Embed Size (px)
DESCRIPTION
fistum
Citation preview

PERKECAMBAHAN DAN DORMANSI
LAPORAN PRAKTIKUM
Untuk Memenuhi Tugas Mata Kuliah Fisiologi Tumbuhan
Yang dibimbing oleh Drs.Sarwono M.Pd
Oleh :
Offering C/ Kelompok 4
1. Hanifa Fitria R. (130341614781)2. Herlizza Basyarotun A. (130341614782)3. Kiki Elita S. (130341614850)4. Mayang Puspa Rena (130341614833)5. Sasty Alvionita (130341614828)6. Shinta Kumalasari (130341614836)
The Learning University
UNIVERSITAS NEGERI MALANG
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
JURUSAN BIOLOGI
NOVEMBER 2014

A. TOPIK : Perkecambahan dan Dormansi
B. TANGGAL : 1 Oktober 2014 – 29 Oktober 2014
C. TUJUAN :
1. Memahami bahwa tidak semua biji dapat langsung tumbuh apabila dikecambahkan.
2. Menduga kondisi dapat disebabkan oleh beberapa faktor baik luar maupun dalam.
3. Dormansi dapat dipecahkan dengan beberapa perlakuan
D. DATA PENGAMATAN
1. Biji Selada
a. Biji selada ( direndam dalam air )
Nama
biji
Hari ke Jumlah biji
Selada 1 29
2 37
3 37
4 39
5 39
6 40
7 40
Ulangan kedua
Nama biji Hari ke Jumlah biji
Selada 1 13
2 15
3 20
4 23
5 26
6 29
7 33

Ulangan ketiga
Nama biji Hari ke Jumlah biji
Selada 1 20
2 21
3 23
4 23
5 28
6 29
7 33
b. Biji selada ( direndam urea )
Ulangan pertama
Nama
biji
Hari ke Jumlah biji
Selada 1 0
2 44
3 50
Ulangan kedua
Nama
biji
Hari ke Jumlah biji
Selada 1 0
2 45
3 50

c. Biji Selada ( direndam 2,4 dinitrophenol )
Ulangan pertama
Nama
biji
Hari ke Jumlah biji
Selada 1 1
2 4
3 9
4 12
5 20
6 35
7 50
Ulangan kedua
Nama
biji
Hari ke Jumlah biji
Selada 1 4
2 18
3 23
4 24
5 32
6 43
7 48

2. Biji Padi
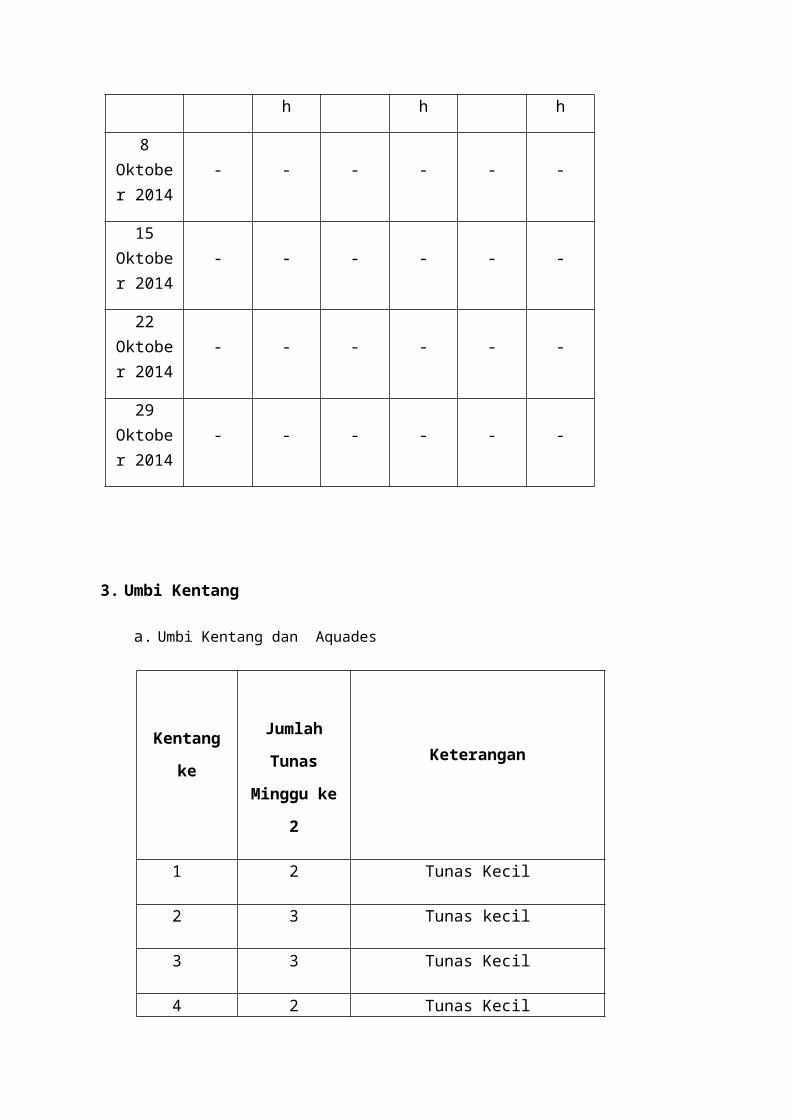
a. Biji Padi ( direndam aquades )
Tanggal
N1 N2 N3
TumbuhTidak
TumbuhBusuk Tumbuh
Tidak Tumbuh
Busuk TumbuhTidak
TumbuhBusuk
8 Oktober
201411 39 - 12 38 - 15 35 -
15 Oktober
201424 26 - 22 28 - 27 23 -
22 Oktober
201434 16 - 36 14 - 32 18 -
29 Oktober
201448 - 2 46 - 4 45 - 5
Persentase Pertumbuhan Kecambah Biji Padi
Tanggal Rata-Rata Biji yang Tumbuh Persentase Biji yang Tumbuh
8 Oktober 2014 13 25 %
15 Oktober 2014 24 48 %
22 Oktober 2014 34 68 %
29 Oktober 2014 46 92 %

