Embed Size (px)
Citation preview

Historical Biology, 1996, Vol 12, pp 93-110 © 1996 OPA (Overseas Publishers Association)Reprints available directly from the publisher Amsterdam B V Published in The NetherlandsPhotocopying available by license only by Harwood Academic Publishers Gmb H
Printed in Malaysia
DIVERSITY AND EXTINCTION PATTERNSOF PERMIAN BRACHIOPODA
OF SOUTH CHINA
SHUZHONG SHEN' and G R SHI 2
'Department of Geology, China University of Mining and Technology,Xuzhou, Jiangsu Province 221008, P R China
2School of Aquatic Science and Natural Resources Management,Deakin University, Rusden Campus, 662 Blackburn Road, Clayton,
Victoria 3168, Australia
(Received January 16, 1996 ; in final form March 20, 1996)
The stratigraphical and geographical distribution of 851 brachiopod species from 216 genera and65 families in the Permian of South China are analysed It is revealed that the brachiopod diversityunderwent two sharp falls during the Permian The first occurred at the end of Maokouan,accompaning the widely recognised, extensive regression across the Maokouan-Wujiapingianboundary Fifty-seven species of 29 genera survived this first major extinction event The secondsharp reduction of brachiopod diversity took place in the later Changhsingian, with only 17 Permian-type brachiopod species of 12 genera straggling into the earliest Triassic Detailed stratigraphicanalysis shows that more than 90 % of the Changhsingian brachiopod species disappeared at differentlevels in the Changhsingian before the widely perceived end-Permian 'mass extinction' occurred.It is also notable that each of the step-wise diversity reduction events was apparently heterochronous.In view of the evidencefrom lithologies, faunal components and geochemical analyses, the twosharp falls of Permian brachiopod diversity in South China are considered to be closely relatedto multiple interactions of an environmental deterioration caused by large-scale regressions.
KEY WORDS: Permian, Brachiopoda, Diversity Patterns, Extinction.
INTRODUCTION
There seems to be a general concensus that the greatest mass extinction event in theearth's history occurred at the Permian-Triassic boundary (Erwin, 1993) Sepkoski ( 1982,1992) showed that 52 %'of marine families and 90 % of marine species became extinctby the end of the Permian Numerous theories have been proposed to explain this Permian-Triassic crisis, ranging from hypotheses of catastrophic events, extra-terrestrial impactsto one or more volcanic eruptions Other scenarios are also available, involving formationof Pangea, global cooling, salinity, and marine anoxia (Kanmera et al , 1976 ; Sun andChai 1984 ; Xu et al , 1987 ; Yang and Yin et al , 1987 ; Yang and Wu et al , 1991 ;Chai et al , 1986, 1992 ; Li et al , 1989 ; He and Shen, 1990 ; Shen et al , 1992 ; Wignalland Hallam, 1993 ; Erwin, 1993, 1994, 1995 ; Stanley and Yang, 1994 ; among manyothers).
South China is perhaps one of the most important regions for studying the Permiandiversity and evolutionary patterns of marine biota where continuous Permian-Triassicsections exist and have been studied intensively over the last two decades Brachiopodsin the Permian sequence of South China are among the most abundant shelly fossils,providing sufficient quantitative data to study the diversity and extinction patterns (Liao,
93

S SHEN AND G R SHI
1980 ; Shen and He, 1991 a-b, 1994 a; Xu and Grant, 1994) However, detailed analysison the stage-by-stage stratigraphical distributions of the Permian brachiopod faunas atspecific, generic and familial levels is lacking, with most previous studies concentratedon the boundary biota itself The present paper is aimed to draw together all publisheddata of Permian Brachiopoda of South China to investigate their changing patterns ofdiversity and extinction through the Permian in an attempt to examine the causes ofthe Permian extinctions.
The data used for this analysis is in part taken from the data base compiled by Shiand Archbold ( 1993 a, b; 1994 a,b), Shen et al ( 1996) and Shi et al ( 1996), supplementedby personal observations and field experience and various unpublished lists compiledby the authors and Professor Jin Yu-gan This data pool represents the most completeinformation available on Permian brachiopod faunas of South China.
The supera-generic classification of Brachiopoda referred to in this study largelyfollows the Treatise on Invertebrate Paleontology (Part H) Brachiopoda (Moore, 1965)but we have incorporated more recent modifications wherever appropriate The newversion of the Treatise that is being revised may have some significant differences inclassification from that of the 1965 version; however, whether or not these potentialbroad-scaled changes in classification of Brachiopoda will alter significantly theconclusions reached here is yet to be seen.
The Permian chronostratigraphic scale used for discussion in this paper follows thatof Jin et al ( 1994), with the Permian System of South China being divided into threeSeries: Chuanshanian, Yanghsingian and Lopingian, in ascending order It must be noted,however, that at the moment no universily accepted Permian time scale is available andcomparable conclusions would have been reached had an alternative Permian time scalebeen used.
FAUNAL CHARACTERISTICS AND CHANGING PATTERNS OF DIVERSITY
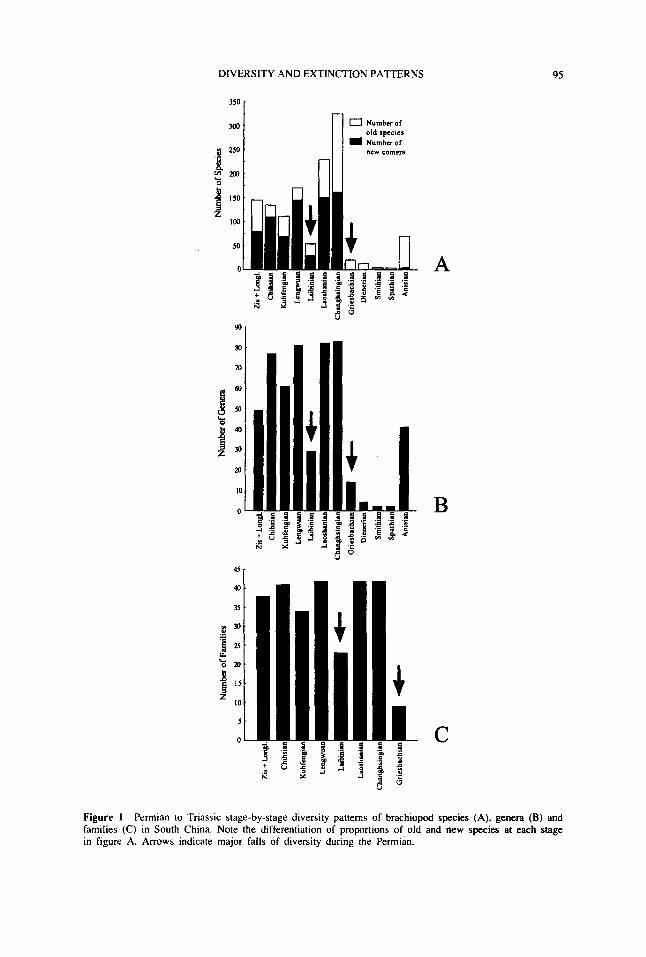
Our data shows that a total of 851 species representing 216 genera and 65 families havebeen recorded (most have been described) from the Permian of South China Simpletabulation of the number of families, genera and species against Permian stages/substagesreveals two sharp falls during the Permian (Figure 1) The first occurred in the lateMaokouan (i e , Lengwuan Stage) of the Middle Permian, followed by a second fallin the late Changhsingian It is of note that most of the Permian brachiopod specieswere extinguished before the end of Changhsingian, with only 17 species of 12 generasurviving into the earliest Triassic (Figure B).
Zisongian and Longlinian Faunas (Early Permian)
The brachiopods between the Late Carboniferous and the Early Permian Zisongian andLonglinian stages have no significant difference in composition and diversity (Liao, 1979 ;Li et al , 1987) Productida is the most prevalent group during the Zisongian andLonglinian In total, 56 productid species have been recorded from the two Early Permianstages, constituting about 34 % of the total species recorded for the Early Permian ofSouth China (Figure 2).
The productids are dominated by the families of Overtoniidae, Echinoconchidae,Buxtoniidae, Marginiferidae, Dictyoclostidae and Linoproductidae, all persisting fromthe Late Carboniferous The most important genera include Krotovia, Echinoconchus,Buxtonia, Linoproductus, Cancrinella and Dictyoclostus, all of which are generally largein size and have fine spines Among the Rugosochonetidae, Neochonetes carbonifera
94
DIVERSITY AND EXTINCTION PATTERNS
I 9
I
A
z I.9
B
aEt~
E
C
Figure I Permian to Triassic stage-by-stage diversity patterns of brachiopod species (A), genera (B) andfamilies (C) in South China Note the differentiation of proportions of old and new species at each stagein figure A Arrows indicate major falls of diversity during the Permian.
95
s.n
..

