Embed Size (px)
Citation preview

The promoter of the Tnt1A retrotransposon is activated by ozone airpollution in tomato, but not in its natural host tobacco
Nathalie Pourtau a,*, Beatrice Lauga a, Colette Audeon b, Marie-Angele Grandbastien b,Philippe Goulas a, Jean-Claude Salvado a
a Laboratoire d’Ecologie Moleculaire, Universite de Pau et des Pays de l’Adour, UFR Sciences et techniques, BP 1155, 64013 Pau, Cedex, Franceb Laboratoire de Biologie Cellulaire, INRA Centre de Versailles, 78026 Versailles Cedex, France
Received 16 May 2003; received in revised form 16 May 2003; accepted 18 June 2003
Plant Science 165 (2003) 983�/992
www.elsevier.com/locate/plantsci
Abstract
The tobacco retrotransposon Tnt1A is one of the few active plant retrotransposon and is known to be transcriptionally activated
in tobacco (Nicotiana tabacum ) and in heterologous species by biotic and abiotic stress factors. It has been previously reported that
Tnt1A expression is linked with the early steps of metabolic pathways leading to the activation of plant defence genes. As ozone is
known to generate an oxidative stress in plant and activate the defence system, we have investigated, using LTR-GUS constructs,
the effect of ozone on the Tnt1A promoter activation in its natural host tobacco and in an heterologous species, tomato
(Lycopersicon esculentum ). Plants cultivated in open top chambers (OTC) were exposed to different ozone concentrations in order to
simulate either different range of a realistic chronic ozone stress or an acute ozone stress. The results show that the Tnt1A promoter
is not activated by ozone in tobacco, whereas dose-dependant and cumulative effects are observed in tomato. This difference
observed between tobacco and tomato is discussed. Moreover the use of such a construct to study both early response to ozone
stress and regulation of the retrotransoposon is examined.
# 2003 Elsevier Ireland Ltd. All rights reserved.
Keywords: Active retrotransposon; Ozone; Tobacco; Tomato; Differential activation; Stress
1. Introduction
Ozone (O3) is the most widespread air pollutant in
many areas of the industrialised world and ozone
concentrations in the tropospheric atmosphere have
increased during the past decades as the result of
anthropogenic activities [1,2]. Ozone is considered to
be the most phytotoxic of the common pollutants [3],
since it reduces photosynthesis, growth, and enhances
premature senescence in plant at concentration not
much excess of maximum natural level [4�/6]. Ozone
pollution, like other environmental stresses including
pathogen attack, drought and heavy metals can generate
an oxidative burst into the plant [7,8]. It is assumed that
O3 enters in the intercellular leaf space of plants through
stomates where it is converted into reactive oxygen
species (ROS), like O2+�, HO+ and H2O2. Once
generated, these compounds can induce structural and
functional alterations of the plasma membrane [9].
Plants respond to O3-induced oxidative stress by acti-
vating a number of antioxidative stress-related defence
mechanisms that, in turn, induce changes in the
biochemical plant machinery [10,11]. Studies have
shown that ROS probably require additional molecules
to transduce and amplify defence signals [12,13]. Others
studies have demonstrated that ozone responses are
similar to those against pathogens [14]. In addition,
Schubert and co-authors have shown using transgenic
tobacco, the presence of an ozone-responsive region
different from the basal pathogen-responsive sequence
in the grapevine resveratrol synthase promoter [15].
Abbreviations: AA, ambient air; GUS, b-glucuronidase; LTR, long
terminal repeat; OMT, ortho-diphenol-O -methyl-transferase; OTC,
open top chambers; ROS, reactive oxygen species.
* Corresponding author. Present address: Department of Biology,
University College London, Gower Street, London WC1E6BT, UK.
Tel.: �/44-20-7679-7275; fax: �/44-20-7679-7096.
E-mail address: [email protected] (N. Pourtau).
0168-9452/03/$ - see front matter # 2003 Elsevier Ireland Ltd. All rights reserved.
doi:10.1016/S0168-9452(03)00278-4

Stress appears as an important factor in the activation
of particular types of genomic sequences like transpo-
sable elements. These mobile genetic elements comprise
different classes, among them retrotransposons, whichare closely related to retrovirus, and transpose via a
RNA intermediate [16]. Representatives of several
classes of retrotransposons have been characterised in
a variety of plant species but only a few plant retro-
transposons are known to be active [17]. Activation is
controlled by the element itself via signals depending on
the host organism, and additionally enhanced by
external factors such as environmental changes or stress[18�/20]. One of the best characterised plant retro-
transposon is the active copia -like Tnt1A element of
tobacco (Nicotiana tabacum ). It is repressed in un-
stressed vegetative tissue, but slightly activated in adult
roots [21]. However the Tnt1A promoter is activated by
various biotic and abiotic factors in tobacco and as well
as in heterologous species, like tomato (Lycopersicon
esculentum ) and Arabidopsis thaliana [22]. Since all thesefactors are known to elicit plant defence response, it has
been suggested that there is a link between Tnt1A
transcript activation and plant defence response [23].
The main objective of this study was to determine
whether O3 realistic exposure could trigger the induction
of the Tnt1A promoter in two different species, tobacco
and tomato. In this aim, experiments have been
performed in open top chambers (OTC), i.e. with plantsgrowing close to natural conditions. Ozone concentra-
tions applied were in the range defined by the United
Nations-Economic Commission for Europe in the ozone
directive (92/72/EEC) [24].
2. Materials and methods
2.1. Fumigation system and measurements of ozone
concentration
Ozone fumigation was carried OTCs, whose technical
characteristics were close to those described by Heagle etal. (1973, 1979) [25,26]. The O3 fumigation and O3
monitoring systems used here were described previously
[27]. The atmospheres tested in OTCs to simulate the
chronic stress of 21 day were the following: ambient air
(AA, ca. 40�/50 nl l�1 in the early summer afternoon),
AA supplied with 20, 40 and 60 nl l�1. Experiments
were also carried out to mimic an acute stress using the
effective amount of 180 nl l�1 of ozone during 7 h. Ineach experiment, extra ozone was equally released each
day from 10:00 to 17:00 h (GMT), to simulate the
natural ozone production, on plants from stage leave 4
to sampling.