b. Biji Padi ( tanpa direndam aquades )
Tanggal
N1 N2 N3
TumbuhTidak
TumbuhTumbuh
Tidak Tumbuh
TumbuhTidak
Tumbuh
8 Oktober
2014- - - - - -
15 Oktober
2014- - - - - -
22 Oktober
2014- - - - - -
29 Oktober
2014- - - - - -
3. Umbi Kentang
a. Umbi Kentang dan Aquades
Kentang ke Jumlah Tunas
Minggu ke 2
Keterangan
1 2 Tunas Kecil
2 3 Tunas kecil
3 3 Tunas Kecil
4 2 Tunas Kecil
5 1 Tunas Kecil
Kentang ke Jumlah Tunas
Minggu ke 4Keterangan
1 4 Tunas besar
2 4 Tunas Besar
3 5 Tunas besar
4 4 Tunas sedang
5 3 Tunas sedang
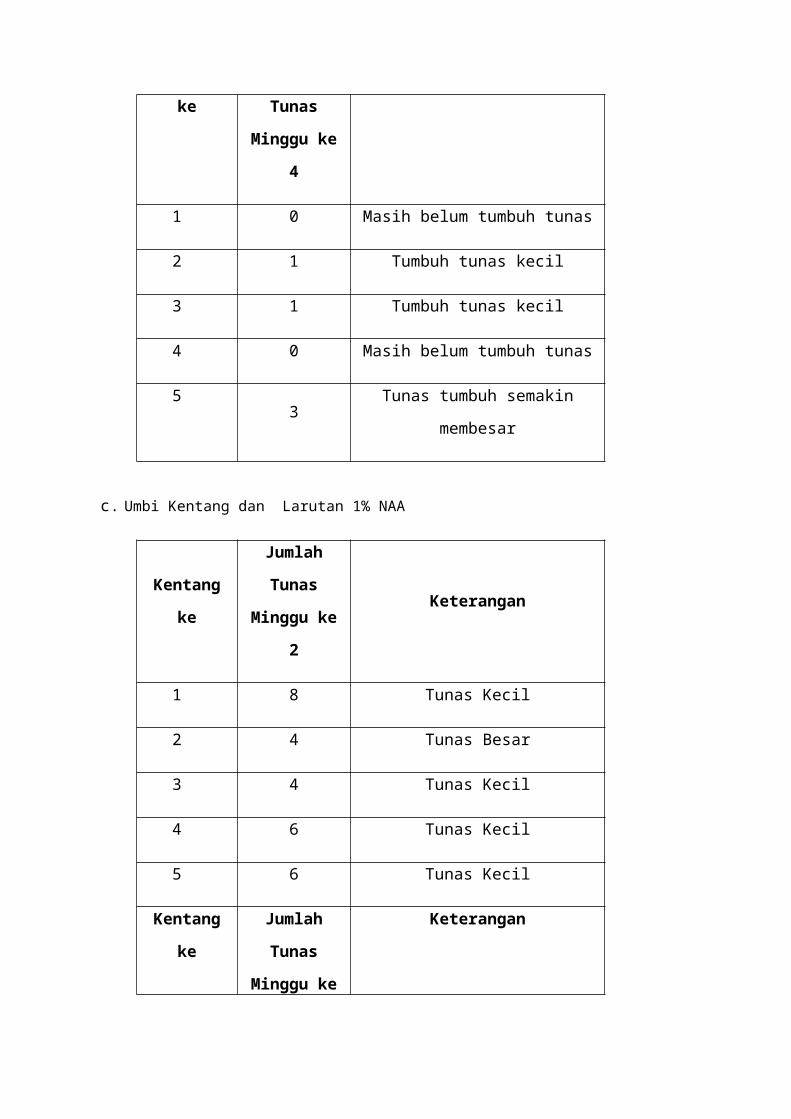
b. Umbi Kentang dan Larutan Thiourea 2%
Kentang keJumlah Tunas
Minggu ke 2Keterangan
1 0 Belum tumbuh Tunas
2 0 Belum Tumbuh Tunas
3 0 Belum Tumbuh Tunas
4 0 Belum Tumbuh Tunas
5 3 Tumbuh tunas kecil
Kentang ke Jumlah Tunas
Minggu ke 4Keterangan
1 0 Masih belum tumbuh tunas
2 1 Tumbuh tunas kecil
3 1 Tumbuh tunas kecil
4 0 Masih belum tumbuh tunas
5 3 Tunas tumbuh semakin membesar

c. Umbi Kentang dan Larutan 1% NAA
Kentang keJumlah Tunas
Minggu ke 2Keterangan
1 8 Tunas Kecil
2 4 Tunas Besar
3 4 Tunas Kecil
4 6 Tunas Kecil
5 6 Tunas Kecil
Kentang ke Jumlah Tunas
Minggu ke 4Keterangan
1 8 Tunas Kecil
2 4 Tunas Besar
3 4 Tunas Kecil
4 6 Tunas Kecil
5 6 Tunas Kecil
E. PEMBAHASAN
1. Biji Selada
Pada percobaan pertama yaitu pengaruh perendaman biji selada (lactuca sativa. L)
dalam aquades terhadap perkecambahan. Pada hari 1 ulangan pertama biji selada yang
mampu berkecambah ada 29, pada ulangan kedua 13. Pada hari 2 ulangan pertama biji
selada yang mampu berkecambah ada 37, ulangan kedua 15. Pada hari 3 ulangan
pertama biji selada yang mampu berkecambah ada 37, ulangan kedua 20. Pada hari 4
ulangan pertama biji selada yang mampu berkecambah ada 39, ulangan kedua 23. Pada
hari 5 ulangan pertama biji selada yang mampu berkecambah ada 39, pada ulangan
kedua 26. Pada hari 6 ulangan pertama biji selada yang mampu berkecambah ada 40,
ulangan kedua 29. Pada hari 7 ulangan pertama biji selada yang mampu berkecambah
ada 40, ulangan kedua 33. Pada hari pertama hingga 7 mengalami peningkatan jumlah
biji selada yang mengalami perkecambahan, hal ini disebabkan kerena aquades memiliki
fungsi sebagai berikut :
1. melunakkan kulit biji, embrio dan endosperm mengembang sehingga kulit biji
robek
2. memfasilitasi masuknya O2 kedalam biji, air imbibisi pada dinding sel sehingga
sel jadi permeabel terhadap gas. Gas masuk secara difusi sehingga suplai O2 pada
sel hidup meningkat dan pernafasan aktif
3. mengencerkan protoplasma, aktivasi macam-macam fungsinya
4. alat transport larutan makanan dari endosperm/kotiledon ketitik tumbuh di
embryonic axis : untuk membentuk protoplasma baru.
Berdasarkan analisis terdapat korelasi positif antara air yang cukup dan
perkecambahan biji , dimana tidak adanya air memicu dormansi (menghambat
perkecambahan). Normalnya, biji yang matang mengalami desikasi (kekurangan air) dari
yang asalnya 80-90 % menjadi hanya 5%. Desikasi ini diinisiasi oleh hormon asam
absisat. Biji yang kekurangan air ini akan terus dorman sampai terdapat faktor-faktor
yang memicu perkecambahan. Faktor yang memicu perkecambahan salah satunya
adalah air yang cukup untuk mengisi kembali kekurangan air pada jaringan biji
(Hopkins, 2008).