S SHEN AND G R SHI
stage
Taxa
Inarticulata
Orthida
Strophomenida
Oldhamidina
Rhynchonellida
rerebratulida
Athyridida
Spiriferida
Chonetida
L Permian
Zisongian
mm
3-
_
_I
Longlinian
M Permian
Chihsian
TM
M^
-_ 2
Maokouan
Kuhf
p 2
M" 1
Hi~
^ 3 w
EaJ 7
n
pa
U PermnianWujiapingian
L.bin
I-
_,
-2 _
Lah
UlaEA
H 3
Changhs.
-__
Il
IBUPU __~_ 3
Triassic
Griesbach
-_
I I I O _II Im I" -I
_M
StrophalosiidaeEchinoconchidae|ulostegidae
rschernyschewidae
|honetellidae
DictyoclostidaeWaagenoconchidaiLinoproductidae
|Marginiferidae
RetariidaeOvertoniidaef HorridoniidaeTeguliferinidaeIpiridiophoridaePaucispiniferidaeMonticuliferidae
Plicatiferidae
|honostegidaeIncisiidaeLamiproductidaetichthofeniidaeZooperidaetratiferidae
BuxtoniidaeAnidanthidaeF Hercosiidaelsogrammidac
_ _
-
-
2-
i_
a:I-
L-
_
-o_
-
I-
mizEu
=l M=M 1
I-
I
I-
I-
-p 2.
2-
I-
_ m
-2-
-2-
we
_ 2 ·
__
mm
I
-21-M 4 M
m·on
I-
n-
2
-I-
-I-
I-
IIIII
_ 2
l
I_
= 3-2-
Figure 2 Changing patterns of species diversity of principal brachiopod higher taxa during the Permian ofSouth China (Note the width of each stage corresponds with its radiometric time duration).
Productida
Ls P
�M
-I
I
l.= al _ II
_ ,msnl
4 mg . _
13 V im
^ l 1 ·-
. I s1 - .
96
I
'- W~= MI 3
_ _
_ _ f I __ _ _
I

DIVERSITY AND EXTINCTION PATTERNS
(Keyserling), N latisinuata Zhan, Chonetes uralicus Moeller, Costachonetes pygmaea(Loczy) are the most abundant species, most of which persisted from the LateCarboniferous Spiriferids are also common and constitute the second largest group forthe Zisongian and Longlinian (Figure 2) Among the spiriferids, Choristites and Martiniaare the most diversified, each represented by 6 species and usually occurring in greatabundance Other characteristic elements of the spiriferid group include Eliva, Neospiriferand Spiriferella In addition, Enteletes of the Enteletidae, Meekella of the Meekellidaeand Terebratuloidea and Stenoscisma of the rhynchonellids are also commonly found.However, lyttoniids are relatively rare, represented by five species each known only froma few specimens Athyridids are seldom found; so far they are known only from twospecies of Cleiothyridina and one species of Composita.
The Chihsian Fauna (Early Middle Permian)
The brachiopods of Chihsian age are slightly less diverse than those in the Early Permian,but there is little change in composition; the predominant group being still the Productida(Figure 2) Dictyoclostidae, Marginiferidae and Linoproductidae families continued toflourish during the Chihsian However, the Chonetellidae had become the most diversegroup among the productids (Figure 2) An examination of all known records revealsthat Haydenella, Ogbinia, Tyloplecta and Linoproductus are the most prevalent generaamong the productids The presence of Haydenella and Tyloplecta is of particular notebecause they typify the Asian Tethyan Region (term defined by Shi et al , 1995) Itis also worth mentioning that Monticulifera and some elements of the Plicatiferidae suchas Neoplicatifera and Paraplicatifera that appeared in the late Chihsian show closemorphological resemblance to those of the Maokouan Subseries.
Athyridids are thesecond largest group for the Chisian (Figure 2), occurring widelyin great abundance and characterised by the radiation of genera Spirigerella and Athyris.On the other hand, spiriferids experienced an evident decline in diversity, with speciesdropping down to 15 compared to 33 in the Early Permian (Figure 2) Choristites andMartinia, the most common genera in the Early Permian, became less diverse in theChihsian and their role as characteristic Chihsian taxa were replaced by Phricodothyris,Squamularia and Neospirifer A slight increase in species diversity from the EarlyPermian is noticable of the strophomenid group, with Meekella and Orthothetina beingmost abundant, seconded by Orthotichia and Acosarina Orthotichia chekiangensis Chaohas long been regarded as one of the most characteristic species of the Chihsian.
Maokouan Faunas (Late Middle Permian)
In this paper, the Maokouan is used as a Subseries rank name, incorporating two stages:Kuhfengian and Lengwuan, following Jin et al ( 1994) By the Kuhfengian Stage, thereexisted 121 species of some 61 genera, in comparison with 146 species of the EarlyPermian (Figure IA) The Kuhfengian brachiopod faunas are characterised by theflourishing of the families Rugosochonetidae, Chonetellidae, Monticuliferidae,Dictyoclostidae, Lochengidae, Chonestegidae and Plicatiferidae Typical genera includeMonticulifera, Uncisteges, Cryptospirifer, Neoplicatifera, Tyloplecta, Spinomarginifera,Urushtenoidea, and Vediproductus The species Urushtenoidea chaoi Ching, U chenanensisChan, and Monticulifera sulcata Yang are restricted to the Kuhfengian Among theSchizophoriidae, Acosarina indica (Waagen) and Orthotichia chekiangensis Chaopersisted from the Chihsian but were significantly less abundant Squamularia, whichflourished in the Late Permian, began to become common during the Kuhfengian.
97

