Embed Size (px)
Citation preview

Tema 2. Organización atómica y molecular de la
materia. Principios de termodinámica
Bioquímica estructural
Grado Ingeniería de la Salud.
Curso 2013-2014
Mohammed Rafii El Idrissi Benhnia, PhD
Departamento de Bioquímica Médica, Biología Molecular e Inmunología
Facultad de Medicina
& Laboratorio de Inumovirologia, Instituto de Biomedicina de Sevilla
SEVILLA, ESPANA

Bioenergética y termodinámica: Energía,
Calor y trabajo La bioenergética describe la forma en la que los organismos adquieren,
canalizan y utilizan la energía: plantas (luz solar) y animales (combustible)
propiedad fundamental de todos los organismos vivos.
estudio cuantitativo de las transducciones de energía, cambio de una
forma de energía en otra.
Utilizan la energía química de los combustibles para poder sintetizar
macromoléculas complejas y altamente ordenadas a partir de precursores.
Convierten esta energía en gradientes de concentración y eléctricos,
en movimientos, calor y en algunos casos en luz (peces abisales)
Los organismos fotosintéticos transducen la energía luminosa en otras
formas de energía.

Bioenergética y termodinámica: Energía,
Calor y trabajo
Dos leys fundamentales de la termodinámica (inter-conversión de las
diferentes formas de energía):
Primera ley: principio de conservación de la energía en el universo,
donde la cantidad total permanece constante.
Segunda ley: el universo tiende hacia un aumento de desorden.
Hay tres cantidades termodinámicas que describen los cambios de
energía en una reacción química: la energía libre G, entalpía H y
entropía S

Bioenergética y termodinámica: Energía,
Calor y trabajo
Las reacciones que tienen lugar espontáneamente suelen ser
exotérmicas, esto es, con desprendimiento de calor.
El calor que se desprende o absorbe en un sistema de reacción a
presión constante recibe el nombre de Entalpía (DH): refleja el
numero y la clase de enlaces químicos entre los reactivos (R) y los
productos (P). El contenido calórico de P es menor que el de R y DH
tiene, por convención, un valor: DH < 0
Sin embargo, hay reacciones endotérmicas que cursan espontá-
neamente (adquieren calor), DH > 0; por ejemplo, la disolución de
sulfato amónico en agua.
¿Cuándo tiene lugar una reacción química?

La disolución de sulfato amónico en agua es un proceso por
el cual se pasa de un sistema altamente ordenado, cual es el
estado cristalino, a otro de mucho mayor desorden molecular.
La función termodinámica que mide el desorden o la aleatoriedad
de un sistema recibe el nombre de Entropía (DS). Puede observarse
que muchas reacciones que cursan espontáneamente lo hacen
con incremento positivo de entropía: DS > 0
(P son menos complejos y mas desordenados que los reactivos)
Sin embargo, hay procesos espontáneos que cursan con dismi-
nución de entropía, p.e.: la solidificación del agua a 0ºC.
Bioenergética y termodinámica.

Ni la entropía ni la entalpía valen como criterio único para definir la
espontaneidad de una reacción.
Existe otra función termodinámica de estado que expresa la cantidad de
energía capaz de realizar trabajos durante una reacción a T y a presión
constante, y agrupa a las dos en procesos, la Energía Libre de
Gibbs, que se define así: DG = DH – TDS
La energía libre de Gibbs es un criterio válido para verificar la
espontaneidad de una reacción. Pueden cursar espontáneamente aquellos
procesos en los que se desprende energía libre (DG < 0, reacción
exergónicas); no pueden hacerlo, sin embargo, aquellos procesos en los
que se absorbe energía libre (DG > 0, reacción endergónica)
Bioenergética y termodinámica.
(Temperatura absoluta, K), DG (J/mol o cal/mol), DH (J/mol)<0, DS (J/mol x K)>0,
1 cal=4,184J; 25C = 298K

