Embed Size (px)
Citation preview

Developmental Biology 242, 96–108 (2002)doi:10.1006/dbio.2001.0544, available online at http://www.idealibrary.com on
Conserved Regulation of the Caenorhabditiselegans labial/Hox1 Gene ceh-13
Adrian Streit,* ,1 Reto Kohler,* ,1,2 Thomas Marty,† ,1,3 Marco Belfiore,*Krisztina Takacs-Vellai,* Maria-Alessandra Vigano,† Ralf Schnabel,‡Markus Affolter,† and Fritz Muller* ,4
*Department of Biology, University of Fribourg, Perolles, CH-1700 Fribourg, Switzerland;†Department of Cell Biology, Biozentrum, University of Basel, Klingelbergstrasse. 70,CH-4056 Basel, Switzerland; and ‡Institut fur Genetik, TU Braunschweig,Spielmannstrasse 7, D-38106 Braunschweig, Germany
Caenorhabditis elegans contains a set of six cluster-type homeobox (Hox) genes that are required during larvaldevelopment. Some of them, but unlike in flies not all of them, are also required during embryogenesis. It has been suggestedthat the control of the embryonic expression of the worm Hox genes might differ from that of other species by beingregulated in a lineal rather than a regional mode. Here, we present a trans-species analysis of the cis-regulatory region ofceh-13, the worm ortholog of the Drosophila labial and the vertebrate Hox1 genes, and find that the molecular mechanismsthat regulate its expression may be similar to what has been found in species that follow a regulative, non-cell-autonomousmode of development. We have identified two enhancer fragments that are involved in different aspects of the embryonicceh-13 expression pattern. We show that important features of comma-stage expression depend on an autoregulatory inputthat requires ceh-13 and ceh-20 functions. Our data show that the molecular nature of Hox1 class gene autoregulation hasbeen conserved between worms, flies, and vertebrates. The second regulatory sequence is sufficient to drive correct earlyembryonic expression of ceh-13. Interestingly, this enhancer fragment acts as a response element of the Wnt/WG signalingpathway in Drosophila embryos. © 2002 Elsevier Science (USA)
Key Words: C. elegans; Drosophila; HOM; Hox genes; ceh-13; labial; transcriptional regulation; autoregulation; Wntpathway.
INTRODUCTION
Hox genes encode a family of helix–turn–helix transcrip-tion factors that are widely conserved within the animalkingdom and that are involved in generating pattern alongthe anterior–posterior (a/p) body axis (for reviews, see Geh-ring et al., 1994; Kenyon et al., 1997; Krumlauf, 1992;McGinnis and Krummlauf, 1992). The genomic organiza-tion of the Hox genes is unique in that they are arrayed inclusters. In Drosophila and in vertebrates, it has been found
1 A.S., R.K., and T.M. contributed equally to this work.2 Present address: Tages Anzeiger, Werdstrasse 21, 8004 Zurich,
Switzerland.3 Present address: Skirball Institute for Biomolecular Medicine,
New York University School of Medicine, 540 First Avenue, NewYork, NY 10016.
4 To whom correspondence should be addressed. Fax: 1141/26-300-97-41. E-mail: [email protected].
96
that the relative position of a gene within these arrayscorresponds to its expression along the a/p axes, such thatthe genes located at the 39 end of a cluster are expressed inthe anterior region of the body and vice versa (for reviews,see Duboule, 1994; Duboule and Morata, 1994).
The genome of Caenorhabditis elegans contains a total ofsix Hox genes. Four of them, ceh-13, lin-39, mab-5, andegl-5, are organized in a loose cluster in the center ofchromosome III (Burglin et al., 1989, 1991; Burglin andRuvkun, 1993), whereas nob-1 and php-3, two AbdB ho-mologs, map to a separate location on the same chromo-some (Ruvkun and Hobert, 1998; Van Auken et al., 2000).Unlike their insect and vertebrate counterparts, the C.elegans genes lin-39, mab-5, and egl-5 do not appear to beessential for embryogenesis. Worms can grow to adulthoodwithout the action of these three Hox genes (Kenyon et al.,1997), and the phenotypes of the corresponding null-mutants only become apparent after embryogenesis, duringlarval stages. Elimination of the function of ceh-13 (the
0012-1606/02 $35.00© 2002 Elsevier Science (USA)
All rights reserved.

97Conserved Inputs Regulate ceh-13 Expression
worm ortholog of the Drosophila gene labial) or of nob-1results in an embryonic lethal phenotype (Brunschwig etal., 1999; Van Auken et al., 2000). Null mutations inceh-13, for example, cause defects in the anterior lateralepidermis and in anterior body wall muscle cells, suggest-ing that this gene plays an important role in the anteriororganization of the C. elegans embryo (Brunschwig et al.,1999).
As stringent spatiotemporal control of Hox gene expres-sion is crucial for their biological function, a large effort hasbeen made to describe the molecular machinery that estab-lishes the expression pattern of these genes. Most of ourcurrent knowledge about the regulation of Hox genes stemsfrom work done in Drosophila and vertebrates. Particularlywell-studied are the events that control the expression ofthe fly Hox1 class gene labial in the embryonic midgut (forreviews, see Bienz, 1994, 1997; Mann and Affolter, 1998).Genetic approaches, combined with an extensive muta-tional analysis of the enhancer region that drives labialexpression in the fly embryonic midgut, led to a complexmodel of the molecular mechanisms that are necessary forproper spatiotemporal control of labial transcription. It hasbeen found that, among others, components of the wingless(wg) and decapentaplegic (dpp) signaling pathways and anautoregulatory input are required for correct labial expres-sion (Immergluck et al., 1990; Hoppler and Bienz, 1995;Grieder et al., 1997; Marty et al., 2001).
Different experimental approaches indicated that theembryonic C. elegans Hox expression patterns are achieved,at least in part, by a lineage-specific control system thatoperates independently of the global a/p position of the cellsin the embryo (Cowing and Kenyon, 1996; Wittmann et al.,1997). Although somehow controversial (Schnabel andSchnabel, 1997), these observations suggested that C. el-egans might use different strategies than those that havebeen characterized previously in Drosophila and verte-brates. Detailed information about the nature of cis-regulatory elements, however, is still lacking for any of theC. elegans Hox genes. Particularly little is known about themechanisms that guide Hox gene expression during earlystages of development. We therefore initiated an analysis ofthe enhancer/promoter region of ceh-13. During these stud-ies, we have characterized two different enhancer fragmentsthat are involved in various aspects of the embryonic ceh-13expression pattern. We show that some features of bean andcomma-stage expression depend on a direct autoregulatoryinput, the molecular nature of which is similar to itscounterparts in mice and flies. We also demonstrate that a740-bp fragment is sufficient to drive expression of a re-porter gene in a pattern indistinguishable from ceh-13expression during early embryogenesis. Interestingly, this740-bp enhancer fragment, when placed in Drosophilamelanogaster acts as a WG-responsive element duringembryogenesis. We propose that this fragment is likely tocontain elements that respond to an early acting Wnt/WGrelated pathway also in C. elegans.
© 2002 Elsevier Science (USA
MATERIALS AND METHODS
Plasmid Constructs
The original ceh-13::gfp reporter construct of pMF1 and the C.elegans strain FR317 carrying pMF1 chromosomally integratedhave been described previously (Wittmann et al., 1997; see alsoFig. 1).
All Drosophila and C. elegans reporter genes were constructedby standard cloning techniques (Sambrooke et al., 1989). Theceh-13 enhancer fragments were generated either by digestion ofpMF1 with HindIII or by PCR amplification, using pMF1 astemplate and primers with suitable restriction sites at their 59 ends.PCR-derived clones were sequenced to exclude mutations due toPCR errors. For the C. elegans GFP reporter constructs, individualenhancer fragments were cloned into the HindIII sites ofpPD107.94 (gift from Andy Fire, Carnegie Institution of Washing-ton, Baltimore, MD; see http://ftp.ciwemb.edu/PNF:byName:/FireLabWeb/FireLabInfo/FireLabVectors/) or pMF1DH3 (a HindIIIdeletion derivative of pMF1 that lacks all sequences upstream ofnucleotide position 22068). (Note that a HindIII site located in thevector 12 bp upstream of the PstI cloning site in pMF1 is not shownin Fig. 1.). Nucleotide positions are indicated with respect to theATG translation start codon of ceh-13 in the wild-type sequence.Drosophila lacZ reporter constructs were made by cloning differentenhancer fragments between the KpnI and BamHI cloning sites ofpCb (a gift of K. Basler, Zurich).
enh3.4 (nucleotide positions 23256 to 26644) was cut frompMF1 for cloning into pPD107.94 (pASF43), or PCR amplified usingprimers 3.3up and 3.3down for cloning into pCb. enh740 (nucleo-tide positions 24001 to 26644) was PCR amplified using primers4192 and 740up for cloning into pPD107.94 (pRK29) or primers770/dros/up and 770/dros/do for cloning into pCb. Note thatenh740 in pCb extends 24 bp further downstream than in pRK29.enh450 (nucleotide positions 23256 to 26172) was amplified byPCR using primers RP3Cel.H.do and RP3Cel.H.up for cloning intopMF1DH3 (pRK24) or pPD107.94 (pRK23), and primers RP3D.K.upand RP3D.B.do for cloning into pCb. Variants of enh450 withmutations in the putative CEH-13 binding site (mutL) or theputative CEH-20 binding site (mutE) or both (mutEL) were con-structed as follows: The point mutation in mutL was introduced bya two-step PCR amplification. The upstream end was amplified byusing the primers RP3Cel.H.up and MMLab2, and the downstreamend by using RP3Cel.H.do and MMLab1. The amplified fragmentswere purified and the full-length fragment was amplified by usingprimers RP3Cel.H.up and RP3Cel.H.do. The other mutant con-structs were generated accordingly, using primers MMExd2 andMMExd1 for mutE, and DS1M2 and DS1M1 for mutE/L. Theresulting fragments were cloned into pMF1DH3 (pLAB for mutL,pEXD for mutE, pDS1M for mutEL).
