Embed Size (px)
Citation preview

Anatomical and Functional Specializations of the Human Gluteus Maximus 1
JACK T . STERN, JR. Department of Anatomy, University of Chicago, Chicago, Illinois 60637
K E Y WORDS Primates . Evolution . Morphology . Muscles . Locomotion.
ABSTRACT Many anthropologists and anatomists have claimed that the human gluteus maximus is a functionally and structurally unique muscle, but there is not agreement on the actual characteristics of the muscle which do distinguish man from other primates. In this paper the superficial gluteus in a wide range of primates is discussed and those traits entirely unique to man are identified. The morphological specializations of the human gluteus maxi- mus are confined to its cranial portion. This part is thicker in man than in any other primate; i t has a new and firm origin, a modified ascending tendon of insertion, and an additional new insertion into the overlying fascia lata. Such changes improve the ability of the gluteus maximus to participate in controlling lateral stability of the trunk, and it suggested that this is the function which has been selected for in human evolution.
Scarcely an author exists who has writ- ten on the hip muscles of primates and failed to place great emphasis on the unique nature of the human gluteus max- imus. This muscle is considered by most to be a hallmark of bipedality. It was noted very early that the most sperficial of the gluteal muscles was relatively much larger in man than in non-human pri- mates, and there is frequent comment that the adjective “maximus” can be ac- curately applied to the muscle in no pri- mate other than man. However, the bulk of the human superficial gluteus, as in- dicative as i t is of uniqueness, should mainly provide us with the incentive to investigate the situation more closely. This has been done from time to time, with the surprising result that there is neither a general consensus on how the function of this muscle differs between man and the lower primates nor even an accurate description of those structural characteristics of the muscle which are distinctly human.
Our story begins, as do so many in com- parative primate morphology, with Edward Tyson (1699). In the first authenticated dissection of an anthropoid ape, the chim- panzee, he observed (p. 89):
The muscles of no part disagreed so much from those in Men, as those of the T h i g h of this Animnl . Here was 110 Glutezis mi- no,-; nor did the Glziteus mas irnus resem- ble the Humane: I t was merely Tendinous at its Origin, from the whole Spine of the 0 s I l ium; it was much longer, and not so thick a s i n Man.
Tyson did not speculate on the func- tional significance of these differences; indeed, he believed that his “pygmie” was probably a biped in its natural environ- ment. Over a century later, Trail1 (1818) agreed that the glutei of the chimpanzee differed much in form from those in man. The externus was very thin and did not come as far forward, nor did it arise from so large a portion of the ilium.
Cuvier (1835) introduced a functional explanation for the differences between man and non-human primates with regard to the superficial gluteus. He attributed the considerable size and strength of the muscle in man to the need for preventing flexion of the trunk under its own weight during standing or during the stance phase of walking, However, Cuvier called attention to the fact that the gluteus
1 Several of the author’s ideas discussed in this paper were presented to the American Association of Anatomists (Chicago, ’ 70 ) and appear in abstract form in the Anat. Rec., 166: 384, 1970.
A M . J. PHYS. ANTHROP., 36; 315-340. 31 5

316 JACK T. STERN, JR.
superficialis of “singes” has an extensive femoral insertion compared to that of man, and he concluded that, as a result of this distal attachment, the muscle perhaps has an action as powerful as in man. Later, Laurillard (Cuvier and Laurillard, 1849) expanded on his colleague’s spec- ulation by stating that a long femoral in- sertion of the gluteus superficialis, as seen in the black lemur, endows this ani- mal with a remarkable agility for leaping.
Burdach (1838) arrived at quite a dif- ferent assessment based on a comparison of man to Old World monkeys. He sug- gested that the action of the gluteal muscles was not the same in these two forms. Burdach emphasized the ability of the human gluteal musculature to rotate the thigh, an action he believed to be very important in bipedal standing and walk- ing. It was his opinion that in monkeys retraction, not rotation, is effected by the gluteal muscles.
In what may be considered the first primarily functional investigation of pri- mate myology, Wilder (1861) discounted the importance of the differences in size of the glutei between man and the Quad- rumana. The real distinction, he asserted, is in the position of these muscles. In man the gluteus maximus does not arise from the ischium and lies equally above and below the center of motion of the hip. In the Quadrumana the muscle arises chiefly from the ischial tuberosities (as we shall see, this is true only in some apes) and inserts much lower on the fe- mur. Although in both forms the super- ficial gluteus is an extensor of the thigh, only in man is it able to bring the lower limb in line with the trunk and can there- fore act with the gluteus medius and psoas “in preserving the balance forward and backward, which is also required in ordinary progression” (p. 372). In the ape, suggested Wilder, this muscle is a power- ful retractor of the whole limb and acts with the gluteus medius in leaping. Alone, the gluteus superficialis would rotate the thigh outward so as to turn the sole of the foot inward, after which it would as- sist the adductors in keeping the foot close against the object grasped in climb- ing.
Both Bischoff (1870, 1880) and Prim-
earlier authors that the giuteus super- ficidis of apes is a powerful extensor of the thigh, but instead of postulating that this is most functional in leaping, they emphasized a role for the muscle in pro- ducing retraction of the limb in climbing. Wiedersheim (1895), though acknowledg- ing the large size and great strength of the human gluteus maximus, agreed with Cuvier and Wilder that the muscle served mainly to steady the trunk on the heads of the femora.
In general, the authors of the nine- teenth century saw the superficial glu- teus as an extensor in both man and non-human primates, the distinction being that in man the gluteus maximus plays an essential role in antero-posterior bal- ance, whereas in other primates the mus- cle functions chiefly in either the leaping or climbing modes of locomotion.
An entirely different approach to the problem was initiated by Klaatsch (‘13). He proposed that the gluteus maximus is a climbing-muscle of the first order, but only in the human manner, this being climbing large vertical tree trunks. Klaatsch believed that such a manner of climbing was preadaptive to erect bipedal- ity. The suggestion that the gluteus max- imus be considered a uniquely powerful extensor in man carrried over to Water- man (‘29). She concluded that the new and firm attachment to the ilium, taken in connection with the sacral origin, en- ables the muscle to act with power in lifting the trunk from the stooping posi- tion. According to Waterman, in monkeys and apes the superficial gluteus is an abductor and relatively much weaker. From Boyer (‘35) came the statement that in the orang-utan the primary action of the gluteus superficialis is abduction, not extension as in man. The trend toward asserting that extension of the thigh is a special function in man continued with Kleinschmidt (‘49, ’51) and reached its culmination with Washburn (‘51). Accord- ing to Washburn, the critical changes in the evolution of erect posture were those in the ilium and gluteus maximus which converted the latter from an abductor in apes to a muscle which is able to complete the powerful extension of the hindlimb necessary in walking (the emphasis on
rose (1898-1899). supported the views of completing extension harkens- back to

SPECIALIZATIONS OF HUMAN GLUTEUS MAXIMUS 317
Wilder). This view has been widely adopted (Tappen, ’55; Breitinger, ’59; Robinson, ’63; Campbell, ’66; Snyder, ’67; Le Gros Clark, ’67).
Howell (‘59) agreed that the superficial gluteus has evolved into a powerful ex- tensor in man, but he suggested that in apes and monkeys it functioned in lateral rotation (a view exactly opposite to that held by Burdach). To Buettner-Janusch (‘66) the extensor action of the gluteus maximus is important more especially for such activities as running, walking up an incline, or climbing stairs, but he did point to a function in bipedal locomotion generally. Napier (‘67) holds a similar view, but specifically discounts the action of the muscle during walking on a level surface.
Thus, in the course of half a century, the opinion that the superficial gluteus is a powerful extensor acting during loco- motion in lower primates, but suited to more refined tasks in man, has been nearly reversed.
Not all recent authors adopt what is the prevailing modern view outlined above. Dart (‘49) emphasizes the rotatory func- tions of the gluteus maximus, as did Burdach. More recently, Sigmon and Rob- inson (‘67) claim that there are no major differences in function of this muscle in apes and man. Robinson (‘68) combines aspects of old and new by concluding that the muscle is a powerful extensor in both groups, but is not able to complete extension in apes. However, he states that the chief importance of the gluteus maxi- mus in man is not during walking but in the more strenuous activities of running, climbing, and rising. Sigmon (‘71) adds that the gluteus maximus in upright bi- peds is more of a speed-of-action muscle compared to that of apes.
It is my belief that although elements of truth are contained in a number of the theories proposed to date, the major mor- phological and functional distinctions be- tween man and non-human primates with regard to the superficial gluteus have not been identified. It is the plan of this paper to determine more carefully how the struc- ture of this muscle differs among primates and to identify those traits entirely unique to man. The results of such an analysis will be correlated with evidence bearing
on the function of the gluteus maximus in order to arrive at a new conclusion about the special role of the muscle in the evolution of human locomotion.
MATERIALS AND METHODS
Observations on the attachments and structure of the gluteus superficialis were made on the following non-human pri- mates (number of specimens in paren- theses): Lemur mongoz, mongoose lemur (1); Lemur fulvus, brown lemur (2) ; Mi- crocebus murinus, lesser mouse lemur (1); Cheirogaleus major, greater dwarf le- mur (1); Propithecus uerreauxi, Ver- raux’s sifaka (1); Nycticebus sp., slow loris (1); Perodicticus potto, potto (1); Galago senegalensis, bushbaby (1); Galago demidovii, dwarf galago (1); Tarsius syr- ichta, philippine tarsier (1); Saguinus ni- grocollis, black and red tamarin (1); Cal- licebus, titi (6); Aotus, owl monkey (7); Saimiri, squirrel monkey (6); Cebus, ca- puchin (7); Cacajao, uakari (6) ; Pithecia, s a h (5); Chiropotes, bearded saki (1); Alouatta, howling monkey (10); Lagothrix, woolly monkey (6); Ateles, spider monkey (6); Cercopithecus aethiops, grivet (1); Macaca mulatta, rhesus monkey (2); Theropithecus gelada, gelada baboon (1); Mandrillus sphinx, mandrill (1); Hylo- bates lar, white-handed gibbon (1); Sym- phalangus syndactylus, siamang (2); Pongo pygmaeus, orang-utan (3); Pan troglodytes, chimpanzee (1); Gorilla gorilla, gorilla (1). Quantification of the level of insertion into the femur was performed on ce- boids Hylobates, Symphalangus, Pan, and two specimens of Pongo. The technique is described by Stern (‘71).
Careful dissection and quantification of insertion of the gluteus maximus was undertaken for two human fetuses (third trimester), and six adult humans. More general confirmatory observations were made on 40 human cadavers used over the last two years in the medical gross anatomy course at the University of Chi- cago.
An excellent review (with some origi- nal data) of the myology of the hip and thigh in Old World primates has recently been published by Uhlmann (‘68). How-
2 More detailed information on the specimens of New World monkeys can be found in Stern (’71).

