Embed Size (px)
Citation preview

Pal&ontologische Zeitschrift 2005, Vol. 79/2, p. 263-284, 30-06-2005
Silurian Leptaeninae (Brachiopoda) from Gotland, Sweden
OLE A. HOEL, Uppsala
with 12 figures
HOEL, O.A. 2005. Silurian Leptaeninae (Brachiopoda) from Gotland, Sweden. - Paliiontologische Zeitschrift 79 (2): 263-284, 12 figs., Stuttgart, 30.6.2005.
Abstract: Strophomenid brachiopods belonging to the genera Leptaena and Lepidoleptaena are described from the uppermost Llandovery - Ludlow succession of Gotland, Sweden. In Gotland, Lepidoleptaena comprises the single species L. poulseni, and Leptaena includes four species: L. rhomboidalis, L. sperion, L. depressa and L. parvirugata n. sp. L. depressa shows a considerable amount of morphological variation, which is recognised in the two new sub- species Leptaena depressa visbyensis n. ssp. from the Llandoverian - Wenlockian Visby Formation, and L. depressa lata n. ssp. from the mid-Wenlockian Slite Group. The distribution of the different species is largely substrate-de- pendent, with Leptaena rhomboidalis and Lepidoleptaena poulseni adapted to high-energy environments with firm substrates, whereas the remaining species preferred fine-grained substrates in low-energy environments.
Keywords: Silurian ° Gotland ° Brachiopoda ° Strophomenida ° Leptaeninae ° Leptaena ° Lepidoleptaena
Kurzfassung: Die zu den Gattungen Leptaena und Lepidoleptaena gehrrenden strophomeniden Brachiopoden aus der Abfolge vom h6chsten Llandovery bis zum Ludlow von Gotland, Schweden, werden beschrieben. Auf Gotland ist Lepidoleptaena nur durch eine einzige Art vertreten: L. poulseni; Leptaena hingegen umfasst dort vier Arten: L. rhomboidalis, L. sperion, L. depressa und L. parvirugata n. sp. Innerhalb von L. depressa gibt es eine betrachtliche morphologische Variabilitat, weshalb hier zwei neue Unterarten aufgestellt werden: L. depressa visbyensis n. ssp. aus der Visby-Formation (Llandoverium - Wenlockium) und L. depressa lata n. ssp. aus der Slite-Gruppe (mittleres Wenlockium). Die Verteilung der unterschiedlichen Arten ist weitgehend vom Substrat abhiingig, wobei Leptaena rhomboidalis und Lepidoleptaena poulseni an energiereiche Lebensr~iume mit festen Substraten angepasst waren; die tibrigen Arten bevorzugten feinkrrnige Substrate in energiearmeren Lebensriiumen.
Schliisselwrrter: Silur • Gotland ° Brachiopoda ° Strophomenida • Leptaeninae ° Leptaena ° Lepidoleptaena
Introduction
The Si lur ian success ion o f Got land is one o f the most in- tensely s tud ied geo log ica l intervals in the world , y e t there are aspec ts o f its fauna that are not well known. One o f these r e l a t ive ly unchar ted areas is the s t ropho- menid b r a c h i o p o d s , wh ich compr ise about 16 % of de - scribed b r a c h i o p o d species f rom the is land (BASSETT & COCKS 1974). A l t h o u g h the divers i ty is high, the number o f spec imens is genera l ly rather smal l , except at a few hor izons and local i t ies . In a study f rom the Visby and H6gk l in t F o r m a t i o n s at Vattenfal let , Visby , BAS- SETT (1979: f igs. 58--61) showed that s t rophomenids oc- cur in mos t hor izons , but do not domina te the fauna at any level .
The long- rang ing genus Leptaena DALMAN, 1828, is widespread in the m i d - O r d o v i c i a n to mid -Devon ian o f Bal t ica , Laurent ia and Ava lon ia . I t appears to have or ig ina ted in eastern Bal t ica , where the subgenus L. (Septomena) ROC)MUSOKS, 1989, is found as ear ly as in the Llanvi rn ian o f Es tonia . In the L o w e r Caradoc o f the Oslo region , SPJELDNJES (1957: 171) found six species o f Leptaena, at least one o f which be longs to L. (Sep- tomena), and two species o f the c lose ly re la ted genus Kiaeromena SPJELDN/ES, 1957.
Spec imens o f Leptaena most ly compr i se a smal l percentage of the total n u m b e r o f spec imens in any fau- na, but can occas iona l ly be fai r ly c o m m o n at some lo- cal i t ies . The smal l number o f spec imens is p robab ly one reason why they have c o m m o n l y been recorded as Lep-
Address of the author: OLE A. HOEL, Institute for Geosciences, Department of Palaeobiology, Uppsala University, Norbyviigen 22, SE-752 36 Uppsala, Sweden; e-mail <o][email protected]>. - Present address: Palaeontological Museum, University of Oslo, Box 1172, Blindern, NO-0318 Oslo, Norway.
0031- 0220/05/0079- 0263 $ 5.50 © 2005 E. Schweizerbart'sche Verlagsbuchhandlung, D-70176 Stuttgart

264 OLE A. HOEL
taena sp. (see e. g., AMSDEN 1978; BOUCOT et al. 1966; VILLAS & COCKS 1996) or as L. "rhomboidalis" (e. g., BOUCOT 1973). The sometimes bewildering morpho- logical variability makes taxonomic studies difficult without relatively large collections and with good local- ity and facies data. Fortunately, such criteria are availa- ble from Gotland with well over a thousand specimens housed in the Swedish Museum of Natural History, Stockholm, the Swedish Geological Survey, Uppsala and the Museum of Evolution, Uppsala.
The name Leptaena was first published by DAL- MAN (1828), based on the species L. rugosa f rom the Ashgill of Sweden. This species was redescribed by SPJELDNAES (1957), who selected a lectotype, and by BERGSTROM (1968). The early history of the generic name was reviewed by SPJELDNtES (1957) and KELLY (1967). In studies published before 1950, many leptae- nids were assigned to Leptaena rhomboidalis (WAHL- ENBERG, 1818), but this species seems to be confined to the lower Wenlock of Gotland. DAVIDSON (1866--1883) described all British strophomenids known at his time, including specimens now known to belong to five spe- cies of Leptaena and one species of Lepidoleptaena HAVLIEK, 1963 (see COCKS 1978); two of late Ordovic- ian age (L. tenuistriata J. de C. SOWERBY, 1839, and L. semiovalis (M'CoY, 1846)), two from the Llandovery (L. contermina COCKS, 1968, and L. zeta LAMONT, 1947), one from the Wenlock (L. depressa (J. de C. SOWERBY, 1824)) and one from the Ludlow (Lepidolep- taena poulseni (KELLY, 1967)). DAVlDSON (1866- 1883) used the generic name Leptaena for the plecto- dontoideans, while the leptaenid specimens were de- scribed under the generic name Strophomena.
The first major work on Scandinavian Silurian stro- phomenids was that of HOLTEDAHL (1916) who de- scribed some 40 different species from the Ordovician/ Silurian of the Oslo Region. Five of these were assigned to Leptaena, and only L. rhomboidalis was named as ranging into the Silurian. This species name was applied to several different forms ranging from the Mid-Ordovi- cian ("etage 4b") to the Upper Wenlock ("etage 9e"). This material evidently contains several species, as shown by BAARLI (1995), who described four species of Leptaena from the Llandovery of the central Oslo re- gion. All of these are also known from the British Isles. In addition, she erected a new genus, Crassitestella, based on the species "Leptaena'" reedi COCKS, 1968, which also occurs in the Oslo region. The specimens fig- ured by HOLTEDAHL (1916) from the Wenlock (his stag- es 8 and 9) are here referred to Leptaena sperion BASSETT, 1977 (see below).
POULSEN (1943: 17, text-figs. 4-7) compared two specimens of Leptaena from the Silurian of Greenland with L. rhomboidalis and L. depressa, and figured sev- eral specimens of both species from Gotland. He was the first to draw attention to the presence of both species in Gotland and made clear the main differences between them.
SPJELDNtES (1957) figured material of Leptaena from Gotland in his study of Upper Ordovician stropho- menids of the Oslo region. His figures show ventral in- teriors of L. depressa (J. de C. SOWERBY) (pl. 7 figs. 9, 15) and L. depressa visbyensis n. subsp, described be- low (pl. 7 fig. 3) from Visby, Gotland.
KELLY (1967), COCKS (1967, 1968) and BASSETT (1974: 1977) described the British Llandoverian to Wenlockian strophomenids, while COCKS (1978) re- viewed all known British early Palaeozoic brachiopods, listing 60 species belonging to 14 genera of the subfami- ly Leptaeninae.
BASSETT & COCKS (1974) figured type specimens of L. rhomboidalis and illustrated the vertical distribu- tion of this species and that of L. depressa in Gotland. They also noted that the diversity of Leptaena was prob- ably higher than previously thought, and the present pa- per shows that this is indeed the case. A range chart for all known leptaenines in Gotland is shown in Fig. 1.
Systematic Palaeontology
The classification used is after WILLIAMS et al. (2000). Abbreviations: BU - Department of Geology, University of Birmingham; NHM - Natural History Museum, London; BGS - British Geological Survey, Keyworth; SGU - Geological Survey of Sweden, Uppsala; SMNH - Naturhistoriska Riks- museet, Stockholm, Sweden. All figured specimens are housed in SMNH.
Order Strophomenida (~PIK, 1934 Superfamily Strophomenoidea KING, 1846 Family Rafinesquinidae SCHUCHERT, 1893
Subfamily Leptaeninae HALL ~,: CLARKE, 1894
Genus Leptaena DALMAN, 1828
Type species: Leptaena rugosa DALMAN, 1828, from the Ash- gill Dalmanitina beds of F~rdalaberg (now Varvsberget), V/~sterg6tland, Sweden. The lectotype (SMNH Br.10435) of this species was figured by SPJELDN~S (1957: pl. 7 fig. 4), who also discussed the history of the generic name.
Remarks: SPJELDN/ES (1957: 173) remarked that most Ordovician leptaenids have flabellate ventral muscle scars, while the Silurian ones have more rounded and well-defined scars. He thus suggested that the name Leptagonia M'CoY, 1844, might be used as a subgenus for the Silurian forms. HAVLi~EK (1967) followed this, raising Leptagonia to generic status. This seems to be an over-splitting approach, based mostly on stratigraphic arguments, and relatively useless as most leptaenine species around the world are known from the exterior only. On the other hand, knowledge of the external or- namention is usually not enough to distinguish species because there is usually a wide variability. This is, e. g., illustrated in Fig. 10 by the ratio "rugae number/disc length" of the Gotland species. This shows that L. rhom- boidalis and L. depressa depressa are tightly clustered, while the other species are widely variable. Fig. 11

Silurian Leptaeninae (Brachiopoda) from Gotland, Sweden 265
©
Lithostratigraphy Graptolite biozones S W 1NE
fo mosu NIIIIIIIIIIIII
Hemse tumescens ~ - - " -
~ scanicus
nilssoni Klinteberg
ludensis
o nassa
lundgreni
ellesae
linnarssoni O O .~ rigidus
m riccartonensis
Slite
H6gldint
%.,
murchisoni Upper Visby crenulata Lower Visby
griestoniensis Not exposed on Gotland
Fig. 1. Stratigraphic distribution of leptaenid species in Gotland.
|
lsp.A
' I Ii ,,spB | • |
m
shows that the length/width ratio also is insufficient as all species are relatively tightly clustered along the same growth trajectory. In the Treatise (COCKS & RONG 2000: 247), Leptagonia is defined as a Devonian to Car- boniferous genus similar to Leptaena, but having raised muscle platforms in both valves. This is not seen in any of the species from Gotland, and they are thus assigned to an undivided genus Leptaena. Llandoverian species of Leptaena fall into three groups (COCKS 1968), name- ly one characterized by an oval ventral muscle area, one with parallel-sided muscle area, and one group of small species that are not well characterized. These may be separable into subgenera, but this is not relevant to this study, as all of the Wenlock and Ludlow species of Brit- ain and Scandinavia (where the interiors are known) have a rounded ventral muscle scar.
Leptaena rhomboidalis (WAHLENBERG, 1818) Figs. 2, 10, 11A
"1818 Anomites rhomboidalis WAHLENBERG: 65. pt 1828 Leptaena depressa (J. de C. SOWERBY, 1825). -
DALMAN: 107, non pl. 1 fig. 8. (non J. de C. SOWER- BY, 1824).
1943 Leptaena rhomboidalis (WILCKENS). - POULSEN: 18, figs. 6b, 7.
1967 Leptaena rhomboidalis (WAHLENBERG). -- KELLY: 594, pl. 98 figs. 1--3.
1974 Leptaena rhornboidalis (WAHLENBERG).- BASSETT & COCKS: 14, pl. 2 figs. 7--8.
Neotype: BU 878, from the Upper Visby or H6gklint Forma- tion, Visby, Gotland; selected by KELLY (1967: 594, pl. 98 fig. 1). The specimen was refigured by BASSETT & COCKS (1974: pl. 2 figs. 7a~l).

