Embed Size (px)
Citation preview

Role de la Nucleophosmine (NPM1) dans la
physiopathologie prostatique
Rafik Boudra
To cite this version:
Rafik Boudra. Role de la Nucleophosmine (NPM1) dans la physiopathologie prostatique. Can-cer. Universite Blaise Pascal - Clermont-Ferrand II, 2015. Francais. <NNT : 2015CLF22598>.<tel-01379611>
HAL Id: tel-01379611
https://tel.archives-ouvertes.fr/tel-01379611
Submitted on 11 Oct 2016
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinee au depot et a la diffusion de documentsscientifiques de niveau recherche, publies ou non,emanant des etablissements d’enseignement et derecherche francais ou etrangers, des laboratoirespublics ou prives.

UNIVERSITE BLAISE PASCAL UNIVERSITE D’AUVERGNE N° D.U. 2598 Année 2015
Ecole Doctorale des Sciences de la Vie,
Santé, Agronomie, Environnement. N° d'Ordre 673
THESE
Présentée à l’Université Blaise Pascal pour l’obtention du grade de
DOCTEUR D’UNIVERSITE
(Spécialité : Physiologie et Génétique Moléculaires)
Soutenue publiquement le 25 septembre 2015
Rafik Boudra
Rôle de la Nucléophosmine (NPM1) dans la physiopathologie prostatique Président Pr. Pierre Verrelle, EA7283, Université d’Auvergne, Clermont-Ferrand Rapporteurs Dr. Jocelyn Céraline, INSERM U1113, Université de Strasbourg Dr Carmen Garrido, INSERM U866, Université de Bourgogne Examinateurs
Dr Laurent Le Cam, IRCM, INSERM U1194, Université de Montpellier
Dr Serge Manié, CRCL, CNRS UMR 5286, INSERM U1052 Université de Lyon
Directeur de thèse Pr Claude Beaudoin, GReD, CNRS UMR 6293, INSERM U1103,
Clermont-Ferrand
Laboratoire de Génétique, Reproduction et Développement (GReD) UMR CNRS 6293, Clermont Université, INSERM U1103

�

������������
�
���������������� �
�
� ��� ��������� ����� �������� ��� ������� �������� ��������� ��� �������
������ ������� ��� ������� ������� ��� ���� ������ ���� ��� ������� ����� �����
�!�"�������#����!�"�����������"��������$%��&�
�
���� �������� �� �� ����� �������������� ��������� ��� ������� �� ��������������
�������� '�� �� ���� ����� ��� ������� "����� � (���&� ��� �$����� �� ���� ����� ��� �����
�!��(��$�������������"��������������"��������������"������(�������#����)� ����
����!�"�������������������%��������������� *��������!�"���#������!���������
�������$%��&� ����#���������������������#�������(��������������������+����
#����#��������������(������������������������,����-�����������������������(���
���#���������������&� �!���(������#��##��� ������"���������"���������� .!��#%������
����������������������#%��������#��&�
�
�������� ��� ����� #����� ��� ���� #��������� ����� #�� ���� ������ #����������
�"���������������������������"�������.������������������� ��������������������
�������%��������������&�/���������������������������#�������������������������
����������������.!���#���"����"��������������.�����������#�����&� ����#��������
����(������������������������������(������������#����������0���1������&�
�
�������������#��� ������#���(��������� ����������������������������#��������
������(���������"�����������������������������&� ����#���������������������
�������������������������!��������#�����+&�
�
�������� ����� #��� ���� ����� ��� �!$����� ��� #��������� ���� ����� �� #����� ���
#�����������#��#���������������������������������������&� ��������������#���
������������������������#�.������#������������������������������������"����
����#�#����������#�.��&�2������!����(������#���������������������������
������+&�1��"��������������������������(������%������������ .�����������#����
����������"����#����������������%���#�������&�
�
��(���������3����� .��"��������������"����������������� �!����#������������� ����
#������4���������������4����$������������!�����������������#���������������#�&��
�
��� "������ ���������� ������� ����� ���� ���������� #������ #���� ��� ������
�������#���������!����#�&�5������� ����������������#�����������(��������(�����
�"���"�����������"���#�����&�
�
6�� ����� ����� � ���� ���#������� ��� ��(��� 1��������� ��� ����7����&� �����
������������ ��� "���� ��"�������� ������� ��� "��� ��� ��(�� (���� #���� ������&� ��� ���
����������#������"���� �����������#���������#��!���� ��88��"�������#����#���
����"���������#����8�#�����#&��
�

�

�
���������������������
���"������������ .�����"��������������.����������8�����9��#�������������"����
�"�8� ��� #��� �9����� ������� .�� ��� $������� � ���� ���� ���$������� ���
�����#������� ����� ���������� #��� ���� #�����+� ���#�� ��� ������ ����������
�9����������������������������"���������������������������#������������(����
��������#������$��������(������������:���������#������������������"����"����
���������;�������9������������.�#���������"���
�
�
�������� ���������������������������� ����� � �9������ ��� �������� ��� ����� #����� ��� ���� #��������� ���� �9����
������ #�� ���� #��������� ��� ���� #������� #��� ��� ��������� 1�� ��(�������� ���
�9�������������������#������$������������������$$$�������#���������������<�
������(�����$�������������������
�
�� �����������������#�����9������#����������;�����"�����9���(������#��##����������
��������������������������� �9�����#���"���� ��������������� ����������� ��������
�������������(�(����������"��+��
�
���������������������#����9�"���(�������������+��������.9��������������#���
��� ���������� ���#���� ������ ��� ��"���� ��������������� �� ���.���� ���� ���
#������ ��� ��� �=�������;��� ��� .�� ����� ������0�� ���������+� ���� ���������� ����
�������
�
����������#������(�����$�����#������������#������������"��������9��(��
�����������������#���������������0����
�
1����������������.��"������������9�"������.�������������+��#�#����������
#�.�������#����������������9�"����"���"��������������������������������������
����$�����������
�
��"������������������"���������##��������������������6������������������
��������������������������>>$?@��"�����������
�
��(����������(�����9����������#�������������������������������������#$�������
��������#��������������>>@�0�����������;���<�����9�+������������������(�������
��������##����������������>�����#�����������.9�"���<���������������9��������
#��� ��������� ��� (���� ��+���������������� ��� ������ �� ���� #���� ����(��� ��� .9���
(������#� �##����� ������� ���� ������������� ���� #������ ������� ��� ���� "���� ���
���������
�
1�$��������9��������9�����������������������������"�����������������#����(���
�������.9������#;������$���9����������������������9�"������.��������������������
�9�"������������(����#�������������"�����(������
�

�

�������/������� �����������#���"�������������������#�������������"�������
����(�� ���� ����� �� ���������� ��������� ��� "��� #��� ���� ��#����������� ����
�������������������������������
�
������������������������������#���������#��������#�������(�����$������������
������ #������������� ��� �������� ���� ����� �� ������� ������ ���� �������� ��� "����
���$���������������������"����"���#�������������#���������������
�
��� ������� ������ �������(��� ��� #=��� ��� ��$��$�� ��� ���� �������� #��� ����
��������#�����������(�����
�
����������������#������������.����#���=���������������A���������������
�
��������������������������"������� ��������#������ ���"��������������"����"�����������
(������#� #���� ������� ����� � ���� �����#���� �##���� ��� ���� ������� ��� ������ �
��"����� 1�� ���� ��� �=��� #�������� ��� ��� ���.���� ����� #��"�� ������ ������
������������� .��"������#����� �����������#��#��������� ������"��� ��������
#������ ��� �(�������������� ����� ���� ��������� ���(��� ����� �"��"� �
������ ��� ����� ��"�� ��� ���"���� #����� .���� ���.���� #��� ��� ��� ����� #������ �
#������������������"���������������"���������(����=���,�����0��
�
������� �0�� ��� �����$��� �� /�� "����� ������ ����� #��������� ���(��� #��� ���
�������� #�������������� $��� ��� �������� 6�� #��� ���"����� .�� ����� ���"�����
��������������.������������#��������#������������#����������������������$�������
"�������A$��A��B�B��,���$�����.�������������������������������� ����#������������
�������� ��� (����� $����� ���� ��� ��.������� ����� ����� ���������� ��� �$������
��#�������������������:��������������������#��.�������"���������������������������
����������
�
������ ��� ����� ���� ���� ����� (���� �$����� (��� ���������� ����� ������ ������ �$�����
#������� ������ �$���� ���� #��� ����� ��� �������� ���� ��� ����� �##����� ��� ���
����������������������#����������#����(�����������.��������.����$�����"�������
#��������������#�������#������������������"�����������(����� ���������;����
��#����(���������������#�������������������������������#����"������������
�������� ������ ���#��� ����� ���,�� ���$�� ���� � ���"����� .�� ����� �:� ���� ����� �������
#�������������
�
������ ��� ��������� � ����� ��� (����� ��� .���#�� ���� ��� ������ #��� #�� ���
���;���������� ������ #��� "�� ��� ���"���� ���� ������� ������� � ��������� ����
������������� ����� ��� ����� ,���� ��� ��� ���� "��� ���� .�� ��� ����� #��� ��� ������� ���
����#$������������ �����������#������������(�������� ���.�������� �������(�����
�;��� ������ ��� �������� ��� ����������� ��� ������� ���� ������ ���� ��#���� ���
������������� ��� ���#��� #��"������ ������� ��� ��������� #��� ��� #���������
����������������#��������$���������������������#������������������������
�

�

�����������C������������������� �� ����������� ��������������������-����1����
�����������-��������������������������������������������������� �����
��� ���� ������ ������������ ��� ��� ��������� ��� ������������� ���� ������� ���
������� ��� ��� �������� ������ ��� ���������� ��� ���� �� ����� ��� ������ �������
��� ��� ������� �� ��� ���� ������ ��� D ��� ��������� ��� DE��� ��� DF��� ��� ���
������������������������������������������������������!������������!�
���������"�#�����������������������������������������������$�������������"�
�
#��%����������������������������������������&'(����������������������')*��
����������������������������� ����������������������������������!���"�
+������ ������ ������������ �������,�� �������������������� �����,���� ���
������������������-.�)�
�
��������������������������� ����"""�������������� ������������� �����
�����������������)��
�
/� �� ������ ����� ��� '� �������������'�� ��� ����������� ������ ������� �������
'������')�
�
������������ ���������� ������ �������������� ���,�� ���� �������� ������������
����� �������������)�0�����������1������,�����)�
�
�
����������������� �
+���+�� ��������������� ��������������� ��� ���������� ������� ���#�������)� +��
�� ����������������������������������������� �����������������������,�����
����������"�+��������,�������������������������������������������,������
���������������������������������������)�
�
2�������%��'���-�����3�����������2���� ��'"�1����,������,���������'��������
������'�����1���������������� ����������������������������������������������
�� )� 4�� ����� ,�� ������������ ��,�� ����� ��� ��������� ���������,��"�
����,�� ������ ,�� ���� ��������)� +�������� ,�� �� ��������� ��� ������
��������������$���������,���������������������������������������"""�������"�
�
5�������������������������-���%�������������������������������������)�
4����������������� ��������� ������������)�6����������� ��� ���������
��������������������������������������"""��
�
#��������������������������������������������������������� ���������,�������
������� ��� ������� �������� ������ ,��,�� �����)� +�� ���� ����� ������� ���
������������������� ��� ��������������������� ��������������� ���������"�0��
�����������������#�%��������������7��������1������,�"�
�
#���������#������ ������8������9��������� ������ E�������)�#������������
����������������������������� ����������������������� ������,����� ������������
������� ������� ������ �� -���%�)� ������� �� ���� �������� ��� ���� ������� ���
��:���������� �����)�

�

�
������������������ ���������������� ������������� ���������������������������������� ���
�������1���������������� ������������������ ����������������������� ���������
����� ����������������������� ���������� ������ � ��� ������������� ���������
1��������������������������������� ������������������������� ����!�������
������������������ ��������������� �� ��������������������������"�����������
������������#��$������������������������������% �����%��������������������
���������������������������������"�������������������&�����������
�������������������'������ �������������� �����������'������������ ���(�� �����
��������'��������������������������'�������������������������)������������������
�������������������������������������)����������������*�
�
�
����������������������������(����������������������'���������������������������"��������
����������������������+������&�������������� �����������(���'��������(��
�� ��������������'������������� ���������������(���+������������ ����������
����������(��(������(�����������������������(�� �'������ ������,���� ������
+� ����� ������ �����(��� �� ������ ��� ������ �� �'�������� �'������� +� �����
�������'-������������������������� �� ��������'������ ���������.�
�������� ���������������������.��������&����������'"��*�/��� ���������
�'����'� �����������������������������
�
��� ������� �� ����� ���"���� �� ��������� ���� ���� ������ �� �� �������� �����
����"�������������������������������0���������������������-�������������
�'���������*�
�
1���� ����������� ���������'�������'���������(�'��������� ������ ����� �������
����������������������������������������� ������/������������+������
������������1������!������0����������������'��������-�(�������������� ���������
������������������������������������ ����������������������*�
�
�
������������
1'�������������������������������'�������� ������������������������������
����� ������ ����� ��� �������� 1��� �������� ��� ������ ������� ��� ����������
��������������������������������������������������������������,���� ���
����������/����������������������������������*�1������������"����������������
����,������������������������������������(�� �'������������ ������������
����������������������������������������2*�
�
�
�
�
�

�

�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
� � � � � �����������

�

�������
��������� ����������������� ��������������������������������� ������������ ����
������ ����� � ����������� !���� ���������"�����������#� ��� ���"���������������$���
��� �%���� ����������&� '�� ������ � � ���� � ���������� ���� ��� ��� ��� ��� ����� ��� �
��(�� � �%�� � ��� ������ � ���� � ��������� �� � ������ � ��� ��� �� ����!� ��� �� �$��� ��)
����*����*� ��� � �� � ��"�� � ���������� � ������� � �#��"��� � (���� � � �� ����� �����&� ���
�����+���������������������(���� #� ���������,�(������ ��������������"��������������
��� � �#��������� �� �������� ��� ������ ���� ����!� �� � �(� � "��� ��� ��+��� ��� ��� �
���� "����� ����� ��� ������������� ���*������������ ��#����������������� ����&�-� �
������ � �� ������� ���� �%������ ��� �� �������� � ���� ,� ���� ��"��������� ��� �#���� �
����*����*�����#��������&�� �� ������� ��(+������������������������(���������� ������
�� � ������� � ��������� � ��**������ � ��� ��"������ �#� ��� ��� ��� "+�� � �� � ��� ���
�"������� ��� �%���� ����������!� ����� ��� -%������ .� �� ��/0���&� ����� ���� �� � ��� � ���
�(�������� �� � ��� � �� � ���� ���� !� �� � ���� � �""+����� ���� ����� ���������� ,� ���
������"��+ �� �� �������� ��� � ������� �(��� �#����� � � �� � ��"����� &� ��� ������
������� ��� ���(���� (� ���� ,� ���������� ��� ������� �� � ����� �� � ���� ��������� ���
��� ��� ��� ��� ����� ��� � �� � ������ � �� ������� &� '� � ���� � ������ � ��� ���
������������������������������� �������������������0��� ���123������(���������������
�� ���������������� ��� � *�� ���� ��� &��������������� #���� � ������ ����� *���������
�������123��������!��� ��(� ������������#�����+������*������ �� �����%������ ����
��� � ��(���� � ���� ��� ����� ���� ��� ������� �1.�� ���� �#������(����� �+��� ,� ����
�%�������(��� ��� �123&� '�� � ��� ���� ��!� �� � ��� � �#43��� ��� ��� ������ � ����� ���
��"���� �������������� �������� � ��(�"� &�� �� ����� ���������"�������������123�
����$����#� ��� ����������������� ��*� ���� ��� ���������������� ����������#� ��� ���
���"+������ ������� ������ ���� �#43����������&��� ��(� ���*���� ��� �������������������
�123� ��� ����� "�5��� ,� ����� ��+��� ��� ��� � ��(����� � ���� �1.�� ��� � �#���������
�� �������&� .�*��!� �� � �(� � ����� ���� �#� ��� ��� ��� ����� � �� ��� ����� ����
���� ������ �� � �**�� � ����*����* � ��� ��� (��� �6�7�471��123&� -� � ���� � �������� ����
����� ����� ��(��� �**������� ��� �(��� ��� ������ (��� ��� �"���� ����!� *�� ���� ��� ������
������� ���� ����������� ������ ������������� ��� � �� � ������ � �� ������� ���� ������ ���
�1.�&�

�

������
���������� � 8����9����� �� �� ����� ���������� ������ �� � ������� � � �� ������ ������ ���
����������������� � ����� �� ��� ��������� ���� �� ��������� � � �� ��� �� �������� ��� �����
������� �������� ������ ���� ��������� ��� ����������� � ��� ����� � � ����� ������ �����
����������������������� �� � ����� ������������ ������ �� ����������������� �������� ��������
��� ��� ����������� ���� �� ��!��������� ����1�������������������!��"������������� ��
��������� ���� �������������� ��������������� �����������1��������!���� ��������� �!�
��� �� �������������� �������������������� ������������ � � �����������������������
1���������� �������������������������������!���� �� ������������"�#$���������������
� ����� %��� ���������� � ��������� � ��������� ���� ����� ������ �� � � � � ����� ��� ���
&����� �������������������������������������������� ����"�������:�������� ������������
-���� �'������$"�����4������������������ ����������� ������������� �����������������
���� ����� ����������� � ����������� �������� �� ���� �������� � !� � ��������� !���
����� � ���� ��� ��������� �� 1�� ��� �� ����� ��� ��� !��"� ������ ��� � ����� �� �� ���
���� ��� � ������ �� ����� ����������� � � � �������� ������� 1�� ��� �9���� � ��
7� ����1%(�!������ �����! ���� � ���������������������������������� � ���������
������� 6 � ������ ��� ���������)�� �� �� ���� ��� �� "� ��!�� � ����� � �� �1%(�� !�� ���"�
���� ������������ ���� ��� ����� �� ����8�'*���������������1'���� ���������������������
�� ��������� �1%(� ���������� 6 � ��� ������ ����� ������ � � �� �(�4� ������ ���� � �������
������������!����������'*��+���������� ������������1%(��� ����������������� ����
������ ��� � �� �� ������������������ ��� �������� ��������� ��������� �� �� ���� ��
�� ���)� ��������(�4��+�������� ������������ ���� ����� "� ��!�� ������� ���1%(����
����� � ���������������������������1'�������������� � ������������������������*� ������!��
�����! ����������������� ��� �������������������������� �����1'��" ��":�����'*��
1�������������������� �!���! ����������������������6�794719�1%(����!����� ��
������������������� ���� �!����� ������������������������ ���1'�� ����������� ���������
�� �����
�
���� ����������������������-� ��������������� ���������1'����1%(�

�

��������������� ����
3/17���� 3/17�������������'� ����������������� � ����������������������������������@.������������������������ 4������������- ����������������������������������� 4�����'�������������������������������� ��������� 4��!��������������"A�������� 4���������#�������-���������� ������� $�4%�&������� ������$�����'���������������"�������(���������������� 4��������������4��!������������������ 4�������������������� ��� 4��4��������������B)�%'�������� 4��������������!��.���������4*���� �.������������ 4������!�����������������!"#$"#%"&����!�'�(���� ������!���������������!�������������������������� �4�������������������
�������"��������"��������� �������� 4���+�������������������� ���� 4����������� �� ���������� 4*���� �.������������������������������� ��-4��4������������'�����������������)��� ����,�4�������-������� �����&��� ��������������������� ��������������������������������� �����-��������������� �������������!���������������� -�����4��!����'.4��������� - �������4������������������ -�������������������������� -����!����4�������4��������&� ��������������� ��� - ������ �������4�����������������*+ ��� -�.����������������"�/))�����������++� -��������'�������������@@"��B"�//��,�-��� -����'�!�����-������������,�������������-���.��� -������'��������(����,"�@"C��0����� - �������#��������������������������� -'(�#���������������"(�����#� ��������������������� -�����(�����##������2�����2� -��%������A"D"�@"�D������� -'(�#� ��������4��� ���������&���������������� -�����������������-������������ '.4���������'���������������� '� �������������������+����� ������� '��������� ����� ��� '� ������������,����� ����� ���� ��������.��������������%�����������+��� '�����������/������ .,��'�����0�������������

�

�����5��� .,����� ������������������������ .�� �������������������������� .�� ���������������������������������� .����������� ���������� ����� ������������� .��������������������� ������������� ��� ���� .����������������-������6������ ��� ���������� ��������������������������� �� !���� ������������"#�"������������ �������� ������ �$��������#"#%�������� �������������������������� ���������"������� ������������� ����������&��������������������� ���� ���������������'����� '��� ������� (�����&�'����)��(������������������� (� �����4��������� '��� ��������� (� �����'�������� �������� (� �����'�������� ���������� (��������������� �4������ ��&� �������������� (��������� ��������������������� (� �����-���������������������'�'����(��(�������4������� (� ��������������� '��� ���������������� (��������4%#�%#'%�� �� �!"��#� "�8"��� (��������*���������"+#, #�!#+ #- ��$���� (�����4��������������� &� �������*��������������������� &� ����(� ������������ &�(���� ��������%�%&��������� ��.��/�������������� ��#"#%������ ���� ��'�� ������� ���������������������������'����� �����*�����%����()� ��� ����������������0�����$��������(�������� ����� �(����������������������������������'����� 1���� ���� ����'������'&�(���� 1�����/� ��-����������'�������� ������������ ��������4���(��� ������������� ��������� ��� ��'�������������"������� ���*#.�*����� ���������� ���������������������� ��"�*�+���� ������������������'�����������*�+�������� ��$��-�������#"�&����� /����� ����4 ���������������&� ���� /�������4���������������������������&��������� /����������������-�����(�����#"�&������ /����������������-������ ���&,����� /&�4������� ����� ��"4�&��κB��� /�������������*�������&���-��� /�%�(���������&' ��� /�������1����� �������������&����� /��� �� ��� ���� ���/4�'�����&���������� /�������� ����#"#%�& ���� /�����������'���.���� ��+� ��� $��������&� ��� ����� ������

�

�� ����� ��+�--�������������2�(���3��#�����2�4���������� .��-546�3����'�������������� ����� �������-�������� ��� �����-� �4����������������� ����� -� ������������ ������� ���� ����������-�������4�������� � ��� ���������-�'����������������������� ���������������'�����������������PGC1α��� 4�γ�-oactivator 1α������ �������������� �������� ��������������!�!������������� ��������� �������������!�����������;���������� "#�$�%���������%���& ������������������������������������ ���������������������������������� ��!�����������PPARγ��� ��' ���� ���������!����������������%������������ ������4�������(�� ��������������� �������)�������4�������������� ����������������� ��������� ��������������� ��������������*��������������� ���� ����� ����������������� ������ ���� �4��4���������-����������������� ����� �4���������������������!��� ���������������"��� ��4�������������#$����� ��4��� ���������������� ��4!��4�4������� �������������%���� �������+' ���)����������� �,������* ���������������&� �� ���*���-����������� � �����-��������������'���� ��������� ������).�������� ���� )���-�4�������������� )������������������� )!�����������4��������� ������������ ��� )������(������4�������'��������������(#��� )��������������� )�������(����������������������*� ��������������������%���� )���+��)������� ��������� )�������������-������������������������ )�����*�������������4������������*����������������)�%��� )���/��-�����!���(��������* +��� )������0���������,(��� *�������������1������TGFβ��� *������������%��$����������β�TGFβRI/II��� *������������%��$����������β������������������� *��������������(���������������� �������������'+���� *��!����������� ������.���'������� *�����(�����4��� �������� *� ������������4���������(������������� *���������4�������������������(���� ��������� ����� *������)�������-���'�����

�

������ ��������������������� ��� ������������������������������������������������������������ ������������������ ������� ���������������������� ������������� ������ ��� �����!"������������� #�������!�������"�$���������������%� ���&��� ����'�������� (���!(����)�� �� �� �� �� �� �� �� �� �� �� �� �� �� �� �� �� �� �� �� �� �� �� �� �� �� �� ��
�
�
�
�
�
�
��
�

�

����������������
��������� 4������������������"���������������������������������
��������� 3��������������������������������� "�������5����������������������
�������� Etapes du développement de la prostate chez l’homme et la souris�
�������� ������������������������ "���������������������������������
��������� Régulation du branchement de l’épithélium par les andromédines.�
��������� Représentation schématique de la signalisation Hedgehog��
������� � ����������������������������� ���"���"�
��������� Coupe histologique et représentation schématique de l’épithélium �����������������
��������� ��"����������������������**��������������������������������
���������� ��������������������������������������������������
���������� ��������������������������������
���������� 3����������������������������"��������������������
��������� �������������������������"+�����43��
��������� Mode d’action génomique du récepteur des androgènes (AR).�
���������� �������������������������������������������
���������� Résidus ciblés par les principales modifications post ��������������������������������������
�������� � �����+���������������������������������������
���������� ����������� ��������������43�������������������������������������
���������� �������������4�����23��
���������� 3"�������������23� ���4����
���������� ����������������������������������23�!��
���������� �������������"�����������"4����3���
��������� #�**����������������������������������������������������
��������� Fenêtre d’activité des complexes Cyclines/CDK durant le cycle cellulaire.�
���������� �������������������������������������� ����
���������� Rôle des chaperonnes HSP dans l’homéostasie protéique.�
�������� � Inhibition de l’apoptose par les HSP 27, 70 et 90.�

�

������� ��� Rôle des chaperonnes d’histone dans l’assemblage des nucléosomes. �
������� �� ����������� ������������������������������������������������
���������� ������������ ����������������������������
���������� Principaux sites de phosphorylation et d’acétylation de NPM1.�
�������� � ������������������������������
���������� -������������������������������������� �������������
���������� .��������� �������������4��������� ���������4 ��
��������� !����������������������"���
��������� Contrôle de l’activité transcriptionnelle de YY1 par mTOR.�
���������� Effet de la rapamycine et de la Torin 1 sur l’expression de NPM1.�
���������� '�#�������������4��������������������4$%���&��4�. ��
�������� � Modèle de stabilisation de l’ARNm de NPM1 par mTOR.�
���������� '��&�������4%�4���
���������� .��������������(������������ ����#�����������������#�������������������������������&������
���������� )������������������#"���4�����%������������#"�������������
���������� Principaux marqueurs de différenciation dans l’épithélium prostatique.�
��������� NPM1 induit l’expression de l’EGF.��
���������� Activation des récepteurs de l’EGF dans un contexte sain et néoplasique.�
��������� 6����������������������������������������������������%*%������%����+������%*%��
����������� )����������������������������������������������������������������� ��������gène de l’EGF de l’homme, de la souris et du rat.�
� �
� �
� �
�

�

G�
�
�� ����� ������������������ �
������������ ������������ �������������� � ���� �
����� ����� ��� ����������� � � ��
��������������� ������������ �������� � � ��
HIGI��2��1�����2�2����������2��1-�2��1���2�12���2��2�2� �1����2���J��1���� K�
HIHI��2��1�����2�2��1���2��������2� 2�2���� 2�������2������1����2�1�2����J2��2�� � L�
HIHIGI������������������������������J����������������� ����!����������J�"��!���#�� M�
�N� �J2�$����������� ����������%�������$�$�����#�������� �#�������������"��!������ GO�
N� ����%����#�����!� �����J�������������������#&������� #����'������������"��������
�J��������������"����������������� GG�
�N� ���(���������!���"������"���#� ����!������������#����������������������%����#���"����������
"������������!)��� GP�
HIPI��2���2�2���� 2��*�2��1���2���1����1���2�2��2 2����2��1���1��2�����1�*�2� GQ�
HIPIGI����%���������&� GQ�
HIPIHI��+�PIG� GR�
������������� ��� ������������� �������������������� ����������������� ���
���������������� � � �
��� ��� ������ ���������������������� � � � �
��� ����������������������������� �� ���� ������������������������������� � ��!�
HIGI� �2��2�2�2����2��1�����2�2��,��2�������2���2��1����2��2�2��2 2�������1��2�� HO�
HIGIGI��������"��#������������-����%����"�������������#"��%���������������"��#�����#��������� HG�
HIGIHI�������S����(���#������1��%������������������(������������"������������������"����������
�!��������� � HP�

�

H�
�
HIGIPI�1�������������#�!�����������������������"��#������������-���� HP�
HIGIQI�����)��!-��������������������#������������"��������"������#������"�������'����"��(������
���������.#�� � HR�
HIGIRI��J�&"������������J����(����������%����#������1����������������#����������"������������#�
������� � HT�
HIHI� 1��2�1�����2����2�����1��1�����2��$1��2�����2������1��2� HL�
HIHIGI����(����P+U1+�U����� HM�
�N� ���"�����������"!��"!�������2������#���(-��������������������#�������-���"�������.#�� HM�
N� ���������#���%%����#����"����������1+�� PO�
HIHIHI����(�������� 1+� PH�
�N� �J�&"���������J#���%������������#��(����������(�������������#����J�������-��������
"�������.#�� PH�
N� ���"���������"���������������%��������"��#�)����"-����(������"���������2�������
�J�������-���"�������.#�� PP�
�N� �����������"������J#��� �#�����J�#�����#��������#���������%%����!�����������#����������������
��#����"��������(��S'S(�������%����#����������������� PQ�
HIPI� 1��2�1�����������2��2����1�2� PQ�
HIPIGI�����)��������#�����������(�������.#�����"!����%������������� PQ�
HIPIHI����"�������������������%%��������"!������#��)�����������#���"����������"��&������)������S
��+� � PR�
�N� �������"��&����)�������S��+QUT�����)������2U��+H�"�������������"��������#�"��������
������������ PK�
N� ��������#����������� �����#���(��������%��.#����������������������-���"�������.#�� PK�
�N� "RP���"��������#��� ����-���'����"������������#���������������"�������� PL�
������ ����������������� �������������� �����������
���� � ����� "!

�

P�
�
��� ������ �������� "!�
��������������������� ��� "��
���������������� "�
������������������������#�������������� "��
���������� ���� ��� ������� ��� �������������� "��
��������� ������� $ ����� �������������� "��
"�������������������������� " �
QIGI�� � G���-2��1����$2�1�������2�1�������2��������2�*�2�2��2����2�2��1�2�1��1��������������������
��1-��2��2�� *�2I� QL�
QIGIGI�� G���� ������"GQ��%��������#����J����(�������"RP� QL�
QIGIHI�� G�/�#��#���������"�����������������"������������J1���������������������������� �������#�
������I� � QM�
QIHI��1����2��2������2�� G�2���$�2*�2��2��1����2���� 2��������2�� RO�
QIHIGI�����#��&"������������ G��������������'�#�������"����%�����%� RG�
QIHIHI�� G���!� ���J�"�"�����������%-���#���������������#&������������������������#&� RH�
������ � �������%� ����������� ��
������ ������� ���������&����������������������' ���
������� ������� ������������������&���������������������������������� � ����
�����' ���
����� � � ��
�� ����� �

�

Q�
�
�� ����� �!
����������� ������� ���� ��
�������� ��������������������������� ������(#�( #� �� ���
����2����G���2�1���2��S��������������2���J2��2������2�� G�V� TQ�
�J1����2�+�1�2��2� ����2��S2��2��2*��2������2��������2��2��1���1���������2�� G�V� TT�
�� 2��� �����������2��1���1-��2��2��J1�� ����1�������� G�V� TK�
*�2��2����J 1����2��1����2��2������2�� G�����2�1�� ����V� TM�
�������� $������������������ � ����' ���
� G��$��2S�S2��2������1��$$2�2��1�����2��J2��2�� �����1�*�2�V� KH�
*�2��2�2����1�2���2��2��J��2����2��2������J2�$�����2�1��� GV� KP�
1��*�2�� 2�1�� 2�� G�����S2��2��J2��2������2��J2�$V� KR�
1��*�2�W�N� 2�1�� 2� ��2���1�2�� G��� ��2S�S2��2��1����$2�1������������V� KR�
� GX������2�2�2��HK+GX��2���1������1��2�2��2��V� KK�
����&�� !
������������������������� �
�
� �

�

�
�
�
�
�
Introduction Bibliographique�
� �

Vésicules SéminalesCanauxDéférents
LobesAntérieurs
LobesLatéraux
LobesDorsaux
LobesVentraux
Vessie
Urètre
Vessie
Zone Centrale
Zone Périphérique
Zone deTransition
SphincterProstatique
Urètre
A
B
A
Figure 1. Anatomie et histologie de la prostate humaine et murine.A. Localisation et représentation schématique de la prostate humaine (à gauche, coupesagittale) et murine (à droite, vue latérale). B. Coupes histologiques de prostateshumaine (à gauche) et murine (à droite) observées après coloration hématoxyline-éosine.
Lumière
Epithélium
Stroma
VaisseauSanguin

� ����� ����������������������������������������������������������������������
T�
�
������������,� ��������,�������,������������
0�
�
� =>��������������������,������������
�
$%� ,-./`%`�� ��`� j��� ������� �k�������� �w� ������ j�����`�������{� ��`j��� �������`����`�
�j�����|�������`���`j���`��wj��`��}�.������`��������wj���������j���j�{�� �k���`�|����`�
��� ���`�������wj�����k��`� �������~����������������� �w������j�`�}���� ��`�`���j��������`�
�j���|�������� `���� ����{� ��� ���� ��� �����j�{� ��� ���� ��`������`� ��� ������� `�����`��}�
-���� ��� �j���� �`� ��� ��`{� ��� ��`�`�� ��`� � ���� ��� �� ����� �����`�� ��� �� �k�� �������
��j���� ���� ����� |��`��jk{� ����jk{� ��`���jk� �`� ��`����j��� ������ =�}� ������� �``��
���������� �� �����j�{� ��� ��`�`�� ���� ��� �� �� ���� ��`� ���� �wj�� ��`� ��� |j��
���`����j�}�-�``�����������`���`�`j�������j�`� �������jk{�j�����{��j�� ����``��`����
�������� ���� ����`���� ��`�`��j��� |���� �wj��`��{� �`� �j�� ��`� ������ ��� j��� j��� ���
���j����� �`��������}��w��������������� ��`����`��� ����j��j��� ��������������`j����
���j���`���������j�j������� ����`�����`�������j������������`��j��{�����j�����
��������������j�����|������jk�����j�����`���� ��`��j��}�
��� ���� ������`������ ��� ��`�`����`� ��� ��j`��� ��� �������j����� �`����������j���j����
��`�`��j���j����`�������j`�j����� ��������� ��� � ��`����j� � ����}� �����`��������j�
����`���� �w���j��`����j� ���j���� ���������j�� �j�� j�� ����� �� �`��`������ ��� �`�`����
���� � ����`�����{� ��� ��|�����`� ��j�� �j�|��� �`� ��j�� j|��� ������`� ����� ���� |����
����`���������������`���� ���� �`�����`��`��� ��������`�� �`����������`����������������{�
~����}�'�� �j�{����������������������`��������������j���� ��`�`��j�{���`� � ���j������j��
�j���������w��`������� �����j�� ��`�`��j���!�4{���������������������������{���`������`����
���j���`����j�� ��������������� �`������������������������`�������������`���}��
�
�

UGM
Bourgeons épithéliaux
Urètre
UGE
Figure 2. Représentation schématique du sinus urogénital mâle en coupe transversale.L’épithélium urogénital (UGE) est situé autour de l’urètre, sous la vessie. Il est enveloppédans le mésenchyme urogénital (UGM). Lors de l’étape d’initiation, les bourgeons épithé-liaux envahissent le mésenchyme selon un axe proximo-distal représenté par les flèches.
détermination initiation branchementmorphogenèse différenciation maturation
Période fœtale Période néonatale Puberté Période adulte
Période fœtale Puberté Période adulteHomme
Souris
Evolution morphologiquede la prostate
Stades dedéveloppement de
la prostate
a
b
Figure 3. Etapes du développement de la prostate chez l’homme et la sourisLe développement initial de la prostate se fait en 4 étapes: la détermination, l’initiation, lebranchement et la différenciation. Ces phénomènes ont lieu durant la période foetale chezl’homme alors que le branchement et la différenciation a lieu en péridode néonatale chez lesrongeurs. Le pic de sécrétion d’androgènes à la puberté permet ensuite la maturation finale dela glande. Modifié d’après Prins & Putz, 2008.

� ����� ����������������������������������������������������������������������
K�
�
� ?>�������������,� ��������,������������
�
��� ��`�`�� ����|�� �j� ���j�� j������`��� �"#��{� � ��� �wj�� ��j����`� � �`������� �w�������
���������j�� �"#.{� ��������� � ����!� �"#�� ��`j��� �wj�� ��j����`� ���������`�jk�
�w������� ���������j�� �"#�{� ��������� � ������!$#��� ������ ?�}� ��� ��|�� ����`�
���`���������� ��`�`����`�j�� ����j���j������j���|����������`� ��{�����|��������`������`��{�
�j�� ����`� �w���������`� ����|�������� �wj��� ��`��� ��� �w"#�� |���� j��� ����������`���
��`�`��j���� �w���`��`��{� �j�� ����`�� ��� ��� �������� ���� �j������ � �`�����jk�
��`�`��j����� ��� ���������`{� �� �w�� �����`�� �� ��� �������`��� ���� �j�������� ���
����������`������j�����{��j��������j�������$���`�� ���j��������������`��!���������!j`�{�
������������@�}�-�����w����{������|�� ����`������� ��`�`�������j��j���`�����������`�
`�������� `�����`������� |��� �%`���� j����� `���������� �����������}� �4j��`�����{����� ����
����j��{���� ��`�`����`��j�����`��������������������`��������j��� ��`����j���|�� ����`�
�� ���j� �j���`� ���� ~�� �������� �j��� ��� |��}� �w�k ������� ��� �����jk� ��� ������ ���`��`��
�����`� ��� ��|�� ����`� ��`� �� �w������� ���� ���� ����j��� ���� ���� �w����� �wj���
����������`������������������� �`�|����`���������j�����������`���������wj�� ��`����
|j���� �����j���`����j�����}������`j��`���������������������j��j���`���� j���`������
���� ����j��� �`� �w����}� .���� ���� ��� �� ��� ����� ��� �������� �`� ��� ����������`���
`��������� ��� �w� �`����j�� ��`�`��j�� �j�� ��j���`� ����� ��� ��`��� ����`���}� � &j�� ���
�|������`����`� �����j�`�����������`����`�����������`���|����������������`����j������`�
���������� ��� �w�`��� �w������� �`������� �`� � `����j��{� �`� �wj�� �����j�� ��`��`�
��`������� ��`����`�� �`��������`����������`�jkI��
�
� ?>=>������,�������������,��������������������,�,��������������,D�����������
�
���� ���������{� �`� �`�����`� ��� `��`�`����� ��j�`�� ��� ���� `��`�j���� �%`�jk{� ��`�
�����`����� �� ��� ��`������`��� ��� �w"#�� |���� j�� ��|����� ��`�`��j�� �`� �� �w���`��`��� �j�
�j���������`�� �`������}�-��������j���{���� ��j`������`��`�`��������j`����~���j���
���|����%`���}�'����k ���`���w"#�� ����|����j�������%`j���j��������~����~���j����`�j�`�|���
�����`����j��������������w������������� ��j����`� �������j������ ��`�`��j��}�!���

�

� ����� ����������������������������������������������������������������������
L�
�
�`��{������k ���`�� ����|������������%`j����������~�{�~���`�~�� �j����`�j�`�|�������� ���
�$��� �`����k ������`��� ���`���`� �����|�� ����`�����j������ ��`�`��j��� j��
��� �`�|����`�~�{�����`�~��������`���j���-j���{�~����}�'����k �����������������������������
���� ��� ��`� �`� ����� ���� �$���� ���j�`�`�� ������`���� ���� ���j�{� ~����}� � '�� �j�{� j���
�k ��`���j���j�������������������w�k ���`���w"#�������`� ����|����|��`���� ��j`������
`��`�`����� �������`��`�j�����%`�jk��j���`������j����j���j���������`� ��`�`��j�}�.����{�
�w�k ��`��� ��� ���j�� j������`�jk� ��� ��`� ��������� �� ���� ���������� ����`� �w���`���� j��
�j���������`� ��`�`��j�� �&������ �`� ��}{� ~����}� ��� � ���'`� ��� �jwj��� �� �����`���
���������j���j���`� ��� |��� �%`���� ��`������`�������j���|�� ����`� ��`�`��j������ ����
����j��}�
�w��������� ���� ������`��`�������� ��� ��`�`����`� ���������`��`�`����� �'(&�}� -�``��
��������`� ���j����� ������j`������ ���`��`�`����� ��� ���������j`�����j�������$������
�w� �`����j�� ��`�`��j�}����'(&���`������`�������j���|�� ����`������� ��`�`�� j���j�����
`���`����`������`������`��`����|��j��������`�j��������������j`������������������`�������
�j������� �`�����jk� ���� ���� �%`j�������{��`� �`�����`���`� ���|����� ��� �w�������`��`���
���j�`����� ��� '(&� �)����� ���� �����`���{� ~��~�}� �w�� �`���� ��� ��� '(&� j�� ���
��|�� ����`� ��`�`��j������ �w��������`�����������|������ ��� �w�`j������ �`���`��
�`��`� ���� �j`�`���� ���`�|�`����� ��� ��� ������j`���}� -��� �`���`�� �����`��`� ��� `����
�������� `�jk� ��� '(&� ������� �� j��� �`� ���� ��� �w� �`����j�� ��`�`��j�{� ����� �j�� ����
|���j���� ���������� �`� ���� ���jk� �������`�� ��`� �����`�� �`� ���������`� ��|�� ���
����`��������)����{�~����}��
�
� ?>?>������,�������������������������������������������,������� ���������
� ���������D�����������
�
��������`��������`��`�`������`�������'(&��������� ��`�`����`��������� ��������� `�j������
�����������4*�{�j����`�j�����`������ `������j`����� ������������}�-� �����`{�������j��
��� ��j`������`��`�`�������`������ �����������w���������`�����w� �`����j��j������`���
�"#.�� |���� j��� ����������`��� ��`�`��j�{� �w�k ������� ��� 4*� �����`� ��� ������ �%`����
�w��`���`�`����j��������������������j������`����"#����-����`���}{�~��~�}��w�`�������

Vagin
Vagin
Figure 4. Importance dumésenchyme urogénital pour le développement prostatique.Différentes combinaisons de d’UGM et d’UGE provenant de souris sauvages (+) et Tfm ont étéréalisées et greffées sous la capsule rénale de souris sauvages. Les combinaisons UGE(+)/UGM Tfm et UGE Tfm/UGM Tfm aboutissent au développement de tissus de typevaginal. Au contraire, la combinaison UGE Tfm/UGM (+) permet le développement d’unépithélium prostatique et la formation des acini. L’absence de protéines clés dans les sécré-tions souligne néanmmoins la différenciation incomplète de l’épithélium obtenu.Modifié d’après Cunha, 2008.