318 JACK T. STERN, JR.
ever, I believe it is necessary to summa- rize below the structure of the gluteus superficialis in non-human primates, espe- cially because my approach is different from Uhlmann’s and I can add extensive personal observations. Much of what is presented is gathered from the literature. In order to conserve space, specific cita- tions will be made only in special cases. However, almost all the references listed by Uhlmann have been personally reviewed by me, and the descriptions which follow are based on this review.
RESULTS
The “primitive” state
There is a basic configuration of the gluteus superficialis common to all pro- simians, callithricids, and the more strict- ly quadrupedal cebids. To this extent, at least, such a configuration may be con- sidered primitive. There is no direct evi- dence that it was present in any or all ancestral primates, but its distribution in living forms leads one to this supposition.
The “primitive” gluteus superficialis is a flat muscle more or less in the shape of a triangle, the base of which is repre- sented by the origin and the apex by the distal point of the insertion (plate 1,A). In the quadrupedal position the hip joint is deeply located approximately at the mid- dle of the triangle.
The gluteus superficialis arises ante- riorly from the strengthened fascia cover- ing the gluteus medius. The extent of this attachment is variable. From the gluteal fascia, the origin passes dorso-caudally to the fascia over the tail muscles and then travels caudally along this fascia up to the region of the first few caudal verte- brae, from the transverse processes of which a considerable portion of the mus- cle directly arises.
The “primitive” gluteus superficialis inserts entirely into the femur. The ante- rior (cranial) half of the muscle goes to a tendon which forms on its deep surface and in turn attaches to the caudo-lateral surface of the femur just below the great- er trochanter (plate 2A,B). In prosimians, the attachment area is marked by a well defined third trochanter, and there is an elevation of less marked prominence in
other forms. The tendon of insertion of the anterior portion of the muscle is called the ascending tendon (Stern, ’71) because its anterior fibers insert most distally, while the more posterior ones attach pro- gressively higher on the femur. The distal edge of the tendon is located at the site where the fascia covering the gluteus superficialis passes distally into the fascia femoris. As a result, it can appear that the anterior fibers of the muscle insert into the fascia femoris, but this quite defi- nitely is not the case.
The caudal portion of the “primitive” gluteus superficialis inserts either direct- ly, or by means of a flat tendon, or by both means, into the posterior surface of the femur from the level of the ascending tendon (or a bit more distally) to well past the middle of the bone. The most anterior fibers of this portion insert proximally, the more posterior fibers attaching pro- gressively further down the shaft. When the caudal portion has a well formed ten- don, its lateral edge is attached to the fascia femoris. However, the tendon fibers do not pass into the fascia femoris, but, on the contrary, proceed to the femur.
Primitively, the anterior portion of the gluteus superficialis is very thin, while the posterior portion, particularly the fi- bers arising from the caudal vertebrae, is thick and seemingly powerful (plate 3,A,C,D). In some cases the division into two portions is not merely topographical; the insertion of the ascending tendon may be quite separate from the remainder of the femoral attachment, and division here can often be carried proximally with very little difficulty. The significance of this is moot.
The descriptions of the gluteus super- ficialis that follow attempt to point out briefly more particular aspects of the structure of this muscle in primate sub- groupings. Emphasis will be placed on deviation from the “primitive” condition.
Prosimians
Among the Lemuroidea, Daubentonin, the Galaginae, and Tursius, the structure of the gluteus superficialis is nearly uni- form and agrees closely with what has been described as the “primitive” state. In all these forms the gluteus superficialis is

SPECIALIZATIONS OF HUMAN GLUTEUS MAXIMUS 319
in close connection with the tensor fasciae femoris, which inserts into the ascending tendon and a tendon fused to it. There is variable evidence of a separation between the two muscles. Fibers arising from the anterior superior angle of the ilium sometimes appear to form a distinct bun- dle which is called tensor fasciae femoris. These fibers are innervated by the superi- or gluteal nerve, but in Lemur, Bluntschli (‘13) and Uhlmann (‘68) found that the superior gluteal nerve also supplied most of the fibers arising from the gluteal fascia. Whether these fibers ought to be assigned to the tensor fasciae femoris (thus leaving us with a gluteus super- ficialis characterized by a very slight or absent origin from the gluteal fascia) or be considered as belonging to the gluteus superficialis but having an “anomalous” innervation (as I believe occurs in some cebids, Stern, ’71), cannot as yet be de- termined. In the specimen of Propithecus available to me (Field Museum of Natu- ral History 85150), there occurred an easily separable tensor fasciae femoris arising from the anterior superior iliac angle, the septum intermusculare iliacum, and adjacent gluteal fascia. It was inner- vated by the superior gluteal nerve, which did not send branches to fibers of the gluteus superficialis arising from the glu- teal fascia. Also in Tarsius, the tensor fasciae femoris is well separated from the gluteus superficialis at their origins.
In lemuroids, Daubentonia, galagines, and Tarsius, the origin of the gluteus superficialis from the tail encompasses the first two, or more usually the first three, caudal vertebrae. The origin here is largely by muscular fibers, though I observed short tendons in Lemur, Propi- thecus, and Tarsius. Galagines differ from the other mentioned forms in having a di- rect muscular origin from the iliac crest (plate 1,A) as opposed to an origin medi- ated by the gluteal fascia. Once again, however, these fibers probably represent tensor fasciae femoris. Murie and Mivart (1872) reported that they inserted into the fascia femoris in one of their speci- mens of G. crassicaudata.
In the four groups of prosimians under discussion, the posterior portion of the gluteus superficialis inserts muscularly into the femur. Only in Lemur has there
been described a small attachment to a lateral intermuscular septum (Bluntschli, ’13) or fascia femoris (Uhlmann, ’68). In my specimen of L. mongoz a tendon was associated with the more proximally in- serting fibers of the caudal portion of the muscle, however, I can confirm the find- ings of most authors who report no true fascia1 insertion in the genus Lemur.
The posterior fibers of the gluteus su- perficialis are said to extend their inser- tion onto the capsule of the knee and tibia1 tuberosity in cheirogalienes (Juof- froy, ’62). I did not observe this, but I did note that the femoral attachment con- tinues further distally than in other pro- simians, reaching the lateral condyle and sesamoid of the lateral head of gastroc- nemius. In lemurines and Daubentonia the posterior fibers of the gluteus super- ficialis fall just short of the lateral con- dyle, and a lesser area of insertion occurs in indriids. Juoffroy (‘62) stated that the distal limit of the femoral attachment in Propithecus was two-thirds the way down the bone, but in my specimen the muscle reached from three-fourths to four-fifths down the femur (plate 3,A). Milne Ed- wards (1875) reported that in Avahi the gluteus superficialis does not insert be- yond the femoral midpoint. Galagines and Tarsius resemble the Indriidae in that the insertion is generally limited to the upper two-thirds of the femur.
In Lemur (Lucae, 1884; Appleton, ’21), Daubentonia (Lucae, 1884), and Tarsius (Appleton, ’21), the weight of the com- bined gluteus superficialis and tensor fas- ciae femoris exceeds that of either of the other two gluteal muscles. Ranke (1897) stated that the gluteus superficialis was stronger than the medius in Lemur. The posterior portion of the gluteus superfici- alis is markedly thicker than the anterior portion in lemuroids (plate 3,A). I found the difference in thickness to be less obvi- ous in Galago, and not at all noticeable in Tarsius. However, the tarsier possesses a powerful caudo-femoralis running along- side the posterior portion of gluteus super- ficialis and, no doubt, having a very simi- lar action.
Among lorises the gluteus superficialis differs in some rather important ways from that in the previously described pro- simians. In the Asian forms there is no

