Embed Size (px)
Citation preview

Dissertation Thesis in Environmental Chemistry
Ecotoxicological Aspects of Humic Substances
Michal Bittner
2007
Masaryk University, Faculty of Science, RECETOX - Research Centre for Environmental Chemistry and Ecotoxicology Brno, Czech Republic
Supervisor: prof. Dr. Ivan Holoubek, Ph.D.

2

Bibliographic identification
Author: Michal Bittner
Title of dissertation: Ecotoxicological aspects of humic substances
Title of dissertation (in Czech): Ekotoxikologické aspekty huminových látek
Ph.D. study program: Chemistry
Specialization: Environmental chemistry
Supervisor: Prof. Dr. Ivan Holoubek, Ph.D.
Year of defense: 2007
Keywords: Humic substances, AhR, ER, in vitro bioassay, photo-transformation, effects on organisms
Keywords (in Czech): Huminové látky, AhR, ER, in vitro biotest, fototransformace, účinky na organismy

© Michal Bittner, Masaryk University, 2007
4

Acknowledgements
I would like to thank:
• my supervisor Prof. Ivan Holoubek for providing me with outstanding research and working conditions;
• my consultants Dr. Klára Hilscherová and Dr. Luděk Bláha for all the invaluable support, motivation, advice and ideas during all my Ph.D. study;
• Dr. Jaroslav Janošek for his help, friendly cooperation and very inspiring ideas;
• all my colleagues and friends from the RECETOX for the help with research, and especially office-mates for the unforgettable working atmosphere;
• Ing. Dagmar Gajdošová and Prof. Josef Havel (Dept. of Analytical Chemistry, Masaryk University) for their willing cooperation;
• Ministry of Education, Youth and Sports of the Czech Republic for financial support (FRVS G4 2510/2006);
Most important, I would like to express my gratitude to my wife, family and friends for support and encouragement during my studies.

Content
6

Content
Content
List of abbreviations ........................................................................................ 9
Abstract............................................................................................................. 11
Abstrakt (Abstract in Czech) ....................................................................... 13
List of original articles................................................................................... 15
The author’s contribution in the articles .................................................. 16
Aims of the dissertation thesis ..................................................................... 17
1. Introduction ............................................................................................ 19
1.1. Origin and classification of humic substances (HS) ........................... 20 1.1.1. Humification process and its theories............................................ 201.1.2. Classification of HS....................................................................... 21
1.2. Molecular structure of HS................................................................... 221.2.1. General structural features............................................................. 221.2.2. Supramolecular structure ............................................................... 23
2. Effect of HS on living organisms (incl. Paper I and II)................. 25
2.1. Indirect effects of HS on organisms .................................................... 27 2.1.1. Binding of metals........................................................................... 27 2.1.2. Binding of organic xenobiotics...................................................... 28
2.2. Direct effects of HS on organisms....................................................... 31 2.2.1. Uptake of HS by organisms........................................................... 31 2.2.2. Toxic effects of HS........................................................................ 32
2.2.2.1. Non-specific mechanisms of toxicity ........................................ 322.2.2.2. Specific mechanisms of toxicity................................................ 322.2.2.3. HA - possible etiological agens of Blackfoot disease ............... 35 2.2.2.4. Hormone-like effects................................................................. 37 2.2.2.5. Interaction of HS with intracellular receptors ........................... 40
2.3. Activation of Ah receptor by HS (Paper I and II) .............................. 41
7
2.3.1. Aryl hydrocarbon receptor (AhR) ................................................. 41

Content
2.3.1.1. Mechanism of AhR-mediated effects........................................ 412.3.1.2. Ligands of AhR ......................................................................... 42 2.3.1.3. Consequences of exposure to weak AhR ligands...................... 45
2.3.2. Results - AhR-mediated activity of HS (Paper I + first part of Paper II ) ....................................................................................... 46
2.3.2.1. HS used in experiments ............................................................. 46 2.3.2.2. Paper I ....................................................................................... 47 2.3.2.3. Paper II - part focused on AhR-mediated activity of HS .......... 49
2.4. Anti/estrogenic activity of HS (Paper II) ............................................ 52 2.4.1. Estrogen receptor (ER) .................................................................. 52
2.4.1.1. Mechanism of ER-mediated effects .......................................... 53 2.4.1.2. Anti/estrogenic compounds....................................................... 54 2.4.1.3. Consequences of exposure to xenoestrogens ............................ 55
2.4.2. Results - ER-mediated activities of HS (Paper II) ......................... 55
3. Interaction of HS with light - ecotoxicological consequences (incl. Paper III) ................................................................................................ 57
3.1. Interaction with light........................................................................... 57 3.1.1. Direct toxicity of HS photo-products to organisms ....................... 58 3.1.2. HS mediated photo-transformations of xenobiotics ...................... 59
3.2. Changes of AhR-med. activity of HS after irradiation (Paper III)...... 61 3.2.1. Results - Decrease of absorbance and AhR-mediated activity of HS
after irradiation ............................................................................. 61
4. Summary and general discussion ....................................................... 65
References......................................................................................................... 69
Appendices ....................................................................................................... 79
I - Paper I
II - Paper II
III - Paper III
8
IV - Curriculum vitae

List of abbreviations
9
List of abbreviations AhR Aryl Hydrocarbon Receptor BaP Benzo[a]pyrene BFD Blackfoot Disease CYP DDT
Cytochrome P450 Dichloro-Diphenyl-Trichloroethane
DNA Deoxyribonucleic Acid DOC Dissolved Organic Carbon DOM Dissolved Organic Matter dpm Disintegrations per Minute E2 17-β-Estradiol ER Estrogen Receptor EROD Ethoxyresorufin O-Deethylase FA Fulvic Acids FW Fresh Weight GC-MS Gas Chromatography - Mass Spectrometry HA Humic Acids HAA Aldrich HA HAH Halogenated Aromatic Hydrocarbons HS HS*
Humic Substances Excited Humic Substances
HS1500 Synthetic Humic Acid HUVECs Human Umbilical Vein Endothelial Cells IC50 Concentration Resulting in 50% Inhibition IHSS International Humic Substances Society KOC Organic Carbon-Water Partition Coefficient LLE Liquid-Liquid Extraction LOEC Lowest Observed Effect Concentration mequ Men+
Miliequivalent Units Generic Metal Cation

List of abbreviations
10
NADPH NF-κB
Nicotinamide Adenine Dinucleotide Phosphate Nuclear Factor κ B
NOM Natural Organic Matter PAH Polycyclic Aromatic Hydrocarbons PCB Polychlorinated Biphenyls PCDF Polychlorinated Dibenzo Furans PCDD Polychlorinated Dibenzo-p-dioxins PCP Pentachlorophenol PeCB 3,3',4,4',5-Pentachlorobiphenyl
POPs Persistent Organic Pollutants PPARγ Peroxisome Proliferator-Activated Receptor γ RBCs Red Blood Cells RT-PCR Real Time Polymerase Chain Reaction ROS RS RRS
Reactive Oxygen Species Reactive Species Very Reactive Species
SD Standard Deviation SEC Size Exclusion Chromatography TEQ TCDD
Toxic (or TCDD) Equivalents 2,3,7,8-Tetrachlorodibenzo-p-dioxin
TCDF 2,3,7,8-Tetrachlorodibenzofuran TOC Total Organic Carbon U.S. EPA United States Environmental Protection Agency UV XAD
Ultraviolet Acrylic Ester Resin; Amberlite

Abstract
11
Abstract
Humic substances (HS) represent ubiquitous products of dead organic matter decomposition, whose chemical constitution depends on the source of parent materials - therefore, exact structure and properties of HS are unpredictable. HS contain great amount of functional groups and hydrophobic structures that implies interaction of HS with surrounding both inorganic and organic compounds. This results in indirect effects of HS on organisms due to decreased bioavailability of affected compounds. Despite the rather large HS-aggregates, it was also shown that HS-like substances can be taken up into the cells of aquatic organisms and subsequently exert direct biological effects - e. g. change expression of heat shock proteins, modulation of biotransformation enzymes, modulation of photosynthetic oxygen release, hormone-like effects on animals, etc. Direct interaction of HS with several intracellular receptors has also been described.
Aryl hydrocarbon receptor (AhR) represents ecotoxicologically really important intracellular receptor, which mediates the toxic effects of environmental contaminants such as polychlorinated dioxins, furans and biphenyls. Nevertheless number of other structurally diverse compounds has also been found to activate AhR. We have evaluated the AhR-mediated activity of various types of HS - humic acids (HA), fulvic acids (FA) and natural organic matter (NOM) - using in vitro bioassay based on transgenic H4IIE-luc cell line. Endpoint of this bioassay is luminescence determination of luciferase that is expressed in the cells after the AhR activation. On account of more detailed characterization of AhR-active HS compartments, activities of aqueous HS solutions, organic extracts and H2SO4-treated HS extracts were examined. Results of our studies revealed significant AhR-mediated activities of five HA out of twelve HS tested, where both alkali HS solutions and organic HS extracts exerted activity, but H2SO4-treated extracts were non-active.
In further study, the interaction of HS with another intracellular receptor - estrogen receptor (ER) - was examined. ER is a target for natural estrogen hormones, but can also be affected by so called “xenoestrogens”, i.e. compounds belonging to the group of endocrine disruptors. Similar to assessment of

Abstract
12
AhR-mediated activity, analogous transgenic cell-based in vitro bioassay was employed. Compared with H4IIE-luc cell line, MVLN cell line gave the luminescence response after the ER activation. None of the twelve tested HS showed any estrogenic activity both in absence and presence of ER-agonist 17-β-estradiol; but ten HS have exerted significant antiestrogenic activity. Compared to AhR-mediated activity, these antiestrogenic effects were observed not only after HA treatment, but also after FA and NOM treatment.
Since HS molecules contain number of chromophores - i.e. aromatic rings and conjugated double bonds, the natural irradiation of dissolved HS can lead to a series of photochemical reactions. These reactions can act on the HS itself, but also on other substances surrounding HS, such as e.g. xenobiotics. Thus, we have also studied the changes in AhR-mediated activities and absorbance of HS after irradiation that simulated natural solar light. After irradiation the absorbance of all of the HS was lower that of the original samples. The AhR-mediated activities of two HA samples (HA-Fluka and HA Sodium Salt) were partially decreased in dependence on irradiation, while activities of other HS, both AhR-active or -inactive, were not changed by irradiation. Results of our measurements demonstrate that AhR-mediated activities of two active HA is caused by both photo-stable and photo-labile AhR activators, while other three active HA contain only photo-stable AhR activators.

Abstrakt (Abstract in Czech)
13
Abstrakt (Abstract in Czech)
Huminové látky (HS) představují všudypřítomné produkty rozkladu odumřelé organické hmoty. Jejich chemické složení závisí na zdroji degradovaného materiálu, a proto není možné určit jejich přesnou chemickou strukturu ani jednotné vlastnosti. HS obsahují velké množství funkčních skupin a hydrofobních struktur zodpovědných za interakci HS s okolními anorganickými i organickými látkami. Důsledkem je snížení biodostupnosti těchto látek, popisované jako nepřímý účinek HS na organismy. Navzdory poměrně velkým agregátům HS však bylo také prokázáno, že látky typu HS mohou pronikat do živých buněk vodních organismů a následně působit přímým účinkem. Popsána tak byla např. změna exprese proteinů teplotního šoku, zvýšení množství biotransformačních enzymů, ovlivnění fotosyntetické tvorby kyslíku u rostlin, účinky hormonálního typu u živočichů, atd. Také již byly popsány případy přímé interakce HS s vnitrobuněčnými receptory.
Receptor pro planární aromatické látky (AhR) představuje ekotoxikologicky velice důležitý vnitrobuněčný receptor, který zprostředkovává toxické účinky kontaminantů životního prostředí, a to především polychlorovaných dioxinů, furanů a bifenylů. Byla již ale také identifikována řada strukturně velmi variabilních látek schopných interakce s AhR. My jsme provedli otestování AhR zprostředkované aktivity různých druhů HS - huminových kyselin (HA), fulvokyselin (FA) a rozpustné organické hmoty (NOM). Toto testování bylo provedeno za použití in vitro biotestu založeného na transgenní buněčné linii H4IIE-luc. Koncovkou tohoto biotestu je luminiscenční stanovení luciferázy, která je v buňkách exprimována po aktivaci AhR. Z důvodu bližší charakterizace AhR-aktivních složek HS jsme otestovali jak samotné vodné roztoky HS, tak také jejich organické extrakty a organické extrakty ošetřené H2SO4. Výsledkem bylo zjištění významné AhR zprostředkované aktivity u pěti vzorků HA a jejich organických extraktů, zatímco organické extrakty ošetřené H2SO4 aktivitu nevykazovaly.
Dále jsme studovali aktivaci dalšího vnitrobuněčného receptoru - estrogenního receptoru (ER). ER představuje cíl pro přirozené estrogenní hormony, avšak jeho funkce může být také ovlivněna tzv. xenoestrogeny, to jest látkami

Abstrakt (Abstract in Czech)
14
patřícími do skupiny endokrinních disruptorů. Podobně jako u testování AhR zprostředkované aktivity jsme i zde využili analogického in vitro biotestu založeného na transgenní buněčné linii. Linie MVLN však narozdíl od linie H4IIE-luc odpovídá luminiscenčním signálem na aktivaci ER. Testováním jsme zjistili významnou antiestrogenitu u deseti z dvanácti testovaných vzorků HS, zatímco estrogenita nebyla zjištěna u žádného vzorku HS, a to jak v přítomnosti tak i nepřítomnosti přirozeného ligandu ER, 17-β-estradiolu. Ve srovnání s AhR zprostředkovanou aktivitou však byl zjištěn významný antiestrogenní účinek nejenom u HA, ale i u FA a NOM.
Jelikož molekuly HS obsahují množství chromoforů, tedy aromatických struktur a konjugovaných dvojných vazeb, vede ozáření rozpuštěných HS k sérii fotochemických reakcí. Těchto reakcí se účastní jak HS samotné, tak i jiné okolní látky jako např. xenobiotika. V další studii jsme se tedy zaměřili na změny absorbance a AhR zprostředkovaných aktivit HS po ozáření simulujícím přirozené sluneční záření. V závislosti na době ozařování došlo k významnému poklesu absorbance u všech vzorků HS. AhR zprostředkovaná aktivita dvou vzorků HA (HA-Fluka a HA Sodium Salt) po ozáření částečně také poklesla, zatímco aktivita ostatních vzorků, jak AhR-aktivních tak i neaktivních, se po ozáření nezměnila. Tyto výsledky ukazují, že aktivita zprostředkovaná AhR je u dvou AhR-aktivních HA způsobena jak fotolabilními, tak i fotostabilními ligandy AhR, zatímco zbývající tři AhR-aktivní HA obsahují pouze fotostabilní ligandy AhR.

List of original articles
15
List of original articles The particular chapters of this thesis correspond with the following papers which are referred to by their Roman numerals in the text:
I. Bittner, M., Janosek, J., Hilscherova, K., Giesy, J., Holoubek, I., and Blaha, L. Activation of Ah receptor by pure humic acids. Environmental Toxicology, 21:338-342; 2006
II. Janosek, J., Bittner, M., Hilscherová, K., Bláha, L., Giesy, J.P., and
Holoubek, I. AhR-mediated and antiestrogenic activity of humic substances. Chemosphere; 2007 (in press)
III. Bittner, M., Hilscherová, K. Giesy J.P. Changes of AhR-mediated
activity of humic substances after irradiation; 2007 (submitted to Environment International)

The author’s contribution in the articles
16
The author’s contribution in the articles
Paper I.
Michal Bittner participated in the experimental design, cultured cell line, carried out all assessments of both reporter gene and EROD bioassays, evaluated data and co-interpreted the results. He wrote the manuscript and finalized the entire article.
Paper II.
Michal Bittner participated in the experimental design, cultured both cell lines, carried out all assessments of both AhR- and ER-mediated activities, evaluated data and co-interpreted the results. He participated in the revision and correction of the manuscript.
Paper III.
Michal Bittner participated in the experimental design, carried out the irradiation experiments and assessed the absorbance and AhR-mediated activities, evaluated data and interpreted the results. He wrote the manuscript and finalized the entire article.

Aims of the dissertation thesis
17
Aims of the dissertation thesis
The main goal of this dissertation thesis was to introduce study of HS to centre RECETOX, and especially to study their ecotoxicological properties. Next goal was to contribute by primary results to rather new area of HS-research - direct effects of HS on living organisms, and extend knowledge on their interaction with intracellular receptors (aryl hydrocarbon and estrogen receptor). In the case of Ah receptor, activities of both native and photo-transformed HS were studied.
Studies to be carried out
• Evaluation of AhR-mediated activity of various HS samples and their extracts using H4IIE-luc cells based in vitro bioassay (Paper I and Paper II, chapter 2.3)
• Evaluation of anti/estrogenic activity of various HS samples (Paper II, chapter 2.4)
• Study of changes in absorbance and AhR-mediated activities of HS after irradiation (Paper III, chapter 3.2)

18

Introduction
19
1. Introduction
What will happen when anything dies - bacteria, plants, trees or animals? Majority of dead organic material stay on/in the soil or water and consequently undergo the procedure called humification process. In this process, all the dead organic carbon is transformed via both degradative and condensation polymerization processes. There is no doubt that humification process - together with photosynthesis - belongs to the most important processes on Earth, because humification ensure the stability of the global cycling of energy and materials (Steinberg, 2003). Products created throughout this process are commonly called humic substances (HS). HS have been found in water, soil and even in the air environment as soil dust particles.
Until recently, HS have been regarded to be refractory or inert in the environment, especially in relation to living organisms. In scientific literature, physico-chemical characteristics of HS such as acid status, light absorption and photolytic release of microbial substrate have been mainly discussed. However, there is few papers dealing with effects of HS to organisms, and the works discussing specially the direct effects of HS are still rare.
The role of HS in both water and terrestrial ecosystems is extensive. In the long term - dissolved HS can determine or strongly change the chemistry of whole lakes and act as the most important natural purification system for xenobiotics in waters. In the mid term -HS can act as a source of (in)organic nutrients and rapidly change the bioconcentration and toxicity of xenobiotic and metals, and finally in the short term - HS can act as natural xenobiotics exerting various biological effects, and thus are able to affect aquatic guilds by various modes of action (Steinberg, 2003). In this thesis, the main goals will be focused both on indirect and direct effects of HS to organisms - the ecotoxicological aspects of HS.

Chapter 1.1.
20
1.1. Origin and classification of humic substances (HS)
1.1.1. Humification process and its theories
Process, in which the HS are formed, is traditionally called “humification”. There are two main different pathways in this process - degradative and condensation polymerization processes (Hatcher et al., 1992).
Throughout degradative process, plant and animal biomacromolecules are degraded to smaller molecules. This is realized by microbial activity, readily accessible labile molecules are degraded and completely mineralized while the more stabile molecules are only partially degraded - e.g. polysaccharides and (lipo)proteins. Inert components such as lignin (primarily), cutin, suberin, N-containing aliphatic and other molecules are selectively conserved (Steinberg, 2003).
In the condensation polymerization process - contradictory to the degradative process - lignin, or slightly degraded lignin and other refractory (macro)molecules react with decomposed proteins or peptides and other large organic polymers e.g. polysaccharides and phenol polymers. This type of humification is described by the “ligno-protein theory” (Chefetz et al., 2002; Stevenson, 1994). The core of humic acids (HA) is formed from almost nondegraded lignin macromolecules, while basic structural elements of fulvic acids (FA) are lignin degradation products - monomers or oligomers of sinapyl-, coniferyl- and coumaryl alcohols. This condensation process takes place mainly in soil, and HA from this matrix are mainly leached to waters. In many instances, both condensation and degradation processes occur simultaneously because humification is a dynamic process with no unique unidirectional vector (Hatcher & Spiker, 1988).
However, there are more humification theories discussed in scientific literature, not only ligno-protein theory. For example, there is no primary source of lignin in marine environment. This fact takes into account “Sugar-amine condensation theory” (or “melanoidin model”) which is based on the reaction between monomer reduced sugars and amino acids, while no lignin derivatives are required (Hatcher et al., 1992).

Introduction
21
“The polyphenol model“ (“phenol-protein theory” or “phenol autoxidation theory”, Stevenson, 1994) is based on the reaction of quinones with amines and ammonia as the most important step of the polymerization. By this route nitrogen is incorporated into HS. Quinones originate from microbial degradation of lignins, which leads to phenols and in a subsequent step to quinones.
“Polyunsaturated structure theory” describes the formation of HS from polyunsaturated compounds, such as fatty acids, carotenoids or alkenones as precursors. The reaction proceeds under UV-light between single fatty acids, so that the molecules become cross-linked (Harvey et al., 1983; Harvey et al., 1984). The weakness of this model is that it does not explain the high nitrogen content in HS.
1.1.2. Classification of HS
There are differences in autochthonous and allochthonous humification pathways in freshwaters, which clearly differ in terms of reaction products. HS are an important component of the total organic carbon (TOC), in aquatic environment dominated by dissolved organic carbon/matter (DOC/M), and also called natural organic matter (NOM). DOC can be isolated from filtered water using reverse osmosis. Technically, DOC is composed of: hydrophobic acids, hydrophobic neutral substances, hydrophilic acids, hydrophilic bases, and hydrophilic neutral substances. The hydrophobic compounds are considered to be HS. Fractionation of DOC into hydrophobic and hydrophilic components (Aiken, 1988) is based on the pH dependent adsorption of organic solutions at the hydrophobic surfaces. According to the definition of the International Humic Substances Society (IHSS), only the adsorbed) material is considered to be HS (IHSS, 2004).
The term HS comprises three groups of substances: fulvic acids (FA), humic acids (HA) and humins. The treatment of total HS with dilute alkali dissolves the HA and FA, leaving the undissolved humins behind. After acidification of this alkaline extract, HA precipitate, leaving FA in solution (Fig. 1). Thus, HA is the fraction that precipitates at pH 2 or less, and FA is the fraction that remains in

Chapter 1.1.
solution at pH 2 or less. HA can be further fractionated into hymatomelin, grey, green and brown humic acids, through extraction in alcohol or redissolving in alkaline solution. This differentiation, however, is applied mainly in soil ecology. In the aquatic environment, HS form approximately 50-70% of NOM (Timofeyev et al., 2004), which is in most natural freshwaters in the concentration range 0.5 to 50 mg L-1, but can reach also more than 100 mg L-1 in raised peat bogs (Steinberg, 2003) or pore water (Zsolnay, 1996). HA alone represent only about 6-8% of the total NOM (Yamamoto et al., 2004).
Fig. 1 Scheme for operational definition of HS (Steinberg, 2003).
1.2. Molecular structure of HS
1.2.1. General structural features
HS are polydispersed polyelectrolytes (Orlov et al., 1995; Wagoner et al., 1997), and as such, they share a characteristically high degree of molecular irregularity and heterogeneity. In addition, almost all HS originate from different parent materials, and in consequence, there are no two identical HS molecules in water or soil. This implies that one cannot give a general structural formula for HS. Usually the HS molecule must be described by statistical features: ratios and variations of the various structural entities (Steinberg, 2003).
22

Introduction
Although no general structural formula for HS can be given, lot of studies have been published on the structural formula of their molecular fragments (Ouatmane et al., 2000; Phillips & Olesik, 2003). The aromatic structures, aliphatic chains, carbohydrates, free radicals and peptide linkages, or chinoide structures are easily identified. The acidic nature of HS is attributable to the many carboxylic (-COO-) and phenolic (-OH-) groups, which are not balanced by the basic functional groups such as amines (-NH2). Comparison of representation of main elements and functional groups in HA and FA is summarized in Tab. 1.
1
Aha1bpmcc
Tab. 1 Elemental composition (weight %) and functional groups content(mequ/g) in HA and FA (Schnitzer, 1978).
.2.2. Supramolecular structure
t low concentrations, HS of aquatic origin exist primarily as dissolved ions. At igher concentrations, colloids and precipitates are formed. The same ggregation takes place when they react with cations and protons (Myneni et al., 999). Aggregation and dis-aggregation have consequences for the iogeochemical behavior of HS and for biological activities such as (food) article filtering (Steinberg et al., 2003). The structure of HS studied by use of ulti-dimensional NMR has been presented as a macromolecular aggregate
omposed from mixture of relatively low-molecular weight (<2 kDa) organic ompounds (aliphatic acids, ethers, esters and alcohols, aromatic lignin derived
23

Chapter 1.2.
fragments; polysaccharides and polypeptides) holding together through a complex combination of hydrophobic, charge-transfer and hydrogen bond interactions, and metal bridging (Simpson et al., 2002). Proposed schema of supramolecular structure of HS is displayed on Fig. 2.
Me n +
Aliphatic chains
Polypeptides
Polysaccharides
A romatic lignin fragments
SimigroumiceWerexpe(SECbutasubsdomlinksalkalsplitexplof onhigh
24
Fig. 2 A concept of HS proposed by Simpson et al. (2002). The major components are of relatively low molecular weight and associate in the presence of metals to form an aggregate. Men+ - generic metal cations.
lar supramolecular structure of HS has been proposed by Picollo and his p (Piccolo, 2001, 2002; Piccolo et al., 1996). They propose irregular lles as the supramolecular structures (in contrast to the regular micelles of shaw’s model (Wershaw, 1986). These ideas are developed on the basis of riments in which HS are fractionated by size exclusion chromatography ) after addition of monocarboxylic acid (formic, acetic, propanoic, and
noic acids) to the HS. Building blocks and non-humic material are equently obtained. Hydrophobic interactions have been suggested as the inant bonding mechanism between the individual micelles. These weak explain the change in HS molecular size under alkaline and weakly ine pH conditions. Suggested supramolecular model explains both the ting and aggregate formation of HS, and it could, for example, clearly ain the aging of FA. However, Piccolo’s model is developed from the results ly one method, namely SEC in two modifications - gel chromatography and
performance size exclusion chromatography.

Effect of HS on living organism
25
2. Effect of HS on living organisms (incl. Paper I and II)
With respect to above mentioned chemical constitution of HS, various effects of HS in the natural environment could be expected. Main environmental issues related to HS are summarized in Tab. 2.
Until recently, mainly indirect effects of HS in the ecosystems have been discussed. Indirect effects of HS on organisms include in particular both heavy metals and nutrients control and modulation of toxicity of pesticides and other xenobiotics. These two issues are discussed in more detail in chapter 2.1. However, in the larger sense, all the issues mentioned in Tab. 2 - except the last item - represent possible indirect effects of HS on living organisms. The last item, direct effects of HS on organisms, is discussed in chapter 2.2.

Chapter 2.
Is C L S S N
M W S Pr
S
PhH P R Ec
D
a
i
oo
N
o
o
ea
26
Tab. 2 Environmental issues involving HS (Tipping, 2002).
sue
e
os
rbon cycling
ght penetration into waters
il warming il and water acidification
utrient source
utrient controlicrobial metabolism eatheringil formation (podzolisation)
operties of fine sediments
il structureotochemistry avy metals
sticides, xenobioticsdioactive waste disposal
ystem buffering
irect biological effect Uptake and direct interaction with living organisms
Role of humic substances
Binding, transport, influence on bioavailability, redox reactions
Binding, transport, influence on bioavailabilityBinding and transport of radionuclide ions in groundwaters
Control of proton and metal ion concentrations, persistence
Translocation of dissolved humic substances and associated metals (Al, Fe)
Adsorption at surfaces and alteration of colloidal properties
Aggregating effect on soil mineral solidsMediation of light-driven reactions
Reservoir of carbon, nitrogen, phosphorus, sulphur and chlorine
Binding of iron and phosphateSubstrate for microbesEnhancement of mineral dissolution rates
Major C pool, transformations, transport and accumulation
Absorption and attenuation of light by humic chromophores
Absorption of solar radiation by soil humic matterBinding of protons, aluminium and base cations in soils and waters

Indirect effects of HS on organisms
27
2.1. Indirect effects of HS on organisms
2.1.1. Binding of metals
Pre-eminent among the interactions of HS with solutes are those involving cations, i.e. protons (H+) and metallic cations such as Na+, K+, Li2+, Al3+, Ca2+, Cu2+, Pb2+ and others. Mechanism, in which ionic structures such as carboxyl-, alcohol- and phenolic hydroxyl groups form complexes with metals (Linnik, 1998; Linnik & Nabivanets, 1984) may function only in high metal concentrations, which seldom or never occur in natural freshwaters. At natural HS concentrations, the metal binding partner is not oxygen, but the, as humus constituents quantitatively unimportant, nitrogen atoms and reduced sulfur atoms (Steinberg, 2003). Thus, the complexity of the issue of metal toxicity and bioavailability in aqueous systems derives both from the high number of natural and anthropogenic organic compounds and from the variability of their structures. In fact, the binding capacity and affinity is dependent on the number and type of ligands, on their position in the structure and on the ligand/metal ratio (Calace & Petronio, 2004).
The interactions Men+-HS involve both immobile organic matter in the solid phase of soils and sediments, and dissolved HS and DOM in general, which may be mobile and transport the ions across the soil or waters. Men+-HS interactions thus controls the reactivity of the cation, including its bioavailability (Tipping, 2002). In metal coordination compounds, metal ion serves as the central atom, i.e. electron-pair acceptor, while HS is the electron-pair donor. Such Men+-HS complex structures subsequently form supramolecular structures of HS (Fig. 2, Pacheco et al., 2003; Simpson et al., 2002).
Binding of metals or nutrients by HS can have direct consequences for both water and soil ecosystems (Garcia-Mina et al., 2004; Heil, 2005; Schulten & Leinweber, 2000). For example, Jones and Huang (2003) have demonstrated a significant relationship between amount of HS and bioavailable arsenic. Different concentrations of arsenic ranging from 0.5 to 5 mg L-1 were associated with HS (up to concentration 42 mg L-1). Result was arsenic toxicity reduction by a factor of 100%.

