Embed Size (px)
Citation preview

Photochemistry and Photobiology, 2006, 82: 981-993
Symposium-in-Print: UV Effects on Aquatic and Coastal Ecosystems
Ecotoxicological Effects of Combined UVB and Organic Contaminants in Coastal Waters: A Review
E. Pelletier*, P. Sargian, J. Payet and S. Demers lnstitut des Sciences de la Mer de Rimouski, Universite du Quebec a Rimouski, Rimouski, Canada
Received 18 September 2005; accepted 4 April 2006; published online 7 April 2006 DOI: 2005-09-18-RA-688
ABSTRACT
Organisms living in coastal waters are exposed to anthropo- genic contaminants from terrestrial drainage, ice melting and maritime traffic and to enhanced UVB radiation (UVBR; 280-320 nm) caused by decreased concentrations of ozone in the stratosphere. This article reviews available information about the combined effects of UVBR and selected hydrosoluble contaminants potentially present in surface waters on marine species and especially on plankton community structure in high-latitude coastal zones. Effects of UVBR on three selected pesticides (Atrazine, carbaryl and Acifluorfen) and possible induction of phototoxicity are reviewed. Most toxicological studies have been conducted under laboratory conditions with questionable relevance for coastal marine ecosystems. Simi- larly, photoactivation of polycyclic aromatic hydrocarbons (PAHs) has been closely examined and reported effects on aquatic species summarized. Experiments with field-sampled communities demonstrated the complexity and the difficulty in determining the impact of multiple stressors on an aquatic ecosystem, even for ecosystems simplified by eliminating large grazers and fish. Nutrient status, specific composition and light history have influenced the different responses of planktonic assemblages exposed to enhanced UVBR and water-soluble fraction (WSF) from crude oil or to tributyltin. Plankton assemblages subjected to changes in the ozone hole were physiologically stressed and more susceptible to WSF tox- icity than communities from less enhanced UVBR-impacted sites. A close relationship between phytoplankton assemblages and bacteria was observed in all experiments in mesocosms. A contaminant-induced phytoplankton crash after a bloom event may release important carbon and nutrient sources for bac- teria. The magnitude of phytoplanktonic mortality induced by a contaminant probably influenced how rapidly bacteria grew over time. The transition from a herbivorous food web to a microbial food web has significant ecological implications for carbon cycling and energy flow in pelagic systems. A high phytoplankton mortality implies a situation in which the potential for downward carbon export from surface waters is high. In contrast, high bacterial enrichment implies that the phytoplankton carbon is largely recycled in surface waters through a microbial loop and does not contribute significantly
*Corresponding author email: [email protected] (E. Pelletier) 0 2006 American Society for Photobiology 003 1 -8655/06
to sinking particle flux. The most ecologically relevant results were obtained with mesocosm studies using field-collected communities. The enhancement of hydrocarbon toxicity in the presence of a high level of UVBR cannot be described as being a synergistic or an additive effect, because the WSF alone is not toxic and may even be beneficial by increasing bacterial activity. This is a case in which one stressor has the ability to modify another stressor to cause it to be toxic to target organ- isms. These abiotically induced interactions may be important for biological communities exposed to extreme conditions when physical, chemical or photochemical reactions modify the nature of environmental stressors before they interact with biological functions. The need for models on the impacts of multiple stressors on biodiversity and ecosystem functioning is emphasized.
INTRODUCTION A decrease in the concentration of ozone in the stratosphere may significantly increase the level of UVB radiation (UVBR; 28G320 nm) reaching the Earth’s surface. These modifications have raised concern among scientists and policy makers during the past 2 decades. Numerous laboratory and field experiments have been conducted illustrating deleterious effects of UVBR on aquatic species and communities (1,2). In addition to UVBR enhancement (mainly at high latitudes), anthropogenic impacts also include the direct release of contaminants (such as heavy metals and organo- metals, chlorinated pesticides, brominated flame retardants, estrogens and complex mixtures of petrogenic and pyrogenic hydrocarbons) into rivers, estuaries and coastal waters. Many of these chemicals are known to have deleterious effects on aquatic organisms because they provide an additional stress on aquatic communities. It is crucial therefore to identify interactions of multiple stressors on biodiversity and ecosystem functioning.
Some recent experimental studies have focused on the toxic effects of simultaneous exposure to solar UV radiation (UVR; natural and/or artificially enhanced) and water-soluble contami- nants and indicate that both stressors can act either additively or synergistically on aquatic microbes (3,4), algae (5-7), plants (8,9), amphipods (1&12), crustaceans (13,14) molluscs (15), echino- derms (4), fish (16-18) and amphibians (19). Laboratory tests involving single species or unrealistic concentrations of contam- inants with artificial UV lamps have been subject to severe criticism (20,21) for their lack of ecological relevance and their lack of usefulness in determining real risks for species and whole
981

982 E. Pelletier et a/.
ecosystems. Very few studies have examined the combined effects of solar radiation and contaminants on field-collected communities (22). Only a recent study examined the effects of water-soluble fractions (WSFs) of' heavy oil on marine enclosed ecosystems in large mesocosms (23) in which experimental tanks were covered with acrylic plates to filter sunlight.
The aim of this article is to provide an overview of reported observations and findings about interactions between UVBR and soluble organic pollutants potentially present in marine coastal waters and their combined effects on marine biota. Special attention has been paid to our recent findings on the combined effects of UVBR and water-soluble hydrocarbons from petroleum (24) and the antifouling agent tributyltin (TBT) (25) on the plankton marine communities sampled in a northern ecosystem (St. Lawrence Estuary, Canada) and sub-Antarctic environment (Southern Argentina).
UVBR AND SELECTED PESTICIDES Pesticides are toxic substances widely used to control or eradicate viruses, fungi and bacteria, algae and plants, insects, molluscs, fish, rodents and many other living organisms considered as detrimental to human activities. More than 6500 chemical pesticides are currently listed in the PAN (Pesticides Action Network) database (available from: http://www.pesticideinfo.org/Index.html). Unlike aromatic hydrocarbons discussed hereafter, aqueous photochemis- try of pesticides has received little attention from research lab- oratories and only a limited number of studies on photomodified toxicity of pesticides by UVBR has been published.
Carbaryl ( 1 -naphthyl-N-methylcarbamate) is an insecticide best known as Sevin@ and is commonly used on fruit and vegetable crops, lawns and ornamental trees. Carbaryl is not considered to be persistent in the environment. Photodegradation of carbaryl in natural coastal waters is unknown but Samanidou et al. (26) determined the photolysis rate of some carbamate pesticides in seawater under laboratory conditions and found a rapid degradation within a few hours or days.
H3% P H I
'-$
Scheme 1: Chemical structure of carbaryl
A recent study by Miller and Chin (27) conducted in wetland surface waters indicated that, when the nitrate concentration was low (which is often the case in coastal waters during summer), natural organic matter played the role of photosensitizer and enhanced the production of unidentified transient species. The main photoproduct is 1-naphthol, a known cytotoxic metabolite that is also a polycyclic aromatic hydrocarbon (PAH) photoproduct requiring high concentrations for toxicity (28). A similar process may apply in brackish and estuarine waters where fulvic and humic acids are known to play an important role in the photochemistry of many natural and anthropogenic chemicals.
In early work by Zaga et al. (19) the photoenhanced toxicity of the insecticide carbaryl was examined in laboratory exposure glass chambers using embryos and tadpoles of two frogs (Xenopus laevis
and Hyla versicolor) under controlled UVBR. All of the UVBR- carbaryl experiments revealed a significant interactive effect between carbaryl, UVBR and day of exposure. Differences ob- served between carbaryl treatments with and carbaryl treatments without UVBR were attributed to the formation of photolytic by- products as the main breakdown product of carbaryl was 1- naphthol. However, Bridges and Boone (29) re-examined the joint toxicity of carbaryl and UVBR in leopard frog (Rana sphenoce- phala) tadpoles in outdoor artificial ponds and found that tadpole survival to metamorphosis was positively influenced by the level of UVBR intensity. Tadpoles in ponds exposed to carbaryl contained more than three times as much algae and yielded larger metamorphs than control ponds. Bridges and Boone (29) suggested that dis- solved organic carbon in the ponds may have played a protective role by increasing photodegradation of carbaryl and naphthol and they emphasized the importance of conducting field studies to more accurately predict what occurs under natural conditions.
Atrazine is a strong herbicide that inhibits photosystem U (PS 11) and is widely used to control broadleaf and some grassy weeds for a vari- ety of major and minor crops ( e g . corn, sugarcane and winter wheat) and nonagricultural uses (e.g. golf courses and residential lawns).
Atrazine and photoproducts have been detected in surface waters
Scheme 2: Chemical structure of atrazine
and coastal waters (30,31) but their toxicity toward marine com- munities is still unexplored. Lin et al. (32) reported the toxicity of atrazine and metolachlor after exposure to simulated sunlight in the marine luminescent bacterium Vibrio fishcheri (known as the Microtox test) and observed a decrease in toxicity with increasing intensity of light and an even lower toxicity in estuarine waters with high concentrations of organic and suspended matter. Their study showed a rapid photodegradation of atrazine and a low toxicity for photoproducts. However, these results were challenged by Kon- stantinou et al. (33), who studied photolysis of pesticides in the laboratory and observed an increased optical filter effect as the concentration of dissolved organic matter (DOM) increased, leading to a lower photodegradation rate in natural waters. The half-life of atrazine in marine waters (using glass beakers and incident solar radiation) reached 56 days, compared with 34 days in distilled water.
A more realistic experiment was conducted in flow-through freshwater mesocosms using atrazine concentrations (5-75 pg/L,) typical of surface waters in the midwestem region of the United States (34). End points measured included nutrients, periphyton biomass and productivity, growth of macrophytes and survival and growth of Daphnia rnagna, Rana pipiens and Pirnephales promelas larvae and adults. The atrazine half-life in the experimental wetlands was 8-14 days. Only periphyton species were signifi- cantly affected by atrazine at any of the concentrations tested but they developed resistance at concentrations greater than 50 pgL. Losses of atrazine from the water column were greater than those predicted by kinetic models and half-lives were ca 3 times shorter

Photochemistry and Photobiology, 2006, 82 983
than calculated values. Both atrazine and sunlight exposures significantly decreased D. magna survival but the authors (34) found no significant interaction between these two stressors. The lack of potentiated atrazine toxicity by sunlight was attributed to a mitigation effect of naturally occurring humic substances (particularly from plant litter), which likely reduced the bio- availability of atrazine as previously observed for anthracene (35).
