Embed Size (px)
Citation preview

COVIRO-298; NO. OF PAGES 11
Retrotransposon replication in plantsAlan H Schulman1,2
Available online at www.sciencedirect.com
ScienceDirect
Retrotransposons comprise the bulk of large plant genomes,
replicating via an RNA intermediate whereby the original,
integrated element remains in place. Of the two main orders,
the LTR retrotransposons considerably outnumber the LINEs.
LINEs integrate into target sites simultaneously with the RNA
transcript being copied into cDNA by target-primed reverse
transcription. LTR retrotransposon replication is basically
equivalent to the intracellular phase of retroviral life cycles. The
envelope gene giving extracellular mobility to retroviruses is in
fact widespread in plants and their retrotransposons.
Evolutionary analyses of the retrotransposons and retroviruses
suggest that both form an ancient monophyletic group. The
particular adaptations of LTR retrotransposons to plant life
cycles enabling their success remain to be clarified.
Addresses1 Institute of Biotechnology, Viikki Biocenter, University of Helsinki, P.O.
Box 65, Helsinki FIN-00014, Finland2 Biotechnology and Food Research, MTT Agrifood Research Finland,
Jokioinen FIN-31600, Finland
Corresponding author: Schulman, Alan H ([email protected])
Current Opinion in Virology 2013, 3:xx–yy
This review comes from a themed issue on Virus replication in
animals and plants
Edited by Ben Berkhout and Kuan-Teh Jeang
1879-6257/$ – see front matter, # 2013 Elsevier B.V. All rights
reserved.
http://dx.doi.org/10.1016/j.coviro.2013.08.009
IntroductionThe sequencing and annotation of increasingly large
plant genomes over the past five years has made clear
that they are comprised mostly of transposable elements
(TEs) rather than the genes directing metabolism,
environmental response, and morphogenesis. TE families
with many thousands of members account for 80% or
more of the total DNA of large genomes such as barley,
wheat or maize [1–3]. Even small genomes such as that of
B. distachyon have hundreds of different TE families
containing several to hundreds of members [4��]. While
many genome projects see TEs only as barriers to assem-
bly and inconveniences to gene annotation, their ubiquity
and abundance makes examining their means of replica-
tion and consequences for the genome worthwhile [5�].The manifold consequences of their replication for the
genome and plant include genome size growth [6,7�],insertional mutagenesis [8,9], imposition of epigenetic
Please cite this article in press as: Schulman AH. Retrotransposon replication in plants, Curr Opi
www.sciencedirect.com
silencing on neighbouring genes [10,11], duplication and
transposition of genes or gene segments [12,13�,14], and
function as promoters and regulatory signals [10,15�,16]
The TEs fall into two main groups [17]. Class I contains
the retrotransposons, which first replicate via an RNA
intermediate and then propagate daughter copies in a
‘copy-and-paste’ process, whereas the Class II transpo-
sons do not replicate as part of their transpositional
process, but rather move as DNA usually in a ‘cut-and-
paste’ process. Although the Class I TEs are united by a
life cycle involving the copying of a genomic RNA into
dsDNA by reverse transcriptase (RT), the five orders of
Class I [17] otherwise differ dramatically in their means of
replication. The LTR (Long Terminal Repeat) retro-
transposon order is the most abundant in most plant
genomes [18]. Among the Class I TEs not containing
LTRs, the LINEs (Long Interspersed Nuclear Elements,
Figure 1) are the best studied due to their importance in
mammals [19,20].
Retrotransposons amount to small genomes within the
organismal genome, containing sufficient information to
replicate using the transcription and translation machin-
ery of their ‘host’ to thrive. The viral research com-
munity thereby has come to regard the retrotransposons
as intracellular viruses and named them accordingly
[21,22�]. Hence, the two major superfamilies of LTR
retrotransposons [17], Gypsy and Copia, respectively
belong to the families Metaviridae and Pseudoviridae.
From the perspective of molecular evolution, however,
the retroviruses (family Retroviridae of the virus
nomenclature) are offshoots of retrotransposons [17],
both being derived from cellular replication machinery.
Both viewpoints shed light on understanding of retro-
transposon replication.
Replication of LINEsWith their simple structure (Figure 1), the LINEs are
thought to be some 600 MY old and the most ancient of
eukaryotic retrotransposons, originating in the Pre-cam-
brian [23]. The basic forms specify only RT and endo-
nuclease activities. Use of RT for replication may be a
vestige of the transition from primordial RNA genomes to
the DNA genomes of current biota [24]. The LINEs
occur virtually ubiquitously in the plants [25,26]; all
appear to belong to the clade formed by the LINE-1
(also called L1) element of mammals, the best studied
LINE [27]. LINEs are generally a minor component in
plant genomes compared to animal genomes, LTR retro-
transposons being the major component in plants. How-
ever, in sugar beet (Beta vulgaris) the L1 group is
n Virol (2013), http://dx.doi.org/10.1016/j.coviro.2013.08.009
Current Opinion in Virology 2013, 3:1–11

2 Virus replication in animals and plants
COVIRO-298; NO. OF PAGES 11
Figure 1
5′ UTR 3′ UTR (A)nape ORF1
LINE(a)
(b) LTR retrotransposon
(A)n
SINE
gag ap in Copia
Gypsygag ap rt-rh in
LTR LTR PBS PPT TRIM
LTR
LTRPBS PPT
env
rt-rh
non-coding LARD
ap in BARE2rt-rh(gag)
LTR
rh-rh
Current Opinion in Virology
Main groups of retrotransposons. (a) Autonomous and non-autonomous non-LTR of the LINE order and the non-autonomous SINE order. A grey bar
indicates a non-coding domain. (b) Autonomous and non-autonomous LTR retrotransposons. An LTR retrotransposon comprises: the long terminal
repeats (LTRs); the primer binding site (PBS), which is the (�)-strand priming site for reverse transcription; the polypurine tract (PPT), which is the (+)-
strand priming site for reverse transcription. The PBS and PPT are part of the internal domain, which in autonomous elements includes the protein-
coding ORFs. Below, the major superfamilies Gypsy and Copia. The position of the envelope (env) domain in those Gypsy and Copia clades that
contain it is shown. Bottom, the non-autonomous retrotransposons. BARE2 is an example of a group having a specific deletion that generates a non-
autonomous subfamily (here, of BARE1). LARD elements have a long internal domain with conserved structure but lacking coding capacity. TRIM
elements have virtually no internal domain except for the PBS and PPT signals.
abundant; in lily (Lilium speciosum), it comprises 4% of the
genome and 250 000 copies [28], which is still short of the
517 000 LINE-1 copies that occupy 17% of the human
genome [29].
Replication of LINEs in plants has not been well studied.
Given the similarity of LINE-1 to plant elements and the
detailed analysis of LINE-1 in mammals, LINE-1 repli-
cation in mammals serves as the model for LINEs in
plants. LINE-1 contains an internal Pol II promoter
within the 50 untranslated region (50 UTR) as well as
two open reading frames, ORF1 and ORF2 [30,31]. Being
internal, the LINE promoter propagates with the rest of
the element and is not lost during replication via RNA.
The LTR retrotransposons, generally lacking internal
promoters as detailed below, have a very different and
arguably more complicated way of solving this problem.
In contrast, the transposase of the directly transposing
Please cite this article in press as: Schulman AH. Retrotransposon replication in plants, Curr Opi
Current Opinion in Virology 2013, 3:1–11
‘cut-and-paste’ Class II transposons merely cuts outside
of the Pol II promoter, mobilizing it together with the
transposase ORF. Once transcribed and transported to
the cytoplasm, ORF1 and ORF2 LINEs are translated
(Figure 2). ORF1 encodes the 40 kDa protein ORF1p,
which is essential for replication and functions as a nucleic
acid chaperone and RNA-binding protein [32]. ORF2
encodes a 150 kDa protein, ORF2p, possessing both
endonuclease and RT activities [30].