2.2. Plant cultivation
L. esculentum cv. UC82B and N. tabacum cv. Xanthi
XHFD8 and Samsun plants carrying the b-glucuroni-dase gene [28] under the control of the Tnt1A promoter
(LTR-GUS) or the CaMV 35S promoter (35S-GUS)
used in this study were described elsewhere [29�/31]. For
the LTR-GUS construct, three independent transgenics
were tested for tobacco plants: LG(7)1, HUT2(1)1 and
HUT2(1)2 and for tomato plants L3(10), L7(6) and
L13(11). All the tomato transformants tested revealed
an activation of LTR-GUS. Therefore, for subsequentanalyses only the tomato transformant LTR-GUS 7(6)
was used. For the control, only one transformant 35S-
GUS was used, PBI121c for tobacco and P(9) for
tomato. One seed of each transgenic plant was sown in
a small plastic pot containing loam, under a plastic
greenhouse. Plantlets with three or four leaves (5 or 6
weeks), were transferred to 2 l plastic pots containing a
loam/peat (50/50) mixture and introduced to each OTCsfor the chronic experiment. For the fumigation with 180
nl l�1 of ozone, plants with at least four fully expanded
leaves (around 6 or 7 weeks) were used.
2.3. GUS assays
Transcriptional regulation of Tnt1A has been studied
by transcript analysis or by analysing the expression of
the LTR-GUS translational fusion. The construct con-sists of the b-glucuronidase reporter gene [28], placed
under the control of sequences composed of Tnt1A
5’LTR and the adjacent untranslated region, as well as
the first nucleotides of the coding domain [21]. Fluori-
metric quantification of GUS activity was carried out
using frozen collected leaves. After grinding, leaf tissues
were resuspended in 200 ml of extraction buffer (50 mM
NaH2PO4, 2 H2O, 10 mM EDTA, 10 mM dithiothreitol(DTT), 10% (v/v) glycerol, pH 7). After a centrifugation
of 20 min, determination of protein concentration in the
extracts was performed using Bradford reagent (Bio-
Rad). Fluorescence of the methyl umbelliferone product
was quantified with a fluorimeter (CytoFluor 4000
multi-well plate reader, PE Biosystems) following the
protocol of Jefferson et al. (1987) [28] using a microliter
plate fluorimeter (Perkin�/Elmer).
2.4. RNA extraction and northern blot hybridisation
Total RNA was extracted following the protocol of
Logemann et al. (1987) [32]. RNA was fractionated in a
denaturing 1.4% agarose gel and transferred onto a
Hybon N� membrane (Schleicher & Shuell). The
Megaprime DNA labelling kit (Amersham) was usedto label the GUS DNA probe with [a-32P]dCTP from
Isotopchim. Hybridisation was carried out at 42 8C in
50% formamide, 6�/SSC, 0.5% S.D.S., 5�/Denhart’s
N. Pourtau et al. / Plant Science 165 (2003) 983�/992984

solution, 100 mg ml�1 sperm salmon DNA. Membranes
were washed at a maximum stringency of 0.1�/SSC,
0.1% S.D.S. at 65 8C.
3. Results
3.1. Effects of chronic ozone stresses on plants
After a 21 days ozone-chronic treatment both tobacco
and tomato plants exhibit a visible reduction in plant
growth and productivity (Fig. 1A). Characteristic re-
sponses are also observed at the foliage level. Initially,
tomato leaflets extremities become curled and whitish
silvered lesions appear. Then, these regions evolvedthrough a yellowish and brownish staining showing
necrosis of the tissue (Fig. 1B). In tobacco, injuries
occurs in the form of small spotted whitish areas
distributed randomly all over the leaf (Fig. 1C). How-
ever, in this species, symptoms appear earlier and
stronger than in tomato during the development, leading
to early plant death. This phenomenon did not allow to
perform the 21 days chronic experiment with the highestozone atmosphere (OTC4).
3.2. Effects of chronic ozone stresses on Tnt1A promoter
After a 21 days exposure to the chronic ozone stress,
leaves of tobacco and tomato were collected and the
GUS activity was determined. No GUS activity was
detected in any of the three tobacco LTR-GUS trans-
genics for all the ozone concentrations applied (data not
shown). In the control 35S-GUS tobacco transgenic,similar level of GUS activity was detected in each OTC
(Fig. 2). The activity measured on 12 tomato plants and
for four ozone concentrations confirmed the induction
of the LTR promoter (Fig. 3A). The synthesis of the
GUS protein is detected as soon as 20 nl l�1 load dose
ozone is added to the AA for the oldest leaves that were
exposed for the longer period to O3. For a AA�/40 nl
l�1 O3 effect is visible on all leaves starting from theyoungest leave (8) to the oldest sampled (leave 4) with an
increase activity from the top to the bottom (Fig. 3A). A
similar pattern is observed for AA�/60 nl l�1 except
that GUS activity is higher for all sampled leaves. For
the control 35S-GUS tomato transgenic the GUS
activity is at a comparable level, whatever the O3
concentration or leaf rank (Fig. 3B). These experiments
were repeated the following year giving similar data.These results suggest that the Tnt1A promoter can be
activated by an ozone treatment in a dose-dependant
manner in tomato leaves.
3.3. Ozone effect according to the leaf development stage
In order to determine whether the response to ozone
stress may vary with development stage, we set up anexperiment in which batches of ten tomato plants at
three different development stages were exposed to an
acute stress of 180 nl l�1 during 7 h (Fig. 4). GUS
activity measured on well-developed leaves without
senescing symptoms showed that the response of the
LTR-GUS construct is similar in all leaf ranks. In
addition, similar results were observed for all three
developmental stages (i.e. from a five to six leaves plantto a plant harbouring the first flower buds). This
suggests that the previous results are the consequence
of a longer exposure to ozone of the oldest leaves rather
than a differential response due to the development
stage of the leaves.
3.4. Kinetic of LTR-GUS activity during a 21 days
chronic ozone stress
The chronic ozone stress of 21 days has been analysed
in a subsequent experiment in which GUS activity was
daily measured starting from the first day of exposure to
the last one for an ozone concentration of AA supplied
with 60 nl l�1 and applied as previously during 7 h per
day. As a control, a similar procedure was conducted in
parallel with AA. Four plants were sampled each day
for each a treatment and a mixture of leaves 3 and 4 wasused to measure GUS activity (Fig. 5). This experiment
showed that the profile response could be decomposed
into two phases. During the first days the activity
increased until it reached a peak after 6 days of
treatment. Moreover it was also noticed that ozone
injury on the leaves start to develop 6 days after
exposure. The GUS activity started to decrease slowly
from the seventh day and was maintained to a lowerlevel until the 21st day. The observed level during this
second phase is higher than the basal level in the absence
of added ozone. Northern blot experiments confirmed
the presence of GUS mRNAs during the first phase
(Fig. 6). Hybridisation signals show that GUS mRNA
accumulation starts at the first day of the kinetic and is
maximal at the fifth. Then, transcripts are no longer
detected. These results highlight a good correlationbetween transcriptional activation of the Tnt1A promo-
ter and GUS activity measurements.