The role of humic substances on the photodegradation of dis- solved chemicals has often been often mentioned in recent years but little is yet known about the molecular mechanisms that could have a beneficial or detrimental effect on the toxicity of metabolites or their parent compounds. It is generally recognized that UVBR induces photochemical reactions in DOM and generates reactive oxygen species that, in turn, may oxidize organic pollutants (36). The photodegradation rate of atrazine in fresh water with different types and concentrations of humic substances under outdoor sunlight conditions was recently reported (37). Increasing concentration of river humic acid induced an increasing photodegradation rate of atrazine although fulvic acid (low molecular weight) had no signi- ficant effect. A comparable field experiment with special attention to DOM has not yet been reported for a marine ecosystem although the presence of atrazine in coastal waters is expected to be too low to produce appreciable phototoxicity except very close to the shore.
The activity of diphenyl-ether herbicides, a family of pesticides introduced in 1990s, may represent a real case of photoinduced toxicity (38). Diphenyl-ethers are broad-spectrum herbicides registered for use on soybeans, peanuts and rice for postemergent weed control. They are light-dependent peroxydizing herbicides (LDPHs) that act in plants by inhibiting the enzyme protopor- phyrinogen oxidase, which is a key enzyme in the chlorophyll biosynthetic pathway. Photochemistry and photoinduced toxicity of Acifluorfen@ were studied in the laboratory by exposing herbicide solutions to a solar simulator (1.1 kW xenon lamp with a passing wavelength of 300-800 nm) for 140 h (38).
Scheme 3: Chemical structure of AcifluorfenB
Toxicity tests using D. magna indicated a significant increase of toxicity toward Acifluorfen@ for solutions in which the parent molecule was entirely transformed in photodegraded by-products. Leo (39) used structural data to calculate that Kow (octanol-water partition coefficients) of some well-identified photoproducts were much higher than that for Acifluorfen@. High Kow provide reasonable indices that photoproducts were much more bioavail- able than the parent compound and were able to interact with biological functions of D. magna. The relevance of these results needs to be assessed by carefully designed field experiments in which a whole freshwater or marine ecosystem is considered.
UVBRANDHYDROCARBONS Hydrocarbons form a very rich and complex family of organic molecules comprising linear and cyclic saturated hydrocarbons, single and polycyclic rings with or without side chains and
heterocycles containing sulfur (thiophenes) and nitrogen (porphyr- ines) atoms. The lighter PAHs exhibit relatively high solubility in water (Table 1) and are readily available (high log Kow) to phy- toplankton and swimming aquatic organisms ranging from micro- zooplankton to fish. Most abundant sources of aromatic hydrocarbons at sea are petroleum residues and crude oil accidentally spilled or intentionally discharged by tankers and other vessels navigating across oceans, along continental coastlines and in narrow estuaries, fjords and river channels.
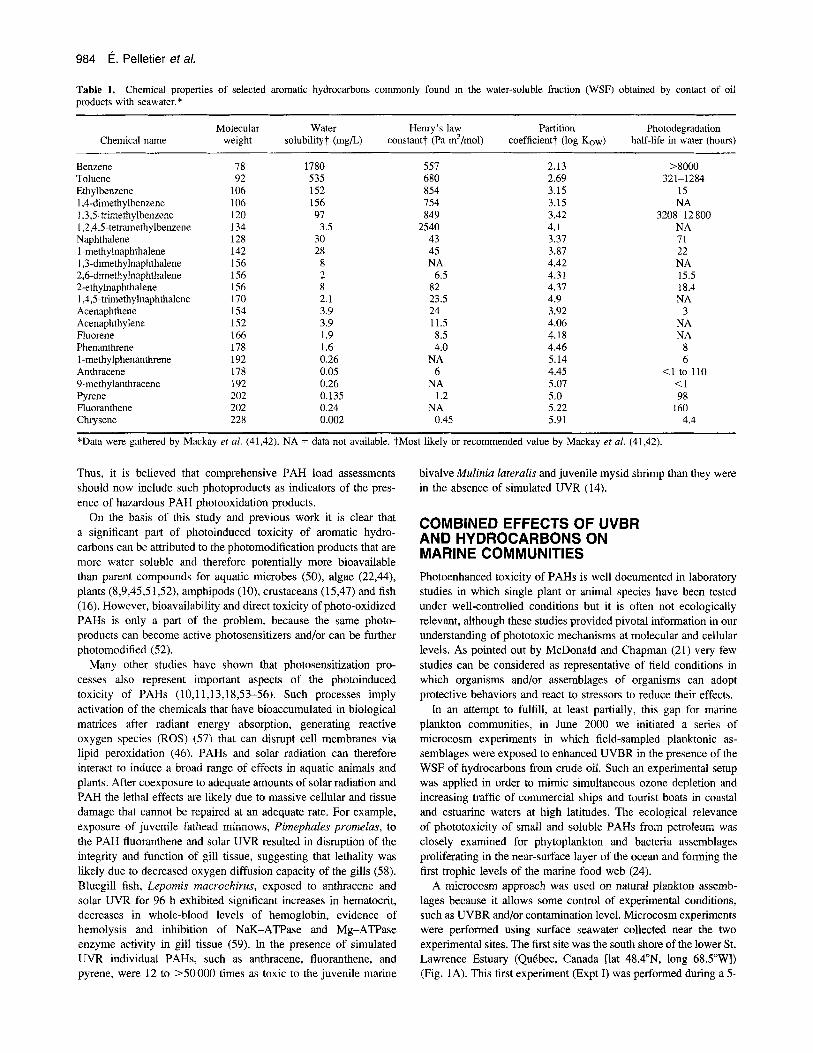
PAHs are also emitted in the air as gases and soot by combustion of fossil fuel, plastics, wood and other natural materials. Most hydrosoluble PAHs (Table 1) can be released in coastal waters by desorption from falling soot particles, suspended particulate matter and sediment carried by storm water runoff and sewage discharges (40). Data on photodegradation of PAHs in water (including photolysis and photooxidation by reaction with hydroxyl radicals) are sparse and often broadly distributed after source and de- termination techniques (41,42). Table 1 shows that smaller molecules are generally more resistant to photodegradation than larger ones are.
UVR activation of PAHs has been intensively studied because of their ability to absorb both UVBR and UVAR, by virtue of their extensive n-orbital system (43). This UVR absorption can lead to the photodegradation of the PAH molecules (photooxidation and photodissociation), thereby resulting in new photoproducts. How- ever, many studies have shown that an important aspect of the photoinduced PAH toxicity is derived from such photodegradation processes (8,22,44-47). In these studies chemical compounds were irradiated before toxicity testing, allowing demonstration of whether photomodified PAHs were contributing to toxicity.
Huang et al. (8) investigated the photoinduced toxicity of anthracene, phenanthrene and benzo(a)pyrene to the aquatic higher plant Lemna gibba (a duckweed) with chemicals irradiated before toxicity testing in which inhibition of growth and the extent of chlorosis were measured. They found that light activated the phy- totoxicity of PAHs (UVR being more effective than visible light), with compounds becoming more toxic as they were converted to new structures by radiation. Comparable results were obtained by Ren et al. (45) with fluoranthene, pyrene and naphthalene, demonstrating that the photomodification process could be rapid under environmentally relevant levels of actinic radiation. So in many instances it was found that photomodification of any given PAH leads to a wide variety of oxidation products that are often difficult to analyze and identify (9,48-50).
The investigation of the toxicity of photoproducts was extended by Mallakin et al. (51), who determined which photoproducts in the mixture of photoproducts made the most significant contribu- tions to toxicity, using anthracene as the test compound. Appli- cation of different radiation conditions demonstrates that the toxicity of the photoproducts of anthracene can also be photo- induced. This is in contrast to phenanthrenequinone (the major photoproduct of phenanthrene), the toxicity of which is greater than that of phenanthrene but not greatly photoinduced (50). Cer- tain anthracene photoproducts are themselves subject to photoox- idation, with a concomitant increase in toxicity. Conversely, some of the photoproducts of anthracene, although not readily photo- oxidized, are nonetheless phototoxic, indicating photosensitization activity. Interestingly, one of these photoproducts (2-hydroxyan- thraquinone), which was not subject to photooxidation, appeared as the most toxic of the compounds tested. As a light-stable com- pound it presents the risk of a persistent environmental hazard.
984 E. Pelletier et a/.
Table 1. products with seawater.*
Chemical properties of selected aromatic hydrocarbons commonly found in the water-soluble fraction (WSF) obtained by contact of oil
Molecular Water Henry's law Partition Photodegradation Chemical name weight solubilityt (mg/L) constant? (Pa m3/mol) coefficient? (log Kow) half-life in water (hours)
Benzene 78 1780 557 2.13 >goo0 Toluene 92 535 680 2.69 Ethylbenzene 106 152 854 3.15 15 1.4-dimethylbenzene 106 156 754 3.15 NA 1,3,5-trimethylbenzene 120 97 849 3.42 3208-12 800 1,2,4,5-tetramethylbenzene 134 3.5 2540 4.1 NA Naphthalene 128 30 43 3.37 71
1,3-dimethylnaphthalene 156 8 NA 4.42 NA 2,6-dimethylnaphthalene 156 2 6.5 4.31 15.5 2-ethylnaphthalene 156 8 82 4.37 18.4 1,4,5-trimethylnaphthaIene 170 2.1 23.5 4.9 NA Acenaphthene 154 3.9 24 3.92 3 Acenaphthylene 152 3.9 11.5 4.06 NA Fluorene 166 1.9 8.5 4.18 NA Phenanthrene 178 1.6 4.0 4.46 8 1 -methylphenanthrene 192 0.26 NA 5.14 6 Anthracene 178 0.05 6 4.45 <1 to 110
Pyrene 202 0.135 1.2 5.0 98 Fluoranthene 202 0.24 NA 5.22 160
321-1284
1 -methylnaphthalene 142 28 45 3.87 22
9-inethylanthracene 192 0.26 NA 5.07 <1
Chrysene 228 0.002 0.45 5.91 4.4
*Data were gathered by Mackay et al. (41,42). NA = data not available. tMost likely or recommended value by Mackay et al. (41,42).
Thus, it is believed that comprehensive PAH load assessments should now include such photoproducts as indicators of the pres- ence of hazardous PAH photooxidation products.
On the basis of this study and previous work it is clear that a significant part of photoinduced toxicity of aromatic hydro- carbons can be attributed to the photomodification products that are more water soluble and therefore potentially more bioavailable than parent compounds for aquatic microbes (50), algae (22,44), plants (8,9,45,51,52), amphipods (lo), crustaceans (15,47) and fish (1 6). However, bioavailability and direct toxicity of photo-oxidized PAHs is only a part of the problem, because the same photo- products can become active photosensitizers and/or can be further photomodified (52).
