Embed Size (px)
Citation preview

Palaeogeography, Palaeoclimatology, Palaeoecology 296 (2010) 185–198
Contents lists available at ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r.com/ locate /pa laeo
Last glacial–interglacial vegetation and environmental dynamicsin southern Siberia: Chronology, forcing and feedbacks
Elena V. Bezrukova a, Pavel E. Tarasov b,⁎, Nadia Solovieva c, Sergey K. Krivonogov d, Frank Riedel b
a Institute of Geochemistry, Siberian Branch Russian Academy of Sciences, Favorsky Str. 1a, Irkutsk, 664033, Russiab Institute of Geological Sciences, Palaeontology, Free University, Malteserstr. 74-100, House D, Berlin, 12249, Germanyc Department of Geography, University College London, Gower Str., London WC1E 6BT, UKd Institute of Geology and Mineralogy, Russian Academy of Sciences, Siberian Branch, Koptug ave. 3, Novosibirsk, 630090, Russia
⁎ Corresponding author. Tel.: +49 30 83870280; fax:E-mail addresses: [email protected] (E.V. Bezruko
[email protected] (P.E. Tarasov), nsolovie@[email protected] (F. Riedel).
0031-0182/$ – see front matter © 2010 Elsevier B.V. Aldoi:10.1016/j.palaeo.2010.07.020
a b s t r a c t
a r t i c l e i n f oArticle history:Received 15 January 2010Received in revised form 18 July 2010Accepted 21 July 2010Available online 25 July 2010
Keywords:EurasiaPollen and diatom recordsQuantitative vegetation reconstructionLate Pleistocene interstadialsLast glacial maximumYounger DryasHolocene climatic optimum
Radiocarbon-dated pollen and diatom records from Lake Kotokel in southern Siberia help to reconstruct theenvironmental history of the area since ~47 kyr BP. Pollen spectra composition and reconstructed biomescores suggest predominance of a tundra–steppe vegetation and variable woody cover (5–20%) between ~47and 30 kyr BP, indicating generally a harsh and unstable climate during this interval, conventionally regardedas the interstadial within the last glacial. The short-term climate amelioration episodes in the glacial part ofthe records are marked by the peaks in taiga and corresponding minima in steppe biome scores and appearsynchronously with the hemispheric temperature and precipitation changes recorded in the Greenland icecores and Chinese stalagmites. Transition to full glacial environments occurred between 32 and 30 kyr BP.The interval at ~30–24 kyr BP was probably the driest and coldest of the whole record, as indicated byhighest scores for steppe biome, woody coverage b5%, absence of diatoms and reduced size of the lake. Aslight amelioration of the regional climate at ~24–22 kyr BP was followed by a shorter than the previous andless pronounced deterioration phase. The late-glacial (~17–11.65 kyr BP) is marked by a gradual increase intree/shrub pollen percentages and re-appearance of diatoms. After 14.7 kyr BP the climate became warmerand wetter than ever during ~47–14.7 kyr BP, resulting in the deepening of the lake and increase in thewoody coverage to 20–30% ~14.5–14 kyr and ~13.3–12.8 kyr BP. These two intervals correspond to theMeiendorf and Allerød interstadials, which until now were interpreted as part of the undifferentiatedBølling/Allerød interstadial complex in the Lake Baikal region. The increase in tundra biome scores andpronounced change in the diatom composition allow (for the first time) the unambiguous identification ofthe Younger Dryas (YD) in the Lake Baikal region at ~12.7–11.65 kyr BP, in agreement with the formaldefinition and dating of the YD based on the Greenland NGRIP ice core records. The maximal spread of thetaiga communities in the region is associated with a warmer and wetter climate than the present prior to~7 kyr BP. This was followed by a wide spread of Scots pine, indicating the onset of modern environments.
+49 30 83870745.va),og.ucl.ac.uk (N. Solovieva),
l rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
Worldwide terrestrial and marine sedimentary archives demon-strate that the last 50-kyr interval in the Earth's history experienced anumber of long- and short-term climatic oscillations (Guiot et al.,1989; Vincens et al., 2005; Heusser et al., 2006; Bout-Roumazeilles etal., 2007; Svensson et al., 2008). High-resolution and accurately datedpollen and sedimentary records of the late-glacial/early Holoceneinterval exist for several regions of Europe (Litt and Stebich, 1999;Allen and Huntley, 2000; Brauer et al., 2008) and East Asia (Stebich
et al., 2009), providing important insight into the environmentaldynamics in the North Pacific and North Atlantic regions. However, arecent global-scale synthesis of the Holocene climatic data (Wanner etal., 2008) demonstrates a lack of palaeorecords of comparable datingquality and resolution from the vast areas of Eurasia, including Siberiaand Central Asia. The dating problem becomes even more obvious,when the pre-Holocene interval of the late Quaternary is considered(Rudaya et al., 2009; Shichi et al., 2009).
Southern Siberia – the region of Russia between ~80–120°E and~50–60°N – consists of numerous sub-latitudinal mountain rangesand lakes, including Lake Baikal in the east (Fig. 1a and b). The lakesediments are one of the most promising sources of detailedpalaeoenvironmental information, which provide an opportunity forbridging the European and Asian palaeoclimate archives and addres-sing critical questions concerning Quaternary climatology, e.g.reconstructing atmospheric circulation patterns (Williams et al.,

Fig. 1. Simplified maps, which show: (a) northern Eurasia and the Lake Baikal region, (b) the Lake Kotokel study area, (c) location of the 47-kyr long KTK2 core (this study) and the15-kyr long KTK1 core (Tarasov et al., 2009). The location of Lake Sihailongwan (SHL) is indicated by a black triangle.
186 E.V. Bezrukova et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 296 (2010) 185–198
2001), and palaeoecology, e.g. discussing local presence/absence ofboreal trees and shrubs during the last glacial (Grichuk, 1984).
Numerous publications on the Lake Baikal region (e.g. Kuzmin etal., 1993; Williams et al., 1997; Prokopenko et al., 2002; Shichi et al.,2009 and references therein) presented coarse-resolution (millenni-al- or multi-century-scale) qualitative reconstructions of the Quater-nary environments. Although the long cores from Lake Baikal spanmillions of years (Williams et al., 1997; Khursevich et al., 2005),research was mainly focused on the Holocene and earlier interglacials(Demske et al., 2005; Tarasov et al., 2005, 2007a; Prokopenko et al.,2007, 2010). However, little is known about glacial intervals due tothe problems associated with very low pollen concentrations, poororganic content, low sedimentation rates and poor dating (Horiuchi etal., 2000; Oda et al., 2000). Until recently, even the YD cooling was notunequivocally identified and dated in the Baikal records. This led tovarious hypotheses suggesting that Lake Baikal could either mask ordelay the effect of global change (Bezrukova et al., 2005; Demske et al.,2005; Shichi et al., 2009).
This studypresents newpollen anddiatomrecords fromLakeKotokel(Fig. 1c) and aims to reconstruct regional vegetation and environ-mental history since ~47 kyr BP (1 kyr=1000 cal yr); to compare itwiththe oxygen isotope records from the North Atlantic and North Pacificregions; and to discuss the underlyingmechanisms of the environmentalchange in the region. Both time resolution and dating control aresubstantially improved in comparison to the earlier studies (e.g. Vipperand Smirnov, 1979; Bezrukova et al., 2008; Shichi et al., 2009).
2. Site setting and environments
Lake Kotokel (458 m a.s.l.) has an area of ~67 km2, a maximumlength of ~15 km and a maximum width of ~6 km (Galaziy, 1993). Inthe west a low-elevated mountain ridge (729 m a.s.l.) separates thelake from Lake Baikal (Fig. 1), and the Ulan-Burgasy Ridge (up to2033 m a.s.l.) bounds it in the east. The average water depth is 5–6 m,and the maximum depth is about 15 m (Shichi et al., 2009; Tarasov etal., 2009 and references therein).
The regional climate is continental with long winters and a well-pronounced summer precipitation maximum (Alpat'ev et al., 1976).The mean January temperature is−20 °C near the lake, but decreasesto below −26 °C with increasing elevation. The mean July temper-ature is 14–16 °C. The annual precipitation reaches 300–400 mm inthe coastal area and above 500 mm in the mountains (Galaziy, 1993).In July and August westerly winds dominating through the yearbecome weak, and southeastern cyclones bringing warm and wetPacific air to the region cause heavy rainfalls at the eastern branch ofthe Polar front (Bezrukova et al., 2008). The Atlantic air massesentering the region with the westerly flowmay bring precipitation allyear round, butmainly during autumn and spring. Dry, cold and sunnywinter weather is associated with the high pressure cell centred overeastern Siberia (Alpat'ev et al., 1976).
Modern vegetation east of Lake Baikal is a mosaic of borealconiferous and deciduous forests (Galaziy, 1993) and swampycommunities, widely distributed in the Selenga River delta and

187E.V. Bezrukova et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 296 (2010) 185–198
south of Lake Kotokel (Takahara et al., 2000; Shichi et al., 2009).Forests are mainly composed of Pinus sylvestris (Scots pine), Larixsibirica (Siberian larch) and Betula (birch) trees, with some admixtureof Populus tremula (aspen) and Alnus fruticosa (shrubby alder). Borealevergreen conifers, including Pinus sibirica (Siberian pine), Abiessibirica (Siberian fir) and Picea obovata (Siberian spruce) grow on theUlan-Burgasy slopes. Above 1800 m this mountain taiga vegetation isreplaced by open birch and larch forests and shrubby sub-alpineassociations represented by Pinus pumila (shrubby pine), Alnusfruticosa and Betula middendorfii (shrubby birch). Alpine tundraoccupies large areas north and northeast of Lake Baikal, while thesteppe is characteristic of Olkhon Island (Fig. 1) and semiariddepressions along the Selenga River, where precipitation drops to~160 mm/yr, (Galaziy, 1993).
3. Data and methods
3.1. Core lithology and age determination
The KTK2 sediment core (52°47′N, 108°07′E; Fig. 1c) was retrievedfrom a depth of ~3.5 m in the southern part of Lake Kotokel in August2005. The Livingston-type piston corer of 7.5 cm diameter was
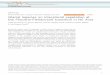
Fig. 2. Lithology column (a), radiocarbon dates (b) and age-de
applied to the upper and softer biogenic sediment (Fig. 2a) and the4.6 cm diameter corer was used to penetrate lower more compactlayers (Shichi et al., 2009). The upper part of the core (Fig. 2a) consistsof the soft brownish black gyttja (0–660 cm) underlined by the greyor blackish slightly laminated silty clay (660–740 cm), laminated greysilty clay (740–1010 cm) and dark-grey or grey silty clay unit (1010–1253 cm).