Sea un sistema de reacción: A B
La reacción cursará de izquierda a derecha cuando las concentraciones
de A y B sean tales que la energía libre del sistema sea negativa. La
energía libre del sistema viene dada por:
DG = DG0 + RT ln [A]
[B]
Donde DG0 es la Energía Libre Estándar de la reacción:
la energía libre del sistema con los reactivos a concentración unidad
(Temperatura absoluta, K), DG (J/mol o cal/mol),
1 cal=4,184J; 25ºC = 298K; R (contante de los gases)= 8,315 J/mol x K (=1,987 cal/mol x K)
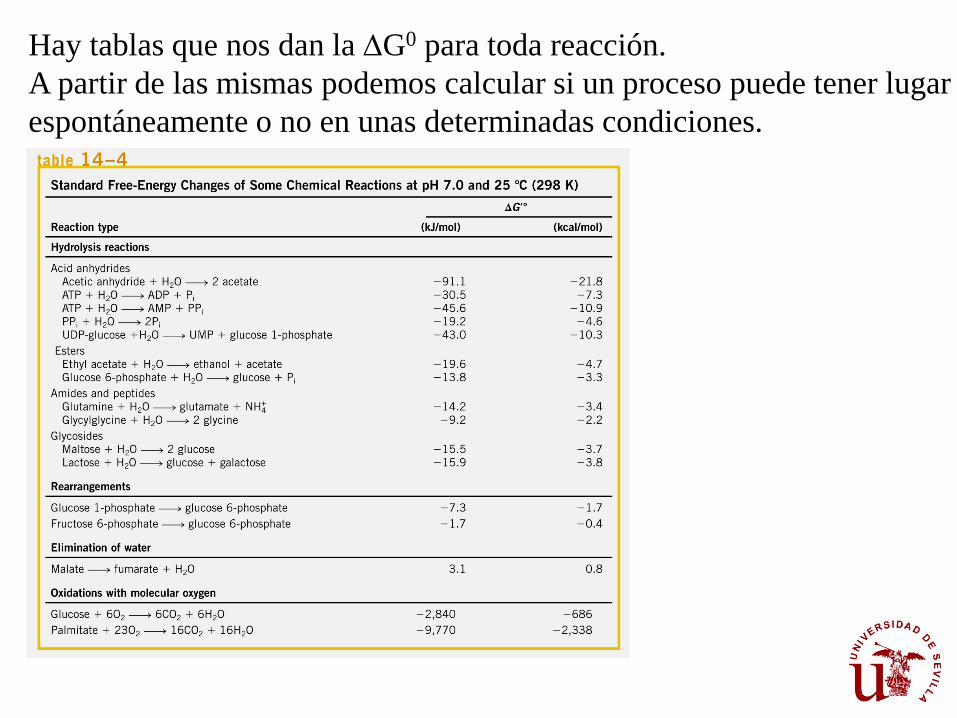
Hay tablas que nos dan la DG0 para toda reacción.
A partir de las mismas podemos calcular si un proceso puede tener lugar
espontáneamente o no en unas determinadas condiciones.

La expresión que nos da la energía libre de un proceso tiene
otra importante derivación. Para la reacción
DG = DG0 + RT ln [A]
[B] Esta expresión es:
En el equilibrio, DG = 0; por tanto,
0 = DG0 + RT ln [Aeq]
[Beq]
Pero [Aeq]
[Beq] = Keq (contante de equilibrio)
Por tanto, DG0 = -RT ln Keq
A B
DG < 0 y se aproxima a 0 a medida que la reacción transcurra debido a que la concentración
real de A será mas pequeña y la de B será cada vez mayor.

Calcular la energía libre del proceso de descomposición del
éster Glucosa-6-fosfato a Glucosa y fosfato inorgánico en las
condiciones intracelulares, que son:
Temperatura: T= 37 ºC, equivalentes a 310 ºK (=273+37)
R = 8,315 J/mol x K (=1,987 cal/mol x K)
[Glucosa-6-fosfato], 1 mM
[Glucosa], 0,01 mM
[fosfato], 10 mM
La reacción es:
G6P + H2O G + Pi
La Energía Libre Estándar de la reacción es de -3250 cal/mol
Ejemplo Bioquímico, cálculos de energía libre:

La expresión que nos da la Energía Libre es:
DG = DG0 + RT ln [Glucosa] [Fosfato]
[Glucosa-6-fosfato]
(No se tiene en cuenta el agua porque su concentración se
considera constante, de forma que su actividad es = a la unidad)
Sustituyendo, obtenemos:
DG = -3250 + 1,98*310* ln 10-5*10-2
10-3 = -8903,3 cal/mol
Por lo tanto, en las condiciones intracelulares el
proceso puede tener lugar espontáneamente.