Proses rehidrasi jaringan dinamakan imbibisi. Tekanan imbibisi oleh biji yang
berkecambah mengakibatkan kulit biji pecah, sehingga embrio (radikula) bisa keluar.
Imbibisi diikuti oleh aktivasi metabolisme biji beberapa menit setelah air masuk
(Hopkins, 2008). Jadi, adanya air dapat membuat radikula dapat keluar menembus kulit
biji dan mengaktifkan metabolisme sehingga embrio dapat tumbuh. Tidak adanya air
menyebabkan biji terus mengalami desikasi dan akan tetap mengalami dormansi.
Pada percobaan kedua yaitu pengaruh berbagai zat kimia terhadap perkecambahan
biji selada (Letuca sativa. L). Perlakuan pertama adalah menggunakan 2,4 Dinitropenol,
pada hari 1 ulangan pertama biji selada yang mampu berkecambah ada 1, pada ulangan
kedua 4. Pada hari 2 ulangan pertama biji selada yang mampu berkecambah ada 4,
ulangan kedua 18. Pada hari 3 ulangan pertama biji selada yang mampu berkecambah
ada 9, ulangan kedua 23. Pada hari 4 ulangan pertama biji selada yang mampu
berkecambah ada 12, ulangan kedua 24. Pada hari 5 ulangan pertama biji selada yang
mampu berkecambah ada 20, pada ulangan kedua 32. Pada hari 6 ulangan pertama biji
selada yang mampu berkecambah ada 35, ulangan kedua 43. Pada hari 7 ulangan
pertama biji selada yang mampu berkecambah ada 50, ulangan kedua 48. Pada hari
pertama hingga 7 mengalami peningkatan jumlah biji selada yang mengalami
perkecambahan. Hal ini dikarenakan penambahan 2,4 Dinitrophenol merupakan salah
satu hormon auksin sintetik yang berfungsi merangsang pertmubuhan akar ( Cistopher,
1992 ). 2,4 2,4- dinitrophenol ( 2,4- DNP ) adalah auksin sintetis, seperti halnya 2,4-
dinitrofenol (2,4-D) sering digunakan secara meluas sebagai herbisida tumbuhan. Pada
Monocotyledoneae, misalnya : jagung dan rumput lainnya dapat dengan cepat men
ginaktifkan auksin sintetik ini, tetapi pada Dicotyledoneae tidak terjadi, bahkan
tanamannya mati karena terlalu banyak dosis hormonalnya. Menyemprot beberapa
tumbuhan serialia ataupun padang rumput dengan 2,4-D, akan mengeliminir gulma
berdaun lebar seperti dandelion. Auksin akan menstimulasi pertumbuhan hanya pada
kisaran konsentrasi tertentu; yaitu antara : 10-8 M sampai 10-4 M. Pada konsentrasi yang
lebih tinggi; auksin akan menghambat perpanjangan sel, mungkin dengan menginduksi
produksi etilen, yaitu suatu hormon yang pada umumnya berperan sebagai inhibitor pada
perpanjangan sel (George, L.W. 1995).
Auksin akan meningkatkan potensial membran (tekanan melewati membran) dan
menurunkan pH di dalam dinding sel. Pengasaman dinding sel ini, akan mengaktifkan
enzim yang disebut ekspansin; yang memecahkan ikatan hidrogen antara mikrofibril

sellulose, dan melonggarkan struktur dinding sel. Ekspansin dapat melemahkan integritas
kertas saring yang dibuat dari sellulose murni. Penambahan potensial membran, akan
meningkatkan pengambilan ion ke dalam sel, yang menyebabkan pengambilan air secara
osmosis. Pengambilan air, bersama dengan penambahan plastisitas dinding sel,
memungkinkan sel untuk memanjang. Auksin juga mengubah ekspresi gen secara cepat,
yang menyebabkan sel dalam daerah perpanjangan, memproduksi protein baru, dalam
jangka waktu beberapa menit. Beberapa protein, merupakan faktor transkripsi yang
secara menekan ataupun mengaktifkan ekspresi gen lainnya. Untuk pertumbuhan
selanjutnya, setelah dorongan awal ini, sel akan membuat lagi sitoplasma dan bahan
dinding sel. Auksin juga menstimulasi respon pertumbuhan selanjutnya (Anna Kasvaa.
2007).
Perlakuan kedua yaitu menggunakan urea, pada hari 1 ulangan pertama biji selada
tidak ada yang berkecambah, pada ulangan kedua juga tidak ada yang berkecambah.
Pada hari 2 ulangan pertama biji selada yang mampu berkecambah ada 44, ulangan
kedua 45. Pada hari 3 ulangan pertama biji selada yang mampu berkecambah ada 50 dan
ulangan kedua 50. Pupuk urea mengandung banyak sekali kandungan nitrogen, yaitu
sebanyak 46 %. Nitrogen yaitu zat yang dapat membantu pertumbuhan tumbuhan.
Pemberian pupuk urea dapat mempengaruhi sifat kimia dan hayati (biologi) tanah.
Fungsi kimia dan hayati yang penting diantaranya adalah selaku penukar ion dan
penyangga kimia, sebagai gudang hara N, P, dan S, pelarutan fosfat dengan jalan
kompleksasi ion Fe dan Al dalam tanah dan sebagai sumber energi mikroorganisme
tanah (Notohadiprawiro, 1998).
Dormansi dapat dikatakan sebagai mekanisme biologis dalam menjamin
perkecambahan biji yang berlangsung pada kondisi dan waktu yang tepat untuk
mendukung pertumbuhan yang tepat. Dormansi bisa diakibatkan karena
ketidakmampuan embrio dalam mengatasi hambatan (Dwidjoseputro, 1983). Dormansi
merupakan suatu keadaan pertumbuhan yang terhambat, dapat disebabkan oleh kondisi
yang kurang baik atau oleh faktor dari dalam tumbuhan itu sendiri. Dormansi dapat
dikatakan sebagai suatu keadaan dimana pertumbuhan tidak dapat terjadi walaupun
kondisi lingkungan mendukung terjadinya perkecambahan (Dartius, 1991).
Dormansi dapat terjadi dalam banyak tipe dan bentuk. Banyak biji dorman untuk
suatu periode tertentu setelah keluar dari buah. Contoh lain dari dormansi adalah