Eleven bulk sediment samples from the KTK2 core were submittedto Poznan Radiocarbon Laboratory. The distribution of obtainedradiocarbon ages versus composite core length (Fig. 2b, Table 1)demonstrates no reversals and a clear trend of increasing age withdepth. The sediment is non-calcareous, thus ensuring that a reservoireffect, which usually complicates accurate dating of the Baikal cores(Colman et al., 1996; Demske et al., 2005), is not an issue in LakeKotokel. The KTK2 core lithology and radiocarbon dates suggest anuninterrupted accumulation of sediment between ~47 kyr BP and thepresent.
The KTK2 14C AMS dates were converted to calendar dates(Table 1), which were then used to construct an age-depth modelbased upon linear interpolation between the neighbouring dates(Fig. 2c). In addition, the KTK2 age model was compared with theannually laminated sedimentary and high-resolution pollen records
pth relationship (c) of the KTK2 core from Lake Kotokel.

Table 1AMS radiocarbon dates from the KTK2 core. Radiocarbon years before present wereconverted to calendar years after Danzeglocke et al. (2008).
Sample depth(cm)
Datedmaterial
14C age(yr BP; 68% range)
Cal. age(yr BP; 68% range)
Laboratorynumber
137–141 Brownishblackgyttja
3855±35 4290±80 Poz-27580
208–212 Brownishblackgyttja
4575±35 5230±110 Poz-27581
289–293 Brownishblackgyttja
5690±35 6480±40 Poz-27582
309–313 Brownishblackgyttja
5890±40 6720±40 Poz-27583
501–505 Brownishblackgyttja
9990±50 11,470±130 Poz-27584
507–511 Brownishblackgyttja
10,190±50 11,890±110 Poz-27593
685–689 Blackishsilty clay
12,680±60 15,190±110 Poz-27585
795–799 Grey siltyclay
18,000±90 21,520±90 Poz-27586
918–922 Grey siltyclay
21,450±110 25,480±230 Poz-27587
1036–1040 Grey siltyclay
27,820±200 32,330±240 Poz-27589
1238–1244 Grey siltyclay
42,600±1100 46,285±1500 Poz-27637
188 E.V. Bezrukova et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 296 (2010) 185–198
from Lake Sihailongwan (SHL: Fig. 1a) and dated by both varve-counting and 40 calibrated AMS dates (Stebich et al., 2009). Theearlier studies suggested that the late-glacial–early Holocene shifts invegetation and climate in the Lake Baikal region were broadlysynchronous to the major oscillations in the oxygen isotope recordsfrom China (Tarasov et al., 2007a, 2009), reflecting changes in theNorthern Hemisphere (NH) temperature and East Asian monsoonintensity (Yuan et al., 2004). For the first time a direct comparison ofthe high-resolution pollen records from both regions became possible.
The characteristic changes in the SHL pollen/climate stratigraphy(Stebich et al., 2009) were compared to the similar events in the KTK2pollen records, and the corresponding ages of these events in the SHLrecordwere then transferred to theKTK2 record (Table 2). Both datasets
Table 2Complementary dates obtained via the pollen-based correlation between the pollen recordsstudy to refine age-depth model for the KTK2 record (Fig. 2).
Depth in the KTK2 core(cm)
Inferred age(cal yr BP)
Dated event in the SHL pollen record(after Stebich et al., 2009)
422.0 ~10,600 Quercus, Acer and Syringa become more commonViburnum reach their empirical limit, high arboreindicate the establishment of dense forest cover,observed in the area today
506.0 ~11,650 Re-increase in thermophyllous tree pollen, and thdecline that indicate a climatic amelioration
541.5 ~12,680 Significant declines in Fraxinus and Ulmus combinsteppe plants and maxima in Betula and Picea higreversal III
561.5 ~13,000 Pronounced minimum in the relative frequenciesFraxinus and increase in the abundance of herb pglacial climate reversal II
591.5 ~13,650 Dynamic increase of Ulmus and Fraxinus, establisbroadleaf forest
609.5 ~13,850 Ulmus minimum, the consequence of a short-term
651.5 ~14,400 Expansion of Ulmus and Fraxinus after the onsetclimatic amelioration at ~14,450 cal yr BP
show a good correspondence (Fig. 2c), suggesting the reliability of theKTK2 age model during the climatically unstable late-glacial interval. Inparticular, the onset of the Holocene interglacial conditions is wellpronounced in both records and canbe independentlydated in theKTK2core to ~11.65 kyr BP (pollen-based correlation with the LSH) and to~11.68 kyr BP (AMS dates from KTK2).
3.2. Pollen extraction and microscopic analysis
The KTK2 core material was stored in plastic liners at the Instituteof Geochemistry (Irkutsk) and sampled there for pollen analysis. Atotal of 212 samples comprising 1 cm3 of thewet sedimentwere takenwith a 6-cm step, yielding an average temporal resolution of220 years. A standard procedure was used to extract pollen, includingHCl and KOH treatment, heavy-liquid separation and acetolysis(Berglund and Ralska-Jasiewiczowa, 1986). Pollen and spores,mounted in glycerin, were counted under the light microscope with×400–1000 magnification. Identification of the pollen and spores wasperformed using regional pollen atlases and the reference pollencollection (Tarasov et al., 2007a and references therein).
All sampleswere rich inpollen, permitting countingup to1800pollengrains per slide. The KTK2 pollen diagram (Figs. 3a and 4) wasconstructed using the Tilia/Tilia-Graph/TGView software (Grimm,2004). Relative percentages for all terrestrial pollen taxa at each levelwere calculated from a terrestrial pollen sum taken as 100%. Percentagesfor aquatic and cryptogam taxa were calculated in relation to the totalsumof counted pollen and spores. The diagramwas subdivided into localpollen zones (PZ), using visual inspection supported by the results ofbiome score calculation described in the section below. This methodtakes into account the ecology of pollen-producing plants (Prentice et al.,1996) and does not entirely rely upon a statistical similarity betweenpollen assemblages composed of numerous taxa representing ecologi-cally different plant functional types. Pollen zones were also definedusing CONISS, which performs a stratigraphically constrained clusteranalysis useful for zonation (for technical details seeGrimm, 1987). In theCONISS analysis only terrestrial pollen taxa with values of 2% or greaterand appearing in more than one sample were considered (Gervais et al.,2002). Results obtained with different methods are shown in Fig. 3.
3.3. Quantitative vegetation reconstruction
The biome reconstruction method (Prentice et al., 1996) permitsthe objective assignment of pollen taxa to major vegetation types/
from Sihailongwan Lake (Stebich et al., 2009) and Kotokel Lake, which are used in this
Comparable event in the KTK2 pollen record(this study)
, Juglans, Tilia andal pollen valuessimilar to that
High arboreal pollen values and minimal values of herbaceouspollen taxa indicate the establishment of dense forest cover,similar to that observed in the area today
e onset of a Larix Reappearance of Abies, decrease in Artemisia reflecting the onsetof the Holocene interglacial conditions
ed with a peak inhlighting a climate
Abrupt decrease in Picea, disappearance of Abies, increase inArtemisia reflecting a considerable reduction in woody coverageand climatic deterioration
of Ulmus andollen marks late-
Short-term decrease in Picea and Abies, increase in theabundance of herb pollen reflecting prominent climaticdeterioration
hment of mixed Increase in Picea, decrease in Artemisia and in total herbs
climatic reversal I Peak in Alnus fruticosa, decrease in total trees reflecting short-term climatic deterioration
of the late-glacial Abrupt increase in Picea and abrupt decrease in Artemisia,appearance of Abies reflecting the late-glacial climaticamelioration

Fig. 3. Zonation of the KTK2 pollen record (g) applied in this study using visual inspection of the pollen percentage diagram (a) and the results of biome score calculation (b) plottedagainst the dendrograms obtained using a stratigraphically constrained cluster analysis CONISS (c–f).
189E.V. Bezrukova et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 296 (2010) 185–198
biomes on the basis of the bioclimatic tolerance, ecology andbiogeography of pollen-producing plants. The method was success-fully tested using extensive regional surface pollen datasets andapplied to the last glacial–interglacial fossil pollen records fromnorthern Eurasia (Edwards et al., 2000; Tarasov et al., 2000; Mokhovaet al., 2009), from Lake Baikal (Tarasov et al., 2005, 2007a), and to thepoorly-dated KTK1 record from Lake Kotokel spanning the past 15 kyr(Tarasov et al., 2009). In the present study all identified terrestrialpollen taxa from the KTK2 record were assigned to regional biomesusing the biome-taxon matrix presented in Table 3.
A complementary approach (Williams et al., 2004), which combinessatellite-based vegetation cover dataset (DeFries et al., 1999) andmodern surface pollen dataset from northern Eurasia (Tarasov et al.,2007b) with the best modern analogue (BMA) approach (Guiot et al.,1989), was applied to the KTK2 pollen record in order to reconstructchanges in the woody cover. The test of the method demonstrated thatpollen-basedmodernwoody cover reconstructions andoriginal satellitemeasurements match well in northern Asia, providing satisfactoryestimates of percent variance explained and root mean square error(RMSE) for both the total woody cover (r2=0.77, RMSE=11.69) andthe broadleaved (r2=0.66, RMSE=3.31), needle-leaved (r2=0.79,RMSE=10.23) woody cover fractions at modern pollen sites (seeTarasov et al., 2007b for the method evaluation and design). Therobustness of the reconstruction for the past 15 kyr was ensured by
comparisonwith the results obtained earlier for the KTK1 pollen record(Tarasov et al., 2009).
3.4. Diatom analysis and pH reconstruction
A total of 125 samples, sampled at 10-cm intervals, were preparedfor diatom analysis using the standard hydrogen peroxide technique(Battarbee et al., 2001). Aliquots of evaporated suspensions wereembedded in Naphrax. Counting of at least 300–600 diatom valves persample along ten horizontal transects using the light microscope withphase-contrast oil immersion objectives at ×400–1000 magnificationwas possible for most of the samples. However, diatom valves wererare or not found at all in 28 samples from the glacial part of therecord. Diatom concentrations (×106 valves per gram of air-driedsediment) and percentage diatom abundances (based on the totalsum of all diatoms counted in each sample) were calculated. Diatomnomenclature followed Krammer and Lange-Bertalot (1986–1991)and AL:PE guidelines (Cameron et al., 1999). The AL:PE diatom–pHcalibration dataset consists of surface-sediment diatom assemblagesfrom 118 lakes and contains 530 taxa. The AL:PE training set is fromhigh-altitude or high-latitude lakes in the Alps, Norway, Svalbard,Kola Peninsula, UK, Slovenia, Slovakia, Poland, Portugal and Spain.
