Embed Size (px)
Citation preview

Plant Molecular Biology 26: 393-402, 1994. © 1994 Kluwer Academic Publishers. Printed in Belgium. 393
Expression of the tobacco Tntl retrotransposon promoter in heterologous species
Peter K. Pauls 1, Karl Kunert 2, Eric Huttner 3 and Marie-Ang+le Grandbastien* Laboratoire de Biologie Cellulaire, INRA-Versailles, F-78026 Versailles Cddex, France (*author for correspondence); Present adresses: 1 Department of Crop Science, University of Guelph, Guelph, Ontario, N1 G2 WI Canada; 2AECI Limited, Research Development Department, P.O. Modderfontein 1645, South Africa; 3 Biosem Pacific, Research school of Biological sciences, The Australian National University, GPO Box 4 75, Canberra A CT 2601, Australia
Received 27 January 1994; accepted in revised form 28 June 1994
Key words: Arabidopsis thaliana, Brassica napus, gene expression, Nicotiana tabacum, retrotransposon
Abstract
The expression of the tobacco (Nicotiana tabacum) retrotransposon Tntl has previously been shown to be strongly regulated and driven from the 5' long terminal repeat (LTR). We report here that the Tntl LTR can promote activity of the fl-glucuronidase (GUS) reporter gene in two heterologous spe- cies of the Brassicaceae family, namely rapeseed (Brassica napus) and Arabidopsis thaliana. The trans- lational LTR-GUS fusion was active in transient expression studies performed with tobacco and rape- seed protoplasts, indicating that the LTR sequences are recognized in heterologous species. Our results also showed that Tntl LTR-promoted GUS expression in transgenic Arabidopsis is strongly regulated, and that, in contrast to tobacco, hormonal activation plays a significant role in the expression of the Tntl LTR in Arabidopsis. LTR sequences were shown to be more effective than the CaMV 35S enhancer region in transient expression studies performed with tobacco or rapeseed protoplasts, and substitution of the LTR sequences upstream from the major transcriptional start with the CaMV 35S enhancer region gave high levels of expression in transgenic tobacco and Arabidopsis leaves, suggesting that a Tntl element with similar substitutions in its 5' LTR might be suited for gene-tagging experiments in heterologous species.
Introduction
Retroelements are defined as mobile DNA se- quences whose structural features suggest that they transpose via a mechanism that includes re- verse transcription of an RNA intermediate (for review see [ 3 ]). Representatives of several classes of retrotransposons have been characterized in a variety of plant species (for review see [13] [26]),
and recent studies have shown that LTR retro- transposons are ubiquitous components of plant genomes [11] [29].
However, to date, direct evidence for trans- positional activity has only been obtained for the maize Bsl element [19], the tobacco Tntl ele- ment [12] and the tobacco Tto elements [16]. Therefore, little is known about the regulation of expression of plant retrotransposons. Never-

394
theless, because of their structural similarities to animal and yeast retrotransposons, the mecha- nisms of transcription and transposition of plant retrotransposons are thought to be similar to those characterized for the former. Animal and yeast retrotransposons replicate through an RNA intermediate whose transcription starts in the 5' LTR and terminates in the 3' LTR [3]. Thus, the LTR functions both as promoter and termi- nator elements.
To date, the only plant retrotransposon for which transcriptional activity has been studied in detail is the Tntl tobacco element. Previous studies showed that Tntl transcription started in the 5' LTR [22]. Constructs containing the Tntl LTR, the untranslated leader sequence and the first 25 amino acids of the element's ORF, which were fused to the fl-glucuronidase (GU S) reporter gene [18], expressed GUS activity in tobacco, both in transient expression assay and transgenic plants [22]. Tntl expression in tobacco was shown to be highly regulated. Tntl transcripts and LTR-GUS gene products are poorly ex- pressed in intact tobacco leaf tissues oftransgenic plants, but strongly induced in leaf mesophyll- derived protoplasts, mostly due to the presence of fungal extracts containing the cell wall hydrolases [22]. A more recent study showed that LTR se- quences located upstream from the first transcrip- tional start contained strong activator elements and were involved in the protoplast-dependent induction of the Tntl promoter expression in tobacco [9].
Transposon-tagging with conventional trans- posable elements, like the maize Ac element, has been very successfully used to identify and clone plant genes [ 1 ]. Retrotransposons, which are ex- pected to transpose to loci that are not linked to the original insertion site, may be interesting as alternatives to conventional plant elements for gene-tagging experiments. Due to the high copy number of Tntl-related elements detected in the tobacco genome, its utilization as a gene tag seems however difficult in its original host. However, Tntl may be useful in heterologous plant species, such as Brassicaceae species which do not contain sequences that are closely related to Tntl
[12]. Details of the functionality of the Tntl pro- moter in heterologous genetic backgrounds needs however to be determined, as well as the condi- tions in which the strong regulation of the expres- sion of Tntl can be modified, in both homologous and heterologous genetic backgrounds.
