Embed Size (px)
Citation preview

Dental Microwear and Stable Isotopes Inform thePaleoecology of Extinct Hominins
Frederick E. Grine,1,2* Matt Sponheimer,3 Peter S. Ungar,4 Julia Lee-Thorp,5 and Mark F. Teaford6
1Department of Anthropology, Stony Brook University, Stony Brook, NY 11794-43642Department of Anatomical Sciences, Stony Brook University, Stony Brook, NY 11794-80813Department of Anthropology, University of Colorado at Boulder, Boulder, CO 80309-02334Department of Anthropology, University of Arkansas, Fayetteville, AR 727015Research Laboratory for Archaeology and the History of Art, University of Oxford, Oxford OX1 3QY, UK6Department of Physical Therapy, School of Health Sciences, High Point University, High Point, NC 27262-3598
KEY WORDS occlusal microwear; carbon isotopes; oxygen isotopes; trace elements; teeth; diet;dietary adaptation; paleoecology; Ardipithecus ramidus; Australopithecus anamensis; Australopithecusafarensis; Australopithecus africanus; Paranthropus robustus; Paranthropus boisei
ABSTRACT Determining the diet of an extinctspecies is paramount in any attempt to reconstruct itspaleoecology. Because the distribution and mechanicalproperties of food items may impact postcranial, cranial,mandibular, and dental morphologies related to their pro-curement, ingestion, and mastication, these anatomicalattributes have been studied intensively. However, whilemechanical environments influence skeletal and dentalfeatures, it is not clear to what extent they dictate partic-ular morphologies. Although biomechanical explanationshave been widely applied to extinct hominins in attemptsto retrodict dietary proclivities, morphology may say asmuch about what they were capable of eating, and per-haps more about phylogenetic history, than about the na-ture of the diet. Anatomical attributes may establish
boundary limits, but direct evidence left by the foods thatwere actually (rather than hypothetically) consumed isrequired to reconstruct diet. Dental microwear and thestable light isotope chemistry of tooth enamel providesuch evidence, and are especially powerful when used intandem. We review the foundations for microwear andbiogeochemistry in diet reconstruction, and discuss thisevidence for six early hominin species (Ardipithecus rami-dus, Australopithecus anamensis, Au. afarensis, Au. afri-canus, Paranthropus robustus, and P. boisei). The dietarysignals derived from microwear and isotope chemistry aresometimes at odds with inferences from biomechanicalapproaches, a potentially disquieting conundrum that isparticularly evident for several species. Am J PhysAnthropol 148:285–317, 2012. VVC 2012 Wiley Periodicals, Inc.
The centrality of diet to nearly every aspect of an ani-mal’s ecology and behavior has meant that its determi-nation is paramount in any attempt to reconstruct thepaleoecology of extinct taxa. The seasonal availabilityand quality of foods impact species attributes rangingfrom mobility patterns to population size and social orga-nization (Clutton-Brock and Harvey, 1977). Moreover,the distribution and mechanical properties of food itemsmay impact postcranial, cranial, mandibular, and dentalmorphologies related to their procurement, ingestion,and mastication (e.g., Hylander, 1975; Kay, 1978; Kinzey,1978; Eaglen, 1984; Daegling, 1992; Anthony and Kay,1993; Ungar, 1996; McGraw, 1998; Daegling andMcGraw, 2001; Taylor, 2002; Wright, 2005; Lucas et al.,2008b; Ungar and Bunn, 2008; Vogel et al., 2008). Conse-quently, these anatomical attributes have been the sub-ject of intense study in attempts to retrodict the dietaryhabits of fossil species.Although strictly adaptationist explanations of ana-
tomical design have been criticized as Panglossian story-telling (Gould and Lewontin, 1979), at least some mor-phological configurations appear to be explicable withreference to the function(s) that they serve. However,exactly how closely the structure of a trait can be relatedto its apparent biological function remains a topic of dis-covery. Mechanical loads clearly influence the skeletalsystem, but it is not clear to what extent mechanicalenvironments dictate particular morphologies. Thus, forexample, mandibular corpus robusticity may be related
more directly to allometric considerations than to dietaryfactors (Ravosa, 2000), and certain aspects of jaw mor-phology (e.g., symphyseal depth) appear to be somaticscaling artifacts that may secondarily affect masticatorybiomechanics (Smith, 1993).Biomechanical explanations for morphological design
have been widely applied to extinct hominins inattempts to understand their dietary proclivities (e.g.,Rak, 1983; Demes and Creel, 1988; Hylander, 1988; Dae-gling and Grine, 1991, 2007; Ungar, 2004, 2007; Machoet al., 2005; Lucas et al., 2008a; Rak and Hylander,2008; Strait et al., 2009; Wroe et al., 2010). However, thesuccess of such applications has been anything but uni-
Grant sponsors: National Science Foundation SBR 9804882,National Research Foundation of South Africa, The Wenner GrenFoundation, The Leakey Foundation, The University of Cape Town,The University of Bradford, The University of Colorado, Boulder,The University of Oxford.
*Correspondence to: Frederick E. Grine, Departments of Anthro-pology & Anatomical Sciences, Stony Brook University, Stony Brook,NY 11794-4364, USA. E-mail: [email protected]
Received 19 February 2011; accepted 30 March 2012
DOI 10.1002/ajpa.22086Published online in Wiley Online Library
(wileyonlinelibrary.com).
VVC 2012 WILEY PERIODICALS, INC.
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 148:285–317 (2012)

form (Grine et al., 2010). They have been limited by themonumental challenge of successfully distinguishingamong the dizzying array of influences on cranial, man-dibular, and dental form, and they are bedeviled by thefact that while craniodental morphology may speak toadaptation, it may tell us more about what an extinctspecies (or individual) was capable of eating, and per-haps more about its phylogenetic history than the consti-tution of its diet.Phylogenetic context provides a valuable source of in-
formation by which to strengthen inferences from com-parative studies, but it can also be a hindrance to directfunction–behavior correlations because genetically deter-mined ancestral patterns of morphology may simply beretained as plesiomorphies in descendant taxa (Ruff andRunestad, 1992; Nee et al., 1996; Ross et al., 2002). Thisis because anatomies can be maintained passively in alineage as the result of a constrained process of inheri-tance that need not involve directional selection. Thisphenomenon is referred to commonly as ‘‘phylogeneticinertia’’—although the concept has meant differentthings to different workers (see Blomberg and Garland[2002] for a review)—and it may be rooted in pleiotropy,genetic linkage, or developmental pathways that canimpose phylogenetic constraints (Cheverud, 1988; Reeveand Sherman, 1993; Hanson and Orzack, 2005).Liem’s (1980) studies of African cichlid fishes led him
to observe that apparently specialized feeders frequentlyconsume nonspecialized resources. This seemingly con-tradictory phenomenon, which has been dubbed ‘‘Liem’sParadox,’’ highlights inconsistencies between trophicmorphologies that are identified as adaptive and ecologi-cal observations (Robinson and Wilson, 1998; Liem andSummers, 2000; Lambert et al., 2004; Binning et al,2009). Animals may actually avoid those foods to whichtheir craniodental traits are seemingly adapted whenmore nutritious or more easily consumed and digesteditems are available to them. Thus, for example, gorillaspossess long molar shearing crests and are capable ofprocessing very tough foods, but they will choose fruitshigh in nonstarch sugars over lower sugar, higher fibersitems when they are able to do so (Remis, 2002). Still, itis the leaves and pith that are consumed when preferredfruits are unavailable to which gorilla teeth are adapted.As noted by Kinzey (1978: 378), ‘‘when a food item is
critical for survival, even though not part of the primaryspecialization, it will influence the selection of dentalfeatures.’’ Even if such fallback food items are consumedonly rarely, they are likely to have significant mechani-cal defenses and lower energy yields, with the resultthat they are the very items that require dental special-izations. The question of whether a given trophic featurein an extinct species reflects phylogenetic inertia, an ad-aptation to preferred foods, or an adaptation to process-ing important albeit less commonly eaten fallback itemsis not a trivial one. In order to address it, measures ofdiet that are independent of an animal’s genetic historyare required.A variety of indirect measures have enabled informed
statements concerning the dietary habits of extincthominin species, and some are obviously more problem-atic than others in the degree to which the sources ofdata are removed from the object of discovery. Thus,inferences concerning paleodiets have been made fromparasite relationships (Hurtado et al., 2008), microbialecology (Ley et al., 2008), and genomic information(Wang et al., 2004; Perry et al., 2007; Tishkoff et al.,
2007; Babbitt et al., 2010). In addition, contextual evi-dence from archaeological refuse, which is certainlyopen to different interpretation and often misses wholeimportant classes of foods (e.g., plants), has beenemployed to infer hominin diets. All such methods, how-ever, require secondary and even tertiary levels of infer-ential argument.Ideally, what is required is direct evidence left by the
foods that were actually (rather than hypothetically)consumed during an individual’s lifetime. In this regard,there are at least two lines of evidence that homininpaleontologists may turn to: dental microwear and thechemical composition (especially the stable light isotoperatios) of tooth enamel. In contrast to biomechanical/adaptive models of jaw and tooth design, chemistry andmicrowear preserve nongenetic signals directly relatedto an individual’s diet. As such, dental microwear andstable isotope chemistry individually, and especially incombination, provide very potent signals relating toearly hominin diet and paleoecology. Understandably,these signals may sometimes be at odds with inferencesderived from strict adaptationist approaches to paleon-tology. That is not to say that dental microwear and sta-ble isotope chemistry are not without their own particu-lar problems and limitations, but if these are recognized,and the data are interpreted in the context of reasonablyinformed paleohabitat reconstructions, they can providea wealth of information pertaining to diet that goesbeyond the anecdotal.We here review the fundamental precepts of dental
microwear and stable isotope chemistry as they havebeen applied to dietary inference in the early homininfossil record. We discuss the potential problems (e.g.,taphonomic or diagenetic artifacts) and the limitationsthat may be encountered with each approach, andthen employ these data to examine the diets of speciesof Ardipithecus, Australopithecus, and Paranthropus.For the purposes of this review, we have chosen toeschew the genus Homo. Although information relatingto the dietary proclivities of some of its members hasbeen obtained from microwear and isotope studies(e.g., Richards, et al., 2000, 2001; Ungar et al.,2006a,b; van der Merwe et al., 2008; El Zaatari et al.,2011), the Early–Middle Pleistocene fossil record forthis genus is beset with problems of alpha taxonomy(e.g., Blumenschine et al., 2003; Spoor et al., 2007;Baab, 2008; Rightmire, 2008; Smith and Grine, 2008;Grine and Fleagle, 2009).We examine the published microwear and/or isotopic
evidence for six Plio-Pleistocene hominin species: Ardipi-thecus ramidus, Australopithecus anamensis, Au. afaren-sis, Au. africanus, Paranthropus robustus, and P. boisei.We have chosen these taxa because microwear and/orstable isotope evidence has been published for each (Ta-ble 1). Unfortunately, both lines of evidence are notavailable in tandem for either Au. anamensis or Au.afarensis (carbon isotope data have not been publishedfor either), and the only microwear information availablefor Ar. ramidus is anecdotal. Moreover, the number ofspecimens that have been sampled for microwear analy-sis (or that even preserve good antemortem molar micro-wear) is often very small. Nevertheless, and notwith-standing these caveats, the microwear and isotopic datathat have been garnered provide potent insights into thedietary habits of these extinct hominins, especially whenthey are considered in the context of paleohabitat recon-structions.
286 F.E. GRINE ET AL.
American Journal of Physical Anthropology

DENTAL MICROWEAR AND DIET
Dental microwear has been extensively studied sincethe early 1980s in attempts to elucidate the dietary hab-its of extinct hominin species (e.g., Grine, 1981, 1986;Walker, 1981; Puech et al., 1983; Puech and Albertini,1984; Grine and Kay, 1988; Ryan and Johanson, 1989;Ungar and Grine, 1991; Teaford et al., 2002; Scott et al.,2005; Grine et al., 2006a,b; Ungar et al., 2006a,b, 2008,2010, in press). The demonstrable relationship betweenocclusal surface wear textures and the properties of die-tary items has been amply documented, such that micro-wear is capable of distinguishing among broad dietarycategories when their constituent items differ in theirfracture properties (Walker et al., 1978; Puech et al.,1981; Teaford and Walker, 1984; Teaford, 1985, 1988a;Teaford and Glander, 1991, 1996). Moreover, subtle dif-ferences in the diets of closely related species, as well asshort-term variations in diet can be detected throughmicrowear analysis, enabling the investigation of sea-sonal and other ecological differences (Teaford and Oyen,1989; Teaford and Robinson, 1989; Teaford and Glander,1991, 1996; Teaford and Runestad, 1992; Mainland,2003; Merceron et al., 2004).The turnover of microwear in extant species, which
has been amply documented in both the laboratory andthe field (Teaford and Oyen, 1989; Teaford and Glander,1991, 1996; Merceron et al., 2010), means that informa-tion pertaining to meals consumed just before an individ-ual’s death will often be fossilized. This is the so-called‘‘Last Supper Effect’’ (Grine, 1986). The amount of timerepresented by this effect is a direct function of the typesof foods consumed. Thus, for example, modern humansconsuming a ‘‘Western diet’’ show a much slower rate ofturnover in dental microwear than do wild-caught howl-ing monkeys (Teaford and Glander, 1991; Teaford andTylenda, 1991). However, all studies of turnover in pri-mates have focused on the ‘‘lifespan’’ of individual micro-wear features during the consumption of specific diets;none has documented the differential ‘‘survival’’ of larger
versus smaller features in response to large-scale dietchanges. Still, microwear fabrics may reflect subtle,short-term variations in dietary items, and this may bea potentially confounding influence in the analysis of fos-sil samples since these may accumulate over a prolongedperiod of time or preferentially represent certain sea-sons, physical geographies, and/or climatic conditions. Assuch, the representation of individuals in a fossil assem-blage may be taphonomically biased (i.e., sampled inunequal proportions) vis-a-vis the parent populationsfrom which they derived.For example, it has been assumed that fallback foods
should be ‘‘over-represented’’ in the paleontological re-cord because of the notion that primates most often dieduring times of food resource stress (Kimbel and Dele-zene, 2009). This is intuitively appealing, and there issome evidence from field studies of modern primates tosupport differential mortality being tied to food abun-dance, although these data are commonly tied to periodsof severe, prolonged drought (e.g., Otis et al., 1981;Gould et al., 1999; Nakagawa et al., 2003). However,other field studies of primate populations have found lit-tle to no association between food availability and mor-tality (e.g., Watts, 1998; Dunbar, 1980; Williams et al.,2008). Seasonal mortality may be driven by predationand/or disease factors (e.g., increased parasitism)(Huffman et al., 1997; Cheney et al., 2004; Chapman etal., 2010) that may be unrelated to dietary stress. Giventhe mixed evidence, it is uncertain that fallback foodsshould be overrepresented in taphonomic assemblages offossil hominins. We clearly need more field studies of dif-ferential mortality in living primate populations, espe-cially those inhabiting the mosaic woodland-savannaenvironments envisioned for early hominins, to begin toaddress this issue.Turnover, of course, means that microwear traces of
diet may be somewhat ephemeral. But rather than mani-festing as a confounding factor, the ‘‘Last Supper Effect’’means that one can decipher the actual dietary habits of
TABLE 1. Pliocene and Pleistocene hominin taxa for which microwear and/or stable carbon isotope data are available (species thathave been attributed to the genus Homo are excluded)
Species d13C data n Microwear data n Reference
Ardipithecus ramidus Yes 5 White et al. (2009b)SEM (anecdotal) 4 Suwa et al. (2009)
Australopithecus anamensis No –SEM 3 Grine et al. (2006a)Texture 3 Ungar et al. (2010)
Australopithecus afarensis No –SEM 19 Grine et al. (2006b)Texture 19 Ungar et al. (2010)
Australopithecus africanus Yes 24 Sponheimer and Lee-Thorp (1999a)van der Merwe et al. (2003)Sponheimer et al. (2005b)Lee-Thorp et al. (2010)
SEM 10 Grine (1986)Texture 10 Scott et al. (2005)
Paranthropus robustus Yes 22 Lee-Thorp et al. (1994)Sponheimer et al. (2005b, 2006b)
SEM 9 Grine (1986)Texture 9 Scott et al. (2005)
Paranthropus boisei Yes 24 van der Merwe et al. (2008)Cerling et al. (2011)
SEM (anecdotal) ? Walker (1981); Suwa et al. (1997)Texture 8 Ungar et al. (2008, in press)
SEM (anecdotal) indicates qualitative observations only. ? refers to the fact that Walker (1981) did not state the number of speci-mens examined, and Suwa et al. (1997) state that they examined ‘‘a few’’ specimens.
287MICROWEAR, ISOTOPES, AND HOMININ PALEOECOLOGY
American Journal of Physical Anthropology

an individual at a given point (or span) of time. As such,the interpretive strength of occlusal microwear is that itrepresents direct evidence left by the foods that wereactually (rather than theoretically, or potentially) con-sumed.The demonstrable relationship between microwear fab-
rics and diet is one between enamel and the properties ofthe foods consumed. More particularly, this relationshipreflects occlusal mechanics, where the angle of approachbetween opposing teeth is guided, if not dictated by thefracture properties of food items. A frequent misunder-standing behind arguments that food items are either toobig or too soft to cause microwear is the notion that thefood item itself is solely responsible. Rather, as demon-strated experimentally, it is the abrasives either in or ona food item that cause microwear, with microwear featuresize, shape, and orientation depending in large measureon whether the abrasives are scraped along a surface(causing striations in the direction of horizontal slip), orpressed into the enamel with force directed normal tothat surface (causing pitting) (Maas, 1994; Gugel et al.,2001). Thus, hard foods likely cause pits because pieces ofthem or their associated abrasives are pressed into theenamel as teeth approach one another with forces normalto the occlusal plane. Tough foods cause scratches as theabrasives are dragged along the occlusal surface as oppos-ing teeth shear past one another. At the same time, how-ever, Brazil nuts fed to laboratory Cebus monkeys havebeen observed to result in the formation of pits as well asscratches on molars (Teaford et al., 2010), even though ithas yet to be determined whether it is the pieces of thenuts, the abrasives on and/or in them, or some combina-tion thereof that cause these features.What is abundantly clear, however, is that microwear
fabrics reflect occlusal movements that relate directly tothe fracture properties of the foods being chewed.Numerous studies have revealed a consistent associationbetween microwear texture patterns and diet across abroad range of mammals that exhibit a great variety ofcraniodental morphologies. Animals that crush hard,brittle foods typically have complex, pitted occlusalsurfaces compared with closely related taxa that sheartough items. The latter tend to show more anisotropicsurfaces with long, parallel striations. This holdswhether the comparisons are between primates thatmasticate tough leaves and those that consume hard-husked fruits, or between carnivores that specialize onmeat and sinew and those that chew bone (e.g., Ungaret al., 2007b; Schubert et al., 2010; Scott et al., 2012).Similarly, the microwear fabrics of tough-grass grazerstend to be characterized by greater homogeneity (paral-lelism or anisotropy) in scratch orientation than those ofmore generalist browsers, whether the comparisons areamong African bovids (e.g., reedbucks and kudus) orAustralian marsupials (e.g., kangaroos and wallabies)(Ungar et al., 2007a; Prieaux et al., 2009).Silica, whether biogenic or pedogenic, is among the abra-
sives that most often have been implicated in the formationof microwear. Whereas pedogenic silica forms quartziticparticles (SiO4) and thus comprises the bulk of ‘‘exogenous’’abrasives, biogenic silica bodies (phytoliths) form in and/orbetween the cell walls of plants as phytoliths as a result ofthe adsorption of orthosilicic acid (H4O4Si) from ground-water (Kalisz and Boettcher, 1990).The role of phytoliths in producing microwear,
although questioned by Sanson et al. (2007), has beenamply documented in numerous experimental and natu-
ralistic studies since the seminal work by Walker et al.(1978) on hyraxes (e.g., Maas, 1994; Lucas and Teaford,1995; Buchet et al., 2001; Gugel et al., 2001; Reinhardand Danielson, 2005; Teaford et al., 2006; Hummel etal., 2010).Phytoliths, which are nearly ubiquitous and may be
particularly abundant in some plant tissues (Epstein,1994), require some 6,000 mega-Pascals (Mpa, one mil-lion Pascals) of force per unit area to deform (Lucas andTeaford, 1995). Even stress-limited foods, such as hardseeds, require significantly less force to break (Rabenoldand Pearson, 2011), and tooth enamel can be scratchedwith about half the force (3,700 Mpa) (Waters, 1980)required to deform a phytolith.Of course, point contacts between enamel surfaces and
hard foods, parts of hard foods, and/or silica particles arealmost certainly going to vary in size and shape, withmany likely under 1 mm2 (where Mpa are defined as N/mm2). As such, varying bite forces and bite points willserve to further complicate the straightforward interpreta-tion of the stiffness–deformation relationship of such itemswith tooth enamel. Moreover, tooth enamel is not a mono-morphic substrate, such as glass, and to treat it as such inmodels of wear and fracture (e.g., Lucas et al., 2008b),while perilous, is perhaps a necessary first step in whatwill undoubtedly become more complex analyses.In addition, alteration of enamel structure through ex-
posure to an acidic environment, such as might easily beproduced by plaque bacteria and/or the consumption ofvarious foodstuffs, renders it more easily indented(Cheng et al, 2009). The effects of the structural modifi-cation of tooth enamel (and dentin) by dilute acid treat-ment to facilitate its subsequent mechanical removal(e.g., using silicon carbamide microparticles) have beenlong recognized by general and cosmetic dentists alike(Maragakis et al., 2001; Ardu et al., 2009). Although che-momechanical methods are used widely in dentistry, che-momechanical processes are not well understood insofaras they relate to enamel wear. This is due, in part, tothe chemical and structural complexity of this tissue,which may be characterized by prismatic and interpris-matic substance with varying degrees of mineralization,and by prisms of differing size, shape, and orientation.In short, full appreciation of the effects of various che-momechanical processes on tooth enamel wear willrequire more work on dental tribology in the context ofthe oral environment. While it is evident that erosionincreases enamel’s susceptibility to abrasion, we havejust begun to scratch the surface of this topic.
Exogenous grit
As noted above, exogenous grit may also play a role inthe formation of dental microwear (Ungar et al., 1995;Daegling and Grine, 1999; Silcox and Teaford, 2002;Mainland, 2003; Nystrom et al., 2004). For example,Nystrom et al. (2004) have shown the effects of seasonand grassland phenology on microwear patterning forbaboons in the Awash National Park, Ethiopia. In someinstances, the role of exogenous agents in the formationof microwear may be detectable in exaggerated levels ofwear feature variability. Thus, in the baboon populationstudied by Daegling and Grine (1999), underground fooditems (especially Eriospermum bulbs) accounted forupwards of 90% of feeding observations during the dryseason, and the abundance of exogenous grit in theirdiet is reflected in microwear feature size variation that
288 F.E. GRINE ET AL.
American Journal of Physical Anthropology

far exceeds that observed range for other primate spe-cies. Exogenous grit has been implicated in the increasedrate of tooth wear in some populations of domestic Ovisaries (Baker et al., 1959), and is strongly correlated withthe degree of hyposodonty in large-bodied African herbi-vores (Hummel et al., 2010). It has even been suggestedthat exogenous grit may have played a selective role inthe evolution of hominin postcanine megadonty (Cud-dahee et al., 2010).
Taphonomic artifacts
Taphonomic artifacts, including environmental ero-sion, etching, and/or abrasion, as well as damageinflicted through the mechanical and/or chemical prepa-ration of fossils, have the potential to confound micro-wear interpretations if they are not properly diagnosed(Teaford, 1988b, King et al., 1999). In particular, theymay severely limit available samples, especially wherefossil assemblages comprise surface collections from flu-vial channel sand deposits. Thus, recent microwear stud-ies found only 18 of 83 early Homo molars (Ungar et al.,2006b), only three Au. anamensis molars (Grine et al.,2006a), and only 23 molars (representing 19 individuals)of the total available sample of 176 specimens of Au.afarensis (Grine et al., 2006b) to be suitable for analysis.Artifacts have rendered some 78% of early Homo, 87% ofAu. afarensis, and 96% of Au. anamensis molars uselessfor microwear study.In this context, it is germane to note that high-magni-
fication (i.e., 3200 to 3500) examination resulted in therejection of a significant number of teeth deemed suita-ble for analysis by lower magnification binocular micros-copy. Thus, for example, all specimens of Au. afarensiswere examined first by binocular light microscopy atmagnifications of up to 350 to determine their suitabil-ity for further study by scanning electron microscopy(SEM), and this resulted in the selection of 37 teeth thatwere apparently artifact free. However, following highermagnification SEM scrutiny, a further 14 teeth wereeliminated from consideration because the occlusalsurfaces were observed to be taphonomically altered(Grine et al., 2006b). As such, high-magnification exami-nation served to reject nearly 40% of the specimens thatwere initially accepted by low-magnification observation.This attests to a significant drawback in the use of low-magnification studies of microwear.
Low-magnification observations
The SEM has approximately 3200 the resolvingpower, 3100 the maximal resolution, 3100 the maxi-mum working distance, and 310 the depth of field atany given magnification when compared with conven-tional optical light microscopy. Although high magnifica-tion (e.g., 3500) may be achieved with conventional lightmicroscopes, depth of field is lost as magnificationincreases such that binocular light microscopy at highmagnification is wholly impractical on anything but aflat surface. More recently, the use of SEM imaging hasbeen augmented by white light confocal microscopy thatdoes not suffer some of the limits of conventional opticalmicroscopy (Ungar et al., 2003). Moreover, the techni-ques of counting and measuring microwear variables(e.g., scratches and pits) employed previously have beenlargely supplanted by more objective texture analyses(Ungar et al., 2003; Scott et al., 2005).
Regardless of the mode of microscopy employed—be itSEM or white light confocal—the high-magnification ex-amination of tooth wear is time consuming, requiringhigh resolution specimen preparation (molding and cast-ing) and the use of specialized equipment. These appa-rent obstacles led Solounias and Semprebon (2002) topropose a return to the use of low-magnification (e.g.,335), binocular light microscopy, with wear featuresbeing classified and tallied by eye. A quantitative variantof this approach introduced by Merceron (2003) and Mer-ceron et al. (2004) involves the use of low-magnificationimaging by light microscopy and the measurement offeatures projected onto a computer screen. Fraser et al.(2009) have proposed that counting features on photo-graphs produced by ‘‘high dynamic range imaging’’improves the poor repeatability of other low magnifica-tion methods. Because it is fast and cheap, low-magnifi-cation light microscopy has gained considerable popular-ity in recent years. Its application has been extendedbeyond its initial application (the reasonably obvious dif-ferentiation of grazing and browsing ungulates) by Solo-unias and Semprebon (2002) to the examination ofextinct primates (Godfrey et al., 2004, 2005; Semprebonet al., 2004; Carter, 2006; Williams et al., 2006; Williamsand Patterson, 2010).Low-magnification studies typically use categories
such as ‘‘puncture pits,’’ ‘‘dentine lakes,’’ ‘‘fine scratch,’’and ‘‘hypercoarse scratch’’ to describe the featuresobserved, but even workers who are experienced in thismethod can have difficulty distinguishing some of these(e.g., Semprebon et al., 2004, 2005). Indeed, as noted byWalker (2007: 9) in a review of the low magnificationstudies by Godfrey et al. (2004, 2005) and Semprebon etal. (2004, 2005),
‘‘the credibility of their tooth wear methods has beenlessened because the authors could not distinguishbetween dentin exposures that would have occurredregardless of the foods eaten and features caused bytooth–food interactions.’’
Although problems relating to replication of resultsusing low magnification have been acknowledged (Scottet al., 2008; Fraser et al., 2009), no study to date hasreported either intraobserver- or interobserver-errorrates. This is a noteworthy omission in light of the errorrate studies that have been undertaken with referenceto high-magnification microwear studies (Grine et al.,2002). This reveals a significant, albeit unacknowledgedshortcoming in the method.Perhaps more importantly, given the problems associ-
ated with the use of light microscopy at high magnifica-tions, it is not possible to resolve the smaller (micronscale) microwear features that dominate wear facets,and these are the very features that are often key tofiner dietary distinctions and the differentiation of ante-mortem microwear from taphonomic artifacts (Teaford etal., 2008). In this context, it is most revealing that thenumbers of specimens that comprise low-magnificationsamples greatly exceed those deemed suitable for inclu-sion in high-magnification analyses.Thus, in the low-magnification studies of fossil papio-
nins from the South African Plio-Pleistocene sites, Wil-liams et al. (2006) included 22 specimens of Papio robin-soni from Swartkrans Member 1 and 18 specimens ofPapio angusticeps from Cooper’s and Kromdraai A,Carter (2006) used 17 specimens of Papio robinsonifrom Swartkrans Member 1 and 12 specimens of Papio
289MICROWEAR, ISOTOPES, AND HOMININ PALEOECOLOGY
American Journal of Physical Anthropology

