Embed Size (px)
Citation preview

Mutation Research 622 (2007) 84–93
Zinc fluxes and zinc transporter genes in chronic diseases
Chiara Devirgiliis a, Peter D. Zalewski b, Giuditta Perozzi a, Chiara Murgia a,∗a INRAN, National Research Institute on Food and Nutrition, Via Ardeatina 546 00178, Roma, Italy
b Department of Medicine, University of Adelaide, The Queen Elizabeth Hospital, Woodville, SA 5011, Australia
Received 24 October 2006; received in revised form 16 January 2007; accepted 24 January 2007Available online 17 February 2007
Abstract
The group IIb metal zinc (Zn) is an essential dietary component that can be found in protein rich foods such as meat, seafood andlegumes. Thousands of genes encoding Zn binding proteins were identified, especially after the completion of genome projects, anindication that a great number of biological processes are Zn dependent. Imbalance in Zn homeostasis was found to be associatedwith several chronic diseases such as asthma, diabetes and Alzheimer’s disease. As it is now evident for most nutrients, bodyZn status results from the interaction between diet and genotype. Zn ions cross biological membranes with the aid of specializedmembrane proteins, belonging to the ZRT/IRT-related Proteins (ZIP) and zinc transporters (ZnT) families. The ZIPs are encodedby the Slc39A gene family and are responsible for uptake of the metal, ZnTs are encoded by the Slc30A genes and are involvedin intracellular traffic and/or excretion. Both ZnTs and Zips exhibit unique tissue-specific expression, differential responsivenessto dietary Zn deficiency and excess, as well as to physiological stimuli via hormones and cytokines. Intracellular Zn concentrationis buffered by metallothioneins (MTs), a class of cytosolic protein with high affinity for metals. Scattered information is availableon the role of proteins responsible for regulating Zn fluxes in the onset and progression of chronic diseases. This paper reviews
reports that link Zn transporter genes, their allelic variants and/or expression profiles in the context of specific diseases. Furtherinvestigation in this direction is very important, since Zn imbalance can result not only from insufficient dietary intake, but alsofrom impaired activity of proteins that regulate Zn metabolism, thus contributing to multifactorial diseases.© 2007 Elsevier B.V. All rights reserved.Keywords: Zinc; Zinc transporters; Nutrigenomics; Chronic diseases
1. Zinc homeostasis
Zinc (Zn) is an essential micronutrient that playsfundamental housekeeping roles in physiology, cellu-lar metabolism and gene expression. Zn is required as
a catalytic cofactor of more than 300 enzymes and itstabilizes the structure of thousands of protein domains.Recently reported results of a global search within the∗ Corresponding author. Tel.: +39 0651494457;fax: +39 0651494550.
E-mail address: [email protected] (C. Murgia).
0027-5107/$ – see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.mrfmmm.2007.01.013
human genome database with a bioinformatics approachestimated that about 10% of the human proteome con-sists of potential Zn-binding proteins [1]. Therefore, agreat number of biological processes are Zn dependent,and an imbalance of Zn homeostasis has complex impli-cations in a number of organs and can contribute to theonset of chronic pathologies [2,3]. The RecommendedDaily Allowance (RDA) for Zn is 12–15 mg/day, in bal-anced diets this amount is obtained by eating meat and
other sources of animal proteins. These foods have a highZn content and the metal is bound to ligands, which facil-itate its absorption. Other sources of Zn are sea-foods,dairy products, cereals and nuts [4]. Most vegetables are
tion Re
ntimiiapa
ttZiao(ota[
tf
TF
*
C. Devirgiliis et al. / Muta
ot ready sources of Zn because of the presence of phy-ate, abundant in legumes and cereals, that chelates metalons and inhibits their absorption [5]. Diets low in ani-al proteins and rich in phytate contribute to the high
ncidence of mild–moderate Zn deficiency in develop-ng countries. Individuals feeding on a balanced diet maylso undergo suboptimal intake of Zn as result of specifichysiological status, for example, during pregnancy orcute inflammation (Table 1).
Intracellular Zn pools can be visualized and quan-ified using autometallography (AMG), a Zn labellingechnique adapted for electron microscopy [6] andn fluorophores [7,8]. These techniques reveal that
ntracellular Zn is found in tight association with met-lloproteins, a largely fixed pool with a slow turnover,r in a more dynamic, free or loosely bound formlabile) that is more readily exchangeable. The amountf intracellular labile Zn differs according to cellype and can participate in specialized functions suchs secretion, cytoprotection and signal transduction
9].Zinquin is the prototype of a panel of Zn fluorophoreshat enabled researchers to map labile Zn and study itsunction in different tissues [2,7,10]. At least half a dozen
able 1eatures of Zn fluxes in tissues displaying alterations associated with disease
Expression of all known members of ZIP and ZnT families has not been inv
search 622 (2007) 84–93 85
other Zn specific fluorophores are now available withdifferent affinities and specificities so that informationon Zn distribution and fluxes is rapidly accumulating[9,11].
Each cell type needs a constant supply of Zn accord-ing to its function; on the other hand, excess of free Znions can be toxic. In order to maintain controlled lev-els and appropriate distribution of this essential metal,tightly regulated homeostatic mechanisms need to bein place. This is achieved through the action of a setof specialized proteins that control Zn uptake, effluxand compartmentalization. These proteins and theircorresponding genes are being rapidly identified andcharacterized [12,13]. Maintenance of body and cellu-lar Zn homeostasis therefore requires a great number ofgenes whose polymorphisms and mutations are likelyto provide complex genetic variations and sensitivityto dietary Zn in health and disease. In this review, wewill focus on the available information on the proteinsresponsible for transmembrane Zn transport (uptake and
intracellular compartmentalization), as well as thoseresponsible for intracellular Zn storage and on theirpotential role in specific diseases associated with alter-ations in body Zn status.estigated in the brain.

tion Re
86 C. Devirgiliis et al. / Muta1.1. Metallothioneins
Metallothioneins (MTs) are small cytosolic proteinswith a high cysteine content (25–30%) that bind Zn, aswell as other metal ions, with high affinity. MTs werethe first group of proteins involved in Zn homeostasis tobe identified and extensively studied. MTs are thought tobe responsible for the intracellular regulation of Zn con-centration and for detoxification of non-essential heavymetals [14]. MT synthesis is strongly induced by metals,mediated by the metal-responsive transcription factor-1 (MTF-1), an essential protein for liver developmentand cell stress response [15]. A single nucleotide poly-morphism (SNP) in the promoter of the MT2A genewas associated with hyperglycemia in a population ofelderly diabetic patients. The same SNP was shown topredispose to Zn deficiency [16]. Another pioneeringstudy on the genetics of Zn metabolism, published byTamura et al. and involving linkage analysis and haplo-type mapping, suggested that a SNP in the MTF-1 genewas associated with lymphoma susceptibility [17]. Theinvolvement of MTF-1 and MT genes in such unrelateddiseases confirms the pleiotropic role of the correspond-ing proteins.
