Embed Size (px)
Citation preview

QUATERNARY RESEARCH 32, 335-341 (1989)
SHORT PAPER
A Last Interglacial Fauna from the Eastern Sahara
KAZIMIERZ KOWALSKI
Institute of Systematic and Experimental Zoology, Polish Academy of Sciences, ul. Stawkowska 17, 31-016 Krakdw, Poland
WIM VAN NEER
Royal Museum of Central Africa, B-1980 Tervuren, Belgium
ZYGMUNT BOCHEASKI, MARIAN MLYNARSKI, BARBARA RZEBIK-KOWALSKA, AND ZBIGNIEW SZYNDLAR
Institute of Systematic and Experimental Zoology, Polish Academy of Sciences, ul. Sfawkowska 17, 31-016 Krakbw, Poland
ACHILLES GAUTIER
Institute of Geology, State University of Ghent, Krtjgslaan 281lS8, B-9000 Ghent, Belgium
ROMUALD SCHILD
Institute for the History of Material Culture, Polish Academy of Sciences, ul. Swierczewskiego 105, 00-140 Warsaw, Poland
AND
ANGELA E. CLOSE AND FRED WENDORF
Department of Anthropology, Southern Methodist University, Dallas, Texas 75275
Received April 24, 1989
Recent work on the middle Paleolithic at Bir Tarfawi, in the hyperarid Eastern Sahara (< 1 mm of ram per annum), has yielded a rich faunaf assemblage, including several thousand remains of fish, amphibians, reptiles, birds, and small mammals. They are derived from the sediments of two consecutive lakes dated by several techniques to about 135,000 yr B.P. Fifty-nine taxa have been identified and indicate that at times during the Last Interglaciation the area received at least 500 mm of rainfall as a result of the northward shift of the monsoon belt, and that, on several occasions, there may have been water connections between Bir Tarfawi and unidentified but permanent bodies of water elsewhere. o 1989 University of Washington.
INTRODUCTION
Bir Tarfawi is a 4 x II-km deflational basin in southwestern Egypt (Fig. l), the geological history of which is tied to fluctu- ations in the water table. During periods of hyperaridity, the water table would fall be- low the surface and deflation would remove the softer sediments, creating hollows of various sizes. During periods of rainfall,
both locally and farther south, the water table would rise and lakes would form in the deflated hollows. There were six middle Pa- leolithic lake episodes in the Tarfawi basin, separated by intervals of hyperaridity and aeolian deflation. From earliest to latest, the Iakes have been named the Silt-Pan, the White Lake, Grey Lakes l-3, and the Green Lake (the last five from the color of their sediments).
335 0033-5894/89 $3.00 Copyright 0 1989 by the University of Washington. All rights of reproduction in any form reserved.