266 OLE A. HOEL
Gotland material: Over 250 specimens, internal as well as ex- ternal preservation common.
Diagnosis: Species of Leptaena with very high and steep rugae separated by deep interspaces. Large fo- ramen. Adductor scars in the ventral valve broad. Cardi- nal process lobes divergent.
Description: Shells are from 0.3 to 1.1 m m thick, the outline is subrectangular to subtriangular, often with wide alae (Figs. 2H, J, S). The disc is usually more or less semi-circular. Ventral disc is usually not flat, but changes growth direction dorsally at each ruga with a different amplitude in each specimen, so that some spec- imens have very convex ventral valves (Figs. 2E-G), while some are flat (Figs. 2A-C) . The trail is usually rather short, but in some specimens it becomes longer than the disc length (Figs. 2C, G, K) (this variation does not seem to be correlated with the ontogenetic stage of the specimens). Geniculation angle is variable, from 70- 90 °. Ventral umbo is usually prominent, but most o f it is always destroyed by the growth of the pedicle foramen, which is always open and large (Figs. 2M, N, P). Ventral interarea is planar and apsacline. Inclination of both in- terareas is about 45 ° (Fig. 2B). Pseudodeltidium is small and convex (Fig. 2X); in most specimens, it is broken off and missing (Figs. 2L, Q, V). Dorsal umbo is con- cave and inconspicuous. Dorsal interarea is planar and anacline. The hinge occupies about half the total width. The notothyrium is relatively narrow, occupying about 5-6 % of total hinge width (Fig. 2R), and covered by a convex chilidium, which is downfolded in a groove be- tween the cardinal process lobes. Concentric ornament consists of usually uninterrupted rugae, which are very prominent with deep trenches between them (Figs. 2K, L). They have a steep side towards the posterior and a gently sloping side towards the margin. Usually about eight rugae were developed before geniculation was in- itiated. Radial ornament of slender, rounded costellae, which cover the whole outer surface including the trail (Figs. 2E, J), and increase in number mainly by interca- lation. Ventral interior: The teeth are relatively small and weak (Fig. 2V), triangular, and somewhat striated on the pos- terior face. The muscle field is well-impressed, and al- most diamond-shaped. The diductor scar area is smooth, while the adductor scar is comparat ively wide, occupy- ing about 25 % of the width of the muscle area (Figs. 2I, V). The bounding ridge around the scars rises quite high above the valve floor, and curves posteriorly at its anter- omedian margin. At this point a myophragm leaves the muscle area, bearing a distinct swelling before it merges with the floor of the extra-muscular area at the point of geniculation. In the extra-muscular area, the inner sur- face is covered by taleolae, arranged in radial rows, probably marking vascular traces, but they are thinner and somewhat sparser than in L. depressa. The rngae are distinct on the inside of both valves, since the secondary shell growth is fairly thin.
Dorsal interior: In most specimens, a bounding ridge (diaphragm) defines the disc (Figs. 2S, T). This ridge is usually of uneven height, and often has a posteriorly di- rected kink anteromedially, truncating the rounded out- line of the shell. The cardinal process lobes are coffee bean-shaped in ventral aspect, strong and divergent at about 45 °. Between the cardinal process lobes, is a deep, rounded depression (Fig. 2U). The diductor muscle at- tachment faces are triangular, pointing posterolaterally, their surface irregular (Figs. 2R,W). Sockets are deep and triangular, and excavated posteriorly well under- neath the interareas. They are covered by secondary shell only on the surface nearest to the muscle platform. The muscle field is raised above the valve floor, with a distinct, rounded rim around the adductor scars, but of- ten not well defined anteriorly, even in gerontic speci- mens (Figs. 2D, O, U). The adductor scars are rounded or triangular, crossed by a straight transmuscular sep- turn, which with the median septum divides the muscle attachment area into four parts. The posterior pair is the largest, and in older specimens this is more deeply im- pressed than the anterior pair. The muscle attachment area occupies about of the disc width. The r im of the muscle platform continues forwards and encircles the structures named "anterior scars" by KELLY (1967). These two variously deep pits are about half as wide as the adductor scars, and are divided by the medial myo- phragm, which does not reach the anterior margin of the disc. Usually the anterior scars are fairly weakly defined and shallow, except in gerontic specimens.
As with the rest of the extramuscular area, the ante- rior scars are mostly covered in taleolae, except for a small, smooth area just in front of the rim around the ad- ductor scars. This is also the most deeply impressed point of the anterior scars in gerontic specimens. Vascu- lar traces are present on the disc in some specimens, but usually they are only visible as sinuous, thin canals on the trail.
Remarks : The history of the species name L. rhomboi- dalis was reviewed by KELLY (1967), who made it clear that the species L. depressa (J. de C. SOWERBY, 1824), which often has been regarded as a junior synonym of L. rhomboidalis, is valid. In fact, L. rhomboidalis is known only from Gotland, where it was confined to moderate energy reef environments during the early Wenlockian. The deep ruga pattern typical ofL. rhomboidalis is prob- ably connected with the environment. The same mor- photype is present, e .g. , in the Prague basin (see HAVL[~EK 1967 and HAVL[(2EK 8~; STORCH 1990), where leptaenids with deep and few rugae (L. rugaurita and L. lemniscata) are found in the shallow-water, higher ener - gy part of the basin, whereas forms with more subdued ornament are found in the deeper-water areas (L. depres- sa vulgaris and L. mendosa).
Occurrence: In the collections of the SMNH, there are only six specimens labelled as coming from the Visby Formation,

Silurian Leptaeninae (Brachiopoda) from Gotland, Sweden 267
Fig. 2. - A-X: Leptaena rhomboidalis (WAHLENBERG, 1818). A-C. SMNH Br116187; unknown locality; upper Visby Formation; conjoined valves. A: Posterior view; note round foramen. B: Posterolateral view. C: Lateral view parallel to the pedicle foramen; note the flat disc. D. SMNH Br120855; Sumpklint, Fleringe parish; HSgklint Formation; dorsal valve interior view. E-G. SMNH Br116186; locality as for Figs. A-C; conjoined valves. E: Posterior view. Note the large foramen. F: Posterolateral view. G: Lateral view parallel to the foramen. Note the dorsal deflection of the growth direction at each ruga, and the probable life orientation vertical to the substrate. H. SMNH Br116122; west of Lansa, F&r6 parish; Slite Group, ventral valve exterior view. I. SMNH Br138044; Irevik, Hangvar parish; H6gklint Formation; incomplete ventral valve interior view; note the large adductor scars. J. SMNH Br137849; Bals klint; upper Visby For- mation; conjoined valves with wide alae; ventral view. K. SMNH Br116283; Storbrut, Stenkyrka parish; HSgklint For- mation; conjoined valves; ventral view. Note the protruding anteromedial portion of the trail. L, Q, V. SMNH Brl 16276; Kopparvik, Visby; HSgklint Formation; ventral valve. L: Exterior view. Q: Interior view. V: Detail of visceral area. Note collapsed pseudodeltidium. M, N, X. SMNH Br116301; Stuguklint, Stenkyrka parish; HSgklint Formation; conjoined valves. M: Ventral view. N: Posterior view. X: Detail of small pseudodeltidium and massive chilidium. O. SMNH Br116123; west of Lansa, F&r5 parish; Slite Group, dorsal valve interior view. P. SMNH Br116300; Stuguklint, Stenkyrka parish; HSgklint Formation; conjoined valves; ventral view. R, S, W. SMNH Brl 16161 ; £)stra Vi, Visby; Slite Group; dorsal valve. R: Posterior view. S: Interior view. W: Detail of cardinal process lobes. T-U. SMNH Br116305; Stuguklint, Stenkyrka parish; HSgklint Formation; dorsal valve. T: Interior view. U: Detail of muscle scars and median and lateral septa. - Scale bar = 10 mm (Figs. A-T), Scale bar = 5 mm (Figs. U-X).