1.2. Zinc transporters
Zn is highly charged and cannot cross biologicalmembranes by passive diffusion, requiring the aid of aset proteins, grouped into two families on the basis oftheir structural and functional features: Solute CarrierFamily 39A (SLC39A) includes mammalian ZRT/IRT-related proteins (ZIPs) [18], and Solute Carrier Family30A (SLC30A), comprises mammalian ZnTs [19]. BothZIPs and ZnTs are multipass transmembrane proteinsand have a high content of lipophilic amino acids. Four-teen ZIP-encoding genes have been identified in thehuman genome and are grouped into four sub-familiesaccording to the molecular features of the encoded pro-teins. All of the Zip proteins mediate Zn uptake fromthe extracellular environment or intracellular vesiclesinto the cytoplasm. Proteins belonging to the ZnT fam-ily were shown to be responsible for Zn efflux from thecytoplasm towards either intracellular vesicles or/and theextracellular space [20], as well as for Zn delivery tospecific metalloproteins [21]. Mutations in two Zn trans-porters have been linked to the Zn deficiency diseasesacrodermatitis enteropathica (ZIP4) in humans [22] and
lethal milk syndrome (ZnT4) in mice [23] (reviewedin ref. [24]). A point mutation in ZnT2 was recentlyassociated with transient Zn deficiency in exclusivelybreast-fed infants [25].search 622 (2007) 84–93
2. Zinc and disease
Several lines of evidence point to Zn and itsmetabolism as important players in the onset and/or pro-gression of multifactorial diseases. Due to its widespreadfunctions in a range of organs, Zn imbalance in specifictissues is likely to contribute to several diseases. In thisreview, we focus on asthma, diabetes and Alzheimer’sdisease, the few pathologies in which a specific role forZn was reported and we discuss disease association withproteins responsible for Zn transport (uptake and intra-cellular compartmentalization), as well as with thoseresponsible for intracellular Zn storage.
2.1. Asthma
Asthma is a chronic airways inflammatory diseasecaused by environmental factors in genetically predis-posed individuals [26]. Asthma affects an increasingproportion of the population and is characterized byrecurrent episodes of wheezing, breathlessness and chesttightness caused by airway inflammation. Growing evi-dence links asthma and Zn deficiency. Case–controlstudies found that the combined risk of atopy, bronchialreactivity and allergic-type symptoms were four- tofive-fold higher in individuals with low Zn intake ascompared to those with higher Zn intakes [27,28].Maternal Zn intake during pregnancy was negativelyassociated with shortness of breath and asthma in astudy on children with a 5 years follow up [29]. Inanother study, hair Zn levels were significantly lowerin wheezy infants than in healthy controls, suggestingthat Zn deficiency may influence the risk of wheez-ing in early childhood [3,30]. The airways epitheliumfrom different species shows intense Zinquin stainingthat is completely quenched in the presence of the Znchelator TPEN (N,N,N′,N′-tetrakis-(2-pyridylmethyl)-ethylenediamine) [10]. In this tissue, labile Zn poolswere largely restricted to apical structures in the proxim-ity of cilia basal bodies [31,32]. Zn deprivation rendersairway epithelial cells highly susceptible to apoptosisinduced by oxidants [32] or by the FAS death receptorpathway [33]. It was suggested that programmed celldeath may play a causative role in acute lung injury duein part to epithelial cell loss. Depletion of labile Zn com-bined with tumor necrosis factor alpha (TNF), interferongamma (IFN) and FAS receptor stimulation acceler-ates caspase-3 activation, proteolysis of E-cadherin and
�-catenin, and cellular apoptosis, leading to increasedparacellular leak across monolayers of both upper airwayand alveolar lung epithelial cultures. Zn supplementationinhibited both apoptosis and paracellular leak [34].
tion Re
mhbahlriarmZindtweacamuTZsiistAabapgctiatttp
2
ifh
C. Devirgiliis et al. / Muta
Features of human asthma can be replicated in animalodels of allergic airway inflammation: Balb/c mice that
ave been sensitised and then aerochallenged with oval-umin (OVA) develop a pronounced airway eosinophiliassociated with airway hyper-responsiveness, mucousyperplasia, damage to the airway epithelium and col-agen deposition [35]. Airway inflammation in miceesults in marked loss of stainable Zn. Mice fed Zn lim-ted diets (ZL, 14 mg/kg Zn), when challenged with thellergen OVA developed greater levels of airways hyper-esponsiveness, as well as increased eosinophilia anducus cell hyperplasia, when compared with mice fed an adequate diet (ZN, 50 mg/kg Zn) [31]. When airway
nflammation was induced by OVA challenge in miceull for the MT-I and MT-II gene functions, the animalsisplayed significant increase of eosinophils and neu-rophils in broncheo-alveolar fluid when compared withild type controls. Expression of interleukin1� was also
nhanced, as were the levels of 8-oxy-deoxyguanosidend nitrotyrosine [36]. These findings led to the con-lusion that Zn and MT exert a protective role againstntigen-induced airway inflammation [9]. Zn supple-entation decreases infiltration of inflammatory cells
pon acute airways inflammation induced by OVA.he expression of genes coding for the full set ofn transporters was investigated in controls and in Znupplemented animals and differences were observed,ncluding increases in ZIP1 and ZIP14 and decreasesn ZIP4 and ZnT4. These changes in gene expres-ion, occurring upon inflammation, were mainly directedowards maintaining or increasing intracellular Zn [37].DAM33 belongs to a subgroup of Zn-dependent met-
lloproteases, it is expressed in lung fibroblasts andronchial smooth muscle, and was proposed to playrole in airways remodelling [38]. A survey of 135
olymorphisms in 23 genes identified the ADAM33ene as being significantly associated with asthma usingase–control, transmission disequilibrium and haplo-ype analyses [22]. These findings indicate that proteinsnvolved in regulating Zn fluxes play an important role inirways physiology, as well as in the onset of inflamma-ory diseases. It becomes therefore relevant to investigateheir genetic variants in order to establish possible rela-ionships between genotype, dietary Zn requirement andredisposition to asthma.