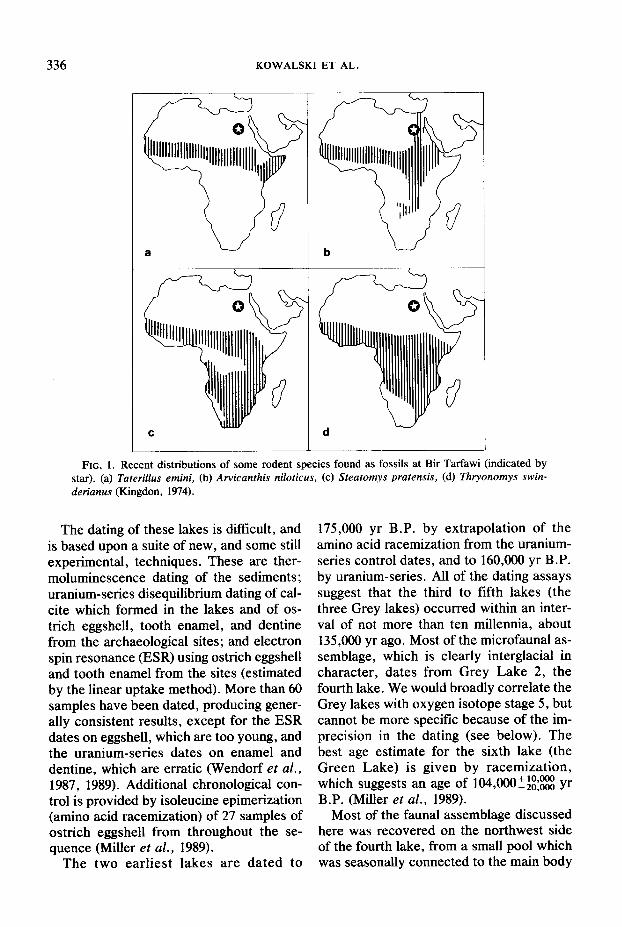
336 KOWALSKI ET AL.
FIG. 1. Recent distributions of some rodent species found as fossils at Bir Tarfawi (indicated by star). (a) Taterillus emini, (b) Arvicanthis niloticus, (c) Steatomys pratensis, (d) Thryonomys swin- derianus (Kingdon, 1974).
The dating of these lakes is difficult, and 175,000 yr B.P. by extrapolation of the is based upon a suite of new, and some still amino acid racemization from the uranium- experimental, techniques. These are ther- series control dates, and to 160,000 yr B.P. moluminescence dating of the sediments; by uranium-series. All of the dating assays uranium-series disequilibrium dating of cal- suggest that the third to fifth lakes (the cite which formed in the lakes and of os- three Grey lakes) occurred within an inter- trich eggshell, tooth enamel, and dentine val of not more than ten millennia, about from the archaeological sites; and electron 135,000 yr ago. Most of the microfaunal as- spin resonance (ESR) using ostrich eggshell semblage, which is clearly interglacial in and tooth enamel from the sites (estimated character, dates from Grey Lake 2, the by the linear uptake method). More than 60 fourth lake. We would broadly correlate the samples have been dated, producing gener- Grey lakes with oxygen isotope stage 5, but ally consistent results, except for the ESR cannot be more specific because of the im- dates on eggshell, which are too young, and precision in the dating (see below). The the uranium-series dates on enamel and best age estimate for the sixth lake (the dentine, which are erratic (Wendorf et al., Green Lake) is given by racemization, 1987, 1989). Additional chronological con- which suggests an age of 104,000~~~~~ yr trol is provided by isoleucine epimerization B.P. (Miller et al., 1989). (amino acid racemization) of 27 samples of Most of the fauna1 assemblage discussed ostrich eggshell from throughout the se- here was recovered on the northwest side quence (Miller et al., 1989). of the fourth lake, from a small pool which
The two earliest lakes are dated to was seasonally connected to the main body

SAHARAN INTERGLACIAL FAUNA 337
of water; the organic fraction of sediments from the pool has yielded a radiocarbon date between 40,000 and 50,000 yr B.P. (H. Haas, personal communication, 1989). The sample is not radiometrically dead, presumably because of contamination, but demonstrates that the pool was not of Ho- locene age. Here, the lower sediments are clays or clay marls, rich in organic material, which laterally grade upward into an or- ganic-rich peaty soil. Some 2 m3 of this peaty sediment from the lake margin was
screened and washed. The recovered re- mains include 51 identified taxa of amphib- ians, reptiles, birds, and small mammals and 8 identified taxa of freshwater fish.
The composition of the nonfish fauna is given in Table 1. The crocodiles and water turtles (Pelusios adansoniz) indicate an im- portant lake, which must at times have been connected to the river net of sub- Saharan Africa, because these species can travel only short distances over land. Waterbirds (Phalacrocorax africanus, Nyc-
TABLE 1. TERRESTRIALVERTEBRATESFROMTHETHIRDMIDDLEPALEOLITHIC LAKE AT BIRTARFAWI
Amphibians cf. Ranidae indet.
Reptiles Lacertidae indet. Agamidae indet. cf. Scincidae indet. Eryx sp. Colubridae sp. 1 Colubridae sp. 2 Colubridae sp. 3 Viperidae indet. Crocodylus sp. Pelusios adansonii
Mammals Crocidura cf. jiilvastra Crocidura cf. j7oweri Crocidura cf. religiosa Chiroptera sp. 1 Chiroptera sp. 2 Chiroptera sp. 3 Thryonomys swinderianus Heliosciurus cf. gambianus Tatera valida Tatera robusta Taterillus cf. emini
Number of fragments MNI”
Number of fragments MN1
2 1
2 1 1 1 2 1 3 1 1 1 2 1 2 1 1 1
Many teeth 455 *30
7 3 Meriones cf. crassus 10 40 27 Desmodilliscus braueri 2 11 6 Gerbil& sp. 1 (large) 41 2 2 Gerbillus sp. 2 (small) 10 1 I Steatomys pratensis 10 1 1 Steatomys parvus 3 2 1 Arvicanthis niloticus 8 1 1 Lemniscomys cf. barbarus 7
17 4 Mus haussa 2 7 2 Acomys wilsoni 11
76 20 Praomys natalensis 6
Birdsb Phalacrocorax africanus Nycticorax nycticorax Ardeola sp. Ardea cinerea Plegadis falcinellus Gyps africanus Circus sp. Turnix cf. sylvatica Porzana cf. pusilla Gallinula chloropus Fulica cristata Chlamydotis undulata Otididae indet. (large) Pterocles sp. Oena capensis Caprimulgidae indet. Riparia sp. Corvus cf. albus Corvus sp.’