Relacionado con el anterior, tendríamos el siguiente problema:
¿Cuál sería la concentración mínima necesaria de glucosa para que,
siendo el resto de las condiciones iguales a las del ejemplo anterior,
en la célula tuviera lugar la formación de éster por reversión de la
hidrólisis?
Esta concentración sería la que diera lugar a un valor de 0 para
DG en el ejemplo anterior. Así, llamando X a la conc. de glucosa,
0 = -3250 + 1,98*310*ln X*10-2
10-3
Despejando obtenemos ln (X*10) = 5,29
X = e(5,29)/10 = 19,93 M

Problemas:

Problemas:
Estructuras
complejas
Estructuras
simples
Catabolismo
Anabolismo
DG
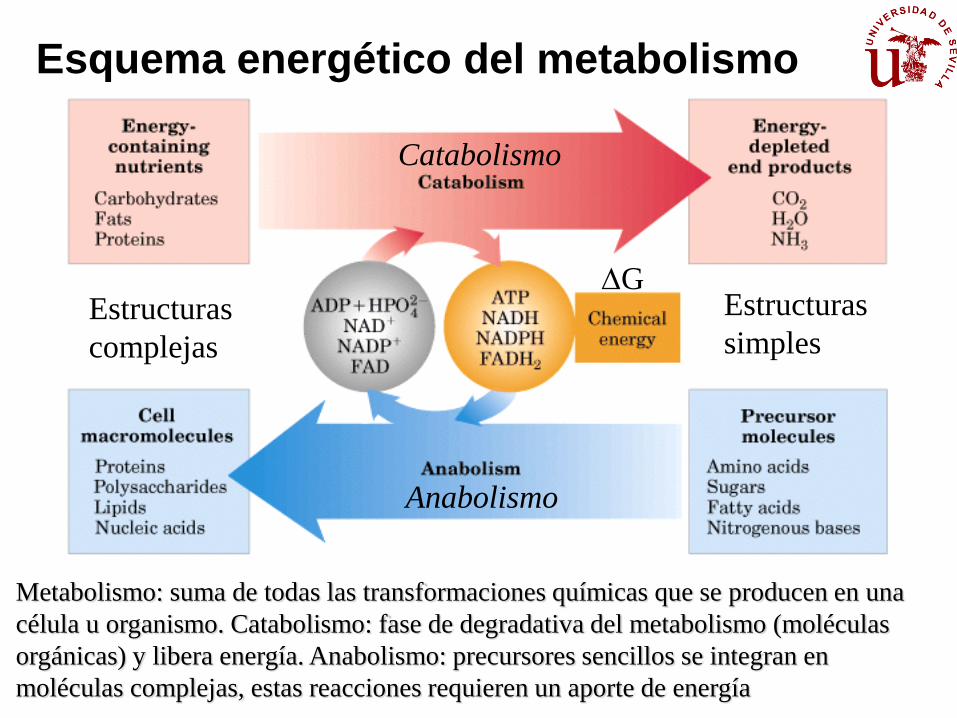
Esquema energético del metabolismo
Metabolismo: suma de todas las transformaciones químicas que se producen en una
célula u organismo. Catabolismo: fase de degradativa del metabolismo (moléculas
orgánicas) y libera energía. Anabolismo: precursores sencillos se integran en
moléculas complejas, estas reacciones requieren un aporte de energía

En general, podemos decir:
Reacciones catabólicas: DG < 0
Reacciones anabólicas: DG > 0
Supongamos una reacción anabólica mediante la cual se forma
un enlace químico entre A y B; esquemáticamente,
A + B A-B (DG > 0)
La reacción no puede tener lugar espontáneamente. ¿Cómo
puede entonces tener lugar en el metabolismo?

Lo que ocurre en el metabolismo es que las reacciones
termodinámicamente desfavorable (endergónicas, DG > 0) se acoplan a
reacciones exergónicas (DG < 0) de manera que :
1. La energía desprendida en una de las reacciones es absorbida por la
otra.
2. La suma total de energías libres de una y otra reacción da una DG <0,
así la reacción global es exergónica y el proceso en conjunto tiene lugar
espontáneamente.