Chapter 2.1.
28
Binding of nutrients with HS can have direct consequences even for mammalian health. Huang et al. (1994) have referred about the effect of HA on thyroidal function. In mice, treatment with low iodine diet significantly increased thyroidal 125I uptake, and additional treatment with HA significantly enhanced the effect of low iodine diet (Huang et al., 1994). Conclusion can be that HA may enhance the goitrogenic effect of low iodine, very likely caused via indirect effect of HA - complexation of iodine by HA. Similar conclusions were also published by Danish scientists (Laurberg et al., 2003).
2.1.2. Binding of organic xenobiotics
Dissolved HS can also physically or chemically interact with organic pollutants in many ways changing many properties of these compounds: partitioning in water (water solubility, Doring & Marschner, 1998; Marschner, 1998), hydrolysis kinetics (Schwarzenbach, 1993), volatilization (Gschwend & Wu, 1985), photolysis rates (Zepp et al., 1981a, b), bioconcentration and toxicity of organic compounds (Bollag & Myers, 1992; Bollag et al., 1992; Cary et al., 1987; Day, 1991; Hodge et al., 1993; Steinberg et al., 1992; Steinberg et al., 1993). Hydrophobic chemicals can be bound with HS by means of various chemical interactions, e.g. occlusion, ion-exchange, hydrogen bonds, charge-transfer (π-π) bonds, covalent bonds, hydrophobic adsorption and partitioning (Martinneto et al., 1994; Piccolo & Celano, 1994; Wijnja et al., 2004). In general, the HS are the most important natural ligands, and the existing stable chemical bonds reduce the toxicity of the particular pollutants. The binding to HS can be initiated through an oxidative coupling reaction catalyzed biologically by polyphenoloxidases and peroxidases or chemically by particular metals and clay minerals. This process can also occur through autoxidation. A coupled reaction requires the release of free radicals, and ends in the formation of C–C–, C–O–, C–N–, and N–N bonds between HS and pollutants (Steinberg, 2003).
For example, Jones and Huang (2003) have studied the interaction between HS extracted from compost NOM and organic pesticide chlorpyrifos. Alternation of pesticides toxicity in the presence of HS in the aqueous phase was measured

Indirect effects of HS on organisms
29
using the Microtox® bioassay. Solutions containing HS in concentrations ranging from 2-42 mg L-1 were added to concentrations of the chlorpyrifos. As a result, pesticide toxicity was reduced from 50 to 100%. These results have demonstrated a significant relationship between HS-pesticide interactions and pesticide toxicity in natural water systems. The same authors subsequently suggested a compost HS addition for contaminated ground waters and surface waters as a mechanism for pesticide toxicity reduction.
The chemical interactions between NOM and chemicals are to a large extent determined by the lipophility of both HS and chemical compound. For compartments such as soil and sediment in which the organic carbon is present in high concentrations as colloids or particles, structural properties in addition to the aromicity are responsible for the accumulation of lipophilic environmental chemicals. A positive correlation is found between the HS aliphaticity, which determines the lipophility, and the partition coefficients organic carbon-water (KOC).
In order to compare the binding capacities of HA and FA, the measurement of KOC for the model compounds benzo[a]pyrene (BaP) and pentachlorophenol (PCP) was performed (DePaolis & Kukkonen, 1997). To study the effect of pH on the binding capacity of HS, KOC values were measured at pH 5, 6.5 and 8. The results have shown that HA have a greater affinity for binding hydrophobic compounds than FA; and change in pH only weakly affects KOC values for the neutral chemical BaP while KOC values for PCP decrease radically with an increasing pH from 5 to 8. Moreover, elemental analysis and spectra indicate that high KOC values can be related to a large aromatic content and to a rather low content of functional groups in the humic structures. These results suggest that only the unionized form of a weak organic acid like PCP can interact with the humic material (DePaolis & Kukkonen, 1997).
The bonding of lipophilic chemicals to HS can also have a kinetic effect. The entrapment and sequestration may result in a gradient of increasingly stable fractions of xenobiotics with simultaneous decrease in bioavailability of the agrochemicals. At the end of this bonding process, covalent bonds are formed with relatively water soluble xenobiotics, such as the triazine herbicide anilazine (Klaus et al., 1998). These covalent bonds can form between ester and ether

Chapter 2.1.
groups via oxygen atoms originating from functional groups of HS (Fig. 3). The depicted model applies not only for soil HS, but also for aquatic HS and shows that anilazine is retained mainly by chemical interactions, charge transfer, and sequestration within the structural voids of the humic material.
30
Fig. 3 Potential major binding interactions between anilazine molecules and aquatic HS (Klaus et al., 1998).
Direct effects of HS on organisms
2.2. Direct effects of HS on organisms
2.2.1. Uptake of HS by organisms
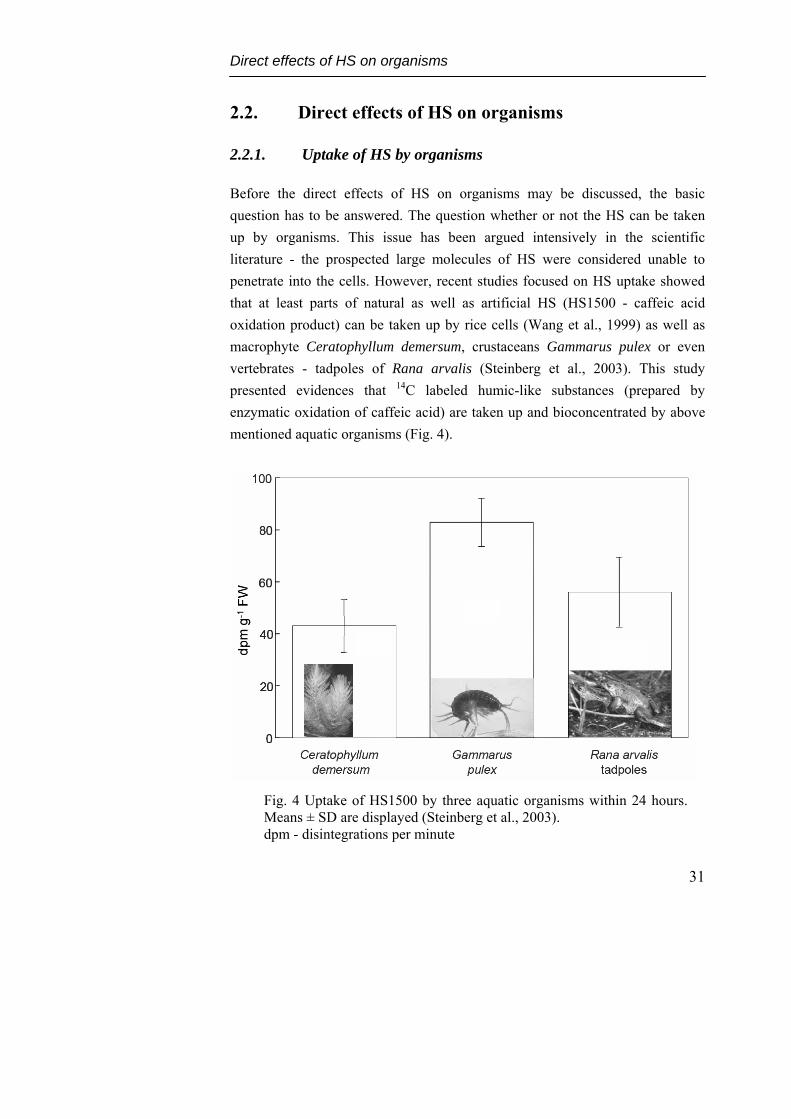
Before the direct effects of HS on organisms may be discussed, the basic question has to be answered. The question whether or not the HS can be taken up by organisms. This issue has been argued intensively in the scientific literature - the prospected large molecules of HS were considered unable to penetrate into the cells. However, recent studies focused on HS uptake showed that at least parts of natural as well as artificial HS (HS1500 - caffeic acid oxidation product) can be taken up by rice cells (Wang et al., 1999) as well as macrophyte Ceratophyllum demersum, crustaceans Gammarus pulex or even vertebrates - tadpoles of Rana arvalis (Steinberg et al., 2003). This study presented evidences that 14C labeled humic-like substances (prepared by enzymatic oxidation of caffeic acid) are taken up and bioconcentrated by above mentioned aquatic organisms (Fig. 4).
31
Fig. 4 Uptake of HS1500 by three aquatic organisms within 24 hours. Means ± SD are displayed (Steinberg et al., 2003). dpm - disintegrations per minute

Chapter 2.2.
32
Furthermore, Beer et al. (2003) have shown that pharmacologically active fractions of fulvic and ulmic (i.e. HA fraction soluble in ethanol) acids can penetrate through human skin and subsequently stimulate response of the spontaneous contractile activity of smooth muscle tissue (chapter 2.2.2.5).
2.2.2. Toxic effects of HS
Per se observable toxicity of natural HS was reported for freshwater snail Lymnea stagnalis. After a 24-h exposure to 0.5 mg L-1 DOC, the activity of the transformation enzyme system was extremely elevated, and Suwannee River NOM and FA even caused death of 10–20% of the exposed snails (Steinberg et al., 2003).
2.2.2.1. Non-specific mechanisms of toxicity
Non-specific markers of toxic effects were studied by analyzing biotransformation enzymes activity and heat shock proteins (HSP). Elevated levels of detoxification enzymes like glutathione S-transferase (GST) and glutathione-peroxidase (GPx) were reported in crustaceans Daphnia magna following HS treatment (Wiegand et al., 2003). Increase in peroxidase activity as well as release of HSP was found also in several amphipod species (Timofeyev et al., 2004) and carp (Wiegand et al., 2003). Exposure to environmentally relevant concentrations of pure HS (0.5 mg L-1) led also to the activation of the microsomal and soluble GST in Ceratophyllum demersum (Pflugmacher et al., 2001).
2.2.2.2. Specific mechanisms of toxicity
There is also evidence that HS can directly affect certain intracellular mechanisms. For example, environmentally relevant concentrations of HS and NOM have induced modulation of photosynthetic oxygen release in alga

Direct effects of HS on organisms
33
Scenedesmus armatus, water moss Vesicularia dubyana and hornwort Ceratophyllum demersum (Fig. 5, Pflugmacher et al., 2006; Pflugmacher et al., 1999; Steinberg et al., 2003).
Influence of HS on sodium metabolism in Daphnia magna was shown by Glover et al. (2005). Environmentally relevant levels of Suwannee River NOM and Aldrich HA (HAA) were observed to significantly enhanced sodium transport in the crustacean. This effect was described as an uncompetitive stimulation of sodium influx, characterized by an increased maximal sodium transport rate, accompanied by a decreased uptake affinity. In the subsequent study, two above mentioned types of HA were compared and mechanism of action was proposed (Glover & Wood, 2005). At pH 4 HAA promoted linear sodium uptake kinetic relationship, attributed to altered membrane permeability due to enhanced membrane binding of HS at low pH. In contrast, a NOM sample elicited no consistent action on sodium influx. These results suggest that impacts on sodium metabolism may be limited only to certain types of humic materials (here HAA), which is nevertheless in accordance with predication on heterogeneous properties of HS.

Chapter 2.2.
Fig. 5 Photosynthetic oxygen production in Ceratophyllum
demersum
after exposure to DO
C sam
ples (0.5 mg L
-1
DO
C) of various origins, show
ing in most cases significant (p=0.05) inhibition (Pflugm
acher et al., 2006).
34

Direct effects of HS on organisms
2.2.2.3. HA - possible etiological agens of Blackfoot disease
HA has also been proposed as a factor that causes Blackfoot disease (BFD), an endemic peripheral vascular disease prevailing in the southwest coast of Taiwan, where well water is rich in HS (Huang et al., 1994). In connection with BFD, there are discussed two main possible etiological factors - arsenic and HA. However, only the possible influence of HS on promotion of BFD will be further discussed.
A possible mechanism leading to anemia in BFD can be damage of human red blood cells (RBCs) by HA-caused oxidative stress, which was manifested as reduction of RBCs deformability and hemolysis (Cheng et al., 1999). HA caused lipid peroxidation in a dose-dependent manner (Fig. 6), and such changes were accompanied by a depletion of glutathione and a reduction in activities of the antioxidant enzymes including catalase, superoxide dismutase, and glucose-6-phosphate dehydrogenase (Cheng et al., 1999).
Rel
ativ
eflu
ores
cenc
e in
tens
ity (%
)
HA concentration µg/mL
HA + Vitamin EHA only
Rel
ativ
eflu
ores
cenc
e in
tens
ity (%
)
HA concentration µg/mL
HA + Vitamin EHA only
Rel
ativ
eflu
ores
cenc
e in
tens
ity (%
)
HA concentration µg/mL
HA + Vitamin EHA only
35
Fig. 6 Lipid peroxidation of C11-fluor-loaded RBCs on exposure to HA in the presence or absence of vitamin E. The lipophilic dye molecules are taken up by RBCs and incorporated into the phospholipid bilayer. On oxidation, the probe molecules turn into nonfluorescent species. Means ± SD are displayed (Cheng et al., 1999).

Chapter 2.2.
36
Other possible mechanisms of HA-induced disorders connected with BFD pathogenesis are suggested further. Cheng et al. (2003) have shown that in vitro HA treatment caused growth arrest and apoptosis in human foreskin fibroblasts. This was accompanied by a significant increase in the level of 8-hydroxy-2'-deoxyguanosine (product of oxidative damage to DNA) in cellular DNA. The increased fluorescence in dichlorofluorescin-stained HA-treated cells again suggests the involvement of reactive oxygen species in HA-induced biological effects.
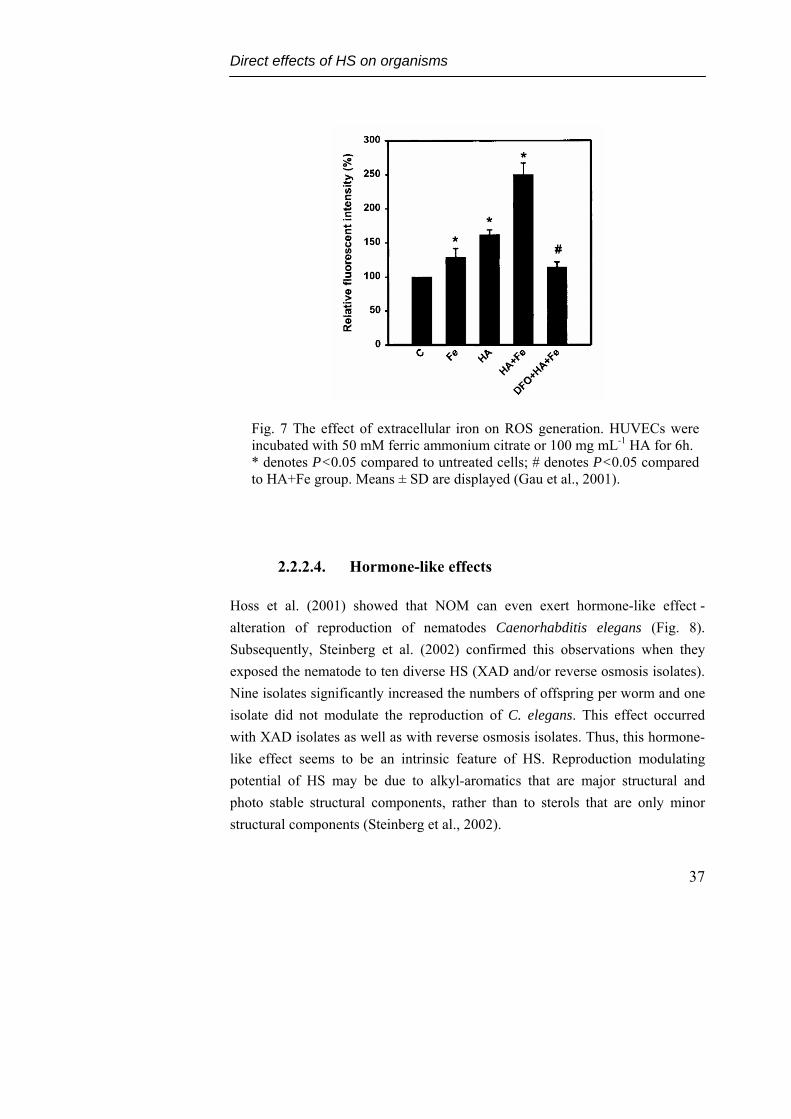
Induction of oxidative stress by HA through increasing intracellular iron could be possible mechanism leading to atherothrombotic vascular disorder in BFD (Gau et al., 2001). HA were observed to enhance the expression of ferritin, induce intracellular chelatable iron and reduce the expression of transferrin receptor in human umbilical vein endothelial cells (HUVECs). Pretreatment of HUVECs with desferrioxamine (iron chelator) inhibited HA-mediated increases of ferritin synthesis and intracellular chelatable iron, but caused recovery of the inhibitory effect on transferrin receptor. Co-treatment with iron and HA induced more ROS and intracellular chelatable iron than iron or HA treatment alone (Fig. 7, Gau et al., 2001).
Direct effects of HS on organisms
Fig. 7 The effect of extracellular iron on ROS generation. HUVECs were incubated with 50 mM ferric ammonium citrate or 100 mg mL-1 HA for 6h. * denotes P<0.05 compared to untreated cells; # denotes P<0.05 compared to HA+Fe group. Means ± SD are displayed (Gau et al., 2001).
2.2.2.4. Hormone-like effects
Hoss et al. (2001) showed that NOM can even exert hormone-like effect - alteration of reproduction of nematodes Caenorhabditis elegans (Fig. 8). Subsequently, Steinberg et al. (2002) confirmed this observations when they exposed the nematode to ten diverse HS (XAD and/or reverse osmosis isolates). Nine isolates significantly increased the numbers of offspring per worm and one isolate did not modulate the reproduction of C. elegans. This effect occurred with XAD isolates as well as with reverse osmosis isolates. Thus, this hormone-like effect seems to be an intrinsic feature of HS. Reproduction modulating potential of HS may be due to alkyl-aromatics that are major structural and photo stable structural components, rather than to sterols that are only minor structural components (Steinberg et al., 2002).
37

Chapter 2.2.
Fig. 8 Modulation of numbers of offsprings in Caenorhabditis elegans exposed to a raised peat bog FA. Means ± SD are displayed (Hoss et al., 2001).
Nevertheless, hormone-like effect of HS is not restricted to the nematodes only (Steinberg et al., 2004), similar effects were observed even with fish (Meinelt et al., 2004) and amphibians (Lutz et al., 2005).
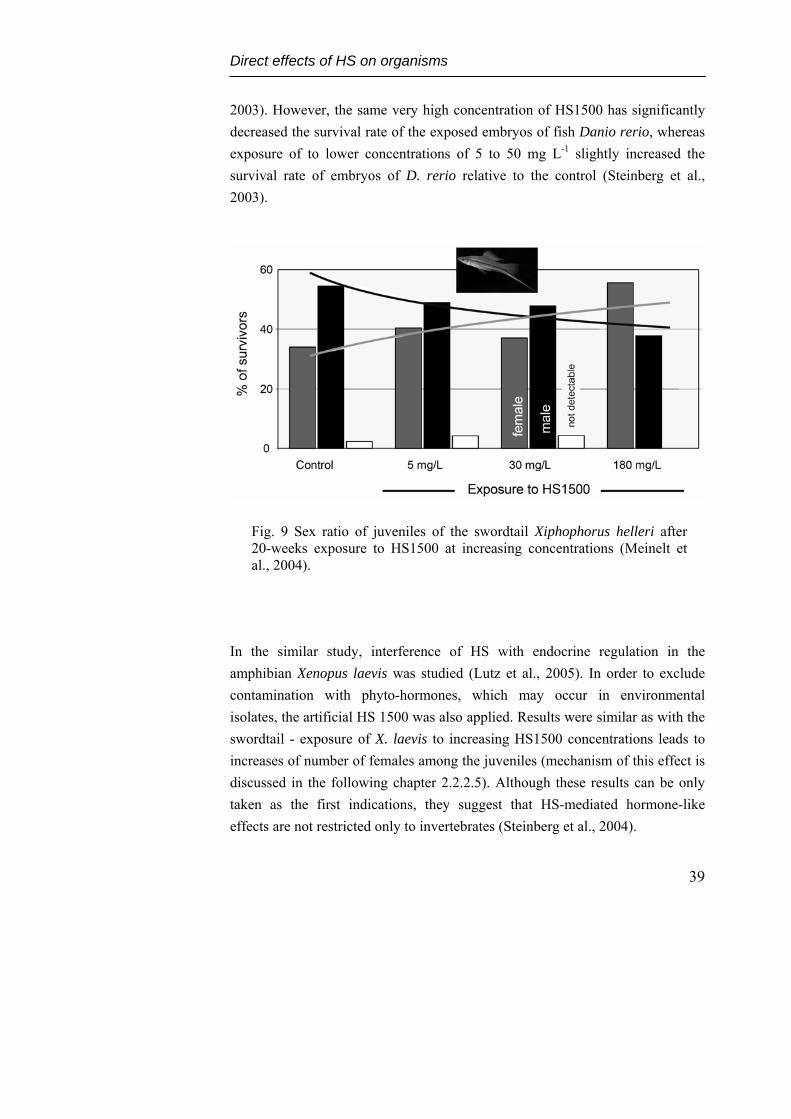
In Meinelt et al. (2004) study, hormone-like effects of HS1500 was studied on swordtail fish Xiphophorus helleri, an ornamental fish which is well known for its hormone inducible sex reversal. In the experiment, two week old juveniles were exposed to increasing HS1500 concentration for 20 weeks, until the sex of most of the juvenile fishes could be identified. Results have shown there is a tendency of feminization with increasing concentrations of HS1500 (Fig. 9). The exposure of 180 mg L-1 HS1500 (unrealistic high concentration) reverses the dominance of male over female, which exists up to 30 mg L-1 HS1500. In addition, this long term exposure to high concentration of HS1500 led also to body mass increase in both males and females (Meinelt et al., 2004), while survival of fry has not decreased even at exposure to 500 mg L-1 (Steinberg et al.,
38
Direct effects of HS on organisms
2003). However, the same very high concentration of HS1500 has significantly decreased the survival rate of the exposed embryos of fish Danio rerio, whereas exposure of to lower concentrations of 5 to 50 mg L-1 slightly increased the survival rate of embryos of D. rerio relative to the control (Steinberg et al., 2003).
Fig. 9 Sex ratio of juveniles of the swordtail Xiphophorus helleri after 20-weeks exposure to HS1500 at increasing concentrations (Meinelt et al., 2004).
In the similar study, interference of HS with endocrine regulation in the amphibian Xenopus laevis was studied (Lutz et al., 2005). In order to exclude contamination with phyto-hormones, which may occur in environmental isolates, the artificial HS 1500 was also applied. Results were similar as with the swordtail - exposure of X. laevis to increasing HS1500 concentrations leads to increases of number of females among the juveniles (mechanism of this effect is discussed in the following chapter 2.2.2.5). Although these results can be only taken as the first indications, they suggest that HS-mediated hormone-like effects are not restricted only to invertebrates (Steinberg et al., 2004).
39

Chapter 2.2.
40
2.2.2.5. Interaction of HS with intracellular receptors
To reveal mechanisms of HS mediated effects observed on whole organisms or tissues, the scientists have also focused on interaction of HS with intracellular receptors. Beer et al. (2000) has described partial agonistic effects of aqueous peat extract and the water-soluble components of FA on the α2-adreno and D2 dopamine receptors. In another studies focused on mammalian receptors, HS have been reported to be involved in gene regulation e.g. inhibiting lipopolysaccharide-induced NF-кB activation in human umbilical vein endothelium cells (Gau et al., 2000), and inducing PPARγ activation in preadipocytes (Lee et al., 1999).
Mechanism of upper mentioned xenoestrogenity of HS1500 (chapter 2.2.2.4) has already been proposed (Lutz et al., 2005). Semi-quantitative RT-PCR has revealed a marked increase of estrogen receptor mRNA in tadpoles exposed to HS1500. Furthermore, preliminary RT-PCR results showed that the thyroid-stimulating hormone has also increased after exposure to HS1500, indicating a weak adverse effect on thyroid hormones availability (Lutz et al., 2005). These findings suggest that HS can act as environmental signal molecules. This assumption was supported by identification of significant transcriptional changes in Caenorhabditis elegans chemosensors, olfactory receptors, as well as enzymes of the biotransformation system after the exposition to HS (Menzel et al., 2005). Finally, recent in vivo experiments on Amazonian fish tambaqui Colossoma macropomum have clearly shown induction of biotransformation enzyme CYP1A, a biomarker of exposure to pollutants that can activate AhR, after the exposure to HAA and NOM samples (Matsuo et al., 2006). These last results are in a good accordance with our in vitro results discussed in following chapter 2.3.