268 OLE A. HOEL
all from the upper subdivision. The species is especially common in a thin bed just at the base of the main reef bodies in the H6gklint Formation in the central part of the northwest coast (personal observation), and most or all of the Visby bed specimens probably occur just at this level. According to BAS- SETT • COCKS (1974), the species is confined to the upper Visby and H6gklint Formations (riccartonensis Biozone), but conspecific specimens also occur in the lower Slite Group on F~6; these rocks are probably coeval with the HOgklint For- mation further to the southwest (M.G. BASSETT pers. com.). Two probably conspecific specimens are also known from Bara Backe at a high level in the Slite Group, probably just be- low the Bara oolite.
1879
1916
* 1977
?pt 1967
?pt 1990
Leptaena sperion BASSETT, 1977 Figs. 3A-R, 10, 11B
Leptaena rhomboidalis WILCKENS. - - BARRANDE: pl. 41 figs. 28, 33 (non pl. 41 figs. 1-27, 29-32, 34- 38). Leptaena rhomboidalis WILCKENS. -- HOLTEDAHL: 70, pl. 12 figs. 1--2, 5, 8--10, (11 ?), 12 (non BASSETT 1977). Leptaena sperion n. sp. BASSETF: 123, pl. 33 figs. 1- 3. Leptaena depressa vulgaris (BARRANDE). -- HAV- LfCEK: 93, pl. 15 figs. 1, 2, 4, 5, 10, 11 (non pl. 15 figs. 8, 9, pl. 16 figs. 10, 12). Leptaena depressa vulgaris (BARRANDE). -- HAV- LICEK &; STORCH: 72, pl. 12 figs. la, b, 2a, b, 6-8 (non pl. 12 figs. 3-5, 9, 10, text-fig. 9).
Holotype: BGS GSM.12683, ventral valve, from the Wool- hope Limestone Formation, Woolhope, Herefordshire, UK. Figured by BASSETT (1977: pl. 33 figs. la~l). Paratypes: BGS GSM.12681-2, from the Woolhope Lime- stone Formation, Malvern area, UK. Hypotypes from Gotland: 41 specimens; both external and internal preservation.
Diagnosis (revised): Large species of Leptaena with large number of irregular, shallow rugae. Dorsal periph- eral rim very low. Chilidium massive, convex; pseudo- deltidium small and flat. Cardinal process lobes large and diverging. Teeth strong, sockets deep, with accesso- ry denticles along their whole length.
Description: The shells are often large (Figs. 3D, H, L), up to 60 mm wide (mean 25 mm), about twice as wide as long in small specimens. Width increases with size, so that the length approaches 40 % of the width. The disc is semicircular, except that a short part of the anterior margin is parallel with the hinge line. The cardinal ex- tremities extend as rather pointed alae. Both discs are flat, with a sharp geniculation at about 90 ° (Figs. 3F, H), and in most specimens the trail is relatively short (50 % of disc length), although it can exceed the disc length in some specimens. There is no peripheral rim. The ventral umbo is small, and there is no trace of any apical fo- ramen in the large specimens. In smaller ones (Figs. 3C, P) an inner opening of a foramen is seen, and the umbo of another specimen has collapsed (Fig. 3G), probably
into an inner remnant of a foramen. The low ventral in- terarea is poorly preserved. The pseudodeltidium is small to vestigial (Fig. 3N). The dorsal interarea is pla- nar, low and tapers to nothing towards the cardinal mar- gins (Fig. 3M). The notothyrium is triangular, relatively narrow, and covered by a slender, convex chilidium, which is downfolded between the cardinal process lobes (Fig. 3Q). The outer surface is covered by from 9-16 (mean 12) small, low, irregular rugae and the costellae are thin and sinuous, the number increasing by bifurca- tion (Figs. 3A, D, E, I). The costellae are usually even, but in some specimens there area more prominent first order costellae (Fig. 3H). Ventral interior: The delthyrial cavity is broad, bisected by a low ridge which extends along the mid-line from the posterior margin and fuses with the adductor scars (Figs. 3N, P). The teeth are thin, almost ridge-like, di- verging about 60 ° from the mid-line, and the anterolater- al ends are abruptly cut off and hollowed out (Fig. 3P).
Fig. 3. - A-R: Leptaena sperion BASSETT, 1977. A. SMNH Br32310; Kappelshamn, L&rbro parish; ventral valve; ventral view. B, N. SMNH Br116271; Ansarve, Tofta parish; HSgklint Formation; incomplete ventral valve. B: Interior view. N: Detail of muscle scars. Note prominent denticles on the anterior faces of the teeth. C, P. SMNH Br116272; Ansarve, Tofta parish; HSgklint For- mation; ventral valve; interior view. note small, open pedicle tube. D. SMNH Brl 16270; Ansarve, Tofta parish; HSgklint Formation; large ventral valve; exterior view. Note irregular rugae. E. SMNH Br116275b; Fleringe par- ish; H6gklint Formation; ventral valve with irregular growth; ventral view. F-G. SMNH Br32309; Kappel- shamn, L&rbro parish; ventral valve. F: Lateral view showing low trail. G: Ventral view. Note collapsed umbo over internal pedicle tube. H. SMNH Br32368; Unknown locality, possibly near Visby; HSgklint Formation(?); con- joined valves; ventral view. Note that the trail of this specimen is unusually tall (1.5 times disc length). I. SMNH Br116275a; Fleringe parish; H6gklint Formation; ventral valve; ventral view. J, K, O. SMNH Br116258; Norderstrand, Visby; lower(?) Visby Formation; dorsal valve. J: Interior view. K: Posterior view. O: Detail of car- dinal process lobes. L-M, Q-R. SMNH Br116273; An- sarve, Tofta parish; HSgklint Formation; dorsal valve. L- M: Interior and posterior views. Q-R: Detail views of the muscle scars and cardinal process lobes. - S-Z: Leptae- na parvirugata n. sp. S. SMNH Br138050, Valleviken 1, Rute parish; exterior view of dorsal valve. T. Holotype SMNH Br138049c, Valleviken 1, Rute parish; interior view of dorsal valve. LI-V. SMNH Br138049b, Valleviken 1, Rute parish; dorsal valve. U: Detail of muscle scars and cardinal process lobes. Note denticles on socket ridges. V: Interior view. Note low bounding ridge. W, Z. SMNH Br138049a; Valleviken 1, Rute parish; ventral valve. W: Interior view. Note the round muscle field and smooth shell surface. Z: Detail of Muscle scars. Note well-developed callist in the delthyrial chamber. X. SMNH Br32641, Follingbo parish. Exterior view of ven- tral valve. Y. SMNH Br129620, Gane, B&I parish. Exteri- or view of ventral valve. Note absence of rugae. - Scale bar = 10 mm (Figs. A-M, S-T, V-Y), Scale bar = 5 mm (Figs. N-R, U, Z).

Silurian Leptaeninae (Brachiopoda) from Gotland, Sweden 269
The teeth bear strong crenulations on both sides. The muscle area is oval to heart-shaped, about as wide as long, occupying about 40 % of disc width. The muscle bounding ridges rise a little above the valve floor; they do not meet in the middle, but bend anteriorly. The ad- ductor scars consist of two lanceolate pairs, occupying about 30 % of the width of the muscle area. The two pairs are separated by a slender myophragm, which in one specimen (Figs. 3B, N) ends anteriorly in a swollen knob, while in another specimen (Figs. 3C, P) it merges
into the extra-muscular area without swelling. Each ad- ductor scar consists of a medial area next to the myo- phragm, and a longer, lateral area, which curves around the medial area and extends further posteriorly. The di- ductor scars are ovate, striated radially, and extend fur- ther posteriorly than the adductor scars; they occupy the anterior 67 % of the muscle area. No specimen showing all of the extra-muscular area is known, but the surface near to the muscle area is covered by radially arranged tubercles. The vascula media is prominent as it extends

270 OLE A. HOEL
forward from the muscle area (Figs. 3B, C), but other vascular traces are not seen. The rugae are quite well re- flected on the inside of the valve Dorsal interior: The disc is defined by a low rim at the point of geniculation (Fig. 3J), even in large specimens; the rim is less than one mm high (Fig. 3L). The cardina- lia are strong, cardinal process lobes are slightly diver- gent, triangular in shape (Figs. 3K, M, O, Q). They rise from a well developed, anchor-shaped notothyrial plat- form; between them is a deep, rounded depression (Fig. 3R). The notothyrial platform continues forward as a rounded median ridge as far as the anterior margin of the muscle scars, where it thins to a slender median septum. Together the length of the ridge and septum is 65-85 % of the disc length. The sockets are deep, rounded trian- gular, and there are denticulated cavities along the sock- et ridge. The adductor scars are prominent on all studied specimens. They are more or less circular, and each is divided by a transmuscle septum inclined at about 60 ° relative to the hinge line. The anterior muscle scar has the largest area. The anterior scars are not developed, even in large specimens, there is only a pair of ridges that originates at the anterior margin of the muscle field, medially to the transmuscle ridge, and diverge anterola- terally about 70 ° to the hinge line (Fig. 3L). These ridges may extend to the ridge at the geniculation, and may be part of the vascular system. Apart from these two ridges, only a few vascular traces are preserved, and then only on the trail. The disc is covered by fine tubercles ar- ranged in radial rows, and the small rugae are reflected on the inside of the shell.
Remarks: The most remarkable feature of this species is its extremely low filtration cavity resulting from the low geniculation rim in the dorsal valve, and the lack, even in large specimens, of the anterior scars so typical in most other Silurian species of Leptaena. It has several morphological characters that are quite similar to L. rhomboidalis: The cardinal process lobes are triangular, quite massive and divergent, rather than small, ridge- like and parallel as seen in L. depressa and L. parviru- gata n. sp. (see below). The poorly developed anterior scars in the dorsal valve are similar to those in non- gerontic specimens of L. rhomboidalis. The ventral muscle field of these two species also have similar dia- mond-shaped outline with very wide adductor scars, while L. depressa and L. parvirugata n. sp. have round- ed or square muscle fields with quite small adductor scars. L. sperion and L. rhomboidalis also have a very similar temporal distribution, but occur in different facies. The morphology indicates that they are closely related, and their distribution in Gotland suggests that they could be ecophenotypes of the same species. A problem with this interpretation is the occurrence of L. sperion in other areas where a morph like L. rhomboida- lis has not been reported (see below), but in those areas reefs are not developed at this time, which might explain the absence of the L. rhomboidalis morphology.
Occurrence: This species occurs along the northwestern coast of Gotland, mostly in inter-reef facies, and other relatively low-energy environments from the Visby Formation (though only one specimen is known from this level), through the H~Sg- klint Formation and into the middle part of the Slite Group. This is a slightly longer range than it has in the British Isles, where it is only found in the lowermost Wenlock Woolhope Limestone Formation (BASSETT 1977). The species also oc- curs in the Oslo region (HOLTEDAHL 1916: 71, pl. 12 figs. 1, 2, 5, 9--12), where it occurs in the Malm0ya Formation, which is coeval with the Hrgklint Formation. The environment in Oslo is a shallow carbonate platform within Benthic Assem- blage 3--4 (WORSLEY et al. 1983), which suggests a depth of less than 50 m (BRETT et al. 1993). Specimens very similar to those from Oslo were described under the name of by BAR- RANDE (1879), HAVLfCEK (1967) and again by HAVLICEK & STORCH (1990) from the Wenlock of the Czech Republic. The species also occurs in Lockport, New York State, USA (per- sonal observation of specimens in SMNH, Stockholm). Other specimens in this museum from Kaugatoma, Saarema Island, Estonia, look very similar to L. sperion, but are probably of early P~fdolf age.
Leptaena parvirugata n. sp. Figs. 3S-Z, 11C
Derivatio nominis: parvus, lat., insignificant, and ruga, lat., wrinkle; from the lack of clear rugae on the shells. Holotype: SMNH Br138049, dorsal valve, from the Slite marl at Valleviken l, Rute parish, Gotland (Fig. 3S) (for locality data see LAUFELD 1974b: 140). Paratypes: At least 50 specimens, from the collections in the SGU, Uppsala and SMNH, Stockholm; mostly conjoined valves.
Diagnosis: Large species of Leptaena with extremely faint to non-existent rugae. Dorsal peripheral rim very low. Ventral muscle field circular, dorsal muscles scars flabellate. Cardinal process lobes relatively small and parallel.
Description: The disc outline is more or less semi-cir- cular (Fig. 3X), but the shells often have pronounced alae, which makes the outline quite transverse (Figs. 3S, T). The geniculation angle is fairly steep, about 90 °, and the trail is long, about 70 % of the disc length. A small peripheral rim is present in the ventral valve (Figs. 3X, Y). The ventral umbo is small and convex, while the dorsal umbo is strongly concave. The foramen is proba- bly small and certainly plugged at a very early stage, as it is not visible at all from the inside. The pseudodeltid- ium is extremely small, and the ventral interarea is apsa- cline and short, only slightly longer near the delthyrium. The dorsal interarea is anacline and as short as the ven- tral area, but appears to be of equal height along the whole width of the shell. The notothyrium is covered by a small chilidium, which is folded medially between the cardinal process lobes. The exterior sculpture is notably subdued (Figs. 3S, X, Y); the costellae are wide, low and rounded, about 3 per mm, the number increasing by in- tercalation. A few (much less than 10) faint rugae are visible in a few specimens. The dorsal valve is so thin