The modern pH of Lake Kotokel water (about 7.0) fits within theAL:PE pH range (4.5–8.0) and the fossil diatom assemblages of Lake

Fig. 4. Simplified pollen percentage diagram of the KTK2 core (analyst E. Bezrukova). See Fig. 2a for the description of lithological units.
190 E.V. Bezrukova et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 296 (2010) 185–198
Kotokel are generally well represented in the AL:PE dataset.Therefore, the AL:PE diatom–pH model was used for pH inferences(see Cameron et al., 1999 for the details of the AL:PE pH diatommodelperformance). The reconstruction was performed using the computerprogramme C2 ver. 1.5 (Juggins, 2007) using jack-knifed cross-validation. The second component in the Weighted Averaging PartialLeast Squares (WAPLS) regression showed the best predictiveperformance (Table 4) and was used for pH reconstruction. Themodernmeasured lake pH varies between 6.8 and 7.3 (Kuzmin, 1988)and the pH predicted by the AL:PE model is 7.8, implying that the
Table 3Terrestrial pollen taxa identified in the KTK2 record (this study) and used in the biomeand woody cover reconstructions (after Tarasov et al., 2000, 2009).
Biome Taxa included
Tundra Alnus fruticosa, Betula sect. Nanae/Fruticosae, Cyperaceae, Ericales,Poaceae, Polemonium, Polygonum, Rumex, Salix, Saxifragaceae,Valeriana
Cold deciduousforest
Alnus, Betula sect. Albae, Ericales, Larix, Pinus sylvestris (subgen.Diploxylon-type), P. pumila (subgen. Haploxylon-type), Salix
Taiga Abies sibirica, Alnus, Betula sect. Albae, Ericales, Larix, Piceaobovata, Pinus sylvestris, P. sibirica (subgen. Haploxylon-type),Ribes, Salix
Cool coniferforest
Abies sibirica, Alnus, Betula sect. Albae, Ericales, Corylus, Larix, Piceaobovata, Pinus sylvestris, P. sibirica (subgen. Haploxylon-type),Ribes, Salix, Ulmus
Steppe Apiaceae, Artemisia, Asteraceae (incl. subfam. Asteroideae andCichorioideae), Brassicaceae, Caryophyllaceae, Chenopodiaceae,Fabaceae, Lamiaceae, Liliaceae, Plantago, Poaceae, Polygonum,Ranunculaceae, Rosaceae, Rubiaceae, Rumex, Scrophulariaceae,Thalictrum, Urtica, Valeriana
Desert Artemisia, Boraginaceae, Chenopodiaceae, Ephedra, Polygonum
model gives slightly overestimated values. The fossil diatom assem-blages between 543 and 501 cm are dominated by Ellerbeckia arenariaand E. arenaria var. teres, which are both absent from the moderndataset. Therefore, the pH inferences between 543 and 501 cm solelyrely on other fossil diatom taxa and samples between 543 and 501 cmprovide poor analogues for pH reconstruction.
Detrended canonical correspondence analysis (DCCA) was used toestimate the overall species turnover measured in standard deviation(SD) units and to generate sample scores for axis 1, which provide anestimate of compositional change along a temporal gradient (terBraak, 1986). The samples' age was used as a sole environmentalvariable in DCCA. In DCCA, species data were square-root trans-formed, no rare species down-weighting was applied, and nonlinearrescaling and detrending by segments were used. DCCA was carriedout using CANOCO 4.5 (ter Braak and Šmilauer, 2002). ZONE software(Juggins, 1991) was used to split the diatom diagram into strati-graphic zones. The statistical significance of the stratigraphic zones
Table 4Apparent and predictive (jack-knifed) statistics for the AL:PE pH–diatommodel appliedto the fossil diatom data from Lake Kotokel.
Performance statistics pH WAPLS, 2nd component
r2 0.928RMSE 0.203Average bias −2.299e−0.005
Maximum bias 0.384r2 (jack-knifed) 0.789RMSEP 0.349Jack-knifed average bias 0.011Jack-knifed maximum bias 0.734

191E.V. Bezrukova et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 296 (2010) 185–198
was then established using the programme BSTICK (Bennett, 1996).Changes in DCCA sample scores on axis 1 were also used as anadditional method of stratigraphic zone boundary identification. Inthe diatom percentage diagram (Fig. 6), the taxa were orderedaccording to their weighted average score, highlighting the diatomfloristic changes. Aulacoseira granulata agg. includes A. granulata var.angustissima, and Staurosira construens agg. includes ecologically closesubspecies, which have identical stratigraphic distribution in theKTK2 core (i.e. S. construens var. construens, S. construens var. venterand S. construens var. binodis). Staurosirella pinnata agg. includes S.pinnata var. pinnata and S. pinnata var. lancetulla for the same reasons.
4. Results and interpretations
4.1. Pollen record and vegetation reconstruction
The pollen diagram (Fig. 4) is subdivided into eight local pollenzones (PZ) on the basis of changing pollen composition and variationsin the dominant biome scores. A brief characteristic of the pollen/biome zones provided here aims to facilitate further discussion.
PZ KTK2-8 (1253–1000 cm; ~47–30 kyr BP) demonstrates highpercentages of herbaceousand shrubby taxa,mainlyArtemisia (30–50%)and Betula sect. Nanae/Fruticosae (up to 40%). Arboreal pollen
Fig. 5. Local pollen assemblages zones (a) and time series of individual vegetation types (biovegetation (c) and quantitative changes in woody cover percentages within a 21×21 km w
percentages reach up to 20–30% in the lower (Picea) and upper (Larix,and Pinus) parts of this zone, but remain at 10%-level in themiddle part.The biome reconstruction shows that steppe and tundra have thehighest scores (Fig. 5b), and scores of the taiga biome are ~5–10, in linewith themoderately lowwoody cover (~10%) around the lake (Fig. 5d).The vegetation was probably a mosaic of scattered trees and/or isolatedforest stands, shrubby and herbaceous tundra communities occupyingwetter habitats and dominant cold steppe communities.
PZ KTK2-7 (1000–850 cm; ~30–23 kyr BP) reveals the highestpercentages of Artemisia (40–50%) and Poaceae (25–40%) andextremely low percentages of boreal tree and shrub pollen. Thebiome reconstruction demonstrates the highest scores for steppe(N20) and lowest taiga scores (Fig. 5), suggesting that cold steppevegetation absolutely predominated in the region. However, thepresence of arboreal pollen, even in very low percentages, andquantitative woody cover reconstruction do not support the totaldisappearance of cold/drought-tolerant boreal trees/shrubs from thevegetation.
PZ KTK2-6 (850–655 cm; ~23–14.5 kyr BP) reveals the highestpercentages of Artemisia (up to 80%) in the lower part of this zone anda progressive increase in the tree/shrub pollen percentages (N40%)towards the uppermost part of this zone. The steppe remains thedominant biome, but the tundra communities once again start to play
mes) dominating the study area since ~47 kyr BP (b), with qualitative characteristics ofindow (d) derived from the KTK2 pollen record following Tarasov et al. (2009).

192 E.V. Bezrukova et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 296 (2010) 185–198
a more important role. Both biome and woody cover reconstructionresults point to a greater-than-present openness of the landscapearound Lake Kotokel.
PZ KTK2-5 (655–540 cm; ~14.5–12.65 kyr BP) demonstrates asharp increase in arboreal taxa percentages, mainly Picea (20–50%),birch and alder shrubs (up to 40%). For the first time, taiga becomesthe dominant biome, closely followed by tundra. Two peaks in woodycover percentages are reconstructed, ~14 kyr (up to 25%) and ~13 kyrBP (up to 35%), separated by an interval with lower woody coverage.The vegetation likely resembled the present-day boreal woodland/forest-tundra with a significant contribution of spruce, fir and larchtrees and shrubs of alder, birch and willow.
PZ KTK2-4 (540–506 cm; ~12.65–11.65 kyr BP) reveals a sharpdecrease in arboreal taxa percentages and an increase in herbaceoustaxa (up to 35%). Abies pollen is not recorded in this zone and pollen ofshrubby birch almost replaces shrubby alder (Fig. 4). The quantitativereconstruction shows that the tundra becomes the dominant biomeand suggests a decrease in the woody cover to ~15%.
PZ KTK2-3 (506–422 cm; ~11.65–10.6 kyr BP) reveals the domi-nance of arboreal taxa (mainly Betula — a well-known woodypioneer) and the return of Abies to the pollen assemblages. Thebiome reconstruction demonstrates equally high scores for the tundraand taiga biomes, suggesting a quick recovery of arboreal communi-ties and boreal woodland vegetation around the lake, in line with thereconstructed increase in the total woody cover up to 40%.
PZ KTK2-2 (422–310 cm; ~10.6–6.8 kyr BP) demonstrates aminimal contribution of herbaceous taxa to the pollen assemblages.The taiga scores are noticeably higher than the scores of non-forestbiomes, suggesting well-established boreal forest in the region. Thetotal woody cover increased to above 50% and exceeded present-dayvalues.
PZ KTK2-1 (310–0 cm; ~6.8–0 kyr BP) shows that Pinus isreplacing Betula as the dominant taxon. This likely reflects change inthe forest composition and the regional spread of Scots pine after7 kyr BP. In this zone taiga scores are noticeably higher than the scoresof non-forest biomes, but the woody cover decreases to its present-day level of ~45%.
4.2. Diatom record and lacustrine environments
The fossil diatom assemblages of the KTK2 core from Lake Kotokelcomprise 143 diatom taxa in total. The percentage diagram (Fig. 6)was divided into seven diatom zones (DZ). The reconstructed pHshows little change throughout the core, fluctuating around 8. No orvery few (1–30 valves) diatoms occurred at 1149.5, 1087.5, 1025–880,865.5, 823.5 and 810–718 cm (Fig. 6), eliminating these samples fromfurther analyses.
DZ KTK2-7 (1240–1069 cm; ~46–34.6 kyr BP) is dominated bysmall benthic Staurosirella pinnata agg., Staurosira construens agg.,Pseudostaurosira brevistriata and Ophephora martyi. The latter isparticularly abundant in the lower and upper parts of this zone. Afew planktonic species, Cyclotella bodanica and Cyclotella ocellata andmeroplanktonic, heavily silicified Aulacoseira ambigua, reach theirhighest abundance in the middle of the zone and disappear in theupper part of the zone. Total diatom concentration (TDC) reaches itshighest values (N100×106 valves g−1) in the lower-middle part ofthe zone and then quickly decreases. DCCA axis 1 sample scoresslightly increase throughout the zone (Fig. 6), tracking the overallmodest floristic changes in the diatom assemblages. The dominance ofpioneer benthic diatom taxa (e.g. Staurosirella pinnata agg. andOphephora martyi) is typical of early stages of lake development.These taxa are commonly found inmany post-glacial lakes and in late-glacial lacustrine sediments (Haworth, 1976; Smol, 1983; Solovievaand Jones, 2002; Finkelstein and Gajewski, 2008) and at present inmany arctic and subarctic lakes with a short open-water season(Smol, 1988; Lotter and Bigler, 2000; Smol and Douglas, 2007).
