Embed Size (px)
Citation preview

VIROLOGY 190, 654-665 (1992)
Genetic Relationships between Bovine Herpesvirus 4 and the Gammaherpesviruses Epstein-Barr Virus and Herpesvirus Saimiri
MICHEL BUBLOT,* PATRICK LOMONTE,” ANNE-SOPHIE LEQUARRE,* JENS-CHRISTIAN ALBRECHT,1- JOHN NICHOLAS,+* BERNHARD FLECKENSTEIN,t PAUL-PIERRE PASTORET,” AND ETIENNE THIRY*v’
*Virology Department, Faculty of Veterinary Medicine, University of Lisge, B6, Sart Tilman, B-4000 Lisge, Belgium; tlnstitut fur Klinische und Molekulare Virologie der Friedrich-Alexander-Universita’t Erlangen-NUrnberg, 7, Loschgestrasse, D-8520 Erlangen, Germany; and *Division
of Virology, National Institute for Medical Research, The Ridgeway, Mill Hill, London NW7 IAA, United Kingdom
Received January 3 1, 1992; accepted June 3, 1992
The overall arrangement of genes in the unique central part of the bovine herpesvirus type 4 (BHV-4) genome has been deduced by analysis of short DNA sequences. Twenty-three genes conserved in at least one of the completely sequenced herpesviruses have been identified and localized. All of these genes encoded amino acid sequences with higher similarity to proteins of the gammaherpesviruses Epstein-Barr virus (EBV) and herpesvirus saimiri (HVS) than to the homologous products of the alphaherpesviruses varicella-zoster virus and herpes simplex virus type 1 or the betaherpesvirus human cytomegalovirus. The genome organization of BHV-4 had also an overall collinearity with that of the gammaherpesviruses EBV and HVS. Furthermore, the BHV-4 genes content and arrangement were more similar to those of HVS than to those of EBV, suggesting that BHV-4 and HVS are evolutionarily more closely related to each other than either are to EBV. BHV-4 DNA sequences were generally deficient in CpG dinucleotide. This CpG deficiency is characteristic of gammaherpesvirus genomes and suggests that the BHV-4 latent genome is extensively methylated. Despite several biological features similar to those of betaherpesviruses, BHV-4 displays the molecular characteristics of the representative members of the gammaherpesvirinae subfamily. 0 1992 Academic PWSS. h.
INTRQDUCTION
Members of the family Herpesviridae are classified, primarily on the basis of biological behavior, into three major subfamilies: the Alphaherpesvirinae, including the neurotropic herpes simplex virus 1 (HSV-1) and 2 (HSV-2) and varicella-zoster virus (VZV); the Betaher- pesvirinae, including cytomegaloviruses; and the Gam- maherpesvirinae, including the lymphotropic Epstein- Barr virus (EBV) and herpesvirus saimiri (HVS) (Honess and Watson, 1977; Roizman, 1982).
Analysis of complete (Baer et a/., 1984; Davison and Scott, 1986; McGeoch et a/., 1988; Chee et al., 1990; J. C. Albrecht, J. Nicholas, D. Biller, K. R. Cameron, B. Biesinger, C. Newman, S. Wittmann, M. A. Craxton, H. Coleman, B. Fleckenstein and R. W. Honess, submit- ted) or partial sequences (see for example, Gompels et al., 1988) of several mammalian herpesviruses has al- lowed the identification of genes common to all these viruses as well as the recognition of genes specific to either a particular virus or virus subfamily. The con-
Sequence data from this article have been deposited with the EMBUGenBank Data Libraries under Accession Nos. M90768 through M90800.
’ To whom reprint requests should be addressed. p Present address: Department of Oncology, Johns Hopkins
School of Medicine, 418 North Bond Street, Baltimore, MD 21231.
served genes are clustered in blocks of genes and the arrangement of these blocks is identical for the members of a subfamily but differs from the other sub- families (Davison and Taylor, 1987; Gompels et a/., 1988; McGeoch, 1989; Kouzarides et al., 1987). An- other molecular criterion which can differentiate the three subfamilies is the CpG dinucleotide frequency: there is a global deficiency in CpG dinucleotide in the genomes of gammaherpesviruses, a local deficiency in the major immediate-early genes of betaherpesvi- ruses, and no deficiency at all in alphaherpesviruses genomes. The CpG deficiency is probably the result of methylation of the latent genome (Honess et al., 1989).
There is a correlation between these biological and molecular criteria for most herpesviruses. Neverthe- less, Marek’s disease virus (MDV), its close relative being the herpesvirus of turkey (HVT), and human her- pesvirus 6 (HHV-6), which present biological features of gammaherpesviruses, do not share the molecular characteristics of these viruses. MDV and HVT ge- names have the same gene content and gene organiza- tion as alphaherpesviruses (Buckmaster et a/., 1988) and HHV-6 is closely related to betaherpesviruses (Lawrence el a/., 1990; Josephs et al., 1991; Neipel et a/., 1991; Martin et al., 1991); furthermore, they are not globally CpG-deficient (Honess et a/., 1989; Lawrence et a/., 1990; Martin et a/., 1991). These results show that the classification on the basis of certain biological
0042.6822/92 $5.00 Copyright 0 1992 by Academic Press, Inc. All rights of reproduction in any form reserved.
654

GENE ORGANIZATION OF BHV-4 655
properties does not necessarily reflect a true evolution- ary relationship.
The group of bovine herpesvirus 4 (BHV-4) includes a collection of antigenically related isolates distinct from other bovine herpesviruses. These viruses are dis- tributed worldwide and are isolated from a variety of clinical diseases and even healthy cattle (reviewed in Thiry et a/., 1990, 1992). It is mainly isolated from cattle and, in some African regions, from buffalo (Syncerus cakier) (Rossiter et al., 1989). It has been also occa- sionally isolated from American bison (Bison bison) (Todd and Storz, 1983) and sheep (Van Opdenbosh et a/., 1986). Feline cell-associated herpesvirus (Fe- CAHV) isolated from cat (Fabricant et a/., 1971) and herpesvirus aotus type 2 (HVA-2) isolated from owl monkey (Aotus trivirgatus) (Barahona et al., 1973) have been shown to be BHV-4 strains (Kit et al., 1986; Kruger et al., 1989; Bublot et al., 1991 b; Dubuisson et al., 1991).
On the basis of some of its biological characteristics, BHV-4 was assigned to be a member of the betaher- pesvirinae subfamily (bovine cytomegalovirus) (Storz et a/., 1984); however, recent molecular data have shown a clear relationship with the members of the gamma- herpesvirinae subfamily (Honess, 1984; Kit et a/., 1986; Bublot et al., 1990, 1991 a; Van Santen, 1991; Thiry et a/., 1992).
The genome structure of BHV-4 (V. Test strain) is similar to that of HVS and consists of a unique coding part (LiDNA) of approximately 108 kb flanked by (G + C)-rich tandem repeats of 2.65 kb called polyrepeti- tive DNA (prDNA or H-DNA) (Ehlers eta/., 1985; Bublot et a/., 1990). The H-DNA contains potential open read- ing frames (M. Goltz and H.-J. Buhk, manuscript in prep- aration) but no RNA derived from these repetitive se- quences has been detected (V. L. Van Santen and L.-Y. Chang, unpublished results).
In order to determine the genetic relationship of BHV-4 to well characterized representative members of the alpha-, beta-, and gamrnaherpesvirinae subfami- lies, an analysis of short nucleotide sequences from cloned restriction fragments distributed across the coding region of the viral genome has been carried out. This analysis has clearly shown a genetic relationship of BHV-4 with gammaherpesviruses EBV and HVS; fur- thermore, BHV-4 is more closely related to HVS than to EBV.
MATERIALS AND METHODS
Recombinant plasmids
Several cloned EcoRl fragments from BHV-4 (V. Test strain) DNA have been chosen (Bublot et al., 1990). Some of them have been subcloned (using HindIll or
BarnHI) in the phagemid Bluescript pSK+ (Stratagene, La Jolla). Orientation of most of these clones was de- termined by restriction analysis (Bublot et al., 1990). Other ones were oriented using homology results of sequences from these clones and those from contigu- ous ones, with genes of well-characterized herpesvi- ruses.
DNA sequencing
Nucleotide sequences were determined on dsDNA by the dideoxynucleotide chain termination method (Sanger et al., 1977) using the T7-sequencing Kit (Pharmacia LKB, Uppsala, Sweden). Sequences were obtained from one or both ends of the cloned frag- ments using appropriate primers recognizing a plasmid sequence and were therefore derived from one strand of the DNA.
Computer analysis of sequence data
The sequence data were compiled using the soft- ware package version 7.0-Unix of the University of Wis- consin Genetic Computer Group installed on Ultrix (Devereux et al., 1984). Nucleotide sequences were translated into all six reading frames and resulting amino acid sequences were compared to protein data- bases (SWISS-PROT, EMBL release 17.0) and to the herpesvirus saimiri amino acid sequences using the FASTA program (Ktuple of 2) (Pearson and Lipman, 1988). Comparisons based on amino acid similarities and identities have been done with the GAP program using a gap weight of 3.0 and a length weight of 0.1 (Devereux et al., 1984). Multiple alignments have been done using the program PILEUP (gap weight of 3.0 and length weight of 0.1) (Feng and Doolittle, 1987; Higgins and Sharp, 1989).