3.5. GUS protein turn over
In order to specify whether the profile response
obtained previously during the second phase was due
to a residual GUS activity in the leaves from the first 6days of treatment or to de novo synthesis of the protein,
we set up an experiment in which tomato plants were
exposed to an acute stress of 180 nl l�1 during 7 h and
N. Pourtau et al. / Plant Science 165 (2003) 983�/992 985

then placed during 20 days in an OTC containing an AA
(Fig. 7). Leaves 3 and 4 were harvested every 2 days
throughout the experiment and GUS activity measured
in the sampled material. The response profile showed
that the synthesis of the GUS protein increases until the
sixth day. The following days, GUS activity decreases
back to the basal level observed in the control plants.
According to this profile, the half-life of GUS protein
seems not to exceed 48 h, as yet estimated by Jefferson et
al. (1987) [28].
3.6. Kinetic of LTR-GUS activity during an acute ozone
stress
We have also studied fine kinetics of the LTR-GUS
response in tobacco and tomato transgenic plants
during an acute stress. Plants were exposed to 180 nl
l�1 of ozone and leaves (3 and 4) were harvested after 1,
3 and 7 h of stress for tobacco plants and after 30 min of
exposure and then every hour until 7 h for tomato
plants. Similarly to the ozone chronic stress, no GUS
Fig. 1. Physiognomy of stressed plants after a 21 days exposure during 7 h per day to different ozone atmospheres: AA (OTC 1), AA supplied with
20 nl l�1 (OTC 2), 40 nl l�1 (OTC 3), and 60 nl l�1 (OTC 4). (A) LTR-GUS tomato plants exposed to the four different ozone atmospheres studied.
(B) Leaves 3 of transgenic tomato plants shown in (A). (C) Leaves 3 of LTR-GUS tobacco grown in OTC 1, OTC 2 and OTC 3.
N. Pourtau et al. / Plant Science 165 (2003) 983�/992986
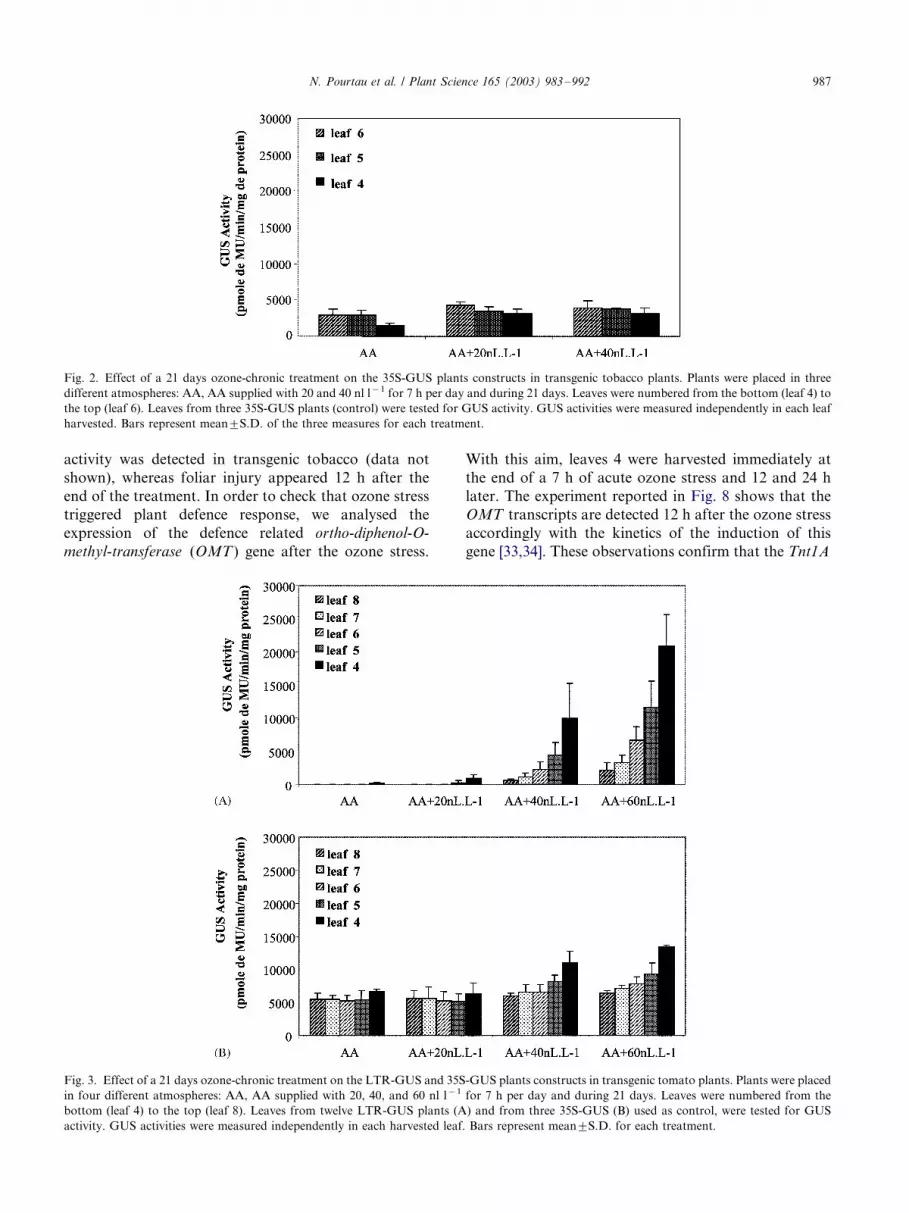
activity was detected in transgenic tobacco (data not
shown), whereas foliar injury appeared 12 h after the
end of the treatment. In order to check that ozone stress
triggered plant defence response, we analysed the
expression of the defence related ortho-diphenol-O-
methyl-transferase (OMT ) gene after the ozone stress.
With this aim, leaves 4 were harvested immediately at
the end of a 7 h of acute ozone stress and 12 and 24 h
later. The experiment reported in Fig. 8 shows that the
OMT transcripts are detected 12 h after the ozone stress
accordingly with the kinetics of the induction of this
gene [33,34]. These observations confirm that the Tnt1A
Fig. 2. Effect of a 21 days ozone-chronic treatment on the 35S-GUS plants constructs in transgenic tobacco plants. Plants were placed in three
different atmospheres: AA, AA supplied with 20 and 40 nl l�1 for 7 h per day and during 21 days. Leaves were numbered from the bottom (leaf 4) to
the top (leaf 6). Leaves from three 35S-GUS plants (control) were tested for GUS activity. GUS activities were measured independently in each leaf
harvested. Bars represent mean9/S.D. of the three measures for each treatment.