LINE RT belongs to a large family of enzymes including
the RTs of LTR retrotransposons, retroviruses, and
pararetroviruses, the PLE elements, as well as telo-
merases, all being descended from a common ancestor
[33��]. Consistent with a common origin, LINE RT can
prime reverse transcription from oligonucleotides that
mimic telomere ends [34]. Moreover, primitive LINEs
without the endonuclease function may have been able
n Virol (2013), http://dx.doi.org/10.1016/j.coviro.2013.08.009
www.sciencedirect.com

Retrotransposon replication in plants Schulman 3
COVIRO-298; NO. OF PAGES 11
Figure 2
TTTTTTAAAAA
AAAAA
AAAAA
(1)
(2)
(3)
(4)
(5)
(6)
(7)
5′ UTR 3′ UTR (A)nape ORF1
rh-rh
TTTTTTAAAAA
TTTTTTAAAAA
TTTTTTAAAAA
Current Opinion in Virology
Replication mechanism of a LINE. The element contains ORF1, specifying an RNA-binding protein, and an open reading frame encoding an apurinic
endonuclease (APE) and the reverse transcriptase (RT) - RNAseH (RH) complex. The LINE is transcribed (Step 1), translated (Step 2; only the RT is
shown), assembled into a ribonucleoprotein particle (Step 3), and transported into the nucleus (not shown). The APE nicks the target site, at which
point the RNA anneals (Step 4). The free 30 hydroxyl group of the nicked target is used to prime reverse transcription by target-primed reverse
transcription (Step 5). The other strand of the target DNA is also nicked, and the second strand of the LINE is synthesized by the RT, generating a target
site duplication (Step 6). The process is completed by strand exchange and the new copy is now inserted at the target site (Step 7). The process is
reviewed elsewhere in detail [20,32].
to replicate as telomeres [35]. Furthermore, mobile orga-
nellar and prokaryotic group II introns, thought to be the
ancestors of sliceosomal introns, specify an RT structu-
rally and functionally similar to that of LINE RT [36].
The ORF1p and ORF2p proteins preferentially associate
with the RNAs from which they are translated, forming
ribonucleoprotein (RNP) particles in the cytoplasm
[37��]. The ORF1p may function analogously to GAG
capsid protein of LTR retrotransposons, described below
[32]. To complete the life cycle, the RNP must be moved
back to the nucleus. The details of that process remain
controversial; active RNP transport across the nuclear
membrane may be required or, alternatively, passive
uptake occurs following disruption of the nuclear envel-
ope during cell division [38,39].
Once at the chromosome, integration of LINE elements
into the genome proceeds via target-primed reverse tran-
scription (TPRT), in which the RNA is converted into
cDNA and the cDNA concomitantly inserted into the
genome [32,40]. The endonuclease domain of ORF2p
Please cite this article in press as: Schulman AH. Retrotransposon replication in plants, Curr Opi
www.sciencedirect.com
nicks a target site in the genome, generally TTTT j AA.
The TTTT 30 OH anneals to the polyA tail of LINE
RNA and is then extended by the RT activity of ORF2p,
copying the LINE RNA in the process (Figure 2). The
second-strand cDNA is then copied from the first strand
by a similar process involving nicking of the opposite host
strand, annealing of the nicked 30 end of the genomic
DNA to the first-strand cDNA, and then extension by
RT. Completion of the LINE replication cycle is essen-
tially double-strand break repair via non-homologous end
joining, an error-prone process involving enzymes not
encoded by the TE itself [41].
Replication of SINEsPlant SINEs (Short INterspersed Elements), like those in
other organisms, constitute a diverse and polyphyletic set
of non-autonomous elements, which can be mobilized
and propagated by the enzymes of LINEs [19,42,43]. The
SINEs include various tRNA, rRNA, and other polymer-
ase III transcripts [42,44] ranging from 75 to 662 bp [43].
SINEs derived from tRNA have the tRNA sequence at
their 50 end and homology at their 30 ends to a LINE from
n Virol (2013), http://dx.doi.org/10.1016/j.coviro.2013.08.009
Current Opinion in Virology 2013, 3:1–11

4 Virus replication in animals and plants
COVIRO-298; NO. OF PAGES 11
the same genome, which is thought to provide binding
sites for LINE-encoded proteins. The 30 tails are often
AT-rich, mirroring their origins as reverse-transcribed
transcripts. Found in up to a million copies in mammalian
genomes [44], they are widespread in plants [43,45], and
tend to number in the hundreds to low thousands (SINE
Base, http://sines.eimb.ru/), the SINEs appearing to be
most prevalent in tobacco and other Solanaceae [42].
Replication of SINE elements has been best examined
for Alu, a 7SL-derived element and the most prevalent of
the SINEs in mammalian genomes [31,46]. Following
transcription by Pol III, Alu RNA is localized to RNP
complexes [47], together with the LINE-1 proteins. The
endonuclease and RT activities of ORF2p cooperate to
replicate Alu in a way analogous to LINEs. The poly(A)
tail needed for this process is already present in the
integrated SINE DNA copy. In the plants, replication
of S1, a tRNA-related SINE of Arabidopsis, has been
most closely examined [48]. The processing of its RNA
appears to be very close to that of mammalian 7SL-related
SINEs such as Alu [48].
Replication of LTR retrotransposonsThe structure and replication of plant LTR retrotranspo-
sons is reminiscent of the related Gypsy and Copiaelements of fungi and animals as well as of the retro-
viruses and endogenous retroviruses. The 50 LTR directs
transcription and contains a Pol II promoter. The 30 LTR
provides the signals for RNA termination and polyade-
nylation. LTRs carry response elements directing the
conditions under which retrotransposon replication can
commence. Plant LTR promoters are activated by a
variety of biotic and abiotic stresses as well tissue culture
and plant hormone treatment [49–53].
Transcription by Pol II begins within the 50 LTR, down-
stream from the promoter, and terminates within the 30
LTR before its 30 end. Transcription has been examined
in detail for BARE1 of barley [54] and Tnt1 of tobacco
[55,56]. In barley, multiple pools of polyadenylated and
non-polyadenylated RNAs are produced, respectively for
translation and reverse transcription [57��]. The tran-
scripts of LTR retrotransposons must serve both trans-
lation and reverse transcription. Transcription yields
RNA lacking complete LTRs at either end, because
the promoter and terminator are within the 50 and 30
LTRs respectively. While this does not hinder trans-
lation, the restoration of both LTRs is needed to produce
an integrationally and replicationally competent cDNA.
This is resolved by the complex reverse transcription
mechanism described below.
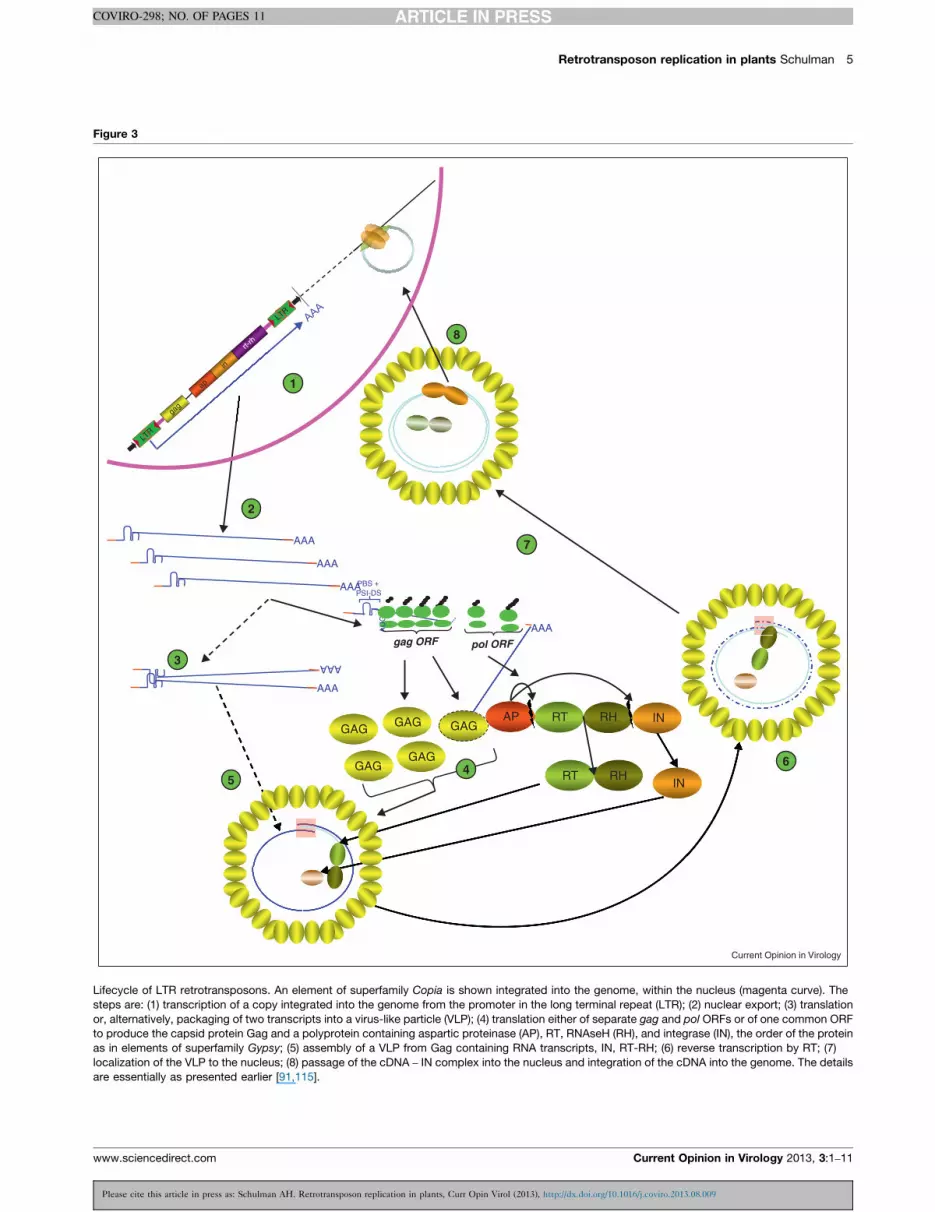
Following transcription, the retrotransposon RNA is
transported to the cytoplasm (Figure 3). For BARE and
most other plant retrotransposons of the Copia super-
family, translation is effected from a single ORF encoding
Please cite this article in press as: Schulman AH. Retrotransposon replication in plants, Curr Opi
Current Opinion in Virology 2013, 3:1–11
both the Gag capsid protein and the polyprotein [58–60].
