Embed Size (px)
Citation preview

Characterization of a Novel Serotonin Receptor fromCaenorhabditis elegans:Cloning and Expression
of Two Splice Variants
Fadi F. Hamdan, *Mark D. Ungrin, *Mark Abramovitz, and Paula Ribeiro
Institute of Parasitology, McGill University, Ste. Anne de Bellevue; and*Department of Biochemistry and Molecular Biology,Merck Frosst Centre for Therapeutic Research, Pointe Claire-Dorval, Quebec, Canada
Abstract: Serotonin [5-hydroxytryptamine (5-HT)] modu-lates feeding activity, egg-laying, and mating behavior inthe free-living nematode, Caenorhabditis elegans. Wehave cloned a novel receptor cDNA from C. elegans(5-HT2Ce) that has high sequence homology with 5-HT2receptors from other species. When transiently ex-pressed in COS-7 cells, 5-HT2Ce exhibited 5-HT bindingactivity and activated Ca21-mediated signaling in a man-ner analogous to other 5-HT2 receptors. However,5-HT2Ce displayed unusual pharmacological properties,which resembled both 5-HT2 and 5-HT1-like receptorsbut did not correlate well with any of the known 5-HT2subtypes. Two splice variants of 5-HT2Ce that differ by 48N-terminal amino acids were identified. The two isoformswere found to have virtually identical binding and signal-ing properties but differed in their levels of mRNA expres-sion, with the longer variant being four times more abun-dant than the shorter species in all developmental stagestested. Taken together, the results describe two variantsof a novel C. elegans 5-HT receptor, which has some ofthe properties of the 5-HT2 family but whose pharmaco-logical profile does not conform to any known class ofreceptor. Key Words: Caenorhabditis elegans—Seroto-nin—5-HT2 receptor—G protein-coupled receptor—Cloning—Expression—Spliced leader—Aequorin.J. Neurochem. 72, 1372–1383 (1999).
Serotonin [5-hydroxytryptamine (5-HT)] is a widelydistributed neuroactive agent of vertebrates and inverte-brates. In the free-living nematodeCaenorhabditis el-egans,5-HT has been identified within several centraland peripheral neurons, including the well-characterizedpharyngeal neurosecretory motorneurons, the hermaph-rodite-specific neurons, and the male-specific CP neu-rons (Horvitz et al., 1982; Desai et al., 1988; Loer andKenyon, 1993). The serotonergic neurosecretory motorneurons may modulate pharyngeal pumping (feeding),locomotion, and egg laying, whereas the hermaphrodite-specific neurons and CP neurons seem to affect egg-laying and male mating behavior, respectively (Horvitzet al., 1982; Desai and Horvitz, 1989; Loer and Kenyon,1993). These effects of 5-HT are mediated, in part, by G
proteins (Bargmann and Kaplan, 1998) and at least onetype of G protein-coupled receptor (GPCR), which isnegatively linked to adenylate cyclase (Olde and Mc-Combie, 1997). It is unknown at present if there are otherreceptors and pathways of signal transduction that me-diate the multiple effects of 5-HT in this animal.
In mammals, where 5-HT is a well-established neuro-transmitter involved in a wide range of physiologicalactivities (Leonard, 1994), as many as seven differentclasses of 5-HT receptors (5-HT1–5-HT7) have beenidentified, all of which are further divided into multiplesubtypes (Hoyer et al., 1994; Gerhardt and Van Heer-ikhuizen, 1997). The vast majority of mammalian 5-HTreceptors belong to the large GPCR superfamily andcouple to adenylate cyclase, either positively (5-HT4,5-HT6, and 5-HT7) or negatively (5-HT1). In contrast, the5-HT2 family has very distinctive structural propertiesand couples to phospholipase C and the phosphoinositol(inositol trisphosphate)/Ca21-mediated pathway of sig-nal transduction (Hoyer et al., 1994; Gerhardt and VanHeerikhuizen, 1997).
Received October 15, 1998; revised manuscript received November30, 1998; accepted December 7, 1998.
Address correspondence and reprint requests to Dr. P. Ribeiro atInstitute of Parasitology, McGill University, Macdonald Campus, 21,111 Lakeshore Road, Ste. Anne de Bellevue, Quebec, Canada H9X3V9.
Abbreviations used:CeIF, Caenorhabditis eleganseukaryotic initi-ation factor 4A homologue; DOI, 2,5-dimethoxy-4-iodoamphetamine;GPCR, G protein-coupled receptor; 5-HT, 5-hydroxytryptamine (sero-tonin); 5-HTAsc, putative 5-hydroxytryptamine2-like sequence recentlycloned from the parasitic nematodeAscaris suum;5-HT2Ce, 5-hydroxy-tryptamine2 receptor from Caenorhabditis elegans;5-HT2CeL and5-HT2CeS, long and short, respectively, 5-hydroxytryptamine2 receptorisoform from Caenorhabditis elegans;5-HT2Dro, 5-hydroxytrypta-mine2 receptor cloned fromDrosophila; 5-HT2Lym, 5-hydroxytrypta-mine2 receptor cloned from the pond snail,Lymnaea;LSD, lysergicacid diethylamide; 8-OH-DPAT, 8-hydroxy-2-(di-n-propylamino)tetra-lin; ORF, open reading frame; PBS, phosphate-buffered saline; RACE,rapid amplification of cDNA ends; SL, spliced leader; TEM buffer, 50mM Tris (pH 7.4), 0.5 mM EDTA, and 10 mM MgCl2; TM, transmem-brane domain; UTR, untranslated region.
1372
Journal of NeurochemistryLippincott Williams & Wilkins, Inc., Philadelphia© 1999 International Society for Neurochemistry

Considerably less is known about the molecular prop-erties and diversity of 5-HT receptors in invertebrates.Many of the invertebrate receptors that have been clonedand characterized, including the previously describedC.elegans receptor (Olde and McCombie, 1997), showstructural homology with the 5-HT1 and, to a lesserextent, 5-HT7 families (Witz et al., 1990; Saudou et al.,1992; Sugamori et al., 1993; Angers et al., 1998). For themost part, these receptors share similar drug binding andsignaling properties with their mammalian counterparts(Hoyer et al., 1994; Gerhardt and Van Heerikhuizen,1997), except for an unusually low affinity (high micro-molarKD) for indolealkylamines, including 5-HT, whichnormally bind to mammalian 5-HT1 and 5-HT7 receptorswith very high affinity (Hoyer et al., 1994). Recently,two 5-HT2 receptors have been cloned fromDrosophila(5-HT2Dro) (Colas et al., 1995) and the pond snail,Lym-naea(5-HT2Lym) (Gerhardt et al., 1996). Each of thesereceptors, in particular, 5-HT2Dro, shares structural andpharmacological properties with the mammalian 5-HT2Breceptor. 5-HT2Lym has been further characterized asbeing linked to the hydrolysis of phosphoinositides andthe Ca21 signaling pathway (Gerhardt et al., 1996), anindication that 5-HT2 receptors and their distinctive cou-pling mechanism are conserved at least in higher inver-tebrates such as molluscs and insects.
Here we report the molecular cloning and functionalcharacterization of a novel 5-HT receptor fromC. el-egans(5-HT2Ce), which has high sequence homologywith mammalian and other invertebrate 5-HT2 receptors,and also appears to be coupled to a Ca21-mediatedsignaling pathway. However, 5-HT2Ce has mixed 5-HT1and 5-HT2 ligand binding properties, which do not con-form to those of any other 5-HT2 receptor, either mam-malian or invertebrate. This study identified two splicevariants of 5-HT2Ce, which are expressed at differentlevels inC. elegansbut otherwise appear to have similarbinding and signaling properties.
EXPERIMENTAL PROCEDURES
ChemicalsThe radioisotope125I-lysergic acid diethylamide (125I-LSD;
2,200 Ci/mmol) was purchased from Mandel (Guelph, ON, Can-ada). 5-HT, histamine, melatonin,N-acetyl-5-HT, and tryptaminewere obtained from Sigma (Oakville, ON, Canada). Dopamine,octopamine, 2,5-dimethoxy-4-iodoamphetamine (DOI), lisuride,8-hydroxy-2-(di-n-propylamino)tetralin (8-OH-DPAT), ketan-serin, metergoline, butaclamol, methiothepin, and cyproheptadinewere purchased from Research Biochemicals International(Natick, MA, U.S.A.). All other chemicals were of the highestreagent grade.
