Embed Size (px)
Citation preview

END-PERMIAN MASS EXTINCTION OF FORAMINIFERS IN THENANPANJIANG BASIN, SOUTH CHINA
HAIJUN SONG,1 JINNAN TONG,1 Z. Q. CHEN,2 HAO YANG,1 AND YONGBIAO WANG1
1Key Laboratory of Biogeology and Environmental Geology of Ministry of Education, Wuhan 430074, PR China, ,[email protected].,,[email protected]., ,[email protected]., ,[email protected].; and 2School of Earth and Environment,
The University of Western Australia, 35 Stirling Highway, Crawley, WA 6009, Australia, ,[email protected].
ABSTRACT—Newly obtained foraminifer faunas from the Permian-Triassic (P-Tr) transition at the Dajiang andBianyang sections in the Nanpanjiang Basin, South China, comprise 61 species in 40 genera. They belong to thePalaeofusulina sinensis Zone, the youngest Permian foraminifer zone in South China. Quantitative analysis revealsthat the last occurrences of more than a half of species (28/54) fall into a 60-cm-interval at the uppermostChanghsingian skeletal packstone unit and thus calibrate the end-Permian extinction to the skeletal packstone-calcimicrobial framestone boundary. About 93% (54/58) of species of the latest Permian assemblage became extinctin the P-Tr crisis. Four major foraminiferal groups, the Miliolida, Fusulinida, Lagenida, and Textulariina, haveextinction rates up to 100%, 96%, 92%, and 50%, respectively, and thus experienced selective extinctions. BothHemigordius longus and ?Globivalvulina bulloides temporarily survived the end-Permian extinction event andextended into the earliest Triassic but became extinct soon after. The post-extinction foraminifer assemblage ischaracterized by the presence of both disaster taxa and Lazarus taxa. Foraminifer distribution near the P-Trboundary also reveals that the irregular contact surface at the uppermost Permian may be created by a massivesubmarine dissolution event, which may be coeval with the end-Permian mass extinction. A new species,Rectostipulina hexamerata, is described here.
INTRODUCTION
T HE MOST severe disaster event in the Phanerozoic historyof Earth’s life happened in the Permian-Triassic (P-Tr)
transition, and it eliminated more than 90% of all marinespecies (Erwin, 1993). However, the causes of this enigmaticcrisis have long been disputed despite the intense study of thisevent. Multiple scenarios suggesting the causes of the P-Trevent have been proposed (see Erwin, 2006 for more details),but there is growing evidence that the following four majortriggers are the most likely causes for this crisis: bolide impact,flood basalt eruption of the Siberian Traps, oceanic anoxia,and catastrophic release of seafloor methane (Knoll et al.,2007). Nevertheless, any reasonable models interpreting the P/Tr biotic crisis need to be tested by analysis of precise bioticextinction patterns and physiological reactions of victims andsurvivors during the P-Tr transition (Knoll et al., 2007).
Most current scenarios interpreting the P-Tr crisis considerbiotic extinction as an abrupt, monoepisodic event, asdocumented by Jin et al. (2000) from the Meishan section,South China. However, Jin et al.’s (2000) extinction pattern isbased on the fossil record from only one P-Tr boundarysection. Even in Meishan, abundant brachiopods and fora-minifer faunas have been obtained from Beds 25–27,immediately above the extinction horizon calibrated to thebase of Bed 25 in Meishan by Jin et al.’s (2000) influentialstudy (Chen et al., 2005, 2006; Song et al., 2006, 2007).Quantitative analysis of the newly obtained foraminifer datafrom Meishan reveals a two-stage extinction pattern near theP-Tr boundary (Song et al., 2009), which agrees well with theepisodic extinction pattern indicated by biomarker analysisfrom the same section (Xie et al., 2005). The two-stageextinction pattern is also confirmed by extremely abundantbenthic fossils obtained from a shallow platform facies of theP-Tr boundary section at Huangzhishan, about 40 km fromMeishan (Chen Z. Q. et al., 2009). Accordingly, bioticextinction patterns based on sound systematic paleontologyacross the P-Tr boundary have been far less studied in
comparison with the growing number of geochemical studiesof this event. Current, updated fossil records from extensive P-Tr boundary sections are crucial to revealing the true bioticextinction pattern during this critical period.
Unlike other fossil groups, foraminifers are one of thecommonest fossil groups through the P-Tr transition and aredistributed in a variety of facies settings. Thus, foraminiferdata can be employed to document the biotic extinctionpattern by using quantitative analysis (Tong and Kuang, 1990;Yang et al., 1993; Stanley and Yang, 1994; Rampino andAdler, 1998; Jin et al., 2000; Groves et al., 2005). In additionto compilation-data-based studies (Tappan and Loeblich,1988; Tong and Shi, 2000; Groves and Altiner, 2005;Marquez, 2005), several recent outcrop-section-based workshave offered new insight into the foraminifer evolutionaryprocess, extinction patterns and subsequent recovery duringthe P-Tr transition (Altiner et al., 2005; Groves et al., 2005,2007; Song et al., 2007, 2009).
In this study, we will undertake quantitative analysis todetermine foraminiferal extinction patterns near the P-Trboundary based on newly obtained foraminifer data from theGreat Bank of Guizhou (GBG) (term of Lehrmann et al.,1998) in the Nanpanjiang Basin, Southwest China (Fig. 1).The GBG records the thickest carbonate successions of the P-Tr transition in the world (Lehrmann et al., 2007), and thusoffers unique opportunity to collect the continuous foramin-ifer samples across the P-Tr boundary. In the GBG, there havebeen great advances in the researches on the P-Tr massextinction and its aftermath in recent years (Lehrmann, 1999;Lehrmann et al., 2003, 2006, 2007; Ezaki et al., 2003, 2008;Krull et al., 2004; Payne et al., 2004, 2006a, b, 2007; Wang etal., 2005; Liu et al., 2007; Chen et al., 2009). However, fewstudies have addressed the P-Tr mass extinction in terms ofpattern and possible horizon except for the studies by Krull etal. (2004) and Payne et al. (2007). In fact, paleontological dataover the P-Tr transition from this region have been poorlydocumented except for conodont faunas (Lehrmann et al.,
J. Paleont., 83(5), 2009, pp. 718–738
Copyright ’ 2009, The Paleontological Society
0022-3360/09/0083-0718$03.00
718

2003; Liu et al., 2007; Ezaki et al., 2008; Chen et al., 2009). Thepotential extinction horizon is usually calibrated to the contactbetween the latest Permian skeletal packstone (SP) unit andthe overlying calcimicrobial framestone (CF) unit. This isbased on both the correlation of carbon isotopic excursioncurves with their counterparts recorded in the Meishan section(Krull et al., 2004) and a sharp turnover from the fossil-enriched skeletal packstone to the poorly fossiliferouscalcimicrobial framestone (Lehrmann et al., 2003). However,the skeletal packstone-calcimicrobial framestone (SP-CF)boundary is an irregular surface, which has been interpretedas sub-aerial exposure (Liu et al., 2007), submarine dissolution(Payne et al., 2007) or both (Collin et al., 2009). Consequently,the interpretations of the geological event associated with theP-Tr mass extinction in the GBG are disputed.
In addition, although various microfossils (i.e., foramini-fers) and macrofossils (sponges, crinoids, gastropods, bra-chiopods, corals, and calcareous algae) have been frequentlyillustrated from the SP unit (Lehrmann et al., 2003; Krull etal., 2004; Payne et al., 2006a; Collin et al., 2009), notaxonomic studies of these fossil groups across the P-Trboundary have been undertaken from the GBG except forconodonts. Thus, we do not know specifically what formswere wiped out and how they became extinct in the great dyingat the end of the Permian. Newly obtained conodont datafrom several different groups working on this problem suggesthighly differing P-Tr boundary positions in the GBG(Lehrmann et al., 2003; Liu et al., 2007; Ezaki et al., 2008;Chen et al., 2009; see below for further discussion), and thuscall into question the previous estimates of the end-Permianextinction horizon in the GBG (Krull et al., 2004). As a result,independent determination of the potential extinction horizonbased on quantitative analysis of the fossil record from GBGbecomes essential in the study of the P-Tr event in this region.This study makes the first effort to document P-Tr foraminifertaxonomy in the GBG and to elucidate foraminiferalevolutionary processes and extinction patterns over the P-Tr
transition. The relevant scenarios interpreting the P-Tr massextinction and associated geological events recorded in theGBG are also evaluated in a broad context based on our newforaminifer data.
THE STUDY SECTIONS, STRATIGRAPHY AND SAMPLING
Foraminifer samples were collected from the Dajiang andBianyang sections in the Nanpanjiang Basin, southernGuizhou Province, Southwest China (Fig. 1). During thePermian and Triassic, the GBG was built in the NanpanjiangBasin (Lehrmann et al., 2003) where the P-Tr carbonatesuccessions are well exposed at a series of sections from west toeast. These are Dawen, Heping, Dajiang, Rongbao, andLangbai sections, approximately one km apart from oneanother. Here, the P-Tr successions are exactly the same in allfive sections. The Upper Permian is comprised mainly ofskeletal packstone of the Wujiagping Formation yieldingabundant fossils of foraminifers, sponges, gastropods, bra-chiopods, corals, and calcareous algae. This formation isoverlain by calcimicrobial framestone of the Daye Formation,yielding Renalcis-like microfossils (Lehrmann, 1999; Wang etal., 2005). The Dajiang section is situated next to the well-studied Heping section (Lehrmann et al., 2003, 2007; Krull etal., 2004; Payne et al., 2007). The SP unit exposed at Dajiang is15 m thick, while the CF unit is 16 m thick.
Lehrmann et al. (2003) reported the conodonts Hindeodusparvus and Isarcicella isarcica Zones from the CF unit at theHeping section and located the P-Tr boundary horizon about65 cm above the SP-CF boundary, based on the firstoccurrence of Hindeodus parvus. These authors have alsofound the fusulinid Palaeofusulina sp. and conodont Clarkinachangxingensis occurring in the SP unit at the Heping andDajiang sections. The last occurrence of the former speciescorresponds to the horizon about 1 m below the SP-CFboundary, while that of the latter is the top of the SP unit(Lehrmann et al., 2003; Krull et al., 2004). Later, Liu et al.(2007) reported H. parvus, together with H. praeparvus, H.
FIGURE 1—Locations of the Dajiang and Bianyang sections, southern Guizhou Province, Southwest China. Early Triassic paleogeographic map ofthe Great Bank of Guizhou as modified from Lehrmann et al. (2001).
SONG ET AL.—PERMIAN EXTINCTION OF CHINESE FORAMINIFERA 719

latidentatus, H. eurypyge, H. typicalis, Clarkina changxingen-sis, and C. deflecta in the topmost skeletal packstone, justbelow the SP-CF boundary at the neighboring Dawen section(Fig. 2). The presence of H. parvus in the SP unit has not beenconfirmed by Chen et al.’s (2009) study from the same section.However, most of the other conodont species of Liu et al.(2007) have also been recovered from the top of the SP unit byChen et al. (2009). Chen (2009) also found C. zhejiangensis inthe same horizon and H. parvus at the middle part of the CFunit, 7.45 m above the base of the CF unit (Fig. 2). Ezaki et al.(2008) reported conodont faunas similar to those of Liu et al.(2007) from the topmost SP unit of the neighboring Langbaisection. However, both Liu et al. (2007) and Ezaki et al. (2008)considered that H. parvus conodont specimens may have beenreworked and deposited secondarily from the overlying CFunit; they placed the P-Tr boundary at the SP-CF boundarysince H. parvus also occurs at the base of the CF unit.Similarly, Chen et al. (2009) assumed that the conodont faunacontaining both C. zhejiangensis and H. eurypyge may haveresulted either from an insoluble residue concentrated at thebedding surface contact due to dissolution of the originalsediments between the SP and CF units or the result ofstratigraphic re-deposition associated with this irregularcontact. The former species is confined to Beds 25 to 27 andthe latter to Bed 27a and higher strata at the Meishan section(Chen et al., 2009, fig. 5). It should be noted that C.zhejiangensis ranges from Bed 24d to Bed 27d at Meishanaccording to Jiang et al. (2007). These authors agree with
Lehrmann et al. (2003) that the P-Tr boundary is placed at thelower part of the microbialite unit. Consequently, there aretwo different views about the position of the P-Tr boundary atthe GBG sections: 1) the boundary coincides with the base ofthe microbialite unit (Liu et al., 2007; Ezaki et al., 2008); 2) theboundary is through the lower part of the microbialite unit,about 0.6–1 m above the SP-CF boundary (Lehrmann et al.,2003; Chen et al., 2009). Regardless the various P-Trboundary positions at the GBG sections, all of these studiescalibrated the end-Permian mass extinction to the SP-CFboundary, marked by a distinct negative-shifting excursion ofd13Ccarb (Krull et al., 2004; Payne et al., 2004). Termination ofthe microbialite unit is marked by the conodont Isarcicellastaeschei Zone followed by the I. isarcica Zone (Liu et al.,2007; Ezaki et al., 2008; Chen et al., 2009), which define Beds27d–28 and Bed 29 respectively at the Meishan section (Jianget al., 2007; see also Fig. 2).
The Bianyang section is located about 15 km northwest ofthe Dajiang section (Fig. 1) and represents the deep-waterbasin facies sedimentation during the P-Tr transition (Enos etal., 2007). Here, the Upper Permian Dalong Formationconsists of thin-bed cherty limestone. Macrofossils are veryrare in the upper Dalong Formation. We have now discoveredthe ammonite Pseudotirolites sp. from the top of theformation. This ammonoid species is characteristic of thelatest Changhsingian ammonoid assemblage in South China(Zhao et al., 1981). The Dalong Formation is overlain by theEarly Triassic Luolou Formation, which consists mainly of
FIGURE 2—Correlation of the P-Tr boundary beds between the GBG and Meishan. The conodont data of the GBG from Lehrmann et al. (2003), Liuet al. (2007), Ezaki et al. (2008), and Chen et al. (2009); the conodont data of Meishan section from Jiang et al. (2007); the foraminifer data of Meishanfrom Song et al. (2009).
720 JOURNAL OF PALEONTOLOGY, V. 83, NO. 5, 2009