JACK T. STERN, JR. 320
muscular bundle arising either tendin- ously or muscularly from the anterior su- perior iliac angle or ventral third of the iliac crest. Instead there is a direct mus- cular origin from the dorsal portion of the iliac crest. These fibers appear to cor- respond to a tensor fasciae femoris; they are innervated by the superior gluteal nerve (Bluntschli, ’13). Perodicticus is similar to most other prosimians in that muscular fibers arise from the anterior superior angle of the ilium and caudal to this from the gluteal fascia.
Bluntschli (’13) reported that in Nycti- cebus muscular fibers arise from the dor- sal edge of the ilium. They were inner- vated by the superior gluteal nerve and possibly could represent a portion of the tensor fasciae femoris. However, I very carefully looked for fibers arising directly from the dorsal edge of the ilium in three specimens of Nycticebus and found no such origin.
The gluteus superficialis arises from caudal vertebrae one to three in Nycti- cebus (Bluntschli, ’1 3). Juoffroy (’62) has reported the same for the potto, but Riska (’36) and Uhlmann (’68) state that the origin also encompasses the fourth and fifth caudal vertebrae in Perodicticus. The latter two authors note that the cau- dal origin in Perodicticus is tendinous, and my observations agree. A similar ten- dinous origin occurs in Nycticebus.
Appleton (‘27) reported that in both Loris and Perodictius, but not in Nycti- cebus, the gluteus superficialis extends its origin to the ischial tuberosity with the formation of a fibrous arcade which he calls a sacro-tuberous ligament. Riska (‘36) noted that in most of his specimens of lorises the posterior portion of the glu- teus superficialis either covered or had a small origin from the ischial tuberosity. He stated that only in these forms and man was the ischial tuberosity covered by this muscle. However, both Uhlmann (‘68) and I found this relationship in Lemur, and I also observed i t in the other lemur- oids. An actual origin from the tuberosity is quite a different matter. Uhlmann (‘68) confirms Appleton’s observation that it occurs in the potto, but Nayak (cited in Juoffroy, ’62) disputes its presence in Loris. No such origin occurred in my specimen of Perodicticus, but in both this
genus and Nycticebus the tendon of origin did adhere strongly to the fibrous tissue over the ischial tuberosity.
As in all prosimians, the ascending ten- don attaches to a narrow region high on the femur, from 5% -1 1% down the shaft in my specimen of slow loris. The inser- tion of the posterior portion of the glu- teus superficialis nearly reaches the lat- eral femoral condyle in lorises. It differs substantially from the condition in other prosimians in that there is a tendon of insertion which also gives origin to fibers of the vastus lateralis (and possibly vastus intermedius). Thus a true lateral inter- muscular septum is formed. I should em- phasize that observations bearing on this point exist only for Nycticebus and Pero- dicticus (Bluntschi, ’13; Uhlmann, ’68; and personal data).
New World monkeys
A detailed description of the gluteus superficialis in cebids can be found in Stern (‘71). The marmosets resemble Cal- licebus.
The structure of the gluteus super- ficialis in marmosets and all cebids ex- clusive of Alouatta and the Atelinae fol- lows closely the primitive pattern. The extent of origin from the gluteal fascia is variable (slight in marmosets, Callicebus, and Pithecia; moderate in Saimiri, Cebus, Chiropotes, and some specimens of Caca- jao; extensive in Aotus and the other spec- imens of uakari). There is a fair degree of interindividual variation in the number of caudal vertebrae giving rise to the posterior portion of the gluteus super- ficialis, but usually the first two or three are involved, with the fourth coming into play often in Saimiri and Cacajao (in this latter form caudal vertebrae may be in- corporated into the sacrum).
The ascending tendon inserts into a short region high upon the femur in those ceboids characterized by a “primitive” gluteus superficialis (plate 2,B). In Cal- licebus, Aotus, Cacajao, and Pithecia the attachment is from 6%-11% down the femur; it lies 1 or 2% further distally in Saimiri and Cebus.
In New World monkeys the posterior portion of the gluteus superficialis inserts largely by means of a tendon, best devel-

SPECIALIZATIONS OF H U M A N GLUTEUS MAXIMUS 321
oped in the larger forms. The muscular fibers continue about half-way down the thigh in Callicebus, Aotus, and Saimiri (plate 2,B); the tendon extends the at- tachment three-fifths down. In Cacajao, Chiropotes, Pithecia, and Cebus, the flesh reaches about two-fifths the way down the femur, and the tendon ends somewhat beyond the midpoint. Therefore, there is a less extensive femoral insertion in “prim- itive” New World monkeys than occurs in almost all prosimians.
Alouatta caraya possesses a gluteus superficialis which differs from the “prim- itive” state by possessing a wider, strong- er, and more distally inserting ascending tendon, and a posterior portion whose tendon of insertion is lightly fused to the aponeurosis of origin of the vastus lat- eralis. The muscular fibers extend about as far down the thigh as in the uakari or capuchin. In Alouatta palliata, there are several additional differences. A well defined tubero-caudal ligament occurs, and from its external surface fibers of the gluteus superficialis arise. The posterior portion of the muscle descends no fur- ther than one-third the way down the thigh. It inserts onto the back of the ascending tendon and also via a clearly developed lateral intermuscular septum. In the woolly monkey no tubero-caudal ligament exists, but the origin from cau- dal vertebrae is tendinous, and in most specimens this tendon extends toward the ischial tuberosity, sometimes nearly reach- ing it. The ascending tendon is well developed and inserts from 14% to 21% down the femur, whereas the posterior portion of the muscle has the same man- ner and extent of attachment as occurs in A. palliata.
The gluteus superficialis of Ateles de- viates very markedly from the “primitive” state. There is a well developed caudal tendon of origin which always extends up to, though not actually reaching the isch- ial tuberosity. Furthermore, the origin from the tail does not go beyond the sec- ond caudal vertebra, and occasionally not beyond the first. The powerful ascending tendon (plate 3,B) attaches from 15% to 24% down the femur. The posterior por- tion of the muscle inserts almost wholly into the back of the ascending tendon, never extending far below it. There is
less distinction between the thicknesses of the caudal and cranial portions of the gluteus superficialis (plate 3,B) but the former remains the thicker. In its shape and localized insertion, the muscle pre- sents an appearance similar to the deltoid of the shoulder.
I have weighed muscles in one speci- men of each cebid genus (Chiropotes and Brachyteles excluded). In Saimiri the glu- teus superficialis weighed slightly more than the medius. The medius was not much heavier than the superficialis in Cebus. In other forms the predominance of the medius was substantial, particularly so in the ateline genera.
Old World monkeys
The gluteus superficialis differs more from the “primitive” state in Old World monkeys than in any other primate group. Furthermore, Klaatsch (‘00) remarked that all Old World monkeys are alike with regard to the configuration of the muscles on the outer part of the thigh, and a re- view of the literature coupled with my own observations leads me to agree. The only point wherein one form may differ significantly from another is in the pres- ence or absence of an origin from a sacro-tuberous ligament and/or ischial tuberosity. Even here differences have not been definitely established.
The origin (plate 1,B) from the gluteal fascia, tail fascia, and caudal vertebrae is not unusual, though in most forms only caudal vertebrae one and two are involved (origin from the first three, or only the first, is not uncommon).
Keith (1894) reported that a great sacro- sciatic ligament, represented by a thick- ened underportion of the sheath of the gluteus superficialis, occurs in Cynopith- ecus (the Celebes ape) and is very strong in Macaca arctoides. Kleinschmidt (‘49) found this structure in Cynopithecus, and a weak one in the Gibraltar macaque. Patterson (‘42) stated that the gluteus superficialis of the Celebes ape arises from the ischial tuberosity. Other authors who have written on macaques report neither a tuberal origin nor a sacro-tuber- ous ligament.
Lachman (‘38) reports finding an origin from the ischial tuberosity in the hamad-

322 JACK T. STERN, JR
ryas baboon, but almost certainly he has mistaken a portion of the flexor cruris lateralis for gluteus superficialis. No other author who has described baboons, including hamadryas, mentions a tuberal origin. On the other hand, Schumacher and Lebzelter (‘24) observed that the cau- dal fibers of the gluteus superficialis of the mandrill arise from a sacro-tuberous ligament (though not from the tuberosity itself). Cuvier and Laurillard (1849, pl. 40) indicate no such origin, Bischoff (1870) does not mention it, and I also found no sacro-tuberous ligament nor any special origin of the posterior portion of the muscle in the mandrill. Furthermore, neither the ligament nor unusual origin occurs in the drill (Kleinschmidt, ’49; Pagenstecher, 1867). Zielinska (‘36, ’37) pictured the gluteus superficialis of Thero- pithecus as arising from a sacro-tuberous ligament, but it is not reported by Apple- ton (‘27) and I found no hint of such a structure (plate 1 ,B).
In Sernnopithecus ( = Presbytis) entellus, Ayer (’48) observed that fibers of the gluteus superficialis arise from an “inter- muscular septum” between the sacrum and ischial tuberosity. Kohlbrugge (1 897) does not discuss this in his description of Sernnopithecus, Polak (’08) did not re- port such a n origin in Colobus, and Pat- terson (‘42) makes no mention of it in Rhinopithecus.
Until further evidence is gathered, one is probably safe in assuming that neither a sacro-tuberous ligament nor an origin of the gluteus superficialis from the isch- ial tuberosity occurs in any Old World monkey with the possible exception of Cynopithecus .
The insertion of the gluteus superficial- is in Old World monkeys is entirely un- like that in any other primates. The anterior fibers insert wholly onto the su- perficial surface of the deep lamina of the fascia femoris (plates 1,B, 2,C). The posterior fibers may do the same, but more usually the deeper of these have a weakly tendinous andlor muscular attach- ment to the posterior surface of the fe- mur. The location of the femoral attach- ment is variable, but usually lies some- where above the midpoint, and often quite proximally. There is no formation of a true lateral intermuscular septum.
The literature is unanimous in empha- sizing the relatively small size of the cer- copithecoid gluteus superficialis. Haugh- ton (1865) found that it weighs less than half the me&us. Nonetheless, the posteri- or portion is markedly thicker than the anterior part of the muscle (plate 3,D).
Hylobatidae
The thin anterior portion of the glu- teus superficialis arises from the gluteal fascia and from an aponeurotic fascia connecting to the lateral crest and spines of the sacrum. This latter fascia is clearly the homologue of the fascia which covers the tail muscles in forms that have them. The posterior portion of the gluteus superficialis arises from the lateral edge of the post-articular sacrum and from caudal vertebra one (van den Broek, ’14; Ranke, 1897). I did not note the exact extent of the caudal origin in my speci- mens, but the muscle did not reach the posterior region of the coccyx. Also attaching to the first caudal vertebra is a tubero-caudal ligament. On cross- section the ligament is “J” shaped, the bend being located along the dorso-caudal edge and the long arm lying deeply. The gluteus superficialis arises from the super- ficial surface of the long arm and from the concavity of the bend. The origin is carried by the ligament to the ischial tuberosity. A substantial portion of the gluteus superficialis arises from the tu- berosity and from the tendon of the long head of biceps femoris.
In the siamang, the tubero-caudal liga- ment attaches also to the caudo-lateral angle of the sacrum and to the posterior inferior iliac spine. Near the ilium, the ligament gives origin to fibers of the glu- teus medius. Further medially, the gluteus superficialis covers the most cranial re- gion of the ligament, but few, if any, muscular fibers arise from that spot.
Keith (cited in Uhlmann, ’68) found that in Hylobates muscular fibers arise directly from the posterior superior spine of the ilium, but this is denied by Kohl- brugge (1890) and Kanagasuntheram (‘52). My observations also do not sup- port Keith’s findings. Kohlbrugge (1890) reported that in the siamang there occurs an iliac origin from the elevated ridge