Regionally, small Fragilariaceae-dominated diatom assemblages oc-curred throughout the Holocene in a small lake from the AltaiMountains, central Siberia (Westover et al., 2006) and in the earlyHolocene in a large deep-water lake from the Mongolian Altai(Rudaya et al., 2009). Diatom assemblages suggest a turbulent andturbid environment implying strongwinds and a high level of erosion.These taxa are indicators of ‘disturbed’ environmental conditions, asthey survive today in harsh arctic environments with low lightpenetration where other more sensitive taxa would not succeed(Smol, 1988). The lake level was high enough to sustain well-established planktonic species at 10–20% abundance. The increase intwo small planktonic Cyclotella species in themiddle of the zone at theexpense of heavily silicified Aulacoseira ambigua and small benthicPseudostaurosira brevistriata and Opeophora martyii might imply aslight amelioration of climate. The 20th century climate warming wasfound to be the most plausible mechanisms underlying the replace-ments of, for example, the larger and heavier Aulacoseira taxa and/orsmall benthic Fragilaria taxa by smaller planktonic Cyclotella taxa inmany arctic, subarctic and temperate lakes (Rühland et al., 2008,Winder et al., 2009).
DZ KTK2-6 (1069–1025 cm; ~34.6–31.5 kyr BP). In this zoneOphephora martyi, Staurosirella pinnata agg. and Staurosira construenscontinue to decline. Cyclotella ocellata, C. bodanica and Aulacoseiraambigua, whichmade a brief appearance in the middle of the previouszone, are replaced by planktonic Aulacoseira subarctica (max 20%) andA. islandica (max 36%). Both A. subarctica and A. islandica are heavierand more silicified taxa compared to small and light Cyclotella andthey require high water turbulence to maintain their position in thewater column and usually prevail during spring/autumn waterturnover periods (e.g. Rühland et al., 2008; Solovieva et al., 2008).This implies that the lake environment might have changed either byshortening the summer period of stratified water, when Cyclotella hasthe competitive advantage, or by increasing the spring/autumn periodof water turbulence. Both changes suggest climate cooling and theextension of the ice-cover season. TDC remains low, fluctuatingbetween 0.16 and 22.73×106 valves g−1. DCCA sample scoresincrease, reaching the highest values (2.3) for the whole core.Diatoms do not occur between 31.5 and 24.2 kyr BP suggestingparticularly harsh lake conditions.
DZ KTK2-5 (880–810 cm; ~24.2–21.9 kyr BP) shows the disap-pearance of meroplanktonic and heavily silicified diatoms (Aulaco-seira subarctica and A. islandica), a considerable decrease in theabundance of Ophephora martyi and the almost complete dominanceof the Staurosirella pinnata agg. complex, which may survive harshenvironmental conditions where other diatom taxa would notsucceed (e.g. Smol, 1988). TDC remains low. DCCA samples scoresdecrease in DZ3 to 0.71 on average. These dramatic changes in thediatom assemblages might have resulted from some catchmentdisturbance or/and a particularly cold/dry climate between 31.5 and24.2 kyr BP, when almost no diatoms were found. As in earlierintervals diatoms were also extremely rare or totally absent between810 and 718 cm (~21.9–16.9 kyr BP), indicating conditions unfavour-able for diatom development.
DZ KTK2-4 (718–550 cm, ~16.9–12.7 kyr BP) follows anotherdisturbance episode coinciding with the LGM, during which almost nodiatoms occurred in the lake. This zone reveals the first occurrence ofplanktonic Aulacoseira granulata, a heavy silicified diatom, which istypical for plankton of large eutrophic and mesotrophic lakes andreservoirs (e.g. Fluin et al., 2010; Ohtaka et al., 2010). Cyclotellaocellata (max 35%) and Aulacoseira ambigua (max 34%) are two otherco-dominant planktonic species in this zone, which made a briefoccurrence in the middle of DZ KTK2-7. The re-occurrence of smallCyclotella taxa might again indicate climate amelioration and a longeropen-water season (e.g. Rühland et al., 2008). A progressive increasein the abundance of planktonic Cyclotella diatoms is accompanied by apronounced decrease in benthic species (Fig. 6). Amphora perpusilla,

Fig. 6. Simplified diatom percentage diagram (analyst N. Ignatova†) with diatom concentration, ALPE-reconstructed pH and DCCA axis 1 sample scores of the KTK2 core from LakeKotokel. See Fig. 2a for a description of lithological units. All diatom taxa with an abundance N5% are shown. The taxa are sorted by their weighted averaging scores from upper left tobottom right to highlight the major changes in the diatom stratigraphy.
193E.V. Bezrukova et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 296 (2010) 185–198
the periphytic diatom, occurs exclusively in this zone, first reaching11% and then decreasing to 3%. Similar to DZ KTK2-7, C. ocellataabruptly disappears in the upper part of the zone replaced by A.ambigua, implying possible deterioration of climate and shortening ofthe open-water season. TDC peaks in the middle of the zone, reaching156.3×106 valves g−1. It decreases to 19.1×106 valves g−1 towardsthe end of the zone implying an abrupt decline in diatom productivity.This decrease is concomitant with the decrease in Cyclotella ocellataand also might suggest deteriorating environmental conditions, inparticular, colder climate. DCCA sample scores range between 0.5 and0.7 throughout the zone. The occurrence and subsequent dominanceof planktonic eutrophic/mesotrophic Aulacoseira granulata, togetherwith relatively high abundances of Cyclotella ocellata in the middle ofthe zone and two peaks of A. ambigua, suggest that the nutrient statusof the lake changed. It is likely that the changes in the catchment soilsand vegetation, and the possible arrival of trees facilitated theincreased nutrient supply to the lake, which sustained high nutrientdemands of A. granulata. A peak in Cyclotella ocellata abundancemightindicate a relatively short-lived climate amelioration episode.
DZ KTK2-3 (550–495 cm; ~12.7–11.6 kyr BP) is absolutely dom-inated by planktonic Aulacoseira granulata and large radial centric andbenthic Ellerbeckia arenaria var. arenaria and E. arenaria var. teres.Aulacoseira ambigua, which prevails in the previous zone, disappearsand Cyclotella ocellata occurs at two levels at 3–4%. The prevalence oftwo Ellerbeckia species also implies the influence of glaciogenicsediments and relatively turbid water (Hickman and Reasoner, 1994).Ellerbeckia is a relatively common diatom in southeastern Siberia. Itoccurs, for instance, in Lake Arachlei located to the east from Kotokel(V. Panizzo, pers. comm.). It was also found in Hoton-Nur lake
sediment in the Mongolian Altai (Rudaya et al., 2009). A few benthicFragilariaceae (Staurosira construens agg., Pseudostaurosira brevis-triata, and Staurosirella pinnata agg.) are present at low abundances.TDC values are very low, and DCCA scores decrease slightly to 0.2 onaverage compared to the previous zone. This short zone reflects thetransition from the late-glacial to the Holocene. It was likely thatfollowing deglaciation catchment erosion increased, which led todecreased water transparency and light penetration and, therefore, asharp decrease in benthic species and the dominance of the aboveAulacoseira taxa.
DZ KTK2-2 (495–130 cm; ~11.6–4 kyr BP) shows continuingdominance of Aulacoseira granulata (45–98%). Staurosira construensis another dominant taxon at some levels with abundances between 2and 40%. Both Ellerbeckia arenaria var. arenaria and Ellerbeckiaarenaria var. teres abruptly decrease down to very low numbers,similarly to Pseudostaurosira brevistriata, Ophephora martyi andStaurosirella pinnata agg. TDC fluctuates frequently between13.3×106 and 136.5×106 valves g−1. DCCA sample scores remainalmost unchanged at about 0.45 on average. The diatom assemblagesof this zone suggest mesotrophic/eutrophic environments, indicatedby dominance of Aulacoseira granulata, a relatively heavy diatomwiththick siliceous frustules, which is common in the large lakes, waterreservoirs and rivers with a relatively high nutrient content (e.g.Kilham and Kilham, 1975; Margalef, 1983; Kuzmin, 1988; Gómez etal., 1995; Fluin et al., 2010; Ohtaka et al., 2010). Similarly to otherAulacoseira taxa from Lake Kotokel, it usually peaks during spring/autumn water turnover periods, requiring high water turbulence tomaintain its position in the water column (Gómez et al., 1995). Thereasons behind the abrupt disappearance of Ellerbeckia are unclear.

194 E.V. Bezrukova et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 296 (2010) 185–198
Most likely it implies that the catchment erosion decreased, andnutrient supply to the lake increased and these favored developmentof competing diatom taxa, namely Aulacoseira granulata.
DZ KTK2-1 (130–0 cm; ~4–0 kyr BP) differs from the previouszone by a slight decrease in Aulacoseira granulata, Staurosiraconstruens agg. and Pseudostaurosira brevistriata, the reappearancein notable abundances of Ellerbeckia arenaria var. teres (up to 11%),and the total disappearance of Staurosirella pinnata agg. TDC remainsat a similar level to DZ6, and DCCA sample scores slightly increasecompared to the previous zone. The further reduction in benthic andperiphytic diatom taxa and the thriving planktonic community implya possible increase in water depth or decrease in water transparency/light penetration, which affected the benthic taxa by reducing theirhabitat availability. Higher values of Staurosira construens agg.compared with other Fragilariaceae might imply warmer climateconditions (e.g. Finkelstein and Gajewski, 2008).
5. Discussion and conclusions
The KTK2 pollen and diatom records span a ~47-kyr interval,which includes the middle glacial interstadial conventionally attrib-uted to the marine isotope stage 3 (MIS3), the last glacial maximum(LGM) followed by the late-glacial climate amelioration (MIS2) andthe Holocene interglacial (MIS1). In this section changes in the pollenand diatom assemblages are discussed in terms of the late Quaternaryclimate dynamics in the Lake Baikal region of southern Siberia.