Dinucleotide frequencies analysis
Observed frequencies (0) of dinucleotides were counted and expected frequencies (e) of dinucleotides were calculated from observed mononucleotide fre- quencies. Deviations of the observed from the ex- pected frequencies of dinucleotides were first dis- played as o/e when o > e or as e/o when e > o (Fig. 4a). In order to examine the correlation between the defi- ciency of CpG dinucleotide and the excess of TpG and CpA dinucleotides, the absolute magnitudes of the dif- ferences o - e were computed (Fig. 4b) (Honess et al,, 1989).

656 BUBLOT ET AL.
EOORI map: O5 kbp
35 31 33
FIG. 1. Localization of sequenced regions on BHV4 (V. Test strain) genome (EcoRI map).
RESULTS
1. Localization of sequenced regions
Thirty-three sequences of the coding part (L-DNA) of BHV-4 (V. Test strain) genome were obtained; they are localized on the EcoRl restriction map in Fig. 1.
The total length of the sequenced regions was 9569 bp (about 9% of the L-DNA) with a (G + C) content of 40.3%.
2. Identification of conserved genes
Homologues of 27 translated sequences were pres- ent in one of the completely sequenced mammalian herpesviruses (EBV, VZV, HSV-1, HCMV, and HVS). The similarity was always greater to EBV and HVS genes than to homologous products of alphaherpes- virusesVZVand HSV-1 or betaherpesvirus human cyto- megalovirus (HCMV). Twenty-three HVS homologues were identified (Table 1). Nineteen of these HVS homo- logues belong to blocks of genes which are conserved among gamma- and alphaherpesviruses and/or beta- herpesviruses.
Five BHV-4 sequences (21,22,23,30, and 31) were found to be homologous to gammaherpesvirus-spe- cific genes. The BHV-4 sequences 21,22, and 23 were homologous to the products of BRRF2, BRRFI, and BLRF2 of EBV (Baer et al., 1984) and those of HVS 48, 49, and 52 (J. C. Albrecht, J. Nicholas, D. Biller, K. R. Cameron, B. Biesinger, C. Newman, S. Wittmann, M. A. Craxton, H. Coleman, 8. Fleckenstein and R. W. Honess, submitted) (corresponding to EDLF5, EDLF4, and EDLFS, respectively, in Nicholas eta/., 199213) (Ta- ble 1). These genes are not conserved in the alpha- and betaherpesviruses. The two other sequences (30 and 31; Table 1) were homologous to the EBV 140,000-M, membrane antigen encoded by BNRFl. This gene is conserved in the gammaherpesviruses HVS (gene 75 or ElLFl in Nicholas eta/., 199213) (Cam- eron et a/., 1987) and murine herpesvirus 68 (Ef- stathiou et a/., 1990b) but no related gene has been identified in the alpha- and betaherpesviruses. The BHV-4 sequences 30 and 3 1 were also homologous to the HVS gene 3, a second BNRFl homologue in the HVS genome (J. C. Albrecht, J. Nicholas, D. Biller, K. R. Cameron, B. Biesinger, C. Newman, S. Wittmann,
M. A. Craxton, H. Coleman, B. Fleckenstein, and R. W. Honess, submitted) (data not shown).
The BHV-4 amino acid sequences were therefore more closely related to that of the gammaherpesvi- ruses than that of the alpha- or betaherpesviruses.
3. Homology between BHV-4, HVS, and EBV sequences
Figure 2 shows alignments of BHV-4 amino acid se- quences with those of homologous products of several herpesviruses. BHV-4 sequence 14a was homologous to the N-terminal part of spliced gene exon 2 which is highly conserved in all herpesviruses subfamilies (Fig. 2a); the DNA sequence of the predicted splice accep- tor site was also very well conserved (data not shown) (Davison and Scott, 1986; Lawrence et al., 1990). BHV-4 sequence 6 was homologous to a part of the gene coding for the HSV-1 glycoprotein B which is also conserved in all herpesviruses subfamilies. This 59- amino-acid BHV-4 sequence contains three potential IV-glycosylation sites; two sites are conserved only with HVS and the third one is conserved with the two gammaherpesviruses as well as with alphaherpesvi- ruses HSV-I and VZV (Fig. 2b).
Alignment of BHV-4 sequence 25, that is homolo- gous to a part of the small subunit of ribonucleotide reductase, is shown in Fig. 2c; the betaherpesvirus HCMV genome does not encode this small subunit, although it encodes the large subunit of this enzyme (Chee et al., 1990). Figure 2d shows alignment of BHV- 4 sequence 14b which was found to be homologous to the gammaherpesviruses HVS gene 26 and EBV BDLF3.5 and the betaherpesvirus HCMV UL91 but not to any alphaherpesvirus gene: EBV BDLF3.5 is a new EBV open reading frame located between BDLF3 and BDLF4 and discovered by analysis of the complete se- quence of HVS (J. C. Albrecht, J. Nicholas, D. Biller, K. R. Cameron, B. Biesinger, C. Newman, S. Wittmann, M. A. Craxton, H. Coleman, B. Fleckenstein, and R. W. Honess, submitted). Alignment of BHV-4 sequence 30 with that of the membrane antigen (HVS 75 and EBV BNRFl genes product), which is a gammaherpesvirus- specific gene, is also shown (Fig. 2e).
Table 1 indicates the percentage of identical and similar amino acids between the homologous se- quences of BHV-4, HVS, and EBV. The most con-
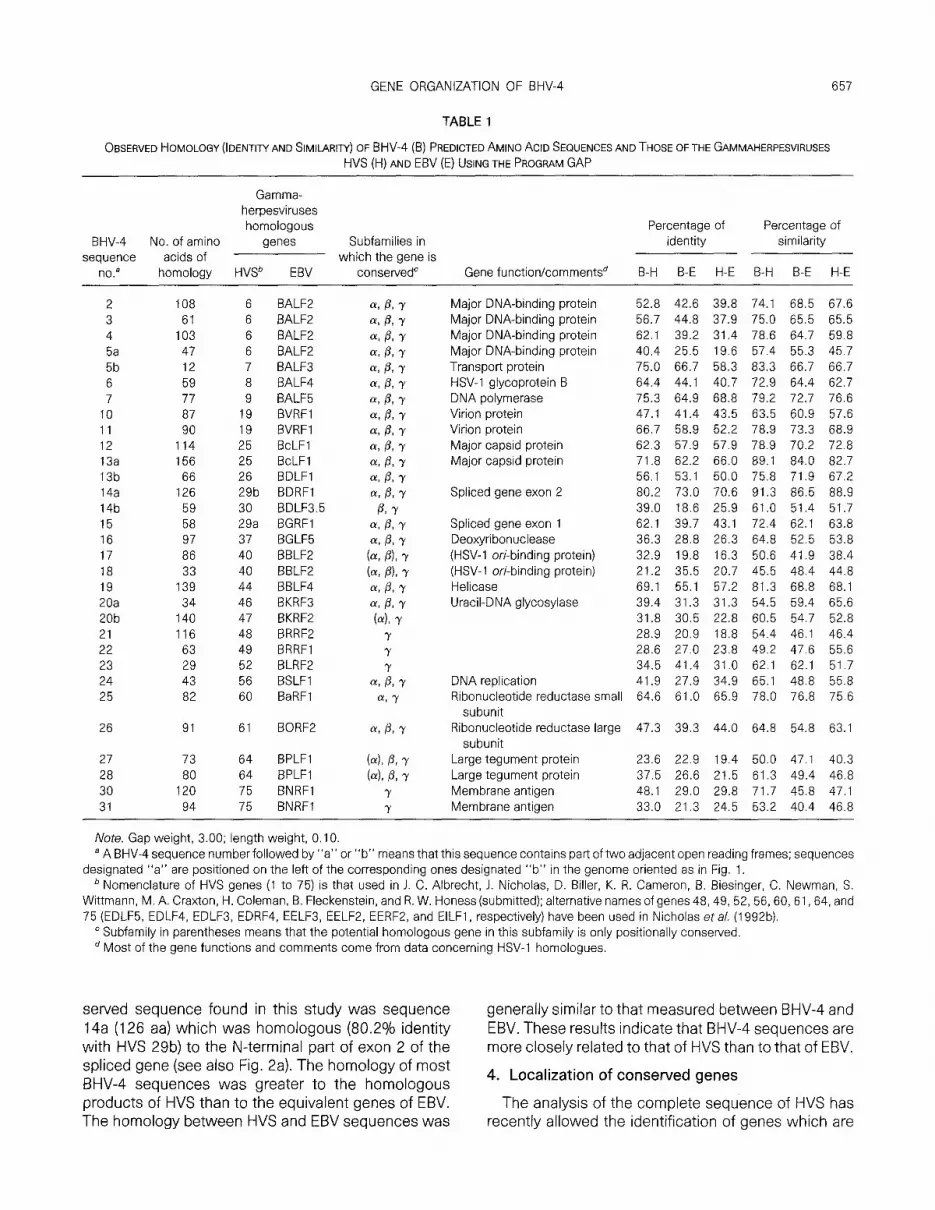
GENE ORGANIZATION OF BHV-4 657
TABLE 1
OBSERVED HOMOLOGY (IDENTIIY AND SIMILARIT/) OF BHV-4 (E?) PREDICTED AMINO ACID SEQUENCES AND THOSE OF THE GAMMAHERPESVIRUSES HVS (H) AND EBV (E) USING THE PROGRAM GAP
Gamma- herpesviruses homologous Percentage of Percentage of
BHV4 No. of amino genes Subfamilies in identity similarity sequence acids of which the gene is
no.8 homology HVSb EBV conservedC Gene function/commentsd B-H B-E H-E B-H B-E H-E
2 108 6 BALF2 3 61 6 BALF2
4 103 6 BALF2 5a 47 6 BALF2 5b 12 7 BALF3
6 59 8 BALF4 7 77 9 BALFS
10 87 19 BVRFl 11 90 19 BVRFl 12 114 25 BcLFl 13a 156 25 BcLFl 13b 66 26 BDLFl 14a 126 29b BDRFl 14b 59 30 BDLF3.5 15 58 29a BGRFl
16 97 37 BGLF5 17 86 40 BBLF2 18 33 40 BBLF2
19 139 44 BBLF4 20a 34 46 BKRF3 20b 140 47 BKRF2 21 116 48 BRRF2 22 63 49 BRRFl 23 29 52 BLRF2 24 43 56 BSLFl 25 82 60 BaRFl