.2. Diabetes
Diabetes is a complex disorder characterized bympaired glucose homeostasis and comprises severalorms, all of them associated with varying degrees ofyperglycemia. The pancreas has a complex architec-
search 622 (2007) 84–93 87
ture. It is mainly composed of exocrine acinar cells,which synthesize and secrete the components of pan-creatic juice. The endocrine component of the pancreasis represented by the islets of Langerhans, interspersedwithin the exocrine tissue. Islet cells control bloodglucose concentration by secreting the two key hor-mones, insulin and glucagon, in response to high or lowglycemia, respectively. Insulin secreting beta cells rep-resent the most abundant islet cell type, while glucagonsecreting alpha cells are fewer. Other two cell types arepresent as minor components of the endocrine pancreas:the delta cells, responsible for synthesis and secretion ofthe inhibitory hormone somatostatin, that exerts multi-ple functions in the body, including downregulation ofboth endocrine and exocrine pancreatic secretions; andthe pancreatic polypeptide producing cells.
Type 1 diabetes, also known as “juvenile” or insulindependent diabetes mellitus (IDDM), is caused by anautoimmune response leading to destruction of islet betacells, which, as a consequence, abolishes insulin produc-tion. On the other hand, Type 2 diabetes is characterizedby hyperinsulinemia caused by a failure in the signalingevents triggered by the activation of the insulin recep-tor [39]. Zn is abundant throughout all pancreas, it isan important component of the pancreatic juice andit is even more concentrated in endocrine islet cells,especially in the secretory vesicles of �-cells where it sta-bilizes the structure of insulin granules [40,41] (Fig. 1).One of the first examples of Zn fluxes in islet cells wasprovided by Zalewski et al. With the aid of the Zn-specific fluorescent probe Zinquin, coupled with digitalimage analysis, labile Zn was visualized and measuredin intact islets and in dissociated islet cells. Labile Znappeared concentrated in secretory vesicles, and a morediffuse cytoplasmic fluorescence signal accounted foran extragranular pool of labile Zn, which could act as areservoir for the newly formed secretory granules [42].
Changes in intracellular free Zn levels occur inresponse to external stimuli such as increasing concen-tration of glucose. Cells incubated in the presence of25.6 mM glucose display decreased Zinquin staining[42]. Zn ions in beta cells are involved in biologicalactivities other than stabilization of insulin hexamers.Released Zn ions were proposed to exert a paracrineeffect in promoting glucagon secretion by alpha cells[43], although this result is controversial. In anotherstudy, Zn was reported to have inhibitory effect onglucagon secretion in isolated pancreata and dissociated
alpha cells [44], while no effect was observed in a thirdreport [45]. Moreover, Zn released by beta cells seemsto contribute to their own destruction under conditionsof insulin hypersecretion [46].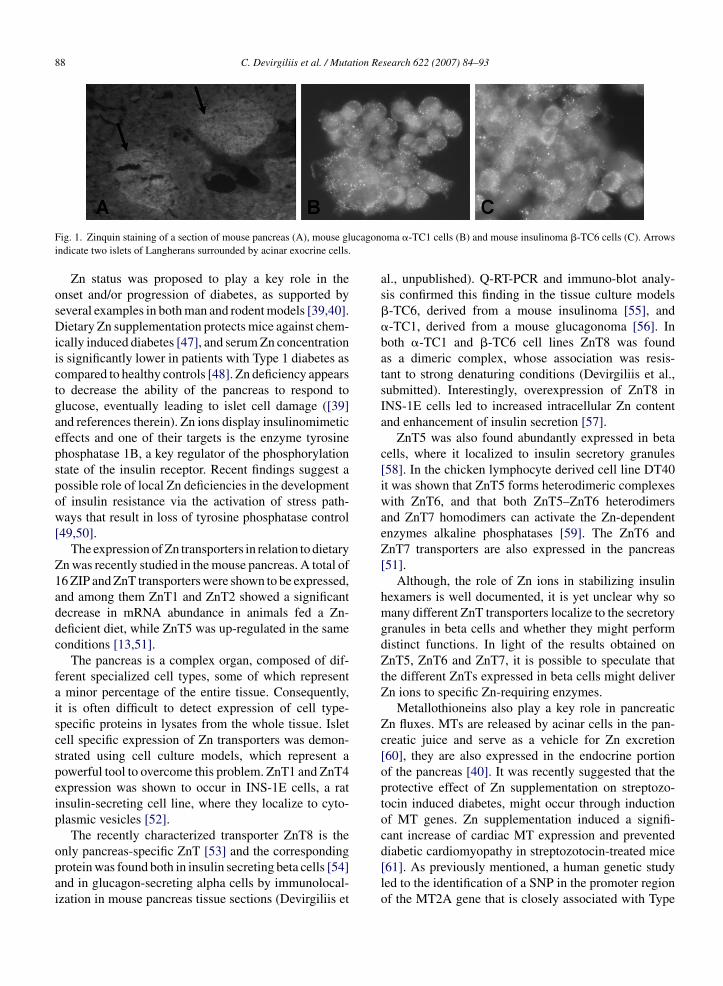
88 C. Devirgiliis et al. / Mutation Research 622 (2007) 84–93
ucagon
Fig. 1. Zinquin staining of a section of mouse pancreas (A), mouse glindicate two islets of Langherans surrounded by acinar exocrine cells.Zn status was proposed to play a key role in theonset and/or progression of diabetes, as supported byseveral examples in both man and rodent models [39,40].Dietary Zn supplementation protects mice against chem-ically induced diabetes [47], and serum Zn concentrationis significantly lower in patients with Type 1 diabetes ascompared to healthy controls [48]. Zn deficiency appearsto decrease the ability of the pancreas to respond toglucose, eventually leading to islet cell damage ([39]and references therein). Zn ions display insulinomimeticeffects and one of their targets is the enzyme tyrosinephosphatase 1B, a key regulator of the phosphorylationstate of the insulin receptor. Recent findings suggest apossible role of local Zn deficiencies in the developmentof insulin resistance via the activation of stress path-ways that result in loss of tyrosine phosphatase control[49,50].
The expression of Zn transporters in relation to dietaryZn was recently studied in the mouse pancreas. A total of16 ZIP and ZnT transporters were shown to be expressed,and among them ZnT1 and ZnT2 showed a significantdecrease in mRNA abundance in animals fed a Zn-deficient diet, while ZnT5 was up-regulated in the sameconditions [13,51].
