Embed Size (px)
Citation preview

JOURNAL OF VIROLOGY,0022-538X/99/$04.0010
May 1999, p. 3913–3919 Vol. 73, No. 5
Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Role of Cellular Tumor Necrosis Factor Receptor-Associated Factorsin NF-kB Activation and Lymphocyte Transformation
by Herpesvirus Saimiri STPHEUIRAN LEE,1,3 JOONG-KOOK CHOI,1 MENGTAO LI,1 KEN KAYE,2
ELLIOTT KIEFF,2 AND JAE U. JUNG1*
Department of Microbiology and Molecular Genetics, New England Regional Primate Research Center, HarvardMedical School, Southborough, Massachusetts 01772-91021; Brigham and Women’s Hospital,
Harvard Medical School, Boston, Massachusetts 021152; and Yonsei Cancer Center,Institute of Cancer Research, Yonsei University College of Medicine,
134 Sinchon-Dong, Seoul, Korea3
Received 1 December 1998/Accepted 15 February 1999
The STP oncoproteins of the herpesvirus saimiri (HVS) subgroup A strain 11 and subgroup C strain 488 arenow found to be stably associated with tumor necrosis factor receptor-associated factor (TRAF) 1, 2, or 3.Mutational analyses identified residues of PXQXT/S in STP-A11 as critical for TRAF association. In addition,a somewhat divergent region of STP-C488 is critical for TRAF association. Mutational analysis also revealedthat STP-C488 induced NF-kB activation that was correlated with its ability to associate with TRAFs. The HVSSTP-C488 P103R mutant was deficient in human T-lymphocyte transformation to interleukin-2-independentgrowth but showed wild-type phenotype for marmoset T-lymphocyte transformation in vitro and in vivo. TheSTP-C488 P103R mutant was also defective in Rat-1 fibroblast transformation, and fibroblast cell transfor-mation was blocked by a TRAF2 dominant-negative mutant. These data implicate TRAFs in STP-C488-mediated transformation of human lymphocytes and rodent fibroblasts. Other factors are implicated inimmortalization of common marmoset T lymphocytes and may also be critical in the transformation of humanlymphocytes and rodent fibroblasts.
Members of the tumor necrosis factor (TNF) receptor(TNFR) superfamily are important for lymphoid organ devel-opment, lymphocyte activation, acute-phase responses, cellgrowth, and apoptosis (3, 22, 49). The TNFR superfamily in-cludes TNFR1, TNFR2, CD27, CD30, CD40, Fas (CD95),4-1BB, and OX40 (45). The cytoplasmic regions of TNFR1,TNFR2, CD30, or CD40 are required for receptor-mediatedsignaling and interact with TNF receptor-associated factors(TRAFs) (8, 23, 43, 48). TRAF2 and TRAF3 have an aminoterminal RING finger structure, and all TRAFs have morethan one zinc finger. TRAFs also have a predicted extendedalpha-helical coiled-coiled hydrophobic heptad repeat domain(8). A carboxyl-terminal TRAF domain of approximately 200amino acids can be further divided into the TRAF-N andTRAF-C subdomains. The highly conserved carboxyl-terminalTRAF-C domain mediates interaction with TNFRs (48). Thecytoplasmic region of LMP1, which is a key effector of Epstein-Barr virus (EBV)-mediated transformation, also binds toTRAFs, and this interaction is essential for B lymphocytegrowth transformation (24, 38). Human and simian LMP1,CD40, and CD30 and a cytoplasmic TRAF-interacting protein,TANK, share a PXQXT/S core sequence through which theyinteract with TRAFs (8, 20, 21). TRAF2 is a mediator ofNF-kB and Jun kinase activation from the TNFRs and LMP1(13, 35, 46, 54).
Herpesvirus saimiri (HVS), a gamma-2 herpesvirus or rhadi-novirus, infects most squirrel monkeys without causing appar-ent disease (11, 17). In other nonhuman primates, however,
HVS induces rapidly fatal T-cell lymphoproliferative diseases(19, 27). Sequence divergence among HVS isolates is mostextensive at the left end of the viral genomic DNA and is thebasis for the classification of HVS into subgroups A, B, and C(9, 36). Sequence variation in the STP gene in this regioncorrelates with different capacities for immortalizing T lym-phocytes in vitro and for inducing lymphoma in nonhumanprimates (5, 9, 12, 32). Subgroup A and subgroup C viruses canimmortalize common marmoset T lymphocytes to interleu-kin-2 (IL-2)-independent proliferation (12, 47). Highly onco-genic subgroup C strains are also able to immortalize human,rabbit, and rhesus monkey lymphocytes and induce fulminantlymphoma in rhesus monkeys (2, 4, 6, 37).
The STPs of subgroup A or C strains (STP-A or STP-C) cantransform rodent fibroblast cells in vitro. STP-C is considerablymore potent (28, 30). STP-C488 associates with cellular Ras intransformed cells (25). Mutations that disrupt Ras associationdisrupt the transforming ability of the STP-C488 oncogene(25). In contrast, STP-A binds to the SH2 domain of Src kinaseand is phosphorylated by the associated Src kinase (34). Trans-genic mice expressing STP-C488 developed invasive epithelialcell tumors (39), while STP-A11 transgenic mice developedperipheral pleomorphic T-cell lymphomas (33). Deletion ofSTP from the group C strain 488 or from the group A strain 11yields viruses that are no longer capable of immortalizing lym-phocytes in vitro or of inducing fatal lymphomas in commonmarmosets (9, 10, 12, 14, 32, 40). Since HVS lacking STP canbe repeatedly isolated from the peripheral blood of commonmarmosets for months or years, STP is not required for viralreplication or persistence in vivo, but it is essential for trans-formation in cell culture and for lymphoma induction in com-mon marmosets (9, 14).
STP-A11 and STP-C488 are similar in genome location and
* Corresponding author. Mailing address: New England RegionalPrimate Research Center, Harvard Medical School, 1 Pine Hill Dr.,P.O. Box 9102, Southborough, MA 01772. Phone: (508) 624-8083. Fax:(508) 624-8190. E-mail: [email protected].
3913
on Decem
ber 3, 2014 by guesthttp://jvi.asm
.org/D
ownloaded from

orientation and have limited sequence similarity (5, 28). Bothopen reading frames encode a highly acidic amino terminus.STP-C488 has 18 direct repeats of a collagen-like motif (Gly-Pro-Pro or Gly-Pro-Gln) that comprises more than 50% of theprotein and is predicted to have an alpha-helical triple struc-ture (5). A mutation which disrupts the collagen repeats hasbeen shown to disrupt the transforming activity of STP-C488(26). STP-A11 is also glycine and proline rich. STP-A11 andSTP-C488 proteins have highly hydrophobic carboxyl terminiwhich are sufficient for membrane interaction (26). SinceSTP-C likely oligomerizes through its collagen-like motif andassociates with cellular membranes, STP-C may mimic a li-gand-independent constitutively active receptor, like the EBVLMP1 protein. STP-A could have similar properties.
In this report, we investigate this hypothesis and find thatSTP-A and STP-C associate with TRAF 1, 2, or 3. Further-more, the STP-C488 TRAF binding site is required for NF-kBactivation and cell growth transformation.