26 91
27 73 28 80 30 120 31 94
61 BORF2
64 BPLFl 64 BPLFl 75 BNRFl 75 BNRFl
Major DNA-binding protein Major DNA-binding protein Major DNA-binding protein Major DNA-binding protein Transport protein HSV-1 glycoprotein B DNA polymerase Virion protein Virion protein Major capsid protein Major capsid protein
Spliced gene exon 2
Spliced gene exon 1 Deoxyribonuclease (HSV-1 ori-binding protein) (HSV-1 ori-binding protein) Helicase Uracil-DNA glycosylase
DNA replication Ribonucleotide reductase small
subunit Ribonucleotide reductase large
subunit Large tegument protein Large tegument protein Membrane antigen Membrane antigen
52.8 42.6 39.8 74.1 68.5 67.6 56.7 44.8 37.9 75.0 65.5 65.5 62.1 39.2 31.4 78.6 64.7 59.8 40.4 25.5 19.6 57.4 55.3 45.7 75.0 66.7 58.3 83.3 66.7 66.7 64.4 44.1 40.7 72.9 64.4 62.7 75.3 64.9 68.8 79.2 72.7 76.6 47.1 41.4 43.5 63.5 60.9 57.6 66.7 58.9 52.2 78.9 73.3 68.9 62.3 57.9 57.9 78.9 70.2 72.8 71.8 62.2 66.0 89.1 84.0 82.7 56.1 53.1 50.0 75.8 71.9 67.2 80.2 73.0 70.6 91.3 86.5 88.9 39.0 18.6 25.9 61.0 51.4 51.7 62.1 39.7 43.1 72.4 62.1 63.8 36.3 28.8 26.3 64.8 52.5 53.8 32.9 19.8 16.3 50.6 41.9 38.4 21.2 35.5 20.7 45.5 48.4 44.8 69.1 55.1 57.2 81.3 68.8 68.1 39.4 31.3 31.3 54.5 59.4 65.6 31.8 30.5 22.8 60.5 54.7 52.8 28.9 20.9 18.8 54.4 46.1 46.4 28.6 27.0 23.8 49.2 47.6 55.6 34.5 41.4 31.0 62.1 62.1 51.7 41.9 27.9 34.9 65.1 48.8 55.8 64.6 61.0 65.9 78.0 76.8 75.6
47.3 39.3 44.0 64.8 54.8 63.1
23.6 22.9 19.4 50.0 47.1 40.3 37.5 26.6 21.5 61.3 49.4 46.8 48.1 29.0 29.8 71.7 45.8 47.1 33.0 21.3 24.5 53.2 40.4 46.8
Note. Gap weight, 3.00; length weight, 0.10. a A BHV-4 sequence number followed by “a” or “b” means that this sequence contains part of two adjacent open reading frames; sequences
designated “a” are positioned on the left of the corresponding ones designated “b” in the genome oriented as in Fig. 1. ’ Nomenclature of HVS genes (1 to 75) is that used in J. C. Albrecht, J. Nicholas, D. Biller, K. R. Cameron, B. Biesinger, C. Newman, S.
Wittmann, M. A. Craxton, H. Coleman, B. Fleckenstein, and R. W. Honess (submitted); alternative names of genes 48,49, 52, 56, 60, 61, 64, and 75 (EDLF5, EDLF4, EDLFB, EDRF4. EELFB, EELF2, EERF2, and ElLFl , respectively) have been used in Nicholas et al. (1992b).
’ Subfamily in parentheses means that the potential homologous gene in this subfamily is only positionally conserved. d Most of the gene functions and comments come from data concerning HSV-1 homologues.
served sequence found in this study was sequence generally similar to that measured between BHV-4 and 14a (126 aa) which was homologous (80.2% identity EBV. These results indicate that BHV-4 sequences are with HVS 29b) to the N-terminal part of exon 2 of the more closely related to that of HVS than to that of EBV. spliced gene (see also Fig. 2a). The homology of most BHV-4 sequences was greater to the homologous
4. Localization of conserved genes
products of HVS than to the equivalent genes of EBV. The analysis of the complete sequence of HVS has The homology between HVS and EBV sequences was recently allowed the identification of genes which are

658 BUBLOT ET AL.
a BEIY-4
EYS EBY EICMY E8Y-6 my-1 YZV
SW-4
EYS EBV EICMY KEY-6 ESY-1 Y&Y
BEY-4
BYS EBY BCMY HRV-6 RSY-1 VZY
b BBV-4
EYS EBV BCMV ESY-1 VZY
C BEV-4
EVS EBY YZV ESY-1
BBY-4
EVS EBY YZY ESV-1
d BEV-4
BYS EBY ECMY
e EVS
BEY-4:
EBV
EVS
BBY-4:
EBV
10 20 30 40 50 60 SIRGQTFNILWDEANFIFX.EALPAILGFMLQKDARIIFISSVNSADKSTSFLFN~NAR IlIIIIII:II:IIIIIIII::IIIIIIIIIIIIII:IIIIIIII:I::IlIIlII:Il:
SIRGOTPNLLYIDEANFIKKDSLPAILGFMLQKD~LIFISSVNSGDRATSFLPNLKNAS SIR~TFELLFVDEANFIKKEALPAILGFMLQKDAKIIFISSVNSADQATSFLYKLKDAQ SIRGQNFBLLLVDEABFIKKEAFNTILGFLAQNTTKIIFISSTNTTSDSTCFL~NNAP SIRGQSFNLLIYDESEFIID~STILGFLPQASTRILLNNSP GIRGQDFNLLFVDEANFIRPDAYQTIHGFLNQANCKIIFVSSTNTGKASTSFLYNLRGAA GIRGQDFNLLRrDEANFIRPDAV~IYGFLNPTNCKII~SSTNT~STSFLYN~GSS
l *** l l l * t t t ** * l l *II t ” l * l
70 80 90 100 110 120 EKMLNVVNYVCPEEKEDFNLQSTLTSCPCYRLBIPTYITIDESIKNTTNLFLDDVFT~L IIIII:III:II:II:II:II::I:IIIIl:I:IIIIIIIII:IIIIIIIIII::IIIII
EKMLNIVNYICPDEKDDFSLQDSLISCPCYLYIPTYITIDETIKNTTNLFLD~T~L ERLLNWSWCOEERODFDMODSMVSCPCPC~EIPSYITMDSNI~TTNLFLDGAFSTEL FDMLNWSWCEEELESFTEKGDATACPCYRLBKPTFISLNSQVRKTANMFMPGAFMDEI FEHLSWSWCEDEABMLNERGNATACSCYRLEKPKFISINAEVKKTANLFLEGAFIEEI DELLNWTYICDDEMPRWTETNATACSCYILKRPVFITnDGI DQLLNWTYVCDDEHPRVLABSDYTACSCYVLNKPYFITMDGAl4RRTADLFMADSFVQEI
l l ** l l * l l t l l l t
MGDIST IIl:l:
NGDMSG (304-429) MGDTSS 1326-4411 IGGTNK (‘297-422) MGGATC (293-418) IGGQAR (344-469) VGGRXQ (353-478)
*
10 20 30 40 50 RSLFFVK~EYFKDYKFVKTMDTN~TLDTFLTL~FIDNIDFKTVKLYSETEP.KM
::I :I1 l::Il:I I:I: :/IIIIIIIII:IIIIII:IIIIl:IIIlI::III: ETFFTAK~YBFKNYVBVETLPVN~TLDTFLAL~FIENIDFFAVELYSSGERKL QYYFQSGNEIBVYNDYBBFKTIELDGIATLQTFISLNTS LKIFIAGNSAYEYVDYLFKRMIDLSSISTVDSMIALDIDPLENTDFRVIXLYSQKELRS RRYFTFGGGYVYFKEYAYSBQLSRADITTVSTFIDL~KLaEDEEE%'PLEYYTREEIKD KRYFLFGBBYVYYEDYRYVREIAVBDYGMISTYVDL~LLKDREFMPLQYYTRDELRD
t t l * l *
10 20 30 40 50 60 VNGVCLANDYISRDELLBTRAAALL YNTMVSGADKPEIILWVBNLFKKAVEVKREFILVKS :ll:illllll:IIl:lil:lllllllll::::::I I:::Il:IlIIlI :/I :I:
MricI~~biriARo~rae~S~~~~~SS~R~SED~I~~~~~~~Fi~G MPGICLANNYISRDELLETRAASLLYNSMTAKADRPRATWIQELFRTAVKVXTAFIEARG FWTCQFNDLISRDEAIBTSASCCIYNNYVP--EKPAITRIEQLFSEAVEIECAFLKSBA LRVTCQSNDLISRDEAYETTASCYIYNNYLGPPPDRYY~~~~IEIGFIRSQA
l t t .** t t t l * l t* t t t l t
70 80 SG----YSAVNVDDIRSFLCATADRI
I II lIl::II Il:llIIII YG----VSLVNVWIRQFLQATADRI EG----YTLVDVRAIKQFLKATADRI PK----l'RLVNVDAITQYVKFSADRL PTDSEILSPAALAAIENYVRFSADRL
* *tt
10 20 30 40 50 MDSGSLTEKDFSDCKBFFSQP~RLIDD~S~NDI-DL~~IEN~~SLLLDLVG~ I :: t:l:Il:Il: Il::II I::: l::l::l I: :I: :::::I:::lII:III
(185-266) (174-255) (179-258) (207-292)
kSKPCIS~R~~?rb~QAiP~~PI~Y~I~KS~i-DW KKRGTIGBREFGELLSWDPTDLPRTVARVYVAVGGLFKQEYSEYQRLKNICTLLDLAGVB