The pancreas is a complex organ, composed of dif-ferent specialized cell types, some of which representa minor percentage of the entire tissue. Consequently,it is often difficult to detect expression of cell type-specific proteins in lysates from the whole tissue. Isletcell specific expression of Zn transporters was demon-strated using cell culture models, which represent apowerful tool to overcome this problem. ZnT1 and ZnT4expression was shown to occur in INS-1E cells, a ratinsulin-secreting cell line, where they localize to cyto-plasmic vesicles [52].
The recently characterized transporter ZnT8 is the
only pancreas-specific ZnT [53] and the correspondingprotein was found both in insulin secreting beta cells [54]and in glucagon-secreting alpha cells by immunolocal-ization in mouse pancreas tissue sections (Devirgiliis etoma �-TC1 cells (B) and mouse insulinoma �-TC6 cells (C). Arrows
al., unpublished). Q-RT-PCR and immuno-blot analy-sis confirmed this finding in the tissue culture models�-TC6, derived from a mouse insulinoma [55], and�-TC1, derived from a mouse glucagonoma [56]. Inboth �-TC1 and �-TC6 cell lines ZnT8 was foundas a dimeric complex, whose association was resis-tant to strong denaturing conditions (Devirgiliis et al.,submitted). Interestingly, overexpression of ZnT8 inINS-1E cells led to increased intracellular Zn contentand enhancement of insulin secretion [57].
ZnT5 was also found abundantly expressed in betacells, where it localized to insulin secretory granules[58]. In the chicken lymphocyte derived cell line DT40it was shown that ZnT5 forms heterodimeric complexeswith ZnT6, and that both ZnT5–ZnT6 heterodimersand ZnT7 homodimers can activate the Zn-dependentenzymes alkaline phosphatases [59]. The ZnT6 andZnT7 transporters are also expressed in the pancreas[51].
Although, the role of Zn ions in stabilizing insulinhexamers is well documented, it is yet unclear why somany different ZnT transporters localize to the secretorygranules in beta cells and whether they might performdistinct functions. In light of the results obtained onZnT5, ZnT6 and ZnT7, it is possible to speculate thatthe different ZnTs expressed in beta cells might deliverZn ions to specific Zn-requiring enzymes.
Metallothioneins also play a key role in pancreaticZn fluxes. MTs are released by acinar cells in the pan-creatic juice and serve as a vehicle for Zn excretion[60], they are also expressed in the endocrine portionof the pancreas [40]. It was recently suggested that theprotective effect of Zn supplementation on streptozo-tocin induced diabetes, might occur through inductionof MT genes. Zn supplementation induced a signifi-cant increase of cardiac MT expression and prevented
diabetic cardiomyopathy in streptozotocin-treated mice[61]. As previously mentioned, a human genetic studyled to the identification of a SNP in the promoter regionof the MT2A gene that is closely associated with Type
C. Devirgiliis et al. / Mutation Re
Fah
2toptatcbZ
2
gbamcppm
oilsmA[op
ig. 2. Schematic view of the effect of the interaction between diet andllelic variations in genes controlling Zn fluxes on the balance betweenealth and disease.
diabetes [16], but no information is yet available onhe occurrence of SNPs in Zn transporter genes andn their potential association with the development ofancreas-related pathologies. The studies described inhis section highlight the tight relationship between Znnd glucose homeostasis, suggesting that further inves-igation in this direction is very important. Zn imbalancean in fact result not only from insufficient dietary intake,ut also from impaired activity of proteins that regulaten metabolism (Fig. 2).
.3. Alzheimer’s disease
Alzheimer’s disease (AD) is a polygenic neurode-enerative disorder associated with aging, characterizedy progressive impairment of memory and cognitivebilities. AD is thought to result from the abnormal accu-ulation and deposition of extracellular senile plaques
omposed of copper-Zn aggregates of the amyloid �-eptide (A�). A� results from proteolytic cleavage of therecursor APP (amyloid �-protein precursor), an integralembrane glycoprotein family.Proper AD diagnosis can only be made post-mortem
n brain sections displaying the specific neuropatholog-cal features of the disease such as selective neuronaloss and accumulation of A� plaques. A useful tool totudy AD is the transgenic mouse model Tg2576. Theseice overexpress the 695-amino acid isoform of human
PP containing the K670N, and the M671L mutations2,62]. These animals have normal learning and mem-ry in spatial reference at 3 months of age, but botharameters are impaired by the age of 10 months. Such
search 622 (2007) 84–93 89
behavioural deficits are accompanied by deposition ofA� plaque [63].
Zn ions bind a cysteine-rich specific domain thatis conserved in all members of the APP superfamily.Increasing evidence suggests that Zn is involved in ADpathogenesis, and that Zn metabolism is altered in thepathology. Excess Zn was found associated with amy-loid plaques, and it was shown that the interaction ofA� with Zn and other biometals promotes its aggrega-tion and decreases its solubility in vitro. Treatment of theTg2576 transgenic mouse model of AD with clioquinol,an orally bioavailable metal chelator, induced markedinhibition of cortical amyloid accumulation [64]. More-over, early-phase clinical trials indicate that A� plaquedeposition is inhibited by Zn chelation [65,66].
Large amounts of Zn are present in the brain, mostlyconcentrated in the glutamate synaptic vesicles of presy-naptic nerve terminals. Zn ions were shown to inhibitpostsynaptic glutamate receptors. Upon excitation withglutamate, cells rapidly take up Zn through a saturabletransport system. Zn is released along with glutamateduring neurotransmission and may reach concentrationsas high as 300 �M in the synapse and, unless promptlyremoved it can damage adjacent neurons ([66] and refer-ences therein). Excessive exposure to extracellular Zn isin fact toxic for neurons and it is believed to contributeto brain damage in transient cerebral ischemia [62,67].
Zn deficiency deeply affects behaviour of infants andchildren. However, brain Zn concentration is maintainedwithin a narrow range, thus indicating a tight regula-tion of brain Zn homeostasis. MT-I and ZnT1 expressiondecreased in mice fed a Zn limiting diet, while the Znimporter Zip6 was upregulated. This mechanism mayaccount for the conservation of Zn brain during Zn defi-ciency [68].