Fig. 3. Effect of a 21 days ozone-chronic treatment on the LTR-GUS and 35S-GUS plants constructs in transgenic tomato plants. Plants were placed
in four different atmospheres: AA, AA supplied with 20, 40, and 60 nl l�1 for 7 h per day and during 21 days. Leaves were numbered from the
bottom (leaf 4) to the top (leaf 8). Leaves from twelve LTR-GUS plants (A) and from three 35S-GUS (B) used as control, were tested for GUS
activity. GUS activities were measured independently in each harvested leaf. Bars represent mean9/S.D. for each treatment.
N. Pourtau et al. / Plant Science 165 (2003) 983�/992 987
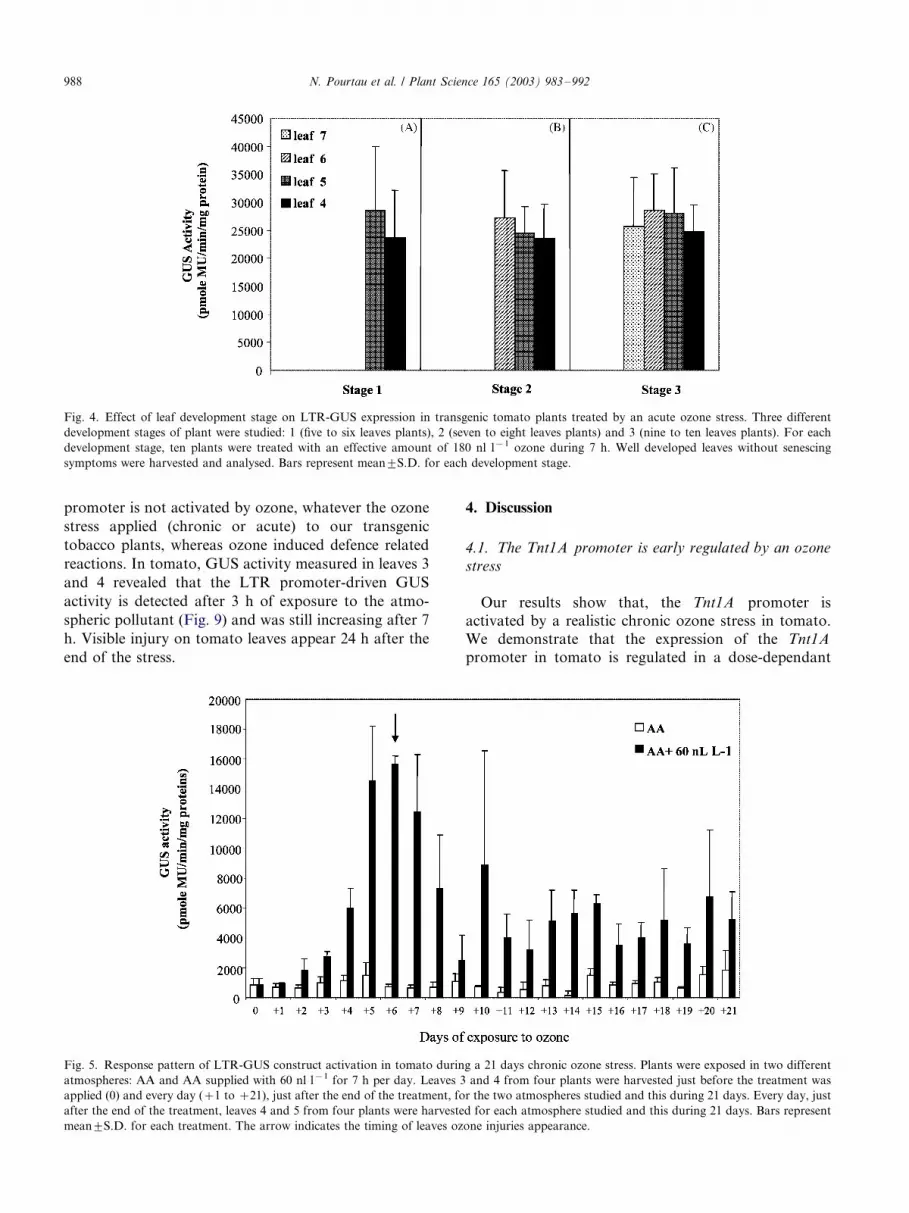
promoter is not activated by ozone, whatever the ozone
stress applied (chronic or acute) to our transgenic
tobacco plants, whereas ozone induced defence related
reactions. In tomato, GUS activity measured in leaves 3
and 4 revealed that the LTR promoter-driven GUS
activity is detected after 3 h of exposure to the atmo-
spheric pollutant (Fig. 9) and was still increasing after 7
h. Visible injury on tomato leaves appear 24 h after the
end of the stress.
4. Discussion
4.1. The Tnt1A promoter is early regulated by an ozone
stress
Our results show that, the Tnt1A promoter is
activated by a realistic chronic ozone stress in tomato.
We demonstrate that the expression of the Tnt1A
promoter in tomato is regulated in a dose-dependant
Fig. 4. Effect of leaf development stage on LTR-GUS expression in transgenic tomato plants treated by an acute ozone stress. Three different
development stages of plant were studied: 1 (five to six leaves plants), 2 (seven to eight leaves plants) and 3 (nine to ten leaves plants). For each
development stage, ten plants were treated with an effective amount of 180 nl l�1 ozone during 7 h. Well developed leaves without senescing
symptoms were harvested and analysed. Bars represent mean9/S.D. for each development stage.
Fig. 5. Response pattern of LTR-GUS construct activation in tomato during a 21 days chronic ozone stress. Plants were exposed in two different
atmospheres: AA and AA supplied with 60 nl l�1 for 7 h per day. Leaves 3 and 4 from four plants were harvested just before the treatment was
applied (0) and every day (�/1 to �/21), just after the end of the treatment, for the two atmospheres studied and this during 21 days. Every day, just
after the end of the treatment, leaves 4 and 5 from four plants were harvested for each atmosphere studied and this during 21 days. Bars represent
mean9/S.D. for each treatment. The arrow indicates the timing of leaves ozone injuries appearance.
