Embed Size (px)
Citation preview

I
ASa
b
c
d
a
ARRAA
KBVbm
1
deci1i(dse
sT
(
h0
Virus Research 197 (2015) 116–126
Contents lists available at ScienceDirect
Virus Research
j ourna l ho me pa g e: www.elsev ier .com/ locate /v i rusres
nteraction of VP8 with mRNAs of bovine herpesvirus-1
zharul Islama, Sandra Schulza, Sharmin Afroza,b, Lorne A. Babiukd,ylvia van Drunen Littel-van den Hurka,b,c,∗
VIDO-Intervac, University of Saskatchewan, 120 Veterinary Road, Saskatoon, SK S7N 5E3, CanadaVaccinology and Immunotherapeutics, University of Saskatchewan, 120 Veterinary Road, Saskatoon, SK S7N 5E3, CanadaMicrobiology and Immunology, University of Saskatchewan, 120 Veterinary Road, Saskatoon, SK S7N 5E3, CanadaUniversity of Alberta, 3-7 University Hall, Edmonton, AB T6G 2J9, Canada
r t i c l e i n f o
rticle history:eceived 6 September 2014eceived in revised form 9 December 2014ccepted 10 December 2014vailable online 18 December 2014
eywords:oHV1P8ICP0RNA
a b s t r a c t
The UL47 gene product, VP8, is one of the most abundant tegument proteins of bovine herpesvirus-1(BoHV-1). Deletion of VP8 leads to impaired growth in tissue culture, and VP8 is indispensable for BHV-1replication in cattle. To elucidate the biological functions of VP8, we explored its interaction with mRNAsof immediate early (bICP0), early (gB, gD) and late (gC) genes of BoHV-1. FLAG-tagged VP8 was pulleddown from COS-7 cells co-transfected with plasmids encoding VP8 and either gB, gC, gD or bICP0. Thiswas followed by RNA extraction, cDNA synthesis and qPCR, which demonstrated binding of VP8 to bICP0,gB, gC and gD mRNAs in the cytoplasm and nucleus. These results were supported by co-localizationof VP8 with bICP0, gB, gC and gD mRNAs in the nucleus as determined by confocal microscopy. Aminoacids 259–342, located in the conserved portion of UL47 homologues, were found to contain the RNAbinding region on VP8. To further characterize these interactions, Northwestern blotting was performedby immobilizing VP8 on a nitrocellulose membrane followed by hybridization with in vitro transcribed
bICP0 mRNA. The results demonstrated binding of VP8 to intron-less mRNA but not intron-containingmRNA of bICP0. In addition, the interaction of VP8 with bICP0 mRNA was confirmed in vitro by RNAelectrophoretic mobility shift assay, which also showed that the zinc finger and acidic domains bothinteract with VP8. Based on these results, we concluded that VP8 binds to intron-less mRNAs of bICP0,gB, gC and gD.© 2014 Elsevier B.V. All rights reserved.
. Introduction
Bovine herpesvirus-1 (BoHV-1) is a member of the herpesviri-ae family (Jones and Chowdhury, 2007; Roizman et al., 1981; Tikoot al., 1995). It is the causative agent of a number of diseases inattle, which cost billions of dollars to the North-American cattlendustry (Jones and Chowdhury, 2007; Tikoo et al., 1995; Yates,982). The BoHV-1 genome is a linear dsDNA molecule of approx-
mately 140 kb with 73 recognized open reading frames (ORFs)Robinson et al., 2008). BoHV-1 genes can be categorized as imme-
iate early (alpha), early (beta) and late (gamma) based on theirynthesis in the infected cells (Jones and Chowdhury, 2007; Tikoot al., 1995). The BoHV-1 virion is composed of the dsDNA genome∗ Corresponding author at: Vaccine and Infectious Disease Organization, Univer-ity of Saskatchewan, 120 Veterinary Road, Saskatoon, SK S7N 5E3, Canada.el.: +1 306 966 1559; fax: +1 306 966 7478.
E-mail address: [email protected]. van Drunen Littel-van den Hurk).
ttp://dx.doi.org/10.1016/j.virusres.2014.12.017168-1702/© 2014 Elsevier B.V. All rights reserved.
surrounded by a capsid, a tegument and an envelope. The tegumentis the most complex compartment of the virion and it is composedof a complex of about 20 virus-encoded proteins, the structures andinteractions of which are not well understood (Mettenleiter et al.,2009). The tegument proteins are among the first proteins to bereleased and interact with the intracellular environment during theinfection process. In addition to their structural role, herpesvirustegument proteins can perform various regulatory functions, suchas transcription modulation (Misra et al., 1995; Triezenberg et al.,1988), kinase activity (Purves and Roizman, 1992), RNase activity(Read et al., 1993) and DNA packaging (Salmon et al., 1998). Thesefunctions are assigned to certain tegument proteins suggesting thatthese virion constituents may function at different stages of viralinfection to establish conditions for efficient viral replication andto promote virus assembly and egress.
VP8, a 97 kDa UL47 gene product, is one of the most abun-
dant tegument proteins of BoHV-1 virions and virus-infected cells(Carpenter and Misra, 1991). Although the UL47 gene is non-essential for viability of BoHV-1 in vitro (Robinson et al., 2008),a UL47 deletion mutant of BoHV1 shows impaired growth in cell
searc
ctLhaeiArVdhLsapnmtH1evaop
ptigtgpt1gtltatVcbdt
2
2
kiH(MH1g1L3(
A. Islam et al. / Virus Re
ulture and is avirulent in cattle (Lobanov et al., 2010) However,he exact role of VP8 during BoHV-1 infection still remains unclear.ike VP13/14 (Meredith et al., 1991), its structural homologue inuman herpesvirus-1 (HHV-1), VP8 is phosphorylated (Carpenternd Misra, 1991) and glycosylated (van Drunen Littel-van den Hurkt al., 1995). Furthermore, VP8 can induce humoral and cellularmmune responses (van Drunen Littel-van den Hurk et al., 1995).t least two kinases [viral US3 and cellular casein kinase 2 (CK2)] areesponsible for the phosphorylation of VP8 (Labiuk et al., 2009). LikeP13/14 (Donnelly and Elliott, 2001a,b), VP8 is found in the nucleusuring the early stages of infection, and the de novo synthesized VP8as been detected in the cytoplasm of infected cells (van Drunenittel-van den Hurk et al., 1995). Both nuclear import and exportignals exist in VP8, which allow VP8 to shuttle between nucleusnd cytoplasm during the course of infection. However, overex-ression of VP8 outside the context of infection leads to primarilyuclear localization in mammalian cells (Zheng et al., 2004). Actino-ycin D inhibits the nucleo-cytoplasmic shuttling of VP8 indicating
hat VP8 may be involved in RNA transport (Verhagen et al., 2006).owever, when expressed in Escherichia coli or insect cells, HHV-
VP13/14, but not BHV-1 VP8, bound directly to RNA (Donnellyt al., 2007). Recently, it was reported that during HHV-1 infection,irion host shutoff (VHS)-RNase interacts with pUL47 (VP13/14),nd that pUL47 plays a role in the attenuation of the degradationf viral and stable host mRNAs through interaction with the VHS inolyribosomes (Shu et al., 2013).