MNSLLARLNRLGVABATTKDVFIFVDRLFQBFSFLFQAEESGPRRXXLYASVFEBLTVS l l
:562-620) 1593-651) (615-673) [ 638-696) 1587-645)
(l-59) (12-71) (l-59)
10 20 30 40 50 60 P~~~~~.~T.~TP~AS~~---RDTPTPDTPIQDTD~~~~~~~--N~TAS~YTS-IERITPA
--I,-,~,,,, :: I::1 II:) l:I(Il:I ::l:Il: IIGLSFTSASDSIPMGE~SMDPTAMDLGIPTFINTPNF
-..-- --_ ..-. - --------- --. I:1 I:I(ItII:I:IIII
EFCRDL1mmm~ : : ::I:::1 :I:I I::I ::: I l:l:Il: : :I\
ASDY~GLcvKLTFGsAs--CPETGSSASNF~---------~~V~~~FSGPLITPV t * l ** l t l *
70 80 90 100 110 LKXAENALYBVCLSKELTLSGSVnNSPThPSSHLPDLDTSKUtDMFYAVKBLISKNLY (768-875) I(:::::l: :::II::II:II:II: )I: : III:::I::I::II Il:l:l:Il:
LKMDGSSLICLSISKQVTLAGSTFKBIFTEQIE~PDYSSSQIRNLFYLVKKLMSENLI ):::II 11 :: :: : :I1 Il::l:: :: :: :t ::I11 I::I:::::: (770-878)
LQRTGSLLIAYRCGDGRIQGGSLFBQLFSDVATTPRAPBALS~NLPaAVPQLVXSGIY l l l * t* * 1 t l
FIG. 2. Amino acid comparisons between BHV-4 sequences and parts of homologous products of other herpesviruses: (a) BHV-4 sequence l4a and the N-terminal parts of the spliced genes exons 2 (conserved in alpha-, beta-, and gammaherpesviruses), (b) BHV-4 sequence 6 and genes coding for HSV-1 glycoprotein B homologues (conserved in alpha-, beta-, and gammaherpesviruses), (c) BHV-4 sequence 25 and

GENE ORGANIZATION OF BHV-4 659
TABLE 2
COMPARISON OF THE ESTIMATED LENGTH OF BHV-4 GENE BLOCKS WITH THOSE OF GAMMAHERPESVIRUSES HVS AND EBV
Genomic regions HVS*
Gene content
EBV” HVS
Length in kbpa
BHV-4 EBV
Left end to block 1 1 to 5 BNLFl a to BALFl 12.6 R, V 5.7 v 5.4 R, V Block 1 6 to 9 BALF2 to BALF5 11.0 11.0 11.1 Block 1 to block 2 10 to 16 BlLFl to BdRFl 7.3 6.8 R, V 4.0 R. V Block 2 17to47 BVRF2 to BKRF2 36.5 37.0 39.8 Block 2 to block 3 BKRFl <O.l <O.l 2.0 R, V Block 3 48 to 50 (EDLF5 to EDRFl) BRRF2 to BRLFl 5.1 R 4.5 4.5 Block 3 to block 4 51 (EDRF2) BZLFl to BLLFla 1.0 1.3 14.0 R, V Block 4 52 to 69 (EDLF3 to ECRF2) BLRF2 to BFLF2 27.9 28.3 33.4 R Block 4 to block 5 70 to 74 (ECLF4 to ECRF3) BHLFl to BCRFl 7.4 R 5.4 v 50.3 R, V Block 5 75 (EILFl) BNRFl 3.9 3.9 4.0 Block 5 to right end 0.2 4.0 1.7
a R, region contains repeats; V, region varies in size between strains or isolates. * Nomenclature of HVS genes (1 to 75) is that used in J. C. Albrecht, J. Nicholas, D. Biller, K. R. Cameron, B, Biesinger, C. Newman, S.
Wittmann, M. A. Craxton, H. Coleman, 6. Fleckenstein, and R. W. Honess (submitted); alternative names of genes used in Nicholas eta/. (1992b) are in parentheses.
c EBV genes are those of the B95-8 strain (Baer et a/., 1984).
conserved with EBV and those which are not. The con- served genes within gammaherpesviruses are clus- tered into five gene blocks (Table 2). The first block contains the well-conserved genes coding for major DNA-binding protein, glycoprotein B homologue, and DNA polymerase; the second block includes genes coding for thymidine kinase, glycoprotein H homo- logue, major capsid protein, the spliced gene, alkaline exonuclease, and uracil-DNA glycosylase; the third block contains three gammaherpesvirus-specific genes (BRRF2, BRRFl, and BRLFl homologues); the fourth block includes genes encoding deoxyuridine tri- phosphatase, ribonucleotide reductase, and the large tegument protein; the gammaherpesvirus-specific gene coding for the membrane antigen constitutes the fifth gene block. Only genes of blocks 1, 2, and 4 are partially conserved with alpha- and betaherpesviruses (Davison and Taylor, 1987; Gompels et a/., 1988; Nich- olas eta/., 1988; Chee eta/., 1990; Albrecht and Fleck- enstein, 1990). These blocks present a similar orienta- tion and localization in the genome of HVS and EBV but the genetic content and the length of regions located between these blocks vary widely; most of the internal repeats are located outside of these blocks.
Localization and orientation of BHV-4 genes in- cluded in these conserved gammaherpesvirus gene blocks were similar to those of HVS and EBV (Fig. 3). The six BHV-4 sequences (1, 8, 9, 29, 32, and 33) which were not found to be homologous to any HVS or EBV genes were located outside of these conserved gene blocks. Genomic regions which were found to vary in size between BHV-4 isolates or strains (Vl , V2, V3, and V4) (Bublot et a/., 1990, 1991 a,b; Thiry et al., 1992) were also located outside of these conserved gene blocks (Fig. 3). The genetic content of these vari- able regions is unknown except for the Vl region which contains tandem repeats (M. Bublot, unpub- lished results).
The length of the BHV-4 genomic regions located between two identified genes of a block was more simi- lar to that of HVS than that of EBV (data not shown). This data allowed the estimation of the length of con- served blocks in the BHV-4 genome. This was done by addition of (1) the length of HVS genome from the left end of a block to the region homologous to the leftmost BHV-4 sequence of the block, (2) the length of BHV-4 genome from the leftmost BHV-4 homologous se- quence to the rightmost one of the block, and (3) the
homologous genes coding for the small subunit of ribonucleotide reductase (conserved in alpha- and gammaherpesviruses), (d) BHV-4 se- quence 14b and EBV BDLF3.5 homologues (conserved in beta- and gammaherpesviruses), and (e) BHV-4 sequence 30 and genes coding for the membrane antigen (HVS 75 and EBV BNRFl) (gammaherpesvirus-specific gene). Identical amino acids are indicated by a vertical bar (I) and similar amino acids are represented by double dots (:). The star (*) indicates an amino acid which is conserved in all the aligned sequences, Gaps introduced into the sequence to maximize overlap are indicated by dashes (-). Numbers in parentheses indicate the start and end positions of the aligned portions of the homologous genes. Potential N-glycosylation sites of gB homologues in (b) are underlined.