Neurons take up Zn and deliver it to intracellular tar-gets, maintaining its homeostasis through the activity ofmetallothioneins, ZnTs and ZIP proteins; as found inother cell types, the majority of Zn in neurons is tightlybound to metallo-proteins. In addition, glutamatergicneurons sequestrate an important fraction of Zn intosynaptic vesicles. Therefore, this cell type deals withlarge amounts of labile Zn that need to be moved fromthe cell bodies to the axon terminals to be packaged intosynaptic vesicles. The ZnT3 transporter was shown tobe required for this latter process [69]. Although, ZnT3ablation does not affect brain development and learn-ing abilities [70], it is significant that the introduction
of the ZnT3(−/−) genotype into Tg2576 mice resultsin an approximately 50% reduction in amyloid burdenwhen compared to control, age-matched Tg2576 mice[71]. The prevalence of AD is substantially higher in
tion Re
90 C. Devirgiliis et al. / Mutapostmenopausal females than in males of the same ageboth in humans and rodents. Consistent with this findingZnT3 expression is regulated by oestrogen. With aging,female Tg2576:ZnT3(+/+) mice manifested higher levelsof synaptic Zn, insoluble amyloid beta, and plaques thanmales; these sex linked differences were not present inTg2576:ZnT3(−/−) [71,72].
Of the nine characterized ZnT proteins, ZnT1, T3,T4, T5, T6 and T7 are expressed in the brain [73,74].ZnT1 expression was shown to be reduced in the hip-pocampus/parahippocampal gyrus of subjects with mildcognitive impairment. In the same study ZnT1 expres-sion increased in early and late stage AD subjects, ascompared with age matched controls [75]. In anotherstudy, the same group showed a significant increase ofZnT4 and ZnT6 proteins in AD [76]. ZnT6 distributionwas further investigated in the brain of AD individu-als by immunohistochemistry and shown to co-localizewith the trans-Golgi network (TGN) marker TGN-38in normal cells. Neurons undergoing degeneration showelevated levels of ZnT6 and significant disruption of theTGN. AD brain displays high levels of free Zn. Modula-tion of transporter expression could therefore reflect anattempt to control and redistribute extracellular Zn poolswhose localization is altered in AD [77].
Neurons express MT-III, a brain-specific form ofmetallothionein that is important in buffering high cyto-plasmatic levels of Zn ions. Overexpression of MT-IIIconfers resistance to elevated concentrations of extra-cellular Zn [78,79].
In conclusion, although Zn transporter studies in rela-tion to neurodegenerative diseases are still few and verypreliminary, it is clear that the activities of these proteinsin controlling brain Zn homeostasis can be importantin the onset and progression of AD. The identificationof genetic polymorphisms in the genes controlling Znhomeostasis is therefore essential to establish possiblefunctional associations with Zn metabolism also in thisdisease.
3. Conclusive remarks
Combinations of genetic and environmental factors,including diet, are likely to influence the risk and/or pro-gression of multifactorial diseases. Linkage studies andmutation screens have recently identified mutations inseveral genes associated with chronic diseases [80]. Inthe case of polygenic, multifactorial diseases affected
by diet, this approach has been particularly effective inidentifying nutrient–gene interactions. Increasing evi-dence points to Zn, and its metabolism, as an importantplayer in the onset of a growing number of chronic dis-search 622 (2007) 84–93
eases. In order to evaluate the influence of dietary Znon disease risk, it is therefore extremely important toimplement research approaches that take into consider-ation the allelic variation in the genes encoding proteinsthat regulate Zn fluxes.
Note added in proof
While this paper was under revision an importantstudy was published in which a genome-wide associ-ation study is reported to identify a number of novel riskloci for type 2 diabetes. These loci include a polymor-phism in SLC30A8 gene that encodes for the pancreasspecific Zn transporter ZnT8 [81].
Acknowledgment
The authors acknowledge support by the FISR grant“Improvement of lipid and mineral contents of milk toenhance its nutraceutical and safety properties” from theItalian Ministry of Research (MIUR).
References
[1] C. Andreini, L. Banci, I. Bertini, A. Rosato, Counting the zinc-proteins encoded in the human genome, J. Proteome Res. 5 (2006)196–201.
[2] W. Maret, H.H. Sandstead, Zinc requirements and the risks andbenefits of zinc supplementation, J. Trace Elem. Med. Biol. 20(2006) 3–18.
[3] C. Murgia, C.J. Lang, A.Q. Truong-Tran, D. Grosser, L. Jayaram,R.E. Ruffin, G. Perozzi, P.D. Zalewski, Zinc and its specific trans-porters as potential targets in airway disease, Curr. Drug Targets7 (2006) 607–627.
[4] I.E. Dreosti, Recommended dietary intakes of iron, zinc, and otherinorganic nutrients and their chemical form and bioavailability,Nutrition 9 (1993) 542–545.
[5] J.E. Arsenault, K.H. Brown, Zinc intake of US preschool chil-dren exceeds new dietary reference intakes, Am. J. Clin. Nutr. 78(2003) 1011–1017.
[6] G. Danscher, The autometallographic zinc-sulphide method. Anew approach involving in vivo creation of nanometer-sized zincsulphide crystal lattices in zinc-enriched synaptic and secretoryvesicles, Histochem. J. 28 (1996) 361–373.
[7] P. Coyle, P.D. Zalewski, J.C. Philcox, I.J. Forbes, A.D. Ward,S.F. Lincoln, I. Mahadevan, A.M. Rofe, Measurement of zinc inhepatocytes by using a fluorescent probe, zinquin: relationshipto metallothionein and intracellular zinc, Biochem. J. 303 (Pt 3)(1994) 781–786.
[8] P.D. Zalewski, I.J. Forbes, W.H. Betts, Correlation of apoptosiswith change in intracellular labile Zn(II) using zinquin [(2-methyl-8-p-toluenesulphonamido-6-quinolyloxy)acetic acid], anew specific fluorescent probe for Zn(II), Biochem. J. 296 (Pt
2) (1993) 403–408.[9] P.D. Zalewski, A.Q. Truong-Tran, D. Grosser, L. Jayaram, C.Murgia, R.E. Ruffin, Zinc metabolism in airway epithelium andairway inflammation: basic mechanisms and clinical targets. Areview, Pharmacol. Ther. 105 (2005) 127–149.

tion Re
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
C. Devirgiliis et al. / Muta
10] A.Q. Truong-Tran, R.E. Ruffin, P.D. Zalewski, Visualization oflabile zinc and its role in apoptosis of primary airway epithelialcells and cell lines, Am. J. Physiol. 279 (2000) L1172–L1183.
11] C. Frederickson, Imaging zinc: old and new tools, Sci. STKE2003 (2003) pe18.
12] D.J. Eide, Zinc transporters and the cellular trafficking of zinc,Biochim. Biophys. Acta 1763 (2006) 711–722.
13] R.J. Cousins, J.P. Liuzzi, L.A. Lichten, Mammalian zinc trans-port, trafficking, and signals, J. Biol. Chem. 281 (2006)24085–24089.