Oligonucleotides
Oligonucleotides used in this study include: 3.3up, GGGGTAC-CTGTCGGTATGTTCAAAAGCTTC; 3.3down, CGGGATCCTG-GATCTTAGGGAATTGTGG; 4192, CCCAAGCTTTCAGATCCC-TCCACATGTC; 740up, TCTGGTAGACTGTGCAAGCAAC; 770/dros/up, GGGGTACCTCAGATCCCTCCACATGTC; 770/dros/do,CGGGATCCTGGATCTTAGGGAATTGTGG; RP3Cel.H.up, TAT-GTTCAAAAGCTTCTTCCCAG; RP3Cel.H.do, CCCAAGCTTGC-GCTAAGAAGGCGTTCGAG; RP3D.K.up, GGGGTACCTTCTTC-CCAGATGCCCTCC; RP3D.B.do, CGGGATCCGGCGCTAAGA-AGGCGTTCG; MMLab1, GCCACCGAACCAACCATCATTTGG-
). All rights reserved.

98 Streit et al.
© 2002 Elsevier Science (USA). All rights reserved.
99Conserved Inputs Regulate ceh-13 Expression
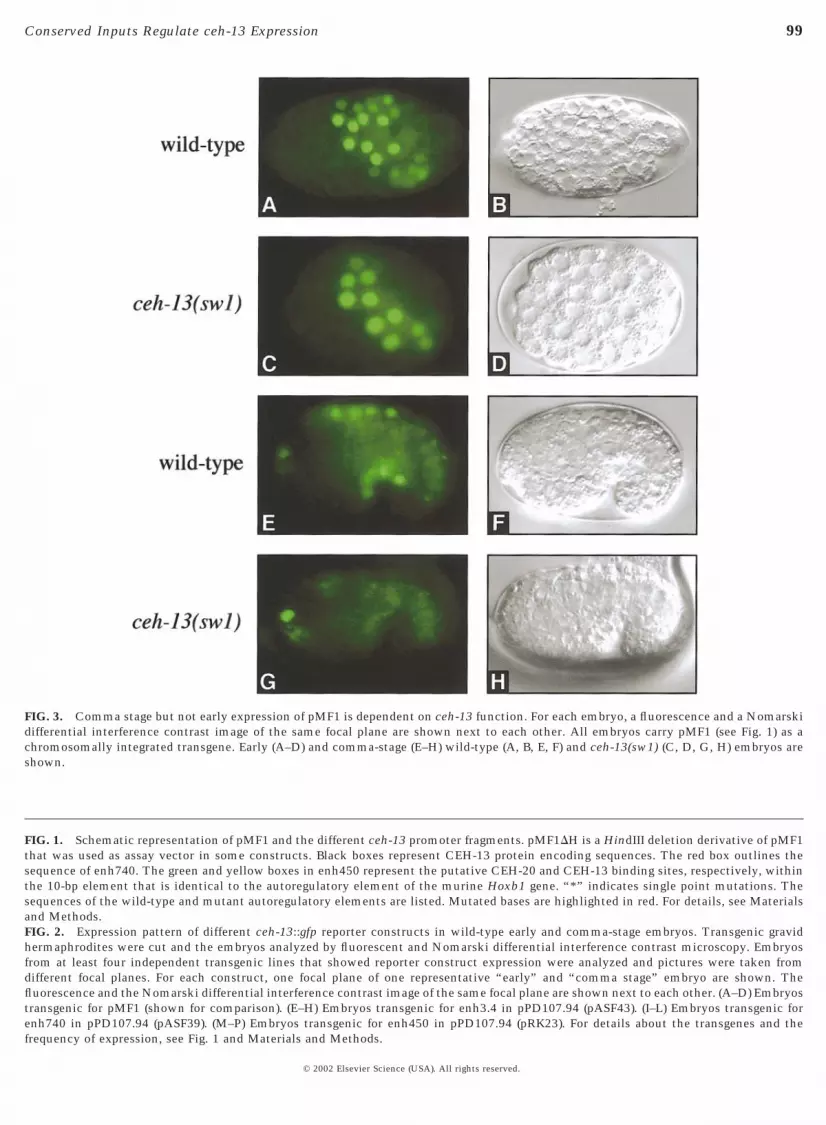
FIG. 3. Comma stage but not early expression of pMF1 is dependent on ceh-13 function. For each embryo, a fluorescence and a Nomarskidifferential interference contrast image of the same focal plane are shown next to each other. All embryos carry pMF1 (see Fig. 1) as achromosomally integrated transgene. Early (A–D) and comma-stage (E–H) wild-type (A, B, E, F) and ceh-13(sw1) (C, D, G, H) embryos areshown.
FIG. 1. Schematic representation of pMF1 and the different ceh-13 promoter fragments. pMF1DH is a HindIII deletion derivative of pMF1that was used as assay vector in some constructs. Black boxes represent CEH-13 protein encoding sequences. The red box outlines thesequence of enh740. The green and yellow boxes in enh450 represent the putative CEH-20 and CEH-13 binding sites, respectively, withinthe 10-bp element that is identical to the autoregulatory element of the murine Hoxb1 gene. “*” indicates single point mutations. Thesequences of the wild-type and mutant autoregulatory elements are listed. Mutated bases are highlighted in red. For details, see Materialsand Methods.FIG. 2. Expression pattern of different ceh-13::gfp reporter constructs in wild-type early and comma-stage embryos. Transgenic gravidhermaphrodites were cut and the embryos analyzed by fluorescent and Nomarski differential interference contrast microscopy. Embryosfrom at least four independent transgenic lines that showed reporter construct expression were analyzed and pictures were taken fromdifferent focal planes. For each construct, one focal plane of one representative “early” and “comma stage” embryo are shown. Thefluorescence and the Nomarski differential interference contrast image of the same focal plane are shown next to each other. (A–D) Embryostransgenic for pMF1 (shown for comparison). (E–H) Embryos transgenic for enh3.4 in pPD107.94 (pASF43). (I–L) Embryos transgenic forenh740 in pPD107.94 (pASF39). (M–P) Embryos transgenic for enh450 in pPD107.94 (pRK23). For details about the transgenes and thefrequency of expression, see Fig. 1 and Materials and Methods.
© 2002 Elsevier Science (USA). All rights reserved.

100 Streit et al.
GTTG; MMLab2, CAACCCAAATGATGGTTGGTTCGGTGGC;MMExd1, CCGAACCATCCATTATTTGGGTTGTGGC; MMExd2,GCCACAACCCAAATAATGGATGGTTCGG; DS1M1, GCCAC-CGAACCAACCATTATTTGGGTTG; DS1M2, CAACCCAAATA-ATGGTTGGTTCGGTGGC; BSWTLEFt, GATTATCCGTCTAC-CTTTTGATGTAAA; BSWTLEFb, CAAAAATTTTACATCAAAA-GGTAGACGGAT; BSMUTLEFt, GATTATCCGTCTACCTGGA-GATGTAAA; and BSMUTLEFb, CAAAAATTTTACATCTCCAGG-TAGACGGAT.
Electrophoretic Mobility Shift Assay with POP-1
Protein preparation. Plasmid pET-21b-popHM6 encodes aminoacids 180–280 of POP-1 tagged with six histidines under thecontrol of a T7 promoter. This fragment contains the DNA bindingdomain of POP-1 and was kindly provided to us by HendrikKorswagen (Korswagen et al., 2000). The protein was expressed inEscherichia coli BL21(DE3) and purified by using the HisTrap kitfrom Amersham Pharmacia Biotech following the manufacturer’sinstructions.
Probe preparation. BSWTLEFt and BSWTLEFb (for wt bindingsite) or BSMUTLEFt and BSMUTLEFb (for mutated binding site)were annealed in TE and labeled by filling in the single-strandedtermini with Klenow enzyme in the presence of 32P a-dATPfollowing standard procedures (Sambrooke et al., 1989) leading tothe double-stranded oligonucleotide wt-pop and mut-pop, respec-tively.
Mobility shift assay. After all the other components had beenpreincubated for 5 min at room temperature (RT), 40,000 cpm ofthe probe (corresponding to about 1.5 ng) were incubated for 25 minat RT in 20 ml of 20 mM Hepes (pH 7.9), 50 mM KCl, 0.5 mg/ml BSA,1 mM DTT, 0.5 mM EDTA, 10% glycerol, 10 ng/mL poly(dI:dC) inthe presence or absence of approximately 100 ng POP-1180–280
protein and of 1 mg of nonlabeled wt or mutated oligonucleotide.Ten microliters of the reaction mix were analyzed on a 5% PAA0.253 TBE gel at 150 V at RT. The gel was fixed in 7% acetic acid,dried, and analyzed with a phosphorimager.