660 BUBLOT ET AL.
v3 Vl v2 v4 lR BHVJ c , , , I I I I I I I I I I
-1 1, .> ; i ,>
El
':P,,. >,,I ,,,, ',. 1.3 ,I. ,.,, I :4 I.. ., i: , m
STP-A T6 GCR HSU-RNAS CD59
cyclin
DHFR CCPH
IR4 I33 EBV
I I I I I I I I I I I I I I I I I I
I I I 1 I I I I I 0 10 20 30 40 50 60 70 60 90 100 110 120 130 140 150 160 170 kb
FIG. 3. Comparison of the order and spacing of gene blocks on the mature, linear genome of BHV-4 (V. Test strain), herpesvirus saimiri (HVS, strain No. 11) (Nicholas eta/., 1992b; J. C. Albrecht, J. Nicholas, D. Biller, K. R. Cameron, B. Biesinger, C. Newman, S. Wittmann, M. A. Craxton, H. Coleman, B. Fleckenstein, and R. W. Honess, submitted) and Epstein-Barr virus (EBV, 695-8 strain) (Baer et al., 1984). The 5 gene blocks, which are conserved within gammaherpesviruses, are indicated by large shaded rectangles. Gene content of each block is reported in Table 2. Genes conserved with beta- and alphaherpesviruses are located in blocks 1, 2, and 4; blocks 3 and 5 include only gammaherpesvirus-specific genes. Also shown as horizontal arrows are the positions of the major immediate-early gene of BHV-4 (IEl) (Van Santen, 1991) and HVS (IE-G) (Nicholas era/., 1990). Vertical arrows indicated important genes located outside of gene blocks and which are not conserved between EBV and HVS; most commonly investigated genes are marked by their abbreviations (STP-A, saimiri transformation-associated protein; HSU-RNAs, herpesvirus saimiri U-RNAs; DHFR, dihydrofolate reductase; CCPH, complement control protein homologue; CD 59, cluster designation 59 homologue; TS, thymidylate synthase; GCR, G protein-coupled receptor homologue; LMP, latent membrane protein; EBNA-2, -3A, B, C, -LP, EBV nuclear antigen 2, 3A, 3B, 3C, leader protein; bcl-2, bcl-2 proto-oncogene homologue; IL1 0, interleukin 10 homologue). EBV origins of replication are indicated by triangles termed “orilyt” (origin of replication for the lytic cycle) and “ori,,” (plasmid origin of replication); terminal repeats (TR) and large internal repeats (IR) are represented by open rectangles; small internal repeated sequences are indicated by vertical lines or black rectangles. Vl , V2, V3, and V4 are genomic regions which vary in size between BHV-4 isolates (Thiry et a/., 1992); the Vl region, at least, contains tandem repeats (M. Bublot, unpublished results). The orientation of the EBV genome is inverted relative to the conventional orientation (Baer et a/., 1984).
length of HVS genome from the region homologous to the rightmost BHV-4 sequence to the right end of the block. Lengths of BHV-4 blocks 3 and 5 have been also estimated by analysis of BHV-4 (DN 599 strain) se- quences kindly provided by V. Van Santen and G. Keil. The results are reported in Table 2. Lengths of blocks 1 and 5 were similar for BHV-4, HVS, and EBV; EBV block 2 and especially block 4 were longer than those of BHV-4 and HVS; and HVS block 3 longer than those of BHV-4 and EBV. This bigger size of HVS block 3 is due to the presence of a repeat in gene 48 which causes an expansion of the acidic C-terminal domain of this BRRF2 homologue (J. C. Albrecht, J. Nicholas, D. Biller, K. R. Cameron, B. Biesinger, C. Newman, S. Wittmann, M. A. Craxton, H. Coleman, B. Fleckenstein, and R. W. Honess, submitted) (Fig. 3; Table 2).
The space between, BHV-4 gene blocks was also more similar to that of HVS than that of EBV. It was very obvious for that between blocks 3 and 4 and that be- tween blocks 4 and 5 which are rich in tandem repeats in EBV (Table 2; Fig. 3). The EBV genomic region lo- cated between blocks 2 and 3 contains the BKRFl
gene coding for EBNA-1 (Baer eta/,, 1984); homologue of this gene has not been found in HVS nor in BHV-4 genome (A.-S. Lequarre and M. Bublot, unpublished results). The intervals between ends of the unique por- tion of the genome and the leftmost and the rightmost block vary widely between these three viruses (Ta- ble 2).
The gene arrangement of BHV-4 is therefore also more similar to that of HVS than to that of EBV.
5. Dinucleotide frequencies analysis
Deviations of observed from expected frequencies of dinucleotides in BHV-4 sequences are presented in Fig. 4a. In total, there was an observed sixfold de- crease in CpG dinucleotide frequency. An increase in TpG and CpA dinucleotides was also detected.
The relationship between deviations of observed from expected frequencies of CpG and the sum of de- viations of observed from expected frequencies of TpG and CpA are presented in Fig. 4b. While the correlation between CpG deficit and TpG + CpA excess was rela-

GENE ORGANIZATION OF BHV-4 661
b a o<e o>e e/o
AG - AT- CA -
t cc- ; CG- $ CT- 3 GA-
2 Gc- a- GT- TA - TC- TG,
I o/e
e-I 765432123456
Deviations of observed (0) from expected (e) frequencies of dinucleotides
-6 -7 -6 -5 -4 -3 -2 -1 0 1 2 Deviations of observed from expected frequencies of CpG
FIG. 4. (a) Deviations of observed (0) from expected (e) frequencies of dinucleotides in BHV-4 sequences. Filled bars indicate the magnitude of deviations of observed occurrence of each dinucleotide from occurrence expected in random DNA sequences with the observed mononucleo- tide compositions [for o c e, e/o = fold deficit (left side) and for o > e, o/e = fold excess (right side)]. Arrows indicate the excess of CpA and TpG dinucleotides. (b) Relationships between deviations of observed (0) from expected (e) frequencies of CpG [CpG(%), - CpG(%),] (abscissa) and the sum of deviations of observed from expected frequencies of TpG and CpA { [TpG(%), - TpG(%),] + [CpA(o/,& - CPA(%),]} (ordinate) in the 33 sequences (filled squares). Dots of the line correspond to an exact correlation between CpG deficiency and TpG + CpA excess. The open square is the mean value for the 33 sequences.
tively weak for individual sequences, it was strong on the average (open square in Fig. 4b). This weak corre- lation for individual sequences was probably due to their short length allowing greater variations between expected and observed values. This deficiency in CpG is likely to be the result of methylation of viral DNA (Bird, 1980).
DISCUSSION
This study has shown that the BHV-4 presents the molecular characteristics of the representative mem- bers of the gammaherpesvirinae subfamily. Indeed, the BHV-4 genes content and organization were simi- lar to those of EBV and HVS and the BHV-4 coding sequence was globally deficient in CpG dinucleotide.
The CpG deficiency suggests that BHV-4 latent ge- nome is methylated and that it is present in dividing cells capable of de nova methylation (Honess et al., 1989). The latency site of BHV-4 has not been charac- terized yet; trigeminal ganglia (Homan and Easterday, 1981; Krogman and McAdaragh; 1982; Castrucci et a/., 1987) and spleen (Osorio and Reed, 1983; Osorio eta/., 1982, 1985) have been proposed to harbor latent BHV-4. The CpG deficit in the BHV-4 genome suggests that the neuronal cells of the trigeminal ganglia are not likely to be the main site of latency because these dif- ferentiated nondividing cells are unable to methylate DNA (Honess et a/., 1989). The spleen, especially di- viding cells, could be a potential latency site of BHV-4 as could be cells of the basal layer of an epithelium or
stem cells of the bone marrow. Alternatively, it is also possible that latently infected cells are immortalized by BHV-4 but such immortalization has never been de- scribed in viva or in vitro. The CpG deficiency suggests also that the BHV-4 latent DNA is replicated (Honess et al., 1989); it could persist as an episome like EBV (Kieff et al., 1983) or HVS (Kaschka-Dierich et a/., 1982).