Silurian Leptaeninae (Brachiopoda) from Gotland, Sweden 271
that the interior radial ridges in front of the muscle area are clearly visible on the outside. Ventral interior (Figs. 3W, Z): The delthyrial cavity is deep. The posterior of the muscle area is filled with sec- ondary shell. The teeth are small. The muscle field is rounded, circular in some specimens, elongated in oth- ers, surrounded by a fairly prominent ridge, which is in- vaginated slightly anteromedially to accommodate the vascula media where they leave the muscle field. The adductor scars are fairly small, with a thin ridge medial- ly. The extra-muscular area is almost smooth, but the costellae are visible on the interior. A thin vascula media is present near the muscle scars, but no other parts of the vascular system are visible. Dorsal interior: The disc is surrounded by an extremely low bounding ridge (Fig. 3V). Seen in profile, the disc bulges ventrally, following the curvature of the ventral valve. The cardinalia are raised on a low, anchor-shaped notothyrial platform. The cardinal process lobes are small and sub-parallel (Fig. 3U). The sockets are narrow and shallow, with denficles along the socket ridge. The muscle field is well defined, the adductor scars are elon- gated anteriorly, each being divided by slender ridges into two subequal parts, one posterolateral and one an- teromedial. There is an extremely faint trace of the an- terior scars, but there are three radial ridges in front of the muscle area, well developed, but thin and low, reaching about halfway to the point of geniculation. On the extra-muscular area, the costellae are visible through the shell. No tubercles protrude from the interi- or surface of the disc, but they are present on the trail and on the alae.
Remarks : This species differs from most other leptae- nids in Gotland by having extremely inconspicuous ru- gae, and by lacking anterior scars and diaphragm in the dorsal valve. It also has an almost circular ventral mus- cle field. The lack of anterior scars in fully grown spec- imens is shared with L. sperion, but it differs from that species in having teeth without denticles and smaller ventral adductor scars, weaker cardinalia, and even low- er dorsal bounding ridge, making the interior volume of a closed shell extremely small.
Occurrence: Within the Slite Group, this form has a distribu- tion similar to Leptaena depressa depressa (i.e., the marl facies), but is confined to that unit, whereas L. depressa de- pressa is found both in older and younger beds.
Leptaena depressa (J. de C. SOWERBY, 1824)
BASSETT & COCKS (1974) recorded this species from the upper Visby Formation, the Slite Group, and the Halla Formation. During the present study, this range has been extended down into the lower Visby Forma- tion and up into the upper Hemse Group. This means that the species has a very long temporal distribution, from the latest Llandoverian to mid-Ludlowian, and the Llandoverian and early Wenlockian specimens could
with some justification be considered a separate spe- cies. In my view, the morphology shows a gradual ev- olution during this interval, and I have therefore chosen to erect two new subspecies to distinguish the earlier specimens f rom the nominal subspecies; L. depressa visbyensis, which is distinguished mainly by its usually small size, numerous, small rugae, and a pear-shaped ventral muscle field, and L. depressa lata, distin- guished by a small trail, but very wide and flattened alae.
Leptaena depressa depressa (J. de C. S O W E R B Y , 1824) Figs. 4, 10, 11D
* 1824 Producta depressa J. de C. SOWERBY: 86, pl. 459 fig. 3, top 2 figures only.
1828 Leptaena depressa (J. de C. SOWERBY). -- DALMAN: 107, pl. 1 fig. 2.
1837 Leptaena depressa (J. de C. SOWERBY). -- HISINGER: pl. 20 fig. 3.
1943 Leptaena depressa (J. de C. SOWERBY). -- POULSEN: 18-19, text-figs. 4, 5, 6A.
1957 Leptaena depressa (J. de C. SOWERBY). -
SPJELDN/ES: 178, text-fig. 39B,pl. 7 figs. 9, 15,pl. 13 figs. 1,3, pl. 14 figs. 3-5, 8.
1967 Leptaena cf. depressa (J. de C. SOWERBY). -- HAV- LfCEK: pl. 16 figs. 8, 9, 13--15.
1974 Leptaena depressa (J. de C. SOWERBY). -- BASSETT & COCKS: 14.
The above list only applies to specimens from Gotland. For a more detailed synonymy, see BASSETT (1974:111). Type material: A "neotype" (NHM B60649), was selected by KELLY (1967: 595, pl. 98 figs. 4, 5). As pointed out by BAS- SETT & COCKS (1974: 14), this is actually the lectotype since it comes from the Sowerby collection. Hypotypes from Gotland: At least 500 specimens, mostly conjoined valves, but also at least 100 specimens with the in- teriors preserved.
Diagnosis: Species of Leptaena with large number of regular rugae and small alae. Chilidium massive, pseu- dodeltidium very small. Inner surface usually strongly tuberculate. Cardinal process lobes relatively very small and parallel. Teeth strong, sockets shallow. See also KELLY (1967: 595).
Remarks : This species was described exhaustively by BASSETT (1974), and the Swedish specimens do not dif- fer much from the British material. There is a degree of variation in the populations, both within the same hori- zon and between different stratigraphic levels. SPJELD- NiES (1984) commented that specimens from the Halla Formation display dimorphism in several features; cur- vature of the discs, the length of the geniculated margin (presumably the vertical height of the trail), sculpture and filter chamber volume.
In a collection of 83 articulated shells from Blhh~ill (Eksta parish) kindly donated by N. SPJELDNeES, O s l o ,
there is dimorphism in the shell shape and the develop- ment of trails: 56 of the specimens have a rectangular

272 OLE A. HOEL
disc, a marked ventral kink at the point of geniculation (similar to that seen in Fig. 4N), and most of these have only a low trail. The remaining 27 specimens have a more semi-circular disc, practically no ventral kink, and most have a large trail (similar to that seen in Figs. 4 A - B, D-F) . In the largest group, many have the lateral mar- gins extended, making the outline more transverse (sim- ilar to that seen in Figs. 4C, G). The specimens were probably collected at the same time, and at the same ap- proximate level, and may be assumed to represent a nor- mal population. I see no discernable dimorphism in sculpture, though there is considerable variation in both groups. Especially in the development of rugae there are specimens with a large number of regular rugae, while others have very few rugae and the costellae are much more prominent, but this cannot be correlated with the dimorphism in shell outline.
Specimens from the Halla Formation usually have a more rounded shell outline, while the specimens from the Hemse Group generally have a more squared off out- line and the anteromedian projection of the trail de- scribed by BASSETT 1974 (Figs. 4 U - V herein). The Hemse specimens also have weaker, and more vertical cardinalia than the specimens from the Halla Formation, and they are often wider, with long alae. The exaggerat- ed length of the trail shown in some British specimens (BASSETT 1974: pl. 29 fig. 4) is more common in the Hemse specimens than in the Halla material. A single specimen from the high-energy environment of the Klinteberg Formation shows that the shell growth can be modified to suit the environment (Figs. 4W-X) . The specimen has deeper rugae than normal, and a large api- cal foramen is open, presumably to allow anchorage in turbulent waters.
Occurrence: L. depressa depressa ranges from the Slite Group (mid-Wenlock) to the upper Hemse Group (lower Ludlow), almost exclusively in marly, fairly low-energy facies (see Fig. 12). This is in contrast to the British Isles, where it seems to have preferred high-energy environments near bioherms (BASSETT 1974). The species also has a wide geographical distribution: In addition to its occurrence in Britain and in Gotland, specimens figured by SOKOLSKAYA (1954: pl. 4 figs. 1--4) from the Jaani Stage (lower Wenlock) of Saarema, Estonia, probably belong to this species. Speci- mens indistinguishable from L. depressa depressa occur at Dalhousie, New Brunswick, Canada (personal observation in SMNH). The internal morphology of L. depressa depressa is almost identical to that of L. oklahornensis AMSDEN, 1951, from the mid(?)-Silurian of Oklahoma, but the external sculpture of L. oklahomensis is more similar to L. rhomboi- dalis.
Leptaena depressa visbyensis n. subsp. Figs. 5, 10, 11E
pt 1879 Leptaena rhomboidalis WILCKENS.- BARRANDE: pl. 41 fig. 31 (non pl. 41 figs. 1--30, 32-38).
? 1916 Leptaena rhomboidalis (WILCKENS).- HOLTEDAHL: 71,pl. 12 fig. 11.
1974 Leptaena sp. BASSETT & COCKS: 14, tab. 1 (list- ed).
Derlvatio nominis: From the Visby Formation. Holotype: SMNH Br137848, a ventral valve from the lower Visby Formation (uppermost Llandovery) of Nyhamn, Lum- melunda parish, Gotland (Figs. 5H-I, P, S). Paratypes: 127 specimens; conjoined and separated valves
Diagnosis: Small to moderately sized subspecies of Leptaena depressa with subrectangular outline, large number of rugae (8-17), pear-shaped ventral muscle field and geniculation angle around 70 °.
Description: Shells are mostly quite small (disc length about 8-9 mm), but can attain disc length of up to 20 mm in rare specimens (Figs. 5A, B). Shape semicir- cular to somewhat rectangular in small specimens, larg- er specimens becoming proportionally wider, mostly because of extension of the alae (Fig. 5C). Ventral disc is moderately convex (Fig. 5D), dorsal disc is planar to slightly concave. Both are more or less quadrate in out- line The angle between the ventral and dorsal discs var- ies from 45 ° to 70 ° degrees, typically around 60 ° . In the ventral valve, there is no peripheral r im at the point of geniculation. Geniculation angle is typically around 70 ° degrees. Trail is often long, varying from 65 to 150 % of disc length, but normally it is the same length as the disc. Some specimens have almost no trail. Maximum length is about 70 % of maximum width. The ventral umbo usually bears a very small foramen (Fig. 5D), which in small, but gerontic specimens, sometimes is sufficiently large to destroy the pseudodeltidium. In larger speci- mens, the foramen is invariably small and open on the outer surface. On the inside it is usually closed by sec- ondary shell (Fig. 5P), though some specimens have a very small internal opening. Umbones are low. Ventral interarea is plane and apsacline (about 45°), delthyrium is wide with rounded apex. Delthyrial angle 80-90 °. Pseudodeltidium is convex, but small, confined to the uppermost margin of the delthyrium (Fig. 5S). Dorsal interarea is plane and anacline (about 45 °) about as long as the ventral interarea. Notothyrium is broad, filled by the convex cardinal process and covered by the chilid- ium (Figs. 5F, R). The chilidium often reaches up under the pseudodeltidium, but sometimes leaves a small gap between the cardinal process lobes. Ornament is multi- costellate, with costellae very much finer than the rugae (3-5 per ram). Rugae are usually continuous, but some bifurcate or are interrupted (Fig. 5C). Number of rugae is variable, mostly from 8 to 13 rugae for a disc length of about 8-10 mm. Maximum number of rugae is 17 for a disc length of 13.8 mm, while minimum is 5 rugae at 4.1 mm. Ventral interior: The delthyrial chamber is relatively narrow. The teeth are prominent, triangular and blunt. Some specimens have crenulations on their posterior faces. The dental plates are solid. The muscle area (Figs. 5H, L, P) is pear-shaped and deeply impressed, narrow in the posterior third, then widening to become twice as

Silurian Leptaeninae (Brachiopoda) from Gotland, Sweden 273
Fig. 4 . - A-V: Leptaena depressa depressa (J. de C. $OWERBY, 1824). A, D, E. SMNH Br112615; Djupvik, Eksta parish; Halla Formation, Djupvik Member; conjoined valves; ventral, posterior and dorsal views. B, F. SMNH Br32441 ; Klintehamn, Klinte parish; Slite Group; conjoined valves; dorsal and posterior views. - C. SMNH Br32440; Klintehamn, Klinte parish; Slite Group; conjoined valves; ventral view. G-H. SMNH Br32248; Mulde Tegelbruk, FrSjel parish; Halla Formation, Mulde Brick-clay Member; conjoined valves, ventral and posterior views. I, M. SMNH Br138045; Mulde Tegelbruk, FrSjel parish; Halla Formation, Mulde Brick-clay Member; ventral interior. J. SMNH Br32377; Mulde Tegelbruk, Fr6jel parish; Halla Formation, Mulde Brick-clay Member; dorsal interior of gerontic spec- imen. K. SMNH Br137872; Mulde Tegelbruk, FrSjel parish; Halla Formation, Mulde Brick-clay Member; ventral inte- rior. L, Q. SMNH Br138047; Mulde Tegelbruk, Fr6jel parish; Halla Formation, Mulde Brick-clay Member; ventral interior. N. $MNH Br32560; Visne myr kanal, Fardhem parish; Hemse Group; ventral view. O, S. SMNH Br138048; Mulde Tegelbruk, FrSjel parish; Halla Formation, Mulde Brick-clay Member; dorsal interior of relatively young speci- men. P, R, T. SMNH Br32558; Visne myr kanal, Fardhem parish; Hemse Group; ventral interior and posterior views. U-M. SMNH Br32557; Visne myr kanal, Fardhem parish; Hemse Group; ventral valve with extremely long trail; lateral and frontal views. - W-X: Leptaena cf. depressa depressa. SMNH Br129104; Fj&le, Anga parish; Klinteberg Forma- tion; posterior and ventral views of specimen with deep rugae, and large pedicle foramen. - Scale bar = 10 mm (Figs. A-L, N-P, R, T-X), Scale bar = 5 mm (Figs. M, Q, S;).