This report provides the first evidence that the Tntl LTR can promote the activity of the GUS reporter gene in heterologous plant systems, and includes transient expression studies performed with rapeseed (Brassica napus) protoplasts, and expression studies of stably transformed Arab# dopsis thaliana plants. We also provide evidence that sequences upstream of the first transcrip- tional start of the LTR function as an activator of transcription in heterologous genetic back- grounds, and we show that Tntl LTR-promoted GUS expression in transgenic Arabidopsis is strongly regulated. Furthermore, we show that replacement of the LTR sequences upstream of the first transcriptional start by the CaMV 35S enhancer sequences induces a high level of con- stitutive activity of the construct, in homologous and heterologous genetic backgrounds.
Materials and methods
Plant materials
Electroporation experiments were performed with Nicotiana tabacum cv. Xanthi XHFD8 [5], and Brassica napus cv. Brutor. Transformations were performed with Nicotiana tabacum cv. Samsun and Arabidopsis thaliana L. genotype C24.
Constructs and plasmids
1. The full-length LTR (DCBA-GUS) constructs used in transient expression studies and in stable transformations are pBMCV102120 and pB- MCV102106 plasmids, respectively, as described [221.
2. Deletion derivatives of the Tntl LTR were made as follow. A Pst I-Eco RI fragment from the plasmid pBI101.2 [18], containing the GUS-

coding sequence and the nopaline synthase (nos) terminator, was cloned into the plasmid Blue- script KS- (Stratagene), to create the plasmid pBMCV102101.2. Different portions of the Tntl LTR [12] were then cloned into the Xba I- Barn HI sites of pBMCV 102101.2 after filling in of the Xba I site by treatment with the Klenow fragment of Polymerase I to create a blunt end: (1) cloning of a Hinc II-Bgl II fragment of Tntl LTR (pos 79 to 766) created the plasmid pB- MCV102150 (CBA-GUS); (2) cloning of a Hind III-Bgl II fragment ofTnt l LTR (194-766), after blunt-ending of the Hind III site by Klenow treatment, created the plasmid pBMCV102145 (BA-GUS); (3) cloning of a Eco RV-BgllI frag- ment ofTnt l LTR (432-766) created the plasmid pBMCV102140 (A-GUS). All these constructs contain also Tntl internal sequences up to the first 25 amino acids of the Tntl open reading frame, fused in frame with the GUS open read- ing frame [22].
3. Substitution of the DC region of the Tntl LTR by the 35S enhancer sequences was per- formed as follows. A Hinc II fragment containing the -343 to + 8 region of the cauliflower mosaic virus (CaMV) 35S promoter was excised from the plasmid pCaMVCN (available from Pharmacia, Uppsala, Sweden), and cloned into the Hinc II site of the vector pUC19 [30] to create the plas- mid pCaMV1. This was followed by introduction of a unique Bgl II restriction site into the Bam HI site, to create the plasmid pCaMV3. The Eco RV- Bgl II fragment of pCaMV3 (containing the -90 to + 8 region of the CaMV 35S promoter) was then replaced by a Hind lII-BgllI (194-766) fragment of Tntl , after rendering the Hind III site of the fragment blunt by Klenow treatment. A Bam HI-EcoRI fragment from the plasmid pBI101.2 [18] was finally cloned between the BgllI and Eco RI (downstream pUC poly- linker) sites. The resulting pBMCV 102166 (35eBA-GUS) plasmid contains the -343 to -90 enhancer region of the 35S promoter, fused to the BA region of the Tntl LTR, as well as Tntl in- ternal sequences up to the 25 first amino acids of the Tntl open reading frame which is fused in frame with the GUS open reading frame. A ver-
395
sion of the 35eBA-GUS construct suitable for stable transformation was made by subcloning a Hind III-Eco RI fragment from pBMCV 102166 into the binary vector pBIN19 [2].
4. Substitution of the 35S CaMV enhancer se- quences by the LTR DC region was performed as follow. The CaMV poly(A) region was excised from the vector pJIT58 [i5] as a Bam HI/Bgl II fragment and cloned into the Barn HI site of the plasmid described above, pCaMV1, to create pCaMV2. An Eco RV-Kpn I fragment from the plasmid pCaMV2, which contained the -90 to + 8 region of the CaMV 35S promoter and a CaMV poly(A) region, was cloned into the Hinc II and Kpn I sites of the plasmid pUC1318 to gen- erate the vector pCaMV4. A Bam HI fragment from the plasmid pJIT58 which contained the GUS-coding sequence was then cloned into the BgllI site of pCaMV4 to create the plasmid pCaMV5. A Sac I fragment from the vector pCaMV5, containing the -90 to + 8 region of the CaMV 35S promoter, the GUS-coding sequence and the CaMV terminator region, was then cloned into the Sac I site of the vector pUC18, to create the vector pCaMV6. The 330 bp Hind III frag- ment containing the DC (pos 1 to 194) region of Tntl LTR was finally subcloned into the unique Hind III restriction site of pCaMV6. The result- ing plasmid pBMCV102186 (DC35p-GUS) con- tained the DC Tntl LTR region upstream of the -90 to + 8 region of the 35S promoter which was fused to the GUS-coding sequence.