SPECIALIZATIONS OF HUMAN GLUTEUS MAXIMUS 323
which medially borders the origin of the gluteus medius. I found no trace of such an origin.
Kanagasuntheram (’52) and Uhlmann (‘68) claimed that the anterior portion of the gluteus superficialis inserts into the fascia femoris in Hylobates, but Kohl- brugge (1880) stated that this part of the muscle inserts by means of a strong ten- don into the femur just distal to the greater trochanter (thus as in the “primi- tive” condition). My observations lend support to Kohlbrugge, but I did find that a few of the most cranial fibers of the gluteus superficialis do attach deeply to the fascia femoris on the lateral side of the thigh. The tensor fasciae femoris and gluteus superficialis are so thorough- ly fused in the siamang that 1 could not determine whether the gluteus has a par- tial fascia1 insertion, or the tensor has a partial attachment to the ascending ten- don, or they each have independent inser- tions into the tendon and fascia respec- tively. In my specimen of the gibbon, the ascending tendon inserted from 9% to 15% down the femur. The corresponding figures for the two siamangs are 11% to 23% and 17% to 34%.
The posterior portion of the gluteus superficialis inserts into the ascending tendon and distally into the femur di- rectly and by means of a tendon which is fused to the aponeurosis of origin of the vastus lateralis. Again, this represents a true lateral intermuscular septum. It is usually reported that the posterior portion inserts as far as one-half to two-thirds the way down the femur. In my specimen of gibbon the muscular fibers continued just beyond the femoral midpoint, but the ten- don of insertion could be traced (with some uncertainty) as far as the lower one- fifth. In Symphalangus , the muscular fibers end at a point roughly 60% down the thigh.
The gluteus superficialis of the gibbon weighs less than half the weight of the gluteus medius (Tappen, ’55).
Pongidae
The general form and relations of the gluteus superficialis of African apes (plate 1,D) is very similar to that in the Hylo- batidae. The orang-utan presents quite a
different picture. A similarity between gibbon and siamang on the one hand, and the chimp and gorilla on the other is somewhat mysterious; presumably it is not based on a particularly close phylo- genetic relationship, nor is the use of the hindlimbs in the extant forms so alike as to lead to convergent morphological specialization. Possibly during some pre- vious time the locomotor behaviors of these‘ animals were more alike than is now the case.
My observations on Pan and Gorilla re- vealed the great resemblance between these forms in the structure of the glu- teus superficialis and showed this struc- ture to be almost identical to that de- scribed by Preuschoft (‘61) for the gorilla. The origin from the gluteal fascia is ex- tensive in the gorilla, and although I found it less so in Pan (plate l,D), many authors report otherwise. There is prob- ably considerable variation in the regard. Posterior to the origin from the gluteal fascia, the muscle arises from the caudal end of the long dorsal sacro-iliac liga- ment, which runs from the posterior su- perior iliac spine to the lower spines of the sacrum. Many authors report a direct origin from the sacrum, but I found that such an origin was from the fascia which covers the multifidus. A direct origin from the ilium is claimed by some (Beddard, 1895; Bischoff, 1880; Miller, ’52), but is not reported by the vast majority of au- thors, and I observed no such attachment.
The origin passes backward onto the coccyx and sacro-tuberous ligament (plate 1,D). This ligament does not have the configuration described above for Hylo- bates. In my specimens of African apes, the sacro-tuberous ligament was a twisted structure. Ligamentous fibers from the medial part of the ischial tuberosity ran deeply upward and forward to attach to the caudo-lateral angle of the sacrum. Fibers from the lateral region of the isch- ial tuberosity cross this group superficially and attach to the coccyx near its tip. These superficial fibers are much better developed than the deeper ones. The glu- teus superficialis reaches the ischial tuber- osity by passing along the superficial fibers. There is an extensive origin of the muscle from the ischial tuberosity and the tendon of the long head of biceps

324 JACK T. STERN, JR.
femoris. A separate ligament runs from the caudo-lateral angle of the sacrum upward to the posterior inferior spine of the ilium, but the sarco-tuberous liga- ment has no direct connection to the ilium.
In the chimp and gorilla, the gluteus superficialis inserts into the femur by means of a well developed ascending ten- don and a tendon fused with the apo- neurosis of origin of the vastus lateralis. In fact it was dissection of these primates that led Champneys (1871), Ranke (1897), and Klaatsch (‘00) to determine the role played by the tendon of insertion of the posterior part of the gluteus superficialis in the formation of the lateral intennus- cular septum.
In my specimen of Pan, the ascending tendon attached from 20% to 33% down the femur. Preuschoft (‘61) reports that it inserted at the end of the proximal third of the shaft in one specimen of gorilla, in another specimen at the femoral mid- point. The posterior portion of the muscle extends its insertion far below the mid- point of the femur (plate 1,D). Many au- thors state that it reaches the lateral condyle, but one cannot always determine whether muscular fibers or tendon are being discussed. It is probably fair to say that the muscle fibers continue at least two-thirds down the thigh, and often beyond.
There are various reports, not repre- senting the majority, of an insertion into the fascia femoris. In one specimen of gorilla, Preuschoft (‘61) noted that a few of the most anterior fibers of the gluteus superficialis did so insert. I found this in both African apes. Neither Preuschoft nor I observed any other mode of insertion into the fascia femoris, and i t is my con- clusion that an insertion into the lateral intermuscular septum has often been mis- interpreted as being into the fascia femo- ris. Also, in the chimp and gorilla, the superficial surface of the ascending ten- don is not entirely covered by muscular fibers (plate 1,D), and the fascia femoris may be bound down to the exposed tendon surface. This could account for some in- correct descriptions of a fascia1 insertion.
In African apes, the anterior portion of the gluteus superficialis is far thinner than the posterior portion. The entire
muscle weighs substantially less than the gluteus medius in Pan (Macalister, 1873; Fick, ’25; personal data), but in Gorilla the two muscles are almost of equal weight (Macalister, 18 73 ; Preuschoft, ’6 1).
The gluteus superficialis of the orang- utan (plate 1,C) arises from the gluteal fascia, the fascia over the multifidus (pos- terior to the sacro-iliac joint), and the coccyx. An origin from a well defined sacro-tuberous ligament is not found. Primrose (1898-1899) stated that the muscle arises from “sacrosciatic” fascia, and van den Broek (’14) described a fra- gile tubero-caudal ligament giving rise to muscle fibers from its external surface. Sonntag (‘24) also reported an origin from a sacrosciatic ligament, but Mysberg (‘1 7) noted that such a structure, though pres- ent, could not be outlined either at its beginning or at its end, where it seemed to lose itself in the perimysium of the gluteus superficialis. Boyer (‘35) noted that the tuberosity of the ischium was connected to the gluteus superficialis by an extension of the latter’s fascia. Op- posed to all these authors are an equal number who do not mention a sacro- tuberous ligament and some (Fick, 1895a; Michalis, ’03; Kleinschmidt, ’49) who deny its presence. One of my specimens was dissected while still fresh. In this animal I found a very weak fibrous band which fits the description given by Mys- berg. I did not notice such a structure in my fixed specimens, but it may have been overlooked because of its extreme tenuosity .
The insertion of the gluteus super- ficialis in Pongo is more similar to the condition in Ateles than in any other pri- mate. The whole muscle inserts into a strong ascending tendon (fig. 2,D); in fact, the only reason for believing that a pos- terior portion exists is that fibers arise from the coccyx. The external surface of the ascending tendon is exposed to the fascia femoris and they adhere, but the fascia may be heavily fat-laden here and no insertion into it occurs. The site of attachment of the tendon is variously reported, but usually stated to lie some- where between one-quarter and one-half the way down the femur. In one of my specimens, the femoral insertion occurred between 17% and 25% down the bone,

