Embed Size (px)
Citation preview

of July 5, 2022.This information is current as
High-Affinity B CellsCells: Implications for Selection of
RIIB on Germinal CenterγFc Receptor FcDifferential Expression of the Inhibitory IgG
Sambasiva P. Rao, Kalpit A. Vora and Tim Manser
http://www.jimmunol.org/content/169/4/1859doi: 10.4049/jimmunol.169.4.1859
2002; 169:1859-1868; ;J Immunol
Referenceshttp://www.jimmunol.org/content/169/4/1859.full#ref-list-1
, 13 of which you can access for free at: cites 42 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Errata
/content/174/8/5133.1.full.pdfor:
next pageAn erratum has been published regarding this article. Please see
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2002 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on July 5, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on July 5, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on July 5, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on July 5, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on July 5, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on July 5, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

Differential Expression of the Inhibitory IgG Fc ReceptorFc�RIIB on Germinal Center Cells: Implications for Selectionof High-Affinity B Cells1
Sambasiva P. Rao,2 Kalpit A. Vora,3 and Tim Manser4
The murine low-affinity receptor for IgG, Fc�RIIB, mediates inhibition of B cell receptor-triggered events in primary B cells. Weinvestigated the expression of Fc�RIIB on germinal center (GC) cells to better understand its role in memory B cell development.Immunohistological analyses demonstrated differential regulation of Fc�RIIB on GC cells. Its levels are markedly down-regulatedon GC B cells and up-regulated on follicular dendritic cells (FDC) at all times during the GC response. Analyses of surfaceexpression of Fc�RIIB by flow cytometry and Fc�RIIB mRNA levels by RT-PCR analysis confirmed that this FcR is down-regulated in GC B cells. In mice lacking Fc�RIIB, the development of the secondary FDC reticulum in GCs is substantiallydelayed, although the overall kinetics of the GC response are unaltered. These findings have direct implications for modelsproposed to account for the selection of high-affinity B cells in the GC and suggest a role for Fc�RIIB in promoting the maturationof the FDC reticulum. The Journal of Immunology, 2002, 169: 1859–1868.
R eceptors for the Fc portion of IgG (Fc�R) are widelydistributed on hemopoietic cells and play an importantrole in immune regulation by linking Ab-mediated re-
sponses with effector functions. The murine high-affinity Fc�RIand the low-affinity Fc�RII and Fc�RIII (1) are distinguished bytheir structure, function, distribution, and affinities for IgG (1, 2).These Fc�R contain homologous extracellular domains; however,they vary considerably in their transmembrane and cytoplasmicregions. Fc�RI and Fc�RIII are associated with a common�-chain, which contains the immunoreceptor tyrosine-based acti-vation motif in its cytoplasmic tail. In contrast, Fc�RII contains inits cytoplasmic tail an immunoreceptor tyrosine-based inhibitorymotif (3). Fc�RIIB is encoded by a single gene in the mouse.However, alternative mRNA splicing results in two Fc�RII iso-forms, Fc�RIIb1 and Fc�RIIb2 (4, 5). Murine B cells exclusivelyexpress Fc�RIIb1 and lack expression of the activation Fc�Rs (6).Coaggregation of the B cell Ag receptor (BCR)5 and Fc�RIIb1 onB cells results in the inhibition of Ag internalization, activation,proliferation, and Ab secretion (7–10). Previous in vitro studieshave also shown that Fc�RIIB is capable of independently gener-ating an apoptotic signal upon clustering (10–12).
Germinal centers (GC) are inducible lymphoid microenviron-ments that arise primarily in response to T-dependent Ags. Duringthe GC response, a high frequency of mutations is introduced inthe variable regions of the BCRs expressed by GC B cells. Agpresent in the form of immune complexes (IC) retained on follic-ular dendritic cells (FDC) within the GC is thought to drive theselection of high-affinity B cell mutants, ultimately resulting in theformation of the memory B cell compartment. During this processof interaction between ICs and GC B cells, it is believed that Bcells bearing high-affinity BCR are preferentially stimulated, re-sulting in the uptake of Ag and its subsequent presentation to Thlymphocytes (13–15). This model considers only the role of BCRbut ignores the role of Fc�RIIB in B cell selection upon interactionwith ICs. Given the general inhibitory role of Fc�RIIB, it is un-clear how GC B cells, on interaction with ICs, could be activatedand internalize the Ag for presentation to Ag-specific Th cells.
In this context, it has been suggested that the consequence ofcoengagement of BCR and Fc�RIIB by ICs retained on FDC inGC is dependent on the balance between concurrent activation andinhibitory signals, leading to stimulatory, inhibitory, or apoptoticresponses (10). Fc�RIIB expression is also induced on FDC withinthe GC, and it has been proposed that this high level expressionallows FDCs to convert ICs to a highly immunogenic form for Bcell activation (16). Interestingly, none of the studies leading tothese models has examined the expression levels of Fc�RIIB onGC B cells. Recent studies have indicated that the surface expres-sion of Fc�RIIB is reduced on GC B cells in autoimmune strainsof mice (17, 18). In contrast, in vitro experiments have indicatedthat activated B cell blasts differ from resting B cells in that theyexpress significantly elevated levels of Fc�RIIB, as well as itsdownstream signaling molecules Src homology domain 2-contain-ing inositol 5-phosphatase and Src homology domain 2-containinginositol 5-phosphatase-2 (19). These data highlight the importanceof elucidating the role of Fc�RIIB in memory B cell developmentand the GC reaction.
In the present study, we examined whether expression ofFc�RIIB is modulated in GC B cells during a T-dependent im-mune response and have characterized the GC response in micelacking this receptor. Our results indicate that both Fc�RIIB
Kimmel Cancer Center and Department of Microbiology and Immunology, ThomasJefferson University, Philadelphia, PA 19107
Received for publication February 22, 2002. Accepted for publication June 5, 2002.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by National Institutes of Health Grant AI 46806 (to T.M.)and by National Institutes of Health Training Grant 5-T32-AI07492 (to S.P.R.).2 Current address: Department of Immunology and Inflammation, Biogen, Cam-bridge, MA 02142.3 Current address: Merck & Co., 126 East Lincoln Avenue, Rahway, NJ 07065.4 Address correspondence and reprint requests to Dr. Tim Manser, Kimmel CancerInstitute, Bluemle Life Sciences Building, 708, 233 South 10th Street, Philadelphia,PA 19107. E-mail address: [email protected] Abbreviations used in this paper: BCR, B cell Ag receptor; GC, germinal center;FDC, follicular dendritic cell; IC, immune complex; NP, 4-hydroxy-3-nitrophenylacetyl; CGG, chicken �-globulin; PNA, peanut agglutinin; AFC, Ab-forming cell;SA, streptavidin.
The Journal of Immunology
Copyright © 2002 by The American Association of Immunologists, Inc. 0022-1767/02/$02.00
by guest on July 5, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

mRNA and protein expression are significantly down-regulated inGC B cells as compared with non-GC B cells. In addition, whereasthe kinetics and magnitude of the GC response appear unaltered inFc�RIIB-deficient mice, the development of FDC reticula in GCsis delayed, supporting a role for this receptor in FDC maturation.The implications of these results are discussed in the context ofmodels proposed to account for selection of high-affinity B cells inthe GC.
Materials and MethodsMice and immunizations
Fc�RIIB�/� mice on a C57BL/6 background (20) and C57BL/6 mice werepurchased from Taconic Farms (Germantown, NY). The generation andcharacterization of common �-chain-deficient mice have been describedpreviously (21). These mice, obtained from Dr. J. Ravetch (RockefellerUniversity, New York, NY), had been backcrossed to C57BL/6 mice foreight generations. All mice were maintained in pathogen-free conditions.Mice (10–12 wk old) were immunized i.p. with 3 � 108 SRBC (LampireBiological Labs, Pipersville, PA) in PBS or 100 �g 4-hydroxy-3-nitrophe-nyl acetyl-chicken �-globulin (NP13-CGG) in alum and were sacrificed atdifferent time intervals for analysis.
Antibodies
The following Abs were used for immunohistology or flow cytometricanalysis: PE-Texas Red-conjugated anti-B220 (clone RA3-6B2; CaltagLaboratories, Burlingame, CA); FITC-labeled GL7; PE-labeled anti-Fc�RII/Fc�RIII (clone 2.4G2); PE-labeled anti-B220 (clone RA3-6B2);biotin-labeled anti-CD35 (clone 8C12) (all purchased from BD PharMin-gen, San Diego, CA); unconjugated rat IgG Ab to mouse FDCs (FDC-M1and FDC-M2, gifts from Dr. M. Kosco-Vilbois, Serono PharmaceuticalResearch Institute, Plan-les-Quates, Switzerland); FITC-labeled MOMA-2(Serotec, Oxford, U.K.); biotin-labeled (Fab�)2 mouse anti-rat IgG (Jack-son ImmunoResearch Laboratories, West Grove, PA); biotin-labeled pea-nut agglutinin (PNA; Sigma-Aldrich, St. Louis, MO); PE or biotin-labeledanti-IgD (clone 11-26; Southern Biotechnology Associates, Birmingham,AL) and streptavidin (SA)-PE (Molecular Probes, Eugene, OR). TheK9.361 hybridoma was a kind gift from Dr. U. Hammerling (Sloan Ket-tering Memorial Hospital, New York, NY).
Histology
Spleens from immunized mice were obtained at various times after immu-nization and processed for histological analysis as previously described(22). Briefly, spleens were embedded in Tissue-Tek OCT compound (Fish-er Scientific, Bridgewater, NJ) by freezing in a 2-methylbutane bath cooledwith liquid nitrogen. Frozen spleens were stored at �70°C until sectioned.Sections of 6 �m were cut on a cryostat microtome and mounted onto0.01% poly-L-lysine (Sigma-Aldrich)-coated slides. The sections were airdried for 1 h, fixed in acetone for 15 min, air dried again for at least 2 h,and stored at �70°C until further analysis.
Frozen sections were rehydrated in TBS for 20 min followed by block-ing in TBS, 5% BSA, 0.05% Tween 20 for another 20 min. Double im-munohistology was then performed by staining with fluorescent Abs de-scribed above for 30 min. For unconjugated Abs, as a secondary step,sections were stained for 30 min with biotinylated mouse anti-rat Ab. Theslides were then washed three times in TBS, 5% BSA, 0.05% Tween 20,and the biotinylated Abs were revealed by SA-PE. The slides were washedand stored in PBS in the dark. The stained sections were analyzed using afluorescence microscope (Leitz Diaplan, Leitz, Wetzlar, West Germany),and digital images were captured using a Kodak DC290 Zoom digital cam-era and MDS-290 software (Eastman Kodak, Rochester, NY).