5. The following plasmids were used as con- trois: (1) the plasmid pBI121 [18] as a CaMV 35S-GUS construct, as well as (2) a fusion of the 35S enhancer region to the GUS gene (35e- GUS). The latter was created as follows: a Eco RV-Xba I fragment was deleted from the plasmid pCaMV1 by digestion with the restric- tion enzymes Eco RV and Xba I and religation after blunt-ending of the Xba I site. This elimi- nates the 35 S promoter region downstream of the -90 Eco RV site. Finally, a Barn HI-Eco RI frag- ment excised from the vector pBI101.2 [ 18], con- taining the GUS-coding sequence and the nos terminator, was cloned between the Barn HI and Eco RI sites to generate the plasmid 35e-GUS.

396
Transgenic plants
The constructs investigated were mobilized into the disarmed Agrobacterium tumefaciens strain C58C1 (pGV2260) by triparental mating [23]. Constructs were used to inoculate either leaf discs from tobacco plants grown under glasshouse con- ditions as described [ 15] oyused to inoculate root explants of Arabidopsis according to the protocol outlined by Valvekens et al. [28]. Tobacco plant- lets 4 to 5 cm high and 8-days old Arabidopsis R1 seedlings were screened for GUS activity.
Callus obtention
To induce callus formation, leaf cuttings were placed on 1 x MS medium containing 0.8~o agar and 1 mg/1 2,4-D. After one week leaf cuttings started to produce calli. Calli were divided every 3 weeks, and maintained on the medium men- tioned above.
Protoplast electroporation
Tobacco protoplasts were prepared in GSG me- dium [ 10] as described [22] and rapeseed proto- plasts were isolated as described [25]. Plasmid DNA was isolated according to Maniatis et al. [20] and protoplasts were electroporated essen- tially as described [ 14]. For each electroporation 10/~g of plasmid DNA was mixed to 50 #g of sonicated calf thymus DNA used as carrier DNA.
Protein and fl-galacturonidase assays
Protein extracts were obtained from protoplasts harvested 20 h after electroporation and from leaf discs of transformed tobacco plants (three 8 mm discs per plant), upper portions of pooled Arabi- dopsis transgenic seedlings that tested positive for GUS activity after histochemical staining, and calli (50 to 100 mg) of Arabidopsis and tobacco plants. The tissues were ground in GUS assay buffer [18] on ice, the protein concentrations of
the extracts from protoplast and plant samples were determined by the Bradford method [7], and the GUS activities in the extracts from pro- toplasts were determined as described [9].
Histochemical staining of plant tissue was per- formed as described by Jefferson [17].
Results
Transient activity in tobacco and rapeseed proto- plasts
Comparative transient expression studies of the LTR-GUS translational fusion previously de- scribed [22] were performed in the present study with tobacco and rapeseed protoplasts. In addi- tion, we analysed the activities of LTR-GUS con- structs with 5' deletions of the LTR (Fig. lb, c, d), of a LTR-GUS construct in which a part of the LTR was replaced with the enhancer se- quences of the CaMV 35S promoter (Fig. le), and of a 35S-GUS construct in which the en- hancer sequences of the CaMV 35S promoter were replaced by a part of the LTR (Fig. if). The Tntl LTR was divided into four blocks D, C, B, and A, that are separated by the restriction sites HinclI, Hind lII, and EcoRV, respectively (Fig. la). Block D contained Tntl LTR 5' up- stream sequences up to position 79 (-155 relative to the major transcriptional start as defined in [22]), as well as 110 bp of a remaining NR se- quence. Block C contains Tntl LTR sequences from position 79 (-155) to 194 (-40). Block B contained sequences from 194 (-40) to 432 (+ 198) and included the major transcriptional start (+ 1) and its corresponding TATA box, as well as a second TATA box, whose correspond- ing transcriptional start is located on block A [22]. Block A contained sequences from posi- tion 432 ( + 198) to 766 ( + 532), and includes the 3' end of Tntl LTR, the untranslated leader se- quence and the first 25 amino acids of Tntl- coding sequence.
Tobacco protoplasts electroporated with the DCBA-GUS construct had levels of GUS activity that were 100-fold higher than controls

Xbal Hinel I Hindl l I EeoRV
A
TRTR TATA
C B
, I
B R
R I I
35e B A L % \ \ ' . .~ \ \ \ \ \ \ % . \ ~. "q~ i ii i!