S SHEN AND G R SHI
The Lenguan Stage was first proposed by Liang ( 1990) on the basis of a large, diversebrachiopod fauna from the Lengwu Formation in Zhejiang Province In southwest China,the Lengwuan Stage is probably represented by the Daozhuanyan Formation of Huangand Zheng ( 1948).
Liang ( 1990) described some 235 brachiopod species of 106 genera from the LengwuFormation, including 153 new species and 30 new genera Unfortunately, the numberof family, genera and species was actually overstated due to the lack of comparisonbetween Liang's 'new' taxa and those previously reported In light of modern taxonomyand detailed revision, we would reduce the Lengwuan fauna to 169 species, representing82 genera and 44 families However, it is clear that the Lengwuan fauna, characterisedby numerous new and endemic taxa, represents a significant change from the Kuhfengian,as well as exhibiting some evidence of faunal transition to the Late Permian Wujiapingian.Productids are still dominant (Figure 2), followed by spiriferids, strophomenids, orthids,terebratulids, and rhynchonellids The Lengwuan Stage saw the last appearance of severalcharacteristic Middle Permian genera including Monticulifera, Neoplicatifera,Vediproductus, Kiangsiella, Uncisteges and also the introduction of a number ofcharacteristic Late Permian taxa such as Peltichia, Edriosteges, Tyloplecta yangtezensis(Chao), Gubleria, and Strophalosiina Another important feature of the Lengwuanbrachiopod fauna is the first abundant concentration of lyttoniids such as Leptodus,Oldhamina and permianellids, strongly suggesting a close relationship with the LatePermian faunas Other important families of the Lengwuan Stage include Meekellidae( 13 species), Spiriferidae ( 8 species), Dielasmatidae ( 8 species) and Notothyridae ( 8species) The stratigraphical distribution of brachiopods within the Lengwuan Stage showsa general decrease upwards in diversity and abundance.
Wujiapingian Fauna (Early Late Permian)
At the end of the Maokouan, South China witnessed an almost complete withdrawalof the sea (the widely known Dongwu Movement), followed by transgression anddeposition of a complex suite of lithofacies now incorporated in the Wujiapingian Stagewhich rests on a major unconformity or sequence boundary Recent palaeontologicalfindings indicate that the Wujiapingian can be readily separated into two parts; the lowerpart, recently named the Laibian by Jin, Zhu and Mei ( 1994), is only found in somenarrow-trending basins or belts in South China, and the upper part, more widelydistributed, is typified by coal-bearing lithofacies.
The Maokouan-Wujiapingian boundary also marks an important discontinuity in thebrachiopod succession and diversity, with many characteristic Early and Middle Permiangenera terminated at the boundary, including Echinoconchus, Linoproductus andCancrinella (which all had a Late Carboniferous history) and Monticulifera, Urushtenoidea,Cryptospirifer and Neoplicatifera of primarily Middle Permian age (Jin, 1993) TheLaibinian brachiopod fauna has yet to be described, but according to unpublished reportsand personal observations, some 57 species of 29 genera may be found in the Laibinianfauna, most of which are typically of Late Permian character Common species of theLaibinian may include Orthothetina rumer (Frech), Acosarina indica (Waagen),Spinomarginifera spp , Tyloplecta yangtzeensis (Chao), Araxathyris araxensis Grunt,Haydenella kiangsiensis (Kayser), Cathaysia chonetoides (Chao) Comparison of thislist with characteristic species of Early and Middle Permian brachiopods previouslymentioned indicates a distinct difference of the Early Wujiapingian brachiopods fromthe Early and Middle Permian faunas.
The continuing transgression in southwest China in the Late Wujiapingian, orLaoshanian as recently named by Jin, Zhu and Mei ( 1994), deposited a coal-bearing
98

DIVERSITY AND EXTINCTION PATTERNS
unit containing and/or interbeded with a number of marine beds Brachiopods areexceedingly abundant and diverse in these marine beds, demonstrating a significantincrease of species, genus and family diversity after the sharp decline at the latestMaokouan (Figure 1) The Laoshanian brachiopod fauna contains 233 species assignedto 82 genera, of which 151 species are new (Figure 2) The species diversity representsa 37 % increase over the Maokouan Stage However, of the 82 genera recognised fromthe Laoshanian, most persisted from the pre-Wujiapingian faunas, with only 17 firstappearing in the Laoshanian Productida attained the highest diversity, with 72 species(Figure 2), of which Chonetellidae, Marginiferidae and Lyttoniidae are most prevalent,typified by Transennatia, Spinomarginifera, Haydenella, Oldhamina, Leptodus, Cathaysia,Edriosteges and Tyloplecta In contrast, Echinoconchidae, Linoproductidae, Buxtoniidae,Dictyoclostidae experienced a rapid reduction in species diversity (Figure 2) Spiriferidaconsititutes the second dominant order of the Late Wujiapingian brachiopod fauna, with36 species of 9 genera recorded, among which Squamularia and Martinia are the twomost diverse genera A significant increase in species diversity is also observed for thechonetids (Figure 2), characterised by Waagenites and Neochonetes This is in contrastwith Pygmochonetes, Tenuichonetes and Songzichonetes which are the dominantchonetacean genera of the pre-Wujiapingian faunas It is of note that no genuineWaagenites has been reported in the Maokouan Stage in South China The explosiveincrease of Strophomenida is another character of the Late Wujiapingian brachiopodfauna, with Orthothetina, Meekella, Streptorhynchus being most abundant and widelyoccurring, followed by Enteletes, Acosarina, Araxathyris and Stenoscisma.
Changhsingian Fauna (Late Late Permian)
Brachiopods continued to increase in generic and specific diversity during this stageand attained acme development during the middle Changhsingian A total of 325 speciesassigned to 83 genera in some 42 families have been reported Taxa that occurred ingreat abundance in the Wujiapingian mostly continued to exist during the Changhsingian(Shen and He, 1991 a) Productida shows a slight reduction in species numbers althoughit is still the most diverse group (Figure 2) The dominant productid genera areSpinomarginifera, Haydenella, Oldhamina, Leptodus and Cathaysia.
On the other hand, Spiriferida, Strophomenida, Orthida and Rhynchonellida alldemonstrate a significant increase in species diversity and abundance (Figure 2) Themost characteristic genera of the Changhsingian Stage include Peltichia, Orthothetina,Meekella, Perigeyerella, Squamularia, Martinia, Prelissorhynchia and Stenoscisma,along with some small and thin-shelled elements such as Waagenities, Paryphella,Crurithyris and Araxathyris, which are all very common in the intra-platform basinalfacies.
Early Triassic Griesbachian Fauna
As shown in Figs 1 and 2, brachiopods suffered another sharp drop in diversity at theend of the Changhsiangian, with more than 93 % species and 85 % genera becoming extinctbefore the P/T boundary (He and Shen, 1990 ; Shen et al , 1992) On the whole, only17 species in 12 genera straggled into the earliest Triassic in what has been known asthe Permian-Triassic 'transitional beds' (Shen and He, 1991 a-b; Shen and He, 1994 a)(Figure 2) With the exception of some new species of Lingula, all recorded Griesbachianspecies are the Late Paleozoic relics, notably Waggenites, Araxathyris, Crurithyris andAcosarina, all with small size and thin shells, are most notable The next recovery ofbrachiopods began in the early Anisian of the Middle Triassic (Figure A and B).
99