Así, la reacción
A + B A-B (DG1 > 0)
Se acopla a
X-Y + H2O X + Y (DG2 < 0)
Dando lugar a una reacción global
A + B + X-Y + H2O A-B + X + Y
Siendo |DG2 | > |DG1|
(DG < 0)

El tipo de reacción
X-Y + H2O X + Y (DG2 < 0)
Que tiene lugar en los seres vivos para acoplarse a procesos ender-
gónicos es, en la mayoría de los casos, la hidrólisis de anhídridos
de ácido, y particularmente, la hidrólisis de polifosfatos :
O-POPO
OO
O-O-
R + H2O O-P
O
O-
HOPO
O
O-
R OH +
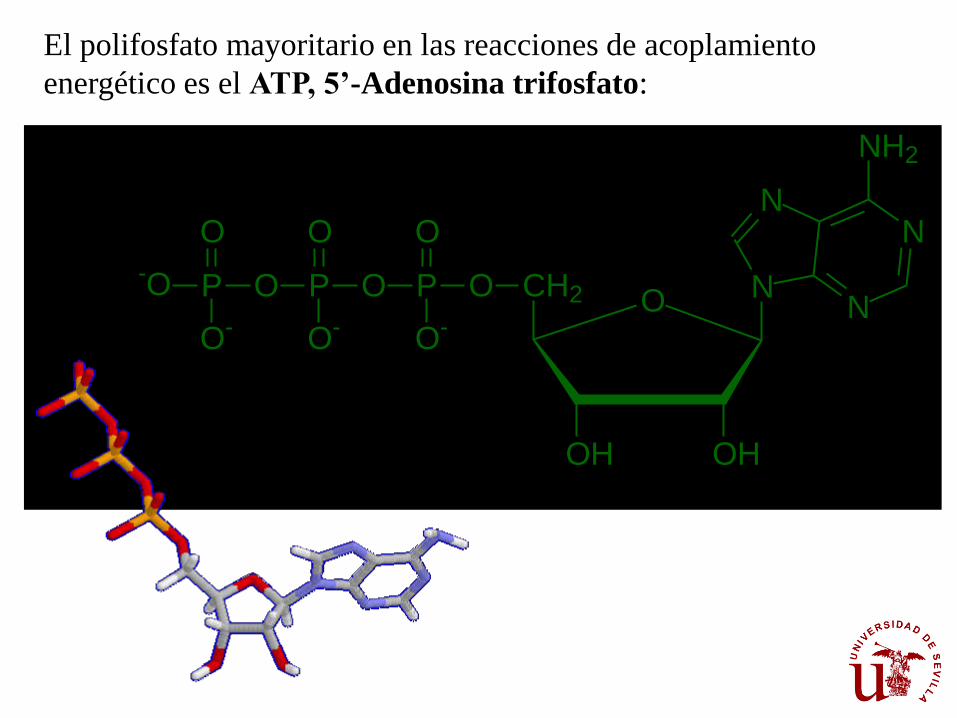
El polifosfato mayoritario en las reacciones de acoplamiento
energético es el ATP, 5’-Adenosina trifosfato:
OCH2 N
N
N
N
NH2
OHOH
OPOPOP-O
O O O
O-O-O-