angusticeps from Cooper’s and Kromdraai A, whileWilliams and Patterson (2010) employed 8 specimens ofParapapio antiquus, 12 of Papio izodi, and 10 of indeter-minate taxonomic attribution from Taung. Their samplefrom Sterkfontein Member 4 included 10 specimens ofParapapio broomi and 5 of Parapapio jonesi. In contrast,the SEM-based study by El-Zaatari et al. (2005: 186)noted that even the initial examination by light micros-copy ‘‘revealed the specimens from Taung, Bolt’s Farmand Coopers to be too poorly preserved for microwearanalysis, and served to exclude many specimens fromthe other sites as well.’’ Indeed, El-Zaatari et al. (2005)excluded some 120 (70%) of the total available sample of170 cercopithecoid fossils from consideration because oftaphonomic artifacts, finding only six specimens of Papiorobinsoni from Swartkrans Member 1 and no specimenof Papio angusticeps to be suitably preserved for analy-sis. It is especially noteworthy that the low-magnifica-tion studies by Williams et al. (2006), Carter (2006), andWilliams and Patterson (2010) included virtually all ofthe available specimens of Papio robinsoni and Papioangusticeps, as well as virtually all of the fossil monkeysfrom Taung as preserving ‘‘pristine’’ antemortemmicrowear. These figures attest to the inability of lowmagnification studies to adequately distinguish actualdiet-related microwear from postmortem taphonomicartifacts.
Buccal microwear
Another approach to diet reconstruction throughmicrowear is the use of wear features on the buccalsurfaces of the molar teeth (Perez-Perez, 2004). Althoughsome studies have reported differences in the frequen-cies, lengths and orientations of striations on thesesurfaces in extant primates that inhabit different envi-ronments and have different diets (Perez-Perez et al.,1994; Galbany and Perez-Perez, 2004; Galbany et al.,2009), the majority of references pertain only to fossils(Lalueza Fox and Perez-Perez, 1993; Lalueza Fox et al.,1996; Perez-Perez et al., 1999, 2003; Estebaranz et al.,2009).However, neither the buccal nor lingual sides of the
molars are employed in food processing, and it has neverbeen made clear why they should preserve wear featuresrelating to food items that are processed occlusally.Rather, it has been merely a matter of belief that as fooditems are masticated, ‘‘the abrasive particles scratch thebuccal enamel surface, where pits are rarely formed andscratches are the only relevant feature’’ (Perez-Perez etal., 1994). The overriding question that has yet to beaddressed is how sufficient force is applied to foods onthe buccal side of a molar to scratch enamel? Hopefully,in vivo investigations instigated by Romero et al. (2009)will begin to address this question.At the same time, however, if buccal microwear is
related to diet, one might reasonably expect that mam-mals that use cheek pouches in food procurement andprocessing should display significantly higher incidencesof features on the buccal surfaces of their molars. How-ever, Ungar and Teaford (1996) found that while cercopi-thecines (which possess cheek pouches that open oppo-site the M2) exhibit somewhat higher incidences of M2
buccal microwear than do colobines and cebids (whichlack cheek pouches), cercopithecines also exhibit higherincidences of lingual surface microwear. The buccal andlingual microwear incidences observed by Ungar and
Teaford (1996) most likely relate to differences in the fre-quency of terrestrial feeding events. As such, the degreeto which a primate feeds on the ground, with the attend-ant opportunity to ingest exogenous grit may relate tothe incidences of nonocclusal microwear. The presence ofmicrowear traces on the buccal and lingual surfaces ofmolars also serves to raise questions about the force thatis actually required to scratch tooth enamel. While it isdifficult to imagine either the tongue or cheek musclesgenerating the force (3,700 Mpa) that has been sug-gested as being required to scratch tooth enamel(Waters, 1980), some chemomechanical environmentsshould render tooth enamel more easily indented thanothers (Cheng et al., 2009). As such, under the right con-ditions, small siliceous bodies might scratch the buccaland lingual surfaces of teeth.It has been argued that, because of its unique forma-
tion dynamics, buccal microwear takes longer to form(and turnover) than occlusal microwear and that ittherefore overcomes the ‘‘Last Supper Effect’’ (Romeroand De Juan, 2007). However, more work must be doneto demonstrate that buccal microwear has any meaning-ful relationship to supper, and that it can be employed toinfer diet in extinct taxa.
STABLE ISOTOPES, TRACE ELEMENTS,AND DIET
Stable light isotope chemistry and trace element anal-ysis are well-established avenues of paleodietaryresearch that have been applied to extinct hominin taxa(Lee-Thorp and Sponheimer, 2006). Two main avenueshave been explored, namely alkaline earth metal concen-trations—especially strontium (Sr) and barium (Ba) —and the stable light isotope ratios of carbon (13C/12C)and nitrogen (15N/14N). All are based upon the notionthat these concentrations reflect dietary composition (theidea that ‘‘you are what you eat’’). However, becausetrace elements and stable isotopes are based on differentbiogeochemical principles they are essentially independ-ent systems. Each approach has its own advantages andits own limitations. In both, constraints are imposed bythe limits of the systems themselves, and our under-standing of them.Because these approaches are based on the chemical
composition of fossilized hard tissues, one common andimportant limitation concerns their diagenetic alteration.The temporal and preservation limits of some hard tis-sue components are closely related issues and, in partic-ular, the main organic compartment of hard tissues, col-lagen, has limited longevity because it is sensitive tomoisture, temperature, and pH, and it dissolves awayrelatively quickly on geological time scales. This limitsapplication of collagen-based carbon and nitrogen iso-topes to the recent past. In contrast, the mineral compo-nent of calcified tissues (bioapatite) can survive far lon-ger, but at the inevitable expense of some degree of post-mortem chemical alteration. Diagenesis is an especiallysevere problem for methods that are reliant on bone ordentine, because they contain poorly crystalline and,therefore, very reactive bioapatites. The problem can belargely, but not completely avoided by using toothenamel, which is a far denser and more crystalline (Lee-Thorp and van der Merwe, 1987; Wang and Cerling,1994; Lee-Thorp, 2002). However, enamel, dentine, andbone are not simply interchangeable; enamel reflects
290 F.E. GRINE ET AL.
American Journal of Physical Anthropology

diet in earlier phases of life, whereas bone variablyreflects earlier and later diets as a result of remodeling.Despite the potential significance of diagenesis, there
are few universally accepted methods that can be usedto demonstrate good preservation. Indeed, the relation-ship between standard indicators of preservation, suchas the infrared splitting factor (a proxy for crystallinity),and the actual state of preservation of the chemical orisotopic dietary indicators is problematic (Trueman etal., 2008). Thus, the demonstration by one or anothermethod that diagenesis has occurred does not necessarilymean that the isotopic composition has been altered.In addition, there are intrinsic constraints to the
amount of information that can be extracted from traceelements and stable light isotopes. Thus, a particularfood source can yield different results depending uponthe environment from which it derives, while differenttypes of foods can have the same appearance insofar astheir trace element and isotopic signatures are con-cerned. For instance, although trace elements may beable to speak to trophic levels given ideal circumstances,the consumption of leaves, or the consumption of ani-mals that have eaten leaves cannot be readily distin-guished using carbon isotope analysis alone. Mostly,however, we are limited by an inadequate understandingof the behavior of trace elements in natural ecosystems,and of stable isotopes in physiological systems (but seeBurton et al., 1999). This is a more significant problemin the case of trace elements, which can vary stronglyboth between and within environments, whereas stableisotopes follow universal principles even though theymay be complex and under different physiological con-straints. Nevertheless, while knowledge of environmen-tal parameters and diet may predict trace element or iso-tope ratios in hominin fossils, the reverse is not neces-sarily the case (Burton and Wright, 1995; Sponheimer etal., 2007).
Trace elements
Alkaline earth metals (Sr and Ba). There is a longhistory of using alkaline earth metal concentrations toinvestigate paleodiet (Toots and Voorhies, 1965; Schoe-ninger, 1979; Sillen, 1981, 1992; Sillen and Kavanagh,1982; Sillen et al., 1995; Safont et al., 1998; Balter et al.,2002; Palmqvist et al., 2003). Adult mammalian diges-tive and renal physiology serves to discriminate againststrontium (Sr) and barium (Ba) with respect to calcium(Ca) in the biopurification of Ca (Lough et al., 1963;Spencer et al., 1973). As a result, herbivore tissues havelower Sr/Ca and Ba/Ca ratios than the plants that theyeat and carnivores have lower Ba/Ca and Sr/Ca ratiosthan the herbivores they consume (Sealy and Sillen,1988; Burton et al., 1999). Because Sr and Ba substitutefor Ca in the calcium phosphate apatite structure ofbone, dentin, and enamel, they can in principle be usedto investigate trophic behavior of extinct animals. Othertrace elements (e.g., zinc) have been used as potential di-etary indicators on occasion, but applications areseverely limited because so little is known about theirdistribution in food webs and their fixation in calcifiedtissues (Ezzo, 1994).As noted above, diagenesis presents a major constraint
in the application of Sr and Ba to paleodietary recon-struction (Boaz and Hampel, 1978; Sillen, 1989),although early researchers were largely unaware of theextent of the problem presented by postmortem chemicalalteration (Toots and Voorhies, 1965; Wyckhoff and
Doberenz, 1968; Brown, 1974). This is particularly prob-lematic with regard to bone and dentin, which are sus-ceptible to chemical alteration that can quickly obliteratethe biological Sr/Ca signal (Tuross et al., 1989). Attemptsto overcome this problem have been devised (e.g., the‘‘solubility profiling’’ technique of Sillen, 1981, 1992), buteven when it is applied, diagenetic strontium often can-not be eradicated (Hoppe et al., 2003; Trickett et al.,2003).Boaz and Hampel (1978) were the first to employ bio-
geochemical methods to investigate the Sr concentra-tions of fossils attributed to Paranthropus and Homofrom the Shungura Formation, southern Ethiopia. Sillenet al. (1995) applied his solubility profiling method toinvestigate the Sr/Ca ratios of P. robustus and earlyHomo bones from the site of Swartkrans, South Africa.Unfortunately, these trace element studies are limitedby the fact that Sr/Ca ratios are highly variable inplants at the base of the food web, and this seriouslycomplicates interpretation of Sr/Ca data in calcified tis-sues (e.g., Bowen and Dymond, 1955; Runia, 1987; Bur-ton et al., 1999). Another problem for Sillen’s (1992;Sillen et al., 1995) study is that it was carried out onbone, which, as noted above, is particularly vulnerable todiagenetic overprinting that cannot be eradicated evenwith the application of the ‘‘solubility profiling’’ tech-nique (Hoppe et al., 2003; Trickett et al., 2003).A recent study by Sponheimer and Lee-Thorp (2006)
was unable to detect evidence for diagenesis in impor-tant trace elements in tooth enamel from several aus-tralopith-bearing sites in South Africa (with the excep-tion of raised levels of manganese [Mn] at the Maka-pansgat Limeworks). Tooth enamel of P. robustus andAu. africanus, which was found to have higher Sr/Caratios than that of contemporaneous baboons and manyassociated herbivores, offers little in the way of evidencefor australopith omnivory (Sponheimer et al., 2005a).However, plotting two physiologically related trace ele-ment ratios (barium/calcium and strontium/barium) forthe Australopithecus teeth reveals distinctive (high) Sr/Ba, suggesting the possibility that they consumed itemswith unusually high Sr and relatively low Ba concentra-tions. Foods that could meet this requirement includegrass seeds and underground storage organs (USOs)(roots, rhizomes, and bulbs), although the evidence forthe latter is indirect (Sponheimer and Lee-Thorp, 2006).Insofar as trace element compositions appear to retainmuch of their fidelity in tooth enamel, studies investigat-ing elemental distribution in modern ecological foodcycles are urgently required.
Stable isotopes
The ratios of several stable isotopes have enjoyedwidespread use in archaeological and paleontologicalstudies. Among those most commonly employed are thenitrogen isotope (15N/14N) composition of bone, and thecarbon isotope (13C/12C) composition of tooth enamel.The determination of the carbon isotope composition ofenamel also generates data pertaining to oxygen isotope(18O/16O) composition, which can provide both ecologicaland climate information.Nitrogen is restricted to the organic phase of hard tis-
sue (i.e., bone collagen), and because this has a maxi-mum preservation limit of some 200 kya even under per-fect conditions (Jones et al., 2001), nitrogen isotope stud-ies are limited to more recent (i.e., Late Pleistocene and
291MICROWEAR, ISOTOPES, AND HOMININ PALEOECOLOGY
American Journal of Physical Anthropology

Holocene) hominin specimens. Its most significant appli-cation to hominin paleodietary research has been in thestudy of Neandertal remains, where relatively high15N/14N ratios suggest that they derived their dietaryprotein primarily from animal rather than plant foods(Bocherens et al., 1991, 1999; Richards et al., 2000; Rich-ards and Schmitz, 2008). Even though nitrogen isotopestudies of archaeological remains are common, the mech-anisms that control their compositions at differenttrophic levels in mammalian communities remain poorlyunderstood (Ambrose, 2000; Vanderklift and Ponsard,2003).Since the isotopes of nitrogen derived from collagen
are not preserved on the timescale of interest for thepurposes of the present review, we restrict the discussionbelow to carbon and oxygen isotopes in tooth enamel.
Carbon (d13C). One of the most widely utilized and per-haps best understood of the biogeochemical approachesto paleodietary reconstruction relies upon stable carbonisotopes. The underlying principle for stable carbon iso-tope analysis relates to the differences between plantsthat follow different photosynthetic pathways (C3, C4,and crassulacean acid metabolism [CAM]). In the C3
photosynthetic pathway, the initial products of CO2 fixa-tion are 3-carbon compounds that are processed throughthe Calvin–Benson cycle, while the C4 pathway initiallyforms 4-carbon compounds via the Hatch–Slack pathway.Both photosynthetic pathways discriminate against13CO2, but to different degrees (Smith and Epstein,1971), resulting in distinct 13C/12C ratios.In comparison to C4 plants (mainly tropical grasses
and many sedges in warm zones, and a restricted num-ber of arid- or saline-adapted shrubs), those that employthe C3 pathway (almost all trees, shrubs and herbs, andtemperate or shade-adapted grasses) are stronglydepleted in 13C relative to atmospheric CO2. Conse-quently, C3 plants have distinctly lower d13C 1 valuesthan C4 plants. Environmental influences acting on C3
plants include the ‘‘canopy effect’’ in dense forests, whichleads to even lower d13C values (van der Merwe and Me-dina, 1989), and aridity/temperature effects that lead toslightly higher d13C values under increased aridity and/or temperature (Tieszen, 1991).Of particular relevance to reconstructing early homi-
nin diets is the observation that in tropical African envi-ronments, virtually all trees, bushes, and forbs utilize C3
photosynthesis, while grasses and a significant numberof sedges use the C4 pathway. The carbon isotopes inthese plants are ultimately incorporated into the tissuesof the animals that consume them, with the result thatC4 grass consumers have very different carbon isotopecompositions than animals that eat C3 vegetation (Vogel,1978; Ambrose and DeNiro, 1986; Lee-Thorp and vander Merwe, 1987).A third photosynthetic pathway, CAM, is biochemically
similar to that employed by C4 plants except that enzy-matic activities are separated diurnally, with CO2 cap-ture occurring at night. CAM photosynthesis results in
d13C values that vary extensively depending upon thespecies of plant and environmental conditions (Winterand Smith, 1996). CAM plants are primarily succulentsand are rare beyond semidesert environments. Moreover,many tend to be toxic, and of little nutritional value. Assuch, CAM plants are rarely consumed by primates,although their occasional use by baboons has been docu-mented (Codron et al., 2006).The carbon isotopes that are incorporated into biologi-
cal apatites in the bones and teeth of an animal canthus be used as dietary proxies (Sullivan and Krueger,1981; Lee-Thorp and van der Merwe, 1987). Even thoughbone mineral persists well beyond bone collagen, it canbe altered postmortem resulting in the loss of the bio-genic dietary signal (Wang and Cerling, 1994; Lee-Thorpand Sponheimer, 2003). Tooth enamel apatite differsfrom that of bone and dentine, particularly in its verylow proportion of organic matrix (phosphoproteins andamelogenins) and its higher crystallinity (LeGeros,1991), with the result that it better retains its biogenicisotope composition.The partitioning, or fractionation of the isotopes of car-
bon (13C and 12C) during bioapatite formation, is a rea-sonably well-understood process (Passey et al., 2005).Some variability in fractionation has been documented(Lee-Thorp et al., 1989; Ambrose and Norr, 1993; Tieszenand Fagre, 1993; Cerling and Harris, 1999; Passey et al.,2005), and this likely reflects differences in metabolismand/or dietary physiology. Nevertheless, bioapatitereflects the d13C values of the bulk diet (both animal andvegetable foods), and not largely the protein componentas is the case with collagen (Lee-Thorp et al., 1989;Ambrose and Norr, 1993). Owing to the problems associ-ated with diagenesis, to which bone and dentin are espe-cially prone, only tooth enamel is sampled for stable iso-tope analysis of specimens that are millions of years old.Analyses of tooth enamel apatite of living species with
known diets have shown that d13C values retain the bio-genic isotope signal; this permits inference about thetypes of plants consumed, or at least the plants at thebase of the food web in paleontological contexts (Lee-Thorp et al., 1989; Cerling et al., 1999). Stable carbonisotope values have been reported for a variety of homi-nin samples ranging from the Pleistocene to the EarlyPliocene.
Oxygen (d18O). Oxygen isotope ratio (18O/16O) data canprovide both ecological and climate information. Deter-mination of d18O values has traditionally been used as apaleoclimate indicator because the primary influence onwater d18O values in the body is the isotopic compositionof available drinking water, which is subject to altitude,climate influences on precipitation, temperature, and rel-ative humidity (Darling et al., 2005). However, the iso-topic mass balance of an animal’s body water, and henceits enamel d18O, is also influenced by factors such as die-tary ecology, drinking behavior, and thermoregulation(Sponheimer and Lee-Thorp, 2001). These factors modu-late the environmental signals such that the distributionof d18O in a faunal assemblage may be strongly differen-tiated at a given time and place. For instance, 18O-enriched plant water and leaf carbohydrates due toevapotranspiration influence herbivore d18O, especiallyin browsers who obtain most of their water from plants(Levin et al., 2006). Trophic behavior may also be impli-cated, especially in species whose diets are high in lipidand protein content. Thus, faunivores such as Otocyon
1By convention, stable light isotope ratios are expressed as d val-ues relative to an international standard in parts per thousand (permil). Thus, for carbon isotopes: d13C (o/oo) 5 (13C/12Csam-
ple/13C/12Cstandard 2 1) 3 1,000, and the international standard is
Vienna Peedee Belemnite (VPDB). VPDB is the international stand-ard for both carbon (13C/12C) and oxygen (18O/16O) isotopes in carbo-nates. For d18O in water, the international standard is StandardMean Ocean Water (SMOW).
292 F.E. GRINE ET AL.
American Journal of Physical Anthropology

megalotis, Crocuta crocuta, and Orycteropus afer aredepleted in 18O compared to herbivores in modern eco-systems (Lee-Thorp and Sponheimer, 2005). Suids andprimates may have relatively low d18O values, but thismay be of uncertain and multiple causes (Sponheimerand Lee-Thorp, 2001). Moreover, the demonstration ofsignificant differences between non-obligate drinkers(e.g., giraffe) versus obligate drinkers (e.g., reduncinesand hippopotamus) has been suggested as an index ofaridity (Levin et al., 2006).Unfortunately, comparison of data from different sites
is hampered by the geographic specificity of local waterd18O. Additionally, interpretation of the distribution ofd18O values amongst many fauna is still insecure, andour understanding of d18O patterning in food websremains very limited.
DENTAL MICROWEAR, STABLE ISOTOPES,AND EXTINCT HOMININ DIETS
Here we provide a review of the evidence from dentalmicrowear and/or stable isotopes that bears on the die-tary habits of six early hominin species (Table 1). Foreach, the geological, faunal, and/or paleosol indicatorsrelating to paleohabitat or paleoenvironmental context(s)are reviewed in order to place the dietary evidence inperspective. In addition, the morphological attributesthat have been invoked to infer the dietary habits oradaptations of each species will be reviewed with a viewto highlighting discrepancies that may relate to speciesadaptations rather than to actual dietary habits.
Ardipithecus ramidus
This early Pliocene species is represented by fossilsrecovered from localities at Aramis in the Middle Awash,Ethiopia, and from sites some 70 km to the north at AsDuma, in the Gona western escarpment of the EthiopianRift (White et al., 1994, 1995, 2009b; Semaw et al.,2005). The former derive from the Lower Aramis Mem-ber of the Sagantole Formation, which is sandwiched bytwo volcanic tuffs of identical age (4.42 Ma) (WoldeGab-riel et al., 1994, 2009; Renne et al., 1999; White et al.,2006). The As Duma specimens are somewhat moreloosely constrained, but appear to date to between 4.51–4.32 Ma, although a younger age (4.2–3.6 Ma) cannot beruled out for some of the fossils (Semaw et al., 2005).Three fragmentary fossils from sites in Kenya (Loth-agam, Tabarin, and Baringo) that are likely penecontem-poraneous with those from Aramis and As Duma arepossibly attributable to Ar. ramidus (White et al., 1994).
Paleoecological context. The Aramis fossils derivefrom silts and clays deposited on a floodplain, and fossil-ized wood, seeds, and phytoliths confirm the presence ofhackberry, fig, and palm trees (WoldeGabriel et al.,2009). The associated fauna is dominated by kudu (Trag-elaphus) and colobine monkeys, and the presence of Can-thium seeds is compatible with the presence of a gallerywoodland/forest (White et al., 2009a,b; WoldeGabriel etal., 2009). There is no evidence of a humid closed-canopytropical rainforest. All told, the associated vertebrateand invertebrate faunal remains, the floral remains, andthe soil carbonate d13C and d18O values suggest a wood-land biotope in the Aramis localities that contain Ar.ramidus (Louchart et al., 2009; WoldeGabriel et al.,2009).
Cerling et al. (2010), however, have noted that thedata presented by WoldeGabriel et al. (2009) on the car-bon isotopic composition of the paleosols and pedogeniccarbonates from Aramis indicate a greater than 40% C4
biomass. This corresponds to woody canopy cover of 5–25%, implying the predominance of tree-bush savanna,arid shrub savanna, edaphic grassland, bush savanna,or open savanna types of vegetation. They also observedthat the water deficit calculated by White et al. (2009a)is compatible with a riparian forest or woodland in anotherwise open bushland or woody grassland. Moreover,Cerling et al. (2010) have observed that the d13C datafrom bovid tooth enamel presented by White et al.(2009a) are also consistent with browsing herbivoresinhabiting environments ranging from woodland to sa-vanna (Cerling et al., 2010). Thus, Cerling et al. (2010)envision the data for Aramis as being consistent withthe presence of relatively open savanna grasslands to-gether with riparian woodlands and forest corridors.The depositional environment of the Ardipithecus fos-
sils at Gona differs somewhat from that at Aramis; atGona there are lake deposits interfingered with smallfluvial channels. Semaw et al. (2005) conclude that thecarbonate-rich paleosols at As Duma formed in a subhu-mid and seasonally dry climate in a landscape coveredby C3-dominated woodlands and grassy woodlands. Assuch, the Aramis setting is envisioned as having beensubstantially drier than at Gona (White et al., 2009a).
Morphological inferences about diet. Ardipithecuspossessed a dentition suggestive of omnivory and frugi-vory (Suwa et al., 2009). In particular, its comparativelysmall and thinly enameled postcanine teeth are seen aslacking the adaptations for heavy chewing and the con-sumption of abrasive foods. The postcranial skeletalremains of Ar. ramidus, especially elements of the fore-limb and foot, are consistent with significant arborealclimbing (Lovejoy et al., 2009a,b), and as such are alsoconsistent with the conclusion of Suwa et al. (2009) con-cerning ‘‘generalized omnivory and frugivory.’’ Thesemorphological attributes and inferences are not in dis-agreement with the isotopic values and microwear sig-nals obtained for Ar. ramidus tooth enamel.
Dental microwear and stable isotopes. With regardto molar occlusal microwear in Ardipithecus, Suwa et al.(2009: 97) examined six teeth from four individuals bySEM, and observed that they ‘‘tend to exhibit finer andmore randomly oriented striae’’ than the molars of Au.afarensis. Unfortunately, Suwa et al. (2009) provide nodata by which the pattern evinced by Ardipithecus canbe assessed in a comparative context. Moreover, theircomparison with Au. afarensis is based on observationsof eight molars from Hadar rather than with the datafor the larger sample presented by Grine et al. (2006b).Nevertheless, it would appear that the microwear of Ar.ramidus molars does not indicate the inclusion of hard,brittle objects in the diets of those individuals examined.Carbon and oxygen isotope data have been recorded
from tooth enamel for five specimens of Ar. ramidusfrom Aramis (White et al., 2009a) (Table 2). The d13Cvalues (mean 5 210.22, SD 5 1.02, n 5 5) fall withinthe ranges for C3 browsers (e.g., tragelaphines and neo-tragines) from Aramis, and White et al. (2009a) sug-gested that Ar. ramidus ‘‘predominantly consumed (ca.,85–90%) C3 plant sources.’’ White et al. (2009a: text S3,p. S9) posited that Ardipithecus consumed ‘‘slightlymore’’ 13C-enriched plants (and/or animals that fed on
293MICROWEAR, ISOTOPES, AND HOMININ PALEOECOLOGY
American Journal of Physical Anthropology