thin-bed limestone and mudstone (Enos et al., 2007). Thepresence of the earliest Triassic ammonoid Ophiceras sp. andbivalve Claraia spp. at the basal Luolou Formation suggeststhe P-Tr boundary is at the base of the formation. The MiddleTriassic sequence, belonging to the Xinyuan Formation, isnicely exposed at the Bianyang section and has been wellstudied previously in terms of biostratigraphy and paleoenvir-onmental analysis (Tong, 1998).
One hundred and seventy-six samples were collected fromthe P-Tr transition of the Dajiang section. Of these, 83 sampleswere taken from the Upper Permian skeletal packstone of theWujiaping Formation and the remaining 93 samples from thecalcimicrobial framestone and overlying packstone of theDaye Formation (Fig. 3). The section was sampled at anaverage interval of about 20 cm. One hundred, seventy-onesamples were collected from the P-Tr transition of theBianyang section. Of these, only five samples were collectedfrom the cherty limestone of the Dalong Formation while 155samples came from the Lower Triassic succession and 11samples from the Middle Triassic Bianyang Formation, butonly 68 samples from the P-Tr boundary beds have beenstudied for foraminifers (Fig. 4).
FORAMINIFER BIOSTRATIGRAPHY
Palaeofusulina species are herein confirmed to occur in thetopmost SP unit, although the genus is reportedly present atthe horizon about 1 m below the top of the unit (Krull et al.,2004, fig. 3). Palaeofusulina sinensis is present at the top of theSP unit at the Dajiang section and at the top of the DalongFormation at the Bianyang section (Figs. 3, 4). At Dajiang,the foraminifers from the SP unit include 58 species in 37genera (Fig. 3). This assemblage is characterized by suchspecies as Colaniella nana, Frondina permica, Nankinella minor,Palaeofusulina sinensis, Partisania typica. Pisolina excessa,Rectostipulina quadrata, Reichelina pulchra, Robuloides lens,Sengoerina argandi, and Tetrataxis planolocula. Of these, P.sinensis is the indicator species of the Palaeofusulina sinensisZone in South China (Zhao et al., 1981; Sheng et al., 1984). Acomparable foraminifer fauna has been described from theuppermost Changxing Formation of the Meishan section(Song et al., 2007). The Meishan fauna comprises 64 species in36 genera (Song et al., 2007) and is assigned to thePalaeofusulina sinensis Zone (Zhao et al., 1981; Sheng et al.,1984; Song et al., 2006). Of these, 70% of genera of theDajiang fauna are also present in the Meishan assemblage and34 species (59%) co-occur in the Meishan fauna, suggestinghigh similarities at both generic and species levels. TheDajiang fauna therefore resembles remarkedly the Meishanassemblage, suggesting that both faunas were contemporane-ous. Thus, the Dajiang fauna is also assignable to the P.sinensis Zone (Fig. 3). In Meishan the P. sinensis Zone isassociated with the conodont Neogondolella yini-H. praeparvus(Jiang et al., 2007) of late Changhsingian age (Zhao et al.,1981; Sheng et al., 1984; Song et al., 2009), just below the end-Permian mass extinction horizon at the base of Bed 25 (Jin etal., 2000). Thus, the foraminifers from the skeletal packstoneunit point to a late Changhsingian age.
A relatively small foraminifer fauna occurs in the upper-most Dalong Formation at the Bianyang section. It is alsocharacterized by the presence of P. sinensis, and thus isassignable to the P. sinensis Zone, strengthening the ageconstraints by the presence of the ammonoid Pseudotirolitessp. at Bianyang.
Foraminifers from the CF unit are very rare; onlyRectocornuspira kalhori occurs in both sections, and Earlandia
sp. is found only in the Dajiang section. The fauna is too smallto establish one biozone. However, R. kalhori is very commonin the lowest Triassic all over the world, for instance, in theeastern Sichuan and Guangxi regions, South China (Ezaki etal., 2003; Galfetti et al., 2008), southern Turkey (Crasquin-Soleau et al., 2004; Altiner et al., 2005; Groves et al., 2005;Payne et al., 2007), northern Italy (Groves et al., 2007), andAustria (Boeckelmann, 1991). The presence of abundant R.kalhori specimens broadly indicates a latest Changhsingian toearliest Griesbachian age, equivalent to the conodontsNeogondelella meishanensis Zone to H. parvus Zone (Jiang etal., 2007).
EXTINCTION PATTERN
Stratigraphic distributions of the P-Tr foraminifers from thestudied sections show a gradual disappearance near the P-Trboundary (Figs. 3, 4). However, the raw data of stratigraphicdistributions do not reveal a true extinction pattern simply dueto the ‘‘Signor-Lipps effect,’’ a common phenomenon blurringthe recognition of true extinction patterns in the studiedsections (Signor and Lipps, 1982). Signor and Lipps (1982)stated that a random distribution of errors at the end points ofstratigraphic ranges can generate artificial range truncationsand apparent gradual decline preceding a sudden massextinction pattern. This phenomenon was termed the Signor-Lipps effect (Raup et al., 1989; Strauss and Sadler, 1989).Meldahl (1990) developed a method to partially negate theSignor-Lipps effect. He identified three kinds of extinctionpatterns (sudden, stepwise, and gradual) by simulating asudden extinction of a modern molluscan biota using corestaken in the intertidal zone of the northern Gulf of California.Since then, Meldahl’s approach has been frequently applied toreveal the true extinction pattern during the P-Tr transitionbased on stratigraphic ranges in fossil record (Rampino andAdler, 1998; Jin et al., 2000; Groves et al., 2005; Groves et al.,2007; Song et al., 2009). Although several studies exploredother ramifications of the Signor-Lipps effect and usedconfidence intervals to place the endpoints of stratigraphicranges and posit the mass extinction boundary (Marshall,1990, 1995; Solow, 1996; Solow and Smith, 2000; Springer,1990; Wang and Marshall, 2004; Wang and Everson, 2007),these approaches require precise radiometric age constraintsfor the fossil horizons and thus are not applicable to the fossilrecord from the Dajiang and Bianyang sections, where noradiometric ages are available for the P-Tr transition.
Meldahl’s (1990) program has been performed to test theextinction pattern of foraminifers at the Dajiang section. Lastoccurrences and stratigraphic abundance of 54 species throughthe P-Tr transition are shown herein (Fig. 5). Last occurrencesof more than half of taxa (28/54) fall in the topmost 60-cm-interval of the Permian packstone unit (Fig. 5). If taking intoaccount the species having stratigraphic abundances .15%,the last occurrences of 11 out of 13 species (85%) fall in the 60-cm-interval just below the SP-CF boundary. Only twospecimens belonging to two species were uncovered from themicrobialite unit. As a result, the possible extinction horizoncan be precisely calibrated to the SP-CF boundary at Dajiang,as recognized previously in the GBG (Krull et al., 2004; Payneet al., 2007).
THE EVOLUTIONARY PROCESS AT THE END OF THE PERMIAN
Of the 58 foraminifer species occurring in the SP unit, 52species (90%) disappear below the SP-CF boundary, the end-Permian mass extinction horizon documented above, at theDajiang section (Fig. 3). Both Hemigordius longus and
SONG ET AL.—PERMIAN EXTINCTION OF CHINESE FORAMINIFERA 721
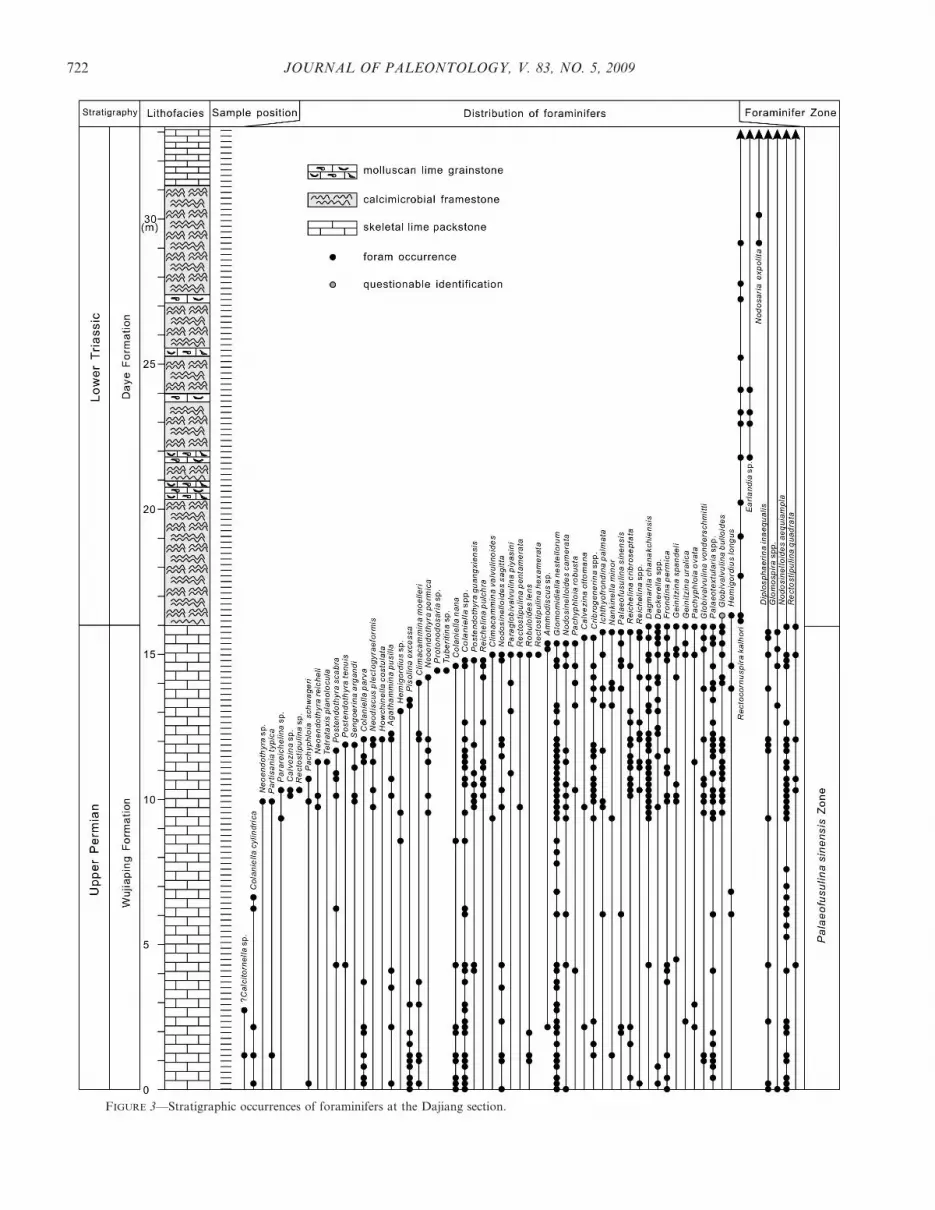
FIGURE 3—Stratigraphic occurrences of foraminifers at the Dajiang section.
722 JOURNAL OF PALEONTOLOGY, V. 83, NO. 5, 2009
?Globivalvulina bulloides cross the SP-CF boundary andappear in the basal part of the microbialite unit about 40 cmabove the SP-CF boundary. These two species, however, donot reappear in the higher strata. Four species, Diplosphaerinainaequalis, Glomospira spp., Nodosinelloides aequiampla, andRectostipulina quadrata, of the latest Permian fauna persistinto the middle Lower and Middle Triassic, although they donot occur in the microbialite unit at the studied sections (Songet al., unpublished data). At Bianyang, 13 out of 16foraminifer species (81%) disappear below the P-Tr boundarywith three species reappearing in the middle Lower andMiddle Triassic (Fig. 4).
Statistical pictures show that almost all of the orders of thePaleozoic foraminifera suffered the severe impact of the end-Permian mass extinction (Tappan and Loeblich, 1988; Tongand Shi, 2000; Tong, 2004; Groves and Altiner, 2005). The sameis also true at the studied sections where the extinction rates ofthe Fusulinida, Lagenida, Textulariina, and Miliolida, fourcommon orders of the Late Paleozoic foraminifera, are up to96% (25/26), 92% (22/24), 50% (1/2), and 100% (6/6),respectively (Fig. 6). It should be noted that the extinct taxacounted here include those species disappearing across the P-Trboundary not only at the studied sections but also worldwide.
The fusulinoidean fusulinides dominated the Carboniferousand Permian foraminifer assemblages but suffered the end-Guadalupian (Middle Permian) mass extinction, and theyfailed to recover in the Late Permian (Lopingian), when non-fusulinoidean fusulinides, which have microgranular tests,seemed to thrive and developed more complex test architecture(Stanley and Yang, 1994; Tong and Shi, 2000; Ota andIsozaki, 2006). The number of the Lagenida taxa increasedrapidly during the Late Permian when this group took overthe dominant role from the Fusulinida and became the mostabundant foraminifer group numerically in the late Changh-singian in the deep-water facies Bianyang section and platformramp facies P-Tr boundary sections such as the Meishansection (Song et al., 2009) and the Liangfengya section (Tongand Kuang, 1990) in South China. In contrast, fusulinidesdominated the Late Permian foraminifer assemblages inshallow platform facies settings such as the Dajiang section.At Dajiang, the upper Changhsingian contains 26 fusulinidspecies in 18 genera (Fig. 3), most of which were wiped out inthe end-Permian crisis. An exception, Diplosphaerina inaequa-lis (Derville, 1931), survived into the middle Lower andMiddle Triassic.
Lagenides were the second largest group among the lateChanghsingian foraminifer assemblages in the NanpanjiangBasin and comprise 24 species in such genera as Calvezina,Colaniella, Cryptoseptida, Frondina, Geinitzina, Howchinella,Ichthyofrondina, Nodosinelloides, Pachyphloia, Partisania,Rectostipulina, and Robuloides. Most of them were widespreadin regions of the world elsewhere, such as in South China(Wang, 1976; Tong and Kuang, 1990; Lin et al., 1990; Shanget al., 2003; Gu et al., 2002, 2005; Zhang and Hong, 2004;Song et al., 2006, 2007), Japan (Kobayashi, 1997, 2004, 2005),Pakistan (Okimura et al., 1985; Mertmann, 2000), India(Okimura and Ishii, 1981), Iran (Bozorgnia, 1973), Russia(Pronina, 1988; Pronina-Nestell and Nestell, 2001; Karavaevaand Nestell, 2007), Greece (Nakazawa et al., 1975; Vachard etal., 2003), Hungary (Berczi-Makk, 1992), Turkey (Altiner,1981; Zaninetti et al., 1981; Koyluoglu and Altiner, 1989; Unalet al., 2003; Groves et al., 2005), and Italy (Jenny-Deshusses etal., 2000; Groves et al., 2007). In Nanpanjiang, other than twospecies, Nodosinelloides aequiampla and Rectostipulina quad-rata, reoccurring in the middle Lower and Middle Triassic, alllagenid taxa disappeared forever near the end-Permianextinction horizons.
When compared with both the Fusulinida and Lagenida,the Textulariina and Miliolida have a small number of species
FIGURE 4—Stratigraphic occurrences of foraminifers at the Bianyangsection.
FIGURE 5—Stratigraphic abundance versus last observed occurrence ofthe latest Permian foraminifer species from the Dajiang section.
SONG ET AL.—PERMIAN EXTINCTION OF CHINESE FORAMINIFERA 723