BHV-4 is genetically related to the gammaherpesvi- ruses HVS and EBV. Furthermore, several data indi- cate that relationship of BHV-4 with HVS is stronger than with EBV: (1) BHV-4 (Ehlers et a/., 1985; Bublot et a/., 1990; this study) and HVS (Fleckenstein et al., 1975; Bornkamm eta/., 1976) possess the same geno- mic structure and a similar (G + C) content differing from those of EBV (Baer et al., 1984); (2) BHV-4 amino acid sequences are more similar to those of HVS; (3) the length of conserved gene blocks and the space between these blocks are more similar between BHV-4 and HVS than between BHV-4 (or HVS) and EBV. An- other finding that underlines the different degrees of relatedness between, on the one hand, BHV-4 and HVS and, on the other hand, EBV is the presence of a major immediate-early gene between block 1 and 2 in BHV-4 (Van Santen, 1991) and HVS (Nicholas et al., 1990) genome but not in EBV genome (Baer et a/., 1984). The two genes are transcribed from similar ge- nomic regions and in identical directions, giving the most abundant immediate-early RNA (Fig. 3); these RNAs are spliced but the potential open reading frame (ORF) of HVS is located in one exon (Nicholas et a/., 1990; L. S. Coles and R. W. Honess, unpublished re-

662 BUBLOT ET AL
sults) and that of BHV-4 in four exons (Van Santen, 1991). These two potential ORFs are not homologous at the amino acid sequence level. Whether they play the same role in the biology of infection is not known. All these results suggest that, in the gammaherpesvi- rinse subfamily, HVS and BHV-4 are evolutionarily more closely related to each other than either are to EBV. Consistent with this division of thegammaherpes- virinae subfamily into two genetically distinct sub- groups, is the observation of Albrecht and Flecken- stein (1990) who have shown that EBV (gamma, proto- type) and HVS (gamma, prototype) (Honess, 1984) were more distinct from each other than VZV and HSV- I, the prototypes of alpha, and alpha, subgroups, re- spectively.
The genomic structure of HVS is also found in other herpesviruses: herpesvirus (h.) ateles (Fleckenstein et al., 1978) herpesvirus sylvilagus (Medveczky et al., 1989) murine herpesvirus 68 (MHV 68) (Efstathiou et al., 1990a) and alcelaphinae herpesvirus 1 (Bridgen et al., 1989). The genetic content has been partially stud- ied for h. ateles (Richter et al., 1988) and MHV 68 (Ef- stathiou et a/., 1990b). These two viruses seem to be also more closely related to HVS than to EBV. All these viruses having the same genomic structure are able to grow in fibroblastoid cell lines unlike EBV. Further stud- ies should be done on these viruses to confirm their closer genetic relationships to HVS and, therefore, their belonging to a gammaherpesvirus subgroup evo- lutionarily distinct from EBV.
Six BHV-4 sequences were not found to be homolo- gous to any HVS or EBV genes. These sequences could contain parts of BHV-4 specific genes but they could as well be parts of noncoding regions of the BHV-4 genome or parts of nonconserved domains of conserved proteins. All these sequences were located outside of conserved gene blocks. These noncon- served regions between HVS and EBV genomes con- tain genes coding for proteins playing an important role in the pathobiology of these viruses. EBV genes coding for the latently expressed proteins such as EBNAs and latent membrane proteins (LMP) as well as the BZLFl gene coding for EBV transactivator ZEBRA and BLLFl coding for the major glycoprotein complex (gp 350/ 220) recognizing EBV cellular receptor CD 21 have no counterpart in HVS (Baer et al., 1984; Kieff and Liebo- witz, 1990; Nicholas et al., 199213; J. C. Albrecht, I. Nicholas, D. Biller, K. R. Cameron, B. Biesinger, C. Newman, S. Wittmann, M. A. Craxton, H. Coleman, B. Fleckenstein and R. W. Honess, submitted). Genes in- volved in lymphocyte immortalization by EBV (those coding for EBNA-2 and LMP and BARFl) (Kieff and Lie- bowitz, 1990; Wei and Ooka, 1989) or by HVS (HVS gene 1 coding for the saimiri transformation-asso-
ciated protein) (Murthy et al., 1989) are also not con- served within these gammaherpesviruses. Further- more, genes homologous to cellular genes are located in these nonconserved regions too; these genes are EBV BCRFl (IL-10 homologue; Moore et a/., 1990), EBV BHRFl (bcl-2 homologue; Cleary et al., 1986), HVS gene 2 (dihydrofolate reductase homologue; Trimble et al., 1988) HVS gene 4 (complement control protein homologue; J.-C. Albrecht and B. Fleckenstein, submitted), HVS gene 15 (Human CD59 homologue; J. C. Albrecht, J. Nicholas, K. R. Cameron, C. Newman, B. Fleckenstein, and R. W. Honess, submitted), HVS gene 70 (ECLF4; thymidylate synthase homologue; Bodemer et a/., 1986; Honess et al., 1986), HVS gene 72 (ECLF2; cyclin homologue; Nicholas et al., 1992a), and HVS gene 74 (ECRF3; protein G-coupled receptor; Nicholas et al., 1992a). HVS U-RNAs (Lee et al., 1988; Wassarman et al., 1989; Albrecht and Fleckenstein, 1992) and EBV EBER RNAs (Baer et al., 1984) are lo- cated between a conserved block and one end of the coding part of the genome and are not conserved (Fig. 3). Genes involved in important biological properties such as latency, immortalization, lytic-cycle transacti- vation, and other virus-host interactions are therefore not conserved between EBV and HVS. The available space for such genes in the BHV-4 genome is roughly similar to that of HVS, and therefore it is likely that these BHV-4 nonconserved regions contain genes in- volved in the specific biology of this virus. Further se- quence analysis of these regions will assess if HVS genes having no counterpart in EBV are conserved in BHV-4 genome, if such regions contain ORFs homolo- gous to cellular genes and therefore if there are prefer- ential sites for insertion of cellular genes into such viral genomes.
Gammaherpesviruses are lymphotropic (Honess and Watson, 1977; Roizman, 1982; Honess, 1984). Nevertheless, BHV-4 infection has some biological characteristics common to members of the Betaher- pesvirinae subfamily (Storz et al., 1984) and the lym- photropic viruses MDV, HVT, and HHV-6 have a gene organization different from that of EBV and HVS. These results show that there is no relationship between lym- photropism and the overall gene organization of her- pesviruses and therefore this biological property can- not be used to identify genetically related herpesvi- ruses. Members of the gammaherpesvirinae subfamily are divided, on the basis of biological behavior, into two subgroups: the gamma,-herpesvirinae subgroup (including EBV and related viruses of Old World mon- keys) and the gamma,-herpesvirinae subgroup (includ- ing viruses of the New World monkeys HVS and h. ateles). The gamma,-herpesviruses are typically asso- ciated with B-lymphocytes and the gamma,-herpesvi-

GENE ORGANIZATION OF BHV-4 663
ruses with T-lymphocytes (Honess, 1984). MHV-68 which has a genome more similar to that of HVS (Ef- stathiou eta/., 1990b) seems to persist in B-cells (N. P. Sun+Chandra, S. Efstathiou, and A. A. Nash, unpub- lished results) and BHV-4 in macrophages at least in the rabbit model of infection (Osorio et al., 1985). Therefore, neither T-cell or B-cell lymphotropism seem to be related to the general gene organization of her- pesviruses.
The genomes of EBV, HVS, MHV-68, and BHV-4 have similar gene organization and are all deficient in CpG dinucleotides (Baer et a/., 1984; Honess et a/., 1989; Efstathiou et a/., 1990b; Albrecht and Flecken- stein, 1990). They all possess tandem repeats at both ends (Fleckenstein eta/., 1975; Bornkamm eta/., 1976; Given et a/,, 1979; Ehlers et al., 1985; Efstathiou et a/., 1990a). Additional studies, especially on the molecular control of latency, will be necessary to assess the rela- tionships between these common molecular charac- teristics and the biological properties of gammaher- pesviruses.
ACKNOWLEDGMENTS
We thank Marc Collet and Robert Herzog of the Free University of Brussels (Department of Molecular Biology) for their help in the com- puter analysis. We thank also Vicky van Santen and Gunther Keil for providing us their BHV-4 (DN 599 strain) sequences, Brigitte Bie- singer and Doris Biller their HVS sequences, and Marie-FranCoise Van Bressem its BHV4 (V. Test strain) sequences. The text presents research results of the Belgian National incentive program on funda- mental research in life sciences initiated by the Belgian State, the Prime Minister’s Office Science Policy Program and was in part sup- ported by the Deutsche Forschungsgemeinschaft, Forschergruppe “DNA-Viren des haematopoetischen Systems.” Scientific responsi- bility is assumed by the authors. We also thank Marinette Muys for the patient application of her secretarial skills.
REFERENCES
ALBRECHT, J. C., and FLECKENSTEIN, B. (1990). Structural organization of the conserved gene block of herpesvirus saimiri coding for DNA polymerase, glycoprotein B, and major DNA binding protein. Virol- ogy 174, 533-542.
ALBRECHT, J. C., and FLECKENSTEIN, B. (1992). Nucleotide sequence of HSURG and HSUR7, two small RNAs of herpesvirus saimiri. Nucleic Acids Res. 20 (in press).
BAER, R., BANKIER, A. T., BIGGIN, M. D., DEININGER, P. L., FARRELL, P. J.. GIBSON, T. J., HATFULL, G., HUDSON, G. S., SATCHWELL, S. C., SEGUIN, C., TUFFNELL, P. S., and BARRELL, B. G. (1984). DNA se- quence and expression of the B95-8 Epstein-Barr virus genome. Nature 310, 207-211.
BARAHONA, H. H., MELENDEZ, L. V., KING, N. W., DANIEL, M. D., FRASER, C. E. O., and PREVILLE, A. C. (1973). Herpesvirus aotus type 2: A new viral agent from owl monkeys (Aotus trivirgatus). J. Infect. Dis. 127, 171-178.