MATERIALS AND METHODS
Cell culture and virus propagation. Rat-1, COS-1, BOSC23, and owl monkeykidney (OMK 1637) cells cultivated in Dulbecco’s modified Eagle’s medium orminimal essential medium supplemented with penicillin, streptomycin, L-glu-tamine, and 10% (vol/vol) heat-inactivated fetal bovine serum (GIBCO BRL,Grand Island, N.Y.) were used for the propagation of the HVS strain C488.Low-passage-number OMK cells (,30 passages) were used for the transfections.Primary human and common marmoset (Callithrix jacchus) peripheral bloodmononuclear cells (PBMCs) were purified by using lymphocyte separation me-dium (Organon Teknika Corp., Malvern, Pa.). Cultures of human and commonmarmoset PBMCs in immortalization assays with HVS recombinants were per-formed in RPMI 1640 medium supplemented with penicillin, streptomycin, Fun-gizone, L-glutamine, 20% (vol/vol) heat-inactivated fetal bovine serum, and 5 mgof b-mercaptoethanol per liter. A DEAE or calcium phosphate transfection wasused for transient expression in COS-1 or BOSC23 cells, respectively. Rat-1 cellswere transfected with plasmid pcDNA3-TRAF2 D6–86 by the calcium phosphateprotocol, followed by the selection with 500 mg of G418 per ml. Subsequently,these cells were transfected with pBabe-puro or pBabe-STP-C488 and selectedwith 5 mg of puromycin/ml.
Plasmid constructions. All STP-C488 mutants have been described previously(26). Mutations in STP-A11 were generated by PCR by using oligonucleotide-directed mutagenesis (16). Oligonucleotide mutant primers from complementarystrands of STP-A11 were synthesized with specific restriction enzyme sites within59 and 39 primers to facilitate cloning into pBluescript KS(1) (Stratagene, SanDiego, Calif.). PCR was carried out with a DNA thermal cycler (Perkin-ElmerCetus Instruments, Norwalk, Conn.) under the following conditions: 30 cycles of1 min at 55°C for annealing, 4 min at 72°C for polymerization, and 1 min at 94°Cfor denaturation. The amplified DNA fragments containing mutations in STP-A11 were purified and cloned into the pBluescript KS(1) vector. Each STP-A11mutant was completely sequenced to verify the presence of the mutation and theabsence of any other changes. After confirmation of the DNA sequence, DNAcontaining the desired STP-A11 mutation was recloned into EcoRI and BglIIcloning sites of the pFJ vector for gene expression. TRAF2 D6–86 and flag-tagged TRAF1, TRAF2 and TRAF3 have been described previously (31). Flag-tagged TRAF3-C and TRAF3 DC were constructed by PCR by using oligonu-cleotide-directed mutagenesis and completely sequenced to verify the presenceof the mutation and the absence of any other changes.
Virion DNA isolation. HVS virion preparations were obtained from the virus-containing medium by removal of OMK cell debris by low-speed centrifugation,followed by pelleting of the virus at 18,000 rpm for 2 h in an SS-34 rotor. Topurify intact virion DNA, the virus was disrupted at 60°C for 2 h in lysis buffercontaining 10 mM Tris (pH 8.5), 1 mM EDTA, 1% (vol/vol) Sarkosyl, and 0.1 mgof proteinase K/ml. Extraction of the aqueous solution, first with an equalvolume of phenol and then twice with chloroform, was sufficient to purify thevirion DNA for use in transfections. Sterile cut pipette tips were used formanipulating virion DNA without shearing.
Construction of recombinant HVS. Generation of the P103R mutation inSTP-C488 by oligonucleotide-directed mutagenesis has been described previ-ously (26). A 3.6-kb clone, pNEB-C488-PX, containing the tyrosine kinase-interacting protein tip, STP-C488, and herpesvirus saimiri U RNAs (HSURs) (5,15), was used to provide a subcloning vector with flanking sequence adequate tofacilitate recombination during cotransfection. Digestion of this vector withEcoRV and SpeI permitted insertion of the corresponding STP fragment con-taining the P103R mutation.
Linearized plasmid DNA containing the 3.6-kb viral DNA with the STP-C488/P103R mutation was cotransfected into OMK cells with HVSDSTP/SV40-SEAPvirion DNA by the calcium phosphate protocol. A pure form of recombinantvirus with the secreted engineered alkaline phosphatase (SEAP) reporter re-
placed with STP-C488/P103R was isolated by limiting dilution and repeatedselection of SEAP-negative virus onto OMK cell monolayers in 48-well tissueculture plates performed as described previously (15). SEAP production wasdetected by a liquid scintillation counter; the chemiluminescence produced incell culture medium was assayed by using Phospha-Light reagents (Tropix Inc.,Bedford, Mass.) according to the manufacturer’s recommendations.
In vitro immortalization of human and common marmoset lymphocytes. As-says of lymphocyte immortalization in vitro have been described previously (14).PBMCs were isolated from heparinized blood specimens from human donorsand common marmosets (C. jacchus) by centrifugation through lymphocyteseparation medium (Organon Teknika Corp.) followed by washing in RPMIculture medium. PBMCs from each donor were individually washed, resus-pended in RPMI, and then distributed in 1-ml volumes containing approximately106 cells into 12-well tissue culture plates. A single well containing PBMCs fromeach donor was then infected at a multiplicity of infection ranging from 1 to 5with 1 ml of fresh, purified HVS viral stocks. Cells were maintained with RPMI1640 growth medium which was changed every 3 to 4 days. Immortalization orcell death was assessed microscopically.
Experimental infection of common marmosets. The in vivo oncogenicity of theHVS-C488 recombinants was assessed by experimental infection of commonmarmosets. Marmosets were injected intramuscularly with 105 50% tissue cultureinfective doses of virus in a volume of 1 ml. Sera and blood cell pellets werecollected and frozen at 270°C weekly during the first 4 weeks and every 2 weeksthereafter. Viral loads in PBMC specimens were assessed periodically by dupli-cate plating of 106 PBMCs and serial threefold dilutions of PBMCs on OMKcells in 24-well tissue culture plates (14). Animals that became moribund wereeuthanized and received complete necropsies. Tissues were fixed in 10% neutralbuffered formalin, embedded in paraffin, sectioned, and stained with hematoxylinand eosin.
Immunoprecipitation, immunoblotting, and antibodies. Cells were harvestedand lysed with lysis buffer (0.15 M NaCl, 0.5% Nonidet P-40, and 50 mM HEPESbuffer [pH 8.0]) containing 1 mM Na2VO3, 1 mM NaF, and protease inhibitors(leupeptin, aprotinin, phenylmethylsulfonyl fluoride, pepstatin, and bestatin).Precleared lysates were used for immunoprecipitation or immunoblot analysis.The rabbit polyclonal antibody 109 and mouse monoclonal antibody A1.4 di-rected against STP-C488 used in these experiments have been described previ-ously (28). Flag and AU-1 antibodies were purchased from KODAK IBI (NewHaven, Conn.) and BABCO Biotech (Berkeley, Calif.), respectively.
Isolation of genomic DNA and PCR analysis. Genomic DNA was isolated witha Qiagene genomic isolation kit according to the manufacturer’s protocol. Fivemicrograms of purified genomic DNA was used for PCR amplification per-formed by using a 59 primer which corresponds to the upstream sequence of theSTP-C488 gene and a 39 primer which corresponds to the downstream sequenceof STP-C488. Amplified DNA was cloned into the TA cloning vector (Invitro-gen, San Diego, Calif.). Both strands from each of five independent clones weresubsequently sequenced using an ABI PRISM 377 automatic DNA sequencer.
Reporter assays. All transfections included pGKbgal, which expresses b-ga-lactosidase from a phosphoglucokinase promoter, together with 3X-kB-luc,which has three copies of the NF-kB binding site from the murine major histo-compatibility complex class I promoter upstream of a minimal fos promoter anda luciferase gene. At 48 h posttransfection, cells were washed once in phosphate-buffered saline and lysed in 200 ml of reporter lysis buffer (Promega, Madison,Wis.). Assays for luciferase were performed with a Luminometer by using aluciferase assay (Promega). Values were normalized to b-galactosidase activity.