SPECIALIZATIONS OF HUMAN GLUTEUS MAXIMUS 325
in another specimen the comparable fig- ures are 19% and 29%.
Due to the great reduction of its poste- rior portion, the gluteus superficialis ap- pears of nearly uniform thickness on cross-section (plate 2,D). A similar phe- nomenon occurs in Ateles (plate 3,B). In the orang-utan, the muscle weighs con- siderably less than the gluteus medius (Fick, 1895b; Langer, 1879).3
Man
Features of the origin, insertion, size, and shape of the gluteus maximus are totally unique to man, and it is just these features which we are seeking to identify and interpret.
Textbooks are nearly uniform with re- gard to the description of the origin of the gluteus maximus. What differences occur between them, and between them and what follows, are of little significance.
There may be an origin from the glu- teal fascia, as is frequent in non-human primates, but this is not a constant fea- ture. When it does occur, however, i t may be extensive. There is a constant origin from the dorsal region of the ilium. This occurs by means of a deep ten- don which attaches to the posterior gluteal line (the tendon being responsible for the line) and by means of a direct fleshy attachment to the ilium behind this line. Immediately inferior to the ilium, the fibers arise from the dorsal surfaces of (a) the sacro-tuberous ligament, which attaches to the posterior spines of the ilium; (b) the long posterior sacro-iliac ligament; and medial to this, (c) the fas- cia covering the multifidus almost up to the sacral spines. The origin passes backward along the multifidus fascia and sacro-tuberous ligament. Where the multi- fidus ends, its covering fascia fuses with the periosteum of the lateral portion of the sacrum, and the gluteus maximus therefore arises from the sacrum. The origin then continues onto the COCCYX nearly to its tip. Fibers arise from the entire superficial surface of the sacro- tuberous ligament, however, the number of such fibers is relatively small. Further- more, since the attachment of the liga- ment to the coccyx does not extend as far caudally as the origin of the gluteus max-
imus from the coccyx, the most inferior fibers of the muscle have no origin from the ligament. This condition is therefore different from what occurs in hylobatids and African apes, and somewhat similar to what is found in Alouatta palliata (and the orang-utan?).
The truly unique attribute of the ori- gin of the gluteus maximus is its strong and extensive attachment to the ilium. Related to this is an origin from the sacro-tuberous ligament as it attaches to the posterior spines of the ilium. Only in man and the siamang does the ligament even reach the ilium, and the gluteus superficialis of the siamang does not arise from either structure in this region. A third unique characteristic of man is the extent of the origin from the fascia over the multifidus. Although an origin from homologous facial layers occurs in all pri- mates, the direct origin from the fascia medial to the long dorsal sacro-iliac liga- ment is strictly human.
Descriptions of the insertion of the glu- teus maximus are at wide variance with each other. Most frequently, detailed ac- counts are not given. A common state- ment is that the entire cranial half of the muscle inserts into the fascia lata where- as only the superficial fibers of the caudal half attach here, the deeper ones going to the lateral intermuscular septum and into a tendon which inserts into the femur. However, many authors report that all of the caudal fibers have a septa1 and femo- ral insertion, while still others state that the deep fibers of the whole muscle have this attachment. Some authors deny a fascia1 insertion altogether.
I have taken particular care in observ- ing the intricacies of the insertion of the
3 The orang-utan is unique among apes in possessing a muscle that runs from the ischial tuberosity to the lower one-half to two-thirds of the femoral shaft and aponeurosis of vastus lateralis (plate 1,C) but is inner- vated by a direct branch from the sciatic nerve and also by a branch of the nerve to the long head of biceps femoris. The muscle may be considered a portion of the biceps femoris, or may be given the name “ischiofemo- ralis.” The action of the ischiofemoralis must be simi- lar to that of the posterior fibers of gluteus superficialis in other apes. However, it is not known if the role played by the ischiofemoralis in the locomotion of the orang is like that played by the posterior fibers of the gluteus superficialis in the movements of the other apes. Also unknown is whether or not any of the apes is descended from a monkey-like ancestor that pos- sessed a long head of biceps which inserted into the thigh, as occurs in cercopithecoids and many ceboids.

STERN. JR.
ing tendon and lateral intermuscular sep- tum. However, a very small number of superficial fibers may gain attachment to the overlying fascia. An examination of the lateral intermuscular septum reveals quite clearly that it is composed of three layers. The most posterior layer contains fibers which run upward and inward to- ward the femur. These represent a tendon of origin of the short head of biceps femoris. Such a tendon is seen as a sepa- rate entity in Pithecia. The middle layer is composed of fibers running downward toward the femur. These can be seen to emanate from the gluteus maximus and represent the tendon of its caudal por- tion. The anterior layer consists of fibers running in the same direction as the short head of biceps tendon. These give origin to the vastus lateralis. Because of the different fiber orientations within the lateral intermuscular septum, the struc- ture has a criss-cross appearance.
In six adult specimens, the average area of attachment of the ascending ten- don extended from 14% to 32% down the femur. This is rather low, though not uniquely so (cf. chimp, gorilla, and sia- mang). However, upon inspection it is clear that the far distal extent of the attachment of the ascending tendon in man is due to the fact that the lower fibers expand markedly near their termi- nation (plate 2,F). The main body of the tendon does not insert much further dis- tally than one-fourth the way down the bone. The expansion results in the distal tendinous fibers of insertion running near- ly parallel to the bone, and in this respect is reminiscent of the insertion of the semitendinosus or semimembranosus. In the latter cases, such a condition has
326 JACK T.
gluteus maximus, and my findings differ only in one important point from the de- tailed account given by Sappey (1876). Since the validity of his observations is not generally acknowledged, I am repro- ducing two of his figures (plate 4) and the following passage from his text (pp. 377- 378):
Les faisceaux superieurs s’inserent a la face profonde de petits tendons rubanes et paralleles, d’autant plus longs qu’ils sont elevees, lesquels contournent le grand tro- chanter en se condensant et en formant une lame epaisse, de plus en plus etroite, pour aller se fixer sur la partie terminale de la branche externe de la ligne Bpre. Cette lame tendineuse est logee dans un dkdouble- ment de l’aponevrose fernorale, dont le feuillet interne passe sur la face profonde en lui adherent de la maniere la plus in- time, tandis que l’externe, beaucoup plus epais et tres-adherent aussi, passe sur la face opposee.
Les faisceaux inferieurs se rendent a un gros et court tendon aplati qui se continue en haut avec la lame preckdente, et en de- hors avec l’aponevrose de la portion externe du triceps crural; ce tendon s’attache a la branche externe de la ligne ipre et a la partie superieure de cette ligne.4
Sappey describes a purely femoral and septa1 insertion of the muscle, and it is with this that I disagree. He stated that the superior portion of the muscle inserts into the deep surface of a small tendon composed of many ribbons. In fact, this “tendon” is a deep layer of the overlying fascia, and the fibrous tissue into which the gluteus maximus inserts is continued into the fascia lata (plate l,E,F). Sap- pey’s observations can be reconciled with mine when it is realized that only the superjicial fibers of the superior half of the gluteus maximus gain an appreci- able attachment to the fascia lata, while the deep fibers go to a typical ascending tendon which in turn inserts on the fe- mur (plate 2,E,F). As in the great apes, the surface of the ascending tendon ad- heres to the overlying fascia lata, and this cannot only lead one astray in the direction of believing that the entire in- sertion is into the bone (as did Sappey), but also in the opposite direction of think- ing that the entire insertion is into fascia. To avoid this, dissections of fetal material are helpful, for here the ascending tendon stands out in greater relief.
The caudal portion of the gluteus max- imus inserts almost totally into the ascend-
4 The superior bundles insert on the deep surface of small ribbon-like and parallel tendons (the more su- perior ones being progressively longer) which twist around the greater trochanter, condense, and form a thick lamina that becomes narrower and narrower to insert on the terminal part of the external bifurcation of the linea aspera. This tendinous lamina is lodged in a doubling of the femoral fascia, the internal layer of which passes on the tendon’s deep surface, adhering to it most closely, whereas the external layer, much thicker and also closely adherent, passes on the ten- don’s outer surface.
The inferior bundles go to a thick and short flat tendon which is continuous superiorly with the lamina discussed above and laterally with the aponeurosis of the vastus lateralis. This tendon attaches to the ex- ternal bifurcation of the linea aspera and to the su- perior part of the linea aspera itself.