Flow cytometry and cell sorting
Cell suspensions prepared from spleens excised from mice on day 8 post-SRBC immunization were depleted of erythrocytes by ammonium chlorideTris lysis (23). For four-color surface staining, 2 � 106 cells resuspendedin PBS containing 2% BSA were incubated with pretitered dilutions ofGL7-FITC, biotinylated anti-IgD, anti-B220-PET-Texas Red, and2.4G2-PE for 30 min at 4°C. SA Red 670 was used as a second-stepreagent. Cells were analyzed using a Coulter Epics XL/MCL analyzer, andthe data were analyzed by WinMDI software (The Scripps Research In-stitute, La Jolla, CA). For sorting, spleen cells prepared from C57BL/6 orFc� chain�/� mice on day 8 post-SRBC immunization were stained withGL7-FITC, anti-B220-PE and anti-IgD-biotin followed by SA Red 670.
GC and non-GC B cells were separated on a Coulter Epics Elite flowcytometric cell sorter.
RT-PCR assay
mRNA was extracted from sorted GC and non-GC B cells using theRNeasy Miniprep kit (Qiagen, Valencia, CA) according to the manufac-turer’s recommendation. Using 0.2 or 0.5 �g of RNA from each sample,cDNA synthesis and Fc�RIIB-specific PCR amplification was performedin a single step using the Superscript one-step RT-PCR system (Life Tech-nologies, Gaithersburg, MD). Fc�RIIB cDNA was amplified using the 5�primer 5�-AAGTCTAGGAAGGACACTGC-3� and the 3� primer 5�-ATCCTGGCCTTCTGGCTTGC-3�. As an internal control, �-actin mRNA wasamplified using primers purchased from Clontech Laboratories (Palo Alto,CA). RT-PCR was conducted in 50-�l volumes with one cycle pro-grammed to perform cDNA synthesis at 50°C for 30 min and 94°C for 2min, followed immediately by PCR amplification for 30 cycles at 95°C for1 min, 55°C for 45 s, and 72°C for 1 min. A final extension was done at72°C for 5 min. One-fifth volume of the PCR was electrophoresed on a1.5% agarose gel, and the bands were visualized under UV light.
Gel hybridization
Agarose gels were dried at 65°C for 2 h under vacuum, denatured in 0.5 MNaOH, 1.5 M NaCl for 1 h, and neutralized for 1 h in 0.5 M Tris, 3 MNaCl. The gels were then prehybridized at 42°C in prehybridization buffersolution containing 6� 0.6 M NaCl, 0.15 M Tris (pH 8.0), 6 mM EDTA;0.05 M Denhardt’s solution; 0.1% SDS; and 250 �g of herring spermDNA. Probes were prepared from PCR products by 32P labeling using arandom primed DNA labeling kit (Roche Molecular Biochemicals, Mann-heim, Germany), denatured, and added to the prehybridization buffer, andthe gels were hybridized overnight at 42°C. The gels were washed at lowstringency in 5� SSC, 5% SDS for 1 h followed by two 30-min washes at43°C. Subsequently, a high stringency wash in 0.1� SSC, 0.1% SDS at60°C was performed. The gels were scanned on a PhosphorImager (Mo-lecular Dynamics, Sunnyvale, CA) after overnight exposure. The bandswere quantitated after subtracting the background signal using ImageQuant software (Molecular Dynamics). The integrated intensity of eachband representing individual PCR products was measured and normalizedusing the �-actin band intensity for each individual sample.
ResultsKinetics of the GC response to SRBC immunization
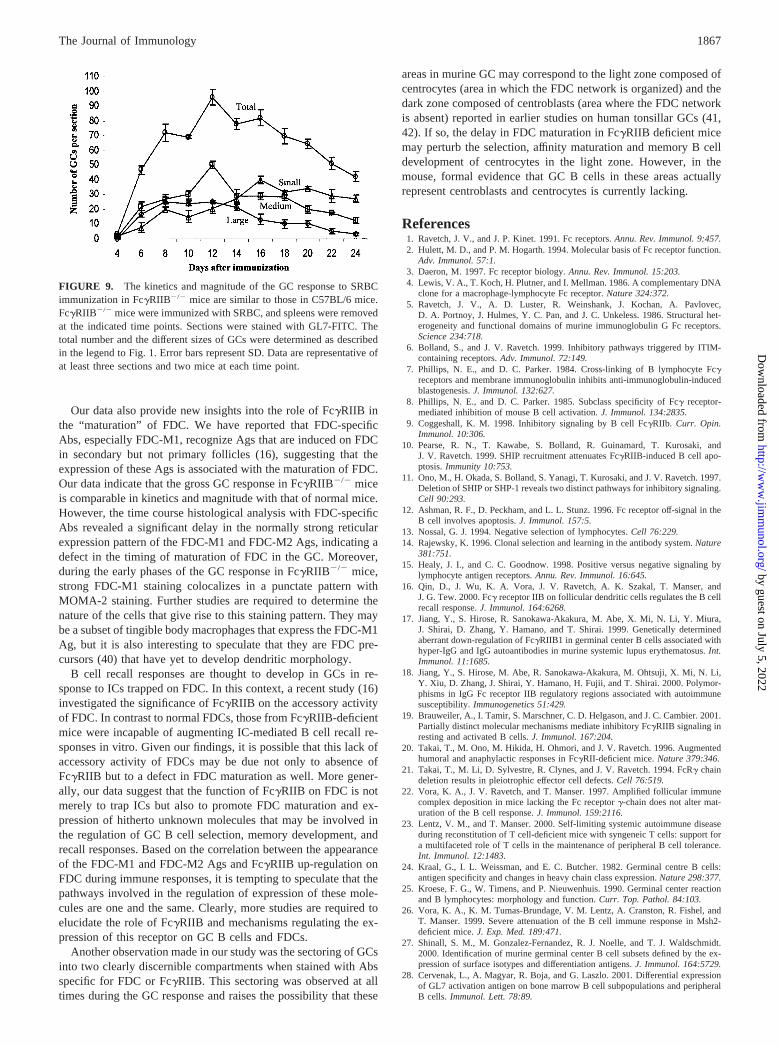
SRBC have been previously shown to induce robust GC responsesin an adjuvant-independent manner (24, 25). However, only lim-ited information was available regarding the magnitude and kinet-ics of this response. Therefore, we first examined the number andsizes of GC in the spleens of SRBC immunized C57BL/6 mice.Spleen sections obtained from days 4–40 after immunization werestained with GL7. The total number of GCs in each section werecounted and categorized into three different sizes by countingGL7� cells as previously described (26). A summary of the resultsis presented in Fig. 1. On day 4 after SRBC immunization, themajority of GL7� cells were scattered in follicles. Small foci ofGL7� cells localized within the FDC network as well as a fewwell-developed small GCs were detectable (data not shown). Byday 6, there was a large expansion in the number of GC comprisingall sizes. This large number of GCs was maintained until day 12,after which the numbers declined. Large GCs were present fromday 6, reached a peak by day 10, and thereafter declined with acorresponding increase in medium and small GCs. Although thenumber of medium sized GCs started to decline after day 12, thesmall GCs continued to increase, perhaps due to the dissolution ofsome of the large and medium sized GCs. The numbers of smallGC gradually declined from day 16 onward, and by day 40 only afew GCs were present, almost all of which were small.
GL7 and PNA staining of GC B cells
Although a strong correlation exists between the patterns of PNAand GL7 staining, earlier studies reported the presence of bothPNA�GL7� as well as PNA�GL7� cell populations in spleen (27,28). Immunohistological analysis of spleen sections from C57BL/6
1860 Fc�RIIB EXPRESSION IS DOWN-REGULATED ON GC B CELLS
by guest on July 5, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
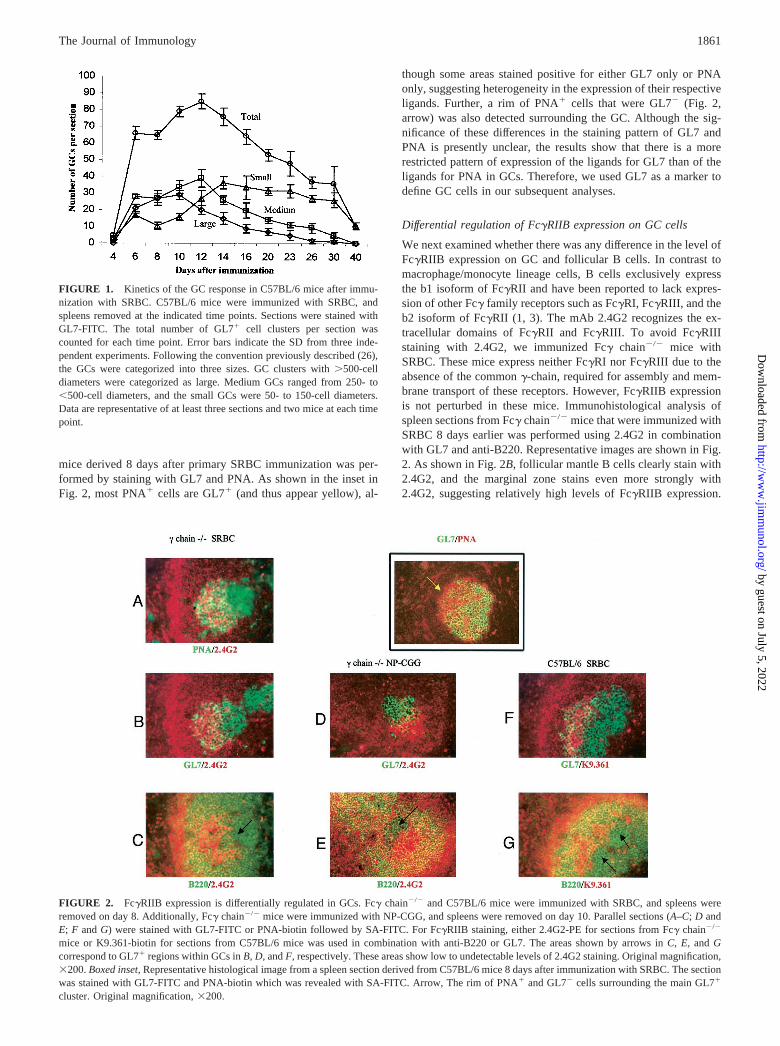
mice derived 8 days after primary SRBC immunization was per-formed by staining with GL7 and PNA. As shown in the inset inFig. 2, most PNA� cells are GL7� (and thus appear yellow), al-
though some areas stained positive for either GL7 only or PNAonly, suggesting heterogeneity in the expression of their respectiveligands. Further, a rim of PNA� cells that were GL7� (Fig. 2,arrow) was also detected surrounding the GC. Although the sig-nificance of these differences in the staining pattern of GL7 andPNA is presently unclear, the results show that there is a morerestricted pattern of expression of the ligands for GL7 than of theligands for PNA in GCs. Therefore, we used GL7 as a marker todefine GC cells in our subsequent analyses.