C. elegansThe wild-type Bristol N2 strain ofC. eleganswas main-
tained at room temperature on 100-mm-diameter NGM agarplates seeded withEscherichia coli(OP50 or NA22). To obtainstage-specific worms,C. elegansnematodes were first synchro-nized and then purified by flotation on a 30% sucrose gradientas described previously (Lewis and Fleming, 1995). Afterpurification, the worms were pelleted by centrifugation and
stored frozen at280°C until needed. For mRNA extraction,aliquots of frozen adult worms were used directly for poly(A)1
RNA purification, using the Fast-Track mRNA isolation kit,according to the recommendations of the manufacturer (In-vitrogen, San Diego, CA, U.S.A.). Total RNA was isolatedfrom different developmental stages with the use of theTRIZOL reagent (GibcoBRL, Burlington, ON, Canada).
Cloning of full-length 5-HT 2Ce cDNAsComputer analyses of data from theC. elegansgenome
sequencing project (Sulston et al., 1992) identified a genomicsequence (cosmid F59C12; GenBank accession no. U41038)that was predicted to encode a protein similar to 5-HT receptorsfrom other species. PCR primers targeting this predicted codingregion (sense, primer F; antisense, primer R; see Fig. 1) wereused in an RT-PCR to amplify a putative 5-HT receptor cDNAsequence. In brief, adultC. elegansmRNA (0.5 mg) wasreverse-transcribed with an oligo(dT) primer and 200 U ofmurine Moloney leukemia virus reverse transcriptase, and1/10th of the resulting cDNA was subjected to 30 cycles ofPCR (30 s at 94°C, 30 s at 54°C, 120 s at 72°C) in a 50-mlreaction mixture containing each primer (F and R) at 0.4mM,according to standard protocols. A PCR product of 1,908 bpwas gel-purified, ligated into the PCR 2.1 vector (Invitrogen),and sequenced by the dideoxy-chain termination method. The59 and 39 ends of the receptor cDNA sequence were obtainedby standard rapid amplification of cDNA ends (RACE) proce-dures using the 59- and 39-RACE systems from GibcoBRL. For59-RACE, adultC. elegansmRNA (0.5 mg/ml) was reverse-transcribed and dC-tailed, before PCR amplification (sameparameters as above) with the kit’s sense oligo(dG)-anchorprimer and a 5-HT2Cegene-specific antisense primer (primer A;see Fig. 1). An aliquot of the resulting PCR product wassubjected to a second PCR procedure using an abridged form ofthe same anchor primer (also supplied by the RACE kit) and anested gene-specific antisense primer (primer B; see Fig. 1).For 39-RACE, RT was performed as described above exceptthat an oligo(dT)-adaptor primer (supplied by the 39-RACE kit)was used to prime the reaction. The resulting cDNA wassimilarly subjected to nested PCR, using two nested gene-specific primers (primers C and D; Fig. 1) and an antisenseadaptor primer. The final 59- and 39-RACE products weregel-purified, cloned into PCR 2.1, and confirmed by DNAsequencing.
Construction of long (5-HT2CeL) and short (5-HT2CeS)5-HT2Ce isoform expression vectors
The complete coding sequences of 5-HT2CeL and 5-HT2CeS
were amplified directly by RT-PCR from adultC. elegans.RTwas performed as described above, and the resulting cDNA wasPCR-amplified using the proofreading DNA polymerase, PWO(Boehringer Mannheim, Laval, QC, Canada), according to thespecifications of the manufacturer. The PCR sense primerswere as follows: 59-acgcgtGCCACCATGCTCATTGAACT-CTTCTCACAC-39 (for 5-HT2CeL) and 59-acgcgtGCCAC-CATGGGAATCTACCATTTCAACGG-39 (for 5-HT2CeS).The antisense primer for both 5-HT2CeL and 5-HT2CeS was59-cgagcggccgcTTACAAGAATGTTTCCTTGATGGC-39. Re-striction sites (shown in italics) were added at the 59 end ofeach primer to facilitate further subcloning into an expressionvector (MluI for the sense primers andNotI for the antisenseprimer). The forward primers also contained a Kozak motif(shown in bold) before the start ATG (underlined) for optimaltranslation in mammalian cells (Kozak, 1986). The resultingPCR products were gel-purified, digested withMluI and NotI,
J. Neurochem., Vol. 72, No. 4, 1999
1373CHARACTERIZATION OF AC. elegansSEROTONIN RECEPTOR

and ligated to the eukaryotic expression vector pCI-neo (Pro-mega, Madison, WI, U.S.A.) that had been linearized by thesame two restriction enzymes. The final constructs were con-firmed by DNA sequencing of at least three separate clones.
Expression of the 5-HT2Ce splice variants in COS-7cells and membrane preparation
The 5-HT2CepCI-neo constructs were transiently transfectedinto COS-7 cells using Lipofectamine (GibcoBRL). In brief,aliquots of 2.5mg of plasmid DNA mixed with 20ml oflipofectamine were added to 10-cm-diameter dishes that hadbeen seeded 10–12 h earlier with;106 cells. The medium waschanged at 5 and 24 h posttransfection, and the cells wereharvested 40–48 h later. For preparation of crude membranefractions, transiently transfected COS-7 cells were washedtwice with ice-cold phosphate-buffered saline (PBS) and thenincubated in a hypotonic solution [15 mM Tris-HCl (pH 7.4),1.25 mM MgCl2, and 1 mM EDTA] for 10–15 min at 4°C.After swelling, the cells were scraped from the plates, centri-fuged at 500g for 5 min, and resuspended in 10 ml of ice-coldTEM buffer [50 mM Tris (pH 7.4), 0.5 mM EDTA, and 10 mMMgCl2] containing 1 mM freshly added phenylmethylsulfonylfluoride. The resuspended cells were lysed on ice by sonication(seven pulses of 15 s on/off), and the lysates were centrifugedat 200g for 5 min. The supernatant was centrifuged at 28,000g for 20 min at 4°C, and the resulting pellet was washed onceby resuspension in ice-cold TEM buffer and centrifugation at28,000g.The final crude membrane pellet was stored at280°Cuntil used in binding assays.
125I-LSD binding assaysFrozen membrane pellets were thawed and resuspended in
TEM buffer at a concentration of 0.5–1.5mg of protein/ml.Protein content was measured according to the method ofBradford (1976), using the Bio-Rad (Mississauga, ON, Canada)protein assay kit.125I-LSD binding was assayed in a 100-mlreaction volume in TEM buffer containing 0.02% ascorbate,COS-7 membranes (5–15mg of protein), and125I-LSD (finalconcentrations, 0.18–5 nM for saturation experiments and 1.5nM for competition assays), in the presence or absence ofvarious unlabeled drugs. Nonspecific binding was measured inthe presence of 10mM cyproheptadine and was typically,10% of total125I-LSD binding. Incubations were performedat room temperature for 50 min in restricted light. Preliminaryexperiments indicated that specific125I-LSD binding increasedlinearly up to 1 h ofincubation under these conditions. Bindingreactions were terminated by dilution with ice-cold TEM bufferfollowed by rapid filtration through Whatman GF/B glass fiberfilters that had been previously soaked in 0.2% polyethylen-imine. The filters were washed twice with 4 ml of the samebuffer, dried, and then radioassayed. All binding data wereanalyzed by the curve-fitting program Prism (GraphPad Soft-ware, San Diego) and were derived from three separate exper-iments, each done in duplicate.
Aequorin assaysThe eukaryotic expression vector pCDM.AEQ (licensed from
the National Institutes of Health, Bethesda, MD, U.S.A.) contain-ing the complete coding sequence of the calcium binding proteinaequorin (Inouye et al., 1985), and either a 5-HT2Ce expressionconstruct or a pCI-neo vector (no insert) control were used in thecotransfection of COS-7 cells. The aequorin assay was performedas described previously (Boie et al., 1997). In brief,;2 days aftertransfection the cells were charged with coelenterazine (8mM).Ninety-six-well plates were prepared containing different concen-
trations of test drugs and placed singly into a Luminoskan RSluminometer (Labsystems, Needham Heights, MA, U.S.A.) fortesting. Charged cells were dispensed at 53 104 cells per wellstarting with well A1. Light emission was recorded for the first30 s (peak 1), after which 100ml of 0.3% (vol/vol) Triton X-100in Hanks’ balanced salt solution was added to solubilize the cells,and the light emission was measured for another 10 s (peak 2).Fractional luminescence was determined at each ligand concen-tration by dividing the area under peak 1 by the sum of the areasunder peak 1 plus peak 2.