at the studied section. The Textulariina contains onlyAmmodiscus sp. and Glomospira sp., while the Miliolidaincludes six species in five genera: Agathammina pusilla,?Calcitornella sp., Glomomidiella nestellorum, Hemigordiuslongus, H. sp., and Neodiscus plectogyraeformis. Of these,only Glomospira spp. of the Textulariina survived into theTriassic.
SURVIVORS, LAZARUS TAXA AND DISASTER TAXA
At Dajiang both Hemigordius longus and ?Globivalvulinabulloides occur on both sides of the SP-CF boundary,corresponding to the mass extinction horizon. H. longus hasa widespread distribution in the Middle and Upper Permian(Altiner et al., 2003) but has never been reported from theLower Triassic strata. G. bulloides is very common in thePermian assemblages but also occurs in the upper part of Bed27 (earliest Triassic), the Survival Fauna Bed 2 (Chen andMcNamara, 2006; Chen et al., 2006) at the Meishan section(Song et al., 2007). Here, both species occurred about 40 cmabove the SP-CF boundary but disappeared soon after andnever reoccurred in the higher horizons of the Induan inNanpanjiang and other regions in the world. These twosurvivors therefore fall into a pattern that was termed ‘DeadClade Walking’ (DCW) by Jablonski (2002), similar to thePermian relict foraminifers and brachiopods occurring in Bed27 of the Meishan section (Chen et al., 2005, 2006; Song et al.,2007, 2009). In contrast, four Permian relict species, Diplo-sphaerina inaequalis, Glomospira spp., Nodosinelloides ae-quiampla, and Rectostipulina quadrata, were absent in theInduan, but reoccurred in the Olenekian and becameabundant in the Middle Triassic in the Nanpanjiang Basin(Song et al., unpublished data). Consequently, they agree wellwith the concept of Lazarus taxa that usually ‘disappeared’ inthe mass extinction interval and ‘re-appeared’ in the post-extinction recovery stage (Jablonski, 1986).
Of these, the newcomers after the end-Permian massextinction in Nanpanjiang, Rectocornuspira kalhori is extraor-dinarily abundant throughout the microbialite unit in the
shallow platform setting of the Dajiang section. This speciesalso occurs in the microbialite unit of the deep-water faciesBianyang section, although it is represented by few specimens(Figs. 3, 4). Outside the Nanpanjiang, R. kalhori is alsoglobally present in the aftermath of the end-Permian massextinction, as mentioned above. Earlandia sp. occurs in themiddle parts of the calcimicrobial framestone at the Dajiangsection. The genus Earlandia is very common in the strataabove the P-Tr mass extinction horizon elsewhere in SouthChina (Ezaki et al., 2003, Song and Tong, in press), southernTurkey (Altiner et al., 2005; Groves et al., 2005; Angiolini etal., 2007), Italy (Groves et al., 2007), and Hungary (Hips andHaas, 2006).
To sum up, these two forms are widespread in rather highabundance in the aftermath of the P-Tr crisis and thus areecologically comparable with the so-called ‘‘disaster taxa’’proliferating in the aftermath of the end-Permian crisis,notably, stromatolites and other calcareous microbialites(Schubert and Bottjer, 1992; Lehrmann, 1999; Baud et al.,2007), bivalves Claraia and Promyalina (Boyer et al., 2004),the brachiopod ‘Lingula’ (Rodland and Bottjer, 2001; Chen etal., 2006), land flora Pleuromeia (Grauvogel-Stamm and Ash,2005), bryozoans Dyscritella, Paralioclema, and Pseudobato-stomella (Powers and Pachut, 2008). Accordingly, we agreewith the previous studies that treated both Earlandia andRectocornuspira, together with Cornuspira and Diplosphaerina,as the disaster taxa of foraminifers in the aftermath of the P-Trcrisis (Hallam and Wignall, 1997; Groves and Altiner, 2005;Groves et al., 2005, 2007).
DISCUSSION
Submarine dissolution event recorded in Dajiang.—In theGBG, the irregular contact between the SP and CF units hasattracted a continuing debate regarding its genesis, as itcoincides with the potential end-Permian mass extinctionhorizon (Krull et al., 2004). This SP-CF contact is character-ized by considerable relief and even overhanging topography(Payne et al., 2007, fig. 3B; Collin et al., 2009, fig. 3A–B). Liuet al. (2007) interpreted the irregular contact as a sub-aerialexposure, which correlates with the exposure surface corre-sponding to the base of Bed 24e at the Meishan section (Zhanget al., 1996; Yin et al., 2001) as the result of regional sea-levelfall across South China. These authors also considered thatthe sub-aerial erosion caused the omission of the strataequivalent to Beds 24e, 25, 26 and 27a–b of Meishan in theGBG.
In marked contrast to the sub-aerial erosion scenario (Liu etal., 2007), Payne et al. (2007) denied the effect of sea-levelchange and interpreted the truncation between the SP and CFunits as a submarine dissolution of carbonate caused bydecreased pH in surface water as a result of increaseddissolved carbon dioxide from volcanic eruption. They alsocorrelated this irregular contact with the minor erosionsurfaces recorded within Bed 27 in Meishan, South China(Cao and Shang, 1998) and erosion surfaces recorded in the P-Tr boundary beds in Turkey, and clarified that a global, or at
FIGURE 6—The extinction rates of four major foraminifer orders.
R
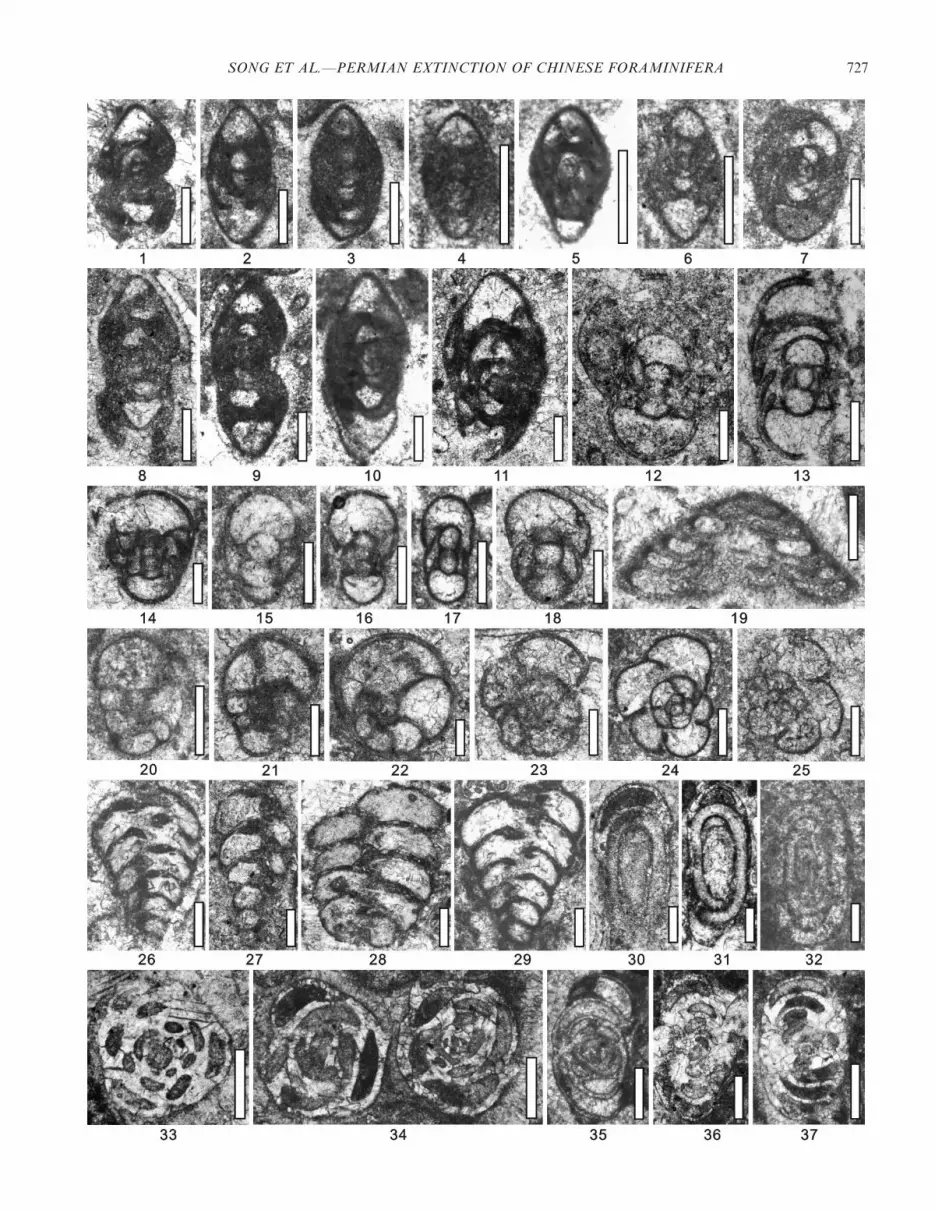
FIGURE 7—1–2, Ammodiscus sp.: 1, DJ-00006; 2, DJ-01380. 3–6, Hemigordius longus Grozdilova, 1956: 3, 00180; 4, DJ-00900; 5, DJ-00980; 6, BY-00001. 7–10, Rectocornuspira kalhori Bronnimann, Zanninetti, and Bozorgnia, 1972, DJ-10800. 11–13, Glomospira spp., DJ-00005. 14–15, Hemigordiussp., DJ-00260. 16–18, Sengoerina argandi Altiner, 1999: 16, DJ-00380; 17, DJ-00580; 18, DJ-00460. 19–22, Earlandia sp.: 19, DJ-10800; 20, DJ-10750; 21,22, Dj-10580. 23–28, Dagmarita chanakchiensis Reytlinger, 1965: 23, 24, 29, DJ-00380; 25, DJ-01160; 27, 28, DJ-00004. 29–35, Globivalvulina bulloides(Brady, 1876), 29, DJ-00520; 30, DJ-00004; 31–33, DJ-00580; 34, DJ-00540; 35, DJ-00360. 36, Tuberitina sp., DJ-00140. 37–43, Diplosphaerina inaequalis(Derville, 1931): 37, DJ-00640; 38, BY-00005; 39, DJ-00360; 40, DJ-00007; 41, DJ-01160; 42, BY-00005; 43, DJ-00380. Scale bar is 100 mm.
724 JOURNAL OF PALEONTOLOGY, V. 83, NO. 5, 2009

SONG ET AL.—PERMIAN EXTINCTION OF CHINESE FORAMINIFERA 725

least Tethys-wide, dissolution event occurred at the end of thePermian.
Based on detailed petrographic and sedimentologic studies,Collin et al. (2009) considered that the genesis mechanism ofthe irregular SP-CF contact is more complex than previouslyexpected, and these three interpretations: 1) sub-aerial physicalerosion coupled with dissolution by acid rains or carbondioxide-enriched mixed water, (2) sub-aerial erosion over-printed by dissolution related to acidified seawater, and 3)submarine dissolution caused by decrease in pH in seawater,cannot be excluded.
Apart from the features showing no sub-aerial erosion signsdocumented by Payne et al. (2007), our observation offoraminifers in thin sections throughout the SP-CF transitionalso show no sign of sub-aerial physical erosion. Contrastingmarkedly with the extraordinarily abundant foraminifer testsat the top of the SP unit (see also Collin et al., 2009, figs. 3F,4B), no foraminiferal test fragments and other reworkedsediments such as intraclasts are seen in thin sections from thebasal CF unit (see also Collin et al., 2009, fig. 4B). If sub-aerialphysical erosion generated the irregular contact surface, theeroded, fragmented foraminifer tests should be reworked and,perhaps bearing microbial envelopes, deposited in the micro-bialite unit, given the pre-erosion sediments rich in foraminifertests and post-erosion seawater rich in microbes. Thissedimentation phenomenon is very common in the Permianoceans (Flugel, 2004). Even if the eroded foraminifer testswere dissolved due to action of the acidified fresh water,brackish water, or seawater, some intraclasts are anticipatedto have been re-deposited after the sub-aerial physical erosionunless these eroded foraminifer tests and intraclasts have beencompletely washed away. However, it is not the case, given thevery short time gap and conspicuous reliefs and evenoverhanging topography (Payne et al., 2007; Collin et al.,2009). As a result, we agree with Payne et al. (2007) thatmassive submarine dissolution under low-energy hydrody-namic conditions may have generated the irregular contact inthe GBG. Nevertheless, we disagree with Payne et al. (2007)that the dissolution event created the minor submarine erosionsurfaces in Bed 27 at Meishan (Cao and Shang, 1998) becausewe detected considerably abundant foraminifer tests on bothsides of the erosion surfaces at the same bed (Song et al., 2007,table 1). Moreover, biogenetic burrows (i.e., Thalassinoidesichnosp.) are common near the erosion surface (Chen et al.,unpublished data). Thus, no dissolution or other chemicalpollution events occurred when Bed 27 was deposited atMeishan.
End-Permian mass extinction event.—Although foraminif-eral quantitative analysis calibrates the P-Tr extinctionhorizon to the SP-CF contact in the GBG, correlation of theextinction event with its counterpart recorded in Meishanremains disputed due to the new findings of conodont faunasfrom the SP unit. As outlined above, two species, H. parvusand H. eurypyge, are important in establishing age constraints
among the conodonts recovered from the topmost SP unit(Liu et al., 2007; Ezaki et al., 2008; Chen J. et al., 2009). Thefirst occurrence of H. parvus suggests that the topmost SP unitcorrelates with the middle part of Bed 27 of Meishan (Fig. 2).This stratigraphic position is clearly higher than the end-Permian mass extinction horizon (base of Bed 25) at Meishan(Jiang et al., 2007). The first occurrence of H. eurypygesuggests a correlation with Bed 27a of Meishan (Jiang et al.,2007), which is also higher than Bed 25 (Jiang et al., 2007).Based on this correlation, Payne et al. (2007) considered thatthe P-Tr extinction associated with the dissolution event in theGBG was younger than the P-Tr crisis recorded in Meishan(Jin et al., 2000). Our study shows that Palaeofusulina sinensisis present just below the dissolution line and some tests havebeen partly dissolved (see Collin et al., 2009, fig. 3B). Lastoccurrence of P. sinensis never reaches Bed 25 of Meishan andits counterparts in South China, according to the past three-decade studies of more than 50 P-Tr boundary sections (Zhaoet al., 1981; Sheng et al., 1984; Yang et al., 1987; Song et al.,2006, 2007). This means that the topmost SP unit cannot beyounger than Bed 24e of Meishan, based on foraminiferalcorrelation. Thus, a discrepancy exists between foraminiferand conodont biostratigraphic correlations. Liu et al. (2007)considered that H. parvus specimens may be derived from theoverlying CF unit and, reworked, diffused through the contactsurface. Chen J. et al. (2009) agree partly with thisinterpretation and also assume that the source of the conodontspecimens at the top of the SP unit may be the undisolvedresidues on the contact surface. This means that a submarinedissolution event took place after the strata equivalent to Beds25–27a of Meishan were deposited. Given such a shortduration (150,000 yr) that Beds 25–28 represent at Meishan(Chen J. et al., 2009, fig. 5), the dissolution of the unlithifiedsediments equivalent to Beds 25–27a is not impossible. Incontrast, a correlation of carbon isotopic excursions betweenthe GBG and Meishan suggests that the end-Permian massextinction coincides with the SP-CF contact (Krull et al., 2004;Chen J. et al., 2009), which is also confirmed by the presentforaminiferal quantitative analysis. As a consequence, weconsider that the topmost SP unit is coeval with Bed 24e ofMeishan. Conodont H. parvus occurred slightly earlier in theGBG than in Meishan and elsewhere in the world. Thesubmarine dissolution event may be coeval with the end-Permian mass extinction in the GBG, but the possibility thatsubmarine dissolution occurred after the end-Permian massextinction cannot be excluded.
Contrasting markedly with the monotone-stage episodicextinction pattern documented here, foraminifers appear in atwo-stage extinction pattern in the Meishan section (Song etal., 2009). The disparency is probably due to two reasons.Firstly, the second extinction event occurred between theconodonts H. parvus and Isarcicella staeschei Zones atMeishan, which corresponds to the top of the CF unit(Fig. 2). The entire CF unit represents an ecologically special
R
FIGURE 8—1, Neoendothyra sp., DJ-00580. 2–7, Neoendothyra reicheli Reytlinger, 1965: 2, 4, DJ-00600; 3, DJ-00440; 5, DJ-00560; 6–7, BY-00002. 8–11, Neoendothyra permica (Lin, 1978): 8, DJ-00440; 9, DJ-00400; 10, DJ-00560; 11, DJ-00140. 12–14, Postendothyra scabra Lin, 1984: 12, DJ-01160; 13,DJ-00560; 14, DJ-00500. 15–18, 23, Postendothyra guangxiensis (Lin, 1978): 15, DJ-00003; 16, DJ-00600; 17, DJ-00580; 18, 23, DJ-00480. 19, Tetrataxisplanolocula Lee and Chen, 1930, DJ-00440. 20–22, Globivalvulina vonderschmitti Reichel, 1945: 20, DJ-01300; 21, DJ-00009; 22, DJ-00004. 24, 25,Postendothyra tenuis Lin, Li and Sun, 1990: 24, DJ-00480; 25, DJ-00980. 26–29, Palaeotextularia spp.: 26, DJ-00340; 27, DJ-00007; 28, DJ-00580; 29, DJ-00380. Scale bar is 200 mm. 30–32, Agathammina pusilla (Geinitz, 1848): 30, DJ-01380; 31, DJ-00340; 32, DJ-01580. 33–34, Glomomidiella nestellorumVachard, Rettori, Angiolini and Checconi, 2008: 33, DJ-00240; 34, DJ-01560. 35–37, Neodiscus plectogyraeformis Lin, Li and Sun, 1990: 35, DJ-01600;36, DJ-00360; 37, DJ-00540. Scale bar is 200 mm.
726 JOURNAL OF PALEONTOLOGY, V. 83, NO. 5, 2009
SONG ET AL.—PERMIAN EXTINCTION OF CHINESE FORAMINIFERA 727