S SHEN AND G R SHI
EXTINCTION PATTERNS
As discussed above, two sharp drops of brachiopod diversity and richness are apparent.The first extinction event occurred at or towards the end of the Maokouan of the MiddlePermian, with 87 % of the then existing species becoming extinct The Middle Permianof South China is characterised by the flourishing of Neoplicatifera, Uncisteges,Monticulifera, Cryptospirifer, Urushtenoidea in South China With the exception of twospecies of Neoplicatifera, these genera did not persist into the Late Permian (Figure3) Also of note is the timing of the disappearance of these genera, whose constituentspecies tend to become extinct in a stepwise fashion rather than just at the end of theMaokouan (Figure 3) For instance, the first occurrence of Cryptospirifer is stratigraphicallyslightly below the Misellina claudae Zone (Li et al , 1987); it attains the highest diversityin the Early Maokouan (Kuhfengian) and became extinct at the end of the EarlyMaokouan Monticuliferidae has a similar extinction pattern; it occurred in the Chihsian,flourished in the Early Maokouan and continued, with only two species (acounting for14 % of the total species of this family), into the Late Maokouan Plicatiferidae appearsto have a relatively longer stratigraphic range Neoplicatifera appeared in the Zisongian,became diverse during the Maokouan and straggled into the Early Wujiapingian withtwo species, Neoplicatifera huangi (Ustrisky) and N sintanensis (Chao) Chonostegidaeunderwent a very short history; it probably appeared in the latest Chihsian, flourishedin the Kuhfengian and became extinct just at the end of the Maokouan.
The observed changing patterns of diversity and extinction of the pre-WujiapingianBrachiopoda are comparable with similar patterns of other fossil groups The mostimportant groups in the Permian are the fusulinid families Schwagerinidae andVerbeekinidae; the former displayed the greatest generic and specific diversity in theEarly Permian and the latter in the Middle Permian The Schubertellidae underwent amarked expansion in the Middle Permian and persisted into Late Permian with only4 genera (Kanmera et al , 1976) According to Jin ( 1993), the succession of fusulinidsin the carbonate sequence of the Late Maokouan and Early Wujiapingian of GuizhouProvince also demonstrates a stepwise process of faunal turnover in both diversity andabundance towards the end of the Maokouan Subseries.
Similarly, Batten ( 1973), on the basis of an examination of all known Guadalupiangastropod faunas, revealed that gastropods were extremely abundant during the MiddlePermian, but underwent a significant drop across the Guadalupian-Dzhulfian boundary,with 62 genera and 12 families extingushed Batten ( 1973) also showed that more than32 Guadalupian genera had been found in the Ladinian of Middle Triassic withoutrepresentatives in the Late Permian and Early Triassic Most of these 'Lazarus' generaare conservative and composed of very few species, but each with a large number ofindividuals.
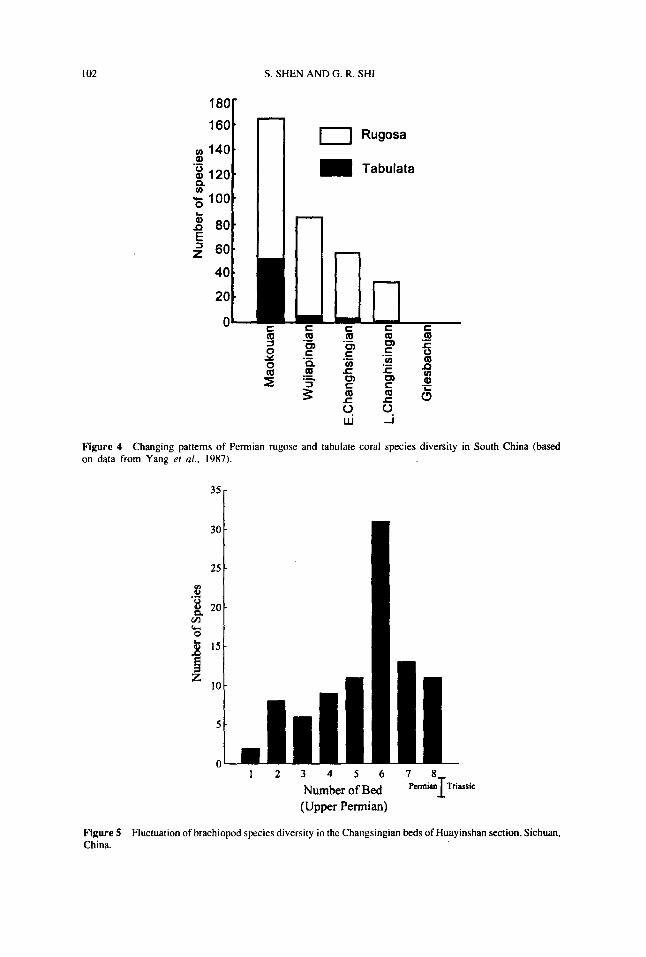
The investigation on the corals of South China made by Wang et al ( 1989) alsosuggests a distinct discontinuity of diversity across the Maokouan-Wujiapingian boundary,with some 14 families and 79 genera among 27 families and 119 genera extinguished.The statistics at specific level made by Yang et al ( 1987) shows that the number ofspecies of rugose corals reduced from 165 species in the Maokouan to 86 species inthe Wujiapingian, among which only 33 species continued to exist during the lateChanghsingian (Figure 4) A comparable reduction of species diversity is also evidentfor the tabulate corals (Figure 4) Similar conclusions have also been made by Ezaki( 1994) who recognised two terminal patterns for the Permian Rugosa, one associatedwith the Middle to Late Permian boundary and characterised by a progressive reductionpattern, and the other towards the end of the Changhsingian and distinguished by thedisappearance of the platform-dwelling colonial corals.
100