Developmental expression of 5-HT2CeL and 5-HT2CeSmRNAs in C. elegans
Expression of the two 5-HT2Ce mRNAs was examined bysemiquantitative RT-PCR. Total RNA (2mg) from three differentdevelopmental stages ofC. elegans(embryos, L2/L3 larvae, andadults) was subjected to RT (as described above) followed by astandard PCR procedure (45 s at 94°C, 45 s at 60°C, 45 s at 72°C)with primers designed to amplify products of 332 and 323 bp for5-HT2CeL and 5-HT2CeS, respectively. The PCR primers spannedintron regions, so as to prevent genomic DNA amplification, andwere selective for the different 5-HT2Ce splice variants. The nu-cleotide sequences of the sense primers were (see Fig. 1) 59-CCCTACTATGTACCTGCAAATGAATC-39 (primer L for5-HT2CeL) and 59-ACCCAAGTTTGAGAATCATTGC-39 (prim-er S for 5-HT2CeS). The antisense primer (CR; 59-CTCCATGTC-CACACCCCGTGCC-39) was common for both splice variants.Each PCR assay was standardized by simultaneous amplificationof a control housekeeping gene sequence [C. eleganseukaryoticinitiation factor 4A homologue (CeIF)] (Roussell and Bennett,1992; Krause, 1995) as described previously (Kinoshita et al.,1992). Standard curves were generated to ensure that the PCRassay was in the exponential phase of synthesis after 32 cycles for5-HT2Ce or 21 cycles for CeIF. The resulting products weresubcloned into PCR2.1 and confirmed by DNA sequencing. Den-sitometric image analyses of RT-PCR products were performedwith the NIH Image program version 1.61.
RESULTS
Isolation of two C. eleganscDNAs encoding 5-HT2CeA computer analysis of genomic sequence from theC.
elegansGenome Data Base (Sulston et al., 1992) iden-tified a predicted coding sequence (cosmid F59C12;GenBank accession no. U41038) that showed homologyto 5-HT receptors from other species. This region wasamplified by RT-PCR using reverse-transcribed adultC.eleganscDNA as a template and primers that targetedthe beginning and end of the predicted coding sequence.A 1,908-bp product was obtained and confirmed by DNAsequencing of four independent clones. DNA sequenceanalysis indicated that the amplified PCR product wasidentical to the predicted sequence obtained from theC.elegansGenome Data Base, except for a stretch of 57nucleotides (positions 3,967–4,023 of cosmid F59C12),which had been predicted to constitute an exon region(see GenBank accession no. U41038) but were lacking inall four independent clones of the RT-PCR product.Additional RACE analyses were performed to confirmthe 59 and 39 ends of the putative receptor sequence. Two59-RACE products of 460 and 546 bp were obtained andfound to be identical except for a variable 86-bp regionat the 59 end. The two 59 RACE products showed sig-
J. Neurochem., Vol. 72, No. 4, 1999
1374 F. F. HAMDAN ET AL.

nificant sequence overlap with the 1,908-bp RT-PCRproduct described above and thus were assumed to rep-resent the 59 ends of two different transcripts of 5-HT2Ce.
The complete cDNA sequence for each 5-HT2Ce tran-script and their respective deduced amino acid sequencesare shown in Fig. 1. The longest of the two sequences(5-HT2CeL) comprises 2,358 bp and has an open readingframe (ORF) that encodes a predicted protein of 683amino acids with a calculated molecular mass of 76.2kDa. The 59 end bears a conserved 22-nucleotide splicedleader (SL) sequence (SL1) (Krause and Hirsh, 1987),suggesting that 5-HT2CeL is trans-spliced at the 59 end toan SL1 sequence, just as described for several otherC.elegansRNAs (Blumenthal and Steward, 1997). A short59 untranslated region (UTR) of 22 bp precedes thepredicted start ATG. At the 39 end, the RACE analysisidentified a 262-bp UTR, which includes a potentialpolyadenylation site (GATAAA) (Blumenthal and Stew-ard, 1997) 21 bp upstream of a poly (A) tail. The shorter5-HT2Cespecies (5-HT2CeS) lacks 86 bp at its 59 end butis otherwise identical to 5-HT2CeL. 5-HT2CeScomprises2,272 bp and has a correspondingly shorter ORF thatencodes a protein of 635 amino acids with a calculatedmolecular mass of 70.8 kDa. The predicted start codonfor this variant is at nucleotide position 189 of the longer5-HT2CeL cDNA sequence (Fig. 1). The two proteinsvary by 48 amino acids at the N-terminal end. As in thecase of the longer species, 5-HT2CeScontains an SL1 atits 59 end (Fig. 1), indicating that the shorter variant of5-HT2Ce is alsotrans-spliced to an SL sequence.
A comparison of the cDNA sequences of 5-HT2CeLand 5-HT2CeSwith the original genomic sequence fromthe C. elegansGenome Data Base provided a genomicmap for 5-HT2Ce and also clarified the origin of the twolong and short transcripts. The organization of the5-HT2Cegene is shown in Fig. 2. The genomic sequenceis ;8 kb long and spans two overlapping cosmids[K08B5 (accession no. U41022) and F59C12 (GenBankaccession no. U41038)] of theC. elegansgenome se-quence. The beginning of the sequence was identified atposition 31,475 of cosmid K08B5, and the potentialpolyadenylation site is at position 6,205 of cosmidF59C12. Six exons separated by five introns of variablelengths were identified, including an unusually long in-tron of 2,117 bp, which separates the first two exons inthe sequence. Further analysis of the genomic sequenceidentified exon I as the variable 86-bp region of the two
FIG. 1. Complete cDNA and predicted amino acid sequence ofthe 5-HT2CeL. Nucleotide sequence that is absent in the shortervariant, 5-HT2CeS, is overlined. Each of the two variants is trans-spliced at its respective 59 end to a conserved C. elegans SL1sequence. Sense (F, C, and D) and antisense (A, B, and R) PCRprimers used for cloning and RACE analyses are marked by a
solid arrow. Selective 5-HT2CeL and 5-HT2CeS primers used forsemiquantitative RT-PCR are indicated by dashed arrows(sense, S and L; antisense, CR). The predicted initiation methi-onine for 5-HT2CeS is marked by a closed circle. Boxed aminoacids represent predicted TMs, which were numbered I–VII.Open squares and circles depict potential N-glycosylation sitesand phosphorylation consensus sites, respectively. The positionof a potential polyadenylation site (GATAAA) is underlined. Thenucleotide sequences for the two splice variants were submittedto GenBank and were designated accession nos. AF031414(5-HT2CeL) and AF031415 (5-HT2CeS).
J. Neurochem., Vol. 72, No. 4, 1999
1375CHARACTERIZATION OF AC. elegansSEROTONIN RECEPTOR

5-HT2Cespecies. Whereas the longer 5-HT2CeLtranscriptretains all six exons, 5-HT2CeS lacks exon I, althoughboth transcripts are similarlytrans-spliced at the 59 endto the SL1 sequence. The predicted translational startcodons for 5-HT2CeL and 5-HT2CeSare located on exon Iand exon II, respectively. The results suggest the longand short species are splice variants of 5-HT2Ce derivedfrom differential splicing of exon I.
Protein sequence analysis of 5-HT2Ce isoformsHydropathy analyses of the deduced 5-HT2CeL and
5-HT2CeS amino acid sequences identified seven pre-dicted transmembrane domains (TMs) connected by pre-dicted intra- and extracellular loops of variable lengths,an organization that is characteristic of all GPCRs. Theanalyses identified a long third intracellular loop (124residues), a region that has been implicated in the cou-pling of GPCRs to intracellular effectors (Wess, 1997),and an unusually long C terminus (253 amino acid res-idues), also predicted to be intracellular. In addition,several distinctive GPCR motifs were identified, includ-ing the highly conserved “DRY” peptide at the C-termi-nal end of TM III (Probst et al., 1992; Wess, 1997).These results suggest that the two forms of 5-HT2Cebelong to the GPCR receptor superfamily. 5-HT2CeL and5-HT2CeS differ only in the length of the predicted N-terminal extracellular domain, which is 48 residuesshorter in the latter species. Protein sequence analysesrevealed that 5-HT2CeL, but not 5-HT2CeS, carries a con-sensus site for N-linked glycosylation (position 21) at itsextracellular N-terminal domain. In addition, the twoisoforms exhibit two consensus phosphorylation sites:one for Ca21/calmodulin-dependent protein kinase oftype II (Thr170), located on the second intracellular loop,and the other for protein kinase C (Ser491), on the pre-dicted intracellular C-terminal tail of the receptor.
BLAST analyses (Altschul et al., 1990) identified highhomology to mammalian 5-HT receptor sequences fromother species, especially those belonging to the 5-HT2family. The dendrogram in Fig. 3 shows that 5-HT2Ceclusters primarily with the two invertebrate 5-HT2 recep-tors,Drosophila5-HT2Dro (Colas et al., 1995) andLym-naea 5-HT2Lym (Gerhardt et al., 1996), and the three
subtypes of mammalian 5-HT2 receptors, 5-HT2A,5-HT2B, and 5-HT2C (Hoyer et al., 1994). In addition,5-HT2Ce is closely related to a putative 5-HT2-like re-ceptor sequence recently cloned from the parasitic nem-atode Ascaris suum(5-HTAsc; Genbank accession no.AF005486). By comparison, 5-HT2Ce shows consider-ably less homology to other 5-HT receptor types, includ-ing the previously described 5-HT1-like receptor fromC.elegans(Olde and McCombie, 1997).