J I ~ a) DCBA-GUS RTG
' i 6us i b) CBA-GUS ' I I I ATG
I 6US I c) BR-GUS I flTG
I ~ d) R-GUS RTG
I I ~ e ) 35eBA-GUS ATG
~ f) DC35p-GUS
397
35e 55p r - - - - - I
35e ~ \ " \ " " " \ " " " " " \ ] 6US i h) 35e-GUS
Fig. 1. Schematic representation of constructs used in this work. a. DCBA-GUS, translational fusion to a promoterless fl-glucuronidase gene [ 18] of the Tntl LTR, to which are attached the untranslated leader sequence and 25 amino acids of the Tntl open reading frame [22]; sites for Hinc II, Hind III and Eco RV restriction enzymes that break up the LTR into blocks A, B, C and D are shown, as well as transcriptions starts (arrows) and corresponding TATA boxes, as determined by Pouteau et al. [22]. b-d. CBA-GUS, BA-GUS and A-GUS, constructs similar to DCBA-GUS but missing block D, block C and block B, respectively, e. 35eBA-GUS, construct similar to DCBA-GUS, but in which the DC fragment has been deleted and replaced by the enhancer region of the 35S CaMV promoter, f. DC35p-GUS, construct similar to the 35S-GUS construct, but in which the 35S enhancer region has been replaced by the LTR DC region, g-h. 35S-GUS and 35e-GUS, control constructs in which the GUS gene is placed under control of the 35S entire promoter or the 35S enhancer region, respectively.
electroporated with carrier DNA (Fig. 2). GUS activity in rapeseed protoplasts electroporated with the DCBA-GUS construct was only 3-fold higher than background levels (Fig. 3). Back- ground levels of GU S activity were however very high in rapeseed, compared to tobacco, and con- trol constructs were poorly expressed. In particu- lar, the rapeseed protoplasts electroporated with the 35S-GUS construct expressed less that two times as much GUS activity as the background level (Fig. 3), while tobacco protoplasts electropo-
rated with the same 35S-GUS construct ex- pressed GUS activities that were almost 6-fold that measured in the controls (Fig. 2). In both tobacco and rapeseed protoplasts, however, tran- sient expression of the DCBA-GUS construct was higher than the 35S-GUS expression.
Removal of the D fragment from the 5' end of the LTR (CBA-GUS construct) eliminated its ac- tivity as a promoter in tobacco (Fig. 2) and rape- seed (Fig. 3) protoplast transient assays. Con- structs with further deletions from the 5' end of
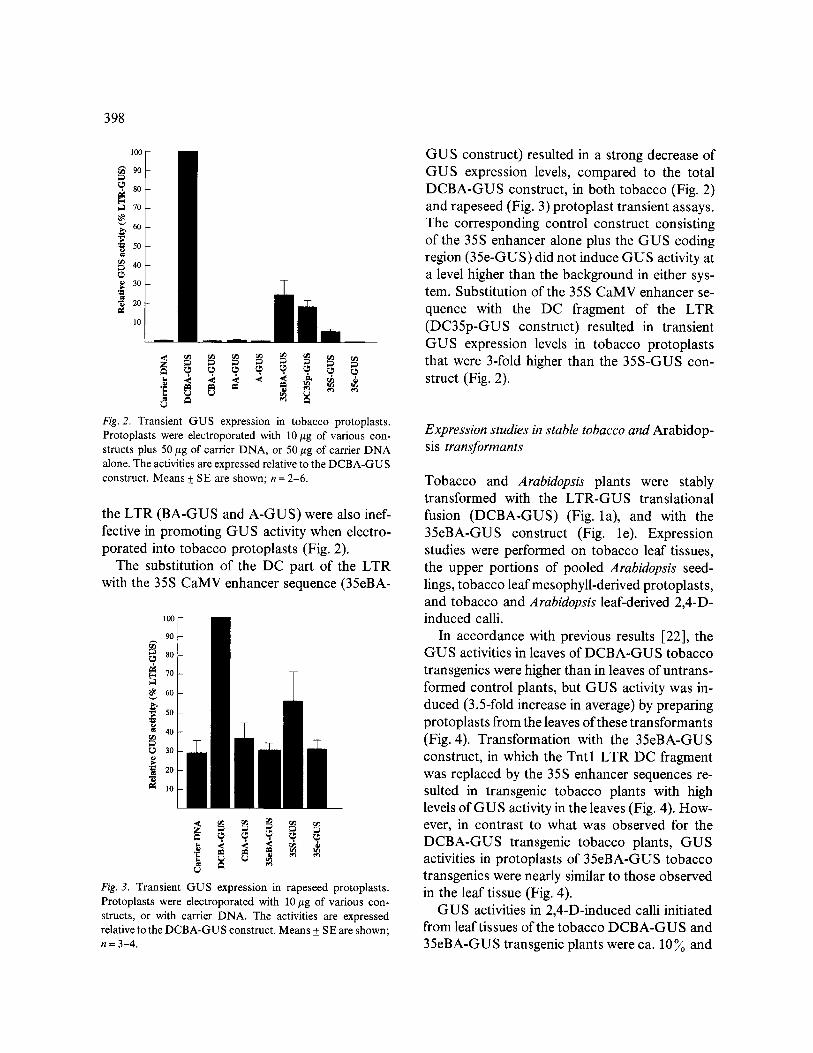
398
100
~" 90
~1 70
~ 50 ~ 4o
~. 3o
~ 2o 10
Fig. 2. Transient G U S expression in tobacco protoplasts . Protoplas ts were electroporated with 10 #g of var ious con-
structs plus 50 #g o f carrier D N A , or 50 #g o f carrier D N A
alone. The activities are expressed relative to the D C B A - G U S construct . M e a n s + SE are shown; n = 2-6 .
the LTR (BA-GUS and A-GUS) were also inef- fective in promoting GUS activity when electro- porated into tobacco protoplasts (Fig. 2).