To test the hypothesis that the nucleocytoplasmic shuttlingrotein VP8 binds RNA, thus potentially influencing viral mRNAransport from the nucleus to the cytoplasm, we investigated thenteraction of VP8 with BoHV-1 immediate early (bICP0), early (gB,D) and late (gC) gene transcripts. The bICP0 protein is a majorranscriptional regulatory protein (Robinson et al., 2008), whilelycoproteins gB, gC and gD play a key role in viral attachment,enetration and cell-to-cell spread of BoHV-1 and can induce pro-ective immune responses (van Drunen Littel-van den Hurk et al.,990, 1994). We demonstrated interactions of VP8 with bICP0, gB,C and gD mRNAs by RNA-immunoprecipitation and qPCR, both inhe nucleus and cytoplasm of COS-7 cells. Amino acids 259–342,ocated in the conserved portion of UL47 homologues, were foundo contain the RNA binding region on VP8. Co-localization of VP8nd mRNAs was visualized by confocal microscopic imaging fur-her supporting their nuclear interaction. A direct interaction ofP8 expressed in mammalian cells with intron-less, but not intron-ontaining, bICP0 mRNA was confirmed in vitro by North-Westernlotting, which also showed that both the zinc finger and the acidicomains of bICP0 interact with VP8. These results support the con-ention that BoHV-1 VP8 binds intron-less viral mRNAs.
. Materials and methods
.1. Cells and viruses
COS-7 (CRL-1651), Hela (CRM-CCL-2) and Madin Darby bovineidney (MDBK) (CCL-22) cells were purchased from the Amer-can Type Culture Collection (Manassas, VA, USA). COS-7 andela cells were grown in Eagle’s minimum essential medium
MEM, Sigma–Aldrich Canada Ltd., Oakville, ON, Canada) andDBK cells in Dulbecco’s (D)MEM, both supplemented with 10 mMEPES buffer (GIBCO/Life Technologies, Burlington, ON, Canada),× non-essential amino acids (GIBCO/Life Technologies), 50 �g/mlentamicin (Hyclone Laboratories Inc, Burlington, ON, Canada) and
0% (vol/vol) gamma-irradiated fetal bovine serum (FBS, SeraCareife Sciences Inc., Oceanside, CA, USA). Cells were cultured in a7 ◦C incubator with constant supply of 5% CO2. BoHV-1 strain 108Jericho and Darcel, 1978) was propagated in MDBK cells.h 197 (2015) 116–126 117
2.2. Antibodies
Fibrillarin (H-140)-specific rabbit polyclonal antibody was pur-chased from Santa Cruz Biotechnology Inc. (Dallas, TX, USA).Tubulin (T6199)-specific and HA-specific monoclonal antibodiesand Anti-FLAG affinity gel were purchased from Sigma–Aldrich.VP8- and gB-specific monoclonal antibodies and gC-, gD- andbICP0-specific polyclonal antibodies were raised as described pre-viously (Labiuk et al., 2009; van Drunen Littel-van den Hurk et al.,1984).
2.3. Plasmid construction
Based on the Gene Bank sequence for BoHV-1 (Gene Bank acces-sion no. NC 001847), the gene encoding bICP0 (NP 045368) wassynthesized by Integrated DNA Technology (IDT, Coralville, IA,USA). During the synthesis of the bICP0 gene, an EcoRI site wasinserted at the 5′end and a XhoI site was inserted at the 3′end forcloning purposes. The synthesized bICP0 was first cloned in a yeastepisomal plasmid. Subsequently, the product was digested withEcoRI and XhoI and cloned into pCDNA3 (Invitrogen), cut with EcoRIand XhoI, to create pCDNA3-bICP0. For cloning of bICP0 with intron,a 10 kb fragment (100475–109190 bp) from the sequence of BoHV-1 (NCBI genome data base) was isolated after digesting the BoHV-1genomic DNA with EcoRV and XbaI. This 10 kb fragment containingthe bICP0 gene with intron was cloned into pCDNA3 cut with EcoRVand XbaI. pCDNA3-actinG1-HA-ds red plasmid (a kind gift from Dr.Suresh Tikoo) was used as a template to amplify the actin G1. AnEcoRI site was inserted at the 5′end of the forward primer (Forward-5′-ACTGACGAATTCATGGAAGAAGAAATTGCCGCCC-3′) and XhoI wasinserted at the 3′end of the reverse primer (Reverse-5′-ACTGACCTCGAGTTAGAAGCATTTGCGGTG GAC-3′). The PCR productwas digested with EcoRI and XhoI, gel purified and clonedinto pCDNA3 digested with the same enzymes. The pFLAG-VP8,pEYFP-N1-VP8, pSLIA-gB, pSLIA-gC and pSLIA-gD constructs weredescribed previously (Labiuk et al., 2009; Lobanov et al., 2010;Zheng et al., 2004). The truncations of VP8 were generated asdescribed by Afroz et al. (manuscript in preparation). Briefly, fivedifferent N-terminally truncated VP8 genes, D1 (aa 121–742), D2(aa 219–742), D3 (aa 343–742), D4 (aa 538–742) and D5 (aa632–742), as well as four C-terminally truncated VP8 genes, DA(aa 1–120), DB (aa 1–258), DC (aa 1–482) and DD (aa 1–631), werecloned with a N-terminal FLAG tag into the pCMV4.1 K vector usingHindIII at the 5′end and XhoI at the 3′end.
2.4. Expression and purification of FLAG-VP8 and FLAG-VP8truncations
COS-7 cells were transfected with pFLAG-VP8 or pFLAG-VP8truncations. At 48 h post-transfection, the cells were washed withPBS, lysis buffer (50 mM Tris–HCl, 150 mM NaCl, 1 mM EDTA, 1%Triton X-100 and 10 �l/ml protease inhibitor, adjusted to pH 7.4)was added, and the cells were incubated at 4 ◦C for 1 h. Untrans-fected COS-7 cells were used as control. The cells were collectedinto an Eppendorf tube and spun at 12,000 × g for 15 min, and thesupernatant was collected into a fresh Eppendorf tube. The super-natant was pre-cleared with 50% prewashed Protein A-Sepharosebeads and then incubated with prewashed Anti-FLAG M2 Affin-ity Gel (Sigma–Aldrich) overnight (ON) at 4 ◦C. Subsequently, thebeads were washed three times with wash buffer (50 mM Tris–HCl,
150 mM NaCl, pH 7.4), and FLAG-VP8 was released by competitivebinding for 1 h with the 3x FLAG peptide. The FLAG peptide wasremoved by dialysis and the FLAG-VP8 protein was concentratedwith a protein concentrator (Millipore, Billerica, MA, USA).
1 esearch 197 (2015) 116–126
2e
CctpAa1opaL2tne1i12Tb
2
pawfbwNoLaCt
2
lep2ipSotzteTlaetqmrp
Table 1List of primers used for real-time PCR.