TABLE 2. Carbon isotope data recorded for individual for Plio-Pleistocene hominin specimens
Specimen d13C Stratigraphic derivation Tooth Source
Ardipithecus ramidusARA-VP-1 700 28.5 Sagantole Fm., Lower Aramis Mb. M2 White et al. (2009b)ARA-VP-1 1818 210.7 Sagantole Fm., Lower Aramis Mb. ? White et al. (2009b)ARA-VP-1 3291 211.2 Sagantole Fm., Lower Aramis Mb. M1 White et al. (2009b)ARA-VP-1 3290 210.3 Sagantole Fm., Lower Aramis Mb. M2 or 3? White et al. (2009b)ARA-VP-6 500a 210.4 Sagantole Fm., Lower Aramis Mb. M1/2/3 White et al. (2009b)Australopithecus africanusMLD 12 27.7 Makapansgat, Member 3 M3 Sponheimer and Lee-Thorp (1999a)MLD 28 28.1 Makapansgat, Member 3 M2 Sponheimer and Lee-Thorp (1999a)MLD 30 25.6 Makapansgat, Member 3 M1 Sponheimer and Lee-Thorp (1999a)MLD 41 211.3 Makapansgat, Member 3 M? Sponheimer and Lee-Thorp (1999a)Sts 31 26.8 Sterkfontein, Member 4 M3 Sponheimer et al. (2005b)Sts 32 27.8 Sterkfontein, Member 4 M3 Sponheimer et al. (2005b)Sts 45 24.0 Sterkfontein, Member 4 M2 Sponheimer et al. (2005b)Sts 72 29.7 Sterkfontein, Member 4 M3 Sponheimer et al. (2005b)Sts 2218 25.9 Sterkfontein, Member 4 M? Sponheimer et al. (2005b)Sts 2253b 26.7 Sterkfontein, Member 4 M1 Lee-Thorp et al. (2010)Sts 2518b 210.0 Sterkfontein, Member 4 M3 Lee-Thorp et al. (2010)Stw 14 26.7 Sterkfontein, Member 4 M1 van der Merwe et al. (2003)Stw 73 28.8 Sterkfontein, Member 4 M2 van der Merwe et al. (2003)Stw 207 22.0 Sterkfontein, Member 4 ? van der Merwe et al. (2003)Stw 211 27.3 Sterkfontein, Member 4 M? van der Merwe et al. (2003)Stw 213i 21.8 Sterkfontein, Member 4 M1 van der Merwe et al. (2003)Stw 229 25.8 Sterkfontein, Member 4 P? van der Merwe et al. (2003)Stw 236 23.7 Sterkfontein, Member 4 P? van der Merwe et al. (2003)Stw 252 27.4 Sterkfontein, Member 4 M1 van der Merwe et al. (2003)Stw 276 28.0 Sterkfontein, Member 4 M1 van der Merwe et al. (2003)Stw 303 24.3 Sterkfontein, Member 4 M2 van der Merwe et al. (2003)Stw 304 27.4 Sterkfontein, Member 4 M? van der Merwe et al. (2003)Stw 309b 26.1 Sterkfontein, Member 4 M1 van der Merwe et al. (2003)Stw 315 25.7 Sterkfontein, Member 4 dm2 van der Merwe et al. (2003)Paranthropus robustusTM 1600 27.9 Kromdraai B, Member 3 M2 Sponheimer et al. (2005b)SK 19 26.3 Swartkrans, Member 1 M3 Sponheimer et al. (2005b)SK 41 26.7 Swartkrans, Member 1 M3 Sponheimer et al. (2005b)SK 57 26.5 Swartkrans, Member 1 M3 Sponheimer et al. (2005b)SK 876 26.7 Swartkrans, Member 1 M? Lee-Thorp et al. (2000)SK 878 26.8 Swartkrans, Member 1 P3 Lee-Thorp et al. (1994)SK 879(a)c 28.5 Swartkrans, Member 1 M1 Lee-Thorp et al. (1994)SK 879(b)c 28.1 Swartkrans, Member 1 M? Lee-Thorp et al. (1994)SK 1512 28.8 Swartkrans, Member 1 P? Lee-Thorp et al. (1994)SK 14000 25.9 Swartkrans, Member 1 M3 Sponheimer et al. (2005b)SK 14132 26.9 Swartkrans, Member 1 M3 Sponheimer et al. (2005b)SK 24605b 26.8 Swartkrans, Member 1 M3 Sponheimer et al. (2006b)SK 24606b 25.6 Swartkrans, Member 1 M2 Sponheimer et al. (2006b)SKW 6 27.0 Swartkrans, Member 1 M3 Sponheimer et al. (2005b)SKW 3068 28.1 Swartkrans, Member 1 M2 Sponheimer et al. (2005b)SKW 4768 27.4 Swartkrans, Member 1 M2 Sponheimer et al. (2005b)SKW 6427b 28.1 Swartkrans, Member 1 M? Sponheimer et al. (2006b)SKX 5015 29.6 Swartkrans, Member 1 M3 Lee-Thorp et al. (1994)SKX 5939b 24.9 Swartkrans, Member 1 M? Sponheimer et al. (2005b)SKX 333 210.0 Swartkrans, Member 2 M1 Lee-Thorp et al. (1994)SKX 1312 28.1 Swartkrans, Member 2 M1 Lee-Thorp et al. (1994)SKX 35025 27.9 Swartkrans, Member 3 M? Lee-Thorp et al. (1994)Paranthropus boiseiOH 5 21.2 Olduvai Gorge, Bed I M2 van der Merwe et al. (2008)NMT-W64–160 20.7 Peninj, Humbu Formation M2 van der Merwe et al. (2008)KNM-CH 302 21.3 Chesowanja, Chemoigut Formation M? Cerling et al. (2011)KNM-ER 1171 20.6 Koobi Fora Fm., upper Burgi Mb. M1 Cerling et al. (2011)KNM-ER 1469 22.3 Koobi Fora Fm., upper Burgi Mb. M3 Cerling et al. (2011)KNM-ER 732 20.1 Koobi Fora Fm., KBS Member P4 Cerling et al. (2011)KNM-ER 802d 21.0 Koobi Fora Fm., KBS Member M1/M3 Cerling et al. (2011)KNM-ER 810 23.4 Koobi Fora Fm., KBS Member P3 Cerling et al. (2011)KNM-ER 816 21.9 Koobi Fora Fm., KBS Member P4 Cerling et al. (2011)KNM-ER 1479 22.3 Koobi Fora Fm., KBS Member M3 Cerling et al. (2011)KNM-ER 1804 21.2 Koobi Fora Fm., KBS Member M3 Cerling et al. (2011)KNM-ER 1806 21.3 Koobi Fora Fm., KBS Member M3 Cerling et al. (2011)KNM-ER 3737 21.6 Koobi Fora Fm., KBS Member M1 Cerling et al. (2011)KNM-ER 3952 21.3 Koobi Fora Fm., KBS Member M3 Cerling et al. (2011)KNM-ER 13750 0.2 Koobi Fora Fm., KBS Member M? Cerling et al. (2011)
294 F.E. GRINE ET AL.
American Journal of Physical Anthropology

such plants) than modern savanna-woodland chimpan-zees. Unlike chimpanzees, however, the d13C values sug-gest that most Ar. ramidus individuals likely consumedat least small quantities of C4 foods. White et al. (2009a)conclude that the low d13C values of four of the five Ar.ramidus individuals from Aramis indicate a diet verydifferent from those of later Plio-Pleistocene hominins,which included substantial quantities of C4 plant foods.The carbon isotope data, coupled with the paleoenvir-
onmental indicators for Aramis (White et al., 2009a;Cerling et al., 2010) suggest that Ar. ramidus had astrong preference for C3 vegetation such as fruits andleaves rather than the possibly abundant C4 resourcesnearby.In addition, the d18O values of the Ar. ramidus speci-
mens are comparable to those of the other Aramis mam-mal fauna (White et al., 2009a). On the other hand, lateraustralopith d18O values in both East and South Africa arelow compared to other fauna (with the exception of carni-vores) (Sponheimer and Lee-Thorp, 1999b; Lee-Thorp etal., 2010; Cerling et al., 2011). However, the causes of therelatively low australopith values remain obscure—theymay be related to water dependence, to frugivory, the useof USOs, or some combination thereof. Our understandingof d18O patterning in the food cycles of ecological commun-ities is, unfortunately, rather limited.
Australopithecus anamensis
Australopithecus anamensis is represented principallyby fossils (mostly isolated teeth and jaw fragments) fromKanapoi and Allia Bay, Kenya (Coffing et al., 1994; Lea-key et al., 1995, 1998). Additional specimens (mostly iso-lated teeth) attributed to Au. anamensis have beenrecovered from Asa Issie, Hana Hari, and Aramis (local-ity 14) in Ethiopia (White et al., 2006). The homininremains from Kanapoi and Allia Bay span a period ofsome 300,000 years, ranging between about 4.2 and 3.9Ma (Leakey et al., 1995, 1998), and those from Asa Issiehave been estimated to be 4.1–4.2 Ma-old on the basis offaunal correlations with the Kenyan sites (White et al.,2006).
Paleoecological context. The bulk of the Kanapoi fos-sils derive from paleosols that formed in an arid to semi-
arid climate with seasonal moisture, and in an environ-ment dominated by a tree-shrub savanna (Wynn, 2000).Accordingly, Wynn (2000) envisions Au. anamensis ashaving inhabited ‘‘a spatially variable ecosystem charac-terized by a mosaic of environments.’’ The paleosols andassociated fauna at Asa Issie reflect humid, grassy,woodland savannah environments, suggesting that thehominins were ‘‘closely and regularly associated with anarrow range of habitats varying from closed to grassywoodlands’’ (White et al., 2006: 885). The fossiliferouslocalities at Allia Bay resulted from the flooding of alarge, meandering proto-Omo River that would have pro-vided a variety of habitats ranging from gallery forest tofloodplain grasslands and swamps (Feibel et al., 1991), areconstruction consistent with the mammalian faunaand the abundance of lungfish (Protopterus) bones.Macho et al. (2003) have argued that there is evidencefor seasonality at Allia Bay from the distribution of‘‘stress lines’’ in the tooth enamel of large-bodied mam-mals. Stable isotope data on soil organics and carbonatesin the Lonyumun Member indicate that 60–80% of thevegetation consisted of C4 grasses, with trees and shrubsconstituting the remaining cover (Cerling et al., 1988).The carbon and oxygen stable isotopes derived frommammalian herbivore tooth enamel are consistent withthe faunal evidence insofar as they suggest the site tohave been better-watered than today, with the presenceof more extensive woodlands (Schoeninger et al., 2003).Schoeninger et al. (2003) have opined that ‘‘such a set-ting matches expectations for the selective advantages ofnut-eating, bipedal hominids.’’
Morphological inferences about diet. Ward et al.(1999, 2001) suggested that Au. anamensis dental mor-phology indicates a ‘‘dietary shift to harder foods’’ thanthose envisaged for the earlier Ar. ramidus. Teaford andUngar (2000) have suggested that its thickly enameledmolars might have rendered it ‘‘the first hominid to beable to effectively withstand the functional demands ofhard and perhaps abrasive objects in its diet.’’ Machoet al. (2005: 310) argued that decussation of this toothenamel, together with its gross dental morphology indi-cate an ‘‘adaptation for habitually consuming a hard-tough diet,’’ and White et al. (2006: 888) have opinedthat its dentognathic morphology indicates ‘‘an adaptive
TABLE 2. (Continued)
Specimen d13C Stratigraphic derivation Tooth Source
KNM-ER 15940 21.1 Koobi Fora Fm., KBS Member M3 Cerling et al. (2011)KNM-WT 37100 21.8 Nachukui Fm., Kaitio Member M2 or 3? Cerling et al. (2011)KNM-WT 37748 22.1 Nachukui Fm., Kaitio Member M3 Cerling et al. (2011)KNM-WT 17396 21.9 Nachukui Fm., Natoo Member M3 Cerling et al. (2011)KNM-ER 729 0.0 Koobi Fora Fm., Okote Member P4 Cerling et al. (2011)KNM-ER 733d 21.0 Koobi Fora Fm., Okote Member P4/M3 Cerling et al. (2011)KNM-ER 818 0.7 Koobi Fora Fm., Okote Member M3 Cerling et al. (2011)KNM-ER 3887 21.6 Koobi Fora Fm., Okote Member M3 Cerling et al. (2011)KNM-ER 6080 22.2 Koobi Fora Fm., Okote Member M2 Cerling et al. (2011)
The d13C values are % where d13C 5 (13C/12Csample/13C/12Cstandard 2 1) 3 1,000, and the international standard is Vienna Peedee
Belemnite (VPDB). Specimens within each species are arranged roughly in ascending chronological sequence, and in numericalorder within that sequence.a Three teeth (M1, M2 and M3; specimen numbers 90, 113 and 115) were sampled for this individual. The three values rangebetween 210.2 and 210.8 (White et al., 2009a; SI: 15). The value recorded here is the average of the three.b Indicates carbon isotope values generated using laser ablation. These have been adjusted following Passey and Cerling (2006) forcomparison with acid hydrolysis values.c Lee-Thorp et al. (1994: 365) record that two separate tooth fragments, both designated SK 879, were analyzed separately ‘‘since their col-our and appearance suggested . . .probably different origins, although this is not certain.’’ These are designated here as SK 879(a) and (b).d Represented by two teeth (M1 and M3) from the same individual. The value recorded here is the average.? indicates that the tooth is unknown; P? indicates an unknown premolar position; M? indicates an unknown molar position.
295MICROWEAR, ISOTOPES, AND HOMININ PALEOECOLOGY
American Journal of Physical Anthropology

shift towards the exploitation of tougher and more abra-sive food resources.’’ However, whereas thickly enam-eled, low-cusped molars would have difficulty processingtough foods, they would easily break down hard, brittleitems (Lucas, 2004).
Dental microwear and stable isotopes. There hasbeen no stable carbon isotope study to date of toothenamel of Au. anamensis. The foregoing discussionmight predict d13C values higher than those reported byWhite et al. (2009a) for Ardipithecus.Occlusal microwear data have been obtained for only
three molars of Au. anamensis representing 4.2% of thetotal worn albeit unfragmented molar sample fromKanapoi and Allia Bay (Grine et al., 2006a; Ungar et al.,2010). The observed microwear fabrics are unexpected inlight of the morphological and paleoecological back-ground discussed above. The SEM-derived occlusalmicrowear data (Table 3) suggest chimpanzees and goril-las as the best modern analogues for dietary preferencein Au. anamensis. The confocal-derived microwear tex-ture data for Au. anamensis (Table 4) are most similarto those of Theropithecus gelada, Alouatta palliata, andTrachypithecus cristatus with respect to the level of com-plexity (Table 5). However, texture anisotropy is lower,suggesting that these three individuals of Au. anamensismay not have had a diet dominated by tough foods.Significantly, there is no evidence for the consumption
of hard, brittle objects. While hypogeous tubers, bulbs,and roots that are taken by an eclectic and opportunisticfeeder like the chacma baboon may have constitutedpart of the dietary repertoire of Au. anamensis, there isno microwear evidence for hard, brittle items (Grineet al., 2006a). Although the microwear texture data forAu. anamensis cannot falsify the notion of rare or fall-back hard-object feeding, it provides no evidence for it(Ungar et al., 2010). The craniodental morphology of Au.anamensis may represent an adaptation for the occa-sional consumption of hard, brittle fallback foods, butthe microwear is consistent with the consumption ofsofter items—at least immediately prior to the deaths ofthe three individuals that have been examined thus far.
Australopithecus afarensis
Australopithecus afarensis is represented principallyby fossils from the Laetoli Beds, Tanzania (Harrison,
2011b and references therein) and the Hadar Formation,Ethiopia (Kimbel and Delezene, 2009 and referencestherein). Additional specimens are known from Dikika(Alemseged et al., 2005, 2006), Maka (White et al.,2000), and Woranso-Mille (Haile-Selassie et al., 2010a,2010b). One skeleton from Woranso-Mille (KSD-VP-1/1)has been attributed to Au. afarensis (Haile-Selassie etal., 2010a), while the bulk of dental remains have beeninterpreted as being intermediate between Au. anamen-sis and Au. afarensis (Haile-Selassie et al., 2010b).A number of isolated teeth that likely represent Au.
afarensis have been described from the Usno and Shun-gura Formations, Ethiopia (Leonard and Hegmon, 1987;Suwa et al., 1996; Hlusko, 2004). A partial cranium(KNM-ER 2602) from the Tulu Bor Member of the KoobiFora Formation, Kenya has been attributed to Au. afar-ensis (Kimbel, 1988), as have two fragmentary mandi-bles (KNM-WT 8556 and KNM-WT 16006) from thelower part of the Lomekwi Member of the Nachukui For-mation (Brown et al., 2001). The jaws were subsequentlyassigned to Kenyanthropus platyops (Leakey et al.,2001), but it has been argued that this taxon is definedon questionable grounds (White, 2003). All but one ofthe Laetoli hominin fossils are constrained between twohorizons dated to 3.85 Ma and 3.63 Ma (Deino, 2011),and the Hadar assemblage is bracketed between hori-zons dated to 3.40 Ma and 2.94 Ma (Kimbel and Dele-zene, 2009 and references therein). The hominin-bearingexposures at Dikika, Maka, and Woranso-Mille fallwithin the age range for the Laetoli—Hadar assemblages(Deino et al., 2010).Just as an ancestor–descendant relationship between
Ar. ramidus and Au. anamensis has been surmised(Ward et al., 1999; White, 2002; Haile-Selassie et al.,2004), so too it has been hypothesized that Au. anamen-sis was ancestral to Au. afarensis (Kimbel et al., 2006).While the former purported relationship is wholly conjec-tural, the latter is at least consistent with the results ofnumerical cladistic studies that have postulated Au. ana-mensis to be the sister taxon to Au. afarensis and allsubsequent hominins (Strait and Grine, 2004; Kimbel etal., 2004). Additional support for this evolutionary rela-tionship has been presented by Ward et al. (2010) andHaile-Selassie et al. (2010b). However, whereas Haile-Selassie et al. (2010b) and Haile-Selassie (2010) arguethat Au. afarensis and Au. anamensis do not appear to
TABLE 3. Summary statistics of molar microwear variables recorded for extinct hominin and extant primate species by scanningelectron microscopy
Taxon n
Pit breadthScratchbreadth Pitting incidence
ReferenceMean SD Mean SD Mean SD
Australopithecus anamensis 3 3.11 0.54a 1.20 0.07a 35.93 4.05a Grine et al. (2006a)Australopithecus afarensis 19 2.99 0.71 1.26 0.24 29.20 9.62 Grine et al. (2006b)Australopithecus africanus 10 5.04 1.31 1.04 0.16 30.80 — Grine (1986)Paranthropus robustus 9 8.18 1.31 1.87 0.41 48.50 — Grine (1986)Cebus apella 5 3.20 0.65 1.20 0.14 59.10 8.74 Ungar et al. (2006b)Lophocebus albigena 5 5.80 1.07 1.90 0.23 59.20 11.03 Ungar et al. (2006b)Papio cynocephalus 16 4.37 1.54 0.82 0.10 23.86 15.09 El Zaatari et al. (2005)Papio ursinus 5 5.90 0.99 1.80 0.81 48.40 16.36 Daegling and Grine (1999)Theropithecus gelada 12 3.85 2.43 0.71 0.16 9.42 6.75 El Zaatari et al. (2005)Colobus guereza 18 4.15 1.72 0.79 0.14 7.94 4.54 El Zaatari et al. (2005)Piliocolobus badius 5 3.58 0.68 1.02 0.19 25.00 20.60 El Zaatari et al. (2005)Gorilla gorilla 5 2.80 0.61 1.30 0.36 25.50 9.32 Ungar et al. (2006b)Pan troglodytes 5 3.70 1.12 1.20 0.09 43.70 5.08 Ungar et al. (2006b)
a Statistics based on a sample of only three individuals.
296 F.E. GRINE ET AL.
American Journal of Physical Anthropology

represent ‘‘distinct taxa,’’ Ward et al. (2010) identify Au.anamensis ‘‘not just as a more primitive version of Au.afarensis, but as a . . . member of [a] . . . lineage leadingto Au. afarensis.’’
Paleoecological context. Paleoenvironmental recon-structions have been proffered for nearly all of the local-ities from which Au. afarensis fossils have been recov-ered. The majority of these are based on associated ver-tebrate faunas, although palynological and
paleobotanical records, as well as data from paleosolsand stable isotopes have featured in some interpreta-tions. The habitat reconstructions for Hadar, Laetoli, theMatabaietu Formation, and the Usno and Shungura for-mations have been reviewed by Grine et al. (2006b).Most reflect a degree of mosaicism (but see White, 1988),and all indicate temporal fluctuations. Subsequent recon-structions of Laetoli confirm a mosaic of closed to openwoodland, shrubland, and grassland with seasonalstreams and/or rivers (see Harrison, 2011a). Similarly,
TABLE 4. Microwear texture data recorded for individual Plio-Pleistocene hominin specimens
Specimen Asfc epLsar Stratigraphic derivation
Australopithecus anamensisKNM-KP 29287 0.8079100 0.0029900 KanapoiKNM-KP 34725 1.3099900 0.0024400 KanapoiKNM-ER 35236 0.9746600 0.0030700 Allia Bay
Australopithecus afarensisLH 4 0.5476675 0.0016300 Laetolil Beds, Tuffs 5–6LH 8/22 0.8227370 0.0031855 Laetolil Beds, Tuffs 7–8LH 15 1.0747980 0.0041870 Laetolil Beds,[ Tuff 8AL 128–23 0.6472790 0.0012170 Hadar Fm., Sidi Hakoma MemberAL 145–35 0.8728000 0.0033735 Hadar Fm., Sidi Hakoma MemberAL 200–1b 0.7034190 0.0057205 Hadar Fm., Sidi Hakoma MemberAL 225–8 1.0619605 0.0059380 Hadar Fm., Sidi Hakoma Member.AL 400–1a 0.7286555 0.0022300 Hadar Fm., Sidi Hakoma MemberAL 188–1 0.5338010 0.0039045 Hadar Fm., Denan Dora MemberAL 333–74 0.5391500 0.0060235 Hadar Fm., Denan Dora MemberAL 333w-1a 0.7115010 0.0055275 Hadar Fm., Denan Dora MemberAL 333w-12 1.1654345 0.0037420 Hadar Fm., Denan Dora MemberAL 333w-57 0.6991250 0.0038215 Hadar Fm., Denan Dora MemberAL 333w-59 0.1907650 0.0028540 Hadar Fm., Denan Dora MemberAL 333w-60 0.7794460 0.0051520 Hadar Fm., Denan Dora MemberAL 366–1 0.8518915 0.0008975 Hadar Fm., Denan Dora MemberAL 486–1 0.9710385 0.0013905 Hadar Fm., Denan Dora MemberAL 487–1c 0.4693200 0.0023570 Hadar Fm., Denan Dora MemberAL 288–1i 0.6880415 0.0018385 Hadar Fm., Kada Hadar Member
Australopithecus africanusSts 12 1.9201785 0.0019650 Sterkfontein, Member 4Sts 17 1.9625975 0.0044405 Sterkfontein, Member 4Sts 22 1.3776680 0.0035720 Sterkfontein, Member 4Sts 28 1.1622040 0.0028475 Sterkfontein, Member 4Sts 30 2.1807300 0.0032890 Sterkfontein, Member 4Sts 31 1.6111305 0.0036850 Sterkfontein, Member 4Sts 52 1.3695840 0.0065155 Sterkfontein, Member 4Sts 53 1.4474865 0.0069480 Sterkfontein, Member 4Sts 61 1.0141410 0.0051305 Sterkfontein, Member 4TM 1511 1.1783785 0.0028195 Sterkfontein, Member 4
Paranthropus robustusTM 1517a 2.8955025 0.0021950 Kromdraai B, Member 3SK 13 2.4917210 0.0026805 Swartkrans, Member 1SK 16–1591 4.1075515 0.0022205 Swartkrans, Member 1SK 42 1.8722350 0.0019165 Swartkrans, Member 1SK 48 5.2165345 0.0024070 Swartkrans, Member 1SK 49 5.0635150 0.0022570 Swartkrans, Member 1SK 834 1.2651385 0.0020110 Swartkrans, Member 1SK 837 4.2899105 0.0008905 Swartkrans, Member 1SK 877 4.6871125 0.0014125 Swartkrans, Member 1
Paranthropus boiseiOmo L7A-125 0.5835735 0.0023900 Shungura Fm., Member GOH 5 1.0453360 0.0013835 Olduvai Gorge, Bed IOH 66 0.3053160 0.0013290 Olduvai Gorge, Bed IKNM-ER 3952 0.4103240 0.0054515 Koobi Fora Fm., KBS MemberKNM-WT 17400 0.3617140 0.0018735 Nachukui Fm., Kaitio MemberKNM-ER 729 0.9471220 0.0033160 Koobi Fora Fm., Okote MemberKNM-ER 3230 0.4479180 0.0065505 Koobi Fora Fm., Okote MemberKNM-CH 1B 0.5821925 0.0020330 Chesowanja, Chemoigut Fm.
Specimens within each species are arranged roughly in ascending chronological sequence, and in numerical order within thatsequence. The sources for these values are recorded in Table 1. Derivation of the Au. afarensis specimens recorded in Grine et al.(2006a); Au. anamensis (Grine et al., 2006b); Au. africanus and P. robustus (Grine, 1986); P. boisei (Ungar et al., 2008, in press).
297MICROWEAR, ISOTOPES, AND HOMININ PALEOECOLOGY
American Journal of Physical Anthropology

the more recent geological and paleontological analysesof the Hadar Formation (Campisano and Feibel, 2008;Reed, 2008) confirm paleoenvironmental oscillationswithin an overall trend from closed to open conditions.Reconstructions at Dikika and Woranso-Mille suggest amosaic of habitats, ranging from rather open environ-ments in the proximity of water, but with some woodedhabitats nearby (Wynn et al., 2006), to a riverine galleryforest extending to closed and open woodland and grass-land (Haile-Selassie et al., 2007).As noted by Grine et al. (2006b), not only were diverse
habitats available to Au. afarensis, this species appears tohave had no marked preference for any single one. Indeed,the existence of this species in diverse habitats, and itspersistence in time from Laetoli through the Hadar For-mation sequence, has led to Au. afarensis being viewed asan ecological (dietary) eurytope (Reed, 2008).
Morphological inferences about diet. White et al.(2000) argued that the postulated evolutionary trajectoryfrom Au. anamensis to Au. afarensis would have involvedpostcanine megadontia and other morphological adapta-tions to a more heavily masticated diet. Similarly, Ward etal. (2010) have conjectured that in the presumptive Au.anamensis—Au. afarensis lineage, ‘‘significant changesappear to occur particularly in the anterior dentition, butalso in jaw structure and molar form, suggesting selectionfor altered diet and/or food processing.’’ In particular, they
suggested a dietary change involving anterior tooth usebecause of less intense wear to the incisors and caninesrelative to the molars in Au. afarensis. Ungar (2004)observed that molar topographic relief in Au. afarensisimplies that these teeth are well-suited to fracture brittle,less deformable foods but not as well-suited to fracturetough, more deformable foods as the crowns of either goril-las or chimpanzees. More bulbous cusps should allowtransmission of higher stresses to a food item withoutthemselves breaking. He suggested that the diet of Au.afarensis may have differed from that of Pan largely inhard, brittle fallback resources. In addition to the dentalfeatures enumerated above, this species possesses verydeep and robust mandibular corpora, tall mandibularrami, and robustly constructed zygomatic arches (Kimbelet al., 2004). As summarized by Kimbel and Delezene(2009: 40), Au. afarensis exhibits a number of craniodentalattributes that are ‘‘conventionally associated with ‘heavymastication’ (however imprecisely defined) compared toextant great apes.’’
Dental microwear and stable isotopes. There hasbeen no stable carbon isotope study to date of the toothenamel of Au. afarensis. The foregoing discussion relat-ing to paleohabitat reconstructions and to morphologicaltraits associated with food procurement and processingwould predict d13C values higher than those reported byWhite et al. (2009a) for Ardipithecus.
TABLE 5. Summary statistics of two molar microwear texture components—complexity (Asfc) and anisotropy (epLsar)—for extincthominin and extant primate species
Taxon Component n mean median SD Min. Max.
Australopithecus anamensis complexity (Asfc) 3 1.031 0.975 0.256 0.808 1.310anisotropy (epLsar) 3 0.003 0.003 0.000 0.002 0.003
Australopithecus afarensis complexity (Asfc) 19 0.740 0.712 0.236 0.191 1.165anisotropy (epLsar) 19 0.003 0.003 0.002 0.001 0.006
Australopithecus africanus complexity (Asfc) 10 1.522 1.413 0.387 1.014 2.181anisotropy (epLsar) 10 0.004 0.004 0.002 0.002 0.007
Paranthropus robustus complexity (Asfc) 9 3.543 4.108 1.449 1.265 5.217anisotropy (epLsar) 9 0.002 0.002 0.001 0.001 0.003
Paranthropus boisei complexity (Asfc) 8 0.585 0.515 0.273 0.305 1.045anisotropy (epLsar) 8 0.003 0.002 0.002 0.001 0.007
Alouatta palliata complexity (Asfc) 31 0.430 0.423 0.210 0.140 0.890anisotropy (epLsar) 31 0.006 0.006 0.002 0.002 0.010
Cebus apella complexity (Asfc) 30 5.816 3.083 6.020 0.458 22.389anisotropy (epLsar) 30 0.004 0.003 0.002 0.001 0.007
Lophocebus albigena complexity (Asfc) 23 2.438 1.640 2.258 0.216 9.003anisotropy (epLsar) 23 0.004 0.003 0.002 0.001 0.009
Cercocebus atys complexity (Asfc) 14 4.076 4.260 1.726 1.256 7.496anisotropy (epLsar) 14 0.004 0.003 0.001 0.001 0.006
Papio cynocephalus complexity (Asfc) 27 3.375 1.945 3.235 0.717 14.134anisotropy (epLsar) 27 0.003 0.003 0.001 0.000 0.007
Papio ursinus complexity (Asfc) 12 2.353 1.901 1.343 0.766 5.909anisotropy (epLsar) 12 0.003 0.003 0.001 0.002 0.006
Theropithecus gelada complexity (Asfc) 12 0.727 0.657 0.210 0.489 1.282anisotropy (epLsar) 12 0.006 0.006 0.002 0.003 0.009
Trachypithecus cristatus complexity (Asfc) 12 0.734 0.514 0.660 0.222 2.665anisotropy (epLsar) 12 0.005 0.004 0.003 0.002 0.100
Pongo pygmaeus complexity (Asfc) 15 1.356 1.122 0.947 0.240 3.627anisotropy (epLsar) 15 0.004 0.004 0.002 0.001 0.006
Gorilla gorilla complexity (Asfc) 15 1.694 1.303 1.045 0.290 3.499anisotropy (epLsar) 15 0.004 0.004 0.002 0.001 0.007
Gorilla beringei complexity (Asfc) 16 1.711 1.182 1.455 0.518 6.124anisotropy (epLsar) 16 0.004 0.004 0.002 0.001 0.006
Pan troglodytes complexity (Asfc) 17 2.246 2.114 1.523 0.395 5.461anisotropy (epLsar) 17 0.003 0.002 0.001 0.001 0.006
Data for Australopithecus africanus and Paranthropus robustus are from Scott et al. (2005); Australopithecus anamensis and Aus-tralopithecus afarensis from Ungar et al. (2010); Paranthropus boisei from Ungar et al. (2008, in press). Individual specimen valuesare recorded in Table 4. Extant species data are from Scott et al. (2006; 2012), Ungar et al. (2008) and Daegling et al. (2011b).
298 F.E. GRINE ET AL.
American Journal of Physical Anthropology