gugurnya daun untuk menghindari terjadinya bahaya waktu udara berubah menjadi
dingin ataupun kemarau. Tanaman bagian atas banyak yang mati selama periode musim
dingin atau kekeringan (Filter & Hay, 1991).
Dormansi benih berhubungan dengan usaha benih untuk menunda perkecambahan,
sehingga waktu dan kondisi lingkungan memungkinkan melangsungkan proses tersebut.
Dormansi dapat terjadi pada kulit biji maupun pada embrio. Biji yang telah masak dan
siap untuk berkecambah membutuhkan kondisi klimatik dan tempat tumbuh yang sesuai
untuk dapat mematahkan dormansi dan memulai proses perkecambahannya. Teknik
skarifikasi, biasa digunakan untuk mematahkan dormansi kulit biji, sedangkan
stratifikasi digunakan dalam mengatasi dormansi embrio (Heddy, 1990).
Dalam Lakitan ( 2007 ) dormansi biji dapat diklasifikasikan berdasarkan bentuk
dormansinyai, yaitu :
a. Dormansi akibat kulit biji impermeabel terhadap air (H2O)
b. Dormansi disebabkan embrio belum masak
c. Biji membutuhkan pemasakan sempuna, sehingga setelah panen dormansi terjadi
dalam penyimpanan kering
d. Biji membutuhkan suhu rendah
e. Biji bersifat sensitif terhadap cahaya
f. Kuantitas cahaya
g. Kualitas cahaya
h. Adanya zat kimia sehingga terjadi dormansi biji
Gejala morfologi dengan pemunculan radikula tersebut, terjadi proses fisiologi-
biokemis yang kompleks, yang dikenal dengan perkecambahan fisiologis. Secara
fisiologis, proses perkecambahan berlangsung dalam beberapa tahapan penting, meliputi
(Sitompul & Guritno, 1995):
a. Absorpsi air
b. Metabolisme pemecahan materi cadanagan makanan
c. Transport materi hasil pemecahan dari endosperm ke embrio yang aktif bertumbuh.
d. Proses-proses pembentukan kembali materi-materi baru.
e. Respirasi dan pertumbuhan
Faktor yang mengontrol proses perkecambahan biji, dapat dibedakan secara
internal dan eksternal. Faktor internal, perkecambahan biji ditentukan oleh keseimbangan
antara promotor dan inhibitor perkecambahan, terutama giberelin (GA) dan asam absisat

(ABA). Faktor eksternal meliputi faktor ekologi yaitu air, suhu, kelembapan, cahaya dan
senyawa-senyawa kimia yang mendukung perkecambahan (Tjitrosoepomo, 1998).
2. Biji Padi
Biji merupakan bagian alat reproduksi. Perkecambahan termasuk proses dimana
dimulainya dengan proses imbibisi air oleh dorman, biasanya kering, biji dan berakhir dengan
proses elongasi dari axis embrionik (H. Lambers et al., 2008). Biji memiliki cadangan
makanan yang membuatnya independen secara luas dari sumber daya lingkungan untuk
bertahan hidup. Perubahan drastis tersebut dalam proses autotropik yang bergantung kepada
suplai cahaya, CO2, air dan nutrisi anorganik dari sekelilingnya untuk pertumbuhan
autotropik. Perkecambahan adalah proses ketika bagian dari embrio, biasanya radikula,
memasuki kulit biji dan mungkin berproses dengan air dan O2 dan pada temperatur yang
stabil.
Dormansi dapat didefiniskan suatu keadaan pertumbuhan dan metabolisme yang
terpendam, dapat disebabkan oleh kondisi lingkungan yang tidak baik atau oleh faktor dari
dalam tumbuhan itu sendiri. Dormansi merupakan suatu mekanisme mempertahankan diri
terhadap suhu yang sangat rendah atau kekeringan di musin panas. Kurangnya air penting
dalam memulai dormansi untuk mempertahankan hidup pada keadaan kering dan panas,
selanjutnya berkurangnya nutrien terutama nitrogen, dapat merupakan penyebab terjadinya
dormansi pada beberapa tumbuhan ( Dahlia, dkk, 2001 ).
Pada percobaan ini bertujuan untuk memecah dormansi pada biji padi dengan
perlakuan perendaman biji padi dengan aquades dengan 3 pengulangan. Pertama biji padi
direndam dalam air selama 1 jam, kemudian menyediakan 6 cawan petri yang telah dialasi
kertas saring untuk 2 perlakuan, pertama 3 pengulangan direndam dengan aquades dan 3
pengulangan tanpa direndam aquades. Setiap pengulangan diletakkan dalam cawan petri
dengan memasukkan 50 butir biji padi. Kemudian cawan petri ditutup dengan meletakkannya
dalam tempat yang gelap.
Pada perlakuan pertama, biji padi direndam dengan aquades. Diamati setiap minggu
perkecambahan biji padi tersebut. Hasilnya pada minggu pertama ( 8 Oktober 2014 )
perendaman diperoleh hasil pada pengulangan 1 ( N1 ), 11 biji padi yang tumbuh dan 39 biji
padi yang tidak tumbuh. Pada pengulangan 2 ( N2 ), 12 biji padi tumbuh dan 38 biji padi
yang tidak tumbuh. Pada pengulangan 3 ( N3 ), 15 biji padi tumbuh dan 35 biji padi yang
tidak tumbuh. Sehingga didapatkan rata-rata biji yang tumbuh pada minggu pertama adalah