XI
q q
6 a~~~~
DIVERSITY AND EXTINCTION PATTERNS 101
-f3av,
a
oa4m
B.e.n
foceo
·r
fioB�so ·-h�P
'Ba"�n�
8-ra
a
e 3 "a.o ·sre" 5
,"
,e
3 E.�Pcau
S SHEN AND G R SHI
1
o
1
.o 1
e)
Ez
Figure 4 Changing patterns of Permian rugose and tabulate coral species diversity in South China (basedon data from Yang et al , 1987).
35
30
25
' 20
b 15
1 o
s 5
.l1 2
.I 13 4 5 6
Number of Bed(Upper Permian)
II7 8Permian I Tria ssic
Figure 5 Fluctuation of brachiopod species diversity in the Changsingian beds of Huayinshan section, Sichuan,China.
102
. 'u_

DIVERSITY AND EXTINCTION PATTERNS
.2
80
z'
I 5 10 15 20 25 29 'Number of Bed Pi- I r Tisic
.0
z
Number of Bed Permianl Triasic
120
100
060
z
20
0
Number of Bed P Ti I Tri ,Sic
Figure 6 Fluctuation patterns of brachiopod species diversity in the Changhsingian beds A Nantong section,Sichuan Province; B Chuanyuan, Sichuan Province; Guiding section, Guizhou Province (note that the bedsare not evenly spaced in the sections)
The second sharp fall of brachiopod diversity in South China occurred during butnot at the end of the Changhsingian, with 308 out of 325 Changhsiangian species, or94 %, and 84 % of Changhsiangian genera eliminated before the end of the Permian.Detailed metre or zonal-scale stratigraphic analysis of several continuous Permian-Triassic sections in South China reveals a multi-phase, stepwise end-Permian extinctionevent extended over the entire interval of the Changhsingian (Figs 5 and 6) This
103

S SHEN AND G R SHI
ciC.
4)* t
ci
4)* 3
z4 o
To
0)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 178 19
Number of Bed Permian I Triassic
Figure 7 Extinction pattern of Changhsingian foraminifer species in Liangfengya section, Sichuan Province.
conclusion may be examplified by the well-known Huayinshan section, Sichuan Province,of a continuous Permian-Triassic sequence with abundant Permian brachiopods (He andShen, 1990) Detailed collecting and stratigraphic analysis of brachiopod species in thesection indicate a stepwise extinction process, with the most severe drop of brachiopodspecies occurring at the middle Changhsingian (bed number 6 in Figure 5), between90-60 m below the recognised Permian-Triassic boundary Similar stepwise extinctionpattern has also been observed in Nantong and Chuanyuan sections of Sichuan Province(Figure 6 A and B) and the Guiding section of Guizhou Province (Figure 6 C).
The stepwise end-Permian extinction pattern demonstrated by the brachiopods is alsoexhibited by foraminiferids A stratigraphic analysis of 83 foraminiferid species in theChanghsing Formation of the Liangfengya section in Zhongliang Hill, Sichuan Province,revealed that 89 % of the 83 species became extinct by th end of the Changhsiangian,with the greatest loss ( 14 species) occurring at O 96 m below the P/T boundary (bed number15 in Figure 7), in comparison with 9 species, or 11 % extinguished just at the boundary.
The stepwise end-Permian extinction pattern is also reflected by the last occurrencesof different taxa or the same taxa at different sections below or near the Permian-Triassicboundary in various Permian-Triassic sequences in South China (Table 1).
SUMMARY AND DISCUSSION
In summary, the so-called end-Paleozoic mass extinction in the marine realm can bedivided into two phases: one occurring at the latest Maokouan and the other towardsthe end of the Changhsingian Both extinction events exhibited a relative short duration,estimated some 3-5 my on average but accelerated towards the end of the two nominalstages (note that the end-Changhsingian extinction could be slightly more rapid thanthe end-Makouan extinction according to the thickness of associated rocks; however,quantification of the exact duration of either extinction event is difficult to achieve).
104
at