Differential regulation of Fc�RIIB expression on GC cells
We next examined whether there was any difference in the level ofFc�RIIB expression on GC and follicular B cells. In contrast tomacrophage/monocyte lineage cells, B cells exclusively expressthe b1 isoform of Fc�RII and have been reported to lack expres-sion of other Fc� family receptors such as Fc�RI, Fc�RIII, and theb2 isoform of Fc�RII (1, 3). The mAb 2.4G2 recognizes the ex-tracellular domains of Fc�RII and Fc�RIII. To avoid Fc�RIIIstaining with 2.4G2, we immunized Fc� chain�/� mice withSRBC. These mice express neither Fc�RI nor Fc�RIII due to theabsence of the common �-chain, required for assembly and mem-brane transport of these receptors. However, Fc�RIIB expressionis not perturbed in these mice. Immunohistological analysis ofspleen sections from Fc� chain�/� mice that were immunized withSRBC 8 days earlier was performed using 2.4G2 in combinationwith GL7 and anti-B220. Representative images are shown in Fig.2. As shown in Fig. 2B, follicular mantle B cells clearly stain with2.4G2, and the marginal zone stains even more strongly with2.4G2, suggesting relatively high levels of Fc�RIIB expression.
FIGURE 1. Kinetics of the GC response in C57BL/6 mice after immu-nization with SRBC. C57BL/6 mice were immunized with SRBC, andspleens removed at the indicated time points. Sections were stained withGL7-FITC. The total number of GL7� cell clusters per section wascounted for each time point. Error bars indicate the SD from three inde-pendent experiments. Following the convention previously described (26),the GCs were categorized into three sizes. GC clusters with �500-celldiameters were categorized as large. Medium GCs ranged from 250- to�500-cell diameters, and the small GCs were 50- to 150-cell diameters.Data are representative of at least three sections and two mice at each timepoint.
FIGURE 2. Fc�RIIB expression is differentially regulated in GCs. Fc� chain�/� and C57BL/6 mice were immunized with SRBC, and spleens wereremoved on day 8. Additionally, Fc� chain�/� mice were immunized with NP-CGG, and spleens were removed on day 10. Parallel sections (A–C; D andE; F and G) were stained with GL7-FITC or PNA-biotin followed by SA-FITC. For Fc�RIIB staining, either 2.4G2-PE for sections from Fc� chain�/�
mice or K9.361-biotin for sections from C57BL/6 mice was used in combination with anti-B220 or GL7. The areas shown by arrows in C, E, and Gcorrespond to GL7� regions within GCs in B, D, and F, respectively. These areas show low to undetectable levels of 2.4G2 staining. Original magnification,�200. Boxed inset, Representative histological image from a spleen section derived from C57BL/6 mice 8 days after immunization with SRBC. The sectionwas stained with GL7-FITC and PNA-biotin which was revealed with SA-FITC. Arrow, The rim of PNA� and GL7� cells surrounding the main GL7�
cluster. Original magnification, �200.
1861The Journal of Immunology
by guest on July 5, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
GL7� GCs revealed an interesting pattern of 2.4G2 staining. Al-though Fc�RIIB expression was undetectable on B cells towardone side of the GC, intense 2.4G2 staining was observed polarizedto the other side of the GC, suggesting up-regulated Fc�RIIB ex-pression in this area. Similar results were obtained when sectionswere stained with PNA and 2.4G2 (Fig. 2A). This sectored patternof Fc�RIIB expression was consistently observed. Parallel sectionsstained with Ab to B220 and 2.4G2 were then analyzed. As shownin Fig. 2C, B220� marginal zone cells exhibited relatively highlevels of Fc�RIIB (and thus appear orange in Fig. 2C), and theB220� follicular mantle cells also coexpressed Fc�RIIB (and thusappear yellow in Fig. 2C). In contrast, the B220� areas corre-sponding to GC exhibited sectoring, with undetectable levels ofFc�RIIB in one area (thus appearing green; Fig. 2C, arrow) andhigh levels of Fc�RIIB in the other area.
We then examined whether the observed differential regulationof Fc�RIIB was also a feature of GC responses to the well-char-acterized model Ag, NP. Day 10 spleen sections from NP13-CGG-immunized Fc� chain�/� mice were stained with 2.4G2 and GL7or anti-B220. These experiments also showed sectoring, with highlevels of Fc�RIIB in one region and low to undetectable levels inthe other region of GCs (Fig. 2, D and E). The analysis was furtherextended by using an Fc�RIIB-specific mAb, K9.361 (29–31),that was made available to us at a later time during the study.K9.361 exclusively recognizes Fc�RIIB (Ly17.2 allotype ofLy17.1/Ly17.2 alloantigen system) in C57BL/6 mice and does notbind to Fc�RIII (31). Immunohistological analysis of day 8 spleensections from SRBC-immunized C57BL/6 mice with K9.361 andGL7 or anti-B220 revealed exactly the same sectored pattern ofGC staining observed with 2.4G2 (Fig. 2, F and G). These datacollectively indicate that the expression of Fc�RIIB is differen-tially modulated in these two areas of GCs.
Fc�RIIB expression is markedly reduced on GC B cells andup-regulated on FDC
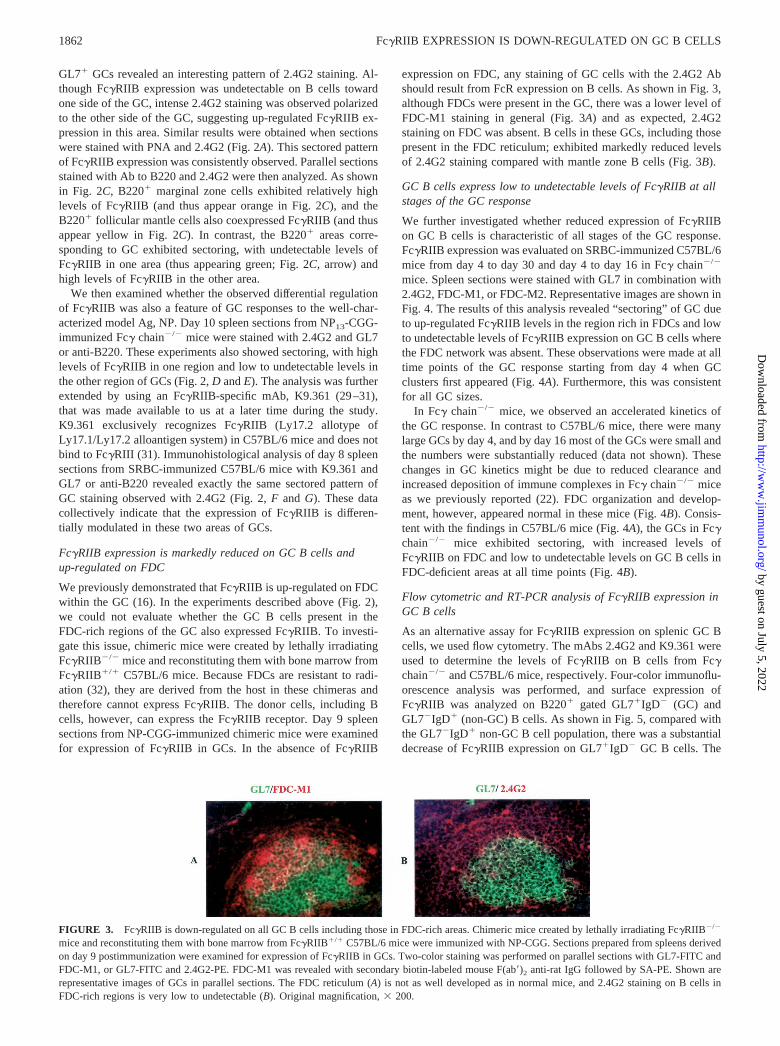
We previously demonstrated that Fc�RIIB is up-regulated on FDCwithin the GC (16). In the experiments described above (Fig. 2),we could not evaluate whether the GC B cells present in theFDC-rich regions of the GC also expressed Fc�RIIB. To investi-gate this issue, chimeric mice were created by lethally irradiatingFc�RIIB�/� mice and reconstituting them with bone marrow fromFc�RIIB�/� C57BL/6 mice. Because FDCs are resistant to radi-ation (32), they are derived from the host in these chimeras andtherefore cannot express Fc�RIIB. The donor cells, including Bcells, however, can express the Fc�RIIB receptor. Day 9 spleensections from NP-CGG-immunized chimeric mice were examinedfor expression of Fc�RIIB in GCs. In the absence of Fc�RIIB
expression on FDC, any staining of GC cells with the 2.4G2 Abshould result from FcR expression on B cells. As shown in Fig. 3,although FDCs were present in the GC, there was a lower level ofFDC-M1 staining in general (Fig. 3A) and as expected, 2.4G2staining on FDC was absent. B cells in these GCs, including thosepresent in the FDC reticulum; exhibited markedly reduced levelsof 2.4G2 staining compared with mantle zone B cells (Fig. 3B).
GC B cells express low to undetectable levels of Fc�RIIB at allstages of the GC response
We further investigated whether reduced expression of Fc�RIIBon GC B cells is characteristic of all stages of the GC response.Fc�RIIB expression was evaluated on SRBC-immunized C57BL/6mice from day 4 to day 30 and day 4 to day 16 in Fc� chain�/�
mice. Spleen sections were stained with GL7 in combination with2.4G2, FDC-M1, or FDC-M2. Representative images are shown inFig. 4. The results of this analysis revealed “sectoring” of GC dueto up-regulated Fc�RIIB levels in the region rich in FDCs and lowto undetectable levels of Fc�RIIB expression on GC B cells wherethe FDC network was absent. These observations were made at alltime points of the GC response starting from day 4 when GCclusters first appeared (Fig. 4A). Furthermore, this was consistentfor all GC sizes.