Activation of Ah receptor by HS
41
2.3. Activation of Ah receptor by HS (Paper I and II)
In these studies, we have investigated a possible biochemical mechanism of HS toxicity via activation of the intracellular aryl hydrocarbon receptor (AhR). AhR mediates the toxic and biological effects of environmental contaminants such as 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD), but a number of structurally diverse compounds has also been found to activate AhR. For better understanding why we have studied AhR-mediated activity of HS, the issue of AhR is further discussed in more detail.
2.3.1. Aryl hydrocarbon receptor (AhR)
AhR belongs among the nuclear receptors that play a key role in toxicity of many persistent organic pollutants (POPs). This cellular protein is located in cytosole until its interaction with the ligand. After activation it enters the nucleus, binds to specific DNA sequences and triggers expression of corresponding genes and consequently causes toxic effects (Janosek et al., 2006).
2.3.1.1. Mechanism of AhR-mediated effects
As a primary target for coplanar aromatic substances (pertaining to the POPs group), the AhR belongs among the most extensively studied nuclear receptors (particularly in studies of environmental xenobiotics, Janosek et al., 2006). The primary biochemical responses to AhR activation are induction of drug metabolizing monooxygenases CYP1A1, CYP1A2 and CYP1B1 (enzymes participating in biotransformation phase I) as well as phase II enzymes such as GST, NADPH-quinone oxidoreductase, xanthinoxidase, etc. (Reen et al., 2002). However, CYP enzymes are playing a key role not only in xenobiotics degradation, but may greatly enhance their toxic and/or mutagenic potency (e.g. metabolic activation of PAH). Beside activation of CYPs, other effects such as modulation of specific cellular signaling pathways are considered another

Chapter 2.3.
42
molecular mechanism of AhR-mediated toxicity. AhR-dependent events include several chronic adverse health effects such as endocrine disruption, changes in proliferation activity, neurotoxicity, embryotoxicity, immunotoxicity as well as carcinogenicity (Parzefall, 2002).
2.3.1.2. Ligands of AhR
The spectrum of chemicals that bind to AhR and subsequently activate the AhR signal transduction pathway, and the identity of materials containing AhR active chemicals is only now being defined. The activators (ligands) of AhR could be divided into two groups - “classical” and “non-classical” ligands - with respect to the structure and affinity to the AhR.
"Classical" AhR ligands and inducers
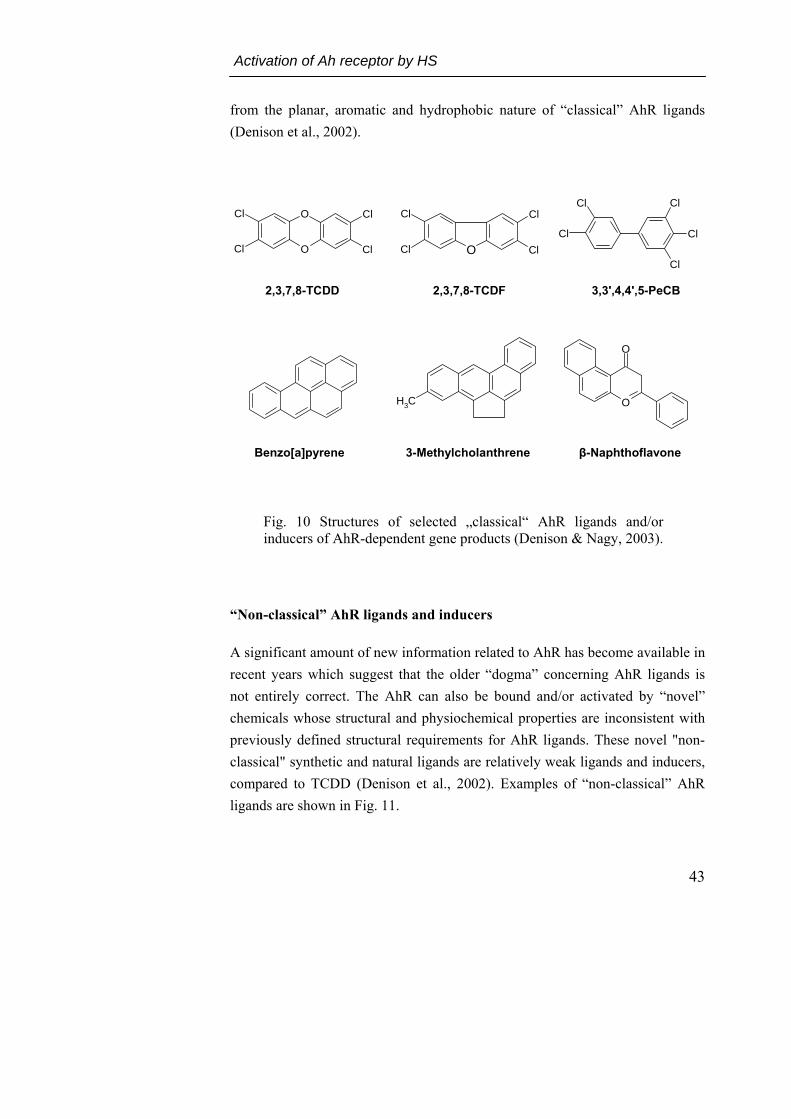
The best studied ligands for the AhR are synthetic environmental contaminants, which include both halogenated aromatic hydrocarbons (HAH), such as the polychlorinated dibenzo-p-dioxins (PCDD), dibenzo furans (PCDF) and biphenyls (PCB), and polycyclic aromatic hydrocarbons (PAH, Fig. 10, (Safe, 1990). HAH have a relatively high binding affinity for the AhR (in the pM to nM range) whereas the PAH have a significantly lower affinity (in the high nM to mM range, Denison et al., 2002). Exposure to these HAH, including 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD, the most potent AhR-activator), produces a wide variety of species- and tissue-specific toxic and biological effects mentioned above.
The physiochemical and structural information derived from analysis of HAH/PAH AhR ligands - i.e. compounds possessing similar characteristic structural features - has provided valuable insights into AhR ligand structure (Waller & McKinney, 1995). However, this structural information was obtained by studies examining the binding of only "classical" ligands (primarily HAH and PAH) to AhR (Safe, 1990). These preconceptions have hampered the search for novel classes of exogenous and endogenous ligands, which may deviate greatly
Activation of Ah receptor by HS
from the planar, aromatic and hydrophobic nature of “classical” AhR ligands (Denison et al., 2002).
Cl
O
O Cl
Cl Cl O
Cl
Cl
Cl
Cl
CH3
Cl
Cl
Cl
Cl
Cl
O
O
2,3,7,8-TCDD 3,3',4,4',5-PeCB 2,3,7,8-TCDF
Benzo[a]pyrene 3-Methylcholanthrene β-Naphthoflavone
“No
A sirecenot chemprevclasscomligan
Fig. 10 Structures of selected „classical“ AhR ligands and/or inducers of AhR-dependent gene products (Denison & Nagy, 2003).
43
n-classical” AhR ligands and inducers
gnificant amount of new information related to AhR has become available in nt years which suggest that the older “dogma” concerning AhR ligands is entirely correct. The AhR can also be bound and/or activated by “novel” icals whose structural and physiochemical properties are inconsistent with
iously defined structural requirements for AhR ligands. These novel "non-ical" synthetic and natural ligands are relatively weak ligands and inducers, pared to TCDD (Denison et al., 2002). Examples of “non-classical” AhR ds are shown in Fig. 11.

Chapter 2.3.
SN
CH3
S
S
O
NH
O
O
Ch3
Ch3 N
CH3CH3
OMe
CH3O
N
NSO
CH2
OCONH CH3
N N NH
CN
F3C
CH3 CH3
CH3
O
CH3CH3
CH3
O
CH3 CH3
CH3 CH3
O
O
OO
CH3 CH3 O
O
CH3
O
O
CH3 CH3
CH3
O
O
O
O
O
CH3
OH
O
CH3
NH2
NH2
YH439
2,3-Diaminotoluene Canthaxanthin
SKF 71739 Omeprazole
Carbaryl Brevetox (PbTX-6)in
Fig. 11 Structures of selected „non-classical“ AhR ligands and/or inducers of AhR-dependent gene products (Denison & Nagy, 2003).
Overall, the ability of a wide range of structurally diverse "non-classical" chemicals to bind AhR, activate AhR dependent DNA binding and/or induce CYP1A1 gene expression, clearly demonstrates that a greater spectrum of chemicals can interact with and activate this receptor than previously thought (Denison & Nagy, 2003). Moreover, experiments with extracts of dietary herbal supplements (e.g. ginseng, white oak bark, licorice, ginkgo biloba, etc.), vegetables and fruits (corn, jalapeno pepper, tomato, etc.) demonstrate that dietary products can be a major source of naturally occurring AhR ligands to which animals and humans are chronically exposed (Jeuken et al., 2003).
44

Activation of Ah receptor by HS
45
2.3.1.3. Consequences of exposure to weak AhR ligands
Many of the toxic effects of TCDD are not observed until several days to weeks following chemical exposure to weak AhR ligands (Devito & Birnbaum, 1995). The adverse effects of these chemicals likely result from the continuous and inappropriate expression of specific genes in responsive cells which ultimately results in the delayed toxic responses. Implicit in this hypothesis is the concept that if the concentrations of PAH or other relatively weak AhR ligands are maintained at appropriately high levels within an organism, toxic effects similar to that caused by TCDD should be observed (Denison & Perdew, 2002).
Consistently with this hypothesis, an earlier study reported that juvenile catfish continually exposed for ninety days to high dietary levels of β-naphthoflavone (prototypical weak PAH AhR ligand, Fig. 10), exhibited a variety of morphological and toxicological effects similar to those produced by exposure to TCDD (Grady et al., 1992).
Although significant advance in the field over the past ten years have clearly defined the role of AhR in the toxic and biological effects of HAH and PAH, the exact biochemical events which lead to the spectrum of species- and tissue-specific toxic responses to these chemicals still remain to be elucidated (Denison & Perdew, 2002).

Chapter 2.3.
46
2.3.2. Results - AhR-mediated activity of HS (Paper I + first part of Paper II )
In these studies, alkali solutions of HA were prepared, and subsequently, lipophilic compounds (including parts of HA) were extracted by liquid-liquid extraction with hexane/dichloromethane. Organic extract of HA was further treated with sulfuric acid to study the role of possible trace persistent contaminants. In vitro dioxin-like activities of obtained extract and HA-solutions have been evaluated using H4IIE-luc cells by determining the induction of AhR-dependent reporter luciferase.
2.3.2.1. HS used in experiments
Regarding the indefinable molecular structure of HS resulting from more or less unique materials and processes participating in the humification process (chapter 1.1.1), the selection of appropriate HS samples for experiments was crucial. Because of reproducibility of our results, we have decided to examine ecotoxicological properties mainly of commercially available HS samples, namely HA, FA and NOM.
Tested substances were isolated from different matrices, and purchased from various sources: HA-Fluka (Prod. No. 53680) from Fluka, Switzerland, HA Sodium Salt (Prod. No. H16752) from Sigma Aldrich, USA, both this HA were derived from coal. The following reference HS were purchased from IHSS (International Humic Substances Society) USA: Suwannee River HA (Prod. No. 2S101H), Florida Peat HA (Prod. No. 1S103H), Waskish Peat HA (Prod. No. 1R107H), Elliot Soil HA (Prod. No. 1S102H), Leonardite HA (Prod. No. 1S104H), Suwannee River FA (Prod. No. 1S101F), Florida Peat FA (Prod. No. 2S103F), Nordic Aquatic FA (Prod. No. 1R105F), Suwannee River NOM (Prod. No. 1R101N), and Nordic Reservoir NOM (Prod. No. 1R108N).
We have also studied HA derived from three types of uncontaminated soils from the Czech Republic: forest soil, permanent grass soil and arable soil. Isolation of

Activation of Ah receptor by HS
these HA was carried out according to IHSS guideline published on internet (IHSS, 2004) - except the last step - dialysis of HA solution against distilled water.
2.3.2.2. Paper I
In this study, we have studied AhR-mediated activity of only one kind of HS - HA-Fluka (i.e. its alkali solution), and its organic extract, alkali HA remaining after LLE and H2SO4-treated organic extracts. This type of "fractionation" is shown in Fig. 12. The three greatest tested concentrations of pure (alkali) HA-Fluka (5, 15 and 50 mg L-1) significantly increased luciferase reporter gene activity in H4IIE-luc cells (Fig. 13) in a dose dependent manner. Activity was related to reference TCDD and resulting relative potency (REP) of HA
determined by luciferase induction assay equaled 6·10-8.
solution in 0.05M NaOH
LLE
HS
alkali HS
remaining alkali HS
HS-extract
H2SO4-treatedHS-extract
LLE+H2SO4
solution in 0.05M NaOH
LLE
HS
alkali HS
remaining alkali HS
HS-extract
H2SO4-treatedHS-extract
LLE+H2SO4
HS
alkali HS
remaining alkali HS
HS-extract
H2SO4-treatedHS-extract
LLE+H2SO4
Fig. 12 Preparation scheme of examined alkali HS solution, organic extract of alkali HS, alkali HS remaining after the extraction, and sulfuric acid treated organic HS extract.
47

Chapter 2.3.
TCDDalkali HAalkali HA remaining after LLEHA-extractH2SO4-treated HA-extract
(% o
f TC
DD
max
ind
uctio
n)
Concentration (mg L-1) 10-8 10-7 10-6 10-5 10-4 10-3 1 10 100
100
60
80
20
40
0
Luci
fera
se a
ctiv
ity
H2SO4
Orgathe opreselucifLLEbut sactivassusolutcom
Our activis nenon-chem
48
Fig. 13 Luciferase activity induced by TCDD and various HA fractions/solutions. The results are related to max. luciferase induction of TCDD. Means ± SD of triplicate determinations are displayed.
nic extract of HA-Fluka elicited practically identical luciferase response as riginal alkali HA (Fig. 13), indicating that almost all of the AhR-activators nt in HA-Fluka were extracted into the organic phase. Induction of
erase activity at maximal tested concentration of "alkali HA remaining after " was about 3.5-fold less than the activity induced by the original alkali HA, till significantly greater than control sample (Fig. 13). However, dioxin-like ity of alkali HA and HA-extract were practically identical. Accordingly, we me that removal of extracted parts of HA has broken the balance in HA ion and thus other AhR-activating compounds could be released from the plex HA.
results showed that an alkali solution as well as organic extract of HA is e in both EROD and luciferase assays while H2SO4-treated extract activity gligible. This findings suggest that AhR-mediated activity is caused by
persistent both hydrophilic and lipophilic HS-compartments. In addition, ical analyses have revealed only non-significant levels of known

Activation of Ah receptor by HS
AhR-inducing contaminants (16 U.S. EPA priority PAH and seven indicator PCB were analyzed using GC-MS). Thus, the results suggest that HA-Fluka or their fragments can elicit significant inductions of AhR mediated effects in vitro. Nevertheless, to decide if this effect can be regarded as a general quality of HS, more HS samples had to be examined (Paper II).
2.3.2.3. Paper II - part focused on AhR-mediated activity of HS
On the basis of the results published in Paper I, we have focused on AhR-mediated activity of wider spectra of commercially available HS. Twelve HS samples were chosen, comprising seven HA samples, three FA samples and two NOM samples (described in chapter 2.3.2.1). Of the twelve HS tested, five elicited a significant AhR-mediated activity in H4IIE-luc cells (Fig. 14).
0
20
40
60
80
100
TCDD
HA - Fluk
a
HA sodiu
m salt
Suwann
ee R
iver H
A
Elliott S
oil H
A
Florida P
eat H
A
Leon
ardite
HA
Waskish
Peat H
A
Suwann
ee R
iver F
A
Florida P
eat F
A
Nordic
Aquatic
FA
Suwann
ee R
iver N
OM
Nordic
Reservo
ir NOM
Luci
fera
se a
ctiv
ity
(% o
f 100
pM
TC
DD
indu
ctio
n)
HS - Humic Substances NOM - Natural Organic Matter
FA - Fulvic AcidsHA - Humic Acids
Fig. 14 AhR-mediated activities of twelve HS determined in H4IIE-luccells; cTCDD= 3.2 x 10-5 mg L-1; cHS=150 mg L-1. Means ± SD of triplicate determinations are displayed.
49

Chapter 2.3.
50
All of the active samples were HA, while no FA and no NOM showed any substantial activity. Relative potencies (related to TCDD) of all the AhR-active HA were in a relatively narrow range of 2.6 – 8.4 x 10-8. All of the active substances elicited significant AhR-mediated effects at concentration as low as 1.9 mg L-1, except for Florida Peat HA (LOEC 16.7 mg L-1).
All AhR-active (alkali) HA were subsequently fractionated in the same way as described in the Paper I (Fig. 12). The organic extracts of AhR-active HA induced significant activities that confirmed presence of non-polar AhR activators. However, compared to the Paper I, the extraction residues elicited relatively great responses as well. Luciferase activities induced by the residues were even greater than those of organic extracts (Tab. 3), except for HA-Fluka and HA Sodium Salt. This result suggests a significant contribution of polar compounds to the AhR-mediated activity of HA.
Extraction with H2SO4 totally suppressed the AhR-mediated activities of all HA. Thus it can be concluded that the compounds responsible for the observed effects were not persistent organic compounds such as PCDD, PCDF or PCB. Moreover, chemical analysis of seven indicator PCB congeners and 16 U.S. EPA priority PAH (indication of possible anthropogenic contamination) did not reveal sufficient quantities of these substances that could be responsible for the relatively great AhR-mediated activities of HA. Toxic equivalents (TEQ) calculated from chemical data for PAH using REPs published by (Machala et al., 2001) was less than 1% of bioassay-derived TEQ for the HA.

Activation of Ah receptor by HS
ToooUnthca(2
Aabh
Tab. 3 Mass balance of extraction procedure
alkali solution extract extr.residue
HA - Fluka 205 70.4 22.2HA sodium salt 219 67.8 31.1Elliot Soil HA 197 37.5 47.1Florida Peat HA 29 w.i. 55.0
(ng TEQ g-1 HA) (% of alkali
solution) (% of alkali
solution)
Leonardite HA 68 30.4 46.1
TEQ – toxic equivalents derived from EC25 obtained from experiments with alkali solutions, their organic extracts and extraction residues; w.i. - weak induction.
reatment with H2SO4 (Fig. 12) totally suppressed the AhR-mediated activities f all HA. Thus it can be concluded that the compounds responsible for the bserved effects were not persistent organic compounds such as PCDD, PCDF r PCB. Moreover, chemical analysis of seven indicator PCB congeners and 16 .S. EPA priority PAH (indication of possible anthropogenic contamination) did ot reveal sufficient quantities of these substances that could be responsible for e relatively great AhR-mediated activities of HA. Toxic equivalents (TEQ) lculated from chemical data for PAH using REPs published by Machala et al., 001) were less than 1% of bioassay-derived TEQ for the HA.
ll the upper mentioned results demonstrate that some HS, particularly HA, are le to activate AhR signaling pathway due to presence of both lipophilic and
ydrophilic AhR activators, while presence of persistent HAH was excluded.
51

Chapter 2.4.
52
2.4. Anti/estrogenic activity of HS (Paper II)
In Paper II we have also studied anti/estrogenic effects of twelve commercially available HS (described in chapter 2.3.2.1) using MVLN cell line stably transfected with luciferase gene under control of estrogen receptor (ER). ER is a target for natural estrogen hormones, but can also be affected by so called “xenoestrogens”, i.e. compounds that belong to the group of endocrine disruptors. For better understanding why we have studied ER-mediated activity of HS, the issue of ER is further discussed in more detail.
2.4.1. Estrogen receptor (ER)
ER plays a key role in female hormone regulation and signaling because mediates the signal transfer of estrogen hormones. Although estrogens are produced mainly in female organisms, mammalian ER were localized in both sexes in numerous tissues (breast, ovaries, brain, liver, bone, cardiovascular system, adrenals, testis, prostate, urogenital or gastrointestinal tract (Jacobs et al., 2003). The major mammalian endogenous estrogen hormones are 17-β-estradiol (E2), estrone and estriol, which are produced to the greatest extent in ovarian cells. Estrogens are responsible for female secondary sex characteristics and regulation of reproduction. E2 stimulates proliferation and growth in the reproductive tract organs, causes the development of the endometrium of the uterus, and influences libido. Estrogens also help maintain pregnancy and prepare the breasts for lactation (Lintelmann et al., 2003). Nevertheless, regarding the presence of ER in males too, abnormal exposure of males to exogenous estrogen-like acting molecules (such as environmental contaminants) can cause a large spectrum of negative effects (chapter 2.4.1.3, Janosek et al., 2006).
The endocrine system is highly conserved in the animal kingdom and estrogens were found to regulate sex characteristics of all vertebrates such as birds, reptiles, amphibians, fish, but also even certain invertebrates such as e.g. mollusks. Nevertheless, inter-species differences exist that may affect sensitivity and response to endocrine disruptors (Lintelmann et al., 2003).

Anti/estrogenic activity of HS
53
2.4.1.1. Mechanism of ER-mediated effects
The mechanism of ER action is similar to that of AhR. In fact, it differs only in DNA binding domain and chaperon proteins – instead of HSP 90 and ILP (for AhR), the DNA binding domain of ER is masked by proteins like HSP70 and/or p60 (Massaad et al., 2002). In mammals, two subtypes of ER have been characterized by molecular cloning, ERα and ERβ. Another subtype ERγ possibly exists in fish (Drummond et al., 2002).
Xenoestrogens belong to the group of endocrine disruptors, i.e. chemicals that can mimic endogenous hormones or interfere with endocrine processes (Singleton & Khan, 2003). Thus, xenoestrogens can exert their effect not only by direct binding to ER and regulating the activity of estrogen-responsive genes, but they can also act through ER-independent mechanisms (Bettin et al., 1996; Zacharewski, 1998). Mode of action of some xenoestrogens can be via inhibition of important enzyme-catalyzed reactions of steroidogenesis. For instance, biosynthesis of estrogens includes the conversion of testosterone to estrogen catalyzed by the enzyme aromatase, and xenobiotic such as tributyltin can inhibit this enzyme, leading to higher testosterone concentrations and to lower estrogen concentrations (Bettin et al., 1996). Interactions of xenobiotics with hormone-binding plasmatic proteins or cross-talk between ER and other receptors as well as disruption of hypothalamo-pituitary axis have also been described (Sovadinova, 2006).
Other ER-independent modes of action of xenoestrogens can be described on the example of TCDD as an exogenous agonist for the AhR (discussed in detail in chapter 2.3.1). Though this receptor is not directly involved in hormone metabolism, its activation can have various influences on the endocrine system by: 1) an increased degradation rate of ER (down-regulation of their levels), 2) induction of E2 metabolizing enzymes, and 3) inhibition of gene expression controlled by E2 or growth promoters (Gillesby & Zacharewski, 1998; Lintelmann et al., 2003; Safe et al., 1998).

Chapter 2.4.
2.4.1.2. Anti/estrogenic compounds
Endocrine disruptors modulating the ER-signaling pathway can be classified into two groups: xenobiotic estrogens and antiestrogens, according to that if the chemicals mimic or antagonize effects of natural estrogen hormones (Sonnenschein & Soto, 1998). Both mentioned groups contain structurally different compounds (Fig. 15 and Fig. 16 - except E2 being natural ER ligand) which demonstrates that a greater spectrum of chemicals can interact with ER (Freyberger & Schmuck, 2005).
OH
CH3
CH3
CH3
OHOH
Cl
Cl
ClClCl
OH
OH
OH
O
OH
OH
OHO
CH2CH3
CH2CH3
OHOH
17-β-Estradiol (E2) Diethylstilbestrol Bisphenol A
Genistein Resveratrol o,p′-DDT
H
54
Fig. 15 Structures of selected potent (17-β-Estradiol, Diethylstilbestrol) and putative (the others) estrogens (Freyberger & Schmuck, 2005).
Tamoxifen
ON
C3
CH3 CH3
ICI 182 780
OH
OH (CH2)9SO(CH2)3CF2CF3
Fig. 16 Structures of selected antiestrogens (Freyberger & Schmuck, 2005).

Anti/estrogenic activity of HS
55
2.4.1.3. Consequences of exposure to xenoestrogens
Exposure to specific xenoestrogenic chemicals both of natural or synthetic nature has been related to cause numerous adverse health effects assessed by both laboratory and field studies. The major impairments include reproduction toxicity, increased incidence of breast cancer, male testes and uterus tumors, delayed male puberty, decreased semen quality and volume, developmental anomalies of the male reproductive system including reduced secondary sex organs size, hypospadias, cryptochordism and enhanced susceptibility to seminomas (Gillesby & Zacharewski, 1998; Mantovani et al., 1999).
However, the reality is such that wildlife is rarely, if ever, exposed to single chemicals, but instead it is exposed to highly complex mixtures of chemicals, including many that are endocrine active in various ways (see chapter 2.4.1.1). Thus, if we would like to prognose all resulting effects of xenoestrogens, we need to understand much better how chemicals interact, and what overall effects will occur upon exposure to such mixtures. Finally, we need to move from assessing effects at the individual organism level, to understanding the consequences of these effects at the population level. (Sumpter, 2003).
2.4.2. Results - ER-mediated activities of HS (Paper II)
Evaluation of anti/estrogenic activity of twelve HS samples (chapter 2.3.2.1) was carried out using MVLN cell line based bioassay. This bioassay (by analogy to H4IIE-luc cells) use cells stably transfected with luciferase gene that is under control of ER. In our experiments, none of the tested HS showed any estrogenic activity both in absence and presence of ER-agonist E2. However, ten out of twelve tested HS caused significant antiestrogenic effect (Fig. 17). Compared to AhR-mediated activity, these antiestrogenic effects were observed not only after HA treatment, but also after FA and NOM treatment. Concentrations causing a 50% decrease of E2-dependent luciferase activity (IC50) were observed for HS in a concentration range of 40-164 mg L-1 (Tab. 4).

Chapter 2.4.
0
20
40
60
80
100
120
E2
Humic
Acid - F
luka
HA Sod
ium Salt
Suwann
ee R
iver H
A
Elliot S
oil H
A
Florida P
eat H
A
Leon
ardite
HA
Waskish
Peat H
A
Suwann
ee R
iver F
A
Florida P
eat F
A
Nordic
Aquatic
FA
Suwann
ee R
iver N
OM
Nordic
Reservo
ir NOM
Luci
56
fera
se a
(%
of
indu
ctio
n of
33
pM E
2)
50 mg L-1 150 mg L-1
ctiv
ity
Fig. 17 Antiestrogenic effects of HS in MVLN cells. HS-dependent inhibition of luciferase activity elicited by 33 pM E2. Means ± SD of triplicate determinations are displayed.
Tab. 4 Antiestrogenic activity of HS alkali solutions; IC50 -concentration causing 50% inhibition of E2-induced luciferase activity.
Antiestrogenic activity
Humic Acid - FlukaHA Sodium Salt
Suwannee River HAElliot Soil HA
Florida Peat HALeonardite HA
Waskish Peat HA Suwannee River FA
Florida Peat FANordic Aquatic FA
Suwannee River NOMNordic Reservoir NOM
LOEC (mg L-1) IC50 (mg L-1)
16.7 43 ± 316.7 40 ± 4n.eff. n.eff.
50 164 ± 9a
150 w.eff.50 132 ± 850 84 ± 650 127 ± 8 50 112 ± 450 83 ± 7150 w.eff.
n.eff. n.eff.
a approximation above measured concentration range n.ind. - no induction; n.eff. - no effect; w.eff. - weak effect

Interaction of HS with light - ecotoxicological consequences
3. Interaction of HS with light - ecotoxicological consequences (incl. Paper III)
3.1. Interaction with light
Dissolved HS are yellow-colored and exert relatively strong absorption especially in the UV region of solar radiation (Hessen & Faerovig, 2001; Rasmussen et al., 1989), but the absorption applies also for visible light. When HS interact with light, series of chemical reactions can occur (Fig. 18). Most of the energy from sunlight absorbed by HS is directly used for warming. But a small proportion of absorbed energy can prompt the HS to promote a chain reaction. The light absorption capacity is, in most cases, linked to the presence of delocalized π-electron systems, which are available from aromatic rings or conjugated double bonds (Steinberg, 2003).
Fig. 18 Pathways of photoreactions of HS (Frimmel, 1994). HS* - excited HS, RS - reactive species, RRS - very reactive species
57

Chapter 3.1.
58
The principal routes of photo-reactions with HS involve the transfer of the energy from excited HS molecules - HS* - on reactive species (or very reactive species, Fig. 18), such as solvated electrons, hydroxyl radicals, singlet oxygen and reactive triplet states (Aguer et al., 1999; Cooper et al., 1989; Frimmel, 1994; Grannas et al., 2006; Paul et al., 2004). The quantum yield of formation of these species depends on such parameters as the radiation wavelength, oxygen concentration, origin of the HS, and also on the metallic cations contained in HS (Aguer & Richard, 1996; Aguer et al., 1999). Through their photo-inductive properties, HS can be used to degrade xenobiotics (chapter 3.1.2).
Strong absorption of HS in UV-region also means that HS provide a natural biogeochemical protection against UV radiation in natural waters. Only a few milligrams of DOC per liter of water are sufficient to act as a biogeochemical shield against UV-B radiation, allowing them to penetrate only a few decimeters into the water column (Bukaveckas & Robbins-Forbes, 2000).
Experiments with irradiation of NOM samples performed by Frimmel (1998) have shown there is a general bleaching effect in the irradiated samples, accompanied by a change in the molecular size distribution of NOM. Thus, irradiation causes a decrease in the higher molecular range and an increase amount of smaller molecules, along with an increase of the assimilable amount of organic matter.
3.1.1. Direct toxicity of HS photo-products to organisms
Hessen and van Donk (1994) performed green alga Ankistrodesmus bibraianus based bioassay for assessment of UV-irradiated surface water from two humic waters. With moderate UV doses (1.1–5.4 J cm-2 at 312 nm), phytoplankton production was stimulated, while higher UV doses (>10 J cm-2) lead to minor to serious growth impairment. Since the phytoplankton was added after the light exposure, the results supported the hypothesis of the long-lived algicidal effect induced by UV iradiation of HS (Hessen & Vandonk, 1994).