The substitution of the DC part of the LTR with the 35S CaMV enhancer sequence (35eBA-
r~
;9 ;9
Fig. 3. Trans ien t G U S expression in rapeseed protoplasts .
Protoplas ts were electroporated with 10 #g of various con-
structs, or with carrier D N A . The activities are expressed relative to the D C B A - G U S construct . M e a n s _+ S E are shown; n = 3-4 .
GUS construct) resulted in a strong decrease of GUS expression levels, compared to the total DCBA-GUS construct, in both tobacco (Fig. 2) and rapeseed (Fig. 3) protoplast transient assays. The corresponding control construct consisting of the 35S enhancer alone plus the GUS coding region (35e-GUS) did not induce GUS activity at a level higher than the background in either sys- tem. Substitution of the 35S CaMV enhancer se- quence with the DC fragment of the LTR (DC35p-GUS construct) resulted in transient GUS expression levels in tobacco protoplasts that were 3-fold higher than the 35S-GUS con- struct (Fig. 2).
Expression studies in stable tobacco and Arabidop- sis transformants
Tobacco and Arabidopsis plants were stably transformed with the LTR-GUS translational fusion (DCBA-GUS) (Fig. la), and with the 35eBA-GUS construct (Fig. le). Expression studies were performed on tobacco leaf tissues, the upper portions of pooled Arabidopsis seed- lings, tobacco leaf mesophyll-derived protoplasts, and tobacco and Arabidopsis leaf-derived 2,4-D- induced calli.
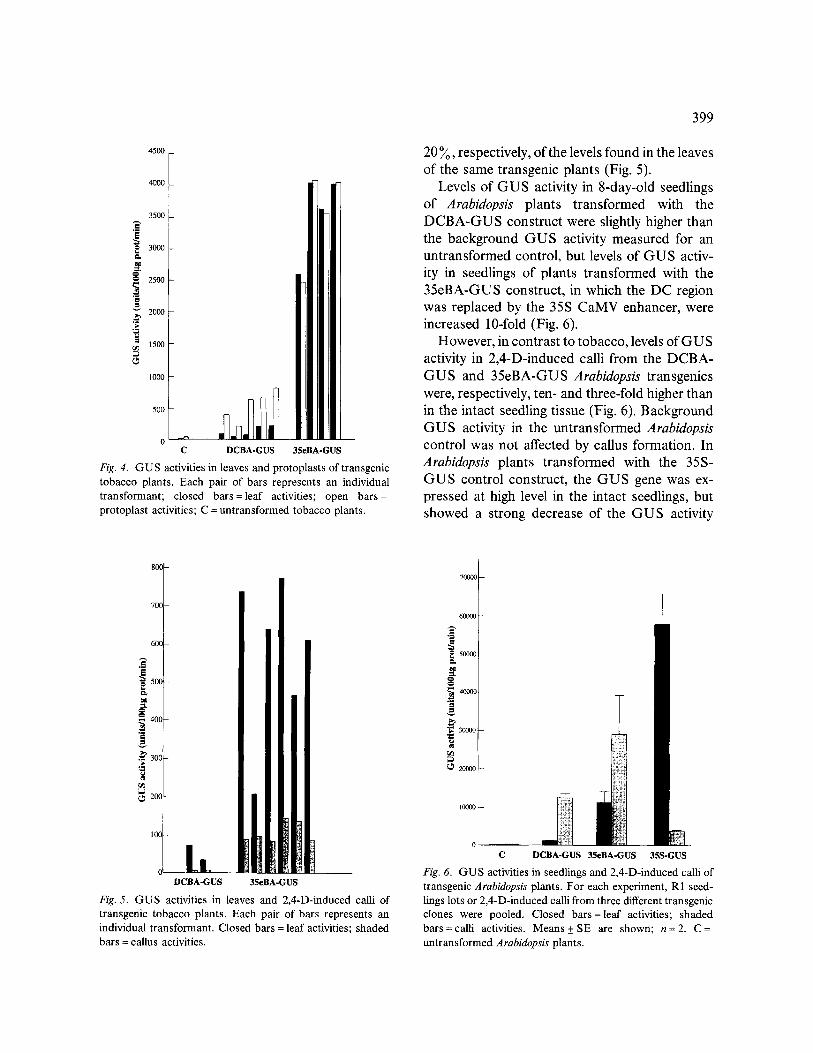
In accordance with previous results [22], the GU S activities in leaves of DCBA-GU S tobacco transgenics were higher than in leaves of untrans- formed control plants, but GUS activity was in- duced (3.5-fold increase in average) by preparing protoplasts from the leaves of these transformants (Fig. 4). Transformation with the 35eBA-GUS construct, in which the Tntl LTR DC fragment was replaced by the 35S enhancer sequences re- sulted in transgenic tobacco plants with high levels of GUS activity in the leaves (Fig. 4). How- ever, in contrast to what was observed for the DCBA-GUS transgenic tobacco plants, GUS activities in protoplasts of 35eBA-GUS tobacco transgenics were nearly similar to those observed in the leaf tissue (Fig. 4).