The polyprotein contains an aspartic proteinase (AP),
integrase (IN), RT, and RNaseH (RH). Antibodies to
BARE1 Gag can detect, in barley and other grasses, both
the polyprotein and the subsequent endoproteolytic clea-
vage products generated by the AP [61,62]. While vir-
tually all animal retrotransposons of the Gypsy superfamily
use �1 translational frameshifting at the end of gag to
translate pol, most plant Gypsy elements appear to employ
a single ORF spanning gag and pol, with a few containing a
+1 frameshift or a stop codon between the two ORFs [60].
In the classical model for retrotransposon and retrovirus
replication (Figure 3), the translated and processed Gag
binds and encapsidates the retrotransposon RNAs into a
virus-like particle (VLP) together with RT-RNAseH and
IN [63,64��]. Two RNA molecules are packaged into each
VLP, directed by the dimerization (DIS) and packaging
(PSI) signals [65]. In plants, these signals have been
identified only for BARE1 and BARE2 of barley [58].
Evidence for formation of virus-like particles has, how-
ever, been demonstrated only for the native BARE1 in
barley [62] and the tobacco Tto1 under an inducible
promoter in Arabidopsis [66].
The encapsidation of the RNA represents a branchpoint
between translation and reverse transcription in the retro-
transposon life cycle (Figure 3). During reverse transcrip-
tion, RNaseH degrades the RNA template. Hence, the
RNA must be translated first if the same molecule is to
serve as a template both for cDNA and protein. Reverse
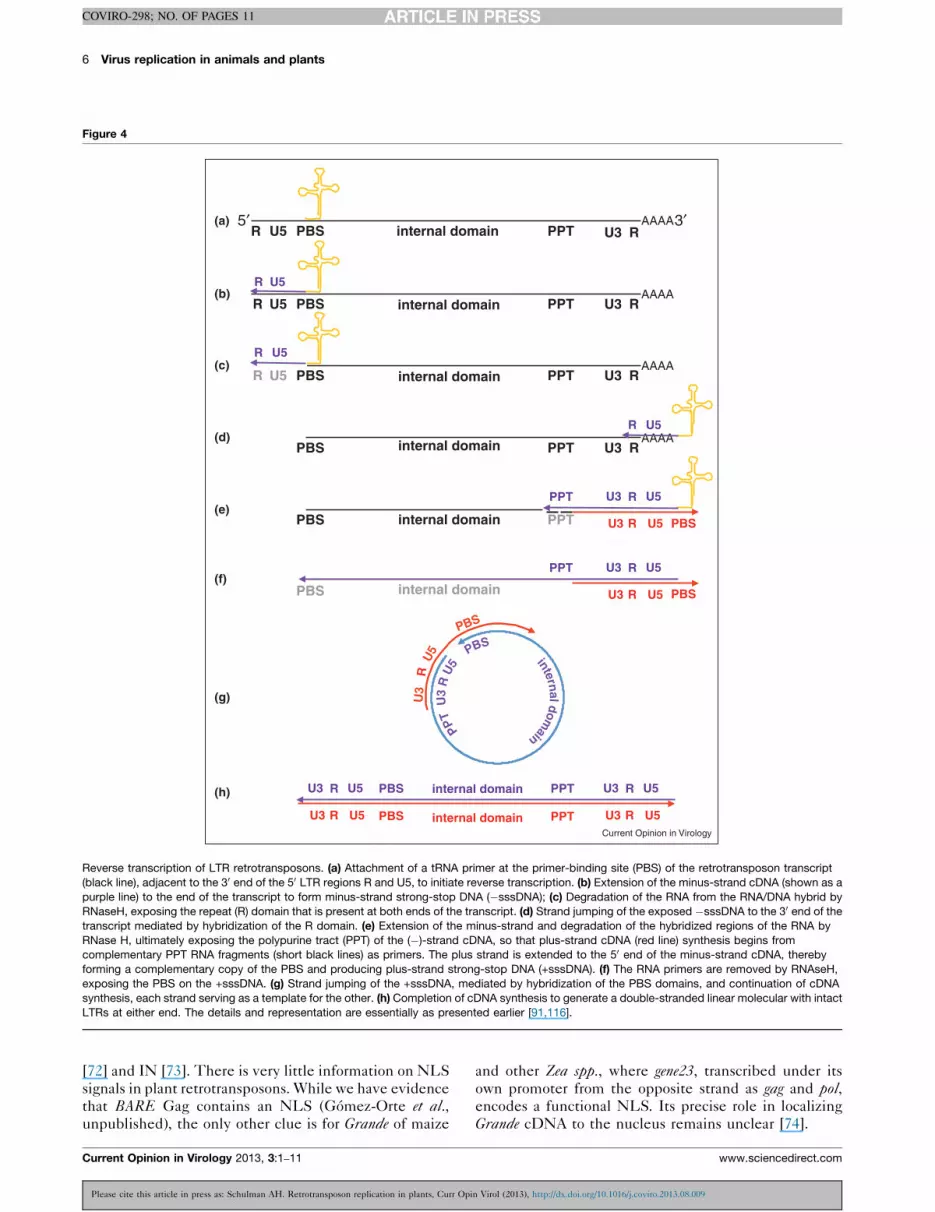
transcription is primed generally from a tRNA matching
the primer binding site (PBS) flanking the 30 end of the 50
LTR (Figures 1 and 4). A (�)-strand strong-stop cDNA is
synthesized, which cannot be further extended without a
jump to the 30 end of the template. Once the (�)-strand
reaches the polypurine tract (PPT) on the 50 side of the 30
LTR within the RNA, synthesis of the (+)-strand cDNA
begins, likewise requiring a template jump in order to
complete (Figure 4). The two strand jumps homogenize
the ends of the final double-stranded cDNA, filling in the
missing 50 and 30 segments of the 50 and 30 LTRs respect-
ively as well as making the two LTRs identical. Detailed
verification of the steps of reverse transcription has not
been made for plant retrotransposons as they have for the
model Ty1 of yeast [67]. However, given both the struc-
tural conservation of plant retrotransposons for features
such as the PBS [68] and the presence of expected cDNA
intermediates in a transgenic test system [69], the mech-
anisms are likely very similar.