Many other studies have shown that photosensitization pro- cesses also represent important aspects of the photoinduced toxicity of PAHs (10,11,13,18,53-56). Such processes imply activation of the chemicals that have bioaccumulated in biological matrices after radiant energy absorption, generating reactive oxygen species (ROS) (57) that can disrupt cell membranes via lipid peroxidation (46). PAHs and solar radiation can therefore interact to induce a broad range of effects in aquatic animals and plants. After coexposure to adequate amounts of solar radiation and PAH the lethal effects are likely due to massive cellular and tissue damage that cannot be repaired at an adequate rate. For example, exposure of juvenile fathead minnows, Pimeplzales promelas, to the PAH fluoranthene and solar UVR resulted in disruption of the integrity and function of gill tissue, suggesting that lethality was likely due to decreased oxygen diffusion capacity of the gills (58). Bluegill fish, Lepomis macrochirus, exposed to anthracene and solar UVR for 96 h exhibited significant increases in hematocrit, decreases in whole-blood levels of hemoglobin, evidence of hemolysis and inhibition of NaK-ATPase and Mg-ATPase enzyme activity in gill tissue (59). In the presence of simulated UVR individual PAHs, such as anthracene, fluoranthene, and pyrene, were 12 to >50000 times as toxic to the juvenile marine
bivalve Mulinia lateralis and juvenile mysid shrimp than they were in the absence of simulated UVR (14).
COMBINED EFFECTS OF UVBR ANDHYDROCARBONSON MARINE COMMUNITIES Photoenhanced toxicity of PAHs is well documented in laboratory studies in which single plant or animal species have been tested under well-controlled conditions but it is often not ecologically relevant, although these studies provided pivotal information in our understanding of phototoxic mechanisms at molecular and cellular levels. As pointed out by McDonald and Chapman (21) very few studies can be considered as representative of field conditions in which organisms and/or assemblages of organisms can adopt protective behaviors and react to stressors to reduce their effects.
In an attempt to fulfill, at least partially, this gap for marine plankton communities, in June 2000 we initiated a series of microcosm experiments in which field-sampled planktonic as- semblages were exposed to enhanced UVBR in the presence of the WSF of hydrocarbons from crude oil. Such an experimental setup was applied in order to mimic simultaneous ozone depletion and increasing traffic of commercial ships and tourist boats in coastal and estuarine waters at high latitudes. The ecological relevance of phototoxicity of small and soluble PAHs from petroleum was closely examined for phytoplankton and bacteria assemblages proliferating in the near-surface layer of the ocean and forming the first trophic levels of the marine food web (24).
A microcosm approach was used on natural plankton assemb- lages because it allows some control of experimental conditions, such as UVBR and/or contamination level. Microcosm experiments were performed using surface seawater collected near the two experimental sites. The first site was the south shore of the lower St. Lawrence Estuary (QuBbec, Canada [lat 48.4"N, long 68.5"WI) (Fig. 1A). This first experiment (Expt I) was performed during a 5-

Photochemistry and Photobiology, 2006, 82 985
Figure 1. Schematic view of the location of the experimental sites of (A) Expt I (Rimouski, Canada) and (B) Expt I1 (Ushuaia, Argentina). The experimental set-up for both studies is also shown (C).
day period in June 2000. The second 5-day experiment (Expt 11) was performed in Beagle Channel Bay near the city of Ushuaia (Tierra del Fuego, Argentina [lat 54.8"S, long 68.2"WI) (Fig. 1B) and was performed in early November 2001. A third experiment (Expt 111) was conducted in mid-August 2001 near Rimouski and used experimental parameters similar to those used in Expt I.
For Expt I and Expt 11, microcosms (91-cylindrical Teflon@ bags [75 cm tall X 25 cm wide]) (Fig. 1C) were placed in larger mesocosms before filling with natural water collected in each specific site and were screened through a 500 pm NitexTM mesh in order to exclude large grazers. Larger mesocosms consisted of clear polyethylene bags (ca 2.3 m deep, with a volume of ca 1800 L) attached to a wharf structure and immersed in the water column. In each experiment microcosms were exposed in triplicate to the following UVR regimes: natural UVBR (NUVBR) and enhanced UVBR (HUVBR). UVBR enhancement was provided by four fluorescent UV lamps (TL40W-l2RS, Philips) with an emission peak at 313 nm. Wavelengths shorter than 290 nm (UVC radiation) were screened out by means of 0.13 mm cellulose acetate films that were changed daily. Compared with the NUVBR, the weighted irradiance in the UV region was 2.15-fold greater for the HUVBR treatment each day for a period of 6 h.
In order to determine possible interactive effects of UVBR and hydrocarbons a stock solution of WSF of crude oil was prepared with crude oil Forties from North Sea oil fields (for the specimens from Rimouski) and an Argentinean crude (for the specimens from Ushuaia). The resulting WSF was then added to the microcosms to yield realistic concentrations according to the experiment and crude oil used (ca 10 mg/L in Expt I and ca 5 mg/L in Expt 11). Microcosms were identified according to their UVBR treatment and WSF addition as follows: NUVBR and HUVBR, respectively,
for microcosms without dissolved hydrocarbons, and NWBR+WSF and HUVBR+WSF, respectively, for microcosms with dissolved hydrocarbons added.
During these 5-day experiments, incident UVBR, WSF and nutrient concentrations, as well as abundance and production of heterotrophic bacteria and phytoplankton were monitored. There- fore, these microcosm experiments, which were performed in different seasons and at different latitudes, revealed the relative short-term impact of UVBR and of WSF on a natural planktonic assemblage. However, the extent of both UVBR effects and WSF effects was different between the two specific sites, which explains the different results observed under simultaneous exposure to dual stress for Expt I and Expt 11.
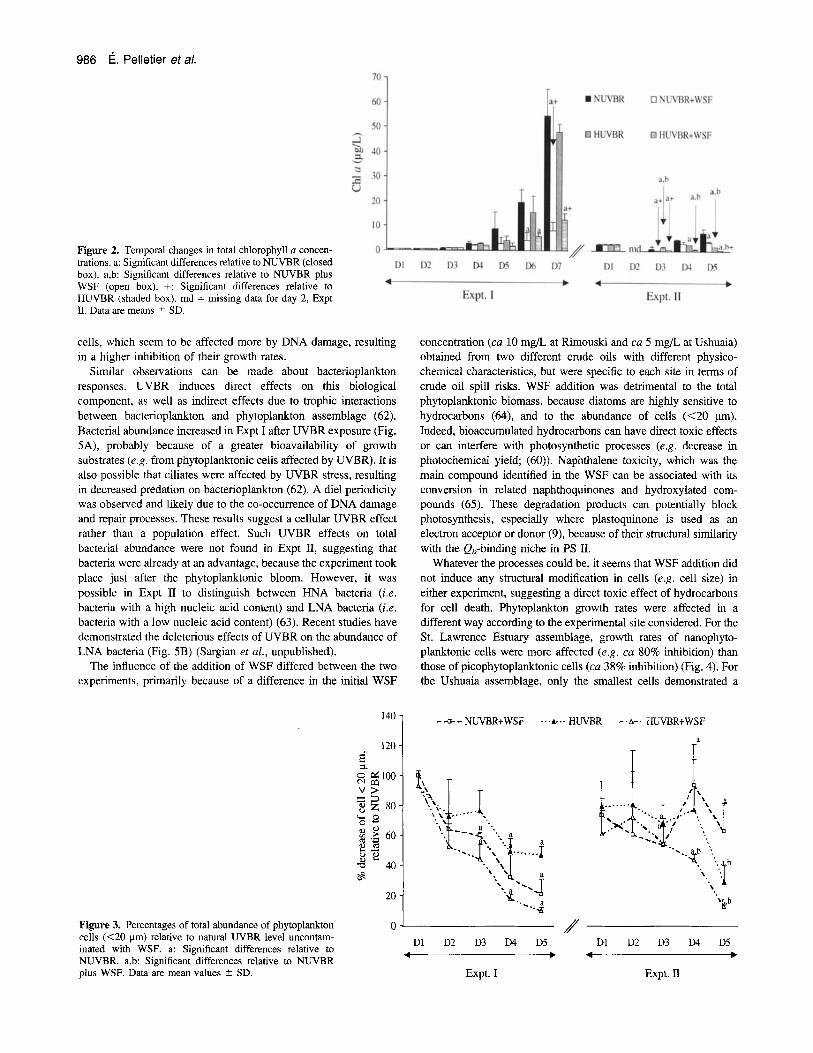
An increase in UVBR alone induced no significant effects on total phytoplanktonic biomass of the St. Lawrence Estuary, as determined by Chl a concentration (Fig. 2). However, a decrease in phytoplankton abundance (as determined by flow cytometry) was observed (Fig. 3), with a greater decrease in growth rates for nanophytoplanktonic cells than for picophytoplanktonic cells (Fig. 4). A decrease in photochemical efficiency given by the FvlFrn value, as determined with a PAM fluorometer on the final day of the experiment, was also observed (60). For Expt I1 phytoplankton was much less abundant and both total biomass and phytoplankton cells abundance were negatively affected (Figs. 2 and 3) with a greater decrease in growth rates for picophytoplankton cells than for nanophytoplankton cells. According to FvlFrn values an inhibition of ca 50% was observed after exposure to enhanced UVBR. However, by the end of the experiment the photochemical yield reached values equal to those observed under natural light conditions.
Many factors can govern the impact of UVBR and may explain differences observed in these two experiments. Nutritional status must be considered first. Initial concentrations were relatively high in Expt I (nitrate-nitrite concentration, 14.15 pmol/L; silicate concentration, 19 pmol/L), allowing the phytoplanktonic bloom (mainly diatoms) to occur. Previous studies have demonstrated the greater resistance of diatoms to deal with UVBR (61). By the end of Expt I the phytoplanktonic bloom had resulted in nutrient depletion and, therefore, an increasing sensitivity to UVBR effects. In the Ushuaia experiment the planktonic assemblage was characterized by lower nutrient concentrations (nitrate-nitrite concentration, 5 pmol/L; silicate concentration, 4.3 pmol/L) because the experiment took place just after the spring bloom. Large cells were less abundant and/or in a senescent state, resulting in a greater UVBR effect on the total biomass. Such limited nutrient status can indeed impede repair mechanisms to cope with UVBR effects, because repair enzyme synthesis and photo- protective pigments require energy and nutrients. Therefore, it is unlikely that phytoplankton assemblages of the St. Lawrence Estuary could have synthesized photoprotective compounds, as revealed by the decrease in photochemical yield by the end of the experiment associated with nutrient depletion.