An amino acid sequence alignment of 5-HT2Ce withother members of the 5-HT2 receptor family is shown inFig. 4. The highest degree of sequence conservationoccurs within the predicted TMs. Based on pairwiseprotein alignments, 5-HT2Ce shares highest TM homol-ogy with Ascaris5-HTAsc (81%), followed byDrosoph-ila 5-HT2Dro (71%) (Colas et al., 1995),Lymnaea5-HT2Lym (67%) (Gerhardt et al., 1996), rat 5-HT2C(66%) (Julius et al., 1988), rat 5-HT2A (65%) (Julius etal., 1990), and rat 5-HT2B (62%) (Foguet et al., 1992).Several of the amino acid residues that have been impli-cated in agonist and antagonist binding to mammalian5-HT2 receptors (Hibert et al., 1991; Choudhary et al.,1992, 1993, 1995; Wang et al., 1993; Roth et al., 1997)are conserved in 5-HT2Ce. In addition, we identifiedseveral amino acid residues that distinguish all 5-HT2sequences, including 5-HT2Ce, from other 5-HT receptortypes. A list of these characteristic residues is describedin Table 1.
Ligand binding properties of 5-HT2CeL and 5-HT2CeSin COS-7 cells
To determine whether the two 5-HT2CecDNAs encodefunctional proteins, constructs of the long and shortvariants of the receptor were transiently transfected intoCOS-7 cells for subsequent measurements of receptorbinding activity. In preliminary separate experiments, thetwo expression constructs were modified to introduce aC-terminal FLAG epitope (Hopp et al., 1988), which wassubsequently targeted with an anti-FLAG antibody formonitoring expression of fusion 5-HT2Ce–FLAG pro-teins in COS-7 cells. The results (data not shown)showed intense immunofluorescence mainly on the sur-face, suggesting that the 5-HT2Ce–FLAG protein was
FIG. 2. Genomic organization of 5-HT2Ce. Thegenomic organization of 5-HT2Ce is shown diagram-maticaly based on genomic sequence from the C.elegans Genome Data Base and cDNA sequenceinformation, as described in Results. Hatched boxesindicate exon sequences that are common to both5-HT2Ce splice variants. The variably spliced exon I,which is present only in 5-HT2CeL transcripts, is rep-resented by an open box. The SL1 sequence, whichis trans-spliced onto each 5-HT2Ce transcript, isshown by a solid box. Introns are represented bydashed horizontal lines. The relative positions of thetwo translational start codons, the common stopcodon (TAA), and the polyadenylation site (GATAAA)are also indicated.
J. Neurochem., Vol. 72, No. 4, 1999
1376 F. F. HAMDAN ET AL.

being expressed at high levels and was targeted to theplasma membrane in COS-7 cells. The same results wereobtained with the short 5-HT2CeSspecies similarly fusedto the FLAG epitope, whereas no immunofluorescencecould be detected in a mock-transfected control.
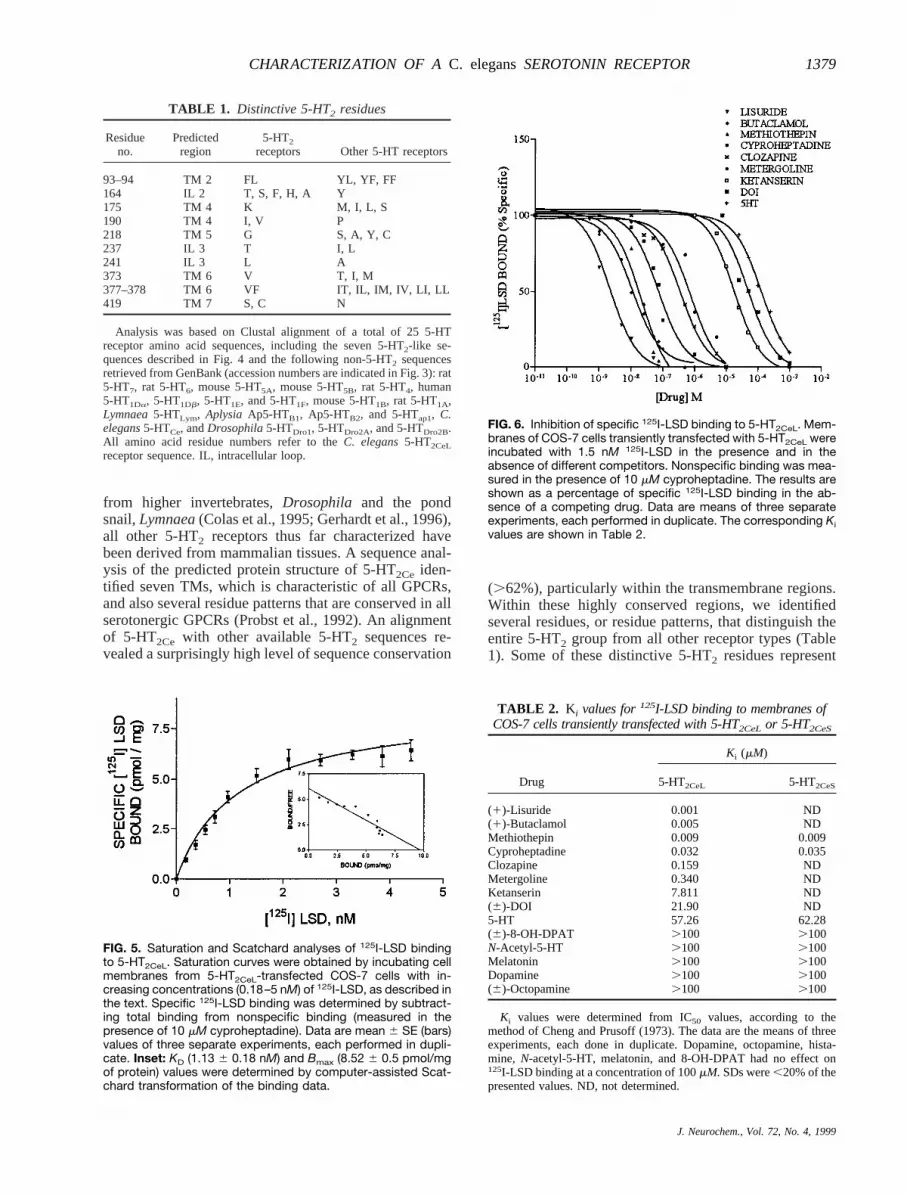
Membranes of transiently transfected COS-7 cellswere isolated and assayed for their ability to bind theserotonergic ligand125I-LSD. Membranes containing5-HT2CeL displayed saturable specific125I-LSD bindingin a concentration range of 0.18–5 nM (Fig. 5). A com-puter-assisted Scatchard analysis of 5-HT2CeL saturationisotherms identified a single class of binding sites withan apparent dissociation constant (KD) of ;1.136 0.18nM and a binding capacity (Bmax) of 8.52 6 0.18pmol/mg of protein. No specific125I-LSD binding wasobserved in untransfected cells or cells transfected withplasmid alone. To determine the pharmacological profileof 5-HT2CeL, we examined the ability of several seroto-nergic and nonserotonergic ligands to compete for125I-LSD-labeled receptor (Fig. 6). The resultingKi values(Table 2) revealed the following order of drug potency:lisuride. butaclamol. methiothepin. cyproheptadine. clozapine. metergoline. ketanserin. DOI . 5-HT. Other biogenic amines, including dopamine, octo-pamine, histamine, and tryptamine, were inactive at con-centrations of 0.1 mM, as were products of 5-HT me-tabolism (N-acetyl-5-HT and melatonin) and the mam-malian 5-HT1A agonist 8-OH-DPAT. Similar experi-ments were repeated with 5-HT2CeSto identify possibledifferences in the pharmacological profile of the shorterreceptor isoform. Competition curves were obtained for5-HT and two of the most effective competitors, methio-thepin and cyproheptadine. All three ligands exhibitedsimilar affinities for the short and long forms of the
receptor (Table 2), with the resulting competition curvesbeing virtually identical (data not shown).