In order to be integrated into the chromosome, the retro-
transposon cDNA must find its way into the nucleus
(Figure 3). Nuclear entry is moderated by the nuclear
localization signal (NLS), which in retroviruses can be
found in various retroviral proteins [70,71]. For fungal
retrotransposons, the NLS has been found in both Gag
n Virol (2013), http://dx.doi.org/10.1016/j.coviro.2013.08.009
www.sciencedirect.com
Retrotransposon replication in plants Schulman 5
COVIRO-298; NO. OF PAGES 11
Please cite this article in press as: Schulman AH. Retrotransposon replication in plants, Curr Opin Virol (2013), http://dx.doi.org/10.1016/j.coviro.2013.08.009
Figure 3
AU
G
PBS + PSI-DS
gag ORFAAA
pol ORF
AAA
AAA
AAA
INRHRT
GAG
GAGGAG
GAG
INRHRTAPGAG
AAA
AAA
1
2
3
46
7
8
5
LTR
LTR
gag
ap
in
rt-rh
Current Opinion in Virology
Lifecycle of LTR retrotransposons. An element of superfamily Copia is shown integrated into the genome, within the nucleus (magenta curve). The
steps are: (1) transcription of a copy integrated into the genome from the promoter in the long terminal repeat (LTR); (2) nuclear export; (3) translation
or, alternatively, packaging of two transcripts into a virus-like particle (VLP); (4) translation either of separate gag and pol ORFs or of one common ORF
to produce the capsid protein Gag and a polyprotein containing aspartic proteinase (AP), RT, RNAseH (RH), and integrase (IN), the order of the protein
as in elements of superfamily Gypsy; (5) assembly of a VLP from Gag containing RNA transcripts, IN, RT-RH; (6) reverse transcription by RT; (7)
localization of the VLP to the nucleus; (8) passage of the cDNA – IN complex into the nucleus and integration of the cDNA into the genome. The details
are essentially as presented earlier [91,115].
www.sciencedirect.com Current Opinion in Virology 2013, 3:1–11
6 Virus replication in animals and plants
COVIRO-298; NO. OF PAGES 11
Figure 4
R AAAA
U5 PBS U3 R PPT
U5 R
5′R
(a)
(b)
(c)
(d)
(e)
(f)
(g)
(h)
AAAA U5 PBS PPT internal domain
3′U3 R
AAAA U3 R PPT U5 R PBS
U5 R
AAAA U3 R PPT PBS
U5 R
U3 R PBS
U5 R U3 PPT
PPT U3 R U5
U3 R PBS
U5 R U3 PPT
PBS
U5
U3
U3 R U5
R
internal domain
internal domain
internal domain
internal domain
internal domain
U5 R U3 PPT
U3 R U5
U5 R U3
U3 R U5
PBS
PBS PPT
internal domain
internal domain
PBS
PBS
PPT
U3
R U
5
PBS internal dom
ain
Current Opinion in Virology
Reverse transcription of LTR retrotransposons. (a) Attachment of a tRNA primer at the primer-binding site (PBS) of the retrotransposon transcript
(black line), adjacent to the 30 end of the 50 LTR regions R and U5, to initiate reverse transcription. (b) Extension of the minus-strand cDNA (shown as a
purple line) to the end of the transcript to form minus-strand strong-stop DNA (�sssDNA); (c) Degradation of the RNA from the RNA/DNA hybrid by
RNaseH, exposing the repeat (R) domain that is present at both ends of the transcript. (d) Strand jumping of the exposed �sssDNA to the 30 end of the
transcript mediated by hybridization of the R domain. (e) Extension of the minus-strand and degradation of the hybridized regions of the RNA by
RNase H, ultimately exposing the polypurine tract (PPT) of the (�)-strand cDNA, so that plus-strand cDNA (red line) synthesis begins from
complementary PPT RNA fragments (short black lines) as primers. The plus strand is extended to the 50 end of the minus-strand cDNA, thereby
forming a complementary copy of the PBS and producing plus-strand strong-stop DNA (+sssDNA). (f) The RNA primers are removed by RNAseH,
exposing the PBS on the +sssDNA. (g) Strand jumping of the +sssDNA, mediated by hybridization of the PBS domains, and continuation of cDNA
synthesis, each strand serving as a template for the other. (h) Completion of cDNA synthesis to generate a double-stranded linear molecular with intact
LTRs at either end. The details and representation are essentially as presented earlier [91,116].
[72] and IN [73]. There is very little information on NLS
signals in plant retrotransposons. While we have evidence
that BARE Gag contains an NLS (Gomez-Orte et al.,unpublished), the only other clue is for Grande of maize
Please cite this article in press as: Schulman AH. Retrotransposon replication in plants, Curr Opi
Current Opinion in Virology 2013, 3:1–11
and other Zea spp., where gene23, transcribed under its
own promoter from the opposite strand as gag and pol,encodes a functional NLS. Its precise role in localizing
Grande cDNA to the nucleus remains unclear [74].
n Virol (2013), http://dx.doi.org/10.1016/j.coviro.2013.08.009
www.sciencedirect.com

Retrotransposon replication in plants Schulman 7
COVIRO-298; NO. OF PAGES 11
In contrast to the LINEs and SINEs, integration of
retrotransposons into the genome is accomplished by a
specialized enzyme, integrase. The IN makes staggered
cuts at the target site, trims extra nucleotides from the 30
termini of the LTRs, and also joins the 30 termini to the
free 50 ends at the staggered cuts. The mechanism of
integration appears to be highly conserved among retro-
transposons and retroviruses, with retroviral IN forming
the interpretive model for the others [75]. Moreover, IN
and bacteriophage Mu transposase share the catalytic
DDE motif and other structural features, implying a
common origin [76–78]. Mechanistic studies on retro-
transposon INs have advanced farthest with those of yeast
Ty1 and Ty3, in keeping with the general advanced state
of these systems [79]. Unravelling the target specificity of
Ty3 IN for integration at Pol III transcription initiation
sites has been of particular interest [80�] as has been,
conversely, the question of substrate specificity for Ty1IN [81].
There has been little functional investigation of plant
INs, although structural predictions for the BARE IN
have been made [82] and processing of cDNA by Tnt1 IN
examined [69]. Plant IN studies have focused, instead, on
evolutionary perspectives for the diversity present in
large retrotransposon families [83]. The INs of the wide-
spread chromovirus group of Gypsy elements [84] are
interesting for possession at their C-terminus of chromo-
domains, structurally similar to members of the hetero-
chromatin protein 1 (HP1) family, which can confer
particular integration patterns to the various clades of
chromoviruses [85]. Functional analysis of the chromo-
domain has been made in a yeast system [86]. For plants,
sequence analysis of chromovirus IN has been the
primary approach [85,87–89], although transcription and
propagation of a whole element, LORE1a, has been
examined in the model legume Lotus japonica [90�].
Replication of non-autonomous LTRretrotransposonsAnalyses of plant genomes tend to reveal large numbers
of LTR retrotransposons that contain stop codons inter-
rupting their ORFs, lack one or more coding domains, or
both. These non-autonomous retrotransposons may none-
theless be transcriptionally competent. For non-autonom-
ous LTR retrotransposons, the steps of packaging,
reverse transcription, nuclear localization, and integration
may be blocked by lack of self-encoded proteins. Never-
theless, all steps potentially can be complemented in
trans if a translationally or enzymatically defective retro-
transposon possesses the correct recognition signals for
proteins encoded by autonomous elements [63,91].
Among non-autonomous elements in plants are con-
served, abundant, and insertionally polymorphic families
such as BARE2, which receives Gag, which it cannot
produce, from BARE1 [58]. In contrast, the Large
Please cite this article in press as: Schulman AH. Retrotransposon replication in plants, Curr Opi
www.sciencedirect.com
Retrotransposon Derivative (LARD) elements have
replaced gag and pol with a long internal domain of
conserved RNA structure having no coding capacity
[92]. The Terminal Repeat retrotransposons In Miniature
(TRIMs) have non-coding internal domains reduced to
just 100–300 bp, but are nevertheless abundant, wide-
spread, active, and insertionally polymorphic in plants
[93–95]. The Cassandra family of TRIMs is both inser-
tionally polymorphic, distributed throughout the plant
kingdom, and transcribed by a Pol III promoter from 5S
sequences found in its LTRs [96,97]. Non-autonomous,
structurally conserved retrotransposons such as BARE2,
TRIMs, and Cassandra appear to represent replicational
strategies based on parasitizing proteins from other retro-
transposons. However, if the host retrotransposons pro-
viding proteins in trans lose their activity, their non-
autonomous partners will become inactive. This appears
to have happened to TRIMs in rice [98].
Control of retrotransposon replicationThe complex lifecycles of LINEs and LTR retrotran-
sposons present multiple points for regulation and epi-
genetic silencing of activity in order to control the many
consequences of replication that were mentioned at the
outset. Indeed, plant retrotransposons can be variously
silenced both transcriptionally by DNA methylation
[99��,100] and chromatin modification [101] and post-
transcriptionally [102]. As with viruses, some retrotran-
sposons are able to evade silencing [103��,104�]. More-
over, retrotransposon activity, which can be sharply
tissue-specific [90�,105��] can conversely lead to post-
transcriptional silencing of neighbouring genes
[10,55,106].
ConclusionsAnalysis of retrotransposon replication shows a unity that
transcends both the borders separating the Kingdoms of
eukaryotic evolution and the diverse forms of these
abundant and ancient entities. The replicational mech-
anisms of retroviruses, LTR retrotransposons, LINEs,
and their non-autonomous partners all rely on RT. RT
appears to originate in an ancient family, rvt, able to
incorporate both ribonucleotides and deoxyribonucleo-
tides [33��]. The great parting of the ways between
LINEs and the retroviruses and LTR retrotransposons
was the innovation of IN for integration of a pre-formed
cDNA. The LTRs of retrotransposons and retroviruses
share a universal 50 TG. . ... CA 30 structure within term-
inal inverted repeats, which provide the recognition and
binding sites for this enzyme. Indeed, a recent analysis
lends weight to the view that LTRs arose once, making
the retrotransposons and retroviruses monophyletic [22�].