5.1. The MIS3 interstadial (~47–30 kyr BP)
The MIS3 interstadial was identified in the palaeorecords from NHand conventionally dated to ~35–25 14C kyr BP (~40–30 kyr BP)(Frenzel et al., 1992). The KTK2 pollen record and quantitativevegetation reconstruction suggest the predominance of tundra-steppe communities with Artemisia, Poaceae and Cyperaceae and amoderately low, but variable woody cover (5 to 20%) during ~47–30 kyr BP, reflecting generally harsh and unstable climatic conditions.Boreal tree and shrub taxa could grow continuously in the locallymoist habitats, expanding rapidly during the intervals of interstadial-like climate amelioration (e.g. ~45–42 and ~35–31 kyr BP), andshrinking in their distribution areas with the onset of the cold/drystadial-like climate (e.g. prior to ~45 and ~42–35 kyr BP). A relativelyshort peak in Cyclotella ocellata, C. bodanica, and Aulacoseira ambiguaand the corresponding peak in TDC between 45 and 42 kyr BPcorroborates a response of the lake system to climate amelioration.Interestingly, another warm event at ~35–31 kyr BP is not as clearlyreflected by the lake diatom assemblages. A distinct Picea pollenmaximum in the KTK2 record at ~45–43 kyr BP allows a more preciseallocation of the similar event found in the low-resolution and poorly-dated pollen record from Lake Baikal (Shichi et al., 2007). It is broadlysynchronous with the larch woodland development in the IndigirkaLowland, northeastern Siberia, ~45 kyr BP (Anderson and Lozhkin,2001), the formation of the black soil in western Siberia (Frechen etal., 2005), and the increased diversity and abundance of testateamoebae in the MIS3 interstadial sediments on the Laptev Sea coast(Müller et al., 2009).
The KTK2 diatom record suggests relatively deep oligo-mesotrophicenvironments in Lake Kotokel prior to ~31.5 kyr BP, in line with thepollen-based interpretation. A pronounced decrease in the diatomconcentration recorded after ~42 kyr and after ~32 kyr BP likelyindicates two major steps in climatic deterioration during the MIS3.
5.2. The full glacial (~30–17 kyr BP)
The full glacial occurs in the KTK2 pollen record at ~30–17 kyr BPand is marked by an increase in herbaceous taxa percentages, thehighest scores for steppe and the lowest scores for taiga biome,
suggesting an increased aridity/continentality of the regional climateand the predominance of open herbaceous communities in thelandscape. The noticeable decrease in diatom concentrations followedby the virtual disappearance of diatoms from the KTK2 sediment(laminated grey silty clay) at ~31.5–17 kyr BP might be anotherindicator of the dry and cold climate with strongly pronouncedseasonality. A similar dramatic reduction in diatom abundanceoccurred in Lake Baikal during periods of climate deterioration duringthe MIS3 interstadial (Swann et al., 2005; Mackay, 2007). The peak incryptogam spore percentages (mainly Equisetum) recorded at ~31–30 kyr BP suggests that the coastal horsetail-covered zone becamecloser to the KTK2 coring site than before. The highest percentages ofRanunculaceae pollen at ~30–23 kyr BP further support a shift ofmeadow/wetland communities closer to the coring point, suggestingthat Lake Kotokel was much smaller in size than today during thisprobably the driest interval of the whole record.
The reappearance of diatoms in small quantities might be a sign ofa slight amelioration of the regional climate at ~24–22 kyr BP, whichmight have influenced the aquatic ecosystem of a small lake, but wasnot strong enough to cause visible changes in the KTK2 core lithologyand pollen assemblages/terrestrial vegetation. Both herb/grass-dom-inated pollen assemblages, the absence of aquatic macrophytes andthe sporadic presence of diatoms in the KTK2 records at ~22–17 kyrBP point to a second phase of climate deterioration, conventionallyassociated with the LGM.
5.3. The late-glacial (~17–11.65 kyr BP)
A gradual increase in tree/shrub pollen percentages after ~18 kyrBP and changes in the diatom and lithology records after ~17 kyr BPindicate the late-glacial climate amelioration in the region. Particu-larly noticeable changes suggested by the KTK2 proxies and proxy-based environmental reconstructions occurred at ~14.7–12.7 kyr BP.At that time the regional climate became warmer and wetter thanduring the MIS3 interval, although it did not yet reach the Holocenelevels. These changes resulted in the spread of forest vegetation(increase in the woody cover percentages up to 20–30%), and apossible increase in lake level/water depth due to the melting of themountain glaciers, thawing of the permafrost active layer and increasein atmospheric precipitation (Bezrukova et al., 2005). The increase insmall planktonic Cyclotella ocellata also suggests climate amelioration(Winder et al., 2009). Recorded increases in arboreal pollenpercentages and associated peaks in taiga scores and woody coverpercentages indicate that trees could have quickly spread out of theirglacial refugia at least twice during the late-glacial interval,corroborating with the results obtained from the poorly-dated KTK1core (Tarasov et al., 2009). This study allows a more accurate datingof the woodland intervals to ~14.5–14 kyr and 13.3–12.8 kyr BP.Chronological comparison of vegetation/environmental changesaround Lake Kotokel with the European records (Sirocko, 2009 andreferences therein) puts the later phasewithin the Allerød interstadial(13.58/13.35–12.7/12.68 kyr BP), in line with other palaeobotanicalrecords from northern Asia (e.g. Andreev and Tarasov, 2007), the LakeBaikal region (e.g. Demske et al., 2005; Bezrukova et al., 2008; Shichiet al., 2009) and northeastern China (Stebich et al., 2009). The earlierphase of forest development seems to be an analogue of theMeiendorf interstadial (14.7–14.05/13.8 kyr BP, Sirocko, 2009),which until now was not distinguished in the lower-resolution pollenrecords from the Lake Baikal region and is likely interpreted as a partof the undifferentiated Bølling/Allerød interstadial complex (e.g.Bezrukova et al., 2005; Demske et al., 2005; Shichi et al., 2009).There is no clear evidence for the short-lived Bølling event (13.67–13.58/13.54 kyr BP, Sirocko, 2009) in the KTK2 records, probablybecause resolution is not high enough.
The pollen and diatom records from Lake Kotokel (e.g. decrease inwoody cover, increase in tundra biome scores and marked changes in

195E.V. Bezrukova et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 296 (2010) 185–198
the diatom assemblages) clearly identify the YD stadial in the LakeBaikal region. The KTK2 age model allocates it within ~12.7–11.7 kyrBP. The YD climate reconstructions suggest lower than presenttemperatures (by 2–4 °С in July and by 8 °С in January) and lowerprecipitation (by 100 mm/yr) around Lake Baikal (Tarasov et al.,2007a, 2009). Very low diatom concentrations and disappearance ofsmall ‘warm’-water planktonic Cyclotella ocellata from the KTK2sediment might imply climate deterioration and shortening of theopen-water season. Predominance of Ellerbeckia arenaria var. arenariaimplies increased amounts of glaciogenic sediments, higher erosionand a turbid water regime during the YD (Hickman and Reasoner,1994). Frustules of finer and thinner diatoms, for instance, smallFragilariaceae, might not have been well-preserved in the highlyminerogenic sediments, whereas robust and thick Aulacoseiragranulata and Ellerbeckia arenaria might have a better chance to bepreserved in the lake sediments and therefore might be over-represented in the fossil record. Overall, it is obvious from the KTK2records that the YD event clearly affected the regional environments.However, the YD climate remained warmer and wetter than at anytime between ~47 and 17 kyr BP.
5.4. The Holocene (11.65 kyr BP — present)
The Holocene vegetation and climate dynamics in the region understudywas recently reconstructed from the KTK1 pollen record (Tarasovet al., 2009). The chronological sequence of environmental changereconstructed from the KTK2 records shows great similarity with theprevious study. The shift to Holocene environments appears in theKTK2 diatom and pollen records at ~11.7–11.65 kyr BP, suggesting analmost synchronous response of terrestrial and aquatic ecosystems tothe global climate change. The KTK2 date agrees with the formaldefinition and dating of the Global Stratotype Section for the base of theHolocene (~11,650 cal yr BP), derived from the Greenland NGRIP icecore and selected regional records (Walker et al., 2009), suggesting asynchronous onset of the Holocene interglacial across Eurasia.
Results of pollen analysis and pollen-based biome andwoody coverreconstructions from the KTK2 core correlate well with the KTK1results (Tarasov et al., 2009), demonstrating that a boreal woodlandspread around Lake Kotokel at ~11.65–10.5 kyr BP in response to anoticeable increase in precipitation and in winter and summertemperatures. The maximal spread of the boreal forest (taiga)communities is associated with higher-than-present July (17–18 °C)and January (−19 °C) temperatures and precipitation (500–550 mm/yr), reconstructed at ~10.5–7 kyr BP (Tarasov et al., 2009). The pollen-based reconstruction of a slight decrease in woody cover around LakeKotokel ~at 7–6.5 kyr BP likely indicates a gradual decrease inprecipitation and temperatures towards their present-day values. Anoticeable increase in Pinus sylvestris pollen recorded after ~7 kyr BPreflects the spread of Scots pine in the Lake Baikal region (Demske etal., 2005; Bezrukova et al., 2008; Shichi et al., 2009), in line with theonset of drier and colder (similar to present) climate. The changes indiatom assemblages support these pollen-based reconstructions. Theabsolute dominance of meroplanktonic Aulacoseira granulata (maxi-mum ~11.6–10 kyr BP), disappearance of Ellerbeckia arenaria var.arenaria by ~11.6 kyr BP and reappearance of benthic taxa suggestestablishment of deep- and transparent-water environments alreadyin the early Holocene. Although this system remained rather stablethroughout the Holocene, minor changes in the diatom assemblageslikely indicate gradual natural enrichment of the lake after ~7 kyr BP.
5.5. Factors controlling the late Quaternary environmental dynamics inthe Lake Baikal region
The Lake Kotokel records contribute to the continuous debateconcerning the degree of climate amelioration and the chronologicalframework of the MIS3 interstadial in Siberia (e.g. Anderson and
Lozhkin, 2001; Kienast et al., 2005; Müller et al., 2009). Frenzel et al.(1992) reconstructed a colder (by 6–8 °C in winter and by 4–5 °C insummer) and drier (by 100–200 mm/yr) than present interstadialclimate in Siberia at ~40–30 kyr BP. Other authors (e.g. Shahgedanova,2002 and references therein) suggested that it could have beenregionally (e.g. in western Siberia) warmer than the present. Thepalaeobotanical (Hubbertenet al., 2004;Kienast et al., 2005) and isotopestudies (Tütken, 2003; Popp et al., 2006) in the arctic parts of easternSiberia led to the reconstruction of an extreme-continental climate,characterised by low precipitation, mean annual and winter tempera-tures, but relatively high summer temperatures and/or a longer snow-free period compared to today. The discontinuous pollen data fromnortheastern Siberia were used to infer a regional climate similar to thepresent at ~43.2–37.5 kyr BP, but colder/drier-than-present environ-ments prior to and after this interval (Anderson and Lozhkin, 2001).