C. elegans Strains and Transformations
C. elegans were grown following standard procedures (Brenner,1974; Stiernagle 1999). C. elegans carrying transgenes on a heri-table extrachromosomal array were constructed by microinjectionas described earlier (Mello et al., 1991; Jin, 1999). The GFP reportergene constructs (50 ng/ml) were coinjected with 200 ng/ml ofplasmid pRF4 carrying the dominant rol-6(su1006) marker, intohermaphrodites of the wild-type strain N2 [except for the wormsshown in Fig. 4, which are derived from strain MT2709 carrying thesilent rol-6(e187n1270) mutation]. The plasmid DNA was preparedby alkaline lysis followed by precipitation with polyethylene glycolas described by Sambrooke et al. (1989) or by using the WizardDNA purification system from Promega. A preliminary inspectionof transgenic F2 animals indicated that, for a given construct, themajority of strains showed an identical GFP expression pattern.The other lines showed little or no expression of the transgene. Forall constructs, at least four independent transgenic lines were keptfor further analysis. Except for construct pRK23 (enh459 inpPD107.94), the number of embryos in a particular transgenic linedisplaying the respective expression pattern was comparable to thetransmission frequency of the transgene in this particular line. ForpRK23, the number of expressing embryos was smaller thanexpected from the transmission frequency and expression in only asmall number of cells was relatively frequent. Therefore, we choose
© 2002 Elsevier Science (USA
construct pRK24 (enh450 in pMF1DH3), which leads to reliableexpression, for further analysis and mutagenesis.
Genotypes of ceh-13 and ceh-20 Mutant Animals
swIs1[rol-6(su1006) 1 ceh-13::gfp] II; ceh-13(sw1)III (Fig. 3).The animals shown were the progeny of rare viable escapers of thesame genotype. Thus, a maternal contribution of ceh-13 function isexcluded.
swEx120[rol-6(su1006) 1 pRK24 ceh-13.-enh450::gfp]; ceh-20(ay38) unc-36(e251) III (Fig. 4). The embryos shown were theprogeny of mothers of the same genotype. Thus, maternal contri-bution of ceh-20 function is excluded. Note that these animals mayhave contained rol-6(n1276 e187) II and/or him-8(e1489) IV ho-mozygously or heterozygously.
RNA Interference
pop-1 RNA interference experiments were performed as de-scribed by Lin et al. (1998), with the exception that double-strandedRNA was used. The RNA was injected into FR317 (Wittmann etal., 1997). All embryos from successful injections showed identicalresults. The pop-1 cDNA that served as template for the synthesisof the dsRNA was kindly provided to us by J. Priess.
Drosophila Strains and Transformations
Drosophila transformations were performed according to stan-dard protocols. For the analysis in null mutant backgrounds, thefollowing alleles were used: wgCX4 (Baker, 1988) and dTCF2 for pan(van de Wetering et al., 1997). Overexpression studies were per-formed by using the following transgenic lines: UAS-armS10(Zecca et al., 1996), UAS-dTcfDN (van de Wetering et al., 1997), and48Y-GAL4 (Martin-Bermudo et al., 1997), 24B-GAL4 (mesoderm-specific), and prd-GAL4 (Brand and Perrimon, 1993). Embryo col-lections, fixations, and immunostainings were performed accord-ing to standard protocols. The anti-WG antibodies were a generousgift from S. Cohen.
Microscopy
Conventional microscopy was performed by using a Leica DMRXA microscope equipped with a Hamamatsu C5810 camera (C.elegans) or a Zeiss Axiophot with DIC optics (Drosophila). Forconfocal microscopy, a Leica DM RXE microscope was used incombination with a TSC NT confocal unit. The desired wave-lengths were provided by an Ar2 and an Ar/Kr2 laser, respectively.The images were processed on Adobe Photoshop 4.0 for Macintosh.
RESULTS
A 3.4-kb Enhancer Fragment Drives Early and LateEmbryonic ceh-13 Expression
Our initial strategy was based on the observation that anintegrated ceh-13::gfp construct (pMF1; Fig. 1) containing8.1 kb of upstream sequences, the first exon, the firstintron, and most of the second exon, mimics endogenousceh-13 expression in transgenic animals. pMF1, like theendogenous ceh-13 gene, is first expressed at the onset ofgastrulation in the posterior daughters of the intestinal
). All rights reserved.

101Conserved Inputs Regulate ceh-13 Expression
precursor cell E (Ep) and of the AB descendants ABxxx(ABxxxp) (Figs. 2A and 2B; Wittmann et al., 1997). Duringlater embryogenesis, CEH-13::GFP is detected in manydifferent tissues and cell types. At the comma stage, forexample, it is expressed in the lateral hypodermal cells H2and V2, in anterior dorsal hypodermal and body wall musclecells and in cells of the prospective ventral nerve cord(VNC) (Figs. 2C and 2D; Brunschwig et al., 1999).
Removal of the most upstream 1.5-kb HindIII fragmentfrom pMF1 (Fig. 1) did not affect the expression pattern,suggesting that this fragment is not required for the properregulation of ceh-13 expression. However, we found the3.4-kb HindIII fragment (enh3.4; Fig. 1) to be responsible fora large part of the normal embryonic ceh-13 expressionpattern. If cloned into pPD107.94, a gfp reporter geneplasmid containing a minimal promoter derived from thepes-10 gene (Fire et al., 1990a,b; see also ftp://ciw2.ciwemb.edu/pub/FireLabVector), enh3.4 was able todrive an early embryonic GFP expression pattern indistin-guishable from that of the endogenous ceh-13 (Figs. 2E and2F). In comma-stage embryos, enh3.4 recapitulated a largepart of the normal ceh-13 expression pattern, includingexpression in cells of the future VNC and embryonic dorsalhypodermal and body wall muscle cells (Figs. 2G and 2H).However, we also observed some ectopic reporter geneexpression in the posterior endoderm and in a few noniden-tified cells of the posterior part of comma-stage embryos.We have also tested the 1.2-kb HindIII fragment (enh1.2)that is located more proximal to the coding region of pMF1(Fig. 1). This fragment appears not to be required forembryonic expression, but drives expression of a GFPreporter gene in larvae and adults, for example, in the maletail (data not shown). Altogether, these results show thatthe enhancer region of ceh-13 contains different regulatoryregions that are responsible for various aspects of thedevelopmental expression pattern of this gene.
enh3.4 Contains Two Separate EnhancerSequences Involved in Early Embryonic- andComma-Stage Expression
Being interested in aspects of embryonic ceh-13 expres-sion, we have further dissected enh3.4. We found thatremoval of the first 2645 bp of enh3.4 had no effect on theonset of GFP expression in the ABxxxp and Ep cells. Indeed,the remaining terminal 740 bp (enh740), when cloned intopPD107.94, were sufficient to initiate the correct earlyembryonic expression pattern (Figs. 2I and 2J). During thecomma stage, however, enh740 failed to induce importantaspects of the GFP expression pattern seen with the entireenh3.4. Particularly, GFP expression in embryonic dorsalbody wall muscle cells and cells of the future VNC wasmissing (Figs. 2K and 2L). Thus, the complete bean andcomma-stage expression pattern of enh3.4 must also de-pend on enhancer elements that are located upstream ofenh740. Indeed, in our deletions analyses, we found that themost upstream located 450 bp (enh450) of enh3.4 weresufficient to drive most aspects of the normal ceh-13
© 2002 Elsevier Science (USA
expression pattern in comma-stage embryos. If cloned intothe vectors pPD107.94 or pMF1DH3 (pMF1 lacking theupstream 1.5-kb fragment, enh3.4, and enh1.2; see Fig. 1),enh450 was able to drive GFP expression in dorsal bodywall muscle cells and cells of the future VNC (Figs. 2O, 2P,and 3A). Interestingly, the strong endodermal expressionobserved with enh740 (Figs. 2K and 2L) was not induced bythese constructs, suggesting that enh740 and enh450 areresponsible for different aspects of ceh-13 expression duringthe comma stage. The most important difference betweenenh740 and enh450, however, is that early embryonic GFPexpression induced by enh740 Figs. 2I and 2J), could not bedetected with the two enh450-containing constructs (Figs.2M and 2N; and results not shown). Altogether, our datasuggest that enh3.4 contains at least two different enhancerelements capable of driving different aspects of earlyembryonic- and comma-stage expression of ceh-13.
ceh-13 Comma-Stage Expression Depends on aDirect Autoregulatory Loop
In Drosophila and mouse, correct labial/Hoxb1 expres-sion at some stages of development relies on autoregulationthat involves cooperative binding of LAB/HOXB-1 andEXD/PBX to a specific autoregulatory DNA element (Chanet al., 1996, 1997; Grieder et al., 1997; Popperl et al., 1995;for a review see Mann and Affolter, 1998). To test whetherceh-13 expression in C. elegans also depends on autoregu-lation, we crossed the integrated pMF1 into a ceh-13 (sw1)mutant background. While early embryonic CEH-13::GFPexpression appeared to be normal (Figs. 3A–3D), we foundthat comma-stage expression in mutant embryos wasgreatly reduced (Figs. 3G and 3H) in comparison with thenormal levels observed in wild-type animals (Figs. 3E and3F). Particularly, GFP expression was absent from dorsalbody wall muscle cells and from the cells of the prospectiveVNC (Figs. 3G and 3H). These results strongly argued for apositive autoregulatory feedback loop that regulates ceh-13expression during the comma stage of C. elegans develop-ment.
