in modern sheep and
goats undertaken to establish the potential range ofdiets, which can be identified for these species within anarchaeological context [18,20–22], and focuses, in par-ticular, on microwear patterns and formation processesin browsing and grazing Gotland sheep from Denmark.
Browsing and grazing were amongst the first dietaryadaptations to be explored using dental microwearanalysis. In the 1970s, Walker et al. [43] demonstratedthat a distinction was evident between microwear pat-terns in browsing and grazing hyrax species, the formerbeing characterised by polished enamel surfaces in whichthe underlying enamel structure was visible and by fewmicrowear defects, the latter by enamel surfaces domi-nated by striations. Subsequent research on a widerrange of browsing and grazing ungulate species bySolounias and co-workers [31–33] confirmed the associ-ation between heavily striated enamel surfaces and graz-ing, but indicated that browsers may be associated with
* Tel.: +44-1274-233541; fax: +44-1274-235190.E-mail address: [email protected] (I.L. Mainland).
Journal of Archaeological Science 30 (2003) 1513–1527
SCIENCE
Journal of
Archaeological
http://www.elsevier.com/locate/jas
SCIENCE
Journal of
Archaeological
http://www.elsevier.com/locate/jas
0305-4403/03/$ - see front matter � 2003 Elsevier Ltd. All rights reserved.doi:10.1016/S0305-4403(03)00055-4

both prism relief and a dominance of pitted micro-wear features. These trends were attributed to dietaryabrasiveness, specifically differences in the phytolithcontent of browse and graze vegetation, with the preva-lence of striations in grazers being equated with theconsumption of phytolith-rich grasses [33,43].
The possibility that microwear could be used todistinguish animals, which had been browsing andgrazing, is of considerable interest for archaeology;utilisation of woodland pasture and the collection ofbrowse vegetation as fodder for domestic stock arewidely assumed to have been critical resources forearly agricultural communities within Europe [10,13],but have been difficult to identify unequivocally owingto the poor preservation of botanical material withinarchaeological contexts [19,22]. Previous research onmicrowear in sheep and goats had identified statisti-cally significant differences between modern grazingovicaprids and those stall-fed on leafy-hay (i.e. a dietof dried browse vegetation), thus indicating the poten-tial existence of a browse–graze dichotomy in themicrowear of these species [2,18,19]. These analysesalso demonstrated that microwear patterning withingrazing sheep is rather variable, and is not typical ofthe grazing ungulates studied by Solounias et al. [33]:only some populations of grazing sheep exhibit heavilystriated enamel surfaces; most are characterised by anemphasis on pits [18,20–22]. This earlier research was,however, limited in that analysis focused only onleafy-hay and did not consider microwear associatedwith the consumption of fresh browse material. More-over, little attempt was made to identify microwearformation processes within the populations studied. Itwas thus unclear why striations are uncommon in graz-ing sheep, whether leafy-hay would be distinguishablefrom a diet of fresh browse or if microwear in freshbrowsing individuals would differ significantly fromgrazers exhibiting pitted microwear patterns [18]. Toaddress these issues, a series of analyses were undertakenon a population of Gotland sheep kept at the HistoricalArchaeological Research Centre at Lejre in Denmark.This research aimed firstly to identify whether differ-ences in microwear were evident between sheep re-stricted to an area of grassland and those kept in awoodland pasture in which the leaves and bark ofdeciduous trees and bushes comprised a high proportionof the diet selected. A secondary aim was to assesswhether any variation in microwear identified could beattributed to the amount and type of abrasives ingestedby the sheep. Finally, to determine whether the con-sumption of fresh browse vegetation could be dis-tinguished both from dried browse and from grazingsheep in which pitted microwear patterns predominated,microwear in the Lejre sheep was compared with thatin leafy-hay-fed sheep and goats from the village ofPlikati in Greece [18,22] and in a large sample of grazing
sheep pastured on diverse northern European grasslandhabitats [18,20,21].
Ruminant species can be classified into three broadcategories on the basis of their digestive system anddietary adaptation [14]: grass and roughage feeders, orgrazers, like the buffalo and cow are adapted to fibrousdiets rich in cell wall contents (lignin, cellulose etc.) andcan eat both young and mature plant species; concentratefeeders, or browsers, e.g. the moose or giraffe, cannottolerate fibre and select easily digestible vegetation,such as fruit and foliage with a high cell content to cellwall ratio; intermediate feeders like the red deer andThomson’s gazelle are adapted to both browsing andgrazing, but prefer less fibrous species and immaturevegetation. Sheep have been classified both as grass pre-ferring intermediate feeders [25,30, pp. 4–5] and grass/roughage feeders [14], and they preferentially eatvegetation with a high nutritional value [2], which mayinclude leaves and fruits from arboreal species [25]. In thefollowing discussion, browsing is used in relation to thesheep and goat populations studied to indicate a diet thatincludes a relatively high proportion of leaves, bark, fruitand twigs from deciduous trees and shrubs and is not acomment on the overall dietary adaptation of sheep.
2. Materials and methods
2.1. The modern sheep and goat populations studied
Gotland sheep, also called the Goth sheep, are asmall, primitive breed classed within the northern short-tailed group and derive originally from the Swedishisland of Gotland in the Baltic Sea [28]. This breed isrenowned for its habit of browsing vegetation and barkfrom deciduous and coniferous trees [28]. Since the1960s, the Archaeological and Research Centre at Lejrehas kept a flock of Gotland sheep, which are pasturedboth on open, semi-indigenous grassland and in an areaof deciduous woodland [11]. The latter was created withthe specific aim of reconstructing prehistoric systems ofwoodland pasture (Rasmussen, personal communi-cation), and consists of a network of open grasslandareas interspersed with wooded stands. A wide range ofshrubs, trees and unimproved grasses are representedincluding wild rose, hawthorn, stinging nettle, elder,beech, ash, willow, elm, birch, lime, hazel and couchgrass. The Gotland sheep select a mixture of browse andgraze species within the woodland pasture, but increas-ingly consume deciduous browse during late autumnand early winter, as the grasses available becomesenescent and less common (Jacobsen, personal com-munication). Ring barking is also observed and againoccurs more frequently in autumn and winter. The opengrassland pastures comprise a mixture of unimprovedand improved grasses, as well as species such as nettlesand thistles.
I.L. Mainland / Journal of Archaeological Science 30 (2003) 1513–15271514

Twenty-two mandibles from the Lejre flock wereavailable for study. These were derived from male andfemale lambs, which had been born in March/April 1996(n�10) and 1997 (n�12) and which were slaughtered atca. 9 months old in the November following their birth(Table 1). Half the lambs had been pastured exclusivelyon the woodland pasture since September, and theremainder had grazed the Lejre common, an area ofopen, unimproved grassland adjacent to the recon-structed Iron-age village. Ad hoc observation by theagricultural managers at Lejre of feeding behaviour inthe woodland pasture suggests that by late October/November, the diet of the lambs consisted of approxi-mately 40% bark, twigs and deciduous browse and 60%graze species (Jacobsen, personal communication).Deciduous browse was not available for the sheeppastured in the common, and the diet of these indi-viduals comprised mainly of grass species (Jacobsen,personal communication).
Microwear in the Lejre sheep was compared with anexisting data base of microwear statistics for grazing andfodder-fed sheep/goats from Europe and the NorthAtlantic islands, specifically sheep grazing indigenous,upland habitats in Greenland (n�56) [21], Orkney(n�11) [20] and the Scottish borders (n�8) [20], andsheep and goats fed a mixture of leafy-hay and grass-hayfrom Greece (n�13) [22].
2.2. Microwear analysis
Araldite replicas (mould=Coltene President’s Jet,Regular Body; cast=Araldite MY753, hardener HY956)were made of the right lower deciduous fourth premolar(dP4) and were examined for microwear at a magnifica-tion of 500�, using a scanning electron microscope(SEM) (Philips XL30). For each individual, an imagewas captured from a specific area of enamel, approxi-mately halfway along the anterior-facing enamel band ofthe bucco-posterior cusp, following procedure outlinedin detail elsewhere [20,21]. Three micrographs wererejected owing to being damaged and/or having brokenenamel surfaces, leaving a total sample of 19: grazing(n�9), browsing (n�10).
The microwear defects visible within each image werecounted and measured for defect orientation (as indi-cated by the angle between the long axis of a feature anda line parallel to the base of the image), maximum lengthand breadth using Microwear 2.2 [40]. As in previousanalyses of ovicaprid microwear [18,20,21,22], defectswere then categorised into feature types (pits, striations,etc.) using the ratio of feature length to breadth (Table2). In two individuals (5w and 3w, Table 1) in which theenamel band did not cover the entire SEM screen at500� (i.e. 236 µm wide), feature frequency was adjustedusing the following formula: number of defects=maximum potential area that could be examinedat 500�/actual area examined�number of defectsrecorded in area examined.