RESULTS
HVS subgroup A and subgroup C STPs interact withTRAFs. HVS STP-A has a PXQXT sequence similar to that inLMP1, CD30, CD40 and TANK (7, 20, 21), while STP-C488and STP-C484M have more divergent sequences with glutamicacid in place of glutamine (Fig. 1). The potential interaction ofSTP with TRAFs was therefore investigated by transfectingBOSC23 cells with expression vectors for STP-A11 or STP-C488 and for TRAF1, TRAF2, or TRAF3. The amino terminiof the TRAFs and STP-A11 were tagged with Flag and AU-1epitopes, respectively. After transfection, TRAF complexeswere precipitated with an anti-Flag antibody and STP-A11 orSTP-C488 was detected by immunoblotting. STP-C488 wasreadily evident at 20 kDa in the TRAF 1, 2, or 3 precipitates(Fig. 2A). STP-A11 was expressed as 26- and 35-kDa proteins(34). STP-A11 and STP-C488 were not detected in precipitatesfrom negative control cell lysates (Fig. 2A and B, lanes 2).These tests demonstrate that STP-A11 and STP-C488 inter-acted specifically with TRAFs in BOSC23 cells.
Different regions of TRAF are required for binding to STP-Aand STP-C. To confirm that TRAFs bind through their
3914 LEE ET AL. J. VIROL.
on Decem
ber 3, 2014 by guesthttp://jvi.asm
.org/D
ownloaded from
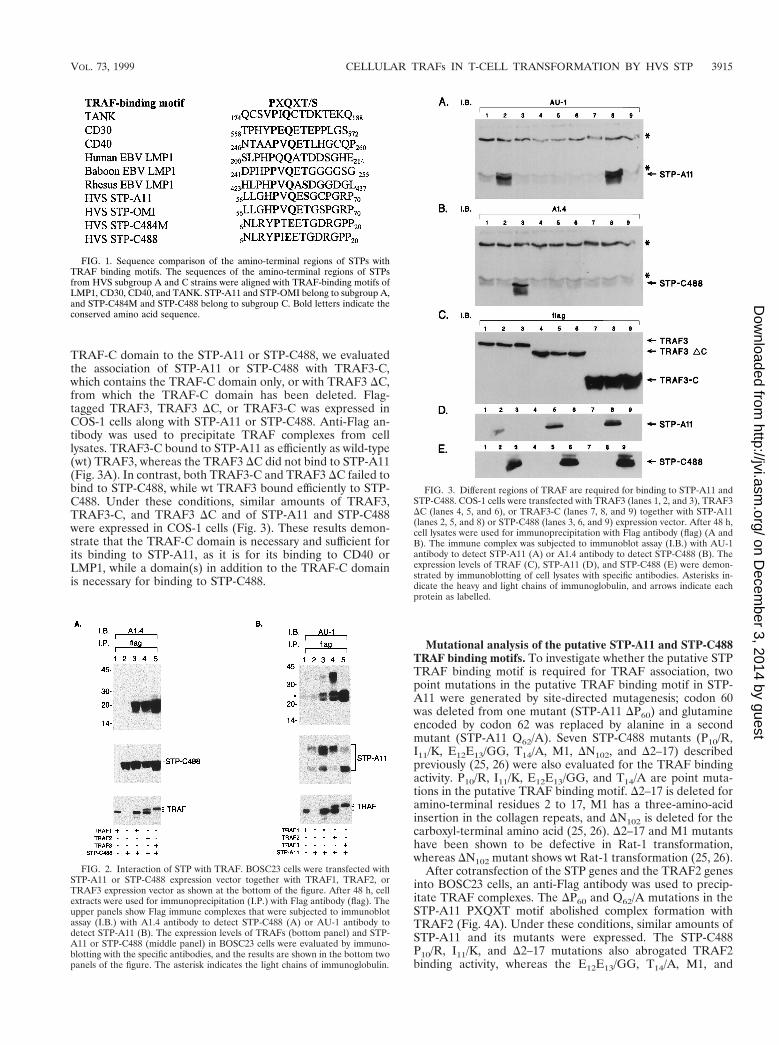
TRAF-C domain to the STP-A11 or STP-C488, we evaluatedthe association of STP-A11 or STP-C488 with TRAF3-C,which contains the TRAF-C domain only, or with TRAF3 DC,from which the TRAF-C domain has been deleted. Flag-tagged TRAF3, TRAF3 DC, or TRAF3-C was expressed inCOS-1 cells along with STP-A11 or STP-C488. Anti-Flag an-tibody was used to precipitate TRAF complexes from celllysates. TRAF3-C bound to STP-A11 as efficiently as wild-type(wt) TRAF3, whereas the TRAF3 DC did not bind to STP-A11(Fig. 3A). In contrast, both TRAF3-C and TRAF3 DC failed tobind to STP-C488, while wt TRAF3 bound efficiently to STP-C488. Under these conditions, similar amounts of TRAF3,TRAF3-C, and TRAF3 DC and of STP-A11 and STP-C488were expressed in COS-1 cells (Fig. 3). These results demon-strate that the TRAF-C domain is necessary and sufficient forits binding to STP-A11, as it is for its binding to CD40 orLMP1, while a domain(s) in addition to the TRAF-C domainis necessary for binding to STP-C488.
Mutational analysis of the putative STP-A11 and STP-C488TRAF binding motifs. To investigate whether the putative STPTRAF binding motif is required for TRAF association, twopoint mutations in the putative TRAF binding motif in STP-A11 were generated by site-directed mutagenesis; codon 60was deleted from one mutant (STP-A11 DP60) and glutamineencoded by codon 62 was replaced by alanine in a secondmutant (STP-A11 Q62/A). Seven STP-C488 mutants (P10/R,I11/K, E12E13/GG, T14/A, M1, DN102, and D2–17) describedpreviously (25, 26) were also evaluated for the TRAF bindingactivity. P10/R, I11/K, E12E13/GG, and T14/A are point muta-tions in the putative TRAF binding motif. D2–17 is deleted foramino-terminal residues 2 to 17, M1 has a three-amino-acidinsertion in the collagen repeats, and DN102 is deleted for thecarboxyl-terminal amino acid (25, 26). D2–17 and M1 mutantshave been shown to be defective in Rat-1 transformation,whereas DN102 mutant shows wt Rat-1 transformation (25, 26).
After cotransfection of the STP genes and the TRAF2 genesinto BOSC23 cells, an anti-Flag antibody was used to precip-itate TRAF complexes. The DP60 and Q62/A mutations in theSTP-A11 PXQXT motif abolished complex formation withTRAF2 (Fig. 4A). Under these conditions, similar amounts ofSTP-A11 and its mutants were expressed. The STP-C488P10/R, I11/K, and D2–17 mutations also abrogated TRAF2binding activity, whereas the E12E13/GG, T14/A, M1, and
FIG. 1. Sequence comparison of the amino-terminal regions of STPs withTRAF binding motifs. The sequences of the amino-terminal regions of STPsfrom HVS subgroup A and C strains were aligned with TRAF-binding motifs ofLMP1, CD30, CD40, and TANK. STP-A11 and STP-OMI belong to subgroup A,and STP-C484M and STP-C488 belong to subgroup C. Bold letters indicate theconserved amino acid sequence.
FIG. 2. Interaction of STP with TRAF. BOSC23 cells were transfected withSTP-A11 or STP-C488 expression vector together with TRAF1, TRAF2, orTRAF3 expression vector as shown at the bottom of the figure. After 48 h, cellextracts were used for immunoprecipitation (I.P.) with Flag antibody (flag). Theupper panels show Flag immune complexes that were subjected to immunoblotassay (I.B.) with A1.4 antibody to detect STP-C488 (A) or AU-1 antibody todetect STP-A11 (B). The expression levels of TRAFs (bottom panel) and STP-A11 or STP-C488 (middle panel) in BOSC23 cells were evaluated by immuno-blotting with the specific antibodies, and the results are shown in the bottom twopanels of the figure. The asterisk indicates the light chains of immunoglobulin.