13 biji dengan presentase 26 % dari keseluruhan biji yang direndam. Pada minggu kedua ( 15
Oktober 2014 ) perendaman diperoleh hasil pada pengulangan 1 ( N1 ), 24 biji padi yang
tumbuh dan 26 biji padi yang tidak tumbuh. Pada pengulangan 2 ( N2 ), 22 biji padi tumbuh
dan 28 biji padi yang tidak tumbuh. Pada pengulangan 3 ( N3 ), 27 biji padi tumbuh dan 23
biji padi yang tidak tumbuh. Sehingga didapatkan rata-rata biji yang tumbuh pada minggu
kedua adalah 24 biji dengan presentase 48 % dari keseluruhan biji yang direndam. Pada
minggu ketiga ( 22 Oktober 2014 ) perendaman diperoleh hasil pada pengulangan 1 ( N1 ),
34 biji padi yang tumbuh dan 16 biji padi yang tidak tumbuh. Pada pengulangan 2 ( N2 ), 36
biji padi tumbuh dan 14 biji padi yang tidak tumbuh. Pada pengulangan 3 ( N3 ), 32 biji padi
tumbuh dan 18 biji padi yang tidak tumbuh. Sehingga didapatkan rata-rata biji yang tumbuh
pada minggu ketiga adalah 34 biji dengan presentase 68 % dari keseluruhan biji yang
direndam. Pada minggu keempat ( 29 Oktober 2014 ) perendaman diperoleh hasil pada
pengulangan 1 ( N1 ), 48 biji padi yang tumbuh dan 2 biji padi busuk. Pada pengulangan 2
( N2 ), 46 biji padi tumbuh dan 4 biji padi yang busuk. Pada pengulangan 3 ( N3 ), 45 biji
padi tumbuh dan 5 biji padi yang busuk. Sehingga didapatkan rata-rata biji yang tumbuh pada
minggu keempat adalah 46 biji dengan presentase 92 % dari keseluruhan biji yang direndam.
Pada perlakuan kedua, biji padi yang sudah direndam selama 1 jam kemudian dimasukkan ke
dalam cawan petri dengan tanpa perendaman lagi. Hasilnya diperoleh, tidak ada biji padi
yang tumbuh atau berkecambah.
Dalam percobaan pada biji padi yang direndam dengan air dapat tumbuh hingga 92 %.
Hal ini membuktikan bahwa dormansi pada biji padi dapat dipatahkan oleh perlakuan
perendaman dengan aquades. Hal ini sesuai dengan Sutopo ( 1988 ), bahwa faktor–faktor
yang meyebabkan hilangnya dorminasi pada benih sangat bervariasi tergantung pada jenis
tanaman dan tentu saja tipe dormansinya, antara lain: karena temperatur, hilangnya
kemampuan untuk menghasilkan zat – zat penghambat perkecambahan, dan imbibisi yang
terjadi pada biji saat direndam. Selain itu terdapat biji padi yang membusuk saat direndam hal
ini disebabkan karena terlalu banyaknya air yang diserap oleh biji. Sedangkan pada biji padi
yang tidak direndam tidak terjadi pertumbuhan atau perkecambahan dari biji padi, hal ini
karena tidak ada faktor yang mematahkan dormansi seperti yang disebutkan dalam Sutopo
(1988) faktor–faktor yang meyebabkan hilangnya dorminasi pada benih sangat bervariasi
tergantung pada jenis tanaman dan tentu saja tipe dormansinya, antara lain: karena
temperatur, hilangnya kemampuan untuk menghasilkan zat–zat penghambat perkecambahan,
dan imbibisi yang terjadi pada biji saat direndam.

Sedangkan tujuan dari penyimpanan ditempat gelap adalah untuk mempercepat proses
perkecambahan dari biji. Sehingga hubungan antara dormansi dengan penyimpanan yaitu
dormansi pada beberapa spesies dapat menghilang apabila disimpan dalam kondisi suhu dan
kelembaban lingkungan yang terkendali. Tempat yang gelap umumnya memiliki tingkat
kelembapan yang tinggi. Ahli fisiologi benih paham benar akan metode-metode terbaik yang
digunakan untuk mematahkan dormansi pada benih yaitu dengan jalan menyimpan pada suhu
lembab ( Jumin, 2002 ).
Dalam perkecambahan biji padi ini memerlukan waktu yang cukup lama. Karena
struktur dari biji padi yang keras. Biji yang mengalami dormansi biasanya disebabkan oleh
rendahnya / tidak adanya proses imbibisi air yang disebabkan oleh struktur benih (kulit
benih) yang keras, sehingga mempersulit keluar masuknya air kedalam benih. Respirasi yang
tertukar, karena adanya membrane atau pericarp dalam kulit benih yang terlalu keras,
sehingga pertukaran udara dalam benih menjadi terhambat dan menyebabkan rendahnya
proses metabolisme dan mobilisasi cadangan makanan dalam benih. Kulit biji yang keras dan
kedap menjadi penghalang mekanis terhadap masuknya air atau gas-gas kedalam bij, yang
menyebabkan impermeabilitas kulit biji terhadap air. Kulit biji yang cukup kuat akan
menghalangi pertumbuhan embrio. Jika kulit biji dihilangkan, maka embrio akan tumbuh
dengan segera. Pada tanama, dormansi sering dijumpai pada benih padi (Salisbury, 1995).
3. Umbi Kentang
Dormansi dapat didefinisikan sebagai suatu keadaan pertumbuhan dan metabolisme
yang terpendam, dapat disebabkan oleh kondisi lingkungan yang tidak baik atau oleh
faktor dari dalam tumbuhan itu sendiri (Sasmitamihardja, 1990). Sedangkan
Perkecambahan merupakan proses pertumbuhan dan perkembangan embrio. Hasil
perkecambahan ini adalah munculnya tumbuhan kecil dari dalam biji. Proses perubahan
embrio saat perkecambahan adalah plumula tumbuh dan berkembang menjadi batang, dan
radikula tumbuh dan berkembang menjadi akar (Syamsuri, 2004).
Pada praktikum ini kami menggunakan 15 umbi kentang (Solanum tuberosum L.),
aquadest, larutan 2% thiourea dan larutan 1% NAA. Menurut Soegihartono (2008 Dalam
Ratnasari, 2010) Solanum tuberosum L. atau yang dikenal dengan kentang merupakan
satu dari lima makanan pokok dunia sebagai sumber karbohidrat. Kelima makanan pokok
tersebut adalah beras, gandum, kentang, sorgum, dan jagung. Disamping beras sebagai