In Fc� chain�/� mice, we observed an accelerated kinetics ofthe GC response. In contrast to C57BL/6 mice, there were manylarge GCs by day 4, and by day 16 most of the GCs were small andthe numbers were substantially reduced (data not shown). Thesechanges in GC kinetics might be due to reduced clearance andincreased deposition of immune complexes in Fc� chain�/� miceas we previously reported (22). FDC organization and develop-ment, however, appeared normal in these mice (Fig. 4B). Consis-tent with the findings in C57BL/6 mice (Fig. 4A), the GCs in Fc�chain�/� mice exhibited sectoring, with increased levels ofFc�RIIB on FDC and low to undetectable levels on GC B cells inFDC-deficient areas at all time points (Fig. 4B).
Flow cytometric and RT-PCR analysis of Fc�RIIB expression inGC B cells
As an alternative assay for Fc�RIIB expression on splenic GC Bcells, we used flow cytometry. The mAbs 2.4G2 and K9.361 wereused to determine the levels of Fc�RIIB on B cells from Fc�chain�/� and C57BL/6 mice, respectively. Four-color immunoflu-orescence analysis was performed, and surface expression ofFc�RIIB was analyzed on B220� gated GL7�IgD� (GC) andGL7�IgD� (non-GC) B cells. As shown in Fig. 5, compared withthe GL7�IgD� non-GC B cell population, there was a substantialdecrease of Fc�RIIB expression on GL7�IgD� GC B cells. The
FIGURE 3. Fc�RIIB is down-regulated on all GC B cells including those in FDC-rich areas. Chimeric mice created by lethally irradiating Fc�RIIB�/�
mice and reconstituting them with bone marrow from Fc�RIIB�/� C57BL/6 mice were immunized with NP-CGG. Sections prepared from spleens derivedon day 9 postimmunization were examined for expression of Fc�RIIB in GCs. Two-color staining was performed on parallel sections with GL7-FITC andFDC-M1, or GL7-FITC and 2.4G2-PE. FDC-M1 was revealed with secondary biotin-labeled mouse F(ab�)2 anti-rat IgG followed by SA-PE. Shown arerepresentative images of GCs in parallel sections. The FDC reticulum (A) is not as well developed as in normal mice, and 2.4G2 staining on B cells inFDC-rich regions is very low to undetectable (B). Original magnification, � 200.
1862 Fc�RIIB EXPRESSION IS DOWN-REGULATED ON GC B CELLS
by guest on July 5, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
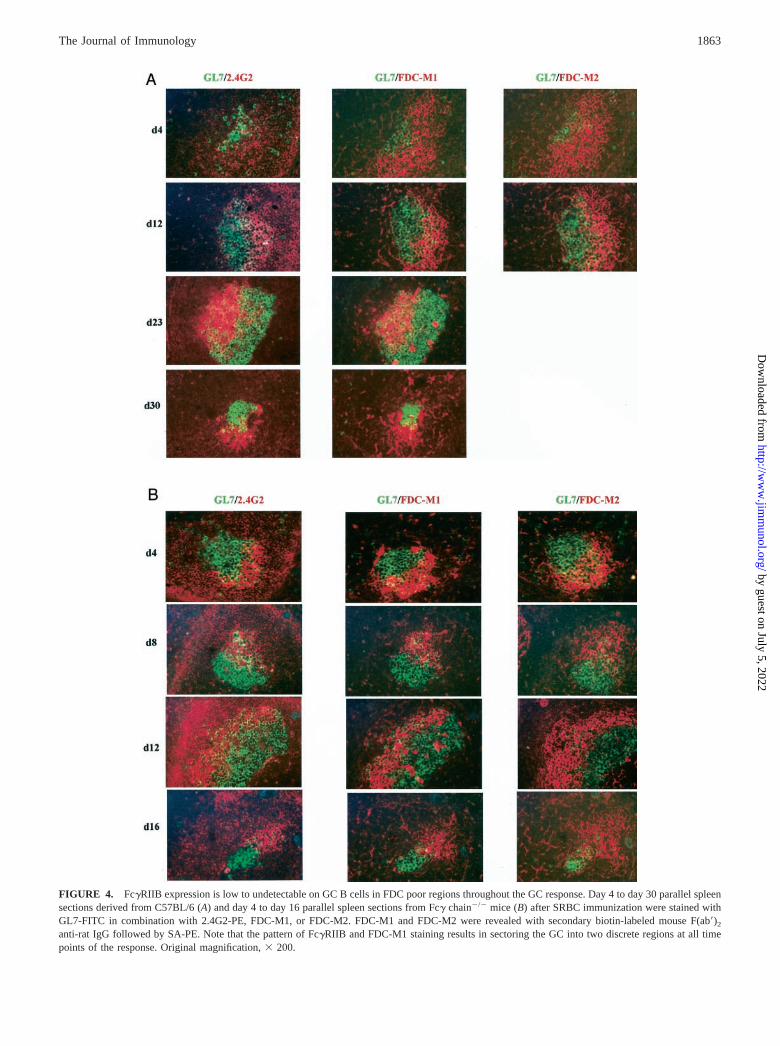
FIGURE 4. Fc�RIIB expression is low to undetectable on GC B cells in FDC poor regions throughout the GC response. Day 4 to day 30 parallel spleensections derived from C57BL/6 (A) and day 4 to day 16 parallel spleen sections from Fc� chain�/� mice (B) after SRBC immunization were stained withGL7-FITC in combination with 2.4G2-PE, FDC-M1, or FDC-M2. FDC-M1 and FDC-M2 were revealed with secondary biotin-labeled mouse F(ab�)2
anti-rat IgG followed by SA-PE. Note that the pattern of Fc�RIIB and FDC-M1 staining results in sectoring the GC into two discrete regions at all timepoints of the response. Original magnification, � 200.
1863The Journal of Immunology
by guest on July 5, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
extent of surface Fc�RIIB down-regulation on GC B cells detectedwith 2.4G2 (Fig. 5A) and K9.361 (Fig. 5B) was similar, thus con-firming our immunohistology observations (Fig. 2). It is possiblethat due to the differences in the sensitivities of flow cytometry andfluorescence microscopy techniques, low level expression ofFc�RIIB on GC B cells was not detectable in immunohistologicalanalyses. However, the unimodal distribution of Fc�RIIB stainingindicates that all GC B cells exhibit this low expression level withno evidence for the presence of any minor population of GC Bcells that express Fc�RIIB at levels similar to GL7�IgD� B cells.
To examine the mechanism and the extent of down-regulation ofFc�RIIB, RNA prepared from sorted GL7�IgD� GC cells fromimmunized C57BL/6 mice was analyzed using RT-PCR to deter-mine the levels of Fc�RIIB encoding mRNA. These levels werecompared with those in the GL7�IgD�-naive B cell population.Fc�RIIB mRNA from both cell populations was amplified usinggene-specific primers (see Materials and Methods). �-ActinmRNA served as an internal control to ensure that equal quantitiesof cDNA were amplified in the reactions. To test the linearity ofPCR amplification and the extent of down-regulation, PCR prod-ucts removed after different numbers of PCR cycles were subjectedto agarose gel electrophoresis. The gel was hybridized with anFc�RIIB-specific probe, and the relative intensity of each bandrepresenting individual PCR products was measured on a Phos-phorImager and expressed as units of integrated band intensity (seeMaterials and Methods). The results of this analysis showed nearlya 6-fold reduction of Fc�RIIB mRNA in GC B cells comparedwith non-GC IgD� B cells (Fig. 6).