In light of the morphological and paleoecological infor-mation for Au. afarensis, it is perhaps surprising thatthe occlusal microwear data recorded by Grine et al.(2006b) and Ungar et al. (2010) for 19 specimens of thisspecies suggest that Gorilla and/or Theropithecus consti-tute the best modern analogs for dietary preference inthose individuals sampled. Suwa et al. (2009) reportedthat their examination of eight molars from Hadar con-firm the results of Grine et al. (2006b). Of course, hypo-geous tubers, bulbs, and roots may have comprised partof the repertoire of Au. afarensis, but there is no micro-wear evidence for the mastication of hard, brittle itemssuch as those processed by Cercocebus atys (Daegling etal., 2011b). Although Au. afarensis possessed the trophicapparatus to process a fairly wide range of foods, includ-ing hard, brittle items such as ‘‘nuts, seeds and hardfruit’’ (Wood and Richmond, 2000: 29), its molar micro-wear suggests that it did not always do so (at least thoseindividuals we have been able to sample do not appearto have done so during the periods in which their micro-wear fabrics were being formed). The constancy of dentalmicrowear in Au. afarensis molars across time and spacemay mean either that this species was able to track itspreferred dietary resources in the face of changing habi-tats and environments, or that environmentally inducedshifts in diet did not involve changes in mechanical prop-erties of the foods typically consumed (Grine et al.,2006b).It is unfortunate that none of the specimens of Au.
afarensis from the Kada Hadar-2 submember of theHadar Formation preserve pristine antemortem micro-wear (Grine et al., 2006b), because it is in this samplethat Lockwood et al. (2000) observed signs of an increasein jaw size. If this signals an attendant increase in bodysize at this time, questions about its potential ecologicalcausation and significance arise (Kimbel and Delezene,2009). Hopefully, carbon isotope data will someday beobtained from teeth that sample this interval.
Australopithecus africanus
Australopithecus africanus is known principally fromdeposits that formed within three paleokarst features inSouth Africa. The type specimen comes from the site ofTaung; a moderate sample is known from Makapansgat,and the bulk of material derives from Sterkfontein.Although these fossils are conventionally regarded asrepresenting a single, albeit variable species (e.g., Whiteet al., 1981; MacLatchy et al., 2010), the possibility thatthe hypodigm comprises more than one species has beena topic of discussion over the past two decades (see Grine[in press] for a detailed treatment of this). While ques-tions persist, studies that have addressed this issue froma statistical perspective have failed to reject the singlespecies hypothesis (Wood, 1991; Ahern, 1998; Moggi-Cec-chi, 2003; Moggi-Cecchi et al., 2006; Grine et al., inpress).
Paleoecological context. A number of paleoenviron-mental reconstructions have been proffered for each ofthe Au. africanus-bearing sites. Although palynologicalanalyses at Makapansgat suggested a closed-forest envi-ronment (Cadman and Rayner, 1989; Rayner et al.,1993), these results may be flawed by contamination.The Makapansgat rodent fauna suggests a significantlywetter environment than today (Levinson 1985); mam-malian community structure, which is dominated by ahigh percentage of browsing species, suggests that this
region was a habitat mosaic that contained riparianwoodland, bushland, and edaphic grassland (Reed, 1997;McKee, 1999). Isotope studies that indicate a woodedmosaic with substantial patches of C4 grasses (Lee Thorpet al., 2007) are consistent with the faunal reconstruc-tions, but isotopic and microwear data suggest a stron-ger browsing component to the bovid fauna than wouldbe implied by either taxonomic uniformitarianism or eco-morphology (Sponheimer et al., 1999; Schubert et al.,2006).The Sterkfontein paleofaunal community suggests a
habitat of open woodland, with areas of bushland andthicket (McKee, 1991; Reed, 1997), and some riverineforest as indicated by the presence of lianas (Bamford,1999). The rodent fauna also indicate an ecotonebetween dry sandy highveld grassland and Kalaharithornveld (Avery, 2001), which is consistent with thepresence of open savanna suggested by the fossilbaboons (Benefit and McCrossin, 1990). Indeed, analysisof d13C of the Sterkfontein Member 4 ungulate faunasuggests a greater abundance of mixed feeders than isfound in the younger deposits at Sterkfontein or at otherSouth African sites (Lee-Thorp et al., 2007).Paleoenvironmental reconstructions for Taung are
bedeviled by the fact that the vast bulk of the faunalassemblage likely derives from the Hrdlicka cave ratherthan being associated with the hominin skull (Butzer,1974). Study of the sediments associated with the Taungskull indicates conditions similar to or slightly wetterthan those of today (Partridge, 2000).
Morphological inferences about diet. Morphologicalvariation in the Au. africanus hypodigm has enabledworkers to view this species in quite different ways,depending upon the features chosen for emphasis. Thus,some have argued that this species lacks features thatare distinctive of Paranthropus (e.g., Robinson, 1967;Olson, 1981, 1985), whereas others have focused on fea-tures regarded as indicating nascent masticatory special-ization to proclaim Au. africanus as a being related tothe ‘‘robust’’ australopiths (e.g., White et al., 1981; Rak,1983; Kimbel et al., 2004).The morphological attributes of Au. africanus have led
to numerous suggestions relating to its dietary procliv-ities (e.g., Robinson, 1954a, 1954b; Jolly, 1970; Wolpoff,1973; Wallace, 1975; Du Brul, 1977; Kay, 1985; Strait etal., 2009). Notwithstanding unusual suggestions of carni-vory (Schaller and Lowther, 1969; Cachel, 1975; Szalay,1975), most reconstructions have envisioned Au. africa-nus as having been primarily, if not exclusively herbivo-rous, with adaptations that permitted the processing, atleast seasonally, of cereals, grains, and/or nuts. In partic-ular, Strait et al. (2009) have argued that the craniofa-cial morphology of Au. africanus attests to the inclusionof hard seeds and nuts in its diet, with these items beingprocessed by the premolars. According to Strait et al.(2009), premolar processing of hard objects is indicatedby the presence of facial biomechanical buttresses (the‘‘anterior pillars’’) that Rak (1983) envisioned as linkingAu. africanus with the enhanced masticatory apparatusof the ‘‘robust’’ australopith, P. robustus. Strait et al.(2009) explained the absence of a ‘‘strong hard-objectmicrowear signal’’ on the molars of Au. africanus by pos-tulating that such objects were initially processed by thepremolars, leaving only the softer remnants to bechewed by the molars. Notwithstanding the observationthat the anterior pillars of Au. africanus and P. robustus
299MICROWEAR, ISOTOPES, AND HOMININ PALEOECOLOGY
American Journal of Physical Anthropology

are not structurally homologous (Villmoare and Kimbel,2011), there are numerous problems manifested by theirfinite element analysis (Grine et al., 2010) and its appli-cation to the world of living organisms (Daegling et al.,2011b). Moreover, because the premolars of Au. africa-nus display the same microwear fabrics as the molars(Grine et al., 2010), there is no evidence at all to supportthe argument by Strait et al. (2009) for premolar nut-cracking in this species.
Dental microwear and stable isotopes. Qualitative aswell as quantitative studies of microwear on Au. africa-nus molars have revealed a predominance of surfaceswith anisotropically oriented striae and small pits(Grine, 1981, 1986; Grine and Kay, 1988; Kay and Grine,1988). The average breadth of wear striae in Au. africa-nus is nearly the same as that recorded for Au. afarensis(1.04 lm vs. 1.26 lm respectively); the incidences of pit-ting in the two species are also nearly identical (31% vs.29%), but the pits tend to be larger in Au. africanus(5.04 lm vs. 3.11 lm respectively) (Table 3). The micro-wear fabric textures of Au. africanus and Au. afarensismolars exhibit comparable degrees of anisotropy,whereas Au. africanus displays notably greater surfacecomplexity (Scott et al., 2005; Ungar et al., 2010) (Table5). The difference in average pit size is wholly consistentwith the higher complexity average for Au. africanus.The greater complexity in Au. africanus may relate to
a somewhat greater degree of variability in its dietaryrepertoire compared to Au. afarensis. Although Au. afri-canus has higher average microwear complexity (i.e., pit-ting) than Au. afarensis, it is still considerably lowerthan that expected of a hard object feeder, such asLophocebus albigena or Cercocebus atys (Table 5). Con-trary to suggestions that the premolars of Au. africanuswere employed to crack open hard objects while theirmolars were employed in the mastication of the softerkernels (Strait et al., 2009), the microwear textures ofthe premolars and molars do not differ from one anotherin this species (Grine et al., 2010).Carbon isotope values of Au. africanus tooth enamel
suggest that some 35–40% of the diet of this species con-sisted of C4 plants, whereas the predominant component(ca., 65%) was C3-based (Sponheimer and Lee-Thorp,1999a; Lee-Thorp and Sponheimer, 2006). Thus, whileAu. africanus consumed primarily C3 vegetation (e.g.,fruits and leaves), it also ingested considerable quanti-ties of C4 resources, possibly grasses or sedges. Ofcourse, given the possibility of some limited faunivory inAu. africanus, these isotope values could have beeninfluenced indirectly through the consumption of ani-mals that ate C3 or C4 plants. These values are interest-ing because even in environments where C4 foods arereadily available to chimpanzees, their d13C values donot indicate any C4 consumption (Schoeninger et al.1999; Carter 2001; Sponheimer et al., 2006a). This sug-gests a fundamental dietary difference between Au. afri-canus and Ar. ramidus (Table 6). Ardipithecus ramidusappears to have been most similar to extant chimpan-zees, although at least some individuals of this extincttaxon seem to have consumed small quantities of C4
foods. As such, Au africanus was not restricted to thekinds of foods that living apes would utilize in the closedwoodland and gallery forest habitats of the BlaaubankValley; rather, this species likely foraged onto the opengrasslands and into local vleis (patches of generally sea-sonal marshland with C4 sedges) for a substantial partof its dietary intake.
At the same time, Au. africanus d13C values exhibitgreater variability than those for virtually all modernand fossil taxa that have been analyzed in South Africa,including P. robustus (Lee-Thorp et al., 1994; Spon-heimer et al., 1999, 2003; Codron, 2003; van der Merweet al., 2003). Indeed, the range of d13C values within theAu. africanus sample (21.8% to 211.3%) is nearly asgreat as the entire combined range for Papio and Thero-pithecus (10.4% to 212.6%) (Codron et al., 2005). Thismay speak to high seasonal and/or interannual variationin food availability. Alternatively, it may be related tothe imprecisely known temporal depth of the Sterkfon-tein Au. africanus sample, or potentially even to its taxo-nomic heterogeneity. However, the variability in d13Cvalues expressed between Au. africanus individuals isalso manifest within individual teeth. Thus, a high reso-lution laser-ablation study of individual teeth fromSterkfontein revealed very high intra-individual d13Cvariability during crown formation (Lee-Thorp et al.,2010). This indicates dietary variability within an indi-vidual over the period of time during which its toothcrowns were forming.The low Ba/Ca ratios in Au. africanus enamel are con-
sistent with the consumption of grass roots and/orUSOs, but probably not consistent with the consumptionof above-ground grass plant parts (Sponheimer and Lee-Thorp, 2006). The high Sr/Ba ratios for Au. africanusare rather striking, being similar to those observed inUSO-consuming mole rats (Cryptomys hottentotus) inSouth Africa today (Sponheimer and Lee-Thorp, 2006).Finally, the low 18O/16O ratios observed for Au. africanusare also consistent with the consumption of USOs, butthey also may reflect a high degree of water dependency(Lee-Thorp et al., 2010).
Paranthropus robustus
This species is known from deposits that formedwithin six geographically proximate paleokarst featuresin South Africa. Skulls and teeth attributable to P.robustus are reasonably abundant, being especially plen-tiful at the sites of Swartkrans (e.g., Broom and Robin-son, 1952; Robinson, 1956; Grine, 1989; Grine andStrait, 1994) and Drimolen (Keyser, 2000; Keyser et al.,2000; Moggi-Cecchi et al., 2010). A smaller, albeit signifi-cant collection, including the type specimen, is knownfrom the nearby site of Kromdraai (Thackeray et al.,2001), and a few isolated specimens are known fromSterkfontein Member 5B (Kuman and Clarke, 2000),Cooper’s (Steininger et al., 2008), and Gondolin (Menteret al., 1999; Grine et al., Grine et al., in press b).Although the notion that the Kromdraai and Swartk-
rans fossils represent two species, namely P. robustusBroom, 1938 and P. crassidens Broom, 1949 respectively,
TABLE 6. Summary statistics of d13C values recorded for Plio-Pleistocene hominin species samples
Taxon n Mean SD SE Min. Max.
Ardipithecus ramidus 5 210.22 1.02 0.46 211.2 28.5Australopithecus africanus 24 26.61 2.33 0.48 211.3 21.8Paranthropus robustus 22 27.39 1.26 0.27 210.0 24.9Paranthropus boisei 24 21.29 0.92 0.19 23.4 10.7
The data upon which these statistics are based are recordedin Table 2. The d13C values are % where d13C 5(13C/12Csample/
13C/12Cstandard 2 1) 3 1,000, and the internationalstandard is Vienna Peedee Belemnite (VPDB).
300 F.E. GRINE ET AL.
American Journal of Physical Anthropology

gained short-lived support from cranial and especiallydeciduous dental comparisons (Howell, 1978; Grine 1982,1985), the discoveries at Drimolen have blurred theseapparent differences (Keyser et al., 2000; Moggi-Cecchiet al., 2010). Taken together, all the fossils from thesevarious sites are conventionally regarded as representinga single species (MacLatchy et al., 2010). Although thegeochronological ages of the sites from which P. robustusfossils derive continue to be a matter of discussion, andnotwithstanding U-Pb determinations for speleothemsthat would see Swartkrans Member 1 and SterkfonteinMember 4 as at least partially if not wholly coeval (Pick-ering et al., 2011), it is abundantly clear that the P.robustus-bearing deposits postdate those that containfossils of Au. africanus.
Paleoecological context. A number of paleoenviron-mental reconstructions have been proffered for the P.robustus-bearing sites of Kromdraai and Swartkrans,and while not all are in strict agreement, most indicatemore open (e.g., grassland) and perhaps drier conditionsthan prevailed earlier during the accumulation of theAu. africanus-bearing deposits. Thus, the large mammalassemblage from Swartkrans Member 1 has been envi-sioned as representing a moderately open grassland sa-vanna (Vrba, 1975; Shipman and Harris, 1988), and sa-vanna woodland with edaphic grasslands (Watson, 1993;Reed, 1997). A grassland component is also suggested bythe micromammals (Avery, 2001). The Swartkrans Mem-ber 2 fauna suggests somewhat drier, moderately openwooded grassland (Vrba, 1975; Shipman and Harris,1988; Reed, 1997; Avery, 2001). Swartkrans Member 3 isbest reconstructed as representing more open, edaphicgrassland conditions than either Member 1 or Member 2(Reed, 1997; Avery, 2001).The Kromdraai B fauna suggests open grassland with
patches of riparian woodland (Vrba, 1975, 1985; Reed,1997). Similarly, the Sterkfontein Member 5B fauna indi-cate open savanna (Vrba, 1975; McKee, 1991) or an opento wooded grassland (Reed, 1997), and certainly drier con-ditions than prevailed during the accumulation of the ear-lier Au. africanus-bearing deposit (Avery, 2001).
Morphological inferences about diet. The morpholog-ical attributes of P. robustus have led a number ofworkers to surmise its dietary proclivities. In particu-lar, its characteristically diminutive incisors and can-ines, large and thickly enameled postcanine teeth,molarized premolars, thick palate, robust mandible,and an orthognathic face with anteriorly positioned,large molars are part of a coherent and distinctivepackage that has been seen as reflecting the generationand distribution of powerful chewing forces in associa-tion with an herbivorous diet (Robinson 1954b, 1967;Grine, 1981; Rak, 1983; Kay, 1985). Most workers haveinterpreted the P. robustus trophic morphology as beingconsistent with a diet that likely included at least somehard foods.
Dental microwear and stable isotopes. Initial micro-wear studies (Grine, 1981, 1986; Grine and Kay, 1988;Kay and Grine, 1988; Ungar and Grine, 1991) indicatedsupport for the notion that the dietary differencesbetween Au. africanus and P. robustus involved differ-ences perhaps not only in the sizes of the foods eaten(with P. robustus eating smaller objects that requiredless ingestive anterior tooth use), but also in the qualityof the items masticated (with harder food objects com-
prising a greater proportion of the diet of P. robustus).Molar microwear, for example, revealed a higher per-centage incidence of pitting (49% compared to 31% inAu. africanus and 29% in Au. afarensis), and signifi-cantly larger pits (8.2 lm compared to 5.0 lm in Au.africanus and 3.0 lm in Au. afarensis) (Table 3). Thesedata, however, also revealed a degree of overlapbetween Au. africanus and P. robustus, which was high-lighted by subsequent analyses employing texture data(Scott et al., 2005) (Table 5).Microwear texture analysis revealed greater average
complexity and lower anisotropy in P. robustus than inAu. africanus, suggesting that this particular ‘‘robust’’australopith chewed more hard and brittle items. At thesame time, there is significantly more individual variationin complexity in P. robustus (Table 5). The variability incomplexity in the P. robustus sample suggests comparisonwith extant primate species such as Lophocebus albigenaand Cebus apella that consume hard objects as fallbackfoods when softer, more preferred foods are unavailable.Scott et al. (2005) suspected that the morphological attrib-utes of P. robustus might well reflect such less commonlyeaten, but critical fracture-resistant foods.With regard to chemical analyses of diet in P. robus-
tus, the pioneering work by Sillen (1992) and Sillen etal., (1995) on Sr/Ca ratios in bones from Swartkrans ledto the conclusion that this species was unlikely to havebeen ‘‘purely herbivorous.’’ However, as noted above,there are serious limitations to these studies, not theleast of which pertains to their having been conductedon bone, which is particularly vulnerable to diageneticoverprinting. Indeed, tooth enamel from P. robustus (likethat of Au. africanus) does not have lower Sr/Ca rationsthan contemporaneous baboons and other herbivores(Sponheimer et al., 2005a). Thus, Sr/Ca data provide noevidence of omnivory in P. robustus.The carbon isotope data from P. robustus tooth enamel
indicate strong differentiation from contemporaneousbovids that were constrained C3 and C4 consumers(Sponheimer et al., 2005b). This distinction cannot beascribed to diagenesis, as there is no evidence that eitherbrowser or grazer d13C has been altered, and diagenesisshould affect all mammalian tooth enamel alike. Thed13C values of P. robustus suggest a diet comprisingsome 35–40% C4 vegetation. Thus, although it consumedprimarily C3 plant material (e.g., fruits, nuts, andleaves), it also ingested significant quantities of C4
resources, possibly grasses or sedges. In this regard, P.robustus cannot be distinguished from Au. africanus,whose d13C values also indicate that 35–40% of the dietconsisted of C4 plants (Table 6).The P. robustus d13C values are notably less variable
than those recorded for Au. africanus, but are more vari-able than most of the extant and fossil taxa that havebeen analyzed in South Africa (Sponheimer et al.,2005b). This might be related to short-term environmen-tal perturbations during the accumulation of the P.robustus-bearing deposits (primarily Swartkrans) thatdrove dietary diversity among an opportunistic homininwith wide habitat tolerances. Moreover, as with Au. afri-canus, the variability is expressed not only between dif-ferent individuals, but is also manifest within individualteeth. A study using laser-ablation mass spectrometrythat sampled the growth lines of individual teeth fromSwartkrans revealed variation of up to 5o/oo in d13C val-ues over intra-annual (seasonal) and inter-annual time-scales (Sponheimer et al., 2006b). Bearing in mind that
301MICROWEAR, ISOTOPES, AND HOMININ PALEOECOLOGY
American Journal of Physical Anthropology

tooth mineralization continues to occur for months afterinitial mineral deposition, which results in isotopic over-printing that dampens apparent dietary perturbations(Passey and Cerling, 2002; Balasse, 2003), a change ofover 5o/oo is quite extraordinary. This almost certainlyindicates that the individual in question changed from aC3- to a C4-dominated diet during the period of enamelformation.While we interpret the P. robustus d13C values in rela-
tion to the predominant if not ubiquitous consumption ofC3 and C4 vegetation, other interpretations are possible.Thus, those who have argued for significant carnivory inthis species (e.g., Cachel, 1975; Sillen, 1992; Sillen et al.,1995) might view the d13C values as reflecting or at leastbeing influenced significantly by the type of fauna con-sumed (i.e., browsers versus grazers). Alternatively, thed13C values and morphological attributes of P. robustushave been interpreted as being consistent with those of amember of a guild of consumers of hard-shelled inverte-brate prey, such as freshwater potamonautid crabs(Shabel, 2006, 2008). According to this ‘‘durophagemodel,’’ the stable carbon isotope values of P. robustusare seen as being similar to those of extant South Afri-can macro-invertebrate consumers such as the marshmongoose (Atilax) and Cape clawless otter (Aonyx), withthe dentognathic morphology of P. robustus being held torepresent an adaptation to cracking crab shells. Whilethis might be viewed as little more than an aside, otters(this time in the form of the sea otter, Enhydra lutris)have once again been proposed as a model by which thetrophic adaptations (specifically the extreme enamelthickness) of Paranthropus to hard object feeding can beexplained (Constantino et al., 2011). In our view, thethin, softer enamel of Enhydra raises questions aboutthe legitimacy of this taxon serving as a model for hard-object feeding in these thick-enameled hominins.The P. robustus d18O values, like those of Au. africa-
nus, are low compared to other paleofauna from SouthAfrica (Sponheimer and Lee-Thorp, 1999b; Lee-Thorp etal., 2010). Here too, however, the causes of these rela-tively low values remain obscure—again, they may belinked to frugivory, the use of USOs, water dependence,or some combination thereof.
Paranthropus boisei
Fossils attributable to P. boisei are known from sitesextending from southern Ethiopia through Kenya andTanzania, and possibly into Malawi. Specimens areknown from Olduvai Gorge (including the type speci-men) and Peninj, Tanzania (see Wood and Constantino,2007 and references therein). A number of spectacular P.boisei fossils have been recovered from the Upper Burgi,KBS, and Okote Members of the Koobi Fora Formation,and from the Kaitio Member of the Nachukui Formationon the eastern and western sides of Lake Turkana,respectively. This species is also represented in the Che-moigut Formation near Chesowanja, Kenya, and insouthern Ethiopia in Members G through L of the Shun-gura Formation and at Konso Gardula.The earliest known occurrences of undoubted P. boisei
fossils are in the Upper Burgi Member of the Koobi ForaFormation and Member G of the Shungura Formation atapproximately 2.3 Ma (Suwa, 1988; Suwa et al., 1994;Wood et al., 1994). Paranthropus fossils from Kenya andTanzania that are older have been attributed to P.aethiopicus on the basis of cranial, dental, and mandibu-
lar differences from P. boisei (Suwa, 1988; Suwa et al.,1994; Wood et al., 1994; Kimbel et al., 2004; Harrison,2011b). A fragment of maxilla from the Chiwondo Bedsof Malawi (Kullmer et al., 1999) may represent P. boiseior perhaps P. aethiopicus if the faunal comparisons thathave been made to Shungura Members D–F (ca., 2.5–2.3Ma) are correct. The last occurrence of P. boisei isrecorded in Member II of Olduvai Gorge at about 1.3Ma. Thus, this species has a geochronological range ofabout one million years. Wood and Constantino (2007)have documented an impressive hypodigm comprisingsome 111 specimens for P. boisei.
Paleoecological context. Given the lengthy geochrono-logical and wide geographic ranges for P. boisei, it is notsurprising that a variety of paleohabitat reconstructionshave been proffered. Those for the earliest sites fromwhich undoubted P. boisei fossils derive (Member G ofthe Shungura Formation and the Burgi Member of theKoobi Fora Formation), suggest a wet habitat, with riv-erine forests or woodlands associated with extensive eda-phic grasslands (Bonnefille, 1985; Shipman and Harris,1988; Reed, 1997). The fauna from the somewhatyounger KBS and Kaitio members have been interpretedby Shipman and Harris (1988) as suggesting a wet habi-tat; Feibel et al. (1991) and Reed (1997) envision scrubwoodland together with wet and dry grasslands. TheOkote Member, which extends from just over 1.6 Ma tosome 1.4 Ma, has been interpreted as preserving a wethabitat, dominated by wetlands and edaphic grasslands,with the presence of some woodland and gallery forests(Shipman and Harris, 1988; Feibel et al., 1991; Reed,1997). By contrast, the extensive 1.4-Ma-old fauna fromKonso indicates the presence of extensive dry grasslands(Suwa et al., 1997), and although there is evidence of alake nearby (Nagaoka et al., 2005), no P. boisei fossil hasbeen recovered from any of the more mesic localities.South of the Turkana Basin, most of the work on pale-
ohabitat reconstruction of relevance to P. boisei hasfocused on Olduvai Gorge. Paleogeographic reconstruc-tions indicate that throughout most of Bed I, the OlduvaiBasin was dominated by a lake of fluctuating size (Hayand Kyser, 2001). Isotopic data indicate generally wetterconditions than prevail today (Cerling and Hay, 1986),and riparian woodlands and wooded grasslands appearto have been evident around the lake (Sikes, 1994).Towards the top of Bed I (ca., 1.75 Ma) there appears tohave been a shift from more closed, wetter habitats tomore open, drier conditions, as evidenced by the rodentfauna (Jaeger, 1976; Kappelman, 1984; Fernandez-Jalvoet al., 1998), pollens (Bonnefille, 1995), and bovids (Gen-try and Gentry, 1978; Kappelman, 1984; Shipman andHarris, 1988; Plummer and Bishop, 1994; Kappelman etal., 1997). During Bed II times (i.e., ca. 1.7–1.3 Ma), thelake margin woodland seems to have given way to opengrasslands (Kappelman, 1984; Cerling and Hay, 1986),and the bovid fauna indicates the increased presence ofopen and arid habitats (Shipman and Harris, 1988).Comparatively little paleohabitat information is avail-
able for the P. boisei-bearing sites of Peninj and Cheso-wanja. For the former, pollen analysis by Dominquez-Rodrigo et al. (2001) indicates the presence of a deltaicenvironment during the deposition of the Upper Sandsunit of the Humbu Formation. The P. boisei mandible,however, derives from the underlying Basal Sands unit,and it is unclear to what degree (if any) the pollen datapertain. Analysis of the Chesowanja fauna by Bishop
302 F.E. GRINE ET AL.
American Journal of Physical Anthropology