Comparative sequence analysis of pMF1 led to the iden-tification of a 10-bp-long sequence motif (TGATGGATGG)in enh450 (starting at nucleotide position 26483 relative tothe ATG start codon of ceh-13) that is identical to that ofthe HOXB1/PBX autoregulatory element of the mouse geneHoxb1 (Chan et al., 1996, 1997; Popperl et al., 1995) anddiffers by only one base pair substitution from that of theLAB/EXD autoregulatory element of the Drosophila genelabial (Grieder et al., 1997). In order to assess the impor-tance of the 10-bp element and to test whether this se-quence represents an autoregulatory element similar to theone described for the mouse and the fly, we performed amutational analysis in the context of the enh450. In Dro-sophila and in mice, it has been shown that the activity ofthe element is sensitive to point mutations in the corebinding sites of the LAB(HOXB1) and EXD(PBX) proteins(Grieder et al., 1997; Popperl et al., 1995). In analogy tothese findings, we introduced two separate 1-bp substitu-
). All rights reserved.
102 Streit et al.
© 2002 Elsevier Science (USA
tions in the 10-bp worm element that were expected todisrupt the potential binding sites for CEH-13 and for aputative worm ortholog of EXD/PBX (see below) (Fig. 1,mutL and mutE). In the double-mutant construct mutE/L,the point mutations in the two half sites of the elementwere combined (Fig. 1).
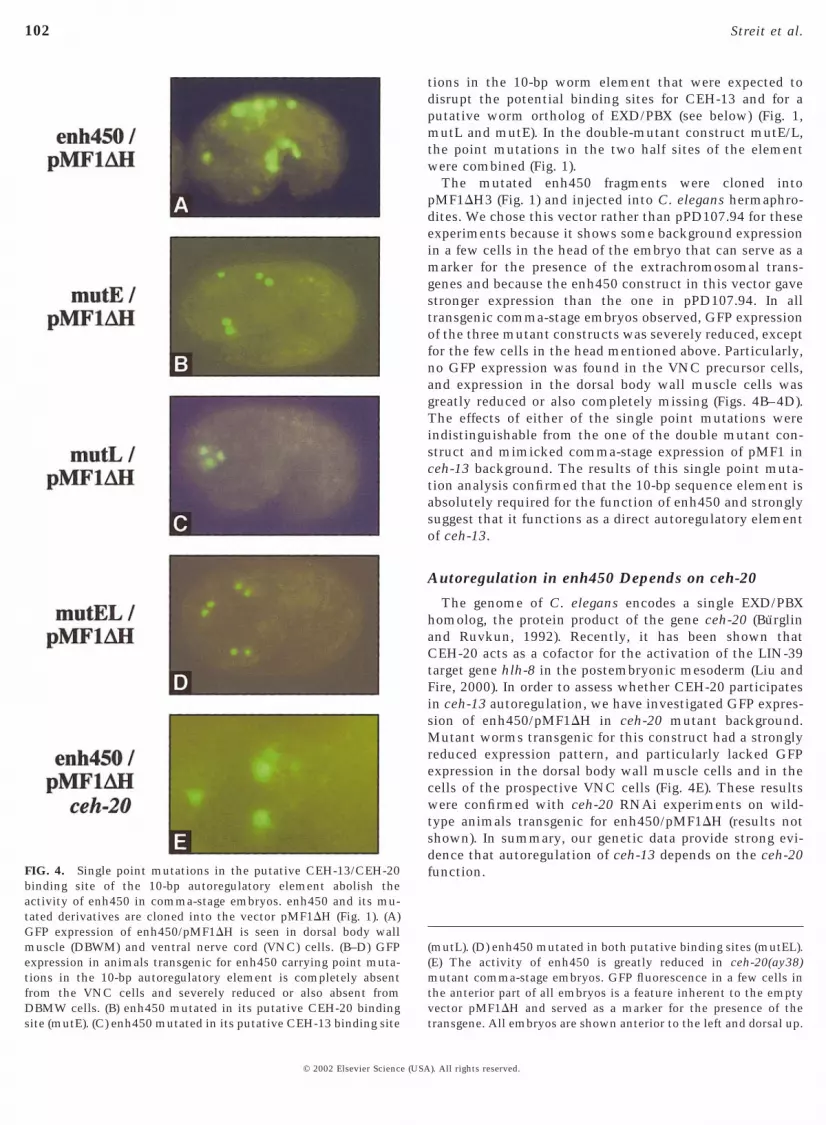
The mutated enh450 fragments were cloned intopMF1DH3 (Fig. 1) and injected into C. elegans hermaphro-dites. We chose this vector rather than pPD107.94 for theseexperiments because it shows some background expressionin a few cells in the head of the embryo that can serve as amarker for the presence of the extrachromosomal trans-genes and because the enh450 construct in this vector gavestronger expression than the one in pPD107.94. In alltransgenic comma-stage embryos observed, GFP expressionof the three mutant constructs was severely reduced, exceptfor the few cells in the head mentioned above. Particularly,no GFP expression was found in the VNC precursor cells,and expression in the dorsal body wall muscle cells wasgreatly reduced or also completely missing (Figs. 4B–4D).The effects of either of the single point mutations wereindistinguishable from the one of the double mutant con-struct and mimicked comma-stage expression of pMF1 inceh-13 background. The results of this single point muta-tion analysis confirmed that the 10-bp sequence element isabsolutely required for the function of enh450 and stronglysuggest that it functions as a direct autoregulatory elementof ceh-13.
Autoregulation in enh450 Depends on ceh-20
The genome of C. elegans encodes a single EXD/PBXhomolog, the protein product of the gene ceh-20 (Burglinand Ruvkun, 1992). Recently, it has been shown thatCEH-20 acts as a cofactor for the activation of the LIN-39target gene hlh-8 in the postembryonic mesoderm (Liu andFire, 2000). In order to assess whether CEH-20 participatesin ceh-13 autoregulation, we have investigated GFP expres-sion of enh450/pMF1DH in ceh-20 mutant background.Mutant worms transgenic for this construct had a stronglyreduced expression pattern, and particularly lacked GFPexpression in the dorsal body wall muscle cells and in thecells of the prospective VNC cells (Fig. 4E). These resultswere confirmed with ceh-20 RNAi experiments on wild-type animals transgenic for enh450/pMF1DH (results notshown). In summary, our genetic data provide strong evi-dence that autoregulation of ceh-13 depends on the ceh-20function.
(mutL). (D) enh450 mutated in both putative binding sites (mutEL).(E) The activity of enh450 is greatly reduced in ceh-20(ay38)mutant comma-stage embryos. GFP fluorescence in a few cells inthe anterior part of all embryos is a feature inherent to the emptyvector pMF1DH and served as a marker for the presence of thetransgene. All embryos are shown anterior to the left and dorsal up.
FIG. 4. Single point mutations in the putative CEH-13/CEH-20binding site of the 10-bp autoregulatory element abolish theactivity of enh450 in comma-stage embryos. enh450 and its mu-tated derivatives are cloned into the vector pMF1DH (Fig. 1). (A)GFP expression of enh450/pMF1DH is seen in dorsal body wallmuscle (DBWM) and ventral nerve cord (VNC) cells. (B–D) GFPexpression in animals transgenic for enh450 carrying point muta-tions in the 10-bp autoregulatory element is completely absentfrom the VNC cells and severely reduced or also absent fromDBMW cells. (B) enh450 mutated in its putative CEH-20 bindingsite (mutE). (C) enh450 mutated in its putative CEH-13 binding site
). All rights reserved.
FIG
.5.
Ace
h-1
3re
gula
tory
elem
ent
indu
ces
WG
-dep
ende
nt
expr
essi
onin
Dro
soph
ila
embr
yos.
Exp
ress
ion
ofC
.ele
gan
sen
han
cers
anal
yzed
inla
cZre
port
erge
ne
assa
ysin
stag
e15
tran
sgen
icD
roso
phil
aem
bryo
s(A
–C).
Exp
ress
ion
was
dete
cted
wit
hen
h3.
4(A
),bo
thin
the
endo
derm
(arr
owh
ead)
and
inec
tode
rmal
stri
pes
(ou
tof
focu
s;se
eD
and
E).
Sim
ilar
expr
essi
onw
asse
enw
ith
enh
740
(C)
but
not
wit
hen
h45
0(B
).L
acZ
expr
essi
onof
enh
740
inth
eec
tode
rmof
stag
e13
embr
yos
was
anal
yzed
byu
sin
gco
nfo
cal
mic
rosc
opy
(D)a
nd
com
pare
dto
the
expr
essi
onof
WG
prot
ein
inth
esa
me
embr
yo(E
).C
lear
ly,a
ctiv
atio
nof
enh
740
isco
nfi
ned
toth
eW
Gex
pres
sion
dom
ain
(F).
Exp
ress
ion
ofen
h74
0w
asab
olis
hed
inw
gm
uta
nts
;on
lysi
ngl
ece
lls
expr
ess
the
tran
sgen
e(G
;ar
row
).In
pan
mu
tan
tem
bryo
s,ex
pres
sion
ofen
h74
0is
not
dete
cted
anym
ore
inth
een
dode
rm(H
).E
ctod
erm
alex
pres
sion
,w
hic
hst
arts
mu
chea
rlie
rth
anen
dode
rmal
expr
essi
on,
albe
itre
duce
d,w
asst
ill
dete
ctab
le.
Los
sof
endo
derm
alex
pres
sion
was
also
seen
upo
ndr
ivin
ga
dom
inan
t-n
egat
ive
vers
ion
ofP
an(d
TC
FDN
)wit
han
endo
derm
-spe
cifi
cdr
iver
lin
e,48
Y-G
AL
4(I
).E
nh
740
resp
onds
toec
topi
cW
Gsi
gnal
ing
inal
lge
rmla
yers
(J–L
).E
xpre
ssio
nof
anac
tiva
ted
vers
ion
ofar
m(a
rmS1
0)in
the
ecto
derm
usi
ng
apr
d-G
AL
4dr
iver
lin
ele
ads
toec
topi
cex
pres
sion
ofen
h74
0in
alte
rnat
ese
gmen
tal
stri
pes
(J).