FDC exhibit maturational defects in Fc�RIIB-deficient mice
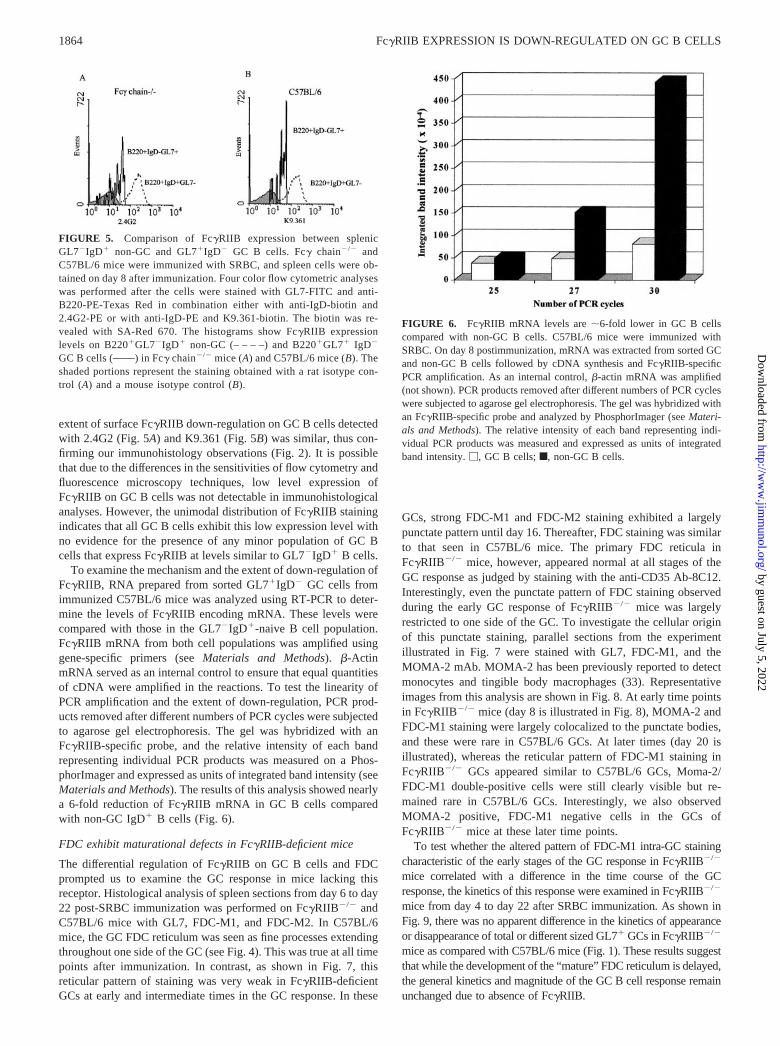
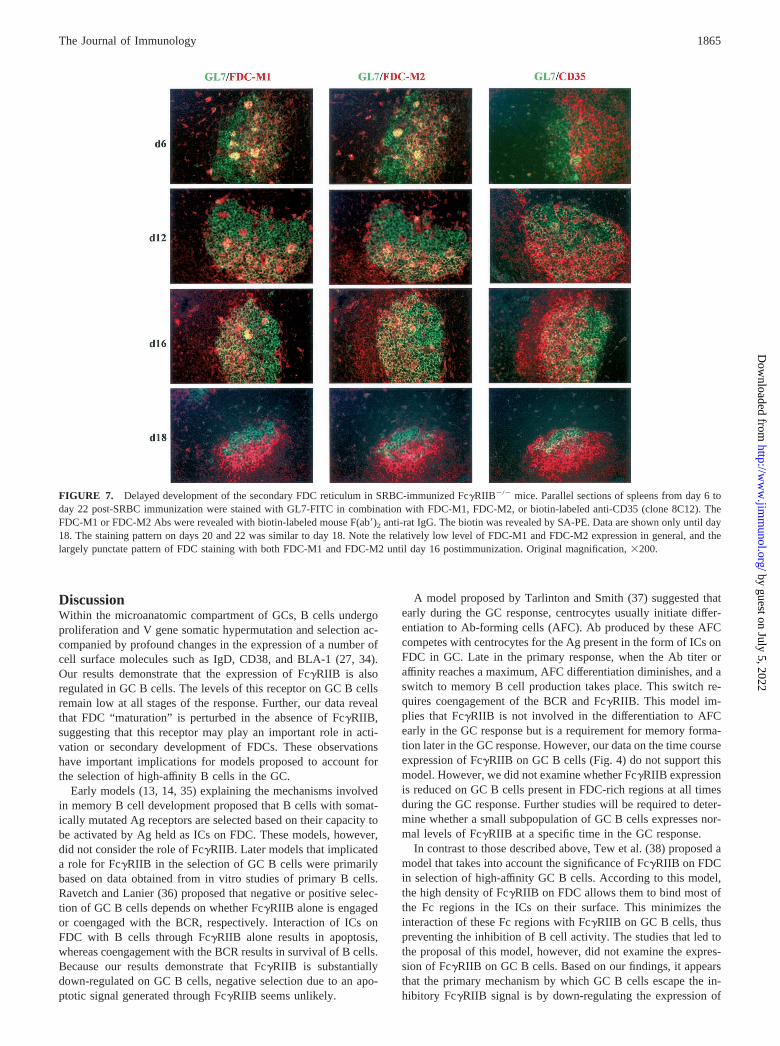
The differential regulation of Fc�RIIB on GC B cells and FDCprompted us to examine the GC response in mice lacking thisreceptor. Histological analysis of spleen sections from day 6 to day22 post-SRBC immunization was performed on Fc�RIIB�/� andC57BL/6 mice with GL7, FDC-M1, and FDC-M2. In C57BL/6mice, the GC FDC reticulum was seen as fine processes extendingthroughout one side of the GC (see Fig. 4). This was true at all timepoints after immunization. In contrast, as shown in Fig. 7, thisreticular pattern of staining was very weak in Fc�RIIB-deficientGCs at early and intermediate times in the GC response. In these
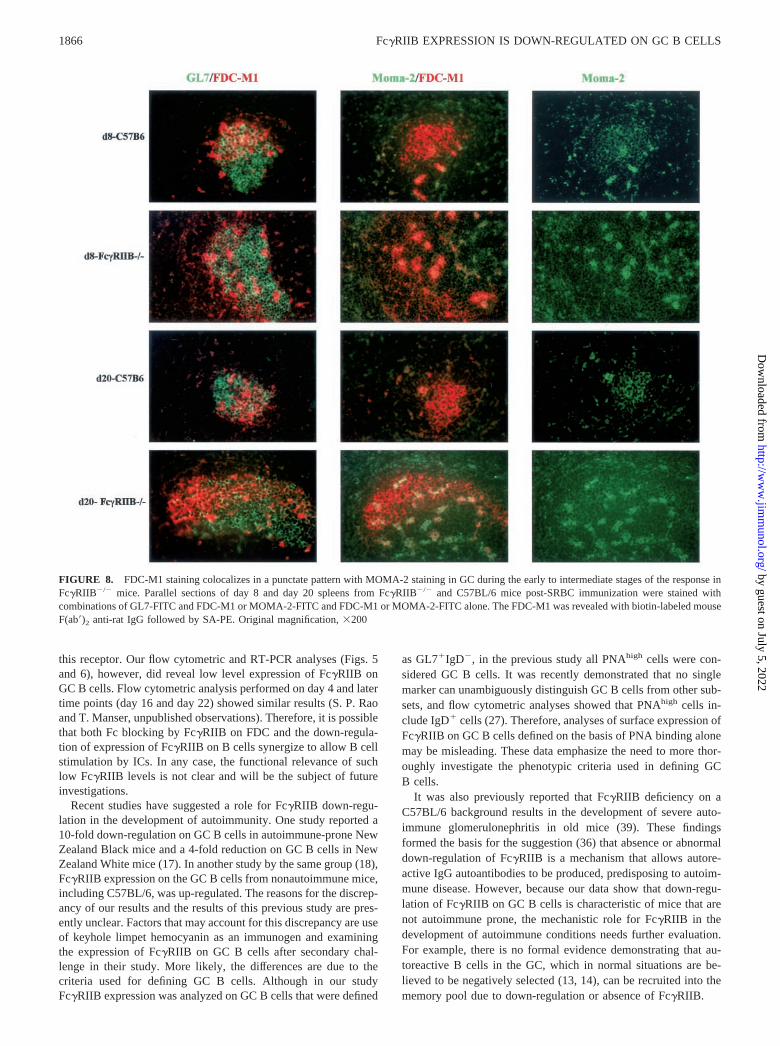
GCs, strong FDC-M1 and FDC-M2 staining exhibited a largelypunctate pattern until day 16. Thereafter, FDC staining was similarto that seen in C57BL/6 mice. The primary FDC reticula inFc�RIIB�/� mice, however, appeared normal at all stages of theGC response as judged by staining with the anti-CD35 Ab-8C12.Interestingly, even the punctate pattern of FDC staining observedduring the early GC response of Fc�RIIB�/� mice was largelyrestricted to one side of the GC. To investigate the cellular originof this punctate staining, parallel sections from the experimentillustrated in Fig. 7 were stained with GL7, FDC-M1, and theMOMA-2 mAb. MOMA-2 has been previously reported to detectmonocytes and tingible body macrophages (33). Representativeimages from this analysis are shown in Fig. 8. At early time pointsin Fc�RIIB�/� mice (day 8 is illustrated in Fig. 8), MOMA-2 andFDC-M1 staining were largely colocalized to the punctate bodies,and these were rare in C57BL/6 GCs. At later times (day 20 isillustrated), whereas the reticular pattern of FDC-M1 staining inFc�RIIB�/� GCs appeared similar to C57BL/6 GCs, Moma-2/FDC-M1 double-positive cells were still clearly visible but re-mained rare in C57BL/6 GCs. Interestingly, we also observedMOMA-2 positive, FDC-M1 negative cells in the GCs ofFc�RIIB�/� mice at these later time points.
To test whether the altered pattern of FDC-M1 intra-GC stainingcharacteristic of the early stages of the GC response in Fc�RIIB�/�
mice correlated with a difference in the time course of the GCresponse, the kinetics of this response were examined in Fc�RIIB�/�
mice from day 4 to day 22 after SRBC immunization. As shown inFig. 9, there was no apparent difference in the kinetics of appearanceor disappearance of total or different sized GL7� GCs in Fc�RIIB�/�
mice as compared with C57BL/6 mice (Fig. 1). These results suggestthat while the development of the “mature” FDC reticulum is delayed,the general kinetics and magnitude of the GC B cell response remainunchanged due to absence of Fc�RIIB.
FIGURE 5. Comparison of Fc�RIIB expression between splenicGL7�IgD� non-GC and GL7�IgD� GC B cells. Fc� chain�/� andC57BL/6 mice were immunized with SRBC, and spleen cells were ob-tained on day 8 after immunization. Four color flow cytometric analyseswas performed after the cells were stained with GL7-FITC and anti-B220-PE-Texas Red in combination either with anti-IgD-biotin and2.4G2-PE or with anti-IgD-PE and K9.361-biotin. The biotin was re-vealed with SA-Red 670. The histograms show Fc�RIIB expressionlevels on B220�GL7�IgD� non-GC (– – – –) and B220�GL7� IgD�
GC B cells (OO) in Fc� chain�/� mice (A) and C57BL/6 mice (B). Theshaded portions represent the staining obtained with a rat isotype con-trol (A) and a mouse isotype control (B).
FIGURE 6. Fc�RIIB mRNA levels are �6-fold lower in GC B cellscompared with non-GC B cells. C57BL/6 mice were immunized withSRBC. On day 8 postimmunization, mRNA was extracted from sorted GCand non-GC B cells followed by cDNA synthesis and Fc�RIIB-specificPCR amplification. As an internal control, �-actin mRNA was amplified(not shown). PCR products removed after different numbers of PCR cycleswere subjected to agarose gel electrophoresis. The gel was hybridized withan Fc�RIIB-specific probe and analyzed by PhosphorImager (see Materi-als and Methods). The relative intensity of each band representing indi-vidual PCR products was measured and expressed as units of integratedband intensity. �, GC B cells; f, non-GC B cells.
1864 Fc�RIIB EXPRESSION IS DOWN-REGULATED ON GC B CELLS
by guest on July 5, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
DiscussionWithin the microanatomic compartment of GCs, B cells undergoproliferation and V gene somatic hypermutation and selection ac-companied by profound changes in the expression of a number ofcell surface molecules such as IgD, CD38, and BLA-1 (27, 34).Our results demonstrate that the expression of Fc�RIIB is alsoregulated in GC B cells. The levels of this receptor on GC B cellsremain low at all stages of the response. Further, our data revealthat FDC “maturation” is perturbed in the absence of Fc�RIIB,suggesting that this receptor may play an important role in acti-vation or secondary development of FDCs. These observationshave important implications for models proposed to account forthe selection of high-affinity B cells in the GC.