De esta manera, los procesos catabólicos productores de energía
generan ATP, que se empleará en todas aquellas reacciones ender-
gónicas en las que sea requerido.
En general, el ATP se produce de dos maneras:
1. Por fosforilación a nivel de substrato
(procesos anaeróbicos, fermentativos)
Por ejemplo: La Glicolisis
2. Por fosforilación oxidativa
(procesos aeróbicos, oxidativos)
Por ejemplo: Ciclo de Krebs, - b oxidación, etc.
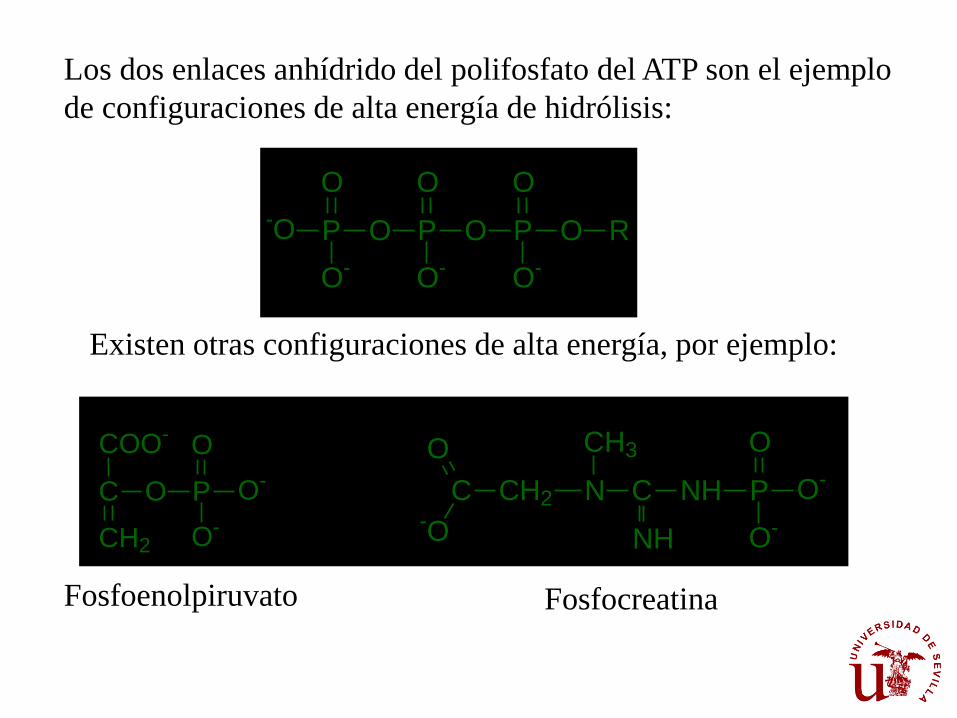