In Expt I1 the die1 periodicity of the photochemical yield (with a significant decrease during the day and recovery at night) could be related to synthesis of photoprotective compounds by nano- phytoplankton, thus explaining the increase in the mean cell size of this phytoplanktonic fraction. However, photoprotective synthesis requires energy that was no longer available for physiological processes, resulting in the observed diminution of growth rates. Synthesis of such compounds is not expected in picophytoplankton
986 E. Pelletier et a/.
Figure 2. Temporal changes in total chlorophyll a concen- trations. a: Significant differences relative to NUVBR (closed box), a,b: Significant differences relative to NUVBR plus WSF (open box). f: Significant differences relative to HUVBR (shaded box). md = missing data for day 2, Expt 11. Data are means 2 SD.
cells, which seem to be affected more by DNA damage, resulting in a higher inhibition of their growth rates.
Similar observations can be made about bacterioplankton responses. UVBR induces direct effects on this biological component, as well as indirect effects due to trophic interactions between bacterioplankton and phytoplankton assemblage (62). Bacterial abundance increased in Expt I after UVBR exposure (Fig. 5A), probably because of a greater bioavailability of growth substrates (e.g. from phytoplanktonic cells affected by UVBR). It is also possible that ciliates were affected by UVBR stress, resulting in decreased predation on bacterioplankton (62). A die1 periodicity was observed and likely due to the co-occurrence of DNA damage and repair processes. These results suggest a cellular UVBR effect rather than a population effect. Such UVBR effects on total bacterial abundance were not found in Expt 11, suggesting that bacteria were already at an advantage, because the experiment took place just after the phytoplanktonic bloom. However, it was possible in Expt I1 to distinguish between HNA bacteria (i.e. bacteria with a high nucleic acid content) and LNA bacteria (i.e. bacteria with a low nucleic acid content) (63). Recent studies have demonstrated the deleterious effects of UVBR on the abundance of LNA bacteria (Fig. 5B) (Sargian et al., unpublished).
The influence of the addition of WSF differed between the two experiments, primarily because of a difference in the initial WSF
concentration (ca 10 mg/L at Rimouski and ca 5 mg/L at Ushuaia) obtained from two different crude oils with different physico- chemical characteristics, but were specific to each site in terms of crude oil spill risks. WSF addition was detrimental to the total phytoplanktonic biomass, because diatoms are highly sensitive to hydrocarbons (64), and to the abundance of cells (<20 pm). Indeed, bioaccumulated hydrocarbons can have direct toxic effects or can interfere with photosynthetic processes ( e g . decrease in photochemical yield; (60)). Naphthalene toxicity, which was the main compound identified in the WSF can be associated with its conversion in related naphthoquinones and hydroxylated com- pounds (65). These degradation products can potentially block photosynthesis, especially where plastoquinone is used as an electron acceptor or donor (9), because of their structural similarity with the Qb-binding niche in PS 11.
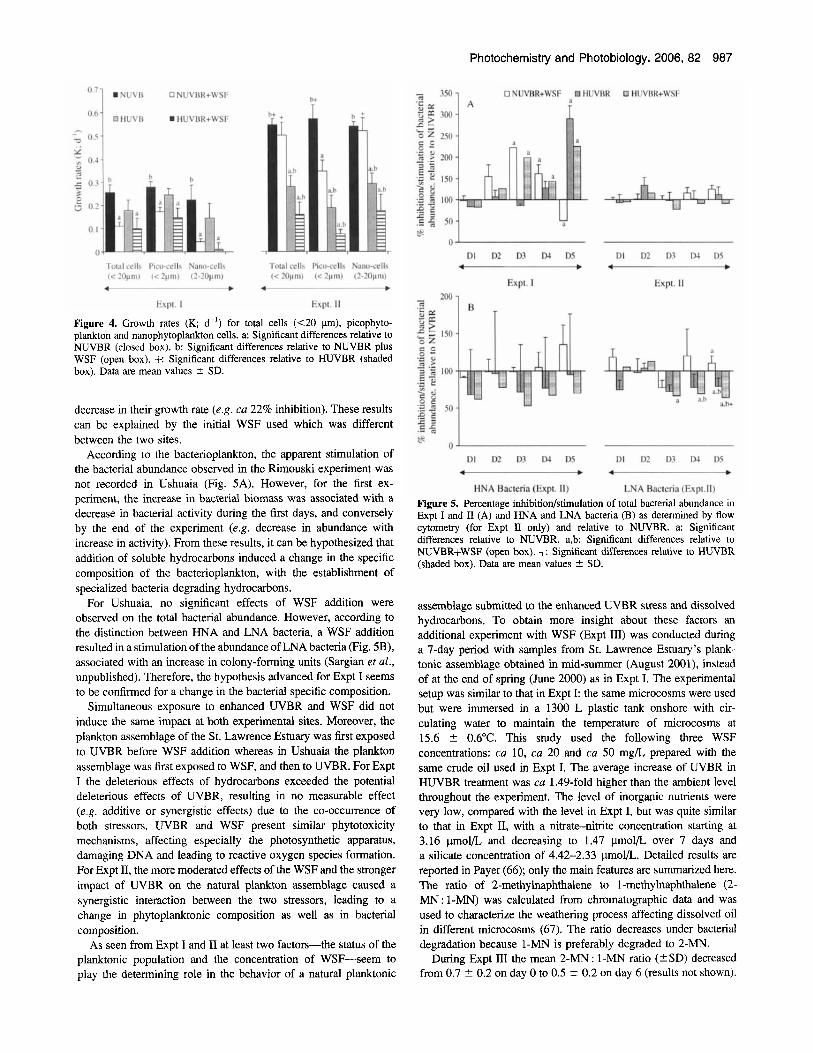
Whatever the processes could be, it seems that WSF addition did not induce any structural modification in cells (e.g. cell size) in either experiment, suggesting a direct toxic effect of hydrocarbons for cell death. Phytoplankton growth rates were affected in a different way according to the experimental site considered. For the St. Lawrence Estuary assemblage, growth rates of nanophyto- planktonic cells were more affected (e.g. ca 80% inhibition) than those of picophytoplanktonic cells (ca 38% inhibition) (Fig. 4). For the Ushuaia assemblage, only the smallest cells demonstrated a
--(I--NUVBR+WSF --.A*.. HUVBR --&, HLJVBR+WSF
T i
Figure 3. Percentages of total abundance of phytoplankton cells (<20 pm) relative to natural UVBR level uncontam- inated with WSF. a: Significant differences relative to NUVBR. a,b: Significant differences relative to NUVBR plus WSF. Data are mean values 5 SD.
D1 D2 D3 D4 D5 0 ‘ //
D1 D2 D3 D4 D5 4 b 4 b
Expt. I Expt. I1
Photochemistry and Photobiology, 2006, 82 987
Figure 4. Growth rates (K; d-') for total cells (<20 p), picophyto- plankton and nanophytoplankton cells. a: Significant differences relative to NUVBR (closed box). b: Significant differences relative to NUVBR plus WSF (open box). f: Significant differences relative to HUVBR (shaded box). Data are mean values 2 SD.
decrease in their growth rate (e.g. ca 22% inhibition). These results can be explained by the initial WSF used which was different between the two sites.
According to the bacterioplankton, the apparent stimulation of the bacterial abundance observed in the Rimouski experiment was not recorded in Ushuaia (Fig. 5A). However, for the first ex- periment, the increase in bacterial biomass was associated with a decrease in bacterial activity during the first days, and conversely by the end of the experiment (e.g. decrease in abundance with increase in activity). From these results, it can be hypothesized that addition of soluble hydrocarbons induced a change in the specific composition of the bacterioplankton, with the establishment of specialized bacteria degrading hydrocarbons.
For Ushuaia, no significant effects of WSF addition were observed on the total bacterial abundance. However, according to the distinction between HNA and LNA bacteria, a WSF addition resulted in a stimulation of the abundance of LNA bacteria (Fig. 5B), associated with an increase in colony-forming units (Sargian et al., unpublished). Therefore, the hypothesis advanced for Expt I seems to be confirmed for a change in the bacterial specific composition.
Simultaneous exposure to enhanced UVBR and WSF did not induce the same impact at both experimental sites. Moreover, the plankton assemblage of the St. Lawrence Estuary was first exposed to UVBR before WSF addition whereas in Ushuaia the plankton assemblage was first exposed to WSF, and then to UVBR. For Expt I the deleterious effects of hydrocarbons exceeded the potential deleterious effects of UVBR, resulting in no measurable effect (e.g. additive or synergistic effects) due to the co-occurrence of both stressors. UVBR and WSF present similar phytotoxicity mechanisms, affecting especially the photosynthetic apparatus, damaging DNA and leading to reactive oxygen species formation. For Expt 11, the more moderated effects of the WSF and the stronger impact of UVBR on the natural plankton assemblage caused a synergistic interaction between the two stressors, leading to a change in phytoplanktonic composition as well as in bacterial composition.
As seen from Expt I and I1 at least two factors-the status of the planktonic population and the concentration of WSF-seem to play the determining role in the behavior of a natural planktonic
Figure 5. Percentage inhibition/stimulation of total bacterial abundance in Expt I and I1 (A) and HNA and LNA bacteria (B) as determined by flow cytometry (for Expt I1 only) and relative to NUVBR. a: Significant differences relative to NUVBR. a,b: Significant differences relative to NUVBRSWSF (open box). +. Significant differences relative to HUVBR (shaded box). Data are mean values 2 SD.