BIRD, A. P. (1980). DNA methylation and the frequency of CpG in animal DNA. Nucleic Acids Res. 8, 1499-l 504.
BODEMER, W., NILLER, H. H., NITSCHE, N., SCHOLT~, B., and FLECKEN-
STEIN, B. (1986). Organization of the thymidylate synthase gene of herpesvirus saimiri. J. Viral. 60, 1 14-l 23.
BORNKAMM, G. W., DELIUS, H., FLECKENSTEIN, B., WERNER, F.-J., and MULDER, C. (1976). Structure of herpesvirus saimiri genomes: ar- rangement of heavy and light sequences in the M genome. J. Viral. 19, 154-161.
BRIDGEN, A., HERRING, A. J., INGLIS, N. F., and REID, H. W. (1989). Preliminary characterization of the alcelaphine herpesvirus 1 ge- nome.1. Gen. Viral. 70, 1141-1 150.
BUBLOT, M., VAN BRESSEM, M.-F., THIRY, E., DUBUISSON, J., and PAS- TORET, P.-P. (1990). Bovine herpesvirus 4 genome: Cloning, map- ping and strain variation analysis. /. Gen. Viral. 71, 133-l 42.
BUBLOT, M., WELLEMANS, G., VAN BRESSEM, M.-F., DUBUISSON. J., PAS- TORET, P.-P., and THIRY, E. (1991 a). Genomic diversity among bo- vine herpesvirus 4 field isolates. Arch. Viral. 116, l-l 8.
BUBLOT, M., DUBUISSON, J., VAN BRESSEM, M.-F., DANYI, S., PASTORET, P.-P., and THIRY, E. (1991 b). Antigenic and genomic identity be- tween simian herpesvirus aotus type 2 and bovine herpesvirus type 4. J. Gen. Viral. 72, 715-719.
BUCKMASTER, A. E., Scorr, S. D., SANDERSON, M. J., BOURSNELL, M. E. G., Ross, N. L. J., and BINNS, M. M. (1988). Gene sequence and mapping data from Marek’s disease virus and herpesvirus of tur- keys: Implications for herpesvirus classification. J. Gen. Viroi. 69, 2033-2042.
CAMERON, K. R., STAMMINGER, T., CRAXTON, M., BODEMER. W., Ho- NESS, R. W., and FLECKENSTEIN, B. (1987). The 160,000-n/r, virion protein encoded at the right end of the herpesvirus saimiri genome is homologous to the 140,000-M, membrane antigen encoded at the left end of the Epstein-Barr virus genome. J. Vkol. 61, 2063- 2070.
CASTRUCCI, G., FRIGERI, F., FERRARI, M., PEDINI, B., ALDROVANDI, V., CILLI, V., RAMPICHINI, L., and GATTI, R. (1987). Reactivation in calves of latent infection by Bovid herpesvirus-4. Microbiologica 10,37-45.
CHEE, M. S., BANKIER, A. T., BECK, S., BOHNI, R., BROWN, C. M., CERNY, R., HORSNELL, T., HUTCHISON, Ill C. A., KOUZARIDES, T., MAR- TIGNETTI, J. A., PREDDIE, E., SATCHWELL, S. C., TOMLINSON, P., WES- TON, K. M., and BARRELL, B. G. (1990). Analysis of the protein-cod- ing content of the sequence of human cytomegalovirus strain AD1 69. Curr. Top. Microbial. Immunol. 154, 125-l 69.
CLEARY. M. L., SMITH, S. D., and SKLAR. J. (1986). Cloning and struc- tural analysis of cDNAs for bcl-2 and hybrid bcl-2/immunoglobulin transcript resulting from the t(14; 18) translocation. Cell 47, 19- 28.
DAVISON, A. J., and SCOTT, J. E. (1986). The complete DNA sequence of varicella-zoster virus. /. Gen. Viral. 68, 1759-l 816.
DAVISON, A. J., and TAYLOR, P. (1987). Genetic relations between varicella-zoster virus and Epstein-Barr virus. f. Gen. Viral. 68, 1067-l 079.
DEVEREUX, J., HAEBERLI, P., and SMITHIES, 0. (1984). A comprehen- sive set of sequence analysis programs for the VAX. Nucleic Acids Res. 12, 387-395.
DUBUISSON, J., DANYI, S., BUBLOT, M., PASTORET, P.-P., and THIRY, E. (1991). Comparison of proteins of simian herpesvirus aotus type 2 and bovine herpesvirus type 4. J. Gen. Viral. 72, 1 145-l 150.
EFSTATHIOU, S., Ho, Y. M., and MINSON, A. C. (1990a). Cloning and molecular characterization of the murine herpesvirus 68 genome. J. Gen. Viral. 71, 1355-l 364.
EFSTATHIOU, S., Ho, Y. M., HALL, S., STYLES, C. J., SCOTT, S. D., and GOMPELS, U. A. (1990b). Murine herpesvirus 68 is genetically re- lated to the gammaherpesviruses Epstein-Barr virus and herpes- virus saimiri. /. Gen. Viral. 71, 1365-l 372.
EHLERS, B., BUHK, H.-J., and LUDWIG, H. (1985). Analysis of bovine cytomegalovirus genome structure: Cloning and mapping of the

664 BUBLOT ET AL.
monomeric polyrepetitive DNA unit, and comparison of European and American strains. J. Gen. Viral. 66, 55-68.
FABRICANT, C. G., GILLEPSIE, J. H., and KROOK, L. (1971). Intracellular and extracellular mineral crystal formation induced by viral infec- tion of cell cultures. Infect. Immun. 3, 416-419.
FENG, D.-F., and DOOLITTLE, R. F. (1987). Progressive sequence alignment as a prerequisite to correct phylogenetic trees. J. Mol. Evol. 25, 351-360.
FLECKENSTEIN, B., BORNKAMM, G. W., and LUDWIG, H. (1975). Repeti- tive sequences in complete and defective genomes of herpesvirus saimiri. 1. Viral. 15, 398-406.
FLECKENSTEIN, B., BORNKAMM, G. W., MULDER, C., WERNER, F.-J., DAN- IEL, M. D., FALK, L. A., and DELIUS, H. (1978). Herpesvirus ateles DNA and its homology with herpesvirus saimiri nucleic acid. J. Viral. 25, 361-373.
GIVEN, D., YEE, D., GRIEM, K., and KIEFF, E. (1979). DNA of Epstein- Barr virus. V. Direct repeats of the ends of Epstein-Barr virus DNA. J. Viral. 30, 852-862.
GOMPELS, U. A., CRAXTON, M. A., and HONESS, R. W. (1988). Conser- vation of gene organization in the lymphotropic herpesviruses, herpesvirus saimiri and Epstein-Barr virus. J. Viral. 62, 757-767.
HIGGINS, D. G., and SHARP, P. M. (1989). Fast and sensitive multiple sequence alignments on microcomputer. C.A.5.I.O.S. 5, 151- 153.
HOMAN, E. J., and EASTERDAY, B. C. (1981). Further studies of natu- rally occurring latent bovine herpesvirus infection. Am. J. Vet. Res. 42, 1811-1813.
HONESS, R. W. (1984). Herpes simplex and “the herpes complex”: Diverse observations and a unifying hypothesis. J. Gen. Viral. 65, 2077-2107.
HONESS, R. W., BODEMER, W., CAMERON, K. R., NILLER, H. H., FLECK- ENSTEIN, B., and RANDALL, R. E. (1986). The A + T-rich genome of herpesvirus saimiri contains a highly conserved gene for thymidy- late synthase. Proc. Natl. Acad. Sci. USA 83, 3604-3608.
HONESS, R. W., GOMPELS, U. A., BARRELL. B. G., CRAXTON, M., CAM- ERON, K. R., STADEN, R., CHANG, Y.-N., and HAYWARD, G. S. (1989). Deviations from expected frequencies of CpG dinucleotides in herpesvirus DNAs may be diagnostic of differences in the states of their latent genomes. J. Gen. Viral. 70, 837-855.
HONES?,, R. W., and WATSON, D. H. (1977). Unity and diversity in the herpesviruses. J. Gen. Viral. 37, 15-37.
JOSEPHS, S. F., ABLASHI, D. V., SALAHUDIN, S. Z., JACODZINSKI, L. L., WONG-STAAL, F., and GALLO, R. C. (1991). Identification of the hu- man herpesvirus 6 glycoprotein H and putative large tegument protein genes. J. V/r-o/. 65, 5597-5604.
KASCHKA-DIERICH, C., WERNER, F. J., BAUER, I., and FLECKENSTEIN, B. (1982). Structure of nonintegrated, circularherpesvirussaimiriand herpesvirus ate/es genomes in tumor cell lines and in vitro-trans- formed cells. J. Viral. 44, 295-310.
KIEFF, E., DAMBAUGH, T., HUMMEL, M., and HELLER, M. (1983). Ep- stein-Barr virus transformation and replication. /n “Advances in Viral Oncology” (G. Klein, Ed.), Vol. 3, pp. 133-l 82. Raven Press, New York.