5-HT induces an increase in intracellular Ca21
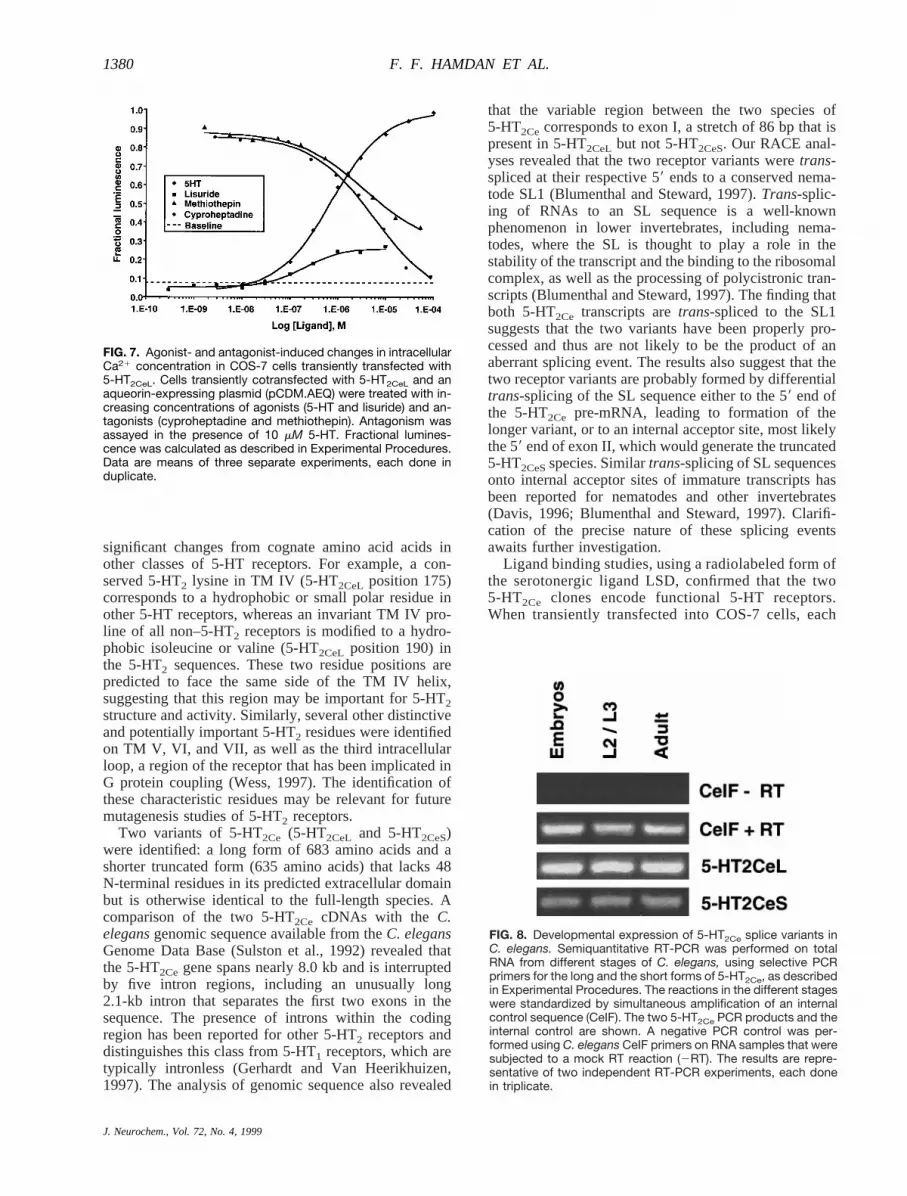
concentration in 5-HT2Ce-transfected COS-7 cellsAgonist activation of serotonergic 5-HT2 receptors is
known to cause an elevation in intracellular Ca21 con-centration, which serves as a second messenger in5-HT2-mediated signaling (Hoyer et al., 1994). We usedan aequorin-based assay (Boie et al., 1997) to investigatewhether 5-HT2Ce mediates a similar intracellular Ca21
concentration increase in COS-7 cells. Cells were tran-siently cotransfected with an aequorin-expressing plas-mid (pCDM.AEQ) and either a 5-HT2Ceexpression con-struct (5-HT2CeL or 5-HT2CeS) or vector alone. An in-crease in aqueorin luminescence, indicative of anelevation in intracellular Ca21 concentration, was seenafter challenge of 5-HT2Ce-transfected cells with 5-HT orthe serotonergic agonist lisuride. In contrast, the mock-transfected control showed no visible response to eitherligand (Fig. 7). Furthermore, no response was detected in5-HT2Ce-transfected cells treated with increasing concen-trations (up to 0.1 mM) of dopamine, octopamine, hista-mine, melatonin, and 8-OH-DPAT (data not shown).5-HT and lisuride elicited identical responses in5-HT2CeL- and 5-HT2CeS-transfected cells. The EC50 val-ues (mean6 SEM) for 5-HT were 0.726 0.09 and 0.806 0.08 mM for the long and short receptor variants,respectively. The corresponding EC50 values for lisuridewere about threefold lower (0.206 0.06 and 0.246 0.13mM for 5-HT2CeL and 5-HT2CeS, respectively), but themaximal response was only;25% of that obtained with5-HT (Fig. 7). Additional experiments with the classical5-HT2 agonist DOI failed to elicit a Ca21 responseexcept for a weak signal at very high concentrations
FIG. 3. Dendrogram shows the structural relation-ship of 5-HT2Ce to other cloned G protein-coupled5-HT receptors. The amino acid sequences of5-HT2CeL and several 5-HT receptors were com-pared and clustered using the program DNASIS ver-sion 3.7 (Hitachi Software, San Francisco, CA,U.S.A.). The lengths of the horizontal lines connect-ing one amino acid sequence to another are in-versely proportional to the percentages of similaritybetween receptors or group of receptors. Receptorsequences used are from the human, rat, mouse,Drosophila (Dro), Lymnaea (Lym), Aplysia (Ap or ap),C. elegans (Ce), and Ascaris (Asc). The correspond-ing GenBank accession no. is indicated next to eachsequence.
J. Neurochem., Vol. 72, No. 4, 1999
1377CHARACTERIZATION OF AC. elegansSEROTONIN RECEPTOR

(.100 mM) of the drug (data not shown). Two classical5-HT receptor antagonists, methiothepin and cyprohep-tadine, were tested for their ability to inhibit the 5-HT-induced elevation in cellular Ca21 concentration. Each ofthe two antagonists was able to inhibit the response in thepresence of a fixed concentration (10mM) of 5-HT inboth 5-HT2CeL-transfected cells (Fig. 7) and cells trans-fected with 5-HT2CeS(data not shown). The IC50 valuesfor methiothepin and cyproheptadine were similar forboth the long and short splice variants of 5-HT2Ce withrespective values (mean6 SEM) of 3.626 2.6 (long)and 2.046 0.45 mM (short) and 5.936 1.4 (long) and3.07 6 0.51 mM (short). It is noteworthy that methio-thepin did not fully reverse the response to 5-HT, even athigh micromolar concentrations, whereas cyprohepta-dine caused almost complete inhibition.
5-HT2Ce developmental expression inC. elegansThe expression of 5-HT2Cesplice variants was examined
by semiquantitative RT-PCR in three different stages ofC.
elegans(embryos, L2/L3 larvae, and adult). As can be seenin Fig. 8, each of the two 5-HT2Ce splice variants wasexpressed at approximately the same level in all stagestested. However, the expression level of 5-HT2CeL wasconsistently fourfold higher than that of 5-HT2CeS in allthree stages. Expression levels were standardized by com-parison with a constitutively expressed control gene (CeIF)(Roussell and Bennett, 1992; Krause, 1995). No CeIF PCRproducts were detected in a negative control that lackedreverse transcriptase, thus confirming the absence of con-taminating genomic DNA (Fig. 8).
DISCUSSION
This study describes the molecular cloning and func-tional expression of a novelC. elegansreceptor (5-HT2Ce) that shows sequence homology with 5-HT2 re-ceptors from other species. This is the most primitive5-HT2-like receptor ever cloned and characterized at themolecular level. With the exception of two receptors
FIG. 4. Amino acid sequence alignment of the TM regions of 5-HT2Ce with members of the 5-HT2 family, including rat 5-HT2A, rat5-HT2C, rat 5-HT2B, Drosophila 5-HT2Dro, Lymnaea 5-HT2Lym, and a putative 5-HT2 receptor sequence from Ascaris, 5-HTAsc (refer to Fig.3 for relevant GenBank accession nos.). Predicted TM I–VII regions are overlined. Numbers in parentheses correspond to the numberof amino acids at the N and C termini and the third intracellular loop that are not represented in the alignment. Dark boxes and boldletters represent identical amino acids in at least four of the sequences, whereas gray boxes indicate conservative amino acid changes.Gaps, indicated by dashes, are included for optimal protein alignment. Amino acid residues that distinguish all 5-HT2 receptors fromother receptor types (see Table 1) are marked by solid circles, whereas those conserved in all GPCRs are indicated by an asterisk.