habitat, which was hospitable for microbial assemblages toproliferate (Ezaki et al., 2008) but was bad for foraminifers tolive, evidenced by the presence of few species in the CF unit(Fig. 3). Thus, the foraminifer data from the CF unit isinadequate to demonstrate an extinction pattern. Secondly,other fossils groups such as ostracods, gastropods, brachio-pods, and bivalves are commonly present in the thin sectionsof thin-bedded bioclastic packstone interbedded in thecalcimicrobial framestone (Payne et al., 2006a). Their taxon-omy however remains unstudied and thus obscures therecognition of the second episode of the P-Tr crisis.
In general, the GBG collections show that the calcareousforaminifers (,97%) governed the Late Permian assemblages,but they suffered an extinction rate of ,95% (56/59) incontrast to an extinction rate of 50% among the agglutinatedtaxa. Moreover, all of the large and architecturally compli-cated elements, mainly fusulinoidean foraminifers, were wipedout in the end-Permian extinction event. The survivors wereusually small and simple forms. Thus, various foraminifergroups experienced selective extinction. This phenomenon isinterpreted as the physiological reactions of foraminifers toseveral defaunation events such as hypercapnia, elevated CO2
content, and global warming that prevailed in the P-Trtransition (Knoll et al., 2007) and massive submarinedissolution event recognized from the GBG (Payne et al.,2007). These devastating events usually resulted in mortalvulnerability to marine ecosystems where organisms arecharacterized by a low basal metabolic rate, limited or nocirculatory system, little elaboration of respiratory surfaces,and precipitation of CaCO3 skeletons under conditions ofminimal physiological buffering (Knoll et al., 2007).
CONCLUSIONS
Latest Permian foraminifers from the Dajiang and Bia-nyang sections of the Great Bank of Guizhou are considerablyabundant and diverse, including 58 species in 37 genera. Theyare assigned to the Palaeofusulina sinensis Zone, the youngestPermian foraminifer zone in South China. Quantitativeanalysis on the stratigraphic distributions of 61 foraminiferspecies shows that the last occurrences of more than half thespecies (28/54) fall into a 60-cm-interval at the topmostskeletal packstone unit and thus calibrates the end-Permianextinction to the skeletal packstone-calcimicrobial framestoneboundary. Most of the Late Permian foraminifers (,93%)became extinct in the P-Tr crisis, various foraminiferal groupssuffering selective extinction in the event. The Miliolida,Fusulinida and Lagenida have been mortally affected with anextinction rate of 100%, 96% and 92%, respectively, while theTextulariina suffered an extinction rate of 50%. Two species,(Hemigordius longus and ?Globivalvulina bulloides, survived thecrisis into the earliest Triassic but disappeared forever soonafter. Four Lazarus species, Diplosphaerina inaequalis, Glo-mospira spp., Nodosinelloides aequiampla, and Rectostipulinaquadrata, and two disaster taxa, Rectocornuspira kalhori andEarlandia sp., characterize the post-extinction foraminiferassemblage. Foraminiferal distributions near the P-Tr bound-
ary also reveal that submarine dissolution could be the favoredcause generating the irregular contact surface associated withthe end-Permian mass extinction horizon. The submarinedissolution event may be coeval with the end-Permian massextinction.
SYSTEMATIC PALEONTOLOGY
Systematic classification of foraminifers has long been asubject of conjecture, although multiple schemes have beenrecently described (Loeblich and Tappan, 1988; Mikhalevich,2000; Armstrong and Brasier, 2005; Groves et al., 2005). Here,the classification of the order Lagenida follows Groves et al.(2005), and that of other orders follow Loeblich and Tappan(1988). Most foraminifer species are illustrated herein(Figs. 7–11), but only new taxa and the poorly known speciesare described below. All described specimens are housed in themuseum of China University of Geosciences, Wuhan, China.
Class FORAMINIFERA Eichwald, 1830Order FUSULINIDA Wedekind, 1937
Family BRADYINIDAE Reytlinger, 1950Genus NEOENDOTHYRA Reytlinger, 1965
NEOENDOTHYRA REICHELI Reytlinger, 1965Figure 8.2–8.7
Neoendothyra reicheli REYTLINGER, 1965, p. 61–62, pl. 1, figs.6–9; BOZORGNIA, 1973, p. 93–94, pl. 35, figs. 2, 4–9, pl. 39,figs. 3, 4; LYS ET AL., 1978, pl. 7, fig. 12; LYS, COLCHEN,BASSOULET, MARCOUX, AND MASCLE, 1980, p. 93, pl. 6,fig. 1; KOBAYASHI, 1986, p. 140, pl. 3, figs. 4, 5, 10; UENO,1992, fig. 9.23; PINARD AND MAMET, 1998, p. 75, pl. 35, fig.13; VACHARD, HAUSER, MARTINI, ZANINETTI, MATTER,AND PETERS, 2002, pl. 1, figs. 3–6; GAILLOT AND VACHARD,2007, pl. 20, fig. 8.
Endothyra sp. SIDO, 1978, pl. 12, figs. 3, 4.Neoendothyra consueta SOSNINA, 1978, p. 29, pl. 1, fig. 22.?Neoendothyra reicheli VACHARD, 1981, pl. 29, fig. 1.Neoendothyra permica KOBAYASHI, 1997, pl. 4, fig. 28; ZHANG
AND HONG, 2000, p. 45, pl. 1, figs. 27, 28; ZHANG AND
HONG, 2004, p. 70, pl. 1, figs. 32, 33; WANG, 2002, p. 118,pl. 2, fig. 5.
Neoendothyra sp. MERTMANN, 2000, fig. 7.7.Neoendothyra parva KOZUR, AYDIN, DEMIR, YAKAR, GON-
CUOGLU, AND KURU, 2000, pl. 2, fig. 10.
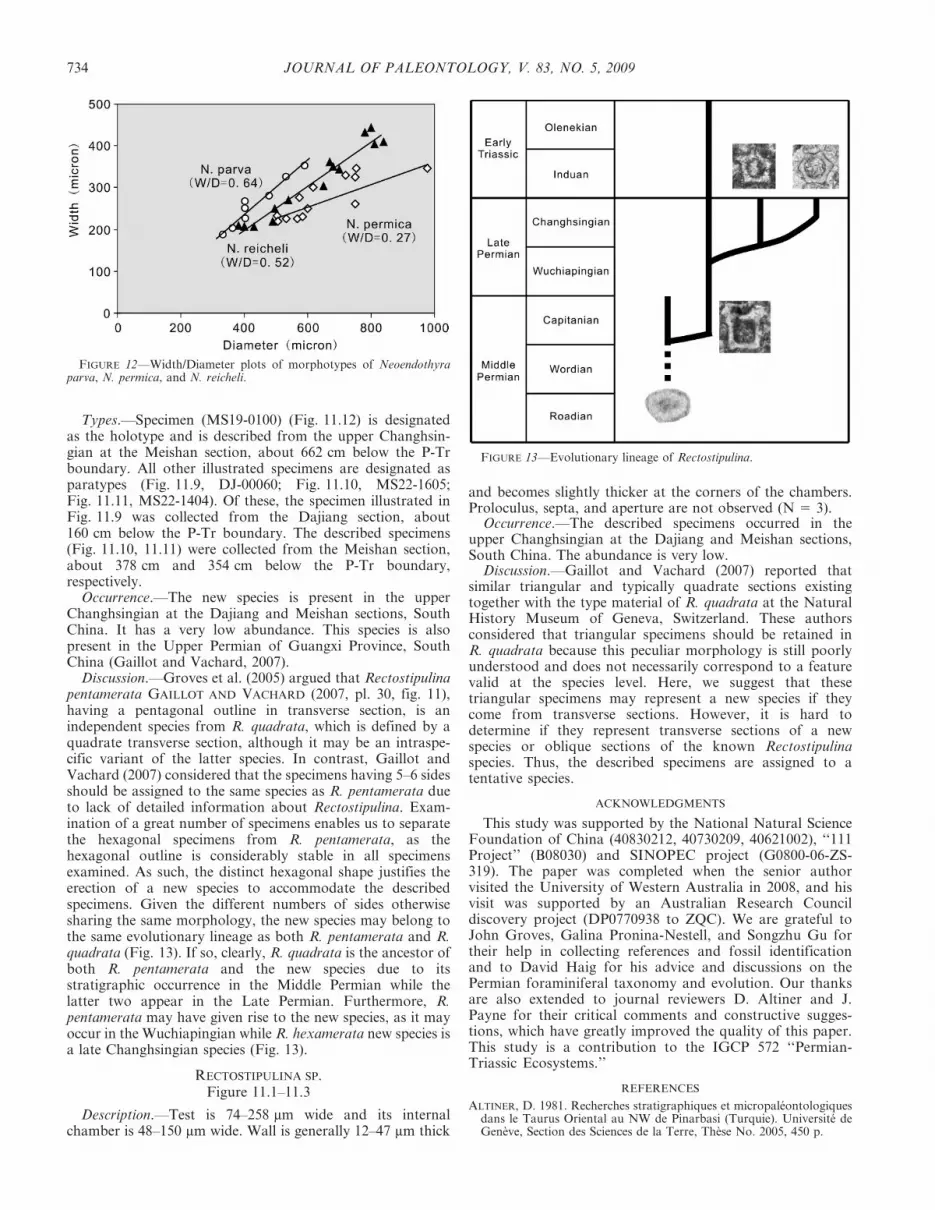
Description.—Tests with 2–2K whorls 148–265 mm in widthand 264–515 mm in diameter. The width/diameter (W/D) ratiois 0.48–0.56. Wall thickness in last whorl is 10–16 mm.Secondary deposits are well developed. (N 5 4)
Occurrence.—Neoendothyra reicheli was first reported fromthe Lower Triassic (Induan) of the Transcaucasus. Later, thisspecies was documented from the Lower to Upper Permianelsewhere in the world such as Iran (Bozorgnia, 1973; Lys etal., 1978), eastern Oman (Vachard et al., 2002), Pakistan(Mertmann, 2000), north Vietnam (Liem, 1971), the Hima-layas (Lys et al., 1980), South China (Zhang and Hong, 2000;Wang, 2002; Zhang and Hong, 2004), Japan (Kobayashi,1986, 1997; Ueno, 1992), south Primorye (Sosnina, 1978),
R
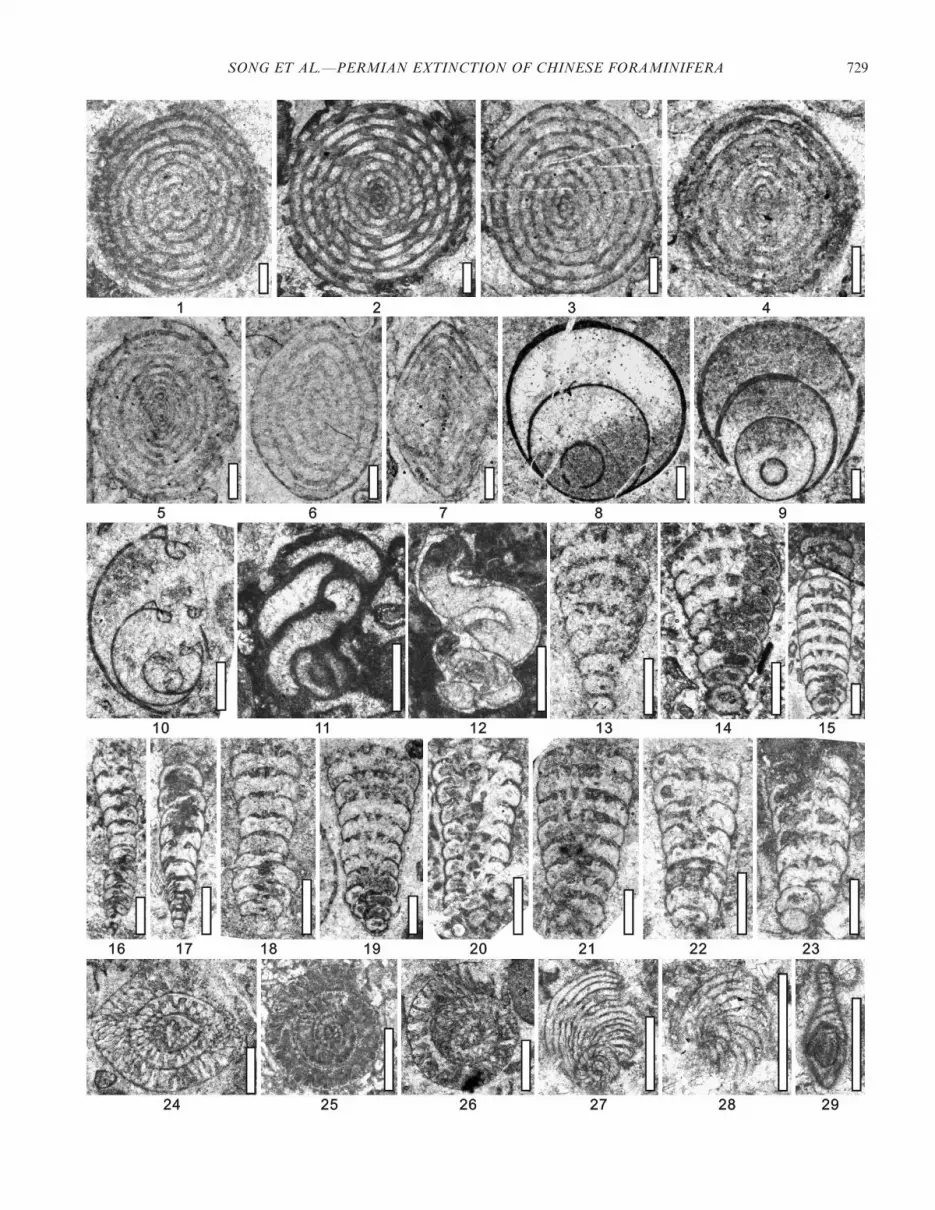
FIGURE 9—1–5, Pisolina excessa Lee, 1934: 1,4, DJ-00220; 2, DJ-01500; 3, 5, DJ-00240. 6–7, Nankinella minor Sheng, 1956: 6, DJ-00640; 7, DJ-00240.8–10, Paraglobivalvulina piyasini Sakagami and Hatta, 1982: 8, DJ-00160; 9, DJ-00004; 10, DJ-00260. 11–12, ?Calcitornella sp.: 11, DJ-01480; 12, DJ-01320.13–15, Cribrogenerina spp.: 13, DJ-01440; 14, DJ-00580; 15, DJ-00460. 16–18, Deckerella spp.: 16, 17, DJ-00004; 18, DJ-00140. 19–20, Climacamminavalvulinoides Lange, 1925: 19, DJ-00640; 20, DJ-00004. 21–23, Climacammina moelleri Reitlinger, 1950: 21, DJ-00160; 22, DJ-01300; 23, DJ-01220. 24–26,Palaeofusulina sinensis, Sheng, 1955: 24, 26, DJ-00180; 25, BY-00001. 27–29, Reichelina pulchra Miklukho-Maklay, 1954: 27, DJ-00560; 28, DJ-00460; 29, DJ-00440. Scale bar is 500 mm.
728 JOURNAL OF PALEONTOLOGY, V. 83, NO. 5, 2009
SONG ET AL.—PERMIAN EXTINCTION OF CHINESE FORAMINIFERA 729