The relationship between diet and dental microwearwas investigated statistically using the Student’s t-test,Mann–Whitney and stepwise discriminant analysis (DA)(Tables 2–7). Inclusion/exclusion of variables in thestepwise DA was based on minimisation of Wilks’lambda [24]. One sample (number 52) did not have avalue for narrow striation length and breadth due to theabsence of this particular feature type. To include thiscase in DA, which will omit all cases with missing values,each missing variable was assigned the average value(per diet group) of the relevant length or breadthvariable [35]. All statistical analyses were undertakenusing SPSS [24].
2.3. Dung analysis
In November 1996, 30 samples of sheep dung con-taining approximately 10–20 individual dung pelletswere collected from each group at Lejre during the weekpreceding slaughter, and a record was maintained of thelambs from which the dung was derived. In all cases, thedung was fresh and did not appear to be contaminatedby wind blown soil or dust.
Analysis of the dung samples aimed firstly to assesswhether variation in the overall level of abrasives in thediet of the Lejre sheep could explain any microwear
Table 1Tooth eruption and wear for the Gotland sheep
Reference number Diet Date of death dP4 M1 M2 M3
46 Grazing November 1996 14l 7a V Ø6g Grazing November 1997 14l 7a 1/2 Ø44 Grazing November 1996 14l 8a E Ø56 Grazing November 1996 14l 8a V Ø2g Grazing November 1997 14l 8a E Ø42 Grazing November 1996 16l 7a V/E Ø4g Grazing November 1997 16l 7a E Ø53 Grazing November 1996 16l 8a V Ø1g Grazing November 1997 16l 8a E Ø5g Grazing November 1997 16l 9a E Ø3g Grazing November 1997 17l 8a E Ø54 Browsing November 1996 14l 7a V/E Ø52 Browsing November 1996 14l 7a V Ø40 Browsing November 1996 14l 7a C/V Ø47 Browsing November 1996 14l 7a V Ø1w Browsing November 1997 14l 8a 1/2 Ø3w Browsing November 1997 14l 8a E Ø4w Browsing November 1997 14l 8a E Ø6w Browsing November 1997 14l 8a E Ø45 Browsing November 1996 16l 8a E Ø2w Browsing November 1997 16l 8a 1/2 Ø5w Browsing November 1997 16l 9 1/2 Ø
Abbreviations: teeth—dP4, deciduous fourth mandibularpremolar; M1–3, first to third mandibular molars; tootheruption—Ø, unerupted; C, crypt visible in bone; V, tooth visiblebelow head of bone; E, tooth emerging through bone; 1/2, tooth halferupted; tooth wear codes are as described by Payne [45].
I.L. Mainland / Journal of Archaeological Science 30 (2003) 1513–1527 1515
trends identified. This was achieved by calculating theinorganic content of dung samples, a technique used forassessing dietary abrasiveness and levels of soil ingestionin agricultural science and wildlife biology [5,12,23]. Theinorganic content of each dung sample was establishedthrough loss-on-ignition (LOI) [3]. Three dung pellets
were selected from each sample, placed in ceramiccrucibles, dried for 12 h at 105 (C and weighed (=dryweight; Table 8). The samples were then ashed at 500 (Cfor a further 12 h and reweighed. The inorganic con-tent was calculated as follows: ashed weight/driedweight�100 (=% inorganic; Table 8).
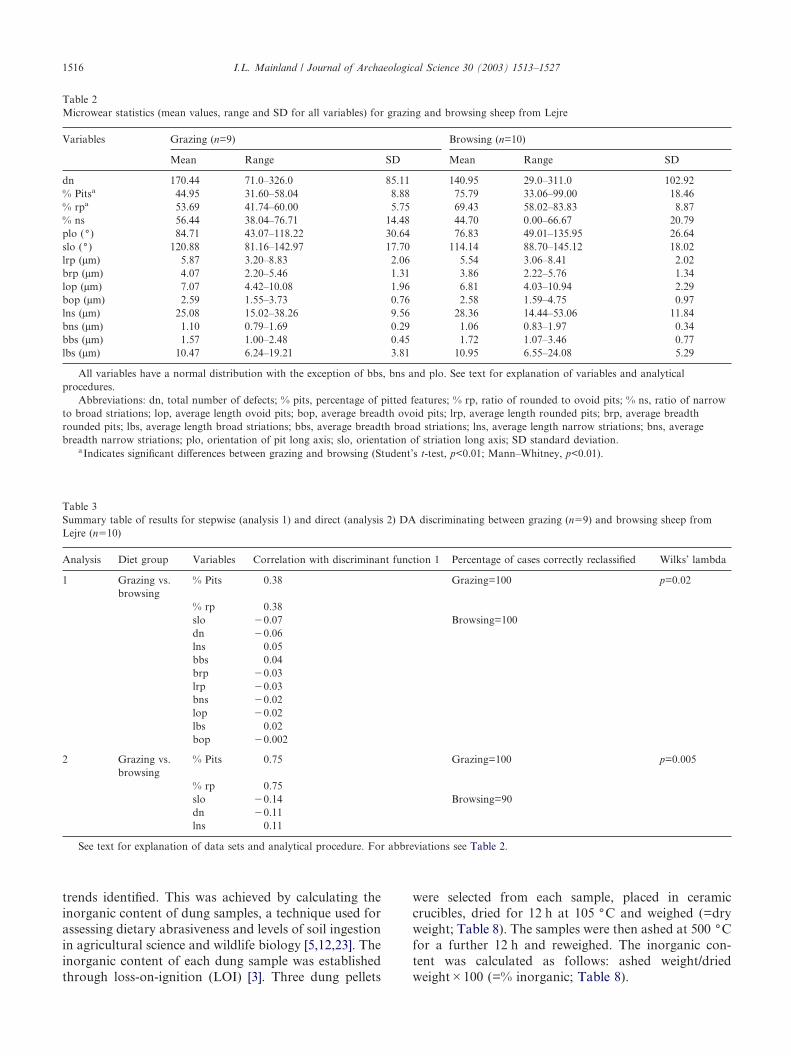
Table 2Microwear statistics (mean values, range and SD for all variables) for grazing and browsing sheep from Lejre
Variables Grazing (n=9) Browsing (n=10)
Mean Range SD Mean Range SD
dn 170.44 71.0–326.0 85.11 140.95 29.0–311.0 102.92% Pitsa 44.95 31.60–58.04 8.88 75.79 33.06–99.00 18.46% rpa 53.69 41.74–60.00 5.75 69.43 58.02–83.83 8.87% ns 56.44 38.04–76.71 14.48 44.70 0.00–66.67 20.79plo (() 84.71 43.07–118.22 30.64 76.83 49.01–135.95 26.64slo (() 120.88 81.16–142.97 17.70 114.14 88.70–145.12 18.02lrp (µm) 5.87 3.20–8.83 2.06 5.54 3.06–8.41 2.02brp (µm) 4.07 2.20–5.46 1.31 3.86 2.22–5.76 1.34lop (µm) 7.07 4.42–10.08 1.96 6.81 4.03–10.94 2.29bop (µm) 2.59 1.55–3.73 0.76 2.58 1.59–4.75 0.97lns (µm) 25.08 15.02–38.26 9.56 28.36 14.44–53.06 11.84bns (µm) 1.10 0.79–1.69 0.29 1.06 0.83–1.97 0.34bbs (µm) 1.57 1.00–2.48 0.45 1.72 1.07–3.46 0.77lbs (µm) 10.47 6.24–19.21 3.81 10.95 6.55–24.08 5.29
All variables have a normal distribution with the exception of bbs, bns and plo. See text for explanation of variables and analyticalprocedures.
Abbreviations: dn, total number of defects; % pits, percentage of pitted features; % rp, ratio of rounded to ovoid pits; % ns, ratio of narrowto broad striations; lop, average length ovoid pits; bop, average breadth ovoid pits; lrp, average length rounded pits; brp, average breadthrounded pits; lbs, average length broad striations; bbs, average breadth broad striations; lns, average length narrow striations; bns, averagebreadth narrow striations; plo, orientation of pit long axis; slo, orientation of striation long axis; SD standard deviation.
aIndicates significant differences between grazing and browsing (Student’s t-test, p<0.01; Mann–Whitney, p<0.01).
Table 3Summary table of results for stepwise (analysis 1) and direct (analysis 2) DA discriminating between grazing (n�9) and browsing sheep fromLejre (n�10)
Analysis Diet group Variables Correlation with discriminant function 1 Percentage of cases correctly reclassified Wilks’ lambda
1 Grazing vs.browsing
% Pits 0.38 Grazing=100 p=0.02
% rp 0.38slo �0.07 Browsing=100dn �0.06lns 0.05bbs 0.04brp �0.03lrp �0.03bns �0.02lop �0.02lbs 0.02bop �0.002
2 Grazing vs.browsing
% Pits 0.75 Grazing=100 p=0.005
% rp 0.75slo �0.14 Browsing=90dn �0.11lns 0.11
See text for explanation of data sets and analytical procedure. For abbreviations see Table 2.