FIG. 3. Different regions of TRAF are required for binding to STP-A11 andSTP-C488. COS-1 cells were transfected with TRAF3 (lanes 1, 2, and 3), TRAF3DC (lanes 4, 5, and 6), or TRAF3-C (lanes 7, 8, and 9) together with STP-A11(lanes 2, 5, and 8) or STP-C488 (lanes 3, 6, and 9) expression vector. After 48 h,cell lysates were used for immunoprecipitation with Flag antibody (flag) (A andB). The immune complex was subjected to immunoblot assay (I.B.) with AU-1antibody to detect STP-A11 (A) or A1.4 antibody to detect STP-C488 (B). Theexpression levels of TRAF (C), STP-A11 (D), and STP-C488 (E) were demon-strated by immunoblotting of cell lysates with specific antibodies. Asterisks in-dicate the heavy and light chains of immunoglobulin, and arrows indicate eachprotein as labelled.
VOL. 73, 1999 CELLULAR TRAFs IN T-CELL TRANSFORMATION BY HVS STP 3915
on Decem
ber 3, 2014 by guesthttp://jvi.asm
.org/D
ownloaded from
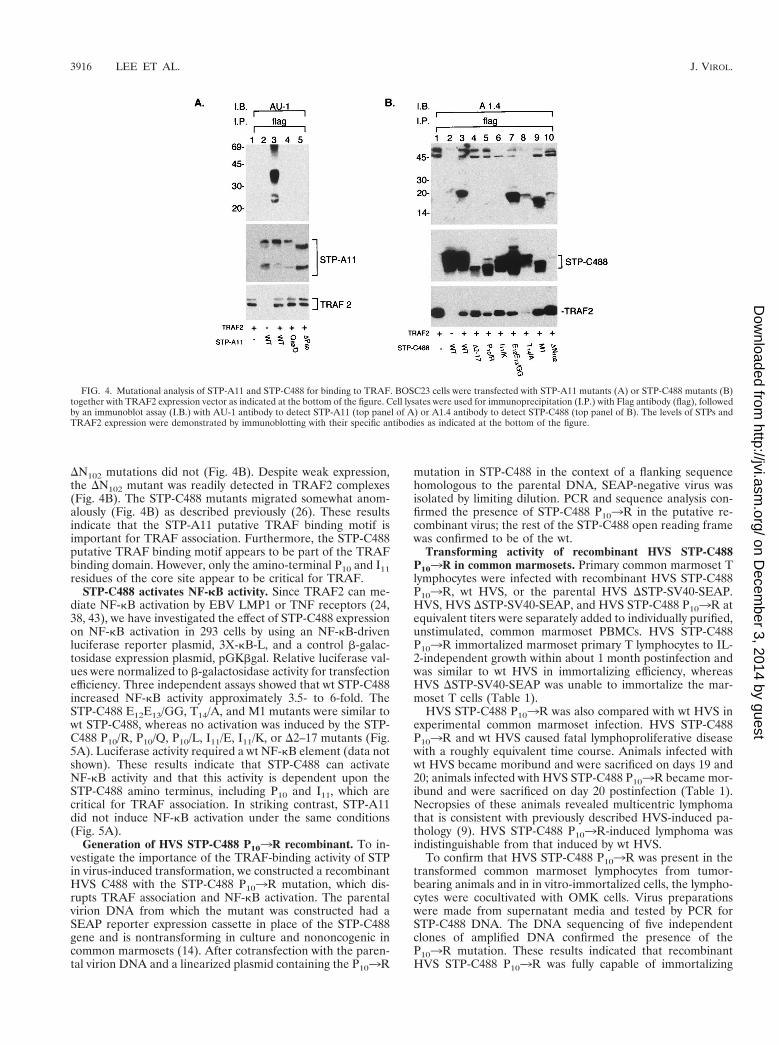
DN102 mutations did not (Fig. 4B). Despite weak expression,the DN102 mutant was readily detected in TRAF2 complexes(Fig. 4B). The STP-C488 mutants migrated somewhat anom-alously (Fig. 4B) as described previously (26). These resultsindicate that the STP-A11 putative TRAF binding motif isimportant for TRAF association. Furthermore, the STP-C488putative TRAF binding motif appears to be part of the TRAFbinding domain. However, only the amino-terminal P10 and I11residues of the core site appear to be critical for TRAF.
STP-C488 activates NF-kB activity. Since TRAF2 can me-diate NF-kB activation by EBV LMP1 or TNF receptors (24,38, 43), we have investigated the effect of STP-C488 expressionon NF-kB activation in 293 cells by using an NF-kB-drivenluciferase reporter plasmid, 3X-kB-L, and a control b-galac-tosidase expression plasmid, pGKbgal. Relative luciferase val-ues were normalized to b-galactosidase activity for transfectionefficiency. Three independent assays showed that wt STP-C488increased NF-kB activity approximately 3.5- to 6-fold. TheSTP-C488 E12E13/GG, T14/A, and M1 mutants were similar towt STP-C488, whereas no activation was induced by the STP-C488 P10/R, P10/Q, P10/L, I11/E, I11/K, or D2–17 mutants (Fig.5A). Luciferase activity required a wt NF-kB element (data notshown). These results indicate that STP-C488 can activateNF-kB activity and that this activity is dependent upon theSTP-C488 amino terminus, including P10 and I11, which arecritical for TRAF association. In striking contrast, STP-A11did not induce NF-kB activation under the same conditions(Fig. 5A).
Generation of HVS STP-C488 P103R recombinant. To in-vestigate the importance of the TRAF-binding activity of STPin virus-induced transformation, we constructed a recombinantHVS C488 with the STP-C488 P103R mutation, which dis-rupts TRAF association and NF-kB activation. The parentalvirion DNA from which the mutant was constructed had aSEAP reporter expression cassette in place of the STP-C488gene and is nontransforming in culture and nononcogenic incommon marmosets (14). After cotransfection with the paren-tal virion DNA and a linearized plasmid containing the P103R
mutation in STP-C488 in the context of a flanking sequencehomologous to the parental DNA, SEAP-negative virus wasisolated by limiting dilution. PCR and sequence analysis con-firmed the presence of STP-C488 P103R in the putative re-combinant virus; the rest of the STP-C488 open reading framewas confirmed to be of the wt.
Transforming activity of recombinant HVS STP-C488P103R in common marmosets. Primary common marmoset Tlymphocytes were infected with recombinant HVS STP-C488P103R, wt HVS, or the parental HVS DSTP-SV40-SEAP.HVS, HVS DSTP-SV40-SEAP, and HVS STP-C488 P103R atequivalent titers were separately added to individually purified,unstimulated, common marmoset PBMCs. HVS STP-C488P103R immortalized marmoset primary T lymphocytes to IL-2-independent growth within about 1 month postinfection andwas similar to wt HVS in immortalizing efficiency, whereasHVS DSTP-SV40-SEAP was unable to immortalize the mar-moset T cells (Table 1).
HVS STP-C488 P103R was also compared with wt HVS inexperimental common marmoset infection. HVS STP-C488P103R and wt HVS caused fatal lymphoproliferative diseasewith a roughly equivalent time course. Animals infected withwt HVS became moribund and were sacrificed on days 19 and20; animals infected with HVS STP-C488 P103R became mor-ibund and were sacrificed on day 20 postinfection (Table 1).Necropsies of these animals revealed multicentric lymphomathat is consistent with previously described HVS-induced pa-thology (9). HVS STP-C488 P103R-induced lymphoma wasindistinguishable from that induced by wt HVS.