Genes Primers Sequences
gB Forward 5′-ACATCGCGCCGTACACGTTC-3′
gB Reverse 5′-AGGCAGCGCCACTTCTTGTC-3′
gC Forward 5′-ACTGGTTCCGCAACGGCTAC-3′
gC Reverse 5′-CGCTCCATGTTAGCGCTCTG-3′
gD Forward 5′-CGAGCCCAGGAAGCACTTTG-3′
gD Reverse 5′-ACCGTGCCGTCGATGTACAG-3′
bICP0 Forward 5′-CGTTTGTGCGCAGCCTGTTG-3′
bICP0 Reverse 5′-GACGACGACTCTTCTGACTC-3′
DWVP Forward 5′-CAGTAGCTTGGGCGATTGTT-3′
18 A. Islam et al. / Virus R
.5. Preparation of total cell lysate and cytoplasmic and nuclearxtract
For RNA immunoprecipitation and protein expression studies,OS-7 cells were transfected with pFLAG-VP8 or pFLAG-VP8 trun-ations. For the in vivo interaction study, pFLAG-VP8 or pFLAG-VP8runcations were co-transfected together with pSLIA-gB, pSLIA-gC,SLIA-gD, pCDNA3-bICP0 or pCDNA3-actin G1-HA into COS-7 cells.t 48 h post-transfection, the cells were washed in ice-cold PBSnd UV-irradiated (900 mJ/cm2) with a UV cross linker (FB-UVXL-000, Fisher Scientific, Ottawa, ON, Canada). For the preparationf total cell lysate, cells were collected in lysis buffer (50 mM Tris,H 7.6, 150 mM NaCl, 2.5 mM MgCl2, 0.1% NP40, 2.5 �l superasend mammalian protease inhibitors) and left on ice for 30 min.ysed cells were centrifuged at 12,000 rpm for 15 min (rotor F45-4-11, Centrifuge 5415, Eppendorf, Mississauga, ON, Canada), andhe supernatant was collected and kept on ice. Cytoplasmic anduclear extracts were prepared as described elsewhere (Abmayrt al., 2006) with cytoplasmic extract buffer (30 mM HEPES, pH 7.9,40 mM KCl, 3 mM MgCl2, 2.5 �l superase and mammalian protease
nhibitors) and high-salt buffer (20 mM HEPES, pH 7.9, 25% glycerol,.5 mM MgCl2, 1.6 M KCl, 0.2 mM EDTA, 0.2 mM PMSF, 0.5 mM DTT,.5 �l superase and mammalian protease inhibitors), respectively.he purity of the nuclear and cytoplasmic extracts was confirmedy immunoblotting with anti-fibrillarin and anti-tubulin antibody.
.6. Western blotting
Total, cytoplasmic and nuclear cell lysates were loaded on 10%olyacrylamide gels and the proteins were separated by SDS-PAGEnd then transferred to nitrocellulose membranes. The membranesere blocked with 5% skim milk for 1 h at room temperature (RT)
ollowed by incubation with appropriately diluted primary anti-odies for 4 h at RT or ON at 4 ◦C. After washing the membraneith PBST (3.2 mM Na2HPO4, 0.5 mM KH2PO4, 1.3 mM KCl, 135 mMaCl, 0.1% Tween 20, pH 7.4), IRDye 680-conjugated anti-mouse IgGr IRDye 800CW-conjugated anti-rabbit IgG (LI-COR Biosciences,incoln, NE, USA) was applied for 1 h. The membrane was washednd analyzed with the Odyssey® CLx Infrared Imaging System (LI-OR Biosciences, Lincoln, NB, USA). Images were processed withhe Odyssey 3.0.16 application software.
.7. RNA immunoprecipitation, cDNA synthesis and qPCR
RNA immunoprecipitation was carried out with COS-7 cellysate transfected with pFLAG-VP8 or pFLAG-VP8 truncationsither alone or in combination with pSLIA-gB, pSLIA-gC, pSLIA-gD,CDNA3-bICP or pCDNA3-actin G1-HA according to (Donnelly et al.,007). One ml of nuclear, cytoplasmic and total cell extract was
ndividually pre-cleared by incubation with 100 �l of a 50% sus-ension of prewashed Protein A-Sepharose beads for 1 h at 4 ◦C.ubsequently, 500 �l prewashed anti-FLAG affinity gel was addedvernight at 4 ◦C. The antibody-protein complex was washed threeimes, and RNA was extracted from the complex with 1 ml Tri-ol reagent (Ambion, Burlington, ON, Canada). Extracted RNA wasreated with Turbo DNase (1 �g/�l) for 15 min at 37 ◦C followed bythanol precipitation and resuspension in 25 �l RNase-free water.he purity of the RNA was determined by Agilent 2100 Bioana-yzer (Santa Clara, USA), and the concentration was measured with
Nanodrop (ND-1000, Wilmington, DE, USA). Five hundred ngxtracted RNA was used to make cDNA with a QuantiTect reverseranscription kit (Qiagen) according to the manufacturer’s protocol.
PCR was carried out using Platinum Sybr green qPCR super-ix UDG (Invitrogen). PCR was run on an iCyclerIQ-multicoloreal time PCR detection system (BioRad, Richmond, CA, USA). TheCDNA3-actinG1-HA construct was used as negative control for
DWVP Reverse 5′-AGCTTCTGGAACGGCAGATA-3′
in vivo interaction studies and the deformed wing virus polypro-tein (DWVP) gene was used as positive control for the qPCR. TheqPCR primers are shown in Table 1. All PCR products were run ina 2% agarose gel, stained with gel red nucleic acid (Biotium, Hay-ward, CA, USA), and recorded with a Molecular Imager® Gel DocTM
(BioRad).
2.8. Preparation of probes for Northwestern blotting andRNA-electrophoretic mobility shift assay (EMSA)
Radiolabeled probes for Northwestern blotting and RNA EMSAwere made by in vitro transcription using the mMESSAGE mMA-CHINE kit (Ambion) and the Megashort Script T7 kit (Ambion),respectively. Radioactivity was measured with a liquid scintilla-tion counter (Beckman Coulter, Indianapolis, IN, USA). To makeNorthwestern blotting probes, pCDNA3-bICP0 (without intron) waslinearized by digesting with XhoI and pCDNA3-bICP0 (with intron)was linearized by digesting with XbaI before in vitro transcription.Probes for RNA-EMSA were made by inserting the T7-promotersequence at the upstream sequence of the zinc finger and acidicdomains and annealed with the opposite strand of the T7 promoterbefore in vitro transcription. The sequences for in vitro transcriptionof the zinc finger and the acidic domain are as follows:
Zinc ring finger domain: 3′-TTAAATTATGCTGAGTGATATCCAG-CACG ACGTAGACGG ACCTGCGCTAGTGGCCGCGCCGCGCGCGAGA-CGGGACGGACGTGCGGAAGACGGACCGGACGTAGGCGGCCACCGA-CCTCCCCGCGGGCTGGACGGGCGACACGTTCCGC-5′ and 5′-AATT-TAATACGACTCACTATAGG-3′
Acidic domain: 3′-TTAAATTATGCTGAGTGATATCCCTCCTGCTG-AGAACCAGCCTCA GTCTTCTCAGCAGCAGCCCCGAGTCGTGGAGG-CTGGACTGGCGGTAACTGCTGTGCCTTTGCCTCGGCCTCTGCCTGCG-CCTCCAACTCAGCTCCGCGTGCGCCCCCCGCAGACCGCGCCGGGCCC-GAGCCGCC and 5′-AATTTAATACGACTCACTATAGG-3′
2.9. Northwestern blotting
Northwestern blotting was performed as described elsewhere(Donnelly et al., 2007). Purified FLAG-VP8 protein was transferredto a nitrocellulose membrane after separation by SDS-PAGE in a10% acrylamide gel. The protein was renatured by soaking themembrane overnight at 4 ◦C in renaturation buffer (10 mM Tris(pH 7.6), 50 mM NaCl, 1 mM EDTA, 0.1% NP40, 0.02% Ficoll, 0.02%polyvinylpyrrolidone, and 0.02% bovine serum albumin). Radio-labeled bICP0 probes (1000,000 cpm; both intron-containing andintron-less) were applied to the membrane for hybridization withVP8 after blocking the membrane with yeast tRNA (0.05 mg/ml).The hybridized membrane was washed three times with renatu-
ration buffer (without tRNA) and the image was transferred ontoa phosphoscreen overnight in a cassette, and developed with aPersonal Molecular Imager FX (BioRad).