J. Neurochem., Vol. 72, No. 4, 1999
1378 F. F. HAMDAN ET AL.
from higher invertebrates,Drosophila and the pondsnail,Lymnaea(Colas et al., 1995; Gerhardt et al., 1996),all other 5-HT2 receptors thus far characterized havebeen derived from mammalian tissues. A sequence anal-ysis of the predicted protein structure of 5-HT2Ce iden-tified seven TMs, which is characteristic of all GPCRs,and also several residue patterns that are conserved in allserotonergic GPCRs (Probst et al., 1992). An alignmentof 5-HT2Ce with other available 5-HT2 sequences re-vealed a surprisingly high level of sequence conservation
(.62%), particularly within the transmembrane regions.Within these highly conserved regions, we identifiedseveral residues, or residue patterns, that distinguish theentire 5-HT2 group from all other receptor types (Table1). Some of these distinctive 5-HT2 residues represent
TABLE 1. Distinctive 5-HT2 residues
Residueno.
Predictedregion
5-HT2
receptors Other 5-HT receptors
93–94 TM 2 FL YL, YF, FF164 IL 2 T, S, F, H, A Y175 TM 4 K M, I, L, S190 TM 4 I, V P218 TM 5 G S, A, Y, C237 IL 3 T I, L241 IL 3 L A373 TM 6 V T, I, M377–378 TM 6 VF IT, IL, IM, IV, LI, LL419 TM 7 S, C N
Analysis was based on Clustal alignment of a total of 25 5-HTreceptor amino acid sequences, including the seven 5-HT2-like se-quences described in Fig. 4 and the following non-5-HT2 sequencesretrieved from GenBank (accession numbers are indicated in Fig. 3): rat5-HT7, rat 5-HT6, mouse 5-HT5A, mouse 5-HT5B, rat 5-HT4, human5-HT1Da, 5-HT1Db, 5-HT1E, and 5-HT1F, mouse 5-HT1B, rat 5-HT1A,Lymnaea5-HTLym, Aplysia Ap5-HTB1, Ap5-HTB2, and 5-HTap1, C.elegans5-HTCe, andDrosophila5-HTDro1, 5-HTDro2A, and 5-HTDro2B.All amino acid residue numbers refer to theC. elegans5-HT2CeL
receptor sequence. IL, intracellular loop.
FIG. 5. Saturation and Scatchard analyses of 125I-LSD bindingto 5-HT2CeL. Saturation curves were obtained by incubating cellmembranes from 5-HT2CeL-transfected COS-7 cells with in-creasing concentrations (0.18–5 nM) of 125I-LSD, as described inthe text. Specific 125I-LSD binding was determined by subtract-ing total binding from nonspecific binding (measured in thepresence of 10 mM cyproheptadine). Data are mean 6 SE (bars)values of three separate experiments, each performed in dupli-cate. Inset: KD (1.13 6 0.18 nM) and Bmax (8.52 6 0.5 pmol/mgof protein) values were determined by computer-assisted Scat-chard transformation of the binding data.
FIG. 6. Inhibition of specific 125I-LSD binding to 5-HT2CeL. Mem-branes of COS-7 cells transiently transfected with 5-HT2CeL wereincubated with 1.5 nM 125I-LSD in the presence and in theabsence of different competitors. Nonspecific binding was mea-sured in the presence of 10 mM cyproheptadine. The results areshown as a percentage of specific 125I-LSD binding in the ab-sence of a competing drug. Data are means of three separateexperiments, each performed in duplicate. The corresponding Kivalues are shown in Table 2.
TABLE 2. Ki values for125I-LSD binding to membranes ofCOS-7 cells transiently transfected with 5-HT2CeL or 5-HT2CeS
Drug
Ki (mM)
5-HT2CeL 5-HT2CeS
(1)-Lisuride 0.001 ND(1)-Butaclamol 0.005 NDMethiothepin 0.009 0.009Cyproheptadine 0.032 0.035Clozapine 0.159 NDMetergoline 0.340 NDKetanserin 7.811 ND(6)-DOI 21.90 ND5-HT 57.26 62.28(6)-8-OH-DPAT .100 .100N-Acetyl-5-HT .100 .100Melatonin .100 .100Dopamine .100 .100(6)-Octopamine .100 .100
Ki values were determined from IC50 values, according to themethod of Cheng and Prusoff (1973). The data are the means of threeexperiments, each done in duplicate. Dopamine, octopamine, hista-mine, N-acetyl-5-HT, melatonin, and 8-OH-DPAT had no effect on125I-LSD binding at a concentration of 100mM. SDs were,20% of thepresented values. ND, not determined.
J. Neurochem., Vol. 72, No. 4, 1999
1379CHARACTERIZATION OF AC. elegansSEROTONIN RECEPTOR
significant changes from cognate amino acid acids inother classes of 5-HT receptors. For example, a con-served 5-HT2 lysine in TM IV (5-HT2CeL position 175)corresponds to a hydrophobic or small polar residue inother 5-HT receptors, whereas an invariant TM IV pro-line of all non–5-HT2 receptors is modified to a hydro-phobic isoleucine or valine (5-HT2CeL position 190) inthe 5-HT2 sequences. These two residue positions arepredicted to face the same side of the TM IV helix,suggesting that this region may be important for 5-HT2structure and activity. Similarly, several other distinctiveand potentially important 5-HT2 residues were identifiedon TM V, VI, and VII, as well as the third intracellularloop, a region of the receptor that has been implicated inG protein coupling (Wess, 1997). The identification ofthese characteristic residues may be relevant for futuremutagenesis studies of 5-HT2 receptors.
Two variants of 5-HT2Ce (5-HT2CeL and 5-HT2CeS)were identified: a long form of 683 amino acids and ashorter truncated form (635 amino acids) that lacks 48N-terminal residues in its predicted extracellular domainbut is otherwise identical to the full-length species. Acomparison of the two 5-HT2Ce cDNAs with the C.elegansgenomic sequence available from theC. elegansGenome Data Base (Sulston et al., 1992) revealed thatthe 5-HT2Ce gene spans nearly 8.0 kb and is interruptedby five intron regions, including an unusually long2.1-kb intron that separates the first two exons in thesequence. The presence of introns within the codingregion has been reported for other 5-HT2 receptors anddistinguishes this class from 5-HT1 receptors, which aretypically intronless (Gerhardt and Van Heerikhuizen,1997). The analysis of genomic sequence also revealed
that the variable region between the two species of5-HT2Cecorresponds to exon I, a stretch of 86 bp that ispresent in 5-HT2CeL but not 5-HT2CeS. Our RACE anal-yses revealed that the two receptor variants weretrans-spliced at their respective 59 ends to a conserved nema-tode SL1 (Blumenthal and Steward, 1997).Trans-splic-ing of RNAs to an SL sequence is a well-knownphenomenon in lower invertebrates, including nema-todes, where the SL is thought to play a role in thestability of the transcript and the binding to the ribosomalcomplex, as well as the processing of polycistronic tran-scripts (Blumenthal and Steward, 1997). The finding thatboth 5-HT2Ce transcripts aretrans-spliced to the SL1suggests that the two variants have been properly pro-cessed and thus are not likely to be the product of anaberrant splicing event. The results also suggest that thetwo receptor variants are probably formed by differentialtrans-splicing of the SL sequence either to the 59 end ofthe 5-HT2Ce pre-mRNA, leading to formation of thelonger variant, or to an internal acceptor site, most likelythe 59 end of exon II, which would generate the truncated5-HT2CeSspecies. Similartrans-splicing of SL sequencesonto internal acceptor sites of immature transcripts hasbeen reported for nematodes and other invertebrates(Davis, 1996; Blumenthal and Steward, 1997). Clarifi-cation of the precise nature of these splicing eventsawaits further investigation.
Ligand binding studies, using a radiolabeled form ofthe serotonergic ligand LSD, confirmed that the two5-HT2Ce clones encode functional 5-HT receptors.When transiently transfected into COS-7 cells, each
FIG. 8. Developmental expression of 5-HT2Ce splice variants inC. elegans. Semiquantitative RT-PCR was performed on totalRNA from different stages of C. elegans, using selective PCRprimers for the long and the short forms of 5-HT2Ce, as describedin Experimental Procedures. The reactions in the different stageswere standardized by simultaneous amplification of an internalcontrol sequence (CeIF). The two 5-HT2Ce PCR products and theinternal control are shown. A negative PCR control was per-formed using C. elegans CeIF primers on RNA samples that weresubjected to a mock RT reaction (2RT). The results are repre-sentative of two independent RT-PCR experiments, each donein triplicate.