DIVERSITY AND EXTINCTION PATTERNS
Table I The last occurrences of different fossil groups in seven sections of South China.
Last occurrenceFossil group point below PI Tr Locality Genus
boundary
7.14 m Chuanyuan Acropyge0.04 m Daijiagou Pseudophillipsia
Trilobites 14 44 m Guiding Pseudophillipsia0.34 m Nantong Genus unidentified0.46 m Zhongliangshan Pseudophillipsia
0.37 m Zhongliangshan LophopyllidiumRugose corals 0 34 m Nantong Genus unidentified
62.13 m Guiding Genus unidentifiedChuanyuan No corals
15.18 m Guiding Genus unidentifiedFusulinids 0 91 m Zhongliangshan Palaeofusulina, Nankinella
Foraminifers 1 78 m Zhongliangshan Glomospira
0.06 m Zhongliangshan Haydenella,0.06 m Nanting Spinomarginifera
Productids 14 44 m Guiding Spinomargini ra2.18 m Chuanyuan Haydenella, Waagenites0.21 m Daijiagou Haydenella,Chonetinella0.50 m Zunyi Neochonetes
Spinomarginifera
Since the discovery of high iridium concentration from the Cretaceous-Tertiaryboundary layers in Gubbio of Italy by Alvarez et al ( 1980), the possibility of an extra-terrestrial impact near the P/T boundary has also been intensively explored in SouthChina An iridium anomaly was subsequently reported by Sun et al ( 1984) and Xu andMa et al ( 1985) from South China, with a scenario of a possible meteorite impact asthe primary cause of the P/T mass extinction However, the analysis by Clark et al.( 1986) on the same southern Chinese sections found far less Ir than that reported bySun et al ( 1984) and Xu and Ma et al ( 1985) As a result, catastrophic hypothesesfor interpreting the end-Palaeozoic mass extinction in South China have been difficultto accept (Orth, 1989 ; Yin et al , 1989 ; He and Shen, 1990 ; Shen et al , 1992) Morerecently, Yin et al ( 1989, 1992) and Yang et al ( 1991) suggested a volcanic originfor the P/T boundary clays and argued that the end-Permian volcanism may have hadfatal effect on the survival of Permian marine faunas However, the importance of theacidic volcanism recorded near the P/T boundary is probably overstated because suchaltered volcanic ashes are common throughout the Carboniferous and Permian sequences.
Another more recently advanced view on the P/T extinction has been related to eustasy.A scenario of a 'global flooding' event has been suggested to relate to the P/T extinction(Hallam, 1989 ; Wignall and Hallam, 1993 ; Jin, Zhang and Shan, 1994) Based on faciesand faunal analyses from Pakistan and China, Wignall and Hallam ( 1993) proposed thatthe P/T extinction of marine invertebrate faunas may have been associated with aspectacularly rapid Griesbachian transgression which led to the widespread establishmentof deep-water anoxic and dysoxic conditions.
105
S SHEN AND G R SHI
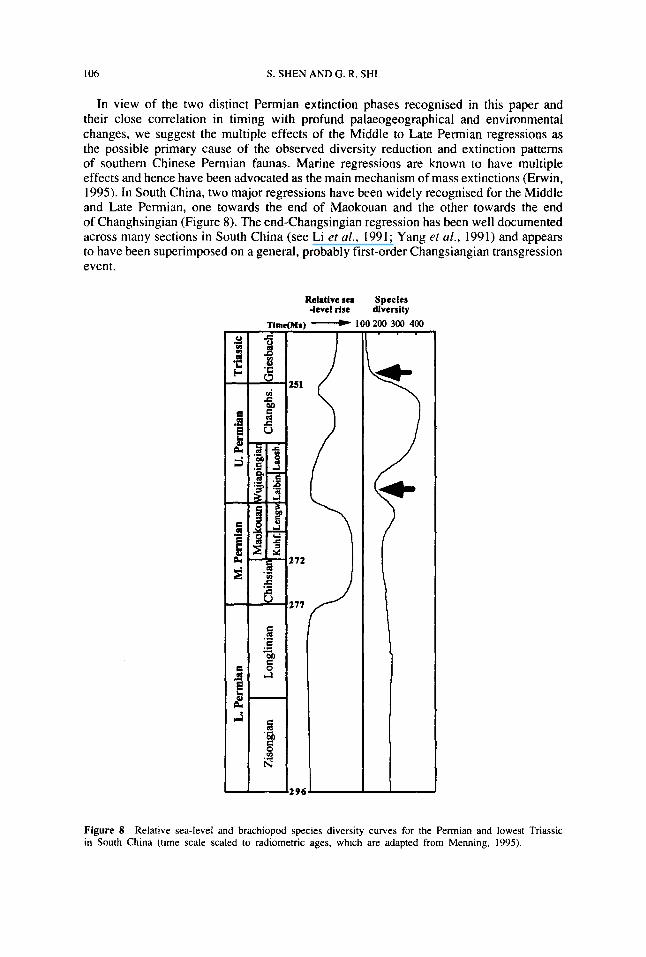
In view of the two distinct Permian extinction phases recognised in this paper andtheir close correlation in timing with profund palaeogeographical and environmentalchanges, we suggest the multiple effects of the Middle to Late Permian regressions asthe possible primary cause of the observed diversity reduction and extinction patternsof southern Chinese Permian faunas Marine regressions are known to have multipleeffects and hence have been advocated as the main mechanism of mass extinctions (Erwin,1995) In South China, two major regressions have been widely recognised for the Middleand Late Permian, one towards the end of Maokouan and the other towards the endof Changhsingian (Figure 8) The end-Changsingian regression has been well documentedacross many sections in South China (see Li et al , 1991 ; Yang et al , 1991) and appearsto have been superimposed on a general, probably first-order Changsiangian transgressionevent.
Relative sea Species-level rise diversity
Figure 8 Relative sea-level and brachiopod species diversity curves for the Permian and lowest Triassicin South China (time scale scaled to radiometric ages, which are adapted from Menning, 1995).
106

DIVERSITY AND EXTINCTION PATTERNS
The end-Maokouan and end-Changsiangian regional regression events may have beenrelated to the widely documented late Permian global mega-regression following theformation of Pangea at the end of Early Permian (Ross and Ross, 1995) The Maokouanregression is clearly related to the Dongwu Movement, which uplifted the southwesternregion of South China, causing a regional unconformity between the Maokouan Formationand the overlying Wujiaping Formation or equivalents However, marine depositioncontinued in limited areas of southeast China where a distinct brachiopod assemblage,the Lenguan fauna, was developed The Maokouan regression was immediately followedby a transgression in the Wujiapingian, resulting in the recovery of a large, diverse LateWujiapingian (Laoshanian) fauna, which persisted and further diversified in the earlyChanghsingian (Figure 8) The end-Changhsingian regression and apparently associatedextinction may have been linked to a significant global sea-level drop, exposing largeareas of continents and severely limiting areas of marine deposition into the Triassic.
Relationship between marine regression and extinction is complex and requires detailedglobal correlations of depositional sequences and bio-events in order to establish thesynchronity of regressions and extinction events So far, a number of effects of regressionshave been identified in relation to the mass extinctions, including the species-area effect(Schopf, 1974), deprovincialisation (Valetine, 1973), habitat destruction (Newell, 1967),nutrient reduction (Erwin, 1995), and deterioration of climatic conditions (Robinson,1973 ; Jablonski, 1986) (see Erwin, 1993 for more extensive reviews and references thereinprovided) Each of these hypotheses is yet to be evaluated against detailed analysis offaunal successions, lithology and geochemistry.
As Erwin ( 1995) pointed out, none of the existing extinction mechanisms seems tobe applicable to all reported Permian extinctions and associated evidence This has ledErwin ( 1995) to suggest a two-phase Permian extinction scenario with each phasedemanding a different mechanism (see also Stanley ahd Yang, 1994) According to Erwin,the first, longer, phase was mainly confined to the Pangean continents, excluding isolatedislands such as South China and Indo-China then scattered within eastern Tethys, andoccurred by the close of Guadalupian This phase clearly corresponds with the end-Maokouan extinction recognised in this paper Erwin's second phase of the Permianextinction began several million years later and was largely restricted to South Chinaand adjacent islands of Cathaysia This phase may be equivalent to the Changhsiangianextinction event recognised in this paper Erwin considers that the first phase of theextinction was most likely to have been related to the formation of Pangea and thatSouth China probably acted as a significant refugium during this phase This latterspeculation was also referred to but rejected by Jablonski ( 1985) and is also contradictoryto the fact that a dramatic reduction of species diversity and richness comparable tothe Pangean extinctions on other continents also occurred in this first phase in SouthChina, as discussed previously in this paper Interpretation of the second phase extinctionis more difficult although we recognise the apparent close relationship between theextinction and the end-Changhsingian regression.
Acknowledgements
Dr G R Shi's work on Late Palaeozoic brachiopod faunas and biogeograpohy issupported by the Australian Research Council Mr R P Irwin drafted the figures.
107