bahan pangan utama, kentang merupakan komoditas pangan yang penting di Indonesia
dan dibutuhkan sepanjang tahun.
Kentang (Solanum tuberosum L.) adalah tanaman dari suku Solanaceae yang
memiliki umbi batang yang dapat dimakan dan disebut "kentang" pula. Tanaman ini
berasal dari daerah subtropika, yaitu dataran tinggi Andes Amerika Utara. Daerah yang
cocok untuk budi daya kentang adalah dataran tinggi atau pegunungan dengan ketinggian
1.000-1.300 meter di atas permukaan laut, curah hujan 1.500 mm per tahun, suhu rata-rata
harian 18-21oC, serta kelembaban udara 80-90 persen (Gklinis, 2009 Dalam Ratnasari,
2010).
Perlakuan yang dilakukan adalah sebagai berikut 5 buah umbi kentang dimasukkan
ke dalam kantung plastik kemudian diberi aquades, 5 umbi kentang lainnya diberi larutan
2% thiourea dan larutan 1% NAA. Umbi umbi kentang direndam dalam larutan tersebut
selama 2 jam dan kemudian dikeluarkan dari kantung plastik di anginkan baru setelah itu
dibungkus rapat menggunakan kertas. Pengamatan dilakukan setiap 2 seminggu sekali
selama 4 minggu.
Perlakuan pertama adalah umbi kentang direndam dalam aquades, kemudian
dibungkus dengan kertas, pengamatan pertama dilakukan saat minggu kedua dan
pengamatan kedua saat minggu ke 4. Pada pengamatan pertama kentang 1 menunjukkan
pertumbuhan 2 tunas kecil, sedangkan pada pengamatan kedua tunas bertambah menjadi
4 dan ukurannya bertambah besar. Pada pengamatan pertama kentang ke 2 jumlah
tunasnya 3 dan ukuran tunas kecil, sedangkan pada pengamatan kedua diamati ada 4
tunas dengan ukuran yang lebih besar. Pada pengamatan pertama kentang ke 3 muncul
3 tunas dengan ukuran yang kecil, kemudian setelah diamati pertumbuhan tunas pada
pengamatan kedua ukurannya lebih besar dan jumlah tunas sebanyak 5. Pada pengamatan
pertama kentang ke 4 tunas muncul sebanyak 2 dengan ukuran yang relatif kecil,
kemudian pada pengamatan kedua jumlah tunas menjadi 4 dengan ukuran tunas yang
tidak besar dan tidak kecil (sedang). Sedangkan pada pengamatan pertama kentang ke 5
jumlah tunas hanya 1 dengan ukuran yang kecil, pengamatan kedua menunjukkan
perubahan jumlah tunas menjadi 3 dengan ukuran sedang. Dari hasil pengamatan
menunjukkan bahwa umbi kentang mengalami pertumbuhan, ditandai dengan
bertambahnya jumlah tunas dan ukuran tunas yang bertambah lebih besar.

Perlakuan merendam biji di dalam air yaitu mengkondisikan daerah di luar biji
potensial airnya tinggi, sedangkan potensial air di dalam biji sendiri rendah. Maka akan
terjadi perpindahan osmosis dari potensial air tinggi ke potensial rendah. Perpindahan ini
akan mengakibatkan lapisan kulit biji yang bersifat keras akan lembek, sehingga yang
semula biji tidak bisa berkecambah akibat terhalang lapisan kulit biji yang keras akan bisa
melakukan fase differensiasi dan fase tumbuh (Campbell, 2002).
Perkecambahan biji tergantung pada imbibisi, penyerapan air akibat potensial air
yang rendah pada biji yang kering. Air yang berimbibisi menyebabkan biji mengembang
dan memecahkan kulit pembungkusnya dan juga memicu perubahan metabolik pada
embrio yang menyebabkan biji tersebut melanjutkan pertumbuhan. Enzim-enzim akan
mulai mencerna bahan-bahan yang disimpan pada endosperma atau kotiledon, dan nutrien-
nutriennya dipindahkan ke bagian embrio yang sedang tumbuh (Campbell, 2002). Dari
teori tersebut dapat diketahui bahwa masuknya air akibat dari perendaman umbi kentang
dalam aquades menyebabkan rusaknya kulit biji sehingga memicu perubahan metabolik,
hal ini menyebabkan pertumbuhan pada embrio yang ditandai dengan munculnya tunas.
Pelakuan kedua dengan merendam umbi kentang didalam larutan thiourea 2%,
seperti pada perlakuan pertama setelah umbi kentang direndam kemudian umbi kentang
dibungkus menggunakan kertas. Pengamatan pertama dilakukan pada minggu ke 2 dan
pengamatan kedua dilakukan pada minggu ke 4. Pada pengamatan pertama umbi kentang
ke 1 menunjukkan tidak munculnya tunas, dan pada pengamatan kedua tunas juga masih
belum tumbuh. Pengataman pertama umbi kentang ke 2 belum ada tunas yang tumbuh,
sedangkan pada pengamatan kedua tunas umbi kentang mulai tumbuh 1 tunas kecil. Pada
pengamatan pertama umbi kentang ke 3 tidak ada tunas yang tumbuh, sedangkan
pengataman kedua umbi kentang menunjukkan ada pertumbuhan 1 tunas yang kecil.
Pengamatan pertama umbi kentang ke 4 menunjukkan belum ada pertumbuhan umbi
kentang yang ditandai tidak munculnya tunas, pada pengamatan kedua juga masih belum
ada pertumbuhan karena masih belum tumbuh tunas. Sedangkan pada pengamatan
pertama umbi kentang ke 5 muncul 3 tunas dengan ukuran yang kecil dan pada
pengamatan kedua menunjukkan bahwa tidak ada pertambahan jumlah tunas tapi ada
perubahan ukuran tunas umbi kentang menjadi lebih besar.
Pada penggunaan thiourea diketahui bahwa umbi kentang ke 1 dan ke 4 tidak
menunjukkan perubahan apapun, atau bisa disebut tidak adanya pertumbuhan pada