Afghanistan (Vachard, 1981), Turkey (Kozur et al., 2000;Altiner and Ozkan-Altiner, 2001; Turhan et al., 2004; Gaillotand Vachard, 2007), western Hungary (Sido, 1978), andCanada (Pinard and Mamet, 1998). In Nanpanjiang, N.reicheli occurs at the upper Changhsingian at both the Dajiangand Bianyang sections, southern Guizhou.
Discussion.—This species is similar to Neoendothyra parvain having a small body size and simple test construction, but ithas a W/D of about 0.64 (Fig. 12) and thus is more flattenedin test shape than N. parva.
NEOENDOTHYRA PERMICA (Lin, 1978)Figure 8.8–8.11
Neoendothyra sp. ISHII, OKIMURA, AND NAKAZAWA, 1975, pl.3, fig. 13; KOTLYAR, ZAKHAROV, KOCZYRKEVICZ, KRO-
PATCHEVA, ROSTOVTSEV, CHEDIJA, AND VUKS, 1983, pl. 1,fig. 5, pl. 7, figs. 3, 4; UENO AND SAKAGAMI, 1993, figs. 3.2–3.3.
Endothyra permica LIN, 1978, p. 31, pl. 6, figs. 4, 5.Neoendothyra permica LIN, 1984, p. 128, pl. 4, figs. 1–2; LIN,
LI, AND SUN, 1990, p. 187, pl. 17, figs. 23, 25, 28–31;KOBAYASHI, 1997, pl.4, figs. 25–27, 29; ZHANG AND HONG,2000, p. 45, pl. 1, figs. 27, 28; ZHANG AND HONG, 2004,p. 70, pl. 1, fig. 31.
Neoendothyra miriformis LIN, 1985, p. 291, pl. 1, figs. 25–27.Frondina permica JENNY, IZART, BAUD, AND JENNY, 2004, pl.
3, fig. 19.Neoendothyra reicheli GAILLOT AND VACHARD, 2007, pl. 20,
fig. 8.
Description.—Tests embrace three whorls and are 323–396 mm in width and 745–854 mm in diameter. The width/diameter (W/D) ratio is about 0.40–0.47. Wall thickness in lastwhorl is about 20–30 mm. Outer diameter of proloculus isabout 40 mm. (N 5 4)
Occurrence.—This species occurs in the Lower-UpperPermian around the world, for instance, northeast Thailand(Ueno and Sakagami, 1993), South China (Lin, 1978, 1984,1985; Lin et al., 1990; Zhang and Hong, 2000, 2004), Japan(Ishii et al., 1975; Kobayashi, 1997). Russia (Kotlyar et al.,1983), and Greece (Jenny et al., 2004). Neoendothyrapermica occurs in the Upper Changhsingian of the Dajiangsection.
Discussion.—Lin (1985) established Neoendothyra mirifor-mis based on foraminifer tests obtained from the LowerPermian Qixia Formation of the Jiahe area, GuangdongProvince, South China. However, the illustrations of thespecies given by Lin (1985, fig. 25–27) are the obliquesections of foraminifer tests. Similar features are also presentin our thin sections. Like Neoendothyra permica, Lin’sspecies possesses three whorls, 300–360 mm in width and540–640 mm in diameter. Thus, all of the illustratedcharacters and description given by Lin (1985, p. 291) justifythe present treatment of N. miriformis as a synonym of N.permica.
NEOENDOTHYRA SP.Figure 8.1
Description.—Test comprises three whorls and is 295 mm inwidth and 510 mm in diameter. The width/diameter (W/D)ratio is about 0.58. Wall thickness in last whorl is about19 mm.
Occurrence and discussion.—Only one specimen from theupper Changhsingian of the Dajiang section is available forstudy. The described specimen agrees well with the concept ofNeoendothyra. When compared with the known species of thegenus, the Dajiang specimen possesses a W/D ratio of about0.58, which is transitional between the W/D ratio of N. reicheliand N. parva. In addition, its umbilicus is significantlynarrower than any other known species within Neoendothyra.Thus, this Dajiang specimen may represent a new species,which however cannot be proposed here due to inadequatematerial.
Genus POSTENDOTHYRA Lin, 1984POSTENDOTHYRA GUANGXIENSIS (Lin, 1978)
Figure 8.15–8.18, 8.23
Endothyranopsis guangxiensis LIN, 1978, p. 34, pl. 7, figs. 1–3;LI AND DING, 1987, pl. 1, fig. 5.
Endothyranopsis guangxiensis obesus LIN, 1978, p. 34, pl. 7,figs. 4, 5.
Bradyina guangxiensis WANG IN ZHAO ET AL., 1981, pl. 1, figs.13–15; PRONINA, 1988, pl. 2, figs. 2, 3.
Postendothyra strena LIN, 1984, p. 137–138, pl. 6, figs. 6–7.Bradyina sp. KOTLYAR, ZAKHAROV, KROPATCHEVA, PRO-
NINA, CHEDIJA, AND BURAGO, 1989, pl. 1, fig. 6; WANG,2002, p. 119, pl. 2, fig. 22.
Postendothyra guangxiensis LIN, LI, AND SUN, 1990, p. 191,pl. 18, figs. 21–26; PRONINA- NESTELL AND NESTELL, 2001,pl. 4, fig. 15; SONG, TONG, AND HE, 2006, p. 94. pl. 2, fig. 1;KOBAYASHI, 2006, p. 183, figs. 4.21–4.24, 4.30, 4.34.
Postendothyra obesa LIN, LI, AND SUN, 1990, p. 191, pl. 19,figs. 1–3.
Description.—Tests have two whorls and are 174–325 mm inwidth and 343–545 mm in diameter. Outer diameter ofproloculus is 62–103 mm. Each whorl has 4–5 chambers. Wallis 15–30 mm thick, becoming slightly thicker in the umbilicus.(N 5 4)
Occurrence.—This species is the most common representa-tive of Postendothyra at the studied sections. Other docu-mented occurrences include the Middle and Upper Permianelsewhere in South China (Lin, 1978, 1984; Li and Ding, 1987;Lin et al., 1990; Wang, 2002; Song et al., 2006), the UpperPermian in the Himalayas and Russia (Lys et al., 1980;Pronina, 1988; Kotlyar et al., 1989), and the Middle Permianin Central Japan (Kobayashi, 2006).
Discussion.—Having small tests, the same number of whorlsand similar outlines to Postendothyra guangxiensis, both P.sterna Lin, 1984 and P. obesa Lin, Li and Sun, 1990 are treatedas junior synonyms of the latter species. Postendothyra
R
FIGURE 10—1–7, Reichelina cribroseptata Erk, 1942: 1, DJ-00520; 2, DJ-00460; 3, DJ-00360; 4, DJ-00002; 5, DJ-00560; 6, DJ-00440; 7, DJ-00420. 8–10, 13, Colaniella nana Miklukho-Maklay, 1954: 8, DJ-01600; 9, DJ-01580; 10, DJ-00720; 13, DJ-01500. 11–12, Parareichelina sp.: 11, DJ-00540; 12, DJ-00640. 14–18, Colaniella parva (Colani, 1924): 14, DJ-01480; 15–16, DJ-01560; 17, DJ-01400; 18, DJ-01380. 19–21, Colaniella cylindrical Miklukho-Maklay, 1954: 19, DJ-01580; 20, DJ-01380; 21, DJ-00960. 22, Howchinella costulata Karavaeva and Nestell, 2007, DJ-00360. 23–25, Calvezina ottomanaSellier de Civrieux and Dessauvagie, 1965: 23, DJ-00600; 24, DJ-01380; 25, DJ-00007. 26–27, Pachyphloia schwageri Sellier de Civrieux and Dessauvagie,1965: 26, BY-00001; 27, DJ-00580. 28–30, Pachyphloia ovata Lange, 1925: 28, DJ-01500; 29, DJ-00560. 31–32, Pachyphloia robusta Miklukho-Maklay,1954: 31, DJ-00004; 32, DJ-00002. 33–35, Robuloides lens Reichel, 1946: 33, DJ-01480; 34, DJ-01500; 35, DJ-01400. Scale bar is 200 mm.
730 JOURNAL OF PALEONTOLOGY, V. 83, NO. 5, 2009
SONG ET AL.—PERMIAN EXTINCTION OF CHINESE FORAMINIFERA 731

guangxiensis is distinguished from P. scabra Lin, 1984 andother species in the genus by its smaller test and fewer whorls.
POSTENDOTHYRA SCABRA Lin, 1984Figure 8.12–8.14
Postendothyra scabra LIN, 1984, p. 137, pl. 6, figs. 1, 2.Postendothyra sinensis LIN, LI, AND SUN, 1990, p. 191–192, pl.
18, figs. 27–33.Postendothyra sp. KOBAYASHI, 1997, pl. 5, figs. 5, 6.
Description.—Tests have 2K–3 whorls and are 458–612 mmwide and 494–748 mm in diameter. Outer diameter ofproloculus is 65–78 mm. Each whorl has 4–5 chambers.Chambers increase rapidly in diameter from inner to outer.Wall is 20–40 mm thick and increases in thickness in theumbilicus. (N 5 3)
Occurrence.—This species occurs in the upper Changhsin-gian at Dajiang section. It has also been reported from theUpper Permian in Guangxi Province (Lin et al., 1990) and theMiddle Permian of the Xintan section, Hubei Province (Lin,1984), South China and the Upper Permian of the westernTokyo, Japan (Kobayashi, 1997).
Discussion.—Lin et al. (1990) differentiated Postendothyrasinensis (Lin et al., 1990, pl. 18, figs. 27–33) from P. scabra(Lin, 1984, pl. 6, figs. 1, 2) by having loose whorls and looselyarranged chambers. However, after inspecting a great numberof similar specimens, we found that the loose whorls andchambers are oblique views obtained when the thin sectionswere cut in various angles. Otherwise, both species share thesame morphology such as the same number of whorls andsimilar body size diagnostic of the genus. These charactersshared between the two justify the treatment of P. sinensis Lin,Li and Sun, 1990 as a junior synonym of P. scabra Lin, 1984,although the former species was described from the Wuchia-pingian and Changhsingian in Guangxi Province and the typespecimens of P. scabra are from the Middle Permian of HubeiProvince.
POSTENDOTHYRA TENUIS Lin, Li and Sun, 1990Figure 8.24, 8.25
Postendothyra tenuis LIN, LI, AND SUN, 1990, p. 192, pl. 19,figs. 4–8; KOBAYASHI, 1997, pl. 5, figs. 1–4; KOBAYASHI,2004, figs. 6.27–6.31; KOBAYASHI, 2006, figs. 4.35–4.38.
Description.—Tests possess 2–2K whorls and are 492–524 mm in diameter. Proloculus is 58 mm in outer diameter.Each whorl has five chambers. Wall is 10–15 mm thick. (N 52)
Occurrence.—This species is described from the upperChanghsingian at the Dajiang section. It also occurs in theUpper Permian in Guangxi, South China (Lin et al., 1990), theMiddle Permian of central Japan (Kobayashi, 2006) and theUpper Permian of west Shikoku and west Tokyo, Japan(Kobayashi, 1997, 2004).
Discussion.—The present species resembles remarkedlyPostendothyra guangxiensis described above in having a smallbody size and simple test construction. However, it has arather thin wall and thus cannot be confused with P.guangxiensis. The relatively thin wall also distinguishes thedescribed specimens from any other Postendothyra species.
Order LAGENIDA Delage and Herouard, 1896IIA4. PALYGONAL Chambers
Genus RECTOSTIPULINA Jenny-Deshusses, 1985
Type species.—Rectostipulina quadrata Jenny-Deshusses,1985
Diagnosis.—Test consists of one or more chambers varyingfrom elongate, columnar, to uniserial outlines. Septa arerather short and nearly perpendicular to the test wall.Chambers are polygonal in transverse section. Wall is atelo-monolamellar. Aperture is a simple opening in the center ofthe apertural face.
Occurrence and discussion.—Rectostipulina comprises threespecies at present, including R. quadrata Jenny-Deshusses,1985, R. pentamerata Groves, Altiner, and Rettori, 2005, andR. syzranaeformis Gaillot and Vachard, 2007. Rectostipulinaquadrata occurs in the upper Middle Permian throughuppermost Permian in Turkey (Lys and Marcoux, 1978;Altiner, 1981; Zaninetti et al., 1981; Koyluoglu and Altiner,1989; Leven and Okay, 1996; Groves et al., 2005), the PersianGulf (Gaillot and Vachard, 2007), northern Italy (Cirilli et al.,1998; Groves et al., 2007), Iran (Jenny-Deshusses, 1985),Greece (Jenny et al., 2004), northeastern Cyprus (Nestell andPronina, 1997), other areas of South China (Song et al., 2007,2009), India (Lys et al., 1980), west Thailand (Tien, 1988), andnorth New Zealand (Vachard and Ferriere, 1991). Rectosti-pulina pentamerata reportedly occurs in the Upper Permian ofTurkey (Zaninetti et al., 1981; Koyluoglu and Altiner, 1989;Groves et al., 2005), northern Italy (Cirilli et al., 1998; Groveset al., 2007), the Persian Gulf (Gaillot and Vachard, 2007),South China (Gaillot and Vachard, 2007; Song et al., 2009),and Cambodia (Tien, 1986). Rectostipulina syzranaeformis hasbeen reported from the upper Midian in Turkey (Gaillot andVachard, 2007). Here, we described one new species belowfrom the Upper Permian in the Nanpanjiang Basin.
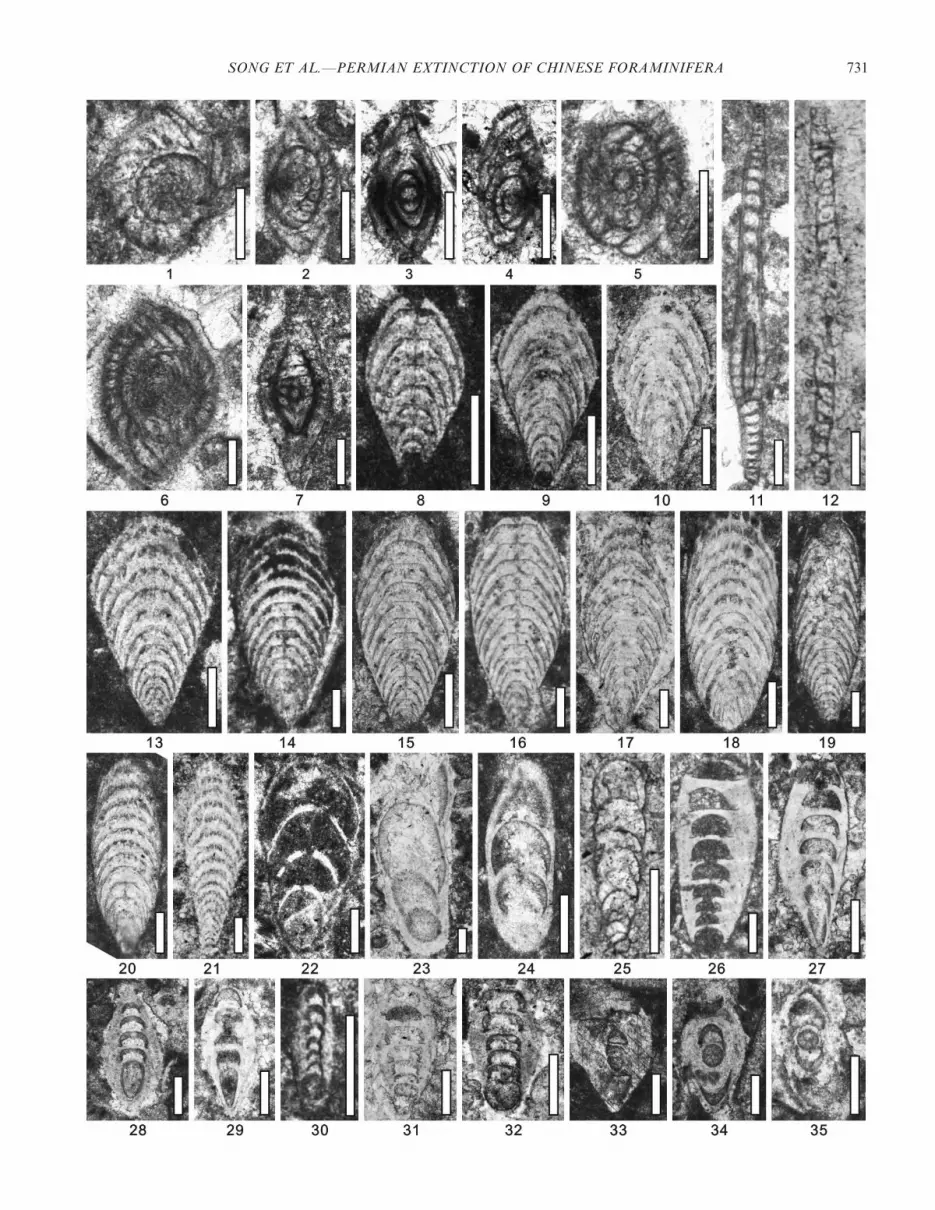
RECTOSTIPULINA HEXAMERATA Song and Tong new speciesFigure 11.9–11.12
Diagnosis.—Chambers are hexagonal in transverse section.Description.—Test is 121–252 mm wide, with the internal
chamber being 71–143 mm wide. Wall thickness varies from12 mm to 38 mm and the wall becomes slightly thicker at thecorners of the chamber. Proloculus, septa, and aperture notobserved. (N 5 4)
Etymology.—From the Greek hexamerus, meaning consist-ing of six parts, in reference to the six-sided columnarchambers, characteristic of the new species.
R
FIGURE 11—1–3, Rectostipulina sp.: 1, MS20-0100; 2, MS22-1201; 3, DJ-00540. 4, 5, Rectostipulina quadrata Jenny-Deshusses, 1985: 4, DJ-00500; 5,DJ-00006. 6–8, Rectostipulina pentamerata Groves, Altiner, and Rettori, 2005: 6, DJ-00600; 7, 8, DJ-00004. 9–12, Rectostipulina hexamerata n. sp.: 9,DJ-00060; 10, MS22-1605; 11, MS22-1404; 12, MS19-0100. 13–16, Nodosinelloides aequiampla (Zolotova in Zolotova and Baryshnikov, 1980): 13, DJ-00820; 14, DJ-00560; 15, BY-00003; 16, BY-00001. 17–21, Nodosinelloides sagitta (Miklukho-Maklay, 1954): 17, DJ-00560; 18, 20, DJ-01360; 19, DJ-00004; 21, DJ-01480. 22–23, Nodosinelloides camerata (Miklukho-Maklay, 1954): 22, DJ-00520; 23, DJ-00002. 24, Protonodosaria sp.: DJ-00140. 25–27,Nodosaria expolita Trifonova, 1978: 25, 26, DJ-1185; 27, DJ-11425. 28–29, Geinitzina uralica Suliemanov, 1949: 28, DJ-00006; 29, DJ-00004. 30–32,Geinitzina spandeli Cherdyntsev, 1914: 30, DJ-00004; 31, DJ-00009; 32, DJ-00560. 33–36, Ichthyofrondina palmata (Wang, 1974): 33, DJ-00600; 34, 35,DJ-00580; 36, DJ-00008. 37–39, Frondina permica Sellier de Civrieux and Dessauvagie, 1965: 37, DJ-01600; 38, DJ-00520; 39, DJ-00009. 40–41,Calvezina sp.: 40, DJ-00540; 41, DJ-00560. 42–43, Partisania typica Sosnina, 1978: 42, DJ-00580; 43, DJ-01480. Scale bar is 100 mm.
732 JOURNAL OF PALEONTOLOGY, V. 83, NO. 5, 2009