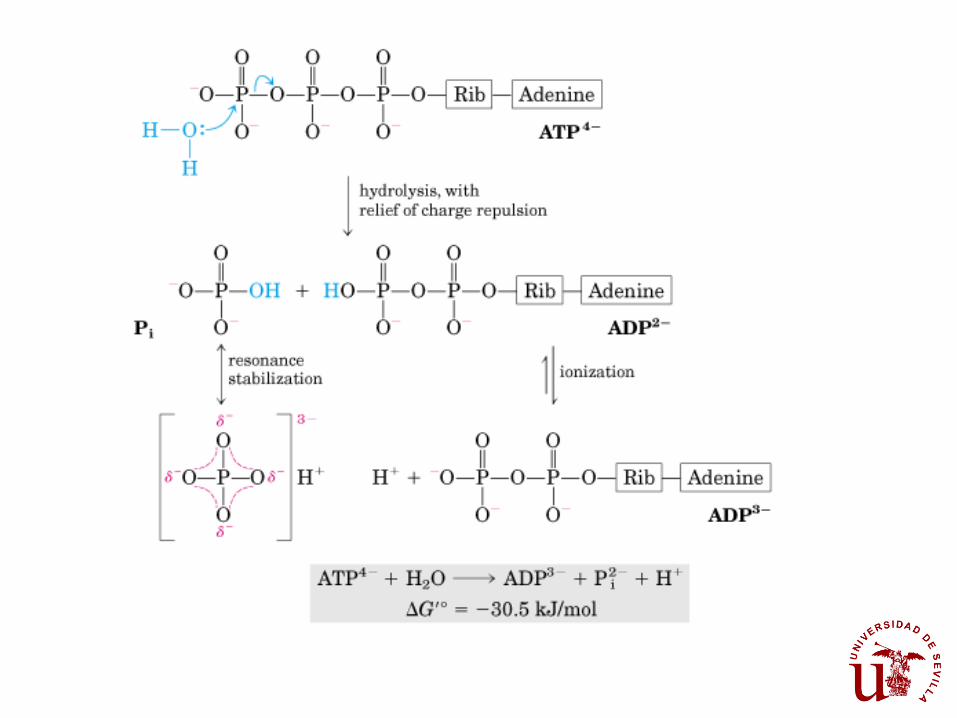
Los dos enlaces anhídrido del polifosfato del ATP son el ejemplo
de configuraciones de alta energía de hidrólisis:
OPOPOP-O
O O O
O-O-O-
R
Existen otras configuraciones de alta energía, por ejemplo:
Fosfoenolpiruvato
O-PO
O
O-
C
CH2
COO-
O-P
O
O-
NHC
NH
N
CH3
CH2C
O
-O
Fosfocreatina
O-P
O
O-
OC
O
H2N
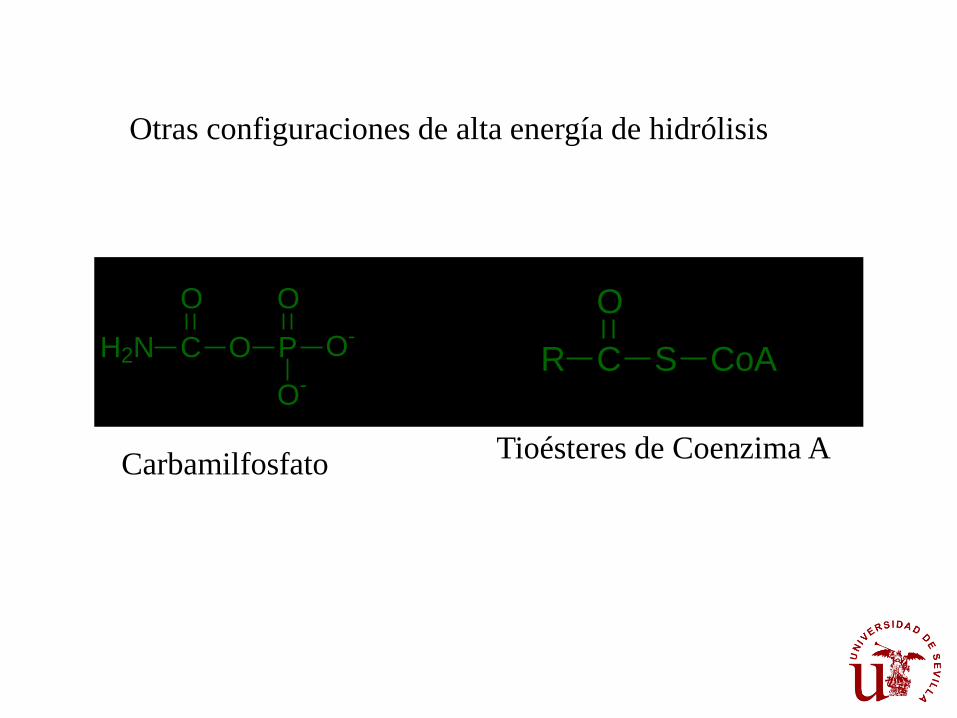
Carbamilfosfato
R C
O
S CoA
Tioésteres de Coenzima A
Otras configuraciones de alta energía de hidrólisis