>loo” 30 2 1 3
id 2 3 1 1 1 1 4 2 1 1 1 3
10 5 1 1 1 1 2 1 2 1 1 1 1 4 1 1 1 1 1
3 2 9 5 5 3 2 2 2 5 1
a Minimum number of individuals. b Several dozen fragments remain unidentifiable. c There are many very small fragments. d These are most probably two different species. e Of the size of Corvus corone.

338 KOWALSKI ET AL.
ticorax nycticorax, Ardeola sp., Ardea ci- nerea, Plegadis falcinellus, Porzana cf. pu- silla, Gallinula chloropus, and Fulica cris- tata) inhabited the emergent vegetation of the lake and bushes or trees around its shore. The shore of the lake was also the biotope of shrews (Crocidura religiosa and C. floweri) and of rodents such as Thryo- nomys swinderianus, Steatomys pratensis, and Arvicanthis niloticus.
Most of the forms present now occur in semiarid environments. Snakes of the ge- nus Eryx live in loose, dry sand. Of the birds, Gyps africanus lives in grassland, sa- vanna, and acacia woodland, Oena capen- sis is a savanna bird, while Chlamydotis un- dulata inhabits arid open plains and grass- land. The shrew Crocidura fulvastra lives in dry savanna. In the rodent fauna, Helios- ciurus gambianus is characteristic of open woodland, but the more numerous gerbil- lids of the genera Tatera, Taterillus, and Desmodilliscus inhabit dry savanna, as do the murids Lemniscomys barbarus and Acomys wilsoni. Only a few elements are true desert animals (Meriones crassus and perhaps Gerbillus spp.), but they also oc- cupy barren areas and dunes in the sa- vanna. Overall, the land vertebrates sug- gest that the lake was large, rather shallow, and eutrophic, with reed beds over much of its surface. The lake most probably was surrounded by savanna and there were clumps of trees close to the water. More arid habitats, like sparse woodland, grass- land, and semidesert, occurred in the vicin- ity.
Excavations elsewhere in deposits of the three Grey and Green lakes have yielded both bones and eggshells of ostrich and re- mains of larger mammals which seem to in- clude white rhinoceros (Ceratotherium si- mum); giraffe (Giraffa camelopardalis sp.); a buffalo (Pelorovis antiquus); wild camel (Camelus thomasi); wild ass (Equus asi- nus); large antelopes, including giant eland (Taurotragus derbianus) and others; a re- duncine antelope, probably kob (Kobus kob); large gazelle (Gazella dama); small
gazelle (G. dorcas?); warthog (Phacochoe- rus aethiopicus); spotted hyena (Crocuta crocuta) and probably striped hyena (Hyaena hyaena); jackal (Canis aureus?); fox (Vulpes vulpes); small cat, probably the wild cat (Felis silvestris); porcupine (Hys- trix cristata); and hare (Lepus capensis). These collections are still being analyzed.
The fauna from the peaty pool sediments contains only extant species (there are ex- tinct forms among the large mammals) and nearly all the nonflying elements now occur in sub-Saharan Africa, with a few (Croci- dura religiosa, Arvicanthis niloticus) along the Nile in Egypt; Crocidura floweri is not now known outside Egypt. The northern limit of the distributions of the rodent spe- cies is now about 250 km south of Bir Tar- fawi, and some of them occur almost 1200 km to the south (Fig. l), which corre- sponds approximately to the 500-mm iso- hyet. We may therefore take 500 mm as our best estimate of the minimum rainfall dur- ing this period. The recent distributions of the birds are more varied (Fig. 2), but there is no evidence of European migrants at Bir Tarfawi for all the European taxa found there breed in Africa.