et al. (1978) suggests a bushy grassland habitat, with riv-erine and lacustrine elements, but it is unclear to whatdegree the fauna is associated with the hominin fossil.While a variety of potential habitats have been recon-
structed for P. boisei, extending from gallery forests, ri-parian woodlands, extensive edaphic grasslands to dryshrub-grasslands, a fairly common theme suggested bymany—at least in the earlier part of the temporal rangeof this species—is the existence of open, well-wateredsites (e.g., deltaic environments and/or edaphic grass-lands with vegetation growth in water-logged soils, suchas might be encountered in wetlands and swamps). Itwould appear that the later sites tend to be somewhatdrier, although even here there is commonly an underly-ing theme of riverine and/or lacustrine elements. Not-withstanding the taphonomic problems that may beassociated with determining to what degree P. boiseiactually preferred to live in such habitats (White, 1988),this species appears to be most commonly associatedwith fairly open and well-watered habitats.
Morphological inferences about diet. Many of thecraniodental features that characterize P. robustus arepossessed also by P. boisei, where the trophic attributesare even more exaggerated. These features include fur-ther molarization of the premolars, an even deeper max-illa, zygomatics that tend to be laterally bowed with a‘‘visor’’-like configuration of the cheek, and temporalisand especially masseter muscles that appear to have hadmore anterior attachments than in P. robustus (Rak,1983, 1988). The dimensions of P. boisei premolars andmolars are the largest recorded for any hominin species.Lucas et al. (1986: 269) have suggested that the M1–
M3 size ratio indicates the consumption of ‘‘small mouth-fuls of leaves and seeds.’’ The low, bulbous cusps of thecheek teeth would seem to be inconsistent with a dietconsisting of leaves and grass blades, unless they wereyoung and less tough than mature leaves. The compara-tively large masticatory muscles would have been able togenerate powerful forces, but these could have beenreduced to unexceptional levels if dissipated uniformlyacross the postcanine occlusal table, such as in the proc-essing of soft, pliant foods (Demes and Creel, 1988; seealso Walker, 1981). The large, thick mandibular corporawould have been able to withstand powerful bendingand twisting moments (Hylander, 1988; Daegling, 1989).Many workers have understandably viewed the cranio-
dental features of P. boisei as specialized adaptations toa diet that likely consisted of small, hard objects. On theother hand, Wood and Strait (2004) envisioned P. boiseias having been an ecological generalist, capable of inhab-iting a variety of environments and consuming a broadarray of foods with the sole exception of ‘‘tough, fibrousfood items’’ such as leaves. The notion, however wide-spread among paleoanthropologists, that the diet of the‘‘hyper-robust’’ P. boisei included hard objects clearlygained indirect support from the microwear of its SouthAfrican ‘‘robust’’ cousin, P. robustus, which indicates theconsumption of hard food items, at least as a significantfallback resource if not as a primary resource (Grine,1986; Scott et al., 2005).
Dental microwear and stable isotopes. The first ex-amination of molar microwear in P. boisei was an SEManalysis undertaken by Walker (1981), who reportedqualitative observations on several specimens from KoobiFora. He remarked that these teeth exhibited finescratches and few, small pits, being reminiscent of the
wear seen in chimpanzees, orangutans, and mandrills.As such, he suggested that the diet of P. boisei may haveentailed prolonged chewing of tough or fibrous vegetablefoods, perhaps with relatively little nutritional value,rather than the consumption of hard food items. As itturns out, this seems to have been a profoundly pro-phetic, albeit largely ignored inference.The first systematic quantitative analysis of microwear
in this species was undertaken nearly thirty years laterby Ungar et al. (2008). That study found that very fewspecimens (only seven of 53 molars) preserved unob-scured, antemortem microwear. Fortunately, this samplespans much of the geochronological and geographicranges of P. boisei; it has been augmented recently withthe addition of a premolar from Bed I of Olduvai Gorge(Ungar et al., in press).2 The microwear fabrics of P. boi-sei teeth are dominated by fine striations; none exhibitthe large, deep pits expected of a hard-object specialistor the uniformly large, deep, and parallel striationsobserved in tough food grazers. The complexity valuesfor these specimens fall near the bottom end of the rangefor extant primates examined thus far, and they exhibitminimal variation (Table 5).The microwear fabrics of the P. boisei specimens show
remarkable uniformity over time (extending from ca.,2.3–1.4 Ma). Although the small size of the sample pre-cludes statistical analysis of its temporal distribution,given the evidence at hand, there is no reason to suspectthat the P. boisei wear signatures will be seen to exhibittemporal or even paleohabitat related change with largersamples. On the other hand, Suwa et al. (2009) reportedthat the Pa. boisei molars from Konso ‘‘tended to includemoderately coarse to broad striae.’’ They remarked that
‘‘this differs from the generally weakly striated and/orpitted molar microwear surfaces of [the]. . . individualsreported by Ungar et al. (2008). The Konso microwearsuggests larger ingested abrasives and/or greater masti-catory force during grinding, indicating a more abrasivefeeding environment involving tough/hard foodstuffs.’’
If quantitative data on microwear for the sample fromthis site are shown to differ from the others published todate (Table 4), this might be attributable to the paleoha-bitat reconstructed for Konso, which features the pres-ence of extensive dry grasslands (Suwa et al., 1997).The microwear results reported by Ungar et al. (2008,
in press), although predicted by the SEM study byWalker (1981), were quite unexpected given the cranio-dental attributes of P. boisei. In particular, the micro-wear fabrics of P. boisei differ markedly from the patternshown by P. robustus, implying that these two congenersprobably had markedly different diets and/or foragingstrategies. Whereas P. robustus wear textures resemblethose of extant hard object eaters (e.g., Lophocebus albi-gena), the wear in P. boisei resembles most closely thatof gelada baboons and extant African apes. There is nomicrowear evidence for the consumption of hard foods,and its microwear fabrics appear to suggest a more spe-cialized diet than that of P. robustus. Perhaps the highlyderived masticatory morphology of this species repre-
2Ungar et al. (in press) recorded microwear data for two P. boiseispecimens from Olduvai Gorge: OH 66 (RP4) and OH 74 (LM2).However, subsequent observations indicate that OH 74 should beexcluded from microwear analysis owing to its overall state of pres-ervation, and the singular nature of its tiny protocone facet. It iseschewed from consideration here.
303MICROWEAR, ISOTOPES, AND HOMININ PALEOECOLOGY
American Journal of Physical Anthropology

sents an adaptation to the biomechanical demands of re-petitive loading of abrasive foods rather than peak load-ing of hard foods.Carbon isotopes have been obtained from two P. boisei
specimens from Olduvai Gorge and Peninj (van derMerwe et al., 2008), one from the Baringo Basin (Chem-eron), and 21 from the Koobi Fora and Nachukui forma-tions (Cerling et al., 2011). The d13C values for these 24specimens (mean 5 21.3 6 0.9%; observed range 5 0.7%to 23.4%) are indistinguishable from those of coeval (andmodern) C4 grass consumers from the region. At thesame time, the P. boisei sample differs in a highly signifi-cant manner from dedicated C3 biomass consumers(Cerling et al., 2011). Like the microwear signatures, thed13C values do not change over the half million years (ca.,1.9–1.4 Ma), or over the sampled geographic range. More-over, the degree of variance among specimens is low, espe-cially when compared to Au. africanus (Tables 2 and 6).The d13C values for P. boisei correspond to a diet
where C4 plants comprise, on average, some 77% of thebiomass consumed, with individual values rangingbetween 61% and 91%. As such, the carbon isotope datafor P. boisei are most consistent with its having been areasonably devoted consumer of C4 plants. To put theseresults in perspective, the mean d13C value for P. boisei(21.3%) is more positive (i.e., indicating more C4) thanthat for a sample of Theropithecus fossils from SouthAfrica (22.4%) (Codron et al., 2005; Fourie et al., 2008).The most likely candidates are C4 grasses, C4 sedges
such as Cyperus papyrus, or some combination thereof.These data are not consistent with these individualshaving consumed more than 20–30% fruits or nuts (atleast during the period during which the tooth crownswere being formed). At this point, it is worth noting thatthe presence of wetlands in most P. boisei habitat recon-structions would be consistent with the consumption ofCyperus papyrus rhizomes and/or culm, a potential foodresource that was emphasized by van der Merwe et al.(2008). While grasses could have comprised part of thediet, these are highly seasonal. By contrast, Cyperaceaeare perennials, being available in all seasons in the vi-cinity of water. Although van der Merwe et al. (2008)argued that the nutritional value of Cyperus papyrusrhizomes and culm compares favorably with that of thedomesticated potato, such that about 2 kg of raw papy-rus rhizome might supply the daily energy requirementsof a human adult, these rhizomes are woody, and theirenergy returns are likely to have been overestimated(Schoeninger et al., 2001). Thus, it is possible, but uncer-tain whether Cyperus papyrus rhizomes and culm wouldhave sufficed for P. boisei, unless it had the digestivecapacity to utilize such foods. Finally, with regard to thepresence of wetlands or proximate water sources inmany paleohabitat reconstructions, it is worthy of notethat the P. boisei d18O values suggest that this specieswas very water dependent (Cerling et al., 2011),although these relatively low values could also be relatedto the use of USOs.Interestingly, the microwear signatures of the P. boisei
specimens measured to date are similar to thoserecorded for the gelada baboon, although Theropithecushas notably higher levels of anisotropy. While we suspectthat P. boisei microwear is consistent also with prolongedbouts of mastication of Cyperus papyrus rhizomes and/orculm, this cannot be tested because no primate (or othermammal) specializes on this resource today.
DISCUSSION
Dental microwear and stable isotope analyses of toothenamel provide the only direct evidence of the diets ofextinct species. In essence, these two methods can beviewed as independent tests of functional morphologicalinferences by revealing clues to the sorts of foods that anindividual may actually have eaten at a specific period(or specific periods) during its lifetime. Microwear andtooth enamel isotopes will only rarely record a synchro-nous dietary signal. For example, it is possible that iso-topic signatures could be obtained for an M3, the enamelcap of which is in the process of forming, and microweardetails recorded for an M1 of the same individual thatwould be in functional occlusion over this period—ca.,10.5–13.5 years in a recent human (AlQahtani et al.,2010). This would, however, represent a rare occurrencein the paleontological record. As such, isotopic andmicrowear data for a single individual will usually repre-sent two discrete periods of time (and of varying dura-tion). Just as it is becoming increasingly clear that themorphological features that are used to infer diet in aspecies probably reflect phylogenetic history and adapt-ive responses in its lineage perhaps as much as, if notmore than the ‘‘average’’ dietary habits of its members(Grine et al., 2010), so too do microwear and biogeochem-ical analyses have their limitations.Some biogeochemical approaches are bedeviled by
problems of diagenetic alteration. Stable carbon isotopes,which are most heavily employed for fossils that ante-date the Middle Pleistocene, may reflect the consumptionof different types of plant foods (C3 or C4), but are silentwith regard to the parts of the plants (leaves, fruit, nuts,etc.) consumed. At the same time, they may be affectedby the hidden veil of trophic level (e.g., consumption ofprey animals that consumed a particular type of vegeta-tion). Moreover, comparisons among species from differ-ent geographic regions may also depend on the environ-ment from which a particular food source derived. Oxy-gen isotopes may reflect differences in diet (e.g.,frugivory), the degree of water dependence, the geo-graphic specificity of local water d18O, or some combina-tion thereof. Unfortunately, as noted previously, ourunderstanding of d18O patterning in modern trophic sys-tems is still rather limited.At the same time, microwear fabrics reflect diets only
insofar as they reflect the fracture properties of its con-stituent items. In other words, the demonstrable rela-tionship of microwear with diet is actually betweenenamel wear, the properties of the foods consumed, andocclusal mechanics. While it is the abrasives—generallyin the form of silica phytoliths or very hard, small exo-carps—that cause microwear, it also reflects occlusalmovements that are affected by the fracture propertiesof the foods being masticated. Microwear analysis alsomay be bedeviled by artifacts produced by exogenousgrit, and even with extreme care it may not be possibleto identify this except in the more extreme cases (e.g.,Daegling and Grine, 1999). Finally, high-magnification(3200–3500) microwear analysis is a slow and painstak-ing task. Attempted shortcuts using low magnification(e.g., 335) methods developed with reference to the rea-sonably obvious differentiation of grazing and browsingungulates (Solounias and Semprebon, 2002) have beenextended to fossil primates (Godfrey et al., 2004, 2005;Semprebon et al., 2004; Carter, 2006; Williams andPatterson, 2010). However, these methods appear to be
304 F.E. GRINE ET AL.
American Journal of Physical Anthropology
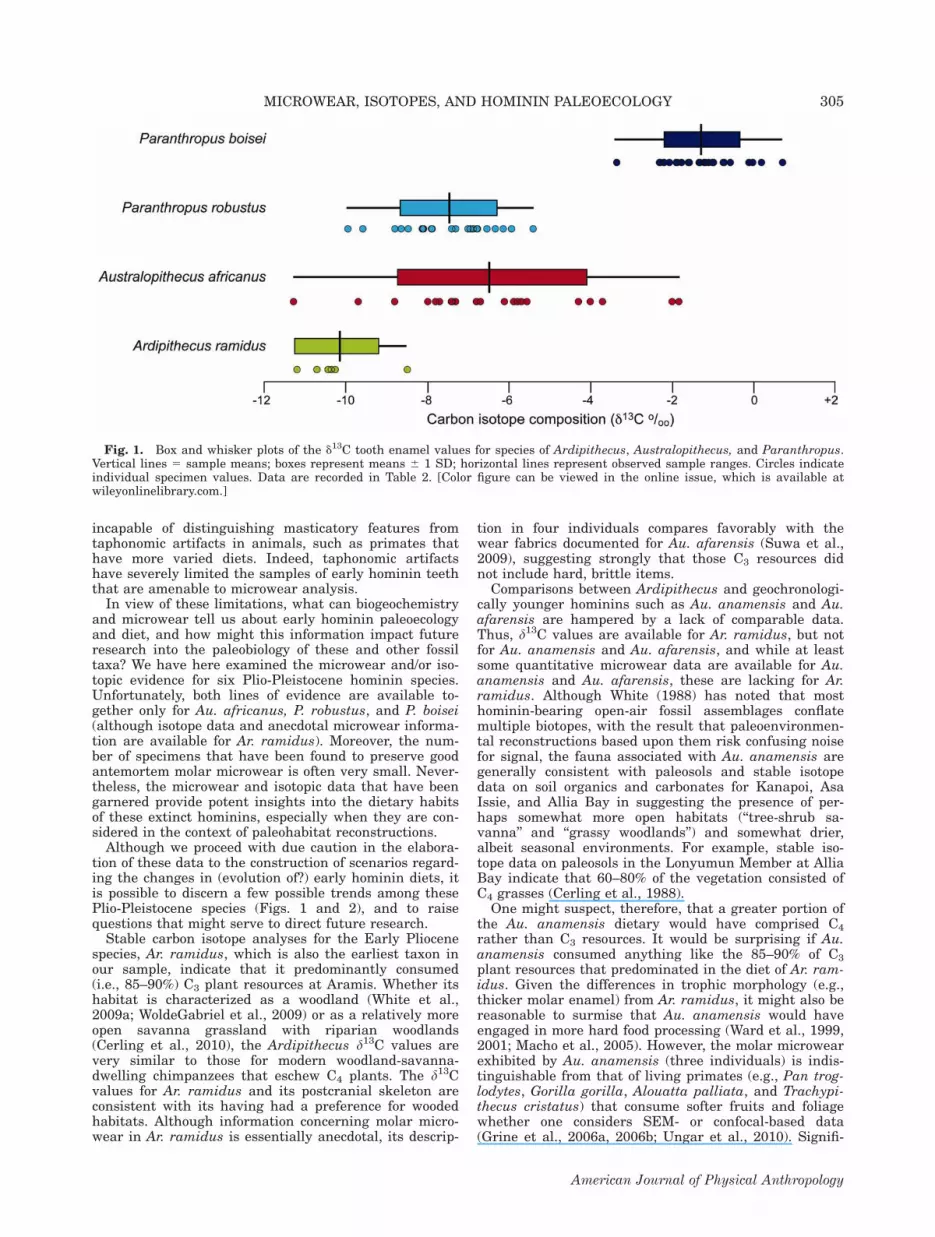
incapable of distinguishing masticatory features fromtaphonomic artifacts in animals, such as primates thathave more varied diets. Indeed, taphonomic artifactshave severely limited the samples of early hominin teeththat are amenable to microwear analysis.In view of these limitations, what can biogeochemistry
and microwear tell us about early hominin paleoecologyand diet, and how might this information impact futureresearch into the paleobiology of these and other fossiltaxa? We have here examined the microwear and/or iso-topic evidence for six Plio-Pleistocene hominin species.Unfortunately, both lines of evidence are available to-gether only for Au. africanus, P. robustus, and P. boisei(although isotope data and anecdotal microwear informa-tion are available for Ar. ramidus). Moreover, the num-ber of specimens that have been found to preserve goodantemortem molar microwear is often very small. Never-theless, the microwear and isotopic data that have beengarnered provide potent insights into the dietary habitsof these extinct hominins, especially when they are con-sidered in the context of paleohabitat reconstructions.Although we proceed with due caution in the elabora-
tion of these data to the construction of scenarios regard-ing the changes in (evolution of?) early hominin diets, itis possible to discern a few possible trends among thesePlio-Pleistocene species (Figs. 1 and 2), and to raisequestions that might serve to direct future research.Stable carbon isotope analyses for the Early Pliocene
species, Ar. ramidus, which is also the earliest taxon inour sample, indicate that it predominantly consumed(i.e., 85–90%) C3 plant resources at Aramis. Whether itshabitat is characterized as a woodland (White et al.,2009a; WoldeGabriel et al., 2009) or as a relatively moreopen savanna grassland with riparian woodlands(Cerling et al., 2010), the Ardipithecus d13C values arevery similar to those for modern woodland-savanna-dwelling chimpanzees that eschew C4 plants. The d13Cvalues for Ar. ramidus and its postcranial skeleton areconsistent with its having had a preference for woodedhabitats. Although information concerning molar micro-wear in Ar. ramidus is essentially anecdotal, its descrip-
tion in four individuals compares favorably with thewear fabrics documented for Au. afarensis (Suwa et al.,2009), suggesting strongly that those C3 resources didnot include hard, brittle items.Comparisons between Ardipithecus and geochronologi-
cally younger hominins such as Au. anamensis and Au.afarensis are hampered by a lack of comparable data.Thus, d13C values are available for Ar. ramidus, but notfor Au. anamensis and Au. afarensis, and while at leastsome quantitative microwear data are available for Au.anamensis and Au. afarensis, these are lacking for Ar.ramidus. Although White (1988) has noted that mosthominin-bearing open-air fossil assemblages conflatemultiple biotopes, with the result that paleoenvironmen-tal reconstructions based upon them risk confusing noisefor signal, the fauna associated with Au. anamensis aregenerally consistent with paleosols and stable isotopedata on soil organics and carbonates for Kanapoi, AsaIssie, and Allia Bay in suggesting the presence of per-haps somewhat more open habitats (‘‘tree-shrub sa-vanna’’ and ‘‘grassy woodlands’’) and somewhat drier,albeit seasonal environments. For example, stable iso-tope data on paleosols in the Lonyumun Member at AlliaBay indicate that 60–80% of the vegetation consisted ofC4 grasses (Cerling et al., 1988).One might suspect, therefore, that a greater portion of
the Au. anamensis dietary would have comprised C4
rather than C3 resources. It would be surprising if Au.anamensis consumed anything like the 85–90% of C3
plant resources that predominated in the diet of Ar. ram-idus. Given the differences in trophic morphology (e.g.,thicker molar enamel) from Ar. ramidus, it might also bereasonable to surmise that Au. anamensis would haveengaged in more hard food processing (Ward et al., 1999,2001; Macho et al., 2005). However, the molar microwearexhibited by Au. anamensis (three individuals) is indis-tinguishable from that of living primates (e.g., Pan trog-lodytes, Gorilla gorilla, Alouatta palliata, and Trachypi-thecus cristatus) that consume softer fruits and foliagewhether one considers SEM- or confocal-based data(Grine et al., 2006a, 2006b; Ungar et al., 2010). Signifi-
Fig. 1. Box and whisker plots of the d13C tooth enamel values for species of Ardipithecus, Australopithecus, and Paranthropus.Vertical lines 5 sample means; boxes represent means 6 1 SD; horizontal lines represent observed sample ranges. Circles indicateindividual specimen values. Data are recorded in Table 2. [Color figure can be viewed in the online issue, which is available atwileyonlinelibrary.com.]
305MICROWEAR, ISOTOPES, AND HOMININ PALEOECOLOGY
American Journal of Physical Anthropology

cantly, the wear fabrics of Au. anamensis provide no evi-dence for the consumption of hard, brittle objects. Hypo-geous tubers, bulbs, and roots that are taken by eclecticand opportunistic feeders like baboons may have consti-tuted part of the dietary repertoire of Au. anamensis,and while the very limited microwear evidence cannotfalsify rare or fallback hard-object feeding, it provides noevidence for it (Grine et al., 2006a; Ungar et al., 2010).At the same time, it is intriguing that the molar
microwear exhibited by Au. anamensis (three individu-als) is indistinguishable from that of Au. afarensis,whether one considers SEM- or confocal-based data(Grine et al., 2006a,b; Ungar et al., 2010), and thatSuwa et al. (2009) have observed that the molar micro-wear in Ardipithecus (four individuals) tends to exhibit‘‘finer and more randomly oriented striae’’ than in (eight)Hadar specimens of Au. afarensis. Insofar as the molarmicrowear fabrics of the Au. afarensis sample examinedby Grine et al. (2006b) and Ungar et al. (2010) also pro-vide no evidence for the consumption of hard, brittleitems, and suggest Gorilla and/or Theropithecus as thebest modern analogs for dietary preference in this Plio-cene hominin, it would appear that the morphologicalchange that characterized evolution in the putative Au.anamensis—Au. afarensis lineage (Kimbel et al., 2004;Ward et al., 2010; Haile-Selassie et al., 2010b) did notinvolve the increased reliance upon or utilization of hardfood items (Fig. 2).Indeed, with reference to Au. afarensis, the number of
specimens sampled for microwear analysis, togetherwith their temporal depth and apparent spread amongdifferent paleohabitats, would seem to make this the ear-
liest taxon for which we might begin to get more broadlyapplicable results. The similarity in Au. afarensis dentalmicrowear across time and space may mean that thisspecies was able to track preferred dietary resources inthe face of altering habitats and environments, or thatenvironmentally induced shifts in diet did not involvechanges in mechanical properties of the foods that weretypically consumed (Grine et al., 2006b). Unfortunately,that portion of the geological range of Au. afarensis (theKada Hadar-2 submember of the Hadar Formation) inwhich a notable increase in mandibular size has beendocumented (Lockwood et al., 2000) has yet to yieldspecimens with antemortem microwear.As with Au. anamensis, no work has been published
on stable light isotope analyses of the teeth of this taxon.However, in light of the paleohabitats that have beenreconstructed for Au. afarensis, it might be suspectedthat a greater portion of its diet, like that of Au. ana-mensis, would have comprised a greater proportion of C4
resources than in Ar. ramidus. Here too, it would bevery surprising if Au. afarensis consumed anything likethe 85–90% of C3 plant resources that predominated inthe diet of Ar. ramidus.Thus, for the geochronologically older part of the homi-
nin species sample in East Africa, our comparisons arebased on combining isotopic data and anecdotal observa-tions about molar microwear in Ar. ramidus, with micro-wear data (however spotty) for Au. anamensis and Au.afarensis. Nevertheless, given the paleohabitats thathave been reconstructed for these three taxa, it may bepossible to provide informed speculation about someaspects of their dietary ecology. In the first instance,
Fig. 2. Box and whisker plots of the complexity (Asfc) component of occlusal microwear for species of Australopithecus and Par-anthropus. Vertical lines 5 sample means; boxes represent means 6 1 SD; horizontal lines represent observed sample ranges.Circles indicate individual specimen values. Data are recorded in Table 5. [Color figure can be viewed in the online issue, which isavailable at wileyonlinelibrary.com.]
306 F.E. GRINE ET AL.
American Journal of Physical Anthropology

Ar. ramidus subsisted on a diet that consisted largely ofsoft or tough C3 foods that would have been available ina wooded environment. Despite habitat shifts to gener-ally more open conditions that would presumably haveindicated a greater abundance and reliance on C4-basedresources, the molar microwear fabrics exhibited by Au.anamensis and Au. afarensis are seemingly like that ofAr. ramidus in being inconsistent with processing hardfoods. These fabrics are somewhat at odds with the mor-phological changes that have been interpreted as havingenhanced their masticatory capabilities to deal withsuch resources. Rather, microwear in Au. anamensis andAu. afarensis is consistent with diets that were perhapsmost similar to those of chimpanzees and gorillas interms of the mechanical properties of the foods con-sumed. It would seem that even if there was a shift tomore open habitats and greater dietary reliance on C4
plants in the Pliocene hominins in East Africa, the me-chanical properties of foods chosen by Ardipithecus andearly australopiths did not differ that much.Certainly a dramatic shift from C3- to C4-based plant
foods occurred at some point (or at several points)between 4.4 Ma and 2.3 Ma among East African austral-opiths. Thus, d13C values reveal that whereas Ar. rami-dus had a diet that consisted of some 85–90% C3 resour-ces, the diet of P. boisei comprised, on average, nearly80% C4 plants. This is not to suppose that we envisionan anagenetic lineage from Ar. ramidus to P. boisei;rather we simply note that at some time (or perhaps atseveral points in time) between 4.4 Ma and 2.3 Ma inEast Africa, there was a radical shift in hominin dietaryhabits (Fig. 1).An anagenetic evolutionary relationship between P.
aethiopicus and its descendant P. boisei is reasonablywell-documented (Walker et al., 1986; Walker and Lea-key, 1988; Kimbel et al., 1988; Suwa, 1988; Wood et al.,1994). If this apparent lineage evolved from Au. afaren-sis or an Au. afarensis-like ancestor, as has been sug-gested by some (e.g., Rak et al., 2007), then it might bereasonable to suggest that the transition to a dietstrongly dominated by C4 plants occurred somewherealong this line.It is interesting to speculate whether this occurred
only with the emergence of P. boisei, whether it beganwith a transitional diet in P. aethiopicus, or whetherthere was a gradual shift along this lineage over the fullcourse of two million years. What will the carbon isotopesignatures of Au. anamensis, Au. Afarensis, and P.aethiopicus look like compared to those of Ardipithecusand P. boisei? Additionally, if there is a steady trendamong these species from C3 to C4, does it fluctuate ran-domly among them, or is P. boisei a unique outlier inEast Africa?What is abundantly apparent, however, is that despite
a profound shift in the plant base of its diet, P. boisei didnot choose C4 items that were hard or brittle. Rather, itsmicrowear signature differs little (if at all) from thosedocumented for Au. anamensis and Au. afarensis (Fig.1). Certainly the specimens from Koobi Fora, Cheso-wanja, and Olduvai Gorge documented by SEM (Walker,1981) and confocal microscopy (Ungar et al., 2008, inpress) and those from Konso examined by SEM (Suwa etal., 1997) do not display the complex, pitted surfacescharacteristic of primates that habitually or even season-ally ingest hard objects.If we turn our attention to South Africa, the geochro-
nologically earliest factual (rather than inferential) indi-
cation of an increased component of C4 vegetation in thediet of an australopith pertains to Au. africanus. Indeed,Au. africanus provides our first reasonable samples forboth isotopic and microwear analyses (Figs. 1 and 2).The latter point to a diet of more variable mechanicalproperties than any earlier australopith and, impor-tantly, this seems to match with stable isotope resultsindicating, for the first time, a significant incorporationof C4 plants in the diet and unprecedented variability ind13C values within and between individuals. When thisis combined with Sr/Ba ratios and d18O values suggest-ing the possible consumption of USO’s, we observe thefirst signs of a consistent dietary change in the earlyhominin record.Paranthropus robustus provides an intriguing mixture
of isotopic and microwear signatures (Figs. 1 and 2). Iso-topically, this species is similar to Au. africanus, whichinhabited the same geographic region just prior to theappearance of the ‘‘robust’’ australopiths. Carbon isotopeanalyses suggest a similar proportion of C3-C4 plants inthe diets of both species, but with slightly less variabilityin d13C values in P. robustus. Specimens of both speciesindicate that the diet was also variable along an ontoge-netic trajectory. At the same time, however, SEM andconfocal data relating to microwear provide consistentevidence that P. robustus did indeed consume hardobjects, at least on occasion within the context of a vari-able diet. The overlap in microwear patterns betweenthese taxa has also yielded clear indications of dietaryoverlap between them, with Au. africanus probablyresorting to tougher foods at times of preferred-food scar-city, and P. robustus resorting to harder foods at suchtimes. This should not be surprising in light of the rangeof isotope work done on these taxa. Observations on theSouth African species suggest that their d18O values arelow compared to other fauna (with the exception of car-nivores) (Sponheimer and Lee-Thorp, 1999b; Lee-Thorpet al., 2010), and the same holds for P. boisei in the Tur-kana Basin (Cerling et al., 2011). While these compara-tive values contrast with those recorded for Ar. ramidus(White et al., 2009a), the causes of these relatively lowvalues remain obscure.
The diet-dietary adaptation conundrum
The microwear and stable carbon isotope results forseveral australopith species have raised questions abouttraditional interpretations of morphology. Indeed, thedata for P. boisei leave us with a remarkable paradox. Inessence, its morphological adaptations suggest hardobject (nut and seed) processing, whereas the microwearand isotope data suggest possibly prolonged masticatorybouts involving tough, abrasive C4 plants. The micro-wear and isotopic signatures for this species are consist-ent in suggesting a diet high in C4 grasses, C4 sedgessuch as Cyperus papyrus, or some combination thereof.Indeed, this interpretation is consistent with the overallpicture that emerges for the preferred habitat of P. boiseias comprising grasslands, usually in association withabundant water.The question then becomes whether we are able to
resolve such a paradox. As noted by many workers (see,for example, the contributions in Plavcan et al., 2002),the best approach is to use multiple lines of evidence todetermine whether any consistent interpretationsemerge. The traditional comparative morphologicalapproach is ultimately based on biomechanical models
307MICROWEAR, ISOTOPES, AND HOMININ PALEOECOLOGY
American Journal of Physical Anthropology