SONG ET AL.—PERMIAN EXTINCTION OF CHINESE FORAMINIFERA 733
Types.—Specimen (MS19-0100) (Fig. 11.12) is designatedas the holotype and is described from the upper Changhsin-gian at the Meishan section, about 662 cm below the P-Trboundary. All other illustrated specimens are designated asparatypes (Fig. 11.9, DJ-00060; Fig. 11.10, MS22-1605;Fig. 11.11, MS22-1404). Of these, the specimen illustrated inFig. 11.9 was collected from the Dajiang section, about160 cm below the P-Tr boundary. The described specimens(Fig. 11.10, 11.11) were collected from the Meishan section,about 378 cm and 354 cm below the P-Tr boundary,respectively.
Occurrence.—The new species is present in the upperChanghsingian at the Dajiang and Meishan sections, SouthChina. It has a very low abundance. This species is alsopresent in the Upper Permian of Guangxi Province, SouthChina (Gaillot and Vachard, 2007).
Discussion.—Groves et al. (2005) argued that Rectostipulinapentamerata GAILLOT AND VACHARD (2007, pl. 30, fig. 11),having a pentagonal outline in transverse section, is anindependent species from R. quadrata, which is defined by aquadrate transverse section, although it may be an intraspe-cific variant of the latter species. In contrast, Gaillot andVachard (2007) considered that the specimens having 5–6 sidesshould be assigned to the same species as R. pentamerata dueto lack of detailed information about Rectostipulina. Exam-ination of a great number of specimens enables us to separatethe hexagonal specimens from R. pentamerata, as thehexagonal outline is considerably stable in all specimensexamined. As such, the distinct hexagonal shape justifies theerection of a new species to accommodate the describedspecimens. Given the different numbers of sides otherwisesharing the same morphology, the new species may belong tothe same evolutionary lineage as both R. pentamerata and R.quadrata (Fig. 13). If so, clearly, R. quadrata is the ancestor ofboth R. pentamerata and the new species due to itsstratigraphic occurrence in the Middle Permian while thelatter two appear in the Late Permian. Furthermore, R.pentamerata may have given rise to the new species, as it mayoccur in the Wuchiapingian while R. hexamerata new species isa late Changhsingian species (Fig. 13).
RECTOSTIPULINA SP.Figure 11.1–11.3
Description.—Test is 74–258 mm wide and its internalchamber is 48–150 mm wide. Wall is generally 12–47 mm thick
and becomes slightly thicker at the corners of the chambers.Proloculus, septa, and aperture are not observed (N 5 3).
Occurrence.—The described specimens occurred in theupper Changhsingian at the Dajiang and Meishan sections,South China. The abundance is very low.
Discussion.—Gaillot and Vachard (2007) reported thatsimilar triangular and typically quadrate sections existingtogether with the type material of R. quadrata at the NaturalHistory Museum of Geneva, Switzerland. These authorsconsidered that triangular specimens should be retained inR. quadrata because this peculiar morphology is still poorlyunderstood and does not necessarily correspond to a featurevalid at the species level. Here, we suggest that thesetriangular specimens may represent a new species if theycome from transverse sections. However, it is hard todetermine if they represent transverse sections of a newspecies or oblique sections of the known Rectostipulinaspecies. Thus, the described specimens are assigned to atentative species.
ACKNOWLEDGMENTS
This study was supported by the National Natural ScienceFoundation of China (40830212, 40730209, 40621002), ‘‘111Project’’ (B08030) and SINOPEC project (G0800-06-ZS-319). The paper was completed when the senior authorvisited the University of Western Australia in 2008, and hisvisit was supported by an Australian Research Councildiscovery project (DP0770938 to ZQC). We are grateful toJohn Groves, Galina Pronina-Nestell, and Songzhu Gu fortheir help in collecting references and fossil identificationand to David Haig for his advice and discussions on thePermian foraminiferal taxonomy and evolution. Our thanksare also extended to journal reviewers D. Altiner and J.Payne for their critical comments and constructive sugges-tions, which have greatly improved the quality of this paper.This study is a contribution to the IGCP 572 ‘‘Permian-Triassic Ecosystems.’’
REFERENCES
ALTINER, D. 1981. Recherches stratigraphiques et micropaleontologiquesdans le Taurus Oriental au NW de Pinarbasi (Turquie). Universite deGeneve, Section des Sciences de la Terre, These No. 2005, 450 p.
FIGURE 12—Width/Diameter plots of morphotypes of Neoendothyraparva, N. permica, and N. reicheli.
FIGURE 13—Evolutionary lineage of Rectostipulina.
734 JOURNAL OF PALEONTOLOGY, V. 83, NO. 5, 2009

ALTINER, D. 1999. Sengoerina argandi n. gen. n. sp., and its position in theevolution of Late Permian biseriamminid foraminifers. Micropaleon-tology, 45:215–220.
ALTINER, D., J. R. GROVES, AND S. OZKAN-ALTINER. 2005. Calcareousforaminiferal recovery from the end-Permian mass extinction, southernTurkey. Albertiana, 33:14–17.
ALTINER, D. AND S. OZKAN-ALTINER. 2001. Charliella rossae n. gen. n.sp., from the Tethyan realm: remarks on the evolution of Late Permianbiseriamminids. Journal of Foraminiferal Research, 31:309–314.
ALTINER, D., R. SAVINI, AND S. OZKAN-ALTINER. 2003. Morphologicalvariation in Hemigordius harltoni Cushman and Waters, 1928: Remarkson the taxonomy of Carboniferous and Permian Hemigordiopsids.Rivista Italiana di Paleontologia e Stratigrafia, 109:195–211.
ANGIOLINI, L., L. CARABELLI, A. NICORA, S. CRASQUIN-SOLEAU, J.MARCOUX, AND R. RETTORI. 2007. Brachiopods and other fossils fromthe Permo-Triassic boundary beds of the Antalya Nappes (SW Taurus,Turkey). Geobios, 40:715–729.
ARMSTRONG, H. A. AND M. D. BRASIER. 2005. Foraminifera, p. 142–187.In H. A. Armstrong and M. D. Brasier (eds.), Microfossils, 2nd ed.Blackwell Publishing, Oxford.
BAUD, A., S. RICHOZ, AND S. PRUSS. 2007. The lower Triassicanachronistic carbonate facies in space and time. Global and PlanetaryChange, 55:81–89.
BERCZI-MAKK, A. 1992. Midian (Upper Permian) foraminifera from thelarge Mihalovits quarry at Nagyvisnyo (North-Hungary). ActaGeologica Hungarica, 35:27–38.
BOECKELMANN, K. 1991. The Permian-Triassic of the Gartnerkofel-1Core and the Reppwand Outcrop section (Carnic Alps, Austria).Abhandlungen der Geologischen Bundesanstalt, 45:17–36.
BOYER, D. L., D. J. BOTTJER, AND M. L. DROSER. 2004. Ecologicalsignature of Lower Triassic shell beds of the Western United States.Palaios, 19:372–380.
BOZORGNIA, F. 1973. Paleozoic foraminiferal biostratigraphy of centraland east Alborz Mountains, Iran. National Iranian Oil Company,Geological Laboratories Publication, 4, 185 p.
BRADY, H. B. 1876. A Monograph of Carboniferous and PermianForaminifera (the Genus Fusulina excepted). PalaeontographicalSociety London, 166 p.
BRONNIMANN, P. AND L. ZANINETTI. 1972. Foraminifera from the basalUpper Muschelkalk at Hyeres, Western Basse-Provence, SouthernFrance. Rivista Italianna di Paleontologia e Stratigrafia, 78:31–64.
CAO, C. AND Q. SHANG. 1998. Microstratigraphy of Permo-Triassictransitional sequence of the Meishan section, Zhejiang, China. Palaeo-world, 9:147–152.
CHEN, J., T. W. BEATTY, C. M. HENDERSON, AND H. ROWE. 2009.Conodont biostratigraphy across the Permian-Triassic boundary at theDawen section, Great Bank of Guizhou, Guizhou Province, SouthChina: Implications for the Late Permian extinction and correlationwith Meishan. Journal of Asian Earth Sciences, 29: (in press).
CHEN, Z. Q., K. KAIHO, AND A. D. GEORGE. 2005. Survival strategies ofbrachiopod faunas from the end-Permian mass extinction. Palaeogeog-raphy, Palaeoclimatology, Palaeoecology, 224:232–269.
CHEN, Z. Q., K. KAIHO, A. D. GEORGE, AND J. TONG. 2006. Survivalbrachiopod faunas of the end-Permian mass extinction from thesouthern Alps (Italy) and South China. Geological Magazine,143:301–327.
CHEN, Z. Q. AND K. J. MCNAMARA. 2006. End-Permian extinction andsubsequent recovery of the Ophiuroidea (Echinodermata). Palaeogeog-raphy, Palaeoclimatology, Palaeoecology, 236:321–344.
CHEN, Z. Q., J. TONG, K. ZHANG, H. YANG, Z. LIAO, H. SONG, AND J.CHEN. 2009. Environmental and biotic turnover across Permian-Triassic boundary from shallow carbonate platform in westernZhejiang, South China. Australian Journal of Earth Sciences, 56(6):775–797.
CHERDYNTSEV, W. 1914. Foraminiferal fauna of the Permian deposits ofthe eastern belt of European Russia. Kazan, Trudy ObshchestvaEstestvoispytateley pri Imperatorskomy Kazanskomy Universitety,46:3–88. (In Russian)
CIRILLI, S., C. P. RADRIZZANI, AND M. E. A. PONTON. 1998.Stratigraphical and palaeoenvironmental analysis of the Permian-Triassic transition in the Badia Valley (Southern Alps, Italy).Palaeogeography, Palaeoclimatology, Palaeoecology, 138:85–113.
COLANI, M. 1924. Nouvelle contribution a l’etude des fusulinides del’Extreme-orient. Memoires du Service Geologique de l’Indo-chine,Hanoi-Haiphong, 11:9–191.
COLLIN, P. Y., S. KERSHAW, S. CRASQUIN-SOLEAU, AND Q. FENG. 2009.Facies changes and diagenetic processes across the Permian-Triassicboundary event horizon, Great Bank of Guizhou, South China: Acontroversy of erosion and dissolution. Sedimentology, 56:677–693.
CRASQUIN-SOLEAU, S., J. MARCOUX, L. ANGIOLINI, S. RICHOZ, A.NICORA, A. BAUD, AND Y. BERTHO. 2004. A new ostracode fauna fromthe Permian-Triassic boundary in Turkey (Taurus, Antalya Nappes).Micropaleontology, 50:281–295.
CUSHMAN, J. A. AND J. A. WATERS. 1928. Some Foraminifera from thePennsylvanian and Permian of Texas. Contributions from the CushmanFoundation for Foraminiferal Research, 4:31–55.
DELAGE, Y. AND E. HEROUARD. 1896. Traite de zoologie concrete, Paris,1, 584 p.
DERVILLE, H. 1931. Les marbres du calcaire Carbonifere en Bas-Boulonnais. Deuxieme partie, pp. 113–142. Etude micrographique desorganismes. O. Boehm, Strasbourg.
EICHWALD, C. E. 1830. Zoologia specialis, Volume 2, 323 pp.ENOS, P., D. L. LEHRMANN, J. WEI, Y. YU, J. XIAO, D. H. CHAIKIN, M.
MINZONI, A. K. BERRY, AND P. MONTGOMERY. 2007. Triassicevolution of the Yangtze platform in Guizhou Province, People’sRepublic of China. The Geological Society of America Special Paper,417:1–105.
ERK, A. S. 1942. Sur la presence du genre Codonofusiella Dunb. et Skin.dans le Permien de Bursa (Turquie). Ecologae Geologicae Helvetiae,34:243–253.
ERWIN, D. H. 1993. The Great Paleozoic Crisis: Life and Death in thePermian. Columbia University Press, New York, 327 p.
ERWIN, D. H. 2006. Extinction: How Life on Earth Nearly Ended 250Million Years Ago. Princeton University Press, Princeton, 306 p.
EZAKI, Y., J. LIU, AND N. ADACHI. 2003. Earliest Triassic microbialitemicro- to megastructures in the Huaying area of Sichuan Province,South China: Implications for the nature of oceanic conditions after theend-Permian extinction. Palaios, 18:388–402.
EZAKI, Y., J. LIU, T. NAGANO, AND N. ADACHI. 2008. Geobiologicalaspects of the earliest Triassic microbialites along the southernperiphery of the tropical Yangtze platform: Initiation and cessation ofa microbial regime. Palaios, 23:356–369.
FLUGEL, E. 2004. Microfacies of Carbonate Rocks: Analysis, Interpreta-tion and Application. Springer, Berlin, 976 p.
GAILLOT, J. AND D. VACHARD. 2007. The Khuff Formation (Middle East)and time-equivalents in Turkey and South China: Biostratigraphy fromCapitanian to Changhsingian times (Permian), new foraminiferal taxa,and palaeogeographical implications. Coloquios de Paleontologıa,57:37–223.
GALFETTI, T., H. BUCHER, R. MARTINI, P. A. HOCHULI, H. WEISSERT, S.CRASQUIN-SOLEAU, A. BRAYARD, N. GOUDEMAND, T. BRUHWILER,AND K. GUODUN. 2008. Evolution of Early Triassic outer platformpaleoenvironments in the Nanpanjiang Basin (South China) and theirsignificance for the biotic recovery. Sedimentary Geology, 204:36–60.
GEINITZ, H. B. AND A. GUTBIER. 1848. Die Versteinerungen desdeutschen Zechsteingebirges und Rothligednen, Heft 1. Arnold, Dresden.
GRAUVOGEL-STAMM, L. AND S. R. ASH. 2005. Recovery of the Triassicland flora from the end-Permian life crisis. Comptes Rendus Palevol,4:593–608.
GROVES, J. R. AND D. ALTINER. 2005. Survival and recovery ofcalcareous foraminifera pursuant to the end-Permian mass extinction.Comptes Rendus Palevol, 4:487–500.
GROVES, J. R., D. ALTINER, AND R. RETTORI. 2005. Extinction, survival,and recovery of lagenide foraminifers in the Permian-Triassic boundaryinterval, Central Taurides, Turkey. Journal of Paleontology, 79, ThePaleontological Society Memoir, 62:1–38.
GROVES, J. R., R. RETTORI, J. L. PAYNE, M. D. BOYCE, AND D. ALTINER.2007. End-Permian mass extinction of Lagenide foraminifers in theSouthern Alps (Northern Italy). Journal of Paleontology, 81:415–434.
GU, S., J. PEI, F. YANG, AND Y. GAO. 2002. Smaller foraminifera faunafrom the Changxingian of the Sidazhai section, Ziyun county, southernGuizhou Province. Acta Micropalaeontologica Sinica, 19:163–169. (InChinese)
GU, S., F. PENG, W. HE, Q. FENG, AND Y. JIN. 2005. Shallow watersmaller foraminifera from the Upper Permian of the Liuqiao area,southeastern Guangxi, South China. Acta Micropalaeontologica Sinica,22:163–172. (In Chinese)
HALLAM, A. AND P. B. WIGNALL. 1997. Mass extinctions and theiraftermath. Oxford University Press, Oxford, 320 p.
HIPS, K. AND J. HAAS. 2006. Calcimicrobial stromatolites at the Permian-Triassic boundary in a western Tethyan section, Bukk Mountains,Hungary. Sedimentary Geology, 185:239–253.
ISHII, K., Y. OKIMURA, AND K. NAKAZAWA. 1975. On the genusColaniella and its biostratigraphic significance. Journal of Geosciences,Osaka City University, 19:107–138.
JABLONSKI, D. 1986. Causes and consequences of mass extinctions: Acomparative approach, p. 183–229. In D. K. Elliott (ed.), Dynamics ofExtinction, John Wiley, New York.
SONG ET AL.—PERMIAN EXTINCTION OF CHINESE FORAMINIFERA 735