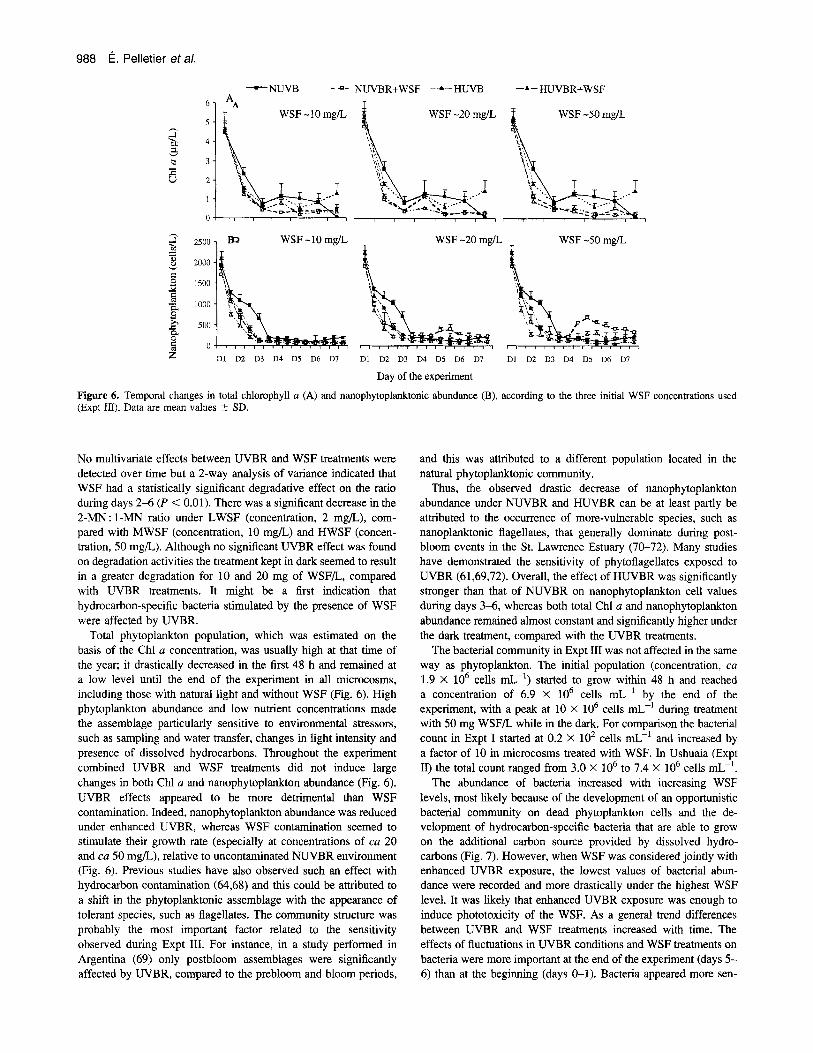
assemblage submitted to the enhanced UVBR stress and dissolved hydrocarbons. To obtain more insight about these factors an additional experiment with WSF (Expt 111) was conducted during a 7-day period with samples from St. Lawrence Estuary's plank- tonic assemblage obtained in mid-summer (August 2001), instead of at the end of spring (June 2000) as in Expt I. The experimental setup was similar to that in Expt I: the same microcosms were used but were immersed in a 1300 L plastic tank onshore with cir- culating water to maintain the temperature of microcosms at 15.6 ? 0.6"C. This study used the following three WSF concentrations: ca 10, ca 20 and ca 50 mg/L prepared with the same crude oil used in Expt I. The average increase of UVBR in HUVBR treatment was ca 1 .@-fold higher than the ambient level throughout the experiment. The level of inorganic nutrients were very low, compared with the level in Expt I, but was quite similar to that in Expt 11, with a nitrate-nitrite concentration starting at 3.16 pmol/L and decreasing to 1.47 ymol/L over 7 days and a silicate concentration of 4.42-2.33 pmol/L. Detailed results are reported in Payet (66); only the main features are summarized here. The ratio of 2-methylnaphthalene to 1 -methylnaphthalene (2- MN: 1-MN) was calculated from chromatographic data and was used to characterize the weathering process affecting dissolved oil in different microcosms (67). The ratio decreases under bacterial degradation because 1-MN is preferably degraded to 2-MN.
During Expt I11 the mean 2-MN : 1-MN ratio (?SD) decreased from 0.7 S 0.2 on day 0 to 0.5 t 0.2 on day 6 (results not shown).
988 E. Pelletier eta/.
T
75flll _I m WSF-10mdL - WSF-20mglL - WSF -50 mgL
DI D2 D3 D4 D5 D6 D7 D1 D2 D3 D4 D5 D6 D7 Dl D2 D3 D4 DS D6 D7
Day of the experiment Figure 6. Temporal changes in total chlorophyll a (A) and nanophytoplanktonic abundance (B), according to the three initial WSF concentrations used (Expt In). Data are mean values i- SD.
No multivariate effects between UVBR and WSF treatments were detected over time but a 2-way analysis of variance indicated that WSF had a statistically significant degradative effect on the ratio during days 2-6 ( P < 0.01). There was a significant decrease in the 2-MN: 1-MN ratio under LWSF (concentration, 2 mg/L), com- pared with MWSF (concentration, 10 mg/L) and HWSF (concen- tration, 50 mg/L). Although no significant UVBR effect was found on degradation activities the treatment kept in dark seemed to result in a greater degradation for 10 and 20 mg of WSF/L, compared with UVBR treatments. It might be a first indication that hydrocarbon-specific bacteria stimulated by the presence of WSF were affected by UVBR.
Total phytoplankton population, which was estimated on the basis of the Chl u concentration, was usually high at that time of the year; it drastically decreased in the first 48 h and remained at a low level until the end of the experiment in all microcosms, including those with natural light and without WSF (Fig. 6). High phytoplankton abundance and low nutrient concentrations made the assemblage particularly sensitive to environmental stressors, such as sampling and water transfer, changes in light intensity and presence of dissolved hydrocarbons. Throughout the experiment combined UVBR and WSF treatments did not induce large changes in both Chl a and nanophytoplankton abundance (Fig. 6). UVBR effects appeared to be more detrimental than WSF contamination. Indeed, nanophytoplankton abundance was reduced under enhanced UVBR, whereas WSF contamination seemed to stimulate their growth rate (especially at concentrations of cu 20 and ca 50 mg/L), relative to uncontaminated NUVBR environment (Fig. 6). Previous studies have also observed such an effect with hydrocarbon contamination (64,68) and this could be attributed to a shift in the phytoplanktonic assemblage with the appearance of tolerant species, such as flagellates. The community structure was probably the most important factor related to the sensitivity observed during Expt III. For instance, in a study performed in Argentina (69) only postbloom assemblages were significantly affected by UVBR, compared to the prebloom and bloom periods,
and this was attributed to a different population located in the natural phytoplanktonic community.
Thus, the observed drastic decrease of nanophytoplankton abundance under NUVBR and HUVBR can be at least partly be attributed to the occurrence of more-vulnerable species, such as nanoplanktonic flagellates, that generally dominate during post- bloom events in the St. Lawrence Estuary (70-72). Many studies have demonstrated the sensitivity of phytoflagellates exposed to UVBR (61,69,72). Overall, the effect of HUVBR was significantly stronger than that of NUVBR on nanophytoplankton cell values during days 3-6, whereas both total Chl u and nanophytoplankton abundance remained almost constant and significantly higher under the dark treatment, compared with the UVBR treatments.
The bacterial community in Expt I11 was not affected in the same way as phytoplankton. The initial population (concentration, cu 1.9 X lo6 cells mL-') started to grow within 48 h and reached a concentration of 6.9 X lo6 cells mL-' by the end of the experiment, with a peak at 10 X lo6 cells mL-' during treatment with 50 mg WSF/L while in the dark. For comparison the bacterial count in Expt I started at 0.2 X lo2 cells mL-' and increased by a factor of 10 in microcosms treated with WSF. In Ushuaia (Expt 11) the total count ranged from 3.0 X lo6 to 7.4 X lo6 cells mL-'.
The abundance of bacteria increased with increasing WSF levels, most likely because of the development of an opportunistic bacterial community on dead phytoplankton cells and the de- velopment of hydrocarbon-specific bacteria that are able to grow on the additional carbon source provided by dissolved hydro- carbons (Fig. 7). However, when WSF was considered jointly with enhanced UVBR exposure, the lowest values of bacterial abun- dance were recorded and more drastically under the highest WSF level. It was likely that enhanced UVBR exposure was enough to induce phototoxicity of the WSF. As a general trend differences between UVBR and WSF treatments increased with time. The effects of fluctuations in UVBR conditions and WSF treatments on bacteria were more important at the end of the experiment (days 5- 6) than at the beginning (days 0-1). Bacteria appeared more sen-
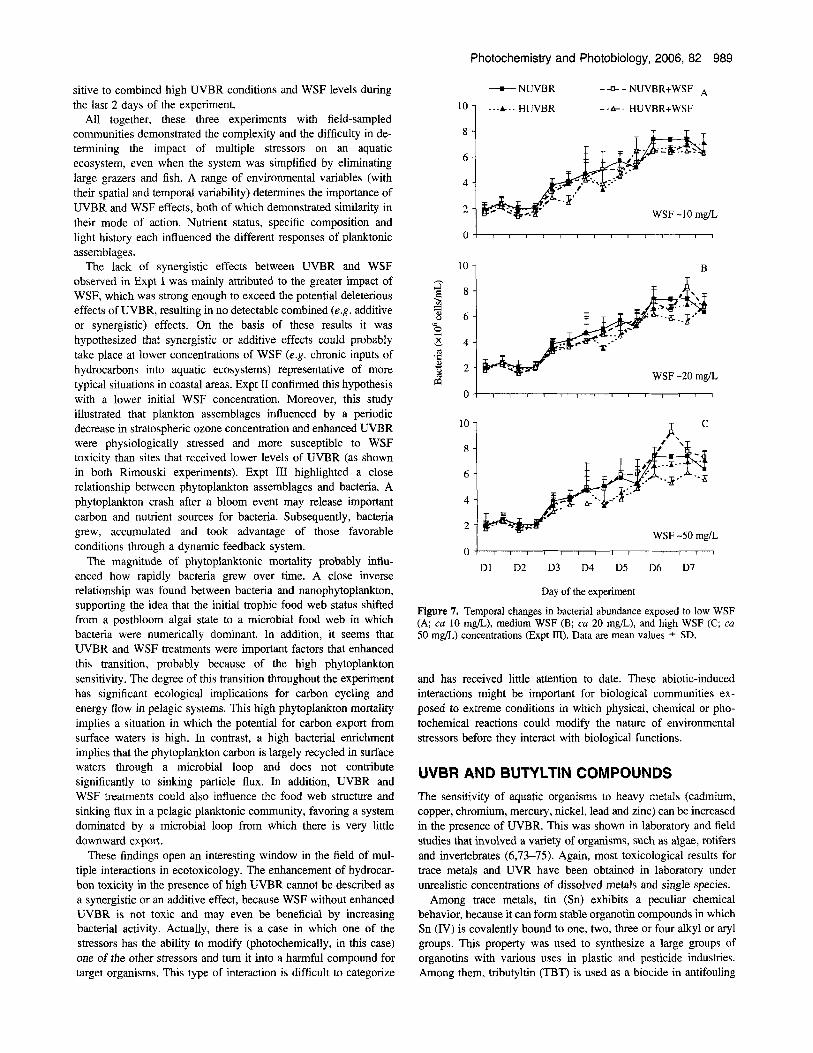
Photochemistry and Photobiology, 2006, 82 989
sitive to combined high UVBR conditions and WSF levels during the last 2 days of the experiment.