Interaction with light
59
Environmentally more relevant changes in irradiated typical surface humus water were shown by Gjessing and Kallovist (1991). Effects comprised the changes in chemical composition of water, including acidity and charge density, and also inhibition of growth of the alga Selenastrum capricornutum. Growth inhibition increased with increased UV-dose and the algicidal effect apparently lasts for long time (several weeks), what is in agreement with above mentioned Hessen and van Donk (1994) observation. The observed action of UV-treated humus water may be explained by photon-initiated interactions of HS and other chemicals in the water which result in the formation of ROS, capable to oxidize organic materials. In another analogous experiment, the toxicity for Daphnia magna was observed to be higher in irradiated samples of HA, especially in deaerated water (Frimmel, 1998).
3.1.2. HS mediated photo-transformations of xenobiotics
As mentioned above, irradiation of dissolved HS lead also to formation of RS (ROS) which can subsequently react with allelochemicals or xenobiotics. This type of reactions is known as indirect or sensitized photolysis that is an important elimination pathway for many persistent xenobiotics in water, which cannot directly absorb radiation with a wavelength >290 nm (Steinberg, 2003). An example of indirect photolysis can be the degradation of the fungicide vinclozolin (Hustert & Moza, 1997). With visible light (UV filtered) and in the absence of HS, there is only a very slow degradation of the fungicide. With visible light (UV filtered) and the presence of HS, there is a 50% degradation in 8 h, while complete sunlight and HS together support another 10% degradation in this time period (Fig. 19).

Chapter 3.1.
Fig. 19 Direct and indirect (+HS) photolysis of the vinclozolin with UV radiation + visible light, or only visible light (+UV filter);cVinclozolin = 1 mg L-1, cHS = 5 mg L-1 (Hustert & Moza, 1997).
HS-induced ROS production has also been shown to oxidize insecticide DDT (Kulovaara et al., 1995) or pesticides Irgarol, Fenuron (Amine-Khodja et al., 2006a; Amine-Khodja et al., 2006b), Carboxin and Oxycarboxin (Hustert et al., 1999). Although photolysis of these compounds can change their properties including toxicity, in some cases it can even significantly increase their toxic potential. In the case of vinclozolin, all three identified photo-products were toxicologically even more hazardous than the parent fungicide (Hustert & Moza, 1997).
60

Changes of AhR-mediated activities of HS after irradiation
61
3.2. Changes of AhR-mediated activities of HS after irradiation (Paper III)
Regarding the polyphenolic structure of humic molecules, HS contain number of aromatic rings and conjugated double bonds - the so called chromophores. That implies irradiation of dissolved HS can lead to a series of photochemical reactions which can act on the HS itself, or on other substances present in aquatic environment along with HS such as e.g. xenobiotics. In Paper III, we have studied the changes in absorption properties and AhR-mediated activities of HS after irradiation simulating natural solar light. Thus, we have linked to our previous studies (Paper I and II) where significant interactions of five HA with AhR were determined.
After the HS irradiation, the absorbance of all of the HS was lower than that of the original materials. In addition, the AhR-mediated activities of two HA were partially decreased by irradiation. The activities of other HS, that were either AhR-active or -inactive, were not affected by irradiation. Potential reasons of the observed irradiation-induced changes in AhR-mediated activities are discussed further (the last paragraph of chapter 3.2.1).
3.2.1. Results - Decrease of absorbance and AhR-mediated activities of HS after irradiation
Twelve different HS (described in chapter 2.3.2.1) were irradiated in Pyrex tubes with a medium-pressure mercury lamp for a duration of from 0 to 52 h (which corresponds to 0-52 days of natural solar irradiation). Original concentrations of HS samples were 50 mg L-1, and the greatest concentration of the HS photo-products subsequently tested in the bioassay was 17 mg L-1, which is an environmentally relevant concentration.

Chapter 3.2.
The absorbance of all twelve irradiated HS samples decreased significantly as a function of duration of irradiation (Fig. 20). The decrease in absorbance of all the HS samples can be described using one phase exponential decay function (Tab. 5). Subsequently calculated t1/2 values of photo-degradation of all irradiated HS are also given.
Tab. 5 Summary of decay equations of both absorbance and AhR-mediatedactivities and calculated t1/2 of photo-degradation of all HS samples.Changes after irradiation
62
Decay equation t1/2 [h] Decay equation t1/2 [h]
HA-Fluka y=0.48e(-0.083x)+0.19 8.4 y=16.25e(-0.20x)+6.72 3.5
HA Sodium Salt y=0.50e(-0.075x)+0.21 9.2 y=6.52e(-0.18x)+5.67 3.9
Elliot Soil HA y=0.53e(-0.076x)+0.57 9.1 - -
Florida Peat HA y=0.64e(-0.095x)+0.23 7.3 - -
Leonardite HA y=0.73e(-0.071x)+0.36 9.8 - -
Suwannee River HA y=0.33e(-0.076x)+0.28 9.1 - -
Waskish Peat HA y=0.41e(-0.012x)+0.27 5.7 - -
Suwannee River FA y=0.19e(-0.015x)+0.15 4.8 - -
Nordic Aquatic FA y=0.27e(-0.014x)+0.17 5.1 - -
Suwannee River NOM y=0.17e(-0.021x)+0.28 3.3 - -
Nordic Reservoir NOM y=0.13e(-0.019x)+0.12 3.6 - -
CompoundAhR-mediated activity Absorbance
t1/2 values were calculated from exponential decay equation.

Changes of AhR-mediated activities of HS after irradiation
0 10 20 30 40 50
0.2
0.4
0.6
0.8
1.0
1.2 Elliot Soil HALeonardite HAFlorida Peat HAHA Sodium SaltHA-Fluka
Time of irradiation [h]
Abs
orba
nce
units
Fig. 20 Decrease of absorbance of irradiated AhR-active HA samples measured at λ = 350 nm. Seven other HS samples exerted analogous exponential decay of absorbance (not shown). Exponential decay equations and t1/2 of absorbing fraction for all HS samples are described in Tab. 5. HS samples were irradiated at concentration 50 mg L-1.
Of the twelve HS samples, irradiation of only two, HA-Fluka and HA Sodium Salt, resulted in a statistically significant decrease of their AhR-mediated activities (Fig. 21). The AhR-mediated activities of other three of the five AhR-active HA samples (Leonardite HA, Elliot Soil HA and Florida Peat HA), was decreased only slightly by irradiation. Because one hour of the simulated irradiation represents approximately one day of direct solar irradiation under natural conditions, this small decrease could be regarded as non-significant. The AhR-mediated activities of the other seven HS (without any substantial AhR-mediated activity) have not changed after irradiation.
63

Chapter 3.2.
0 10 20 30 40 500
5
10
15
20
25
HA-FlukaHA Sodium Salt
Time of irradiation [h]
Luci
fera
se a
ctiv
ity(%
of T
CD
Dm
ax in
duct
ion)
Fig. 21 Decrease of AhR-mediated activities of irradiated HA-Fluka and HA Sodium Salt. The results are related to maximal standard TCDD (100 pM) induction. Means ± SD of triplicate determinations are displayed. Concentration of tested HA samples was 17 mg L-1.
Nevertheless, even after 52 h of laboratory irradiation, the photo-induced decay of HA-Fluka and HA Sodium Salt AhR-mediated activities did not reach zero. The AhR-mediated activity of both HA samples reached significant plateau values - 6.7% for HA-Fluka and 5.7% for HA Sodium Salt (i.e. % of maximal standard TCDD induction), which corresponds to 29% of original activity of HA-Fluka sample, resp. 46% of original activity of HA Sodium Salt sample. These findings suggest a combination of photo-stable and photo-labile AhR activators in both HA samples with decrease of their AhR-mediated activity (HA-Fluka and HA Sodium Salt, Fig. 21). The other three AhR-active HA samples without any significant decrease in AhR-mediated activity (Elliot Soil HA, Leonardite HA and Florida Peat HA) then contain only photo-stable AhR-activators.
64

Summary and general discussion
65
4. Summary and general discussion
Presented dissertation thesis is focused on ecotoxicological properties of humic substances (HS), especially on their direct effects on living organisms. Regarding the natural and ubiquitous occurrence of HS both in water and terrestrial environments, it is very important to assess their ecotoxicological properties. Until recently, the main point of interest was focused on their indirect effects on organisms, such as modulation of bioavailability (i.e. toxicity as well) both of inorganic and organic compounds by HS. Nevertheless, certain studies have demonstrated uptake of HS both into plant and animal cells, which makes relevant following observed direct effects of HS: induction of biotransformation enzymes in amphipods, modulation of photosynthetic oxygen release in plants, hormone-like effects of HS on nematode, swordtail fish and clawed frog, or even induction of human Black foot disease. In our research, we have focused mainly on interaction of HS with intracellular receptors, particularly with aryl hydrocarbon receptor (AhR) and estrogen receptor (ER). Study of interaction of HS with these receptors was carried out using modern in vitro bioassays based on permanent transfected cell lines: H4IIE-luc cell line for assessment of AhR-mediated effects, and MVLN cell line for assessment of ER-mediated effects.
The first two papers - Paper I and part of the Paper II - deal with interaction of wide spectra of HS with AhR, i.e. with receptor that play a key role in toxicity of many mainly persistent organic pollutants. Our results have shown significant activation of AhR by five humic acid samples out of twelve HS tested. More throughout investigation has revealed that activation of AhR is caused both by hydrophilic and lipophilic compounds, excluding significant contribution of man-made persistent AhR-ligands such as PCDD or PCB. Similar results of significant AhR-mediated activity were also obtained by assessment of all three HA samples we isolated from soils (chapter 2.3.2.1). The interpretation of results of these soil HA samples, however, can be complicated by the skipping of the last cleaning step of HA isolation - dialyses. Thus, this samples can not be regarded as reliable HA samples, compared with commercially available HS. In addition, further research would be necessary to examine role of the possible

Chapter 4.
66
anthropogenic contamination of these soil HA, e.g. by analyses for the content of indicator PCB congeners and U.S. EPA priority PAH.
The traditional view of HS structures suggested relatively large rigid molecules that are not only unlikely to bind to the ligand binding region of the AhR, but also unlikely to enter cells. However, recent studies have shown that HS are able to cross both plant and animal cell membranes (chapter 2.2.1) and interact with receptors (chapter 2.2.2.5). Moreover, certain studies of HS structure suggest it consists of aggregates of relatively low-molecular weight (<2 kDa) organic compounds and metal ions bound together by non-covalent interactions (chapter 1.2.2). This leads to a hypothesis that the AhR-active compounds may be rather small molecules released from this complex.
Although the molecular structure of HS (or their compartments) is probably different from the structure of HAH - well known AhR ligands, AhR can be bound and activated by structurally diverse ligands (unlike most ligand-dependent receptors, chapter 2.3.1.2). This diversity is evident by comparison of the naturally occurring „non-classical“ AhR ligands (Fig. 11) to that of synthetic „classical“ AhR ligands (Fig. 10). „Non-classical“ AhR ligands exert relatively low affinity to AhR (compared to HAH), however, exposure of animals to these ligands can be in greater amounts by reason of their occurrence e.g. in dietary products. Since some HS are AhR ligands (although relatively weak - results from the Papers I and II), we assume that chronic exposure to high levels of HS (environmentally relevant concentrations are in the order of mg L-1, chapter 1.1.2) could potentially produce adverse effects similar to those produced by TCDD.
Interaction of HS with estrogen receptor, which is described in the second part of Paper II, represents another specific mode of HS biological action. Disruption of estrogens activity by xenobiotic chemicals can be either by activating or blocking of ER, but also by number of various mechanisms that do not involve interaction of xenobiotic with ER. Our results, obtained using in vitro bioassay, have shown relatively weak antiestrogenic effect of twelve HS. On the other hand, this in vitro indication of antiestrogenic effect is in inconsistence with in vivo results where was observed more likely opposite estrogenic effect of HS (chapter 2.2.2.4). However, these in vivo studies were conducted with a synthetic

Summary and general discussion
67
HS1500 and whole organisms, which may cause the differences in results on account of various modes of possible xenoestrogenic action (chapter 2.4.1.1). Other effects on reproduction have been observed in nematodes (chapter 2.2.2.4) where natural peat FA strongly inhibited number of offspring of exposed individuals. A recent in vitro study with oocytes of frog Rana temporaria (Zenkevics et al., 2005) showed that HS reduced sensitivity of oocytes to gonadotropic hormone and thus delayed their maturation. These authors attribute this effect to blocking membrane transport of gonadotropic hormone by HS. Thus, HS-caused changes of membrane permeability for E2 may also be the reason of antiestrogenic effects observed in our study. Another reason may be the simple sorption of relatively hydrophobic molecule of E2 to carbon-rich HS in test medium. This hypothesis is supported by a recent study on influence of NOM on partitioning of E2 between water phase and an artificial membrane, where the researchers have observed 30% decrease of membrane/water partition coefficients of E2 at concentrations as low as 3 mg L-1 of HAA and 20% decrease for 8 mg L-1 of Suwannee River FA (Yamamoto et al., 2004). That corresponds with our finding that both HA-Fluka and HA Sodium Salt elicited much higher antiestrogenic activity than other HS. On the other hand, sorption is a relatively non-specific process what would suggest all HS to be active. However, our four samples (two NOM and two HA) elicited no or only weak antiestrogenic activity, even at high concentrations (Tab. 4). Question of some more specific mechanism of action thus still remains unanswered.
The last part of thesis discusses photochemical aspects of HS and is related to the Paper III. Our results demonstrate that all twelve HS elicited significant photo-bleaching effect when exposed to simulated irradiation. These findings have confirmed well known observations that HS represent photo-active compounds. AhR-mediated activity assessment of irradiated HS has shown a combination of photo-stable and photo-labile AhR activators in two HA samples that exerted decrease of their AhR-mediated activity while the other HS both AhR-active or -inactive were not changed their activity after irradiation. Results of our measurements demonstrate that AhR-mediated activities of two active HA is caused by both photo-stable and photo-labile AhR activators, while other three active HA contain only photo-stable AhR activators.

Chapter 4.
68
Based on the results of our study, we suppose that decrease of AhR-mediated activity of both HA-Fluka and HA Sodium Salt after simulated irradiation could be related to indirect photolysis of AhR-active parts of HA by reactive oxygen molecules, by analogy with observations described in chapter 3.1.2. Results in Paper I and II suggest the AhR-activating agents of HS are both hydrophilic and lipophilic non-persistent molecules, such as e.g. PAH (chapter 2.3.1.2) and their various, often more hydrophilic derivatives, e.g. aza-PAHs (Sovadinova et al., 2006). The results in Paper III shown that potency of HA to act through the specific mechanism of action - activation of AhR - either stay the same or decreases after irradiation. Thus, in some cases, irradiation of dissolved HA can be beneficial in the sense of decrease of AhR active substances based stress to aquatic vertebrates.
Overall summarized, we have to keep still in mind that generalization of all upper mentioned ecotoxicological conclusions is limited to fact that each HS is of different origin and thus can elicit rather specific properties. Nevertheless, based on the scientific observations, we can at ease agree with the statement: "HS are controlling factors in aquatic ecology as important, for instance, as nutrients, temperature or even light. We are just at the beginning of an understanding of the complete function of HS in freshwater ecosystems" (Steinberg et al., 2006).

References
69
References Aguer, J.P. & Richard, C. (1996) Reactive species produced on irradiation at 365 nm of
aqueous solutions of humic acids. Journal of Photochemistry and Photobiology a-Chemistry, 93, 193-198.
Aguer, J.P., Richard, C., & Andreux, F. (1999) Effect of light on humic substances: Production of reactive species. Analusis, 27, 387-390.
Aiken, G.R. (1988). A critical evaluation of the use of macroporous resins for the isolation of humic substances. In Humic substances and their role in the environment (eds F.H. Frimmel & R.F. Christman), pp. 15-27. Wiley-Interscience, Berlin.
Amine-Khodja, A., Richard, C., Lavedrine, B., Guyot, G., Trubetskaya, O., & Trubetskoj, O. (2006a) Water-soluble fractions of composts for the photodegradation of organic pollutants in solar light. Environmental Chemistry Letters, 3, 173-177.
Amine-Khodja, A., Trubetskaya, O., Trubetskoj, O., Cavani, L., Ciavatta, C., Guyot, G., & Richard, C. (2006b) Humic-like substances extracted from composts can promote the photo degradation of Irgarol 1051 in solar light. Chemosphere, 62, 1021-1027.
Beer, A.M., Junginger, H.E., Lukanov, J., & Sagorchev, P. (2003) Evaluation of the permeation of peat substances through human skin in vitro. International Journal of Pharmaceutics, 253, 169-175.
Beer, A.M., Lukanov, J., & Sagorchev, P. (2000) The influence of fulvic and ulmic acids from peat, on the spontaneous contractile activity of smooth muscles. Phytomedicine, 7, 407-415.
Bettin, C., Oehlmann, J., & Stroben, E. (1996) TBT-induced imposex in marine neogastropods is mediated by an increasing androgen level. Helgolander Meeresuntersuchungen, 50, 299-317.
Bollag, J.M. & Myers, C. (1992) Detoxification of Aquatic and Terrestrial Sites through Binding of Pollutants to Humic Substances. Science of the Total Environment, 118, 357-366.
Bollag, J.M., Myers, C.J., & Minard, R.D. (1992) Biological and Chemical Interactions of Pesticides with Soil Organic-Matter. Science of the Total Environment, 123, 205-217.
Bukaveckas, P.A. & Robbins-Forbes, M. (2000) Role of dissolved organic carbon in the attenuation of photosynthetically active and ultraviolet radiation in Adirondack lakes. Freshwater Biology, 43, 339-354.
Calace, N. & Petronio, B.M. (2004) The role of organic matter on metal toxicity and bio-availability. Annali Di Chimica, 94, 487-493.

References
70
Cary, G.A., McMahon, J.A., & Kuc, W.J. (1987) The Effect of Suspended-Solids and Naturally-Occurring Dissolved Organics in Reducing the Acute Toxicities of Cationic Polyelectrolytes to Aquatic Organisms. Environmental Toxicology and Chemistry, 6, 469-474.
Cooper, W.J., Zika, R.G., Petasne, R.G., & Fischer, A.M. (1989) Sunlight-Induced Photochemistry of Humic Substances in Natural-Waters - Major Reactive Species. Acs Symposium Series, 219, 333-362.
Day, K.E. (1991) Effects of Dissolved Organic-Carbon on Accumulation and Acute Toxicity of Fenvalerate, Deltamethrin and Cyhalothrin to Daphnia-Magna (Straus). Environmental Toxicology and Chemistry, 10, 91-101.
Denison, M.S. & Nagy, S.R. (2003) Activation of the aryl hydrocarbon receptor by structurally diverse exogenous and endogenous chemicals. Annual Review of Pharmacology and Toxicology, 43, 309-334.
Denison, M.S., Pandini, A., Nagy, S.R., Baldwin, E.P., & Bonati, L. (2002) Ligand binding and activation of the Ah receptor. Chemico-Biological Interactions, 141, 3-24.
Denison, M.S. & Perdew, G.H. (2002) The Ah receptors - Molecular mechanisms and signal transduction - Introductory comments. Chemico-Biological Interactions, 141, 1-1.
DePaolis, F. & Kukkonen, J. (1997) Binding of organic pollutants to humic and fulvic acids: Influence of pH and the structure of humic material. Chemosphere, 34, 1693-1704.
Devito, M.J. & Birnbaum, L.S. (1995) Dioxins - Model Chemicals for Assessing Receptor-Mediated Toxicity. Toxicology, 102, 115-123.
Doring, U.M. & Marschner, B. (1998) Water Solubility Enhancement of Benzo(a)pyrene and 2,2′,5,5′-terachlorobiphenyl by Dissolved Organic Matter (DOM). Physics and Chemistry of The Earth, 23, 193-197.
Drummond, A.E., Britt, K.L., Dyson, M., Jones, M.E., Kerr, J.B., O'Donnell, L., Simpson, E.R., & Findlay, J.K. (2002) Ovarian steroid receptors and their role in ovarian function. Molecular and Cellular Endocrinology, 191, 27-33.
Freyberger, A. & Schmuck, G. (2005) Screening for estrogenicity and anti-estrogenicity: a critical evaluation of an MVLN cell-based transactivation assay. Toxicology Letters, 155, 1-13.
Frimmel, F.H. (1994) Photochemical Aspects Related to Humic Substances. Environment International, 20, 373-385.
Frimmel, F.H. (1998) Impact of light on the properties of aquatic natural organic matter. Environment International, 24, 559-571.

References
71
Garcia-Mina, J.M., Antolin, M.C., & Sanchez-Diaz, M. (2004) Metal-humic complexes and plant micronutrient uptake: a study based on different plant species cultivated in diverse soil types. Plant and Soil, 258, 57-68.
Gau, R.J., Yang, H.L., Chow, S.N., Suen, J.L., & Lu, F.J. (2000) Humic acid suppresses the LPS-induced expression of cell-surface adhesion proteins through the inhibition of NF-kappa B activation. Toxicology and Applied Pharmacology, 166, 59-67.
Gau, R.J., Yang, H.L., Suen, J.L., & Lu, F.J. (2001) Induction of oxidative stress by humic acid through increasing intracellular iron: A possible mechanism leading to atherothrombotic vascular disorder in blackfoot disease. Biochemical and Biophysical Research Communications, 283, 743-749.
Gillesby, B.E. & Zacharewski, T.R. (1998) Exoestrogens: Mechanisms of action and strategies for identification and assessment. Environmental Toxicology and Chemistry, 17, 3-14.
Gjessing, E.T. & Kallovist, T. (1991) Algicidal and Chemical Effect of Uv-Radiation of Water Containing Humic Substances. Water Research, 25, 491-494.
Glover, C.N., Pane, E.F., & Wood, C.M. (2005) Humic substances influence sodium metabolism in the freshwater crustacean Daphnia magna. Physiological and Biochemical Zoology, 78, 405-416.
Glover, C.N. & Wood, C.M. (2005) The disruption of Daphnia magna sodium metabolism by humic substances: Mechanism of action and effect of humic substance source. Physiological and Biochemical Zoology, 78, 1005-1016.
Grady, A.W., Fabacher, D.L., Frame, G., & Steadman, B.L. (1992) Morphological deformities in brown bullheads administered dietary b-naphthoflavone. Journal Of Aquatic Animal Health, 7-16.
Grannas, A.M., Martin, C.B., Chin, Y.P., & Platz, M. (2006) Hydroxyl radical production from irradiated arctic dissolved organic matter. Biogeochemistry, 78, 51-66.
Gschwend, P.M. & Wu, S.C. (1985) On the Constancy of Sediment Water Partition-Coefficients of Hydrophobic Organic Pollutants. Environmental Science & Technology, 19, 90-96.
Harvey, G.R., Boran, D.A., Chesal, L.A., & Tokar, J.M. (1983) The Structure of Marine Fulvic and Humic Acids. Marine Chemistry, 12, 119-132.
Harvey, G.R., Boran, D.A., Piotrowicz, S.R., & Weisel, C.P. (1984) Synthesis of Marine Humic Substances from Unsaturated Lipids. Nature, 309, 244-246.
Hatcher, P.G., Faulon, J.-L., Clifford, D.A., & Matthews, J.P. (1992) A three-dimensional structural model for humic acids from oxidized soil. In Humic Substances in the Global Environment and Implications on Human Health (eds N. Senesi & T.M. Miano), pp. 133-138. Elsevier, Italy.

References
72
Hatcher, P.G. & Spiker, E.C. (1988). Selective degradation ofplant biomolecules. In Humic Substances and their Role in the Environment (ed F.H. Frimmel, Christman, R.F.), pp. 59±74. Wiley, New York.
Heil, C.A. (2005) Influence of humic, fulvic and hydrophilic acids on the growth, photosynthesis and respiration of the dinoflagellate Prorocentrum minimum (Pavillard) Schiller. Harmful Algae, 4, 603-618.
Hessen, D.O. & Faerovig, P.J. (2001) The photoprotective role of humus-DOC for Selenastrum and Daphnia. Plant Ecology, 154, 261-+.
Hessen, D.O. & Vandonk, E. (1994) Effects of Uv-Radiation of Humic Water on Primary and Secondary Production. Water Air and Soil Pollution, 75, 325-338.
Hodge, V.A., Fan, G.T., Solomon, K.R., Kaushik, N.K., Leppard, G.G., & Burnison, B.K. (1993) Effects of the Presence and Absence of Various Fractions of Dissolved Organic-Matter on the Toxicity of Fenvalerate to Daphnia-Magna. Environmental Toxicology and Chemistry, 12, 167-176.
Hoss, S., Bergtold, M., Haitzer, M., Traunspurger, W., & Steinberg, C.E.W. (2001) Refractory dissolved organic matter can influence the reproduction of Caenorhabditis elegans (Nematoda). Freshwater Biology, 46, 1-10.
Huang, T.S., Lu, F.J., Tsai, C.W., & Chopra, I.J. (1994) Effect of Humic Acids on Thyroidal Function. Journal of Endocrinological Investigation, 17, 787-791.
Hustert, K. & Moza, P.N. (1997) Photochemical degradation of dicarboximide fungicides in the presence of soil constituents. Chemosphere, 35, 33-37.
Hustert, K., Moza, P.N., & Kettrup, A. (1999) Photochemical degradation of carboxin and oxycarboxin in the presence of humic substances and soil. Chemosphere, 38, 3423-3429.
Chefetz, B., Tarchitzky, J., Deshmukh, A.P., Hatcher, P.G., & Chen, Y. (2002) Structural characterization of soil organic matter and humic acids in particle-size fractions of an agricultural soil. Soil Science Society of America Journal, 66, 129-141.
Cheng, M.L., Ho, H.Y., Huang, Y.W., Lu, F.J., & Chiu, D.T.Y. (2003) Humic acid induces oxidative DNA damage, growth retardation, and apoptosis in human primary fibroblasts. Experimental Biology and Medicine, 228, 413-423.
Cheng, M.L., Ho, H.Y., Chiu, D.T.Y., & Lu, F.J. (1999) Humic acid-mediated oxidative damages to human erythrocytes: A possible mechanism leading to anemia in Blackfoot disease. Free Radical Biology and Medicine, 27, 470-477.
IHSS (2004) Isolation of IHSS Samples, Vol. 2006. International Humic Substances Society, http://www.ihss.gatech.edu, on-line 8.1.2006
Jacobs, M.N., Dickins, M., & Lewis, D.F.V. (2003) Homology modelling of the nuclear receptors: human oestrogen receptor beta (hER beta), the human pregnane-X-receptor (PXR), the Ah receptor (AhR) and the constitutive androstane receptor

References
73
(CAR) ligand binding domains from the human oestrogen receptor alpha (hER alpha) crystal structure, and the human peroxisome proliferator activated receptor alpha (PPAR alpha) ligand binding domain from the human PPAR gamma crystal structure. Journal of Steroid Biochemistry and Molecular Biology, 84, 117-132.
Janosek, J., Hilscherova, K., Blaha, L., & Holoubek, I. (2006) Environmental xenobiotics and nuclear receptors - Interactions, effects and in vitro assessment. Toxicology in Vitro, 20, 18-37.
Jeuken, A., Keser, B.J.G., Khan, E., Brouwer, A., Koeman, J., & Denison, M.S. (2003) Activation of the Ah receptor by extracts of dietary herbal supplements, vegetables, and fruits. Journal of Agricultural and Food Chemistry, 51, 5478-5487.
Jones, K.D. & Huang, W.H. (2003) Evaluation of toxicity of the pesticides, chlorpyrifos and arsenic, in the presence of compost humic substances in aqueous systems. Journal of Hazardous Materials, 103, 93-105.
Klaus, U., Mohamed, S., Volk, M., & Spiteller, M. (1998) Interaction of aquatic humic substances with anilazine and its derivatives: The nature of the bound residues. Chemosphere, 37, 341-361.
Kulovaara, M., Backlund, P., & Corin, N. (1995) Light-Induced Degradation of Ddt in Humic Water. Science of the Total Environment, 170, 185-191.
Laurberg, P., Andersen, S., Pedersen, I.B., Ovesen, L., & Knudsen, N. (2003) Humic substances in drinking water and the epidemiology of thyroid disease. Biofactors, 19, 145-153.
Lee, Y., Huang, T.S., Yan, M.L., Huang, L.R., Chen, C.H., & Lu, F.J. (1999) Peroxisome proliferation, adipocyte determination and differentiation of C3H10T1/2 fibroblast cells induced by humic acid: Induction of PPAR in diverse cells. Journal of Cellular Physiology, 179, 218-225.
Linnik, P.M. (1998) The state of heavy metals in the interstitial solutions as an important characteristic of their migration mobility. International Review of Hydrobiology, 83, 239-247.
Linnik, P.N. & Nabivanets, B.I. (1984) The State of Metal-Ions in Natural-Waters. Acta Hydrochimica Et Hydrobiologica, 12, 335-361.
Lintelmann, J., Katayama, A., Kurihara, N., Shore, L., & Wenzel, A. (2003) Endocrine disruptors in the environment (IUPAC Technical Report). Pure and Applied Chemistry, 75, 631–681.
Lutz, I., Jie, Z., Opitz, R., Kloas, W., Ying, X., Menzel, R., & Steinberg, C.E.W. (2005) Environmental signals: Synthetic humic substances act as xeno-estrogen and affect the thyroid system of Xenopus laevis. Chemosphere, 61, 1183-1188.
Machala, M., Vondracek, J., Blaha, L., Ciganek, M., & Neca, J. (2001) Aryl hydrocarbon receptor-mediated activity of mutagenic polycyclic aromatic