JABLONSKI, D. 2002. Survival without recovery after mass extinctions.Proceedings of the National Academy of Sciences of the United Statesof America, 99:8139–8144.
JENNY, C., A. IZART, A. BAUD, AND J. JENNY. 2004. The Permiandeposits of Hydra Island (Greece), micropaleontology, sedimentologyand paleoenvironments. Revue de Paleobiologie, 23:275–312.
JENNY-DESHUSSES, C. 1985. Rectostipulina n. gen. (5Stipulina Lys, 1978),un organisme incertae sedis du Permien superieur de la Tethys moyen-orientale: description morphologique et remarques stratigraphiques.Revue de Paleobiologie, 4:153–158.
JENNY-DESHUSSES, C., R. MARTINI, AND L. ZANINETTI. 2000. Firstoccurrence of the foraminifer Colaniella Likharev in the Upper Permianfrom the Sosio Valley (Sicily). Comptes Rendus de l’Academie desSciences - Series IIA - Earth and Planetary Science, 330:799–804.
JIANG, H., X. LAI, G. LUO, R. ALDRIDGE, K. ZHANG, AND P. WIGNALL.2007. Restudy of conodont zonation and evolution across the P/Tboundary at Meishan section, Changxing, Zhejiang, China. Global andPlanetary Change, 55:39–55.
JIN, Y. G., Y. WANG, W. WANG, Q. H. SHANG, C. Q. CAO, AND D. H.ERWIN. 2000. Pattern of marine mass extinction near the Permian-Triassic boundary in South China. Science, 289:432–436.
KARAVAEVA, N. I. AND G. P. NESTELL. 2007. Permian foraminifers of theOmolon Massifm northeastern Siberia, Russia. Micropaleontology,53:161–211.
KNOLL, A. H., R. K. BAMBACH, J. L. PAYNE, S. PRUSS, AND W. W.FISCHER. 2007. Paleophysiology and end-Permian mass extinction.Earth and Planetary Science Letters, 256:295–313.
KOBAYASHI, F. 1986. Middle Permian foraminifers of the GozenyamaFormation, Southern Kwanto Mountains, Japan. Bulletin of theNational Science Museum, Series C (Geology and Palaeontology),12:131–163.
KOBAYASHI, F. 1997. Upper Permian foraminifers from the Iwai-Kanyoarea, west Tokyo, Japan. Journal of Foraminiferal Research, 27:186–195.
KOBAYASHI, F. 2004. Late Permian foraminifers from the limestone blockin the southern Chichibu terrane of West Shikoku, SW Japan. Journalof Paleontology, 78:62–70.
KOBAYASHI, F. 2005. Permian foraminifers from the Itsukaichi-OmeArea, west Tokyo, Japan. Journal of Paleontology, 79:413–432.
KOBAYASHI, F. 2006. Middle Permian foraminifers of Kaize, southernpart of the Saku Basin, Nagano Prefecture, central Japan. Paleonto-logical Research, 10:179–194.
KOTLYAR, G. V., Y. D. ZAKHAROV, B. V. KOCZYRKEVICZ, G. S.KROPATCHEVA, K. O. ROSTOVTSEV, I. O. CHEDIJA, AND G. P. VUKS.1983. Evolution of the Latest Permian Biota. Dzhulfian and Dora-shamian Regional Stages in the USSR. Izdatelstvo Nauka, Leningrad,200 p. (In Russian)
KOTLYAR, G. V., Y. D. ZAKHAROV, G. S. KROPATCHEVA, G. P.PRONINA, I. O. CHEDIJA, AND V. I. BURAGO. 1989. Evolution of theLatest Permian Biota. Midian Regional Stage in the USSR. IzdatelstvoNauka, Leningrad, 185 p. (In Russian)
KOYLUOGLU, M. AND D. ALTINER. 1989. Micropaleontologie (foramini-feres) et biostratigraphie du Permien superieur de la region d’Hakkari(SE Turquie). Revue de Paleobiologie, 8:467–503.
KOZUR, H. W., M. AYDIN, O. DEMIR, H. YAKAR, M. C. GONCUOGLU,AND F. KURU. 2000. New stratigraphic and palaeogeographic resultsfrom the Palaeozoic and early Mesozoic of the Middle Pontides(Northern Turkey) in the Azdavay, Devrekani, Kyre and Inebolu areas:Implications for the Carboniferous-Early Cretaceous geodynamicevolution and some related remarks to the Karakaya oceanic rift basin.Geologia Croatica, 53:209–268.
KRULL, E. S., D. J. LEHRMANN, D. DRUKE, B. KESSEL, Y. YU, AND R.LI. 2004. Stable carbon isotope stratigraphy across the Permian-Triassicboundary in shallow marine carbonate platforms, Nanpanjiang Basin,South China. Palaeogeography, Palaeoclimatology, Palaeoecology,204:297–315.
LANGE, E. 1925. Eine Mittelpermische Fauna von Guguk Bulat (PadangerOberland, Sumatra), Verhandelingen van het Geologisch-Mijnbouw-kundig Genootschap voor Nederland en Kolonien. Geologische serie,7:213–295.
LEE, J. S. 1934. Taxonomic criteria of Fusulinidae with notes on sevennew Permian genera. The National Research Institute of Geology,Memoirs, 14:1–32.
LEE, J. S., S. CHEN, AND S. CHU. 1930. The Huanglung Limestone andits fauna. The National Research Institute of Geology, Memoirs, 9:85–143.
LEHRMANN, D. J. 1999. Early Triassic calcimicrobial mounds andbiostromes of the Nanpanjiang basin, south China. Geology, 27:359–362.
LEHRMANN, D. J., J. L. PAYNE, S. V. FELIX, P. M. DILLETT, H. WANG,Y. YU, AND J. WEI. 2003. Permian-Triassic boundary sections fromshallow-marine carbonate platforms of the Nanpangjiang Basin, southChina: Implications for oceanic conditions associated with the end-Permian extinction and its aftermath. Palaios, 18:138–152.
LEHRMANN, D. J., J. L. PAYNE, D. PEI, P. ENOS, D. DRUKE, K. STEFFEN,J. ZHANG, J. WEI, M. J. ORCHARD, AND B. ELLWOOD. 2007. Record ofthe end-Permian extinction and Triassic biotic recovery in theChongzuo-Pingguo platform, southern Nanpanjiang basin, Guangxi,south China. Palaeogeography, Palaeoclimatology, Palaeoecology,252:200–217.
LEHRMANN, D. J., J. RAMEZANI, S. A. BOWRING, M. W. MARTIN, P.MONTGOMERY, P. ENOS, J. L. PAYNE, M. J. ORCHARD, H. WANG, AND
J. WEI. 2006. Timing of recovery from the end-Permian extinction:Geochronologic and biostratigraphic constraints from South China.Geology, 34:1053–1056.
LEHRMANN, D. J., Y. WAN, J. WEI, Y. YU, AND J. XIAO. 2001. LowerTriassic peritidal cyclic limestone: an example of anachronisticcarbonate facies from the Great Band of Guizhou, Nanpanjiang Basin,Guizhou province, South China. Palaeogeography, Palaeoclimatology,Palaeoecology, 173:103–123.
LEHRMANN, D. J., J. WEI, AND P. ENOS. 1998. Controls on faciesarchitecture of a large Triassic carbonate platform: the Great Bank ofGuizhou, Nanpanjiang Basin, South China. Journal of SedimentaryResearch, 68:311–326.
LEVEN, E. AND A. I. OKAY. 1996. Foraminifera from the exotic Permo-Carboniferous limestone blocks in the Karakaya Complex, northwest-ern Turkey. Rivista Italiana di Paleontologia e Stratigrafia, 102:139–174.
LI, X. AND M. DING. 1987. Correlation and analysis of the UpperPermian Changxingian foraminifer and conodont zones (or assemblag-es) in south China. Geological Review, 33:395–401. (In Chinese)
LIEM, N. V. 1971. New Late Permian species of Neoendothyra(Endothyrida) from north Vietnam. Paleontologicheskiiy Zhurnal,3:110–111.
LIN, J. 1978. Foraminifera (Carboniferous-Permian), p. 4–43. In ThePalaeontological Atlas of Central-South China, Part 4: Microfossils.Geological Publishing House, Beijing. (In Chinese)
LIN, J. 1984. Foraminifera, p. 110–177, 328–330. In Yichang Institute ofGeology and Minerals Resources (ed.), Biostratigraphy of the YangtzeGorge area (3): Late Palaeozoic Era. Geological Publishing House,Beijing. (In Chinese)
LIN, J. 1985. Foraminifera from the Lower Permian Qixia Formation ofJiahe, Guangdong. Geological Review, 31:289–294. (In Chinese)
LIN, J., J. LI, AND Q. SUN. 1990. Late Paleozoic Foraminifera in SouthChina. Science Press, Beijing, 297 p. (In Chinese)
LIU, J. B., Y. EZAKI, S. R. YANG, H. F. WANG, AND N. ADACHI. 2007.Age and sedimentology of microbialites after the end-Permian massextinction in Luodian, Guizhou Province. Journal of Palaeogeography,9:473–486. (In Chinese)
LOEBLICH, A. R., JR. AND H. TAPPAN. 1988. Foraminiferal Genera andTheir Classification. Van Nostrand Reinhold Co., New York, 970 p.
LYS, M., M. COLCHEN, J. P. BASSOULET, J. MARCOUX, AND G. MASCLE.1980. La biozone a Colaniella parva du Permien superieur et samicrofaune dans le bloc calcaire exotique de Lamayuru, Himalaya duLadakh. Revue de Micropaleontologie, 23:76–108.
LYS, M. AND J. MARCOUX. 1978. Les niveaux du Permian superieur desNappes d’Antalya (Taurides occidentales, Turquie). Compte RenduHebdomadaire des Seances de l’ Academie des Sciences, Paris, ser. D,286:1417–1420.
LYS, M., G. STAMPFLI, AND J. JENNY. 1978. Biostratigraphie duCarbonifere et du Permien de I’Elbourz oriental (Iran du N. E.). Notesdu Laboratoire de Paleontologie de l’Universite de Geneve, 2:63–99.
MARQUEZ, L. 2005. Foraminiferal fauna recovered after the Late Permianextinctions in Iberia and the westernmost Tethys area. Palaeogeogra-phy, Palaeoclimatology, Palaeoecology, 229:137–157.
MARSHALL, C. R. 1990. Confidence intervals on stratigraphic ranges.Paleobiology, 16:1–10.
MARSHALL, C. R. 1995. Distinguishing between sudden and gradualextinctions in the fossil record: predicting the position of theCretaceous-Tertiary iridium anomaly using the ammonite fossil recordon Seymour Island, Antarctica. Geology, 23:731–734.
MELDAHL, K. H. 1990. Sampling, species abundance, and the strati-graphic signature of mass extinction: A test using Holocene tidal flatmolluscs. Geology, 18:890–893.
MERTMANN, D. 2000. Foraminiferal assemblages in Permian carbonatesof the Zaluch Group (Salt Range and Trans Indus Ranges, Pakistan).Neues Jahrbuch fur Geologie und Palaontologie-Monatshefte, 3:129–146.
736 JOURNAL OF PALEONTOLOGY, V. 83, NO. 5, 2009