Consideremos ahora la reacción de degradación aeróbica de la glucosa:
C6H12O6 + 6O2 6CO2 + 6H2O DG0’ = -684 kcal/mol
Según lo hasta ahora expuesto, esta reacción es fuertemente
exergónica, por lo que debería cursar espontáneamente.
Sin embargo, la glucosa en presencia de oxígeno es perfectamente
estable y no entra espontáneamente en combustión.
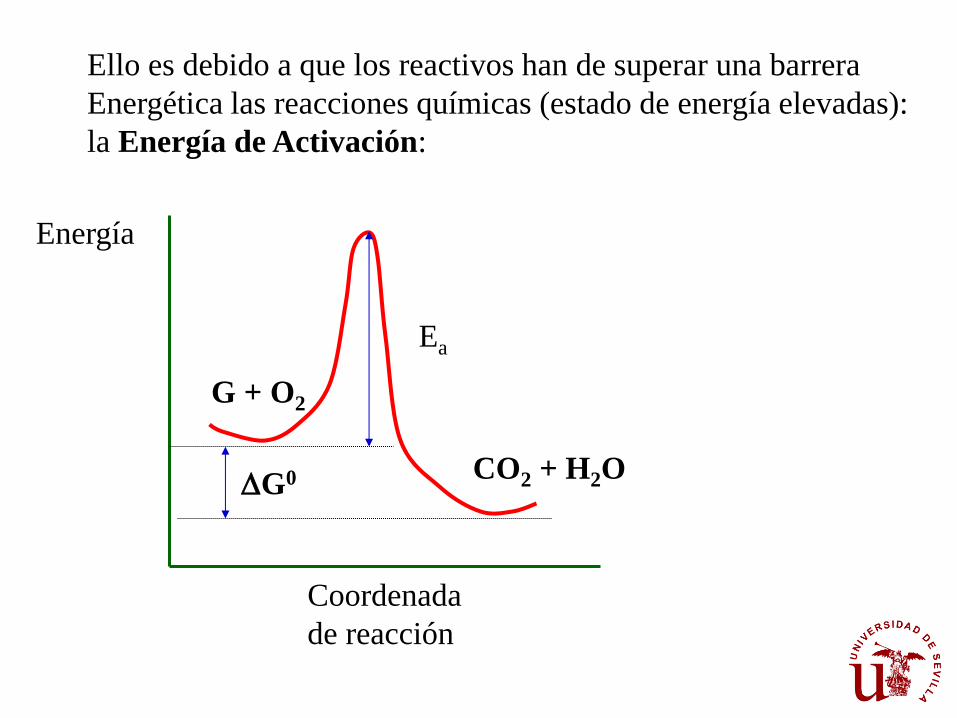
Ello es debido a que los reactivos han de superar una barrera
Energética las reacciones químicas (estado de energía elevadas):
la Energía de Activación:
Energía
Coordenada
de reacción
Ea
G + O2
CO2 + H2O DG0
Progreso de la reacción
S
P
AGo’ DGo´=-RTLnKeq
Progreso de la reacción
S
P
X‡ AG‡
AGo’
S X‡
K++eq =
[X‡]
[S] DG‡=-RTLnK‡eq
AG‡ = Energía de activación
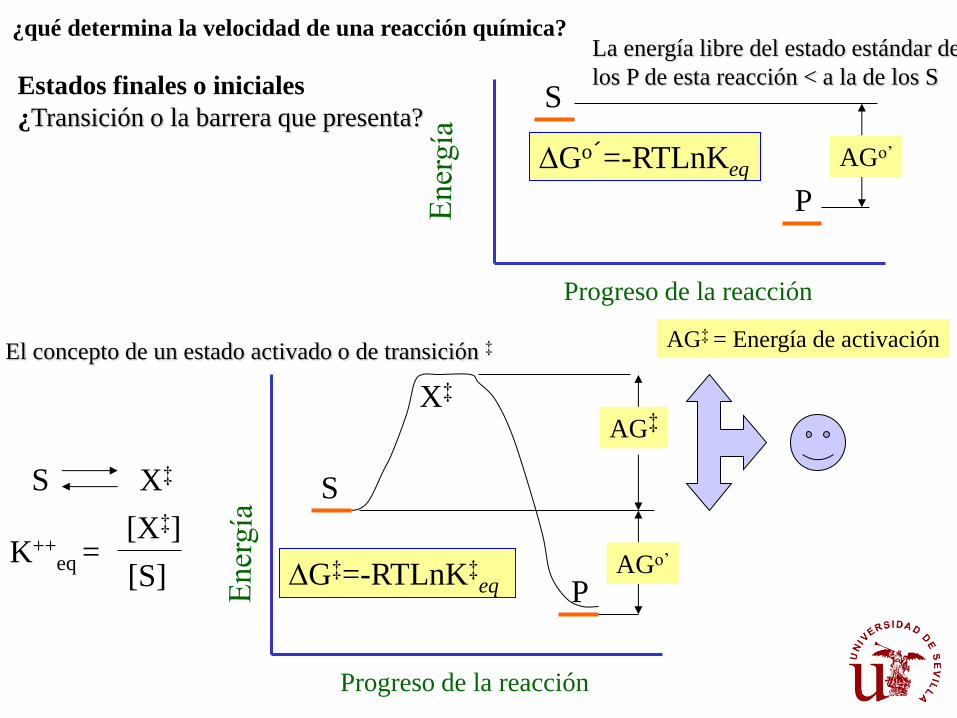
¿qué determina la velocidad de una reacción química? La energía libre del estado estándar de
los P de esta reacción < a la de los S Estados finales o iniciales
¿Transición o la barrera que presenta?
El concepto de un estado activado o de transición ‡
DG‡=-RTLnK‡eq