N. Pourtau et al. / Plant Science 165 (2003) 983�/992988
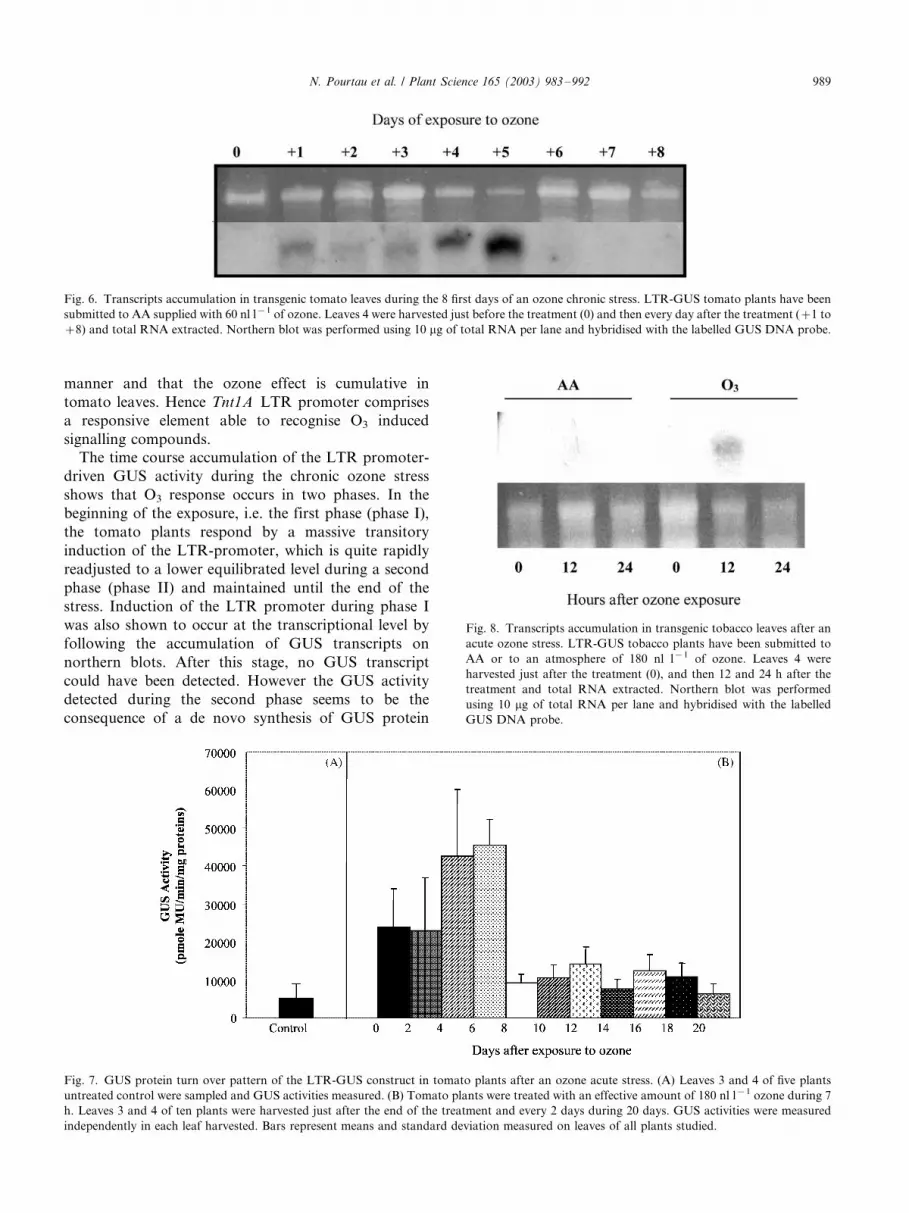
manner and that the ozone effect is cumulative in
tomato leaves. Hence Tnt1A LTR promoter comprises
a responsive element able to recognise O3 induced
signalling compounds.
The time course accumulation of the LTR promoter-
driven GUS activity during the chronic ozone stress
shows that O3 response occurs in two phases. In the
beginning of the exposure, i.e. the first phase (phase I),
the tomato plants respond by a massive transitory
induction of the LTR-promoter, which is quite rapidly
readjusted to a lower equilibrated level during a second
phase (phase II) and maintained until the end of the
stress. Induction of the LTR promoter during phase I
was also shown to occur at the transcriptional level by
following the accumulation of GUS transcripts on
northern blots. After this stage, no GUS transcript
could have been detected. However the GUS activity
detected during the second phase seems to be the
consequence of a de novo synthesis of GUS protein
Fig. 6. Transcripts accumulation in transgenic tomato leaves during the 8 first days of an ozone chronic stress. LTR-GUS tomato plants have been
submitted to AA supplied with 60 nl l�1 of ozone. Leaves 4 were harvested just before the treatment (0) and then every day after the treatment (�/1 to
�/8) and total RNA extracted. Northern blot was performed using 10 mg of total RNA per lane and hybridised with the labelled GUS DNA probe.
Fig. 7. GUS protein turn over pattern of the LTR-GUS construct in tomato plants after an ozone acute stress. (A) Leaves 3 and 4 of five plants
untreated control were sampled and GUS activities measured. (B) Tomato plants were treated with an effective amount of 180 nl l�1 ozone during 7
h. Leaves 3 and 4 of ten plants were harvested just after the end of the treatment and every 2 days during 20 days. GUS activities were measured
independently in each leaf harvested. Bars represent means and standard deviation measured on leaves of all plants studied.
Fig. 8. Transcripts accumulation in transgenic tobacco leaves after an
acute ozone stress. LTR-GUS tobacco plants have been submitted to
AA or to an atmosphere of 180 nl l�1 of ozone. Leaves 4 were
harvested just after the treatment (0), and then 12 and 24 h after the
treatment and total RNA extracted. Northern blot was performed
using 10 mg of total RNA per lane and hybridised with the labelled
GUS DNA probe.