The KTK2 records presented here point to a drier/colder-than-present late Pleistocene climate in the southern part of eastern Siberiaeven during the interstadial episodes of climatic amelioration that arevisible, for example, in the taiga (maxima) and steppe (minima)biome scores (Fig. 7d). The reconstructed pattern of changes in theregional environments demonstrates that the late Pleistocene climatedynamics in southern Siberia were more complex than previouslythought, and resemble the temperature variations (e.g. Greenlandinterstadials and Heinrich events) expressed in the δ18O record fromGreenland ice (Fig. 7a) and the East Asian Monsoon intensity signal inthe δ18O record from Chinese stalagmites (Fig. 7e). Minor incon-sistencies could be explained by the lower accuracy of the KTK2 agemodel and the lower resolution of the KTK2 record in comparison tothe above δ18O records. Wang et al. (2001) demonstrated that theisotope records from Hulu Cave near Nanjing and from Greenland icecores bear a remarkable resemblance, suggesting that the intensity ofthe summer Pacific monsoon, which brings moisture to the easternpart of Asia, changed in accordance with North Atlantic temperaturesbetween 75 and 11 kyr BP. Evidence for climatic teleconnectionsbetween Europe and Lake Baikal duringMIS3was discussed by Swannet al. (2005) and Mackay (2007). The Lake Kotokel records presentedhere link changes in the North Atlantic and North Pacific climate,which are responsible for the sub-latitudinal transport of heat andmoisture to the central parts of Eurasia.
Further comparison of the KTK2 records with orbitally-inducedinsolation variations (Fig. 7b) and with the global sea levelreconstruction (Fig. 7c) suggests that the phase with the maximalspread of cold steppe communities and minimal representation ofwoody taxa in the regional vegetation (Fig. 7d) occurred during theinterval, when the NH summer insolation and global sea level reachedtheir minima, with the global ice volume consequently reaching itsmaximum. The records summarized in Fig. 7 and the KTK2 diatomrecord imply that the LGM in the Lake Baikal region and probably onthe global scale should be placed at ~25–24 kyr BP and not ~21 kyr BP,as conventionally used in the palaeoenvironmental studies (e.g.Tarasov et al., 2000; Yokoyama et al., 2000; Clark et al., 2006) and inmodelling experiments (e.g. Kageyama et al., 2001). Our conclusionbased upon the Lake Kotokel data is in line with the growing body ofevidence from elsewhere that suggests an earlier onset of the LGM(e.g. Allen and Huntley, 2000; Peltier and Fairbanks, 2006; Genty,2008, Veres et al., 2008; Sirocko, 2009).
The relatively high temporal resolution and reliable AMS-basedage model of the KTK2 pollen record enable its comparison with thereference palaeoclimatic archives representing North Atlantic (e.g. Littand Stebich, 1999; Brauer et al., 2008; Svensson et al., 2008; Sirocko,2009) and North Pacific regions (Yuan et al., 2004; Stebich et al.,2009). This comparison suggests that the reconstructed shifts in latePleistocene–Holocene vegetation and environments in the Lake Baikalregion could have been controlled by the major factors controlling NHclimate (Fig. 7). The correspondence between the KTK2 record andvery high-resolution isotope and pollen records from far distant North

Fig. 7. Comparison of (a) the δ18O records from Greenland, as indicator of the Northern Hemisphere (NH) air temperature (after Svensson et al., 2008) and (e) from Chinesestalagmites (afterWang et al., 2001; Yuan et al., 2004), as indicator of the Pacificmonsoon intensity, (b) the NH June and December insolation at 60°N (after Berger and Loutre, 1991),(c) global sea level (after Yokoyama et al., 2007), as indicator of the global ice volume, and (d) numerical biome scores for steppe and taiga, as palaeoclimatic indicators in the LakeBaikal region (this study), plotted along their respective time scales. The arrows refer to the Greenland Interstadials (GI1 to 12) shown in the left graph. Grey bars indicateapproximate positions of the Younger Dryas (YD) and Heinrich events H1 to H5 (after Tierney et al., 2008).
196 E.V. Bezrukova et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 296 (2010) 185–198
Atlantic and North Pacific regions implies that southern Siberia,despite its location in the interior of the Eurasian landmass, respondedswiftly to global change. This conclusion is a key point in the ongoingdebate on the synchronic/non-synchronic environmental dynamics ofterrestrial environments within the Lake Baikal region during the last15 kyr, based upon less accurately dated pollen records (see Demskeet al., 2005; Shichi et al., 2009 for discussion and references).Moreover, in the current absence of better-resolved and datedsequences, the KTK2 core from Lake Kotokel may serve as a regionalstratotype section (Bezrukova et al., 2008), which allows one to revisitthe chronologies of the earlier published pollen/environmentalrecords from the Lake Baikal region and possibly reconsider some ofthe earlier interpretations.
Acknowledgements
This paper is a contribution to the German Research Foundation(DFG) project TA-540/1. The study was also partly supported by theRussian Foundation for Basic Research (RFBR) project 09-05-00123.We would like to acknowledge Prof. H. Takahara and all teammembers for participating in the coring campaign at Lake Kotokel in2005, Prof. T. Goslar (Poznan, Poland) for the AMS dating of the KTK2samples, and Dr. E. Schalk for polishing the English. We are grateful to
Prof. J.P. Smol, Dr. K. Kremenetski and Prof. A.P. Kershaw for theirthorough review, which helped to improve the manuscript. Theauthors like to acknowledge fruitful discussions of the resultspresented in the current study during the RFBR- and DFG-sponsored(TA-540/4) International Workshop “Bridging Eurasia” (Freie Uni-versität Berlin, April 28–May 2 2010). The paper is dedicated toNatalia V. Ignatova, who passed away in August 2008.
References
Allen, J.R.M., Huntley, B., 2000. Weichselian palynological records from southernEurope: correlation and chronology. Quat. Int. 111–125 73/74.
Alpat'ev, A.M., Arkhangel'skii, A.M., Podoplelov, N.Y., Stepanov, A.Y., 1976. Fizicheskayageografiya SSSR (Aziatskaya chast'). Vysshaya Shkola, Moscow.
Anderson, P.M., Lozhkin, A.V., 2001. The Stage 3 interstadial complex (Karginskii/middle Wisconsinan interval) of Beringia: variations in paleoenvironments andimplications for paleoclimatic interpretations. Quat. Sci. Rev. 20, 93–125.
Andreev, A.A., Tarasov, P.E., 2007. Pollen records, postglacial: Northern Asia. In:Elias, S.A. (Ed.), Encyclopedia of Quaternary Science, 4. Elsevier, Amsterdam,pp. 2721–2729.
Battarbee, R.W., Jones, V.J., Flower, R.J., Cameron, N.G., Bennion, H., Carvalho, L., Juggins,S., 2001. Diatoms. In: Smol, J.P., Birks, H.J.B. (Eds.), Tracking Environmental Changeusing Lake Sediments. Terrestrial, Algal, and Siliceous Indicators, 3. KluwerAcademic Publishers, Dordrecht, pp. 171–205.
Bennett, K.D., 1996. Determination of the number of zones in a biostratigraphicsequence. New Phytol. 132, 155–179.

197E.V. Bezrukova et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 296 (2010) 185–198
Berger, A., Loutre, M.F., 1991. Insolation values for the climate of the last 10 millionyears. Quat. Sci. Rev. 10 (4), 297–317.
Berglund, B.E., Ralska-Jasiewiczowa, M., 1986. Pollen analysis and pollen diagrams. In:Berglund, B.E. (Ed.), Handbook of Holocene Palaeoecology and Palaeohydrology.Interscience, New-York, pp. 455–484.
Bezrukova, E.V., Abzaeva, A.A., Letunova, P.P., Kulagina, N.V., Vershinin, K.E., Belov, A.V.,Orlova, L.A., Danko, L.V., Krapivina, S.M., 2005. Post-glacial history of Siberianspruce (Picea obovata) in the lake Baikal area and the significance of this species asa paleo-environmental indicator. Quat. Int. 136, 47–57.
Bezrukova, E.V., Krivonogov, S.K., Takahara, H., Letunova, P.P., Shichi, K., Abzaeva, A.A.,Kulagina, N.V., Zabelina, Yu.S., 2008. Lake Kotokel as a Stratotype for the Late Glacialand Holocene in Southeastern Siberia. Doklady Earth Sci. 420, 658–663.
Bout-Roumazeilles, V., Combourieu Nebout, N., Peyron, O., Cortijo, E., Landais, A.,Masson-Delmotte, V., 2007. Connection between South Mediterranean climate andNorth African atmospheric circulation during the last 50, 000 yr BP North Atlanticcold events. Quat. Sci. Rev. 26, 3197–3215.
Brauer, A., Haug, G.H., Dulski, P., Sigman, D.M., Negendank, J.F.W., 2008. An abrupt windshift in western Europe at the onset of the Younger Dryas cold period. NatureGeosci. 1, 520–523.
Cameron, N.G., Birks, H.J.B., Jones, V.J., Berge, F., Catalan, J., Flower, R.J., Garcia, J.,Kawecka, B., Koinig, K.A., Marchetto, A., Sanchez-Castillo, P., Schmidt, R., Sisko, M.,Solovieva, N., Stefkova, E., Valasquez, T., 1999. Surface-sediment and epilithicdiatom calibration set for remote European mountain lakes (AL:PE Project) andtheir comparison with the Surface Waters Acidification Programme (SWAP)calibration set. J. Paleolimn. 22, 291–317.
Clark, P.U., Archer, D., Pollard, D., Blum, J.D., Rial, J.A., Brovkin, V., Mix, A.C., Pisias, N.G., Roy,M., 2006. The middle Pleistocene transition: characteristics, mechanisms andimplications for long-term changes in atmospheric CO2. Quat. Sci. Rev. 25, 3150–3184.
Colman, S.M., Jones, G.A., Rubin, M., King, J.W., Peck, J.A., Orem, W.H., 1996. AMSradiocarbon analyses from Lake Baikal, Siberia: challenges of dating sedimentsfrom a large, oligotrophic lake. Quat. Sci. Rev. 15, 669–684.
Danzeglocke, U., Jöris, O., Weninger, B., 2008. CalPal-2007online. http://www.calpal-online.de.
DeFries, R.S., Townshend, J.R.G., Hansen, M.C., 1999. Continuous fields of vegetationcharacteristics at the global scale at 1-km resolution. J. Geophys. Res. 104,16911–16923.