SPECIALIZATIONS OF HUMAN GLUTCUS MAXIMUS 327
been interpreted (Mollier, '37) as being a correlate of the nearly parallel pull exerted on the tibia by these muscles when the leg is extended. Possibly the expansion of the ascending tendon of the gluteus maximus indicates a pull on the femur nearly parallel to its shaft.
The muscular fibers of the caudal por- tion of the gluteus maximus insert, on the average, as far as 33% down the thigh. Such a limited distal extent is found also in prehensile-tailed cebids. Even though the muscular fibers reach little or no further down the thigh than does the ascending tendon, the caudal fibers of the muscle are able to gain broad attachment to the lateral intermus- cular septum because they insert lateral to the expansion of the ascending ten- don (plate 2,F).
The unique characteristics of the in- sertion of the gluteus maximus are (a) the extensive attachment of the super- ficial fibers of the cranial portion into the deep surface of the overlying fascia, and (b) the broad expansion of the lower fibers of the ascending tendon. All other aspects of the insertion in man can be duplicated in the lower primates, though the limited distal extent of the insertion of the cqudal fibers is uncommon.
The gluteus maximus weighs more than twice the medius (Voss, '56; Schumacher and Wolff, '66). This is unparalleled in other primates. But it is not sufficient to state this fact without inquiring wherein the extra bulk of muscle is concentrated. Ranke (1897) noted that in lower pri- mates the superficial gluteus comes pre- dominantly from the tail vertebrae, where- as in man only the slightest portion of the fibers arise from the coccyx. He em- phasized the great mass of the part aris- ing from the sacrum. Cross-sections of the bellies of the gluteus superficialis and maximus (plate 3 ) provide us with a good indication of where the additional fibers in man are located. It can be seen that the anterior portion of the muscle is equal to or greater in thickness than the posterior portion in man, but is thin- ner, most often markedly so, than the posterior portion in all other primates. The large size of the cranial portion of the gluteus maximus correlates well with the new firm origin from the ilium, the
more extensive origin from the multifidus fascia medial to the long dorsal sacro- iliac ligament, and the origin from the sacro-tuberous ligament near the iliac spines. Whether or not the relatively large weight of the gluteus maximus is en- tirely attributable to the increased cranial portion has not been determined.
DISCUSSION
The evidence accumulated above dem- onstrates quite clearly that the unique morphological specializations of the hu- man gluteus maximus are confined to its cranial portion. This part of the mus- cle is much thicker than in any other primate. It has a new and firm origin, a modified ascending tendon of insertion, and an entirely new insertion into the overlying fascia continuous distally with the fascia lata. Of interest now, is the identification of those functional abilities which are made possible by the unique anatomical configuration. Having done this, we can consider a possible selective pressure favoring such abilities and look for evidence in fossil man of the changes which were occurring.
The cranial portion of the gluteus max- imus is certainly not that segment which is topographically best suited to produce extension of the thigh. On the contrary, the caudal fibers of the muscle have the most favorable moment arm for extension, yet they exhibit no peculiarity which can- not also be seen in other primates. Of course, these fibers have been adjusted so that their physiological resting length occurs when the leg is in line with the trunk, as opposed to what can be pre- sumed to be the case in quadrupedal forms in which the resting length is likely to occur when the femur is positioned at about 90" to the trunk in a sagittal plane. Furthermore, the angulation of the human sacrum (probably an obstetri- cal necessity) grants to the caudal portion of the gluteus maximus a leverage which would have been markedly reduced had not the position of the sacrum been changed from that seen in non-human primates. These two modifications, one physiological, the other morphological, en- able the gluteus maximus to extend the lower limb beyond the resting position

328 JACK T. STERN, JR.
with power. However, it can certainly be assumed that in most other primates the gluteus superficialis can extend the lower limb beyond the resting position with power. Thus, the changes in man have been directed to the preservation of an old ability, rather than to the creation of a new one.
That the extensor activity of the glu- teus maximus is primarily important as an aid to the hamstrings during activities which require special effort has been demonstrated by Wheatley and Jahnke (‘51), Joseph and Williams (’57), Karlsson and Jonsson (‘65), and Pauly and Schev- ing (‘68). It has no specific role as an extensor in erect posture or locomotion. Duchenne (1867) observed that paralysis of the gluteus maximus does not impair standing or walking. Subsequently, it has been shown (Joseph and Williams, ’57; Karlsson and Jonsson, ’65) that the mus- cle is not electrically active in standing at ease. If, during standing, the center of gravity of the trunk is shifted forward, the hamstrings are the first to be called into action, and most often they alone suffice to maintain equilibrium. The great importance of the hamstrings in erect posture is suggested by the nature of the changes in the ischial tuberosity which have occurred in human evolution.
The gluteus maximus is active during walking (Eberhart, Inman and Bresler, ’54; Battye and Joseph, ’66), but not in all subjects (Sutherland, Bost and Schott- staedt, ’60). When it is active, it func- tions at the beginning of the stance phase, presumably to assist the hamstrings in preventing jacknife of the trunk (as Cu- vier suggested).
Most of the electromyographic studies of the gluteus maximus have involved recording from the “middle” of the mus- cle. However, because of the very special nature of the cranial portion, i t is essen- tial to learn whether this behaves differ- ently from the remainder. Jonsson and Steen (’63) and Karlsson and Jonsson (‘65) provide the essential information. Although both parts are equally active during extension of the thigh or trunk, a difference occurs during abduction. Pre- vious authors have speculated on a role of the cranial portion of the gluteus maxi- mus in abduction; some believed it was
possible, others denied it. The recent elec- tromyographic evidence indicates quite clearly that the gluteus maximus, p i n - cipnlly its csnninl past, is often called into action during forceful abduction of the thigh and during support on one leg. That the portion of the muscle most modi- fied morphologically should be the por- tion active in forceful abduction leads me to suggest that the evolutionary history of the gluteus maximus was directed to- ward improving the all important ability to control lateral stability of the trunk. All the changes which characterize the anterior portion are such as to improve its strength and leverage for abduction.
The abducting role of the gluteus maxi- mus in locomotion has not been studied, and it certainly should be. However, sev- eral interesting clues can be gathered from the literature. In walking on a level, the period of activity of the gluteus maxi- mus coincides with that of the gluteus medius. The two muscles have been thought to perform different functions during this period, but the possibility that they perform similar functions must exist. It is also of note that the glutei maxi- mus and medius contract simultaneously during climbing and descending stairs (Joseph and Watson, ’67). In bicycle pedal- ling, neither of these muscles are con- sistently active except when resistance to movement is great (Houtz and Fischer, ’59). Then, both muscles act during the same phase of the pedalling cycle. These various facts do not represent strong evi- dence that the functions of the glutei maximus and medius are often the same, or even partly so, but investigation of this possibility would seem worthwhile.
Although the gluteus maximus may function partially as a n abductor in bi- pedal walking, the evidence leads us to believe that such action is not essential for this mode of locomotion. I would like to speculate that the abductor function of the gluteus maximus may be more im- portant in modes of locomotion in which the unsupported weight of the body falls onto one limb from some height. Running is such a mode of locomotion. Slow run- ning, or jogging, would seem particularly noteworthy, since at low speeds the ver- tical oscillations of the center of gravity are greater than at high speeds. Also,

SPECIALIZATIONS OF HUMAN GLUTEUS MAXIMUS 329
in running, the trunk is somewhat flexed on the femur and the cranial fibers of the gluteus maximus would be in a more fa- vorable position for abduction.
The speculation can be carried further: is jogging such an important mode of loco- motion that the evolution of the gluteus maximus could be in response to its demands? An answer might be provided by considering primitive hunting beha- vior. Life studies of Bushmen (Garst, '69; and personal communication from R. Sing- er) indicate that jogging is the mode of locomotion used to track wounded prey for many miles. May not the necessity for the protracted hunt have placed selec- tive value on changes in the gluteus maximus which would enable it better to participate in controlling lateral sta- bility of the trunk? If so, then the develop- ment of the posterior gluteal line of the ilium and the expanded portion of the iliac blade behind this line may be im- portant osteological evidence of well estab- lished hunting behavior.
Dart ('49) stated that a posterior gluteal line could be traced with tolerable accu- racy on the left ilium from Makapans- gat. He noted that the posterior gluteal line passed much closer to the posterior iliac spines in Australopithecus prometh- eus ( = africanus) than in modern man, and, therefore, that the area of origin for the gluteus maximus was much small- er in the fossil. My examination of the Wenner-Gren Foundation cast (F-ST6) of the right innominate from A. ufricanus at Sterkfontein disclosed nothing which re- sembles a posterior gluteal line. Unfor- tunately, a portion of the fossil ilium just anterior to the posterior inferior spine is damaged. As a result, the possibility of a very atypical posterior gluteal line can- not be dismissed. The differences between the Makapansgat and Sterkfontein ilia suggest that the Makapan ape-man was practicing a more advanced bipedality than Australopithecus at Sterkfontein. This confirms the evidence provided by the ischia of the two forms. The Maka- pansgat specimen shows the greater simi- larity to modern man in possessing an ischial tuberosity that is more rounded and lies more dorsally and superiorly than in the Sterkfontein innominate (Broom and Robinson, '50; Broom, '50). If the
speculations offered above are correct, the Makapansgat inhabitants were venturing into the role of long distance predators.
Recently the major portion of a left in- nominate (Olduvai hominid 28) attrib- utable to Homo erectus has been described by Day ('71). It is stated that a posterior gluteal line is present. However, the pub- lished photographs of this specimen indi- cate some damage to the outer surface of the ilium just antero-superior to the posterior superior 'spine. This is the region that in modern man shows the clearest indication of the posterior gluteal line and enables one to estimate the size of the area devoted to the origin of the glu- teus maximus. We will probably need to await the discovery of a well-preserved ilium of Home erectus before we can find evidence that nature has completed those very inportant changes in the anterior portion of the superficial gluteus which are the hallmark of mankind.
ACKNOWLEDGMENTS
I wish to thank Drs. Ronald Singer, Charles Oxnard, and Mr. Timothy Strick- ler for reading this manuscript and pro- viding thoughtful comments and criti- cisms, I also wish to thank the Field Museum of Natural History (Dr. Karl Liem, Curator of Vertebrate Anatomy) for providing numerous valuable specimens for dissection. This work was supported by General Research Support grant PHS- RR-05367 and by National Sciepce Foun- dation grant GB-29296.
LITERATURE CITED
Appleton, A. B. 1921 The gluteal region of Tarsius spectrum. Proc. Cambr. Philos. Soc., 20: 466-474.
1927 The muscles and nerves of the post-axial region of the tetrapod thigh. J. Anat., Lond., 62: 364438.
Ayer, A. A. 1948 The anatomy of Semnopithe- cus entellus. The Indian Publ. House, Madras.
Battye, C. K., and J. Joseph 1966 A n investi- gation by telemetering of the activity of some muscles in walking. Med. & Biol. Engng., 4: 125-135.
Beddard, F. E. 1895 Contributions to the anat- omy of the anthropoid apes. Trans. Zool. SOC. Lond., 13: 177-219.
Bischoff, T. L. W. von 1870 Beitrage zur Anat- omie des Hylobotes lrirciscits und zu einer vergleichenden Anatomie der Muskeln der Af- fen und des Menschen. Abh. bayer. Akad. Wiss., math.-phys. Kl. , 10 (Abt. 3): 198-297.