N. Pourtau et al. / Plant Science 165 (2003) 983�/992 989
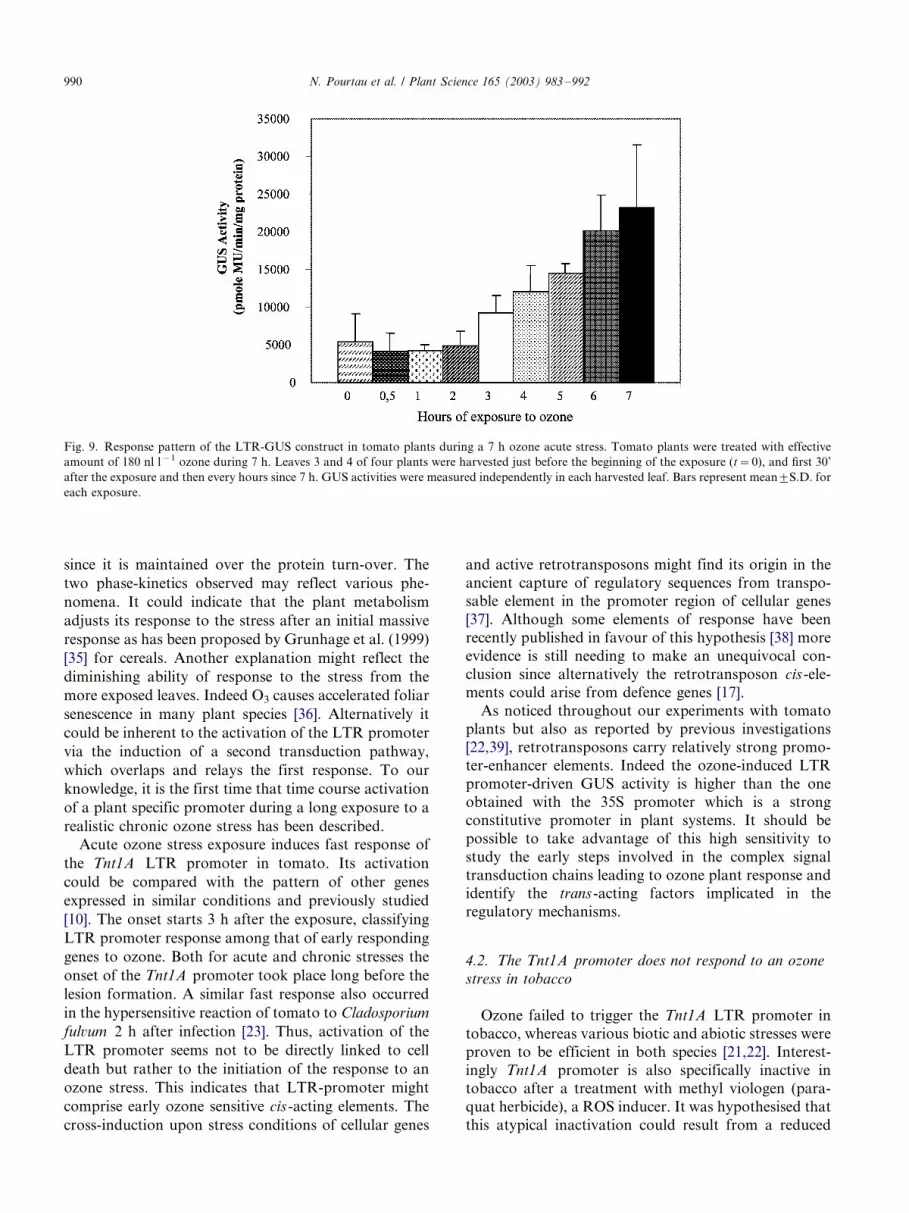
since it is maintained over the protein turn-over. The
two phase-kinetics observed may reflect various phe-
nomena. It could indicate that the plant metabolism
adjusts its response to the stress after an initial massive
response as has been proposed by Grunhage et al. (1999)
[35] for cereals. Another explanation might reflect the
diminishing ability of response to the stress from the
more exposed leaves. Indeed O3 causes accelerated foliar
senescence in many plant species [36]. Alternatively it
could be inherent to the activation of the LTR promoter
via the induction of a second transduction pathway,
which overlaps and relays the first response. To our
knowledge, it is the first time that time course activation
of a plant specific promoter during a long exposure to a
realistic chronic ozone stress has been described.
Acute ozone stress exposure induces fast response of
the Tnt1A LTR promoter in tomato. Its activation
could be compared with the pattern of other genes
expressed in similar conditions and previously studied
[10]. The onset starts 3 h after the exposure, classifying
LTR promoter response among that of early responding
genes to ozone. Both for acute and chronic stresses the
onset of the Tnt1A promoter took place long before the
lesion formation. A similar fast response also occurred
in the hypersensitive reaction of tomato to Cladosporium
fulvum 2 h after infection [23]. Thus, activation of the
LTR promoter seems not to be directly linked to cell
death but rather to the initiation of the response to an
ozone stress. This indicates that LTR-promoter might
comprise early ozone sensitive cis -acting elements. The
cross-induction upon stress conditions of cellular genes
and active retrotransposons might find its origin in the
ancient capture of regulatory sequences from transpo-sable element in the promoter region of cellular genes
[37]. Although some elements of response have been
recently published in favour of this hypothesis [38] more
evidence is still needing to make an unequivocal con-
clusion since alternatively the retrotransposon cis -ele-
ments could arise from defence genes [17].
As noticed throughout our experiments with tomato
plants but also as reported by previous investigations[22,39], retrotransposons carry relatively strong promo-
ter-enhancer elements. Indeed the ozone-induced LTR
promoter-driven GUS activity is higher than the one
obtained with the 35S promoter which is a strong
constitutive promoter in plant systems. It should be
possible to take advantage of this high sensitivity to
study the early steps involved in the complex signal
transduction chains leading to ozone plant response andidentify the trans -acting factors implicated in the
regulatory mechanisms.
4.2. The Tnt1A promoter does not respond to an ozone
stress in tobacco
Ozone failed to trigger the Tnt1A LTR promoter in
tobacco, whereas various biotic and abiotic stresses were
proven to be efficient in both species [21,22]. Interest-
ingly Tnt1A promoter is also specifically inactive intobacco after a treatment with methyl viologen (para-
quat herbicide), a ROS inducer. It was hypothesised that
this atypical inactivation could result from a reduced
Fig. 9. Response pattern of the LTR-GUS construct in tomato plants during a 7 h ozone acute stress. Tomato plants were treated with effective
amount of 180 nl l�1 ozone during 7 h. Leaves 3 and 4 of four plants were harvested just before the beginning of the exposure (t�/0), and first 30’
after the exposure and then every hours since 7 h. GUS activities were measured independently in each harvested leaf. Bars represent mean9/S.D. for
each exposure.
N. Pourtau et al. / Plant Science 165 (2003) 983�/992990

sensitivity of this plant to the herbicide [40]. However, in
the ozone case, a similar hypothesis cannot be proposed,
since tobacco showed a high sensitivity to ozone stress
as noticed previously [41] but also as evidenced throughthe injuries we have observed on the leaves as well as the
activation of defence related genes.
The differential behaviour of Tnt1A promoter in
tobacco and tomato raises the question of whether
similar pathways are induced during ozone stress in the
two species. Indeed discrepancies have been noticed
previously among species since accumulation of H2O2 in
response to O3 has been reported in tobacco [41,42] andbirch [43] whereas in Arabidopsis O3 induced mainly
accumulation of O2+� and H2O2 to a lesser extent
[13,42]. Recent studies suggest that more than one
pathway of ROS generation could exist in plants, even
within a single species [42,44].
Downstream from this complex pattern it has been
suggested that one or multiple targets could be oxidised
upon an ozone exposure resulting in various responsesdepending upon which oxidation events occurred and
which oxidation products were formed [7,8,36]. Addi-
tionally, these target sites could be, eventually, species
dependent. However N. tabacum and L. esculentum
belong to the same taxonomic family (Solanaceae) and it
seems possible that mechanisms conferring plant ozone
response are conserved among these plants [42].