2 7 4 OLE A. HOEL
Fig. 5. - A-U: Leptaena depressa visbyensis n. subsp. A. SMNH Br32290; Gnisv&rd, Tofta parish; Upper Visby For- mation; conjoined valves; ventral view showing repaired, large shell damage. B. SMNH Brl 16195; Norderstrand, Vis- by; Lower Visby Fm.;dorsal valve; interior view. C-D. SMNH Br116196; Paratype; Norderstrand, Visby; Lower Visby Fm.; conjoined valves; ventral and posterior views showing the small pedicle foramen. E-F, Q-R. SMNH Br116262; Paratype; Norderstrand, Visby; Lower Visby Fm.; dorsal valve; interior view. G. SMNH Br116261 Norderstrand, Vis- by; Lower Visby Fro.; dorsal valve; interior view. H-I, P, S. Hoiotype SMNH Br137848; Nyhamn, Lummelu nda parish; Lower Visby Fm.; ventral valve; interior and posterior views. K. SMNH Br116260; Norderstrand, Visby; Lower Visby Fm.; dorsal valve; interior view. L-M. SMNH Br116199; Norderstrand, Visby; Lower Visby Fm.; ventral valve; interior and posterior views. N-O. SMNH Br32627; Nyhamn, Lummelunda parish; Lower Visby Fm. ;dorsal valve; interior and frontal views. "r-u. SMNH Br116194; Norderstrand, Visby; Lower Visby Fm.; dorsal valve; interior and posterior views. - Scale bar = 10 mm (Figs. A-O, T-U), Scale bar = 5 mm (Figs. P-S).

Silurian Leptaeninae (Brachiopoda) from Gotland, Sweden 275
wide in the anterior part. It occupies about 67 % of the ventral disc length, and from 33 to 50 % of maximum shell width. The muscle bounding ridges are quite low; most of the depth of the muscle field is below the surface of the extra-muscular area (Figs. 5I, M, S). The adductor scars are oval and relatively wide, occupying about 25 - 33 % of the width of the muscle field. They are set on a flat platform. Posteriorly this is not higher than the di- ductor scars, but anterior of the adductor scars the plat- form rises and becomes knoblike. The vascula media originate immediately anterior to this. The diductor scars are each as wide as the adductor scars, deeply stri- ated radially, almost twice as long as the adductors. The anterior ends of the diductor muscle scars have irregular margins. The extra-muscular area and most of the trail is covered by tubercles, in a somewhat radial pattern. The mantle canal system is saccate, with vascula media fair- ly prominent, running straight forwards and usually dis- appearing near the edge of the trail. Thin, radial vascular traces are visible on the trail in some specimens, but there is no obvious connection to the vascula media; in- stead they continue the radial trend of the tubercles on the disc. Dorsal interior: The disc is bounded by a well defined rim at the point of geniculation. This is usually low (Figs. 5E, G, U), but in some gerontic specimens is quite high (Fig. 50) . Cardinalia are strong, on a low, rounded notothyrial platform. The platform bears a deep, round- ed depression between the cardinal process lobes and continues forwards as a rounded ridge separating the ad- ductor scars (Fig. 5Q). The cardinal process lobes are moderately strong and slightly divergent anterolaterally. The gap between them is small. The attachment faces are oval (Figs. 5R, T). Their lower portions are covered by the chilidium. Sockets are deep and triangular and slightly excavated beneath the dorsal interarea. They are underlain by a tongue-like sheet of secondary shell, which extends anterolaterally a short way onto the extra- muscular area, and sometimes forms a ridge dividing the socket f rom this area (Fig. 5R). The muscle field is raised on a platform, into which the muscle scars are deeply impressed in mature specimens. This platform occupies about 40 % of total disc width and 60-70 % of disc length. The posterior scars are more or less circular, divided into two pairs of subequal size by thin ridges, which curve anterolaterally. The scars are limited poste- riorly by the notothyrial platform, and laterally by the socket ridges and anteromedially curving muscle bounding ridges. Anteriorly, the platform continues and tapers to a point. It has broad bounding ridges enclosing two shallow, oval depressions divided by a myophragm, which rises and forms a knob at its anterior end (Figs. 5K, O). These depressions (or anterior scars of KELLY 1967) obviously held no muscular tissue, as they bear the same tubercles as the rest of the extra-muscular area. The extra-muscular area bears prominent tubercles on the whole surface, also on the trail, though the latter are smaller (Figs. 5J, K, N). The vascular system is only
weakly preserved in all specimens, but appears to be lemniscate in shape, which is unusual for Leptaena. It consists mostly of very fine radial canals, especially vis- ible on the trail.
Remarks : This subspecies is most probably directly an- cestral to L. depressa depressa. This earlier form differs mainly in having a larger number of rugae for its usually smaller adult size, a pear-shaped (rather than a subcircu- lar) ventral muscle area with slightly flabellate diductor scars (compare Figs. 41, L, M, P - R with Figs. 5H, L, P), and an apical foramen which is open on the outside. Its cardinal process lobes are slightly less robust and more elongated postero-anteriorly, the shell shape is more subcircular, and the amount of secondary shell secretion is less. The dorsal adductor scar halves are of equal size, whereas in L. depressa depressa the posterolateral half is larger than the anteromedial half (Figs. 4J, O, S). All these differences are of relatively small significance, and thus only warrant differentiation at a subspecific level. The subspecies differs from L. depressa kathekta BASSETT, 1974 (new name in COCKS 1978), which has a less well developed dorsal muscle field and no anterior scars in the dorsal valve, and a rounded ventral muscle scar (see also JIN & CHATTERTON 1997).
Among earlier species, L. depressa visbyensis can be compared with L. purpurea COCKS, 1968, from the late Llandoverian of Shropshire, which it resembles in the shell ornamentation. The most apparent difference is in the different size and thickness of the shell: L. purpu- rea is only about 10 m m wide, while L. depressa visby- ensis is up to 40 m m wide. Another possibly closely allied form is L. cf. depressa from the upper Llandovery - lower Wenlock of Nova Scotia (HARPER 1973: pl. 14 figs. 9-16). This form also has a pear-shaped ventral muscle field, but it is slightly more elongated. At the same time, the ventral muscle field in most of HARPER' s figured specimens (1973: pl. 14 figs. 9-11) is shorter compared to the disc than in L. depressa visbyensis.
Occurrence: In Gotland, this subspecies occurs both in the lower and upper divisions of the Visby Formation, and some specimens probably belonging to this subspecies occur in the Hrgklint Formation. This indicates a terminal Llandoverian to earliest Wenlockian age. A specimen figured by BARRANDE (1879: pl. 41 fig. 31) from the ?Llandovery of the Prague Ba- sin at Tachlowice is remarkably similar to the specimen fig- ured herein (Fig. 5K) and probably belongs to the subspecies. Some specimens of broadly similar age from the Oslo region (e. g., HOLTEDAHL 1916: pl. 12 fig. 11) may belong to this sub- species, but they are usually not sufficiently well preserved for a detailed comparison. Specimens of this subspecies also oc- cur in the Jaani Stage (lower Wenlock) at Jaani church, Esto- nia (personal observation in SMNH, Stockholm)
Leptaena depressa lata n. subsp. Figs. 6, 7, 10, 11F
Derivatio nominis: latus, lat., wide; from the wide lateral alae.

276 OLE A. HOEL
Holotype: SMNH Br137854 (dorsal interior) from the upper- most Slitc Group (early Homerian, Wenlock) at Robbj~ins 2, Klinte parish, Gotland (Fig. 6A). Paratypes: 58 specimens, mostly conjoined valves in SMNH.
Diagnosis: Subspecies of L. depressa with greatly ex- tended alae and lateral part of trail. Pedicle foramen open. Cardinal process lobes weak and slightly diverg- ing. Interior of disc and trail with strong vascular traces.
Description: This subspecies is somewhat similar to L. depressa visbyensis in retaining an externally open pedi- cle foramen on the apex of a fairly protruding ventral beak (Fig. 6D). It also has similar external ornamenta- tion (Fig. 6G), dorsal muscle field (Fig. 6B) and cardi- nalia (Fig. 6C), but differs notably in the shape of the posterolateral areas of the trail. These are laterally elon- gated, sometimes considerably so (Figs. 6E, 7). The trail is usually low and developed only directly in front of the disc (Figs. 6A, F), while the lateral areas are horizontal. This makes the outline very transverse and rectangular (Fig. 6H). In some specimens, the pseudodeltidium is larger than the chilidium. The disc is more or less pen- tagonal in some specimens, not round as in L. depressa
visbyensis or rounded square as in L. depressa depressa, with a raised rim around it in the dorsal valve. In the ventral valve there is no trace of such a marginal rim. It may be a variant of L. depressa visbyensis specialised for living on thixotropic substrates, and the specimen in Figs. 6B-C is more or less transitional between the two forms, but the differences in morphology and distribu- tion are distinct enough for a separation.
Occurrence: This subspecies occurs only in the Slite Group, in a small area around Klintehamn in the west central part of the Island, in the deeper, more muddy part of the platform.
Genus Lepidoleptaena HAVL~EK, 1963
Type species: Strophomena rhomboidalis WILCK. sp. var. lepidula BARRANDE (1879: pl. 41 figs. 10a, b); from the Kone- prusy Limestone (Lower Devonian, Pragian Stage) of Kone- prusy, Bohemia, Czech Republic; by original designation of HAVLi~EK (1963: 224).
Remarks : In the diagnosis of this genus given by HAV- Lf~EK (1967: 107), the only clearly unique characters for definition are the presence of a high bounding ridge
F i g . 6 . - A-H: Leptaena depressa lata n. subsp.; all from the Slite Group. A. Holotype SMNH Br137854; Robbj&ns 2, Klinte parish; dorsal valve with wide, square alae; interior view. B-C. SMNH Br32745; V&stergarn, Sanda parish; dorsal valve with almost normal alae; interior and posterior views. Note the concentric arrangement of the tubercles on the disc. D. SMNH Br32748; V&stergarn, Sanda parish; conjoined valves; posterior view parallel to the probable orientation in life. Note the small apical foramen similar to the condition in L. depressa visbyensis. E-F. SMNH Br134087; Klintehamn, Klinte parish; incomplete dorsal valve with extreme development of the alae. E: Interior show- ing the well-developed mantle canals. F: Frontal view showing the small height of the anterior geniculated margin. G-H. SMNH Br32747; V&stergarn, Sanda parish; conjoined valves. G: Ventral view perpedicular to the disc. H: Ven- tral view perpendicular to the surface of the alae. - Scale bar = 10 ram.