Demske, D., Heumann, G., Granoszewski, W., Nita, M., Mamakowa, K., Tarasov, P.E.,Oberhansli, H., 2005. Late glacial and Holocene vegetation and regional climatevariability evidenced in high-resolution pollen records from Lake Baikal. Glob.Planet. Change 46, 255–279.
Edwards, M.E., Brubaker, L.B., Ager, T.A., Andreev, A.A., Bigelow, N.H., Cwynar, L.C.,Eisner, W.R., Harrison, S.P., Hu, F.-S., Jolly, D., Lozhkin, A.V., MacDonald, G.M., Mock,C.J., Ritchie, J.C., Sher, A.V., Spear, R.W., Williams, J.W., Yu, G., 2000. Pollen-basedbiomes for Beringia 18, 000, 6000 and 0 14C yr BP. J. Biogeogr. 27, 521–554.
Finkelstein, S.A., Gajewski, K., 2008. Responses of Fragilarioid-dominated diatomassemblages in a small Arctic lake to Holocene climatic changes, Russell Island,Nunavut. Canada. J. Paleolimnol. 40, 1079–1095.
Fluin, J., Tibby, J., Gell, P., 2010. The palaeolimnological record from Lake Culluleraine,lower Murray River, (south-east Australia): implications for understandingriverine histories. J. Paleolimnol. 43, 309–322.
Frechen, M., Zander, A., Zykina, V., Boenigk, W., 2005. The loess record from the sectionat Kurtak in Middle Siberia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 228, 228–244.
Frenzel, B., Pecsi, B., Velichko, A.A., 1992. Atlas of Palaeoclimates and Palaeoenviron-ments of the Northern Hemisphere. INQUA/Hungarian Academy of Sciences,Budapest.
Galaziy, G.I. (Ed.), 1993. Baikal Atlas. Federal Agency for Geodesy and Cartography ofRussia, Moscow.
Genty, D., 2008. Palaeoclimate research in Villars Cave (Dordogne, SW-France). Int. J.Speleol. 37 (3), 173–191.
Gervais, B.R., MacDonald, G.M., Snyder, J.A., Kremenetski, C.V., 2002. Pinus sylvestristreeline development and movement on the Kola Peninsula of Russia: pollen andstomate evidence. J. Ecology 90, 627–638.
Gómez, N., Riera, J.L., Sabater, S., 1995. Ecology and morphological variability ofAulacoseira granulata (Bacillariophyceae) in Spanish reservoirs. J. Plankton Res. 17,1–16.
Grichuk, V.P., 1984. Late Pleistocene vegetation history. In: Velichko, A.A., Wright Jr.,H.E., Barnosky, C.W. (Eds.), Late Quaternary Environments of the Soviet Union.University of Minnesota Press, Minneapolis, MN, pp. 155–178.
Grimm, E.C., 1987. CONISS: A Fortran 77 program for stratigraphically constrainedcluster analysis by the method of incremental sum of squares. Comput. Geosci. 13,13–35.
Grimm, E.C., 2004. TGView. Illinois State Museum, Research and Collections Center,Springfield.
Guiot, J., Pons, A., de Beaulieu, J.L., Reille, M.A., 1989. 140, 000-year continental climatereconstruction from two European pollen records. Nature 338, 309–313.
Haworth, E.Y., 1976. Two late-glacial (Late Devensian) diatom assemblage profiles fromNorthern Scotland. New Phytol. 77 (1), 227–256.
Heusser, L., Heusser, C., Pisias, N., 2006. Vegetation and climate dynamics of southernChile during the past 50, 000 years: results of ODP Site 1233 pollen analysis. Quat.Sci. Rev. 25, 474–485.
Hickman, M., Reasoner, M.A., 1994. Diatom responses to late Quaternary vegetation andclimate change, and to deposition of two tephras in an alpine and a subalpine lakein Yoho National Park, British Columbia. J. Paleolimnol. 11, 173–188.
Horiuchi, K., Minoura, K., Hoshino, K., Oda, T., Nakamura, T., Kawai, T., 2000.Palaeoenvironmental history of Lake Baikal during the last 23000 years. Palaeo-geogr. Palaeoclimatol. Palaeoecol. 157, 95–108.
Hubberten, H.-W., Andreev, A.A., Astakhov, V.I., Demidov, I., Dowdeswell, J.A.,Henriksen, M., Hjort, C., Houmark-Nielsen, M., Jakobsson, M., Kuzmina, S., Larsen,E., Pekka Lunkka, J., Lyså, A., Mangerud, J., Möller, P., Saarnisto, M., Schirrmeister, L.,Sher, A.V., Siegert, C., Siegert, M.J., Svendsen, J.I., 2004. The periglacial climate andenvironment in northern Eurasia during the Last Glaciation. Quat. Sci. Rev. 23,1333–1357.
Juggins, S., 1991. ZONE. Unpublished computer program, version 1.2. Department ofGeography, University of Newcastle, Newcastle-upon-Tyne.
Juggins, S., 2007. C2 version 1.5 User guide. Software for ecological and palaeoecologicaldata analysis and visualisation. Department of Geography, Newcastle University,Newcastle upon Tyne.
Kageyama, M., Peyron, O., Pinot, S., Tarasov, P., Guiot, J., Joussaume, S., Ramstein, G.,PMIP participating groups, 2001. The Last Glacial Maximum climate over Europeand western Siberia: a PMIP comparison between models and data. Clim. Dyn. 17(1), 23–43.
Khursevich, G.K., Prokopenko, A.A., Fedenya, S.A., Tkachenko, L.I., Williams, D.F., 2005.Diatom biostratigraphy of Lake Baikal during the past 1.25 Ma: new results fromBDP-96-2 and BDP-99 drill cores. Quat. Int. 36, 95–104.
Kienast, F., Schirrmeister, L., Siegert, C., Tarasov, P., 2005. Palaeobotanical evidence forwarm summers in the East Siberian Arctic during the last cold stage. Quat. Res. 63,283–300.
Kilham, S.S., Kilham, P., 1975.Melosira granulata (Ehr) Ralfs, morphology and ecology ofa cosmopolitan freshwater diatom. Verh. Int. Ver. Limnol. 19, 2716–2721.
Krammer, K., Lange-Bertalot, H., 1986–1991. Bacillariophyceae. Gustav Fisher Verlag,Stuttgart.
Kuzmin, V.N., 1988. Bioproduktivnost' evtrofnykh ozer Irkana i Kotokel basseina ozeraBaikal. Promrybvod, Leningrad.
Kuzmin, M.I., Williams, D.F., Logachev, N.A., Colman, S.M., Khakhaev, B.N., Kawai, T.,Hearn, P.P., Horie, S., Pevzner, L.A., Bukharov, A.A., Fialkov, V.A., 1993. The BaikalDrilling Project; scientific objectives and recent results. Russ. Geol. Geophys. 34(10–11), 5–15.
Litt, T., Stebich, M., 1999. Bio- and chronostratigraphy of the lateglacial in the Eifelregion, Germany. Quat. Int. 61, 5–16.
Lotter, A.F., Bigler, C., 2000. Do diatoms in the Swiss Alps reflect the length of ice-cover?Aquat. Sci. 62, 125–141.
Mackay, A.W., 2007. The paleoclimatology of Lake Baikal: a diatom synthesis andprospectus. Earth Sci. Rev. 82, 181–215.
Margalef, R., 1983. Limnologia. Omega, Barcelona.Mokhova, L., Tarasov, P., Bazarova, V., Klimin, M., 2009. Quantitative biome
reconstruction using modern and late Quaternary pollen data from the southernpart of the Russian Far East. Quat. Sci. Rev. 28, 2913–2926.
Müller, S., Bobrov, A.A., Schirrmeister, L., Andreev, A.A., Tarasov, P.E., 2009. Testateamoebae record from the Laptev Sea coast and its implication for thereconstruction of Late Pleistocene and Holocene environments in the ArcticSiberia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 271, 301–315.
Oda, T., Minoura, K., Fujimura, C., Nakamura, T., Kawai, T., 2000. Vegetation history ofthe Baikal drainage basin during the last 250, 000 years. Japan J. Palynol. 46, 81–92.
Ohtaka, A., Watanabe, R., Im, S., Chhay, R., Tsukawaki, S., 2010. Spatial and seasonalchanges in net plankton and zoobenthos in Lake Tonle Sap, Cambodia. Limnology11, 85–94.
Peltier, W.R., Fairbanks, R.G., 2006. Global glacial ice volume and Last Glacial Maximumduration from an extended Barbados sea level record. Quat. Sci. Rev. 25, 3322–3337.
Popp, S., Diekmann, B., Meyer, H., Siegert, C., Syromyatnikov, I., Hubberten, H.-W., 2006.Palaeoclimate signals as inferred from stable-isotope composition of ground ice inthe Verkhoyansk Foreland, Central Yakutia. Permafrost Periglac. Process. 17,119–132.
Prentice, I.C., Guiot, J., Huntley, B., Jolly, D., Cheddadi, R., 1996. Reconstructing biomesfrom palaecological data: a general method and its application to European pollendata at 0 and 6 ka. Clim. Dyn. 12, 185–194.
Prokopenko, A.A., Williams, D.F., Kuzmin, M.I., Karabanov, E.B., Khursevich, G.K., Peck, J.A.,2002. Muted climate variations in continental Siberia during the mid-Pleistoceneepoch. Nature 418, 65–68.
Prokopenko, A.A., Khursevich, G.K., Bezrukova, E.V., Kuzmin, M.I., Boes, X., Williams, D.F.,Fedenya, S.A., Kulagina, N.V., Letunova, P.P., Abzaeva, A.A., 2007. Paleoenvironmentalproxy records from Lake Hovsgol, Mongolia, and a synthesis of Holocene climatechange in the Lake Baikal watershed. Quat. Res. 68, 2–17.
Prokopenko, A.A., Bezrukova, E.V., Khursevich, G.K., Solotchina, E.P., Kuzmin, M.I.,Tarasov, P.E., 2010. Climate in continental interior Asia during the longestinterglacial of the past 500, 000 years: the new MIS 11 records from Lake Baikal,SE Siberia. Clim. Past 6, 31–48.
Rudaya, N., Tarasov, P., Dorofeyuk, N., Solovieva, N., Kalugin, I., Andreev, A., Daryin, A.,Diekmann, B., Riedel, F., Tserendash, N., Wagner, M., 2009. Holocene environmentsand climate in the Mongolian Altai reconstructed from the Hoton-Nur pollen anddiatom records: a step towards better understanding climate dynamics in CentralAsia. Quat. Sci. Rev. 28, 540–554.
Rühland, K.M., Paterson, A.M., Smol, J.P., Pienitz, R., 2008. Hemispheric-scale patterns ofclimate-related shifts in planktonic diatoms from North American and Europeanlakes. Global Change Biology 14, 2740–2754.