and assumptions about evolutionary mechanisms. Thisindirect approach has given us the time-honored imageof P. boisei as a creature capable of generating (and dis-sipating) large masticatory forces in the pursuit of punc-ture-resistant foods. However, two very different formsof data—from dental microwear and stable isotope analy-ses—provide results that should give us pause forthought. While both of these approaches have underlyingassumptions and limitations, they have independentlyyielded corroborative results. Assuming that the resultsfrom both methods can be taken at face value, the ques-tion to be addressed is why might the traditional view ofP. boisei be incorrect?To make inferences about the craniodental adaptations
of this (or any extinct) creature, researchers need tobuild upon close, replicable associations between formand function in extant taxa (Kay and Cartmill, 1977;Ross et al., 2002), but the measurement of both may befraught with difficulties (Smith, 1996; Plavcan et al.,2002). Even if a one-to-one correspondence between formand function were within our grasp for something likedental morphology, what is most striking about P. boiseiis the combination of features in its cranial vault, face,palate, mandible, and teeth. It is a unique combina-tion—certainly one that is not seen (or probably evenapproximated) in any living species. Deciphering thefunction(s) of such a suite of characters requires anunderstanding of the biomechanical basis for every mor-phological idiosyncrasy in its anatomy. For example, con-sider the large, low-relief molars of P. boisei. Theoreti-cally, one might expect them to have been ideally suitedfor processing soft fruits and small seeds (Lucas, 2004).However, while experimental work has shown that ani-mals with more occlusal relief can subdivide tough foodsmore finely than can animals with less occlusal relief(e.g., Sheine and Kay, 1977), many leaves are not tough,and whether large flat teeth can process leaves of vary-ing properties is an unknown. Moreover, perhaps thecraniodental features of Paranthropus could have facili-tated a unique way of grinding tougher foods (Rak andHylander, 2008), which might facilitate a corroborativelink between traditional morphological interpretationsand those from dental microwear and stable isotopeanalyses. As another example, the shape of the mandibu-lar symphysis and corpus of P. boisei has traditionallybeen thought to have provided resistance to powerfulbending and twisting moments (Hylander, 1988). This,in conjunction with the aforementioned assumptionsabout dental morphology, swung the focus of discussionto the processing of hard objects. Yet, as noted byHylander himself (1988), such a symphysis could just aseasily have resisted the strains associated with process-ing tough foods. Finer-resolution analyses of the spatialpatterning of morphological properties in the jaw (e.g.,Daegling et al., 2011a) may lead to a better understand-ing of mandibular responses to different feeding regimes.This, in turn, may lead to a better link between morpho-logical interpretations and those from dental microwearand stable isotope analyses. These sorts of insights couldbe garnered from the full range of features exhibited byP. boisei.Thus, even though a great deal of work has been done
on the australopiths, we are only beginning to understandhow each of the components of their craniodental anato-mies might have functioned. For example, finite elementanalysis (FEA) has the potential to yield new perspectiveson the way forces are dissipated in the skull (Huiskes and
Chao, 1983; Rayfield, 2007; Strait et al., 2009; Wroe et al.,2010), but as a model it is only as strong as its underlyingassumptions (Grine et al., 2010). Moreover, its use in theinterpretation of extinct and morphologically unique fos-sil species requires a number of crucial assumptions. Itwould be folly to portray FEA or any other biomechanicalmethod as the ultimate arbiter of australopith functionalmorphology and adaptation. Finite element modeling canbe a powerful tool, and one source of information pertain-ing to anatomical structure, but ultimately it is a tooldesigned to test hypotheses about how structures behavebiomechanically under certain conditions. Like every tool,it has its strengths as well as its limitations. Recognizingthese enables us to move to meaningful discussions ofhow such morphological complexes might develop evolu-tionarily.Arguments for the evolution of morphological differen-
tiation have customarily involved inferences about thepossible survival/reproduction benefits of specific pheno-typic traits or feature complexes that are coupled withassumptions about the genetic basis for their expression.Relating morphological differences to differential sur-vival and/or reproduction is difficult albeit not impossible(see King et al., 2005), but all too often this is manifestsimply as an intuitively pleasing ‘‘just so’’ story. Underly-ing it all has been the debate over whether it is frequentuse, or critical use, that determines which roles are themost important for the evolution of morphological differ-ences. It has long been known that the most importantrole for a trait may not be the one performed most fre-quently (Kinzey, 1978; Rosenberger and Kinzey, 1976;Stern and Oxnard, 1973). However, does that mean thatfrequent use may not be just as important in other situa-tions (Kay and Hiiemae, 1974; Kay and Hylander, 1978)?More than likely, it is not an either-or alternative, andthe answer will differ for individual species and circum-stances. In some situations, animals may be forced toresort to foods that test the physical limits of their food-processing ability in order to survive; in other instances,it may be the most frequently eaten foods which allowthem to attain sufficient body mass to survive duringtimes of food scarcity. The key point for the present dis-cussion is that in order to sort through these alterna-tives, we need every possible indicator of what extinctanimals actually did during their lifetimes.We have attempted here to summarize evidence from
two such sources—biogeochemistry and occlusal micro-wear—and both provide corroborative results. Someearly hominins were not doing what we might haveexpected based on inferences from functional morphol-ogy. As a result, we need to take a closer look at whatwe know and do not know about the functional morphol-ogy of the masticatory apparatus in extant taxa.
CONCLUSIONS
Determining the diet of an extinct species is para-mount in any attempt to reconstruct its paleoecology.Because the distribution and mechanical properties offood items may impact postcranial, cranial, mandibular,and dental morphologies related to their procurement,ingestion, and mastication, these anatomical attributeshave been studied intensively. However, while mechani-cal environments influence skeletal and dental features,it is not clear to what extent they dictate particular mor-phologies. While biomechanical explanations have beenwidely applied to extinct hominins in attempts to retro-
308 F.E. GRINE ET AL.
American Journal of Physical Anthropology

dict dietary proclivities, morphology may say as muchabout what they were capable of eating and about phylo-genetic history than about the nature of the diet. Whileanatomical attributes may establish boundary limits,direct evidence left by the foods that were actually(rather than hypothetically) consumed is required toreconstruct diet.Dental microwear and the stable light isotope chemis-
try of tooth enamel provide such evidence, and are espe-cially powerful when used in tandem. We review thefoundations for microwear and biogeochemistry in dietreconstruction, and discuss this evidence for six earlyhominin species—Ar. ramidus, Au. anamensis, Au. afar-ensis, Au. africanus, P. robustus, and P. boisei. The die-tary signals derived from microwear and isotope chemis-try are sometimes at odds with inferences from biome-chanical approaches. This potentially disquietingconundrum is particularly evident for several species,and highlights the need for additional biomechanicalanalyses of living animals for its resolution.
ACKNOWLEDGMENTS
FEG, PSU, and MFT thank the National Museums ofKenya, the National Museum of Ethiopia, the DitsongMuseum of Natural History, Pretoria and the Universityof the Witwatersrand, Johannesburg for permission toexamine specimens in their care. Authors thank W.H.Kimbel and D.C. Johanson for access to unpublishedHadar fossils. Authors also thank the curators at theUnited States National Museum, the Cleveland Museumof Natural History, and the Ditsong Natural History Mu-seum for access to extant primate collections. FEG thanksJ.F. Gogarten for discussions relating to seasonality ofdeath among primates, and PSU thanks A. Perez-Perezand L. Martınez for suggesting that OH 74 be re-exam-ined. All authors are grateful to Frank Cuozzo andMichelle Sauther for co-organizing a stimulating and col-legial symposium at the 2010 Annual Meeting of theAmerican Association of Physical Anthropologists, fromwhich this contribution evolved, and for their patienceover the completion of the manuscript. Authors wouldlike to express their gratitude to C. Ruff, the associateeditor and two anonymous reviewers for their cogent andinsightful comments on this manuscript.
LITERATURE CITED
Ahern JCM. 1998. Underestimating intraspecific variation: theproblem with excluding Sts 19 from Australopithecus africa-nus. Am J Phys Anthropol 105:461–480.
Alemseged Z, Spoor F, Kimbel WH, Bobe R, Geraads D, Reed D,Wynn JG. 2006. A juvenile early hominin skeleton fromDikika, Ethiopia. Nature 443:296–301.
Alemseged Z, Wynn JG, Kimbel WH, Reed D, Geraads D, BobeR. 2005. A new hominin from the Basal member of the HadarFormation, Dikika, Ethiopia, and its geological context. JHum Evol 49:499–514.
AlQahtani SJ, Hector MP, Liversidge HM. 2010. The Londonatlas of human tooth development and eruption. Am J PhysAnthropol 142:481–490.
Ambrose SH. 2000. Controlled diet and climate experiments onnitrogen isotope ratios of rats. In: Ambrose SH, KatzenbergMA, editors. Biogeochemical approaches to paleodietary anal-ysis. New York: Kluwer Plenum. p 243–259.
Ambrose SH, DeNiro MJ. 1986. Reconstruction of Africanhuman diet using bone collagen carbon and nitrogen isotoperatios. Nature 319:321–324.
Ambrose SH, Norr L. 1993. Experimental evidence for the rela-tionship of the carbon isotope ratios of whole diet and dietaryprotein to those of bone collagen and carbonate. In: LambertJB, Grupe G, editors. Prehistoric human bone: archaeology atthe molecular level. New York: Springer-Verlag. p 1–37.
Anthony MRL, Kay RF. 1993. Tooth form and diet in atelineand alouattine primates: reflections on the comparativemethod. Am J Sci 293A:356–382.
Ardu S, Benbachir N, Stavridakis M, Dietschi D, Krejci I,Feilzer A. 2009. A combined chemo-mechanical approach foraesthetic management of superficial enamel defects. Br DentJ 206:205–208.
Avery DM. 2001. The Plio-Pleistocene vegetation and climate ofSterkfontein and Swartkrans, South Africa, based on micro-mammals. J Hum Evol 41:113–132.
Baab KL. 2008. A re-evaluation of the taxonomic affinities ofthe early Homo cranium KNM-ER 42700. J Hum Evol55:741–746.
Babbitt CC, Warner LR, Fedrigo O, Wall CE, Wray GA. 2010.Genomic signatures of diet-related shifts during human ori-gins. Proc R Soc B 278:961–969.
Baker G, Jones LHP, Wardrop ID. 1959. Cause of wear insheep’s teeth. Nature 184:1583–1584.
Balasse M. 2003. Reconstructing dietary and environmental his-tory from enamel isotopic analysis: time resolution of intra-tooth sequential sampling. Int J Osteoarchaeol 12:155–165.
Balter V, Bocherens H, Person A, Labourdette N, Renard M,Vandermeersch B. 2002. Ecological and physiological variabili-ty of Sr/Ca and Ba/Ca in mammals of West European mid-Wurmian food webs. Paleogeogr Paleoclimatol Paleoecol186:127–143.
Bamford M. 1999. Pliocene fossil woods from an early hominidcave deposit, Sterkfontein, South Africa. S Afr J Sci 95:231–237.
Benefit BR, McCrossin ML. 1990. Diet, species diversity, anddistribution of African fossil baboons. Kroeber Anthropol SocPap 71:79–93.
Binning SA, Chapman LJ, Cosandey-Godin A. 2009. Specializedmorphology for a generalist diet: evidence for Liem’s Paradoxin a cichlid fish. J Fish Biol 75:1683–1699.
Bishop WW, Hill A, Pickford M. 1978. Chesowanja: a revised ge-ological interpretation. In: Bishop WW, editor. Geologicalbackground to fossil man. London: Scottish Academic Press. p309–327.
Blomberg SP, Garland T. 2002. Tempo and mode in evolution:phylogenetic inertia, adaptation and comparative methods. JEvol Biol 15:899–910.
Blumenschine RJ, Peters CR, Masao FT, Clarke RJ, Deino AL,Hay RL, Swisher CC, Stanistreet IG, Ashley GM, McHenryLJ, Sikes NE, van der Merwe NJ, Tactikos JC, Cushing AE,Deocampo DM, Njau JK, Ebert JI. 2003. Late Pliocene Homoand hominid land use from western Olduvai Gorge, Tanzania.Science 299:1217–1221.
Boaz NT, Hampel J. 1978. Strontium content of fossil toothenamel and diet of early hominids. J Paleontol 52:928–933.
Bocherens H, Billiou D, Mariotti A, Patou-Mathis M, Otte M,Bonjean D, Toussaint M. 1999. Palaeoenvironmental andpalaeodietary implications of isotopic biogeochemistry of lastinterglacial Neandertal and mammal bones in Scladina Cave(Belgium). J Archaeol Sci 26:599–607.
Bocherens H, Fizet M, Mariotti A, Lange-Badre B, Vander-meersch B, Borel JP, Bellon G. 1991. Isotopic biochemistry(13C, 15N) of fossil vertebrate collagen: implications for thestudy of fossil food web including Neandertal man. J HumEvol 20:481–492.
Bonnefille R. 1985. Evolution of the continental vegetation: thepalaeoecological record for East Africa. S Afr J Sci 81:267–270.
Bonnefille R. 1995. A reassessment of the Plio-Pleistocene pollenrecord of East Africa. In: Vrba ES, Denton GH, Partridge TC,Burckle LH, editors. Paleoclimate and evolution with empha-sis on human origins. New Haven, CT: Yale University Press.p 299–310.
Bowen HJM, Dymond JA. 1955. Strontium and barium inplants and soils. Phil Trans R Soc Lond B 144:355–368.
309MICROWEAR, ISOTOPES, AND HOMININ PALEOECOLOGY
American Journal of Physical Anthropology

Broom R, Robinson JT. 1952. Swartkrans Ape-man Paranthro-pus crassidens. Pretoria: Transvaal Museum Memoir, no. 6.
Brown AB. 1974. Bone strontium as a dietary indicator inhuman skeletal populations. Contrib Geol 13:47–48.
Brown B, Brown FH, Walker A. 2001. New hominids from theLake Turkana Basin, Kenya. J Hum Evol 41:29–44.
Buchet L, Cremoni N, Rucker C, Verdin P. 2001. Comparisonbetween the distribution of dental micro-striations and plantmaterial included in the calculus of human teeth. In: MeunierJD, Colin F, editors. Phytoliths: applications in earth sciencesand human history. Rotterdam: A.A. Balkema. p 107–117.
Burton JH, Price TD, Middleton WD. 1999. Correlation of boneBa/Ca and Sr/Ca due to biological purification of calcium. JArchaeol Sci 26:609–616.
Burton JH, Wright LE. 1995. Nonlinearity in the relationshipbetween bone Sr/Ca and diet: paleodietary implications. Am JPhys Anthropol 96:273–282.
Butzer KW. 1974. Paleoecology of the South African australopi-thecines: Taung revisited. Curr Anthropol 15:367–382.
Cachel S. 1975. A new view of speciation in Australopithecus.In: Tuttle R, editor. Paleoanthropology, morphology and paleo-ecology. The Hague: Mouton. p 183–201.
Cadman A, Rayner R. 1989. Climatic change and the appear-ance of Australopithecus africanus in the Makapansgat sedi-ments. J Hum Evol 18:107–113.
Campisano CJ, Feibel CS. 2008. Depositional environments andstratigraphic summary of the Pliocene Hadar Formation atHadar, Afar Depression, Ethiopia. Geol Soc Am Special Pap446:179–201.
Carter B. 2006. Paleoecological reconstructions of the SouthAfrican Plio-Pleistocene based on low-magnification dentalmicrowear of fossil primates. MA Thesis, Georgia State Uni-versity.
Carter ML. 2001. Sensitivity of stable isotopes (13C, 15N, and18O) in bone to dietary specialization and niche separationamong sympatric primates in Kibale National Park, Uganda.PhD Dissertation, University of Chicago.
Cerling TE, Bowman JR, O’Neil JR. 1988. An isotopic study of afluvial-lacustrine sequence: the Plio-Pleistocene Koobi-Forasequence, East Africa. Palaeogeogr Palaeoclimatol Palaeoecol63:335–256.
Cerling TE, Harris JM. 1999. Carbon isotope fractionationbetween diet and bioapatite in ungulate mammals and impli-cations for ecological and paleoecological studies. Oecologia120:247–263.
Cerling TE, Harris JM, Leakey MG. 1999. Browsing and graz-ing in elephants: the isotope record of modern and fossil pro-boscideans. Oecologia 120:364–374.
Cerling TE, Hay RL. 1986. An isotopic study of paleosol carbo-nates from Olduvai Gorge. Quatern Res 25:63–78.
Cerling TE, Levin NE, Quade J, Wynn JG, Fox DL, KingstonJD, Klein RG, Brown FH. 2010. Comment on the paleoenvir-onment of Ardipithecus ramidus. Science 328:1105.
Cerling TE, Mbua E, Kirera FM, Manthi FK, Grine FE, LeakeyMG, Sponheimer M, Uno K. 2011. Diet of Paranthropus boiseiin the early Pleistocene of East Africa. Proc Natl Acad SciUSA 108:9337–9341.
Chapman CA, Speirs ML, Hodder SAM, Rothman JM. 2010.Colobus monkey parasite infections in wet and dry habitats:implications for climate change. Afr J Ecol 48:555–558.
Cheney DL, Seyfarth RM, Fischer J, Beehner J, Bergman T,Johnson SE, Kitchen DM, Palombit RA, Rendall D, Silk JB.2004. Factors affecting reproduction and mortality amongbaboons in the Okavango Delta, Botswana. Int J Primatol25:401–428.
Cheng ZJ, Wang XM, Cui FZ, Yan JX. 2009. The enamel soften-ing and loss during early erosion studied by AFM, SEM andnanoindentation. Biomed Mater 4:015020.
Cheverud J. 1988. A comparison of genetic and phenotypic cor-relation. Evolution 42:958–968.
Clutton-Brock TH, Harvey PH. 1977. Primate ecology and socialorganization. J Zool 183:1–39.
Codron D, Lee-Thorp JA, Sponheimer M, de Ruiter D, CodronJ. 2006. Inter- and intra-habitat dietary variability of Chacma
baboons (Papio ursinus) in South African savannas based onfecal d13C, d15N and %N. Am J Phys Anthropol 129:195–204.
Codron D, Luyt J, Lee-Thorp JA, Sponheimer M, de Ruiter D,Codron J. 2005. Utilization of savanna based resources byPlio-Pleistocene baboons. S Afr J Sci 101:245–248.
Codron DM. 2003. Dietary ecology of chacma baboons (Papioursinus (Kerr, 1792)) and Pleistocene cercopithecoidea in sa-vanna environments of South Africa. M.Sc. Thesis, Universityof Cape Town.
Coffing K, Feibel C, Leakey M, Walker A. 1994. Four-million-year-old hominids from East Lake Turkana Kenya. Am JPhys Anthropol 93:55–65.
Constantino PJ, Lee JJW, Morris D, Lucas PW, Hartstone-RoseA, Lee WK, Dominy NJ, Cunningham A, Wagner M, LawnBR. 2011. Adaptation to hard-object feeding in sea otters andhominins. J Hum Evol 61:89–96.
Cuddahee R, Madden R, Churchill S, Bobe R. 2010. Non-dietaryabrasives and the evolution of hominin megadonty. Am JPhys Anthropol 141 (Suppl 50):88–89.
Daegling DJ. 1989. Biomechanics of cross-sectional size andshape in the hominoid mandibular corpus. Am J Phys Anthro-pol 80:91–106.
Daegling DJ. 1992. Mandibular morphology and diet in the ge-nus Cebus. Int J Primatol 13:545–570.
Daegling DJ, Granatosky MC, McGraw WS, Rapoff AJ. 2011a.Spatial patterning of bone stiffness variation in the colobinealveolar process. Archs Oral Biol 56:220–230.
Daegling DJ, Grine FE. 1991. Compact bone distribution andbiomechanics of early hominid mandibles. Am J Phys Anthro-pol 86:321–339.
Daegling DJ, Grine FE. 1999. Terrestrial foraging and dentalmicrowear in Papio ursinus. Primates 40:559–72.
Daegling DJ, Grine FE. 2007. Mandibular biomechanics and thepaleontological evidence for the evolution of human diet. In:Ungar PS, editor. Evolution of the human diet: the known,the unknown, and the unknowable. New York: Oxford Univer-sity Press. p 106–131.
Daegling DJ, McGraw WS. 2001. Feeding, diet and jaw form in WestAfrican Colobus and Procolobus. Int J Primatol 22:1033–1055.
Daegling DJ, McGraw WS, Ungar PS, Pampush JD, Vick AE,Bitty EA. 2011b. Hard object feeding in sooty mangabeys(Cercocebus atys) and the interpretation of early homininfeeding ecology. PLoS One 6:e23095 (1–12).
Darling WG, Bath AH, Gibson JJ, Rozanski K. 2005. Isotopes inwater. In: Leng MJ, editor. Isotopes in palaeoenvironmentalresearch. Dordrecht: Springer. p 1–66.
Deino A. 2011. 40Ar/39Ar dating of Laetoli, Tanzania. In: Harri-son T, editor. Paleontology and geology of Laetoli: human evo-lution in context. Vol.1: Geology, geochronology, paleoecologyand paleoenvironment. Dordrecht: Springer. p 77–97.
Deino A, Scott GR, Saylor B, Alene M, Angelini J, Haile-Selas-sie Y. 2010. 40Ar/39Ar dating, paleomagnetism, and tephros-tratigraphy of Pliocene strata of the hominid-bearing Wor-anso-Mille area, west-central Afar Rift, Ethiopia. J Hum Evol58:111–126.
Demes B, Creel N. 1988. Bite force, diet, and cranial morphol-ogy of fossil hominids. J Hum Evol 17:657–670.
Dominguez-Rodrigo M, Lopez-Saez JA, Vincens A, Alcala L,Luque L, Serralonga J. 2001. Fossil pollen from the upperHumbu Formation of Peninj (Tanzania): hominid adaptationto a dry open Plio-Pleistocene savanna environment. J HumEvol 40:151–157.
Du Brul EL. 1977. Early hominid feeding mechanisms. Am JPhys Anthropol 47:305–320.
Dunbar RIM. 1980. Demographic and life history variables of apopulation of gelada baboons (Theropithecus gelada). J Ani-mal Ecol 49:485–506.
Eaglen RH. 1984. Incisor size and diet revisited: the viewfrom a platyrrhine perspective. Am J Phys Anthropol64:263–275.
El-Zaatari S, Grine FE, Teaford MF, Smith HF. 2005. Molarmicrowear and dietary reconstructions of fossil Cercopithecoi-dea from the Plio-Pleistocene deposits of South Africa. J HumEvol 49:180–205.
310 F.E. GRINE ET AL.
American Journal of Physical Anthropology

El Zaatari S, Grine FE, Ungar PS, Hublin J-J. 2011. Ecogeo-graphic variation in Neandertal dietary habits: evidence fromocclusal molar microwear texture analysis. J Hum Evol61:411–424.
Epstein E. 1994. The anomaly of silicon in plant biology. ProcNatl Acad Sci USA 91:11–17.
Estebaranz F, Martınez LM, Galbany J, Turbon D, Perez-PerezA. 2009. Testing hypotheses of dietary reconstruction frombuccal dental microwear in Australopithecus afarensis. J HumEvol 57:739–750.
Ezzo JA. 1994. Putting the ‘‘chemistry’’ back into archaeologicalbone chemistry analysis: modeling potential dietary indica-tors. J Anthropol Archaeol 13:1–34.
Feibel CS, Harris JM, Brown FH. 1991. Palaeoenvironmentalcontext for the Late Neogene of the Turkana Basin. In: HarrisJM, editor. Koobi Fora Research Project, Vol. 3. Oxford:Clarendon Press. p 321–346.
Fernandez-Jalvo Y, Denys C, Andrews P, Williams T, DauphinY, Humphrey L. 1998. Taphonomy and palaeoecology of Oldu-vai Bed I (Pleistocene, Tanzania). J Hum Evol 34:137–172.
Fourie NH, Lee-Thorp JA, Ackermann RR. 2008. Biogeochemi-cal and craniometric investigation of dietary ecology, nicheseparation, and taxonomy of Plio-Pleistocene cercopithecoidsfrom the Makapansgat Limeworks. Am J Phys Anthropol135:121–135.
Fraser D, Mallon JC, Furr R, Theodor JM. 2009. Improving therepeatability of low magnification microwear methods usinghigh dynamic range imaging. Palaios 24:818–825.
Galbany J, Estebaranz F, Martınez LM, Perez-Perez A. 2009.Buccal dental microwear variability in extant African Homi-noidea: taxonomy versus ecology. Primates 50:221–230.
Galbany J, Perez-Perez A. 2004. Buccal enamel microwear vari-ability in Cercopithecoidea primates as a reflection of dietaryhabits in forested and open savannah environments. Anthro-pologie 42:13–19.
Gentry AW, Gentry A. 1978. Fossil Bovidae (Mammalia) of Old-uvai Gorge, Tanzania. Parts 1 and 2. Bull Brit Mus Nat Hist29:289–446;30:1–83.
Godfrey LR, Semprebon GM, Jungers WL, Sutherland MR,Simons EL, Solounias N. 2004. Dental use wear in extinctlemurs: evidence of diet and niche differentiation. J HumEvol 47:145–169.
Godfrey LR, Semprebon GM, Schwartz GT, Burney DA, JungersWL, Flanagan EK, Cuozzo FP, King SJ. 2005. New insightsinto old lemurs: the trophic adaptations of the Archaeolemuri-dae. Int J Primatol 26:825–854.
Gould L, Sussman RW, Sauther ML. 1999. Natural disastersand primate populations: the effects of a 2-year drought ona naturally occurring population of ring-tailed lemurs(Lemur catta) in southwestern Madagascar. Int J Primatol20:69–84.
Gould SJ, Lewontin RC. 1979. The spandrels of San Marcos andthe Panglossian paradigm, a critique of the adaptationist pro-gramme. Proc R Soc Lond B 205:581–598.
Grine FE. 1981. Trophic differences between ‘gracile’ and ‘ro-bust’ australopithecines: a scanning electron microscope anal-ysis of occlusal events. S Afr J Sci 77:203–230.
Grine FE. 1982. A new juvenile hominid (Mammalia: Primates)from Member 3, Kromdraai Formation, Transvaal, SouthAfrica. Ann Transvaal Mus 33:163–239.
Grine FE. 1985. Australopithecine evolution: the deciduous den-tal evidence. In: Delson E, editor. Ancestors: the hard evi-dence. New York: Alan R. Liss. p 153–167.
Grine FE. 1986. Dental evidence for dietary differences in Aus-tralopithecus and Paranthropus: a quantitative analysis ofpermanent molar microwear. J Hum Evol 15:783–822.
Grine FE. 1989. New hominid fossils from the SwartkransFormation (1979–1986 excavations): craniodental specimens.Am J Phys Anthropol 79:409–449.
Grine FE. The alpha taxonomy of Australopithecus africanus.In: Reed KE, Fleagle JG, editors. The paleobiology of Austral-opithecus. Dordrecht: Springer, in press.
Grine FE, Delanty MM, Wood BA. Variation in mandibularpostcanine dental morphology and hominin species represen-
tation in Member 4, Sterkfontein, South Africa. In: Reed K,Fleagle JG, editors. The paleobiology of Australopithecus.Dordrecht: Springer, in press, a.
Grine FE, Fleagle JG. 2009. The first humans: a summary per-spective on the origin and early evolution of the genus Homo.In: Grine FE, Fleagle JG, Leakey RE, editors. The firsthumans: origin and early evolution of the genus Homo.Dordrecht: Springer. p 197–207.
Grine FE, Jacobs RL, Reed KE, Plavcan JM. The enigmaticmolar from Gondolin, South Africa: implications forParanthropus paleobiology. J Hum Evol, in press, b.
Grine FE, Judex S, Daegling DJ, Ozcivici E, Ungar PS, TeafordMF, Sponheimer M, Scott J, Scott RS, Walker A. 2010. Cra-niofacial biomechanics and functional and dietary inferencesin hominin paleontology. J Hum Evol 58:293–308.
Grine FE, Kay RF. 1988. Early hominid diets from quantitativeimage analysis of dental microwear. Nature 333:765–768.
Grine FE, Strait DS. 1994. New hominid fossils from the Mem-ber 1 ‘‘Hanging Remnant’’, Swartkrans Formation, SouthAfrica. J Hum Evol 26:57–75.
Grine FE, Ungar PS, Teaford MF. 2002. Error rates in dentalmicrowear quantification using SEM. Scanning 24:144–153.
Grine FE, Ungar PS, Teaford MF. 2006a. Was the Early Plio-cene hominin ‘Australopithecus’ anamensis a hard objectfeeder? S Afr J Sci 102:301–310.
Grine FE, Ungar PS, Teaford MF, El Zaatari S. 2006b. Molarmicrowear in Praeanthropus afarensis: evidence for dietarystasis through time and under diverse paleoecological condi-tions. J Hum Evol 51:297–319.
Gugel IL, Grupe G, Kunzelmann KH. 2001. Simulation of den-tal microwear: characteristic traces of opal phytoliths giveclues to ancient human dietary behavior. Am J Phys Anthro-pol 114:124–138.
Haile-Selassie Y. 2010. Phylogeny of early Australopithecus:new fossil evidence from the Woranso-Mille (central Afar,Ethiopia). Phil Trans R Soc B 365:3323–3331.
Haile-Selassie Y, Deino A, Saylor B, Umer M, Latimer B. 2007.Preliminary geology and paleontology of new hominid-bearingPliocene localities in the central Afar region of Ethiopia.Anthropol Sci 115:215–222.
Haile-Selassie Y, Latimer BM, Alene M, Deino AL, Gibert L,Melillo SM, Saylor BZ, Scott GR, Lovejoy CO. 2010a. An earlyAustralopithecus afarensis postcranium from Woranso-Mille,Ethiopia. Proc Natl Acad Sci USA 107:12121–12126.
Haile-Selassie Y, Saylor B, Deino A, Alene M, Latimer B. 2010b.New hominid fossils from Woranso-Mille (Central Afar, Ethio-pia) and taxonomy of early Australopithecus. Am J PhysAnthropol 141:406–417.
Haile-Selassie Y, Suwa G, White TD. 2004. Late Miocene teethfrom Middle Awash, Ethiopia, and early hominid dental evolu-tion. Science 303:1503–1505.
Hanson TF, Orzack SH. 2005. Assessing current adaptationand phylogenetic inertia as explanations for trait evolution:the need for controlled comparisons. Evolution 59:2063–2072.
Harrison T, editor. 2011a. Paleontology and geology of Laetoli:human evolution in context. Vol.1: Geology, geochronology,paleoecology and paleoenvironment. Dordrecht: Springer.
Harrison T. 2011b. Hominins from the Upper Laetolil andUpper Ndolanya Beds, Laetoli. In: Harrison T, editor. Paleon-tology and geology of Laetoli: human evolution in context. Vol.2: Fossil hominins and the associated fauna. Dordrecht:Springer. p 141–188.
Hay RL, Kyser TK. 2001. Chemical sedimentology and paleoen-vironmental history of Lake Olduvai, a Pliocene lake in north-ern Tanzania. Geol Soc Am Bull 133:1505–1521.
Hlusko LJ. 2004. Protostylid variation in Australopithecus.J Hum Evol 46:579–594.
Hoppe KA, Koch PL, Furutani TT. 2003. Assessing the preser-vation of biogenic strontium in fossil bones and tooth enamel.Int J Osteoarchaeol 13:20–28.
Howell FC. 1978. Hominidae. In: Maglio VJ, Cooke HBS, edi-tors. Evolution of African mammals. Cambridge, MA: HarvardUniversity Press. p 154–248.
311MICROWEAR, ISOTOPES, AND HOMININ PALEOECOLOGY
American Journal of Physical Anthropology