A. Islam et al. / Virus Research 197 (2015) 116–126 119
Fig. 1. Expression of FLAG-VP8, gB, gC, gD and bICP0 proteins in COS-7 cells. (a) Expression plasmids (pFLAG-VP8, pSLIA-gB, pSLIA-gC, pSLIA-gD and pCDNA3-bICP0) weret protew nd 50t in wa
2
la5KwgsT(
2
Hw11ipp
CCAC
ransfected into COS-7 cells. Cell lysates were prepared 48 h post-transfection andere transfected with pFLAG-VP8. Cell lysate was prepared 48 h post-transfection, a
he complex by competitive binding with the 3xFLAG peptide. The eluted VP8 prote
.10. RNA electrophoretic mobility shift assay (EMSA)
RNA EMSA was carried out following a modification of a pub-ished protocol (Yakhnin et al., 2012). Radiolabeled zinc finger andcidic domain probes (10,000 cpm) were allowed to bind with00 ng VP8 in binding buffer (100 mM HEPES, 200 mM KCl, 10 mMCl and 10 mM DTT, pH 7.4) at RT for 30 min. The reaction mixtureas run in a 4% acrylamide native gel for 4–6 h at 150 V, and the
el was dried on Whatman paper. The dried gel was kept in a cas-ette with an upside down phosphoscreen to transfer the image.he image was developed with a Personal Molecular Imager FXTM
BioRad).
.11. Confocal microscopy
Confocal microscopy was performed as described previously.ela cells were plated at a density of 1 × 105 cells per well in two-ell chamber slides. One �g pGFP-N1-VP8 was transfected with
�g pSLIA-gB, pSLIA-gC, pSLIA-gD or pCDNA3-bICP0 along with �g corresponding DNA-probe. As a negative control, cells were
ndividually transfected into Hela cells with 1 �g of pGFP-N1, DNArobes, pGFP-N1-VP8 or a combination of pGFP-N1-VP8 and DNArobe. The sequences of the DNA probes (oligos) are as follows:
gB:5cy3/AGCGCGGCGCGCACGTCTGTGCTGTTGTCGGGGCTGG-
GGCGTCGTCGCCGTCGGGG*C*C*G-3, gC:5cy3/AGGGGTTGCGCA-GGCCTCGTCGTCGCCGCGCGTGGAGCGGAACCTGCCCACGCGCT*G**A-3, gD:5cy3/AAGCTGTCCCAAAACGGGGGTGTGCGGTAGCGG-AGTACCCAAAGTGCTTCCTGGGC*T*C*G-3 and bICP0:5cy3/in expression was determined by SDS-PAGE and Western blotting. (b) COS-7 cells0 �l prewashed anti-FLAG affinity gel was added to bind VP8. VP8 was eluted froms analyzed by SDS-PAGE and staining with CBB.
CTCCGACCAAGAGTCGTCCTCCGAGCCCTCCGAGTCCGAGTCCGAGG-TGTCGAGAAA*C*A*C-3.
The probes were labeled with Cy3 at the 5′end and phospho-rothioates were inserted at the 3′end, indicated by stars. At 48 hpost transfection, cells were fixed with 3.9% paraformaldehyde andwashed three times with PBS. Cells were stained with DAPI anddried for 24 h at RT, and the image was developed using a LeicaMicrosystems (Leica SP5) confocal microscope. The microscope wasset up for DAPI (461 nm), Cy3 (633 nm) and EYFP (488 nm), andimages were processed by ImageJ (Collins, 2007).
3. Results
3.1. VP8 interacts with mRNAs of immediate early (bICP0), early(gB, gD) and late (gC) genes in the cytoplasm and nucleus ofCOS-7 cells
As plasmids pFLAG-VP8, pSLIA-gB, pSLIA-gC, pSLIA-gD andpCDNA3-bICP0 were used to study the in vivo interactions of VP8with mRNAs of gB, gC, gD and bICP0, expression of the respec-tive proteins in COS-7 cells was assessed by Western blotting. VP8,gB, gC, gD and bICP0 were expressed with the expected apparentmolecular weights indicating that the constructs are suitable forin vivo interaction studies with VP8 (Fig. 1a). To examine the in vivo
interactions of VP8 with mRNAs of BoHV-1, VP8 was expressedwith a FLAG-tag and purified on anti-FLAG resin. The purity ofVP8 was confirmed by coomassie brilliant blue (CBB) staining(Fig. 1b).
120 A. Islam et al. / Virus Research 197 (2015) 116–126
Fig. 2. VP8 interacts with mRNAs of bICP0, gB, gC and gD in the cytoplasm and nucleus of COS-7 cells. COS-7 cells were co-transfected with pFLAG-VP8 together with pCDN3-bICP0, pSLIA-gB, pSLIA-gC, pSLIA-gD or pCNDA3-actinG1-HA (negative control; −). The DWVP gene was used as positive control for the qPCR (+). At 48 h post transfection,nuclear, cytoplasmic and total cell extracts were prepared. Anti-FLAG affinity gel was applied to pull down the mRNAs attached to VP8, the mRNAs were extracted andtreated with Turbo DNase, and cDNAs were synthesized. The bICP0, gB, gC and gD genes were amplified by real-time PCR using synthesized cDNAs as template. (a) CT valuesof PCR products; (b) Real-time PCR products run in a 2% agarose gel, stained with gel red and recorded with the Molecular Imager-GelDoc XR+ imaging system. The purityof cytoplasmic and nuclear extracts was confirmed by Western blotting with anti-Tubulin (c) and anti-Fibrillarin (d) antibodies. Two cytoplasmic and one nuclear extractsw d antI
p1egppHipawt
ere run in two separate 10% polyacrylamide gels and probed with anti-tubulin anmaging System.
Being a nucleocytoplasmic shuttling protein, VP8 may have theotential to bind and export RNA from the nucleus (Fischer et al.,994; Phelan et al., 1996; Sandri-Goldin, 1998). To test this hypoth-sis, we investigated the interaction of VP8 with mRNAs of gB,C, gD and bICP0 in total, nuclear and cytoplasmic cell extractsrepared after co-transfection of COS-7 cells with pFLAG-VP8 andSLIA-gB, pSLIA-gC, pSLIA-gD or pCDN3-bICP0. pCNDA3-actinG1-A was used as negative control in this experiment. To determine
f mRNAs of gB, gC, gD and bICP0 were bound to VP8, 500 �l of
rewashed anti-FLAG antibody attached to the affinity bead waspplied to pull down VP8 such that maximum amounts of mRNAsould be obtained. One �l cDNA was used as template to amplifyhe gB, gC, gD and bICP0 mRNAs using the primers shown in Table 1.
i-fibrillarin antibody, and images were developed with the Odyssey® CLx Infrared
The DWVP gene was used as positive control for qPCR. A perfectmelting curve was observed for gB, gC, gD and bICP0 (not shown).In real-time PCR, we noticed that the melting temperature wasrelatively high (over 90 ◦C) for all these genes, which was drivenby their high GC content (over 65%). The CT values of the real-time PCR products (Fig. 2a) clearly indicate binding of gB, gC, gDand bICP0 mRNAs to VP8. The PCR products were also run in a 2%agarose gel and stained with gel red as shown in Fig. 2b, whichconfirmed binding of VP8 to the gB, gC, gD and bICP0 mRNAs in
both nuclear and cytoplasmic fractions. The size of the amplifiedfragments of the gB, gC, gD and bICP0 genes was 150–200 bp asexpected. To confirm the purity of the cytoplasmic and nuclearextracts, they were probed by anti-tubulin and anti-fibrillarin
A. Islam et al. / Virus Research 197 (2015) 116–126 121
Fig. 3. Amino acids 259–342 interact with bICP0, gB, gC and gD mRNA. (a) Expression of FLAG-tagged VP8 truncations D1 (aa 121–742), D2 (aa 219–742), D3 (aa 343–742),D4 (aa 538–742), DA (aa 1–120), DB (aa 1–258), DC (aa 1–482) and DD (aa 1–631) was confirmed by Western blotting with anti-FLAG antibody. (b–e) CT values of real-timebICP0, gB, gC and gD PCR products bound to VP8 truncations. COS-7 cells were co-transfected with pFLAG-VP8 truncations together with pSLIA-gB, pSLIA-gC, pSLIA-gD orpCDNA3-bICP0. At 48 h post transfection total cell extracts were prepared. Anti-FLAG affinity gel was applied to pull down the mRNAs attached to the VP8 truncations, them ed. Thc
antVlc
RNAs were extracted and treated with Turbo DNase, and cDNAs were synthesizDNAs as template.