I.L. Mainland / Journal of Archaeological Science 30 (2003) 1513–15271516
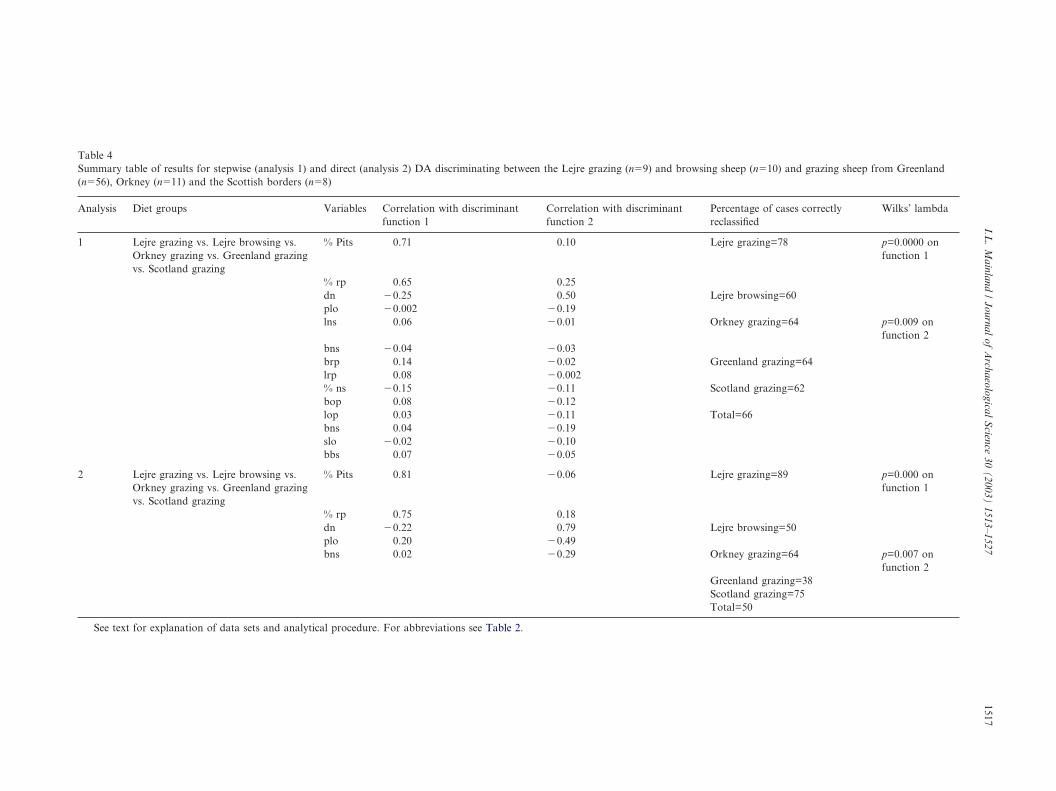
Table 4Summary table of results for stepwise (analysis 1) and direct (analysis 2) DA discriminating between the Lejre grazing (n�9) and browsing sheep (n�10) and grazing sheep from Greenland(n�56), Orkney (n�11) and the Scottish borders (n�8)
Analysis Diet groups Variables Correlation with discriminantfunction 1
Correlation with discriminantfunction 2
Percentage of cases correctlyreclassified
Wilks’ lambda
1 Lejre grazing vs. Lejre browsing vs.Orkney grazing vs. Greenland grazingvs. Scotland grazing
% Pits 0.71 0.10 Lejre grazing=78 p=0.0000 onfunction 1
% rp 0.65 0.25dn �0.25 0.50 Lejre browsing=60plo �0.002 �0.19lns 0.06 �0.01 Orkney grazing=64 p=0.009 on
function 2bns �0.04 �0.03brp 0.14 �0.02 Greenland grazing=64lrp 0.08 �0.002% ns �0.15 �0.11 Scotland grazing=62bop 0.08 �0.12lop 0.03 �0.11 Total=66bns 0.04 �0.19slo �0.02 �0.10bbs 0.07 �0.05
2 Lejre grazing vs. Lejre browsing vs.Orkney grazing vs. Greenland grazingvs. Scotland grazing
% Pits 0.81 �0.06 Lejre grazing=89 p=0.000 onfunction 1
% rp 0.75 0.18dn �0.22 0.79 Lejre browsing=50plo 0.20 �0.49bns 0.02 �0.29 Orkney grazing=64 p=0.007 on
function 2Greenland grazing=38Scotland grazing=75Total=50
See text for explanation of data sets and analytical procedure. For abbreviations see Table 2.
I.L.
Mainland
/Journal
ofA
rchaeologicalS
cience30
(2003)1513–1527
1517
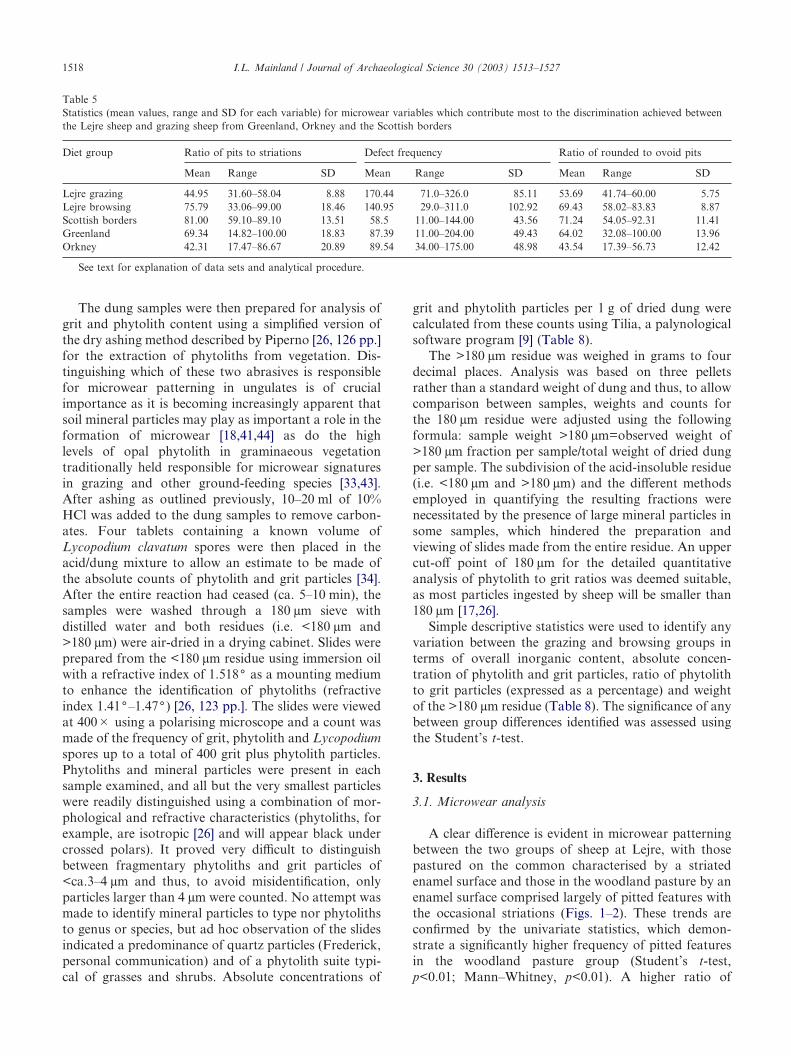
The dung samples were then prepared for analysis ofgrit and phytolith content using a simplified version ofthe dry ashing method described by Piperno [26, 126 pp.]for the extraction of phytoliths from vegetation. Dis-tinguishing which of these two abrasives is responsiblefor microwear patterning in ungulates is of crucialimportance as it is becoming increasingly apparent thatsoil mineral particles may play as important a role in theformation of microwear [18,41,44] as do the highlevels of opal phytolith in graminaeous vegetationtraditionally held responsible for microwear signaturesin grazing and other ground-feeding species [33,43].After ashing as outlined previously, 10–20 ml of 10%HCl was added to the dung samples to remove carbon-ates. Four tablets containing a known volume ofLycopodium clavatum spores were then placed in theacid/dung mixture to allow an estimate to be made ofthe absolute counts of phytolith and grit particles [34].After the entire reaction had ceased (ca. 5–10 min), thesamples were washed through a 180 µm sieve withdistilled water and both residues (i.e. <180 µm and>180 µm) were air-dried in a drying cabinet. Slides wereprepared from the <180 µm residue using immersion oilwith a refractive index of 1.518( as a mounting mediumto enhance the identification of phytoliths (refractiveindex 1.41(–1.47() [26, 123 pp.]. The slides were viewedat 400� using a polarising microscope and a count wasmade of the frequency of grit, phytolith and Lycopodiumspores up to a total of 400 grit plus phytolith particles.Phytoliths and mineral particles were present in eachsample examined, and all but the very smallest particleswere readily distinguished using a combination of mor-phological and refractive characteristics (phytoliths, forexample, are isotropic [26] and will appear black undercrossed polars). It proved very difficult to distinguishbetween fragmentary phytoliths and grit particles of<ca.3–4 µm and thus, to avoid misidentification, onlyparticles larger than 4 µm were counted. No attempt wasmade to identify mineral particles to type nor phytolithsto genus or species, but ad hoc observation of the slidesindicated a predominance of quartz particles (Frederick,personal communication) and of a phytolith suite typi-cal of grasses and shrubs. Absolute concentrations of
grit and phytolith particles per 1 g of dried dung werecalculated from these counts using Tilia, a palynologicalsoftware program [9] (Table 8).