Early models (13, 14, 35) explaining the mechanisms involvedin memory B cell development proposed that B cells with somat-ically mutated Ag receptors are selected based on their capacity tobe activated by Ag held as ICs on FDC. These models, however,did not consider the role of Fc�RIIB. Later models that implicateda role for Fc�RIIB in the selection of GC B cells were primarilybased on data obtained from in vitro studies of primary B cells.Ravetch and Lanier (36) proposed that negative or positive selec-tion of GC B cells depends on whether Fc�RIIB alone is engagedor coengaged with the BCR, respectively. Interaction of ICs onFDC with B cells through Fc�RIIB alone results in apoptosis,whereas coengagement with the BCR results in survival of B cells.Because our results demonstrate that Fc�RIIB is substantiallydown-regulated on GC B cells, negative selection due to an apo-ptotic signal generated through Fc�RIIB seems unlikely.
A model proposed by Tarlinton and Smith (37) suggested thatearly during the GC response, centrocytes usually initiate differ-entiation to Ab-forming cells (AFC). Ab produced by these AFCcompetes with centrocytes for the Ag present in the form of ICs onFDC in GC. Late in the primary response, when the Ab titer oraffinity reaches a maximum, AFC differentiation diminishes, and aswitch to memory B cell production takes place. This switch re-quires coengagement of the BCR and Fc�RIIB. This model im-plies that Fc�RIIB is not involved in the differentiation to AFCearly in the GC response but is a requirement for memory forma-tion later in the GC response. However, our data on the time courseexpression of Fc�RIIB on GC B cells (Fig. 4) do not support thismodel. However, we did not examine whether Fc�RIIB expressionis reduced on GC B cells present in FDC-rich regions at all timesduring the GC response. Further studies will be required to deter-mine whether a small subpopulation of GC B cells expresses nor-mal levels of Fc�RIIB at a specific time in the GC response.
In contrast to those described above, Tew et al. (38) proposed amodel that takes into account the significance of Fc�RIIB on FDCin selection of high-affinity GC B cells. According to this model,the high density of Fc�RIIB on FDC allows them to bind most ofthe Fc regions in the ICs on their surface. This minimizes theinteraction of these Fc regions with Fc�RIIB on GC B cells, thuspreventing the inhibition of B cell activity. The studies that led tothe proposal of this model, however, did not examine the expres-sion of Fc�RIIB on GC B cells. Based on our findings, it appearsthat the primary mechanism by which GC B cells escape the in-hibitory Fc�RIIB signal is by down-regulating the expression of
FIGURE 7. Delayed development of the secondary FDC reticulum in SRBC-immunized Fc�RIIB�/� mice. Parallel sections of spleens from day 6 today 22 post-SRBC immunization were stained with GL7-FITC in combination with FDC-M1, FDC-M2, or biotin-labeled anti-CD35 (clone 8C12). TheFDC-M1 or FDC-M2 Abs were revealed with biotin-labeled mouse F(ab�)2 anti-rat IgG. The biotin was revealed by SA-PE. Data are shown only until day18. The staining pattern on days 20 and 22 was similar to day 18. Note the relatively low level of FDC-M1 and FDC-M2 expression in general, and thelargely punctate pattern of FDC staining with both FDC-M1 and FDC-M2 until day 16 postimmunization. Original magnification, �200.
1865The Journal of Immunology
by guest on July 5, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
this receptor. Our flow cytometric and RT-PCR analyses (Figs. 5and 6), however, did reveal low level expression of Fc�RIIB onGC B cells. Flow cytometric analysis performed on day 4 and latertime points (day 16 and day 22) showed similar results (S. P. Raoand T. Manser, unpublished observations). Therefore, it is possiblethat both Fc blocking by Fc�RIIB on FDC and the down-regula-tion of expression of Fc�RIIB on B cells synergize to allow B cellstimulation by ICs. In any case, the functional relevance of suchlow Fc�RIIB levels is not clear and will be the subject of futureinvestigations.
Recent studies have suggested a role for Fc�RIIB down-regu-lation in the development of autoimmunity. One study reported a10-fold down-regulation on GC B cells in autoimmune-prone NewZealand Black mice and a 4-fold reduction on GC B cells in NewZealand White mice (17). In another study by the same group (18),Fc�RIIB expression on the GC B cells from nonautoimmune mice,including C57BL/6, was up-regulated. The reasons for the discrep-ancy of our results and the results of this previous study are pres-ently unclear. Factors that may account for this discrepancy are useof keyhole limpet hemocyanin as an immunogen and examiningthe expression of Fc�RIIB on GC B cells after secondary chal-lenge in their study. More likely, the differences are due to thecriteria used for defining GC B cells. Although in our studyFc�RIIB expression was analyzed on GC B cells that were defined
as GL7�IgD�, in the previous study all PNAhigh cells were con-sidered GC B cells. It was recently demonstrated that no singlemarker can unambiguously distinguish GC B cells from other sub-sets, and flow cytometric analyses showed that PNAhigh cells in-clude IgD� cells (27). Therefore, analyses of surface expression ofFc�RIIB on GC B cells defined on the basis of PNA binding alonemay be misleading. These data emphasize the need to more thor-oughly investigate the phenotypic criteria used in defining GCB cells.
It was also previously reported that Fc�RIIB deficiency on aC57BL/6 background results in the development of severe auto-immune glomerulonephritis in old mice (39). These findingsformed the basis for the suggestion (36) that absence or abnormaldown-regulation of Fc�RIIB is a mechanism that allows autore-active IgG autoantibodies to be produced, predisposing to autoim-mune disease. However, because our data show that down-regu-lation of Fc�RIIB on GC B cells is characteristic of mice that arenot autoimmune prone, the mechanistic role for Fc�RIIB in thedevelopment of autoimmune conditions needs further evaluation.For example, there is no formal evidence demonstrating that au-toreactive B cells in the GC, which in normal situations are be-lieved to be negatively selected (13, 14), can be recruited into thememory pool due to down-regulation or absence of Fc�RIIB.
FIGURE 8. FDC-M1 staining colocalizes in a punctate pattern with MOMA-2 staining in GC during the early to intermediate stages of the response inFc�RIIB�/� mice. Parallel sections of day 8 and day 20 spleens from Fc�RIIB�/� and C57BL/6 mice post-SRBC immunization were stained withcombinations of GL7-FITC and FDC-M1 or MOMA-2-FITC and FDC-M1 or MOMA-2-FITC alone. The FDC-M1 was revealed with biotin-labeled mouseF(ab�)2 anti-rat IgG followed by SA-PE. Original magnification, �200
1866 Fc�RIIB EXPRESSION IS DOWN-REGULATED ON GC B CELLS
by guest on July 5, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
Our data also provide new insights into the role of Fc�RIIB inthe “maturation” of FDC. We have reported that FDC-specificAbs, especially FDC-M1, recognize Ags that are induced on FDCin secondary but not primary follicles (16), suggesting that theexpression of these Ags is associated with the maturation of FDC.Our data indicate that the gross GC response in Fc�RIIB�/� miceis comparable in kinetics and magnitude with that of normal mice.However, the time course histological analysis with FDC-specificAbs revealed a significant delay in the normally strong reticularexpression pattern of the FDC-M1 and FDC-M2 Ags, indicating adefect in the timing of maturation of FDC in the GC. Moreover,during the early phases of the GC response in Fc�RIIB�/� mice,strong FDC-M1 staining colocalizes in a punctate pattern withMOMA-2 staining. Further studies are required to determine thenature of the cells that give rise to this staining pattern. They maybe a subset of tingible body macrophages that express the FDC-M1Ag, but it is also interesting to speculate that they are FDC pre-cursors (40) that have yet to develop dendritic morphology.
B cell recall responses are thought to develop in GCs in re-sponse to ICs trapped on FDC. In this context, a recent study (16)investigated the significance of Fc�RIIB on the accessory activityof FDC. In contrast to normal FDCs, those from Fc�RIIB-deficientmice were incapable of augmenting IC-mediated B cell recall re-sponses in vitro. Given our findings, it is possible that this lack ofaccessory activity of FDCs may be due not only to absence ofFc�RIIB but to a defect in FDC maturation as well. More gener-ally, our data suggest that the function of Fc�RIIB on FDC is notmerely to trap ICs but also to promote FDC maturation and ex-pression of hitherto unknown molecules that may be involved inthe regulation of GC B cell selection, memory development, andrecall responses. Based on the correlation between the appearanceof the FDC-M1 and FDC-M2 Ags and Fc�RIIB up-regulation onFDC during immune responses, it is tempting to speculate that thepathways involved in the regulation of expression of these mole-cules are one and the same. Clearly, more studies are required toelucidate the role of Fc�RIIB and mechanisms regulating the ex-pression of this receptor on GC B cells and FDCs.
Another observation made in our study was the sectoring of GCsinto two clearly discernible compartments when stained with Absspecific for FDC or Fc�RIIB. This sectoring was observed at alltimes during the GC response and raises the possibility that these
areas in murine GC may correspond to the light zone composed ofcentrocytes (area in which the FDC network is organized) and thedark zone composed of centroblasts (area where the FDC networkis absent) reported in earlier studies on human tonsillar GCs (41,42). If so, the delay in FDC maturation in Fc�RIIB deficient micemay perturb the selection, affinity maturation and memory B celldevelopment of centrocytes in the light zone. However, in themouse, formal evidence that GC B cells in these areas actuallyrepresent centroblasts and centrocytes is currently lacking.
References1. Ravetch, J. V., and J. P. Kinet. 1991. Fc receptors. Annu. Rev. Immunol. 9:457.2. Hulett, M. D., and P. M. Hogarth. 1994. Molecular basis of Fc receptor function.
Adv. Immunol. 57:1.3. Daeron, M. 1997. Fc receptor biology. Annu. Rev. Immunol. 15:203.4. Lewis, V. A., T. Koch, H. Plutner, and I. Mellman. 1986. A complementary DNA
clone for a macrophage-lymphocyte Fc receptor. Nature 324:372.5. Ravetch, J. V., A. D. Luster, R. Weinshank, J. Kochan, A. Pavlovec,
D. A. Portnoy, J. Hulmes, Y. C. Pan, and J. C. Unkeless. 1986. Structural het-erogeneity and functional domains of murine immunoglobulin G Fc receptors.Science 234:718.
6. Bolland, S., and J. V. Ravetch. 1999. Inhibitory pathways triggered by ITIM-containing receptors. Adv. Immunol. 72:149.