All together, these three experiments with field-sampled communities demonstrated the complexity and the difficulty in de- termining the impact of multiple stressors on an aquatic ecosystem, even when the system was simplified by eliminating large grazers and fish. A range of environmental variables (with their spatial and temporal variability) determines the importance of UVBR and WSF effects, both of which demonstrated similarity in their mode of action. Nutrient status, specific composition and light history each influenced the different responses of planktonic assemblages.
The lack of synergistic effects between UVBR and WSF observed in Expt I was mainly attributed to the greater impact of WSF, which was strong enough to exceed the potential deleterious effects of UVBR, resulting in no detectable combined (e.g. additive or synergistic) effects. On the basis of these results it was hypothesized that synergistic or additive effects could probably take place at lower concentrations of WSF (e.g. chronic inputs of hydrocarbons into aquatic ecosystems) representative of more typical situations in coastal areas. Expt 11 confirmed this hypothesis with a lower initial WSF concentration. Moreover, this study illustrated that plankton assemblages influenced by a periodic decrease in stratospheric ozone concentration and enhanced UVBR were physiologically stressed and more susceptible to WSF toxicity than sites that received lower levels of UVBR (as shown in both Rimouski experiments). Expt III highlighted a close relationship between phytoplankton assemblages and bacteria. A phytoplankton crash after a bloom event may release important carbon and nutrient sources for bacteria. Subsequently, bacteria grew, accumulated and took advantage of those favorable conditions through a dynamic feedback system.
The magnitude of phytoplanktonic mortality probably influ- enced how rapidly bacteria grew over time. A close inverse relationship was found between bacteria and nanophytoplankton, supporting the idea that the initial trophic food web status shifted from a postbloom algal state to a microbial food web in which bacteria were numerically dominant. In addition, it seems that UVBR and WSF treatments were important factors that enhanced this transition, probably because of the high phytoplankton sensitivity. The degree of this transition throughout the experiment has significant ecological implications for carbon cycling and energy flow in pelagic systems. This high phytoplankton mortality implies a situation in which the potential for carbon export from surface waters is high. In contrast, a high bacterial enrichment implies that the phytoplankton carbon is largely recycled in surface waters through a microbial loop and does not contribute significantly to sinking particle flux. In addition, UVBR and WSF treatments could also influence the food web structure and sinking flux in a pelagic planktonic community, favoring a system dominated by a microbial loop from which there is very little downward export.
These findings open an interesting window in the field of mul- tiple interactions in ecotoxicology. The enhancement of hydrocar- bon toxicity in the presence of high UVBR cannot be described as a synergistic or an additive effect, because WSF without enhanced UVBR is not toxic and may even be beneficial by increasing bacterial activity. Actually, there is a case in which one of the stressors has the ability to modify (photochemically, in this case) one of the other stressors and turn it into a harmful compound for target organisms. This type of interaction is difficult to categorize
- NUVBR
. . .I.. HUVBR
- Z- - NUVBR+WSF A
- .P . HUVBR+WSF
z. WSF -10 mg/L
I
B T
O J I I I I I I I I I I I I I I
D1 D2 D3 D4 D5 D6 D7
Day of the experiment
Figure 7. Temporal changes in bacterial abundance exposed to low WSF ( A cu 10 mg/L), medium WSF (B; cu 20 rng/L), and high WSF ( C cu 50 mgb) concentrations (Expt m). Data are mean values 5 SD.
and has received little attention to date. These abiotic-induced interactions might be important for biological communities ex- posed to extreme conditions in which physical, chemical or pho- tochemical reactions could modify the nature of environmental stressors before they interact with biological functions.
UVBR AND BUTYLTIN COMPOUNDS The sensitivity of aquatic organisms to heavy metals (cadmium, copper, chromium, mercury, nickel, lead and zinc) can be increased in the presence of UVBR. This was shown in laboratory and field studies that involved a variety of organisms, such as algae, rotifers and invertebrates (6,73-75). Again, most toxicological results for trace metals and UVR have been obtained in laboratory under unrealistic concentrations of dissolved metals and single species.
Among trace metals, tin (Sn) exhibits a peculiar chemical behavior, because it can form stable organotin compounds in which Sn (IV) is covalently bound to one, two, three or four alkyl or aryl groups. This property was used to synthesize a large groups of organotins with various uses in plastic and pesticide industries. Among them, tributyltin (TBT) is used as a biocide in antifouling

990 E. Pelletier et a/.
Table 2. Summary of effects of tributyltin (TBT) and UVBR on field-collected plankton community exposed over 5 days in mesocosms.*
Measured parameter Observations Ecotoxicological significance
Nutrients Significant differences in N and P concentrations between TBT treatments and normal UVBR where nutrients are depleted in 5 days
Steady decrease of Chl a in all TBT treatments and increase of Chl a in TBT-free treatments; significant photoinhibition of PS 11, especially in TBT-positive enhanced UVBR.
Significant decrease of total abundance of cells in TBT treatments with a worse situation in
TBT increased cell size of nanoautotrophs and induced high SSC values; indices of picoautotrophs not significantly affected in the first days
Growth rates of larger cells are much more affected than those of smaller cells
Bacterial abundance was significantly higher in all TBT treatments toward normal UVBR
Inhibition of the specific activity in TBT treatments
A rapid shift of the community from autotrophs to heterotrophs and low assimilation of nitrate
Chlorophyll a Highly significant inhibitory effect induced by TBT on phytoplankton cells
Abundance of Synergetic effects of TBT-positive enhanced levels of UVBR on the abundance of cells
TBT inhibition of photophosphorylation and subsequent inhibition of ATP-positive Pi exchange activities
Shift from large autotrophs to small autotrophs as a transition step before heterotrophy takes place
Respiration of suddenly available organic carbon by bacteria
Adaptation of the bacterial community to a toxic environment
phytoplankton cells ((20 pm) enhancedUVBRtreatment
Cell characteristics
Growth rates
Bacterial abundance
Bacterial-specific activity
*Data are from Sargian et al. (25). SSC = side-light scattering related to refractive index, granularity and cell shape.
paints for ship hulls. TBT exhibits a high toxic activity toward a variety of aquatic nontargeted organisms (76), including phytoplankton (77,78) and bacterioplankton (79). Despite the current restrictions on TBT in antifouling paints and a total ban expected by January 2008, this organometallic compound persists in coastal areas at concentrations considered to be chronically toxic (80). Butyltin compounds absorb radiant energy only in the UV region and essentially in the UVBR component of sunlight (280- 320 nm), which can cause photodegradation of butyltins in surface waters (81). It can be hypothesized that contamination of estuaries and coastal environments by TBT and its degradation by-products (dibutyltin and monobutyltin) can increase the susceptibility of planktonic assemblages to enhanced UVBR levels.
Using the previously described setup for Expt I on UVBR and PAHs, we exposed a field-collected planktonic community to dissolved TBT (0.3 nM) and UVBR for 5 days (25). Single and combined toxic effects were investigated by determining bacterial abundance and productivity, phytoplankton abundance, cellular characteristics and growth rates and Chl a fluorescence. The main findings and their interpretation are summarized in Table 2. The results clearly demonstrated that TBT addition resulted in a decrease in photosynthesis, an increase in the mean phytoplankton cell size and changes in cell shape, with ultimately a significant decrease in cell abundance. The toxic effects of TBT were magnified when cells were jointly exposed to enhanced UVBR. Sargian et al. (25) have described cellular mechanisms by which UVBR and TBT lead to these catabolic cellular changes.
The bacterial component of the natural plankton assemblage showed a different behaviour, with an apparent stimulation of the bacterial abundance (i.e. additional available carbon) associated with an inhibition of the bacterial activity, suggesting a shift in the specific composition of the bacterial community toward a consor- tium more resistant to butyltins.
The synergistic mechanism of TBT and UVBR on a natural plankton community could imply either a UVBR-associated increase in TBT sensitivity or a TBT-associated increase in UVBR sensitivity of cells but it was not possible to define the exact synergistic mechanism involved. However, it was evident that both
environmental stressors acted together to induce negative effects reflected in the continuously decreasing total abundance of phytoplankton cells (<20 pm), as well as in the inhibition of the bacterial productivity. As previously mentioned for PAHs, the combined effects of UVBR and TBT favor the microbial food web characterized by ammonium-based production (i.e. “regenerated production”), dominance of small organisms (flagellates, bacteria and microzooplankton) and low carbon export.
PERSPECTIVES Identifying interactions between stressors is a challenging task because the potential effects of two or more stressors acting simultaneously are often astonishingly complex and could include abiotic induced interactions, biodegradation and bioactivation, food availability, seasonal and annual cycles, interspecific interactions, tolerance and protective behavior, population shifts and biodiversity losses. Models have been developed to assess effects of multiple stressors (82-85) but most authors agree that the predictive success of a given model is first governed by the quality of data used to test the model. Many results reported in our review show how problematic the extrapolation of laboratory results is, because most laboratory experiments do not account for in- terspecific interactions, autoprotective behavior of sensitive species and chemical interactions with naturally occurring organic matter. Even water temperature is often neglected, On the other hand, results obtained with field-sampled communities and mesocosms appear to be much more relevant to a proper evaluation of the ecological impacts of UVBR and anthropogenic pollutants released in coastal waters. A shift toward small species seems to be a generalized stress response because the ability to survive various stressors might depend on a rapid reproductive rate and a small body size (86,87). A shift toward heterotrophic assemblages is an unsuspected adverse effect that reduces the sink of C02 and most probably reduces the biodiversity of coastal waters. The tolerance of plankton species to multiple stressors, particularly enhanced UVBR and pollutants, should be determined and models on impacts of multiple stressors on biodiversity and ecosystem functioning (88) should be developed in the near future.

Photochemistry and Photobiology, 2006, 82 991
Acknowledgements-This work was supported by the National Sciences and Engineering Research Council of Canada, the Fonds Qudbecois de Recherche sur la Nature et les Technologies, the Inter American Institute for Global Change Research and the Canada Research Chairs Program.
REFERENCES 1.
2.
3.
4.
5.
6.
7.
8.
9.