The >180 µm residue was weighed in grams to fourdecimal places. Analysis was based on three pelletsrather than a standard weight of dung and thus, to allowcomparison between samples, weights and counts forthe 180 µm residue were adjusted using the followingformula: sample weight >180 µm=observed weight of>180 µm fraction per sample/total weight of dried dungper sample. The subdivision of the acid-insoluble residue(i.e. <180 µm and >180 µm) and the different methodsemployed in quantifying the resulting fractions werenecessitated by the presence of large mineral particles insome samples, which hindered the preparation andviewing of slides made from the entire residue. An uppercut-off point of 180 µm for the detailed quantitativeanalysis of phytolith to grit ratios was deemed suitable,as most particles ingested by sheep will be smaller than180 µm [17,26].
Simple descriptive statistics were used to identify anyvariation between the grazing and browsing groups interms of overall inorganic content, absolute concen-tration of phytolith and grit particles, ratio of phytolithto grit particles (expressed as a percentage) and weightof the >180 µm residue (Table 8). The significance of anybetween group differences identified was assessed usingthe Student’s t-test.
3. Results
3.1. Microwear analysis
A clear difference is evident in microwear patterningbetween the two groups of sheep at Lejre, with thosepastured on the common characterised by a striatedenamel surface and those in the woodland pasture by anenamel surface comprised largely of pitted features withthe occasional striations (Figs. 1–2). These trends areconfirmed by the univariate statistics, which demon-strate a significantly higher frequency of pitted featuresin the woodland pasture group (Student’s t-test,p<0.01; Mann–Whitney, p<0.01). A higher ratio of
Table 5Statistics (mean values, range and SD for each variable) for microwear variables which contribute most to the discrimination achieved betweenthe Lejre sheep and grazing sheep from Greenland, Orkney and the Scottish borders
Diet group Ratio of pits to striations Defect frequency Ratio of rounded to ovoid pits
Mean Range SD Mean Range SD Mean Range SD
Lejre grazing 44.95 31.60–58.04 8.88 170.44 71.0–326.0 85.11 53.69 41.74–60.00 5.75Lejre browsing 75.79 33.06–99.00 18.46 140.95 29.0–311.0 102.92 69.43 58.02–83.83 8.87Scottish borders 81.00 59.10–89.10 13.51 58.5 11.00–144.00 43.56 71.24 54.05–92.31 11.41Greenland 69.34 14.82–100.00 18.83 87.39 11.00–204.00 49.43 64.02 32.08–100.00 13.96Orkney 42.31 17.47–86.67 20.89 89.54 34.00–175.00 48.98 43.54 17.39–56.73 12.42
See text for explanation of data sets and analytical procedure.
I.L. Mainland / Journal of Archaeological Science 30 (2003) 1513–15271518
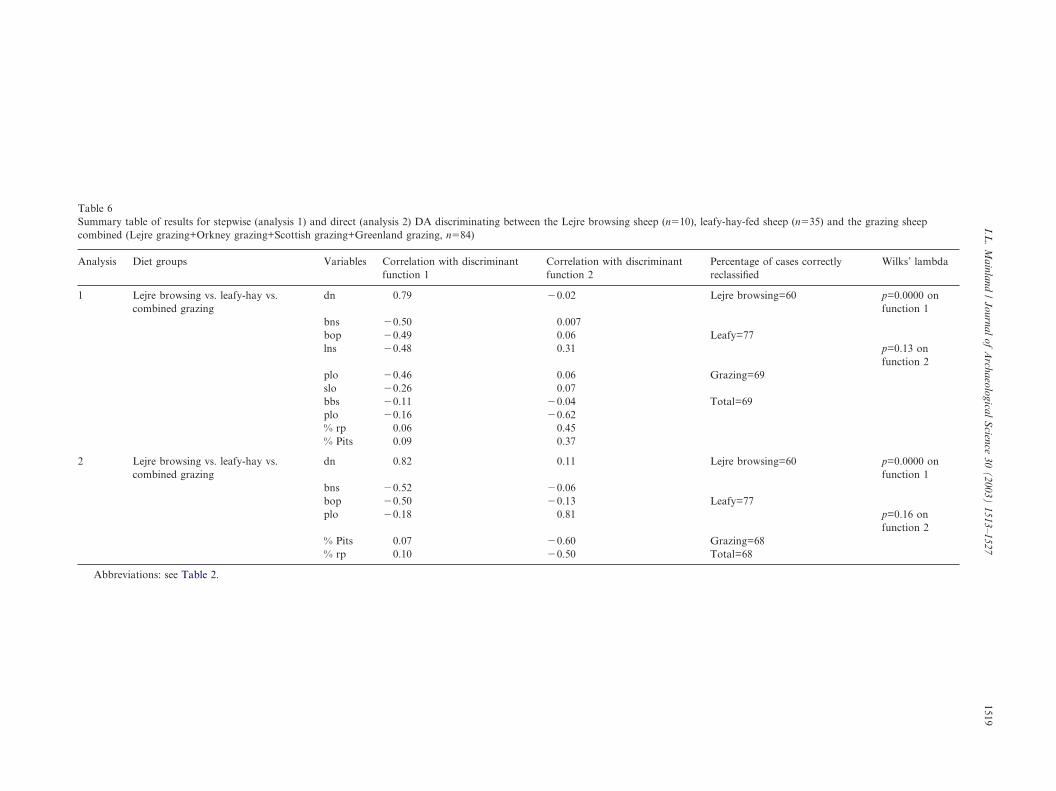
Table 6Summary table of results for stepwise (analysis 1) and direct (analysis 2) DA discriminating between the Lejre browsing sheep (n�10), leafy-hay-fed sheep (n�35) and the grazing sheepcombined (Lejre grazing+Orkney grazing+Scottish grazing+Greenland grazing, n�84)
Analysis Diet groups Variables Correlation with discriminantfunction 1
Correlation with discriminantfunction 2
Percentage of cases correctlyreclassified
Wilks’ lambda
1 Lejre browsing vs. leafy-hay vs.combined grazing
dn 0.79 �0.02 Lejre browsing=60 p=0.0000 onfunction 1
bns �0.50 0.007bop �0.49 0.06 Leafy=77lns �0.48 0.31 p=0.13 on
function 2plo �0.46 0.06 Grazing=69slo �0.26 0.07bbs �0.11 �0.04 Total=69plo �0.16 �0.62% rp 0.06 0.45% Pits 0.09 0.37
2 Lejre browsing vs. leafy-hay vs.combined grazing
dn 0.82 0.11 Lejre browsing=60 p=0.0000 onfunction 1
bns �0.52 �0.06bop �0.50 �0.13 Leafy=77plo �0.18 0.81 p=0.16 on
function 2% Pits 0.07 �0.60 Grazing=68% rp 0.10 �0.50 Total=68
Abbreviations: see Table 2.
I.L.
Mainland
/Journal
ofA
rchaeologicalS
cience30
(2003)1513–1527
1519
rounded to ovoid pits is also evident in this group,but no further statistically significant differences inmicrowear pattern are noted (Table 2).
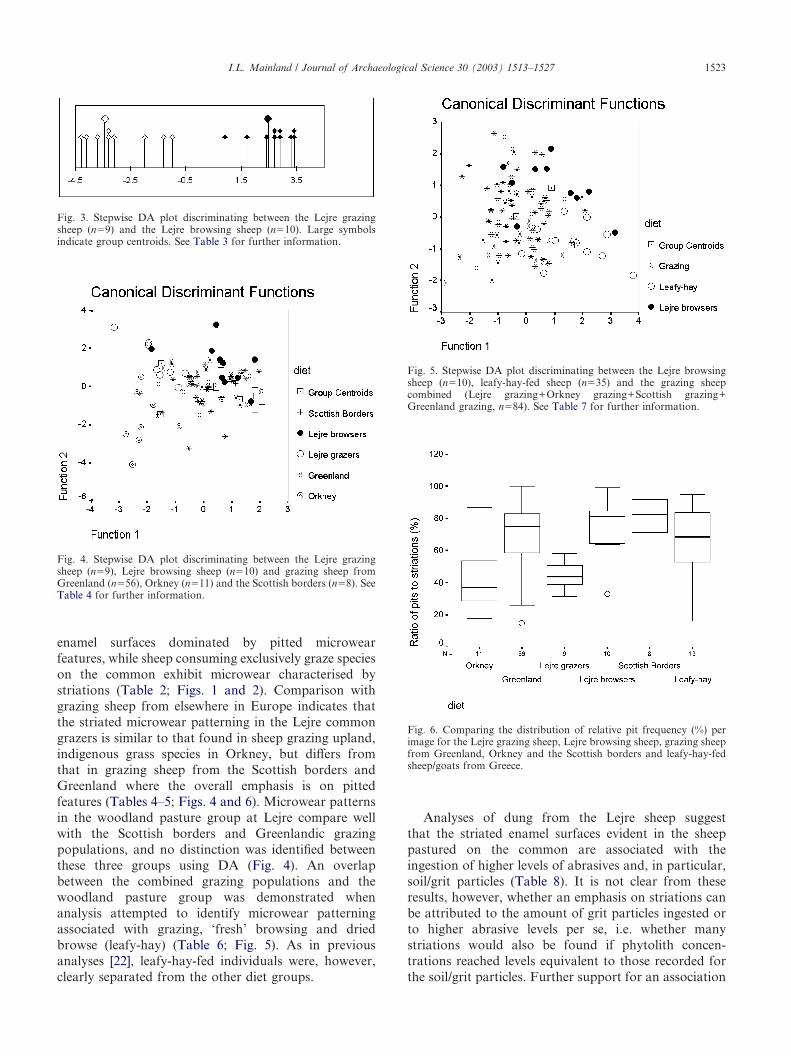
Stepwise DA selected 12 variables to discriminatebetween the ‘browsing’ and ‘grazing’ groups (analysis 1,Table 3). A good discrimination is achieved with a clearseparation of groups evident on the discriminant func-tion and 100% of cases assigned to the correct group(analysis 1, Table 3; Fig. 3). Wilks’ lambda indicates thatthese differences are significant (p#0.01). The pooledwithin-group correlation between discriminating vari-ables and discriminant function indicates that two vari-ables, the ratio of pits to striations and of rounded toovoid pits, contribute most to between group differences(Table 3). To explore whether the differences identifiedmight be attributable not to actual variation in micro-wear patterning but to chance due to a rather high ratioof variables to cases [35], a further direct DA wasundertaken using a restricted number of variables(n�6). The variables selected were those that con-tributed most to the discriminant function, as indicatedby the pooled within-group correlation between dis-criminating variables and discriminant function. Theclassification results obtained from this analysis com-pare well with the stepwise DA (analysis 2, Table 3)and reinforce the suggestion that the trends identifiedrepresent real differences in microwear patterning.