GUS activities in 2,4-D-induced calli initiated from leaf tissues of the tobacco DCBA-GUS and 35eBA-GUS transgenic plants were ca. 10~ and
4500
41300
3500
~ 3000 eL
~ 2500
1500
1000
500
O - - C DCBA-GUS 3SeBA-GUS
Fig. 4. G U S activities in leaves and protoplasts of t ransgenic tobacco plants. Each pair of bars represents an individual t ransformant ; closed b a r s = l e a f activities; open b a r s = protoplas t activities; C = unt ransformed tobacco plants.
399
20 ~ , respectively, of the levels found in the leaves of the same transgenic plants (Fig. 5).
Levels of GUS activity in 8-day-old seedlings of Arabidopsis plants transformed with the DCBA-GUS construct were slightly higher than the background GUS activity measured for an untransformed control, but levels of GUS activ- ity in seedlings of plants transformed with the 35eBA-GUS construct, in which the DC region was replaced by the 35S CaMV enhancer, were increased 10-fold (Fig. 6).
However, in contrast to tobacco, levels of GU S activity in 2,4-D-induced calli from the DCBA- GUS and 35eBA-GUS Arabidopsis transgenics were, respectively, ten- and three-fold higher than in the intact seedling tissue (Fig. 6). Background GUS activity in the untransformed Arabidopsis control was not affected by callus formation. In Arabidopsis plants transformed with the 35S- GUS control construct, the GUS gene was ex- pressed at high level in the intact seedlings, but showed a strong decrease of the GUS activity
800
70(
6~
g ~ 40C
30C
~ 20(
10C
DCBA-GUS 35eBA-GUS
Fig. 5. G U S activities in leaves and 2,4-D-induced calli of transgenic tobacco plants. Each pair of bars represents an individual t ransformant . Closed bars = leaf activities; shaded bars = callus activities.
70000
600o0
.o 50oo0 e~
~00
.~ 30000
r~
200O0
1001
C DCBA-GUS 35eBA-GUS 35S-GUS
Fig. 6. G U S activities in seedlings and 2,4-D-induced calli of transgenic Arabidopsis plants. For each experiment, R1 seed- lings lots or 2,4-D-induced calli from three different transgenic clones were pooled. Closed bars = leaf activities; shaded b a r s = c a l l i activities. Means + SE are shown; n = 2. C = unt ransformed Arabidopsis plants.

400
in 2,4-D-induced calli derived from these trans- genics.
Discussion
The results presented in this report show that the LTR of the tobacco retrotransposon Tntl can promote transcription of the GUS reporter gene in transient expression studies performed with rapeseed protoplasts, as well as in 2,4-D-derived calli from stable transgenic Arabidopsis plants, thus demonstrating that the tobacco Tntl LTR is also active in heterologous species. However, the level of LTR-GUS activity, relative to the back- ground level, in the electroporated rapeseed pro- toplasts was lower than that measured for to- bacco protoplasts. The transient expression experiments with rapeseed protoplasts were dif- ficult to carry out because the GUS background activity was high and expression of the 35S-GUS construct was very low in these protoplasts rela- tive to what is typically observed in other species. Low levels of transient expression for numerous promoters have previously been observed in tran- sient expression assays performed with rapeseed protoplasts and are thought to result from a higher level of DNA degradation in these protoplasts [24]. Nevertheless, our results clearly demon- strate that the Tntl LTR promoter is expressed at higher levels than the commonly used CaMV 35S promoter, in homologous and heterologous species. This conclusion is based on the present observations that the transient GUS activities in the tobacco and rapeseed protoplasts were higher for the full-length LTR-GUS construct than for the 35S-GUS construct. These results are also in agreement with previous reports of high promoter activity of the LTR in tobacco transient expres- sion studies [22, 9]. Furthermore, the present and previous [9] observations that the replacement of the 35S enhancer region by the DC LTR fragment induces higher levels of GUS expression in to- bacco protoplasts than the intact 35S promoter demonstrate that at least a portion of the pro- moting activity of the LTR is located in the DC fragment and that this activity is retained beyond the LTR context.
Our studies also demonstrate that sequences upstream from the LTR major transcription start are important for the promoter to function in rapeseed. In both tobacco and rapeseed transient expression studies, the removal of the D region (sequences up to -155), which has been previ- ously shown to contain activator sequences [9], eliminates LTR promoter activity. However, it should be noted that the C region, which is re- tained in this construct, also contains elements able to activate a heterologous promoter in to- bacco transient expression studies [9]. The likely explanation for our result is that the properties of the activator elements present in the C region are different in homologous or heterologous promoter contexts.