FIG. 7. Agonist- and antagonist-induced changes in intracellularCa21 concentration in COS-7 cells transiently transfected with5-HT2CeL. Cells transiently cotransfected with 5-HT2CeL and anaqueorin-expressing plasmid (pCDM.AEQ) were treated with in-creasing concentrations of agonists (5-HT and lisuride) and an-tagonists (cyproheptadine and methiothepin). Antagonism wasassayed in the presence of 10 mM 5-HT. Fractional lumines-cence was calculated as described in Experimental Procedures.Data are means of three separate experiments, each done induplicate.
J. Neurochem., Vol. 72, No. 4, 1999
1380 F. F. HAMDAN ET AL.

receptor variant was targeted to the cell membrane andwas shown to bind125I-LSD specifically and with highaffinity. In addition, 5-HT and other known serotoner-gic agents were able to inhibit fully125I-LSD binding,whereas other related monoamines, such as dopamineand octopamine, which are also present inC. elegans,were inactive. These findings demonstrate specificityof 5-HT2Ce for 5-HT. Competition studies of the longreceptor isoform revealed some of the binding char-acteristics of mammalian 5-HT2 receptors, including alow affinity for the endogenous ligand (5-HT) andcomparatively higher affinities for the classic 5-HT2antagonists ketanserin, metergoline, and cyprohepta-dine (Hoyer et al., 1994). However, the present phar-macological characterization also unveiled severalsurprising properties of 5-HT2Ce, which are inconsis-tent with those of either mammalian or other inverte-brate 5-HT2 receptors. Most notable is that 5-HT2Ceexhibited much higher affinity for ergolines, such asLSD and lisuride, and the nonspecific antagonist me-thiothepin than any of the 5-HT2 ligands tested. Alsosurprising was the relative low affinity of 5-HT2Ce forDOI, a selective high-affinity agonist of all known5-HT2 receptors, and the lack of discrimination be-tween metergoline, a 5-HT2 antagonist, and clozapine,a relatively nonspecific antagonist of 5-HT6/5-HT7receptors (Hoyer et al., 1994). Taken together, theresults indicate that 5-HT2Ce is a novel pharmacolog-ical receptor type, which is structurally related to the5-HT2 family but has a distinctive drug binding pro-file.
The binding specificity of 5-HT2Ce, including its ap-parent preference for ergot derivatives and the exception-ally low affinity for the natural ligand (5-HT) is reminis-cent of invertebrate 5-HT1-like receptors (Gerhardt andVan Heerikhuizen, 1997), in particular,Lymnaea5-HTLym (Sugamori et al., 1993) and the previouslyclonedC. elegans5-HTCe receptor (Olde and McCom-bie, 1997). In the latter case, the receptor also showedpreferential affinity for methiothepin and clozapine, aswell as butaclamol, with the overall order of drug po-tency being very similar to that reported here for5-HT2Ce. These observations suggest that 5-HT2Ce hasinvertebrate 5-HT1-like binding determinants, despite theoverall lack of structural homology with the 5-HT1 class.The results also raise the interesting possibility that thismost primitive member of the 5-HT2 family may haveretained characteristics of an ancestral 5-HT1/5-HT2 pro-totype that existed before the separation of the two re-ceptor families. Thus, 5-HT2Ce may be be more closelyrelated to an earlier form of 5-HT2 receptor than anyother known 5-HT2 subtype.
We have shown that 5-HT can activate 5-HT2Ce ex-pressed in COS-7 cells and that the activation leads to arise in intracellular Ca21 concentration. These findingsindicate that 5-HT2Ce is able to couple to intracellulareffectors in the COS-7 environment and probably actsthrough the same Ca21-mediated pathway of signaltransduction as other 5-HT2 receptors. Although it is
unknown if 5-HT2Ce acts in the same manner in vivo, itis noteworthy that the key elements needed for 5-HT2signaling are present inC. elegans,including Gaq (Barg-mann and Kaplan, 1998), to which 5-HT2 receptors areknown to couple, and a phosphoinositide-specific phos-pholipase C (Shibatohge et al., 1998). Additional phar-macological characterization of the 5-HT2Ce-mediatedCa21 response revealed that cyproheptadine and, to alesser extent, methiothepin are effective antagonists,whereas lisuride has agonist activity and DOI is essen-tially inactive. The lack of response to the classic 5-HT2agonist DOI illustrates once again the unusual pharma-cological characteristics of this 5-HT2-like C. elegansreceptor. It is noteworthy that the effect of lisuride wasonly ;25% of that obtained with 5-HT. Thus, 5-HTappears to be a much more efficacious agonist despite itsrelatively lower affinity for the receptor.
A parallel characterization of 5-HT2CeL and 5-HT2CeShas shown that the two splice variants have essentiallyidentical ligand binding and signaling properties. 5-HT,in particular, displayed the same binding affinity and thesame agonist activity at both forms of the receptor. Thissimilarity is not surprising because the two receptorforms differ only in their N-terminal extracellular do-main, a region that is not believed to be essential forfunctional activity of the receptor (Buck et al., 1991). Itis noteworthy, however, that the N-terminal region con-tains a potentialN-glycosylation site (position 21), whichis lacking in the shorter species. Thus, there may bedifferences in the posttranslational modification of thetwo C. elegansreceptor isoforms, which have not beenidentified in this study but may be of significance in vivo.To learn more about these two receptor species, weinvestigated their relative expression at the mRNA levelin different developmental stages ofC. elegans.Theresults revealed that the level of expression for the longerreceptor variant is about fourfold higher than that of theshorter species in all developmental stages tested, includ-ing embryos, L2/L3 larvae, and adults. Additional re-search is needed to elucidate the biological role of5-HT2Ceand to determine if differential expression of thetwo receptor variants has functional significance inC.elegans.
Acknowledgment: We thank Dr. G. O’Neill and Dr. D.Cully for providing C. elegansmRNA and for helpful discus-sions on the heterologous expression of the 5-HT2Ce receptor.F. F. Hamdan is the recipient of a Ph.D. Scholarship fromFonds Pour la Formation de Chercheurs et l’Aide a la Recher-che du Quebec. This work was supported by a grant from theNatural Sciences and Engineering Research Council of Canada.
REFERENCES
Almaula N., Ebersole B. J., Zhang D., Weinstein H., and Sealfon S. C.(1996) Mapping the binding site pocket of the serotonin5-hydroxytryptamine2A receptor. Ser3.36(159) provides a secondinteraction site for the protonated amine of serotonin but not oflysergic acid diethylamide or bufotenin.J. Biol. Chem.271,14672–14675.
J. Neurochem., Vol. 72, No. 4, 1999
1381CHARACTERIZATION OF AC. elegansSEROTONIN RECEPTOR

Altschul S. F., Gish W., Miller W., Myers E. W., and Lipman D. J.(1990) Basic local alignment search tool.J. Mol. Biol. 215,403–410.
Angers A., Storozhuk M. V., Duchaine T., Castellucci V. F., andDesGroseillers L. (1998) Cloning and functional expression of anAplysia 5-HT receptor negatively coupled to adenylate cyclase.J. Neurosci.18, 5586–5593.
Bargmann C. I. and Kaplan J. M. (1998) Signal transduction in theCaenorhabditis elegansnervous system.Annu. Rev. Neurosci.21,279–308.
Blumenthal T. and Steward K. (1997) RNA processing and genestructure, inC. elegans II(Riddle D. L., Blumenthal T., MeyerB. J., and Priess J. R., eds), pp. 117–145. Cold Spring HarborLaboratory Press, Cold Spring Harbor, New York.
Boie Y., Stocco R., Sawyer N., Slipetz D. M., Ungrin M. D., Neus-chafer-Rube F., Puschel G. P., Metters K. M., and Abramovitz M.(1997) Molecular cloning and characterization of the four ratprostaglandin E2 prostanoid receptor subtypes.Eur. J. Pharmacol.340,227–241.
Bradford M. M. (1976) A rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principle ofprotein–dye binding.Anal. Biochem.72, 248–254.
Buck F., Meyerhof W., Werr H., and Richter D. (1991) Characteriza-tion of N- and C-terminal deletion mutants of the rat serotoninHT2 receptor inXenopus laevisoocytes.Biochem. Biophys. Res.Commun.178,1421–1428.
Cheng Y. and Prusoff W. H. (1973) Relationship between the inhibitionconstant (Ki) and the concentration of inhibitor which causes 50per cent inhibition (I50) of an enzymatic reaction.Biochem. Phar-macol.22, 3099–3108.