To assess how the microwear patterning in the Lejresheep compared with that evident in other grazing andfodder-fed sheep/goat populations, two further stepwisediscriminant analyses were undertaken. In the first ofthese, where the aim was to assess variation within andbetween different grazing populations, analysis focusedon five groups: the Lejre browsers (n�10), the Lejregrazers (n�9) and grazing sheep from Greenland(n�56), the Scottish borders (n�8) and Orkney (n�11).Fourteen variables were selected by stepwise DA (analy-sis 1, Table 4) and four functions were derived, with thefirst two accounting for 45 and 33% of the variance,respectively. Although the overall rate of reclassification
was low (66%), some trends are evident (Fig. 4). On thefirst function, some separation is evident between theLejre and Orkney grazers and the other groups; thiscan be attributed to two variables, the ratio of pitsto striations and the ratio of rounded to ovoid pits(Table 4), with a higher frequency of striated featuresand lower frequency of rounded pits evident in the Lejreand Orkney grazers (Table 5). No clear differences areevident between the Lejre browsers and the Greenlandicor Scottish grazers (Table 4). These trends are main-tained when a direct DA is undertaken using a reducedset of variables (analysis 2, Table 4).
The second stepwise DA aimed to assess whetherdifferences could be identified between grazing, freshbrowsing and animals fed leafy-hay and was based onthree groups; all grazers combined (n�84), animalsfed leafy-hay (n�13) and Lejre browsers (analysis 1,Table 6). Ten variables were selected and the twofunctions derived account for 78 and 22% of the vari-ance. Wilks’ lambda indicates that only the first functioncontributes significantly to group discrimination (analy-sis 1, Table 7). A reasonably good discrimination isachieved for leafy-hay, with 77% of cases correctlyassigned to this group (analysis 1, Table 6; Fig. 5).Greater overlap is evident between the combined grazersand the Lejre browsers (Fig. 5). This is reflected in therather low overall classification rate for the two func-tions (69%). Five variables are associated with the firstdiscriminant function: feature frequency, narrow stria-tion breadth, ovoid pit breadth, narrow striation lengthand ovoid pit length. Feature frequency is higher in theleafy-hay-fed sheep/goat, while feature dimensions aresmaller (Table 7). Once again these trends were main-tained under a direct DA using a reduced set of variables(analysis 2, Table 6).
3.2. Dung analysis
The dung of the ‘common’ grazing sheep differs fromthat of the woodland pasture sheep in having a higher
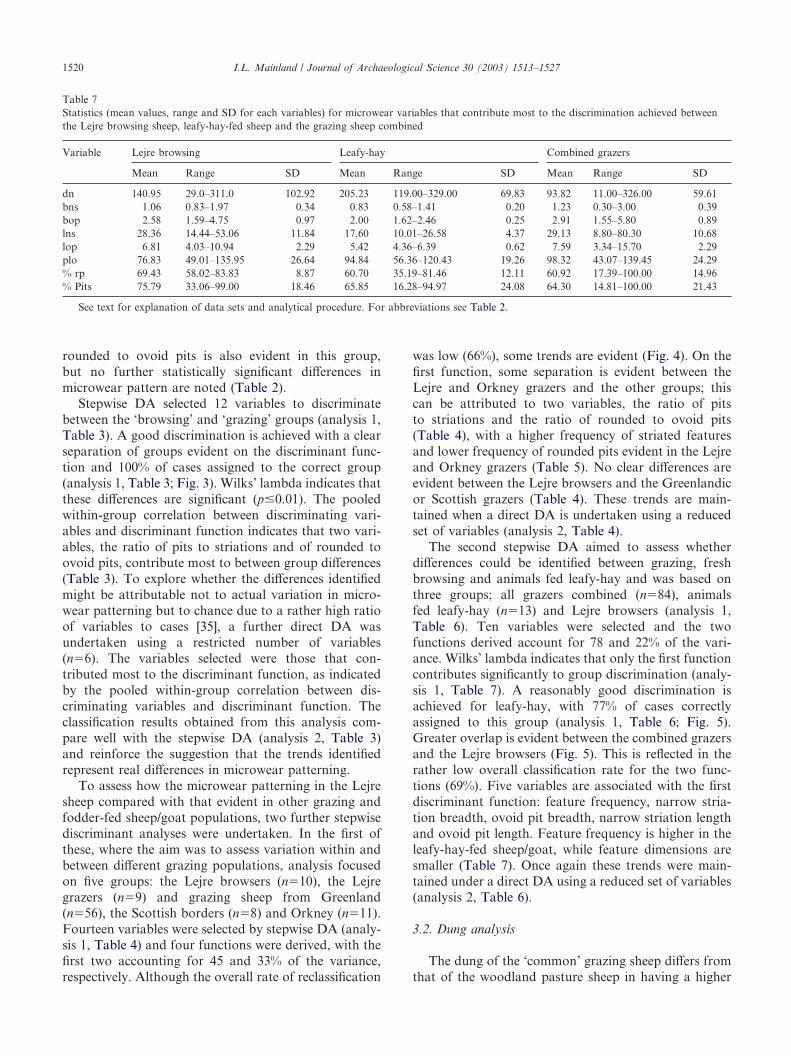
Table 7Statistics (mean values, range and SD for each variables) for microwear variables that contribute most to the discrimination achieved betweenthe Lejre browsing sheep, leafy-hay-fed sheep and the grazing sheep combined
Variable Lejre browsing Leafy-hay Combined grazers
Mean Range SD Mean Range SD Mean Range SD
dn 140.95 29.0–311.0 102.92 205.23 119.00–329.00 69.83 93.82 11.00–326.00 59.61bns 1.06 0.83–1.97 0.34 0.83 0.58–1.41 0.20 1.23 0.30–3.00 0.39bop 2.58 1.59–4.75 0.97 2.00 1.62–2.46 0.25 2.91 1.55–5.80 0.89lns 28.36 14.44–53.06 11.84 17.60 10.01–26.58 4.37 29.13 8.80–80.30 10.68lop 6.81 4.03–10.94 2.29 5.42 4.36–6.39 0.62 7.59 3.34–15.70 2.29plo 76.83 49.01–135.95 26.64 94.84 56.36–120.43 19.26 98.32 43.07–139.45 24.29% rp 69.43 58.02–83.83 8.87 60.70 35.19–81.46 12.11 60.92 17.39–100.00 14.96% Pits 75.79 33.06–99.00 18.46 65.85 16.28–94.97 24.08 64.30 14.81–100.00 21.43
See text for explanation of data sets and analytical procedure. For abbreviations see Table 2.