Silurian Leptaeninae (Brachiopoda) from Gotland, Sweden 277
Fig. 7. Reconstruction by photomosaic of the interior of the dorsal valve of Leptaena depressa lata n. subsp. based on the specimens figured in Figs. 6B and E. - Scale bar = 10 mm.
ably two species of Lepidoleptaena from the Lochkov- ian - Pragian of Yukon, Canada. BOUCOT (1973: pl. 6 fig. 16) figured a specimen with Lepidoleptaena-like or- namentation from beds of probable P~fdolian age in the Moose River area, Maine, USA. The specimens de- scribed by AMSDEN (1958: 86, pl. 3 figs. 10-14) from the lower Devonian Bois d 'Arc Formation, Oklahoma, as Leptaena cf. rhomboidalis probably also belong to Lepidoleptaena and are very similar to L. poulseni, ex- cept that the ventral valve seemingly lacks the high bounding ridge around the disc; this might indicate that the latter is not a stable generic character, in which case the two genera may be distinguishable only on a subge- neric level.
around the ventral disc and the strong socket ridges. In addition, the following characters distinguish Lepido- pt 1871 leptaena from Leptaena: The shape of the disc-bound- ing ridges in both valves are different: In Leptaena, the ridges make an even curve inwards posteriorly, whereas * 1967 in Lepidoleptaena, the ridges curve inwards first, but 1974 then bends outwards again as they approach the interar- 1977 ea (Figs. 8I-L, N). In Gotland at least, the genus can be
? 1989 distinguished from Leptaena also by the ornamentation; the notably large number of regular rugae in Lepidolep- 1997 taena (Figs. 8A, E, H) differs from the smaller number of rather irregular rugae in Leptaena (e. g., Fig. 4N). In smaller specimens of Lepidoleptaena, the pseudodelti- dium is much larger than the chilidium (Fig. 9E). This changes during growth, as the pseudodeltidium is con- sumed by the growth of the large apical foramen (Fig. 8F).
Distribution of Lepidoleptaena: BASSETT (1977) assigned the following species to Lepidoleptaena: • Strophomena rhomboidalis var. lepidula BARRANDE
(1879: pl. 41 figs. 10a-b), Gedinnian, Prague. • Lepidoleptaena sp. JOHNSON, BOUCOT & MURPHY (1973:
36), Gedinnian, Nevada. • Lepidoleptaena sp. BASSETT & COCKS (1974: 15), Hemse-
Sundre Formation (Ludlow), Gotland. • Leptaena poulseni KELLY (1967: 597), Much Wenlock
Limestone Formation, Dudley.
Several other authors have figured specimens here con- sidered to belong to Lepidoleptaena: ZHANG (1989: pl. 4 figs. 13, 17, 21, 26, 31, 33, 34) figured specimens of Leptaena sp. Form 1, which is very similar to Lepido- leptaena in the wide alae (figs. 17, 21), the angled cardi- nal process lobes (figs. 26, 31, 33), the curved teeth ridges (straight in Form 2), and especially in the shape of the ridge around the dorsal disc (fig. 31), and the large, open pedicle foramen (figs. 13, 34). This might be the earliest known member of the genus as it is probably of early Wenlockian age. The species described by ZHANG as Leptaena Form 2 is more similar to ordinary species of Leptaena. LENZ (1977: 70, pl. 8 figs. 42, 45-- 55, pl. 9 figs. I--31) figured specimens referable to prob-
Lepidoleptaena poulseni (KELLY, 1967) Figs. 8, 9A-G, 10, 11G
Strophomena rhomboidalis ( W I L C K E N S ) . - D A V I D -
SON: 281, pl. 39 figs. 1,10, 11, 15 (non pl. 39 figs. 2- 9, 12--14, 16-21). Leptaenapoulseni KELLY: 597, pl. 98 figs. 10-13. Lepidoleptaena sp. BASSETT & COCKS: 15. Lepidoleptaena poulseni ( K E L L Y ) . -- BASSETT: 1 3 2 ,
pl. 35 figs. 1-9. Leptaena sp. Form 1 ZHANG: 106, pl. 4 figs. 8--10, 13, 17, 18, 21,22, 25, 26, 29--34. Lepidoleptaena poulseni ( K E L L Y ) . -- JIN & C H A T -
TERTON: 30, pl. 19 figs. 1-20.
Holotype: BU 889, dorsal valve (showing interior only), fig- ured by KELLY (1967: pl. 98 fig. 12). It was refigured by BAS- SETT (1977: pl. 35 figs. la, b). KELLY's paratypes (BU 890 and BU 891) were figured by BASSETT (1977 pl. 35 figs. 2, 3). Hypotypes from Gotland: Over 300 specimens, conjoined valves and interiors.
Diagnosis: see KELLY (1967: 597).
Remarks: BASSETT (1977: 132) described the species in detail from South Wales and the Welsh Borderland, and the Swedish specimens do not exhibit any great differ- ence other than to emphasize the generally robust mor- phology (Figs. 8J-N) and the large amount of encrustation suffered by the Gotland specimens. The number of rugae is about 11-12 in adults (12-16 in Brit- ish material) (Figs. 8A, E, G). The Swedish specimens are from the late Ludlow, while the British material is from the late Wenlockian. Many Swedish specimens reach geniculation (maturity?) at a width of only about 15 mm (Figs. 9D, F, G), even though the largest individ- uals have a width of up to 50 mm (Figs. 9A-C). The trail is often quite long in large specimens (Figs. 8B, D), but most small specimens have very short trails (Fig. 9F). An exception to this is seen in Fig 9G, where the brachi- opod grew a long trail in an ultimately unsuccessful at- tempt to avoid being smothered by an encrusting bryozoan colony. Another encrusting bryozoan colony has preserved the shape of the brachiopod's pedicle (Fig. 9A). The specimens figured by JIN & CHATTER- TON (1997) are also of late Wenlockian age, and show
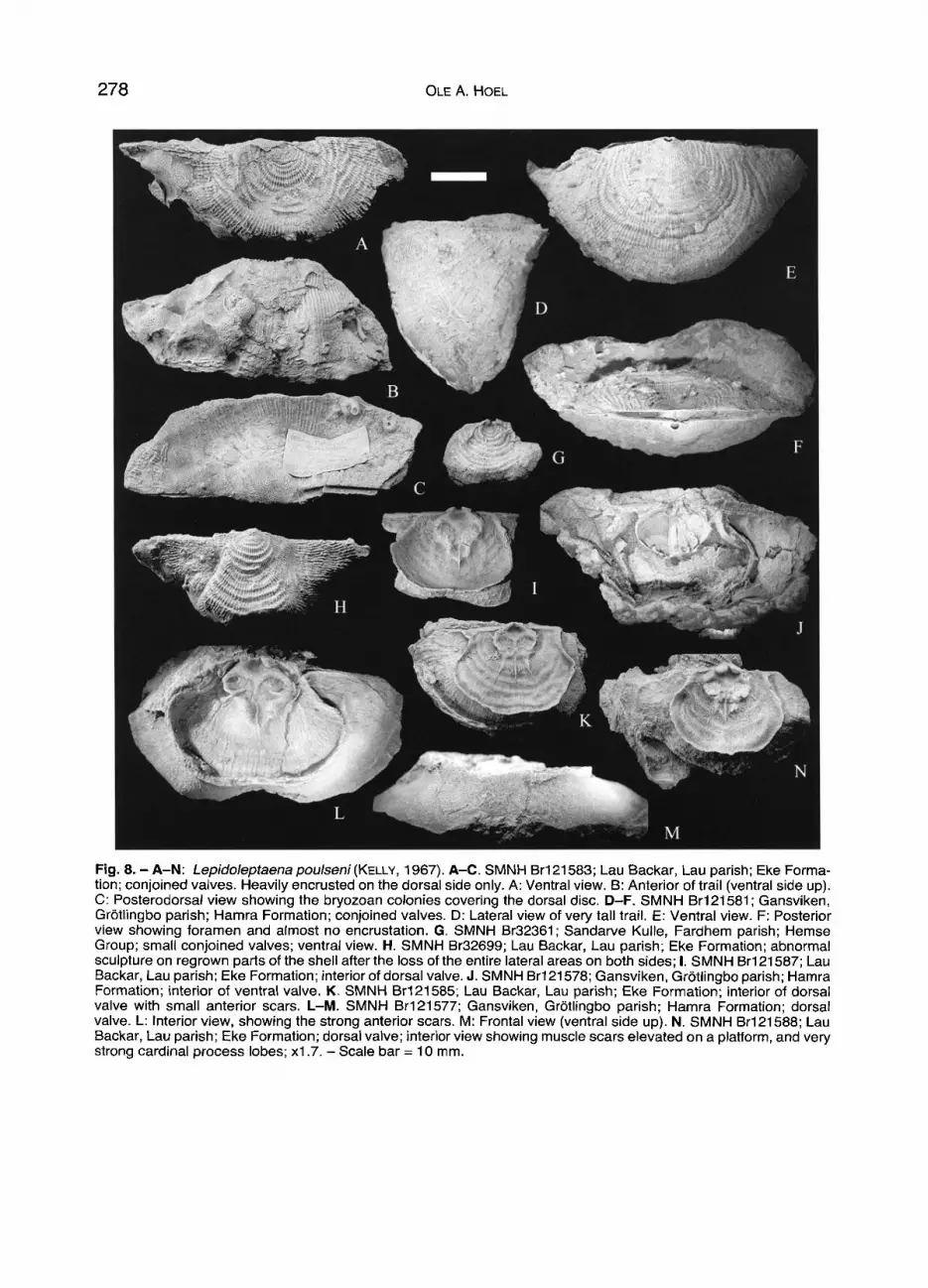
278 OLE A. HOEL
Fig. 8 . - A-N: Lepido/eptaena pou/seni (KELLY, 1967). A-C. SMNH Br121583; Lau Backar, Lau parish; Eke Forma- tion; conjoined valves. Heavily encrusted on the dorsal side only. A: Ventral view. B: Anterior of trail (ventral side up). C: Posterodorsal view showing the bryozoan colonies covering the dorsal disc. D-F. SMNH Br121581; Gansviken, GrStlingbo parish; Hamra Formation; conjoined valves. D: Lateral view of very tall trail. E: Ventral view. F: Posterior view showing foramen and almost no encrustation. G. SMNH Br32361; Sandarve Kulle, Fardhem parish; Hemse Group; small conjoined valves; ventral view. H. SMNH Br32699; Lau Backar, Lau parish; Eke Formation; abnormal sculpture on regrown parts of the shell after the loss of the entire lateral areas on both sides; I. SMNH Br121587; Lau Backar, Lau parish; Eke Formation;interior of dorsal valve. J. SMNH Br121578; Gansviken, GrStlingbo parish; Hamra Formation; interior of ventral valve. K. SMNH Br121585; Lau Backar, Lau parish; Eke Formation; interior of dorsal valve with small anterior scars. L-M. SMNH Br121577; Gansviken, GrStlingbo parish; Harnra Formation; dorsal valve. L: Interior view, showing the strong anterior scars. M: Frontal view (ventral side up). N. SMNH Br121588; Lau Backar, Lau parish; Eke Formation; dorsal valve; interior view showing muscle scars elevated on a platform, and very strong cardinal process lobes; x1.7. - Scale bar = 10 mm.