Eight taxa of freshwater fish have been identified from the third and fourth middle Paleolithic lakes, and at least one more spe- cies remains unidentifiable (Table 2). The fish were very small and it is therefore likely that many of them represent individ- uals that died in a seasonally dry pool. They could also have been partly deposited by the local colony of cormorants.
Like the crocodiles and water turtles in the fourth lake, the presence of fish in at least two of the lakes means either that they survived the intervening arid phases from a pre-middle Paleolithic period or that they recolonized Bir Tarfawi. The Nile perch, Lutes niloticus, requires deep, well- oxygenated water, and adult Bagrus also prefers deep water, whereas Alestes is a pe- lagic fish. Thus, any residual pond would have had to be of considerable extent to maintain such a varied fauna. However,

SAHARAN INTERGLACIAL FAUNA 339
FIG. 2. Recent distributions of some bird species found as fossils at Bir Tarfawi (indicated by star). (a) Gallinula chloropus: Europe, Asia (Cramp, 1980), and Africa (Urban et al., 1986); (b) Chlamydotis undulatn: Asia (Cramp, 1980) and Africa (Urban et al., 1986); (c) Phalacrocorax africanus: subsaharan Africa (water bird) (Brown et al., 1982); (d) Oena cupensis: subsaharan Africa (savanna) (Urban et al., 1986).
stratigraphic evidence suggests that there was no surface water during the arid phases, so recolonization must have oc- curred for each lake episode.
In general, fishes can colonize an area only through water connections. Other kinds of dispersal are theoretically possible (Gudger, 1929; Roberts, 1975), but they are unlikely to play a role in long-distance movement and, in any case, do not apply to all the taxa found at Tarfawi. The fish must have come by water from areas where they survived the preceding arid periods. The Tarfawi fish taxa are typical of the present- day Nile, Chad, and Niger basin faunas (Daget and Durand, 1981), which were probably dispersed across Africa during the
Tertiary. There are no effects of geograph- ical separation, such as speciation, visible today between the various North African basins, which also indicates that exchanges occurred during the Pleistocene. The routes of these exchanges remain unknown, but may have followed the buried channels dis- covered by ground-penetrating radar (Mc- Cauley et al., 1982).
The fauna from the third and fourth mid- dle Paleolithic lakes at Bir Tarfawi indicate that the Eastern Sahara enjoyed rainfall of at least 500 mm per annum. This is consid- erably more than fell during the humid pe- riods of the Holocene; Ritchie et al. (1985) have estimated that as much as 400 mm of rain may have fallen to the south in north-

340 KOWALSKI ET AL.
TABLE 2. FISH TAXA FROM THE THIRD MIDDLE PALEOLITHIC LAKE AT BIR TARFAWI
Fragments
toric Expedition, which is jointly sponsored by the Polish Academy of Sciences, the Geological Survey of Egypt, and Southern Methodist University. The field- work was supported by National Science Foundation
Number % MNI Grant BNS-8518574 to Fred Wendorf.
Tilapia 1849 Clarias 1452 Synodontis 303 Lutes 127 Alestes 12 Mormyridae 10 Bagrus 2 Polypterus 1
49.2 81 38.7 116 8.1 139 3.4 15 0.3 2 0.3 1 + 1 + 1
ern Sudan in the early Holocene, but in southern Egypt the corresponding figure is less than 100 mm per annum (Neumann, 1987; Peters, 1988). The middle Paleolithic environments were therefore much more lush than those witnessed by Neolithic in- habitants of the Eastern Sahara.
Wetter conditions in the Holocene south- ern Sahara were a response to intensified monsoons (COHMAP Members, 1988) and the southern character of the terrestrial fauna suggests that the same was true in the middle Paleolithic; the lack of nonflying Mediterranean forms indicates that there was an uninterrupted desert belt to the north. This presents a problem in dating, since Prell and Kutzbach (1987) have found evidence for strong, monsoon-related events 126,000, 104,000,82,000, and 10,000 yr ago, corresponding to the maxima of northern hemisphere summer radiation (Imbrie et al., 1984). Green Lake might cor- respond to the 104,000 yr B.P. event, but the Grey lakes (third to fifth lakes), as cur- rently dated, seem somewhat older than the 126,000 yr B.P. event. In spite of this ap- parent discrepancy, which is most probably a reflection of the remaining imprecision in the dating of the lakes, the dates and the interglacial character of the associated fauna do suggest that the Grey lakes corre- late with some part of oxygen isotope stage 5.