The activator ability of the DC region of the LTR seen in transient expression assays was not evident in leaf tissues of the tobacco LTR-GUS transgenic plants. In these plants, the GUS ac- tivity was 3-fold higher than background levels, while expression in transient expression studies was 100-fold higher than the background levels. Therefore, as observed previously [22], the ex- pression of the Tntl LTR is strongly regulated when it is stably incorporated into the tobacco genome. Also, as observed by Pouteau et al. [22], the activity of the LTR was enhanced by proto- plastization in the tobacco LTR-GUS transgen- ics. Our results indicated that the protoplast- specific regulation is mediated by the DC LTR fragment, because GUS levels, in the transgenics obtained with the construct where the DC region was substituted with the CaMV 35S enhancer region (35eBA-GUS), were not affected by the protoplasting procedure. This conclusion is in ac- cordance with recent observations from transient expression studies that showed that the DC re- gion of the LTR is involved in protoplast- dependent expression [9].
An intriguing observation made with the LTR- GUS and 35eBA-GUS Arabidopsis transgenics was that the GUS levels were considerably en- hanced in 2,4-D-induced calli derived from them. This induction was not observed in 2,4-D-induced calli obtained from transgenic tobacco plants. The stimulation of GUS activity in the transgenic

Arabidopsis calli was clearly not a property of the 35S enhancer because the GUS activities in the 2,4-D-induced calli from the 35eBA-GUS trans- genics were higher than the activities in 2,4-D- induced calli from the 35S-GUS transgenics. Moreover, in the latter case, levels of GUS ac- tivities were reduced in 2,4-D-derived calli, com- pared to intact leaves or plantlets. This reduction is consistent with observations that protein levels driven from the 35S promoter are constantly higher in older than in younger tissues because of stable accumulation of protein in older tissues [8]. This reduction could also be attributed to a dilution of the GUS protein due to a general en- hancement of protein synthesis in actively divid- ing tissues, such as young tissues or calli. If the latter is true, the stimulations of GUS activity observed in 2,4-D-derived calli of LTR-GUS and 35eBA-GUS transgenics are probably un- derestimated.
Hormonal activation appears to play a signif- icant role in the expression of the Tntl LTR in Arabidopsis but not in tobacco. Preliminary ex- periments of treatments with different auxin-type compounds on leaf pieces of Arabidopsis and to- bacco suggest that GUS activation in the first species is a hormonal effect and confirm that these treatments have no effect on tobacco (J. Casacuberta, unpublished). The latter conclusion is in accordance with previous observations that showed that the auxin content of the protoplas- ting media has no effect on Tntl activation in tobacco (S. Pouteau, unpublished results). These results suggest that Arabidopsis possesses specific transactivating factors that are sensitive to auxins, and are missing or ineffective in tobacco. Because the BA LTR region was sufficient to make the Arabidopsis transgenics sensitive to 2,4-D, it is likely that this region is involved in the hor- mone-specific activation of the Tntl LTR in Ara- bidopsis. Host-specific differential control of ex- pression has been also reported for the copia element, for which different regulatory regions are required for optimal expression in different Droso- phila species [27]. Therefore, further studies will likely be required to determine if specific se- quences are required for optimal expression and
401
specific regulation of Tntl in heterologous plant species.
Conclusion
This work is the first report of the ability of a plant retrotransposon promoter to function in hetero- logous species, namely B. napus and A. thaliana. This paper also demonstrates that the sequences involved in the expression of Tnt 1 in heterologous plant species are the same as those that are ac- tive in tobacco. Furthermore, we show that, as in tobacco, expression driven from the Tntl LTR is strongly regulated in a heterologous species, but that, in contrast to what is found in tobacco, hormonal activation seems to play a significant role in the expression of the Tntl LTR in Arabi- dopsis.
Our work also show that substitution of the Tntl LTR region upstream from the first tran- scriptional start with the 35 S enhancer sequences induces high levels of expression in transgenic tobacco and Arabidopsis leaves. These sequences correspond to the U3 region of the LTR, and their replacement should therefore not affect the trans- positional ability of the element, as described for other retrotransposon systems [4]. Thus, a Tntl element in which the 5' LTR carries a substitu- tion of the U3 region with the 35S enhancer se- quences, or with other regulatory sequences, might give high frequencies of transposition throughout the plant, or in specific tissues, and be particularly suited for gene tagging. The next step in the development of Tntl-based constructs for gene tagging will be to determine whether expres- sion levels are related to transposition levels.
Acknowledgements
We are grateful to P. Guerche for advice on iso- lating protoplasts in the electroporation experi- ments. We also thank J. Casacuberta, H. Lucas and C. Meyer for critical reading of the manu- script.

402
References
1. Balcells L, Swinburne J, Coupland G: Transposons as tools for the isolation of plant genes. Trends Biotechnol 9:31-36 (1991).