KIEFF, E., and LIEBOWITZ, D. (1990). Epstein-Barrvirus and its replica- tion. /n “Virology” (Fields et a/., Eds.), 2nd ed., Vol. 2, pp. 889- 1958. Raven Press, New York.
KIT, S., KIT, M., ICHIMURA, H., CRANDELL, R., and MCCONNELL, S. (1986). Induction of thymidine kinase activity by viruses with group B DNA genomes: Bovine cytomegalovirus (bovine herpesvirus 4). Virus Res. 4, 197-212.
KOUZARIDES, T., BANKIER, A. T., SATCHWELL, S. C., WESTON, K., TOM- LINSON, P., and BARRELL, B. G. (1987). Large-scale rearrangement of homologous regions in the genomes of HCMV and EBV. Virol- ogy 157, 397-413.
KROGMAN, L. A., and MCADARAGH, J. P. (1982). Recrudescence of bovine herpesvirus- in experimentally infected calves. Am. J. Vet. Res. 43, 336-338.
KRUGER, J. M., OSBORNE, C. A., WHETSTONE, C. A., GOYAL, S. M., and SEMLAK, R. A. (1989). Genetic and serologic analysis of feline cell- associated herpesvirus-induced infection of the urinary tract in conventionally reared cats. Am. J. Vet Res. 50, 2023-2027.
LAWRENCE, G. L., CHEE, M., CRAXTON, M. A., GOMPELS, U. A., HONES& R. W., and BARRELL, B. G. (1990). Human herpesvirus 6 is closely related to human cytomegalovirus. J. Viral. 64, 287-299.
LEE, S. I., MURTHY, S. C. S., TRIMBLE, J. J., DESROSIERS, R. C., and STEITZ, J. A. (1988). Four novel U RNAs are encoded by a herpes- virus. Cell 54, 599-607.
MARTIN, M. E. D., NICHOLAS, J., THOMSON, B. J., NEWMAN, C., and HONES& R. W. (1991). Identification of a transactivating function mapping to the putative immediate-early locus of human herpes- virus 6.1. Viral. 65, 5381-5390.
MCGEOCH, D. J., DALRYMPLE, M. A., DAVISON, A. J., DOLAN, A., FRAME, M. C., MCNAB, D., PERRY, L. J., Scorr, J. E., and TAYLOR, P. (1988). The complete DNA sequence of the long unique region in the genome of herpes simplex virus type 1. J. Gen. Viral. 69, 1531- 1574.
MCGEOCH, D. J. (1989). The genomes of the human herpesviruses: Contents, relationships and evolution. Annu. Rev. Microbial. 43, 235-265.
MEDVECZKY, M., GECK, P., CLARKE, C., BYRNE% J., SULLIVAN, J. L., and MEDVECZKY, P. G. (1989). Arrangement of repetitive sequences in the genome of herpesvirus sylvilagus. 1. Viral. 63, 101 O-l 014.
MOORE, K. W., VIEIRA, P., FIORENTINO, D. F., TROUNSTINE, M. L., KHAN, T. A., and MOSMANN, T. R. (1990). Homology of cytokine synthesis inhibitoty factor (IL-l 0) to the Epstein-Barr virus gene BCRFl Science 248, 1230-l 234.
MURTHY, S. C. S., TRIMBLE, J. J., and DESROSIERS, R. C. (1989). Dele- tion mutants of herpesvirus saimiri define an open reading frame necessary for transformation. J. Viral. 63, 3307-3314.
NEIPEL, F., ELLINGER, K., and FLECKENSTEIN, B. (1991). The unique region of the human herpesvirus 6 genome is essentially collinear with the U, segment of human cytomegalovirus. 1. Gen. Viral. 72, 2293-2297.
NICHOLAS, J., GOMPELS, U. A., CRAXTON, M. A., and HONES% R. W. (1988). Conservation of sequence and function between the prod- uct of the 52-kilodalton immediate-early gene of herpesvirus sai- miri and the BMLFl-encoded transcriptional effector (EB2) of Ep- stein-Barr virus. 1. Viral. 62, 3250-3257.
NICHOLAS, J., SMITH, E. P., COLES, L., and HONES% R. (1990). Gene expression in cells infected with gammaherpesvirus saimiri: Prop- erties of transcripts from two immediate-early genes. Virology 179, 189-200.
NICHOLAS, J., COLES, L. S., NEWMAN, C., and HONES% R. W. (1991). Regulation of the herpesvirus saimiri (HVS) delayed-early 1 1 O-ki- lodalton promoter by HVS immediate-early gene products and a homolog of the Epstein-Barr virus R fransactivator. J. Viral. 65, 2457-2466.
NICHOLAS, J., CAMERON, K. R., and HONES& R. W. (1992a). Herpes- virus saimiri encodes homologues of G protein-coupled receptors and cyclins. Nature 355, 362-365.
NICHOLAS, J., CAMERON, K. R., COLEMAN, H., NEWMAN, C., and Ho- NESS, R. W. (1992b). Analysis of nucleotide sequence of the right- most 43kbp of herpesvirus saimiri (HVS) L-DNA: General conser- vation of genetic organization between HVS and Epstein-Barr virus. Virology 186, 296-310.
OSORIO, F. A., REED, D. E., and ROCK, D. L. (1982). Experimental infection of rabbits with bovine herpesvirus-4: Acute and persis- tent infection. Vet Microbial. 7, 503-513.

GENE ORGANIZATION OF BHV-4 665
OSORIO, F. A., and REED, D. E. (1983). Experimental inoculation of cattle with bovine herpesvirus-4: Evidence for a lymphoid-asso- ciated persistent infection. Am. J. Vet. Res. 44, 975-980.
OSORIO, F. A.. ROCK, D. L., and REED, D. E. (1985). Studies on the pathogenesis of a bovine cytomegalo-like virus in an experimental host. J. Gen. Viral. 66, 1941-l 951.
PEARSON, W. R., and LIPMAN, D. J. (1988). Improved tools for biologi- cal sequence comparisons. Proc. Nat/. Acad. Sci. USA 85, 2444- 2448.
RICHTER, J., PUCHTLER, I., and FLECKENSTEIN, B. (1988). Thymidylate synthase gene for herpesvirus ateles. J. Viral. 62, 3530-3535.
ROIZMAN, B. (1982). The family Herpesviridae: General description, taxonomy and classification. In “The Herpesviruses” (B. Roizman, Ed.), Vol. 1, pp. l-23, Plenum, New York/London.
ROSSITER, P. B., GUMM, I. D., STAGG, D.A., CONRAD, P. A., MUKOLWE, S., DAVIES, F. G., and WHITE, H. (1989). Isolation of bovine herpes- virus-3 from African buffaloes (Syncerus caffer). Res. Vet. Sci. 46, 337-343.
SANGER, F., NICKLEN, S., and COULSON, A. R. (1977). DNA sequenc- ing with chain terminating inhibitors. Proc. Nat/. Acad. Sci. USA 74, 5463-5468.
STORZ. J., EHLERS, B., TODD, W. J., and LUDWIG, H. (1984). Bovine cytomegaloviruses: Identification and differential properties. J. Gen. Viral. 65, 697-706.
THIRY, E., DUBUISSON, J., BUBLOT, M., VAN BRESSEM, M.-F., and PAS- TORET, P.-P. (1990). The biology of bovine herpesvirus 4 infection of cattle. Dtsch. Tier%ztl. Wochenschr. 97, 72-77.
THIRY, E., BUBLOT, M., DUBUISSON, J., VAN BRESSEM, M.-F., LEQUARRE, A.-S., LOMONTE, P., VANDERPLASSCHEN, A., and PASTORET, P.-P. (1992). Molecular biology of bovine herpesvirus type 4. Adv. Vet. Viral., in press.
TODD, W. J., and STORZ, J. (1983). Morphogenesis of a cytomegalovi- rus from an American bison affected with malignant catarrhal fever. J. Gen. Viral. 64, 1025-l 030.
TRIMBLE, J. J., MURTHY, S. C. S., BAKKER, A., GRASSMANN, R., and DESROSIERS, R. C. (1988). A gene for dihydrofolate reductase in a herpesvirus. Science 239, 1 145-l 147.
VAN OPDENBOSCH, E., WELLEMANS, G., and OUDEWATER, J. (1986). Toevallige isolatie van het boviene herpesvirus 4 uit de long van een schaap. Vlaams Diergeneesk. Tijdschr. 55, 432-433.
VAN SANTEN, V. L. (1991). Characterization of bovine herpesvirus 4 major immediate-early transcript. 1. Viral. 65, 521 1-5224.
WASSARMAN, D. A., LEE, S. I., and STEITZ, J. A. (1989). The nucleotide sequence of HSUR 5 RNA from herpesvirus saimiri. Nucleic Acids Res. 17, 1258.
WEI, X. M., and OOKA, T. (1989). A transforming function of the BARFl gene encoded by Epstein-Barr virus. EMBO 1. 8, 2897- 2903.





