7. Phillips, N. E., and D. C. Parker. 1984. Cross-linking of B lymphocyte Fc�receptors and membrane immunoglobulin inhibits anti-immunoglobulin-inducedblastogenesis. J. Immunol. 132:627.
8. Phillips, N. E., and D. C. Parker. 1985. Subclass specificity of Fc� receptor-mediated inhibition of mouse B cell activation. J. Immunol. 134:2835.
9. Coggeshall, K. M. 1998. Inhibitory signaling by B cell Fc�RIIb. Curr. Opin.Immunol. 10:306.
10. Pearse, R. N., T. Kawabe, S. Bolland, R. Guinamard, T. Kurosaki, andJ. V. Ravetch. 1999. SHIP recruitment attenuates Fc�RIIB-induced B cell apo-ptosis. Immunity 10:753.
11. Ono, M., H. Okada, S. Bolland, S. Yanagi, T. Kurosaki, and J. V. Ravetch. 1997.Deletion of SHIP or SHP-1 reveals two distinct pathways for inhibitory signaling.Cell 90:293.
12. Ashman, R. F., D. Peckham, and L. L. Stunz. 1996. Fc receptor off-signal in theB cell involves apoptosis. J. Immunol. 157:5.
13. Nossal, G. J. 1994. Negative selection of lymphocytes. Cell 76:229.14. Rajewsky, K. 1996. Clonal selection and learning in the antibody system. Nature
381:751.15. Healy, J. I., and C. C. Goodnow. 1998. Positive versus negative signaling by
lymphocyte antigen receptors. Annu. Rev. Immunol. 16:645.16. Qin, D., J. Wu, K. A. Vora, J. V. Ravetch, A. K. Szakal, T. Manser, and
J. G. Tew. 2000. Fc� receptor IIB on follicular dendritic cells regulates the B cellrecall response. J. Immunol. 164:6268.
17. Jiang, Y., S. Hirose, R. Sanokawa-Akakura, M. Abe, X. Mi, N. Li, Y. Miura,J. Shirai, D. Zhang, Y. Hamano, and T. Shirai. 1999. Genetically determinedaberrant down-regulation of Fc�RIIB1 in germinal center B cells associated withhyper-IgG and IgG autoantibodies in murine systemic lupus erythematosus. Int.Immunol. 11:1685.
18. Jiang, Y., S. Hirose, M. Abe, R. Sanokawa-Akakura, M. Ohtsuji, X. Mi, N. Li,Y. Xiu, D. Zhang, J. Shirai, Y. Hamano, H. Fujii, and T. Shirai. 2000. Polymor-phisms in IgG Fc receptor IIB regulatory regions associated with autoimmunesusceptibility. Immunogenetics 51:429.
19. Brauweiler, A., I. Tamir, S. Marschner, C. D. Helgason, and J. C. Cambier. 2001.Partially distinct molecular mechanisms mediate inhibitory Fc�RIIB signaling inresting and activated B cells. J. Immunol. 167:204.
20. Takai, T., M. Ono, M. Hikida, H. Ohmori, and J. V. Ravetch. 1996. Augmentedhumoral and anaphylactic responses in Fc�RII-deficient mice. Nature 379:346.
21. Takai, T., M. Li, D. Sylvestre, R. Clynes, and J. V. Ravetch. 1994. FcR� chaindeletion results in pleiotrophic effector cell defects. Cell 76:519.
22. Vora, K. A., J. V. Ravetch, and T. Manser. 1997. Amplified follicular immunecomplex deposition in mice lacking the Fc receptor �-chain does not alter mat-uration of the B cell response. J. Immunol. 159:2116.
23. Lentz, V. M., and T. Manser. 2000. Self-limiting systemic autoimmune diseaseduring reconstitution of T cell-deficient mice with syngeneic T cells: support fora multifaceted role of T cells in the maintenance of peripheral B cell tolerance.Int. Immunol. 12:1483.
24. Kraal, G., I. L. Weissman, and E. C. Butcher. 1982. Germinal centre B cells:antigen specificity and changes in heavy chain class expression. Nature 298:377.
25. Kroese, F. G., W. Timens, and P. Nieuwenhuis. 1990. Germinal center reactionand B lymphocytes: morphology and function. Curr. Top. Pathol. 84:103.
26. Vora, K. A., K. M. Tumas-Brundage, V. M. Lentz, A. Cranston, R. Fishel, andT. Manser. 1999. Severe attenuation of the B cell immune response in Msh2-deficient mice. J. Exp. Med. 189:471.
27. Shinall, S. M., M. Gonzalez-Fernandez, R. J. Noelle, and T. J. Waldschmidt.2000. Identification of murine germinal center B cell subsets defined by the ex-pression of surface isotypes and differentiation antigens. J. Immunol. 164:5729.
28. Cervenak, L., A. Magyar, R. Boja, and G. Laszlo. 2001. Differential expressionof GL7 activation antigen on bone marrow B cell subpopulations and peripheralB cells. Immunol. Lett. 78:89.
FIGURE 9. The kinetics and magnitude of the GC response to SRBCimmunization in Fc�RIIB�/� mice are similar to those in C57BL/6 mice.Fc�RIIB�/� mice were immunized with SRBC, and spleens were removedat the indicated time points. Sections were stained with GL7-FITC. Thetotal number and the different sizes of GCs were determined as describedin the legend to Fig. 1. Error bars represent SD. Data are representative ofat least three sections and two mice at each time point.
1867The Journal of Immunology
by guest on July 5, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

29. Kimura, S., N. Tada, E. Nakayama, Y. Liu, and U. Hammerling. 1981. A newmouse cell-surface antigen (Ly-m20) controlled by a gene linked to Mls locus anddefined by monoclonal antibodies. Immunogenetics 14:3.
30. Holmes, K. L., R. G. Palfree, U. Hammerling, and H. C. Morse III. 1985. Allelesof the Ly-17 alloantigen define polymorphisms of the murine IgG Fc receptor.Proc. Natl. Acad. Sci. USA 82:7706.
31. Schiller, C., I. Janssen-Graalfs, U. Baumann, K. Schwerter-Strumpf, S. Izui,T. Takai, R. E. Schmidt, and J. E. Gessner. 2000. Mouse Fc�RII is a negativeregulator of Fc�RIII in IgG immune complex-triggered inflammation but not inautoantibody-induced hemolysis. Eur. J. Immunol. 30:481.
32. Tew, J. G., J. Wu, D. Qin, S. Helm, G. F. Burton, and A. K. Szakal. 1997.Follicular dendritic cells and presentation of antigen and costimulatory signals toB cells. Immunol. Rev. 156:39.
33. Kraal, G., M. Rep, and M. Janse. 1987. Macrophages in T and B cell compart-ments and other tissue macrophages recognized by monoclonal antibodyMOMA-2: an immunohistochemical study. Scand J. Immunol. 26:653.
34. Oliver, A. M., F. Martin, and J. F. Kearney. 1997. Mouse CD38 is down-regu-lated on germinal center B cells and mature plasma cells. J. Immunol. 158:1108.
35. Liu, Y. J., G. D. Johnson, J. Gordon, and I. C. MacLennan. 1992. Germinalcentres in T-cell-dependent antibody responses. Immunol. Today 13:17.
36. Ravetch, J. V., and L. L. Lanier. 2000. Immune inhibitory receptors. Science290:84.
37. Tarlinton, D. M., and K. G. Smith. 2000. Dissecting affinity maturation: a modelexplaining selection of antibody-forming cells and memory B cells in the ger-minal centre. Immunol. Today 21:436.
38. Tew, J. G., J. Wu, M. Fakher, A. K. Szakal, and D. Qin. 2001. Follicular dendriticcells: beyond the necessity of T-cell help. Trends Immunol. 22:361.
39. Bolland, S., and J. V. Ravetch. 2000. Spontaneous autoimmune disease inFc(�)RIIB-deficient mice results from strain-specific epistasis. Immunity 13:277.
40. Szakal, A. K., K. L. Holmes, and J. G. Tew. 1983. Transport of immune com-plexes from the subcapsular sinus to lymph node follicles on the surface ofnonphagocytic cells, including cells with dendritic morphology. J. Immunol. 131:1714.
41. Hardie, D. L., G. D. Johnson, M. Khan, and I. C. MacLennan. 1993. Quantitativeanalysis of molecules which distinguish functional compartments within germinalcenters. Eur. J. Immunol. 23:997.
42. Kosco-Vilbois, M. H., H. Zentgraf, J. Gerdes, and J. Y. Bonnefoy. 1997. To “B”or not to “B” a germinal center? Immunol. Today 18:225.
1868 Fc�RIIB EXPRESSION IS DOWN-REGULATED ON GC B CELLS
by guest on July 5, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
CORRECTIONSRoderich E. Schwarz and John C. Hiserodt. The Expression and Functional Involvement of Laminin-Like Molecules inNon-MHC Restricted Cytotoxicity by Human Leu-19�/CD3� Natural Killer Lymphocytes. The Journal of Immunology,1988, 141: 3318–3323.