ntibody. Tubulin was detected in the cytoplasm but not in theucleus, while fibrillarin was detected in the nucleus but not inhe cytoplasm (Fig. 2c and d). This result clearly indicates that
P8 binds mRNAs of immediate early (bICP0), early (gB, gD) andate (gC) genes in the cytoplasm and nucleus of transfected COS-7ells.
e bICP0, gB, gC and gD genes were amplified by real-time PCR using synthesized
3.2. Amino acids 259–342 of VP8 bind to bICP0, gB, gC and gDmRNA
To further define the RNA-interacting domain on VP8, a seriesof N-terminal and C-terminal truncations of FLAG-VP8 were gener-ated. The truncated versions of VP8 were expressed in COS-7 cells

122 A. Islam et al. / Virus Researc
Fig. 4. VP8 interacts with bICP0 mRNA in vitro. (a) Intron-containing (i) and intron-less (ii) radiolabeled bICP0 probes were synthesized by in vitro transcription; (b)4 �g BVDV E2, and 1 �g, 2 �g or 4 �g of VP8-FLAG (lanes 1, 2, 3 and 4) were sub-jected to SDS-PAGE in two separate 10% polyacrylamide gels and transferred tonitrocelullose membrane. After blocking with yeast tRNA (0.05 mg/ml), the mem-brane was probed with intron-less (b. i) and intron-containing (b. ii) 32P-labeledbICP0 probes as described in Section 2. Labeled probes (10,000 cpm) were appliedON for hybridization. After washing the membrane with renaturation buffer (with-oM
tatvctab(2
3
wgicmovmRama
ut tRNAs), the hybridized bands were analyzed by phosphoimaging with a Personalolecular Imager FX.
ogether with bICP0, gB, gC or gD, and pulled down on anti-FLAGffinity beads. Binding of bICP0 gB, gC and gD mRNA to each of theruncations was analyzed by qRT-PCR. Expression of the truncatedersions of VP8 with the expected apparent molecular weights wasonfirmed in Fig. 3a. As shown in Fig. 3b–e, the CT values of the real-ime PCR products indicate binding of deletions D1 (aa 121–742)nd D2 (aa 219–742) as well as DC (aa 1–482) and DD (aa 1–631),ut not D3 (aa 343–742), D4 (aa 538–742), D5 (aa 632–742), DAaa 1–120) and DB (aa 1–258), which indicates that amino acids59–342 of VP8 bind bICP0, gB, gC and gD RNA.
.3. VP8 interacts with mRNAs of bICP0 in vitro
To further confirm the interactions of VP8 with viral mRNA,e performed Northwestern blotting. Out of the four genes (gB,
C gD and bICP0) studied, we selected bICP0 for the in vitronteraction study, because it has an intron, which allows us toompare binding of intron-containing vs. intron-less RNA; further-ore, bICP0 is the only protein that regulates the transcription
f all BoHV-1 genes, and in fact is the key regulator for the acti-ation of all viral promoters of BoHV-1 (Inman et al., 2001). Weade both intron-containing and intron-less radiolabeled bICP0
NA probes by in vitro transcription (Fig. 4a). One �g, two �gnd four �g VP8 were transferred to two separate nitrocelluloseembranes and hybridized individually with the intron-containing
nd intron-less radiolabeled bICP0 probes to explore their binding.
h 197 (2015) 116–126
Phospho-imaging analysis clearly indicated that VP8 binds tointron-less bICP0 probe but not to intron-containing RNA probe(Fig. 4b). Furthermore, neither of the probes bound to the E2 pro-tein of bovine viral diarrhea virus used as control. These resultsindicate that VP8 interacts with intron-less bICP0 mRNA in vitro.
3.4. VP8 interacts with zinc finger and acidic domains of bICP0
Several virus-encoded RNA-binding proteins have been shownto bind to specific RNA sequences that often have defined secondarystructure (Daly et al., 1989). Based on the observed interactionof VP8 with intron-less bICP0 mRNA as determined by North-western blotting and RNA immunoprecipitation, we addressed thequestion which domain of the bICP0 mRNA is responsible for itsinteraction with VP8. There are four domains (zinc finger domain,acidic domain, transactivation domain and nuclear localization sig-nal domain) in the structure of bICP0 (Zhang et al., 2005). The zincfinger and acidic domains of bICP0 were tested to explore theirinteraction with VP8 by RNA EMSA. Radiolabeled probes of thezinc finger and the acidic domains were synthesized by in vitrotranscription using the T7 promoter, and 32P-labeled CTP (Fig. 5a).Three different buffer systems were used to explore the binding.Phosphoimaging data suggest that while VP8 binds to both thezinc finger and the acidic domain, the interaction with the formerappears to be slightly stronger (Fig. 5b and d). To confirm theseinteractions, we performed a competitive binding assay. In thisexperiment, a higher concentration of unlabeled zinc finger probeand acidic probe compared to the corresponding labeled probe wasadded in the reaction mixture and allowed to bind to VP8 at thesame experimental condition. Binding of the labeled probes to VP8was prevented by the unlabeled probes (Fig. 5c and e). Based onthese experimental results, we conclude that VP8 interacts withbICP0 through both the zinc finger and acidic domains.
3.5. Co-localization of VP8 and gB, gC, gD and bICP0 mRNAs
In support of the interactions between VP8 and intron-less gB,gC, gD and bICP0 mRNAs, we investigated the co-localization ofVP8 and the mRNAs of the gB, gC, gD and bICP0 genes by co-transfecting 1 �g pGFP-N1-VP8 plasmid with 1 �g of pSLIA-gB,pSLIA-gC, pSLIA-gD or pCDNA3-bICP0 and 1 �g of the correspond-ing DNA-probes. The DNA probes were modified by inserting threephosphothiolates at the 3′end so that they can penetrate into thenucleus. Transfection with GFP plasmid, DNA probes, VP8-GFP plas-mid or VP8-GFP plasmid and bICP0 DNA probe demonstrated theabsence of non-specific binding of the DNA probe to any cellu-lar components (Fig. 6a). Co-localization of VP8 and mRNAs wasdetermined based on merging green fluorescence-stained VP8 andred fluorescence-stained mRNAs with nuclear (DAPI) staining. Theresults clearly showed that VP8 and the mRNAs were co-localizedin the nucleus (Fig. 6b). However, no obvious co-localization of VP8with mRNAs was observed in the cytoplasm, which may be due tothe diffuse distribution of VP8 in the cytoplasm in contrast to thenucleus where it seems to co-localize in a concentrated mannerin/with nuclear dots.