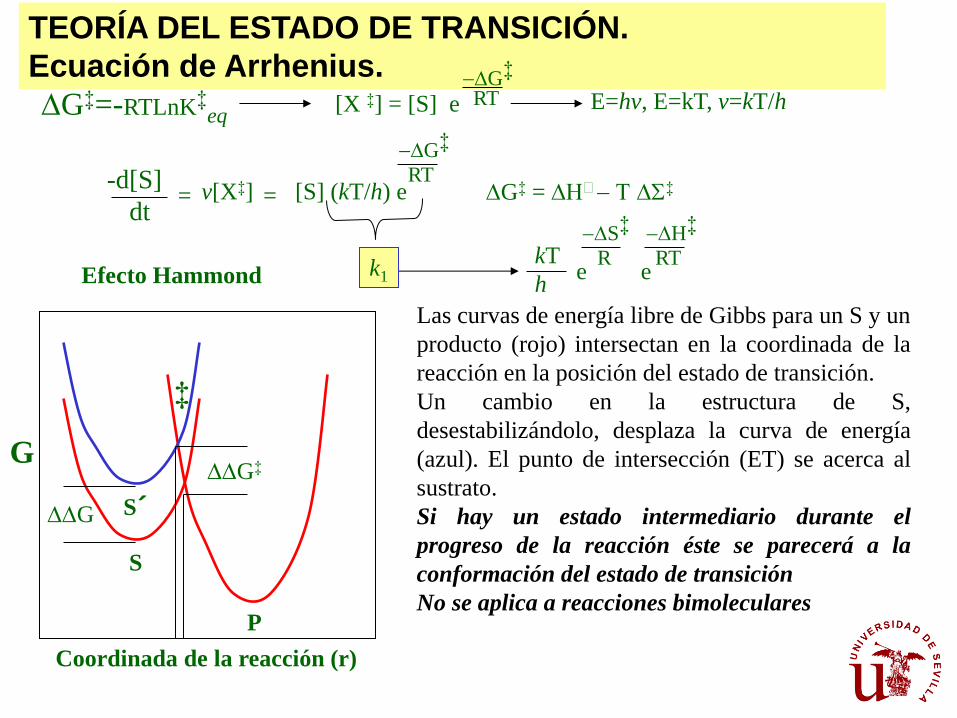
TEORÍA DEL ESTADO DE TRANSICIÓN.
Ecuación de Arrhenius. [X ‡] = [S] e RT
-DG‡
E=hv, E=kT, v=kT/h
-d[S]
dt = = v[X‡] [S] (kT/h) e
RT
-DG‡
k1
DG‡ = DH - DS‡
kT
h e e
R
-DS‡
RT
-DH‡
G
Coordinada de la reacción (r)
DDG‡
S
P
S´ DDG
‡
Efecto Hammond
Las curvas de energía libre de Gibbs para un S y un
producto (rojo) intersectan en la coordinada de la
reacción en la posición del estado de transición.
Un cambio en la estructura de S,
desestabilizándolo, desplaza la curva de energía
(azul). El punto de intersección (ET) se acerca al
sustrato.
Si hay un estado intermediario durante el
progreso de la reacción éste se parecerá a la
conformación del estado de transición
No se aplica a reacciones bimoleculares
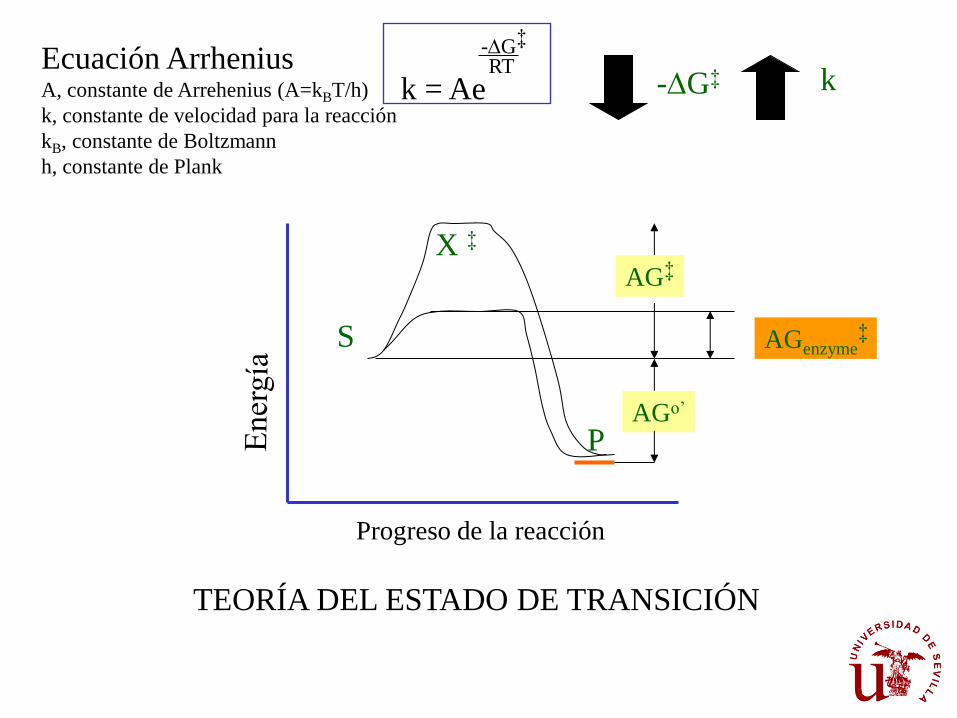
k = Ae
-DG‡
RT Ecuación Arrhenius A, constante de Arrehenius (A=kBT/h)
k, constante de velocidad para la reacción
kB, constante de Boltzmann
h, constante de Plank
-DG‡ k
Progreso de la reacción
S
P
X ‡
AG‡
AGo’
AGenzyme‡
TEORÍA DEL ESTADO DE TRANSICIÓN





