JACK T. STERN. JR. 330
1880 Beitrage zur Anatomie des Go- rilla. Abh. bayer. Akad. Wiss., math-phys. KI., 13 (Abt. 3): 1-48.
Bluntschli, H. 1913 Die Fascia lata und ihre Bedeutung fiir die Umbildung des Gefassap- parates a n der unteren Gliedmasse in der Primatenreihe. Verh. Anat. Gesell. (27th Vers., Greifswald). Anat. Anz., 44 (suppl.): 43-66.
Boyer, E. L. 1935 The musculature of the in- ferior extremity of the orang-utan S imin scity- rus. Am. J. Anat., 56: 193-256.
Breitinger, E. 1959 On the earliest phase of hominid evolution. In: Ideas on Human Evolu- tion. 1962. W. W. Howells. ed. Harvard Univ. Press, Cambridge, pp. 172-202.
Broek, A. J . P. van den 1914 Studien zur Morphologie des Primatenbeckens. Morph. Jb. 49: 1-118.
Broom, R. 1950 The genera and species of the South African fossil ape-men. Am. J. Phys. Anthrop., 8: 1-13.
Broom, R., and J. T. Robinson 1950 Notes on the pelves of the fossil ape-men. Am. J . Phys. Anthrop., 8: 489-494.
Buettner-Janusch, J . 1966 Origins of man. John Wiley & Sons, N. Y.
Burdach, E. 1838 Beitrag zur vergleichenden Anatomie der Affen. Ber. Kon. anat. Anstalt Konigsberg, 9: 1-103.
Campbell, B. G. 1966 Human evolution. Al- dine, Chicago.
Champneys, F. 1871 On the muscles and nerves of a chimpanzee (Troglodytes niger) and a Cynocephal i is aizubis. J. Anat., Lond., 6: 1 76-21 1.
Cuvier, G. 1835 Leqons d’anatomie comparke. Second ed. Crochard, Paris.
Cuvier, G.. and C. L. Laurillard 1849 Anato- mie comparee recueil de planches myologie. Chez Dusacq, Paris.
Dart, R. 1949 Innominate fragments of AILS- trulopitlzrcits prometheus. Am. J. Phys. An- throp., 7: 301-334.
Day, M. H. 1971 Postcranial remains of Ho??ao erectits from Bed IV, Olduvai Gorge, Tanzania. Nature, 232. 383-387.
Duchenne, G. B. 1867 Physiology of motion. Lippincott, Phila., 1949.
Eberhart, H. D.: V. T. Inman and B. Bresler 1954 The principal elements in human loco- motion. In: Human Limbs and Their Substi- tutes. P. E. Klopsteg and P. D. Wilson, eds. McGraw-Hill, N. Y., pp. 437-471.
Fick, R. 1895a Vergleichend anatomische Stu- dien a n einem erwachsenen Oran-Utang. Arch. Anat. Physiol., Lpz., Anat. Abt., pp. 1-100.
189513 Beobachtungen an einem zweiten erwachsenen Orang-Utan und einem Schimpan- sen. Arch. Anat. Physiol., Lpz., Anat. Abt., pp. 289-318.
1925 Anatomische Untersuchungen an einigen der Teneriffaschimpansen, namentlich iiber die Gewichts- und Querschnitts-verhalt- nisse der Muskeln. S. B. yreuss. Akad. Wiss. Berlin, phys.-math. Kl., pp. 162-197.
Garst, W. E. 1969 Rulers of the Kalahari. A film distributed by Don Meier Productions, Chi- cago.
Haughton, S. 1865 Notes on animal mechanics. No. VIII-Further comparison of the hip joint
and knee joint muscles in the Cercopithecus, Cynocephalus and Macacus. Proc. Roy. Irish Acad., 9: 287-294.
Houtz, S. J., and F. J . Fischer 1959 An analysis of muscle action and joint excursion during ex- ercise on a stationary bicycle. J . Bone Jt. Surg.,
Howell, F. C. 1959 The Villafranchian and hu- man origins. Science, 130: 831-844.
Jonsson, B., and B. Steen 1963 Function of the hip and thigh muscles in Romberg’s test and “standing at ease.” An electromyographic study. Acta Morph. Neer1.-Scand., 5: 269-276.
Joseph, J., and R. Watson 1967 Telemetering electromyography of muscles used in walking up and down stairs. J. Bone Jt. Surg., 49B: 774-780.
Joseph, J., and P. L. Williams 1957 Electro- myography of certain hip muscles. J . Anat., Lond., 91: 286-294.
Juoffroy, F. K . 1962 La musculature des mem- bres chez les Lemuriens de Madagascar. Mam- nialia, 26: suppl. no. 2, 326 pp.
Kanagasuntheram, R. 1952 Observations on the anatomy of the hoolock gibbon. Ceylon J. Sci., Sect. G, 5: 11-64.
Karlsson, E., and B. Jonsson 1965 Function of the gluteus maximus muscle. An electromyo- graphic study. Acta Morph. Neal.-Scand., 6: 161-169.
Keith, A. 1894 The ligaments of the catarrhine monkeys, with reference to corresponding struc- tures in man. J. Anat. Physiol., Lond., 28: 149-169.
Klaatsch, H. 1900 Der kurze Kopf des Muscu- lus biceps femoris und der Tenuissimus. Morpb.
Die Erwerbung der aufrecten Hal- tung und ihre Folgen. Verh. Anat. Gesell. (27th Vers., Greifswald), Anat. Anz., 44 (suppl.): 161- 186.
Kleinschmidt, A. 1949 Funktionell-molpholo- gische Beobachtungen am Becken der Sauger unter besonderer Berucksichtingungender Prima- ten. Verh. Deutsch. Zoologen (1948, Kiel), pp. 95-103.
1951 Vergleichende Untersuchungen an der Extremitaten-muskulatur des Menschen und der Anthropoiden. Anat. Anz., 98 (suppl.): 153-1 63.
Kohlbrugge, 1. H. F. 1897 Muskeln und periph- ere Nerven der Primaten, mit besonderer Ber- ucksichtigung ihrer Anomalien. Verh. Kon. Akad. Weten., Amsterdam, Sect. 2, 5(6), 246 pp.
Lachman, 0. 1938 Zur topographischen Anat- omie der Hintergliedmasse des Hamadryas- Pavinns. Z . Morph. Anthrop., 37: 188-217.
Langer, C. 1879 Die Musculature der Extremi- taten des Orang als Grundlage einer vergleich- end-myologischen Untersuchung. S. B. Kais. Akad. Wiss. Wien, math.-naturw. Cl., 79 (Abt. 3): 177-222.
Le Gros Clark, W. E. 1967 Man-apes or ape- men? Holt, Rinehart and Winston, N. Y .
Lucae, J. C. G. 1884 Statik und Mechanik der Quadrupen a n dem Skelett und den Muskeln des Lemur und Clzoloepus. Abh. Senck. Naturf. Gesell. Frankfurt, 13: 1-92.
Macalister, A. 1873 The muscular anatomy of the gorilla. Proc. Roy. Irish Acad., I 1 : 501-506.
41Ar 123-131.
Jb., 29: 217-281. 1913