I.L. Mainland / Journal of Archaeological Science 30 (2003) 1513–15271520
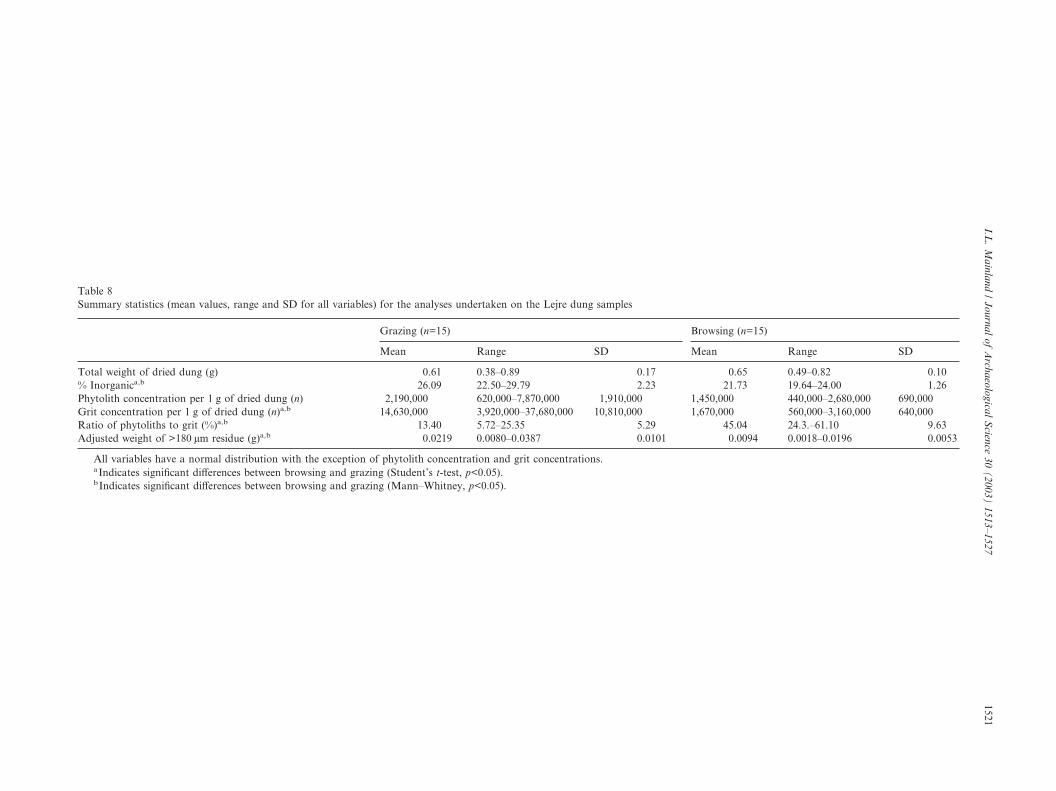
Table 8Summary statistics (mean values, range and SD for all variables) for the analyses undertaken on the Lejre dung samples
Grazing (n=15) Browsing (n=15)
Mean Range SD Mean Range SD
Total weight of dried dung (g) 0.61 0.38–0.89 0.17 0.65 0.49–0.82 0.10% Inorganica,b 26.09 22.50–29.79 2.23 21.73 19.64–24.00 1.26Phytolith concentration per 1 g of dried dung (n) 2,190,000 620,000–7,870,000 1,910,000 1,450,000 440,000–2,680,000 690,000Grit concentration per 1 g of dried dung (n)a,b 14,630,000 3,920,000–37,680,000 10,810,000 1,670,000 560,000–3,160,000 640,000Ratio of phytoliths to grit (%)a,b 13.40 5.72–25.35 5.29 45.04 24.3.–61.10 9.63Adjusted weight of >180 µm residue (g)a,b 0.0219 0.0080–0.0387 0.0101 0.0094 0.0018–0.0196 0.0053
All variables have a normal distribution with the exception of phytolith concentration and grit concentrations.aIndicates significant differences between browsing and grazing (Student’s t-test, p<0.05).bIndicates significant differences between browsing and grazing (Mann–Whitney, p<0.05).
I.L.
Mainland
/Journal
ofA
rchaeologicalS
cience30
(2003)1513–1527
1521

inorganic content, a greater concentrations of gritparticles in the <180 µm fraction, a higher concentrationof abrasives in the >180 µm fraction and a lower ratioof phytolith to grit particles (Table 8). Student’s t-testand Mann–Whitney indicates that these differencesare significant (p<0.05). Phytolith concentrations werecomparable between the two groups (Table 8).
4. Discussion
Microwear patterning in the Goth sheep fromLejre varies with diet/habitat. Sheep pastured on thewoodland area, in which deciduous browse species areestimated to have comprised at least 40% of vegetationconsumed in the weeks preceding slaughter, exhibit
Fig. 1. The microwear evident on the enamel surface of a grazing Gotland sheep at Lejre (top, anterior; left, lingual).
Fig. 2. The microwear evident on the enamel surface of a browsing Gotland sheep at Lejre (top, anterior; left, lingual).
I.L. Mainland / Journal of Archaeological Science 30 (2003) 1513–15271522
enamel surfaces dominated by pitted microwearfeatures, while sheep consuming exclusively graze specieson the common exhibit microwear characterised bystriations (Table 2; Figs. 1 and 2). Comparison withgrazing sheep from elsewhere in Europe indicates thatthe striated microwear patterning in the Lejre commongrazers is similar to that found in sheep grazing upland,indigenous grass species in Orkney, but differs fromthat in grazing sheep from the Scottish borders andGreenland where the overall emphasis is on pittedfeatures (Tables 4–5; Figs. 4 and 6). Microwear patternsin the woodland pasture group at Lejre compare wellwith the Scottish borders and Greenlandic grazingpopulations, and no distinction was identified betweenthese three groups using DA (Fig. 4). An overlapbetween the combined grazing populations and thewoodland pasture group was demonstrated whenanalysis attempted to identify microwear patterningassociated with grazing, ‘fresh’ browsing and driedbrowse (leafy-hay) (Table 6; Fig. 5). As in previousanalyses [22], leafy-hay-fed individuals were, however,clearly separated from the other diet groups.
Analyses of dung from the Lejre sheep suggestthat the striated enamel surfaces evident in the sheeppastured on the common are associated with theingestion of higher levels of abrasives and, in particular,soil/grit particles (Table 8). It is not clear from theseresults, however, whether an emphasis on striations canbe attributed to the amount of grit particles ingested orto higher abrasive levels per se, i.e. whether manystriations would also be found if phytolith concen-trations reached levels equivalent to those recorded forthe soil/grit particles. Further support for an association
Fig. 3. Stepwise DA plot discriminating between the Lejre grazingsheep (n�9) and the Lejre browsing sheep (n�10). Large symbolsindicate group centroids. See Table 3 for further information.
Fig. 4. Stepwise DA plot discriminating between the Lejre grazingsheep (n�9), Lejre browsing sheep (n�10) and grazing sheep fromGreenland (n�56), Orkney (n�11) and the Scottish borders (n�8). SeeTable 4 for further information.
Fig. 5. Stepwise DA plot discriminating between the Lejre browsingsheep (n�10), leafy-hay-fed sheep (n�35) and the grazing sheepcombined (Lejre grazing+Orkney grazing+Scottish grazing+Greenland grazing, n�84). See Table 7 for further information.
Fig. 6. Comparing the distribution of relative pit frequency (%) perimage for the Lejre grazing sheep, Lejre browsing sheep, grazing sheepfrom Greenland, Orkney and the Scottish borders and leafy-hay-fedsheep/goats from Greece.
I.L. Mainland / Journal of Archaeological Science 30 (2003) 1513–1527 1523

between heavily striated enamel surfaces and soilingestion in sheep is provided by analyses of abrasives inthe dung of the Greenlandic population (Table 9) [21].Microwear in these sheep exhibits few striated featuresand cannot be distinguished from that evident in theLejre woodland ‘browsers’ (Fig. 4). Dung analysis wasundertaken using identical procedures as those describedpreviously, and observation of minerals present in theslides again suggests an emphasis on quartz. Phytolithconcentrations for the Greenlandic sheep are com-parable with grit concentrations in the Lejre grazers(Tables 8 and 9). Moreover, no appreciable differencein overall quantities of abrasives is evident betweeneither of the Lejre groups and the Greenlandic sheep. Itcan be inferred, therefore, that it is not merely the totalamount of abrasives ingested, but the type of abrasivepresent, namely grit particles, that is responsible forstriations in the Lejre sheep (Tables 8 and 9). Neverthe-less, that a certain level of grit particles have to beingested before an emphasis on striations is evident issuggested by the relatively low frequencies of striationsin the Greenlandic grazing sheep, despite the presence ofgrit particles in the dung of this population.
The ingestion of soil particles has been equated withlarge pits in some grazing ungulate species, includingOvis canadensis and Kobus ellipsiprymus [31]. Microwearwill reflect both the nature of the abrasives consumedand the force and direction of jaw movements requiredto comminute food [8,37]. In primates and hominids, forexample, the ingestion of grit particles is reflected bylarge pitted features on the crushing facets of occlusalsurfaces, where predominately vertical jaw movementsprevail [36,39], but by striations on incisors where foodis pulled across teeth [41]. In sheep, transverse jawmovements predominate during the comminution andrumination of grasses [15,42], and it can be argued thatstriations rather than pits are likely to be formed by anygrit particles trapped between the occluding enamelsurfaces during grazing. Nevertheless, that soil ingestionmay in some circumstances be reflected by features other
than striations is suggested by microwear patterning inseaweed eating populations where a high incidence oflarge pitted features has been attributed to a combina-tion of the ingestion of sand along with seaweed and thepredominately vertical jaw movements required to com-minute seaweed, a gelatinous and bulky foodstuff [20].