Although retroviruses and superfamily Gypsy retrotran-
sposons share a common order of their gag and pol genes,
the env gene is generally thought to distinguish the
retroviruses. It encodes the envelope protein enabling
n Virol (2013), http://dx.doi.org/10.1016/j.coviro.2013.08.009
Current Opinion in Virology 2013, 3:1–11

8 Virus replication in animals and plants
COVIRO-298; NO. OF PAGES 11
retroviruses to leave the host cell and infect other cells.
However, env-bearing clades of Gypsy [107] and Copiaelements [108,109] are widespread in plants. The intra-
cellular or extracellular role of their env domains never-
theless remains to be established. Taken together, the
evidence suggests that the viral forms — the retro-
viruses — represent a gain of function within the Gypsyretrotransposons. Indeed, an analysis of gag and polsuggests that the Retroviridae is polyphyletic, having
arisen at least three times from within the Gypsy clade
[110]. In this context, the endogenous retroviruses
(ERVs), which have lost their infectivity, have merely
returned to their old life style [111].
The role of the envelope protein in retroviral infectivity
and its widespread occurrence in plants raises the question
of its role in ‘horizontal transfers’ of plant retrotransposons
between reproductively isolated species. Horizontal trans-
fer is often invoked when a group of retrotransposons is
found in an organism phylogenetically distant from the
clade in which it is expected. For example, TRIM
elements were heretofore found only in plants, but have
been recently discovered in red harvester ants [112].
Claims of horizontal transfer need to be considered in light
of the availability of sequence data bridging the phyloge-
netic divide. Nonetheless, well-supported cases of hori-
zontal transfer of retrotransposons in plants have been
reported [113]. Evidence for horizontal transfer of env-
bearing elements in plants is, however, still lacking, sharply
different from the situation in Drosophila [114].
For the plants, what remains unresolved, aside from many
mechanistic details, is how retrotransposon replication
takes place within the context of the plant life cycle so
that new copies are passed through the germline to the
next generation. The plant life cycle differs greatly from
that of animals or fungi. In plants, the germline is laid
down late in development, following many somatic cell
divisions and the eventual transition of an apical meris-
tem to a floral meristem. The reasons that LTR retro-
transposons in particular have been so successful in plants
remain to be established.
AcknowledgementsThomas Wicker, Francois Sabot, and Jan Buchmann are thanked for usefuldiscussions. Funding from Academy of Finland (Decision 134079) isgratefully acknowledged.
References and recommended readingPapers of particular interest, published within the period of review,have been highlighted as:
� of special interest
�� of outstanding interest
1. International Barley Genome Sequencing Consortium, Mayer KF,Waugh R, Brown JW, Schulman A, Langridge P, Platzer M,Fincher GB, Muehlbauer GJ, Sato K et al.: A physical, genetic andfunctional sequence assembly of the barley genome. Nature2012, 491:711-716.
Please cite this article in press as: Schulman AH. Retrotransposon replication in plants, Curr Opi
Current Opinion in Virology 2013, 3:1–11
2. Schnable PS, Ware D, Fulton RS, Stein JC, Wei F, Pasternak S,Liang C, Zhang J, Fulton L, Graves TA et al.: The B73 maizegenome: complexity, diversity, and dynamics. Science 2009,326:1112-1115.
3. Tenaillon MI, Hufford MB, Gaut BS, Ross-Ibarra J: Genome sizeand transposable element content as determined by high-throughput sequencing in maize and Zea luxurians. GenomeBiol Evol 2011, 3:219-229.
4.��
International Brachypodium Initiative: Genome sequencing andanalysis of the model grass Brachypodium distachyon. Nature2010, 463:763-768.
Using a high-quality assembly, this study was able to show differingretrotransposon dynamics between chromosomes and regions andestablish how the genome stays small despite active retrotransposonfamilies.
5.�
Vesely P, Bures P, Smarda P, Pavlıcek T: Genome size and DNAbase composition of geophytes: the mirror of phenology andecology? Ann Bot 2012, 109:65-75.
The authors revisit the long-debated question of the selective advantageof genome size, to which retrotransposon replication contributes, thougha survey of genome size, morphology, and anatomy Increased genomesize among 219 land plants. Associations were found between genomesize, flowering time, growth in humid conditions, and stomatal size.
6. Estep MC, DeBarry JD, Bennetzen JL: The dynamics of LTRretrotransposon accumulation across 25 million years ofpanicoid grass evolution. Heredity 2013, 110:194-204.
7.�
Park MJP, Kim S, Kwon JK, Park HM, Bae IH, Yang TJ, Lee YH,Kang BC, Choi D: Evolution of the large genome in Capsicumannuum occurred through accumulation of single-type longterminal repeat retrotransposons and their derivatives. Plant J2012, 69:1018-1029.
A demonstration of how a single family of retrotransposons, Del of theGypsy superfamily, expanded the pepper genome to its current size of2700 Mb.
8. Xiao H, Jiang N, Schaffner E, Stockinger EJ, van der Knaap E: Aretrotransposon-mediated gene duplication underliesmorphological variation of tomato fruit. Science 2008,319:1527-1530.
9. Duangpan S, Zhang W, Wu Y, Jansky SH, Jiang J: Insertionalmutagenesis using Tnt1 retrotransposon in potato. PlantPhysiol 2013, 163:21-29 10.1104/pp.1113.221903.
10. Kashkush K, Khasdan V: Large-scale survey of cytosinemethylation of retrotransposons and the impact of readouttranscription from long terminal repeats on expression ofadjacent rice genes. Genetics 2007, 177:1975-1985.
11. Lisch D, Bennetzen JL: Transposable element origins ofepigenetic gene regulation. Curr Opin Plant Biol 2011, 14:156-161.
12. Jiang N, Ferguson AA, Slotkin RK, Lisch D: Pack-Mutator-liketransposable elements (Pack-MULEs) induce directionalmodification of genes through biased insertion and DNAacquisition. Proc Natl Acad Sci U S A 2011, 108:1537-1542.
13.�
Barbaglia AM, Klusman KM, Higgins J, Shaw JR, Hannah LC,Lal S: Gene capture by Helitron transposons reshuffles thetranscriptome of maize. Genetics 2012, 190:965-975.
The first evidence that Helitrons not only modify genes by insertiontogether with exon segments, but also produce use transcriptional read-through into exons outside the gene in which they are inserted. In maize,at least 11 000 new transcripts are expressed by Helitrons.
14. Wicker T, Buchmann JP, Keller B: Patching gaps in plantgenomes results in gene movement and erosion of colinearity.Genome Res 2010, 20:1229-1237.
15.�
Studer A, Zhao Q, Ross-Ibarra J, Doebley J: Identification of afunctional transposon insertion in the maize domesticationgene tb1. Nat Genet 2011, 43:1160-1163.
A clear case where insertion of a transposable element (Hopscotch)caused a change of function of crucial importance for agriculture. Theinsertion into the domestication gene of teosinte, ancestor of maizepredated domestication by �10 000 years and provided the plant formselected by early farmers.
16. Xu Z, Rafi S, Ramakrishna W: Polymorphisms and evolutionaryhistory of retrotransposon insertions in rice promoters.Genome 2011, 54:629-638.
n Virol (2013), http://dx.doi.org/10.1016/j.coviro.2013.08.009
www.sciencedirect.com

Retrotransposon replication in plants Schulman 9
COVIRO-298; NO. OF PAGES 11
17. Wicker T, Sabot F, Hua-Van A, Bennetzen J, Capy P, Chalhoub B,Flavell AJ, Leroy P, Morgante M, Panaud O et al.: A unifiedclassification system for eukaryotic transposable elements.Nat Rev Genet 2007, 8:973-982.
18. Gaut BS, Ross-Ibarra J: Selection on major components ofangiosperm genomes. Science 2008, 320:484-486.
19. Goodier JL, Kazazian HHJ: Retrotransposons revisited: therestraint and rehabilitation of parasites. Cell 2008, 135:23-35.
20. Han JS, Boeke JD: LINE-1 retrotransposons: modulators ofquantity and quality of mammalian gene expression?BioEssays 2005, 27:775-784.