SPECIALIZATIONS OF HUMAN GLUTEUS MAXIMUS 331
Michaelis, P. 1903 Beitrage zur vergleichenden Myologie des Cynocephalus babuin, Simia saty- ms, Troglodytes niger. Archiv f. Anat. Physiol., Anat. Abt., pp. 205-256.
Miller, R. A. 1952 The musculature of Pan panisczis. Am. J. Anat., 91: 183-232.
Milne Edwards, A,, and A. Grandidier 1875 Histoire naturelle des mammiferes. Volume VI of Histoire physique, naturelle et politique de Madagascar (A. Grandidier). Imprimerie Na- tionale, Paris.
Mollier, G. 1937 Beziehungen zwischen Form und Funktion der Sehnen im Muskel-Sehnen- Knochen-System. Morph. Jb., 79: 161-199.
Murie, J., and St. G. Mivart 1872 On the . . . . .. - anatomy of the Lemuroidea. Trans. Zool. SOC. Lond.. 7: 1-114.
Mysberg, W. A. 1917 Uber die Verbindungen zwischen dem Sitzbeine und der Wirbelsaule bei den Saugetieren. Anat. Hefte, Abt. 1, 54: 64 1-668.
Napier, J. R. 1967 The antiquity of human walking. Sci. Amer., 21 6: 5-6.
Pagenstecher, H. A. 1867 Mensch und Affe. Ein Vergleich der Muskulatur des Drill mit der des Menschen, unter Beriicksichtigung allge- meiner Gesichtspunkte der Muskellehre und der Unterschiede von Hand und Fuss. Zool. Garten (Frankfurt), 8: 121-137.
Patterson, E. L. 1942 The myology of Rhino- pi thecus roxellanae and Cynopithecus niger. Proc. 2001. SOC. Lond., 112(B): 31-104.
Pauly, J. E., and L. E. Scheving 1968 An elec- tromyographic study of some hip and thigh muscles. in man. Electromyography, 8 (suppl.
Polak, C. 1908 Die Anatomie des Genus Colo- bus. Verh. Kon. Akad. Weten., Amsterdam, Sect. 2, 14(2), 247 pp.
Preuschoft, H. 1961 Muskeln und Glelenke der Hinterextremitat des Gorillas. Morph. Jb., 101: 432-540.
Primrose, A. 1898-1899 The anatomy of the Orang outang (S imia satyrus), a n account of some of its external characteristics: and the myology of the extremities. Trans. Canad. Inst. (Toronto), 6: 507-598.
Ranke, K. 1897 Muskel- und Nervenyariationen der dorsalen elemente des Plexus ischiadicus der Primaten. Arch. f. Anthrop., 24: 117-144.
Riska, N. 1936 Om glutealregionens muskler hos Primates. Finska Lak. Handl., 79: 637-658.
Robinson, J. T. 1963 Adaptive radiation in the australopithecines and the origin of man. In: African Ecology and Human Evolution. F. C. Howell and F. Bourliere, eds. Aldine, Chicago, pp. 385416.
The origin and adaptive radiation of the australopithecines. In: Evolution and Hominisation. G. Kurth, ed. Gustav Fischer, Stuttgart, pp. 150-175.
Sappey, Ph. C. 1876 Trait6 danatomie descrip- tive. Tome 2. Delahaye, Paris.
Schumacher, G. H., and E. Wolff 1966 Trock- engewicht und physiologischer Querschnitt der
1 ) : 131-147.
1968
menschlichen Skelettmuskulatur. I . Trockenge- wicht. Anat. Anz., 118: 317-330.
Schumacher, O., and V. Lebzelter 1924 Zur Kenntniss der Oberschenkelmuskulatur bei Mor- m o n sphinx . 2. Anat. EntwGesch., 71: 282- 303.
Sigmon, B. A. 1971 Bipedal behavior and the emergence of erect posture in man. Am. J . Phys. Anthrop., 34: 5.560.
Sigmon, B. A,, and J. T. Robinson 1967 On the function of the M. gluteus maximus in apes and man. Am. J. Phys. Anthrop., 27: 245-246 (Abstract).
Snyder, R. C. 1967 Adaptive values of biped- alism. Am. J. Phys. Anthrop., 26: 131-134.
Sonntag, C. F. 1924 On the anatomy, physiol- ogy, and pathology of the orang-outan. Proc. Zool. SOC. Lond., pp. 349-450.
Stern, J. T., Jr. 1971 Functional myology of the hip and thigh of cebid monkeys and its impli- cations for the evolution of erect posture. Bib- lio. Primatol., No. 14. s. Karger, Basel.
Sutherland, D. H., F. C. Bost and E. R. Schott- staedt 1960 Electromyographic study of transplanted muscles about the knee in polio- myelitic patients. J. Bone Jt. Surg., 42A: 919-938.
Tappen, N. C. 1955 Relative weights of some functionally important muscles of the thigh, hip and leg in a gibbon and in man. Am. J . Phys. Anthrop., 13: 415420.
Trail], T. S. 1818 Observations on the anatomy of the Orang Outang. Mem. Wernerian Nat. Hist. SOC., 3: 1 4 9 .
Tyson, E. 1969 Orang-Outang, sive Homo Syl- vestris: or, the anatomy of a pygmie compared with that of a m o n k e y , an ape, and a Man. Thomas Bennet, London.
Uhlmann, K. 1968 Huft- und Oberschenkel- muskulatur. Systematische und vergleichende Anatomie. Primatologia, 4 , No. 10,442 pp.
Voss, H. 1956 Tabelle der Muskelgewichte des Mannes, berechnet und zusammengestelt nach dem Untersuchungen von W. Theile (1884). Anat. Anz., 103: 356-360.
Washburn, S. L. 1951 The analysis of primate evolution with particular reference to the origin of man. Cold Spring Harb. Sympos. Quant. Biol., IS: 67-78.
Waterman, H. C. 1929 Studies on the evolu- tion of the pelvis of man and other primates. Bull. Amer. Mus. Nat. Hist., 58: 585639.
Wheatley, M. D., and W. D. Jahnke 1951 Elec- tromyographic study of the superficial thigh and hip muscles in normal individuals. Arch. Phys. Med., 32: 508-515.
Wiedersheim, R. 1895 The structure of man. Translated by H. and M. Bernhard. Macmillan, London.
Wilder, B. G. 1861 Contributions to the com- parative myology of the chimpanzee. Bost. J. Nat. Hist., 7: 353-384.
Zielinska, H. 1936-1937 Antropomorfologja M. glutaei maximi. Folia Morph. (Warszawa), 7: 84-1 13.

PLATE 1
EXPLANATION OF FIGURE
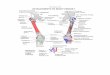
1 Lateral view of the right hip and thigh demonstrating the general con- figuration of the gluteus superficialis (GS) or gluteus maximus (GM). BLH, long head of biceps femoris; BSH, short head of biceps femoris; FCL, flexor cruris lateralis; STL, sacro-tuberous ligament; TFF, tensor fasciae femoris. A - Galago senegalensis; B - Theropithectis gelada; C - Pongo pygmaeiis; D - P a n troglodytes; E - H o m o snpiens fetus; F - H o m o sapiems, adult. The “primitive” condition is demonstrated in Gnlngo. Note that in Old World monkey ( B ) almost the entire muscle inserts into the fascia of the thigh. In the orang-utan, the entire muscle inserts tendinously into the proximal region of the femur. The origin of the gluteus superfacialis in the chimpanzee differs from the “primitive” state in that the muslce arises largely from the sacro- tuberous ligament and ischial tuberosity. Note that in man the ante- rior fibers of the gluteus maximus insert into the overlying fascia and that the posterior fibers do not extend far down the thigh.
332

SPECIALIZATIONS OF HUMAN GLUTEUS MAXIMUS Jack T. Stern, Jr.
PLATE I

PLATE 2
EXPLANATION O F FIGURE
2 Lateral view of the right hip and thigh with gluteus superficialis (GS) or gluteus maximus (GM) cut to show insertion of the anterior portion via the ascending tendon (AT) in all forms except Old World monkeys. GMp, posterior (inferior) portion of gluteus maximus; GSp, posterior portion of gluteus superficialis; TFF, tensor fasciae femoris. A - Ga- lago senegalensis; B - Sa imir i sciureus; C - M a n d r i l l u s s p h i n x ; D - Pongo p y g m a e u s ; E - H o m o sup iens fetus; F - Homo sap iens adult. The “primitive” condition, characterized by a narrow ascending ten- don inserting quite proximally on the femur and a large posterior muscular portion inserting over the major portion of the femoral shaft, is seen in Galago and Saimiri . Note the absence of an ascending tendon in the Old World monkey (C), and the distal expansion of this structure in man. The insertion of the posterior portion of the gluteus maximus into the aponeurosis of the vastus lateralis can be seen in the adult specimen of man.
334

SPECIALIZATIONS OF HUMAN GLUTEUS MAXIMUS Jack T. Stern, Jr.
PLATE 2

PLATE 3
EXPLANATION O F FIGURE
3 Lateral view of the right hip and thigh with bisected gluteus super- ficialis (GS) or gluteus maximus (GM) demonstrating the relative thicknesses of the anterior and posterior portions of the muscle. AT, ascending tendon; GSp, posterior portion of gluteus super- ficialis. A - Propithecus uerrauxi; B - Ateles geoffroyi; C - Alounttu cnraya; D - Cercopithecus aethiops; E - Homo sapiens fetus; F - Homo sapiens adult. Note that the anterior portion of the muscle is thicker, usually markedly so, than the posterior portion in all pri- mates except man.
336

SPECIALIZATIONS OF HUMAN GLUTEUS MAXIMUS Jack T. Stern, Jr.
PLATE 3

w w m
PLATE 4
EXPLANATION OF FIGURE
4 Reproduction of figures 315 (left) and 316 (right) from Sappey (1876). The gluteal region of man is depicted. Sappey employs the following labels: Figure 315-1 = gluteus maximus; 2 = inferior portion of gluteus maximus; 3 = tendinous bundles by which the inferior por- tion attaches to the whole extent of the external bifurcation of linea aspera; 4 = superior part of gluteus maximus; 5 = tendinous rih- bons by which superior part also goes to insert on external bifurca- tion of linea aspera; 6 = superior part of the femoral aponeurosis; 7 = doubling of this aponeurosis at the level of the superior border of the gluteus maximus; 8 = portion of its superficial lamina that ad- heres tightly to the tendon ribbons; 9 = inferior extremity of tensor fasciae latae; 10 = portion of the femoral aponeurosis continuous with the tendinous bundles of the tensor, excised in order to show insertion of gluteus maximus to femur. Figure 316 - 1 = gluteus max- imus; 2 = insertion of gluteus maximus to external bifurcation of linea aspera; 3 = triangular aponeurosis formed by converging ten- dinous bundles of the superior part of gluteus maximus.

SPE
CIA
LIZ
AT
ION
S O
F H
UM
AN
GLUTEUS M
AX
IMU
S Ja
ck T
. S
tern
, Jr.
PL
AT
E 4