The Journal received the following letter from Roderich E. Schwarz requesting correction of this article, which waspublished in the November 15, 1988 issue:
To the editor:
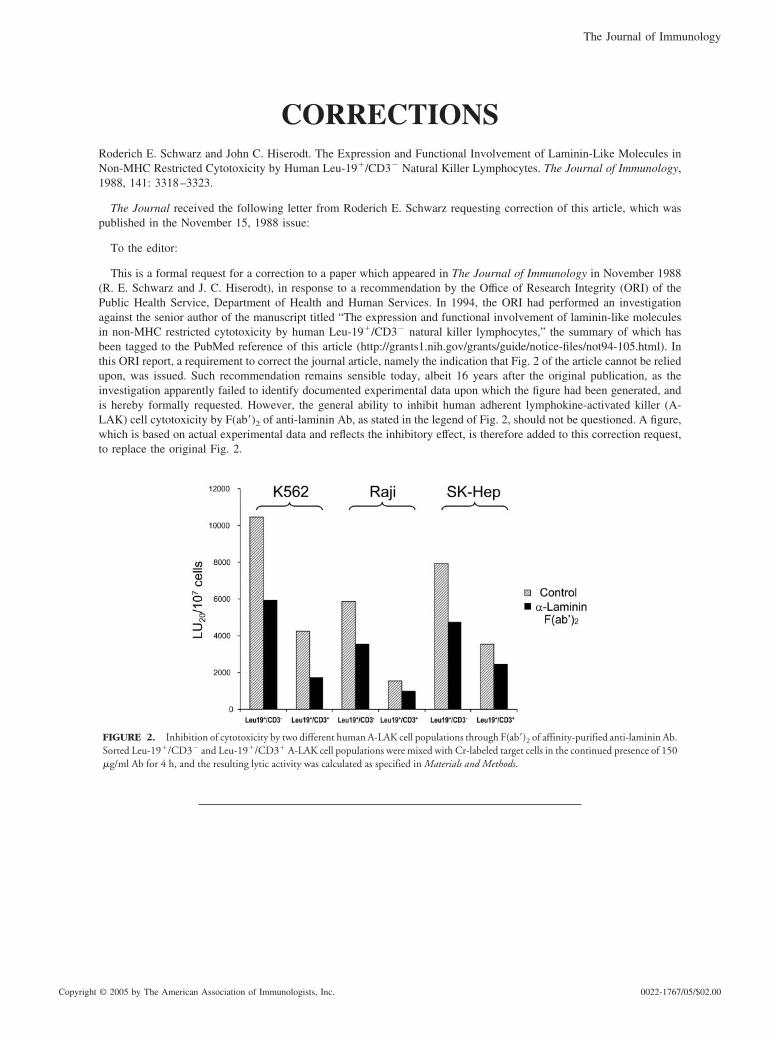
This is a formal request for a correction to a paper which appeared in The Journal of Immunology in November 1988(R. E. Schwarz and J. C. Hiserodt), in response to a recommendation by the Office of Research Integrity (ORI) of thePublic Health Service, Department of Health and Human Services. In 1994, the ORI had performed an investigationagainst the senior author of the manuscript titled “The expression and functional involvement of laminin-like moleculesin non-MHC restricted cytotoxicity by human Leu-19�/CD3� natural killer lymphocytes,” the summary of which hasbeen tagged to the PubMed reference of this article (http://grants1.nih.gov/grants/guide/notice-files/not94-105.html). Inthis ORI report, a requirement to correct the journal article, namely the indication that Fig. 2 of the article cannot be reliedupon, was issued. Such recommendation remains sensible today, albeit 16 years after the original publication, as theinvestigation apparently failed to identify documented experimental data upon which the figure had been generated, andis hereby formally requested. However, the general ability to inhibit human adherent lymphokine-activated killer (A-LAK) cell cytotoxicity by F(ab�)2 of anti-laminin Ab, as stated in the legend of Fig. 2, should not be questioned. A figure,which is based on actual experimental data and reflects the inhibitory effect, is therefore added to this correction request,to replace the original Fig. 2.
FIGURE 2. Inhibition of cytotoxicity by two different human A-LAK cell populations through F(ab�)2 of affinity-purified anti-laminin Ab.Sorted Leu-19�/CD3� and Leu-19�/CD3� A-LAK cell populations were mixed with Cr-labeled target cells in the continued presence of 150�g/ml Ab for 4 h, and the resulting lytic activity was calculated as specified in Materials and Methods.
The Journal of Immunology
Copyright © 2005 by The American Association of Immunologists, Inc. 0022-1767/05/$02.00

Sambasiva P. Rao, Kalpit A. Vora, and Tim Manser. Differential Expression of the Inhibitory IgG Fc Receptor Fc�RIIBon Germinal Center Cells: Implications for Selection of High-Affinity B Cells. The Journal of Immunology, 2002, 169:1859–1868.
The Journal received the following letter requesting correction of this article, which was published in the August 15,2002 issue:
The authors would like to alert the scientific community to the fact that we have been unable to reproduce one of thefindings presented in this manuscript. In Fig. 5B of this manuscript we showed the results of flow cytometric studiesdesigned to measure the levels of surface expression of Fc�RIIB on splenic germinal center (GC) B cells (defined asB220�, IgD�, GL7�) as compared with splenic non-GC B cells (defined as B220�, IgD�, GL7�) using the anti-Fc�RIIBmAb K9.361. These cells were isolated from C57BL/6 mice that had been immunized i.p. 8 days earlier with 3 � 108
sheep RBC (SRBC) per mouse. Fig. 5B illustrated �5-fold lower levels of K9.361 staining on GC B cells as comparedwith non-GC B cells. In Fig. 6, we presented the results of the semiquantitation of Fc�RIIB mRNA levels, via RT-PCRand in gel hybridization, in these two populations of B cells that had been purified by FACS. This figure indicated �6-foldlower levels of Fc�RIIB mRNA in GC, as compared with non-GC B cells.
In multiple recent experiments designed to extend these published studies, neither the reduced levels of K9.361 surfacestaining of B220�, IgD�, GL7� splenic B cells detected by flow cytometry or the reduced levels of Fc�RIIB mRNA insuch cells isolated by FACS 8 days after i.p. immunization of C57BL/6 mice with SRBCs (evaluated via real-timeRT-PCR) have been observed.
In several other figures in the above-referenced manuscript, the results of immunohistological analysis of Fc�RIIBexpression in the GCs of SRBC immunized C57BL/6 mice were illustrated and interpreted to corroborate the results ofthe studies presented in Figs. 5 and 6. Due to the relative insensitivity of immunohistology as compared with flowcytometry, whether GC B cells stained 5- to 6-fold less intensely with anti-Fc�RIIB mAbs as compared with non-GC Bcells could not have been unequivocally determined using the former approach. Nonetheless, our previous interpretationsof these immunohistological data with regard to levels of Fc�RIIB on GC B cells appear to have been incorrect. Inaddition, arguments we forwarded in Discussion based on the conclusion that GC B cells express lower levels of Fc�RIIBthan non-GC B cells may no longer hold merit.
We currently can provide no compelling explanation for why our previously published results on the expression levelsof Fc�RIIB on GC B cells and the results of our more recent studies differ, but are actively investigating severalpossibilities. We should hasten to point out that our failure to reproduce the results presented in Figs. 5B and 6 does notinfluence the validity of any of the data or conclusions presented in the above-referenced manuscript regarding theexpression and function of Fc�RIIB on follicular dendritic cells.
Per O. Anderson, Anette Sundstedt, Zihni Yazici, Sophie Minaee, Richard Woolf, Kirsty Nicolson, Nathaniel Whitley,Li Li, Suling Li, David C. Wraith, and Ping Wang. IL-2 Overcomes the Unresponsiveness but Fails to Reverse theRegulatory Function of Antigen-Induced T Regulatory Cells. The Journal of Immunology, 2005, 174: 310–319.
The fifth author’s name, Emma J. O’Neill, was inadvertently omitted. The correct list of authors and affiliations isshown below.
Per O. Anderson,*† Anette Sundstedt,§ Zihni Yazici,§ Sophie Minaee,§ Emma J. O’Neill,§ Richard Woolf,§ KirstyNicolson,§ Nathaniel Whitley,§ Li Li,*† Suling Li,†‡ David C. Wraith,§ and Ping Wang*†
*Institute of Cell and Molecular Science, Barts and The London School of Medicine and Dentistry, London, UnitedKingdom; †Tumor Immunology, Lund University, Lund, Sweden; ‡Department of Biological Sciences, Brunel University,Uxbridge, London, United Kingdom; and §Department of Pathology and Microbiology, School of Medical Sciences,University of Bristol, Bristol, United Kingdom
5133The Journal of Immunology

Winifred Huang, Hans D. Ochs, Bo Dupont, and Yatin M. Vyas. The Wiskott-Aldrich Syndrome Protein RegulatesNuclear Translocation of NFAT2 and NF-�B (RelA) Independently of Its Role in Filamentous Actin Polymerization andActin Cytoskeletal Rearrangement. The Journal of Immunology, 2005, 174: 2602–2611.
In Results, Fig. 2A1 was incorrectly published in black and white. The error has been corrected in the online version,which now differs from the print version as originally published. The correct color figure is shown below.
Also in Results, Fig. 3 fails to mention the headings “NFAT2” and “NF-�B (RelA)” on both panels a and b. The legendto this figure is written with the presumption that the headings are present on each of the two panels. The correct figureis shown below.
Elizabeth P. Ryan, Stephen J. Pollack, Thomas I. Murant, Steven H. Bernstein, Raymond E. Felgar, and Richard P.Phipps. Activated Human B Lymphocytes Express Cyclooxygenase-2 and Cyclooxygenase Inhibitors Attenuate AntibodyProduction. The Journal of Immunology, 2005, 174: 2619–2626.
The second author’s last name is misspelled. The correct name is Stephen J. Pollock.
5134 CORRECTION

In This Issue. The Journal of Immunology, 2005, 174: 3135–3136.
In the last summary of In This Issue titled “Helicobacter pylori-neutrophil interactions,” an error was made in citingthe page number of the original article titled “Helicobacter pylori disrupts NADPH oxidase targeting in human neutrophilsto induce extracellular superoxide release” by Lee-Ann H. Allen, Benjamin R. Beecher, Jeffrey T. Lynch, Olga V. Rohner,and Lara M. Wittine. The correct page number associated with the article is 3658. The error has been corrected in theonline version, which now differs from the print version as originally published.
Soren Schenk, Danielle D. Kish, Chunshui He, Tarek El-Sawy, Eise Chiffoleau, Chuangqui Chen, Zihao Wu, SigridSandner, Anton V. Gorbachev, Kiyotaka Fukamachi, Peter S. Heeger, Mohamed H. Sayegh, Laurence A. Turka, andRobert L. Fairchild. Alloreactive T Cell Responses and Acute Rejection of Single Class II MHC-Disparate Heart Allo-grafts Are under Strict Regulation by CD4�CD25� T Cells. The Journal of Immunology, 2005, 174: 3741–3748.
The sixth author’s first name is misspelled. The correct name is Chuangqi Chen.
5135The Journal of Immunology





![[Expression of Fc receptors for IgG in peripheral blood leucocytes from hepatitis C virus infected individuals]](https://img.dokumen.tips/doc/110x75/635775e2ea2708a6d3021420/expression-of-fc-receptors-for-igg-in-peripheral-blood-leucocytes-from-hepatitis.jpg)

![[Implications of receptors for the Fc portion of IgG (FcgammaRs) in mechanism of action of therapeutic antibodies]](https://img.dokumen.tips/doc/110x75/63452682596bdb97a908cbba/implications-of-receptors-for-the-fc-portion-of-igg-fcgammars-in-mechanism-of.jpg)





