To confirm that HVS STP-C488 P103R was present in thetransformed common marmoset lymphocytes from tumor-bearing animals and in in vitro-immortalized cells, the lympho-cytes were cocultivated with OMK cells. Virus preparationswere made from supernatant media and tested by PCR forSTP-C488 DNA. The DNA sequencing of five independentclones of amplified DNA confirmed the presence of theP103R mutation. These results indicated that recombinantHVS STP-C488 P103R was fully capable of immortalizing
FIG. 4. Mutational analysis of STP-A11 and STP-C488 for binding to TRAF. BOSC23 cells were transfected with STP-A11 mutants (A) or STP-C488 mutants (B)together with TRAF2 expression vector as indicated at the bottom of the figure. Cell lysates were used for immunoprecipitation (I.P.) with Flag antibody (flag), followedby an immunoblot assay (I.B.) with AU-1 antibody to detect STP-A11 (top panel of A) or A1.4 antibody to detect STP-C488 (top panel of B). The levels of STPs andTRAF2 expression were demonstrated by immunoblotting with their specific antibodies as indicated at the bottom of the figure.
3916 LEE ET AL. J. VIROL.
on Decem
ber 3, 2014 by guesthttp://jvi.asm
.org/D
ownloaded from
common marmoset lymphocytes to continuous growth and ofinducing lymphoma in common marmosets. Thus, the TRAFbinding activity of STP-C488 is not essential for virus-inducedtransformation of primary marmoset lymphocytes in vitro andin vivo.
Transformation of human lymphocytes by recombinantHVS STP-C488 P103R. To further examine the role of STP-TRAF interaction in transformation, HVS STP-C488 P103Rwas compared with wt HVS and HVS DSTP-SV40-SEAP inprimary human T-lymphocyte immortalization assays. Equiva-
lent titers of HVS STP-C488 P103R, wt HVS, and HVSDSTP-SV40-SEAP were used to infect unstimulated, humanPBMCs from five donors. HVS transformed the primary hu-man lymphocytes from three of the five donors to IL-2-inde-pendent growth within about 2 months postinfection, whileHVS STP-C488/P103R and parental HVSDSTP-SV40-SEAPfailed to immortalize primary human T lymphocytes to IL-2-independent growth (Table 1). These results indicate that STP-TRAF interaction is required for primary human lymphocytetransformation.
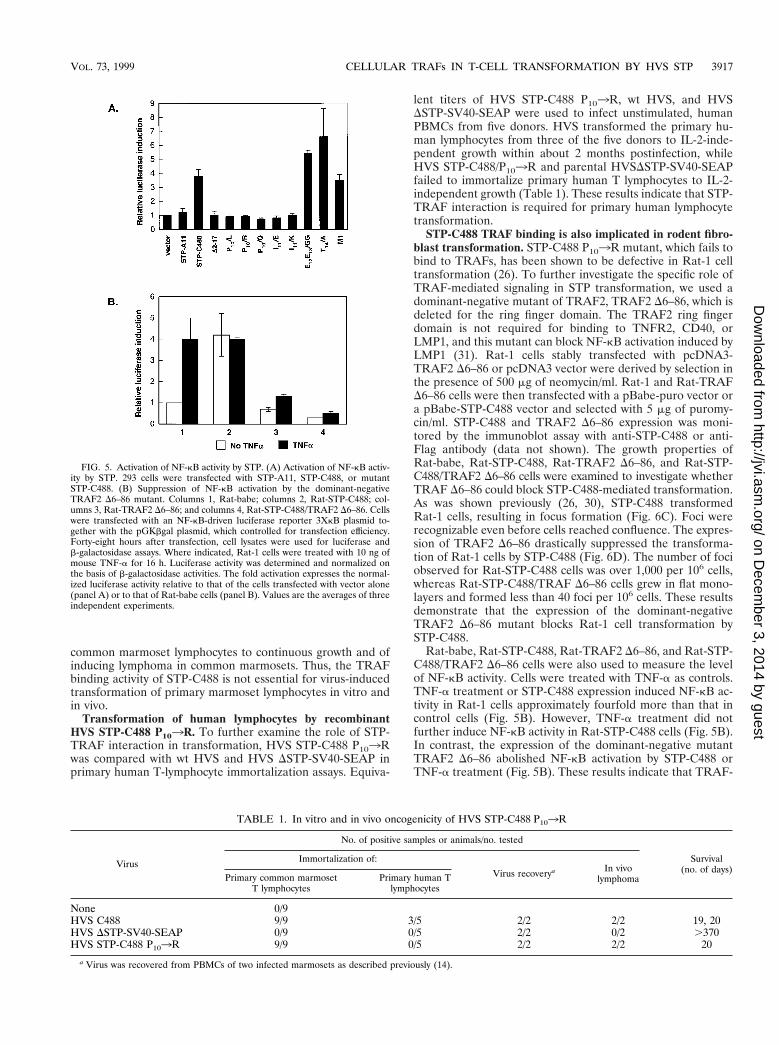
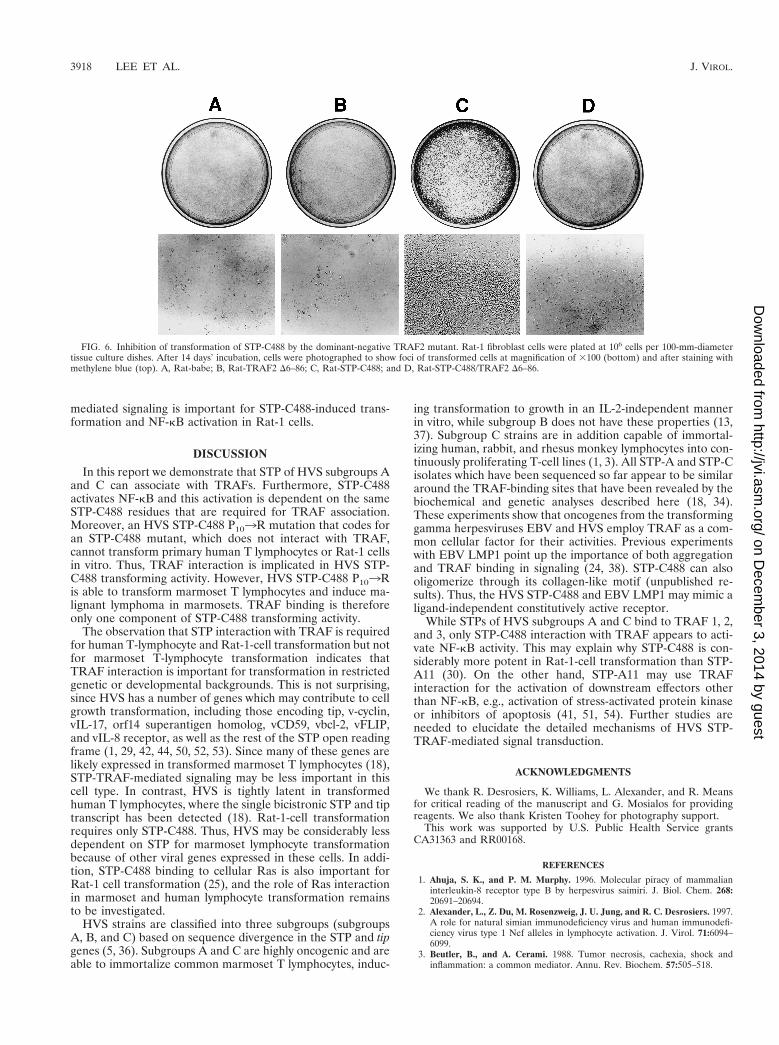
STP-C488 TRAF binding is also implicated in rodent fibro-blast transformation. STP-C488 P103R mutant, which fails tobind to TRAFs, has been shown to be defective in Rat-1 celltransformation (26). To further investigate the specific role ofTRAF-mediated signaling in STP transformation, we used adominant-negative mutant of TRAF2, TRAF2 D6–86, which isdeleted for the ring finger domain. The TRAF2 ring fingerdomain is not required for binding to TNFR2, CD40, orLMP1, and this mutant can block NF-kB activation induced byLMP1 (31). Rat-1 cells stably transfected with pcDNA3-TRAF2 D6–86 or pcDNA3 vector were derived by selection inthe presence of 500 mg of neomycin/ml. Rat-1 and Rat-TRAFD6–86 cells were then transfected with a pBabe-puro vector ora pBabe-STP-C488 vector and selected with 5 mg of puromy-cin/ml. STP-C488 and TRAF2 D6–86 expression was moni-tored by the immunoblot assay with anti-STP-C488 or anti-Flag antibody (data not shown). The growth properties ofRat-babe, Rat-STP-C488, Rat-TRAF2 D6–86, and Rat-STP-C488/TRAF2 D6–86 cells were examined to investigate whetherTRAF D6–86 could block STP-C488-mediated transformation.As was shown previously (26, 30), STP-C488 transformedRat-1 cells, resulting in focus formation (Fig. 6C). Foci wererecognizable even before cells reached confluence. The expres-sion of TRAF2 D6–86 drastically suppressed the transforma-tion of Rat-1 cells by STP-C488 (Fig. 6D). The number of fociobserved for Rat-STP-C488 cells was over 1,000 per 106 cells,whereas Rat-STP-C488/TRAF D6–86 cells grew in flat mono-layers and formed less than 40 foci per 106 cells. These resultsdemonstrate that the expression of the dominant-negativeTRAF2 D6–86 mutant blocks Rat-1 cell transformation bySTP-C488.