MIKHALEVICH, V. I. 2000. The phylum Foraminifera d’Orbigny, 1826 -Foraminifers, p. 533–623. In G. P. Protista (ed.), Manual on Zoology.Part 1. Russian Academy of Sciences, Zoological Institute, Sankt-Petersburg, Nauka.
MIKLUKHO-MAKLAY, K. V. 1954. Foraminifers of the Upper PermianDeposits of the Northern Caucasus. Trudy Vsesoyuznogo Nauchno-Issledovatel’skogo Geologicheskogo Instituta (VSEGEI). MinisterstvaGeologii i Okhrani Nedr, 163 p.
NAKAZAWA, K., H. M. KAPOOR, K. ISHII, Y. BANDO, Y. OKIMURA, AND
T. TOKUOKA. 1975. The Upper Permian and Lower Triassic inKashmir. Memior of Faculty of Science, Kyoto University, SeriesGeology and Mineralogy, 42:1–106.
NESTELL, M. AND G. P. PRONINA. 1997. The age and distribution of thegenus Hemigordiopsis, p. 105–110. In C. A. Ross, J. R. P. Ross, andP. L. Breckle (eds.), Late Paleozoic Foraminifera-their biostratigra-phy, evolution, and paleoecology and the Mid-Carboniferousboundary. Volume 36. Cushman Foundation for ForaminiferalResearch.
OKIMURA, Y. AND K. ISHII. 1981. Upper Permian and Lower Triassicforaminifera from Guryul Ravine and the spur three kilometers north ofBarus. In K. Nakazawa and H. M. Kapoor (eds.), The Upper Permianand Lower Triassic faunas of Kashmir. Palaeontologica Indica,Memoirs of the Geological Survey of India, New Series, 46:25–38
OKIMURA, Y., K. ISHII, AND C. A. ROSS. 1985. Biostratigraphicalsignificance and faunal provinces of Tethyan Late Permian smallerForaminifera, p. 115–1158. In K. Nakazawa and J. M. Dickins (eds.),The Tethys, her paleogeography and paleobiogeography from Paleozoicto Mesozoic. Tokai University Press, Tokyo.
OTA, A. AND Y. ISOZAKI. 2006. Fusuline biotic turnover across theGuadalupian-Lopingian (Middle-Upper Permian) boundary in mid-oceanic carbonate buildups: Biostratigraphy of accreted limestone inJapan. Journal of Asian Earth Sciences, 26:353–368.
PAYNE, J. L., D. J. LEHRMANN, S. CHRISTENSEN, J. WEI, AND A. H.KNOLL. 2006b. Environmental and biological controls on the initiationand growth of a Middle Triassic (Anisian) reef complex on the GreatBank of Guizhou , China. Palaios, 21:325–343.
PAYNE, J. L., D. J. LEHRMANN, D. FOLLETT, M. SEIBEL, L. R. KUMP, A.RICCARDI, D. ALTINER, H. SANO, AND J. WEI. 2007. Erosionaltruncation of uppermost Permian shallow-marine carbonates andimplications for Permian-Triassic boundary events. GSA Bulletin,119:771–784.
PAYNE, J. L., D. J. LEHRMANN, J. WEI, AND A. H. KNOLL. 2006a. Thepattern and timing of biotic recovery from the end-Permian extinctionon the Great Bank of Guizhou, Guizhou Province, China. Palaios,21:63–85.
PAYNE, J. L., D. J. LEHRMANN, J. WEI, M. J. ORCHARD, D. P. SCHRAG,AND A. H. KNOLL. 2004. Large perturbations of the carbon cycleduring recovery from the end-Permian mass extinction. Science, 305:506–509.
PINARD, S. AND B. L. MAMET. 1998. Taxonomie des petits foraminiferesdu Carbonifere superieur-Permien inferieur du bassin de Sverdrup,Arctique Canadien. Palaeontographica Canadiana, 15:1–253.
POWERS, C. M. AND J. F. PACHUT. 2008. Diversity and Distribution ofTriassic Bryozoans in the Aftermath of the End-Permian MassExtinction. Journal of Paleontology, 82:362–371.
PRONINA, G. P. 1988. The Late Permian smaller foraminifers ofTranscaucasus. Revue de Paleobiologie, 2:89–96.
PRONINA-NESTELL, G. P. AND M. K. NESTELL. 2001. Late Changhsingianforaminifers from the northwestern Caucasus. Micropaleontology,47:205–234.
RAMPINO, M. R. AND A. C. ADLER. 1998. Evidence for abrupt latestPermian mass extinction of foraminifera: Results of tests for the Signor-Lipps effect. Geology, 26:415–418.
RAUP, D. M., P. A. SABINE, P. ASHMOLE, P. W. KING, AND A. W.WOLFENDALE. 1989. The case for extraterrestrial causes of extinction.Philosophical Transactions of the Royal Society of London - Series B:Biological Sciences, 325:421–435.
REICHEL, M. 1945. Sur un Miliolide nouveau du Permien de l’ıle dechypre. Verhandlungen der Naturforschenden Gesellschaft in Basel,56:521–530.
REICHEL, M. 1946. Sur quelques foraminiferes nouveaux du Permienmediterraneen. Eclogae Geologicae Helvetiae, 38:524–560.
REYTLINGER, E. A. 1950. Foraminifera from the Middle Carboniferousdeposits of the central part of the Russian Platform (exclusive of thefamily Fusulinidae). Akademiya Nauk SSSR, Trudy Instituta Geolo-gicheskikh Nauk, vypusk 126, Geologicheskaya Seriya, 47:1–127. (InRussian)
REYTLINGER, E. A. 1965. Foraminiferal development in the Late Permianand Early Triassic epochs in the territory of Transcaucasia. VoprosyMikropaleontologii, 9:45–70.
RODLAND, D. L. AND D. J. BOTTJER. 2001. Biotic recovery from the End-Permian mass extinction: Behavior of the inarticulate brachiopodLingula as a disaster taxon. Palaios, 16:95–101.
SAKAGAMI, S. AND A. HATTA. 1982. On the Upper Permian Palaeofu-sulina-Colaniella fauna from Khao Doi Pha Phlung, north Thailand.Geology and Palaeontology of Southeast Asia, 24:1–14.
SCHUBERT, J. K. AND D. J. BOTTJER. 1992. Early Triassic stromatolites aspost-mass extinction disaster forms. Geology, 20:883–886.
SELLIER DE CIVRIEUX, J. M. AND T. F. J. DESSAUVAGIE. 1965.Reclassification de quelques Nodosariidae, particulierement du Permienau Lias. Maden Tetkik ve Arama Enstitusu Yayinlarindan, Ankara,124:1–178.
SHANG, Q., D. VACHARD AND M. CARDROIT. 2003. Smaller foraminiferafrom the Late Changhsingian (Latest Permian) of southern Guangxiand discussion on the Permian-Triassic boundary. Acta Micropalaeon-tologica Sinica, 20:377–388. (In Chinese)
SHENG, J. 1955. Fusulinids from Changxing limestone. Acta Palaeonto-logica Sinica, 3:287–297. (In Chinese)
SHENG, J. 1956. Permian fusulinids from Liangshan, Hanchung, southernShensi. Acta Palaeontologica Sinica, 4:173–227. (In Chinese)
SHENG, J., C. CHEN, Y. WANG, L. RUI, Z. LIAO, Y. BANDO, K. ISHII, AND
K. NAKAZAWA. 1984. Permian-Triassic boundary in middle and easternTethys. Journal of Faculty of Sciences, Hokkaido University, Series 4,21:133–181.
SIDO, M. 1978. Lower Carboniferous Foraminifera from bituminouslimestones recovered by an ore exploratory pir near Szabadbattyan,western Hungary. Bulletin of the Hungarian Geological Society,108:172–193.
SIGNOR, P. W. AND J. H. LIPPS. 1982. Sampling bias, gradual extinctionpatterns, and catastrophes in the fossil record, p. 291–296. In L. T.Silver and P. H. Schultz (eds.), Geological implications of impacts oflarge asteroids and comets on the Earth. The Geological Society ofAmerica Special Paper 190.
SOLOW, A. R. 1996. Tests and confidence intervals for a common upperendpoint in fossil taxa. Paleobiology, 22:406–410.
SOLOW, A. R., AND W. K. SMITH. 2000. Testing for a mass extinctionwithout selecting taxa. Paleobiology, 26:647–650.
SONG, H. AND J. TONG. in press. Foraminifera, In S. Xie (ed.),Geobiology: Interaction of Life and Environment on Earth. ChinaUniversity of Geosciences Press, Wuhan. (In Chinese)
SONG, H., J. TONG, AND Z. Q. CHEN. 2009. Two episodes of foraminiferalextinction near the Permian-Triassic boundary at the Meishan section,South China. Australian Journal of Earth Sciences, 56(6):764–774.
SONG, H., J. TONG, AND W. HE. 2006. Latest Permian small foraminiferalfauna at the Meishan Section, Zhejiang Province. Acta Micropalaeon-tologica Sinica, 23:87–104. (In Chinese)
SONG, H., J. TONG, K. ZHANG, Q. WANG, AND Z. Q. CHEN. 2007.Foraminifers surviving from the end-Permian mass extinction atMeishan, Changxing, China. Palaeoworld, 22:105–119.
SOSNINA, M. I. 1978. On foraminifera of the late Permian ChandalazskHorizon of South Primorye, p. 24–41. In L. I. Popeko (ed.), UpperPaleozoic of northeast Asia. Instituta Tektoniki i Geofiziki, AkademiyaNauk SSSR, Dal’ nevostochnyy Nauchnyy Tsentr, Vladivostok.
SPRINGER, M. S. 1990. The effect of random range truncations on patternsof evolution in the fossil record. Paleobiology, 16:512–520.
STANLEY, S. M. AND X. YANG. 1994. A double mass extinction at the endof the Paleozoic Era. Science, 266:1340–1344.
STRAUSS, D. AND P. M. SADLER. 1989. Classical confidence intervals andBayesian probability estimates for ends of local taxon ranges.Mathematical Geology, 21:411–427.
SULIEMANOV, I. S. 1949. Some smaller foraminifers from the UpperPaleozoic deposits of Bashkiria. Akademiya Nauk SSSR, TrudyInstituta Geologicheskikh Nauk, vypusk 105, Geologicheskaya Seriya,35:236–243. (In Russian)
TAPPAN, H. AND A. R. LOEBLICH, JR. 1988. Foraminiferal evolution,diversification, and extinction. Journal of Paleontology, 62:695–714.
TIEN, N. D. 1986. Foraminifera and algae from the Permian ofKampuchea, p. 116–137. In H. Fontaine (ed.), The Permian ofSoutheast Asia. COOP Technical Bulletin, 18.
TIEN, N. D. 1988. Note on two ‘‘incertae sedis’’ from the Permian of westThailand, p. 109–111. In H. Fontaine and V. Suteethorn (eds.), LatePalaeozoic and Mesozoic fossils of west Thailand and their environ-ments. CCOP Technical Bulletin, 20.
TONG, J. 1998. The Middle Triassic environstratigraphy of central-southGuizhou, Southwest China. Palaeogeography, Palaeoclimatology,Palaeoecology, 143:293–305.
TONG, J. 2004. Evolution of foraminiferid groups through the Permian-Triassic transition in South China., p. 701–708, 1070. In J. Rong and Z.Fang (eds.), Mass extinction and recovery—Evidences from the
SONG ET AL.—PERMIAN EXTINCTION OF CHINESE FORAMINIFERA 737

Palaeozoic and Triassic of South China. University of Science andTechnology of China Press, Hefei. (In Chinese)
TONG, J. AND W. KUANG. 1990. A study of the Changxingianforaminifera and microfacies in Liangfengya, Chongqing, SichuanProvince. Earth Science—Journal of China University of Geosciences,15:337–344. (In Chinese)
TONG, J. AND G. R. SHI. 2000. Evolution of the Permian and TriassicForaminifera in South China, p. 291–307. In H. Yin, J. M. Dickins, G.R. Shi, and J. Tong (eds.), Permian-Triassic evolution of Tethys andwestern circum Pacific. Elsevier, Amsterdam.
TRIFONOVA, E. T. 1978. New foraminifera species from the Lower-Triassic and Middle Triassic in Bulgaria. Dokl Bolg Akad Nauk,31:1151–1154.
TURHAN, N., C. OKUYUCU, AND C. GONCUOGLU. 2004. AutochthonousUpper Permian (Midian) carbonates in the western Sakarya compositeTerrane, Geyve area, Turkey: Preliminary data. Turkish Journal ofEarth Sciences, 13:215–229.
UENO, K. 1992. Permian foraminifers from the Takakurayama Group ofthe southern Abukuma Mountains, northeast Japan. Transactions andProceedings of the Palaeontological Society of Japan, 168:1265–1295.
UENO, K. AND S. SAKAGAMI. 1993. Middle Permian foraminifers fromBan Nam Suai Sa-At, Changw at Loei, Northeast Thailand. Transac-tions and Proceedings of the Palaeontological Society of Japan,172:277–291.
UNAL, E., D. ALTINER, O. YILMAZI, AND S. OZKAN-ALTINER. 2003.Cyclic sedimentation across the Permian-Triassic boundary (CentralTaurides, Turkey). Rivista Italiana di Paleontologia e Stratigrafia,109:359–376.
VACHARD, D. 1981. Tethys et Gondwana au Paleozoıque superieur. Lesdonnees afghanes-biostratigrphie, micropaleontologie, paleontologie.Documents et travaux de l’Istitute geologique Albert de Lapparent(I.G.A.L.), 2, 463 p.
VACHARD, D. AND J. FERRIERE. 1991. Une association a Yabeina(foraminifere fusulinoıde) dans le Midien (Permien superieur) de laregion de Whangaroa (Baie d’Orua, Nouvelle-Zelande). Revue deMicropaleontologie, 34:201–230.
VACHARD, D., M. HAUSER, R. MARTINI, L. ZANINETTI, A. MATTER,AND T. PETERS. 2002. Middle Permian (Midian) foraminiferalassemblages from the Batain Plain (eastern Oman): Their significanceto Neotethyan paleogeography. Journal of Foraminiferal Research,32:155–172.
VACHARD, D., R. RETTORI, L. ANGIOLINI, AND A. CHECCONI. 2008.Glomomidiella gen. n. (Foraminifera, Miliolata, Neodiscidae): A newgenus from the Late Guadalupian-Lopingian of Hydra Island (Greece).Rivista Italiana di Paleontologia e Stratigrafia, 114:349–361.
VACHARD, D., A. ZAMBETAKIS-LEKKAS, E. SKOURTSOS, R. MARTINI,AND L. ZANINETTI. 2003. Foraminifera, algae and carbonate micro-problematica from the Late Wuchiapingian/Dzhulfian (Late Permian)of Peloponnesus (Greece). Rivista Italiana di Paleontologia e Strati-grafia, 109:339–358.
WANG, K. 1974. Carboniferous and Permian foraminifera, p. 248–288. InNanjing Institute of Geology and Palaeontology, Acedia Sinica (ed.),Palaeontological Atlas of Southwest China. Science Press, Bejing. (InChinese)
WANG, K. 1976. The foraminifera from the Changhsing formation inwestern Guizhou. Acta Palaeontologica Sinica, 15:186–194. (InChinese)
WANG, K. 2002. Carboniferous and Permian foraminifera from theHengduan Mountain region. Acta Micropalaeontologica Sinica,19:112–133. (In Chinese)
WANG, S. C. AND P. J. EVERSON. 2007. Confidence intervals for pulsedmass extinction events. Paleobiology, 33:324–336.
WANG, S. C. AND C. R. MARSHALL. 2004. Improved confidence intervalsfor estimating the position of a mass extinction boundary. Paleobiology,30:5–8.
WANG, Y., J. TONG, J. WANG, AND X. ZHOU. 2005. Calcimicrobialiteafter end-Permian mass extinction in South China and its palaeoenvi-ronmental significance. Chinese Science Bulletin, 50:665–671.
WEDEKIND, R. 1937. Einfuhrung in die Grundlagen der historischenGeologie. II. Mikrobiostratigraphie: Die Korallen-und Foraminifer-enzeit. Ferdinand Enke, Stuttgart, 136 p.
XIE, S., R. D. PANCOST, H. YIN, H. WANG, AND R. P. EVERSHED. 2005.Two episodes of microbial change coupled with Permo/Triassic faunalmass extinction. Nature, 434:494–497.
YANG, Z., H. YIN, S. WU, F. YANG, M. DING, AND G. XU. 1987.Permian-Triassic Boundary Stratigraphy and Fauna of South China.Geological Publishing House, Beijing, 379 p. (In Chinese with Englishsummary)
YANG, Z. Y., S. B. WU, H. F. YIN, G. R. XU, K. X. ZHANG, AND X. M.BI. 1993. Permo-Triassic Events of South China. Geological PublishingHouse, Beijing, 153 p.
YIN, H., K. ZHANG, J. TONG, Z. YANG, AND S. WU. 2001. The GlobalStratotype Section and Point (GSSP) of the Permian-Triassic boundary.Episodes, 24:102–114.
ZANINETTI, L., D. ALTINER, AND E. CATAL. 1981. Foraminiferes etbiostratigraphie dans le Permian superieur du Taurus oriental, Turquie.Notes du Laboratoire de Paleontologie de l’Universite de Geneve, 7:1–37.
ZHANG, K. X., M. H. DING, X. L. LAI, AND J. H. LIU. 1996. Conodontsequences of the Permian-Triassic boundary strata at Meishan Section,South China, p. 57–64. In H. F. Yin (ed.), The Palaeozoic-MesozoicBoundary candidates of Global Stratotype Section and Point of thePermian-Triassic Boundary. China University of Geosciences Press,Wuhan.
ZHANG, Z. AND Z. HONG. 2000. The middle Permian smaller foraminif-eral from the Tungtzeyen form ation of Wuping, Fujian. ActaMicropalaeontologica Sinica, 17:39–56. (In Chinese)
ZHANG, Z. AND Z. HONG. 2004. Smaller Foraminifera Fauna from theChanghsing Formation of Datian, Fujian. Acta MicropalaeontologicaSinica, 21:64–84. (In Chinese)
ZHAO, J., J. SHENG, Z. YAO, X. LIANG, C. CHEN, L. RUI, AND Z. LAO.1981. The Changhsingian and Permian-Triassic boundary of SouthChina. Bulletin of Nanjing Institute of Geology and Paleontology,Academia Sinica, 2:1–112. (In Chinese)
ZOLOTOVA, V. P. AND V. V. BARYSHNIKOV. 1980. Foraminifers from thestratotype area of the Kungurian Stage, p. 72–105. Akademiya NaukSSSR, Ural’skii Nauchnyi Tsentr. (In Russian)
ACCEPTED 9 MAY 2009
738 JOURNAL OF PALEONTOLOGY, V. 83, NO. 5, 2009





