Huffman M A, Gotoh S, Turner LA, Hamai M, Yoshida K. 1997.Seasonal trends in intestinal nematode infection and medici-nal plant use among chimpanzees in the Mahale Mountains,Tanzania. Primates 38:111–125.
Huiskes R, Chao EYS. 1983. A survey of finite element analysisin orthopedic biomechanics: the first decade. J Biomech16:385–409.
Hummel J, Findeisen E, Sudekum KH, Ruf I, Kaiser TM,Bucher M, Clauss M, Codron D. 2010. Another one bites thedust: faecal silica levels in large herbivores correlate withhigh-crowned teeth. Proc R Soc B 278:1742–1747.
Hurtado AM, Frey MA, Hurtado I, Hill K, Baker J. 2008. Therole of helminthes in human evolution. Implications for globalhealth in the 21st century. In: Elton S, O’Higgins P, editors.Medicine and evolution: current applications, future pros-pects. Boca Raton: CRC Press. p 153–180.
Hylander WL. 1975. Incisor size and diet in anthropoids withspecial reference to Cercopithecidae. Science 189:1095–1098.
Hylander WL. 1988. Implications of in vivo experiments forinterpreting the functional significance of ‘‘robust’’ australopi-thecine jaws. In: Grine FE, editor. Evolutionary history of the‘‘robust’’ australopithecines. New York: Aldine de Gruyter. p55–83.
Jaeger JJ. 1976. Les rongeurs (Mammalia, Rodentia) du Pleisto-cene inferieur d’Olduvai Bed I (Tanzanie), 1ere Partie: LesMurides. In: Savage RJG, Coryndon SC, editors. Fossil verte-brates of Africa 4. London: Academic Press. p 57–120.
Jolly CJ. 1970. The seed eaters: a new model for hominid differ-entiation based on a baboon analogy. Man 5:5–26.
Jones AM, O’Connell TC, Young ED, Scott K, Buckingham CM,Iacumin P, Brasier MD. 2001. Biogeochemical data from wellpreserved 200 ka collagen and skeletal remains. Earth PlanetSci Lett 193:143–149.
Kalisz PJ, Boettcher SE. 1990. Phytolith analysis of soils at Buf-falo Beats, a small forest opening in southeastern Ohio. Tor-rey Bot Club Bull 117:445–447.
Kappelman J. 1984. Plio-Pleistocene environments of Bed I andLower Bed II, Olduvai Gorge, Tanzania. Palaeogeog Palaeocli-matol Palaeoecol 48:171–196.
Kappleman J, Plummer T, Bishop L, Duncan A, Appleton A.1997. Bovids as indicators of Plio-Pleistocene paleoenviron-ments in East Africa. J Hum Evol 32:229–256.
Kay RF. 1978. Molar structure and diet in extant Cercopitheci-dae. In: Butler PM, Joysey KA, editors. Development, func-tion, and evolution of teeth. New York: Academic. p 309–339.
Kay RF. 1985. Dental evidence for the diet of Australopithecus.Ann Rev Anthropol 14:15–41.
Kay RF, Cartmill M. 1977. Cranial morphology and adaptationsof Palaechthon nacimienti and other Paromomyidae (Plesiada-poidea, ?Primates), with description of a new genus and spe-cies. J Hum Evol 6:19–53.
Kay RF, Grine FE. 1988. Tooth morphology, wear and diet inAustralopithecus and Paranthropus from southern Africa. In:Grine FE, editor. Evolutionary history of the ‘‘robust’’ austral-opithecines. New York: Aldine de Gruyter. p 427–448.
Kay RF, Hiiemae KM. 1974. Jaw movement and tooth use inrecent and fossil primates. Am J Phys Anthropol 40:227–256.
Kay RF, Hylander WL. 1978. The dental structure of mamma-lian folivores with special reference to primates and phalange-roids (Marsupialia). In: Montgomery GG, editor. The ecologyof arboreal folivores. Washington, DC: Smithsonian Institu-tion Press. p 173–192.
Keyser AW. 2000. The Drimolen skull: the most complete aus-tralopithecine cranium and mandible to date. S Afr J Sci96:189–193.
Keyser AW, Menter CG, Moggi-Cecchi J, Pickering TR, BergerLR. 2000. Drimolen: a new hominid-bearing site in Gauteng,South Africa. S Afr J Sci 96:193–197.
Kimbel WH. 1988. Identification of a partial cranium of Austral-opithecus afarensis from the Koobi Fora Formation, Kenya. JHum Evol 17:647–656.
Kimbel WH, Delezene LK. 2009. ‘‘Lucy’’ redux: a review ofresearch on Australopithecus afarensis. Yearb Phys Anthropol52:2–48.
Kimbel WH, Lockwood CA, Ward CV, Leakey MG, Rak Y,Johanson DC. 2006. Was Australopithecus anamensis ances-tral to A. afarensis? A case for anagenesis in the hominin fos-sil record. J Hum Evol 51:134–152.
Kimbel WH, Rak Y, Johanson DC. 2004. The skull of Australo-pithecus afarensis. Oxford: Oxford University Press.
Kimbel WH, White TD, Johanson DC. 1988. Implications ofKNM-WT 17000 for the evolution of ‘‘robust’’ australopithe-cines. In: Grine FE, editor. Evolutionary history of the ‘‘ro-bust’’ australopithecines. New York: Aldine. p 259–268.
King SJ, Arrigo-Nelson SJ, Pochron ST, Semprebon GM, God-frey LR, Wright PC, Jernvall J. 2005. Dental senescence in along-lived primate links survival to rainfall. Proc Natl AcadSci USA 102:16579–16583.
King T, Andrews P, Boz B. 1999. Effect of taphonomic processeson dental microwear. Am J Phys Anthropol 108:359–373.
Kinzey WG. 1978. Feeding behavior and molar features in twospecies of Titi Monkey. In: Chivers DJ, Herbert J, editors.Recent advances in primatology. Vol.1: Behavior. New York:Academic. p 373–385.
Kullmer O, Sandrock O, Abel R, Schrenk F, Bromage TG,Juwayeyi YM. 1999. The first Paranthropus from the MalawiRift. J Hum Evol 37:121–127.
Kuman K, Clarke RJ. 2000. Stratigraphy, artefact industriesand hominid associations for Sterkfontein, Member 5. J HumEvol 38:827–847.
Lalueza Fox C, Perez-Perez A. 1993. The diet of the Neander-thal child Gibraltar 2 (Devils Tower) through the study of thevestibular striation pattern. J Hum Evol 24:29–41.
Lalueza Fox C, Perez-Perez A, Turbon D. 1996. Dietary infer-ences through buccal microwear analysis of Middle and UpperPleistocene human fossils. Am J Phys Anthropol 100:367–387.
Lambert JE, Chapman CA, Wrangham RW, Conklin-BrittainNL. 2004. Hardness of cercopithecines foods: implications forthe critical function of enamel thickness in exploiting fallbackfoods. Am J Phys Anthropol 125:363–368.
Leakey MG, Feibel CS, McDougall I, Walker A. 1995. New four-million-year old hominid species from Kanapoi and Allia Bay,Kenya. Nature 376:565–571.
Leakey MG, Feibel CS, McDougall I, Ward CV, Walker A. 1998.New specimens and confirmation of an early age for Australo-pithecus anamensis. Nature 393:62–66.
Leakey MG, Spoor F, Brown FH, Gathogo PN, Kiarie C, LeakeyLN, McDougall I. 2001. New hominin genus from eastern Africashows diverse middle Pliocene lineages. Nature 410:433–440.
Lee-Thorp JA, Sealy JC, van der Merwe NJ. 1989. Stable carbonisotope ratio differences between bone collagen and bone apa-tite, and their relationship to diet. J Archaeol Sci 16:585–599.
Lee-Thorp JA. 2002. Two decades of progress towards under-standing fossilization processes and isotopic signals in calcifedtissue minerals. Archaeometry 44:435–446.
Lee-Thorp JA, Sponheimer M. 2003. Three case studies used toreassess the reliability of fossil bone and enamel isotope signalsfor palaeodietary studies. J Anthropol Archaeol 22:208–216.
Lee-Thorp JA, Sponheimer M. 2005. Opportunities and con-straints for reconstructing palaeoenvironments from stablelight isotope ratios in fossils. Geol Q 49:195–204.
Lee-Thorp JA, Sponheimer M. 2006. Biogeochemical approaches toinvestigating hominin diets. Yearb Phys Anthropol 49:131–48.
Lee-Thorp JA, Sponheimer M, Luyt JC. 2007. Tracking chang-ing environments using stable carbon isotopes in fossil toothenamel: an example from the South African hominin sites. JHum Evol 53:595–601.
Lee-Thorp JA, Sponheimer M, Passey BH, de Ruiter DJ, CerlingTE. 2010. Stable isotopes in fossil hominin tooth enamel sug-gest a fundamental dietary shift in the Pliocene. Phil Trans RSoc B 365:3389–3396.
Lee-Thorp JA, Thackeray JF, van der Merwe NJ. 2000. Thehunters and the hunted revisited. J Hum Evol 39:565–576.
Lee-Thorp JA, van der Merwe NJ. 1987. Carbon isotope analy-sis of fossil bone apatite. S Afr J Sci 83:712–715.
Lee-Thorp JA, van der Merwe NJ, Brain CK. 1994. Diet of Aus-tralopithecus robustus at Swartkrans from stable carbon iso-topic analysis. J Hum Evol 27:361–372.
312 F.E. GRINE ET AL.
American Journal of Physical Anthropology

LeGeros RZ. 1991. Calcium phosphates in oral biology and med-icine. Basel: Karger.
Leonard WR, Hegmon M. 1987. Evolution of P3 morphology inAustralopithecus afarensis. Am J Phys Anthropol 73:41–63.
Levin NE, Cerling TE, Passey BH, Harris JM, Ehleringer JR.2006. A stable isotope aridity index for terrestrial environ-ments. Proc Natl Acad Sci USA 103:11201–11205.
Levinson M. 1985. Are fossil rodents useful in palaeo-ecologicalinterpretations? Ann Geo Surv S Afr 19:53–64.
Ley RE, Hamady M, Lozupone C, Turnbaugh PJ, Ramey RR,Bircher JS, Schlegel ML, Tucker TA, Schrenzel M, Knight R,Gordon JI. 2008. Evolution of mammals and their gutmicrobes. Science 320:1647–1651.
Liem KF. 1980. Adaptive significance of intraspecific and inter-specific differences in the feeding repertoires of cichlid fishes.Am Zool 20:295–314.
Liem KF, Summers AP. 2000. Integration of versatile functionaldesign, population ecology, ontogeny and phylogeny. Nether-lands J Zool 50:245–259.
Lockwood CA, Kimbel WH, Johanson DC. 2000. Temporaltrends and metric variation in the mandibles and dentition ofAustralopithecus afarensis. J Hum Evol 39:23–55.
Louchart A, Wesselman H, Blumenschine RJ, Hlusko LJ, NjauJK, Black MT, Asnake M, White TD. 2009. Taphonomic,avian, and small-vertebrate indicators of Ardipithecus rami-dus habitat. Science 326:66e1–66e4.
Lough SA, Rivera J, Comar CL. 1963. Retention of strontium,calcium and phosphorous in human infants. Proc Soc ExpBiol Med 112:631–636.
Lovejoy CO, Latimer B, Suwa G, Asfaw B, White TD. 2009a.Combining prehension and propulsion: the foot of Ardipithe-cus ramidus. Science 326:72e1–72e8.
Lovejoy CO, Simpson SW, White TD, Asfaw B, Suwa G. 2009b.Careful climbing in the Miocene: the forelimbs of Ardipithe-cus ramidus and humans are primitive. Science 326:70e1–70e8.
Lucas PW. 2004. Dental functional morphology: how teeth work.Cambridge: Cambridge University Press.
Lucas PW, Constantino PJ, Wood BA. 2008a. Inferences regard-ing the diet of extinct hominins: structural and functionaltrends in dental and mandibular morphology within the homi-nin clade. J Anat 212:486–500.
Lucas PW, Constantino P, Wood B, Lawn B. 2008b. Dentalenamel as a dietary indicator in mammals. Bioessays 30:374–385.
Lucas PW, Corlett RT, Luke DA. 1986. Postcanine tooth sizeand diet in anthropoid primates. Zeit Morphol Anthropol76:253–276.
Lucas PW, Teaford MF. 1995. Significance of silica in leaves tolong-tailed macaques (Macaca fascicularis). Folia Primatol64:30–36.
Maas MC. 1994. A scanning electron-microscope study of in vivoabrasion of mammalian tooth enamel under compressiveloads. Archs Oral Biol 39:1–11.
Macho GA, Leakey MG, Williamson DK, Jiang Y. 2003. Palae-oenvironmental reconstruction: evidence for seasonality atAllia Bay, Kenya, at 3.9 million years. Palaeogeog Palaeocli-matol Palaeoecol 199:17–30.
Macho GA, Shimizu D, Jiang Y, Spears IR. 2005. Australopithe-cus anamensis: a finite-element approach to studying thefunctional adaptations of extinct hominins. Anat Rec A283:310–318.
MacLatchy LM, DeSilva JM, Sanders WJ, Wood BA. 2010.Hominini. In: Werdelin L, Sanders WJ, editors. Cenozoicmammals of Africa. Berkeley: University of California Press.p 471–540.
Mainland IL. 2003. Dental microwear in grazing and browsingGotland sheep (Ovis aries) and its implications for dietaryreconstruction. J Archaeol Sci 30:1513–1527.
Maragakis GM, Hahn P, Hellwig E. 2001. Chemomechanicalcaries removal: a comprehensive review of the literature. IntDent J 51:291–299.
McGraw WS. 1998. Posture and support use of Old World mon-keys (Cercopithecidae): the influence of foraging strategies,
activity patterns, and the spatial distribution of preferredfood items. Am J Primatol 46:229–250.
McKee JK. 1991. Paleo-ecology of the Sterkfontein hominids: areview and synthesis. Palaeontol Afr 28:41–51.
McKee JK. 1999. The autocatalytic nature of hominid evolutionin African Plio-Pleistocene environments. In: Bromage TG,Schrenk F, editors. African biogeography, climate change, andhuman evolution. New York: Oxford University Press. p 57–75.
Menter CG, Kuykendall KL, Keyser AW, Conroy GC. 1999. Firstrecord of hominid teeth from the Plio-Pleistocene site of Gon-dolin, South Africa. J Hum Evol 37:299–307.
Merceron G. 2003. Une nouvelle methodologie pour la quantifi-cation de la micro-usure dentaire: application a l’hominoıdefossile Ouranopithecus macedoniensis (Miocene, Grece). BullMem Soc Anthropol Paris 15:300–310.
Merceron G, Escarguel G, Angibault JM, Verheyden-Tixier H.2010. Can dental microwear textures record inter-individualdietary variations? PLoS One 5:e9542.
Merceron G, Viriot L, Blondel C. 2004. Tooth microwear patternin roe deer (Capreolus capreolus L.) from Chize (westernFrance) and relation to food composition. Small Rum Res53:125–132.
Moggi-Cecchi J. 2003. The elusive ‘second species’ in Sterkfon-tein Member 4: the dental metrical evidence. S Afr J Sci99:268–270.
Moggi-Cecchi J, Grine FE, Tobias PV. 2006. Early hominid den-tal remains from Members 4 and 5 of the Sterkfontein Forma-tion (1966–1996 excavations): catalogue, individual associa-tions, morphological descriptions and initial metrical analysis.J Hum Evol 50:239–328.
Moggi-Cecchi J, Menter C, Boccone S, Keyser A. 2010. Earlyhominin dental remains from the Plio-Pleistocene site of Dri-molen, South Africa. J Hum Evol 58:374–405.
Nagaoka S, Katoh S, WoldeGabriel G, Sato H, Nakaya H,Beyene Y, Suwa G. 2005. Lithostratigraphy and sedimentaryenvironments of the hominid-bearing Pliocene-PleistoceneKonso Formation in the southern Main Ethiopian Rift, Ethio-pia. Palaeogeogr Palaeoclimatol Palaeoecol 216:333–357.
Nakagawa N, Ohsawa H, Muroyama Y. 2003. Life-history pa-rameters of a wild group of West African patas monkeys(Erythrocebus patas patas). Primates 44:281–290.
Nee S, Read AF, Harvey PH. 1996. Why phylogenies are neces-sary for comparative analysis. In: Martins EP, editor. Phyloge-nies and the comparative method in animal behavior. NewYork: Oxford University Press. p 399–413.
Nystrom P, Phillips-Conroy JE, Jolly CJ. 2004. Dental micro-wear in anubis and hybrid baboons (Papio hamadryas, sensulato) living in Awash National Park, Ethiopia. Am J PhysAnthropol 125:279–291.
Olson TR. 1981. Basicranial morphology of the extant hominoidsand Pliocene hominids: the new material from the Hadar For-mation, Ethiopia, and its significance in early human evolu-tion and taxonomy. In: Stringer CB, editor. Aspects of humanevolution. London: Taylor and Francis. p 99–128.
Olson TR. 1985. Cranial morphology and the systematic of theHadar Formation hominids and ‘‘Australopithecus’’ africanus.In: Delson E, editor. Ancestors: the hard evidence. New York:Alan R. Liss. p 102–119.
Otis JS, Froehlich JW, Thorington RW. 1981. Seasonal and age-related differential mortality by sex in the mantled howlermonkey, Alouatta palliata. Int J Primatol 2:197–205.
Palmqvist P, Grocke DR, Arribas A, Farina RA. 2003. Paleoeco-logical reconstruction of a lower Pleistocene large mammalcommunity using biogeochemical (d13C, d15N, Sr/Zn) and eco-morphological approaches. Paleobiology 29:205–229.
Partridge TC. 2000. Hominid-bearing cave and tufa deposits. In:Partridge TC, Maud RR, editors. The Cenozoic of SouthernAfrica. New York: Oxford University Press. p 100–125.
Passey BH, Cerling TE. 2002. Tooth enamel mineralization inungulates: implications for recovering a primary isotopictime-series. Geochim Cosmochim Acta 66:3225–3234.
Passey BH, Cerling TE. 2006. In situ stable isotope analysis(d13C, d18O) of very small teeth using laser ablation GC/IRMS. Chem Geol 235:238–249.
313MICROWEAR, ISOTOPES, AND HOMININ PALEOECOLOGY
American Journal of Physical Anthropology

Passey BH, Robinson TF, Ayliffe LK, Cerling TE, SponheimerM, Dearing MD, Roeder BL, Ehleringer JR. 2005. Carbon iso-tope fractionation between diet breadth, CO2, and bioapatitein different mammals. J Archaeol Sci 32:1459–1470.
Perez-Perez A. 2004. Why buccal microwear? Anthropologie42:1–3.
Perez-Perez A, Bermudez de Castro JM, Arsuaga JL. 1999.Non-occlusal dental microwear analysis of 300,000 year-oldHomo heidelbergensis teeth from Sima de los Huesos (Sierrade Atapuerca, Spain): implications of intrapopulation variabil-ity for dietary analysis of hominid fossil remains. Am J PhysAnthropol 108:433–457.
Perez-Perez A, Espurz V, Bermudez de Castro JM, de LumleyMA, Turbon D. 2003. Non-occlusal dental microwear variabilityin a sample of Middle and Upper Pleistocene human popula-tions from Europe and the Near East. J Hum Evol 44:497–513.
Perez-Perez A, Lalueza C, Turbon D. 1994. Intraindividual andintragroup variability of buccal tooth striation pattern. Am JPhys Anthropol 94:175–187.
Perry GH, Dominy NJ, Claw KG, Lee AS, Fiegler H, Redon R,Werner J, Villanea FA, Mountain JL, Misra R, Carter NP, LeeC, Stone AC. 2007. Diet and the evolution of human amylasegene copy number variation. Nat Genet 39:1256–1260.
Pickering R, Kramers JD, Hancox PJ, de Ruiter DJ, WoodheadJD. 2011. Contemporary flowstone development links earlyhominin bearing cave deposits in South Africa. Earth PlanetSci Lett 306:23–32.
Plavcan JM, Kay RF, Jungers WL, van Schaik CP, editors.2002. Reconstructing behavior in the primate fossil record.New York: Plenum.
Plummer TW, Bishop LC. 1994. Hominid paleoecology at Oldu-vai Gorge, Tanzania as indicated by antelope remains. J HumEvol 27:47–75.
Prideaux GJ, Ayliffe LK, DeSantis LRG, Schubert BW, MurrayPF, Gagan MK, Cerling TE. 2009. Extinction implications of achenopod browse diet for a giant Pleistocene kangaroo. ProcNatl Acad Sci USA 106:11646–11560.
Puech PF, Albertini H. 1984. Dental microwear and mechanismsin early hominids from Laetoli and Hadar. Am J Phys Anthro-pol 65:87–91.
Puech PF, Albertini H, Serratrice C. 1983. Tooth microwear anddietary patterns in early hominids from Laetoli, Hadar andOlduvai. J Hum Evol 12:721–729.
Puech PF, Prone A, Albertini H. 1981. Reproduction experimen-tale des processus d-alteration de la surface dentaire par fric-tion non abrasive et non adhesive: application a l’etude de ali-mentation de l’Homme fossile. C R Acad Sci Paris 293:729–734.
Rabenold D, Pearson OM. 2011. Abrasive, silica phytoliths andthe evolution of thick molar enamel in primates, with implica-tions for the diet of Paranthropus boisei. PLoS ONE 6(12):e28379. doi:10.1371/journal.pone.0028379.
Rak Y. 1983. The australopithecine face. New York: Academic.RakY. 1988.On variation in themasticatory systemofAustralopithe-
cus boisei. In: Grine FE, editor. Evolutionary history of the ‘‘robust’’australopithecines.NewYork: Aldine deGruyter. p 193–198.
Rak Y, Ginzburg, A, Geffen E. 2007. Gorilla-like anatomy onAustralopithecus afarensis mandibles suggests Au. afarensislink to robust australopiths. Proc Natl Acad Sci USA104:6568–6572.
Rak Y, Hylander WL. 2008. What else is the tall mandibularramus of the robust australopiths good for? In: Vinyard CJ,Wall CE, Ravosa MJ, editors. Primate craniofacial functionand biology: papers in honor of Bill Hylander. Dordrecht:Springer. p 431–442.
Ravosa MJ. 2000. Size and scaling in the mandible of living andextinct apes. Folia Primatol 71:305–322.
Rayfield E. 2007. Finite element analysis and understandingthe biomechanics and evolution of living and extinct organ-isms. Ann Rev Earth Planet Sci 35:541–576.
Rayner RJ, Moon BP, Masters JC. 1993. The Makapansgat aus-tralopithecine environment. J Hum Evol 24:219–231.
Reed KE. 1997. Early hominid evolution and ecological changethrough the African Plio-Pleistocene. J Hum Evol 32:289–322.
Reed KE. 2008. Paleoecological patterns at the Hadarhominin site, Afar Regional State, Ethiopia. J Hum Evol 54:743–768.
Reeve HK, Sherman PW. 1993. Adaptation and the goals of evo-lutionary research. Q Rev Biol 68:1–32.
Reinhard KJ, Danielson DR. 2005. Pervasiveness of phytolithsin prehistoric southwestern diet and implications for regionaland temporal trends for dental microwear. J Archaeol Sci32:981–988.
Remis MJ. 2002. Food Preferences among captive western goril-las (Gorilla gorilla gorilla) and chimpanzees (Pan troglodytes).Int J Primatol 23:231–249.
Renne PR, WoldeGabriel G, Hart WK, Heiken G, White TD.1999. Chronostratigraphy of the Miocene-Pliocene SagantoleFormation, Middle Awash Valley, Afar Rift, Ethiopia. Geol SocAm Bull 111:869–885.
Richards MP, Pettett PB, Stiner MC, Trinkaus E. 2001. Stableisotope evidence for increasing dietary breadth in the Euro-pean mid-Upper Paleolithic. Proc Natl Acad Sci USA98:6528–6532.
Richards MP, Pettitt PB, Trinkaus E, Smith FH, Paunovic M,Karavanic I. 2000. Neanderthal diet at Vindija and Neander-thal predation: the evidence from stable isotopes. Proc NatlAcad Sci USA 97:7663–7666.
Richards MP, Schmitz RW. 2008. Isotopic evidence for the dietof the Neanderthal type specimen. Antiquity 82:553–559.
Rightmire GP. 2008. Homo in the Middle Pleistocene: hypodigms,variation, and species recognition. Evol Anthropol 17:8–21.
Robinson BW, Wilson DS. 1998. Optimal foraging, specializa-tion, and a solution to Liem’s paradox. Am Nat 151:223–235.
Robinson JT. 1954a. The genera and species of the Australopi-thecinae. Am J Phys Anthropol 12:181–200.
Robinson JT. 1954b. Prehominid dentition and hominid evolu-tion. Evolution 8:324–334.
Robinson JT. 1956. The Dentition of the Australopithecinae.Transvaal Mus Mem 9:1–179.
Robinson JT. 1967. Variation and taxonomy of the early homi-nids. In: Dobzhansky T, Hecht MK, Steere WC, editors. Evo-lutionary biology. Vol.1. New York: Meredith. p 69–100.
Romero A, De Juan J. 2007. Intra- and interpopulation humanbuccal tooth surface microwear analysis: inferences about dietand formation processes. Anthropologie 45:61–70.
Romero A, Galbany J, Martinez-Ruiz N, De Juan J. 2009. Invivo turnover rates in human buccal dental-microwear. Am JPhys Anthropol 138 (Suppl 48):223–224.
Rosenberger AL, Kinzey WG. 1976. Functional patterns ofmolar occlusion in platyrrhine primates. Am J Phys Anthro-pol 45:281–298.
Ross CF, Lockwood CA, Fleagle JG, Jungers WL. 2002. Adapta-tion and behavior in the primate fossil record. In: PlavcanJM, Kay RF, Jungers WL, van Schaik CP, editors. Recon-structing behavior in the primate fossil record. New York:Kluwer. p 1–41.
Ruff CB, Runestad JA. 1992. Primate limb bone structuraladaptations. Ann Rev Anthropol 21:407–433.
Runia LJ. 1987. Strontium and calcium distribution in plants:effect on paleodietary studies. J Archaeol Sci 14:599–608.
Ryan AS, Johanson DC. 1989. Anterior dental microwear inAustralopithecus afarensis: comparisons with human and non-human primates. J Hum Evol 18:235–268.
Safont S, Malgosa A, Subira ME, Gilbert J. 1998. Can trace ele-ments in fossils provide information about palaeodiet? Int JOsteoarchaeol 8:23–37.
Sanson GD, Kerr SA, Gross KA. 2007. Do silica phytoliths reallywear mammalian teeth? J Archaeol Sci 34:526–531.
Schaller GB, Lowther GR. 1969. The relevance of carnivorebehaviour to the study of early hominids. Southwest JAnthropol 25:307–341.
Schoeninger MJ. 1979. Diet and status at Chalcatzingo: someempirical and technical aspects of strontium analysis. Am JPhys Anthropol 51:295–310.
Schoeninger MJ, Bunn HT, Murray SS, Marlett JA. 2001. Com-position of tubers used by Hadza foragers of Tanzania. J FoodComp Analysis 14:15–25.
314 F.E. GRINE ET AL.
American Journal of Physical Anthropology