References
74
hydrocarbons determined using in vitro reporter gene assay. Mutation Research-Genetic Toxicology and Environmental Mutagenesis, 497, 49-62.
Mantovani, A., Stazi, A.V., Macri, C., Maranghi, F., & Ricciardi, C. (1999) Problems in testing and risk assessment of endocrine disrupting chemicals with regard to developmental toxicology. Chemosphere, 39, 1293-1300.
Marschner, B. (1998) DOM-Enhanced Mobilization of Benzo(a)pyrene in a Contaminated Soil Under Different Chemical Conditions. Physics and Chemistry of The Earth, 23, 199-203.
Martinneto, L., Vieira, E.M., & Sposito, G. (1994) Mechanism of Atrazine Sorption by Humic-Acid - a Spectroscopic Study. Environmental Science & Technology, 28, 1867-1873.
Massaad, C., Entezami, F., Massade, L., Benahmed, M., Olivennes, F., Barouki, R., & Hamamah, S. (2002) How can chemical compounds alter human fertility? European Journal of Obstetrics Gynecology and Reproductive Biology, 100, 127-137.
Matsuo, A.Y.O., Woodin, B.R., Reddy, C.M., Val, A.L., & Stegeman, J.J. (2006) Humic substances and crude oil induce cytochrome P450 1A expression in the Amazonian fish species Colossoma macropomum (Tambaqui). Environmental Science & Technology, 40, 2851-2858.
Meinelt, T., Schreckenbach, K., Knopf, K., Wienke, A., Stuber, A., & Steinberg, C.E.W. (2004) Humic substances affect physiological condition and sex ratio of swordtail (Xiphophorus helleri Heckel). Aquatic Sciences, 66, 239-245.
Menzel, R., Sturzenbaum, S., Barenwaldt, A., Kulas, J., & Steinberg, C.E.W. (2005) Humic material induces behavioral and global transcriptional responses in the nematode Caenorhabditis elegans. Environmental Science & Technology, 39, 8324-8332.
Myneni, S.C.B., Brown, J.T., Martinez, G.A., & Meyer-Ilse, W. (1999) Imaging of humic substance macromolecular structures in water and soils. Science, 286, 1335-1337.
Orlov, D.S., Sadovnikova, L.K., & Savrova, A.L. (1995) Comparative study copper, zinc and lead adsorption by humic acids of different origin. Doklady Akademii Nauk, 345, 535-537.
Ouatmane, A., Dorazio, V., Hafidi, M., Revel, J.C., & Senesi, N. (2000) Elemental and spectroscopic characterization of humic acids fractionated by gel permeation chromatography. Agronomie, 20, 491-504.
Pacheco, M.L., Pena-Mendez, E.M., & Havel, J. (2003) Supramolecular interactions of humic acids with organic and inorganic xenobiotics studied by capillary electrophoresis. Chemosphere, 51, 95-108.
Parzefall, W. (2002) Risk assessment of dioxin contamination in human food. Food and Chemical Toxicology, 40, 1185-1189.

References
75
Paul, A., Hackbarth, S., Vogt, R.D., Roder, B., Burnison, B.K., & Steinberg, C.E.W. (2004) Photogeneration of singlet oxygen by humic substances: comparison of humic substances of aquatic and terrestrial origin. Photochemical & Photobiological Sciences, 3, 273-280.
Pflugmacher, S., Pietsch, C., Rieger, W., & Steinberg, C.E.W. (2006) Dissolved natural organic matter (NOM) impacts photosynthetic oxygen production and electron transport in coontail Ceratophyllum demersum. Science of the Total Environment, 357, 169-175.
Pflugmacher, S., Spangenberg, M., & Steinberg, C.E.W. (1999) Dissolved organic matter (DOM) and effects on the aquatic macrophyte Ceratophyllum demersum in relation to photosynthesis, pigmentpattern and activity of detoxication enzymes. Journal of Applied Botany-Angewandte Botanik, 73, 184-190.
Pflugmacher, S., Tidwell, L.F., & Steinberg, C.E.W. (2001) Dissolved humic substances can directly affect freshwater organisms. Acta Hydrochimica Et Hydrobiologica, 29, 34-40.
Phillips, S.L. & Olesik, S.V. (2003) Initial characterization of humic acids using liquid chromatography at the critical condition followed by size-exclusion chromatography and electrospray ionization mass spectrometry. Analytical Chemistry, 75, 5544-5553.
Piccolo, A. (2001) The supramolecular structure of humic substances. Soil Science, 166, 810-832.
Piccolo, A. (2002). The supramolecular structure of humic substances: A novel understanding of humus chemistry and implications in soil science. In Advances in Agronomy, Vol 75, Vol. 75, pp. 57-134.
Piccolo, A. & Celano, G. (1994) Hydrogen-Bonding Interactions between the Herbicide Glyphosate and Water-Soluble Humic Substances. Environmental Toxicology and Chemistry, 13, 1737-1741.
Piccolo, A., Nardi, S., & Concheri, G. (1996) Macromolecular changes of humic substances induced by interaction with organic acids. European Journal of Soil Science, 47, 319-328.
Rasmussen, J.B., Godbout, L., & Schallenberg, M. (1989) The Humic Content of Lake Water and Its Relationship to Watershed and Lake Morphometry. Limnology and Oceanography, 34, 1336-1343.
Reen, R.K., Cadwallader, A., & Perdew, G.H. (2002) The subdomains of the transactivation domain of the aryl hydrocarbon receptor (AhR) inhibit AhR and estrogen receptor transcriptional activity. Archives of Biochemistry and Biophysics, 408, 93-102.
Safe, S. (1990) Polychlorinated-Biphenyls (Pcbs), Dibenzo-Para-Dioxins (Pcdds), Dibenzofurans (Pcdfs), and Related-Compounds - Environmental and Mechanistic

References
76
Considerations Which Support the Development of Toxic Equivalency Factors (Tefs). Critical Reviews in Toxicology, 21, 51-88.
Safe, S., Wang, F., Porter, W., Duan, R., & McDougal, A. (1998) Ah receptor agonists as endocrine disruptors: antiestrogenic activity and mechanisms. Toxicology Letters, 103, 343-347.
Schnitzer, M. (1978). Humic substances: Chemistry and Reactions. In Soil Organic Matter (eds M. Schnitzer & S.U. Khan). Elsevier, New York.
Schulten, H.R. & Leinweber, P. (2000) New insights into organic-mineral particles: composition, properties and models of molecular structure. Biology and Fertility of Soils, 30, 399-432.
Schwarzenbach, R.P. (1993) Organic Contaminants in Water. Abstracts of Papers of the American Chemical Society, 205, 278-CHED.
Simpson, A.J., Kingery, W.L., Hayes, M.H.B., Spraul, M., Humpfer, E., Dvortsak, P., Kerssebaum, R., Godejohann, M., & Hofmann, M. (2002) Molecular structures and associations of humic substances in the terrestrial environment. Naturwissenschaften, 89, 84-88.
Singleton, D.W. & Khan, S.A. (2003) Xenoestrogen exposure and mechanisms of endocrine disruption. Frontiers in Bioscience, 8, S110-S118.
Sonnenschein, C. & Soto, A.M. (1998) An updated review of environmental estrogen and androgen mimics and antagonists. Journal of Steroid Biochemistry and Molecular Biology, 65, 143-150.
Sovadinova, I. (2006). Nuclear receptor-mediated toxicity of xenobiotics. Dissertation thesis. Masaryk University, Brno.
Sovadinova, I., Blaha, L., Janosek, J., Hilscherova, K., Giesy, J.P., Jones, P.D., & Holoubek, I. (2006) Cytotoxicity and aryl hydrocarbon receptor-mediated activity of N-heterocyclic polycyclic aromatic hydrocarbons: Structure-activity relationships. Environmental Toxicology and Chemistry, 25, 1291-1297.
Steinberg, C.E.W. (2003) Ecology of Humic Substances in Freshwaters - Determinants from Geochemistry to Ecological Niches Springer, BERLIN.
Steinberg, C.E.W., Hoss, S., & Bruggemann, R. (2002) Further evidence that humic substances have the potential to modulate the reproduction of the nematode Caenorhabditis elegans. International Review of Hydrobiology, 87, 121-133.
Steinberg, C.E.W., Hoss, S., Kloas, W., Lutz, I., Meinelt, T., Pflugmacher, S., & Wiegand, C. (2004) Hormonelike effects of humic substances on fish, amphibians, and invertebrates. Environmental Toxicology, 19, 409-411.
Steinberg, C.E.W., Kamara, S., Prokhotskaya, V.Y., Manusadzianas, L., Karasyova, T.A., Timofeyev, M.A., Jie, Z., Paul, A., Meinelt, T., Farjalla, V.F., Matsuo, A.Y.O., Burnison, B.K., & Menzel, R. (2006) Dissolved humic substances -

References
77
ecological driving forces from the individual to the ecosystem level? Freshwater Biology, 51, 1189-1210.
Steinberg, C.E.W., Paul, A., Pflugmacher, S., Meinelt, T., Klocking, R., & Wiegand, C. (2003) Pure humic substances have the potential to act as xenobiotic chemicals - A review. Fresenius Environmental Bulletin, 12, 391-401.
Steinberg, C.E.W., Sturm, A., Kelbel, J., Lee, S.K., Hertkorn, N., Freitag, D., & Kettrup, A.A. (1992) Changes of Acute Toxicity of Organic-Chemicals to Daphnia-Magna in the Presence of Dissolved Humic Material (Dhm). Zeitschrift Fur Wasser- Und Abwasser-Forschung-Journal for Water and Wastewater Research-Acta Hydrochimica Et Hydrobiologica, 326-332.
Steinberg, C.E.W., Xu, Y., Lee, S.K., Freitag, D., & Kettrup, A. (1993) Effect of Dissolved Humic Material (Dhm) on Bioavailability of Some Organic Xenobiotics to Daphnia-Magna. Chemical Speciation and Bioavailability, 5, 1-9.
Stevenson, F.J. (1994) Humus Chemistry. Genesis, Composition, Reaction, Second edn. John Wiley and Sons, New York.
Sumpter, J.R. (2003) Endocrine disruption in wildlife: The future? Pure and Applied Chemistry, 75, 2355-2360.
Timofeyev, M.A., Wiegand, C., Burnison, B.K., Shatilina, Z.M., Pflugmacher, S., & Steinberg, C.E.W. (2004) Impact of natural organic matter (NOM) on freshwater amphipods. Science of the Total Environment, 319, 115-121.
Tipping, E. (2002) Cation binding by humic substances Press Syndicate of the University of Cambridge, Cambridge.
Wagoner, D.B., Christman, R.F., Cauchon, G., & Paulson, R. (1997) Molar mass and size of Suwannee River natural organic matter using multi-angle laser light scattering. Environmental Science & Technology, 31, 937-941.
Waller, C.L. & McKinney, J.D. (1995) 3-Dimensional Quantitative Structure-Activity-Relationships of Dioxins and Dioxin-Like Compounds - Model Validation and Ah Receptor Characterization. Chemical Research in Toxicology, 8, 847-858.
Wang, W.H., Bray, C.M., & Jones, M.N. (1999) The fate of C-14-labelled humic substances in rice cells in culture. Journal of Plant Physiology, 154, 203-211.
Wershaw, R.L. (1986) A New Model for Humic Materials and their Interaction with Hydrophobic Organic Chemicals in Soil-Water or Sediment-Water Systems. Journal of Contaminant Hydrology 1, 29-45.
Wiegand, C., Meems, N., Timoveyev, M.A., Steinberg, C.E.W., & Pflugmacher, S. (2003). More evidence for humic substances acting as biogeochemicals on organisms. In Humic Substances: Nature's Most Versatile Materials (eds E.A. Ghabbour & G. Davies), pp. 349-361, New York.

References
78
Wijnja, H., Pignatello, J.J., & Malekani, K. (2004) Formation of pi-pi complexes between phenanthrene and model pi-acceptor humic subunits. Journal of Environmental Quality, 33, 265-275.
Yamamoto, H., Liljestrand, H. M. & Shimizu, H. (2004). Effects of dissolved organic matter surrogates on the partitioning of 17 beta-estradiol and p-nonylphenol between synthetic membrane vesicles and water. Environmental Science & Technology, 38:2351-2358.
Zacharewski, T. (1998) Identification and assessment of endocrine disruptors: Limitations of in vivo and in vitro assays. Environmental Health Perspectives, 106, 577-582.
Zenkevics, H., Klavins, M., Vose, V. & Bucena, A. (2005). Humic acid reduces gonadotropin activity and hormonal sensitivity of frog oocytes. Aquat. Toxicol. 75, 380-383.
Zepp, R.G., Baughman, G.L., & Schlotzhauer, P.F. (1981a) Comparison of Photochemical Behavior of Various Humic Substances in Water .1. Sunlight Induced Reactions of Aquatic Pollutants Photosensitized by Humic Substances. Chemosphere, 10, 109-117.
Zepp, R.G., Baughman, G.L., & Schlotzhauer, P.F. (1981b) Comparison of Photochemical Behavior of Various Humic Substances in Water .2. Photosensitized Oxygenations. Chemosphere, 10, 119-126.
Zsolnay, A. (1996). Dissolved humus in soil waters. Pages 171– 224 in A. Piccolo, editor. Humic Substances in Terrestrial Ecosystems. Elsevier, Amsterdam.

Appendices
I. Bittner, M., Janosek, J., Hilscherova, K., Giesy, J., Holoubek, I. and Blaha, L.
Activation of Ah receptor by pure humic acids. Environmental Toxicology,
21:338-342; 2006
II. Janosek, J., Bittner, M., Hilscherová, K., Bláha, L., Giesy, J.P. and
Holoubek, I. AhR-mediated and antiestrogenic activity of humic substances.
Chemosphere; 2006 (in press)
III. Bittner, M., Hilscherová, K. and Giesy J.P. Changes of AhR-mediated
activity of humic substances after irradiation; 2006 (submitted to Environment
International)
IV. Curriculum vitae


Paper I.
Bittner, M., Janosek, J., Hilscherova, K., Giesy, J., Holoubek, I. and Blaha, L.
Activation of Ah receptor by pure humic acids. Environmental Toxicology,
21:338-342; 2006


Activation of Ah Receptor by Pure Humic Acids
Michal Bittner,1 Jaroslav Janosek,1 Klara Hilscherova,1 John Giesy,2,3
Ivan Holoubek,1 Ludek Blaha1
1Masaryk University in Brno, RECETOX, Kamenice 126/3, 625 00 Brno, Czech Republic
2National Food Safety and Toxicology Center, Center for Integrative Toxicology,Department of Zoology, Michigan State University, East Lansing, Michigan 48824-1222, USA
3Department of Biology and Chemistry, City University of Hong Kong, 83 Tat Chee Avenue,Kowloon, Hong Kong SAR, China
Received 17 June 2005; accepted 21 March 2006
ABSTRACT: Humic substances (HS) are ubiquitous in the environment. However, some studies indicatethat HS could induce direct adverse effects on human health and hormone-like effects in fish, amphibians,and invertebrates. In this study we investigated a possible biochemical mechanism of HS toxicity via acti-vation of the aryl hydrocarbon receptor (AhR). AhR mediates the toxic and biological effects of environ-mental contaminants such as 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD), but a number of structurallydiverse compounds has also been found to activate AhR. Alkali solutions of humic acids (HA) were pre-pared, and subsequently, lipophilic compounds (including parts of HA) were extracted by liquid–liquidextraction with hexane/dichloromethane. Organic extract of HA was further treated with sulfuric acid tostudy the role of possible trace persistent contaminants. In vitro dioxin-like activities of obtained extractand HA solutions have been evaluated using H4IIE.luc cells by determining the ethoxyresorufin-O-deethy-lase (EROD) activity and induction of AhR-dependent reporter luciferase. Traces of nonpersistent residuesin HA with known AhR activity were identified and quantified by GC-MS. Our results show that an alkalisolution as well as organic extract of HA were active in both EROD and luciferase assays, while H2SO4-treated extract activity was negligible. Only nonsignificant levels of AhR-inducing contaminants (PAHs andPCBs) were found in the HA samples. Our results indicate that HA or their fragments can elicit significantinductions of AhR-mediated effects in vitro. To our best knowledge, this study is the first in providing directevidence of dioxin-like effects of HA. Further efforts should focus on detailed characterization of potentialtoxic effects of various HSs. # 2006 Wiley Periodicals, Inc. Environ Toxicol 21: 338–342, 2006.
Keywords: aryl hydrocarbon receptor; H4IIE.luc; luciferase; EROD; humic substances
INTRODUCTION
Humic acids (HA) and fulvic acids are certain fractions of
the group of organic compounds called humic substances
(HS). HS are ubiquitous organic compounds that occur in
soils and aquatic ecosystems. In aquatic ecosystems, they
are the main components of dissolved organic matter that is
found in most natural freshwaters in concentrations 0.5–
50 mg L�1 (Steinberg, 2003). Until recently, HS were gen-
erally considered to be inert and only the sorption of metals
and organic contaminants on HS was considered to be im-
portant (McCarthy, 1987; McGeer et al., 2002; Mezin and
Hale, 2004). However, some recent works have shown that
HS can act as xenobiotic chemicals. Meinelt et al. (2001)
found that synthetic HA (HS1500) affects physiological
condition and slightly also the sex ratio of the fish Xipho-phorus helleri. Furthermore, the hormone-like effect of HS
on nematode Caenorhabditis elegans (Hoss et al., 2001;
Correspondence to: J. Janosek; e-mail: [email protected]
Supported by: The Ministry of Education, Czech Republic (No. FRVS
2510/05).
Published online in Wiley InterScience (www.interscience.wiley.com).
DOI 10.1002/tox.20185
�C 2006 Wiley Periodicals, Inc.
338

Steinberg et al., 2002), and increase in amphipod mortality
and change of biochemical parameters (Timofeyev et al.,
2004) were shown. Recent studies conducted in Taiwan
have suggested that HS can even be the causative agents
of endemic human diseases such as Blackfoot disease
(Cheng et al., 1999; Gau et al., 2001; Hseu et al., 2002;
Huang et al., 2003) or Kashin-Beck disease (Liang et al.,
1999).
So far, two types of mechanisms of HS action have been
studied. Nonspecific mechanisms of action were examined
with Daphnia magna, three other amphipod species, and
common carp Cyprinus carpio (Wiegand et al., 2003).
Dose–response relationships between HS and/or natural or-
ganic matter (NOM) exposure and modulation of heat
shock proteins 70 (Hsp 70) were found in carp. Addition-
ally, HS induced Hsp 70 as well as the biotransformation
enzymes glutathione S-transferases, oxidative-stress en-
zymes peroxidase and glutathione-peroxidase in amphipods
and D. magna (Wiegand et al., 2003). A more specific
mechanism of action is modulation of photosynthetic oxy-
gen release induced by HS and NOM in alga Scenedesmusarmatus, water moss Vesicularia dubyana, and hornwort
Ceratophyllum demersum (Pflugmacher et al., 1999; Stein-
berg et al., 2003).
Another specific mechanism of direct action of HS, the
activation of cytosolic aryl hydrocarbon receptor (AhR),
was suggested by Pflugmacher (unpublished results, from
Steinberg, 2003). The hypothesis of binding interaction
between HS and AhR was derived from indirect evidence
that Hsp 70 might be induced after activation of the AhR.
However, we are not aware of any research evidence of this
fact. The aim of this study was to examine whether the HA
is able to act as an activator of the AhR. For this purpose,
we used two different endpoints (reporter luciferase and
ethoxyresorufin-O-deethylase (EROD) assays) for determi-
nation of the ability of HA to elicit AhR-mediated response
in vitro in H4IIE.luc cell line. The advantage of the EROD
assay is a natural AhR-mediated expression of measured
protein activity, while the expression of luciferase, being
foreign to the cell, is probably not affected by posttranscrip-
tional and -translational events which influence CYP1A1
expression measured as EROD (Sanderson et al., 1996).
MATERIALS AND METHODS
Materials
Humic acids (HA) was purchased from Fluka, Switzerland
(prod. no. 53680). HA dissolved in 0.05 M NaOH (‘‘alkali
HA’’) was used for testing. Extraction of lipophilic organic
compounds from alkali HA solution (fragments of HA,
but also possible AhR-active organic contaminants like
PCDDs, PCDFs, PCBs, PAHs, etc.) was carried out by
liquid–liquid extraction (LLE; 3 mL of alkali HA solution,
10 g L�1, was extracted with 5 mL of hexane/dichlorome-
thane, 3:1 v/v). This LLE was repeated three times and all
organic extracts were pooled and concentrated under a
stream of nitrogen to a volume of 1 mL (‘‘HA-extract’’).
One half of the concentrated organic extract was transferred
into 1 mL dimethyl sulfoxide (DMSO) and prepared for
bioassay analysis; another half was analyzed for the pres-
ence of 16 US EPA priority PAHs using GC-MS (Hewlett-
Packard, USA) and 7 indicator PCBs (IUPAC No. 28, 52,
101, 118, 138, 153, and 180) using GC-ECD (Hewlett-
Packard, USA). One-milliliter aliquot of remaining alkali
HA solution after extraction (‘‘alkali HA remaining after
LLE’’) was also stored for subsequent bioassay analysis.
An extraction of only persistent organic compounds (e.g.,
PCDDs, PCBs, and PCDFs) from alkali HA was conducted
in a manner similar to that described earlier, but with the
addition of 5 mL of H2SO4 to the extraction mixture with
only 0.25 mL of HA (i.e., HA þ H2SO4 þ hexane/DCM)
and incubation of the mixture in water bath. The scheme
of preparation for all of the tested samples is shown in
Figure 1. Appropriate extraction control samples were pre-
pared to preclude any possible contamination with AhR
activators from the used solvents or equipment.
Cell Culture and Exposure
The H4IIE.luc cells are stably transfected with DRE-driven
firefly luciferase reporter gene construct; its transcriptional
activation occurs in an AhR-dependent manner (Sanderson
et al., 1996). Recombinant rat hepatoma H4IIE.luc cells
were grown and maintained in DMEM medium containing
10% fetal calf serum (PPA laboratories, Austria) at 5%
CO2 and 378C. Cells were grown to about 70% confluence
in a sterile 96-well plate for 24 h, and subsequently incu-
bated with reference tetrachlorodibenzo-p-dioxin (TCDD)
and tested samples for up to 24 h at 378C (final volume of
vehicle was 0.5% v/v). Cells exposed to DMEM with 0.5%
DMSO or 0.5% 0.05 M NaOH were used for the appropri-
ate vehicle controls.
Fig. 1. Preparation scheme of tested solutions as des-cribed in Materials and Methods.
339Ah RECEPTOR ACTIVATION BY PURE HUMIC ACIDS
Environmental Toxicology DOI 10.1002/tox