21. King AMQ, Adams MJ, Carstens EB, Lefkowitz EJ (Eds): Virustaxonomy: classification and nomenclature of viruses: Ninth Reportof the International Committee on Taxonomy of Viruses. ElsevierAcademic Press; 2012.
22.�
Benachenhou F, Sperber GO, Bongcam-Rudloff E, Andersson G,Boeke JD, Blomberg J: Conserved structure and inferredevolutionary history of long terminal repeats (LTRs). Mob DNA2013, 4:5 http://dx.doi.org/10.1186/1759-8753-4-5.
Ambitious attempt to use Hidden Markov models on LTRs to infer theevolutionary origin of all LTR retrotransposons and retroviruses. The datasupport a monophyletic origin for all, from within the non-LTR transpo-sons related to LINE and Penelope elements.
23. Eickbush TH, Jamburuthugoda VK: The diversity ofretrotransposons and the properties of their reversetranscriptases. Virus Res 2008, 34:221-234.
24. Brosius J: Echoes from the past—are we still in an RNP world?Cytogenet Genome Res 2005, 110:8-24.
25. Noma K, Ohtsubo E, Ohtsubo H: Non-LTR retrotransposons(LINEs) as ubiquitous components of plant genomes. Mol GenGenet 1999, 261:71-79.
26. Schmidt T: LINEs, SINEs and repetitive DNA: non-LTRretrotransposons in plant genomes. Plant Mol Biol 1999,40:903-910.
27. Hancks DC, Kazazian HHJ: Active human retrotransposons:variation and disease. Curr Opin Genet Dev 2012, 22:191-203.
28. Wenke T, Holtgrawe D, Horn AV, Weisshaar B, Schmidt T: Anabundant and heavily truncated non-LTR retrotransposon(LINE) family in Beta vulgaris. Plant Mol Biol 2009, 71:585-597.
29. Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, Baldwin J,Devon K, Dewar K, Doyle M, FitzHugh W et al.: Initial sequencingand analysis of the human genome. Nature 2001, 409:860-921.
30. Beck CR, Garcia-Perez JL, Badge RM, Moran JV: LINE-1elements in structural variation and disease. Annu Rev GenomHum Genet 2011, 12:187-215.
31. Kroutter EN, Belancio VP, Wagstaff BJ, Roy-Engel AM: The RNApolymerase dictates ORF1 requirement and timing of LINE andSINE retrotransposition. PLoS Genet 2009, 5:e1000458 http://dx.doi.org/10.1371/journal.pgen.1000458.
32. Martin SL: Nucleic acid chaperone properties of ORF1p from thenon-LTR retrotransposon, LINE-1. RNA Biol 2010, 7:706-711.
33.��
Gladyshev EA, Arkhipova IR: A widespread class of reversetranscriptase-related cellular genes. Proc Natl Acad Sci U S A2011, 108:20311-20316.
The authors have discovered a group of cellular genes, rvt, that are similarto RT of LINEs, which are widely distributed in protists, fungi, animals, andplants. The rvt genes are evolving under purifying selection, and thecorresponding protein from at least one, from Neurospora crassa is ableto polymerize NTPs as well as dNTPs in vitro. The findings indicate thatRTs play a role in cellular processes and raise evolutionary questionsconcerning the origins of RT and the evolution of retrotransposons.
34. Kopera HC, Moldovan JB, Morrish TA, Garcia-Perez JL, Moran JV:Similarities between long interspersed element-1 (LINE-1)reverse transcriptase and telomerase. Proc Natl Acad Sci U S A2011, 108:20345-20350.
35. Morrish TA, Garcia-Perez JL, Stamato TD, Taccioli GE,Sekiguchi J, Moran JV: Endonuclease-independent LINE-1retrotransposition at mammalian telomeres. Nature 2007,446:208-212.
Please cite this article in press as: Schulman AH. Retrotransposon replication in plants, Curr Opi
www.sciencedirect.com
36. Gu SQ, Cui X, Mou S, Mohr S, Yao J, Lambowitz AM: Geneticidentification of potential RNA-binding regions in a group IIintron-encoded reverse transcriptase. RNA 2010, 16:732-747.
37.��
Doucet AJ, Hulme AE, Sahinovic E, Kulpa DA, Moldovan JB,Kopera HC, Athanikar JN, Hasnaoui M, Bucheton A, Moran JVet al.: Characterization of LINE-1 ribonucleoprotein particles.PLoS Genet 2010, 7:6 http://dx.doi.org/10.1371/journal.pgen.1001150.
The authors have developed a system for tagging L1 LINE components todissect the retrotransposition process on the cellular level. Their methodswill help overcome the difficulty of following replication of the �80–100active L1 elements among the half-million copies in the human genome.
38. Mathias SL, Scott AF, Kazazian HH Jr, Boeke JD, Gabriel A:Reverse transcriptase encoded by a human transposableelement. Science 1991, 254:1808-1810.
39. Kubo S, Seleme MC, Soifer HS, Perez JL, Moran JV, Kazazian HHJr, Kasahara N: L1 retrotransposition in nondividing andprimary human somatic cells. Proc Natl Acad Sci U S A 2006,103:8036-8041.
40. Cost GJ, Feng Q, Jacquier A, Boeke JD: Human L1 elementtarget-primed reverse transcription in vitro. EMBO J 2002,21:5899-5910.
41. Puchta H: The repair of double-strand breaks in plants:mechanisms and consequences for genome evolution. J ExpBot 2005, 56:1-14.
42. Wenke T, Dobel T, Sorensen TR, Junghans H, Weisshaar B,Schmidt T: Targeted identification of short interspersednuclear element families shows their widespread existenceand extreme heterogeneity in plant genomes. Plant Cell 2011,23:3117-3128.
43. Vassetzky NS, Kramerov DA: SINEBase: a database and tool forSINE analysis. Nucleic Acids Res 2013, 41:D83-D89.
44. Kramerov D, Vassetzky N: Short retroposons in eukaryoticgenomes. Int Rev Cytol 2005, 247:165-221.
45. Deragon J, Zhang X: Short Interspersed Elements (SINEs) inplants: origin, classification, and use as phylogenetic markers.Sys Biol 2006, 55:949-956.
46. Dewannieux M, Esnault C, Heidmann T: LINE-mediatedretrotransposition of marked Alu sequences. Nat Genet 2003,35:41-48.
47. West N, Roy-Engel AM, Imataka H, Sonenberg N, Deininger P:Shared protein components of SINE RNPs. J Mol Biol 2002,321:423-432.
48. Pelissier T, Bousquet-Antonelli C, Lavie L, Deragon JM: Synthesisand processing of tRNA-related SINE transcripts inArabidopsis thaliana. Nucleic Acids Res 2004, 32:3957-3966.
49. Ramallo E, Kalendar R, Schulman AH, Martınez-Izquierdo JA:Reme1, a Copia retrotransposon in melon, is transcriptionallyinduced by UV light. Plant Mol Biol 2008, 66:137-150.
50. Salazar M, Gonzalez E, Casaretto JA, Casacuberta JM, Ruiz-Lara S: The promoter of the TLC1.1 retrotransposon fromSolanum chilense is activated by multiple stress-relatedsignaling molecules. Plant Cell Rep 2007, 26:1861-1868.
51. Ansari KI, Walter S, Brennan JM, Lemmens M, Kessans S,McGahern A, Egan D, Doohan FM: Retrotransposon and geneactivation in wheat in response to mycotoxigenic and non-mycotoxigenic-associated Fusarium stress. Theor Appl Genet2007, 114:927-937.
52. Grandbastien MA, Audeon C, Bonnivard E, Casacuberta JM,Chalhoub B, Costa AP, Le QH, Melayah D, Petit M, Poncet C et al.:Stress activation and genomic impact of Tnt1 retrotransposonsin Solanaceae. Cytogenet Genome Res 2005, 110:229-241.
53. Kalendar R, Tanskanen J, Immonen S, Nevo E, Schulman AH:Genome evolution of wild barley (Hordeum spontaneum) byBARE-1 retrotransposon dynamics in response to sharpmicroclimatic divergence. Proc Natl Acad Sci U S A 2000,97:6603-6607.
54. Chang W, Schulman AH: BARE retrotransposons producemultiple groups of rarely polyadenylated transcripts
n Virol (2013), http://dx.doi.org/10.1016/j.coviro.2013.08.009
Current Opinion in Virology 2013, 3:1–11

10 Virus replication in animals and plants
COVIRO-298; NO. OF PAGES 11
from two differentially regulated promoters. Plant J2008, 56:40-50.
55. Hernandez-Pinzon I, de Jesus E, Santiago N, Casacuberta JM:The frequent transcriptional readthrough of the tobacco Tnt1retrotransposon and its possible implications for the controlof resistance genes. J Mol Evol 2009, 68:269-278.