Silurian Leptaeninae (Brachiopoda) from Gotland, Sweden 279
the internal m o r p h o l o g y o f small spec imens , which is not wel l k n o w n f rom Got l and (Figs. 9 B - D ) .
Measu remen t s on the popula t ions f rom the Eke Forma t ion (100 spec imens) and Hamra Format ion (220 spec imens) indicate d imorph i sm; smal l specimens are t ight ly c lus te red a round a 7 x 11.5 m m disc area, while a group o f large spec imens are more loose ly clustered around a disc area o f about 19 x 25 mm. This pattern is also seen in other ratios: max . length /max, width, max. length/disc length and disc length /number of rugae (see Fig . 11G). A s imi lar pat tern is also seen in Leptaena de- pressa depressa (Fig . 11D).
Occurrence: This species is the only leptaenid occurring in the upper Ludlow of Gotland. At the boundary between se- quences III and IV (CHERNS t~z BASSET]? in press), Leptaena depressa depressa has its last occurrence and Lepidoleptaena poulseni appears. L. poulseni occurs in the uppermost part of
the Hemse Group in the east, and in the Eke, Hamra and Sun- dre formations. It appears to be typical of shallow, but not the shallowest water: After both emergence events of the Ludlow of Gotland (top of the Hemse Group and top of the Burgsvik Formation), it occurs a few metres above each boundary, prob- ably below normal wave base. In Gotland, Lepidoleptaena poulseni occurs in younger rocks than in the British Isles (BASSETI" 1977) and in the Mackenzie District, Canada (JIN & CHATTERTON 1997). In both areas, it occurs in the Wenlocki- an.
?Lepidoleptaena sp. A Fig. 9H
A few spec imens ques t ionably referable to Lepidolep- taena have been found at local i t ies in the Kl in teberg Fo rma t ion at Kl in teberge t and in the lower part o f the H e m s e G r o u p at Ka t thammarsv ik , Br~innklint, L inde Kl int , Sandarve Kul le , Rudvie r and Mannag~rda . These
Fig. 9 . - A-G: Lepidoleptaena poulseni (KELLY, 1967). A. SMNH Br129751; Burgsvik, Oja parish; Hamra Formation; Pedicle shape revealed by bryozoan overgrowth. B. SMNH Br131993; K&ttelviken, Sundre parish; Hamra Formation; fragmentary ventral valve showing the shape of the muscle scars. C. SMNH Br137895; K&ttelviken, Sundre parish; Hamra Formation; young dorsal valve; interior view. D. SMNH Br131994; K&ttelviken, .S. undre parish; Hamra Forma- tion; abnormal growth of dorsal trail on the right side. E. SMNH Br129752; Burgsvik; Oja parish; Hamra Formation; posterior view of small specimen showing the large pseudodeltidium before it is consumed by the growth of the fo- ramen. F-G. Posterolateral views of two small specimens showing the effect on shell growth by encrustation; Burgs- vik, (3ja parish; Hamra Formation. F: SMNH Br129748; without bryozoan encrustation, the trail is low. G: With bryozoan encrustation, the trail has grown tall in an ultimately unsuccessful attempt to escape being overgrown. - H: ?Lepidoleptaena sp. A. SMNH Br32357; Sandarve Kulle, Fardhem parish; Hemse Group; small specimen with few, irregular and deep rugae. - I-M: ?Lepidoleptaena sp. B. Stora KarlsS; Slite Group. I-K. SMNH Br32196; Conjoined valves. I: Posterior view. J: Ventral view. K: Dorsal view. I_-M. SMNH Br32197; ventral valve with parts of the cardinal processes adhering to the inside of the muscle area. L: Internal view. M: External view. Scale bar = 15 mm.

2 8 0 OLE A. NOEL
specimens are relatively small, often with wide alae and few rugae. They occur in red, fine-grained limestone, whereas specimens ofL. poulseni occur mostly in marl- stones and bioclastic limestones.
?Lepidoleptaena sp. B Figs. 9I-M
Specimens from the Slite Group on Stora Karls6 have a sculpture typical of Lepidoleptaena (Figs. 9J, K, M), and a large pseudodeltidium (Fig. 9I), although the ven- tral interiors do not have the high rim around the disc (Fig. 9L). If these specimens belong to Lepidoleptaena, their Sheinwoodian age makes them the earliest known representatives of that genus.
G e o l o g i c a l s e t t i n g a n d d i s t r i b u t i o n of l e p t a e n i d s in G o t l a n d
The rocks exposed in Gotland range in age from latest Llandoverian to latest Ludlowian. The whole succession dips slightly (1-2 °) to the SE, so the youngest beds are exposed in the southeast. The facies comprise mostly marly limestones and marlstones, with more marly facies to the souhwest. At certain levels (H6gklint, Klin- teberg and Hamra-Sundre formations), extensive reefs are developed, especially in the northeast region of each unit. Towards the northeast and the East Baltic states, the two upper reef sequences continue as patch reef complexes. These have been mapped by the seismic studies of FLODt~N et al. (2001) and BJERKI~US c~: ERIKS-
SON (2001). The succession in Gotland shows at least three shallowing upward sequences (Visby to top of H6gklint; Tofta to top of Slite; Halla to Burgsvik). The geology is reviewed by HEDE (1921), MUNTHE et al. (1925), MANTEN ( 1971), LAUFELD ( 1 9 7 4 a ) , LAUFELD & BASSETT (1981) and SAMTLEBEN et al. (1996).
The ecological distribution of the leptaenids in Gotland is illustrated in Fig. 9 and their stratigraphic ranges are illustrated in Fig. 10. They are mostly con- fined to the marlstones. L. depressa visbyensis n. subsp. occurs almost exclusively in the Visby marlstones, which crop out along the NW coast; it is common at all localities of these beds. Some specimens are found in the H6gklint and Tofta formations in the westernmost part of the island. Here, it probably gives rise to L. de- pressa lata n. subsp., with its distinctive wide and flat alae. This subspecies occurs only in the extreme SW margin of the Slite Group, in the deepest water area, and the morphology was probably an adaptation for an am- bitopic lifestyle ("floating" in soft substrates). L. rhom- boidalis occurs in the H6gklint Formation, especially in the lowermost parts of each reef body, and in the high- energy Slite Group of Fgtr6, which is probably of similar age to the H6gklint Formation to the SW. Specimens probably belonging to this species are also found in a few reef localities in the uppermost part of the Slite Group. L. sperion has a similar temporal distribution to L. rhomboidalis and occurs in marlstones and in bedded inter-reef facies. L. parvirugata has an almost smooth shell and is found in marly facies of the Slite Group. In this unit, it has a distribution similar to that ofL. depres-
30
25
2O
"o 15 o
5 ¸
• +
+
. % •
• L. depressa depressa
-- L. rhomboidalis
• L. sperion
+ Lepidoleptaena o L. depressa lata
L. depressa visbyensis
+ •
L. rhombo tdahs ^ , * • _ • o ,"
\ / = = I ° K 12"i o , l ,
• xl
L. depressa v isbyens is
L. depressa depressa
5 10 15 20 25
Fig. 10. Scatter diagram plotting the length of the disc against the number of rugae on the disc in all lep- taenids in Gotland (with the exception for the non-rugate L. parvirugata). Tight group- ings are seen in L. rhomboi- da/is, L. depressa depressa and L. depressa visbyensis. The other taxa display much more variation.
n u m b e r o f rugae on disc

Silurian Leptaeninae (Brachiopoda) from Gotland, Sweden 281
sa depressa, except that it occurs further to the NW than L. depressa depressa, indicating a slight preference for higher water energy. It disappears after the sea level drops to expose the platform in the Mulde Event (CAL- NER & JEPPSSON 2003). L. depressa depressa occurs in marlstones from the upper part of the Tofta Formation to
the upper part of the Hemse Group. Its disappearance coincides with another lowstand non-depositional se- quence boundary with some evidence of sub-aerial ex- posure (CHERNS 1983). The transgression following this event was not sufficiently large to submerge the plat- form as deeply as before, and the platform thus re-
70
60
50
40
30
20
t0
0
A Leptaena rhomboidalis
• 4 b ~ l l ~ "
0 5 10 15 20 25 30 35
50,
40.
30.
20.
1 0
ol
Leptaena sperion
?;:., t4l,
° 4 e.
5 10 15 20 25 30 35
70
60
50
40
30
20
10
0
C Leptaena parvirugata
0 5 10 15 20 25 30 35
7 0
6 0
5 0
4 0
3 0
2 0
10"
0
D Leptaena depressa depressa
: ~ p ; q t ' : . - - .
5 10 15 20 25 30 35
70
60
50
40
30
20
10
0
E Leptaena depressa visbyensis
o ~ . ' t . "t:'"
0 5 10 15 20 25 30 35
F Leptaena depressa lata 5O
4 0 -
3 0
2 0
1 0
0
-
#t%
5 10 15 20 25 30 35
70
60
50
40
30
20
10
0
G Lepidoleptaenapoulseni
:: % * *
0 5 10 15 20 25 30 35
IT v
Max length (ram)
Fig. 11. Scatter diagrams plotting maximum length against maximum width in each leptaenid taxa occur- ring in Gotland. Two size groups are evident in L. de- pressa depressa and Lepidoleptaena pou/seni.

282 OLE A. HOEL
X / '
/
,, ',
~xv
b D
v t
~ Q
o
Q
~J
• Leptaena rhomboidal&
[] Leptaena sperion
Leptaena depressa visbyensis
0 Leptaena depressa lata
A Leptaena depressa depressa
• Leptaenaparvirugata
Lepidoleptaena sp. A and t3
Lepidoleptaena poulseni
0 5 lOkm I J r ~ p I [
0
Facies types D Limestone
D Calcareous sandstone (Slite Formation)
Marls
D Siliceous sandstone (Burgsvik Formation)
Reef areas
Fig. 12. Map of Gotland with the main facies types and the distribution of the leptaenid species.