4. Discussion
In this study, we demonstrated the interaction of VP8 withmRNAs of BoHV-1. Since VP8 is post-translationally modified(phosphorylated, glycosylated and ubiquitinated), which takesplace in mammalian cells, it was expressed and purified from tran-
siently transfected COS-7 cells. For interaction studies in tissueculture, immunoprecipitated and extracted RNAs were analyzedby Bioanalyzer. The data showed that the structural integrity andpurity of RNAs were intact; however, a pool of RNAs was observed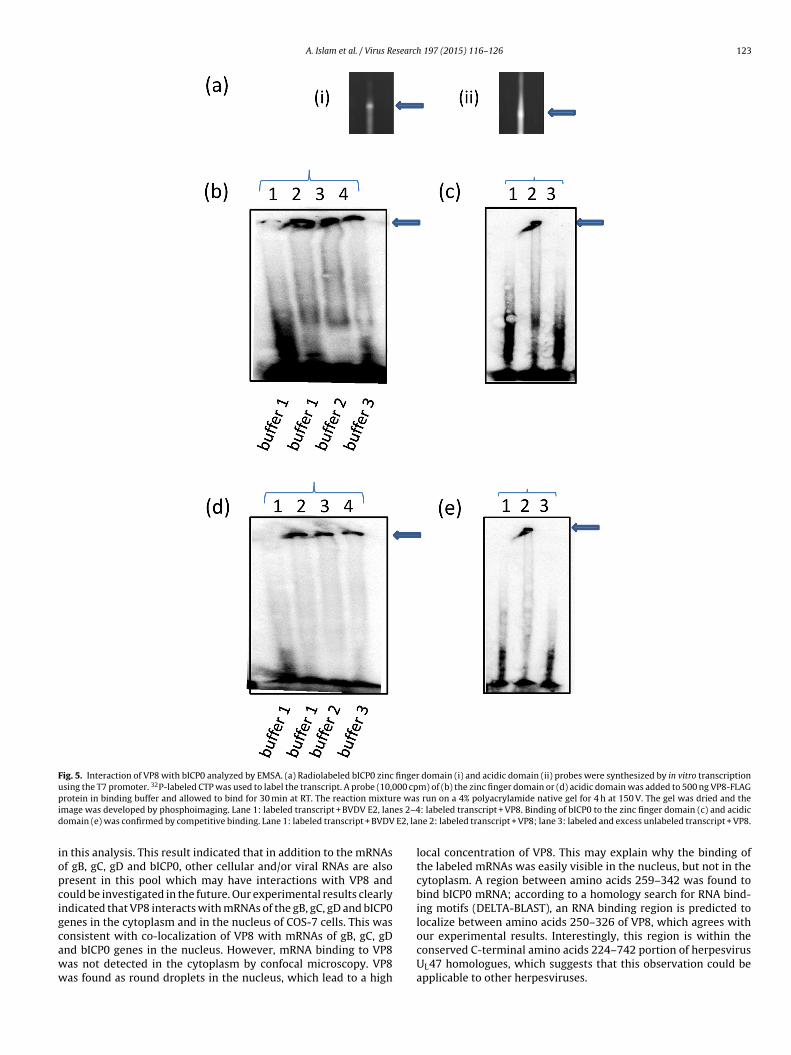
A. Islam et al. / Virus Research 197 (2015) 116–126 123
Fig. 5. Interaction of VP8 with bICP0 analyzed by EMSA. (a) Radiolabeled bICP0 zinc finger domain (i) and acidic domain (ii) probes were synthesized by in vitro transcriptionusing the T7 promoter. 32P-labeled CTP was used to label the transcript. A probe (10,000 cpm) of (b) the zinc finger domain or (d) acidic domain was added to 500 ng VP8-FLAGp re wasi es 2–4d E2, la
iopcigcaww
rotein in binding buffer and allowed to bind for 30 min at RT. The reaction mixtumage was developed by phosphoimaging. Lane 1: labeled transcript + BVDV E2, lanomain (e) was confirmed by competitive binding. Lane 1: labeled transcript + BVDV
n this analysis. This result indicated that in addition to the mRNAsf gB, gC, gD and bICP0, other cellular and/or viral RNAs are alsoresent in this pool which may have interactions with VP8 andould be investigated in the future. Our experimental results clearlyndicated that VP8 interacts with mRNAs of the gB, gC, gD and bICP0enes in the cytoplasm and in the nucleus of COS-7 cells. This was
onsistent with co-localization of VP8 with mRNAs of gB, gC, gDnd bICP0 genes in the nucleus. However, mRNA binding to VP8as not detected in the cytoplasm by confocal microscopy. VP8as found as round droplets in the nucleus, which lead to a highrun on a 4% polyacrylamide native gel for 4 h at 150 V. The gel was dried and the: labeled transcript + VP8. Binding of bICP0 to the zinc finger domain (c) and acidic
ne 2: labeled transcript + VP8; lane 3: labeled and excess unlabeled transcript + VP8.
local concentration of VP8. This may explain why the binding ofthe labeled mRNAs was easily visible in the nucleus, but not in thecytoplasm. A region between amino acids 259–342 was found tobind bICP0 mRNA; according to a homology search for RNA bind-ing motifs (DELTA-BLAST), an RNA binding region is predicted tolocalize between amino acids 250–326 of VP8, which agrees with
our experimental results. Interestingly, this region is within theconserved C-terminal amino acids 224–742 portion of herpesvirusUL47 homologues, which suggests that this observation could beapplicable to other herpesviruses.
1 esearc
dcsiapstmtpaCifa
Fppwo
24 A. Islam et al. / Virus R
Hybridization with radiolabeled probe in Northwestern blottingemonstrated the binding of intron-less mRNA, but not intron-ontaining mRNA, of bICP0. Although the same coding regionequence exists in intron-containing RNA of bICP0 in addition tots long (about 5 kb) intron, it was still unable to bind to VP8. Asccording to previous reports several virus-encoded RNA-bindingroteins bind to specific RNA sequences that often have definedecondary structure (Dingwall et al., 1990), this might be due tohe secondary structure of bICP0 intron-containing RNA, which
ight prevent binding to VP8. Both the zinc finger domain andhe acidic domain of bICP0 interacted with VP8. To address theotential reason for an interaction of both domains with VP8, wenalyzed the bICP0 sequence and observed homologies betweenCGACCTGCCCGC in the zinc finger domain and CCGACCTGACCGC
n the acidic domain. This thirteen base-sequence homology dif-ered by only one base, C in the zinc finger domain and A in thecidic domain. If these sequences indeed are responsible for the
ig. 6. Subcellular localization of VP8 and bICP0, gB, gC and gD mRNAs. Hela cells were tGFP-N1-VP8 plasmid or a combination of pGFP-N1-VP8 and bICP0 DNA probe, or with (bSLIA-gC, or 1 �g pSLIA-gD along with 1 �g corresponding DNA-probe. At 48 h post-tranere then stained with DAPI and images were acquired individually using green (488 nm
f all three colors were merged, and pictures were taken at 1024 × 1024 pixels.
h 197 (2015) 116–126
interaction of bICP0 mRNA with VP8, the C base in the zinc fingerdomain may be more favorable whereas the A base in the acidicdomain may be less favorable.