umbi tersebut. Menurut Haber (1960) Thiourea menghambat mitosis pada percobaan
ini meskipun merangsang perkecambahan, kami menyimpulkan thiourea adalah
inhibitor mitosis dalam biji karena mitosis tidak menyebabkan pertumbuhan dan
perkembangan sel. Dari pernyataan tersebut meskipun thiourea menyebabkan
perkecambahan tetapi thiourea juga menghambat proses mitosis sehingga
perkecambahan yang terjadi tidak begitu maksimal.
Perlakuan ketiga sama dengan perlakuan pertama dan kedua hanya beda pada
larutan yang digunakan, pada perlakuan ketiga larutan yang digunakan adalah larutan
1% NAA. NAA (napthalene acetic acid) merupakan auksin sintetik (Dewi, 2008).
Pengamatan pertama umbi kentang ke 1 menunjukkan adanya pertumbuhan 8 tunas
dengan ukuran kecil, sedangkan pada pengamatan kedua jumlah tunas umbi kentang
tidak berubah dan ukurannya masih sama. Pengamatan pertama umbi kentang ke 2
menunjukkan muncul 4 tunas dengan ukuran yang besar, dan pada pengamatan kedua
hasil pengamatan kedua sama dengan pengamatan pertama yaitu tunas umbi umbi
kentang yang muncul ada 4 dengan ukuran tunas yang besar. Pengamatan pertama
umbi kentang ke 3 muncul 4 tunas dengan ukuran yang kecil, pada pengamatan kedua
tidak ada perubahan pada tunas umbi kentang.,Pengamatan pertama umbi kentang ke 4
muncul 6 tunas dengan ukuran yang kecil, pada pengamatan kedua masih sama dengan
pengamatan pertama muncul 4 tunas kecil. Sedangkan pada pengamatan pertama umbi
kentang ke 5 menunjukan munculnya 6 tunas dengan ukuran yang kecil, dan saat
pengamatan kedua dilakukan tidak ada perubahan pada umbi kentang tunas yang
muncul masih 6 dan ukurannya masih kecil.
Dari ketiga perendaman yang digunakan dalam praktikum ini NAA sangat
efektif dalam memecahkan dormansi dalam hal ini diketahui banyaknya tunas yang
tumbuh setelah direndam dalam larutan 1% NAA. Tetapi data pengamatan
menunjukkan bahwa tidak ada perubahan baik jumlah maupun ukuran pada
pengamatan pertama dan kedua. Hal ini kemungkinan ada satu zat yang perlu
menunjang pemecahan dormansi selain menggunakan NAA, hal ini merujuk pada
pernyataan Santoso (2003) yang menyatakan bahwa zat pengatur tumbuh adalah
senyawa organik ataupun anorganik yang hanya dibutuhkan tanaman dalam
konsentrasi yang sangat sedikit. Zat pengatur tumbuh yang sering digunakan untuk
menginduksi pertumbuhan pada teknik mikropropagasi adalah kombinasi golongan

auksin dan sitokinin dimana pada penelitian ini jenis yang digunakan adalah NAA
yang dikombinasikan dengan BAP.
Pada praktikum ini ditemui adanya beberapa ganjalan seperti ada umbi
kentang yang tidak tumbuh dan pertumbuhan pada umbi yang tidak terlihat nyata
seperti tidak bertambahnya jumlah tunas dan ukuran tunas. Kami memperkirakan hal
ini disebabkan karena bebapa hal :
a. Umbi kentang yang digunakan belum memenuhi kematangan yang sesuai, hal ini
bisa dilihat dengan ukuran umbi kentang yang relatif masih kecil.
b. Berat benih dalam hal ini merujuk pada pernyataan Sutopo (2002) yang
menyatakan bahwa Berat benih berpengaruh terhadap kecepatan pertumbuhan
dan produksi karena berat benih menentukan besarnya kecambah pada saat
permulaan dan berat tanaman pada saat dipanen
c. Adanya zat penghambat (inhibitor) Menurut Kuswanto (1996), penghambat
perkecambahan benih dapat berupa kehadiran inhibitor baik dalam benih maupun
di permukaan benih, adanya larutan dengan nilai osmotik yang tinggi serta bahan
yang menghambat lintasan metabolik atau menghambat laju respirasi.
d. Suhu saat perkecambahan yang kurang optimal. Menurut Sutopo (2002) Suhu
optimal adalah yang paling menguntungkan berlangsungnya perkecambahan
benih dimana presentase perkembangan tertinggi dapat dicapai yaitu pada kisaran
suhu antara 26.5 sd 35°C
e. Oksigen, Menurut Sutopo (200) Saat berlangsungnya perkecambahan, proses
respirasi akan meningkat disertai dengan meningkatnya pengambilan oksigen dan
pelepasan CO2, air dan energi panas. Terbatasnya oksigen yang dapat dipakai
akan menghambat proses perkecambahan benih. Pada perkecambahan karena
pada waktu mematahkan dormansi umbi kentang dibungkus dengan kertas
sehingga kemungkinan pertumbuhan tunas umbi kentang menjadi terhambat.
f. Cahaya, menurut Kamil (1979) Adapun besar pengaruh cahanya terhadap
perkecambahan tergantung pada intensitas cahaya, kualitas cahaya, lamanya
penyinaran
g. Medium, Medium yang baik untuk perkecambahan haruslah memiliki sifat fisik
yang baik, gembur, mempunyai kemampuan menyerap air dan bebas dari
organisme penyebab penyakit terutama cendawan (Sutopo, 2002).