De Mora, S., S. Demers and M. Vernet (2000) The Effects of UV Radiation in the Marine Environment. Cambridge University Press Environmental Chemistry Series, Cambridge. Helbling, E. W. and H. Zagarese (2003) UV Effects in Aquatic Organisms and Ecosystems. Springer-Verlag. New York. El-Alawi, Y. S., D. G. Dixon and B. M. Greenberg (2001) Effects of pre-incubation period on the photoinduced toxicity of polycyclic aromatic hydrocarbons to the luminescent bacterium Vibrio fischeri. Environ. Toxicol. 16, 277-286. Steevens, J. A., M. Slateny, D. Schlenk, A. AryI and W. H. Benson ( I 999) Effects of ultraviolet-B light and polyaromatic hydrocarbon exposure on sea urchin development and bacterial bioluminescence. Mar. Environ. Res. 48, 439-457. Gala, W. R. and J. P. Giesy (1992) Photo-induced toxicity of anthracene to the green alga, Selenastrum capricornutum. Arch. Environ. Contam. Toxicol. 23, 316-323. Rai, P. K. and L. G. Rai (1997) Interactive effects of UV-B and Cu on photosynthesis, uptake and metabolism of nutrients in a green alga Chlorella vulgaris under simulated ozone column. J . Gen. Appl. Microbiol. 43, 281-288. Southerland, H. A. and A. J. Lewitus (2004) Physiological responses of estuarine phytoplankton to ultraviolet light-induced fluoranthene toxicity. J . Exp. Mar. Biol. Ecol. 298, 303-322. Huang, X. D., D. G. Dixon and B. M. Greenberg (1993) Impacts of UV radiation and photomodification on the toxicity of PAHs to the higher plant Lemna gibba (duckweed). Environ. Toxicol. Chem. 12, 1067-1 077. Huang, X. D., B. J. McConkey, S. T. Babu and B. M. Greenberg (1997) Mechanisms of photoinduced toxicity of photomodified anthracene to plants: Inhibition of photosynthesis in the aquatic higher plant Lemna gibba (duckweed). Environ. Toxicol. Chem. 16, 1707-1715.
10. Ankley, G. T., S. A. Collyard, P. D. Monson and P. A. Kosian (1994) Influence of ultraviolet light on results of toxicity tests conducted with sediments contaminated with polycyclic aromatic hydrocarbons. Environ. Toxicol. Chem. 13, 1791-1796.
11. Boese, B. L., J. 0. Lamberson, R. C. Swartz, R. Ozretich and F. Cole (1998) Photoinduced toxicity of PAHs and akylated PAHs to a marine infaunal amphipod (Rhepoxynius abronius). Arch. Environ. Contam. Toxicol. 34, 235-240.
12. Diamond, S. A., N. J. Milroy, V. R. Mattson, L. J. Heinis and D. R. Mount (2003) Photoactivated toxicity in amphipods collected from polycyclic aromatic hydrocarboncontaminated sites. Environ. Toxicol. Chem. 22, 2752-2760.
13. Cleveland, L., E. E. Little, R. D. Calfee and M. G. Barron (2000) Photoenhanced toxicity of weathered oil to Mysidopsis bahia. Aquat. Toxicol. 49, 63-76.
14. Pelletier, M. C., R. M. Burgess, K. T. Ho, A. Kuhn, R. A. McKinney and S. A. Ryba (1997) Phototoxicity of individual polycyclic aromatic hydrocarbons and petroleum to marine invertebrate larvae and juveniles. Environ. Toxicol. Chem. 16, 219g2199.
15. Peachey, R. B. M. (2005) The synergism between hydrocarbon pollutants and UV radiation: Potential link between coastal pollution and larval mortality. J . Exp. Mar. Biol. Ecol. 315, 103-114.
16. Barron, M. G., M. G. Carls, J. W. Short, S. D. Rice, R. A. Heintz, M. Rau and R. Di Giulio (2005) Assessment of the phototoxicity of weathered Alaska North Slope crude oil to juvenile pink salmon. Chemosphere 60, 105-1 10.
17. Bowling, J. W., G. J. Leversee, P. F. Landrum and J. P. Giesy (1983) Acute mortality of anthracene-contaminated fish exposed to sunlight. Aquat. Toxicol. 3, 79-90.
18. Little, E. E., L. Cleveland, R. Calfee and M. G. Barron (2000) Assessment of the photoenhanced toxicity of a weathered oil to the tidewater silverside. Environ. Toxicol. Chem. 19, 926-932.
19. Zaga, A,, E. E. Little, C. F. Rabeni and M. R. Ellersieck (1998) Photoenhanced toxicity of a carbamate insecticide to early life stage anuran amphibians. Environ. Toxicol. Chem. 17, 2543-2553.
20.
21.
22.
23.
24.
25,
26.
27,
28.
29.
30
Boese. B. L., J. 0. Lamberson, R. C. Swartz, F. A. Cole, J. Pelletier and J. Jones (1999) Toxicity and phototoxicity of mixture of highly lipophilic PAH compounds in marine sediment: Can the sum PAH model be extrapolated? Arch. Environ. Contam. Toxicol. 36, 270-280. McDonald, B. G. and P. M. Chapman (2002) PAH phototoxicity- An ecologically irrelevant phenomenon? Mar. Pollut. Bull. 44,
Manvood, C. A., R. E. H. Smith, K. R. Solomon, M. N. Charlton and B. M. Greenberg (1999) Intact and photomodified polycyclic aromatic hydrocarbons inhibit photosynthesis in natural assemblage of Lake Erie phytoplankton exposed to solar radiation. Ecotoxicol. Environ. Sac 44, 322-327. Ohwada, K., M. Nishumura, M. Wada, H. Nomura, A. Shibata, K. Okamoto, K. Toyoda, A. Yoshida, H. Takada and M. Yamada (2003). Study of the effect of water-soluble fractions of heavy-oil on coastal marine organisms using enclosed ecosystems, mesocosms. Mar. Pollut.
Sargian, P., B. Mostajir, K. Chatila, G. A. Ferreyra, E. Pelletier and S. Demers (2005a) Non-synergistic effects of water-soluble crude oil and enhanced ultraviolet-B radiation on a natural plankton assemblage. Mar. Ecol. Prog. Ser. 294, 63-77. Sargian, P., E. Pelletier, B. Mostajir, G. A. Ferreyra and S. Demers (2005b) TBT toxicity on a natural planktonic assemblage exposed to enhanced ultraviolet-B radiation. Aquat. Toxicol. 73, 299-3 14. Samanidou, V., K. Fytianos, G. Pfister and M. Bahair (1988). Photo- chemical decomposition of carbamate pesticides in natural waters of Northern Greece. Sci. Total Environ. 76, 85-92. Miller, P. L. and Y. P. Chin (2002) Photoinduced degradation of carbaryl in a wetland surface water. J . Agric. Food Chem. 50, 6758-6765. Tringle, M. D., M. Pirmohamed, E. Templeton, A. S. Wilson, S. Wadden, N. R. Kitteringham and B. K. Park (1993) An investigation of the formation of cytotoxic, genotoxic, protein-reactive and stable metabolites from naphthalene by human liver microsomes. Biochem. Pharmacol. 46, 1529-1538. Bridges, C. M. and M. D. Boone (2003) The interactive effects of UV-B and insecticide exposure on tadpole survival, growth and development. Biol. Conserv. 113, 49-54. Readmen, J. W., T. A. Albanis, D. Barcelo, S. Galissi, J. Tronczynski and G. P. Gabrielides (1993) Herbicide contamination in Mediterra- nean estuarine waters. Results from a MEDPOL pilot survey. Mar. Pollut. 26, 613419.
1321-1326.
Bull. 47, 78-84.
31. Albanis, T. A. and D. G. Hela (1998) Pesticide concentrations in Louros river and their fluxes into the marine environment. Int. .I. Environ. Anal. Chem. 70, 105-120.
32. Lm, Y.-J., M. Karuppiah, A. Shaw and G. Gupta (1999) Effect of simulated sunlight on atrazine and metolachlor toxicity of surface waters. Ecotoxicol. Environ. Sac 43, 35-37.
33. Konstantinou, 1. K., A. K. Zarkadis and T. A. Albanis (2001) Photodegradation of selected herbicides in various natural waters and soils under natural conditions. J . Environ. Qual. 30, 121-130.
34. Detenbeck, N. E., R. Hermanutz, K. Allen and M. C. Swift (1996) Fate and effects of the herbicide atrazine in flow-through wetland mesocosms. Environ. Toxicol. Chem. 15, 937-946.
35. Oris, J. T., A. T. Hall and J. D. Tylka (1990) Humic acids reduce the photo-induced toxicity of anthracene to fish and daphnia. Environ. Toxicol. Chem. 9, 575-583.
36. Zepp, R. G. (1988) Environmental photoprocesses involving natural organic matter. In Humic Substances and their Role in the Environ- ment (Edited by F. H. Frimmel R. F. Christman), pp. 193-214. Wiley, New York.
37. Zeng, K., H. Hwang and H. Yu (2002) Effect of dissolved humic substances on the photochemical degradation rate of 1-aminopyrene and atrazine. Int. J . Mol. Sci. 3, 1048-1057.
38. Scrano, L., S. A. Bufo, M. D’Auria, 0. Meallier, A. Behechti and K. W. Shramm (2002) Photochemistry and photoinduced toxicity of Acifluorfen, a diphenyl-ether herbicide. J . Environ. Qual. 31, 268-274.
39. Leo, A. J. (1993) Calculating log P,, from structures. Chem. Rev. 93, 128 1-1 306.
40. Neff, J. (1979) Polycyclic Aromatic Hydrocarbons in the Aquatic Environment. Applied Science Publishers, Essex, United Kingdom.
41. Mackay, D., W. Y. Shiu and K. C. Ma (1991a) Illustrated Handbook of Physical-chemical Properties and Environmental Fate for Organic

992 E. Pelletier eta/,
Chemicals. Volume I : Monoaromatic Hydrocarbons, Chlorobenzenes and PCBs. Lewis Publishers, Chelsea, MI.
42. Mackay, D., W. Y. Shiu and K. C. Ma (1991b) IllustratedHandbook of Physical-chemical Properties and Environmental Fate for Organic Chemicals. Volume II : Polycyclic Aromatic Hydrocarbons, Polychlori- nated Dioiins, and Dibenzofurans. Lewis Publishers, Chelsea, Michigan.
43. Dabestani, R. and I. N. Ivanov (1999) A compilation of physical, spectroscopic and photophysical properties of polycyclic aromatic hydrocarbons. Photochem. Photobiol. 70, 10-34.
44. Karydis, M. (1982) Toxicity of a photooxidised crude oil on two marine microalgae. Bot. Mar. 25, 25-29.
45. Ren, L., X.-D. Huang, B. J. McConkey, D. G. Dixon and B. M. Greenberg (1994) Photoinduced toxicity to three polycyclic aro- matic hydrocarbons (fluoranthene, pyrene and naphthalene) to the duckweed Lemna gibba L. (3-3. Ecotoxicol. Environ. Saf. 28, 16@171.