Alternatively, some host-specific regulators of theexpression of the Tnt1A promoter during ozone abiotic
stress could be generated. In tobacco, they could be
responsible for the inactivation of the Tnt1A promoter
under the ozone stress but also after the methylviologen
treatment. This inactivation would take place very early
in the signalling network induced by ozone and methyl-
viologen without affecting plant defence response.
Indeed, previous results have suggested that Tnt1A
activation could be attributed to ROS and not to the
activation of plant defence response in tomato plant
[23]. Moreover, SA, a major phenylpropanoid com-
pound which would constitute a second messenger
downstream in the defence signalling pathway induced
by ozone [45�/47], has been showed to activate Tnt1A
promoter in tobacco [40].
The lack of response of the LTR-promoter to ozonein tobacco might be interpreted as an adaptive phenom-
enon, which would protect against intensive transcrip-
tion and eventually deleterious transposition effect of
the retrotransposon in stress conditions that could be
too prevalent. Indeed we know that ROS are ubiquitous
in plant systems and that they are often form as
byproducts of normal metabolism [48]. The molecular
pathway responsible for this adaptation may have beenselected during coevolution of the Tnt1A retrotranspo-
son and its natural host in order to shut off Tnt1 in the
face of ubiquitous stress conditions such as an oxidative
stress generated by ozone.
Acknowledgements
This research was supported by ‘Elf Aquitaine’, the
‘Region Aquitaine’ and the ‘Conseil General des Pyr-enees Atlantiques’. B.L. was supported by a fellowship
from the ‘Societe de Secours des Amis des Sciences’. We
wish to thank the ‘Laboratoires Departementaux 64’ for
their technical assistance in the OTC maintenance. We
are especially grateful to Cedric Feschotte, Astrid
Wingler, Alec Forsyth and Nigel Bell for critical reading
of the manuscript.
References
[1] S.V. Krupa, W.J. Manning, Atmospheric ozone-formation and
effects on vegetation, Environ. Pollut. 50 (1988) 101�/137.
[2] A. Volz, D. Kley, Evaluation of the Montsouris series of ozone
measurements made in the 19th-century, Nature 332 (1988) 240�/
242.
[3] R. Guderian, Effects of photochemical oxidants on plants, in: R.
Guderian (Ed.), Air Pollution by Photochemical Oxidants:
Formation, Transport, Control and Effects on Plants, Springer,
Berlin, 1985, pp. 129�/333.
[4] N.M. Darrall, The effect of air-pollutants on physiological
processes in plants, Plant Cell Environ. 12 (1989) 1�/30.
[5] P.B. Reich, Quantifying plant response to ozone: a unifying
theory, Tree Physiol. 3 (1987) 63�/91.
[6] E.J. Pell, M.S. Dann, Multiple stress induced foliar senescence
and implications for whole plant, in: H.A. Mooney, W.E. Winner,
E.J. Pell (Eds.), Response of Plants to Multiples Stress, Academic
Press, San Diego, CA, 1991, pp. 189�/204.
[7] O. Blokhina, E. Virolainen, K. Fagerstedt, Antioxidants, oxida-
tive damage and oxygen deprivation stress: a review, Ann. Bot. 91
(2003) 179�/194.
[8] R. Mittler, Oxidative stress, antioxidants and stress tolerance,
Trends Plant Sci. 7 (2002) 405�/410.
[9] H. Moldau, Ozone detoxification in the mesophyll cell wall during
a simulated oxidative burst, Free Radic. Res. 31 (1999) S19�/S24.
[10] H. Sandermann, Ozone and plant health, Annu. Rev. Phyto-
pathol. 34 (1996) 347�/366.
[11] Y.K. Sharma, K.R. Davis, The effects of ozone on antioxidant
responses in plants, Free Radic. Biol. Med. 23 (1997) 480�/488.
[12] M.V. Rao, H. Lee, R.A. Creelman, J.E. Mullet, K.R. Davis,
Jasmonic acid signalling modulates ozone-induced hypersensitive
cell death, Plant Cell 12 (2000) 1633�/1646.
[13] K. Overmyer, H. Tuominen, R. Kettunen, C. Betz, C. Lange-
bartels, H. Sandermann, J. Kangasjarvi, Ozone-sensitive Arabi-
dopsis rcd1 mutant reveals opposite roles for ethylene and
jasmonate signalling pathways in regulating superoxide-depen-
dent cell death, Plant Cell 12 (2000) 1849�/1862.
[14] H. Eckey-Kaltenbach, D. Ernst, W.A. Heller, J. Sandermann,
Cross-induction of defensive phenylpropanoid pathways in pars-
ley, Acta Hort. 381 (1994) 192�/198.
[15] R. Schubert, R. Fischer, R. Hain, P.H. Schreier, G. Bahnweg, D.
Ernst, H. Sandermann, An ozone-responsive region of the
grapevine resveratrol synthase promoter differs from the basal
pathogen-responsive sequence, Plant Mol. Biol. 34 (1997) 417�/
426.
[16] J.D. Boeke, V.G. Corces, Transcription and reverse transcription
of retrotransposons, Ann. Rev. Microbiol. 43 (1989) 403�/434.
[17] A. Kumar, J.L. Bennetzen, Plant retrotransposons, Ann. Rev.
Genet. 33 (1999) 479�/532.
N. Pourtau et al. / Plant Science 165 (2003) 983�/992 991

[18] M.-A. Grandbastien, Activation of plant retrotransposons under
stress conditions (reviews), Trends Plant Sci. 3 (1998) 181�/187.
[19] H. Hirochika, Activation of tobacco retrotransposons during
tissue culture, EMBO J. 12 (1993) 2521�/2528.
[20] R. Kalendar, J. Tanskanen, S. Immonen, E. Nevo, A.H. Schul-
man, From the cover: genome evolution of wild barley (Hordeum
spontaneum ) by BARE-1 retrotransposon dynamics in response to
sharp microclimatic divergence, Proc. Natl. Acad. Sci. USA 97
(2000) 6603�/6607.
[21] S. Pouteau, E. Huttner, M.A. Grandbastien, M. Caboche,
Specific expression of the tobacco Tnt1 retrotransposon in
protoplasts, EMBO J. 10 (1991) 1911�/1918.
[22] M.A. Grandbastien, H. Lucas, J.B. Morel, C. Mhiri, S. Vernh-
ettes, J.M. Casacuberta, The expression of the tobacco Tnt1
retrotransposon is linked to plant defense responses, Genetica 100
(1997) 241�/252.
[23] C. Mhiri, P.J.G.M. De Wit, M.A. Grandbastien, Activation of the
promoter of the Tnt1 retrotransposon in tomato after inoculation
with the fungal pathogen Cladosporium fulvum , Mol. Plant-
Microbe Interact. 12 (1999) 592�/603.