ACKNOWLEDGMENTS
This study was conducted by the Combined Prehis-
REFERENCES
Brown, L. H., Urban, E. K., and Newman, K. (Eds.) (1982). “The Birds of Africa I.” Academic Press, Orlando, FL.
COHMAP Members (1988). Climatic changes of the last 18,000 years: Observations and model simula- tions. Science 241, 1043-1052.
Cramp, S. (Ed.) (1980). “Handbook of the Birds of Europe, the Middle East and North Africa. The Birds of the Western Palearctic. II. Hawks to Bustards.” Oxford Univ. Press, Oxford.
Daget, J., and Durand, J.-R. (1981). Poissons. In “Flore et faune aquatiques de 1’Afrique sahelo- soudanienne” (J.-R. Durand and C. Lev@que, Eds.), pp. 687-771. ORSTOM, Paris.
Gudger, E. W. (1929). More rains of fishes. Annual Magazine of Natural History 10(3), l-24.
Imbrie, J., Hayes, J. D., Martinson, D. G., McIntyre, A., Mix, A. C., Morley, J. J., Pisias, N. G., Prell, W. L., and Shackleton, N. J. (1984). The orbital theory of Pleistocene climate: Support from a re- vised chronology of the marine 6i*O record. In “Mi- lankovitch and Climate Part 1” (A. Berger, J. Im- brie, J. Hays, G. Kukla, and B. Saltzmann, Eds.), pp. 269-305. Reidel, Hingham, MA.
Kingdon, J. (1974). “East African Mammals IIB.” Ac- ademic Press, New York.
McCauley, J. F., Schaber, G. G., Breed, C. S., Gro- lier, M. J., Haynes, C. V., Issawi, B., Elachi, C., and Blom, R. (1982). Subsurface valleys and geoar- chaeology of the Eastern Sahara revealed by shuttle radar. Science 218, 1004-1020.
Miller, G. H., Wendorf, F., Ernst, R., Close, A. E., Schild, R., Friedman, I., and Schwartz, H. P. (1989). Racemization chronology of high lake level events in the Eastern Sahara. Palaeogeography, Palaeoclimatology, Palaeoecology.
Neumann, K. (1987). Middle Holocene vegetation of the Gilf Kebir/SW Egypt-A reconstruction. In “Palaeoecology of Africa” (J. A. Coetzee, Ed.), Vol. 18, pp. 17%188. Balkema, Rotterdam.
Peters, J. (1988). The palaeoenvironment of the Gilf Kebir-Jebel Uweinat area during the first half of the Holocene: The latest evidence. Sahara 1, 73-76.
Prell, W. L., and Kutzbach, J. E. (1987). Monsoon variability over the past 150,000 years. Journal of Geophysical Research 92(D7), 8411-8425.
Ritchie, J. C., Eyles, C. H., and Haynes, C. V. (1985). Sediment and pollen evidence for an early to

SAHARAN INTERGLACIAL FAUNA 341
mid-Holocene humid period in the eastern Sahara. Nature (London) 314, 352-355.
Roberts, T. R. (1975). Geographical distribution of Af- rican freshwater fishes. Zoological Journal of the Linnaean Society 51, 249-319.
Urban, E. K., Fry, C. H., and Keith, S. (Eds.) (1986). “The Birds of Africa II.” Academic Press, Orlando, FL.
Wendorf, F., Close, A. E., and Schild, R. (1987). Re-
cent work on the Middle Palaeolithic of the Eastern Sahara. African Archaeological Review 5, 49-63.
Wendorf, F., Close, A. E., Schild, R., Gautier, A., Schwartz, H. P., Miller, G. H., Kowalski, K., KnYik, H., Bluszcz, A., Robins, D., Griln, R., and McKinney, C. (1989). Chronology and stratigraphy of the middle Paleolithic at Bir Tarfawi, Egypt. In “Lower and Middle Palaeolithic of Africa” (J. D. Clark, Ed.). UISPP, Mainz.





