Previously, another group reported that unlike VP13/14 of HSV-1, VP8 of BoHV1 may require a virus-specific modification to enableits RNA binding activity (Donnelly et al., 2007) as VP8 expressedand purified from E. coli or insect cells as recombinant protein wasunable to bind RNAs (Donnelly et al., 2007). Here we show for thefirst time the interaction of VP8 with single-stranded mRNAs ofBoHV-1 genes in vitro and in vivo outside the context of infection,suggesting that other viral proteins are not needed for its RNA bind-ing activity. This may be explained by the fact that the VP8 that weused for in vitro studies was produced in transfected COS-7 cells, inwhich all post-translational modifications occur. Based on this phe-
nomenon, we can conclude that mammalian cell post-translationalmodifications of VP8 are probably required for its interactionwith mRNAs of BoHV-1for VP8 to bind to RNA, either directly, orransfected in two-well chamber slides with (a) 1 �g of pGFP-N1, bICP0 DNA probe,) 1 �g pGFP-N1-VP8 in combination with 1 �g pCDNA3-bICP0, 1 �g pSLIA-gB, 1 �gsfection, cells were fixed with 3.9% paraformaldehyde and washed with PBS. Cells), blue (461 nm) and red (633 nm) laser conditions by confocal microscopy. Images

A. Islam et al. / Virus Research 197 (2015) 116–126 125
(cotin
iRb(twncq
tcttonosnb2
Fig. 6.
ndirectly through influence on its structure. We localized theNA binding site between amino acids 259–342; there appear toe no sites that are targets for phosphorylation in that regionmanuscript submitted). However, there are several serines andhreonines that can then be targets for O-linked glycosylation. Thisould further support why VP8 from E coli or insect cells doesot bind RNA. Generation of a mutant VP8 without O-linked gly-osylation sites in that region would be required to answer thisuestion.
As VP8 is a well-established nucleocytoplasmic shuttling pro-ein, it may play a role in transport of RNAs from the nucleus to theytoplasm. The concept of RNA transport is supported by the facthat we found interactions between VP8 and viral mRNAs in bothhe nucleus and the cytoplasm, as well as the experimental resultsf another group who showed that actinomycin D inhibits theucleocytoplasmic shuttling of VP8 (Verhagen et al., 2006). Somether herpesvirus tegument proteins such as HVS (herpesvirus
aimiri) ORF59 (Williams et al., 2005) indeed transport VP8 fromucleus to the cytoplasm. Similarly, ICP27 of HSV shuttles mRNAsetween the nucleus and cytoplasm (Chen et al., 2002; Koffa et al.,001).ued.)
Binding and transport of mRNA may occur to prevent the cellularmechanism of mRNA export (Dimaano and Ullman, 2004). Alterna-tively, the nucleocytoplasmic shuttling of VP8 and RNA transportmay be a simultaneous process, and as actinomycin D is an inhibitorof RNA polymerase II, nucleocytoplasmic shuttling of VP8 may alsobe involved in the biogenesis of viral RNAs. Furthermore, it hasbeen reported that both cellular and viral transcripts are pack-aged into HCMV virion particles (Bresnahan and Shenk, 2000). Theviral RNAs are released to the freshly infected cells and allow theexpression of viral genes prior to the transcription of viral genome.Packaged polyadenylated viral RNAs, as well as cellular transcripts,have also been detected in HSV1 virions (Sciortino et al., 2001).The incorporation of these RNAs into viral particles is mediatedby interactions with tegument proteins such as US11, UL47, andUL49 of HSV1 (Sciortino et al., 2002). Based on these publishedreports, it is possible that the interaction of VP8 with viral RNAs maypromote the assembly of viral RNAs, in particular regulatory trans-
cripts (bICP0 and bICP4) that may be immediately translated uponinfection.In conclusion, our experimental results clearly demonstrate theinteraction of VP8 with intron-less mRNAs of BoHV-1 both in vivo

1 esearc
atbTop
A
RCS
R
A
B
C
C
CD
D
D
D
D
D
F
I
J
J
K
L
L
M
Zhang, Y., Zhou, J., Jones, C., 2005. Identification of functional domains within the
26 A. Islam et al. / Virus R
nd in vitro, which suggests a new potential functional charac-eristic of VP8. The RNA binding region was found to be locatedetween amino acids 259–342 in the conserved portion of VP8.his interaction may suggest VP8 to be involved in the transportf intron-less mRNAs from the nucleus to the cytoplasm and couldossibly facilitate the infection process of BoHV-1.
cknowledgements
This work was funded by the Natural Science and Engineeringesearch Council and the Canadian Institute of Health Research,anada. Published as Vaccine and Infectious Disease Organizationeries Number 721.
eferences
bmayr, S.M., Yao, T., Parmely, T., Workman, J.L., 2006. In: Ausubel, F.M., et al. (Eds.),Preparation of nuclear and cytoplasmic extracts from mammalian cells. Currentprotocols in molecular biology. (Chapter 12, Unit 12.11).
resnahan, W.A., Shenk, T., 2000. A subset of viral transcripts packaged withinhuman cytomegalovirus particles. Science 288 (5475), 2373–2376.
arpenter, D.E., Misra, V., 1991. The most abundant protein in bovine herpes 1 viri-ons is a homologue of herpes simplex virus type 1 UL47. J. Gen. Virol. 72 (12),3077–3084.
hen, I.H., Sciabica, K.S., Sandri-Goldin, R.M., 2002. ICP27 interacts with the RNAexport factor Aly/REF to direct herpes simplex virus type 1 intronless mRNAs tothe TAP export pathway. J. Virol. 76 (24), 12877–12889.
ollins, T.J., 2007. ImageJ for microscopy. Biotechniques 43 (Suppl. 1), 25–30.aly, T.J., Cook, K.S., Gray, G.S., Maione, T.E., Rusche, J.R., 1989. Specific binding of
HIV-1 recombinant Rev protein to the Rev-responsive element in vitro. Nature342 (6251), 816–819.
imaano, C., Ullman, K.S., 2004. Nucleocytoplasmic transport: integrating mRNAproduction and turnover with export through the nuclear pore. Mol. Cell. Biol.24 (8), 3069–3076.
ingwall, C., Ernberg, I., Gait, M.J., Green, S.M., Heaphy, S., Karn, J., Lowe, A.D., Singh,M., Skinner, M.A., 1990. HIV-1 tat protein stimulates transcription by binding toa U-rich bulge in the stem of the TAR RNA structure. EMBO J. 9 (12), 4145–4153.
onnelly, M., Elliott, G., 2001a. Fluorescent tagging of herpes simplex virus tegumentprotein VP13/14 in virus infection. J. Virol. 75 (6), 2575–2583.
onnelly, M., Elliott, G., 2001b. Nuclear localization and shuttling of herpes simplexvirus tegument protein VP13/14. J. Virol. 75 (6), 2566–2574.
onnelly, M., Verhagen, J., Elliott, G., 2007. RNA binding by the herpes simplex virustype 1 nucleocytoplasmic shuttling protein UL47 is mediated by an N-Terminalarginine-rich domain that also functions as its nuclear localization signal. J. Virol.81 (5), 2283–2296.
ischer, U.S., Meyer, S., Teufel, M., Heckel, C., Luhrmann, R., Rautmann, G., 1994.Evidence that HIV-1 Rev directly promotes the nuclear export of unspliced RNA.EMBO J. 13 (17), 4105–4112.
nman, M., Zhang, Y., Geiser, V., Jones, C., 2001. The zinc ring finger in the bICP0protein encoded by bovine herpesvirus-1 mediates toxicity and activates pro-ductive infection. J. Gen. Virol. 82 (3), 483–492.
ericho, K.W., Darcel, Q.C., 1978. Response of the respiratory tract of calves kept atcontrolled climatic conditions to bovine herpesvirus 1 in aerosol. Can. J. Comp.Med. 42 (2), 156–167.
ones, C., Chowdhury, S., 2007. A review of the biology of bovine herpesvirus type1 (BHV-1), its role as a cofactor in the bovine respiratory disease complex anddevelopment of improved vaccines. Anim. Health Res. Rev. 8 (2), 187–205.