Rat-babe, Rat-STP-C488, Rat-TRAF2 D6–86, and Rat-STP-C488/TRAF2 D6–86 cells were also used to measure the levelof NF-kB activity. Cells were treated with TNF-a as controls.TNF-a treatment or STP-C488 expression induced NF-kB ac-tivity in Rat-1 cells approximately fourfold more than that incontrol cells (Fig. 5B). However, TNF-a treatment did notfurther induce NF-kB activity in Rat-STP-C488 cells (Fig. 5B).In contrast, the expression of the dominant-negative mutantTRAF2 D6–86 abolished NF-kB activation by STP-C488 orTNF-a treatment (Fig. 5B). These results indicate that TRAF-
FIG. 5. Activation of NF-kB activity by STP. (A) Activation of NF-kB activ-ity by STP. 293 cells were transfected with STP-A11, STP-C488, or mutantSTP-C488. (B) Suppression of NF-kB activation by the dominant-negativeTRAF2 D6–86 mutant. Columns 1, Rat-babe; columns 2, Rat-STP-C488; col-umns 3, Rat-TRAF2 D6–86; and columns 4, Rat-STP-C488/TRAF2 D6–86. Cellswere transfected with an NF-kB-driven luciferase reporter 3XkB plasmid to-gether with the pGKbgal plasmid, which controlled for transfection efficiency.Forty-eight hours after transfection, cell lysates were used for luciferase andb-galactosidase assays. Where indicated, Rat-1 cells were treated with 10 ng ofmouse TNF-a for 16 h. Luciferase activity was determined and normalized onthe basis of b-galactosidase activities. The fold activation expresses the normal-ized luciferase activity relative to that of the cells transfected with vector alone(panel A) or to that of Rat-babe cells (panel B). Values are the averages of threeindependent experiments.
TABLE 1. In vitro and in vivo oncogenicity of HVS STP-C488 P103R
Virus
No. of positive samples or animals/no. tested
Survival(no. of days)
Immortalization of:
Virus recoverya In vivolymphomaPrimary common marmoset
T lymphocytesPrimary human T
lymphocytes
None 0/9HVS C488 9/9 3/5 2/2 2/2 19, 20HVS DSTP-SV40-SEAP 0/9 0/5 2/2 0/2 .370HVS STP-C488 P103R 9/9 0/5 2/2 2/2 20
a Virus was recovered from PBMCs of two infected marmosets as described previously (14).
VOL. 73, 1999 CELLULAR TRAFs IN T-CELL TRANSFORMATION BY HVS STP 3917
on Decem
ber 3, 2014 by guesthttp://jvi.asm
.org/D
ownloaded from
mediated signaling is important for STP-C488-induced trans-formation and NF-kB activation in Rat-1 cells.
DISCUSSION
In this report we demonstrate that STP of HVS subgroups Aand C can associate with TRAFs. Furthermore, STP-C488activates NF-kB and this activation is dependent on the sameSTP-C488 residues that are required for TRAF association.Moreover, an HVS STP-C488 P103R mutation that codes foran STP-C488 mutant, which does not interact with TRAF,cannot transform primary human T lymphocytes or Rat-1 cellsin vitro. Thus, TRAF interaction is implicated in HVS STP-C488 transforming activity. However, HVS STP-C488 P103Ris able to transform marmoset T lymphocytes and induce ma-lignant lymphoma in marmosets. TRAF binding is thereforeonly one component of STP-C488 transforming activity.
The observation that STP interaction with TRAF is requiredfor human T-lymphocyte and Rat-1-cell transformation but notfor marmoset T-lymphocyte transformation indicates thatTRAF interaction is important for transformation in restrictedgenetic or developmental backgrounds. This is not surprising,since HVS has a number of genes which may contribute to cellgrowth transformation, including those encoding tip, v-cyclin,vIL-17, orf14 superantigen homolog, vCD59, vbcl-2, vFLIP,and vIL-8 receptor, as well as the rest of the STP open readingframe (1, 29, 42, 44, 50, 52, 53). Since many of these genes arelikely expressed in transformed marmoset T lymphocytes (18),STP-TRAF-mediated signaling may be less important in thiscell type. In contrast, HVS is tightly latent in transformedhuman T lymphocytes, where the single bicistronic STP and tiptranscript has been detected (18). Rat-1-cell transformationrequires only STP-C488. Thus, HVS may be considerably lessdependent on STP for marmoset lymphocyte transformationbecause of other viral genes expressed in these cells. In addi-tion, STP-C488 binding to cellular Ras is also important forRat-1 cell transformation (25), and the role of Ras interactionin marmoset and human lymphocyte transformation remainsto be investigated.
HVS strains are classified into three subgroups (subgroupsA, B, and C) based on sequence divergence in the STP and tipgenes (5, 36). Subgroups A and C are highly oncogenic and areable to immortalize common marmoset T lymphocytes, induc-
ing transformation to growth in an IL-2-independent mannerin vitro, while subgroup B does not have these properties (13,37). Subgroup C strains are in addition capable of immortal-izing human, rabbit, and rhesus monkey lymphocytes into con-tinuously proliferating T-cell lines (1, 3). All STP-A and STP-Cisolates which have been sequenced so far appear to be similararound the TRAF-binding sites that have been revealed by thebiochemical and genetic analyses described here (18, 34).These experiments show that oncogenes from the transforminggamma herpesviruses EBV and HVS employ TRAF as a com-mon cellular factor for their activities. Previous experimentswith EBV LMP1 point up the importance of both aggregationand TRAF binding in signaling (24, 38). STP-C488 can alsooligomerize through its collagen-like motif (unpublished re-sults). Thus, the HVS STP-C488 and EBV LMP1 may mimic aligand-independent constitutively active receptor.
While STPs of HVS subgroups A and C bind to TRAF 1, 2,and 3, only STP-C488 interaction with TRAF appears to acti-vate NF-kB activity. This may explain why STP-C488 is con-siderably more potent in Rat-1-cell transformation than STP-A11 (30). On the other hand, STP-A11 may use TRAFinteraction for the activation of downstream effectors otherthan NF-kB, e.g., activation of stress-activated protein kinaseor inhibitors of apoptosis (41, 51, 54). Further studies areneeded to elucidate the detailed mechanisms of HVS STP-TRAF-mediated signal transduction.