[24] J. Benton, J. Fuhrer, L. Skarby, G. Sanders, Results from the
UN/ECE ICP-crops indicate the extent of exceedance of the
critical levels of ozone in Europe, Water Air Soil Pollut. 85 (1995)
1473�/1478.
[25] A.S. Heagle, D.E. Body, W.W. Heck, An open-top chamber field
chamber to assess the impact of air pollution on plants, J.
Environ. Qual. 2 (1973) 365�/368.
[26] A.S. Heagle, R.B. Philbeck, H.H. Rogers, M.B. Letchworth,
Dispensing and monitoring ozone in open-top field chambers for
plant effects studies, Phytopathology 69 (1979) 15�/20.
[27] J.P. Biolley, B. Lauga, C. Cagnon, R. Duran, J.C. Salvado, P.
Goulas, Phenolic pattern of bean (Phaseolus vulgaris L.) as an
indicator of chronic ozone stress, Water Air Soil Pollut. 106
(1998) 355�/368.
[28] R.A. Jefferson, T.A. Kavanagh, M.W. Bevan, GUS fusions: beta-
glucuronidase as a sensitive and versatile gene fusion marker in
higher plants, EMBO J. 6 (1987) 3901�/3907.
[29] P.K. Pauls, K. Kunert, E. Huttner, M.A. Grandbastien, Expres-
sion of the tobacco Tnt1 retrotransposon promoter in hetero-
logous species, Plant Mol. Biol. 26 (1994) 393�/402.
[30] C. Moreau-Mhiri, J.-B. Morel, C. Audeon, M. Ferault, M.-A.
Grandbastien, H. Lucas, Regulation of expression of the tobacco
Tnt1 retrotransposon in heterologous species-related stresses,
Plant J. 9 (1996) 409�/419.
[31] H. Vaucheret, A. Marionpoll, C. Meyer, J.D. Faure, E. Marin,
M. Caboche, Interest in and limits to the utilization of reporter
genes for the analysis of transcriptional regulation of nitrate
reductase, Mol. Gen. Genet. 235 (1992) 259�/268.
[32] J. Logemann, J. Schell, L. Willmitzer, Improved method for the
isolation of RNA from plant-tissues, Anal. Biochem. 163 (1987)
16�/20.
[33] M. Biagioni, C. Nali, D. a. Heimler, G. Lorenzini, PAL activity
and differential ozone sensitivity in tobacco, bean and poplar, J.
Phytopathol. 145 (1997) 533�/539.
[34] J. Riehl Koch, A.J. Scherzer, S.M. Eshita, K.R. Davis, Ozone
sensitivity in hybrid poplar is correlated with a lack of defense-
gene activation, Plant Physiol. 118 (1998) 124312�/124352.
[35] L. Grunhage, H.J. Jager, H.D. Haenel, F.J. Lopmeier, K.
Hanewald, The European critical levels for ozone: improving
their usage, Environ. Pollut. 105 (1999) 163�/173.
[36] E.J. Pell, C.D. Schlagnhaufer, R.N. Arteca, Ozone-induced
oxidative stress: mechanisms of action and reaction, Physiol.
Plant. 100 (1997) 264�/273.
[37] J.F. McDonald, Macroevolution and retroviral elements: inser-
tion of viral-like DNA segments may bring about rapid and
dramatic changes in gene regulation and development, Bioscience
40 (1990) 183�/191.
[38] S. Takeda, K. Sugimoto, H. Otsuki, H. Hirochika, A 13-bp cis-
regulatory element in the LTR promoter of the tobacco retro-
transposon Tto1 is involved in responsiveness to tissue culture,
wounding, methyl jasmonate and fungal elicitors, Plant J. 18
(1999) 383�/393.
[39] H. Hirochika, H. Otsuki, M. Yoshikawa, Y. Otsuki, K. Sugi-
moto, S. Takeda, Autonomous transposition of the tobacco
retrotransposon Tto1 in rice, Plant Cell 8 (1996) 725�/734.
[40] C. Mhiri, J.B. Morel, S. Vernhettes, J.M. Casacuberta, H. Lucas,
M.A. Grandbastien, The promoter of the tobacco Tnt1 retro-
transposon is induced by wounding and by abiotic stress, Plant
Mol. Biol. 33 (1997) 257�/266.
[41] M. Schraudner, W. Moeder, C. Wiese, W. Van Camp, D. Inze, C.
Langebartels, H. Sandermann, Ozone-induced oxidative burst in
the ozone biomonitor plant, tobacco Bel W3, Plant J. 16 (1998)
235�/245.
[42] H. Wohlgemuth, K. Mittelstrass, S. Kschieschan, J. Bender, H.J.
Weigel, K. Overmyer, J. Kangasjarvi, H. Sandermann, C.
Langebartels, Activation of an oxidative burst is a general feature
of sensitive plants exposed to the air pollutant ozone, Plant Cell
Environ. 25 (2002) 717�/726.
[43] R. Pellinen, T. Palva, J. Kangasjarvi, Short communication:
subcellular localization of ozone-induced hydrogen peroxide
production in birch (Betula pendula ) leaf cells, Plant J. 20
(1999) 349�/356.
[44] C. Langebartels, H. Wohlgemuth, S. Kschieschan, S. Grun, H.
Sandermann, Oxidative burst and cell death in ozone-exposed
plants, Plant Physiol. Biochem. 40 (2002) 567�/575.
[45] M.V. Rao, K.R. Davis, The physiology of ozone induced cell
death, Planta 213 (2001) 682�/690.
[46] M.V. Rao, H. Lee, K.R. Davis, Ozone-induced ethylene produc-
tion is dependent on salicylic acid, and both salicylic acid and
ethylene act in concert to regulate ozone-induced cell death, Plant
J. 32 (2002) 447�/456.
[47] W. Moeder, C.S. Barry, A.A. Tauriainen, C. Betz, J. Tuomainen,
M. Utriainen, D. Grierson, H. Sandermann, C. Langebartels, J.
Kangasjarvi, Ethylene synthesis regulated by biphasic induction
of 1- aminocyclopropane-1-carboxylic acid synthase and 1-
aminocyclopropane-1-carboxylic acid oxidase genes is required
for hydrogen peroxide accumulation and cell death in ozone-
exposed tomato, Plant Physiol. 130 (2002) 1918�/1926.
[48] G.P. Bolwell, Role of active oxygen species and NO in plant
defence responses, Curr. Opin. Plant Biol. 2 (1999) 287�/294.
N. Pourtau et al. / Plant Science 165 (2003) 983�/992992





