Ect
opic
expr
essi
onof
arm
S10
inth
em
esod
erm
usi
ng
24B
-GA
L4
lead
sto
the
acti
vati
onof
enh
740
inm
ost
ofth
ece
lls
ofth
evi
scer
alm
esod
erm
(K).
Exp
ress
ion
ofar
mS1
0in
the
endo
derm
wit
h48
Y-G
AL
4re
sult
sin
stro
ng
and
ecto
pic
acti
vati
onof
enh
740
inth
ean
teri
oren
dode
rm(L
).Fo
rre
ason
sw
edo
not
un
ders
tan
d,th
een
han
cer
can
not
beac
tiva
ted
byA
rmS1
0in
the
post
erio
rpa
rtof
the
endo
derm
.
103Conserved Inputs Regulate ceh-13 Expression
© 2002 Elsevier Science (USA). All rights reserved.

104 Streit et al.
A ceh-13 Regulatory Region Induces WG-Dependent Expression in the Drosophila Embryo
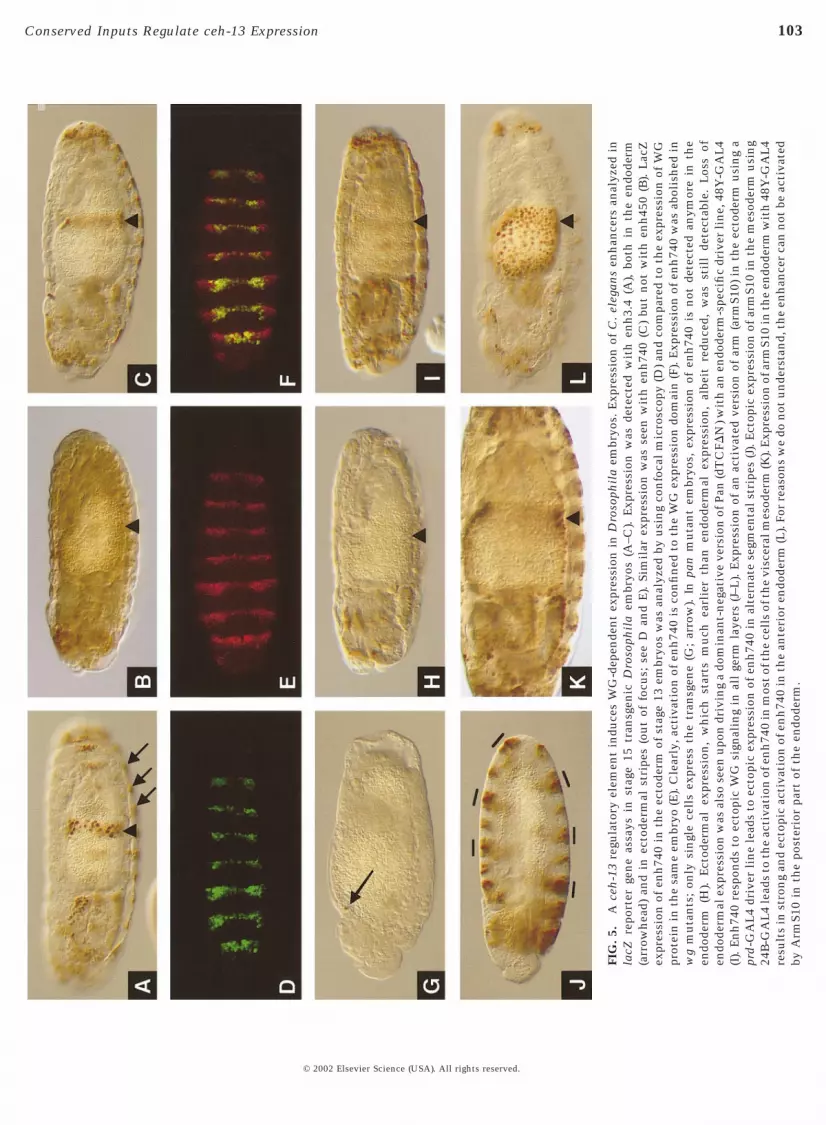
Considering that a plethora of genetic and molecular dataabout lab expression in the embryonic midgut of the fly hasaccumulated (Bienz, 1994, 1997; Grieder et al., 1997; Hop-pler et al., 1995; Ryoo et al., 1999; Szuts et al., 1998), weassayed the activity of the C. elegans enhancer elements inthe fly. In an initial series of experiments, we made fliestransgenic for nuclear LacZ-encoding constructs bearingthree different portions of the ceh-13 enhancer, namelyenh1.2, enh3.4, and a combined 4.6-kb-long fragment(enh4.6; Fig. 1). No reliable expression pattern was detectedwith enh1.2 (data not shown). However, the two largerfragments (enh3.4 and enh4.6) were able to induce a promi-nent domain of LacZ expression around the second midgutconstriction in early stage 15 embryos (shown only forenh3.4 in Fig. 5A, arrowhead), in a region in which endog-enous lab is expressed. However, colocalization studiesusing antibodies against LacZ and LAB showed that thedomain of lacZ expression driven by the ceh-13 cis-regulatory regions is shifted a few cells toward posteriorwith respect to the expression of the endogenous, LABprotein (data not shown; see Discussion).
Expression of lab in the embryonic midgut depends on anautoregulatory feedback mechanism and on input from thedecapentaplegic (dpp) and wingless (wg) signaling cascades(for review, see Bienz, 1997). We could not find any indica-
FIG. 6. POP-1 binds to a fragment of enh740 in vitro. (A) Sequencesof the wild-type (wt-pop) and the mutated (mut-pop) oligonucleotidesused for electrophoretic mobility shift assay with POP-1(180–280) protein.wt-pop contains a putative POP-1 binding site indicated in bold. Thenucleotides mutated in the putative POP-1 binding site of mut-popare underlined. (B) Radiolabeled double-stranded oligonucleotide wt-pop (lanes 1–4) was incubated with POP-1(180–280) protein (lanes 2–4) inthe absence (lanes 1 and 2) or presence (lanes 3 and 4) of an excess ofnonlabeled wt-pop (lane 3) or mut-pop (lane 4). For details, see text andMaterials and Methods.
© 2002 Elsevier Science (USA
tion of a direct effect of dpp on the activity of enh3.4. Onthe other hand, expression of enh3.4 was dependent on thepresence of endogenous labial function and on the wgsignaling cascade, indicating that enh3.4 responds to atleast two of the regulatory inputs that control endogenouslabial (data not shown, but see below). To find out which, ifany, of the isolated enhancer elements functional in C.elegans development were active in the Drosophila embry-onic nuclei, we assayed both the autoregulatory elementcontaining enh450 and the early acting element enh740individually in transgenic embryos. While no expressionwas seen with enh450 in five independent transgenic lines(Fig. 5B; see Discussion) we found that expression similar oridentical to enh3.4 was driven by enh740 (Fig. 5C). Inter-estingly, enh740 was also expressed in lab mutant embryosin the same pattern as in wt embryos, except that thenumber of cells expressing enh740 in a lab mutant mayhave been somewhat reduced. This indicated that enh740,in contrast to enh3.4, was not dependent on endogenous lab(data not shown).
In addition to the endodermal expression, enh3.4 andenh740 also directed expression to a number of cell groupsin other germ layers, most prominently in stripes of ventralepidermal cells (see arrows in Fig. 5A, and below). In fact,LacZ expression of enh3.4 and enh740 was highly reminis-cent of the distribution of the Wingless (WG) signalingmolecule during Drosophila development. Therefore, weconcomitantly analyzed the temporal and spatial distribu-tion of WG and LacZ in transgenic embryos. As shown inFigs. 5D–5F, we observed a virtual overlap between the twoexpression domains; this was obvious not only for thestriped ectodermal expression but also for all the otherexpression patterns driven by enh740, including the devel-oping midgut (data not shown). LacZ expression was exclu-sively seen in nuclei surrounded by extracellular accumu-lation of WG protein; nuclei elsewhere in the embryo didnot accumulate LacZ. The LacZ staining was somewhatvariable from embryo to embryo so that not always allnuclei showed the same strong staining.
To determine whether the WG pathway was indeedinvolved in the activation of the ceh-13 enhancer elementin the Drosophila embryo, we crossed the enh740 con-structs into various genetic backgrounds that modulate WGsignaling. In a wg null mutant embryo, virtually all expres-sion observed in the presence of the enh740 construct inwild-type embryos was missing and only very few cellsretained the LacZ protein (Fig. 5G). In a mutant defective inthe pangolin (pan) gene, which encodes the Drosophilahomolog of vertebrate LEF1 the transcription factor thatmediates WG signaling (Brunner et al., 1997; van de Weter-ing et al., 1997), ectodermal expression was not abolished(presumably due to perdurance of the maternally contrib-uted pan activity), but expression in the gut was absent (Fig.5H). The same phenotype was observed when a dominant-negative version of the PAN protein (van de Wetering et al.,1997) was expressed in the endoderm of wild-type embryos,bearing the enh740 reporter construct (Fig. 5I). Altogether,
). All rights reserved.

105Conserved Inputs Regulate ceh-13 Expression
these results clearly demonstrate that enh740 expression isdependent on WG signaling and its nuclear effector PAN.