ACKNOWLEDGMENTS
We thank R. Desrosiers, K. Williams, L. Alexander, and R. Meansfor critical reading of the manuscript and G. Mosialos for providingreagents. We also thank Kristen Toohey for photography support.
This work was supported by U.S. Public Health Service grantsCA31363 and RR00168.
REFERENCES
1. Ahuja, S. K., and P. M. Murphy. 1996. Molecular piracy of mammalianinterleukin-8 receptor type B by herpesvirus saimiri. J. Biol. Chem. 268:20691–20694.
2. Alexander, L., Z. Du, M. Rosenzweig, J. U. Jung, and R. C. Desrosiers. 1997.A role for natural simian immunodeficiency virus and human immunodefi-ciency virus type 1 Nef alleles in lymphocyte activation. J. Virol. 71:6094–6099.
3. Beutler, B., and A. Cerami. 1988. Tumor necrosis, cachexia, shock andinflammation: a common mediator. Annu. Rev. Biochem. 57:505–518.
FIG. 6. Inhibition of transformation of STP-C488 by the dominant-negative TRAF2 mutant. Rat-1 fibroblast cells were plated at 106 cells per 100-mm-diametertissue culture dishes. After 14 days’ incubation, cells were photographed to show foci of transformed cells at magnification of 3100 (bottom) and after staining withmethylene blue (top). A, Rat-babe; B, Rat-TRAF2 D6–86; C, Rat-STP-C488; and D, Rat-STP-C488/TRAF2 D6–86.
3918 LEE ET AL. J. VIROL.
on Decem
ber 3, 2014 by guesthttp://jvi.asm
.org/D
ownloaded from

4. Biesinger, B., I. Muller-Fleckenstein, B. Simmer, G. Lang, S. Wittmann, E.Platzer, R. C. Desrosiers, and B. Fleckenstein. 1992. Stable growth trans-formation of human T lymphocytes by herpesvirus saimiri. Proc. Natl. Acad.Sci. USA 89:3116–3119.
5. Biesinger, B., J. J. Trimble, R. C. Desrosiers, and B. Fleckenstein. 1990. Thedivergence between two oncogenic herpesvirus saimiri strains in a genomicregion related to the transforming phenotype. Virology 176:505–514.
6. Broker, B. M., A. Y. Tsygankov, I. Muller-Fleckenstein, A. H. Guse, N. A.Chitaev, B. Biesinger, B. Fleckenstein, and F. Emmrich. 1993. Immortalizationof human T cell clones by Herpesvirus saimiri. J. Immunol. 151:1184–1192.
7. Cheng, G., and D. Baltimore. 1996. TANK, a co-inducer with TRAF2 ofTNF- and CD40L-mediated NF-kB activation. Genes Dev. 10:963–973.
8. Cheng, G., A. M. Cleary, Z.-S. Ye, D. I. Hong, S. Lederman, and D. Balti-more. 1995. Involvement of CRAF1, a relative of TRAF, in CD40 signaling.Science 267:1494–1497.
9. Desrosiers, R. C., A. Bakker, J. Kamine, L. A. Falk, R. D. Hunt, and N. W.King. 1985. A region of the herpesvirus saimiri genome required for onco-genicity. Science 228:184–187.
10. Desrosiers, R. C., R. L. Burghoff, A. Bakker, and J. Kamine. 1984. Construc-tion of replication-competent herpesvirus saimiri deletion mutants. J. Virol.49:343–348.
11. Desrosiers, R. C., and L. A. Falk. 1982. Herpesvirus saimiri strain variability.J. Virol. 43:352–356.
12. Desrosiers, R. C., D. Silva, L. M. Waldron, and N. L. Letvin. 1986. Nonon-cogenic deletion mutants of herpesvirus saimiri are defective for in vitroimmortalization. J. Virol. 57:701–705.
13. Devergne, O., E. C. McFarland, G. Mosialos, K. M. Izumi, C. F. Ware, andE. Kieff. 1998. Role of the TRAF binding site and NF-kB activation inEpstein-Barr virus latent membrane protein 1-induced cell gene expression.J. Virol. 72:7900–7908.
14. Duboise, S. M., J. Guo, S. Czajak, R. C. Desrosiers, and J. U. Jung. 1998.STP and tip are essential for herpesvirus saimiri oncogenicity. J. Virol.72:1308–1313.
15. Duboise, S. M., J. Guo, R. C. Desrosiers, and J. U. Jung. 1996. Use of virionDNA as a cloning vector for the construction of mutant and recombinantherpesviruses. Proc. Natl. Acad. Sci. USA 93:11389–11394.
16. Du, Z., D. A. Regier, and R. C. Desrosiers. 1995. Improved recombinant PCRmutagenesis procedure that uses alkaline-denatured plasmid template. Bio-Techniques 18:376–378.
17. Falk, L., L. Wolfe, and F. Deinhardt. 1972. Isolation of herpesvirus saimirifrom blood of squirrel monkeys (Saimiri sciureus). J. Natl. Cancer Inst.48:1499–1505.
18. Fickenscher, H., B. Biesinger, A. Knappe, S. Wittmann, and B. Fleckenstein.1996. Regulation of the herpesvirus saimiri oncogene stpC, similar to that ofT-cell activation genes, in growth-transformed human T lymphocytes. J. Vi-rol. 70:6012–6019.
19. Fleckenstein, B., and R. C. Desrosiers. 1982. Herpesvirus saimiri and her-pesvirus ateles, p. 253–332. In B. Roizman (ed.), The herpesviruses, vol. 1.Plenum Publishing Corporation, New York, N.Y.
20. Franken, M., O. Devergne, M. Rosenzweig, B. Annis, E. Kieff, and F. Wang.1996. Comparative analysis identifies conserved tumor necrosis factor recep-tor-associated factor 3 binding sites in the human and simian Epstein-Barrvirus oncogene LMP1. J. Virol. 70:7819–7826.
21. Gedrich, R. W., M. C. Gilfillan, C. S. Duckett, J. L. Van Dongen, and C. B.Thompson. 1996. CD30 contains two binding sites with different specificitiesfor members of the tumor necrosis factor receptor-associated factor family ofsignal transducing proteins. J. Biol. Chem. 271:12852–12858.
22. Goeddel, D. V., B. B. Aggarwal, P. W. Gray, D. W. Leung, G. E. Nedwin, G. E.Palladino, J. S. Patton, D. Pennica, H. M. Shepard, B. J. Sugarman, andG. H. W. Wong. 1986. Tumor necrosis factors: gene structure and biologicalactivities. Cold Spring Harbor Symp. Quant. Biol. 51:597–609.
23. Hu, H. M., K. O’Rourke, M. S. Boguski, and V. M. Dixit. 1994. A novelRING finger protein interacts with the cytoplasmic domain of CD40. J. Biol.Chem. 269:30069–30076.
24. Izumi, K. M., K. M. Kaye, and E. D. Kieff. 1997. The Epstein-Barr virusLMP1 amino acid sequence that engages tumor necrosis factor receptorassociated factors is critical for primary B lymphocyte growth transforma-tion. Proc. Natl. Acad. Sci. USA 94:1447–1452.
25. Jung, J. U., and R. C. Desrosiers. 1995. Association of the viral oncoproteinSTP-C488 with cellular ras. Mol. Cell. Biol. 15:6506–6512.
26. Jung, J. U., and R. C. Desrosiers. 1994. Distinct functional domains ofSTP-C488 of herpesvirus saimiri. Virology 204:751–758.
27. Jung, J. U., and R. C. Desrosiers. 1994. Herpesvirus saimiri and ateles, p.614–622. In R. Webster and A. Granoff (ed.), Encyclopedia of virology.Saunders Scientific Publications, Inc., Philadelphia, Pa.
28. Jung, J. U., and R. C. Desrosiers. 1991. Identification and characterization ofthe herpesvirus saimiri oncoprotein STP-C488. J. Virol. 65:6953–6960.