Luciferase Reporter Gene Assayand EROD Activity
After incubation with test substances, cells were washed
twice with phosphate-buffered saline, and luminescence
was measured with Steady-Glo Kit (Promega, USA)
according to manufacturer’s instructions, using an auto-
mated microplate fluoro-/luminometer GENios (TECAN,
Switzerland). Final values are expressed as percentage of
maximal TCDD induction (after subtraction of the solvent
control activity). EROD activity was measured by a proce-
dure slightly modified from methods described by Jung
et al. (2001). We measured fluorescence at 550 nm excita-
tion and 612 nm emission. Fluorescence was measured
using a microplate fluoro-/luminometer GENios.
Statistical Analysis
Data were examined statistically using Statistica for Win-
dows 6.0 (StatSoft, USA) and a level of significance P ¼0.05. In figures, mean 6 SD of triplicate determinations are
shown. Simple log-linear regression models were calcu-
lated for linear portions of the dose–response curves of
standard TCDD and tested HA samples. Relative lucifer-
ase-induction potency of HA (expressed as relative po-
tency, REP) was calculated using the equi-effective
approach (Jones and Anderson, 1999; Villeneuve et al.,
2002). Concentrations of the HA inducing 50% of the
TCDD-max response (CEQ-50) were compared with the
EC50 of the reference TCDD, and the REPHA ¼ CEQ-50/
EC50 was derived for the HA. For risk characterization of
AhR-active compounds concept of toxic equivalents (TEQ,
Van der Berg et al., 1998) was used. Bioassay-determined
TEQs of HA (bioassay-TEQHA) were calculated using the
equation bioassay-TEQHA ¼ [HAFluka] � REPHA. Chemi-
cal analyses-derived TEQs (chem-TEQ) of determined
PAHs were calculated using the equation chem-TEQ ¼S [PAHi � REPi], using the REP values for individual
PAHs suggested by Machala et al. (2001).
RESULTS AND DISCUSSION
The three greatest tested concentrations of HA (5, 15, and
50 mg L�1) significantly increased luciferase reporter gene
activity in H4IIE.luc cells (Fig. 2) in a dose-dependent
manner. Activity was related to reference TCDD and result-
ing REP of HA determined by luciferase induction assay
equaled 6 � 10�8. HA extract elicited practically an identi-
cal luciferase response as the original alkali HA (Fig. 2).
This indicates that almost all of the AhR activators present
in HA were extracted into the organic phase. Induction of
luciferase activity at maximal tested concentration of alkali
HA remaining after LLE was about 3.5-fold less than the
activity induced by the original alkali HA, but still signifi-
cantly greater than control sample (Fig. 2). However,
dioxin-like activity of alkali HA and HA extract were prac-
tically identical. Accordingly, we assume that removal of
extracted parts of HA has broken the balance in HA solu-
tion and thus other AhR-activating compounds could be
released from the complex HA. It was not possible to deter-
mine the REP of alkali HA remaining after LLE because
even the greatest tested concentration (50 mg L�1) did not
achieve 50% maximal standard induction. Data obtained
from EROD assay (Fig. 3) corresponded very well with the
results from luciferase assay.
The structure of humic substances (HS) studied using
multidimensional NMR is presented as a macromolecular
aggregate composed from a mixture of relatively low-mo-
lecular weight (<2kDa) organic compounds holding to-
gether through a complex combination of hydrophobic,
Fig. 2. Luciferase activity induced by TCDD, alkali HA, al-kali HA remaining after LLE, HA-extract, and H2SO4-treatedHA extract. The results are related to max. luciferase induc-tion of TCDD. Values represent the mean 6 SD of triplicatedeterminations.
Fig. 3. EROD activity induced by TCDD, alkali HA, HAextract, and alkali HA remaining after LLE related to max.EROD induction by TCDD. Values represent the mean 6 SDof triplicate determinations.
340 BITTNER ET AL.
Environmental Toxicology DOI 10.1002/tox

charge-transfer and hydrogen bond interactions, and metal
bridging (Simpson et al., 2002). This finding, in conjunc-
tion with our results, implies that activation of AhR in
H4IIE.luc cells could be caused mainly by lipophilic parts
of macromolecular ‘‘humic aggregate’’ that were released
from HA macromolecule.
Possible false-positive results caused by the contamina-
tion of HA with classical AhR activators (such as PCDDs,
PCDFs, PCBs, and PAHs) were investigated by both
H2SO4-treatment of HA extract as well as chemical analy-
ses. No significant luciferase activity of HA extract after
H2SO4 treatment ruled out possible contribution of persis-
tent AhR activators to observed luciferase activity of alkali
HA and HA extract. The bioassay data were confirmed by
chemical analyses. None of the 7 indicator PCB congeners
was detected (<LOD), and only trace concentrations of
some PAHs known to activate AhR were determined (con-
centrations in ng g�1): Fluoranthen-80, Pyren-376, Indeno
(123cd)pyren-160. Chem-TEQ calculated from chemical
data for detected PAHs equaled 4.8 � 10�2 ng TCDD g�1,
what accounted for only about 0.1% of total observed AhR-
mediated activity of HAFluka (bioassay-TEQHA �60 ng
TCDD g�1).
The ability of alkali HA and HA extracts to induce AhR
observed in our study is, at the first sight, surprising
because HA (or their fragments) has apparently different
molecular structure than the prototypical and most potent
AhR agonists such as PCDDs, PCDFs, PCBs, or PAHs
(Safe, 1990; Simpson et al., 2002). However, several stud-
ies have revealed AhR activators with very diverse struc-
tures (Denison and Heath-Pagliuso, 1998; Denison and
Nagy, 2003; Jeuken et al., 2003), and consequently AhR
might be considered as a relatively nonspecific receptor.
In conclusion, the results of our study have shown that
both alkali HA and its organic extract cause significant
inductions of AhR-mediated responses in vitro (as shown
by both reporter luciferase inductions and EROD bioas-
says). Possible false-positives due to the presence of AhR-
active contaminants were excluded by chemical analyses.
In vitro dioxin-like effects of HA were observed at environ-
mentally relevant concentrations and the determined
REPHA was 6 � 10�8. Therefore, our study indicates that
HS (or their constituents) might act as AhR-inducing sub-
stances in vivo, and can thus significantly affect various
toxicological processes. However, HS are not uniform com-
pounds; their structure and properties differ with regard to
the place of origin. Our further experiments focus on
detailed characterization of AhR modulations by a wider
range of available HS.
REFERENCES
Cheng ML, Ho HY, Chiu DTY, Lu FJ. 1999. Humic acid-medi-
ated oxidative damages to human erythrocytes: A possible
mechanism leading to anemia in Blackfoot disease. Free Radic
Biol Med 27:470–477.
Denison MS, Heath-Pagliuso S. 1998. The Ah Receptor: A Regu-
lator of the biochemical and toxicological actions of structurally
diverse chemicals. Bull Environ Contam Toxicol 61:557–568.
Denison MS, Nagy SR. 2003. Activation of the aryl hydrocarbon
receptor by structurally diverse exogenous and endogenous
chemicals. Ann Rev Pharmacol Toxicol 43:309–334.
Gau RJ, Yang HL, Suen JL, Lu FJ. 2001. Induction of oxidative
stress by humic acid through increasing intracellular iron: A
possible mechanism leading to atherothrombotic vascular disor-
der in blackfoot disease. Biochem Biophys Res Commun 283:
743–749.
Hoss S, Bergtold M, Haitzer M, Traunspurger W, Steinberg CEW.
2001. Refractory dissolved organic matter can influence the
reproduction of Caenorhabditis elegans (Nematoda). Fresh-
water Biol 46(1):1–10.
Hseu YC, Huang HW, Wang SY, Chen HY, Lu FJ, Gau RJ,
Yang HL. 2002. Humic acid induces apoptosis in human en-
dothelial cells. Toxicol Appl Pharmacol 182(1):34–43.
Huang YL, Huang TS, Lu FJ. 2003. Humic acid induces the
expression of ox-LDL receptor in HL-60 cells through activa-
tion of PPAR gamma. Bull Environ Contam Toxicol 71:429–
436.
Jeuken A, Keser BJG, Khan E, Brouwer A, Koeman J, Denison MS.
2003. Activation of the Ah receptor by extracts of dietary herbal
supplements, vegetables, and fruits. J Agric Food Chem
51:5478–5487.
Jones JM, Anderson JW. 1999. Relative potencies of PAHs and
PCBs based on the response of human cells. Environ Toxicol
Pharmacol 7:19–26.
Jung DKJ, Klaus T, Fent K. 2001. Cytochrome P450 induction by
nitrated polycyclic aromatic hydrocarbons, azaarenes, and bi-
nary mixtures in fish hepatoma cell line PLHC-1. Environ Toxi-
col Chem 20:149–159.
Liang HJ, Tsai CL, Chen PQ, Lu FJ. 1999. Oxidative injury
induced by synthetic humic acid polymer and monomer in cul-
tured rabbit articular chondrocytes. Life Sci 65:1163–1173.
Machala M, Vondracek J, Blaha L, Ciganek M, Neca J. 2001. Aryl
hydrocarbon receptor-mediated activity of mutagenic polycy-
clic aromatic hydrocarbons determined using in vitro reporter
gene assay. Mutat Res – Genet Toxicol Environ Mutagen
497(1/2):49–62.
McCarthy JF. 1987. Humic substances reduce bioavailability and
toxicity of contaminants. Abstracts of Papers of the American
Chemical Society 193:100-ENVR.
McGeer JC, Szebedinszky C, McDonald DG, Wood CM. 2002.
The role of dissolved organic carbon in moderating the bio-
availability and toxicity of Cu to rainbow trout during chronic
waterborne exposure. Comparative Biochem Physiol C 133:
147–160.
Meinelt T, Playle R, Schreckenbach K, Pietrock M. 2001. The tox-
icity of the antiparasitic mixture FMC is changed by humic sub-
stances and calcium. Aquaculture Res 32:405–410.
Mezin LC, Hale RC. 2004. Effect of humic acids on toxicity of
DDT and chlorpyrifos to freshwater and estuarine invertebrates.
Environ Toxicol Chem 23:583–590.
341Ah RECEPTOR ACTIVATION BY PURE HUMIC ACIDS
Environmental Toxicology DOI 10.1002/tox

Pflugmacher S, Spangenberg M, Steinberg CEW. 1999. Dissolved
organic matter (DOM) and effects on the aquatic macrophyte
Ceratophyllum demersum in relation to photosynthesis, pigment
pattern and activity of detoxication enzymes. J Appl Bot-
Angew Bot 73:184–190.
Safe S. 1990. Polychlorinated-biphenyls (Pcbs), dibenzo-para-di-
oxins (Pcdds), dibenzofurans (Pcdfs), and related compounds—
Environmental and mechanistic considerations which support
the development of toxic equivalency factors (TEFs). Crit Rev
Toxicol 21(1):51–88.
Sanderson JT, Aarts J, Brouwer A, Froese KL, Denison MS, Giesy
JP. 1996. Comparison of Ah receptor-mediated luciferase and
ethoxyresorufin-O-deethylase induction in H4IIE cells: Implica-
tions for their use as bioanalytical tools for the detection of
polyhalogenated aromatic hydrocarbons. Toxicol Appl Pharma-
col 137:316–325.
Simpson AJ, Kingery WL, Hayes MHB, Spraul M, Humpfer E,
Dvortsak P, Kerssebaum R, Godejohann M, Hofmann M.
2002. Molecular structures and associations of humic substan-
ces in the terrestrial environment. Naturwissenschaften 89(2):
84–88.
Steinberg CEW. 2003. Ecology of Humic Substances in Fresh-
waters—Determinants from Geochemistry to Ecological Niches.
Berlin: Springer. 432 p.
Steinberg CEW, Hoss S, Bruggemann R. 2002. Further evidence
that humic substances have the potential to modulate the repro-
duction of the nematode Caenorhabditis elegans. Int Rev Hy-
drobiol 87:121–133.
Steinberg CEW, Paul A, Pflugmacher S, Meinelt T, Klocking R,
Wiegand C. 2003. Pure humic substances have the potential to
act as xenobiotic chemicals—A review. Fresenius Environ Bull
12:391–401.
Timofeyev MA, Wiegand C, Burnison BK, Shatilina ZM, Pflug-
macher S, Steinberg CEW. 2004. Impact of natural organic matter
(NOM) on freshwater amphipods. Sci Total Environ 319(1/3):
115–121.
Van der Berg M, Birnbaum L, Bosveld ATC, Brunstrom B, Cook P,
Feeley M, Giesy JP, Hanberg A, Hasegawa R, Kennedy S, Larsen
JC, van Leeuwen FXR, Djien Liem AK, Nolt C, Peterson RE,
Poellinger L, Safe S, Schrenk S, Tillitt D, Tysklind M, Younes M,
Waern F, Zacharewski T. 1998. Toxic equivalency factors (TEFs)
for PCBs, PCDDs, PCDFs for humans and wildlife. Environ
Health Perspect 106:775–792.
Villeneuve DL, Khim JS, Kannan K, Giesy JP. 2002. Relative
potencies of individual polycyclic aromatic hydrocarbons to
induce dioxinlike and estrogenic responses in three cell lines.
Environ Toxicol 17:128–137.
Wiegand C, Meems N, Timoveyev MA, Steinberg CEW, Pflug-
macher S. 2003. More evidence for humic substances acting as
biogeochemicals on organisms. In: Ghabbour EA, Davies G,
editors. Humic substances: nature’s most versatile materials.
New York: Taylor & Francis. pp 349–361.
342 BITTNER ET AL.
Environmental Toxicology DOI 10.1002/tox

Paper II.
Janosek, J., Bittner, M., Hilscherová, K., Bláha, L., Giesy, J.P. and Holoubek, I.
AhR-mediated and antiestrogenic activity of humic substances. Chemosphere;
2006 (in press)


1
2
3
45678
910
11
1213141516171819202122
2324
25
2627282930313233343536
CHEM 7930 No. of Pages 6, Model 5+
21 December 2006; Disk UsedARTICLE IN PRESS
www.elsevier.com/locate/chemosphere
Chemosphere xxx (2006) xxx–xxx
OO
F
AhR-mediated and antiestrogenic activity of humic substances
J. Janosek a, M. Bittner a,*, K. Hilscherova a, L. Blaha a, J.P. Giesy b,c,d, I. Holoubek a
a RECETOX, Masaryk University, Brno, 625 00 Brno, Czech Republicb Department Biomedical Veterinary Sciences and Toxicology Centre, University of Saskatchewan, Saskatoon, Saskatchewan, Canada
c Zoology Department, National Food Safety and Toxicology Center, Center for Integrative Toxicology, Michigan State University,
E. Lansing, MI 48823, USAd Biology and Chemistry Department, City University of Hong Kong, Kowloon, Hong Kong, SAR, China
Received 19 May 2006; received in revised form 23 November 2006; accepted 24 November 2006
R
EC
TED
PAbstract
Humic substances (HS) were for decades regarded as inert in the ecosystems with respect to their possible toxicity. However, HS havebeen recently shown to elicit various adverse effects generally attributed to xenobiotics. In our study, we used MVLN and H4IIE-luc celllines stably transfected with luciferase gene under control of estrogen receptor (ER) and Ah receptor (AhR; receptor connected with so-called dioxin-like toxicity) for assessment of anti/estrogenic and AhR-mediated effects of 12 commercially available humic substances. Outof those, five humic acids were shown to induce AhR-mediated activity with relative potencies related to TCDD 2.6 · 10�8–7.4 · 10�8.Organic extracts of HS solutions also elicited high activities what means that lipophilic molecules are responsible for a great part of effect.However, relatively high activity remaining in extracted solution suggests also presence of polar AhR-agonists. Contribution of persistentorganic compounds to the observed effects was ruled out by H2SO4 treatment. Eight out of twelve HS elicited significant antiestrogeniceffects with IC50 ranging from 40 to 164 mg l�1. The possible explanations of the antiestrogenic effect include sorption of 17-b-estradiol(E2) on HS, changes in membrane permeability for E2 or another specific mechanism.� 2006 Elsevier Ltd. All rights reserved.
Keywords: Ah receptor; AhR-mediated activity; Antiestrogenic effects; Endocrine disruption
R
37383940414243444546474849
UN
CO
R1. Introduction
Humic substances (HS) are ubiquitous natural productsof decomposition of dead organic matter. Generally, HScan be divided into three groups: humic acids (HA), fulvicacids (FA) and humins. In the aquatic environment, HSform approximately 50–70% of dissolved/natural organicmatter (D/NOM, Timofeyev et al., 2004), which is in mostnatural freshwaters in the concentration range 0.5–50 mg l�1 (Steinberg, 2003).
Until recently, only indirect effects of HS in the ecosys-tems such as changes in bioavailability of organic pollu-tants and/or complexation of inorganic ions, particularly
50515253
0045-6535/$ - see front matter � 2006 Elsevier Ltd. All rights reserved.
doi:10.1016/j.chemosphere.2006.11.045
* Corresponding author. Tel.: + 420 549491462; fax: + 420 549492840.E-mail address: [email protected] (M. Bittner).
Please cite this article in press as: Janosek, J. et al., AhR-mediated(2006), doi:10.1016/j.chemosphere.2006.11.045
heavy metals, have been reported (Steinberg, 2003). Thepresumed large molecules of HS were not considered tobe able to penetrate into cells. However, recent studiesfocused on HS uptake showed that at least parts of naturalas well as artificial HS can be assimilated by plant cells inculture (Wang et al., 1999) as well as crustaceans (Gamma-
rus pulex) or even vertebrates (Rana arvalis tadpoles; Stein-berg et al., 2003). Furthermore, biological effects of HShave been reported for algae, invertebrates as well as verte-brates (Pflugmacher et al., 2001; Steinberg et al., 2003).
Direct toxic effects of HS were reported for the freshwa-ter snail Lymnea stagnalis. Exposure to natural concentra-tions of HS from the Suwannee River (0.5 mg l�1) causeddeath of up to 20% of individuals (Steinberg et al., 2003).Responses of nonspecific markers of toxic effects ofHS were also observed. Elevated levels of detoxifica-tion enzymes glutathione-S-transferase and glutathione
and antiestrogenic activity of humic substances, Chemosphere

T
54555657585960616263646566676869707172737475767778798081828384858687888990
91
92
93949596979899
100101102103104105106107
108
109110111112113114115116117118119120121122123124125126127128129130
131
132133134135136137138139140141142143144145146147148149150151152153154155156157158159160161
2 J. Janosek et al. / Chemosphere xxx (2006) xxx–xxx
CHEM 7930 No. of Pages 6, Model 5+
21 December 2006; Disk UsedARTICLE IN PRESS
UN
CO
RR
EC
peroxidase were reported in crustaceans Daphnia magna
following HS treatment at concentration range 0.5–100 mg l�1 (Wiegand et al., 2003). An increase in peroxi-dase activity as well as release of heat shock proteins wasalso observed in several amphipod species (Timofeyevet al., 2004) and carp (Wiegand et al., 2003). Hoss et al.(2001) reported that natural organic matter (NOM) alteredreproduction of nematodes Caenorhabditis elegans whatwas further confirmed in a study with broader spectra ofHS (Steinberg et al., 2002).
A synthetic humic substance HS1500 (a polyphenol oxi-dation product) was shown to elicit significant estrogenicand anti-thyroidal effects on Xenopus laevis tadpoles (Lutzet al., 2005). Furthermore, long-term exposure of swordtailfish Xiphophorus helleri fry to a high concentration ofHS1500 (from 5 to 180 mg l�1) lead to changes of sex ratioin favor of females as well as to body mass increase in bothmales and females (Meinelt et al., 2004), while survival offry has not decreased even at exposure to 500 mg l�1 (Stein-berg et al., 2003). However, the same very high concentra-tion of HS1500 has significantly decreased the survival rateof the exposed embryos of fish Danio rerio, whereas expo-sure to lower concentrations of 5–50 mg l�1 slightlyincreased the survival rate of embryos relative to the con-trol (Steinberg et al., 2003).
In spite of these in vivo experiments, no mechanisticin vitro studies had been conducted to determine the modeof action of direct effects. With respect to observed estro-genic effects as well as increase in levels of detoxificationenzymes, we decided to investigate possible interactions ofa wide spectrum of HS (including HA, FA and NOMobtained from various sources) with estrogen receptor(ER) and aryl hydrocarbon receptor (AhR). The latter isresponsible for so-called dioxin-like mode of action result-ing in a wide range of in vivo effects in vertebrates including,e.g. enormous enhancement of activity of detoxificationenzymes.
2. Material and methods
2.1. Chemicals
HS isolated from different matrices were purchased fromvarious sources: HA-Fluka (Product No. 53680; Fluka,Switzerland), HA sodium salt (Product No. H16752;Sigma–Aldrich, Prague, Czech Republic). The followingreference HS were purchased from the International HumicSubstances Society (IHSS, Denver, USA): Suwannee RiverHA (Product No. 2S101H), Suwannee River FA (1S101F)and Suwannee River NOM (1R101N), Florida Peat HA(1S103H) and Florida Peat FA (2S103F), Nordic AquaticFA (1R105F), Nordic Reservoir NOM (1R108N), WaskishPeat HA (1R107H), Elliot Soil HA (1S102H) and Leonar-dite HA (1S104H). Reference compounds were purchasedfrom Dr. Ehrenstorfer, Augsburg, Germany (2,3,7,8-tetra-chlorodibenzo-p-dioxin, TCDD) and Sigma–Aldrich (17-b-estradiol, E2).
Please cite this article in press as: Janosek, J. et al., AhR-mediated(2006), doi:10.1016/j.chemosphere.2006.11.045
ED
PR
OO
F
2.2. Sample preparation
HS were dissolved in 0.05 M NaOH and final concentra-tions ranging from 1.9 to 150 mg l�1 were studied. To eval-uate the nature of active molecules, further investigationswere conducted. The alkaline solution of HS was extractedthree times with a 3:1 hexane:dichloromethane mixture.Half of this extract was transferred into DMSO forin vitro assays (further regarded as organic extract); theother half was analyzed using GC–MS for known xenobiot-ics such as seven indicator PCB (IUPAC No. 28, 52, 101,118, 138, 153 and 180) and 16 PAH monitored by theUnited States Environmental Protection Agency (USEPA).To determine possible contributions of other persistentorganic compounds that might have been present in stan-dard materials, the alkaline solutions (200 ll) were trans-ferred into excess of 96% H2SO4 (5 ml) and liquid–liquidextraction with hexane:dichloromethane (3 · 5 ml) wasconducted. Using this method, only persistent compoundssuch as PCDD, PCDF and PCB were extracted. The origi-nal alkaline solutions were examined for both AhR-medi-ated and anti/estrogenic activity, organic extracts as wellas extraction residues of alkali solutions and H2SO4-treatedextracts were tested for AhR-mediated activity.
2.3. In vitro assays
H4IIE-luc (rat hepatocarcinoma) and MVLN (humanbreast carcinoma) cells stably transfected with luciferasegene under control of AhR and ER, respectively, were usedfor analysis of receptor activation. Both cell lines are wellestablished models for evaluation of AhR-mediated(H4IIE-luc) and anti/estrogenic (MVLN) activities of puresubstances as well as environmental samples (Demirpenceet al., 1993; Machala et al., 2001; Villeneuve et al., 2002).
H4IIE-luc cells: Cells were grown and maintained inDulbecco’s Modified Eagle’s Medium (DMEM) containing10% fetal calf serum (both PAA laboratories, Pasching,Austria) at 5% CO2 and 37 �C. Once the cells reachedabout 70% confluence they were passaged and seeded intoa sterile 96-well plate at density 15000 cells/well. After24 h, the cells were exposed in triplicates to the tested sam-ples (concentrations 1.9, 5.6, 16.7, 50 and 150 mg l�1) orreference compound (dilution series 0.1–100 pM TCDD)for 24 h at 37 �C (final vehicle concentration was 0.5%v/v). Cells exposed to DMEM with 0.5% DMSO and0.5% 0.05 M NaOH were used for the appropriate vehiclecontrols. Intensity of luciferase luminescence was measuredusing Promega Steady Glo Kit (Promega, Mannheim,Germany) after 24 h exposure. At least three independentassays have been conducted for each concentration tested.
MVLN cells: Cells were grown in DMEM-F12 withoutphenol red (Sigma Aldrich, USA) containing 10% fetal calfserum. For experiments, cells were grown in medium con-taining fetal calf serum treated with dextran-coated char-coal (strongly reduced concentrations of natural steroids).For studies of anti/estrogenic effects of HS, cells were
and antiestrogenic activity of humic substances, Chemosphere

T
PR
OO
F
162163164165
166
167168169170171172173174
175
176
177178179180181182183184185186187188189190191192193
194195196197198199200201202203204205
Table 1AhR-mediated and antiestrogenic activity of crude alkali solutions of HS
AhR-mediated activity Antiestrogenic activity
LOEC(mg l�1)
REP LOEC(mg l�1)
IC50
(mg l�1)
HA – Fluka 1.9 4.5 · 10�8 16.7 43 ± 3HA sodium salt 1.9 7.4 · 10�8 16.7 40 ± 4Suwannee River
HAn.ind. n.ind. n.eff. n.eff.
Elliot soil HA 1.9 8.4 · 10�8 50 164 ± 9a
Florida Peat HA 16.7 2.6 · 10�8 150 w.eff.Leonardite HA 1.9 4.3 · 10�8 50 132 ± 8Waskish Peat HA n.ind. n.ind. 50 84 ± 6Suwannee River
FAn.ind. n.ind. 50 127 ± 8
Florida Peat FA n.ind. n.ind. 50 112 ± 4Nordic Aquatic
FAn.ind. n.ind. 50 83 ± 7
Suwannee RiverNOM
n.ind. n.ind. 150 w.eff.
Nordic ReservoirNOM
n.ind. n.ind. n.eff. n.eff.
REP – relative potencies related to TCDD; IC50 – concentration causing50% inhibition of E2-induced luciferase activity; n.ind. – no induction;n.eff. – no effect; w.eff. – weak effect.
a Approximation above measured concentration range.
J. Janosek et al. / Chemosphere xxx (2006) xxx–xxx 3
CHEM 7930 No. of Pages 6, Model 5+
21 December 2006; Disk UsedARTICLE IN PRESS
REC
exposed to HS with/without simultaneous exposure to33 pM E2 with E2 calibration series ranging from 1 to100 pM. Other conditions were identical with thosedescribed for H4IIE-luc cells.
2.4. Calculations
Results were calculated using equi-effective approachdescribed by Villeneuve et al. (2000) and expressed as rela-tive potencies (REP) with respect to TCDD. REP were cal-culated from EC50 values. For determination of massbalance of extracts, toxic equivalents (TEQ) expressed asng TEQ g-1 HA were calculated from EC25 because someof the extracts and extraction residues did not reach 50%effect of maximal TCDD induction.
3. Results
3.1. AhR-mediated activity
Of the 12 HS tested, five elicited a significant AhR-med-iated activity in H4IIE-luc cells (Fig. 1). All of the activesamples were HA, while no FA and no NOM samplesshowed any substantial activity. Relative potencies (relatedto TCDD) of all the AhR-active HA were in a relativelynarrow range of 2.6–8.4 · 10�8. All of the active substanceselicited significant AhR-mediated effects at concentrationas little as 1.9 mg l�1, except for Florida Peat HA (LOEC16.7 mg l�1; see Table 1). The organic extracts of HAinduced significant activities what confirmed presence ofnon-polar AhR activators. However, the extraction resi-dues also elicited relatively great responses (Fig. 2).Increase in luciferase activities caused by the residues waseven greater than that of organic extracts (Table 2) exceptfor HA-Fluka and HA sodium salt. This result suggests asignificant contribution of polar compounds to the AhR-mediated activity of HA.
UN
CO
R
0
20
40
60
80
100
TCDD
HA -Fluk
a
HAso
dium
salt
Suwann
eeRive
r HA
Elliott S
oil H
A
Florid
a Pea
t HA
Leon
ardit
e
Was
Luci
fera
se a
ctiv
ity
(% o
f 100
pM
TC
DD
indu
ctio
n)
HS -NOM
Fig. 1. AhR-mediated activity of 12 samples of HS determined in H4IIE-luc
mean ± SD of triplicate determinations.
Please cite this article in press as: Janosek, J. et al., AhR-mediated(2006), doi:10.1016/j.chemosphere.2006.11.045
ED
Extraction with H2SO4 totally suppressed the AhR-mediated activity of HA. Thus it can be concluded thatthe compounds responsible for the observed effects werenot persistent organic compounds such as PCDD, PCDFor PCB. Moreover, chemical analysis of seven indicatorPCB congeners and 16 US EPA priority PAH (indicationof possible anthropogenic contamination) did not revealsufficient quantities of these substances that could beresponsible for the relatively great AhR-mediated activitiesof HA. TEQ calculated from chemical data for PAH usingREPs published by Machala et al. (2001) was less than 1%of bioassay-derived TEQ for the HA.
HA
kish
Peat H
A
Suwann
ee R
iver F
A
Florida
Peat F
A
Nordic
Aqu
aticFA
Suwann
ee R
iver N
OM
Nordic
Reser
voir N
OM
Humic Substances - Natural Organic Matter
FA - Fulvic AcidsHA - Humic Acids
cells; cTCDD = 3.2 · 10�5 mg l�1; cHS = 150 mg l�1. Values represent the
and antiestrogenic activity of humic substances, Chemosphere

EC
T
RO
OF
206
207208
209210211212213214215216217
218
219220221222223224
0
20
40
60
80
100
120
140TCDDLeonardite HA- alkali solutionLeonardite HA- org. extractLeonardite HA- extr. residueLeonardite HA- H2SO4 treated
Luci
fera
se a
ctiv
ity (
% o
f 100
pM
TC
DD
)
10-9 10-8 10-7 10-6 10-5 10-4 1 10 100 1000
Concentration (mg l-1)
Fig. 2. Dose–response curves of TCDD, Leonardite HA and its different solutions and extracts in H4IIE-luc cells. Values represent the mean ± SD oftriplicate determinations.
Table 2Mass balance of extraction procedure
Alkali solution(ng TEQ g�1 HA)
Extract (% ofalkali solution)
Extract residue(% of alkalisolution)
HA – Fluka 205 70.4 22.2HA sodium salt 219 67.8 31.1Elliot soil HA 197 37.5 47.1Florida Peat
HA29 w.i. 55.0
Leonardite HA 68 30.4 46.1
TEQ – toxic equivalents derived from EC25 obtained from experimentswith alkali solutions, their organic extracts and extraction residues; w.i. –weak induction.
4 J. Janosek et al. / Chemosphere xxx (2006) xxx–xxx
CHEM 7930 No. of Pages 6, Model 5+
21 December 2006; Disk UsedARTICLE IN PRESS
RR3.2. Interaction with ER pathway
None of the tested HS showed any estrogenic activityboth in absence and presence of ER-agonist E2. However,
UN
CO
0
20
40
60
80
100
120
E2
Humic
Acid- F
luka
HASod
iumSal
t
Suwann
ee R
iver HA
Elliot S
oilHA
Florid
a Pea
t HA
Leon
ardi
te
Was
Luci
fera
se a
ctiv
ity
(% o
f in
duct
ion
of 3
3 pM
E2)
Fig. 3. Antiestrogenic effects of HS in MVLN cells – HS-dependent inhibition oof triplicate determinations.
Please cite this article in press as: Janosek, J. et al., AhR-mediated(2006), doi:10.1016/j.chemosphere.2006.11.045
ED
Pten out of twelve tested HS caused significant decreases ofER-dependent gene expression (Fig. 3). Unlike the AhR-mediated activity, antiestrogenic effects were observed afteradministration of HA, FA and NOM. Concentrationscausing a 50% decrease of E2-dependent luciferase activity(IC50) were observed for HS in a concentration range of40–164 mg l�1 (see Table 1). The strongest suppression ofER signaling was caused by HA-Fluka and HA sodiumsalt.
4. Discussion
The results of this study demonstrates that some HS,particularly HA, are able to activate ‘‘dioxin-like’’ signalingpathway. However, as the phrase ‘‘dioxin-like’’ also sug-gests persistence, here we refer to this activity as AhR-med-iated. FA were found to be inactive in these assays.Accordingly, since HA represents only about 6–8% of the
HA
kish
Peat H
A
Suwann
ee R
iver FA
Florid
a Pea
t FA
Nordic
Aqu
atic F
A
Suwann
ee R
iver NOM
Nordic
Reser
voir
NOM
50 mg l -1 150 mg l -1
f luciferase activity elicited by 33 pM E2. Values represent the mean ± SD
and antiestrogenic activity of humic substances, Chemosphere