Shahgedanova, M. (Ed.), 2002. The Physical Geography of Northern Eurasia. OxfordUniversity Press, Oxford.
Shichi, K., Kawamuro, K., Takahara, H., Hase, Y., Maki, T., Miyoshi, N., 2007. Climate andvegetation changes around Lake Baikal during the last 350, 000 years. Palaeogeogr.Palaeoclim. Palaeoecol. 248, 357–375.
Shichi, K., Takahara, H., Krivonogov, S.K., Bezrukova, E.V., Kashiwaya, K., Takehara, A.,Nakamura, T., 2009. Late Pleistocene and Holocene vegetation and climate recordsfrom Lake Kotokel, central Baikal region. Quat. Int. 205 (1–2), 98–110.

198 E.V. Bezrukova et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 296 (2010) 185–198
Sirocko, F. (Ed.), 2009. Wetter, Klima, Menschheits-Entwicklung von der Eiszeit bis ins21. Jahrhundert. Wissenschaftliche Buchgesellschaft, Darmstadt.
Smol, J.P., 1983. Palaeophycology of a high Arctic lake near Cape Herschel, EllesmereIsland. Can. J. Botany 61, 2195–2204.
Smol, J.P., 1988. Palaeoclimate proxy data from freshwater diatoms. Vern. Int. Ver.Limnol. 23, 837–844.
Smol, J.P., Douglas, M.S.V., 2007. From controversy to consensus: making the case forrecent climatic change in the Arctic using lake sediments. Frontiers in Ecology andthe Environment 5, 466–474.
Solovieva, N., Jones, V.J., 2002. A multiproxy record of Holocene environmental changesin the central Kola Peninsula, northwest Russia. J. Quat. Sci. 17, 303–318.
Solovieva, N., Jones, V.J., Birks, J.H.B., Appleby, P., Nazarova, L., Brooks, S., 2008. Diatomresponses to 20th century climate warming in lakes from the northern Urals.Russia. Palaeogeogr. Palaeoclim. Palaeoecol. 259, 96–106.
Stebich, M., Mingram, J., Han, J., Liu, J., 2009. Late Pleistocene spread of (cool-)temperateforests in Northeast China and climate changes synchronous with the NorthAtlantic region. Glob. Planet. Change 65, 56–70.
Svensson, A., Andersen, K.K., Bigler, M., Clausen, H.B., Dahl-Jensen, D., Davies, S.M.,Johnsen, S.J., Muscheler, R., Parrenin, F., Rasmussen, S.O., Rothlisberger, R., Seierstad, I.,Steffensen, J.P., Vinther, B.M., 2008. A 60 000 year Greenland stratigraphic ice corechronology. Clim. Past 4, 47–57.
Swann, G.E.A., Mackay, A.W., Leng, M.J., Demory, F., 2005. Climatic change in CentralAsia during MIS 3/2: a case study using biological responses from Lake Baikal. Glob.Planet. Change 46, 235–253.
Takahara, H., Krivonogov, S.K., Bezrukova, E.V., Miyoshi, N., Morita, Y., Nakamura, T.,Hase, Y., Shinomiya, Y., Kawamuro, K., 2000. Vegetation history of the southeasternand eastern coasts of Lake Baikal from bog sediments since the last interstade. In:Minoura, K. (Ed.), Lake Baikal: AMirror in Time and Space for Understanding GlobalChange Processes. Elsevier, Amsterdam, pp. 108–118.
Tarasov, P.E., Volkova, V.S., Webb III, T., Guiot, J., Andreev, A.A., Bezusko, L.G., Bezusko, T.V.,Bykova, G.V., Dorofeyuk, N.I., Kvavadze, E.V., Osipova, I.M., Panova, N.K., Sevastyanov,D.V., 2000. Last Glacial Maximum biomes reconstructed from pollen and plantmacrofossil data from Northern Eurasia. J. Biogeogr. 27 (3), 609–620.
Tarasov, P., Granoszewski, W., Bezrukova, E., Brewer, S., Nita, M., Abzaeva, A.,Oberhänsli, H., 2005. Quantitative reconstruction of the Last Interglacial vegetationand climate based on the pollen record from Lake Baikal, Russia. Clim. Dyn. 25 (6),625–637.
Tarasov, P., Bezrukova, E., Karabanov, E., Nakagawa, T., Wagner, M., Kulagina, N.,Letunova, P., Abzaeva, A., Granoszewski, W., Riedel, F., 2007a. Vegetation andclimate dynamics during the Holocene and Eemian interglacials derived from LakeBaikal pollen records. Palaeogeogr. Palaeoclim. Palaeoecol. 252, 440–457.
Tarasov, P., Williams, J.W., Andreev, A., Nakagawa, T., Bezrukova, E., Herzschuh, U.,Igarashi, Y., Müller, S., Werner, K., Zheng, Z., 2007b. Satellite- and pollen-basedquantitative woody cover reconstructions for northern Asia: verification andapplication to late-Quaternary pollen data. Earth Planet. Sci. Lett. 264, 284–298.
Tarasov, P.E., Bezrukova, E.V., Krivonogov, S.K., 2009. Late Glacial and Holocene changesin vegetation cover and climate in southern Siberia derived from a 15 kyr longpollen record from Lake Kotokel. Clim. Past 5, 285–295.
ter Braak, C.F.J., 1986. Canonical correspondence analysis: a new eigenvector techniquefor multivariate direct gradient analysis. Ecology 67, 1167–1179.
ter Braak, C.F.J., Šmilauer, P., 2002. CANOCO Reference manual and CANODRAWfor Windows User's guide: Software for Canonical Community Ordination(version 4.5), Ithaca, NY.
Tierney, J.E., Russell, J.M., Huang, Y., Sinninghe Damsté, J.S., Hopmans, E.C., Cohen, A.S.,2008. Northern hemisphere controls on tropical southeast African climate duringthe past 60, 000 years. Science 322 (5899), 252–255.
Tütken, T., 2003. The implications of early diagenesis to the preservation of in vivoelemental and isotopic compositions in fossil bones. PhD Thesis, Eberhard-KarlsUniversität Tübingen. Online publication, laste access 9 December 2009, http://w210.ub.uni-tuebingen.de/dbt/volltexte/2003/962/.
Veres, D., Lallier-Verges, E., Wohlfarth, B., Lacourse, T., Keravis, D., Bjorck, S., Preusser, F.,Andrieu-Ponel, V., Ampel, L., 2008. Climate-driven changes in lake conditionsduring late MIS 3 and MIS 2: a high-resolution geochemical record from Les Echets,France. Boreas 38, 230–243.
Vincens, A., Buchet, G., Williamson, D., Taieb, M.A., 2005. 23, 000 yr pollen record fromLake Rukwa (88 S, SW Tanzania): new data on vegetation dynamics and climate inCentral Eastern Africa. Rev. Palaeobot. Palynol. 137, 147–162.
Vipper, P.B., Smirnov, N.N., 1979. The studying of biogeocenozes history by the freshwater lakes bottom deposits, Obshchie metody izucheniya istorii sovremennykhekosistem, Nauka, Moscow, pp. 14–39 (in Russian).
Walker, M., Johnsen, S., Rasmussen, S.O., Popp, T., Steffensen, J.-P., Gibbard, P., Hoek, W.,Lowe, J., Andrews, J., Björck, S., Cwynar, L.C., Hughen, K., Kershaw, P., Kromer, B.,Litt, T., Lowe, D.J., Nakagawa, T., Newnham, R., Schwander, J., 2009. Formaldefinition and dating of the GSSP (Global Stratotype Section and Point) for the baseof the Holocene using the Greenland NGRIP ice core, and selected auxiliary records.J. Quat. Sci. 24 (1), 3–17.
Wang, Y.J., Cheng, H., Edwards, R.L., An, Z.S., Wu, J.Y., Shen, C.-C., Dorale, J.A., 2001. Ahigh-resolution absolute-dated Late Pleistocene Monsoon record from Hulu Cave,China. Science 294, 2345–2348.
Wanner, H., Beer, J., Bütikofer, J., Crowley, T.J., Cubasch, U., Flückiger, J., Goosse, H.,Grosjean, M., Joos, F., Kaplan, J.O., Küttel, M., Müller, S.A., Prentice, I.C., Solomina, O.,Stocker, T.F., Tarasov, P., Wagner, M., Widmann, M., 2008. Mid- to late Holoceneclimate change — an overview. Quat. Sci. Rev. 27 (19–20), 1791–1828.
Westover, K.S., Fritz, S.C., Blyakharchuk, T.A., Wright, H.E., 2006. Diatom paleolimno-logical record of Holocene climatic and environmental change in the AltaiMountains, Siberia. J. Paleolim. 35, 519–541.
Winder, M., John, E., Reuter, J.E., Schladow, G.S., 2009. Lake warming favours small-sized planktonic diatom species. Proc. R. Soc. B. 276, 427–435.
Williams, D.F., Peck, J., Karabanov, E.B., Prokopenko, A.A., Kravchinsky, V., King, J.,Kuzmin, M.I., 1997. Lake Baikal record of continental climate response to orbitalisolation during the past 5 million years. Science 278, 1114–1117.
Williams, D.F., Kuzmin, M.I., Prokopenko, A.A., Karabanov, E.B., Khursevich, G.K.,Bezrukova, E.V., 2001. The Lake Baikal drilling project in the context of a global lakedrilling initiative. Quat. Int. 3–18 80/81.
Williams, J.W., Shuman, B.N.,Webb III, T., Bartlein, P.J., Leduc, P.L., 2004. Late Quaternaryvegetation dynamics in North America: scaling from taxa to biomes. Ecol. Monogr.74, 309–334.
Yokoyama, Y., Lambeck, K., De Deckker, P., Johnston, P., Fifield, L.K., 2000. Timing of theLast Glacial Maximum from observed sea-level minima. Nature 406, 713–716.
Yokoyama, Y., Kido, Y., Tada, R., Minami, I., Finkel, R.C., Matsuzaki, H., 2007. Japan Seaoxygen isotope stratigraphy and global sea-level changes for the last 50, 000 yearsrecorded in sediment cores from the Oki Ridge. Palaeogeogr. Palaeoclimatol.Palaeoecol. 247, 5–17.
Yuan, D.X., Cheng, H., Edwards, R.L., Dykoski, C.A., Kelly, M.J., Zhang, M.L., Qing, J.M., Lin,Y.S., Wang, Y.J., Wu, J.Y., Dorale, J.A., An, Z.S., Cai, Y.J., 2004. Timing, duration, andtransitions of the Last Interglacial Asian monsoon. Science 304, 575–578.