29. Jung, J. U., M. Stager, and R. C. Desrosiers. 1994. Virus-encoded cyclin.Mol. Cell. Biol. 14:7235–7244.
30. Jung, J. U., J. J. Trimble, N. W. King, B. Biesinger, B. W. Fleckenstein, andR. C. Desrosiers. 1991. Identification of transforming genes of subgroup A
and C strains of herpesvirus saimiri. Proc. Natl. Acad. Sci. USA 88:7051–7055.31. Kaye, K. M., O. Devergne, J. N. Harada, K. M. Izumi, R. Yalamanchili, E.
Kieff, and G. Mosialos. 1996. Tumor necrosis factor receptor associatedfactor 2 is a mediator of NF-kB activation by latent infection membraneprotein 1, the Epstein-Barr virus transforming protein. Proc. Natl. Acad. Sci.USA 93:11085–11090.
32. Koomey, J. M., C. Mulder, R. L. Burghoff, B. Fleckenstein, and R. C.Desrosiers. 1984. Deletion of DNA sequences in a nononcogenic variant ofherpesvirus saimiri. J. Virol. 50:662–665.
33. Kretschmer, C., C. Murphy, B. Biesinger, J. Beckers, H. Fickenscher, T. Kirch-ner, B. Fleckenstein, and U. Ruther. 1996. A Herpes saimiri oncogene causingperipheral T-cell lymphoma in transgenic mice. Oncogene 12:1609–1616.
34. Lee, H., J. J. Trimble, D.-W. Yoon, D. Regier, R. C. Desrosiers, and J. U.Jung. 1997. Genetic variation of herpesvirus saimiri subgroup A transform-ing protein and its association with cellular src. J. Virol. 71:3817–3825.
35. Lee, S. Y., A. Reichlin, A. Santana, K. A. Sokol, M. C. Nussenzweig, and Y.Choi. 1997. TRAF2 is essential for JNK but not NF-kappaB activation andregulates lymphocyte proliferation and survival. Immunity 7:703–713.
36. Medveczky, P., E. Szomolayi, R. C. Desrosiers, and C. Mulder. 1984. Clas-sification of herpesvirus saimiri into three groups based on extreme variationin a DNA region required for oncogenicity. J. Virol. 52:938–944.
37. Mittrucker, H.-W., I. Muller-Fleckenstein, B. Fleckenstein, and B. Fleish-cher. 1995. CD2-mediated autocrine growth of herpes virus saimiri-trans-formed human T lymphocytes. J. Exp. Med. 176:900–913.
38. Mosialos, G., M. Birkenbach, R. Yalamanchili, T. VanArsdale, C. Ware, andE. Kieff. 1995. The Epstein-Barr virus transforming protein LMP1 engagessignaling proteins for the tumor necrosis factor receptor family. Cell 80:389–399.
39. Murphy, C., C. Kretschmer, B. Biesinger, J. Beckers, J. Jung, R. C. Desro-siers, H. K. Muller-Hermelink, B. W. Fleckenstein, and U. Ruther. 1994.Epithelial tumors induced by a herpesvirus oncogene in transgenic mice.Oncogene 9:221–226.
40. Murthy, S. C. S., J. J. Trimble, and R. C. Desrosiers. 1989. Deletion mutantsof herpesvirus saimiri define an open reading frame necessary for transfor-mation. J. Virol. 63:3307–3314.
41. Natoli, G., A. Costanzo, A. Ianni, D. J. Templeton, J. R. Woodgett, C.Balsano, and M. Levrero. 1997. Activation of SAPK/JNK by TNF receptor1 through a noncytotoxic TRAF2-dependent pathway. Science 10:200–203.
42. Nava, V. E., E. H.-Y. Cheng, M. Veliuona, S. Zou, R. J. Clem, M. L. Mayer,and J. M. Hardwick. 1997. Herpesvirus saimiri encodes a functional homologof the human bcl-2 oncogene. J. Virol. 71:4118–4122.
43. Rothe, M., S. C. Wong, W. J. Henzel, and D. V. Goeddel. 1994. A novel familyof putative signal transducers associated with the cytoplasmic domain of the75 kDa tumor necrosis factor receptor. Cell 78:681–692.
44. Rother, R. P., S. A. Rollins, W. L. Fodor, J.-C. Albrecht, E. Setter, B.Fleckenstein, and S. P. Squinto. 1994. Inhibition of complement-mediatedcytolysis by the terminal complement inhibitor of herpesvirus saimiri. J. Vi-rol. 68:730–737.
45. Smith, C. A., T. Farrah, and R. G. Goodwin. 1994. The TNF receptorsuperfamily of cellular and viral proteins: activation, costimulation, anddeath. Cell 76:959–962.
46. Song, H. Y., C. H. Regnier, C. J. Kirschning, D. V. Goeddel, and M. Rothe.1997. Tumor necrosis factor (TNF)-mediated kinase cascades: bifurcation ofnuclear factor-kappaB and c-jun N-terminal kinase (JNK/SAPK) pathways atTNF receptor-associated factor 2. Proc. Natl. Acad. Sci. USA 94:9792–9796.
47. Szomolanyi, E., P. Medveczky, and C. Mulder. 1987. In vitro immortalizationof marmoset cells with three subgroups of herpesvirus saimiri. J. Virol.61:3485–3490.
48. Takeuchi, M., M. Rothe, and D. V. Goeddel. 1996. Anatomy of TRAF2.J. Biol. Chem. 271:19935–19942.
49. Tartaglia, L. A., and D. V. Goeddel. 1992. Two TNF receptors. Immunol.Today 13:151–153.
50. Thome, M., P. Schneider, K. Hofmann, H. Fickenscher, E. Meinl, F. Neipel, C.Mattmann, K. Burns, J.-L. Bodmer, M. Schroter, C. Scaffidi, P. H. Krammer,M. E. Peter, and J. Tschopp. 1997. Viral FLICE-inhibitory proteins (FLIPs)prevent apoptosis induced by death receptors. Nature 386:517–521.
51. Wang, C. Y., M. W. Mayo, R. G. Korneluk, D. V. Goeddel, and A. S. J. Baldwin.1998. NF-kappaB antiapoptosis: induction of TRAF1 and TRAF2 and c-IAP1and c-IAP2 to suppress caspase-8 activation. Science 281:1680–1683.
52. Yao, Z., W. C. Fanslow, M. F. Seldin, A. M. Rousseau, S. L. Painter, M. R.Comeau, J. I. Cohen, and M. K. Spriggs. 1995. Herpesvirus saimiri encodesa new cytokine IL-17, which binds to a novel cytokine receptor. Immunity3:811–821.
53. Yao, Z., E. Maraskovsky, M. K. Spriggs, J. I. Cohen, R. J. Armitage, andM. R. Alderson. 1996. Herpesvirus saimiri open reading frame 14, a proteinencoded by a T lymphotropic herpesvirus, binds to MHC class II moleculesand stimulates T cell proliferation. J. Immunol. 156:3260–3266.
54. Yuasa, T., S. Ohno, J. H. Kehrl, and J. M. Kyriakis. 1998. Tumor necrosisfactor signaling to stress-activated protein kinase (SAPK)/Jun NH2-terminalkinase (JNK) and p38. Germinal center kinase couples TRAF2 to mitogen-activated protein kinase/ERK kinase kinase 1 SAPK while receptor inter-acting protein associates with a mitogen-activated protein kinase kinasekinase upstream of MKK6 and p38. J. Biol. Chem. 273:22681.
VOL. 73, 1999 CELLULAR TRAFs IN T-CELL TRANSFORMATION BY HVS STP 3919
on Decem
ber 3, 2014 by guesthttp://jvi.asm
.org/D
ownloaded from





