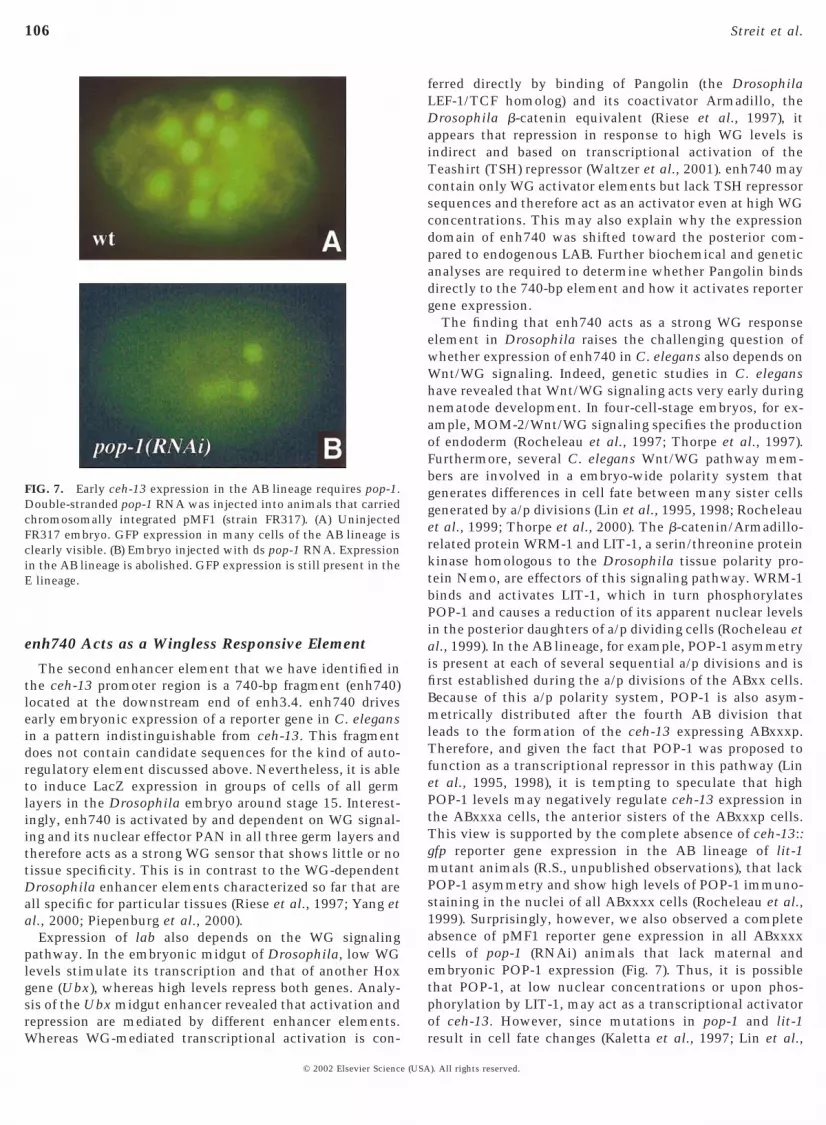
To assess whether WG signaling was not only requiredbut also sufficient to activate transcription via the ceh-13regulatory sequences, we expressed an activated version ofthe WG pathway component Armadillo (ArmS10; van deWetering et al., 1997) in each of the three germ layers andanalyzed whether the corresponding expression domainexpanded or not (see Materials and Methods for details). Inall three germ layers, we found a significant expansion ofthe expression pattern; ectodermal stripes expressingArmS10 (see Materials and Methods) were considerablybroader (Fig. 5J), and expression was seen throughout mostof the visceral mesoderm (Fig. 5K) and the endoderm (Fig.5L). We conclude that the enh740 ceh-13 element harbors astrong WG-responsive element, which responds to signal-ing in all germ layers of the fly embryo. These results raisedthe interesting possibility that the 740-bp element mightalso be controlled by wg signaling in the early C. elegansembryo. Consistent with such a model, we found thatexpression of CEH-13::GFP was severely reduced in em-bryos with a defective WNT signaling cascade. We per-formed RNA interference (RNAi) experiments with pop-1,the worm pangolin ortholog (Fig. 7) (Lin et al., 1995).Double-stranded pop-1 RNA was injected into the gonads ofworms that carried pMF1 (Fig. 1) as chromosomally inte-grated transgene (strain FR317), and their progeny wereanalyzed. In these pop-1(RNAi) embryos (Fig. 7B), expres-sion from pMF1 was absent from all cells of the AB celllineage (Fig. 7A). Expression in the E lineage was stillpresent, although it appeared to be somewhat delayed andprobably quantitatively reduced.
While these results show that pop-1 is required for properceh-13 expression, it is still possible that the effect is highlyindirect, for example, due to cell fate changes that happenearlier in development (see Discussion). An argument for adirect involvement of POP-1 in the control of ceh-13 isshown in Fig. 6. Sequence inspection of enh740 showed thatit contains several sites that resemble the one that has beenshown to bind POP-1180–280 efficiently (Korswagen et al.,2000). POP-1180–280 is a 101-amino-acid fragment of POP-1that contains the HMG DNA binding domain (Korswagenet al., 2000). We choose one of these putative binding sites[CTTTTGATG contained in the double-stranded oligonu-cleotide wt-pop (Fig. 6A) that corresponds to position 3443to 3476 upstream of the ceh-13 translation start codon] andtested whether it binds POP-1180–280 in vitro (Fig. 6B). Radio-labeled wt-pop formed a complex with POP-1180–280 (Fig. 6B,lane 2) This complex could be competed for with unlabeledwt-pop (Fig. 6B, lane 3) but not with mut-pop (Fig. 6B, lane4), an oligo identical with wt-pop except for a 3-bp substi-tution in the putative core binding site (Fig. 6A), indicatingthat POP-1180–280 binds specifically to this sequence. Whenmut-pop was used as a labeled probe, no complex wasformed but the oligo ran at the expected position in the gel,indicating that it was intact (data not shown).
© 2002 Elsevier Science (USA
DISCUSSION
Here, we show that a 3.4-kb enhancer fragment of the C.elegans labial homolog ceh-13 (enh3.4), located 3.2–6.6 kbupstream of the translational start site, is capable of drivingembryonic ceh-13 expression. Dissection of enh3.4 allowedthe identification of two cis-regulatory elements, enh740and enh450, that are able to induce different aspects ofembryonic ceh-13 expression.
Comma-Stage Expression of enh450 Is Controlledby a Highly Conserved Autoregulatory Loop
We show that comma-stage expression of pMF1 dependson the presence of a functional ceh-13 gene. Furthermore,we found that enh450, a subfragment of pMF1 that issufficient to drive ceh-13-like comma-stage expression of areporter, contains a 10-bp-long DNA element whose se-quence, TGATGGATGG, is identical to that of the mouseHoxb1 autoregulatory element (Popperl et al., 1995) anddiffers in only one position from that of the correspondingelement in the midgut enhancer region of the Drosophilagene labial (Grieder et al., 1997). Single point mutations inthe C. elegans element at positions that have been shown tobe crucial for cooperative LAB/EXD binding in the fly andin the mouse elements (Chan et al., 1997; Grieder et al.,1997) resulted in a strong reduction or complete loss of theenh450 activity. Furthermore, we have shown that enh450activity also depends on the presence of CEH-20, the onlyEXD/PBX ortholog encoded by the genome of C. elegans(Burglin and Ruvkun, 1992; Burglin, 1997; Liu and Fire,2000). Taken together, our data strongly suggest that con-trol of ceh-13 involves a positive autoregulatory feedbackmechanism that is similar to the one found in Drosophilalab and mammalian hoxb1 and that enh450 contains anelement that participates in this process. LAB/EXD bindingto the Drosophila lab autoregulatory element involves athird protein, Homothorax (HTH) (Ryoo et al., 1999). Thegenome of C. elegans encodes a single HTH ortholog(ceh-25; Burglin, 1997), but at present it is not knownwhether it participates in the regulation of ceh-13 expres-sion.
The full-length enh3.4 appears to be under the control ofthe lab autoregulatory loop in Drosophila, since its expres-sion was dependent on lab. The fact that enh450 on its ownwas not able to drive reporter gene expression in Drosophilais not really surprising. The mouse Hoxb1 autoregulatoryelement was only active in Drosophila when present inmultiple copies (Chan et al., 1996; Popperl et al., 1995).Similarly, multiple copies of the CEH-13/CEH-20 bindingsites might be necessary to activate reporter gene expres-sion in Drosophila. Alternatively, the CEH-13/CEH-20binding site might require additional synergistic inputs forwhich only the full-length enh3.4, but not enh450, offersthe sequences necessary.
). All rights reserved.
106 Streit et al.
enh740 Acts as a Wingless Responsive Element
The second enhancer element that we have identified inthe ceh-13 promoter region is a 740-bp fragment (enh740)located at the downstream end of enh3.4. enh740 drivesearly embryonic expression of a reporter gene in C. elegansin a pattern indistinguishable from ceh-13. This fragmentdoes not contain candidate sequences for the kind of auto-regulatory element discussed above. Nevertheless, it is ableto induce LacZ expression in groups of cells of all germlayers in the Drosophila embryo around stage 15. Interest-ingly, enh740 is activated by and dependent on WG signal-ing and its nuclear effector PAN in all three germ layers andtherefore acts as a strong WG sensor that shows little or notissue specificity. This is in contrast to the WG-dependentDrosophila enhancer elements characterized so far that areall specific for particular tissues (Riese et al., 1997; Yang etal., 2000; Piepenburg et al., 2000).