14] M. Vasak, D.W. Hasler, Metallothioneins: new functional andstructural insights, Curr. Opin. Chem. Biol. 4 (2000) 177–183.
15] P. Lichtlen, W. Schaffner, The “metal transcription factor” MTF-1: biological facts and medical implications, Swiss Med. Wkly.131 (2001) 647–652.
16] R. Giacconi, C. Cipriano, E. Muti, L. Costarelli, C. Maurizio,V. Saba, N. Gasparini, M. Malavolta, E. Mocchegiani, Novel-209A/G MT2A polymorphism in old patients with type 2 diabetesand atherosclerosis: relationship with inflammation (IL-6) andzinc, Biogerontology 6 (2005) 407–413.
17] Y. Tamura, M. Maruyama, Y. Mishima, H. Fujisawa, M. Obata, Y.Kodama, Y. Yoshikai, Y. Aoyagi, O. Niwa, W. Schaffner, R. Kom-inami, Predisposition to mouse thymic lymphomas in responseto ionizing radiation depends on variant alleles encoding metal-responsive transcription factor-1 (Mtf-1), Oncogene 24 (2005)399–406.
18] D.J. Eide, The SLC39 family of metal ion transporters, PflugersArch. 447 (2004) 796–800 (Epub 2003 May 2014).
19] R.D. Palmiter, L. Huang, Efflux and compartmentalization of zincby members of the SLC30 family of solute carriers, Pflugers Arch.447 (2004) 744–751 (Epub 2003 May 2014).
20] J.P. Liuzzi, R.J. Cousins, Mammalian zinc transporters, Annu.Rev. Nutr. 24 (2004) 151–172.
21] K. Ishihara, T. Yamazaki, Y. Ishida, T. Suzuki, K. Oda, M.Nagao, Y. Yamaguchi-Iwai, T. Kambe, Zinc transport complexescontribute to the homeostatic maintenance of secretory path-way function in vertebrate cells, J. Biol. Chem. 281 (2006)17743–17750.
22] P. Van Eerdewegh, R.D. Little, J. Dupuis, R.G. Del Mastro, K.Falls, J. Simon, D. Torrey, S. Pandit, J. McKenny, K. Braun-schweiger, A. Walsh, Z. Liu, B. Hayward, C. Folz, S.P. Manning,A. Bawa, L. Saracino, M. Thackston, Y. Benchekroun, N. Cap-parell, M. Wang, R. Adair, Y. Feng, J. Dubois, M.G. FitzGerald,H. Huang, R. Gibson, K.M. Allen, A. Pedan, M.R. Danzig, S.P.Umland, R.W. Egan, F.M. Cuss, S. Rorke, J.B. Clough, J.W. Hol-loway, S.T. Holgate, T.P. Keith, Association of the ADAM33gene with asthma and bronchial hyperresponsiveness, Nature 418(2002) 426–430.
23] L. Huang, J. Gitschier, A novel gene involved in zinc transport isdeficient in the lethal milk mouse, Nat. Genet. 17 (1997) 292–297.
24] M.L. Ackland, L. Michalczyk, Zinc deficiency and its inheriteddisorders—a review, Genes Nutr. 1 (2006) 41–50.
25] W. Chowanadisai, B. Lonnerdal, S.L. Kelleher, Identification ofa mutation in SLC30A2 (ZnT-2) in women with low milk zincconcentration that results in transient neonatal zinc deficiency, J.Biol. Chem. 281 (2006) 39699–39707.
26] D.L. Duffy, N.G. Martin, D. Battistutta, J.L. Hopper, J.D. Math-
ews, Genetics of asthma and hay fever in Australian twins, Am.Rev. Respir. Dis. 142 (1990) 1351–1358.27] D.A. de Luis, O. Izaola, R. Aller, A. Armentia, L. Cuellar, Antiox-idant and fat intake in patients with polinic asthma, Med. Clin.(Barc) 121 (2003) 653–654.
[
search 622 (2007) 84–93 91
28] A. Soutar, A. Seaton, K. Brown, Bronchial reactivity and dietaryantioxidants, Thorax 52 (1997) 166–170.
29] G. Devereux, S.W. Turner, L.C. Craig, G. McNeill, S. Martin-dale, P.J. Harbour, P.J. Helms, A. Seaton, Low maternal vitaminE intake during pregnancy is associated with asthma in 5-year-oldchildren, Am. J. Respir. Crit. Care Med. 174 (2006) 499–507.
30] F. Tahan, C. Karakukcu, Zinc status in infantile wheezing, Pediatr.Pulmonol. 41 (2006) 630–634.
31] A.Q. Truong-Tran, R.E. Ruffin, P.S. Foster, A.M. Koskinen, P.Coyle, J.C. Philcox, A.M. Rofe, P.D. Zalewski, Altered zinchomeostasis and caspase-3 activity in murine allergic airwayinflammation, Am. J. Respir. Cell Mol. Biol. 27 (2002) 286–296.
32] J.E. Carter, A.Q. Truong-Tran, D. Grosser, L. Ho, R.E. Ruffin,P.D. Zalewski, Involvement of redox events in caspase activationin zinc-depleted airway epithelial cells. Visualization of labilezinc and its role in apoptosis of primary airway epithelial cellsand cell lines, Biochem. Biophys. Res. Commun. 297 (2002)1062–1070.
33] S. Bao, D.L. Knoell, Zinc modulates airway epithelium suscepti-bility to death receptor-mediated apoptosis, Am. J. Physiol. LungCell Mol. Physiol. 290 (2006) L433–L441.
34] S.L. Bao, D.L. Knoell, Zinc modulates cytokine-induced lungepithelial cell barrier permeability, Am. J. Physiol. Lung CellMol. Physiol. (2006).
35] P.S. Foster, S.P. Hogan, A.J. Ramsay, K.I. Matthaei, I.G. Young,Interleukin 5 deficiency abolishes eosinophilia, airways hyperre-activity, and lung damage in a mouse asthma model, J. Exp. Med.183 (1996) 195–201.
36] K. Inoue, H. Takano, R. Yanagisawa, M. Sakurai, T. Ichinose,K. Sadakane, K. Hiyoshi, M. Sato, A. Shimada, M. Inoue, T.Yoshikawa, Role of metallothionein in antigen-related airwayinflammation, Exp. Biol. Med. (Maywood) 230 (2005) 75–81.
37] C.J. Lang, C. Murgia, M. Leong, L.W. Tan, G. Perozzi, D. Knight,R.E. Ruffin, P.D. Zalewski, Anti-inflammatory effects of zinc andalterations in zinc transporter mRNA in mouse models of allergicinflammation, Am. J. Physiol. Lung Cell Mol. Physiol. (2006).