Ground-feeding animals such as grazing sheep areknown to ingest significant quantities of soil along withvegetation during feeding [5]. The differing levels ofmineral/grit particles ingested by grazing sheep fromGreenland and Denmark can be attributed to factors,such as vegetation cover, grazing pressure and seasonof grazing: in grazing sheep, soil ingestion is reduced inpastures with a low stocking rate and/or dense vegeta-tion and is higher in winter when grass cover is patchier[6,12,17,23]. The Greenlandic flock was grazed exten-sively, at low stocking rates, on indigenous, uplandvegetation communities with good all year vegetationcover [21] and demonstrates low levels of soil ingestionin comparison with other populations of grazing sheep(e.g. Ref. [12]). The Lejre grazing sheep in contrast werepastured in an area of closely cropped grassland, whichis utilised fairly heavily throughout the year by bothsheep and cattle; this population exhibits levels of soilingestion within the middle–upper ranges for sheep (e.g.Ref. [12]). Although it was not possible to analyse dungsamples from Orkney, management practices are con-sistent with high levels of soil ingestion; overgrazing bysheep is a well-attested problem in Orkney [29], andpastures are very often heavily grazed, particularly dur-ing winter/spring [18,29]. Like the Greenlandic popula-tion, the sheep from the Scottish borders were pasturedat low stocking levels on upland grassland communitieswith dense vegetation cover and, consequently, arelikely to have ingested limited amounts of soil/mineralparticles [18]. Levels of soil ingestion will in generalbe lower in browsing animals than in grazers, sincebrowsers tend to forage for vegetation above groundlevel. The differences observed in the frequency of soilabrasives in the dung of the two groups of Goth sheep at
Table 9Summary statistics (mean values, range and SD for all variables) for the analyses undertaken on the Greenlandic dung samples
Greenland (n�40)
Mean Range SD
Total weight of dried dung (g) 0.58 0.35–1.05 0.155% Inorganica,b 9.01 2.04–14.63 2.45Phytolith concentration per 1 g of dried dung (n)a,b 5,150,000 1,050,000–15,380,000 3,650,000Grit concentration per 1 g of dried dung (n)b 2,560,000 470,000–8,700,000 1,950,000Ratio of phytoliths to grit (%)a,b 66.17 31.11–88.89 12.05Adjusted weight of >180 µm residue (g)a 0.0012 0.0000–0.0166 0.003
All variables have a non-normal distribution with the exception of percentage inorganic and ratio of phytoliths to grit.aIndicates significant differences between Greenland and Lejre browsers (Mann–Whitney, p<0.05).bIndicates significant differences between Greenland and Lejre grazing (Mann–Whitney, p<0.05).
I.L. Mainland / Journal of Archaeological Science 30 (2003) 1513–15271524

Lejre may, therefore, reflect differences in the height offoraging between the sheep grazing on the common andthose selecting a higher proportion of browse species inthe woodland pasture.
Analyses of microwear pattering and dung samples ingrazing, browsing and fodder-fed sheep from a widerange of different habitats in Europe have thus demon-strated an association between high levels of soilingestion and striated enamel surfaces and low levels ofsoil ingestion and pitted enamel surfaces. The presenceof striations under conditions of high soil ingestion canbe explained by the predominately transverse jaw move-ment exhibited by grazing sheep; variation in level of soilingestion is attributed to both grassland managementpractices (e.g. stocking levels, season of grazing) and tofeeding behaviour, specifically browsing/grazing heightwithin the vegetation canopy. The apparent associationbetween pitting and a diet low in abrasives is moredifficult to interpret. Teaford and Oyen [38] have sug-gested that a high frequency of small pitted features inprimates can be attributed to the close proximityachieved by teeth in the presence of soft food, whichconcentrates occlusal forces on small areas of the toothsurface and brings about the removal of tiny ‘prism-sized’ fragments of enamel through either an adhesivewear or by the creation of minute cracks in the enamelsurface. The comminution and/or rumination of a foodbolus containing few abrasive particles may have asimilar affect on ovicaprid tooth surfaces.
5. Conclusions: implications for dietary reconstruction
The associations evident in the Goth sheep, between adiet of predominately graze species and striated enamelsurfaces and between browse species and pitted enamelsurfaces parallels trends identified in various browsingand grazing ungulate species [31,33,43]. In these studies,the heavily striated surfaces evident in grazers wereattributed to the high phytolith content of grasses,and on the basis of Walker et al.’s [43] unquantifiedobservations of greater amounts of phytoliths in thedung of grazing than of browsing hyrax species. Analy-ses undertaken on dung from the Lejre and Greenlandicsheep, however, found no significant differences inphytolith concentrations between the ‘browsing’ andgrazing populations at Lejre and suggested ratherthat striations are formed through the ingestion of soilabrasives. Differences in microwear pattern between the‘browsing’ and grazing sheep at Lejre were attributed toforaging height and not variation in phytolith contentbetween browse and graze species. These findings thusquestion previous models for microwear formation pro-cesses in grazing and browsing ungulates and havesignificant implications for dietary reconstruction insuch species. As soil ingestion primarily occurs inground-feeding animals [5], it can be argued that striated
enamel features should be more commonly associatedwith grazing and are thus a fairly reliable ‘signature’ forthis dietary adaptation in species, which have predomi-nately lateral mandibular movements during the com-minution of grasses and forbs, such as the bovids [7].The sheep from Greenland and the Scottish bordersdemonstrate, however, that an emphasis on striationswill not necessarily be found in all grazing individualsand, moreover, that an absence of striations may reflectlow levels of soil ingestion rather than a browsingadaptation. These observations are, however, based onevidence for microwear formation processes in a singlespecies, the sheep, and, as such, may not be applicable toall ungulates owing to variations in feeding behaviour(e.g. whether grazing involves consumption of the entireplant, including the roots, or merely leaves and buds)and/or masticatory mechanics (e.g. degree of lateralmovement during mastication). Nevertheless, it is clearthat further research into microwear formation pro-cesses in the ungulates studied by Solounies et al. andWalker et al. [33,43] is now necessary to establish morefully the relationship between soil and phytolith levelsand microwear patterning in these species.
Research on microwear patterning in the Goth sheepat Lejre and on the wide range of grazing and fodder-fedsheep and goats described elsewhere [18,20–22] aimsultimately to evaluate the potential of dental microwearanalysis for exploring questions relating to the manage-ment and provision of grassland and fodder resourcesfor domesticated sheep and goat populations within anarchaeological context [19,21]. The research presentedhere indicates potential limitations associated with themethod, but equally has identified aspects of pasthusbandry practices on which microwear analysis willpotentially allow invaluable insights. The contrastingmicrowear patterns evident in grazing sheep populationskept at low stocking levels where soil ingestion is lowand the Lejre sheep where grazing is more intensive andsoil ingestion higher, for example, has important impli-cations for understanding animal husbandry practices inthe past, specifically in allowing insight into stockinglevels, overgrazing and pasture degradation [21]. Like-wise, the distinction evident between leafy-hay, i.e. a dietof dried browse and other diet groups indicates that thearchaeological recognition of this important fodder maybe achievable and further confirms earlier observa-tions on microwear patterning in leafy-hay-fed ovi-caprines [18,19,22]. The overlap observed between thewoodland pasture sheep and the combined grazingpopulation suggests, however, that for sheep at least itmay not always be possible to distinguish between‘fresh’ browsing and grazing individuals on microwearalone. Although it will undoubtedly limit archaeologicalapplication for this species in some contexts, in manycases, consideration of microwear patterning togetherwith other sources of evidence for foddering and grazing
I.L. Mainland / Journal of Archaeological Science 30 (2003) 1513–1527 1525

practices (archaeobotany, entomology, isotopes, etc.)would allow some indication of whether browsing orgrazing is the more likely interpretation. Nevertheless,these results are based on one population of browsingsheep which were, moreover, consuming some grazespecies and were observed during late autumn, a periodin which the amount of leafy material consumed is likelyto have been reduced in comparison to twigs and bark(Rasmussen and Jacobsen, personal communication).To assess more fully the potential of dental microwearanalysis for identifying fresh browsing in ovicaprines,further research is, therefore, required to determine howmicrowear is affected by season of browsing, the speciesbrowsed (e.g. coniferous vs. deciduous), browse materialselected (e.g. leaves vs. buds vs. bark) and intensity ofbrowsing.
Acknowledgements
This research was made possible through collabor-ation with the Historical Archaeological ResearchCentre at Lejre in Denmark. I would like to thank thestaff at Lejre for their help and advice throughout, and,in particular, Marianne Rasmussen for organising theproject, and Niels Jacobsen and Christian Kjargaard forsupplying information on the feeding behaviour of thesheep and collecting dung samples. I would also liketo thank Nils Andreasen for helping me in collectingthe mandibles from Lejre, Dr Charles Frederick andRobert Craigie of the Department of Archaeology andPrehistory, University of Sheffield, for advice regardingthe analysis of the dung samples and the staff of theMulti-Imaging Centre in Cambridge, where the SEMwork was undertaken. I am also grateful to Dr PaulHalstead and Dr Keith Dobney for invaluable criticalcomment on an earlier version of this article. Thecollection of the modern mandibles from Lejre and theanalysis of the dung samples were undertaken duringa British Academy Postdoctoral Fellowship in theDepartment of Archaeology and Prehistory, SheffieldUniversity.