F. KESIMPULAN
Dormansi dapat didefinisikan sebagai suatu keadaan pertumbuhan dan metabolisme
yang terpendam, dapat disebabkan oleh kondisi lingkungan yang tidak baik atau oleh
faktor dari dalam tumbuhan itu sendiri.
Perkecambahan biji tergantung pada imbibisi, penyerapan air akibat potensial air yang
rendah pada biji yang kering
Air yang berimbibisi menyebabkan biji mengembang dan memecahkan kulit
pembungkusnya dan juga memicu perubahan metabolik pada embrio yang
menyebabkan biji tersebut berkecambah.
Dormansi dapat dipatahkan dengan memberikan perlakuan perendaman biji dengan
aquades atau dengan larutan kimia lainnya ( larutan thiourea, larutan urea).
Tanda dormansi dapat dipatahkan dengan tumbuhnya kecambah pada biji yang
diberikan perlakuan.
Pematahan dormansi pada biji padi lebih lama dibandingkan dengan pematahan
dormansi pada biji selada, hal ini disebabkan struktur dari kulit biji padi lebih keras.
Pada biji padi mengalami dormansi disebabkan oleh rendahnya / tidak adanya proses
imbibisi air yang disebabkan oleh struktur benih (kulit benih) yang keras, sehingga
mempersulit keluar masuknya air kedalam benih.
Faktor yang mengontrol proses perkecambahan biji yaitu, faktor internal meliputi
perkecambahan biji ditentukan oleh keseimbangan antara promotor dan inhibitor
perkecambahan, terutama giberelin (GA) dan asam absisat (ABA) dan faktor eksternal
meliputi faktor ekologi yaitu air, suhu, kelembapan, cahaya dan senyawa-senyawa
kimia yang mendukung perkecambahan
Pada perendaman umbi kentang dengan larutan thiourea. Thiourea menghambat
mitosis pada percobaan ini meskipun merangsang perkecambahan, kami
menyimpulkan thiourea adalah inhibitor mitosis sehingga perkecambahan tidak
maksimal.
Pada perendaman umbi kentang dengan aquades, larutan Thiourea, dan larutan NAA.
NAA sangat efektif dalam memecahkan dormansi dalam hal ini diketahui banyaknya
tunas yang tumbuh setelah direndam dalam larutan 1% NAA.

DAFTAR RUJUKAN
Anna Kasvaa. 2007. The growth enhancing effects of triacontanol. Online (htttp://www.carbonkick.fi) Diakses tanggal 11 November 2014.
Campbell, Neil A. 2002. Biologi. Jakarta : Erlangga
Cistopher, T. K . H. 1992. Pengenalan teknologi kultur tisu tumbuhan. Pusat kajian sains Hayat. Universiti Sains Malaysia.
Dahlia, Betty Lukiaty, Leily T. Kusumaputri. 2001. Petunjuk Praktikum Fisiologi Tumbuhan. FMIPA : Universitas Negeri Malang. Malang
Dartius. 1991. Dasar-dasar Fisiologi Tumbuhan. USU-Press. Medan.
Dewi, Intan Ratna. 2008. Peranan Dan Fungsi Fitohormon Bagi Pertumbuhan Tanaman. Bandung : Universitas Padjajaran
Dwijoseputro, D. 1983. Pengantar Fisiologi Tumbuhan. Gramedia. Jakarta.
Filter, A. H. dan R. K. M. Hay. 1991. Fisiologi Lingkungan Tanaman. UGM Press. Yogyakarta.
George, L.W. 1995. Teknik Kultur In Vitro dalam Holtikultura. Penebar Swadaya. Jakarta
Haber, Alan H. & Luippold, Helen J. 1960. Effects Of Gibberellin, Kinetin, Thiourea, And Photomorphogenic Radiation On Mitotic Activity In Dormant Lettuce Seed. (Online) Http://Www.Ncbi.Nlm.Nih.Gov/Pmc/Articles/Pmc405991/Pdf/Plntphys00473-0074.Pdf Diakses Pada 10 November 2014
Heddy, S. 1990. Biologi Pertanian. Rajawali Press. Jakarta.
Hopkins, William. 2008. Introduction to Plant Physiology Fourth Edition. London
Jumin, Hasan Basri. 2002. Dasar-Dasar Agronomi. Edisi Revisi. PT RajaGrafindo Persada, Jakarta.
Kamil. 1979. Teknologi Benih 1. Padang : Angkasa Raya
Kuswanto H. 1996. Dasar-Dasar Teknologi Produksi Dan Sertifikasi Benih. Yogyakarta : Andi
Lakitan, B. 2007. Dasar-Dasar Fisiologi Tumbuhan. Raja Grafindo Persada. Jakarta.
Lambers, H; Chapin III S. F. And Pons L.T.2008. Plant Physiology Ecology 2rd Edition. 2008 Springer Science and Business Media, LLC. Shanmugavalli, M; Renganayaki, PR; Menka, C. Seed Dormancy and Germination Improvement Treatment in Fodder Sorghum. An open access journal published by ICRISAT. Seed Germination and Dormancy
Notohadiprawiro, T. 1998. Tanah dan Lingkungan. Dirjen Pendidikan Tinggi. Depdikbud. Jakarta.

Ratnasari, Tuti. 2010. Kajian Pembelahan Umbi Benih Dan Perendaman Dalam Giberelin Pada Pertumbuhan Dan Hasil Tanaman Kentang (Universitas Sebelas Maret (Solanum Tuberosum L.). Solo : Universitas Sebelas Maret
Salisbury.1995. Fisiologi Tumbuhan jilid 3. Bandung : ITB.Santoso, U. Dan Nursandi, F. 2003. Kultur Jaringan Tanaman. Malang : Universitas
Muhammadiyah Malang
Sasmitamihardja, dkk., 1990, Fisiologi Tumbuhan, Departemen Pendidikan dan Kebudayaan, FMIPA-ITB, Bandung.
Sitompul, S. M. dan Guritno. B. 1995. Pertumbuhan Tanaman. UGM Press. Yogyakarta.
Sutopo L. 1988. Teknologi Benih. Jakarta : Rajawali
Sutopo, L. 2002. Teknologi Benih. Jakarta : PT. Raja Grafindo Persada.
Syamsuri, Istamar. 2004. Biologi. Jakarta : Erlangga
Tjitrosoepomo, H.S. 1998. Botani Umum. UGM Press. Yogyakarta.





