S SHEN AND G R SHI
References
Alvarez, L W , Alvarez, W , Asaro, F and Michel, H V ( 1980) Exterrestrial cause of theCretaceous/Tertiary extinction Science, 208, 1094-1108
Batten, R L ( 1973) The vicissitude of the gastropods during the interval of Guadalupian-Ladiniantime In: The Permian and Triassic Systems and their mutual boundary, edited by A Loganand L V Hills, pp 596-607 Canadian Society of Petroleum Geologists Memoir 2.
Chai, C F , Ma, S L , Mao, X Y , Sun, Y Y , Xu, D Y , Zhang, Q W and Yang, Z Z ( 1986)Elemental geochemical characters at the Permian-Triassic boundary section in Changxin,Zhejiang, China Acta Geologica Sinica, 60 ( 2), 139-150.
Chai, C F , Ma, S L , Mao, X Y , Sun, Y Y , Xu, D Y , Zhang, Q W and Yang, Z Z ( 1992)Geochemical constraits on the Permo-Triassic boundary events in South China In: Permo-Triassic events in the eastern Tethys An overview, edited by W C Sweet, Z Y Yang,J.M Dickens and H F Yin, pp 158-168 Cambridge: Cambridge univ Press.
Clark, D L , Wang, C Y , Charles, J O and James, S G ( 1986) Conodont survival and lowiridium abundances across the Permian-Triassic boundary in South China Science, 233,984-986.
Erwin, D H ( 1993) The great Paleozoic Crisis: Life and Death in the Permian New York:Columbia University Press, 323 pp.
Erwin, D H ( 1994) The Permo-Triassic extinction Nature, 367 ( 6460), 231-235.Erwin, D H ( 1995) The end-Permian mass extinction In The Permian of Northern Pangea.
1 Paleogeography, Paleoclimates, Stratigraphy, edited by P A Scholle, T M Peryt andD.S Ulmer-Scholle, pp 20-34 Berlin: Springer-Verlag.
Ezaki, Y ( 1994) Patterns and palaeoenvironmental implications of end-Permian extinction ofRugosa in South China Palaeogeography, Palaeoclimatology, Palaeoecology, 107 ( 1-2),165-177.
Hallam, A ( 1989) The case for sea-level change as a dominant causal factor in mass extinctionof marine invertebrates Philosophical transactions of the Royal Society of London B 325,437-455.
He, X L and Shen, S Z ( 1990) A research into the biotic alternation near the Permo-Triassicboundary in Zhongliang Hill, Chongqing City and its vicinity Documenta Nature, 61, 6-14.
Huang, T K and Zheng, D Q ( 1948) Subdivision of the Permian in Huayinshan, Sichuan.Geological review, 13 ( 3-4), 253-255.
Jablonski, D ( 1985) Marine regressions and mass extinctions: a test using the modern biota InPhanerozoic Diveristy Patterns, edited by J W Valentine pp 335-354 Princeton:Princeton University Press.
Jablonski, D ( 1986) Causes and consequences of mass extinctions: a comparative approach InDynamics of Extinctions, edited by D K Elliot, pp 183-229 New York: John Wiley.
Jin, Y G ( 1993) The pre-Lopingian benthos Crisis Compte Rendu, XII ICCP, Buenos Aries, 2,629-278.
Jin, Y G , Glenister, B F , Kotlyar, G V and Sheng, J Z ( 1994) An operational scheme of Permianchronostratigraphy Palaeoworld, 4, 1-13.
Jin, Y G , Zhang, J and Shang, Q H ( 1994) Two phases of the end Permian mass extinction.In Pangea: Global Environments and resources, edited by B Beauchamp Canadian Societyof Petroleum Geologists Memoir 17, 813-822.
Jin, Y G , Zhu, Z L and Mei, S R ( 1994) The Maokouan-Lopingian boundary sequences inSouth China Paleoworld, 4, 138-152.
Kanmera, K , Ishii, K and Toriyama,R ( 1976) The evolution and extinction patterns of PermianFusulinaceans Contribution to the Geology and Palaeontology of Southeast Asia, 17,129-154.
Li, L , Yang, D L and Feng, R L ( 1987) The brachiopods and the boundary of the LateCarboniferous-Early Permian in longlin Region, Guangxi Bulletin of the Yichang Institute ofGeology and mineral resouces, Chinese Academy of Geological Science, 11, 239-276.
Li, Z S , Zhan, L P , Yao, J X and Zhou, Y Q ( 1991) On the Permian-Triassic events in SouthChina probe into the end Permian abrupt extinction and its possible causes Saito Ho-on KaiSpecial Publication No 3 (Proceedings of Shallow Tethys 3, Sendai, Japan, 1990), 371-385.
108