References
[1] W. Anyonge, Microwear on canines and killing behaviour in largecarnivores: Saber function in Smilodon fatalis, Journal ofMammology 77 (1996) 1059–1067.
[2] G.W. Arnold, Factors within plant associations affecting thebehaviour and performance of grazing animals, in: D.J. Crisp(Ed.), Grazing in Terrestrial and Marine Environments,Blackwell, Oxford, 1964, pp. 133–154.
[3] B.E. Berglund, Handbook of Holocene Palaeoecology andPalaeohydrology, Wiley, Chichester, 1986.
[4] I. Beuls, B. De Cupere, P. Van Mele, M. Vermoere, M. Waelkens,Present-day traditional ovicaprine herding as a reconstructional
aid for understanding herding at Roman Sagalassos, in: M.Mashkour, A.M. Choyke, H. Buitenhuis, F. Poplin (Eds.), Pro-ceedings of the Fourth International Symposium on the Archaeo-zoology of Southwestern Asia and Adjacent Areas, ARCPublication 32, ARC, Groningen, 2000, pp. 216–223.
[5] W.N. Beyer, E.E. Connor, S. Gerould, Estimates of soil ingestionby wildlife, Journal of Wildlife Management 58 (1994) 375–382.
[6] A.C. Field, D. Purves, The intake of soil by grazing sheep,Proceedings of the Nutrition Society 23 (1964) 24.
[7] M. Fortelius, Ungulate cheek teeth: developmental, functionaland evolutionary interrelations, Acta Zoologica Fennica 180(1985) 1–76.
[8] K.D. Gordon, A study of microwear on Chimpanzee molars:implications for dental microwear analysis, American Journal ofPhysical Anthropology 59 (1982) 195–215.
[9] E.C. Grimm, Tilia and Tilia-Graph, Illinois State Museum,Springfield, 1991.
[10] P. Halstead, Ask the fellows who lop the hay: leaf-fodder in themountains of Northwest Greece, Rural History 9 (1998) 211–234.
[11] T. Hatting, Faret i oldtid and nutid, Forsøg Med Fortiden 4,Historisk Arckæologisk Forsøgscenter, Lejre, 1983.
[12] W.B. Healy, T.G. Ludwig, Wear of sheep’s teeth. I. The role ofingested soil, New Zealand Journal of Agricultural Research 8(1965) 737–752.
[13] H.M. Heybroek, Diseases and lopping for fodder as possiblecauses of a prehistoric decline of Ulmus, Acta BotanicaNeerlandica 12 (1963) 1–11.
[14] R.R. Hofmann, Evolutionary steps of ecophysiological adapta-tion and diversification in ruminants: a comparative view of theirdigestive system, Oecologia 78 (1989) 443–457.
[15] H.J. de Jongh, R. Dantuma, Electromyographic analysis ofmastication and rumination in sheep, Acta MorphologicaNeerlando-Scandinavica 24 (1987) 26–27.
[16] P.J. Lewis, M. Gutierrez, E. Johnson, Ondatra zibethicus(Arvicolinae, Rodentia) dental microwear patterns as apotential tool for palaeoenvironmental reconstruction, Journal ofArchaeological Science 27 (2000) 789–798.
[17] T.G. Ludwig, W.B. Healy, T.W. Cutress, Wear of sheep’s teeth.III Seasonal variation in wear and ingested soil, New ZealandJournal of Agricultural Research 9 (1966) 157–164.
[18] I.L. Mainland, Dental microwear and diet in domestic sheep(Ovis aries) and goats (Capra hircus): distinguishing grazing andfodder-fed ovicaprids using a quantitative analytical approach,Journal of Archaeological Science 25 (1998) 1259–1271.
[19] I.L. Mainland, The lamb’s last supper: the role of dental micro-wear analysis in reconstructing livestock diet in the past, Environ-mental Archaeology 1 (1998) 55–62.
[20] I.L. Mainland, A dental microwear study of seaweed-eatingand grazing sheep from Orkney, International Journal ofOsteoarchaeology 10 (2000) 93–107.
[21] I.L. Mainland, The potential of dental microwear for exploringseasonal aspects of sheep husbandry and management in NorseGreenland, Archaeozoolgia 11 (2001) 79–100.
[22] I.L. Mainland, Dental microwear in modern Greek ovicaprids:identifying microwear signatures associated with a diet of leafy-hay, in: E. Kotjabopoulou, Y. Hamilakis, P. Halstead, C. Gamble(Eds.), Zooarchaeology in Greece: Recent Advances, in press.
[23] T. Nolan, W.J.M. Black, Effect of stocking rate on tooth wear inewes, Irish Journal of Agricultural Research 9 (1970) 187–196.
[24] M.J. Norusis, SPSS/PC+Advanced Statistics 4.0, SPSS Inc,Chicago, 1990.
[25] J.A. Pfister, J.C. Malechek, Dietary selection by goats and sheepin a deciduous woodland of northeastern Brazil, Journal of RangeManagement 39 (1986) 24–28.
[26] D. Piperno, Phytolith Analysis. An Archaeological andGeological Perspective, Academic Press, London, 1988.
I.L. Mainland / Journal of Archaeological Science 30 (2003) 1513–15271526

[27] J.C. Rose, P.S. Ungar, Gross dental wear and dental microwearin historical perspective, in: K.W. Alt, F.W. Rosing, M.Teschler-Nicola (Eds.), Dental Anthropology, Springer, Vienna,1998.
[28] M.L. Ryder, Sheep and Man, Duckworth, London, 1983.[29] I. Simpson, L. Scott, A.H. Kirkpatrick, A.J. MacDonald, Sheep
grazing on the moorland landscapes of Orkney and its implica-tions for nature conservation, in: G.H. Griffiths (Ed.), LandscapeEcology: Theory and Application, IALE, Aberdeen, 1995,pp. 40–48.
[30] P.J. van Soest, Nutritional Ecology of the Ruminant, OB Books,Corvallis, 1982.
[31] N. Solounias, L.A.C. Hayek, New methods of tooth microwearanalysis and application to dietary determination of two extinctungulates, Journal of Zoology 229 (1993) 421–445.
[32] N. Solounias, S.M.C. Moelleken, Tooth microwear analysis ofEotragus sansaniensis (Mammalia Ruminantia): one of the oldestknown bovids, Journal of Vertebrate Palaeontology 12 (1992)113–121.
[33] N. Solounias, M. Teaford, A. Walker, Interpreting the diet ofextinct ruminants: the case of a non-browsing giraffid,Paleobiology 14 (1988) 287–300.
[34] J. Stockmarr, Tablets with spores used in absolute pollen counts,Pollen and Spores 13 (1971) 615–621.
[35] B.G. Tabachnick, L.S. Fidell, Using Multivariate Statistics, thirded., Harper Collins, New York, 1996.
[36] M.F. Teaford, Dental microwear and diet in extant and extinctTheropithecus: preliminary analyses, in: N.G. Jablonski (Ed.),Theropithecus: The Life and Death of a Primate Genus,Cambridge University Press, Cambridge, 1993, pp. 331–349.
[37] M.F. Teaford, Dental microwear and dental function,Evolutionary Anthropology 17 (1994) 17–30.
[38] M.F. Teaford, O.J. Oyen, Differences in the rate of molar wearbetween monkeys raised on different diets, Journal of DentalResearch 68 (1989) 1513–1518.
[39] M.F. Teaford, C. Spencer, R.F. Pastor, V.E. Noble, Pits andscratches: microscopic evidence of tooth use and masticatorybehaviour in La Florida, in: C.S. Larsen (Ed.), Bioarchaeologyof La Florida: Human Biology in the Northern FrontierNew Spain, University Press of Florida, Florida, 2001,pp. 82–112.
[40] P.S. Ungar, Microwear Image Analysis Software. Version 2.2,1995.
[41] P.S. Ungar, M.F. Teaford, Preliminary examination of non-occlusal dental microwear in Anthropoids: implications for thestudy of fossil primates, American Journal of PhysicalAnthropology 100 (1996) 101–113.
[42] F.L. de Vree, C. Gans, Masticatory responses of pygmy goats(Capra hircus) to different foods, American Zoologist 25 (1976)1342–1343.
[43] A.C. Walker, H. Hoek, L. Perez, Microwear of mammalian teethas an indicator of diet, Science 201 (1978) 908–910.
[44] J. Ward, I.L. Mainland, Microwear in modern free-ranging andstall-fed pigs: the potential of dental microwear analysis forexploring pig diet and management in the past, EnvironmentalArchaeology 4 (1999) 25–32.
[45] S. Payne, Reference codes for wear states in the mandibular cheekteeth of sheep and goats, Journal of Archaeological Science 14(1987) 609–614.
I.L. Mainland / Journal of Archaeological Science 30 (2003) 1513–1527 1527





