T
225226227228229230231232233234235236237238239240241242243244245246247248249250251252253254255256257258259260261262263264265266267268269270271272273274275276277278279280281
282283284285286287288289290291292293294295296297298299300301302303304305306307308309310311312313314315316317318319320321322323324325326327328329330331332333334335336337338
J. Janosek et al. / Chemosphere xxx (2006) xxx–xxx 5
CHEM 7930 No. of Pages 6, Model 5+
21 December 2006; Disk UsedARTICLE IN PRESS
UN
CO
RR
EC
total NOM (Yamamoto et al., 2004), tested NOM samplesalso did not elicit any AhR-mediated activity at any of thestudied concentrations.
The traditional view of HA structures suggests relativelylarge molecules that are not only unlikely to bind to theligand-binding region of the AhR but also unlikely to entercells. However, recent studies have shown that HS are ableto cross plant (Nardi et al., 2002) and mammalian cellmembranes and interact with some receptors (Beer et al.,2000). Moreover, recent studies of HS structure suggest itconsists of aggregates of relatively low-molecular weight(<2 kDa) organic compounds and metal ions boundtogether by non-covalent interactions (Simpson et al.,2002). This leads to a hypothesis that the AhR-active com-pounds may be rather small molecules released from thiscomplex. As for structural dissimilarity of HA with ‘‘clas-sical’’ high affinity AhR ligands such as PCDD, PCDFand PCB, a variety of AhR activators structurally very dif-ferent from the ‘‘classical’’ ligands have been identifiedrecently (Denison and Heath-Pagliuso, 1998). These‘‘non-classical’’ ligands, such as oxidized carotenoids,indoles and heterocyclic amines, are usually very weak acti-vators when compared to the halogenated aromatic hydro-carbons (HAH). HA also seem to belong to this group of‘‘non-classical’’ AhR ligands.
The adverse biological effects of HAH attributed to theAhR activation are usually thought to be due to their highaffinity to AhR and low susceptibility to biodegradation.Many of these effects can be observed only after severaldays to weeks of exposure what suggests that their causelies in long-term continuous activation of AhR-signalingpathway (Denison and Heath-Pagliuso, 1998). Thus, con-tinuous exposure to low-affinity ligands may result ineffects similar to those caused by HAH, as was shown inbrown bullhead Ameiurus nebulosus (Grady et al., 1992).On the other hand, a single activation of AhR-signalingpathway by the non-persistent ligands may also not onlycause no harm but even produces some beneficial effects(Jeuken et al., 2003). Thus, AhR-mediated activity of HAmay be of importance namely for vertebrates (i.e. animalswith ligand-binding domain of AhR) living in aquatic sys-tems with great concentrations of HS.
Concentrations of DOM in surface water are in range of afew mg l�1, in eutrophic lakes and marshes up to tens ofmg l�1 and can reach more than 100 mg l�1 in raised peatbogs (Steinberg, 2003) or pore water (Zsolnay, 1996). Asmentioned above, HA form only a minor fraction of DOM(6–8%). Since LOEC was 1.9 mg l�1 for all AhR-active HAexcept for peat HA (16.7 mg l�1), the AhR-mediated activityof HA may be of environmental concern particularly for ani-mals living in eutrophic aquatic systems, peat bogs or for soilorganisms exposed to pore water. Furthermore, ecotoxico-logical significance of AhR activity of HA can be alsoemphasized by the fact that LOEC values of all AhR-activeHA are much lower than, e.g. above mentioned HS1500 con-centration significantly reducing the survival rate of theexposed embryos of fish Danio rerio,which was 500 mg l�1,
Please cite this article in press as: Janosek, J. et al., AhR-mediated(2006), doi:10.1016/j.chemosphere.2006.11.045
ED
PR
OO
F
while survival of Xiphophorus helleri fry has not been influ-enced even at such high concentration of HS1500 (Steinberget al., 2003).
It is also important to emphasize that maximal responseobtained with AhR-active HS samples, even at very highconcentrations (150 mg l�1), reached only about half ofmaximal induction caused by standard agonist TCDD.Thus, AhR-active HA samples can be considered ‘‘partialagonists’’ because they are unable to elicit a full response.According to pharmacologic receptor principles, partialagonists could act as antagonists against potent full agonists.It is possible that AhR-active HS (weak agonists) could actu-ally antagonize the toxicity of potent AhR agonists such asTCDD, potentially reducing its toxicity in the analogousway such as, e.g. weak AhR-agonists a-naphthoflavone(Santostefano et al., 1993) or 4 0-iodoflavone (Lu et al.,1996). From this point of view, HS might help to protectwater organisms from toxicity of full potent AhR agonists.
Extraction of the alkali solution with organic solventsresulted in an interesting outcome – the AhR-mediatedactivity was distributed between both organic extract andextraction residue. The mass balance was quite good forthe more active compounds (Table 2). However, in the leastactive sample (Florida Peat HA) activity of the organicextract did not reach the 25% of induction caused by 100pM TCDD. Thus, its activity could not be calculated. Any-way, induction of AhR-mediated effects by both organicand water solutions demonstrate presence of both lipo-philic and hydrophilic AhR activators.
Antiestrogenic effects of HS are not restricted only toHA; FA as well as NOM were also shown to negativelyaffect E2 induced luciferase activity. However, this findingis not consistent with the recent results of studies on sword-tail fish Xiphophorus helleri (Meinelt et al., 2004) and Afri-can clawed frog Xenopus laevis (Lutz et al., 2005), wherefeminization of tadpoles was observed after exposure to asynthetic HS1500. However, these studies were conductedwith a synthetic humic substance (an oxidation productof caffeic acid) and whole organisms, which may causethe differences in results. Other effects on reproductionhave been observed in nematodes (Steinberg et al., 2004)where natural peat FA strongly inhibited number of off-springs of exposed individuals. A recent in vitro study withoocytes of frog Rana temporaria (Zenkevics et al., 2005)showed that HS reduced sensitivity of oocytes to gonado-tropic hormone and thus delayed their maturation. Theseauthors attribute this effect to blocking membrane trans-port of gonadotropic hormone. Changes of membrane per-meability for E2 may also be the reason of antiestrogeniceffects observed in our study. Another cause may be thesimple sorption of relatively hydrophobic molecule of E2to carbon-rich HS in test medium. This hypothesis is sup-ported by a recent study on influence of NOM on parti-tioning of E2 between water phase and an artificialmembrane (Yamamoto et al., 2004). The researchers haveobserved �30% decrease of membrane/water partitioncoefficients of E2 at concentrations as low as 3 mg l�1 of
and antiestrogenic activity of humic substances, Chemosphere

T
339340341342343344345346347348349350351
352
353354355356357358359360361362363364365
366
367368369370371372373
374
375376377378379380381382383384385386387388389390391392
393394395396397398399400401402403404405406407408409410411412413414415416417418419420421422423424425426427428429430431432433434435436437438439440441442443444445446447448449450451452453454455456457458459460
6 J. Janosek et al. / Chemosphere xxx (2006) xxx–xxx
CHEM 7930 No. of Pages 6, Model 5+
21 December 2006; Disk UsedARTICLE IN PRESS
UN
CO
RR
EC
HA (Sigma) and 20% decrease for 7.7 mg l�1 of SuwanneeRiver FA, respectively. That corresponds with our findingthat HA (Sigma) elicits much higher antiestrogenic activity.Unfortunately, Yamamoto et al. (2004) did not studygreater concentrations of HS, and therefore we cannotdirectly compare their results with ours. At such low con-centrations we have not observed greater effect than 10%,if any. On the other hand, sorption is a relatively non-spe-cific process what would suggest all samples to be active.However, four samples (two samples of NOM, one HAand one FA) elicited no or only weak antiestrogenic activ-ity even at high concentrations. Question of some morespecific mechanism of action thus still remains unsettled.
5. Conclusions
The results of this study demonstrate interactions of HSwith AhR signaling pathway. Some HA but no FA wereable to elicit significant AhR-mediated effects with relativepotencies in range of 2.6–8.4 · 10�8. Thus, although beingnatural compounds, HS may cause some adverse effects onorganisms that are continuously exposed to high concen-trations of HS, such as soil organisms or organisms frompeat bogs. In vitro antiestrogenic effects have also beendetected that may be caused by simple sorption of E2 onHS, by reduced membrane permeability for E2 caused byparts of HS molecules, or by another more specific mecha-nism. However, determination of the mode of actionrequires further studies.
Acknowledgements
We highly acknowledge financial support from researchgrants of Czech Ministry of Education (INCHEMBIOLMSM 0021622412) and Grant Agency of the Czech Repub-lic (525/05/P160). We would like to thank Dr. JanaKlanova for the analysis of HS for PCB and PAH content,and we also thank kind reviewers for their valuablecomments.
References
Beer, A.M., Lukanov, J., Sagorchev, P., 2000. The influence of fulvic andulmic acids from peat on the spontaneous contractile activity ofsmooth muscles. Phytomedicine 7, 407–415.
Demirpence, E., Duchesne, M.J., Badia, E., Gagne, D., Pons, M., 1993.MVLN cells – a bioluminescent Mcf-7-derived cell line to study themodulation of estrogenic activity. J. Steroid. Biochem. 46, 355–364.
Denison, M.S., Heath-Pagliuso, S., 1998. The Ah Receptor: a regulator ofthe biochemical and toxicological actions of structurally diversechemicals. B. Environ. Contam. Tox. 61, 557–568.
Grady, A., Fabacher, D., Frame, G., Steadman, B., 1992. Morphologicaldeformities in brown bullheads administered dietary a-Naphthoflav-one. J. Aquat. Anim. Health. 4, 7–16.
Hoss, S., Bergtold, M., Haitzer, M., Traunspurger, W., Steinberg, C.E.W.,2001. Refractory dissolved organic matter can influence the reproduc-tion of Caenorhabditis elegans(Nematoda). Freshwater Biol. 46, 1–10.
Jeuken, A., Keser, B.J.G., Khan, E., Brouwer, A., Koeman, J., Denison,M.S., 2003. Activation of the Ah receptor by extracts of dietary herbalsupplements, vegetables, and fruits. J. Agr. Food Chem. 51, 5478–5487.
461
Please cite this article in press as: Janosek, J. et al., AhR-mediated(2006), doi:10.1016/j.chemosphere.2006.11.045
ED
PR
OO
F
Lu, Y.F., Santostefano, M., Cunningham, B.D.M., Threadgill, M.D.,Safe, S., 1996. Substituted flavones as aryl hydrocarbon (Ah) receptoragonists and antagonists. Biochem. Pharmacol. 51, 1077–1087.
Lutz, I., Jie, Z., Opitz, R., Kloas, W., Ying, X., Menzel, R., Steinberg,C.E.W., 2005. Environmental signals: synthetic humic substances actas xeno-estrogen and affect the thyroid system of Xenopus laevis.Chemosphere 61, 1183–1188.
Machala, M., Vondracek, J., Blaha, L., Ciganek, M., Neca, J., 2001. Arylhydrocarbon receptor-mediated activity of mutagenic polycyclic aro-matic hydrocarbons determined using in vitro reporter gene assay.Mutat. Res. Gen. Tox. En. 497, 49–62.
Meinelt, T., Schreckenbach, K., Knopf, K., Wienke, A., Stuber, A.,Steinberg, C.E.W., 2004. Humic substances affect physiological con-dition and sex ratio of swordtail (Xiphophorus helleri Heckel). Aquat.Sci. 66, 239–245.
Nardi, S., Pizzeghello, D., Muscolo, A., Vianello, A., 2002. Physiologicaleffects of humic substances on higher plants. Soil Biol. Biochem. 34,1527–1536.
Pflugmacher, S., Tidwell, L.F., Steinberg, C.E.W., 2001. Dissolved humicsubstances can directly affect freshwater organisms. Acta Hydroch.Hydrob. 29, 34–40.
Santostefano, M., Merchant, M., Arellano, L., Morrison, V., Denison,M.S., Safe, S., 1993. a-Naphthoflavone-induced Cyp1A1 gene-expres-sion and cytosolic aryl hydrocarbon receptor transformation. Mol.Pharmacol. 43, 200–206.
Simpson, A.J., Kingery, W.L., Hayes, M.H.B., Spraul, M., Humpfer, E.,Dvortsak, P., Kerssebaum, R., Godejohann, M., Hofmann, M., 2002.Molecular structures and associations of humic substances in theterrestrial environment. Naturwissenschaften 89, 84–88.
Steinberg, C.E.W., 2003. Ecology of Humic Substances in Freshwaters –Determinants from Geochemistry to Ecological Niches. Springer,Berlin.
Steinberg, C.E.W., Hoss, S., Bruggemann, R., 2002. Further evidence thathumic substances have the potential to modulate the reproduction ofthe nematode Caenorhabditis elegans. Int. Rev. Hydrobiol. 87, 121–133.
Steinberg, C.E.W., Paul, A., Pflugmacher, S., Meinelt, T., Klocking, R.,Wiegand, C., 2003. Pure humic substances have the potential to act asxenobiotic chemicals – a review. Fresen. Environ. Bull. 12, 391–401.
Steinberg, C.E.W., Hoss, S., Kloas, W., Lutz, I., Meinelt, T., Pflugmacher,S., Wiegand, C., 2004. Hormonelike effects of humic substances onfish, amphibians, and invertebrates. Environ. Toxicol. 19, 409–411.
Timofeyev, M.A., Wiegand, C., Burnison, B.K., Shatilina, Z.M., Pflugm-acher, S., Steinberg, C.E.W., 2004. Impact of natural organic matter(NOM) on freshwater amphipods. Sci. Total Environ. 319, 115–121.
Villeneuve, D.L., Blankenship, A.L., Giesy, J.P., 2000. Derivation andapplication of relative potency estimates based on in vitro bioassayresults. Environ. Toxicol. Chem. 19, 2835–2843.
Villeneuve, D.L., Khim, J.S., Kannan, K., Giesy, J.P., 2002. Relativepotencies of individual polycyclic aromatic hydrocarbons to inducedioxin-like and estrogenic responses in three cell lines. Environ.Toxicol. 17, 128–137.
Wang, W.H., Bray, C.M., Jones, M.N., 1999. The fate of C-14-labelledhumic substances in rice cells in culture. J. Plant Physiol. 154, 203–211.
Wiegand, C., Meems, N., Timoveyev, M.A., Steinberg, C.E.W., Pflugm-acher, S., 2003. More evidence for humic substances acting asbiogeochemicals on organisms. In: Ghabbour, E.A., Davies, G.(Eds.), Humic Substances Nature’s Most Versatile Materials. Taylorand Francis, Inc., New York, pp. 349–361.
Yamamoto, H., Liljestrand, H.M., Shimizu, Y., 2004. Effects of dissolvedorganic matter surrogates on the partitioning of 17 b-estradiol and p-nonylphenol between synthetic membrane vesicles and water. Environ.Sci. Technol. 38, 2351–2358.
Zenkevics, H., Klavins, M., Vose, V., Bucena, A., 2005. Humic acidreduces gonadotropin activity and hormonal sensitivity of frogoocytes. Aquat. Toxicol. 75, 380–383.
Zsolnay, A., 1996. Dissolved humus in soil waters. In: Piccolo, A. (Ed.),Humic Substances in Terrestrial Ecosystems. Elsevier, Amsterdam,pp. 171–224.
and antiestrogenic activity of humic substances, Chemosphere

Paper III.
Bittner, M., Hilscherová, K. and Giesy J.P. Changes of AhR-mediated activity of
humic substances after irradiation; 2006 (submitted to Environment
International)


Changes of AhR-mediated Activity of Humic Substances after
Irradiation
M. Bittner1*, K. Hilscherová1, J. P. Giesy2,3,4
1 RECETOX, Masaryk University, Brno, Czech Republic, Kamenice 126/3, 625 00
Brno, Czech Republic
2 Dept. Biomedical Veterinary Sciences and Toxicology Centre, University of
Saskatchewan, Saskatoon, Saskatchewan, Canada
3 Zoology Department, National Food Safety and Toxicology Center, Center for
Integrative Toxicology, Michigan State University, E. Lansing, MI 48823
4 Biology and Chemistry Department, City University of Hong Kong, Kowloon, Hong
Kong, SAR, China
* Corresponding Author: Address:
RECETOX
Kamenice 126/3
625 00 Brno
Czech Republic
E-mail: [email protected]
Phone: +420 549491462
Fax: +420 549492840

Abstract
Humic substances (HS) are natural organic compounds ubiquitous in the
environment. However, some studies indicate that HS can act as xenobiotics, e.g. induce
hormone-like effects in fish, amphibians and invertebrates. HS molecules contain
number of aromatic rings and conjugated double bonds - the so called chromophores.
That implies irradiation of dissolved HS can lead to a series of photochemical reactions
which can act on the HS itself, or on other substances present in aquatic environment
along with HS such as e.g. xenobiotics. In our previous study, we have found significant
interactions of five humic acids (HA) with cytosolic aryl hydrocarbon receptor (AhR) in
a in vitro bioassay based on H4IIE-luc cells. In the present study, we have studied the
changes in AhR-mediated activities of HS after irradiation that simulated natural solar
light. Twelve different HS were irradiated in Pyrex tubes with a medium-pressure
mercury lamp for a duration of from 0 to 52 h (which corresponds to 0-52 d natural
solar radiation). Original concentrations of HS samples were 50 mg L-1, and the greatest
concentration of HS photoproducts subsequently tested in the bioassay was 17 mg L-1,
which is an environmentally relevant concentration. After irradiation the absorbances of
all of the HS were less than the original materials. The AhR-mediated activity of the
HA-Fluka and HA Sodium Salt were partially decreased by irradiation. The activities of
other HS, that were either AhR-active or -inactive were not changed by irradiation. The
results of the study demonstrate that AhR-mediated activities of two active HA is
caused by both photo-stable and photo-labile AhR activators, while other three active
HA contain only photo-stable AhR activators. Potential mechanisms of the observed
irradiation-induced changes in AhR-mediated activities are discussed.
Keywords: Humic acids, Ah Receptor, H4IIE-luc, Absorbance

1. Introduction
Humic acids (HA) and fulvic acids (FA) are important fractions of the group of
organic compounds called humic substances (HS) that are ubiquitous natural products
of decomposition of dead organic matter. In the aquatic environment, HS form
approximately 50-70% of dissolved/natural organic matter (D/NOM, Timofeyev et al.,
2004) that is found in most natural freshwaters at concentrations of 0.5 to 50 mg L-1, but
can occur at concentrations as great as 100 mg L-1 in raised peat bogs (Steinberg, 2003),
where the HA fraction forms about 6-8% of the total NOM (Yamamoto et al., 2004).
Although HS have been regarded to be without any direct biological activity,
recently it has been found that HS alone can act as xenobiotic chemicals. One of the
rather non-specific mechanisms of action is NOM-dependent modulation of heat shock
proteins 70 in carp and Daphnia magna, and induction of biotransformation enzymes
glutathione-S-transferases, oxidative stress enzymes peroxidase and glutathione
peroxidase in amphipods (Wiegand et al., 2003). A more specific mechanism of action
is both HS and NOM-dependent modulation of photosynthetic oxygen release in alga
Scenedesmus armatus, water moss Vesicularia dubyana and hornwort Ceratophyllum
demersum (Pflugmacher et al., 2006; Steinberg et al., 2003). Additionally, it has been
reported that synthetic HA (HS1500) affect physiological condition and slightly the sex
ratio of the swordtail fish (Xiphophorus helleri, Meinelt et al., 2001). Furthermore,
hormone-like effect of HS on the nematode Caenorhabditis elegans (Hoss et al., 2001;
Steinberg et al., 2002), and increased mortality of amphipods and biochemical changes
have been found (Timofeyev et al., 2004).
Dissolved HS are yellow colored and exert relatively strong absorption in the
UV region of solar radiation (Hessen and Faerovig, 2001; Rasmussen et al., 1989) that
is linked to the presence of delocalized π-electron systems, which are available from
aromatic rings or conjugated double bonds (Steinberg, 2003). Photo-transformation of
HS in surface waters is a natural biogeochemical process that leads to changes of their
biological, chemical, and physical properties (Amador et al., 1991; Frimmel, 1994,
1998; Polewski et al., 2005). Experiments with irradiation of NOM samples performed
by Frimmel et al. (1998) have shown that there is a general bleaching effect (i.e.
decrease of absorbance) in the irradiated samples accompanied with a change in the
molecular size distribution of NOM. From an ecotoxicological point of view, photo-
degradation of HS can lead to either beneficial or harmful effects on water organism.

The beneficial effect has been shown for bacterial population since photo-degradation
of HS leads to the release of more easily bioavailable low-molecular organic molecules
that can stimulate bacterial production (Bertilsson and Tranvik, 1998; Wetzel et al.,
1995). On the other hand, irradiation of HS induces production of reactive oxygen
molecules (ROS, Frimmel, 1994) that can restrict viability of aquatic organisms, such as
bacteria (Scully et al., 2003), invertebrates such as the water flea (Daphnia magna,
Frimmel, 1998) and algae Selenastrum capricornutum (Gjessing and Kallovist, 1991).
However, ecotoxicological studies on the effects of irradiated HS on aquatic organisms
are still rare.
In our previous in vitro studies (Bittner et al., 2006; Janosek et al., 2006), we
have described specific mechanism of HS action, such as induction of biotransformation
enzymes via activation of intracellular aryl hydrocarbon receptor (AhR) by HA. The
activation of AhR by HA was also confirmed by in vivo experiments using the
Amazonian fish tambaqui (Colossoma macropomum, Matsuo et al., 2006). In that
study, induction of Cytochrome P450 1A (the biomarker of exposure to pollutants) by
HA-Fluka was shown by use of Western blot analysis, catalytic assay, and
immunohistochemistry.
The traditional view of HS structures suggests relatively large molecules that are
not only unlikely to bind with the ligand binding region of the AhR but also unlikely to
enter cells. However, recent studies have shown that HS are able to cross plant (Nardi et
al., 2002) and mammalian cell membranes and interact with some receptors (Beer et al.,
2000). Moreover, recent studies suggest that the HS structure consists of aggregates of
relatively low-molecular weight (<2 kDa) organic compounds and metal ions bound
together by non-covalent interactions (Simpson et al., 2002). This leads to a hypothesis
that the AhR-active compounds may be rather small molecules released from this
complex. As mentioned above, irradiation of HS leads to decrease of their mean
molecular weight. Thus, we hypothesized irradiation of HS could enhance amount of
smaller molecules that can easier penetrate cell membrane and subsequently interact
with intracellular AhR. In our present study, we have evaluated the changes in ability
of irradiated HS to activate the AhR. First of all, we irradiated 12 HS samples under
laboratory conditions. After that, subsequent changes in both biological and absorption
properties of irradiated HA samples were studied.

2. Materials and methods
2.1. Materials Humic substances (HS) isolated from different matrices were purchased from
various sources: Humic acid (HA-Fluka; product No. 53680) from Fluka, Switzerland,
Humic acid sodium salt (HA Sodium Salt; Product No. H16752) from Sigma Aldrich,
USA. The following reference HS were purchased from IHSS, USA: Suwannee River
HA (Product No. 2S101H), Suwannee River FA (1S101F) and Suwannee River Natural
Organic Matter (NOM; 1R101N), Florida Peat HA (1S103H) and Florida Peat FA
(2S103F), Nordic Aquatic FA (1R105F), Nordic Reservoir NOM (1R108N), Waskish
Peat HA (1R107H), Elliot Soil HA (1S102H) and Leonardite HA (1S104H). Reference
compound 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) was purchased from Dr.
Ehrenstorfer, Germany. HS used for irradiation were first dissolved in 0.05 M NaOH to
obtain stock concentration 2 g L-1, and after that these solutions were diluted by distilled
water to obtain final concentration 50 mg L-1 used for irradiation experiments.
2.2. Cell Culture and Exposure The H4IIE-luc cells are stably transfected with a DRE-driven firefly luciferase
reporter gene construct; the transcriptional activation of which occurs in an AhR-
dependent manner (Sanderson et al., 1996). Recombinant rat hepatoma H4IIE-luc cells
were grown and maintained in Dulbecco's Modified Eagle's Medium (DMEM)
containing 10% fetal calf serum (PAA laboratories, Austria) at 5% CO2 and 37 °C.
Cells were grown to about 70% confluence in a sterile 96-well plate for 24 h, and
subsequently incubated with reference TCDD (dissolved in DMSO) and tested samples
(with addition of corresponding amount of DMSO) for up to 24 h at 37 °C.
Concentration of irradiated HS tested was 17 mg L-1, that is concentration relevant for
environmental conditions. Cells exposed to DMEM with 0.5% DMSO were used for the
appropriate vehicle controls.
2.3. Luciferase Reporter Gene Assay After incubation with test substances, cells were washed twice with phosphate-
buffered saline, and luminescence was measured with Steady-Glo Kit (Promega, USA)
according to manufacturer’s instructions using an automated microplate luminometer