56. Beguiristain T, Grandbastien MA, Puigdomenech P,Casacuberta JM: Three Tnt1 subfamilies show different stress-associated patterns of expression in tobacco. Consequencesfor retrotransposon control and evolution in plants. PlantPhysiol 2001, 127:212-221.
57.��
Chang W, Jaaskelainen M, Li S-P, Schulman AH: BAREretrotransposons are translated and replicated via distinctRNA pools. PLoS ONE 2013, 8:e72270 http://dx.doi.org/10.1371/journal.pone.0072270.
The first demonstration of distinct RNA pools for translation and tran-scription in retrotransposon replication. The BARE retrotransposon pro-duces three classes of transcripts from one ORF. The capped andpolyadenylated mRNAs are translated, of which some are spliced toproduce only Gag; RNAs that are not capped or polyadenylated arepackaged for reverse transcription.
58. Tanskanen JA, Sabot F, Vicient C, Schulman AH: Life withoutGAG: the BARE-2 retrotransposon as a parasite’s parasite.Gene 2007, 390:166-174.
59. Moisy C, Garrison KE, Meredith CP, Pelsy F: Characterization often novel Ty1/copia-like retrotransposon families of thegrapevine genome. BMC Genomics 2008, 9:469 http://dx.doi.org/10.1186/1471-2164-9-469.
60. Gao X, Havecker ER, Baranov PV, Atkins JF, Voytas DF:Translational recoding signals between gag and pol in diverseLTR retrotransposons. RNA 2003, 9:1422-1430.
61. Vicient CM, Jaaskelainen M, Kalendar R, Schulman AH: Activeretrotransposons are a common feature of grass genomes.Plant Physiol 2001, 125:1283-1292.
62. Jaaskelainen M, Mykkanen A-H, Arna T, Vicient C, Suoniemi A,Kalendar R, Savilahti H, Schulman AH: Retrotransposon BARE-1: expression of encoded proteins and formation of virus-likeparticles in barley cells. Plant J 1999, 20:413-422.
63. Sabot F, Schulman AH: Parasitism and the retrotransposon lifecycle in plants: a hitchhiker’s guide to the genome. Heredity2006, 97:381-388.
64.��
Lee SK, Potempa M, Swanstrom R: The choreography of HIV-1proteolytic processing and virion assembly. J Biol Chem 2012,287:40867-40874.
This is the best current review processing and assembly of HIV-1 virions,summing the results of the last 25 years of research. The HIV-1 systemforms a model for understanding the life cycle of plant LTR retrotran-sposons.
65. Lanchy JM, Ivanovitch JD, Lodmell JS: A structural linkagebetween the dimerization and encapsidation signals in HIV-2leader RNA. RNA 2003, 9:1007-1018.
66. Bohmdorfer G, Luxa K, Frosch A, Garber K, Tramontano A,Jelenic S, Weber M, Bachmair A: Virus-like particle formationand translational start site choice of the plant retrotransposonTto1. Virology 2008, 372:437-446.
67. Pochart P, Agoutin B, Rousset S, Chanet R, Doroszkiewicz V,Heyman T: Biochemical and electron microscope analyses ofthe DNA reverse transcripts present in the virus-like particlesof the yeast transposon Ty1. Identification of a second originof Ty1 DNA plus strand synthesis. Nucleic Acids Res 1993,21:3513-3520.
68. Kalendar R, Antonius K, Smykal P, Schulman AH: iPBS: auniversal method for DNA fingerprinting and retrotransposonisolation. Theor Appl Genet 2010, 121:1419-1430.
69. Feuerbach F, Drouaud J, Lucas H: Retrovirus-like endprocessing of the tobacco Tnt1 retrotransposon linearintermediates of replication. J Virol 1997, 71:4000-4005.
70. Suzuki Y, Craigie R: The road to chromatin—nuclear entry ofretroviruses. Nat Rev Microbiol 2007, 5:187-196.
Please cite this article in press as: Schulman AH. Retrotransposon replication in plants, Curr Opi
Current Opinion in Virology 2013, 3:1–11
71. Mullers E, Stirnnagel K, Kaulfuss S, Lindemann D: Prototypefoamy virus gag nuclear localization: a novel pathway amongretroviruses. J Virol 2011, 85:9276-9285.
72. Kim MK, Claiborn KC, Levin HL: The long terminal repeat-containing retrotransposon Tf1 possesses amino acids in gagthat regulate nuclear localization and particle formation. J Virol2005, 79:9540-9555.
73. Kenna MA, Brachmann CB, Devine SE, Boeke JD: Invading theyeast nucleus: a nuclear localization signal at the C terminusof Ty1 integrase is required for transposition in vivo. Mol CellBiol 1998, 18:1115-1124.
74. Gomez-Orte E, Vicient CM, Martınez-Izquierdo JA: Granderetrotransposons contain an accessory gene in the unusuallylong 30-internal region that encodes a nuclear proteintranscribed from its own promoter. Plant Mol Biol 2013, 81:541-551.
75. Krishnan L, Engelman A: Retroviral integrase proteins and HIV-1DNA integration. J Biol Chem 2012, 287:40858-40866.
76. Hickman AB, Chandler M, Dyda F: Integrating prokaryotes andeukaryotes: DNA transposases in light of structure. Crit RevBiochem Mol Biol 2010, 45:50-69.
77. Montano SP, Rice PA: Moving DNA around: DNA transpositionand retroviral integration. Curr Opin Struct Biol 2011, 21:370-378.
78. Montano SP, Pigli YZ, Rice PA: The m transpososome structuresheds light on DDE recombinase evolution. Nature 2012,491:413-417.
79. Lesage P, AL T: Happy together: the life and times of Tyretrotransposons and their hosts. Cytogenet Genome Res 2005,110:70-90.
80.�
Qi X, Vargas E, Larsen L, Knapp W, Hatfield GW, Lathrop R,Sandmeyer S: Directed DNA shuffling of retrovirus andretrotransposon integrase protein domains. PLoS ONE 2013,8:e63957 http://dx.doi.org/10.1371/journal.pone.0063957.
To address why retrotransposons can be display greater integration sitespecificity than do retroviruses, the authors constructed shuffled amino-terminal, catalytic core, and carboxyl-terminal domains of the integrasegene from HIV, prototype foamy virus, and retrotransposon Ty3. In vitroassays produced results consistent with insertion specificities observedin vivo.
81. Friedl AA, Kiechle M, Maxeiner HG, Schiestl RH, Eckardt-Schupp F: Ty1 integrase overexpression leads to integrationof non-Ty1 DNA fragments into the genome ofSaccharomyces cerevisiae. Mol Genet Genomics 2010,284:231-242.
82. Suoniemi A, Tanskanen J, Pentikainen O, Johnson MS,Schulman AH: The core domain of retrotransposon integrase inHordeum: predicted structure and evolution. Mol Biol Evol1998, 15:1135-1144.
83. Gomez E, Schulman AH, Martınez-Izquierdo JA, Vicient CM:Integrase diversity and transcription of the maizeretrotransposon Grande. Genome 2006, 49:558-562.
84. Gorinsek B, Gubensek F, Kordis D: Phylogenomic analysis ofchromoviruses. Cytogenet Genome Res 2005, 110:543-552.
85. Weber B, Heitkam T, Holtgrawe D, Weisshaar B, Minoche AE,Dohm JC, Himmelbauer H, Schmidt T: Highly diversechromoviruses of Beta vulgaris are classified bychromodomains and chromosomal integration. Mob DNA2013, 4:8 http://dx.doi.org/10.1186/1759-8753-1184-1188.
86. Nakayashiki H, Awa T, Tosa Y, Mayama S: The C-terminalchromodomain-like module in the integrase domain is crucialfor high transposition efficiency of the retrotransposonMAGGY. FEBS Lett 2005, 579:488-492.
87. Wolfgruber TK, Sharma A, Schneider KL, Albert PS, Koo DH, Shi J,Gao Z, Han F, Lee H, Xu R et al.: Maize centromere structure andevolution: sequence analysis of centromeres 2 and 5 revealsdynamic Loci shaped primarily by retrotransposons. PLoSGenetics 2009, 5:e1000743 http://dx.doi.org/10.1371/journal.pgen.1000743.
n Virol (2013), http://dx.doi.org/10.1016/j.coviro.2013.08.009
www.sciencedirect.com

Retrotransposon replication in plants Schulman 11
COVIRO-298; NO. OF PAGES 11
88. Sharma A, Presting GG: Centromeric retrotransposon lineagespredate the maize/rice divergence and differ in abundanceand activity. Mol Genet Genomics 2008, 279:133-147.