Silurian Leptaeninae (Brachiopoda) from Gotland, Sweden 283
mained near the surface for the rest of the Ludlowian. This high-energy environment evidently was not suita- ble for Leptaena, and Lepidoleptaena poulseni is the only leptaenid in these beds. It was apparently well adapted to the high water energy by having a large pedi- cle and thick shell.
Acknowledgements
I e x t e n d m y t h a n k s to MIKE BASSETT, N a t i o n a l M u s e u m o f
W a l e s , a n d LARS HOLMER, U p p s a l a U n i v e r s i t y , for g iv ing m e
a lot o f i n f o r m a t i o n a n d f i e ld e x p e r i e n c e in G o t l a nd , and fo r
grea t ly i m p r o v i n g th is m a n u s c r i p t . JAN OVE EBBESTAD, U p p -
sa la U n i v e r s i t y , h e l p e d in p r e p a r i n g the p h o t o g r a p h i c f igu res .
CHRISTINA FRANZI~N BENGTSSON ( S M N H ) a n d LARS KARIS
( S G U ) m a d e the l a rge c o l l e c t i o n s ava i l ab le for s tudy . I a l so
t h a n k CARSTEN BRAUCKMANN, T e c h n i s c h e Universit~it
C l a u s t h a l , a n d t w o a n o n y m o u s r e v i e w e r s for cri t ical c o m -
m e n t s o n the m a n u s c r i p t .
References
AMSDEN, T.W. 1951. Brachiopods of the Henryhouse Formation (Silur- ian) of Oklahoma. - Journal of Paleontology 25: 69-96.
AMSDEN, T.W. 1958. Stratigraphy and Paleontology of the Hunton Group in the Arbuckle Mountain Region. - Oklahoma Geological Survey Bulletin 78: 1-199.
AMSDEN, T.W. 1978. Articulate Brachiopods of the Quarry Mountain Formation (Silurian), Eastern Oklahoma. - Oklahoma Geological Survey Bulletin 125: 1-75.
BAARLI, B.G. 1995. Orthacean and strophomenid brachiopods from the Lower Silurian of the central Oslo Region. - Fossils and Strata 39: 1-93.
BARRANDE, J. 1879. Systrme Silurien du centre de la Bohrme. 16re par- tie. Recherches palrontologiques, 5. Classe des Mollusques. Or- dre des Brachiopodes. - 226 p., 153 pls., Prague & Paris.
BASSETT, M.G. 1974. The articulate Brachiopods from the Wenlock Se- ries of the Welsh Borderland and South Wales, 3. - Palaeonto- graphical Society Monographs 128 (541): 79-122, pls. 18-32.
BASSETT, M.G. 1977. The articulate Brachiopods from the Wenlock Se- ries of the Welsh Borderland and South Wales, 4. - Palaeonto- graphical Society Monographs 130 (547): 123-176, pls. 33-47.
BASSETT, M.G. 1979. Brachiopods. - In: JAANUSSON, V.; LAUFELD, S. ~; SKOGLUND, R., eds., Lower Wenlock faunal and floral dynam- ics - Vattenfallet section, Gotland. - Sveriges Geologiska Under- srkning C 762: 185-206.
BASSETT, M.G. & COCKS, L.R.M. 1974. A review of Silurian brachio- pods from Gotland. - Fossils and Strata 3: 1-56.
BERGSTROM, J. 1968. Upper Ordovician Brachiopods from V~isterg/3t- land, Sweden. - Geologica et Pal~eontologica 2: 1-35.
BJERKI~US, M. & ERIKSSON, M. 2001. Late Silurian reef development in the Baltic Sea. - Geologiska Frreningens I Stockholm Frr- handlingar 123:169 179.
BOUCOT, A J . 1973. Early Paleozoic Brachiopods of the Moose River Synclinorium. - United States Geological Survey, Professional Paper 784: 1-81.
BoucoT, A.J.; JOHNSON, J.G.; HARPER, C. ~; WALMSLEY, V.G. 1966. Silurian brachiopods and gastropods of southern New Bruns- wick. - Geological Survey of Canada, Bulletin 140: 1-45.
CALNER, M. & JEPPSSON, L. 2003. Carbonate platform evolution and conodont stratigraphy during the middle Silurian Mulde Event, Gotland, Sweden. - Geological Magazine 140 (2): 173-203.
CHERNS, L. 1983. The Hemse-Eke Boundary. Facies relationships in the Ludlow Series of Gotland, Sweden. - Sveriges Geologiska Undersrkning C 800: 1 4 4 .
CHERNS, L. & BASSET]', M.G. in press. A framework sequence strati graphy for the Silurian of Gotland. - Journal of the Geological Society of London.
COCKS, L.R.M. 1967. Llandovery stropheodontids from the Welsh Bor derland. - Palaeontology 10 (2): 245-265.
COCKS, L.R.M. 1968. Some strophomenacean brachiopods from the British Lower Silurian. - Bulletin of the British Museum (Natural History), (Geology) 19 (4): 142-203.
COCKS, L.R.M. 1978. A review of British Lower Palaeozoic brachio- pods, including a synoptic revision of Davidson 's monograph. - Palaeontographical Society Monographs 131 (549): 1-256.
COCKS, L.R.M. & RONG, JIA-YU 2000. Strophomenida. - In: WILLIAMS, A.; BRUNTON, C.H.C. & CARLSON, S.J., eds., Treatise on Inverte brate Paleontology. Part H. Brachiopoda (revised), 2-3 :216-348, Lawrence, Kans., Boulder, Colo. (University of Kansas Press).
DALMAN, J.W. 1828. Uppst~illning och Beskrifning af de i Sverige fun ne Terebratuliter. - Kungliga Svenska Vetenskapsakademiens Handlingar [fOr 1827]: 85-155.
DAVIDSON, T. 1866-1871. A monograph of the British fossil Brachiop- oda, Vol. 3 (7). Silurian: No. 1. Palaeontographical Society Monograph 19 (84): 1-88, pls. 1 - t 2 (1866); No. 2. Palaeonto graphical Society Monograph 20 (87): 89-168, pls. 13-22 (1867); No. 3. Palaeontographical Society Monographs 22 (96): 169-248, pls. 23-37 (1869); No. 4. Palaeontographical Society Monograph 24 (108): 249-397, pls. 38-50 (1871).
FLODI~N, T., BJERKt~US, M.; TUULING, I. & ERIKSSON, M. 2001. A Sil uriah reefal succession in the Gotland area, Baltic Sea. - Geolo- giska F~ireningens I Stockholm Frrhandlingar 123: 137-152.
HALL, J. & CLARKE, J.M. 1894. An introduction to the study of the gen era of Palaeozoic Brachiopoda. Natural History of New York. Palaeontology, 8 (2). - xvi + 394 p., pls. 21-84, Albany (New York Geological Survey. Charles van Benthuysen & Sons).
HARPER, C.W. 1973. Brachiopods of the Arisaig Group (Silurian - low er Devonian) of Nova Scotia. - Geological Survey of Canada. Bulletin 215: 1-163.
HAVLI~EK, V. 1963. eled Leptaenidae (Brachiopoda) v eskfm siluru a devonu. - asopis Narodni Museum 132 (4): 220-225.
HAVLICEK, V. 1967. Brachiopoda of the suborder Strophomenidina in Czechoslovakia. - Rozpravy l)strednl'ho I)stavu Geologickrho 33: 1-235.
HAVL~CEK, V. & STORCH, P. 1967. Silurian brachiopods and benthic communit ies in the Prague Basin (Czechoslovakia). - Rozpravy l)stredniho lQstavu Geologickrho 48: 1-275.
HEDE, J.E. 1921. Gottlands Silurstratigrafi.- Sveriges Geologiska Un- dersrkning C 305: 1-100.
HISINGER, W. 1837-1841. Lethaea suecica seu Petrificata Suecica. iconibus et characteribus illustrata. - 1. (1837): 124 pp., pls. 1- 36; 2. (1840): 11 pp., pls. 37-39; 3. (1841): 6 pp., pls. 40-42, Stockholm.
HOLTEDAHL, O. 1916. The Strophomenidae of the Kristiania Region. - Det Norske Vitenskaps-Akademi i Oslo. I. Skrifter. Matematisk Naturvitenskapelig Klasse 1915 (12): 1-117.
JIN JISOU & CHATTERTON, B.D.E. 1997. Latest Ordovician - Silurian articulate brachiopods and biostratigraphy of the Avalanche Lake area, southwestern District of Mackenzie, Canada. - Palaeonto graphica Canadiana 13: 1-169.
JOHNSON, J.G.; BOUCOT, A.J. & MURPHY, M. 1973. Pidolian and early Gedinnian age brachiopods from the Roberts Mountains Forma tion of Central Nevada. - University of California Publications in Geological Sciences 100: 1-75.
KING, W. 1846. Remarks on certain genera belonging to the class Pal- liobranchiata. - Annals and Magazine of Natural History (1) 18: 2 6 4 2 , 83-94.
KELLY, F.B. 1967. Silurian Leptaenids (Brachiopoda). - Palaeontology 10 (4): 590~602.
LAMONT, A. 1947. Gala-Tarannon Beds in the Pentland Hills, near Ed inburgh. - Geological Magazine 84: 193-208,289-303.
LAUFELD, S. 1974. Silurian Chitinozoa from Gotland. - Fossils and Strata 5: 1-130.

284 OLE A. HOEL
LAUFELD, S. & BASSETT, M.G. 1981. Gotland: The anatomy of a Silu- rian carbonate platform. - Episodes 1981 (2): 1-5.
LENZ, A.C. 1977. Upper Silurian and Lower Devonian brachiopods of Royal Creek, Yukon, Canada, 1. Orthida, Strophomenida, Pen- tamerida, Rhynchonellida. - Palaeontographica (A) 159: 37-109.
M'CoY, F. 1844. A synopsis of the characters of the Carboniferous Limestone fossils of Ireland. - 207 p., 29 pls., London (Williams & Norgate).
M'CoY, F. 1846. A synopsis of the Silurian fossils of Ireland collected from the several districts by Richard Griffith, F.G,S. - 72 pp., 5 pls., Dublin.
MANTEN, A.A. 1971. Silurian reefs of Gotland. - Developments in Sed- imentology 13: 1-539, Amsterdam (Elsevier).
MUNTHE, H.; HEDE, J.E. & von POST, L. 1925. Gottlands Geologi. En 6versikt. - Sveriges Geologiska Unders6kning C 331: 1- 130.
OPIK, A.A. 1934. Uber Klitamboniten. - Universitatis Tartuensis (Dor- patensis) Acta et Commentationes (A) 26 (5): 1-239.
POULSEN, C. 1943. The Silurian faunas of north Greenland. 2. The fau- na of the Offley Island Formation. 2. Brachiopoda. - Meddelelser om GrCnland 72 (3): 1-60.
ROOMUSOKS, A.K. 1989. f0ber die Divergenz der Leptaenidae (Brachi- opoda) in der Viru- und Harju-Zeit in Baltoskandia. - Proceed- ings of the Academy of Sciences in the Estonian SSR, Geology 38:112-117.
SAMTLEBEN, C.Z MUNNECKE, A.; BICKERT, T. & PATZOLD, J. 1996. The Silurian of Gotland (Sweden): Facies interpretation based on sta- ble isotopes in brachiopod shells. - Geologische Rundschau 85: 278-292.
SCHUCHERT, C. 1893. Classification of the Brachiopoda. - American Geologist 11: 141-167.
SOKOLSKAYA, A.N. 1954. Strophomenidij Russkogo Platformi [Stro- phomenids of the Russian Platform]. - Akademija Nauk SSSR. Trudy Paleontologijeskogo lnstituta 51: 1-191.
SOWERBY, J. 1812-22. The Mineral Conchology of Great Britain. 1. 234 p., pls. 1-102 (1812-1815); 2. 251 p., pls. 103-203 (1815- 1818); 3. 194 p., pls. 204-306 (1818-22), London.
SOWERBY, J. ~; SOWERBY, J. de C. 1821-1823. The Mineral Conchol- ogy of Great Britain. 4. 160 p., pls. 307-407 (1821-23), London.
SOWERBY, J. de C. 1823-1846. The Mineral Conchology of Great Brit- ain. 5. 171 p., pls. 408-503 (1823-25); 6. 235 p., pls. 504-609 (1826-29); 7 .80 p., pls. 610-648 (1840-46), London.
SPJELDN/ES, N. 1957. The Middle Ordovician of the Oslo Region, Nor- way. 8. Brachiopods of the Suborder Strophomenida. - Norsk Geologisk Tidsskrift 37: 1-214.
SPJELDN/ES, N. 1984. Epifauna as a tool in autecological analysis of Silurian brachiopods. - Special Papers in Palaeontology 32: 225- 235.
VILLAS, E. 8~; COCKS, L.R.M. 1996. The first Early Silurian brachiopod fauna from the Iberian Peninsula. - J o u m a l of Paleontology 70 (4): 571-588.
WAHLENBERG, G. 1818. Petrificata Telluris Svecanae. - Acta Societatis Regiae Scientiarium Uppsaliensis 8 :1 -116 .
WORSLEY, D.; AARHUS, N.; BASSETT, M.G.; HOWE, M.P.A.; MORK, A. 8,: OLAUSSEN, S. 1983. The Silurian succession of the Oslo Re- gion. - Norges Geologiske Undersckelse Bulletin 72: 1-57.
ZHANG, N. 1989. Wenlockian (Silurian) brachiopods of the Cape Phil- lips Formation, Baillie Hamilton Island, Arctic Canada, 2. - Pa- laeontographica (A) 206: 99-135.
Manuskripteingang/manuscript received 23 .9 . 2003; Manuskriptannahme/manuscript accepted 19.7. 2004.





