offa, M.D., Clements, J.B., Izaurralde, E., Wadd, S., Wilson, S.A., Mattaj, I.W., Kuersten,S., 2001. Herpes simplex virus ICP27 protein provides viral mRNAs with accessto the cellular mRNA export pathway. EMBO J. 20 (20), 5769–5778.
abiuk, S.L., Babiuk, L.A., van Drunen Littel-van den Hurk, S., 2009. Major tegumentprotein VP8 of bovine herpesvirus 1 is phosphorylated by viral US3 and cellularCK2 protein kinases. J. Gen. Virol. 90 (Pt 12), 2829–2839.
obanov, V.A., Maher-Sturgess, S.L., Snider, M.G., Lawman, Z., Babiuk, L.A., vanDrunen Littel-van den Hurk, S., 2010. A UL47 gene deletion mutant of bovine
herpesvirus type 1 exhibits impaired growth in cell culture and lack of virulencein cattle. J. Virol. 84 (1), 445–458.eredith, D., Lindsay, J., Halliburton, I., Whittaker, G., 1991. Post-translational mod-ification of the tegument proteins (VP13 and VP14) of herpes simplex virus type1 by glycosylation and phosphorylation. J. Gen. Virol. 72 (11), 2771–2775.
h 197 (2015) 116–126
Mettenleiter, T.C., Klupp, B.G., Granzow, H., 2009. Herpesvirus assembly: an update.Virus Res. 143 (2), 222–234.
Misra, V., Walker, S., Hayes, S., O’Hare, P., 1995. The bovine herpesvirus alphagene trans-inducing factor activates transcription by mechanisms different fromthose of its herpes simplex virus type 1 counterpart VP16. J. Virol. 69 (9),5209–5216.
Phelan, A., Dunlop, J., Clements, J.B., 1996. Herpes simplex virus type 1 protein IE63affects the nuclear export of virus intron-containing transcripts. J. Virol. 70 (8),5255–5265.
Purves, F.C., Roizman, B., 1992. The UL13 gene of herpes simplex virus1 encodesthe functions for posttranslational processing associated with phosphoryla-tion of the regulatory protein alpha 22. Proc. Natl. Acad. Sci. U. S. A. 89 (16),7310–7314.
Read, G.S., Karr, B.M., Knight, K., 1993. Isolation of a herpes simplex virus type 1mutant with a deletion in the virion host shutoff gene and identification ofmultiple forms of the vhs (UL41) polypeptide. J. Virol. 67 (12), 7149–7160.
Robinson, K., Meers, J., Gravel, J., McCarthy, F., Mahony, T., 2008. The essen-tial and non-essential genes of bovine herpesvirus I. J. Gen. Virol. 89 (11),2851–2863.
Roizman, B., Carmichael, L., Reinhardt, F., de-The, G., Nahmias, A., Plowright, W.,Rapp, F., Sheldrick, P., Takahashi, M., Wolf, K., 1981. Herpesviridae. Definition,provisional nomenclature, and taxonomy. The Herpesvirus Study Group, theInternational Committee on Taxonomy of Viruses. Intervirology 16 (4), 201–217.
Salmon, B., Cunningham, C., Davison, A.J., Harris, W.J., Baines, J.D., 1998. The herpessimplex virus type 1 U(L)17 gene encodes virion tegument proteins that arerequired for cleavage and packaging of viral DNA. J. Virol. 72 (5), 3779–3788.
Sandri-Goldin, R.M., 1998. ICP27 mediates HSV RNA export by shuttling through aleucine-rich nuclear export signal and binding viral intronless RNAs through anRGG motif. Genes Dev. 12 (6), 868–879.
Sciortino, M.T., Suzuki, M., Taddeo, B., Roizman, B., 2001. RNAs extracted from her-pes simplex virus 1 virions: apparent selectivity of viral but not cellular RNAspackaged in virions. J. Virol. 75 (12), 8105–8116.
Sciortino, M.T., Taddeo, B., Poon, A.P., Mastino, A., Roizman, B., 2002. Of the threetegument proteins that package mRNA in herpes simplex virions, one (VP22)transports the mRNA to uninfected cells for expression prior to viral infection.Proc. Natl. Acad. Sci. U. S. A. 99 (12), 8318–8323.
Shu, M., Taddeo, B., Zhang, W., Roizman, B., 2013. Selective degradation of mRNAs bythe HSV host shutoff RNase is regulated by the UL47 tegument protein. J. Virol.87 (24), 13569–13578.
Tikoo, S.K., Campos, M., Babiuk, L.A., 1995. Bovine herpesvirus 1 (BHV-1): biology,pathogenesis, and control. Adv. Virus Res. 45, 191–223.
Triezenberg, S.J., Kingsbury, R.C., McKnight, S.L., 1988. Functional dissection of VP16,the trans-activator of herpes simplex virus immediate early gene expression.Genes Dev. 2 (6), 718–729.
van Drunen Littel-van den Hurk, S., Garzon, S., van den Hurk, J.V., Babiuk, L.A., Tijssen,P., 1995. The role of the major tegument protein VP8 of bovine herpesvirus-1 ininfection and immunity. Virology 206 (1), 413–425.
van Drunen Littel-van den Hurk, S., Gifford, G.A., Babiuk, L.A., 1990. Epitopespecificity of the protective immune response induced by individual bovineherpesvirus-1 glycoproteins. Vaccine 8 (4), 358–368.
van Drunen Littel-van den Hurk, S., van den Hurk, J.V., Gilchrist, J.E., Misra, V., Babiuk,L.A., 1984. Interactions of monoclonal antibodies and bovine herpesvirus type 1(BHV-1) glycoproteins: characterization of their biochemical and immunologi-cal properties. Virology 135 (2), 466–479.
van Drunen Littel-van den Hurk, S., Van Donkersgoed, J., Kowalski, J., van den Hurk,J.V., Harland, R., Babiuk, L.A., Zamb, T.J., 1994. A subunit gIV vaccine, producedby transfected mammalian cells in culture, induces mucosal immunity againstbovine herpesvirus-1 in cattle. Vaccine 12 (14), 1295–1302.
Verhagen, J., Hutchinson, I., Elliott, G., 2006. Nucleocytoplasmic shuttling of bovineherpesvirus 1 UL47 protein in infected cells. J. Virol. 80 (2), 1059–1063.
Williams, B.J., Boyne, J.R., Goodwin, D.J., Roaden, L., Hautbergue, G.M., Wilson, S.A.,Whitehouse, A., 2005. The prototype gamma-2 herpesvirus nucleocytoplasmicshuttling protein, ORF 57, transports viral RNA through the cellular mRNA exportpathway. Biochem. J. 387 (Pt 2), 295–308.
Yakhnin, A.V., Yakhnin, H., Babitze, P., 2012. Gel mobility shift assays to detectprotein-RNA interactions. Methods Mol. Biol. 905, 201–211.
Yates, W.D., 1982. A review of infectious bovine rhinotracheitis, shipping fever pneu-monia and viral-bacterial synergism in respiratory disease of cattle. Can. J. Comp.Med. 46 (3), 225–263.
bICP0 protein encoded by bovine herpesvirus 1. J. Gen. Virol. 86 (4), 879–886.Zheng, C., Brownlie, R., Babiuk, L.A., van Drunen Littel-van den Hurk, S., 2004. Char-
acterization of nuclear localization and export signals of the major tegumentprotein VP8 of bovine herpesvirus-1. Virology 324 (2), 327–339.





