DIVERSITY AND EXTINCTION PATTERNS
Li, Z S , Zhan, L P , Zhu, X F , Chang, J H and others ( 1989) Mass extinction and geologicalevents between Paleozoic and Mesozoic Era Acta Geologica Sinica, 60 ( 1), 1-15.
Liang, W P ( 1990) Lengwu Formation of Permian and its brachiopod fauna in Zhejiang province.P.R China Ministry of Geology and Mineral Resources Geological Memoirs, Series 2 ( 10),1-522.
Liao, Z T ( 1979) Uppermost Carboniferous brachiopods from western Guizhou Acta Palaeon-tological Sinica, 18 ( 6), 527-546.
Menning, M ( 1995) A numerical time scale for the Permian and Triassic periods: An integratedtime analysis The Permian of Northern Pangea 1 Paleogeography, Paleoclimates, Stratigraphy,edited by P A Scholle, T M Peryt and D S Ulmer-Scholle, pp 77-97 Berlin: Springer-Verlag.
Moore, R C (Editor) ( 1965) Treatise on Invertebrate Paleontology Part H Brachiopoda.Geological Society of America and the University of Kansas, 927 pp ( 2 vols).
Newell, N D ( 1967) Revolutions in the history of life Geological Society of America SpecialPaper, 89, 63-91.
Orth, C J ( 1989) Geochemistry of the bio-event horizons In Mass Extinctions: Processes andEvidence, edited by S K Donovan, pp 37-72 London: Belkhaven.
Robinson, P L ( 1973) Palaeoclimatology and continental drift In Implications of Continental Driftto the Earth Sciences, edited by D H Darlington and S K Runcorn, pp 451-476 London:Acdemic Press.
Ross, C A and Ross, J R P ( 1995) Permian sequence stratigraphy In The Permian of NorthernPangea 1 Paleogebgraphy, Paleoclimates, Stratigraphy, edited by P A Scholle, T M Perytand D S Ulmer-Scholle, pp 98-123 Berlin: Springer-Verlag.
Schopf, T J M ( 1974) Permian-Triassic extinctions: relation to sea-floor spreading Journal ofGeology, 82, 129-143.
Sepkoski, J J ( 1982) Mass extinction in the Phanerozoic oceans: A review Geological Societyof America Special Paper, 190, 283-289.
Sepkoski, J J ( 1992) A compendium of fossil marine animal families 2nd ed Milwankee PublicMuseum, Contribution in Biology and Geology 83, 1-156.
Shen, S Z and He,s X L ( 1991 a) Changhsingian brachiopod assemblages in Zhongliang Hill,Chongqing Journal of Stratigraphy, 15 ( 3), 189-196.
Shen, S Z and He, X L ( 1991 b) Permo-Triassic "mixed fauna" in South China and Permo-Triassicboundary Journal of Changchun University of Earth Science, 21 ( 2), 121-128.
Shen, S Z and He, X L ( 1994 a) Brachiopod assemblages from the Changhsingian to lowerestTriassic of Southwest China and correlations over the Tethys Newsletters on Stratigraphy 31 ( 3),151-165.
Shen, S Z and He, X L ( 1994 b) Changhsingian brachiopod fauna from Guiding, Guizhou ActaPalaeontologica Sinica 33 ( 4), 440-454.
Shen, S Z , Shi, G R and Archbold, N W ( 1996) A Compendium of the Permian BrachiopodFaunas of the Western Pacific Region 6 Wujiapingian Deakin University, School of AquaticScience and Natural Resources Management, Technical Paper, 1996/2 (in press).
Shen, S Z , Zhang, X P and Xue, C J ( 1992) Permo-Triassic biotic mass extinction-Relation tothe environmental changes In Symposium on coal geology and exploration, edited by Cheng,J.Z and Shen, S Z pp 198-204 Jilin: Jilin University Press, Jilin, China.
Shi, G R and Archbold, N W ( 1993 a) A Compendium of the Permian Brachiopod Faunas ofthe Western Pacific Region 1 Asselian-Tastubian Deakin University, School of Aquatic Scienceand Natural Resources Management, Technical Paper, 1993/1, iv + 68 pp.
Shi, G R and Archbold, N W ( 1993 b) A Compendium of the Permian Brachiopod Faunas ofthe Western Pacific Region 2 Baigendzhinian-Early Kungurian Deakin University, School ofAquatic Science and Natural Resources Management, Technical Paper, 1993/2, iv + 74 pp.
Shi, G R and Archbold, N W ( 1994 a) A Compendium of the Permian Brachiopod Faunas ofthe Western Pacific Region 3 Kazanian-Midian Deakin University, School of Aquatic Scienceand Natural Resources Management, Technical Paper, 1994/2, iii + 89 pp.
Shi, G R and Archbold, N W ( 1994 b) A Compendium of the Permian Brachiopod Faunas ofthe Western Pacific Region 4 Sterlitamakian-Aktastinian Deakin University, School of AquaticScience and Natural Resources Management, Technical Paper, 1994/3, iii + 52 pp.
109

S SHEN AND G R SHI
Shi, G R , Archbold, G R and Zhan, L P ( 1995) Distribution and characteristics of mixed(transitional) mid-Permian (Late Artinskian-Ufimian) marine faunas in Asia and theirpalaeogeographical implications Palaeogeography, Palaeoclimatology, Palaeoecology, 114, -241-271.
Shi, G R , Shen, S Z and Archbold, N W ( 1996) A Compendium of the Permian BrachiopodFaunas of the Western Pacific Region 5 Changhsiangian Deakin University, School of AquaticScience and Natural Resources Management, Technical Paper, 1996/1 (in press).
Stanley, S M and Yang, X N ( 1994) A double mass extinction at the end of the Paleozoic Era.Science 266 ( 5189), 1340-1344.
Sun, Y Y and Chai, Z F ( 1984) The discovery of Ir anomaly in the P/Tr boundary clay inChangxing, Zhejiang, China and its signicance Developments in Geoscience, Contribution to27th International Geological Congress Science Press, bejing, 391 p.
Valentine, J W ( 1973) Evolutionary Paleoecology of the Marine Biosphere Englewood Cliffs:Prentice-Hall, 511 pp.
Wang, H Z , He, X Y and Chen, J Q ( 1989) Classification, evolution and biogeography of thePaleozoic corals of China Beijing: Science Press.
Wignall, P B and Hallam, A ( 1993) Griesbachian (Earliest Triassic) palaeoenvironmental changesin the Salt Range, Pakistan and Southeast China and their bearing on the Permo-Triassicextinction Palaeogeography, Palaeoclimatology, Palaeoecology, 102, 215-237.
Xu, D Y , Ma, S L , Chai, Z F , Sun, X Y , Zhang, Y Y and Yang, Z Z ( 1985) Aundance variationof iridium and trace elements at the Permian-Triassic boundary at Shangsi in China Nature,314, 154-156.
Xu, D Y , Zhang, Q W and Sun, Y Y ( 1987) Biomass extinction A fundamental indicator formajor natural divisions of geological history Acta Geologica Sinica, 61 ( 3), 195-204.
Xu, G R and Grant, R E ( 1994) Brachiopods near the Permian-Triassic boundary in South China.Smithsonian Contribution to Paleobiology, 76, 1-68.
Yang, Z Y , Wu, S B , Yin, H F , Xu, G R and Zhang, K X ( 1991) Geological events of Permo-Triassic transitional period in South China Geological Publishing House, Bejing, 190 pp.
Yang, Z Y , Yin, H F , Wu, S B , Yang, F Q , Ding, M H and Xu, G R ( 1987) Permian-Triassicboundary stratigraphy and ftuna of the South China Geological Publishing House, Bejing,435 pp.
Yin, H F , Huang, S , Zhang, K X , Yang, H Q , Ding, M H , Bi, X M and Zhang S X ( 1989)Volcanism at the Permian-Triassic boundary in South China and its effects on mass extinction.Acta Geologica Sinica, 2, 417-431.
Yin, H F , Huang, S J , Zhang, K X , Hansen, H J , Yang, F Q , Ding, M H and Bie, X M ( 1992)The effects of volcanism on the Permo-Triassic events in the eastern Tethys An overview.In Permo-Triassic Events in the Eastern Tethys: Stratigraphy, Classification, and Relations withthe Western Tethys, edited by W C S Sweet, Z Y Yang, Dickins, J M and H F Yin, pp.146-157 Cmabridge: Cambridge University Press.
110





