Schoeninger MJ, Moore J, Sept M. 1999. Subsistence strategiesof two ‘savanna’ chimpanzee populations: the stable isotopeevidence. Am J Primatol 49:297–314.
Schoeninger MJ, Reeser H, Hallin K. 2003. Paleoenvironment ofAustralopithecus anamensis at Allia Bay, East Turkana,Kenya: evidence from mammalian herbivore enamel stableisotopes. J Anthropol Archaeol 22:200–207.
Schubert BW, Ungar PS, DeSantis LRG. 2010. Carnassialmicrowear and dietary behaviour in large carnivorans. J Zool280:257–263.
Schubert BW, Ungar PS, Sponheimer M, Reed KE. 2006. Micro-wear evidence for Plio-Pleistocene bovid diets from Makapans-gat Limeworks Cave, South Africa. Palaeogeogr Palaeoclima-tol Palaeoecol 241:301–319.
Scott RS, Schubert B, Grine FE, Teaford M. 2008. Low magnifi-cation microwear: questions of precision and repeatability. JVert Paleontol 28 (Suppl 3):139A.
Scott RS, Teaford MF, Ungar, PS. 2012. Dental microwear tex-ture and anthropoid diets. Am J Phys Anthropol 147:551–579.
Scott RS, Ungar PS, Bergstrom TS, Brown CA, Grine FE, Tea-ford MF, Walker A. 2005. Dental microwear texture analysisreflects diets of living primates and fossil hominins. Nature436:693–695.
Sealy JC, Sillen A. 1988. Sr and Sr/Ca in marine and terrestrialfoodwebs in the southwestern Cape, South Africa. J ArchaeolSci 15:425–438.
Semaw S, Simpson SW, Quade J, Renne PR, Butler RF, McIntoshWC, Levin N, Dominguez-Rodrigo M, Rogers MJ. 2005. EarlyPliocene hominids from Gona, Ethiopia. Nature 433:301–305.
Semprebon GM, Godfrey LR, Solounias N, Sutherland MR,Jungers WL. 2004. Can low-magnification stereomicroscopyreveal diet? J Hum Evol 47:115–144.
Semprebon GM, Godfrey LR, Solounias N, Sutherland MR,Jungers WL. 2005. [Erratum] Can low-magnification stereomicroscopy reveal diet? J Hum Evol 49:662–663.
Shabel AB. 2006. The paleobiology of the robust australopithecines(Paranthropus): a preliminary test of the durophage model usingcarbon isotope analysis. Am J Phys Anthropol S42:163.
Shabel AB. 2008. The paleobiology of the robust australopithe-cines (Paranthropus): a test of the durophage model with amorphometric analysis of carnivoran skulls. Am J PhysAnthropol S46:190–191.
Sheine WS, Kay RF. 1977. An analysis of chewed food particlesize and its relationship to molar structure in the primatesCheirogaleus medius and Galago senegalensis and the insecti-voran Tupaia glis. Am J Phys Anthropol 47:15–20.
Shipman P, Harris JM. 1988. Habitat preference and paleoecol-ogy of Australopithecus boisei in eastern Africa. In: Grine FE,editor. Evolutionary history of the ‘‘robust’’ australopithecines.New York: Aldine de Gruyter. p 343–381.
Sikes NE. 1994. Early hominid habitat preferences: paleosol car-bon isotopic evidence. J Hum Evol 27:25–45.
Silcox MT, Teaford MF. 2002. The diet of worms: an analysis ofmole dental microwear. J Mammal 83:804–814.
Sillen A. 1981. Strontium and diet at Hayonim Cave. Am JPhys Anthropol 56:131–137.
Sillen A. 1989. Diagenesis of the inorganic phase ofcortical bone. In: Price TD, editor. The chemistry of prehis-toric human bone. Cambridge: Cambridge University Press.p 211–299.
Sillen A. 1992. Strontium–calcium ratios (Sr/Ca) of Australopi-thecus robustus and associated fauna from Swartkrans. JHum Evol 23:495–516.
Sillen A, Hall G, Armstrong R. 1995. Strontium–calcium ratios(Sr/Ca) and strontium isotope ratios (87Sr/86Sr) of Australopi-thecus robustus and Homo sp. from Swartkrans. J Hum Evol28:277–286.
Sillen A, Kavanagh M. 1982. Strontium and paleodietaryresearch. Yearb Phys Anthropol 25:67–90.
Smith BN, Epstein S. 1971. Two categories of 13C/12C ratios forhigher plants. Plant Physiol 47:380–384.
Smith HF, Grine FE. 2008. Cladistic analysis of early Homo cra-nia from Swartkrans and Sterkfontein, South Africa. J HumEvol 54:684–704.
Smith RJ. 1993. Categories of allometry: body size versus bio-mechanics. J Hum Evol 24:173–182.
Smith RJ. 1996. Biology and body size in human evolution:statistical inference misapplied. Curr Anthropol 37:451–481.
Solounias N, Semprebon G. 2002. Advances in the reconstruc-tion of ungulate ecomorphology with application to early fossilequids. Am Mus Novitiates 3366:1–49.
Spencer H, Warren JM, Kramer L, Samachson J. 1973. Passageof calcium and strontium across the intestine in man. ClinOrthop 91:225–234.
Sponheimer M, de Ruiter D, Lee-Thorp J, Spath A. 2005a. Sr/Ca and early hominin diets revisited: new data from modernand fossil tooth enamel. J Hum Evol 48:147–156.
Sponheimer M, Lee-Thorp JA. 1999a. Isotopic evidence for thediet of an early hominid, Australopithecus africanus. Science283:368–370.
Sponheimer M, Lee-Thorp JA. 1999b. Oxygen isotopes inenamel carbonate and their ecological significance. J ArchaeolSci 26:723–728.
Sponheimer M, Lee-Thorp JA. 2001. The oxygen isotope compo-sition of mammalian enamel carbonate: a case study fromMorea Estate, Mpumalanga Province, South Africa. Oecologia126:153–157.
Sponheimer M, Lee-Thorp JA. 2006. Enamel diagenesis atSouth African australopith sites: implications for paleoecologi-cal reconstruction with trace elements. Geochim CosmochimActa 70:1644–1654.
Sponheimer M, Lee-Thorp JA, de Ruiter D. 2007. Icarus, iso-topes and australopith diets. In: Ungar PS, editor. Evolutionof the human diet: the known, the unknown and the unknow-able. Oxford: Oxford University Press. p 132–149.
Sponheimer M, Lee-Thorp JA, de Ruiter D, Codron D, CodronJ, Baugh A, Thackeray JF. 2005b. Hominins, sedges and ter-mites: new carbon isotope data for the Sterkfontein Valley. JHum Evol 48:301–312.
Sponheimer M, Lee-Thorp J, de Ruiter D, Smith J, van derMerwe N, Reed K, Ayliffe L, Heidelberger C, Marcus W. 2003.Diets of southern African Bovidae: stable isotope evidence. JMammal 84:471–479.
Sponheimer M, Loudon JE, Codron D, Howells ME, Pruetz JD,Codron J, de Ruiter D, Lee-Thorp JA. 2006a. Do ‘savanna’chimpanzees consume C4 resources? J Hum Evol 51:128–133.
Sponheimer M, Passey BH, de Ruiter DJ, Guatelli-Steinberg D,Cerling TE, Lee-Thorp JA. 2006b. Isotopic evidence for die-tary variability in the early hominin Paranthropus robustus.Science 314:980–982.
Sponheimer M, Reed KE, Lee-Thorp JA. 1999. Combining iso-topic and ecomorphological data to refine bovid paleodietaryreconstruction: a case study from the Makapansgat Lime-works hominin locality. J Hum Evol 36:705–718.
Spoor F, Leakey MG, Gathogo PN, Brown FH, Anton SC,McDougall I, Kiarie C, Manthi FK, Leakey LN. 2007. Implica-tions of new early Homo fossils from Ileret, east of Lake Tur-kana, Kenya. Nature 448:688–691.
Steininger CM, Berger LR, Kuhn BF. 2008. A partial skull ofParanthropus robustus from Cooper’s Cave, South Africa. SAfr J Sci 104:143–146.
Stern JT, Oxnard CE. 1973. Primate locomotion: some linkswith evolution and morphology. Folia Primatol 4:1–93.
Strait DS, Grine FE. 2004. Inferring hominoid and early homi-nid phylogeny using craniodental characters: the role of fossiltaxa. J Hum Evol 47:399–452.
Strait DS, Weber GW, Neubauer S, Chalk J, Richmond BG,Lucas PW, Spencer MA, Schrein C, Dechow PC, Ross CF,Grosse IR, Wright BW, Constantino P, Wood BA, Lawn B,Hylander WL, Wang Q, Byron C, Slice DE, Smith AL.2009. The feeding biomechanics and dietary ecology of Aus-tralopithecus africanus. Proc Natl Acad Sci USA 106:2124–2129.
Sullivan CH, Krueger HW. 1981. Carbon isotope analysis of sep-arate chemical phases in modern and fossil bone. Nature292:333–335.
Suwa G. 1988. Evolution of the ‘‘robust’’ australopithecines in theOmo succession: evidence from mandibular premolar morphology.
315MICROWEAR, ISOTOPES, AND HOMININ PALEOECOLOGY
American Journal of Physical Anthropology

In: Grine FE, editor. Evolutionary history of the ‘‘robust’’ australo-pithecines. New York: Aldine de Gruyter. p 199–222.
Suwa G, Asfaw B, Beyene Y, White T, Katoh S, Nagaoka S,Nakaya H, Uzawa K, Renne P, WoldeGabriel G. 1997. Thefirst skull of Australopithecus boisei. Nature 389:489–492.
Suwa G, Kono RT, Simpson SW, Asfaw B, Lovejoy CO, WhiteT.D. 2009. Paleobiological implications of the Ardipithecusramidus dentition. Science 326:94–99.
Suwa G, White TD, Howell FC. 1996. Mandibular postcaninedentition from the Shungura Formation, Ethiopia: crown mor-phology, taxonomic allocations, and Plio-Pleistocene hominidevolution. Am J Phys Anthropol 101:247–282.
Suwa G, Wood BA, White TD. 1994. Further analysis of man-dibular molar crown and cusp areas in Pliocene and earlyPleistocene hominids. Am J Phys Anthropol 93:407–426.
Szalay FS. 1975. Hunting-scavenging protohominids: a modelfor hominid origins. Man 10:420–429.
Taylor AB. 2002. Masticatory form and function in the Africanapes. Am J Phys Anthropol 117:133–156.
Teaford MF. 1985. Molar microwear and diet in the genusCebus. Am J Phys Anthropol 66:363–70.
Teaford MF. 1988a. A review of dental microwear and diet inmodem mammals. Scanning Microsc 2:1149–66.
Teaford MF. 1988b. Scanning electron microscope diagnosis ofwear patterns versus artifacts on fossil teeth. ScanningMicrosc 2:1167–1175.
Teaford MF, Glander KE. 1991. Dental microwear in live, wild-trapped Alouatta palliata from Costa Rica. Am J PhysAnthropol 85:313–319.
Teaford MF, Glander KE. 1996. Dental microwear and diet in awild population of mantled howling monkeys (Alouatta pal-liata). In: Norconk MA, Rosenberger AL, Garber PA, editors.Adaptive radiations of Neotropical primates. New York: Ple-num. p 433–439.
Teaford MF, Grine FE, Kay RF, Schubert B. 2008. Low magnifi-cation dental microwear: the problem of postmortem artefacts.J Vert Paleontol 28 (Suppl 3):151A.
Teaford MF, Lucas PW, Ungar PS, Glander KE. 2006. Mechani-cal defenses in leaves eaten by Costa Rican Alouatta palliata.Am J Phys Anthropol 129:99–104.
Teaford MF, Oyen OJ. 1989. In vivo and in vitro turnover indental microwear. Am J Phys Anthropol 80:447–460.
Teaford MF, Robinson JG. 1989. Seasonal or ecological differen-ces in diet and molar microwear in Cebus nigrivittatus. Am JPhys Anthropol 80:391–401.
Teaford MF, Runestad JA. 1992. Dental microwear and diet inVenezuelan primates. Am J Phys Anthropol 88:347–364.
Teaford MF, Tylenda CA. 1991. A new approach to the study oftooth wear. J Dent Res 70:204–207.
Teaford MF, Ungar PS. 2000. Diet and the evolution of the earliesthuman ancestors. Proc Natl Acad Sci USA 97:13506–13511.
Teaford MF, Ungar PS, Grine FE. 2002. Paleontological evi-dence for the diets of African Plio–Pleistocene hominins withspecial reference to early Homo. In: Ungar PS, Teaford MF,editors. Human diet: its origin and evolution. Westport, CT:Bergin and Garvey. p 143–166.
Teaford MF, Ungar PS, Ross CF, Vinyard CJ. 2010. The dentalmicrowear of hard-object feeding in laboratory primates. AmJ Phys Anthropol 50:228.
Teaford MF, Walker A. 1984. Quantitative differences in dentalmicrowear between primate species with different diets and acomment on the presumed diet of Sivapithecus. Am J PhysAnthropol 64:191–200.
Thackeray JF, de Ruiter D, Berger L, van der Merwe N. 2001.Hominid fossils from Kromdraai: a revised list of specimensdiscovered since 1938. Ann Transvaal Mus 38:43–56.
Tieszen LL. 1991. Natural variations in the carbon isotope val-ues of plants: implications for archaeology, ecology, and paleo-ecology. J Archaeol Sci 18:227–248.
Tieszen LL, Fagre T. 1993. Effect of diet quality and composi-tion on the isotopic composition of respiratory CO2, bone colla-gen, bioapatite, and soft tissues. In: Lambert JB, Grupe G,editors. Prehistoric human bone: archaeology at the molecularlevel. Berlin: Springer. p 121–155.
Tishkoff SA, Reed FA, Ranciaro A, Voight BF, Babbitt CC, Sil-verman JS, Powell K, Mortensen HM, Hirbo JB, Osman M,Ibrahim M, Omar SA, Lema G, Nyambo TB, Ghori J, Bump-stead S, Pritchard JK, Wray GA, Deloukas P. 2007. Conver-gent adaptation of human lactase persistence in Africa andEurope. Nat Genet 39:31–40.
Toots H, Voorhies MR. 1965. Strontium in fossil bones and thereconstruction of food chains. Science 149:854–855.
Trickett MA, Budd P, Montgomery J, Evans J. 2003. An assess-ment of solubility profiling as a decontamination procedurefor the 87Sr/86Sr analysis of archaeological human skeletal tis-sue. Appl Geochem 18:653–658.
Trueman CN, Privat K, Field J. 2008. Why do crystallinity val-ues fail to predict the extent of diagenetic alteration of bonemineral? Palaeogeog Palaeoclimatol Palaeoecol 266:160–167.
Tuross N, Behrensmeyer AK, Eanes ED. 1989. Sr increase andcrystallinity changes in taphonomic and archaeological bones.J Archaeol Sci 16:661–672.
Ungar PS. 1996. Relationship of incisor size to diet and anteriortooth use in sympatric Sumatran anthropoids. Am J Primatol38:145–156.
Ungar PS. 2004. Dental topography and diets of Australopithe-cus afarensis and early Homo. J Hum Evol 46:605–622.
Ungar PS. 2007. Dental topography and human evolution withcomments on the diets of Australopithecus africanus and Par-anthropus robustus. In: Bailey S, Hublin J-J, editors. dentalperspectives on human evolution: state of the art research indental anthropology. Dordrecht: Springer. p 321–344.
Ungar PS, Brown CA, Bergstrom TS, Walker A. 2003. Quantifica-tion of dental microwear by tandem scanning confocal micros-copy, and scale sensitive fractal analyses. Scanning 25:185–93.
Ungar PS, Bunn JM. 2008. Primate dental topographic analysisand functional morphology. In: Irish JD, Nelson GC, editors.Technique and application in dental anthropology. New York:Cambridge University Press. p 253–265.
Ungar PS, Grine FE. 1991. Incisor size and wear in Australopi-thecus africanus and Paranthropus robustus. J Hum Evol20:313–40.
Ungar PS, Grine FE, Teaford MF. 2006a. Diet in early Homo: areview of the evidence and a new model of adaptive versatil-ity. Ann Rev Anthropol 35:209–228.
Ungar PS, Grine FE, Teaford MF. 2008. Dental microwear indi-cates that Paranthropus boisei was not a hard-object feeder.PLoS One 3:e2044:1–6.
Ungar PS, Grine FE, TeafordMF, El Zaatari S. 2006b. Dental micro-wear and diets of African earlyHomo. J HumEvol 50:78–95.
Ungar PS, Krueger KL, Blumenschine RJ, Njau J, Scott RS.Dental microwear texture analysis of hominins recovered bythe Olduvai Landscape Paleoanthropology Project, 1995–2007.J Hum Evol, in press.
Ungar PS, Merceron G, Scott RS. 2007a. Dental microwear tex-ture analysis of Varswater bovids and early Pliocene paleoe-cology of Langebaanweg, Western Cape Province, SouthAfrica. J Mammal Evol 14:163–181.
Ungar PS, Scott RS, Grine FE, Teaford MF. 2010. Molar micro-wear textures and the diets of Australopithecus anamensis andAustralopithecus afarensis. Phil Trans R Soc B 365:3345–3354.
Ungar PS, Scott RS, Scott JR, Teaford MF. 2007b. Dental micro-wear analysis: historical perspectives and new approaches. In:Irish JD, Nelson GC, editors. Technique and application indental anthropology. Cambridge: Cambridge University Press.p 389–425.
Ungar PS, Teaford MF. 1996. Preliminary examination of non-occlusal dental microwear in anthropoids: implications for thestudy of fossil primates. Am J Phys Anthropol 100:101–113.
Ungar PS, Teaford MF, Glander KE, Pastor RF. 1995. Dustaccumulation in the canopy: a potential cause of dental micro-wear in primates. Am J Phys Anthropol 97:93–99.
Vanderklift MA, Ponsard S. 2003. Sources of variation in consumer-diet d15N enrichment: a meta-analysis. Oecologia 136:169–182.
van der Merwe NJ, Masao FT, Bamford MK. 2008. Isotopicevidence for contrasting diets of early hominins Homo habilisand Australopithecus boisei of Tanzania. S Afr J Sci 104:153–155.
316 F.E. GRINE ET AL.
American Journal of Physical Anthropology

van der Merwe NJ, Medina E. 1989. Photosynthesis and 13C/12Cratios in Amazonian rain forests. Geochim Cosmochim Acta53:1091–1094.
van der Merwe NJ, Thackeray JF, Lee-Thorp JA, Luyt J. 2003.The carbon isotope ecology and diet of Australopithecus afri-canus at Sterkfontein, South Africa. J Hum Evol 44:581–597.
Villmoare BA, Kimbel WH. 2011. CT-based study of internalstructure of the anterior pillar in extinct hominins and itsimplications for the phylogeny of robust Australopithecus.Proc Natl Acad Sci USA 108:16200–16205.
Vogel ER, van Woerden JT, Lucas PW, Atmoko SSU, van SchaikCP, Dominy NJ. 2008. Functional ecology and evolution ofhominoid molar enamel thickness: Pan troglodytes schwein-furthii and Pongo pygmaeus wurmbii. J Hum Evol 55:60–74.
Vogel JC. 1978. Isotopic assessment of the dietary habits ofungulates. S Afr J Sci 74:298–301.
Vrba ES. 1975. Some evidence of chronology and palaeoecologyof Sterkfontein, Swartkrans, and Kromdraai from the fossilBovidae. Nature 254:301–304.
Vrba ES. 1985. Early hominids in southern Africa: updatedobservations on chronological and ecological background. In:Tobias PV, editor. Hominid evolution: past, present andfuture. New York: Alan R. Liss. p 165–170.
Walker AC. 1981. Diet and teeth. Dietary hypotheses andhuman evolution. Phil Trans R Soc Lond 292:57–64.
Walker AC. 2007. Early hominin diets. Overview and historicalperspectives. In: Ungar PS, editor. Evolution of the humandiet. The known, the unknown, and the unknowable. NewYork: Oxford University Press. p 3–10.
Walker A, Hoeck HN, Perez L. 1978. Microwear of mammalianteeth as an indicator of diet. Science 201:908–910.
Walker AC, Leakey RE. 1988. The evolution of Australopithecusboisei. In: Grine FE, editor. Evolutionary history of the ‘‘robust’’australopithecines. New York: Aldine de Gruyter. p 247–258.
Walker AC, Leakey RE, Harris JM, Brown FH. 1986. 2.5 MyrAustralopithecus boisei from west of Lake Turkana, Kenya.Nature 322:517–522.
Wallace JA. 1975. Dietary adaptations of Australopithecus andearly Homo. In: Tuttle R, editor. Paleoanthropology, morphol-ogy and paleoecology. The Hague: Mouton. p 203–223.
Wang XX, Thomas SD, Zhang JZ. 2004. Relaxation of selectiveconstraint and loss of function in the evolution of human bit-ter taste receptor genes. Hum Mol Genet 13:2671–2678.
Wang Y, Cerling TE. 1994. A model of fossil tooth and bone dia-genesis: implications for paleodiet reconstruction from stableisotopes. Palaeogeog Palaeoclimatol Palaeoecol 107:281–289.
Ward CV, Leakey MG, Walker A. 2001. Morphology of Australo-pithecus anamensis from Kanapoi and Allia Bay, Kenya. JHum Evol 41:255–368.
Ward CV, Leakey MG, Walker AC. 1999. The new hominid spe-cies, Australopithecus anamensis. Evol Anthropol 7:197–205.
Ward CV, Plavcan JM, Manthi FK. 2010. Anterior dental evolu-tion in the Australopithecus anamensis—afarensis lineage.Phil Trans R Soc B 365:3333–3344.
Waters NE. 1980. Some mechanical and physical properties ofteeth. In: Vincent JFV, Currey JD, editors. The mechanicalproperties of biological materials. Cambridge: Cambridge Uni-versity Press. p 99–135.
Watson V. 1993. Composition of the Swartkrans bone accumula-tion in terms of skeletal parts and animals represented. In:Brain CK, editor. Swartkrans: a cave’s chronicle of early Man.Transvaal Museum Monograph 8. Pretoria: Transvaal Mu-seum. p 35–74.
Watts DP. 1998. Seasonality in the ecology and life histories ofmountain gorillas (Gorilla gorilla beringei). Int J Primatol19:929–948.
White TD. 1988. The comparative biology of ‘‘robust’’ Australopi-thecus: clues from context. In: Grine FE, editor. Evolutionaryhistory of the ‘‘robust’’ australopithecines. New York: Aldinede Gruyter. p 449–483.
White TD. 2002. Earliest hominids. In: Hartwig WC, editor. Theprimate fossil record. Cambridge: Cambridge UniversityPress. p 407–417.
White TD. 2003. Early hominids - diversity or distortion? Sci-ence 299:1994–1997.
White TD, Ambrose SH, Suwa G, Su DF, DeGusta D, BernorRL, Boisserie JR, Brunet M, Delson E, Frost S, Garcia N,Giaourtsakis IX, Haile-Selassie Y, Howell FC, Lehmann T,Likius A, Pehlevan C, Saegusa H, Semprebon G, Teaford M,Vrba E. 2009a. Macrovertebrate paleontology and the Pliocenehabitat of Ardipithecus ramidus. Science 326:87–93.
White TD, Asfaw B, Beyene Y, Haile-Selassie Y, Lovejoy CO,Suwa G, WoldeGabriel G. 2009b. Ardipithecus ramidus andthe paleobiology of early hominids. Science 326:75–86.
White TD, Johanson DC, Kimbel WH. 1981. Australopithecus afri-canus: its phyletic position reconsidered. S Afr J Sci 77:445–470.
White TD, Suwa G, Asfaw B. 1994. Australopithecus ramidus, anew species of early hominid from Aramis, Ethiopia. Nature371:306–312.
White TD, Suwa G, Asfaw B. 1995. Australopithecus ramidus, anew species of early hominid from Aramis, Ethiopia. [Corri-gendum]. Nature 375:88.
White TD, Suwa G, Simpson S, Asfaw B. 2000. Jaws and teethof Australopithecus afarensis from Maka, Middle Awash,Ethiopia. Am J Phys Anthropol 111:45–68.
White TD, WoldeGabriel G, Asfaw B, Ambrose S, Beyene Y, Ber-nor RL, Boisserie JR, Currie B, Gilbert H, Haile-Selassie Y,Hart WK, Hlusko LJ, Howell FC, Kono RT, Lehmann T, Lou-chart A, Lovejoy CO, Renne PR, Saegusa H, Vrba ES, Wessel-man H, Suwa G. 2006. Asa Issie, Aramis and the origin ofAustralopithecus. Nature 440:883–889.
Williams FL, Clymer G, Proctor D. 2006. ‘‘Is Papio robinsoni orPapio angusticeps a better dietary ancestor for Papio ursinus?Am J Phys Anthropol 129 (Suppl 42):188.
Williams JM, Lonsdorf EV, Wilson ML, Schumacher-Stankey J,Goodall J, Pusey AE. 2008. Causes of death in the Kasekelachimpanzees of Gombe National Park, Tanzania. Am J Prima-tol 70:766–777.
Williams FL, Patterson JW. 2010. Reconstructing the paleoecol-ogy of Taung, South Africa from low magnification of dentalmicrowear features in fossil primates. Palaios 25:439–448.
Winter K, Smith JAC, editors. 1996. Crassulacean acid metabo-lism: biochemistry, ecophysiology and evolution. Berlin:Springer-Verlag.
WoldeGabriel G, Ambrose SH, Barboni D, Bonnefille R, Bre-mond L, Currie B, DeGusta D, Hart WK, Murray AM, RennePR, Jolly-Saad MC, Stewart KM, White TD. 2009. The geolog-ical, isotopic, botanical, invertebrate, and lower vertebratesurroundings of Ardipithecus ramidus. Science 326:65–69.
WoldeGabriel G, White TD, Suwa G, Renne P, de Heinzelin J,Hart WK, Heiken G. 1994. Ecological and temporal placementof early Pliocene hominids at Aramis, Ethiopia. Nature371:330–333.
Wolpoff MH. 1973. Posterior tooth size, body size and diet inSouth African gracile australopithecines. Am J Phys Anthro-pol 39:375–394.
Wood BA. 1991. A palaeontological model for determining the limitsof early hominid taxonomic variability. Paleontol Afr 28:71–77.
Wood BA, Constantino P. 2007. Paranthropus boisei: fifty yearsof evidence and analysis. Yearb Phys Anthropol 50:106–132.
Wood BA, Richmond BG. 2000. Human evolution: taxonomy andpaleobiology. J Anat 196:19–60.
Wood BA, Strait DS. 2004. Patterns of resource use in earlyHomo and Paranthropus. J Hum Evol 46:119–162.
Wood BA, Wood CW, Konigsberg LW. 1994. Paranthropus boisei:an example of evolutionary stasis? Am J Phys Anthropol95:117–136.
Wright BW. 2005. Craniodental biomechanics and dietary tough-ness in the genus Cebus. J Hum Evol 48:473–492.
Wroe S, Ferrara TL, McHenry CR, Curnoe D, Chamoli U. 2010.The craniomandibular biomechanics of being human. Proc RSoc B 277:3579–3586.
Wyckoff RWG, Doberenz AR. 1968. The strontium content of fos-sil teeth and bones. Geochim Cosmochim Acta 32:109–115.
Wynn JG. 2000. Paleosols, stable carbon isotopes, and paleoen-vironmental interpretation of Kanapoi, Northern Kenya. JHum Evol 39:411–432.
Wynn JG, Alemseged Z, Bobe R, Geraads D, Reed D, Roman DC.2006. Geological and palaeontological context of a Pliocene juve-nile hominin at Dikika, Ethiopia. Nature 443:332–336.
317MICROWEAR, ISOTOPES, AND HOMININ PALEOECOLOGY
American Journal of Physical Anthropology





