Expression of lab also depends on the WG signalingpathway. In the embryonic midgut of Drosophila, low WGlevels stimulate its transcription and that of another Hoxgene (Ubx), whereas high levels repress both genes. Analy-sis of the Ubx midgut enhancer revealed that activation andrepression are mediated by different enhancer elements.Whereas WG-mediated transcriptional activation is con-
FIG. 7. Early ceh-13 expression in the AB lineage requires pop-1.Double-stranded pop-1 RNA was injected into animals that carriedchromosomally integrated pMF1 (strain FR317). (A) UninjectedFR317 embryo. GFP expression in many cells of the AB lineage isclearly visible. (B) Embryo injected with ds pop-1 RNA. Expressionin the AB lineage is abolished. GFP expression is still present in theE lineage.
© 2002 Elsevier Science (USA
ferred directly by binding of Pangolin (the DrosophilaLEF-1/TCF homolog) and its coactivator Armadillo, theDrosophila b-catenin equivalent (Riese et al., 1997), itappears that repression in response to high WG levels isindirect and based on transcriptional activation of theTeashirt (TSH) repressor (Waltzer et al., 2001). enh740 maycontain only WG activator elements but lack TSH repressorsequences and therefore act as an activator even at high WGconcentrations. This may also explain why the expressiondomain of enh740 was shifted toward the posterior com-pared to endogenous LAB. Further biochemical and geneticanalyses are required to determine whether Pangolin bindsdirectly to the 740-bp element and how it activates reportergene expression.
The finding that enh740 acts as a strong WG responseelement in Drosophila raises the challenging question ofwhether expression of enh740 in C. elegans also depends onWnt/WG signaling. Indeed, genetic studies in C. eleganshave revealed that Wnt/WG signaling acts very early duringnematode development. In four-cell-stage embryos, for ex-ample, MOM-2/Wnt/WG signaling specifies the productionof endoderm (Rocheleau et al., 1997; Thorpe et al., 1997).Furthermore, several C. elegans Wnt/WG pathway mem-bers are involved in a embryo-wide polarity system thatgenerates differences in cell fate between many sister cellsgenerated by a/p divisions (Lin et al., 1995, 1998; Rocheleauet al., 1999; Thorpe et al., 2000). The b-catenin/Armadillo-related protein WRM-1 and LIT-1, a serin/threonine proteinkinase homologous to the Drosophila tissue polarity pro-tein Nemo, are effectors of this signaling pathway. WRM-1binds and activates LIT-1, which in turn phosphorylatesPOP-1 and causes a reduction of its apparent nuclear levelsin the posterior daughters of a/p dividing cells (Rocheleau etal., 1999). In the AB lineage, for example, POP-1 asymmetryis present at each of several sequential a/p divisions and isfirst established during the a/p divisions of the ABxx cells.Because of this a/p polarity system, POP-1 is also asym-metrically distributed after the fourth AB division thatleads to the formation of the ceh-13 expressing ABxxxp.Therefore, and given the fact that POP-1 was proposed tofunction as a transcriptional repressor in this pathway (Linet al., 1995, 1998), it is tempting to speculate that highPOP-1 levels may negatively regulate ceh-13 expression inthe ABxxxa cells, the anterior sisters of the ABxxxp cells.This view is supported by the complete absence of ceh-13::gfp reporter gene expression in the AB lineage of lit-1mutant animals (R.S., unpublished observations), that lackPOP-1 asymmetry and show high levels of POP-1 immuno-staining in the nuclei of all ABxxxx cells (Rocheleau et al.,1999). Surprisingly, however, we also observed a completeabsence of pMF1 reporter gene expression in all ABxxxxcells of pop-1 (RNAi) animals that lack maternal andembryonic POP-1 expression (Fig. 7). Thus, it is possiblethat POP-1, at low nuclear concentrations or upon phos-phorylation by LIT-1, may act as a transcriptional activatorof ceh-13. However, since mutations in pop-1 and lit-1result in cell fate changes (Kaletta et al., 1997; Lin et al.,
). All rights reserved.

107Conserved Inputs Regulate ceh-13 Expression
1998) and start to act earlier in development than ceh-13 isfirst expressed, other explanations for these results can notbe excluded. An argument for a direct involvement ofPOP-1 in control of ceh-13 is the fact that enh740 doescontain several putative POP-1 binding sites and at leastone of them does bind POP-1 in vitro (Fig. 6). Altogether, wethink that WNT signaling, or more specifically POP-1, isvery likely to be one of the spatiotemporal cues involved incontrol of ceh-13 expression in the early embryo. However,early embryonic ceh-13/enh740 expression cannot dependon WG mediated polarity cues only: POP-1 asymmetry inthe AB lineage is established already in the daughters of theABxx cells, but onset of ceh-13 expression occurs only onecell division later. Regardless of the POP-1 distribution,ceh-13 continues to be expressed in both the anterior andposterior daughters of almost all of the ABxxxp cells (Witt-mann et al., 1997). Furthermore, ceh-13 is not expressed inthe cells of the early C lineage. Thus, we propose that, inaddition to Wnt/WG-mediated polarity cues, other yetunidentified factors participate in the regulation of ceh-13/enh740 early expression and provide the cells with tempo-ral and lineage-specific inputs. Further work will be re-quired to confirm the involvement of the worm Wnt/WGpathway and its nuclear effector POP-1 on enh740 and toidentify other regulatory proteins that bind to it.
Earlier experiments have suggested that the pattern ofexpression of the worm Hox genes are determined, at leastin part, by mechanisms independent of the global positionof the cells along the a/p axis (Cowing and Kenyon, 1996;Wittmann et al., 1997). This is in contrast to Drosophilaand vertebrates, where the Hox genes are controlled byglobal positional mechanisms. Based on these observations,it has been suggested that Hox gene regulation in C. elegansmay rely on different strategies than those previouslycharacterized in Drosophila and mammalians. Neverthe-less, we show here that the molecular nature of the mecha-nisms that control Hox 1 class genes may be more con-served between flies and worms than previously assumed.
ACKNOWLEDGMENTS
We thank J. Priess for providing the pop-1 cDNA, A. Fire for thevector kits, H. Korswagen for pET-21b-popHM6, and Steve Cohen foranti-WG antibodies. Some C. elegans strains were supplied by theCaenorhabditis Genetics Center, which is funded by the NationalInstitutes of Health (NIH) National Center for Research Resources.We are in debt to A. Puoti, P. Pugnale, T. Burglin, G. Cassata, T.Vellai, W. B. Wood, and Y. Suzuki for their help and scientificadvice. We are also grateful to L. Buillard, H. Gachoud, and Y.Molleyres and C. Folly for excellent technical help. This researchwas supported by Swiss National Science Foundation (SNSF)Grants 31-001.91, 31-40776.94, 31-539829.98, by the EU TMRGrant FMRXCT980217, and the Kantons Basel-Stadt and Basel-Land and Fribourg.
© 2002 Elsevier Science (USA
REFERENCES
Baker, N. E. (1988). Transcription of the segment-polarity genewingless in the imaginal discs of Drosophila, and the phenotypeof a pupal-lethal wg mutation. Development 102, 489–497.
Bienz, M. (1994). Homeotic genes and positional signaling in theDrosophila viscera. Trends Genet. 10, 22–26.
Bienz, M. (1997). Endoderm induction in Drosophila: The nucleartargets of the inducing signals. Curr. Opin. Genet. Dev. 7,683–688.
Brand, A. H., and Perrimon, N. (1993). Targeted gene expression asa means of altering cell fates and generating dominant pheno-types. Development 118, 401–415.
Brenner, S. (1974). The genetics of Caenorhabditis elegans. Genet-ics 77, 71–94.
Brunner, E., Peter, O., Schweizer, L., and Basler, K. (1997). pangolinencodes a Lef-1 homologue that acts downstream of Armadillo totransduce the Wingless signal in Drosophila. Nature 385, 829–833.
Brunschwig, K., Wittmann, C., Schnabel, R., Burglin, T. R., Tobler,H., and Muller, F. (1999). Anterior organization of the Caeno-rhabditis elegans embryo by the labial-like Hox gene ceh-13.Development 126, 1537–1546.
Burglin, T. R., Finney, M., Coulson, A., and Ruvkun, G. (1989).Caenorhabditis elegans has scores of homeobox-containinggenes. Nature 341, 239–243.
Burglin, T. R., Ruvkun, G., Coulson, A., Hawkins, N. C., McGhee,J. D., Schaller, D., Wittmann, C., Muller, F., and Waterston, R. H.(1991). Nematode homeobox cluster. Nature 351, 703.
Burglin, T. R., and Ruvkun, G. (1992). New motif in PBX genes.Nat. Genet. 1, 319–320.
Burglin, T. R., and Ruvkun, G. (1993). The Caenorhabditis eleganshomeobox gene cluster. Curr. Opin. Genet. Dev. 3, 615–620.
Burglin, T. R. (1997). Analysis of TALE superclass homeobox genes(MEIS, PBC, KNOX, Iroquois, TGIF) reveals a novel domainconserved between plants and animals. Nucleic Acids Res. 25,4173–4180.
Chan, S. K., Popperl, H., Krumlauf, R., and Mann, R. S. (1996). Anextradenticle-induced conformational change in a HOX proteinovercomes an inhibitory function of the conserved hexapeptidemotif. EMBO J. 15, 2476–2487.
Chan, S. K., Ryoo, H. D., Gould, A., Krumlauf, R., and Mann, R. S.(1997). Switching the in vivo specificity of a minimal Hox-responsive element. Development 124, 2007–2014.