38] S.T. Holgate, D.E. Davies, R.M. Powell, J.W. Holloway,ADAM33: a newly identified protease involved in airway remod-elling, Pulm. Pharmacol. Ther. 19 (2006) 3–11.
39] A.B. Chausmer, Zinc, insulin and diabetes, J. Am. Coll. Nutr. 17(1998) 109–115.
40] C.G. Taylor, Zinc, the pancreas, and diabetes: insights from rodentstudies and future directions, Biometals 18 (2005) 305–312.
41] L.H. Kristiansen, J. Rungby, L.G. Sondergaard, M. Stoltenberg,G. Danscher, Autometallography allows ultrastructural monitor-ing of zinc in the endocrine pancreas, Histochem. Cell Biol. 115(2001) 125–129.
42] P.D. Zalewski, S.H. Millard, I.J. Forbes, O. Kapaniris, A.Slavotinek, W.H. Betts, A.D. Ward, S.F. Lincoln, I. Mahade-van, Video image analysis of labile zinc in viable pancreatic isletcells using a specific fluorescent probe for zinc, J. Histochem.Cytochem. 42 (1994) 877–884.
43] H. Ishihara, P. Maechler, A. Gjinovci, P.L. Herrera, C.B. Woll-heim, Islet beta-cell secretion determines glucagon release fromneighbouring alpha-cells, Nat. Cell Biol. 5 (2003) 330–335.
44] I. Franklin, J. Gromada, A. Gjinovci, S. Theander, C.B. Wollheim,Beta-cell secretory products activate alpha-cell ATP-dependent
potassium channels to inhibit glucagon release, Diabetes 54(2005) 1808–1815.45] M.A. Ravier, G.A. Rutter, Glucose or insulin, but not zinc ions,inhibit glucagon secretion from mouse pancreatic alpha-cells,Diabetes 54 (2005) 1789–1797.

tion Re
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
92 C. Devirgiliis et al. / Muta
46] B.J. Kim, Y.H. Kim, S. Kim, J.W. Kim, J.Y. Koh, S.H. Oh, M.K.Lee, K.W. Kim, M.S. Lee, Zinc as a paracrine effector in pancre-atic islet cell death, Diabetes 49 (2000) 367–372.
47] E. Ho, N. Quan, Y.H. Tsai, W. Lai, T.M. Bray, Dietary zinc sup-plementation inhibits NFkappaB activation and protects againstchemically induced diabetes in CD1 mice, Exp. Biol. Med. (May-wood) 226 (2001) 103–111.
48] C. Terres-Martos, M. Navarro-Alarcon, F. Martin-Lagos,G.d.l.S.H. Lopez, V. Perez-Valero, M.C. Lopez-Martinez, Serumzinc and copper concentrations and Cu/Zn ratios in patients withhepatopathies or diabetes, J. Trace Elem. Med. Biol. 12 (1998)44–49.
49] H. Haase, W. Maret, Fluctuations of cellular, available zincmodulate insulin signaling via inhibition of protein tyrosine phos-phatases, J. Trace Elem. Med. Biol. 19 (2005) 37–42.
50] H. Haase, W. Maret, Protein tyrosine phosphatases as targets of thecombined insulinomimetic effects of zinc and oxidants, Biometals18 (2005) 333–338.
51] J.P. Liuzzi, J.A. Bobo, L.A. Lichten, D.A. Samuelson, R.J.Cousins, Responsive transporter genes within the murineintestinal-pancreatic axis form a basis of zinc homeostasis, Proc.Natl. Acad. Sci. U.S.A. 101 (2004) 14355–14360.
52] L.G. Sondergaard, B. Brock, M. Stoltenberg, A. Flyvbjerg, O.Schmitz, K. Smidt, G. Danscher, J. Rungby, Zinc fluxes duringacute and chronic exposure of INS-1E cells to increasing glucoselevels, Horm. Metab. Res. 37 (2005) 133–139.
53] M. Seve, F. Chimienti, S. Devergnas, A. Favier, In silico iden-tification and expression of SLC30 family genes: an expressedsequence tag data mining strategy for the characterization of zinctransporters’ tissue expression, BMC Genomics 5 (2004) 32.
54] F. Chimienti, S. Devergnas, A. Favier, M. Seve, Identification andcloning of a beta-cell-specific zinc transporter, ZnT-8, localizedinto insulin secretory granules, Diabetes 53 (2004) 2330–2337.
55] D. Knaack, D.M. Fiore, M. Surana, M. Leiser, M. Laurance,D. Fusco-DeMane, O.D. Hegre, N. Fleischer, S. Efrat, Clonalinsulinoma cell line that stably maintains correct glucose respon-siveness, Diabetes 43 (1994) 1413–1417.
56] A.C. Powers, S. Efrat, S. Mojsov, D. Spector, J.F. Habener, D.Hanahan, Proglucagon processing similar to normal islets in pan-creatic alpha-like cell line derived from transgenic mouse tumor,Diabetes 39 (1990) 406–414.
57] F. Chimienti, S. Devergnas, F. Pattou, F. Schuit, R. Garcia-Cuenca,B. Vandewalle, J. Kerr- Conte, L. Van Lommel, D. Grunwald, A.Favier, M. Seve, In vivo expression and functional characteri-zation of the zinc transporter ZnT8 in glucose-induced insulinsecretion, J. Cell Sci. (2006).
58] T. Kambe, H. Narita, Y. Yamaguchi-Iwai, J. Hirose, T. Amano,N. Sugiura, R. Sasaki, K. Mori, T. Iwanaga, M. Nagao, Cloningand characterization of a novel mammalian zinc transporter, zinctransporter 5, abundantly expressed in pancreatic beta cells, J.Biol. Chem. 277 (2002) 19049–19055.
59] T. Suzuki, K. Ishihara, H. Migaki, M. Nagao, Y. Yamaguchi-Iwai, T. Kambe, Two different zinc transport complexes of cationdiffusion facilitator proteins localized in the secretory pathwayoperate to activate alkaline phosphatases in vertebrate cells, J.Biol. Chem. 280 (2005) 30956–30962.
60] R.C. De Lisle, M.P. Sarras Jr., J. Hidalgo, G.K. Andrews, Met-
allothionein is a component of exocrine pancreas secretion:implications for zinc homeostasis, Am. J. Physiol. 271 (1996)C1103–C1110.61] J. Wang, Y. Song, L. Elsherif, Z. Song, G. Zhou, S.D. Prabhu,J.T. Saari, L. Cai, Cardiac metallothionein induction plays the
[
search 622 (2007) 84–93
major role in the prevention of diabetic cardiomyopathy by zincsupplementation, Circulation 113 (2006) 544–554.