89. Neumann P, Navratilova A, Koblızkova A, Kejnovsky E, Hribova E,Hobza R, Widmer A, Dolezel J, Macas J: Plant centromericretrotransposons: a structural and cytogenetic perspective.Mob DNA 2011, 2:4 http://dx.doi.org/10.1186/1759-8753-2-4.
90.�
Fukai E, Umehara Y, Sato S, Endo M, Kouchi H, Hayashi M,Stougaard J, Hirochika H: Derepression of the plantChromovirus LORE1 induces germline transposition inregenerated plants. PLoS Genet 2010, 6:e1000868 http://dx.doi.org/10.1371/journal.pgen.1000868.
Activity of chromovirus group of Gypsy retrotransposons had not beenanalyzed in depth before. The authors show that chromovirus LORE1afrom the model plant Lotus japonica replicates in the male germlinefollowing loss of epigenetic silencing in tissue culture. No strong insertionsite specificity was found, raising the potential for use of LORE1a as agenetic tool.
91. Schulman AH: Hitching a ride: nonautonomousretrotransposons and parasitism as a lifestyle. In PlantTransposable Elements, Topics in Current Genetics, vol 24. Editedby Grandbastien M-A, Casacuberta JM. Springer Verlag; 2012:71-88.
92. Kalendar R, Vicient CM, Peleg O, Anamthawat-Jonsson K,Bolshoy A, Schulman AH: LARD retroelements: novel, non-autonomous components of barley and related genomes.Genetics 2004, 166:1437-1450.
93. Witte CP, Le QH, Bureau T, Kumar A: Terminal-repeatretrotransposons in miniature (TRIM) are involved inrestructuring plant genomes. Proc Natl Acad Sci U S A 2001,98:13778-13783.
94. Kwon SJ, Kim DH, Lim MH, Long Y, Meng JL, Lim KB, Kim JA,Kim JS, Jin M, Kim HI et al.: Terminal repeat retrotransposon inminiature (TRIM) as DNA markers in Brassica relatives. MolGenet Genom 2007, 278:361-370.
95. Kraitshtein Z, Yaakov B, Khasdan V, Kashkush K: Genetic andepigenetic dynamics of a retrotransposon afterallopolyploidization of wheat. Genetics 2010, 186:801-812.
96. Antonius-Klemola K, Kalendar R, Schulman AH: TRIMretrotransposons occur in apple and are polymorphicbetween varieties but not sports. Theor Appl Genet 2006,112:999-1008.
97. Kalendar R, Tanskanen JA, Chang W, Antonius K, Sela H, Peleg P,Schulman AH: Cassandra retrotransposons carryindependently transcribed 5S RNA. Proc Natl Acad Sci U S A2008, 105:5833-5838.
98. Baruch O, Kashkush K: Analysis of copy-number variation,insertional polymorphism, and methylation status of thetiniest class I (TRIM) and class II (MITE) transposable elementfamilies in various rice strains. Plant Cell Rep 2012, 31:885-893.
99.��
Cui X, Jin P, Cui X, Gu L, Lu Z, Xue Y, Wei L, Qi J, Song X, Luo Met al.: Control of transposon activity by a histone H3K4demethylase in rice. Proc Natl Acad Sci U S A 2013, 110:1953-1958.
The first report that histone demethylation can be involved in TE silencing.Here, elevated levels of H3K4me3 caused through decreased activity ofrice protein JMJ703, an H3K4-specific demethylase, led to increasedexpression of many genes and reactivation of two families of non-LTRretrotransposons. However, other TEs silenced by other epigenetic path-ways were not affected.
100. Lee TF, Gurazada SG, Zhai J, Li S, Simon SA, Matzke MA, Chen X,Meyers BC: RNA polymerase V-dependent small RNAs inArabidopsis originate from small, intergenic loci includingmost SINE repeats. Epigenetics 2012, 7:781-795.
101. Eichten SR, Ellis NA, Makarevitch I, Yeh CT, Gent JI, Guo L,McGinnis KM, Zhang X, Schnable PS, Vaughn MW et al.:Spreading of heterochromatin is limited to specific families ofmaize retrotransposons. PLoS Genet 2012, 8:e1003127 http://dx.doi.org/10.1371/journal.pgen.1003127.
Please cite this article in press as: Schulman AH. Retrotransposon replication in plants, Curr Opi
www.sciencedirect.com
102. McCue AD, Nuthikattu S, Reeder SH, Slotkin RK: Geneexpression and stress response mediated by the epigeneticregulation of a transposable element small RNA. PLoS Genet2012, 8:e1002474 http://dx.doi.org/10.1371/journal.pgen.1002474.
103��
Marı-Ordonez A, Marchais A, Etcheverry M, Martin A, Colot V,Voinnet O: Reconstructing de novo silencing of an active plantretrotransposon. Nat Genet 2013, 45:1029-1039 10.1038/ng.2703.
An innovative approach using recombinant inbred Arabidopsis thalianaepigenomes (epiRILs) to track the invasion, activity, proliferation andsilencing of the retrotransposon Evade EVD.
104�
Hernandez-Pinzon I, Cifuentes M, Henaff E, Santiago N,Espinas ML, Casacuberta JM: The Tnt1 retrotransposonescapes silencing in tobacco, its natural host. PLoS ONE 2012,7:e33816 http://dx.doi.org/10.1371/journal.pone.0033816.
The authors establish that the well-studied Tnt1 retrotransposon oftobacco remains active even when Tnt1 promoters driving transgenesbecome silenced by methylation.
105��
Jaaskelainen M, Chang W, Moisy C, Schulman AH:Retrotransposon BARE displays strong tissue-specificdifferences in expression. New Phytol 2013. in press.
The authors address retrotransposon BARE replication at the RNA andprotein levels through immunolocalization and in situ hybridization. BAREprotein and RNA expression are seen in apical meristems only followingfloral transition, arguing that this way to pass new copies into gameteshelps explain the replicative success of BARE.
106. McCue AD, Nuthikattu S, Slotkin RK: Genome-wideidentification of genes regulated in trans by transposableelement small interfering RNAs. RNA Biol 2013, 10:76-92.
107. Vicient CM, Kalendar R, Schulman AH: Envelope-containingretrovirus-like elements are widespread, transcribed andspliced, and insertionally polymorphic in plants. Genome Res2001, 11:2041-2049.
108. Havecker ER, Gao X, Voytas DF: The Sireviruses, a plant-specific lineage of the Ty1/copia retrotransposons, interactwith a family of proteins related to dynein light chain 8. PlantPhysiol 2005, 139:857-868.
109. Bousios A, Darzentas N, Tsaftaris A, Pearce SR: Highlyconserved motifs in non-coding regions of Sirevirusretrotransposons: the key for their pattern of distributionwithin and across plants? BMC Genomics 2010, 11:89.
110. Llorens C, Fares MA, Moya A: Relationships of gag-pol diversitybetween Ty3/Gypsy and Retroviridae LTR retroelements andthe three kings hypothesis. BMC Evol Biol 2008, 8:276 http://dx.doi.org/10.1186/1471-2148-8-276.
111. Bannert N, Kurth R: The evolutionary dynamics of humanendogenous retroviral families. Annu Rev Genom Hum Genet2006, 7:149-173.
112. Zhou Y, Cahan SH: A novel family of terminal-repeatretrotransposon in miniature (TRIM) in the genome of the redharvester ant, Pogonomyrmex barbatus. PLoS ONE 2012,7:e53401 http://dx.doi.org/10.1371/journal.pone.0053401.
113. Roulin A, Piegu B, Wing RA, Panaud O: Evidence of multiplehorizontal transfers of the long terminal repeatretrotransposon RIRE1 within the genus Oryza. Plant J 2008,53:950-959.
114. Ludwig A, Valente VL, Loreto EL: Multiple invasions ofErrantivirus in the genus Drosophila. Insect Mol Biol 2008,17:113-124.
115. Schulman AH, Wicker T: A field guide to transposable elements.In Plant transposons and genome dynamics in evolution. Edited byFedoroff NV. John Wiley and Sons; 2013:15-40.
116. Telesnitsky A, Goff SP: Reverse transcriptase and thegeneration of retroviral DNA. In Retroviruses. Edited by CoffinJM, Hughes SH, Varmus HE. Cold Spring Harbor LaboratoryPress; 1997:121-160.
n Virol (2013), http://dx.doi.org/10.1016/j.coviro.2013.08.009
Current Opinion in Virology 2013, 3:1–11





