2. Bevan M: Binary Agrobacterium vectors for plant trans- formation. Nucl Acids Res 12:8711-8721 (1984).
3. Boeke JD, Corces VG: Transcription and reverse tran- scription ofretrotransposons. Annu Rev Microb 43: 403- 434 (1989).
4. Boeke JD, Garfinkel DJ, Styles CA, Fink GR: Ty ele- ments transpose through an RNA intermediate. Cell 40: 491-500 (1985).
5. Bourgin JP, Missonier C: Vegetative propagation and cold preservation of haploid plants of Nicotiana tabacum and Nicotianapaniculata. Haploid Inf Serv 8:7 (1973).
6. Bourgin JP, Chupeau Y, Missonier C: Plant regeneration from mesophyll protoplasts of several Nicotiana species. Physiol Plant 45:288-292 (1979).
7. Bradford MM: A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248-254 (1976).
8. Carozzi NB, Warren GW, Desai N, Jayne SM, Lotstein R, Rice DA, Evola S, Koziel MG: Expression of a chi- meric CaMV 35 S Bacillus thuringiensis insecticidal protein gene in transgenic tobacco. Plant Mol Biol 20:539-548 (1992).
9. Casacuberta JM, Grandbastien MA: Characterisation of LTR sequences involved in the protoplast specific expres- sion of the tobacco Tntl retrotransposon. Nucl Acids Res 21:2087-2093 (1993).
10. Chupeau Y, Bourgin JP, Missonier C, Dorion N, Morel G: Pr6paration et culture de protoplastes de divers Nicotiana. CR Acad Sci Paris (D) 278:1565-1568 (1974).
11. Flavell AJ, Dunbar E, Anderson R, Pearce SR, Hartley R, Kumar A: Tyl-copia group retrotransposons are ubiq- uitous and heterogeneous in higher plants. Nucl Acids Res 20:3639-3644 (1992).
12. Grandbastien MA, Spielmann A, Caboche M: Tntl, a mobile retroviral-like element of tobacco isolated by plant cell genetics. Nature 337:376-380 (1989).
13. Grandbastien MA: Retroelements in higher plants. Trends Genet 8:103-108 (1992).
14. Guerche P, Bellini C, LeMoullec JM, Caboche M: Use of a transient expression assay for the optimization of direct gene transfer into tobacco mesophyll protoplasts by electroporation. Biochimie 69:621-628 (1987).
15. Guerineau F, Brooks L, Meadows J, Lucy A, Mullineaux
P: Sulfonamide resistance gene for plant transformation. Plant Mol Biol 15:127-136 (1990).
16. Hirochika H: Activation of tobacco retrotransposons during tissue culture. EMBO J 12:2521-2528 (1993).
17. Jefferson RA: Assaying chimeric genes in plants: the GU S gene fusion system. Plant Mol Biol Rep 5:387-405 (1987).
18. Jefferson RA, Kavanagh TA, Bevan MW: GUS fusions: fl-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J 6:3901-3907 (1987).
19. Johns MA, Mottinger J, Freeling M: A low copy number, copia-like transposon in maize. EMBO J 4:1093-1102 (1985).
20. Maniatis T, Fritsch EF, Sambrook J: Molecular Cloning: A Laboratory manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1982).
21. Murashige T, Skoog F: A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473-497 (1962).
22. Pouteau S, Huttner E, Grandbastien MA, Caboche M: Specific expression of the tobacco Tntl retrotransposon in protoplasts. EMBO J 10:1911-1918 (1991).
23. Rogers SG, Horsch RB, Fraley RT: Gene transfer in plants: Production of transformed plants using Ti plas- mid vectors. Meth Enzymol 118:627-640 (1986).
24. Rouan D: Transfert direct de g6nes chez les Brassica: application ~t la r6sistance au virus de la mosaJque du chou-fleur. Th6se, Toulouse, France (1991).
25. Rouan D, Guerche P: Transformation and regeneration of oilseed rape protoplasts. In: Lindsey K (ed) Plant Tissue Culture Manual, B4: 1-24. Kluwer Academic Publishers, Dordrecht, Netherlands (1991).
26. Smyth DR: Plant retrotransposons. In: Verma DPS (ed) Control of Plant Gene Expression, pp 1-15. CRC Press, Boca Raton, FL (1993).
27. Sneddon A, Flavell AJ: The transcriptional control re- gions of the copia retrotransposon. Nucl Acids Res 17: 4125-4035 (1989).
28. Valvekens D, Van Montagu M, Van Lijsebettens M: Agrobacterium tumefaciens-mediated transformation of Arabidopsis thaliana root explants by using kanamycin selection. Proc Natl Acad Sci USA 85:5536-5540 (1988).
29. Voytas DF, Cummings MP, Konieczny A, Ausubel F, Rodermel C: Copia-like retrotransposons are ubiquitous among plants. Proc Natl Acad Sci USA 89:7124-7128 (1993).
30. Yanisch-Perron C, Vieira J, Messing J: Improved M13 phage cloning vectors and host strains: Nucleotide se- quences of the M13mpl8 and pUC19 vectors. Gene 33: 103-119 (1985).