62] N. Tonder, F.F. Johansen, C.J. Frederickson, J. Zimmer, N.H.Diemer, Possible role of zinc in the selective degeneration of den-tate hilar neurons after cerebral ischemia in the adult rat, Neurosci.Lett. 109 (1990) 247–252.
63] K. Hsiao, P. Chapman, S. Nilsen, C. Eckman, Y. Harigaya, S.Younkin, F. Yang, G. Cole, Correlative memory deficits, Abetaelevation, and amyloid plaques in transgenic mice, Science 274(1996) 99–102.
64] R.A. Cherny, C.S. Atwood, M.E. Xilinas, D.N. Gray, W.D. Jones,C.A. McLean, K.J. Barnham, I. Volitakis, F.W. Fraser, Y. Kim,X. Huang, L.E. Goldstein, R.D. Moir, J.T. Lim, K. Beyreuther,H. Zheng, R.E. Tanzi, C.L. Masters, A.I. Bush, Treatment with acopper-zinc chelator markedly and rapidly inhibits beta-amyloidaccumulation in Alzheimer’s disease transgenic mice, Neuron 30(2001) 665–676.
65] C.W. Ritchie, A.I. Bush, A. Mackinnon, S. Macfarlane, M. Mas-twyk, L. MacGregor, L. Kiers, R. Cherny, Q.X. Li, A. Tammer,D. Carrington, C. Mavros, I. Volitakis, M. Xilinas, D. Ames, S.Davis, K. Beyreuther, R.E. Tanzi, C.L. Masters, Metal-proteinattenuation with iodochlorhydroxyquin (clioquinol) targetingAbeta amyloid deposition and toxicity in Alzheimer disease: apilot phase 2 clinical trial, Arch. Neurol. 60 (2003) 1685–1691.
66] C.J. Frederickson, J.Y. Koh, A.I. Bush, The neurobiology of zincin health and disease, Nat. Rev. Neurosci. 6 (2005) 449–462.
67] J.Y. Koh, S.W. Suh, B.J. Gwag, Y.Y. He, C.Y. Hsu, D.W. Choi,The role of zinc in selective neuronal death after transient globalcerebral ischemia, Science 272 (1996) 1013–1016.
68] W. Chowanadisai, S.L. Kelleher, B. Lonnerdal, Zinc deficiency isassociated with increased brain zinc import and LIV-1 expressionand decreased ZnT-1 expression in neonatal rats, J. Nutr. 135(2005) 1002–1007.
69] R.D. Palmiter, T.B. Cole, C.J. Quaife, S.D. Findley, ZnT-3, a puta-tive transporter of zinc into synaptic vesicles, Proc. Natl. Acad.Sci. U.S.A. 93 (1996) 14934–14939.
70] T.B. Cole, A. Martyanova, R.D. Palmiter, Removing zinc fromsynaptic vesicles does not impair spatial learning, memory, or sen-sorimotor functions in the mouse, Brain Res. 891 (2001) 253–265.
71] J.Y. Lee, T.B. Cole, R.D. Palmiter, S.W. Suh, J.Y. Koh, Contribu-tion by synaptic zinc to the gender-disparate plaque formation inhuman Swedish mutant APP transgenic mice, Proc. Natl. Acad.Sci. U.S.A. 99 (2002) 7705–7710.
72] J.Y. Lee, J.H. Kim, S.H. Hong, R.A. Cherny, A.I. Bush, R.D.Palmiter, J.Y. Koh, Estrogen decreases zinc transporter 3 expres-sion and synaptic vesicle zinc levels in mouse brain, J. Biol. Chem.279 (2004) 8602–8607.
73] R.A. Colvin, C.P. Fontaine, M. Laskowski, D. Thomas, Zn2+
transporters and Zn2+ homeostasis in neurons, Eur. J. Pharmacol.479 (2003) 171–185.
74] Z.H. Chi, X. Wang, Z.Y. Wang, H.L. Gao, A. Dahlstrom, L.Huang, Zinc transporter 7 is located in the cis-Golgi appara-tus of mouse choroid epithelial cells, Neuroreport 17 (2006)1807–1811.
75] M.A. Lovell, J.L. Smith, S. Xiong, W.R. Markesbery, Alterationsin zinc transporter protein-1 (ZnT-1) in the brain of subjects withmild cognitive impairment, early, and late-stage Alzheimer’s dis-
ease, Neurotox. Res. 7 (2005) 265–271.76] J.L. Smith, S. Xiong, W.R. Markesbery, M.A. Lovell, Alteredexpression of zinc transporters-4 and -6 in mild cognitive impair-ment, early and late Alzheimer’s disease brain, Neuroscience 140(2006) 879–888.

tion Re
[
[
[
[
[
C. Devirgiliis et al. / Muta
77] M.A. Lovell, J.L. Smith, W.R. Markesbery, S. Xiong, Elevatedzinc transporter-6 in mild cognitive impairment, Alzheimer dis-ease, and pick disease Altered expression of zinc transporters-4and -6 in mild cognitive impairment, early and late Alzheimer’sdisease brain, J. Neuropathol. Exp. Neurol. 65 (2006) 489–498.
78] R.D. Palmiter, The elusive functions of metallothionenins, Proc.Natl. Acad. Sci. U.S.A. 95 (1998) 8230–8248.
79] H. Tapiero, K.D. Tew, Trace elements in human physiology andpathology: zinc and metallothioneins, Biomed. Pharmacother. 57(2003) 399–411.
search 622 (2007) 84–93 93
80] A.J. Brookes, J.A. Prince, Genetic association analysis: lessonsfrom the study of Alzheimers disease, Mutat. Res. 573 (2005)152–159.
81] R. Sladek, G. Rocheleau1, J. Rung, C. Dina, L. Shen, D. Serre,P. Boutin, D. Vincent, A. Belisle, S. Hadjadj, B. Balkau, B.
Heude, G. Charpentier, T.J. Hudson, A. Montpetit, A.V. Pshezhet-sky10, M. Prentki, B.I. Posner, D.J. Balding, D. Meyre, C.Polychronakos, P. Froguel, A genome-wide association studyidentifies novel risk loci for type 2 diabetes, Nature 445 (2007)881–885.




