46. Arfsten, D. P., D. J. Schaffer and D. C. Mulveny (1996) The effects of near ultraviolet radiation on the toxic effects of PAH in animals and plants: A review. Ecotox. Environ. Saf, 33, 1-24.
47. Nikkila, A., S. Penttinen and J. V. K. Kukkonen (1999) UV-B induced acute toxicity of pyrene to the waterflea Daphnia magna in natural freshwaters. Ecotoxicol. Environ. Saf, 44, 27 1-279.
48. Duxbury, C. I., D. G. Dixon and B. M. Greenberg (1997) The effects of simulated solar radiation on the bioaccumulation of polycyclic aromatic hydrocarbons by the duckweed Lemna gibba. Environ. Toxicol. Chem. 16, 1739-1748.
49. Krylov, S. N., X.-D. Huang, L. F. Zeiler, D. G. Dixon and B. M. Greenberg (1997) Mechanism quantitative structure-activity relation- ship model for the photoinduced toxicity of polycyclic aromatic hydrocarbons. I. Physical model based on chemical kinetics in a two- compartment system. Environ. Toxicol. Chem. 16, 2283-2295.
50. McConkey, B. J., C. L. Duxbury, D. G. Dixon and B. M. Greenberg (1997) Toxicity of a PAH photooxidation product to the bacteria Photoharterium phosphoreum and the duckweed Lemna gibba: Effects of phenanthrene and its primary photoproduct, phenanthrenequinone. Environ. Toxicol. Chem. 16, 892-899.
51. Mallakin, A., B. J. McConkey, G. Miao, B. McKibben, V. Snieckus, D. G. Dixon and B. M. Greenberg (1999) Impacts of structural photomodification on the toxicity of environmental contaminants: Anthracene photooxidation products. Ecotoxicol. Environ. Sab 43, 204-2 12.
52. Newsted, J. L. and J. P. Giesy (1987) Predictive models for photo- induced acute toxicity of polycyclic aromatic hydrocarbons to Daphnia magna, Strauss (Cladocera, Crustacea). Environ. Toxicol. Chem. 6, 445-46 1.
53. Boese, B. L., J. 0. Lamberson, R. C. Swartz and R. J. Ozretich (1997) Photoinduced toxicity of fluoranthene to seven benthic crustaceans. Arch. Environ. Contam. Toxicol. 32, 389-393.
54. Huovinen, P. S., M. R. Soimasuo and A. 0. J. Oikari (2001) Photoinduced toxicity of retene to Daphnia magna under enhanced UV-B radiation. Chemosphere 45, 683-691.
55. Monson, P. D., G. T. Ankley and P. A. Kosian (1 995) Phototoxic response of Lumbriculus variegates to sediments contaminated by polycyclic aromatic hydrocarbons. Environ. Toxicol. Chem. 14,891694.
56. Swartz, R. C.. S. P. Ferraro, J. 0. Lamberson, F. A. Cole, R. J. Ozretich, B. L. Boese, D. W. Schults, M. Behrenfeld and G. T. M l e y (1997) Photoactivation and toxicity of mixtures of polycyclic aromatic hydrocarbon compounds in marine sediment. Environ. Toxical. Chem. 16, 2151-2157.
57. Foote, C. S. (1991) Definition of type I and type I1 photosensitized oxidation. Photochem. Photobiol. 54, 659.
58. Weinstein, J. W., J. T. Oris and D. H. Tayler (1997) An ultrastructural examination of the mode of UV-induced toxic action of fluoranthene in the fathead minnow, Pimephales promelas. Aquat. Toxicol. 39, 1-22.
59. McCloskey, J. T. and J. T. Oris (1993) Effects of anthracene and solar ultraviolet radiation exposure on gill ATPase and selected hematologic measurements in bluegill sunfish (Lepomis macrochirus). Aquar. Toxicol. 24, 207-218.
60. Sargian, P. (2005) Comportement des premiers maillons du dseau trophique marin exposis au rayonnement ultraviolet-B et aux contaminants dissous. Ph.D. thesis, UniversitC du Quebec h Rimouski, Quebec.
61. Vernet, M. (2000) Effects of UV radiation on the physiology and ecology of marine phytoplankton. In The Effects of UV Radiation in rhe Marine Environment (Edited by S. De Mora, S. Demers and M. Vernet), pp. 237-278. Cambridge Environmental Chemistry, University Press.
62. Mostajir, B., S. Demers, S. de Mora, C. Belzile, J.-P. Chanut, M. Gosselin, S. Roy, P. Zulema Villegas, J. Fauchot, J. Bouchard, D. Bird, P. Monfort and M. Levasseur (1999) Experimental test of the effect of ultraviolet-B radiation on a planktonic community. Limnol. Oceanogr. 44, 586-596.
63. Lebaron, P., P. Servais, H. AgoguC, C. Courties and F. Joux (2001) Does the high nucleic acid content of individual bacterial cells allow us to discriminate between active cells and inactive cells in aquatic systems? Ap 1 Environ. Microbiol. 67, 1775-1782.
Pelletier and S. Roy (1996) Effects of dispersed and adsorbed crude oil on microalgal and bacterial communities of cold seawater. Ecotaxicol. 5, 229-25 1.
65. Bagchi, M., D. Bagchi, J. Balmoori, X. Ye and S. J. Stohs (1998) Naphthalene-induced oxidative stress and DNA damage in cultured macrophage J774A.1 cells. Free Rad. Bid . Med. 25, 137-143.
66. Payet, J. (2003) Effets combinks du rayonnement ultraviolet-B et des hydrocarbures dissous sur les communautCs microbiennes d’un Ccosysttme cBtier naturel. M.S. thesis, UniversitC du QuCbec B Rimouski, Quebec.
67. Siron, R., 6. Pelletier, D. Delille and S. Roy (1993). Fate and effects of dispersed crude oil under icy conditions simulated in mesocosms. Mar. Environ. Res. 35,273-302.
68. Roy, S., R. Siron and E. Pelletier (1991) Comparison of radiocarbon uptake and DCMU-fluorescence techniques in evaluating dispersed oil effects on phytoplankton photosynthetic activity. Water Res. 25, 1249-1254.
69. Barbieri, E., V. Villafaiie and E. W. Helbling (2002) Experimental assessment of UV effects on temperate marine phytoplankton when exposed to variable radiation regimes. Limnol. Oceanogr. 47, 1648- 1655.
70. Levasseur, M., J.-C. Therriault and L. Legendre (1984) Hierarchical control of phytoplankton succession by physical factors. Mar. Ecol. Prog. Ser. 19, 21 1-222.
71. Ferreyra, G. (1995) Effets du rayonnement ultraviolet sur le plancton des kgions froides tempCr6es et polaires. Ph.D. thesis, UniversitC du QuCbec B Rimouski, Quebec.
72. Roy, S., J.-P. Chanut, M. Gosselin and T. She-Ngando (1996) Characterization of phytoplankton communities in the lower St. Lawrence Estuary using HPLC-detected pigments and cell microscopy. Mar. Ecol. Prog. Ser. 142, 55-73.
73. Duquesne, S. and M. Liess (2003) Increased sensitivity of the macroinvertebrate Paramorea walkeri to heavy-metal contamination in the presence of solar UV radiation in Antarctic shoreline waters. Mar. Ecol. Prog. Ser. 255, 183-191.
74. Preston, B. L., T. W. Snell and R. Kneisel (1999) UV-B exposure increases acute toxicity of pentachlorophenol and mercury to the rotifer Brachionus calycyjorus. Environ. Pollut. 106, 23-3 1.
75. Ray, L. C., B. Tyagi, P. K. Ray and N. Mallick (1998) Interactive effects of UV-B and heavy-metals (Cu and Pb) on nitrogen and phosphorus metabolism of a Nz-fixing cyanobacterium Anabaena doliolum. Environ. Exp. Bot. 39, 221-23 1.
76. Maguire, R. J. (2000) Review of the persistence, bioaccumulation and toxicity of uibutyltin in aquatic environments in relation to Canada’s Toxic Substances Management Policy. Wat. Qual. Res. J . Can. 35, 633-679.
77. Huang, G., S. Dai and H. Sun (1996) Toxic effects of organotin species on algae. Appl. Organornet. Chem. 10, 377-387.
78. Sidharthan, M., K. S. Young, L. H. Woul, P. K. Soon and H. W. Shin (2002) TBT toxicity on the marine microalga Nannochloropsis oculata. Mar. Pollut. Bull. 45, 177-182.
79. Cooney, J. J. and S. Wuertz (1989) Toxic effects of tin compounds on microorganisms. J . Ind. Microbial. 4, 375402.
80. Viglino, L., 6. Pelletier and R. St-Louis (2004) Highly persistent butyltins in northern marine sediments: A long-term threat for the Saguenay Fjord (Canada). Environ. Toxicol. Chem. 23, 2673-268 1.
81. Navio, J. A,, F. J. Marchena and C. Cerrillos (1993) UV photolytic degradation of butyltin chlorides in water. J . Phorochem. Phorobiol. A. Chem. 71, 97-102.
64. Siron, R.,

82. Bruland, K. W., J. R. Donat and D. A. Hutchins (1991) Interactive influences of bioactive trace metals on biological production in oceanic waters. Limnol. Oceunogr. 36, 1555-1577.
83. Hay, M. E., Q. E. Kappel and W. Fenical (1994). Synergisms in plant defenses against herbivores: Interactions of chemistry, calcification, and plant quality. Ecology 75, 1714-1727.
84. Soluk, D. A. (1993) Multiple predator effects: Predicting combined functional response of stream fish and invertebrate predators. Ecology. 74, 219-225
85. Folt, C. L., C. Y. Chen, M. V. Moore and J. Bumaford (1999) Synergism and antagonism among multiple stressors. Limnol. Ocean- ogr. 44, 864-877.
Photochemistry and Photobiology, 2006, 82 993
86. Moore, M. V. and C. L. Folt (1993) Zooplankton body size and community structure: Effects of thermal and contaminant stress. Trends
87. Hanazota, T. (1998) Response of a zooplankton community to insecticide application in experimental ponds: A review and the implications of the effects of chemicals on the structure and functioning of freshwater communities. Environ. Pollur. 101, 361-373.
88. Vinebrooke, R. D., K. L. Cottingham, J. Norberg, M. Scheffer, S. I. Dodson, S. C. Maberly and U. Sommer (2004) Impacts of multiple stressors on biodiversity and ecosystem functioning: The role of species co-tolerance. Oikos 104, 45 1-457.
E d . E d . 8, 178-183.





