Ascent (Thermo Electron Corp., USA). Final values are expressed as a percentage of
maximal TCDD induction (after subtraction of the solvent control activity).
2.4. Irradiation and measurement of absorbance HS samples were irradiated in 13 × 100 mm Pyrex (eliminates wavelengths
below 300 nm) tubes, sealed with septa using a water-cooled 125W medium-pressure
mercury lamp (Teslamp, Czech Rep.). The concentration of all irradiated HS samples in
aqueous solution was 50 mg L-1. Sub-samples (2 mL) of the irradiated samples were
taken at the following times [h]: 0, 0.5, 1, 3, 6, 12, 20, 37 and 52, where approximately
one hour of lab-irradiation represents one day of direct natural solar irradiation. All
these irradiated samples were used for both bioassay and spectro-photometric
measurement. Absorbance of irradiated HS samples was measured in plastic cuvettes
using UV-VIS spectrophotometer Cary 50 Bio (Varian, USA) at range of wavelengths
250 - 650 nm.
2.5. Statistical analysis and calculations Bioassay-derived data were examined statistically using Statistica for Windows
6.0 (StatSoft, USA) at level of significance of P = 0.05. In figures, means ± SD of
triplicate determinations are shown. The same software was used for calculation of one
phase exponential decay equations for both absorbance and bioassay data. Half-life
values (t1/2) of both absorbance and AhR-mediated activity decreases were obtained
from the calculated one phase exponential decay equations, and mean rate of decrease
from the highest value (time 0) of absorbance/AhR-mediated activity to plateau value
(not to zero). Plateau values of one phase exponential decay equations are the intercepts
of the equations in Table 1.
3. Results
Based on our previous findings (Janosek et al., 2006), only five HA out of 12 HS
tested elicited a significant AhR-mediated activity in H4IIE-luc cells. Relative potencies
(REP) and LOEC values of these five HA samples are summarized in Table 1. The
AhR-active samples were HA-Fluka, HA Sodium Salt, Elliot Soil HA, Florida Peat HA
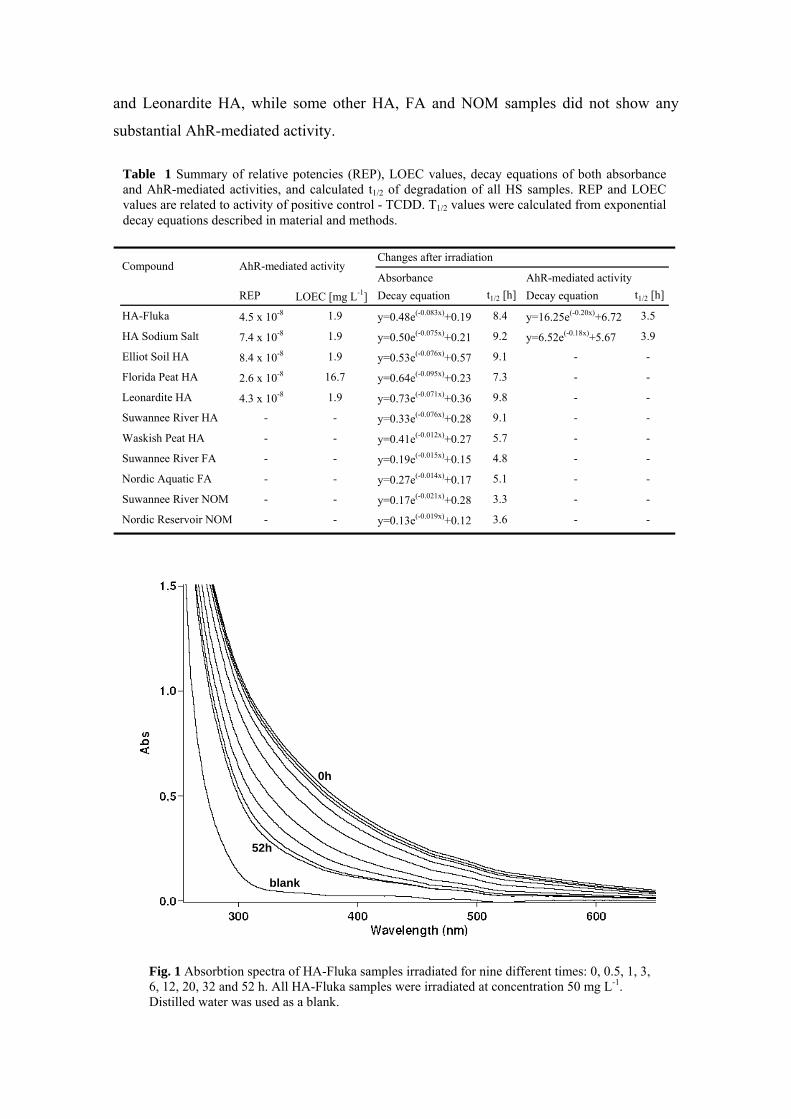
and Leonardite HA, while some other HA, FA and NOM samples did not show any
substantial AhR-mediated activity.
REP LOEC [mg L-1] Decay equation t1/2 [h] Decay equation t1/2 [h]
HA-Fluka 4.5 x 10-8 1.9 y=0.48e(-0.083x)+0.19 8.4 y=16.25e(-0.20x)+6.72 3.5
HA Sodium Salt 7.4 x 10-8 1.9 y=0.50e(-0.075x)+0.21 9.2 y=6.52e(-0.18x)+5.67 3.9
Elliot Soil HA 8.4 x 10-8 1.9 y=0.53e(-0.076x)+0.57 9.1 - -
Florida Peat HA 2.6 x 10-8 16.7 y=0.64e(-0.095x)+0.23 7.3 - -
Leonardite HA 4.3 x 10-8 1.9 y=0.73e(-0.071x)+0.36 9.8 - -
Suwannee River HA - - y=0.33e(-0.076x)+0.28 9.1 - -
Waskish Peat HA - - y=0.41e(-0.012x)+0.27 5.7 - -
Suwannee River FA - - y=0.19e(-0.015x)+0.15 4.8 - -
Nordic Aquatic FA - - y=0.27e(-0.014x)+0.17 5.1 - -
Suwannee River NOM - - y=0.17e(-0.021x)+0.28 3.3 - -
Nordic Reservoir NOM - - y=0.13e(-0.019x)+0.12 3.6 - -
CompoundAhR-mediated activity Absorbance
Changes after irradiationAhR-mediated activity
Table 1 Summary of relative potencies (REP), LOEC values, decay equations of both absorbance and AhR-mediated activities, and calculated t1/2 of degradation of all HS samples. REP and LOEC values are related to activity of positive control - TCDD. T1/2 values were calculated from exponential decay equations described in material and methods.
52h
blank
0h
Fig. 1 Absorbtion spectra of HA-Fluka samples irradiated for nine different times: 0, 0.5, 1, 3,6, 12, 20, 32 and 52 h. All HA-Fluka samples were irradiated at concentration 50 mg L-1. Distilled water was used as a blank.
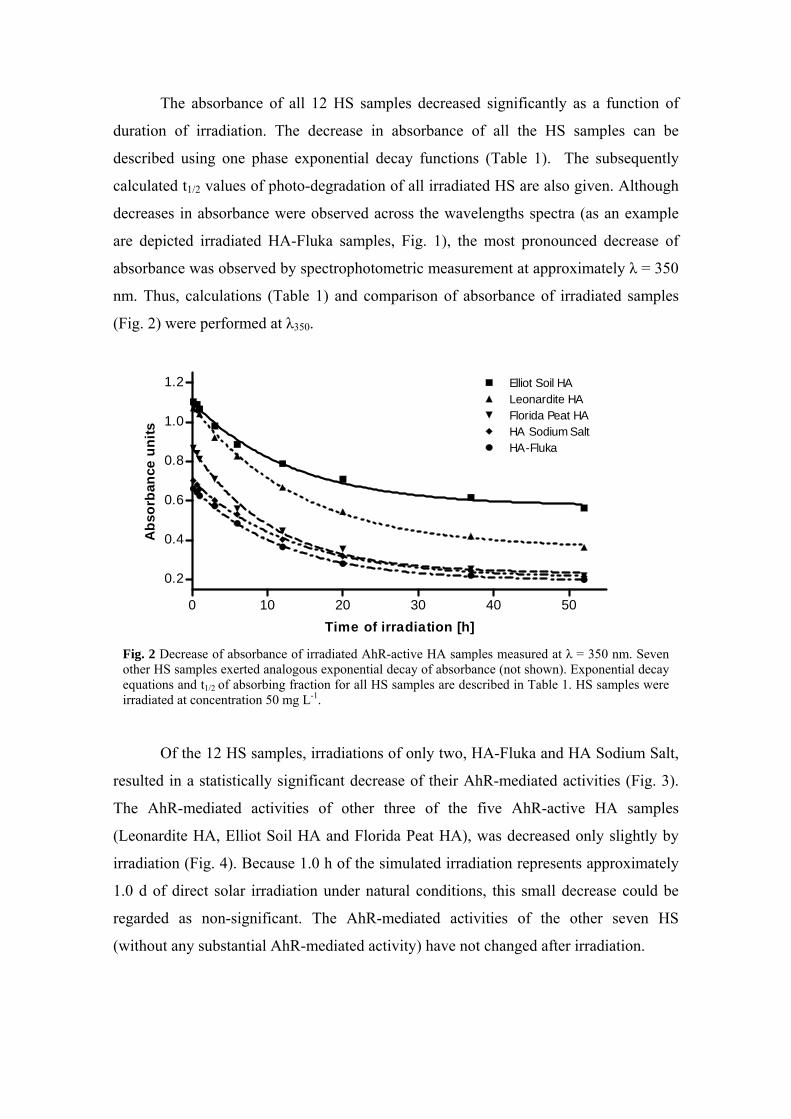
The absorbance of all 12 HS samples decreased significantly as a function of
duration of irradiation. The decrease in absorbance of all the HS samples can be
described using one phase exponential decay functions (Table 1). The subsequently
calculated t1/2 values of photo-degradation of all irradiated HS are also given. Although
decreases in absorbance were observed across the wavelengths spectra (as an example
are depicted irradiated HA-Fluka samples, Fig. 1), the most pronounced decrease of
absorbance was observed by spectrophotometric measurement at approximately λ = 350
nm. Thus, calculations (Table 1) and comparison of absorbance of irradiated samples
(Fig. 2) were performed at λ350.
0 10 20 30 40 50
0.2
0.4
0.6
0.8
1.0
1.2 Elliot Soil HALeonardite HAFlorida Peat HAHA Sodium SaltHA-Fluka
Time of irradiation [h]
Abs
orba
nce
units
Fig. 2 Decrease of absorbance of irradiated AhR-active HA samples measured at λ = 350 nm. Sevenother HS samples exerted analogous exponential decay of absorbance (not shown). Exponential decayequations and t1/2 of absorbing fraction for all HS samples are described in Table 1. HS samples wereirradiated at concentration 50 mg L-1.
Of the 12 HS samples, irradiations of only two, HA-Fluka and HA Sodium Salt,
resulted in a statistically significant decrease of their AhR-mediated activities (Fig. 3).
The AhR-mediated activities of other three of the five AhR-active HA samples
(Leonardite HA, Elliot Soil HA and Florida Peat HA), was decreased only slightly by
irradiation (Fig. 4). Because 1.0 h of the simulated irradiation represents approximately
1.0 d of direct solar irradiation under natural conditions, this small decrease could be
regarded as non-significant. The AhR-mediated activities of the other seven HS
(without any substantial AhR-mediated activity) have not changed after irradiation.

0 10 20 30 40 500
5
10
15
20
25
HA-FlukaHA Sodium Salt
Time of irradiation [h]
Luci
fera
se a
ctiv
(% o
f TC
DD
max
indu
cton
)ity
i
AhR-m
Salt (F
58% o
74% o
irradia
activit
signif
Fig. 3 Decrease of AhR-mediated activities of irradiated HA-Fluka and HA Sodium Salt.The results are related to maximal standard (100 pM TCDD) induction. Values represent themean ± SD of triplicate determinations. Exponential decay equations and t1/2 of AhR-activefraction are described in Table 1. Concentration of tested HA samples was 17 mg L-1.
0 10 20 30 40 500
5
10
15
20
25 Leonardite HAElliot Soil HAFlorida Peat HA
Time of irradiation [h]
Luci
fera
se a
ctiv
ity(%
of T
CD
Dm
ax in
duct
ion)
Fig. 4 AhR-mediated activities of irradiated Leonardite HA, Elliot Soil HA and Florida Peat HA. The results are related to maximal standard (100 pM TCDD) induction. Values represent the mean ± SD of triplicate determinations. Concentration of tested HA samples was 17 mg L-1.
The intrinsic AhR-mediated activity of HA-Fluka as well as the decrease of its
ediated activity after irradiation was greater compared to those of HA Sodium
ig. 3). After 6 h irradiation, the AhR-mediated activity of HA-Fluka decreased to
f original activity of non-irradiated sample, while HA Sodium Salt decreased to
f original activity after the same time. Nevertheless, even after 52 h of laboratory
tion, the photo-induced decay of HA-Fluka and HA Sodium Salt AhR-mediated
ies did not reach zero. The AhR-mediated activity of both HA samples reached
icant plateau values - 6.7% for HA-Fluka and 5.7% for HA Sodium Salt (i.e. % of

maximal standard TCDD induction), which corresponds to 29% of original activity of
HA-Fluka sample, resp. 46% of original activity of HA Sodium Salt sample. The rate of
decrease of the AhR-mediated activity of both HA samples exhibited t1/2 values of 3.5 h
and 3.9 h, for the HA-Fluka and sodium salt, respectively. This t1/2 values are
equivalent to approximately 3.5 and 3.9 d under natural solar irradiation.
4. Discussion
The fate and properties of HS as naturally occurring compounds are significantly
affected by the environmental conditions common in the water environment, including
solar irradiation. Since HS also exert significant biological activity, we have evaluated
impact of irradiation on both absorption properties and AhR-mediated activity.
The results of the studies which are reported here, demonstrate that all 12 of the HS
elicited significant photo-bleaching effect, as measured by absorbance, when exposed to
simulated irradiation. These results are similar to those reported by other researchers
(Frimmel, 1998; Grzybowski, 2000). Decoloration of HS-solutions is caused by
decrease of the mean molecular size of light absorbing organic matter with release of
molecules with lower mean molecular size (Bertilsson and Tranvik, 1998; Opsahl and
Benner, 1998). Based on this fact, we developed the hypothesis that smaller molecules
produced by photo-degradation of HS could be more able to activate AhR than the
original HS samples. This hypothesis can be rejected, since either no change or in some
cases even a decrease in AhR-mediated activities after irradiation were observed.
Based on the results of our study, we now suppose that decrease of AhR-
mediated activity of both HA-Fluka and HA Sodium Salt after simulated irradiation
could be related to indirect photolysis of AhR-active parts of HA by reactive oxygen
molecules (ROS). Our previous results (Janosek et al., 2007) suggest the AhR-
activating agents of HS are both hydrophilic and lipophilic non-persistent molecules,
such as polycyclic aromatic hydrocarbons (PAHs, Grady et al., 1992; Safe, 1990) and
their various, often more hydrophilic derivatives e.g. aza-PAHs (Sovadinova et al.,
2006).
It is well known that ROS are generated in surface water after photo-chemical
excitation of HS (Aguer et al., 1999; Frimmel, 1994; Hoigne et al., 1989), and are able
to oxidize surrounding dissolved organic molecules. HS-induced ROS production has
been shown to oxidize numerous chemicals including DDT (Kulovaara et al., 1995) or

pesticides Irgarol, fenuron (Amine-Khodja et al., 2006), carboxin, oxycarboxin, and
vinclozolin (Hustert and Moza, 1997; Hustert et al., 1999). The photolysis of these
compounds can change their properties including toxicity, in some cases it can even
significantly increase their toxic potential, such as in the case of vinclozolin. These
observations together with our results indicate that AhR-active agents within HS could
be degraded by ROS leading to products with different toxicological properties.
Either a decrease or no significant changes of AhR-mediated activity of HS
samples after irradiation were observed in our studies. Nevertheless, the photo-induced
decrease of AhR-mediated activity of HA-Fluka and HA Sodium Salt also show photo-
induced degradation of AhR-active fraction or compartments of HA is not complete, but
only to a certain plateau of the AhR-mediated activity. Then the activity did not
decrease further, which was similar that observed for the other three AhR active HA
samples. These findings suggest a combination of photo-stable and photo-labile AhR
activators in both HA samples with decrease of their AhR-mediated activity (HA-Fluka
and HA Sodium Salt, Fig. 3), and presence of only photo-stable AhR-activators in the
other three AhR-active HA samples without any significant decrease in AhR-mediated
activity (Elliot Soil HA, Leonardite HA and Florida Peat HA, Fig.4).
Overall, the results of our studies suggest that potency of HA to act through this
specific mechanism of action - activation of AhR - either stays the same or decreases
after irradiation. Thus, in some cases, irradiation of dissolved HA can be beneficial in
the sense of decrease of AhR active substances based stress to aquatic vertebrates.
However, generalization of this ecotoxicological conclusion is strongly limited to fact
that each HS is of different origin and thus can elicit rather specific properties.
Acknowledgement
We highly acknowledge scientific support of Dr. Jaroslav Janosek, Dr. Ludek Blaha and
prof. Ivan Holoubek. This work was supported by grant GACR 525/05/P160 and
Ministry of Education Grant (Project "INCHEMBIOL" VZ0021622412 of RECETOX,
Masaryk University).

References
Aguer, J. P., Richard, C., and Andreux, F. Effect of light on humic substances: Production of reactive species. Analusis 27:387-390; 1999
Amador, J. A., Alexander, M., and Zika, R. G. Degradation of Aromatic-Compounds Bound to Humic-Acid by the Combined Action of Sunlight and Microorganisms. Environ. Toxicol. Chem. 10:475-482; 1991
Amine-Khodja, A., Richard, C., Lavedrine, B., Guyot, G., Trubetskaya, O., and Trubetskoj, O. Water-soluble fractions of composts for the photodegradation of organic pollutants in solar light. Environ. Chem. Lett. 3:173-177; 2006
Beer, A. M., Lukanov, J., and Sagorchev, P. The influence of fulvic and ulmic acids from peat, on the spontaneous contractile activity of smooth muscles. Phytomedicine 7:407-415; 2000
Bertilsson, S., and Tranvik, L. J. Photochemically produced carboxylic acids as substrates for freshwater bacterioplankton. Limnol. Oceanogr. 43:885-895; 1998
Bittner, M., Janosek, J., Hilscherova, K., Giesy, J., Holoubek, I., and Blaha, L. Activation of Ah receptor by pure humic acids. Environ. Toxicol. 21:338-342; 2006
Frimmel, F. H. Photochemical Aspects Related to Humic Substances. Environ. Int. 20:373-385; 1994
Frimmel, F. H. Impact of light on the properties of aquatic natural organic matter. Environ. Int. 24:559-571; 1998
Gjessing, E. T., and Kallovist, T. Algicidal and Chemical Effect of Uv-Radiation of Water Containing Humic Substances. Water Res. 25:491-494; 1991
Grady, A. W., Fabacher, D. L., Frame, G., and Steadman, B. L. Morphological deformities in brown bullheads administered dietary b-naphthoflavone. J. Aquat. Anim. Health, 4:7-16; 1992
Grzybowski, W. Effect of short-term sunlight irradiation on absorbance spectra of chromophoric organic matter dissolved in coastal and riverine water. Chemosphere 40:1313-1318; 2000
Hessen, D. O., and Faerovig, P. J. The photoprotective role of humus-DOC for Selenastrum and Daphnia. Plant Ecol. 154:261-+; 2001
Hoigne, J., Faust, B. C., Haag, W. R., Scully, F. E., and Zepp, R. G. Aquatic Humic Substances as Sources and Sinks of Photochemically Produced Transient Reactants. Acs Sym. Ser. 219:363-381; 1989
Hoss, S., Bergtold, M., Haitzer, M., Traunspurger, W., and Steinberg, C. E. W. Refractory dissolved organic matter can influence the reproduction of Caenorhabditis elegans (Nematoda). Freshwater Biol. 46:1-10; 2001
Hustert, K., and Moza, P. N. Photochemical degradation of dicarboximide fungicides in the presence of soil constituents. Chemosphere 35:33-37; 1997
Hustert, K., Moza, P. N., and Kettrup, A. Photochemical degradation of carboxin and oxycarboxin in the presence of humic substances and soil. Chemosphere 38:3423-3429; 1999

Janosek, J., Bittner, M., Hilscherová, K., Bláha, L., Giesy, J.P., and Holoubek, I. AhR-mediated and antiestrogenic activity of humic substances. Chemosphere; 2006 (in press)
Kulovaara, M., Backlund, P., and Corin, N. Light-Induced Degradation of Ddt in Humic Water. Sci. Total Environ. 170:185-191; 1995
Matsuo, A. Y. O., Woodin, B. R., Reddy, C. M., Val, A. L., and Stegeman, J. J. Humic substances and crude oil induce cytochrome P450 1A expression in the Amazonian fish species Colossoma macropomum (Tambaqui). Environ. Sci. Technol. 40:2851-2858; 2006
Meinelt, T., Playle, R., Schreckenbach, K., and Pietrock, M. The toxicity of the antiparasitic mixture FMC is changed by humic substances and calcium. Aquac. Res. 32:405-410; 2001
Nardi, S., Pizzeghello, D., Muscolo, A., and Vianello, A. Physiological effects of humic substances on higher plants. Soil Biol. Biochem. 34:1527-1536; 2002
Opsahl, S., and Benner, R. Photochemical reactivity of dissolved lignin in river and ocean waters. Limnol. Oceanogr. 43:1297-1304; 1998
Pflugmacher, S., Pietsch, C., Rieger, W., and Steinberg, C. E. W. Dissolved natural organic matter (NOM) impacts photosynthetic oxygen production and electron transport in coontail Ceratophyllum demersum. Sci. Total Environ. 357:169-175; 2006
Polewski, K., Slawinska, D., Slawinski, J., and Pawlak, A. The effect of UV and visible light radiation on natural humic acid EPR spectral and kinetic studies. Geoderma 126:291-299; 2005
Rasmussen, J. B., Godbout, L., and Schallenberg, M. The Humic Content of Lake Water and Its Relationship to Watershed and Lake Morphometry. Limnol. Oceanogr. 34:1336-1343
Safe, S. Polychlorinated-Biphenyls (Pcbs), Dibenzo-Para-Dioxins (Pcdds), Dibenzofurans (Pcdfs), and Related-Compounds - Environmental and Mechanistic Considerations Which Support the Development of Toxic Equivalency Factors (Tefs). Crit. Rev. Toxicol. 21:51-88; 1990
Scully, N. M., Cooper, W. J., and Tranvik, L. J. Photochemical effects on microbial activity in natural waters: the interaction of reactive oxygen species and dissolved organic matter. Fems Microbiol. Ecol. 46:353-357; 2003
Simpson, A. J., Kingery, W. L., Hayes, M. H. B., Spraul, M., Humpfer, E., Dvortsak, P., Kerssebaum, R., Godejohann, M., and Hofmann, M. Molecular structures and associations of humic substances in the terrestrial environment. Naturwissenschaften 89:84-88; 2002
Sovadinova, I., Blaha, L., Janosek, J., Hilscherova, K., Giesy, J. P., Jones, P. D., and Holoubek, I. Cytotoxicity and aryl hydrocarbon receptor-mediated activity of N-heterocyclic polycyclic aromatic hydrocarbons: Structure-activity relationships. Environ. Toxicol. Chem. 25:1291-1297; 2006
Steinberg, C. E. W. Ecology of Humic Substances in Freshwaters - Determinants from Geochemistry to Ecological Niches. Springer, BERLIN. 2003

Steinberg, C. E. W., Hoss, S., and Bruggemann, R. Further evidence that humic substances have the potential to modulate the reproduction of the nematode Caenorhabditis elegans. Int. Rev. Hydrobiol. 87:121-133; 2002
Steinberg, C. E. W., Paul, A., Pflugmacher, S., Meinelt, T., Klocking, R., and Wiegand, C. Pure humic substances have the potential to act as xenobiotic chemicals - A review. Fresen. Environ. Bull. 12:391-401; 2003
Timofeyev, M. A., Wiegand, C., Burnison, B. K., Shatilina, Z. M., Pflugmacher, S., and Steinberg, C. E. W. Impact of natural organic matter (NOM) on freshwater amphipods. Sci. Total Environ. 319:115-121; 2004
Wetzel, R. G., Hatcher, P. G., and Bianchi, T. S. Natural photolysis by ultraviolet irradiance of recalcitrant dissolved organic matter to simple substrates for rapid bacterial metabolism. Limnol. Oceanogr. 40:1369-1380; 1995
Wiegand, C., Meems, N., Timoveyev, M. A., Steinberg, C. E. W. and Pflugmacher, S. More evidence for humic substances acting as biogeochemicals on organisms. In E. A. Ghabbour and G. Davies (eds). Humic Substances Nature's Most Versatile Materials, Taylor and Francis, Inc. New York, pp. 349-361; 2003
Yamamoto, H., Liljestrand, H. M., and Shimizu, Y. Effects of dissolved organic matter surrogates on the partitioning of 17 beta-estradiol and p-nonylphenol between synthetic membrane vesicles and water. Environ. Sci. Technol. 38:2351-2358; 2004

IV. Curriculum Vitae - Michal Bittner


IV. Curriculum Vitae - RNDr. Michal Bittner
RECETOX, Kamenice 126/3, CZ-625 00 Brno, Czech Republic,
phone: +420 549 491 462, fax: +420 549 492 840,
e-mail: [email protected]
Personal data
Nationality: Czech
Date and birth-place: July 28, 1979, Náchod, Czech Republic
Education
Since 2003 Ph.D. candidate in Environmental Chemistry
RECETOX - Research Centre for Environmental Chemistry and Ecotoxicology, Faculty of Science, Masaryk University, Brno, Czech Republic
(www.recetox.muni.cz)
2006 RNDr. degree in Ecotoxicology
RECETOX - Research Centre for Environmental Chemistry and Ecotoxicology, Faculty of Science, Masaryk University, Brno, Czech Republic
1998 - 2003 M.Sc. degree in Ecotoxicology
RECETOX - Research Centre for Environmental Chemistry and Ecotoxicology, Faculty of Science, Masaryk University, Brno, Czech Republic

Professional
Since 2005 research scientist, part-time
RECETOX, Masaryk University, Czech Republic
Research interests
humic substances, in vitro toxicology, persistent organic pollutants
Languages
Czech (native), English (good), German (passive)
Research projects
2006 “Unexpected effects of humic substances and their photo-transformation products” Project of Ministry of Education, Youth and Sports of the Czech Republic FRVS TO G4 2510/2006 - principal investigator.
Project EC (2005-2008). "ECODIS" (6th FWP STREP contract No. 518043-1) Dynamic Sensing of Chemical Pollution Disasters and Predictive Modelling of their Spread and Ecological Impact - participation.
Awards:
Best poster presentation
International Symposium on Toxicity Assessment (ISTA12): Bittner, M., Janošek, J., Bláha, L., Hilscherová, K., Holoubek, I., Giesy, J.P. Application of in vitro bioassay for study of dioxin-like activity of humic acids. ISTA12 International Symposium on Toxicity Assessment, Greece, June 12-17, 2005, Book of Abstracts: 65, ISBN 960-88067-6-3 (poster presentation)

Publications
Theses
Bittner, M. 2006. Direct effects of humic substances on organisms.
RNDr. Thesis. Masaryk University: 29 pp (in Czech).
Bittner, M. 2003. HPLC determination of atrazine, its degradation products, and chlorophenols in the form of fluorescent derivatives with PIBCl.
M.Sc. Thesis. Masaryk University: 77 pp (in Czech).
Journal articles - published
Bittner, M., Janosek, J., Hilscherova, K., Giesy, J., Holoubek, I., and Blaha, L. Activation of Ah receptor by pure humic acids. Environmental Toxicology, 21:338-342; 2006
Janosek, J., Bittner, M., Hilscherová, K., Bláha, L., Giesy, J.P., and Holoubek, I. AhR-mediated and antiestrogenic activity of humic substances. Chemosphere; 2006 (in press)
Journal articles - submitted
Bittner, M., Hilscherová, K. Giesy J.P. Changes of AhR-mediated activity of humic substances after irradiation; 2006 (submitted to Environment International)
International conferences
Bittner, M., Hilscherová, K., Janošek, J., Bláha, L., Giesy, J.P., Holoubek, I.. Decrease of humic acids AhR-mediated activity after irradiation. International Conference on Ecotoxicology: Trends and Perspectives, Wisla, Poland, September 17-20, 2006, Book of Abstracts: 22 (platform presentation).

Bittner, M., Janošek, J., Hilscherová, K., Giesy, J.P. Dioxinová aktivita huminových kyselin a jejich fotodegradačních produktů. Študentská vedecká konferencia, Bratislava, Slovak Republic, April 26, 2006, Book of Reviewed Abstracts: 206-208, ISBN: 80-88870-58-5 (platform presentation).
Bittner, M., Janošek, J., Hilscherová, K., Bláha, L., Holoubek, I., Giesy, J.P. AhR-mediated activity of humic acids - changes after irradiation, SETAC Europe - 16th annual meeting, SETAC Europe, The Hague, The Netherlands, May 7-11, 2006, Book of Abstracts: 112 (poster presentation)
Janošek, J., Bittner, M., Hilscherová, K., Bláha, L., Holoubek, I., Giesy, J.P. Humic acids as dioxin-like acting chemicals. In Joint Conference of Scandinavian Society of Cell Toxicology and Estonian Society of Toxicology. Toila, Estonsko: ETS-SSCT, October 20-23, 2005. Book of Abstracts: 102, ISBN 9949-13-297-5 (poster presentation)
Bittner, M., Janošek, J., Bláha, L., Hilscherová, K., Holoubek, I., Giesy, J.P. Effect of Humic Substances on Intracellular Receptors. SETAC Europe, 15th annual meeting, SETAC Europe, France, May 22-26, 2005, Book of Abstracts: 277 (poster presentation)
Bittner, M., Janošek, J., Bláha, L., Hilscherová, K., Holoubek, I., Giesy, J.P. Application of in vitro bioassay for study of dioxin-like activity of humic acids. ISTA12 International Symposium on Toxicity Assessment, Greece, June 12-17, 2005, Book of Abstracts: 65, ISBN 960-88067-6-3 (poster presentation) - ISTA12 Award winner - Best poster presentation
Bittner, M., Janošek, J., Bláha, L., Hilscherová, K., Holoubek, I., Giesy, J.P. Activation of Ah-receptor by humic substances. ECOTOX 2005, Advances and Trends in Ecotoxicology, Masaryk University, Brno, Czech Republic, September 5-7, 2005, Book of Abstracts: 115, ISBN 80-210-3799-7 (poster presentation)
Bittner, M., Janošek, J., Bláha, L., Hilscherová, K., Holoubek, I., Giesy, J.P Humic Acids as Potential Activators of the Aryl Hydrocarbon Receptor. First Biennial Central & Eastern European Environmental Health Conference, International Health Sciences Solving Common Problems, Prague, October 24-27, 2004, Book of Abstracts: 22-23 (poster presentation)

Bittner, M., Janošek, J., Bláha, L. (Eko)toxikologické vlastnosti huminových látek a jejich další neočekávané efekty. Česko-Slovenská Studentská Vědecká Konference, Brno, May 1th, 2004, Book of Abstracts: 19, ISBN80-210-3376-2 (platform presentation)





