Cowing, D., and Kenyon, C. (1996). Correct Hox gene expressionestablished independently of position in Caenorhabditis elegans.Nature 382, 353–356.
Duboule, D. (1994). Temporal colinearity and the phylotypic pro-gression: a basis for the stability of a vertebrate Bauplan and theevolution of morphologies through heterochrony. Dev. Suppl.,135–142.
Duboule, D., and Morata, G. (1994). Colinearity and functionalhierarchy among genes of the homeotic complexes. TrendsGenet. 10, 358–364.
Fire, A., Harrison, S. W., and Dixon, D. (1990a). A modular set oflacZ fusion vectors for studying gene expression in Caenorhab-ditis elegans. Gene 93, 189–198.
Fire, A., Kondo, K., and Waterston, R. (1990b). Vectors for low copytransformation of C. elegans. Nucleic Acids Res. 18, 4269–4270.
Gehring, W. J., Affolter, M., and Burglin, T. (1994). Homeodomainproteins. Annu. Rev. Biochem. 63, 487–526.
). All rights reserved.

108 Streit et al.
Grieder, N. C., Marty, T., Ryoo, H. D., Mann, R. S., and Affolter, M.(1997). Synergistic activation of a Drosophila enhancer by HOM/EXD and DPP signaling. EMBO J. 16, 7402–7410.
Hoppler, S., and Bienz, M. (1995). Two different thresholds ofwingless signaling with distinct developmental consequences inthe Drosophila midgut. EMBO J. 14, 5016–5026.
Immergluck, K., Lawrence, P. A., and Bienz, M. (1990). Inductionacross germ layers in Drosophila mediated by a genetic cascade.Cell 62, 261–268.
Jin., Y. (1999). Transformation. In “C. elegans. A Practical Ap-proach” (I. A. Hope, Ed.), pp.69–96. Oxford Univ. Press, NewYork.
Kaletta, T., Schnabel, H., and Schnabel, R. (1997). Binary specifica-tion of the embryonic lineage in Caenorhabditis elegans. Nature390, 294–298.
Kenyon, C. J., Austin, J., Costa, M., Cowing, D. W., Harris, J. M.,Honigberg, L., Hunter, C. P., Maloof, J. N., Muller-Immergluck,M. M., Salser, S. J., et al. (1997). The dance of the Hox genes:Patterning the anteroposterior body axis of Caenorhabditis el-egans. Cold Spring Harbor Symp. Quant. Biol. 62, 293–305.
Korswagen, H. C., Herman, M. A., and Clevers, H. C. (2000).Distinct b-catenins mediate adhesion and signaling functions inC. elegans. Nature 406, 527–531.
Krumlauf, R. (1992). Evolution of the vertebrate Hox homeoboxgenes. BioEssays 14, 245–252.
Lin, R., Thompson, S., and Priess, J. R. (1995). pop-1 encodes anHMG box protein required for the specification of a mesodermprecursor in early C. elegans embryos. Cell 83, 599–609.
Lin, R., Hill, R. J., and Priess, J. R. (1998). POP-1 and anterior–posterior fate decisions in C. elegans embryos. Cell 92, 229–239.
Liu, J., and Fire, A. (2000). Overlapping roles of two Hox genes andthe exd ortholog ceh-20 in diversification of the C. eleganspostembryonic mesoderm. Development 127, 5179–5190.
Mann, R. S., and Affolter, M. (1998). Hox proteins meet morepartners. Curr. Opin. Genet. Dev. 8, 423–429.
Martin-Bermudo, M. D., Dunin-Borkowski, O. M., and Brown,N. H. (1997). Specificity of PS integrin function during embryo-genesis resides in the alpha subunit extracellular domain. EMBOJ. 16, 4184–4193.
Marty, T., Vigano, M. A., Ribeiro, C., Nussbaumer, U., Grieder,N. C., and Affolter, M. (2001). A HOX complex, a repressorelement and a 50 bp sequence confer regional specificity to aDPP-responsive enhancer. Development 128, 2833–2845.
McGinnis, W., and Krummlauf, R. (1992). Homeobox genes andaxial patterning. Cell 68, 283–302.
Mello, C. C., Kramer, J. M., Stinchcomb, D., and Ambros, V. (1991).Efficient gene transfer in C. elegans: Extrachromosomal mainte-nance and integration of transforming sequences. EMBO J. 10,3959–3970.
Piepenburg, O., Vorbruggen, G., and Jackle, H. (2000). Drosophilasegment borders result from unilateral repression of hedgehogactivity by wingless signaling. Mol. Cell 6, 203–209.
Popperl, H., Bienz, M., Studer, M., Chan, S. K., Aparicio, S.,Brenner, S., Mann, R. S., and Krumlauf, R. (1995). Segmentalexpression of Hoxb-1 is controlled by a highly conserved auto-regulatory loop dependent upon exd/pbx. Cell 81, 1031–1042.
Riese, J., Yu, X., Munnerlyn, A., Eresh, S., Hsu, S. C., Grosschedl,R., and Bienz, M. (1997). LEF-1, a nuclear factor coordinatingsignaling inputs from wingless and decapentaplegic. Cell 88,777–787.
Rocheleau, C. E., Downs, W. D., Lin, R., Wittmann, C., Bei, Y.,Cha, Y. H., Ali, M., Priess, J. R., and Mello, C. C. (1997). Wnt
© 2002 Elsevier Science (USA
signaling and an APC-related gene specify endoderm in early C.elegans embryos. Cell 90, 707–716.
Rocheleau, C. E., Yasuda, J., Shin, T. H., Lin, R., Sawa, H., Okano,H., Priess, J. R., Davis, R. J., and Mello, C. C. (1999). WRM-1activates the LIT-1 protein kinase to transduce anterior/posteriorpolarity signals in C. elegans. Cell 97, 717–726.
Ruvkun, G., and Hobert, O. (1998). The taxonomy of developmen-tal control in Caenorhabditis elegans. Science 282, 2033–2041.
Ryoo, H. D., Marty, T., Casares, F., Affolter, M., and Mann, R. S.(1999). Regulation of Hox target genes by a DNA boundHomothorax/Hox/Extradenticle complex. Development 126,5137–5148.
Sambrooke, J., Fritsch, E. F., and Maniatis, T. (1989). “MolecularCloning: A Laboratory Manual, Second Edition.” Cold SpringHarbor Laboratory Press, Cold Spring Harbor, NY.
Schnabel, R., and Schnabel, H. (1997). Hox genes misled by localenvironments. Nature 385, 588–589.
Stiernagle, T. (1999). Maintenance of C. elegans. In “C. elegans. APractical Approach” (I. A. Hope, Ed.), pp.51–67. Oxford Univ.Press, New York.
Sulston, J. E., Schierenberg, E., White, J. G., and Thomson, J. N.(1983). The embryonic cell lineage of the nematode Caenorhab-ditis elegans. Dev. Biol. 100, 64–119.
Szuts, D., Eresh, S., and Bienz, M. (1998). Functional intertwiningof Dpp and EGFR signaling during Drosophila endoderm induc-tion. Genes Dev. 12, 2022–2035.
Tremml, G., and Bienz, M. (1992). Induction of labial expression inthe Drosophila endoderm: Response elements for dpp signalingand for autoregulation. Development 116, 447–456.
Thorpe, C. J., Schlesinger, A., Carter, J. C., and Bowerman, B.(1997). Wnt signaling polarizes an early C. elegans blastomere todistinguish endoderm from mesoderm. Cell 90, 695–705.
Thorpe, C. J., Schlesinger, A., and Bowerman, B. (2000). Wntsignaling in Caenorhabditis elegans: Regulating repressors andpolarizing the cytoskeleton. Trends Cell Biol. 10, 10–17.
Waltzer, L., Vandel, L., and Bienz, M. (2001). Teashirt is required fortranscriptional repression mediated by high Wingless levels.EMBO J. 20, 137–145.
Van Auken, K., Weaver, D. C., Edgar, L. G., and Wood, W. B. (2000).Caenorhabditis elegans embryonic axial patterning requires tworecently discovered posterior-group Hox genes. Proc. Natl. Acad.Sci. USA 97, 4499–4503.
Van de Wetering, M., Cavallo, R., Dooijes, D., van Beest, M., vanEs, J., Loureiro, J., Ypma, A., Hursh, D., Jones, T., Bejsovec, A.,Peifer, M., Mortin, M., and Clevers, H. (1997). Armadillo coacti-vates transcription driven by the product of the Drosophilasegment polarity gene dTCF. Cell 88, 789–799.
Wittmann, C., Bossinger, O., Goldstein, B., Fleischmann, M.,Kohler, R., Brunschwig, K., Tobler, H., and Muller, F. (1997). Theexpression of the C. elegans labial-like Hox gene ceh-13 duringearly embryogenesis relies on cell fate and on anteroposterior cellpolarity. Development 124, 4193–4200.
Yang, X., van Beest, M., Clevers, H., Jones, T., Hursh, D. A., andMortin, M. A. (2000). decapentaplegic is a direct target of dTcfrepression in the Drosophila visceral mesoderm. Development127, 3695–3702.
Zecca, M., Basler, K., and Struhl, G. (1996). Direct and long-rangeaction of a wingless morphogen gradient. Cell 87, 833–844.
Received for publication May 25, 2001Revised November 16, 2001
Accepted November 16, 2001Published online January 15, 2002
). All rights reserved.





