Choudhary M. S., Craigo S., and Roth B. L. (1992) Identification ofreceptor domains that modify ligand binding to 5-hydroxytrypta-mine2 and 5-hydroxytryptamine1c serotonin receptors.Mol. Phar-macol.42, 627–633.
Choudhary M. S., Craigo S., and Roth B. L. (1993) A singlepoint mutation (Phe340 3 Leu340) of a conserved phenylala-nine abolishes 4-[125I]iodo-2,5-dimethoxy)phenylisopro-pylamine and [3H]mesulergine but not [3H]ketanserin bindingto 5-hydroxytryptamine2 receptors.Mol. Pharmacol.43, 755–761.
Choudhary M. S., Sachs N., Uluer A., Glennon R. A., Westkaemper R. B.,and Roth B. L. (1995) Differential ergoline and ergopeptine bindingto 5-hydroxytryptamine2A receptors: ergolines require an aromaticresidue at position 340 for high affinity binding.Mol. Pharmacol.47,450–457.
Colas J. F., Launay J. M., Kellermann O., Rosay P., and Maroteaux L.(1995) Drosophila 5-HT2 serotonin receptor: coexpression withfushi-tarazu during segmentation.Proc. Natl. Acad. Sci. USA92,5441–5445.
Davis R. E. (1996) Spliced leader RNAtrans-splicing in metazoa.Parasitol. Today12, 33–40.
Desai C. and Horvitz H. R. (1989)Caenorhabditis elegansmutantsdefective in the functioning of the motor neurons responsible foregg laying.Genetics121,703–721.
Desai C., Garriga G., McIntire S. L., and Horvitz H. R. (1988) Agenetic pathway for the development of theCaenorhabditis el-egansHSN motor neurons.Nature336,638–646.
Foguet M., Hoyer D., Pardo L. A., Parekh A., Kluxen F. W., KalkmanH. O., Stuhmer W., and Lubbert H. (1992) Cloning and functionalcharacterization of the rat stomach fundus serotonin receptor.EMBO J.11, 3481–3487.
Gerhardt C. C. and Van Heerikhuizen H. (1997) Functional character-istics of heterologously expressed 5-HT receptors.Eur. J. Phar-macol.334,1–23.
Gerhardt C. C., Leysen J. E., Planta R. J., Vreugdenhil E., and VanHeerikhuizen H. (1996) Functional characterisation of a 5-HT2
receptor cDNA cloned fromLymnaea stagnalis. Eur. J. Pharma-col. 311,249–258.
Hibert M. F., Trumpp-Kallmeyer S., Bruinvels A., and Hoflack J.(1991) Three-dimensional models of neurotransmitter G-bindingprotein-coupled receptors.Mol. Pharmacol.40, 8–15.
Hopp T. P., Prickett K. S., Price V. L., Libby R. T., March C. J.,Cerretti D. P., Urdal D. L., and Conlon P. J. (1988) A shortpolypeptide marker sequence useful for recombinant protein iden-tification and purification.Biotechnology6, 1204–1210.
Horvitz H. R., Chalfie M., Trent C., Sulston J., and Evans P. D. (1982)Serotonin and octopamine in the nematodeCaenorhabditis el-egans. Science216,1012–1014.
Hoyer D., Clarke D. E., Fozard J. R., Hartig P. R., Martin G. R.,Mylecharane E. J., Saxena P. R., and Humphrey P. P. (1994)International Union of Pharmacology classification of receptorsfor 5-hydroxytryptamine (serotonin).Pharmacol. Rev.46, 157–203.
Inouye S., Noguchi M., Sakaki Y., Takagi Y., Miyata T., Iwanaga S.,and Tsuji F. I. (1985) Cloning and sequence analysis of cDNA forthe luminescent protein aequorin.Proc. Natl. Acad. Sci. USA82,3154–3158.
Johnson M. P., Loncharich R. J., Baez M., and Nelson D. L. (1994)Species variations in transmembrane region V of the 5-hydroxy-tryptamine type 2A receptor alter the structure–activity relation-ship of certain ergolines and tryptamines.Mol. Pharmacol.45,277–286.
Julius D., MacDermott A. B., Axel R., and Jessell T. M. (1988)Molecular characterization of a functional cDNA encoding theserotonin 1c receptor.Science241,558–564.
Julius D., Huang K. N., Livelli T. J., Axel R., and Jessell T. M. (1990)The 5-HT2 receptor defines a family of structurally distinct butfunctionally conserved serotonin receptors.Proc. Natl. Acad. Sci.USA87, 928–932.
Kinoshita T., Imamura J., Nagai H., and Shimotohno K. (1992) Quan-tification of gene expression over a wide range by the polymerasechain reaction.Anal. Biochem.206,231–235.
Kozak M. (1986) Point mutations define a sequence flanking the AUGinitiator codon that modulates translation by eukaryotic ribo-somes.Cell 44, 283–292.
Krause M. (1995) Techniques for analyzing transcription and trans-lation, in Methods in Cell Biology, Vol. 48: Caenorhabditiselegans: Modern Biological Analysis of an Organism(EpsteinH. F. and Shakes D. C., eds), pp. 513–529. Academic Press,San Diego.
Krause M. and Hirsh D. (1987) Atrans-spliced leader sequence onactin mRNA inC. elegans. Cell49, 753–761.
Leonard B. E. (1994) Serotonin receptors—where are they going?Int.Clin. Psychopharmacol.9 (Suppl. 1), 7–17.
Lewis J. A. and Fleming J. T. (1995) Basic culture methods, inMethodsin Cell Biology, Vol. 48: Caenorhabditis elegans: Modern Bio-logical Analysis of an Organism(Epstein H. F. and Shakes D. C.,eds), pp. 3–29. Academic Press, San Diego.
Loer C. M. and Kenyon C. J. (1993) Serotonin-deficient mutants andmale mating behavior in the nematodeCaenorhabditis elegans.J. Neurosci.13, 5407–5417.
Olde B. and McCombie W. R. (1997) Molecular cloning and functionalexpression of a serotonin receptor fromCaenorhabditis elegans. J.Mol. Neurosci.8, 53–62.
Probst W. C., Snyder L. A., Schuster D. I., Brosius J., and Sealfon S. C.(1992) Sequence alignment of the G-protein coupled receptorsuperfamily.DNA Cell Biol.11, 1–20.
Roth B. L., Shoham M., Choudhary M. S., and Khan N. (1997) Identifi-cation of conserved aromatic residues essential for agonist bindingand second messenger production at 5-hydroxytryptamine2A recep-tors.Mol. Pharmacol.52, 259–266.
Roussell D. L. and Bennett K. L. (1992)CaenorhabditiscDNA en-codes an eIF-4A-like protein.Nucleic Acids Res.20, 3783.
Saudou F., Boschert U., Amlaiky N., Plassat J. L., and Hen R. (1992)A family of Drosophilaserotonin receptors with distinct intracel-lular signalling properties and expression patterns.EMBO J.11,7–17.
Shibatohge M., Kariya K., Liao Y., Hu C. D., Watari Y., Goshima M.,Shima F., and Kataoka T. (1998) Identification of PLC210, aCaenorhabditis elegansphospholipase C, as a putative effector ofRas.J. Biol. Chem.273,6218–6222.
J. Neurochem., Vol. 72, No. 4, 1999
1382 F. F. HAMDAN ET AL.

Sugamori K. S., Sunahara R. K., Guan H. C., Bulloch A. G., TensenC. P., Seeman P., Niznik H. B., and Van Tol H. H. (1993)Serotonin receptor cDNA cloned fromLymnaea stagnalis. Proc.Natl. Acad. Sci. USA90, 11–15.
Sulston J., Du Z., Thomas K., Wilson R., Hillier L., Staden R., HalloranN., Green P., Thierry-Mieg J., Qiu L., et al. (1992) TheC. elegansgenome sequencing project: a beginning.Nature356,37–41.
Wang C. D., Gallaher T. K., and Shih J. C. (1993) Site-directedmutagenesis of the serotonin 5-hydroxytrypamine2 receptor: iden-
tification of amino acids necessary for ligand binding and receptoractivation.Mol. Pharmacol.43, 931–940.
Wess J. (1997) G-protein-coupled receptors: molecular mechanismsinvolved in receptor activation and selectivity of G-protein recog-nition. FASEB J.11, 346–354.
Witz P., Amlaiky N., Plassat J. L., Maroteaux L., Borrelli E., and HenR. (1990) Cloning and characterization of aDrosophilaserotoninreceptor that activates adenylate cyclase.Proc. Natl. Acad. Sci.USA87, 8940–8944.
J. Neurochem., Vol. 72, No. 4, 1999
1383CHARACTERIZATION OF AC. elegansSEROTONIN RECEPTOR





