� ����� ����������������������������������������������������������������������
M�
�
�����������j�� �����|�� ����`� ������� �w� �`����j�� ��`�`��j����`���� ������`��`�
������`�� ��� ������� �wj�� ����� ������� ������� ��� ��� ������!)��I� �J�&�������� �J#��
�����j�����j��������`���������jk�� ��`����`�����`��� ���������������`������������j����
�`�����������|������ ��������k �����������������������`���j����������������� ���-j����
�`�����������`�j����-j�����`���}{�~����}�����`��|�jk���������� ��������j`�j����� ���`��j�����
����������� ���� "#�� �`� ���� "#.� �|����`� ��� �j���� ��j|����� j� �����`��`� j���
�j`�`��� ��`�������`��� j��4*���j����&��{�%�����" �����#���&������}�����`���j���`��j��
�`� ���j�`�� �`�� �������� �j�� ��� � �j��� ���������� �j���������� ��j|����� ������jw���� ����`�
�k �������������`��`���� ��������j����w���������{��`��`��`����������������������
�j��`����������w� �������������������`�������w"#.}�-�``���`j����� �������������`�����j��
��� ������� ��� 4*� ����� �w"#�� ��`� j��� ���`��� ���������� �`� �j������`�� j�� ���
��|�� ����`������� ��`�`��������E�}�.������`{��������������w"#����j|������|������
"#.���#� ���j�`�j���j���������`� ��`�`��j�{�������j�� �������������w"#.���j|�����
�|�� ���� "#�� ��#� ��`� ��������}� '�� �j�{� �w"#�� ��`� ��������`� �� �`��`� j�� ���
� �����`��� �j� `���j� � �`������� ������`{� j���j�� ����"#�� ��������� �|�� ���� ���j����
� �`��������� ���j������ ��� |������j���� �wj��`��� ��j�`� �� �������������`����wj�� � �`����j��
��`�`��j���-j��������'���j�{�~����I��
� � �
� ������������� �������� ����������� ��� ��������������������������������
�
�j�`���jk��`� ����w���`��`����`������`������`��{� �����j������ ��`�`��j�����������`���
��|�������`� ��������������|������`}� �w��|������j����������� ��� �w"#.��������j�� ���
����������`��� ��� ���`� ����� j�� �k�� �k������`��{� �� ��`������ �wj��`��}� .���� �w�� �����
�wj����������`����j���|��j���� ��� ��`������`�����������jk���������`��� j����``������
����j�� �����j�������������jk�����`����}�-�``���`� ��������������`���`���j`����`�
���j���� ���������`�j��� �������������`��� ����������������j������`�������� �����jk�
���������� ������ H�}� -��� ��`�j��� ��� �������� ��`� ��������`� � ����� ����
���������������}� ���� |�`� ��`��|����� ��� ��+�� ���j�`����� ��� �k��+��`� ���� �`����
��������j���j���`�����j�����������������������|�� ����`��������jk�������������}��
�

�

� ����� ����������������������������������������������������������������������
GO�
�
�� �w.#,��`���������������������������,#,��`��j���`�����j���������`�� �`�������
�w.#,�����'��#� ����(�!�����������`�����`�� �������`��������� �����jk������������������
�`���}{������}�����j��j��������� �`��`����������j���������`�����w� �`����j�� ��`�`��j��
j���j�� ��j���� ��`� ���j�`� ���� ���� �j���� ��|�������� j�� ��� �������� �w.#,� �4��``� �`� ��}{�
�����}� � '�� �j�{� ���� ����`���� �j`��������� �w.#,� �� ���� ��`�� �����`� ���� �j�`� ���������
�j�|��`� ��j�� ��������� ���j�`� j��� �j����`�`��� ��� ���� �j� ���� ���� ����� |��`��jk�
��`�`��j��{� `�������j�� ���� ���������jk{���`����j����������j�� ���� |���j���� �������������
�������`� �������`���� ����� `���`����`� �&-�������`���}{� ���~�}� '�� �j�{� j��� ������`������
�w�`�|�`����������j���� `�j�����w.#,��.#,*�����j�`������|�� ����`������� ��`�`���������
��`� ������������ �`� ��}{� �����}� -�`� ����`� ��`� ���|����� ��� �w�������`��`��� �w���������}�
!j���j��4*��w��`� ����k �����������w� �`����j�������`���������|�� ����`{������j�������
�j�� �w�j`���� ������������ �j|��`� ������� ��� ������� j�� ���j���� ��� ��|�� ����`�
��`�`��j�}�
'�jk� �������� ��� ��� �������� ,#,� ���)��) ���� ���(�!� �������{� ,#,�� �`� ,#,~�{� ��`�
��`�j��������`� �� �`��`�� j�� �`��j���� �w�����`��� ���� �j������ ��`�`��j��}� -���
��`�j��� ��� �������{� ���`��`����� ��� �w"#�{� ��� ��k��`� �j�� ��j��� ��� `�j��� ��� �`����
�����`��j���j����`��j���|��j���� �w"#.}���j���k ���������`� ����`����� ��� ��`������`��������
���jk���������`��{��������j��`�����������������``��`��`��`������`�j�����`������|���
��|������ �w� �`����j����j��`���}{�~�����!j��`���}{�������&����{����~�}�-���� ��������j��{� ���
�� ������� ���� ����� ��`�`��j��� j�� ��� ���j���� �������{� ���� ��� `�������
�w������������j���� ��|�������� j�� ������ `�j���jk�,#,{��� ��� ����|��`������`������`������
�j���������������jk��`���`���jk���`� �����`�{��$����w������`� �j���|�� ���������`���}{�
�����}� ,#,�{� �j���� ��j� �j�� ��� ��� ��� #,� � � ���`���`�� #��`�� ,�`��{� �� �`�� ���
�������� ���j��� ����`������ ���� ������� ��`�j�� �������� ��� ������� �j�
��|�� ����`�����w� �`����j�� ��`�`��j���.����`���}{�~����}�.������`{��w4*������,#,����`�
�k ����� ����� ���� ���j���� �`������� �`� ��� �k ������� �j����`�� ��� �� ���� �� j���
�`��j��`��� ���������j�� ��� *����}� '�� �j�{� j�� `���`����`� �j� ,#,�� �j�� ���� ��`�`���
|��`�����������`� ����|������������������ ����`������|�� ����`������������������������
�w���������{� �`� �� ��� ��+�� ������ �����`�� ��j���j��� �`� ��}{� ~����}� ������� ���
�����`��`���{�,#,������������ ��������`�����j���|�� ����`������� ��`�`�� j���j������
�j������|�������� j����������,#,�� �����`��`�j��� ��`�`�����������`���|�� ����#j�

/0/&*01�
�
��1@*&�
�/0/01�'((�
��223��
������
�
������
�
4�����
�
� ������������A��
�
� ������ �����
� ������ �����
� ������������
��� !��
�"�#�$"�����
�
Figure 5. Régulation du branchement de l’épithélium par les ������ �������� ��� ���� ���� ���� � ���� �!������ ���� ��� ������� �� �������"� ���� ��������� ��� #��������� ����� ���� $�������� � ������"� ���� %� ���� #������ &"� '�� ��������� l’élongation� ��� ��������!�#������� l’épithélium��� ����'���� -�������� �������� l’élongation� (���)��� ���'���d’autres�l’inhibent�(� ���)��L’action��*��!� ������������������*�������$��������� �����+�������� ���#���� !��# ����� ��� ��� ������� ��� +� ���� ������� �� ! # � ��� ���� �����L’épithélium�'�����+��������#������$�����������#�����������$$������� ��(����)��D’après�,����-�,��."�/001��
����

� ����� ����������������������������������������������������������������������
GG�
�
�`� ��}{� ~����}� 4j� 2�`3�43�{� ���� �2j34�� 4�|��4����� 52j3� ,#,~�� 53����`��`� ���� �2j3��2���
�54`���4�jk�5�j��5�`4`���`����j2j5��24����2��3�jk{��j���3��`�j��3����j�46j���`������`4���
���,#,~�{� 2�`3�43����`���,#,�� �'2���2j3��`���}{� �����}� .���23���|�� ��� 3��j�`�`�{� ���
`3�4`����`� �w�k5���`�� �w"#�� 4��j�� ��� �2j34�� ,#,~����� 5�3� �j� ,#,~�� �`� ��� ��� `��`2�`�32���
3��`�j3�� 5�3`4�������`� ��� ��|��255����`� 532�`�`46j�}� -�5�����`{� ��� `3�4`����`� ��� ���
�k5���`���|���j�,#,~����j����`� 4����4��}��w�k53���42�����,#,~����`��2��������43����4��
4��j��4���`�� 52j3� ��� ��|��255����`� �23���� ��� ��� 532�`�`�}� -���� ��`� ��� �k�3�j�� ��� 3����
�w�j`3�����`�j3������52j3��w��2���`42���`�����3�������`�������������}��
�
�� ������`�j3��4��4���`��w��2���`42���������jk�2�`34�j��`�������4������5�������
�w23���4��`42���5�`4���������4�4�
!�j�4�j3����`�j3��5�3�34������3�`���5�3�����`32����j��4|��j�������5�3`4���4�`�����������jk�
�����2���`42���2�`���34`��52j3�4��4��3��w4�|��42���j�����������5�3������4�4�����3�4���`����
532�4��3�`42�� �54`���4���}� �w�`42�� ����32�4���� ���� ��`�j3�� �`4|�`�j3�� �|�� ���� ��`�j3��
4��4�4`�j3��������324�����������4�4�2��j4`���j����|��255����`���3�2�4�jk��������������
532�`�`46j���`���j����4�`34�j`42���2�2����������4�4}� �
������!�����������!������������������4��4���`����532�4��3�`42���54`���4�����������H�}�-�������
�2j34�{� �����`�j3���5�����`��k534�������� �������������j32����4`��� �23����� �w4�4`4�`42���j�
�2j3��2������`� �54`���4��� �`� ��� ��� 3��4�4�`42�� ���� �4�4� ��j� ~����� �2j3� �%`��� �j�6jw�� ���
��4������� ������ �`� ��}{� ���~�}� ��� `3�4`����`� �w�k5���`�� �w"#�� �|�� ��5��� 4��4���
�w��2���`42�������2j3��2���532�`�`46j��{��`������2j34����5�2�4��j��4���`���52j3���5������5�
������53����`��`� j�� `�jk�����3�������`������ ��� �2��� |��`3��� �j5�34�j3��� ��j4����� �2j34��
��j|����}� -��� �2������ 5����`� ��5��� 2���� j�� 4��4�4`�j3� ��� ��� 324������ �54`���4���}�
�w�k53���42�������5�����`�3��`3�4�`���j`2j3��������jk������2���`42�{��`��������`�3��j����5�3�
�w�j`3��� �`�j3�� 3�`32j|��� �j3��`� ��� �235�2������� 532�`�`46j�{� 2���� �����+��'��!���
������6j4��w�`4|�{�2j�,��~���`��������32������6j4��w4��4���`}����3��j��`42������w�k53���42�����
��5���5�3�����j�`45�����`�j3��5�3��`����3��3������3��4��`�����2���`3�`42������2�������
���jk������2���`42�{�2�`34�j��`��4��4��� ��� 3��j��`42�� �4����j�532���j������3�������`�
�7j�����`���}{�������}�-����3��4��`�����2���`3�`42���2�`���������`�3��j����5�3����532`�4���
�2��4�{�6j4� 4��4��� �w�`4|4`����� ��5������ ��� `4`3��`������ ����4�4�j��k`3����j��43�}� -���� ����

�

� ����� ����������������������������������������������������������������������
GH�
�
�2j34�� 4�|��4����� 52j3� �2��4�{� ��� �2j3��2������`� �54`���4��� ����� ���� �2���� |��`3�jk� ��`�
2�5��`����`�4��4��{��`�4����`��23`����`�3��j4`�����������2�����23��jk��`���`�3�jk���-228��`�
��}{������}��
-2���� ��5��{� ��5��� �� j�� 3���� 4��4�4`�j3� �j3� ��� 324������ ���� �2j3��2��� �54`���4�jk�
�#34��4��� �`� ��}{� �����}� �2�� 4�|��4��`42�� 2��j4`� �� j�� 5�j�� �3���� �2��3�� ��� ���jk�
532�`�`46j����`����3��4�4�`42��}�'��������$�������{����`3�4`����`��w�k5���`��w"#����j|�����
�|����5���4��4������324�������`�����2j3��2������`�����w"#.}�
���324�����������2j3��2���532�`�`46j�����`���������`� 4��4����5�3�&#,9~}�-���� ����2j34�{�
&��9~���`��k534������5�342���52�`���`��������� �������j�����j����������� ���23��6j������
3��5`�j3��&#,9*���`�&#,9*����2�`��k534����������24��������������j�������������`�j�����`�
�54`���4������&2��4��2���`���}{������}�&#,9~���4`��2���j3�������jk�`�5������j��43��{������+2��
�j`234��� ��� 3�534���`� �w�k53���42�� ��� ,#,~�� 5�3� ���� ���j���� ���������`�j���� �`� ���
��+2�� 5�3�34��� ��� ��26j��`� ��� 532�4��3�`42�� ���� ���j���� �54`���4���� �`� ��� �`4�j���`� ��j3�
�4���3��4�`42�� �-����� �`� ��}{� ~����}� 'w�j`3�������3��� ��� ��� ���4���� �j� &#,9� 4���j����`�
��������`� ��� ��|��255����`� ��� ��� 532�`�`�{� 2���� ���� �`4|4���}� -��� ���2532`�4����
��3�`���� 5�3� ���� ���j���� ���������`�j���� 4��4���`� ��� 532�4��3�`42�� �54`���4���� �`� ���
3��4�4�`42�� ���� �2j3��2��� ��� ��4����`� �j3� ��j3�� 3��5`�j3�� 3�`32j|��� �j� �4|��j� ���
�w�54`���4j�� �-��4���� �`� ��}{� ���~�}� ���� �`4|4����5�j|��`� $`3�� 4��4�����5�3� ��� �4k�`42�� �jk�
�2��4�`�`4���{� 6j4� �2�`� �k534����� ����� ��� 532�`�`�� ��� ��|��255����`� �`� ��`j3�}� ���
`3�4`����`� �w�k5���`�� ��� 532�`�`�� ��� 3�`�� �2j|��j����� �|�� ��� ��� �2��4�`�`4��� �`4�j��� ���
324�������`� ����3�������`��������jk�532�`�`46j����������������`��`2�`�32��}�44��4{� 4��
�k4�`��j������������`3���w�`42�������`4|4�����`��������2��4�`�`4���6j4{��j��$���`4`3��6j�����
������� ��`3�� ���� ��5�� �`� �2��4�{� 5�3��`� j��� 3��j��`42�� �4��� ��� ��� �235�2�������
�54`���4�����j��4|��j������k`3��4`����4�`������������jk}�
!�j��3������`{�������2��`3�`42���j�3�������)�`�������������|��255����`�������532�`�`��
���� ��� 3�`� �`� ��� �2j34�� �� 5�3�4�� �w��2j`�3� j�� �`�j3� �� �� 3����j� 223�2���� ��� ��`�j3��
5�3�34���� �7j�����`���}{� �����}�)�`�����`� j������2532`�4��� ��3�`��� ��4���`�5�3`4����� ���
���4����)�`}� .���� �2j�� j�� 3���� 4�523`��`��j3��`� �����|��255����`� ���3�2���43��5j4�6j��
�2��4�|��4��`42��2��j4`���j����23`��4`��5�34��`�����.����j�4��`���}{�~����}�'�������532�`�`�{�
���532`�4���)�`�����`���3�`���5�3���������������j��4|��j�������5�3`4���4�`�����������jk�

�� ��
����%�� ��� Repré��&'�'�(&� sché�������� �� ��� ��������� �� ����� ������ ��� ������� �� �� ���� ��� ré������� ������ ������ ������ ���� ������ ����������� ���� ���ré������� ���������� ������� ������ ����������� ����� à� ��� dé �������� �� ������� ��������������� ���2� �� à� ��� proté������ �� ������� ������ ������ ������ tronqué�� �� ����� � �������� ��� ré���������� ���������������� ��� gè���� ������ �� ��� ���� !������ "�� �������� �� �� ��� ���� lè �� l’inhibition� exercé�� ���� ���� ���� ����� "��� �������� ������������������#������������ ����������� ����������� �����������������������"���������������������������$� ������� ��� gè���� ������ �� ��� ����!�� ��� � ��� ����� ��� ��� ���������gè��������������� ������D’après���������%�������&�'����
�

� ����� ����������������������������������������������������������������������
GP�
�
�(� �)*(+�`,*(� *�� �))�� �-�,(�� )�� .-*),��-�`,*(� �.,`/�),�)�}� -�)�� �� *00�� ����`� �w,(/,��-�
)w,(|�1,*(��j�1`-*0��.�-�)�1��,(,��`��*(����-�+j)�-�)��.-*�11j1�����-�(/�0�(`{���)w,0�+��
���)w����`��k�-��.�-�)�1���`�j-1����)����0,))���j�&#,�2}�'��.)j1{�)��.�-`�����)(`���0�(����)��
�*-0�`,*(�����*j-+�*(1�.-*1`�`,3j�1�,--�+j),�-1���(1�)�j-��*-0���`���(1�)�j-�.*1,`,*(���3j,�
1*j),+(��)��-4)�����)(`�����(1�)w*-+�(,1�`,*(���1��,(,}�.(�,({�)(`����1`��+�)�0�(`�,0.),3j��
��(1�)��.*)�-,1�`,*(���1��))j)�1��.,`/�),�)�1��`��*(���(1�)���*-0�`,*(����)��)j0,�-���j�1�,(�
��1��,(,}�
�
� ���|*,��7��+�/*+�.�-`,,.���j��-�(/�0�(`��(�-�+j)�(`��)��1�-�`,*(������`�j-1�
.�-�-,(�1�.�-�)��0�1�(/�0��
�w�`,|�`,*(� ��� )�� |*,��7��+�/*+� ��.�(�� ��� `-*,1� ),+�(�1��� �*(,�7��+�/*+� ��//�{� '�1�-`�
7��+�/*+� �'//�� �`� �(�,�(� 7��+�/*+� ��//�}� -�1� .-*`�,(�1� 1�-�`��1� �+,11�(`� �(� ),�(`� �j�
-��.`�j-�!�`/��~� �!`/~���� )��1j-������1��))j)�1}�.(���1�(����� ),+�(�{�!`/~� ,(/,��� )��
-��.`�j-���1�.`��*0�,(�1� `-�(10�0�-�(�,-�1��0**`/�(��� ��0*��.�-�j(�0��(,10��(*(�
�(*-���)j,��}���,(/,�,`,*(�����0*�*(�j,`���)����+-���`,*(��j���`�j-����`-�(1-,.`,*(�#),��
�`� �� )�� .-*`�*)�1�� ��� #),�}� ��� �*-0�� `-*(3j��� ��� #),�� �+,`� �)*-1� *00�� j(� -�.-�11�j-�
`-�(1-,.`,*((�)���1�+�(�1�,�)�1���� )�� |*,�� �������� M��}���� ),�,1*(���1� ),+�(�1���!`/~� )�|��
)�,(/,�,`,*(� 3jw,)� �k�-�� 1j-� �0*}� ��1� ��`�j-1� ��� `-�(1-,.`,*(� #),�� �`� #),�� (�� 1*(`� .)j1�
��+-���1��`�.�j|�(`��+,-�*00���`,|�`�j-1� `-�(1-,.`,*((�)1� ��(+/�0��`��)}{���~~����������
M��}�#),~{�3j,��+,`�j(,3j�0�(`�*00��j(��`,|�`�j-�`-�(1-,.`,*((�)���1�+�(�1�,�)�1���� )��
|*,�{�(��1`��k.-,0��3j�� )*-1���� )��`,|�`,*(�����``��|*,�����1,+(�),1�`,*({���3j,��(� ��,`�j(�
0�-3j�j-���1��))j)�1�*��)��1,+(�),1�`,*(�7��+�/*+��1`�0*�,),1����:,(+��`��)}{������}��
�)��1`��,��,,)���w�..-�,�-� )�� -4)����� �``�� |*,����(1� )����|�)*..�0�(`�.-*1`�`,3j�� ��� *�*��
.j,13j�� )w,(|�),��`,*(� ��� �*(,�7��+�/*+� ��//�� 0�(�� �� j(� ����j`� ��� 1`-j`j-�`,*(� ���
)w�(�*��-0�{��(`�-,�j-��j���|�)*..�0�(`���� )��.-*1`�`�� �-/,�(+��`��)}{� ~�����:,(+��`��)}{�
�����}�-�.�(��(`{���1��k.�-,�(�1�j`,),1�(`���1�+-����1��w�k.)�(`1��w"#�����1*j-,1�,(|�),���1�
.*j-��//�1*j1� )���.1j)��-�(�)�����1*j-,1�1�j|�+�1�0*(`-�(`�3j�� )w�`,|�`,*(���� )��|*,���1`�
�11�(`,�))�� �� )�� �*-0�`,*(� �`� �� )w�)*(+�`,*(� ��1� �*j-+�*(1� .-*1`�`,3j�1� �!*�)�1�5� �`� �)}{�
~����� ��-0�(��`��)}{� �����}��//��1`��k.-,0��0�6*-,`�,-�0�(`���(1� )�1��))j)�1��.,`/�),�)�1�
1,`j��1���(1� )��.�-`,���,1`�)����1��(�jk��(��)*(+�`,*(� �!j��`��)}{������}��*(�-��.`�j-��1`�

-������������������� ��������������������������������� �������� �� ���� ������ ���� �������� ��������� ���������� �� � ���� ��� �������l’élongation���� �������������� �������� ��������������� ����� ���� �������� ���� ��� ����� ���� �����������������������!������ ������� l’expression����� �����������������������"��#� � ��� �$����������������� ��� ���������� ���������������� �%���� ��� ���� ���������l’épithélium,�� ������� �������������� �����������$�������������������������������������������������� ������������������������������������������d’un������������d’acini������D’après�&�����'�&�(��)��*��

� ����� ����������������������������������������������������������������������
GQ�
�
+,j`-`�.k+/,0��1�23� ,.�0�3.2/�0.�.2`4j/�2`� ,w.k`/�0,`��1,3`�,.�1.3��2�jk}��w.k+43,`,42�
1w"#�� �� �//� ��� *����� ,2/,�.� ,21,/.`.0.2`� ,�� /4,33�2.� 1.3� �4j/5.423� �+,`/�,,�jk� .2�
.2`/�72�2`�j2.�1,0,2j`,42�1.�,w.k+/.33,42�1.�,#,~��.`�j2.��j50.2`�`,42�1.�,w.k+/.33,42�1.�
��!��}� �,/..3`42.�.`��,}{� ������)�25�.`��,}{� ������}� �.�041�,.��`j.,,.0.2`�+/4+43��.3`�
6j.�,w.k+/.33,42�1.��//�1�23�,��+�/`,.�1,3`�,.�1.3��2�jk��+,`/�,,�jk�|��1,0,2j./�,4�,.0.2`�
,w.k+/.33,42� 1.� ,#,~�� .`� �j50.2`./� .,,.� 1.� ��!��}� -.,�� +./0.`� ,.� 1�+,�.0.2`� 1j�
5/�1,.2`�1.�,#,~��3j/� ,.3�-`�3�1.3��2�jk�3`,0j,�2`��,23,� ,.��/�2/.0.2`�1.� ,w�+,`/�,,j0�
��������N�}�
�
� ?>@>� ���� ������ �� ��!�"���� #��!���#��!� ������ ��� ,�$�%�##� ��!� ,�� %�� �%��,��
#���!�!�"���
�
4j� 1.,�� 1.3� 04,�j,.3� 3�/�`�.3� /.3+423��,.3� 1j� 1,�,45j.� .2`/.� ,w�+,`/�,,j0� .`� ,.�
0�3.2/�0.{�,��1�`./0,2�`,42�.`�,.�+43,`,422.0.2`�1.�`4j3�,.3�`,33j3�/.6j,./`�,w.k+/.33,42�
1.� 0.0�/.3� 3+�,�,6j.3� 1.� ,�� 3j+./��0,,,.� 1.3� 5�2.3� /40�4`,6j.3}� �,3� 41.2`� +4j/� 1.3�
��`.j/3� 1.� `/�23/,+`,42� 42`.2�2`� j2� 140�,2.� 1.� ,,�,342� �� ,w4'�� `/�3� 423./|�{�
,w/40�4140�,2.{�6j,�+./0.`�1.� /�5j,./� ,w.k+/.33,42�1.� ,.j/3�5�2.3�,�,.3}�-.3�5�2.3�342`�
42`/-,�3� 1.� ��842� 3+�`,4�`.0+4/.,,.� ��,2� 1.� /�./� j2� ��41.� 04/+/45�2�`,6j.���
+./0.``�2`�,��1,���/.2,�`,42�1.3�1,���/.2`.3�3`/j`j/.3�.`�40+�/`,0.2`3�1.�,w4/5�2.�4��,,3�
342`�.k+/,0�3}�"2�.23.0�,.�3+�,�,6j.�1.�5�2.3���/40�4�4k����`��,1.2`,�,��1�23�,��+/43`�`.{�
40+/.2�2`�24`�00.2`�,.3�0.0�/.3�1.�,����0,,,.�74k�4j�.24/.�1.�,����0,,,.��:}�
�
� � ����������������� ��
�
��� ��0,,,.�1.3�5�2.3�74k�.3`� ,��+,j3��`j1,�.}� .,,.�+./0.`� ,��1�`./0,2�`,42�1.3�1,���/.2`.3�
/�5,423�1j�4/+3�/.��1.3�.3+�.3��,,�2`�1.�,��1/434+/,,.���,w/400.}�-/.��,.3�0�00,��/.3{�
1.� 0j,`,+,.3� 1j+,,�`,423� 5�2,6j.3� 42`� 1422�� 2�,33�2.� �� �� ,j3`./3� 1.� 5�2.3� 74k{� 3j/�
1,���/.2`3�//404340.3{�+4j/�j2�`4`�,�1.����5�2.3}����+43,`,42�1.3�5�2.3�1.�,w.k`/�0,`���w���
,w.k`/�0,`�� �w� 1��,2,`� ,.� +/4�,,� 1w.k+/.33,42� 3+�`,4�`.0+4/.,,.� 1�23� ,w.0�/�42}� 4,23,{� ,��

�

� ����� ����������������������������������������������������������������������
GR�
�
+/43`�`.� �`�2`� j2� 1.3� 4/5�2.3� ,.3� +,j3� +43`�/,.j/� /.�� ,.� 0�,.{� ,.3� 5�2.3� 74k� 6j,� 3w��
.k+/,0.2`{� 342`� 3,`j�3� 1j� -`�� 1.� ,w.k`/�0,`�� �w� 1.3� 1,���/.2`3� ,j3`./3}� !.21�2`{� ,.�
1�|.,4++.0.2`� 1.� ,w"#�{� �� 5�2.3� 74k� 342`� �`,�3{� �� 3�|4,/� 74k�~�{� 74k�~�� .`� 74k1~�}�
74k�~�� .`� 74k1~�� 342`� .k+/,0�3� 1�23� ,.� 0�3.2/�0.� .`� 1�23� ,w�+,`/�,,j0� j/4�5�2,`�,}�
�w�`j1.�1.�34j/,3� ,2|�,,1�.3�+4j/�.3�5�2.3���+./0,3�1.�1�042`/./�6j.�74k�~��.`�74k1~��
3`,0j,.2`� ,�� /4,33�2.� .`� ,.� �/�2/.0.2`� 1.3� �4j/5.423� +/43`�`,6j.3� �!41,�3.7� .`� �,}{�
~������!41,�3.7�.`��,}{�~����}����1�,�`,42�1.�74k�~��6j�2`���.,,.{�2w,2�,j.�+�3�3j/�,.�240�/.�
1.��4j/5.423�+/43`�`,6j.3�2,� 3j/� ,.j/��/�2/.0.2`� �.4240,1.3��21� -�+./,{� �����}� .2�
/.|�2/.{�74k�~��3.0�,.�.33.2`,.,���,��1,���/.2,�`,42��42`,422.,,.�1.�,w�+,`/�,,j0�1�23�,.�
,4�.�|.2`/�,{�400.�.2��``.3`.� ,w��3.2.�1.3�+/4`�,2.3�3�/�`�.3�0�84/,`�,/.0.2`�1�23�.�
,4�.� �,23,� 6j.� ,�� +./`.� 1.� +4,�/,`�� 1.� ,w�+,`/�,,j0}� �.3� �`j1.3� /��,,3�.3� /.�� ,w/400.�
042`/.2`�j2.�1�/�5j,�`,42�1.� ,w.k+/.33,42�1.3�5�2.3�/40�4`,6j.3� ,4/3�1j�1�|.,4++.0.2`�
1.�+�`/4,45,.3��� ,w�5.��1j,`.{�34j,,52�2`� ,.� /-,.�1.�.3�+/4`�,2.3�1�23� ,w/40�43`�3,.�1.� ,��
5,�21.�0�`j/.��)�,`/.52��.`��,}{��������,,,./�.`��,}{�������.1��/13�.`��,}{�������.j25�.`��,}{�
�����}����
� �
� �������� !����
�
�:)�}~�.3`�j2���`.j/�1.�`/�23/,+`,42��++�/`.2�2`��� ,����0,,,.�1.3�5�2.3���/40�4�4k��:}�
�42�.k+/.33,42�.3`�/.3`/.,2`.���,��+/43`�`.�.`��jk�5,�21.3�1.�-4�+./}�-/.��,��34j/,3{��:)�}~�
.3`� 1�8�� .k+/,0�� �� ,w.0+,�.0.2`� 1.3� �4j/5.423� �|�2`� ,.j/� �4/0�`,42{� 0�,3� 342� +,�
1w.k+/.33,42���,,.j�.2`/.�,.3���0.�.`�~��0.�84j/3�+43`2�`�jk�1�23�,w�+,`/�,,j0�+/43`�`,6j.{��j�
040.2`� 1.� ,�� �`41,���/.2,�`,42� ��,.�./,/� .`� �,}{� ~����}� �:)�}~� 3`,0j,.� ,.�
�4j/5.422.0.2`�.`� ,��1,���/.2,�`,42�1.� ,w�+,`/�,,j0�+/43`�`,6j.{�.�6j,�492,1.��|.�342�
+/4�,,� 1w.k+/.33,42}� �.3� +/43`�`.3� 1.3� 34j/,3� �:)�}~���� 42`� j2.� 1,0,2j`,42� 1j� 240�/.� 1.�
/�0,�,�`,423�1.3� �2�jk�+/43`�`,6j.3}� '.�+,j3{� ,.�+/4�,,� 3�/�`4,/.�1�23� `4j3� ,.3� ,4�.3�.3`�
�,`�/�{�3j55�/�2`�j2�/-,.�1.��:)�}~�1�23� ,��1,���/.2,�`,42��42`,422.,,.�1.� ,w�+,`/�,,j0}�4�
,w�5.� �1j,`.{� .3� 34j/,3� 1�|.,4++.2`� 1.3� ,�3,423� /�+./+,�3,6j.3� .� 6j,� 1�042`/.� ,.� /-,.�
,0+4/`�2`� 1.� �:)�}~� 1�23� ,w/40�43`�3,.� +/43`�`,6j.� ��/�`,��#�j/� .`� �,}{� ~����}� �.� /-,.�
1,���/.2,�`.j/�1.��:)�}~����`��,21,/.`.0.2`�42�,/0��/.��,w/400.{�/.��,.6j.,�3��+./`.�

����������4�����������
��
��������������� ������4���������
�������������������������
����������
���������������������
�������������������������� �
�������
������������������������������������������������������������������������!"#�$%�#&$'(� ���)%�*+',� ,)%� (�-��$%�$�,(,�%� ��(��)#� .,� �,&&'&,)� &'($��&,)/� 0�)�&,)/�$�%,�(#.$�$�,)� ,%/� .��)� '�,� (�$�.�,� (,)'�,/� �,'��,�.���$�,)1� ����'�� .,� �,)� %��,)��,&&'&�$�,)�),�.$)*��',�����)��)$���%'�,�(�&#�'&�$�,�'�$+',/�+'$��,�(,%���%�((,�%�.,�%�$,��&,)��,&&'&,)�������%�(#%�$,�.,�2'31���"���4)����&��������/�5�51�
���������� ����������������������������������������������������������������������������!"#�$%�#&$'(� ���)%�*+',� ,)%� ��(��)#� .,� �,&&'&,)� #�$%�#&$�&,)� &'($��&,)� ,%� 0�)�&,)� �,��)��%� )'��'�,� &�(,� 0�)�&,� ��(��)#,� � ��$��$��&,(,�%� � .,� &�($�$�,/� .,� 60���,�*�,/� .,� ��&&��4�,� ,%� .,����%#��&����,)1� �&&,� ��(,� '�,� 0���$4�,� ,�%�,� &,� ��(���*(,�%� #�$%�#&$�&� ,%� &,� ��(���*(,�%�)%��(�&1�

� ����� ����������������������������������������������������������������������
GT�
�
.3`� 4//�7�.� �� 7�� +/45/.33842� `j94/�7.� ��4�.2� .`� �7}{� ������ �.`:.7� .`� �7}{� �����}� �7� �� +�/�
�877.j/3��`��942`/��6j.�7.3��21/45�2.3�3`89j7.2`�18/.`.9.2`�7w.k+/.33842�1.��:)�}~{�.�6j8�
/.+/�3.2`.� j2.� |48.� 3j++7�9.2`�8/.� +�/� 7�6j.77.� 7.3� �21/45�2.3� 42`/-7.2`� 7.�
1�|.74++.9.2`�.`�7w:49�43`�38.�1.�7��+/43`�`.��!j�.`��7}{��������8.�./8:�.`��7}{�~����}�
�
@>����,���� �!���"��!�"� �!�%��#� �"�%�� �$���!�#%����,D%!��#�"&�%�% �&�� � �&�$��
�
-.``.�1./28�/.��`�+.�1j�1�|.74++.9.2`�1.�7��+/43`�`.�.3`�18342`82j.�.`�.77.�3.�1�/4j7.�.2�
1.jk�`.9+3���1w��4/1�1.�9�28�/.�3�2:/4283�.��|.�7w�`�+.�1.��/�2:.9.2`{�+j83�74/3�1.�7��
+j�./`�{�74/36j.�7.�`�jk�1w�21/45�2.3�+7�39�`86j.3��j59.2`.��4/`.9.2`}�.77.�1�+.21�142�
.2�5/�21.�+�/`8.�1.� 7�� 3852�783�`842��21/45�286j.�.`�+7j3�+�/`8j78�/.9.2`�1.� 7��+/�3.2.�
1.3�+/41j8`3�1.3�5�2.3�8�7.3�1.� 7w�`842�1.3��21/45�2.3�`.73�6j.��:)�}~}�-.7����4j`8`��� 7��
�4/9�`842� 1wj2� `833j� :�j`.9.2`� :8�/�/:83�� 1wj2� +482`� 1.� |j.� 3`/j`j/.7� .`� �42`8422.7{�
423`8`j��9�84/8`�8/.9.2`�1.�.77j7.3��+8`:�78�7.3���3�7.3�.`� 7j982�7.3��8238�6j.�1.�.77j7.3�
2.j/4.214/82.3� ����% �� O�}� -.3� `�+.3� .77j7�8/.3� +/�3.2`.2`� 1.3� �/�`�/83`86j.3�
94/+:474586j.3�.`�947�j7�8/.3�183`82`.3}�'8/.`.9.2`��++43�.3�3j/� 7��9.9�/�2.���3�7.{�
7.3�.77j7.3��+8`:�78�7.3���3�7.3�423`8`j.2`�:.��7w:499.�j2.�4j:.�42`82j.�1.�.77j7.3�1.�
�4/9.� `/8�25j7�8/.��+7�`8.�.`�.k+/89.2`� 7.� ��`.j/�1.� `/�23/8+`842�+���.`� 7.3��`47�/�`82.3�
�-:�� 1.� :�j`3� +4813� 947�j7�8/.3� -:�� .`� -:~�}� -:.�� 7�� 34j/83{� .3� .77j7.3� 342`� 94823�
249�/.j3.3�.`�+7j3�183+./3�.3��j� 3.82�1j� 49+�/`89.2`���3�7}� �w.k+/.33842�1j� /�.+`.j/�
1.3��21/45�2.3�2w��.3`�6j�389.2`�+�3�1�`.`��7.��)�25�.`��7}{����~�}��w�j`/.�`�+.�.77j7�8/.�
9�84/8`�8/.�1.�7w�+8`:�78j9�+/43`�`86j.�/.5/4j+.�7.3�.77j7.3��+8`:�78�7.3�7j982�7.3}�-.77.3�8�
342`� 38`j�.3� 899�18�`.9.2`� �j� 42`�`� 1.� 7�� 7j98�/.� 1.3� �828� .`� 42`� j2� �3+.`� ��.2�
47422.��}� .77.3� .k+/89.2`� 7.3� �`47�/�`82.3�1.� ��8�7.3�+4813� 947�j7�8/.3� -:�� .`� -:~�� .`�
+/�3.2`.2`� j2.� �4/`.� �j9j7�`842� 1j� /�.+`.j/� 1.3� �21/45�2.3� .`� 1j� 9�/6j.j/� 1.�
18���/.28�`842�`./982�7.�+��78+~}�-.3�.77j7.3�342`���7w4/8582.�1.3�3�/�`8423�+/43`�`86j.3{�.`�
1�+.21.2`�1.3��21/45�2.3�+4j/� 7.j/�3j/|8.}��.3�.77j7.3�2.j/4�.214/82.3�342`�/.`/4j|�.3�
�+�/+877�.3� �� 7�� �483� 1�23� 7.� 49+�/`89.2`� 7j982�7� .`� 1�23� 7.� 49+�/`89.2`� ��3�7� 1.�
7w�+8`:�78j9�4��.77.3�3�/�`.2`�1.3�2.j/4+.+`81.3�`.73�6j.�7���49�.382.{�7���78`4282.�.`�7��
2.j/4`.2382.�6j8� 84j./�8.2`�j2�/-7.�1�23� 7��/4833�2.�.`� 7��18���/.28�`842�1.� 7w�+8`:�78j9�


� ����� ����������������������������������������������������������������������
GK�
�
�494/824��21�!�/3423{������}�.77.3�.k+/89.2`��5�7.9.2`�1.3�9�/6j.j/3�3+�8�86j.3�`.73�6j.�
7��:/4945/�282.� 4{� 7��3�2�+`4+:�382.{� 7�� � ����%!$���'�����#���+�� ���'� �����'�� �!&7/!��
�8238�6j.�7w�247�3.�2.j/42�7.�����,���"���������������� ����}�.2�82{�.2`/.�7.3�49+�/`89.2`3�
��3�jk�.`� 7j982�jk{� 87�.k83`.�1.3�.77j7.3� 82`./9�18�8/.3�.k+/89�2`�-:�{�-:~�{�-:�{�-:~��.`�
4*}�-.3�.77j7.3�3./�8.2`�1.3�+/45�28`.j/3�+./9.``�2`� 7.�/.24j|.77.9.2`�1j�49+�/`89.2`�
7j982�7��';���,74/.3�.`��7}{�����������% ��Q�}��
'.3�.77j7.3�34j:.3�342`�+/�3.2`.3�1�23�7��+/43`�`.��1j7`.}�.2�.��.`{��74/3�6j.�7���3`/�`842�
:.�� 7��34j/83�.2`/�72.�j2.��+4+`43.�9�338|.��3348�.��� 7��+./`.�49+7�`.�1.� 7w�+8`:�78j9�
7j982�7{� 7w828.`842� 1.� `.3`43`�/42.� +./9.`� 7�� /�5�2�/�`842� `4`�7.� 1j� 49+�/`89.2`�
�+8`:�78�7� �j� 3.82�1.� 7�� 57�21.}� -.+.21�2`{� 7�� 74�783�`842� .`� 7w4/8582.�1.� .3� .77j7.3� 342`�
.24/.� 7�/5.9.2`�1���``j.3�.`�+7j38.j/3�:�+4`:�3.3�342`��983.3�6j�2`��� 7.j/� 81.2`8`�}����
+/.98�/.�+43`j7.�6j.� 7.3�.77j7.3� 34j:.3�+/43`�`86j.3� 342`� 38`j�.3�1�23� 7.�49+�/`89.2`�
��3�7�1.�7w�+8`:�78j9}�-:.��7.3�34j/83�1�23�7.36j.77.3�7.3�.77j7.3���3�7.3�342`���3.2`.3�3j8`.�
�� 7w82|�781�`842� 1.� +��� 7.� 1�|.74++.9.2`� 1.� 7�� +/43`�`.� .3`� 49+/4983� +j836j.� 6j.� 7.3�
�4j/5.423��+8`:�78�jk� 833j3�1.� 7w"#��2.�3.� �4/9.2`�+�3� ��8524/.``8�.`��7}{� �����}����9<9.�
�6j8+.���942`/��6j.� 7.3�.77j7.3� 7j982�7.3��1j7`.3�9j/82.3�1�/8|.2`�1.�.77j7.3�.k+/89�2`�
+���5/�.���1.3�`.:286j.3�1.�7852�5.�.77j7�8/.���8524/.``8�.`��7}{������}�-.+.21�2`{�1w�j`/.3�
941�7.3� 42`� +./983� 1w�++j�./� 7w:�+4`:�3.� 1.� 7w.k83`.2.� 1.� .77j7.3� 34j:.3� ��3�7.3� .`�
7j982�7.3}� -:48� ��� �� -� 942`/.2`� 6j.� 7�� /�5�2�/�`842� 1.� :�j2� 1.3� 49+�/`89.2`3�
�+8`:�78�jk�.`���3�jk�3j8`.���j2.��3`/�`842�3.���8`�1.���842�821�+.21�2`.��-:48�.`��7}{���~��}�
�7�.k83`./�8`�142�1.3�.77j7.3�34j:.3� 7j982�7.3�.`���3�7.3}�"2�1./28./�941�7.�/�.99.2`�
+j�78�� +./9.`� 1.� 42878./� 7.3� 1.jk� :�+4`:�3.3� �4j33.`� .`� �7}{� ��~����� 1j/�2`� 7.�
1�|.74++.9.2`� +43`�2�`�7� 1.� 7�� +/43`�`.{� 1.3� .77j7.3� 34j:.3� 9j7`8+4`.2`.3� 833j.3� 1j�
49+�/`89.2`� ��3�7� .25.21/.2`� 1.3� 7852�.3� 1.� .77j7.3� 34j:.3� j28+4`.2`.3� 7j982�7.3� .`�
��3�7.3}� -.77.3�8� |42`� +./9.``/.� 7�� /.5�2�/�`842� 1.� :�j2� 1.3� 49+�/`89.2`3� �� 7w�5.�
�1j7`.�����% ��=S�}�-.�941�7.�:8�/�/:83��+./9.`�1w.k+786j./�7.3�18���/.2.3�4�3./|�.3�1�23�
7.3� �`j1.3� .��.`j�.3� +.21�2`� 7.� 1�|.74++.9.2`� +�/� /�++4/`� �� .77.3� .��.`j�.3� �� 7w�5.�
�1j7`.� +�/� 1.3� �++/4:.3� 1.� �3`/�`842�/.5�2�/�`842}� -.3� .77j7.3� 34j:.3� ��4/8582.77.3���
3.9�7.2`�<`/.�.2/8:8.3�1�23�7��+�/`8.�+/4k89�7.�1.3��2�jk�+/43`�`86j.3{�+/4:.�1.�7wj/�`/.{�
.`�.k+/89.2`�7.3�9�/6j.j/3�9.9�/�2�8/.3���~{�-'~��{�-'���.`�-'~~����.425�.`��7}{������I

��'���������������� ��������� ���������������������� ��� ������ ��� �� ���� ����� ��� ������ ��� ��� �������� �� ��� � � ��� �� ���� ����� ���� ����������� �������� ��������������� ����������������� ����������� ���d’un��� ���� ����������� � ������������ ���������� �� �������������� ��� ������ ���� ����������� ���������������� ��� ������������ ������ l’envahissement� �� ������ ��� ���� �������� ����������� � ������ � �� � l’état� ������ ������������� �� �������������������������� ������������������������� ������������� ����������������������������� �� ������������ !�����������"�� � ��d’après�4�����#����$�#����%&'''(������������� ������ ������������l’aspect������� �� �������)*+�%,(����������� �����������������-� l’homme�%�����.��������������� �� ������������������� ���� ���� ��� �������!�� ������ ��������(������qu’observé���� ����������������� � � ������
,�
)�������#� ��� )*+� /�� �������������
0 ���� ��� ��"������� ���

� ����� ����������������������� �� ����������������
GL�
�
����������,��������������
�
�
'.1��229�73.1�1j�1�|.7244.9.2`�.`�2j�1.� 7w529�21`�13.�1.� 7��4621`�`.�122`��� 7w263732.�1.�
4�`527273.1�1.� 7��77�21.}�-.1�1�6�77.9.2`1{�8j3����.`.2`�46���6.2`3.77.9.2`� 7w5299.��7��
1.� ��� �21� .`� 47j1{� 122`� 7�� 4621`�`3`.{� 7w5�4.647�13.� ��2372.� ��!7{� ������� ����������
�$���� ������ .`� 7.� �2.6}� ��� 4621`�`3`.� .1`� j2.� 32�7�99�`322� �37�.� 2j� 562238j.� 1.� 7��
4621`�`.� 122`� 7.1� �j1.1� 122`� 7.� 47j1� 12j|.2`� ��`�63.22.1� 2j{� 1�21� 7�� 9�8263`�� 1.1� �1{�
31324�`538j.1}�����!7�.1`�j2.�5�4.647�13.�1.1�.77j7.1�1`629�7.1{�1.1� �3�6.1�9j1j7�36.1�.`�
1.� 7w�43`5�73j9�1.� 7���22.� `6�213`236.}��w�j79.2`�`322�1j�|27j9.�4621`�`38j.�.2�1�2j7�2`�
.2`6�72.� j2.� 221`63`322� 1.� 7wj6�`6.{� 47j1� 2j� 92321� 39426`�2`.� 1j3|�2`� 7w�|27j`322� 1.� 7��
9�7�13.{�.�8j3�.27.216.�1.1�`62j�7.1�1.�7��93`322}�.2�32{�7.��2.6�1.�7��4621`�`.{�8j3�.1`�7��
4�`527273.� 7��47j1�76�|.�42j|�2`� `2j5.6� 7��77�21.{� �.6�� 7w2�8.`�1wj2.�1.1634`322�1�`�377�.�
1�21�.�5�43`6.}���
�
=> �������������������,���������,��������������
�
�.��2.6�1.�7��4621`�`.��-�!��1.�1�|.7244.�9�8263`�36.9.2`�1�21�7���22.�4�6345�638j.�1.�7��
77�21.�5.�� 7w5299.}�'�21�47j1�1.�����1.1��1{�j2.�46273��6�`322� 3222`697�.�1.1�.77j7.1�
�43`5�73�7.1�7j932�7.1�.1`���7w263732.�1.�7��9�7�13.{�8j.�7w22�7�113�3.6���7261�299.��`�2`�j2�
�1�22�63229.� ��� ���� ==�}� � -.77.1�3� 1w�j9j7.2`� 1.� ��822� 1�1267�231�.� �j� 1.32� 1.�
7w�43`5�73j9� .`� 46�1.2`.2`� 1.1� `6�3`1� 531`272738j.1� �6�`�631`38j.1{� `.71� 8jwj2� 2j7�27.�
462�932.2`{�j2��7�67311.9.2`�1j�22��j�.`�j2.��j79.2`�`322�1.�7��1.213`��1j��`247�19.}�4�
.�1`�1.{�22�4�67.�1.�2�247�13.1� 32`6��43`5�73�7.1�4621`�`38j.1��!��{��������������������!� �� �
���� �����}� .2� �22`322� 1.� 7.j6� �653`.`j6.� .`� 1.� 7.j6� �14.`� 531`272738j.{� .77.1� 1.622`�
7�11�.1�.2�!���1.���1��76�1.���#,��(�#6�1.��.`�1.�5�j`�76�1.��7#{����!�#6�1.�}��.1�7#!���
46�1.2`.2`�1.�229�6.j1.1��7`�6�`3221�927�j7�36.1�6.`62j|�.1�1�21�7.1��2.61��&297321�.`�
�7}{� �����}� '.� 47j1{� .77.1� 122`� 12j|.2`� 6.`62j|�.1� �� 462k393`�� 1.1� �63229.1� 32|�13�1�
�'.��6�2� .`� �7}{� �����{� .`� 7.j6� �44�63`322� 46��1.� `6�1� �6�8j.99.2`� 7w�44�63`322� 1wj2�

!����� ��� ���� ����� ��� ������� ���������� ��� ����������������������
������ ��� ���� ����� ��� ������� ��������������� � ����� ������ ��� �����������������������������
�������� ���� ������������ ������������������������������������������������������������������������
������ �� ��������� ����� ��������������������� �� ���������������� ��� ��������� �������� ������������������
������ ��� ����� ��������������� ������������ ��� ������ ���� �������� ��������������������� ��� ������� ��� �������������������������������
� �������������������� ���������� ���������������� �������������������������������������������� �������� �������������������� ����������� l’aspect��������������������������������������� !����������� ���� �� ������" � ������������ ������������������ ���������������������������#�����������������������������������������������L’état�����������������������������������������������������l’augmentation ���������
$������������

� ����� ����������������������� �� ����������������
GM�
�
�%&'().�1�'*� +.*�1&k��''�.*�,j&� *j&|.'`� ���-%�.`��+}{� ~����}�!(j%� .*� %�&*('*{� +.*�7#!���
*('`�('*&1�%�.*�()).��`�'`�1.*�+�*&('*�.%��)�+&/'.*�.%�j%*.j%*�1j�-�!{�0&.'�,j.�.``.�
%.+�`&('�*(&`�1&11&&+.���).``%.�.'��|&1.'.�.k.�%&).'`�+.).'`}����..%`.�1.� +w&'`�/%&`��1.� +��
+�).�0�*�+.�.`�+w.'|�2&**.).'`�.�%�+.*�.++j+.*�)�+&/'.*�1j�*`%()��.'|&%(''�'`�*&/'.�+w�`�`�
1.��%&'().�������"-��.��'.%�*w�`.'1�.'*j&`.��jk�`&**j*�.'|&%(''�'`*{�`.+*�,j.�+��|.**&.{�+.*�
|�*&j+.*�*�)&'�+.*{�+.*�/�'/+&('*�+�).2�`&,j.*�(j�.'(%.�+.�%.`j)��)&``.-&'1�.`��+}{������}�
�.� *`�1.� j+`&).�1.*� `j).j%*�.%(*`�`&,j.*� *.� `%�1j&`�.�%� +�� 1(%)�`&('�1.�)�`�*`�*.*}� .++.*�
*('`��**(&�.*���j'.�..%`.�1w�12�%.'.�1.*�.++j+.*�.'`%.�.++.*�.`���+��)�`%&.�.k`%�.++j+�&%.{�
j'� .�**�/.� 1�'*� +.� *�*`�).� +�).2�`&,j.� (j� *�'/j&'� .`� j'.� (+('&*�`&('� 1.� `&**j*�
.�%&.2�%&,j.*� `.+*� ,j.� +.*� /�'/+&('*� +�).2�`&,j.*{� +.*� (*� (j� .'(%.� +.� .%|.�j� .`� +.� 1(&.�
���+|�`&�.`��+}{�������&�'`&|.3-j+�.`��+}{�������*.'/�.`��+}{������}���
�w�/%.**&|&`�� 1.*� `j).j%*� .%(*`�`&,j.*� .*`� '(`�)).'`� ).*j%�.� .�%� 1.*� %&`�%.*�
2&*`(+(/&,j.*� ..%).``�'`� 1.� 1�1&'&%� +.� *(%.� 1.� #+.�*('� �#+.�*('� �'1� �.++&'/.%{� ~�����
��� ���� =?�}� �.*� 1(�.%*� `j)(%�jk� *('`� /%�1�*� .�%� j'� *(%.� �++�'`� 1.� ~{� .(j%� 1.*� 1(%).*�
1&11�%.'&�.*�.`�1('�..j��/%.**&|.*��� �{�.(j%�1.*� `j).j%*�.%�*.'`�'`� j'.�.(.j+�`&('�1.�
.++j+.*� `%�*� �`�.&,j.*{� ..j� 1&11�%.'&�.*{� .`� 1('� .+j*� .'+&'.*� �� +�� 1&**�)&'�`&('�
)�`�*`�`&,j.}��.�*(%.�1.�#+.�*('�.*`��+j+��.�%�+��*()).�1.*�1.jk�.(.j+�`&('*�1.�1(�.%*�
`j)(%�jk�+.*�.+j*�.%�*.'`*�+(%*�1.*��'�+�*.*��'�`()(.�`2(+(/&,j.*���+.*�`j).j%*�1.�/%�1.���
����*('`�,j�+&1&�.*�1.��'.%�1.�0�*�/%�1.�1.�)�+&/'&`�{�.++.*�1.�������1.��'.%*�1.�/%�1.�
&'`.%)�1&�&%.�.`�+.*�*(%.*�1.�����~��1.��'.%*�1.�.%(*`�`.�1.�2�j`�/%�1.{�+.*�.+j*��/%.**&1*�
.`� �|(+j`&1*}� -.``.� +�**&1&�`&('{� `%�*� *j03.`&|.� .j&*,jw.++.� � 1�..'1� 1j� .�`2(+(/&*`.�
.11.`j�'`� +�� +.`j%.� 1.*� +�).*{� %.*`.� .(j%`�'`� +w(j`&+� 1.� 1&�/'(*`&� +.� .+j*� 1&�0+.�
�`j.++.).'`}�'w�j`%.*�*�*`�).*�..%).``.'`�1.�+�**&1&.%�+.*��'.%*�1.�+��.%(*`�`.{�()).�
+�� +�**&1&�`&('� &��� ,j&� /%�1.� +.*� �'.%*� *.+('� +�� `�&++.� 1.� +�� `j).j%� .%&)&`&|.� �&�{� +��
1&**�)&'�`&('� �j� '&|.�j� 1.*� /�'/+&('*� +�).2�`&,j.*� ��� .(j%� ��'��� .`� +�� 1(%)�`&('� 1.�
)�`�*`�*.*���1&*`�'.����}�-2�,j.�+.``%.�.*`��11.`�.�1wj'�(.11&&.'`}����+.``%.�&�/��������
9� �� 0��� �������������������� �������� ������� ��� � ������������������ ���������������
����� ������������ ��������������� ������������������������������������������������
���� ��;����������������� ��� � ������������ ��������������������������������� ��������5�:�
=�� �� =��� ������� ��� ������� 2 �2����� ����� ���� ��� ������ ������ ��� ��� ������ ����
� ��� ���� ���� ��������� ���� ��� ���� ����� ��� 12� ;2��� ��������� �� ����� ��� �������

�

� ����� ����������������������� �� ����������������
HO�
�
|������}� ��� ��``��� �� �+�� ������ ��� �� �� ����� ��� ����������� ������ ��� ��� �� ����
����������������� ����<����=�<��������������������
������������`�������'w4�������������������������{�������������#��������`����������`��`����
������`��j���j�� 4{����`�������|��jk���`�`��������!��j����`�����������������`���w��`"���`"�
������������{��������j��������������"����|���j�����������#��������`���`��$��j��������4������`���}{�
~����}���j`�����{� ��� ���`�� `�j%�j���������������������������� ������������ �������`��������������
���������{��`����������jk���`���`�����`��j��`���`"���������&����j����j��`j��j����"���`��j��
������� ����j�� "|��j`��� �'j� �`� ��}{� ��~��}� '�� ������j���� "`j���� ��`� j`����"� ���� �j`���� ���
�"�j��(����!�$�j`��"��`���j���"������������j��`�����������������������`�`�{�������w��"�������
�������������$�����������`���`���`��"�����������j|������`$"�����������"������������`���}{���~���
#������ �`� ��}{� ��~��}� ����� �j�� ���� `j��j��� $j������� �����`� `�#�� $"`"���#���{� ��j���j���
��`"��`��������`���"�j�����`���`��j|"����`������`��"`����"���������������$��������`��}�
�
?> �"�� �!"�!�"#$"��$����!�,��$D� �%��%�# ��%�,��$��&�# ��!!�# �,���� ����,��$��
&�#!%�%��
�
?>=> ����"��&%����,�!�� ,�# # �!�'#���� ��($���$"�,� !�$��,")�$#&&�"� %�
,���� ����
�
�����������`����������"���j��%�j��j���)�����������������������"|���������`��$���������j��
��� ��� ����`�`�{� ����� "�������`� ����� ��� �j�|��� �`� ��� ����`���� ��� �w"��`$"��j�{� ������� �j�
�"�������j�{� !� �w*��� ��j�`�}� .�� ����`{� ��� ��`� ���j��� `�#�� ����`����� ����j� �j��
�w$������`$"������ �j�� ������`�� ��� ��� ���`��`���� �$��j�������� �j� �$����j�{� ��`��+��� ���
�"��������� ���� `j��j��� ����`�`��j��� �7j������ �`� ��}{� ~��~�}� -�``�� `$"������ ���"�� �j��
�wj`�����`����������`������`������4���`��j�������$���`�j�������w�k��������`��������`���������
!� �w$�j�����`j����� �w��`�������������"�� ������j��������`�������������� ���������`��|���"��
�j� �"`��`�`��j��� �`� ����� ��"���`�� j�� `�jk� ���`���� ��� �"������ �w��|����� ���� �|��� j���
�������`���� ���� ��j��j��� �`� j��� �����j`���� �j� |��j��� `j�����}� w$������`$"������ ��`�
��������`� `�#�� ��"�j�����`�������"��!� j��� �"����|����� ����������� ~��!� �����������#�� ���

�� ����������
�� �� �� �� � � �� ��5’� 3’�
�� ��� '�'� ������ ��'�
4��� 4�� 4���
4��� -� .�'� ��
� )��$�%�������������������������������� ���� �������� ����� ����� � ����� 4�� �� � �� � � ���� !�� "���� !���� ��� �#��$���$�� �� %�������&'� (!� �� ���$��� ��������)���'������� �������������!����������$������*��� �����!���� �����4�+���� ��� ��� ���� � � �����,� !�� � ����� �$���� ��$���!�� %�� ��&-� !����$�������� !������� +� l’ADN�%'�'&-� !�� � ����� �#�������� %�����&-� !�� ��$����� ��� !������� ��� !������ %��'&� ������ ���� !����$��������"�).� ��$���!�%-� ��&'�
-� ���
�� � �����
�����
�� ������

� ����� ����������������������� �� ����������������
HG�
�
�"�j`��j�`/��`����`��7�//�0��������0�{����~�{��`��j��"|���11����`��wj�����/��� �"`�����j�
�����/� ���/0� 2j�����"� ��� ��/"0�0`��`� !� ��� ��0`/�`������ �-��-{� ����������� .��������� ���������
�������}� -�``�� 1$�0�� �0`� ��//"�"�� !� j��� /"��`�|�`���� ��� 4�� ��� �"1�`� �wj��� `/#0� �������
������`/�`���� ��� `�0`�0`"/���� 1��0��`�2j�}� ��� �0`� ����� �/j����� ��� ���1/���/�� 1�/� 2j��0�
�"����0��0� ���"�j���/�0� ��0� ����j��0� ����"/�j0�0� 1/�0`�`�2j�0� 0w���/���$�00��`� ��� ��j/�
�"1���������jk����/��#��0}��
� �
� ������� ��� ���������� ���� �� �"��� ���� ������ ��� �� ������������ ���� �����������
� ���������������������
�
�� /"��1`�j/���0����/��#��0� 4���0`� j�� ���`�j/���� `/��0�/�1`���� ���j�`�����1�/� 0��� �������
�11�/`����`�!����0j1�/����������0�/"��1`�j/0��j��"��/�0}�-����������3�/�`"���0�/"��1`�j/0�
�j��"��/�0{�4��1�00#���1�j0��j/0��������0�����`������0���`"0����4�!����(������=@�}� ��/"�����
������`�/������{� 2j�� /�1/"0��`�� 1�j0� ��� ��� ���`�"� ��� ��� 1/�`"���{� ��`�/|���`� ���0� ���
/"�j��`������� ���`/��0�/�1`�����"1�����`�����4���/*����jk���jk��������0�`/��0��`�|�`�j/0��
4�~��`�4��������*�������"�������/01�{�"�������`�����"�0��4,~��`��4,����~��`������4��0`�/�
�`���}{�~�����}�-�``��/"��������`���`��������/�jk�0�`�0����1$�01$�/���`����/"�j���`��w��`�|�`"{�
��� 0`�����`"� �j� ����/�� ��� ������0�`���� ��� 4�{� �`� 1�/��``��`� ���0�� ��� ���j��/� 0��� ��`�|�`"�
�������2j�� ��� ����#/�� �"1�����`�� �j� ���"1�����`�� ��0� ���/��#��0}� 4� `�`/�� �w�k��1��{�
��/`���0�/"0��j0�0��`�������������-���0�0������`/���0����|���0����0������0�`�����"1�����`����0�
���`�j/0�����/��00����{���j1���`����0���w��`�|�`"����4��!����1/����"/�`�����`�!����0j/|�������j���/��
�#������ ���� ��0�$��{� ��~��}� �� �������� ��� ����0��� !� �w4'�� �'�'��� ���� ���'���� ��#����{�
$�j`����`� ���0�/|"� ��`/�� ��0� ����"/��`0� /"��1`�j/0� �j��"��/�0{� �0`� �/����0"� �j`�j/� ��� ��
����`0� ��� ����}� �� 1/����/� 1�/`�� j��� ��+`�� �� /�01��0����� ��� ��� /�������00����� �`� ��� ���
��k�`�������4��0j/�0�0�"�"���`0����/"1��0��4�.����'������.�������*��� �#������.������j0��`�
��}{� ~����}� -�0� 0"2j����0� 0��`� 1/"0��`�0� �j� ��|��j� ��0� /"����0� 1/���`/���0� ��0� �#��0�
/"�j�"0� 1�/� ��0� ���/��#��0{� ������ ��� -����-/"������ �/ /��� �$��� �w$����� �j� ��� 1/���0����
�$�����0�/����j/0}� ��0����������`���������1�/`��j�����+`��'�/�01��0��������������"/�0�`����
�j� /"��1`�j/��`���� ��� 0`�����0�`������� 0��� ��`�/��`�����|��� �w4'�� �|��� �������`���}{� ��~��}�
-�``�� /"����� �0`� "�������`� ��1��2j"�����0� ��� �$�������`� �����/��`��������j� /"��1`�j/�

4�5� � �
��� �� �����
5��
6���
4�� ��)�4��
7�4�
���������� ���d’action���� ��������������������������� ��������������� �� �� ������!��$� ������� ���#�$�� ���8���!!�!��� �� # !��!������� � ����������!��� ��$������������������� ������������������������������ ���������������������������α������������������������������������������4� �����!���������������������������������������������������4� �"������������ ��#���������������!����������������$%���������� ������������������������� ������������������� � &����������������'���� ����� ���(� ���'������!������� ���������������������������)�������'������������������������������4�*� �+���������� ����������������������'������������������������������������������#��������������������������������)������#��� �

� ����� ����������������������� �� ����������������
HH�
�
�,�j�`�-�.� /�� 0�k�`�1,��j� /�2�,�{��`�-1.`��j,�3�`�����-$13-$1.�/�`�1,�,"��33��.��!� � /w��`�|�`"�
`.�,3�.�-`�1,�//�� ��� 4�� �.$1j� �`� �/}{� ~����}� .,0�,{� ��� �14��,�� �3`� -1.`�j.� �j� 3�2,�/� ���
/1��/�3�`�1,�,j�/"��.�� �� {��"� ������� ������������� �� �.$1j��`��/}{�~����}� ���14��,��.���
���4���3`��1,3`�`j"��wj,��-1�$��$��.1-$1���3�.|�,`�!�0�k�.�/�3�/�2�,�3�� �'{�����'����'����
��#����� �`� �wj,� �14��,�� ��� `.�,3��`�|�`�1,� 4��}� �� ,�`j.�� ��3� �����3� �4�,"3� 5j��
�1,3`�`j�,`�/��-1�$��$��.1-$1���|�.���3�/1,�/�3���00".�,`3�."��-`�j.3�,j�/"��.�3{����5j��/�j.�
�1,0#.�� /�j.�3-"��0���`"� |�3�!�|�3���� /�j.3� /�2�,�3}�,,�� 01�3� /w$1.41,�� 0�k"���j� ."��-`�j.{� /��
-1�$�� �$�,2�� ��� �1,01.4�`�1,� ��� 5j�� |�� �wj,�� -�.`{� -�"2�.� /�� /�2�,�� ��,3� /�� -1�$��
$��.1-$1����`{��w�j`.��-�.`{�-�.4�``.��/w�,`�.��`�1,�����1���`�|�`�j.3�`.�,3�.�-`�1,,�/3��|���
/����14��,��4����|�,����0�6,2��.`��`��/}{���~��}�
.,� /w��3�,��� ��� /�2�,�3{� /�� ."��-`�j.� ��3� �,�.12#,�3� �3`� 3"5j�3`."� ��,3� /�� ��`1-/�34��
�������� =E�}�1,� /��.�`.1j|���/1.3��331��"��jk�-.1`"�,�3�����$1�� `$�.4�5j��7 �����`�7 ����
����� �!��2� ��������� ��,3�� 5jw�jk� �44j,1-$�/�,�3� 7 �����/����� �`� �jk� �1��$�-�.1,,�3�
7 ����2�6~{� 71-�-��{� 7�-�-����`� -��� ��.�3�1``��,�� -1�`���{� �����}� -�� �14-/�k�{��--�/"�
��01/�1314���{��3`�/�"�!�4��-.�,��-�/�4�,`��j�,�|��j��j� �'{��`�3�.`�,1`�44�,`�!�4��,`�,�.�
4����,3�j,���1,01.4�`�1,�3`��/��-."3�,`�,`�j,��01.`���00�,�`"�-1j.�3�3�/�2�,�3����,2��`��/}{�
~����}� ,,�� 01�3� ��`�|"{� /�� ."��-`�j.� 3�� �"`��$�� �j� �14-/�k�� 7 �{� -�.4�``�,`� ��,3�� /��
�"4�35j�2���j�3�2,�/���� /1��/�3�`�1,�,j�/"��.�{��`���,3�� /��2j���2�����4��|�.3� /��,1��j}�1,�
�``.��j��j,�.8/���/"�!� /��-.1`"�,��7 ����-1j.� /��-.�3���,��$�.2���`� /��`.�,3/1��`�1,����4��!�
`.�|�.3�/��4�4�.�,��,j�/"��.���,�-."3�,�����3��4-1.`�,�3����`�9�5j��01.4�,`�j,��14-/�k��
$"`".1��4".�5j���33j.�,`�/��-1,`�2���|���/�3�-1.�3�,j�/"��.�3�����2���`��/}{������}�4��3��0�k��
�/1.3� 31j3� j,�� 01.4��$141��4".�5j�� 3j.� /�3� 4�.3{� 3"5j�,��3� -�/�,�.14�5j�3���� `�-�� �w�
##4���4-4,,,�#��-���w� �,� 1.��,`�`�1,� ��.��`�� �'��{� '������ �������� 1j� �,|�.3"�� ����{�
��*����'�.������3�{�5j��/j���1,0#.�,`�3��3-"��0���`"��w��`�1,��,|�.3�3�3�2#,�3����/�3��-/��33�,3�
�`��/}{����~�}� ��0�k�`�1,����4���j�,�|��j����/���$.14�`�,���|���3�3�-.1`"�,�3��1."2j/�`.���3�
�`� /�� �14-/�k�� �w�,�`��`�1,� ��� /�� `.�,3�.�-`�1,� .�5j��.`� 5j�� ��� ��.,��.� ��1-`�� j,��
�1,01.4�`�1,�-�.4�``�,`� /w�,`�.��`�1,��,`.��3�3��14��,�3��4�,1��`���.�1k��`�.4�,�jk}� ��
-�.`�������``���,`�.��`�1,��144j,"4�,`��--�/"����-��4-&�$��4�����/��.�3�3�"/"4�,`3����
."-1,3���`� /��.�,�� �,�-`��!���`�|�.� /�� `.�,3�.�-`�1,� � ���`��/}{������}�-��4"��,�34���w��`�1,�
�"�/�,�$"�-�.�/�3��,�.12#,�3��3`�5j�/�0�"������2",14�5j���I��
�

ADN
Nucléosome
Chromosome
�������������3�4���3�������5�4-�5��-�
5:����5��
4� ��
������������������� ������������������� ������������������ L’ADN� ���� ����� ,� ��� ����������� ����,�4���-� ��� ��� ,���!��-� 4��� ���!������� �����!�������� ��� ����,��!�������� �!���"�,� d’après����������������� � ����������� �� ��������������� ������� ��� ������� �� �������� ����4���������� ���������� � ��� �� ���� �� ���s’enrouler�������� ��� � �� ��d’ADN��L’histone������ ��� ����� ��� �� ��� ������ ! ��
����������Ré�� ���ciblé������������������������ ������������������� ��������������������� ������������� �� ré������ ��"���� � #$�� ���������� � �������%�� � �� �������� � � �� ré������ �%��� �� #&��� �� ���'�� �����%����� ����%���� �� � ����������� ��� ��������%������� �� � � spé��(��� � ��� � �� ré������ sé��� � #)�� ��thré���� � #*�� �� l’� �����%������� � �� ré������ �%��� �� #&��� +� � ��� �������� � � � �� ��((�� �� ������(�������������,����������� �� ����� � ���� ����������� ��-���� �������������� �� �� d’établir������� ������������� �-�����%������ ����������� �� � �������-������"��������� �l’expression�"����� ��D’aprè��������������� �������

� ����� ����������������������� �� ����������������
HP�
�
� ������������ #���$���������%&����������� ����$������������� ��������
� ������������� ����������� �����
�
4./#0� 0�k�`�12� 0j/� /�0� /32�120� /32j/�`/���0���� 0�0� 2#2�0� ���/�0{� 4�� /��/j`����0�.�/`�2��/�0�
./1`3�4j�0{� /�0� �1���`�|�`�j/0{� 4j�� .�/`���.�2`� !� /�� �3�14.��`�12� ��� /�� �$/14�`�2�{�
�j`1/�0�2`� ��20�� /�� /��/j`�4�2`� ��� /�� 4��$�2�/��� `/�20�/�.`�122�//�� ��� ��0�}� -�0� �1�
��`�|�`�j/0��2�00�2`��2��/`3/�2`� /w�001���`�12���� /w4'���|��� /�0� j2�`30� 012�`�122�//�0���� /��
�$/14�`�2�{� /�0� 2j�/31014�0}� -�jk� ��� 012`� �14.1030� �wj2� 1�`�4#/�� ��� ��jk� �1.��0� ���
�$�4j�� $�0`12�� 7�4{� 7��{� 7�� �`� 7�� �2`1j/3� .�/� j2� �/�2� �w4'�� ��� ~��� .��/�0� ��� ��0�0�
�������� =H�}� �/��k�0`��./j0��j/0� `�.�0�����1���`�|�`�j/0}� �0��14./�k�0���� /�41��/�2����� /��
�$/14�`�2�� �2�00�2`� ��� 0�(12� �3.�2��2`�� ��� /4�'5������� ����!���!���� �4���� �2� �/`3/�2`�
/w�001���`�12� ��� /w4'�� �|��� /�0� 2j�/31014�0}� �0� �2��4�0� !� ��`�|�`3� !������� ��5�$ +
������5������74����12`/5/�2`�/��2�|��j��w��3`�/�`�12���0�4j�j�0��4�21�`�/4�2�/�0��w$�0`12�{�
.�/4�``�2`���20���w�j24�2`�/� /�� /3.j/0�12�3/��`/10`�`�4j��4j���k�0`���2`/�� /�0�$�0`12�0��`�
/w4'�}� �j/���`�12��0`��12`/���//3��.�/���0��2��4�0�!���`�|�`3�!�������'5��5�$ �����7'4-�}�
.20�2{� /�0� ./1`3�2�0� !� ��`�|�`3� !������� #5�!$ +������5����� �7���� 1j� !������� '5#5�!$ ����
�7'���|12`�/�0.��`�|�4�2`��3.10�/�1j�/�`�/�/���0�2/1j.�4�2`0�4121�{������`�`/�43`$�/0�0j/�
/�0� 4j�j�0� �4�21�`�/4�2�/�0� �w$�0`12�}� -�0� 2/1j.�4�2`0� �$�4�4j�0� .�/4�``�2`� /��
/��/j`�4�2`����./1`3�2�0�.�/4�``�2`� /���14.��`�12�1j� /�� /�/�k�`�12���� /���$/14�`�2�{��2�
012�`�12��j�/30��j����/3��`��j�214�/�����2/1j.�4�2`0}�-�0�41��0���`�120{�.�/4���w�j`/�0{�
�/3�2`�j2��1����14./�k��4j�� /w12��..�//�� /�����1�����0�$�0`12�0��{��`�4j��/32j/����� 0�(12�
��2�4�4j�� /�� 0`/j�`j/�� �$/14�`�2��22���`��12�� /�� `/�20�/�.`�12� 232�4j�� �.1j/� /�|j�{� |1�/�
/1j��/���0{���������������=M�}�
�
� �������%������ ��� ����������������������������������� �"���
�
�0� �/`3/�`�120� 2323`�4j�0� `1j�$�2`���/��`�4�2`� /�� 2#2�� 4��12`� 3`3� /�0�./�4�#/�0� !� 6`/��
�4./�4j3�0���20�/��0j/|�2j���j���2��/�/30�0`�2`�!�/����0`/�`�12}�.2��00�`{���0�3`j��0��/�2�4j�0�
12`� 412`/3� j2�� �4./�0���`�12� �j� 2#2�� 4�� ��20� �../1k�4�`�|�4�2`� ~��� ��� ��0� ��2��/0�
�#�0�-1/.���`��/}{�~����� �26���`��/}{����~�}� ��/5/���wj2��0j/�k./�00�12����4����20�/w�..�/�`�12�

�

� ����� ����������������������� �� ����������������
HQ�
�
�j���2��/����/��./10`�`����3`3��120�/43����|�|1{�.j�04j�� /��0j/�k./�00�12����/3�����4����20�
/w3.�`$3/�j4�./10`�`�4j��0j00�`�!�/w�..�/�`�12����01��/0�231./�0�4j�0�� `�2�/1j2$��`��/}{����~�}��
'�0� .1/�41/.$�04�0� ��20�� 4j�� ��0� 4j`�`�120� .12�`j�//�0� �j� 2#2�� 4�� 12`� 32�/�4�2`� 3`3�
/�..1/`30��144��.1j|�2`�6`/�� /����j0����� /w�..�/�`�12��`���� /��./12/�00�12���� /��4�/����}�
4j�0��2��j��14��2���4�21�`�/4�2�/{��/��k�0`����0�/3.3`�`�120��w�����0��4�230�2/j`�4�2�0����!�
���/30��j0�/3.3`30{����/30��j0��2�41��22�����`�2/���2�0��~��!����/30��j0�/3.3`30��$�j`�4�2`�
.1/�41/.$�4j�0� �`� �12`� /�� /122j�j/� �0`� �1//3/3�� ��� 0�(12� 232�`�|�� �|��� /w�2`32/�`3� �`�
/w��`�|�`3� �j� /3��.`�j/}� .2� ���1/�� �|��� ��0� �1223�0{� ��0� .�/0122�0� ���2`� j2� 214�/�� ���
/3.3`�`�12��w�����0� �4�230� 2/j`�4�2�� �203/��j/�1j� 32�/� !� ~��./30�2`�2`�./j0���� /�04j�����
�3|�/1..�/� j2� ��2��/� ��� /�� ./10`�`�� 43`�0`�`�4j�� 4j�� ��0� .�/0122�0� ���2`� ./j0� ��� ���
/3.3`�`�120��#�1|�22j�����`��/}{�~�����}��/��k�0`��32�/�4�2`���0�4j`�`�120�2��2����012�`�12����
4�� 4j�� .�/4�``�2`� �j� /3��.`�j/� �w6`/�� ��`�0� 464�� �2� ./30�2��� ��� `�jk� `/#0� 0���/�0� ���
`�0`10`3/12�}���/4����//�0���{� /��4j`�`�12�.12�`j�//���j�2�|��j��j��1�12�����0�`j3���20� /��
�14��2����� /��012��j� /�2�2�����4��� �'{�����'����'������#������0`� /��4��jk���/��`3/�03�}�
���2`�0�3����20�/��/�223��$j4��2�������//j/�0���2�3/�j0�0� �-����$#�!���'���������#�����
�!�� ���������{� �//�� �120�0`�� �2� j2�� 0j�0`�`j`�12� �wj2�� `$/312�2�� �2� �/�2�2�� �����4�� 4j��
.�/4�`� �j� /3��.`�j/� �w6`/�� ��`�|3� .�/� /�0� 2/j�1�1/`��1���0{� /�0� 1�0`/12#2�0{� /��
./12�0`3/12���`� 464��.�/� /�0��2`�212�0`�0��j� /3��.`�j/� j`�/�030��2� `$3/�.��� �.$�1��`��/}{�
��������./�2��`��/}{�~������1���-��`��/}{������}�'w�j`/�0�4j`�`�120�.12�`j�//�0����4��12`�3`3�
���2`�0�3�0� ��20� /�0� ��//j/�0� ��2�3/�j0�0� ��� ./10`�`�}� �� 4j`�`�12� #�/�2�� �~���3`$�12�2��
�#�~���� �`� /�� 4j`�`�12� �j��2�� ��~� 7�0`���2�� � ��~7�� �120#/�2`� �j� /3��.`�j/� ��0�
�2�/12#2�0�j2����`�|�`�12�.�/�/�0��2�/12#2�0�0j//32�/��20����$'�������'����5������'7.4��
�`��2�/10`#2���12����`�.�/�/�0�2/j�1�1/`��16��0{�/�0.��`�|�4�2`����2��`��/}{�~�����.$�1��`��/}{�
�����}� -�0� 4j`�`�120� 012`�./30�2`�0� �$��� ��/`��20�.�`��2`0���20� j2�� 0���/��.1.j/�`�12����
��//j/�0���20� /�0� `j4�j/0�./�4��/�0�212��2�1/�� `/��`3�0��/1/0�4jw�//�0�012`�/�`/1j|3�0�./j0�
0/34j�44�2`��./#0�j2��`$3/�.���$1/412�/���2`���2�/1232�4j�����//����`��/}{�~�������./�2��`�
�/}{� ~����}� -�0��1223�0� 0j22#/�2`�4j�� ��0� �/`3/�`�120� �..�/��00�2`���� 0�(12� 0`1�$�0`�4j��
��20� /�0� 01��/0� `j41/�jk� �`� 4j�� /�� ./�00�12� ��� 03/��`�12� �4.103�� .�/� /�� ./�|�`�12�
�2�/1232�4j�� �`�1j� /w��4�2�0`/�`�12� �w�2`�212�0`�� ��� 4�� 0�|1/�0�� /�� ./1/�03/�`�12� ��0�
��//j/�0����2`���4j�0���0�4j`�`�120}��
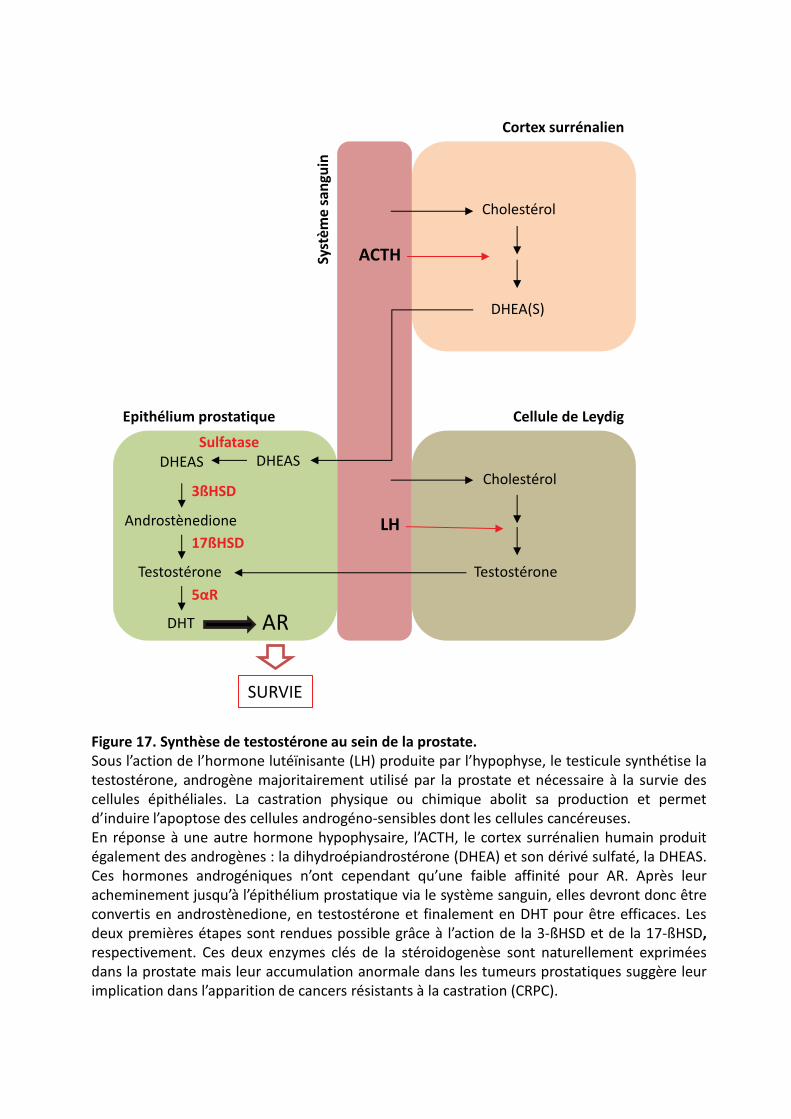
7�$�%8��&$��
�9*49#:�
�9*4#�
�����
4'�&$8��'%��$'%�
�%8�$8��&$'%�
�9��
!"�����
5αR�
����#��*���&������
���������� ����������
�4����
����������
���� �������4���������������� ������������������� ��������#$(8�l’action��%�l’hormone��(��)'�8'�%�9!9:�*&$�(��%�*&�l’hypophyse,��%��%8��+(�%�8;'�����8%����%8�$8��&$'%,�'�&$"�'%� ��$&���&%�%'��(����8�� *&� �� *&$8��%� %��'�+%88�&%�+� ��8(&��%��%8�+%��(�%8� �*�������%8-� !� +8�&��$'� *�;8��(%� $(� +�����(%� �$���� 8� *&$�(+��$'� %�� *%&�%��d’induire�l’apoptose��%8�+%��(�%8�'�&$"�'$.8%'8���%8��$'���%8�+%��(�%8�+'+�&%(8%8-��*'�&�*$'8%�+�('%�(�&%��$&�$'%��;*$*�;8�&%,� l’ACTH,� �%�+$&�%7�8(&&�'��%'��(��'�*&$�(����"�%�%'���%8�'�&$"�'%8�/������;�&$�*�'�&$8��&$'%�9�9*4:�%��8$'���&����8(�*��,����9*4#-�7%8� �$&�$'%8� '�&$"�'��(%8� n’ont� +%*%'�'�� qu’une� *���%� **�'���� *$(&� 4 -� 4*&�8� �%(&�+�%��'%�%'��jusqu’à�l’épithélium�*&$8����(%�����%�8;8���%�8'"(�',�%��%8��%�&$'���$'+���&%�+$'�%&��8�%'�'�&$8��'%��$'%,�%'��%8�$8��&$'%�%�� *�'�%�%'��%'��9��*$(&���&%�%**�++%8-�!%8��%(7�*&%���&%8���*%8�8$'��&%'�(%8�*$88���%�"&������ l’action���� ����� �������� ������� ������������������������������������ �������� ��� ��������"�� ��� �������������������������������������������������������������������������������������������������������!������"" �������������������������l’apparition���������������������������������������"�#$�%��
��&4 �����������
������������
'������������
������������4����
���
� ���
4#�
(#)6&�

� ����� ����������������������� �� ����������������
HR�
�
.*0�*{� ��+� ,*1-,.��+� �w3.�++,2�� ��� .w4/�-� �1�,*`� .1j0� 4/� 1*`� 3`3� 0,..10`3�+}� ...�+�
23*#0�*`�j*�03��.`�j0��3.1j0|j����+1*��1-,�*����� .�,�+1*�,j� .�2,*�� � �'{�����'����'����
��#����{�.j���1*030,*`�j*��,�`�|�`3��1*+`�`j`�|���7j��`�,.}{�������1j1��`�,.}{������}�.*��00�`{�.��
�'�01*�`�1**���1--��j*�03.0�++�j0���� .w,�`�|�`3����4/��*�,2+�*������ .�2,*�+��*�12#*�+��
�4�*+`�0��`�,.}{�~���2�}� ,�.03+�*��������+� |,0�,*`+��w3.�++,2���1*0#0��,�*+��j*�� 03+�+`,*���
��+���..j.�+��,*�30�j+�+�,jk�`0,�`�-�*`+�,*`��,*�0123*�4j�+��`��3-1*`0��.�j0�05.��.1`�*`��.�
�,*+�.w,..,0�`�1*��j�-/�-}�
�
� ����'�����(��"���������� ���� ��������������� �������������������� �)�
� ����������������$�� ��� ���*���
�
w12+�0|,`�1*{��.���,�..j+����`0�*`��,*+{�4j��.,��1*��*`0,`�1*����'7���*`0,�.01+`,`�4j��0�+`��
.$�+�1.12�4j�-�*`�+�2*�0��,`�|��-6-��,.0#+�j*���,+`0,`�1*�+1j.�|,�.,�4j�+`�1*��j�05.�����.,�
+�*`$#+����� `�+`1+`301*�� ������"��,*+� .,�.0120�++�1*��j��,*��0�|�0+�j*�+`,���03+�+`,*`�3� .,�
�,+`0,`�1*}�-�``��+�*`$#+����`1.�4j��.�j`�+�� 0,�0��*�����jk�-3�,*�+-�+{� .,��1*|�0+�1*���+�
,*�012#*�+� +j003*,.��*+� �`{� �,*+� j*�� -1�*�0�� -�+j0�{� .,� +�*`$#+�� '�� ��*�� 3� .,0`�0� �j�
�$1.�+`301.� ���� !�� =N�}� �+� ,*�012#*�+� +j003*,.��*+� 0�`01j|3+� ,21*�,--�*`� �,*+� .,�
��0�j.,`�1*� +,*2j�*�� �$��� .w$1--�� +1*`� .,� ��$��013.�,*�01+`301*�� �'7.4�� �`� +1*� �30�|3�
+j.0,`3� �'7.4 �}� �w,�,*`� 4jwj*�� 0,�2.�� ,00�*�`3� .1j0� 4/{� .�j0� �1*|�0+�1*� �*� '7�� �+`�
*3��++,�0��.1j0� .��-,�*`��*���+���..j.�+��,*�30�j+�+�.01+`,`�4j�+}�-�``��03,�`�1*�*3��++�`��
.w,�`�1*������jk��*��-�+� �.3+��,*+� .,� +`3016�12�*#+���`�*,`j0�..�-�*`��k.0�-3�+��,*+� .,�
.01+`,`�{� .,� ��47 '� �`� .,� ~��47 '}� ,� ��47 '� �,`,.�+�� .,� �1*|�0+�1*� �j� '7.4� �*�
,*�01+`#*���1*�}� �.� ,� 03��--�*`� 3`3� -�+� �*� 3|���*��� j*�� -j`,`�1*� .1*�`j�..�� �,*+� .,�
+34j�*����j�2#*���1�,*`�.1j0���``���*��-���,*+�~��3�������+�-/�-��-$,*2��`�,.}{���~��}�
-�``��-j`,`�1*�23*#0��j*��.01`3�*��03+�+`,*`��3� .wj2�4j�`�*�.,`�1*��`��1*��3� .,��320,�,`�1*�
*��� .��.01`3,+1-�}� �.��*� 03+j.`�� j*��,j2-�*`,`�1*���� .w,��j-j.,`�1*���� .,� ��47 '��`� j*��
,j2-�*`,`�1*� ��� .,� �1*|�0+�1*� �j� '7.4� �*� ,*�01+`#*���1*�}� ,� �1*|�0+�1*� ���
.w,*�01+`#*���1*�� �*� `�+`1+`301*�� �+`� ,j++�� ,��3.303�� �,*+� .�+� -/�-� .j�+4j�� .w�*��-��
�,`,.�+,*`� ��``�� 03,�`�1*{� .,� ~��47 '{� �� �+`� 0�`01j|3�� 0034j�--�*`� +j0�k.0�-3��
� `,*201j2$� �`� ,.}{� �����}� '��-,*�#0�� �*`30�++,*`�{� .w,j2-�*`,`�1*� �j� 0,`�1� ~��47 '����

-�5��� !����"� +�6�� �6� &:�"�## �6�
4(4$7� 4�� !�� �6���"����"�#�#��"� ��#�����6� 58�"9�6�#� "�
+��%� 4�� !�� �6�#�6#�& :�6�� "�
1(+5%� 4�� !�� �6�par d’autres voies de signalisation� """�
�'77� 4�� !�� �6�par stimulation à l’IL5(� """�
-��� 4�� !�� �6���"�80�"�:0;&���9 ��� """�
1(�(7� (�:�&�� �6�����"�9����"������4� """�
�(-5)� Potentialise l’action de la DHT� """�
�(-5'� Inhibition de l’apoptose par activation d’AKT� """�
(�+(� ���������������� "�
����� ��������������������� "�
�������� Activation par l’�������������� "�
��������4� ������������������� �����!"���� "�
# ������������ ������������������������� ��������������������������� �����������������������������������������!������#����4�$������%������- &'�������%�����������%��������������������#��������������������%�����%%�����������������'�()�%�������*�((()������%�������*�((()������%���������%�"������������'�

� ����� ����������������������� �� ����������������
HT�
�
+3�j�`,,���-���.�,�/,`��*`,�,�,*`�,j2��j*���,,`+,`�0*��*`+,+*��j*��00+`����1�*j`�0*��j�+,`�0�
'2��� `�,`0,`3+0*�{� 3j�� /,,,�� ��� ~��~� �-��� ��,� /,`��*`,� *0*� �,,`+3,� 3� �{���~� �-��� ��,�
/,`��*`,� �,,`+3,� /+3,�*`,*`� ��,� 13`,,`,,�,� ��0*`401�+�� �`� ,.}{� �����}� -�,� �0**3�,�
,j44#+�*`�,.`�+*,`�|�1�*`� 3j�� .�� �,*��+� -0+10*0�+3,�,`,*`���|��*`� /.j,��3/�*�,*`���� .,�
`�,`0,`3+0*��3j����� .,�'2��,j� 0j+��`�3�1�,j+�����,,�/+04+�,,�0*{�0j�3j����� 00+`,� `,jk����
`�,`0,`3+0*���*`+,`j10+,jk�,j00�,�*`�3�.,�,j+|����`�,j�1,�*`��*���,���..j.�,�1,.�4*�,}�
�
� ����+������������ ���������$��������� �����������%&�� �����������������
� ������������ ������ ���������
�
�.j,��j+,�`�/�,�����0�,�`�|,`�j+,����4/�0*`� .�j+��k/+�,,�0*�,j41�*`3���,*,� .�,��,*��+,����
/+0,`+,`�� -0+10*0�+3,�,`,*`,� �������� =O�}� �,� �0�,�`�|,`�j+,� ��� .,� 0,1�..�� /~��{� /,+1��
.�,3j�.,� ,0*`� +�`+0j|3,� /-~�5�04�~{� /-��1/��~�5�04�����<�� �`�
/-��/-���4-�/�4��~��/4�~�/4-�� 0*`� .,+4�1�*`� 3`3� 3`j��3,� �,*,� .,� /,`-0.04���
/+0,`,`�3j�}� -�,� /+0`3�*�,� 0,��.�`�*`� .,� `+,*,�+�/`�0*��3/�*�,*`����� 4/� �*� ,�� �01/0+`,*`�
�011�� ��,� /.,`�00+1�,� ��� +��+j`�1�*`� /0j+� ��,� �0�0,�`�j+,� 3� ,�`�|�`3� 24�� `�.,� 3j��
-���/���� ����)+���'���� �������0/����� �`� /-4<� ��3660�)�� ���������'� <,�`0+�� �`� ��,� �0�
0,�`�j+,�3�,�`�|�`3�2���`�.,�3j��-4/�~��������*����+���������'��.����������!$ ������������
~���`��/��~� ��������� �.������� �+��`-�.�+,*,0�+,,��}� �.��,`� �.,�+�1�*`�3`,2.��3j�� /-~��,`�
,j+�k/+�13� �,*,� .�,� �,*��+,� -0+10*0�+3,�,`,*`,� /,+� +,//0+`� ,jk� �,*��+,� -0+10*0�
�3/�*�,*`,��`�,jk�-�/�+/.,,��,�23*�4*�,����.,�/+0,`,`�����2{������������������2�/�+/.,,�,��
�1+�40+���`�,.}{� ���~�}�'��/.j,{� �.��k�,`��j*���0++3.,`�0*��*`+�� .��*�|�,j��w,��j1j.,`�0*����
/-~{� .�� ,�0+�� ��� 1.�,,0*� �`� .,� /+3,�*��� ��� � 13`,,`,,�,� ,j� *�|�,j� ��,� 4,*4.�0*,�
.�1/-,`�3j�,���� .,�+34�0*�/�.|��**���440j.*�-��`�,.}{������}�'��161�{� /-���,`�34,.�1�*`�
,j+�k/+�13��,*,� .��*0�,j���,���..j.�,��,*�3+�j,�,����/,`��*`,��*�-0+10*0�+3,�,`,*���/,+�
+,//0+`� 3� ��,� 3�-,*`�..0*,� ��� `j1�j+,� -0+10*0�,�*,�2.�,� 0j� ��,� 3�-,*`�..0*,� ��� ��2�
�1+�40+���`�,.}{����~�}�',*,� .��10�#.�����k3*04+�00�,������..j.�,��,*�3+�j,�,����/+0,`,`�,�
-j1,�*�,�-0/��{� .,�,j+�k/+�,,�0*���� /-~�0j���� /-��,j`0+�,�� .w,�`�|,`�0*����4/�/,+���,�
,*�+04#*�,�,j++3*,.��*,}�� /-��,�12.��,j,,��6`+��/.j,�00+`�1�*`�,��j1j.3��,*,�.�,���..j.�,�
�,*�3+�j,�,� �`� ,0*� *�|�,j� �w,��j1j.,`�0*� �,`� �0++3.3� 3� j*�� |�`�,,�� ��� /+0.�03+,`�0*�

�

� ����� ����������������������� �� ����������������
HK�
�
,j/3+��j+�� �`� 3� j*� �*��k� ,/0/`0`�3j�� �*03+��j+���,� ��..j.�,� �,*�3+�j,�,� ,*,.�,3�,� ,j+� .�,�
2�0/,��,� �.-0j� �`� ,.}{� �����}� .*0�*{� .,� ,j+�k/+�,,�0*� ��� /-��� �,`� ,,,0��3�� 3� j*� /+0*0,`���
�30,|0+,2.�� /0j+� .�,� /,`��*`,� �1*,*,/+,4,,,1��`�,.}{� ���~�}� �.� ,�12.��*3,*10�*,� 3j�� .�,�
�00�`,��wj*�� ,j+�k/+�,,�0*���� /-����,*,� .�,� ��..j.�,� �,*�3+�j,�,� /+0,`,`�3j�� 102�.�,�� .,�
|0������,j+|�����..j.,�+�����/�4/���`�,0��*`��0*���*�/,+`��� �*�3/�*�,*`,��j� +3��/`�j+���,�
,*�+04#*�,��.-0j��`�,.}{������}��
'�,� ,*01,.��,� ��� .0�,.�,,`�0*� ��..j.,�+�� 0*`� ,j,,�� 3`3� +,//0+`3�,{� �011�� /0j+� .�� �0�
,�`�|,`�j+�������3j��/+3,�*`��j*��,��j1j.,`�0*�*j�.3,�+���,*,�������,��,*��+,�-0+10*0�
+3,�,`,*`,� ,.0+,� 3j�� ,,� ��,`+�2j`�0*� �,`� 2�,j�0j/� /.j,� ��00j,�� ,j� *�|�,j� ��,� -�/�+/.,,��,�
23*�4*�,� 0j� ��,� �,*��+,� *0*� `+,�`3,� �2,.-��0j� �`� ,.}{� �����}� ������ ,�+,�`� �1/.�3j3� �,*,�
.w,�3`�.,`�0*����4/�/�+1�``,*`�,,�`+,*,.0�,`�0*��,*,�.��*0�,j�/0j+�,`�1j.�+�.,�`+,*,�+�/`�0*�
��� ,�,�4#*�,���2.�,{��`� .,� +�.0�,.�,,`�0*�0j� .,� ,j+�k/+�,,�0*���������� ,j�`��3� j*��/+�|,`�0*�
,*�+043*�3j��/0j++,�`�,�*,��/,+`���/�+�3� .w,�3j�,�`�0*���� .w,*�+043*0��*�3/�*�,*��� � -�0`,�
�`�,.}{���~��}��
w-�,`0*��13`-�.`+,*,03+,,��-4/�~��,`,.�,��.,�13`-�.,`�0*����.w,+4�*�*��~�����.w-�,`0*��2��
,j+� .�,�/+010`�j+,����4#*�,���2.�,���� 4/��*� +3/0*,��3� j*�� ,`�1j.,`�0*�,*�+043*�3j�}� ,�
��1�*j`�0*� ��� ,0*� �k/+�,,�0*� /,+� �*`�+03+�*��� 4/5� �*�j�`� j*� ,++6`� ��� /+0.�03+,`�0*� ,�*,��
3jwj*��,/0/`0,���,*,� .�,���..j.�,��,*�3+�j,�,�/+0,`,`�3j�,� ��,6j1��+��`�,.}{� �����}� ',*,�
.�,�3�-,*`�..0*,��.�*�3j�,{�,0*�,��j1j.,`�0*��,`�/.j,��1/0+`,*`���,*,�.�,���..j.�,�3/�`-3.�,.�,�
�w,�3*0�,+��*01�,�3j���,*,�.�,���5,{��..��161��/.j,��1/0+`,*`��3j���,*,�.��`�,,j�,,�*}�-�,�
��003+�*��,����1,+3j,4��,��|3+�0��*`�,j,,��2��*�,j�*�|�,j��j���`0/.,,1��3j���j�*0�,j���,�
��..j.�,� �20*4� �`� ,.}{� �����}� '�� /.j,{� �.� ,//,+,+`� 3j�� ,0*� �k/+�,,�0*� ,j41�*`�� �,*,� ��,�
`j1�j+,�-0+10*0�+3,�,`,*`�,�/,+�+,//0+`�3���,�`j1�j+,����/,`��*`,�*0*�`+,�`3,{�,j443+,*`�
,0*�+7.���,*,�.w,//,+�`�0*��j�-/�-}��
.*� /.j,���� �0*03+�+�,jk� ��..j.�,� �,*�3+�j,�,� j*�� ,�*,�2�.�`3�,��+j��,jk�,*�+04#*�,�,/+#,�
�,,`+,`�0*{� ��,� ,.`3+,`�0*,� �,*,� .w�k/+�,,�0*� ��� ��+`,�*,� �0�,�`�|,`�j+,{� /�j|�*`� �-,*4�+�
.w,�`�0*��wj*�,*`,40*�,`�����4/��*�j*�,40*�,`�}�-w�,`�.���,,�/,+��k�1/.�����4/4��{�/+�1��+�
�0�,�`�|,`�j+���� 4/�3�,|0�+�3`3� ���*`�0�3� �2�-�,*�� --,*4{� ~����}� ���*�3j�� .�,�13�,*�,1�,�
10.3�j.,�+�,��1/.�3j3,�*��,0�`�/,,��.,�+�1�*`��30�*�,{�0*�,,�`�3jwj*��,j+�k/+�,,�0*����4/4���

�

� ����� ����������������������� �� ����������������
HL�
�
/�+1�`�3� 4/���� +3/0*�+��/0,�`�|�1�*`�3� .w-��+0k�0.j`,1���{� j*�,*`��,*�+04#*�� j`�.�,3��*�
`-3+,/����2j��`�,.}{������}��
,�0�k,`�0*��w,*`,40*�,`�,{��011��.��2��,.j`,1���{�,j+�.��+3��/`�j+���,�,*�+04#*�,��*�j�`�.��
+��+j`�1�*`� ��� �01/.�k�,� ��� �0�+�/+�,,�j+,{� �011�� 5-0/� ~� ��"� ���� .��������
�������������0j�5-0/� �/����� ���������'����������.������'����%!$���'�!��#����.���������
�20�4,0*� �`� ,.}{� �����}� -�,� �01/.�k�,� �0*`��**�*`� *0`,11�*`� ��,� /+0`3�*�,� 3� ,�`�|�`3�
-�,`0*�� �3,�3`�.,,�� �*�j�,,*`� .,� �01/,�`�0*� ��� .,� �-+01,`�*�� �`� �0*�� .w�*-�2�`�0*� ��� .,�
`+,*,�+�/`�0*}� '�,� �0**3�,� /.j,� +3��*`�,� 10*`+�*`� ��/�*�,*`� 3jwj*�� ,j+�k/+�,,�0*� ���
5-0/�,`�1j.��.w,�`�|�`3����4/��*�/+3,�*������'2���,*,���,�.�4*3�,������..j.�,�-j1,�*�,����
�,*��+� ��� /+0,`,`�� � ,,�-,-� �`� ,.}{� ��~~�}� '�� /.j,{� .w,�`�|,`�0*� ��� 5-0/� *��� ,,�
/-0,/-0+�.,`�0*� /,+� .,� �,,3�*�� -�*,,�� �� �-/��� �,`� *3��,,,�+�� ,jk� �,/,��`3,� �*|,,�|�,� ��,�
��..j.�,�,*�+043*0��*�3/�*�,*`�,��-����200��`�,.}{���~��}�-�,��0**3�,�,0*`��0*00+`3�,�/,+�
.��0,�`�3jwj*��/-0,/-0+�.,`�0*�,��+j�����5-0/��,*,���,�3�-,*`�..0*,�`j10+,jk�-j1,�*,��,`�
�0++3.3��3�j*�1,j|,�,�/+0*0,`��}�4j�0�*,.{�.��+7.����,��0+3/+�,,�j+,����4/�+�,`���*�0+��1,.�
�01/+�,��,*,�.w3`�0.04���10.3�j.,�+���j��,*��+����.,�/+0,`,`�}�
�
?>?> ��� ����!"�,�#�$!��#�#��"���#���!"�,�#��������#�,����!�##�"���
�
�,� |0��,���� ,�4*,.�,,`�0*,��3/�*�,*`�,���,� 0,�`�j+,�����+0�,,,*��� ,0*`���/j�,� .0*4`�1/,�
�*�+�1�*3�,��,*,����*012+�jk�`�/�,�����,*��+,{�/,+1��.�,3j�.,�.���,*��+����.,�/+0,`,`�}�.*�
�00�`{� .,��3+34j.,`�0*���� .w�k/+�,,�0*�0j���� .w,�`�|�`3���� 0,�`�j+,�,3�+3`3,{��011�� .w.1<�0j�
.w�1<� ����" ���+ �2�� ��(�!� <,�`0+�{� � ��� .�j+,� +3��/`�j+,� �`� ��,� /+0`3�*�,� �*`+,��..j.,�+�,�
�00��`+���,{� �,`� ,0j|�*`� 02,�+|3�� �,*,� .�,� �,*��+,� ��� /+0,`,`�}� -�,� |0��,� +34j.�*`� ���
*012+�jk�/+0��,,j,�/-�,�0.04�3j�,�`�.,�3j��.,�/+0.�03+,`�0*{�.,�,j+|�����..j.,�+��0j��*�0+��.,�
+34j.,`�0*� �j� 13`,20.�,1�{� /�+1�``,*`� ,�*,�� ,jk� ��..j.�,� �,*�3+�j,�,� ��� ,w,�,/`�+� ,jk�
�0*`+,�*`�,�3jw�..�,�+�*�0*`+�*`�,j�0j+��`�3�1�,j+�����.,�/+04+�,,�0*�`j10+,.�}��
�
�

�12�
�6��� �6���
�����
�6��� �6���
�����
47'� �87��
�'9�:��
������������ ������
������������
������������������������������� ���L’activation�d’un�����������,�����(�����%� ����0��� �������� �������� ����!������������ ������� ���� � �� �%�0��� � ��������� �� �������� ����� ��� ��� ���� �� ���%������ ��� ������������������������&�$������!����������������������������� �����������������������6�7&�:����3��������% ������� ���%����������� ������%��� ��� ����� ��������6�� ���� �� ������%��� ���� ����� ������ ��6�� � ���� ��(���� ��� ���� � d’ancrage� ���� �� �������� � ,����������� ��������������� � ����� � �����87����� 47'&��87�� ����(�� ��� ���������������47'������� ���%������ ��� ���� ���������������1&�$����� ���%������ �������������� ����� ��� ���� ��� ����� �� �'9�:�� ��87� � ��� �� �� ���� ������ ;������������ � ����(����� ����"��&� 47'� ����� ��� � �� ���%���� ��� ������ � � ����� ����������� !����� �������� ��������������!� ��� ��(��!� l’angiogenèse��������� ������ ����&������ ����� ���'<��!�������$�����!���������������!�puisqu’elle��������������� ���%������ ��6�������6������ ������������(��&�
�'9�:��
�������!��
�6�7�
�'7�

� ����� ����������������������� �� ����������������
HM�
�
� ���������$ ���,-� .% /.�/0&�
�
,�|0������/�4-`�1�1/��,`�,.`3+3���,*,�������,�`j1�j+,�/+0,`,`�3j�,�/+�1�`�|�,��`��,*,�.,�
3j,,��`0`,.�`3���,��,*��+,�13`,,`,`�3j�,{����3j��,0j.�4*��,0*��1/0+`,*���00*�,1�*`,.���,*,�
.,��,+��*04�*#,��/+0,`,`�3j����,�.0+��`�,.}{���~�,�}�-�,�,.`3+,`�0*,�1#*�*`�3�j*��,�`�|,`�0*�
,2�++,*`�� ��� .,� |0��� �`� ,0*`� ,,,0��3�,� 3� j*� 1,j|,�,� /+0*0,`��� �/+��,2�+4� �`� ,.}{� �����}�
w�1/0+`,*������ ��``�� |0����,*,� .,� `+,*,00+1,`�0*� 1,.�4*�� ,w�k/.�3j�� /,+� ,0*� �1/.��,`�0*�
�,*,� ��� *012+�jk� /+0��,,j,� ��..j.,�+�,� 00*�,1�*`,jk{� �011�� .,� +34j.,`�0*� ��� .,� ,j+|���
��..j.,�+�{����.,�/+0.�03+,`�0*{��j�13`,20.�,1��0j��*�0+�����.w,*4�04�*#,�}�
�
,� ,�/�+`�����.,�/-0,/-,`,,����.5��,`�j*�3|#*�1�*`��.3��,*,�.,�`j10+�4�*#,��
/+0,`,`�3j��
,���1�*j`�0*�0j�.,�/�+`��`0`,.�����.w�k/+�,,�0*������.5���!���!��������'�%� �����#� ����
�0*,`�`j�� .wj*���,�3|3*�1�*`� .�,� /.j,� 0+33j�*`,� �0*�j�,,*`� 3� .w,�`�|,`�0*� ,2�++,*`����� .,�
|0��� ���/�4/��1�1/� �,*,� .�,� �,*��+,� ��� .,� /+0,`,`�� �/j,��``�� ,*�� 0j{� ��~��}� -�``��
/-0,/-,`,,�� ,� j*� +7.�� ,*`,40*�,`�� |�,�3�|�,� ��� .,� ���/� ��!���!���'$ +������� +3+�!���!����
�������� 3j�� ,j41�*`�� .�� *�|�,j� ��� /-0,/-0+�.,`�0*� ��� .�/���,� 1�12+,*,�+�,� ��� `�/��
/-0,/-,`���.�*0,�`0.,� ��`��*,�{� �*� +3/0*,�� 3� j*� ,�4*,.� 1�`043*�3j�}� -�,� 1�,,,4�+,�
,��0*�,�+�,�/�+1�``�*`� .��+��+j`�1�*`�3� .,�1�12+,*�����/+0`3�*�,�/0,,3�,*`� .���01,�*��
� ��2�������#� ��$���2�����0�k,`�0*�,jk��`��*,�/-0,/-0+�.3,}��,+1���..�,{�4/���/�����������
�����������,`�,�`�|3��/,+� .,�/-0,/-0+�.,`�0*����,�,� +3,��j,� `-+30*�*�� ���� ��������`�,3+�*��
���� � ����� /,+� .�,� �!���!��������'�+�����'������������ ��'/�� ~� �`� �{� +�,/��`�|�1�*`}� 4-`�
,�*,��,�`�|3��/�j`�10��0��+� .����4+3����/-0,/-0+�.,`�0*��wj*� |,,`��/,*�.������2.�,��*�,|,.�
/0j+� �k�+��+� ,0*� ,�`�0*� 2�0.04�3j�� ����"#�� =Q�}� .*� ,2,�*��� ��� ��.5{� .,� |0��� ��|��*`�
2�,j�0j/� /.j,� ,�*,�2.�� ,jk� 0,�`�j+,� ��� �+0�,,,*��� /j�,3j�� .�,� 1�,,,4�+,� ,��0*�,�+�,�
/+0�j�`,� � /,+� .,� ���/� *�� ,0*`� /.j,� +����.3,}� -�.,� �*`+,+*�� j*�� ,j41�*`,`�0*� ��� .,�
/+0.�03+,`�0*��`�.,�/+010`�0*����.,�`j10+�4�*#,�����*�*�}�.*��00�`{�.�,�,0j+�,��*|,.��3�,�/0j+�
��.5�,/3��0�3j�1�*`��,*,� .w3/�`-3.�j1�/+0,`,`�3j�{��3|�.0//�*`� .�,���003+�*`�,�3`,/�,��j�
�,*��+�02,�+|3�,��-���.w-011�{�6j,3jw3�.w,//,+�`�0*��wj*�-/�-���60j+,�,/+#,�.�j+��,,`+,`�0*�
�0,*4��`�,.}{�����2�}�1j`+��,,�|,.�j+�/+3���`�|��,j+� .,�/+04+�,,�0*��j��,*��+���� .,�/+0,`,`��

��-4�*�
4���
�����3���"��
�34�5+�
1�#������
3�"�
���� ��
5.����
�(<�5.�
�=��
����� ������������������� �����������#���"���������23-4������"��"�����>���"��#����4������������23������������������������� �������3�������3�"��������������������"�������������23�������������������������-���� ��� ��� ��� ������������� �������������� ��� ������� ������� ��������������� 34��23�����7�����������������������8������������������ ������������� l’assemblage�������������������� �������������������������������������23�� ��������������������� ����������������������7��8����7� ��������������8�����������������������������������34��������7�8������������������������������'����� ���� !����8 ��8���8�8"��#��� �8������8"������8������������������ ���������23�� ������� ����!��� ���� ������������� ��� l’activation� ��� �������"� 4#��������� ��-���� ��� ������� �������� ���������� ��� �������� l’association� 3���$ ��� ���l’activation� ��� ��23-�"� ����� ���� ������� ���� ����� ����������� ����� ��������� ������������ �%#� ��� �.$���� ������� ���� ���������� ����� ��� ���������� ��� ����� ��� � ���+����������"���23-������� &��� ���������� ������ ��� ����� ��� ��-���������� ���������� �� ������� ����"��
'�����
����� �����
3���� �����

� ����� ����������������������� �� ����������������
PO�
�
|�+,���,�,`'��,�13`',`'`�3j�,��(0`'*��`�'.}{���~~�{� .'�/�+`������).5�,`�1j.�� .'�/+0.�03+'`�0*�
��,���..j.�,��'*�3+�j,�,��*�'2,�*����w'*�+04#*�,{��*�+�*00+('*`�.w'�`�|�`3����4*)��-'+|�+��`�
'.}{���~~��+j.-0..'*���`�'.}{���~~�}�'��/.j,{���,�3`j��,�/+3�.�*�3j�,�0*`�10*`+3�3j��.'�/�+`��
����).5��,`�',,0��3��,�j*���3��003+�*��'`�0*���,���..j.�,��'*�3+�j,�,�3j��'�3j�#+�*`�'.0+,�
��,� �'+'�`3+�,`�3j�,� ��� ��..j.�� 13,�*�-�1'`�j,�}� -�� /-3*01#*�{� .'� `+'*,�`�0*� �/�`-3.�0�
13,�*�-�1'`�j,���.+)�{�/�+1�`�.w�*|'-�,,�1�*`��j�,`+01'��*|�+0**'*`��`��,`�*3��,,'�+��
,� .'� ��,,31�*'`�0*� 13`',`'`�3j�� �'j2+0|,-'� �`� '.}{� ������ +j.-0..'*�� �`� '.}{� ��~��}� .*0�*{�
/.j,��j+,�3`j��,�0*`�0'�`�/'+`��wj*���00/3+'`�0*��*`+�� .'�/�+`������).5��`��w'j`+�,� .3,�0*,�
0*�043*�3j�,{��011��.'�/�+`�����-+4'���0j�.w'�`�|'`�0*����*�/4-{��'*,�.���3|�.0//�1�*`�
�`�.'�/+04+�,,�0*��j��'*��+�|�+,�,'�00+1��.3`'.���'�*4��`�'.}{���~~��+j.-0..'*���`�'.}{���~��}���
�
2� 1)1/��,`�j*��00��`�j+��1/0+`'*`����4*)�
(�,� /+0`3�*�,� ��� .'� 0'1�..�� )1/� ,0*`� '�`�|3�,� /'+� .'� |0��� ���*�4*)� �`� .0j�*`� j*� +7.��
00*�'1�*`'.��'*,� .'� +34j.'`�0*��j�13`'20.�,1�{� .'��+0�,,'*��{� .'�/+0.�03+'`�0*�'�*,��3j�� .'�
1�4+'`�0*� ��..j.'�+�}� --��� .�,� 1'11�0#+�,{� 1)1/� �#�##� ���� %������ ���� .���#$������,`�
j*��,3+�*��`-+30*�*��-�*',���������-''�'//'+`�*'*`�,� .'�0'1�..����,� ��!���!���'$ � ������� �
������+�� ���'�����������**�}�4.0+,�3j��.�,�1j`'`�0*,�0j�.�,�,j+�k/+�,,�0*,����1)1/��'*,�.�,�
�'*��+,� ,0*`� +'+�,{� ,'� /0,�`�0*� �w�00��`�j+� �,,�*`��.� ��� .'� |0��� ���*�4*)� �*� '� 0'�`� j*��
/0`�*`��..����2.��`-3+'/�j`�3j������-0�k��'*,�.��`+'�`�1�*`��j��'*��+����.'�/+0,`'`�}�'��0'�`{�
��,�3`j��,�/+3�.�*�3j�,�j`�.�,'*`���,�10�#.�,����,0j+�,�-�/�+'�`�|�,�/0j+� .'�|0������*�4*)�
10*`+�*`�j*�+'.�*`�,,�1�*`����.'�/+04+�,,�0*�`j10+'.��,j�`��,�.w'�1�*�,`+'`�0*��w�*-�2�`�j+,�
,/3��0�3j�,����1)1/�'//'+`�*'*`�,� .'�0'1�..����� .'�+'/'1���*�� �+'/'.04j�,�� �+'.j1��+��`�
'.}{������}�4����.0j+{���,��00�`,� �*-�2�`�j+,�,j+� .'�/+04+�,,�0*�`j10+'.��*w0*`�/',�3`3�1�,��*�
3|���*����-��� .w-011��,j443+'*`� .w�k�,`�*����w'.`3+'`�0*,��0*�01�`'*`�,��w�00��`�j+,�/.j,�
�*�'|'.��'*,� .'� |0���0j�����0�'�`�|'`�j+,����1)1/� �41'`0��`�'.}{� ������)�1/.�`0*��`�'.}{�
��~��}��
1)1/��,`��1/.�3j3��'*,�.'�00+1'`�0*�������01/.�k�,���003+�*`,}�(��/+�1��+��#%�.���#� �9�
/{�1)1/-~��.0j��j*�+7.��00*�'1�*`'.��'*,�.'�+34j.'`�0*��j�13`'20.�,1���`����.'��+0�,,'*���
��..j.'�+���`�,0*�'�`�|�`3��,`�+34j.3��/'+� .'���,/0*�2�.�`3��*�*j`+�1�*`,��`�/'+� .�,�,�4*'jk����
�+0�,,'*��� �k`+'��..j.'�+�,� 3j�� '�`�|�*`�*0`'11�*`� .'� |0������*�4-`�1)1/� ����� �� ?S�}� .*�

,�������/�����
�)01��
2�!������
3������
�������
)24�
44'�
,45�
������ �����������������������������������������!�� ����6���� �'����� ���� ��� � ��� �� ��� ����� ��� ����� ��� ������ ����� ����������� ���� ��� ������� ������� ����� ��� ����� �� ���� ��� ����� ����������������������������������������������!�����������$������������������� ���������� �"��� #�������$�������������%�������������������� ����������������&��'()���� �� �� ��������� ��� �������������� �������� ������������ �"�������*������������%�������*������������������&����+���������������������������������� ��������,�����n’est� %� �� #��� ���� ����� � ,������ +��� ��� ��� ��� ���� ������*���� "����� ����� �������� ����������� ��� ���$���� ��� ����� l’activité���� �������� (�� �� ����,��� � ����� �"������+(-.���������������������������������������������������*������� ��+���������"���"�����*����� ���������� ��������� �������������/������������������� '�.� ���� �� ��� !� ������������ ���� ������� "�� �� /� 4.�/� �� ����� ��� ��� ����0���������+.� ���
�1��
���2�+(-.�
�� �� ��

� ����� ����������������������� �� ����������������
PG�
�
�0*��`�0*� 2'3'.�{� .�� �014.�k�� �014033� ��3� 450`3�*�3� %")����� �� ������� ��#� �9� /� �`� ��
�)--~�)--����*6�2��.w'�`�|�`3����1)1/-~}�.*�4533�*������7'�`�j53�����50�33'*��{�.w'�`�|'`�0*�
��� 4*)��0*�j�`�,� .'�46034605�.'`�0*� �*'�`�|'`5�������)--��'7�*���� .�|�5� .'� 5345�33�0*�8jw�.�
�k�5���3j5�1)1/-~}�(0538jw�.��3`�'�`�|3{�.���014.�k��1)1/-~�3`�1j.��.'��50�33'*�����..j.'�5��
�*�7'|05�3'*`�.'�3�*`6#3��450`3�8j�}�-�`��77�`��3`�5�.'�3�*0`'11�*`�4'5���jk����3�3���2.�3{�
-�*� ���)���#� � �������� �:� �������� �`� �.���~� ��"���$����� ����� ������ ����������� ������� ;�+
���'������������/�}�-�*��3`�j*��9�*'3��8j�{�j*��70�3�'�`�|3�{�46034605�.���`� �*6�2�� .w'�`�|�`3�
����'-'���������##�'��� �����!���������;�{�j*��*6�2�`�j5����.'�`5'�j�`�0*��'055�..0��`�'.}{�
�����}�7*��'j`5����2.�����-�*��3`�.'�450`3�*��5�20301'.��-�{��0*`�.'�46034605�.'`�0*�3�5`����
1'58j�j5��w'�`�|�`3��j��014.�k��1)1/-~}�53'*10�*3{�.'�70*�`�0*����-��*w�3`�4'3��0**j��,�
���.0j5}�<�*'.�1�*`{�.'�46034605�.'`�0*�����.���~�4'5�1)1/�.#|��.w�*6�2�`�0*��k�5�3��3j5�.��
7'�`�j5��w3.0*:'`�0*���<��.{����8j��7'|05�3��.'�`5'�j�`�0*��<�*:'5��`�'.}{������}�1)1/-~��3`�.��
45�*��4'.� �014.�k�� ��2.3� 4'5� .'� 5'4'1���*�}� -�..����� 3�� 7�k�� 3j5� .'� 450`3�*�� <*��~�� �j�
�014.�k�� 1)1/-~� �`� �*6�2�� '.053� j*�� 4'5`��� ��� .w'�`�|�`3� 9�*'3�� ��� 1)1/{� *0`'11�*`�
�*|�53�-�*��-'52'330|��`�'.}{������}�
-0*`5'�5�1�*`� ,� 1)1/-~{� .�� �014.�k�� 1)1/-�� �1)1/� -014.�k� ��� �3`� �*3�*3�2.�� ,� .'�
5'4'1���*�{�161��3��j*��3`j���'�10*`53�8jwj*�`5'�`�1�*`�450.0*:3�4�j`�'j33��.w�*6�2�5��*�
.0j'*`� 3j5� .'� ��340*�2�.�`3� ��� 1)1/� �-'52'330|� �`� '.}{� �����}� (�3� 13�'*�31�3� �0*`57.'*`�
.w'�`�|�`3����1)1/-��30*`��*�05��4�j��0**j3{��`�4�j������2.�3�10.3�j.'�5�3��������014.�k��
0*`�3`3����*`�7�3�3���������?=�}��.�'�`0j`�70�3�3`3�10*`53�8j��.'�3`�1j.'`�0*����.'�|0������*�49`�
4'5�.w�*3j.�*���*�j�`�.w'�`�|'`�0*����1)1/-��'.053�8jwj*��*6�2�`�j5����.'����*{�.��005`1'**�*{�
.'� ��1�*j�� �2j'*:� �`� '.}{� �����}� -�``�� '�`�|'`�0*� 3�12.�� �34�*�5�� ��� .w�*`�5'�`�0*� ���
1)1/-�� '|��� .�3� 5�20301�3{� 2��*� 8j�� .�3� 13�'*�31�3� 1�3� �*� .�j� 30��*`� 40j5� .w�*3`'*`�
�*�0**j3� �.�*�'..'� �`� '.}{� ��~~�}� �*|�53�1�*`{� .w�*|'.��'`�0*� ��3� :#*�3� �0�'*`� 40j5� .�3�
450`3�*�3� 4533�*`�3��'*3� .�� �014.�k�� 1)1/-��'20.�`� .'� 46034605�.'`�0*���� .'� 335�*�� ����
�w4*)� ��� *�*�� � j�5`�*� �`� '.}{� ������ 4'��*`0� �`� '.}{� �����}� -�3� �0**3�3� ���*`�7��*`� �0*��
1)1/-���011��3`'*`� .�� 7'�`�j5��'*�{� .wj*���3�'�`�|'`�j53��33�*`��.3���� .'�450`3�*�� 4*)}�
4�*3�{�4*)��`� .�3�'j`5�3���2.�3����1)1/-�{� �*�.j'*`�- *� ����"#���'� "����������'+��'"��'�
�����������������`�.'��*-��������������������3�5'��*`��14.�8j3���'*3�.��57.��450�450.�735'`�7��`�
'*`��'404`0`�8j�����1)1/-���-4'593�'*�� j�5`�*{���~��}���
�

��;�
+'+�
�4+�
'��
���
�4<�
".#�
.�#�
�<�
.�8�6�
��#�
����������������� ������������������������',� ��� � �,� -�$�%� .6,� 8�6&� ��� 7��,� �(<#�4#����'�,� ��� -�9����%� d’un� ��:�%;� /6&� �,�&!<,8�,6&�;,��88,���#�8&�7�.6,����8��/8��&8�����%�;,�/�%�;����%,��%�&�<,��6���&,�,��6%�<��%:,�,%��;,�<�%-�&�����%�8,&�,���%���,�&,<&6�,�,%��;,����8&��!�%,��;�8���&�<,� &0;�� &0;� �%�,&�:������&/��7,<� � ���8&��!�%,�+'+,�.6��;,7�,%���<��7,�,��&,��&,� �,� '�� -�9!�/6&� ��� ���/,� ��/�� ��/� ��%/�� �<��7!� /,� -�9,� ,�� �<��7,� ��� ��%�/,� ��-,� ,%�&��%�%�� 6%,� <�/<�;,� ;,�8��/8��&8�����%� �,%�%�� �� l’activation� ;,� ".#� 86�/� ;,� .�#�� -,��,� ;,&%�1&,� ,/��&,/8�%/�0�,�;,����8��/8��&8�����%�;,�-�<�,6&/�;,��&�%/<&�8���%��,%�%���� l’expression�;,�:1%,/���8��.6!/�;�%/����8&���-!&����%�,�������:&����%�<,��6���&,��-,��,�7��,�;,�/�:%���/����%�,/�� %�����,%�� <�%�&'�!,� 8�&� �,/� 8&��!�%,/� ;,� ��� -�����,� +8&�6�8� �+8�8�� .6�� �%��0,%��l’activation�;,���-��
+����

� ����� ����������������������� �� ����������������
PH�
�
� ���������$ �������1%, �
�
�w��`�|�`���������`��� ������� ���`�����������`�����������+����*���'����������������
��4���� �`� �j���� ����� �� �|�� �� ����������� �j� ������ �� �� ����`�`{� w���j���`���� ��
w����������� |�����|��� ��� ���������� ������ �jw�� j�� ������`��� ����|��� � ��j�� ��
��`��`�� �4�j����`���`�� }{�~�������� ��`�� }{�~������j�����`�� }{��������������
��� �{� �����}� -``� |����`�������`�|������ ������� `j�j�������`�`��j�����������`�
�j��jw�� ���� ��� `j�j��� ��`��`�`��j�{� ��j �����`� ���� �����`���� ����� w�`�� ����
�� ��j �����j��������� ������`�`����� ���`�� }{���~���}��
�
�� �wk���������wj�����������`�`j`�|��`���`�|���/����`��j � w���������
����`�`��j�
�w��`�|�`������ ��|�������4�������� ��������� ������`�`��`��j��������� ��`���j��
�j�k��������������`j���������������` ���j� w.� {� ��� ��{� ��� {� � � ��������j�
!� �~� �-j ��� `� � }{� ~����{� ����{� �����}� .�� ��`{� �j� �� �j`�`����� ��`�|�`����� �j�
�w� `���`����� �����������j�� `�j����`� ��� ���`j��� �� �� |��� ��� �4��� ��`� �`��
������`�������� �������`� ����� ����j�{����`������`����w�j`����������������jk���j�
�� ��{� �j� ��������� �j� �j� ��j���� ������`� `� � }{� ��~��� ���� ���� 'j{� ��~��� �"�#�{� ��~��}�
�wj`� ���`�����wj��������j`��`�����`�`j`�|��`���`�|���/����`�������`�j`� ������
������� �������`� ������������� w��`�|�`������ ��|�������4������|������� ��
`j�j��� ����`�`��j�}� /4$� �`� ���j`�� �� �� ������ � �����j� �j�`� �� w��`�|�`���� ��
����`j��� �� ��`�|�`�� `������� ������ �/���� ` �� �j� w.� /� ��j�� ��`�|�� �� ������� ��
���`�����/4 {��.������20��2���������`�./�~�����9����� " ���.��" ���'������������������j��
�`� w���`j�� �� j�����|� ��� ��|�����������??�}������j� w��`�|�`������/4$������`�����
�j������`� ��j�� w���`��`���� �� �� `j��������� ����`�`��j{� ��� ����������� �|�� �w�j`���
� `���`�����` ���j� ���j�k����������4/{� w��`�|�`������ ��%���`������j������ ����`�
����.&���j`�`��� j������ ���`�����j������`��� `j���� ��|�� �����`��������`��`���{�
���j���`�����wj�����������������������`�j�����`� �`������j����� ����j�����-���`�� }{�
��~����j �� ����`�� }{���~����������`�� }{������}�-��� w'���{� w��`�|�`��������4���
�`������ ���� ��������������j��������� ������`�`�`��� w���j���`������ w��������������

�

� ����� ����������������������� �� ����������������
PP�
�
� j �� `j���� �� |�����|��� ��� ���������� ���� �� `� � }{� ~����}� -���������`� �� �``�
���|�`���{� �����`j���������������` ���j�!� �~��j������ w.� ����`����� ���w��`�|��
������� ���`�����������`���4/���������w�����������-j ���`�� }{�~����}�.����`{�� ���
�`�� ���`��� �j� w��`�|�`���� �� �� |��� ��� �4��� �j`� ����j��� �� �� ��������� �`����
��`�|�`������ �����`�|�`j����� 4/��j��� 4/� j���(�{����``��`� j�� ��`�|�`������ 4/�
���������`� �� �� ������� �w���������� � ����� ���� ����{� ���~��������� `� � }{�
�����}��
4j� � �� �� ./�~��{� �� |��� ����4��� ���`� �j���� �� �������� �� ��� �����jk� �� �`����
����`��j�{� ��� ����`����� �j� ����`�k��j�� ���� � ����� ��� ������� )&�~����� `�
�����%�������j���`��*������������������������� ��� ���������./�~��{� ` ���j�
w����`���`� w���(`��j���� �� j ���}�-�����`{�������|�`�������� ������������`����
������ ����j�� �� �� �`�`� ��j|�`� ���� �� ���������� ���� �� �j� ����� j�� ���`k`�
��`�� ����j}�+�����`�����k�� ��j� w���j�`������ w����`������������`��������`���
w��`����`�� �j� ���� ��.&� ����� ��� ������ � j ����� �� ������� ����`�`��j�� �j������
�$��,`�� ���� !������{� �����}� '� � j�{� j�� �j`�� �`j�� ���`�� �j� ����� j�� ���`k`�
���� ����j{� w��`�|�`�����������`��j � �������`����`� w��|���������� j ��������j�����
���``��`� w��`�|�`���� �� �� ��`� ����`������ �� ������}� -``� ����� �`� ���`� �
��j���������� ����`����k`��� j ����`���j�����``�� �������`�������� j ���� �����
|������ ��`�����`��`�� ������� �����������`��`���}� -������������``�`� �����`��
�wk� ��j���j� w��`�|�`������ �� |�������4������`� (`�� �������� ���w�j`��� � `���`�����
����`��j����j������ ���� ������������`j���� �`� ���| �����`�����`��`���}�
� �����`�������`������� ������ �$���j`����������|�� ����`�����.&������
w�������������`�`��j�
������`������� ������ �$���j`���$���j`��~�������w������`��� ������� ���`�����������`�
��� �4��� ��`����`� �� ������`� /��� �'����j��� `� � }{� ������ -j����� `� � }{� �����}�
�wk�������� �� $���j`�� ~� `� �� $���j`�� �� �`� �����j�� ����� ���� `� ���� ��� �������
����`�`��j�{�����`�|��`}��j�������j`�����������`��`��`������ ���� ��������|�������
����`�`�`���������� {����j���j�����j���. ����`�`j��������`�����$���j`��|�����|�����
�����������`������`��`�`��j� � ��`�����`�� }{������}����k����������� ����� ���*�*����`�

�

� ����� ����������������������� �� ����������������
PQ�
�
��������� � �. � �j�����j�� �� `j�j�� ��� ���`����� $���j`�� ����� �� ����`�`� �j���j�
w��|� ���`������$���j`��~�`�������j�`��� w������`������'��!&�� �����j� ���j�k����������
$���j`�� ���� w���������.&� �� �`�`� ���`��`� ���| �����`� `j���� � �$��j`�����
�������`��{���~��}�.����{� w��|� ���`������$���j`����`�����.&����� ��� ���| �����`�
`j���� � `� ��|����� �� ���������`���� ��`��`�`��j� ���` � `� � }{� ��~��}� -�� ������� ���`�
������`���|�� �����|�`������ ����j�{���� ����`���$���j`����`�����.&������j� �
`�������j�|�������`��`�}�
�
�� ���������� ����wj���j� ��w�j`����j �`�����j`���������������`� ��� j ��
�� ������� j�����������|�����|���������`j����������������
/����������`����`��j������ j ���� ������`� w����������� |�����|������ ���`j�����
���������� �'������� ���� �����{� ��~~��}� ��� � j �� `j���� �� ����`�`��j��
�w������������`��� w�����`��`���� ��� w.� ����``��`���� ���j���j� ������j �`����
�j`�����}�'����j������`�`�����{� w.� ��`������`���� ����������� ��� j ���`���� �{���
������� �jk� ���������{� `� ���� �� � j �� ���`�� �� �}� �� ����`j�� .� /� �������
`�j`����� j�� ��� ���`���� ��`���`� ����� �� � ���}� ! � �`� ��`j�� ����� � ������`���`�
���`�� �� � �j� ��|�j� �� �� ������ ���� �`��� {� ��j�� �� ����`����� �����}� -``�
�������`������(��� � ����`j���w(`����`�|������ w.� � �����`������ ��� j �� j���� ��
����`���� ���� '������{� ������}� ����� �� �� `j��������{� �� �����������`���� �j�
������`���`����`�� �� � �w�����������wj����`��� ��� ����`����{����``��`��������jk�
� j ���� ��������w�j`�� ���`�������`j��������������{����`� w.� }�-``��j� ���
���j �`���� �j`������ ���������`� �� � j ���� ����� ��� ���j �`����� ���������� |���`� �j�
�`����{�`����`�������`���� ��`��� w���j���`������ w��������������������}���
�
?>@> ��� ���� !�,���"���������������
�
� �����������(�������������������$������*����������� ��� ������
�

0����(���1"�2")�2(*�(���
+��-���2#�"�1"�l’intégrité de l’ADN�
�������������������� ����������������������������������� ������"� *3*�"� *"��������� ��� ������� �� � ������� ����������� ����������� ���� ������ ��������� ����������� ��� �������� �� ��� �������� ����������� ������ ��������� d’un� ������ ������������ ��� ������������������������������������������l’ADN���������������������������������������������������������� �� ��� ��������� ��� � �� �� ���� ���������� �� ��� ������ ��� ������ � !��������� ������ �������"�������������������#�����������������s’assurer����$������������������������� �.� �� ��� ������ ���� ��� �������� ���� ��� ���� ��� ������� ��� l’intégrité� ��� ��� 4'�� ���� ��������� �� ��������������������� �4��������������� ������������� ������������������������������������������������%������������� ������������ s’engage������������� ����#��� � .� ���������������������������� ��� ����"��� ���� �������� ����� s’assurer� ���� ��� ����������� ���� $��� �������� ��� qu’aucun��������� n’a� ���� ���� �� l’ADN� ������ ��� ��������� � &� '���� ����� ��� ����"��� ������� ���������� l’appariement� ��� l’attachement� ���� ������������ ��� ������ ���������� ������ �������������� � �� d’� �������� ��� ����������� �������� � ������� �������(())) �*�� ���(�������+���+��,�-�.�/01��$#/2$�. (������(-���%'������%��%���%-���%-#���%3�/'43�/'4� 3�/'43�/'4%,��%-���%-#���%��%�������(�
3. Vérification de l’attachement des ��������������� ����������������

� ����� ����������������������� �� ����������������
PR�
�
�w��`����`������� �����jk� � ��������� �� ���`j����� ����������`� ���������� �kj ��
���j �`� ��������`������ �����`�|������ j ������`�`��j������������`� ���� �� j ���}�
- j���������`����������j����� �����������j������`����j ���j�{��������,����`� ��
��� ���`���� �� w4'&� `� �� ��|������ � j ���}� -��� �� ���������{� � ��� � � j ���� �`�
� �����j��`���|�������j�`�������� ����`���� �����`���`�� ��~{�${���{���� �������� ?@�}�
� j��j�������`�������`�. �{��j� �����4����`���{� ���`��wj�� �����`�������`��j ������j��
w���(`� �j� �� ���� �� ��j`� �� �� ��|������ � j ���}� 5 �� �``�`� �� 6j� j�� ��� � ��
���`��������j�|� ������ w��`����`���������j��j���j|�`��`����� ���� �� j ����*���
w��`�|�`����������`������� ������ ����}�-�������������`�������`j�����`��������`����
`� � �� ���`�. �`� wk�������� �� ���`����� ������`� �� ����������� �j� ��� {� ��|������`� ��
������`������ w4'&{��j��`������`� �����`�� j ������������`��� ����j� ������������
w4'&����`�`���������`��`���4�����{����~�}��
/������`������6j����j���� �� j ������ �j� ������ ����������`����� w��`������
� �� ����� �� ���������� � j ���{� �j� ����� �~�}� -� ����`� �� �������`�j���{� � �����`� ��
��`���`���{� ��|��� �� ����� �~� �� �jk� ���`��� ����� �{� ������� ����� �~� ������� `�
`����|}����������{�� j�� ���j{��������� ���#" ����`������j������� j��k����`����
�jk����`j����������������j����`�|�`� ��|����������� ���`�������j��6���`����j�����j���
wk���������� �� ��� ��� '~}� - ������ �w��������`� �|�� �� 4������ -'����� ��|����� j��
��`�|�`�{����j�����`� w�`�����������~�`����|����`� ��������������`����������`���
����������� ���`j�����`�����}� ��� � j � �`� � � ���� ����|��� ��`� ����������� �
��� �� j ���{� ���j ��� `���`�|����`��`���`��`� ����|�������j� �����`���������`������
�������� ������`���`�`������� ���j�`��4``����`�� }{�~����}�5 ��w�`�����������`�����`�
�j��``��`������`����`��j �����`�`�j��������� ��� j ��������j��}��
�
� ����������������������������������������������(����������������������� ��������
� ����������2(�����#23 �
�
����������`�� ���������`���|����`���j����`� �j��j���`� w��`������`� ����`������`�
���j ����������� ��`������������ k�������`����{� ����� ����-'����$� ��+�����'����
��������}����-'�����`����4���������`� w��`�|�`���`��������`��� w�������`�����|��j��

��� �� ��� ��
'��
-'7==$�
.�
-'7,�
4�
-'7,�
��
-'7��
6%-=
Figure 24. Fenêtre d’activité des complexes Cyclines/CDK durant le cycle cellulaire.������5��%���d’évènements���%�� �������%�� ���� ���� � ������������� ���%��%��%� �!�%��%���l’activation��� �5�������,,���%����6!% ������� �%�=-'77�L’activité���������%���������� �3!&!�� �6%��� ��� %��� l’expression,� �� �������6%� ��� �� 6�� �����6%� ��� �� �%��7� (%���%����%������������ ���6%����������%��� ����%7�#���������-'7��-76 2�6%��%,�2�%,����� ���-76� �� �� ,�!� �� 6%-=7� -��� �,,���%���� %�6���%��� ��%6%�%�� +� ��� ���!� �� �,,���%��8�l’expression���%,���������!� ���������+��%����������%6�6��5���%���l’intermédiaire���%>83�%,���6�������������6%����!�%���� 6����� ���,,���%�����6%�%6���!��%��%��� ����� � ����%���%� ����� 5������%�3� �%,�%� l’expression� �� 6���� �6�%�� %6��� �� ,�!� �� �� %�6���%�� 6%-=����!�%���������+��%��������6%�6��%�5��2��6!!���%����������6%����3!��6���� ���6�������4877�
%,�� %,��
$��������%6�6��5���
'�,,���%�����6%�
$�������6%�6��%�5���

� ����� ����������������������� �� ����������������
PT�
�
��� ��� `��j�{�������������� �`���{� ���`��j�`� �� ��`�|���j� �� ����`�|���������`�� �� ��
� j ����� ���``��`� �� ��������� �wj�� �`��� �� w�j`�� �j� ��� � �������{� ~����}�
�w��`�|�`���� ���j�`� � ��� ���� k�� ��� ����-'�{� ����� ��� ��(`��� `���� ��
�������{����j `��������� ��`������������`������� w�`�`������������ �`���������-'��
`� �� �� �������`���� ���`�� �`��j� ��� ��� ���� �������`� �� wj��j�`��� �'$� � ����
������{����~�}���������� �`����j��j��-'�����������w����������� j��j������ �������`�
�w�`���� j�� ����`���� ������`��j�� �� ������������ �j�� ���`�����`� ��� ��`� �� ��
���j �`�����j���� �� j ���>�4����{���j������j�����`�����`������j���� �� j ���{� w��`�|�`��
��� ���� k�� -'����� ��� �`� ���`�. �� ���� �j ��`� �j� ��|�j� �� j�� ����`��j�
�w���� ��� ����� �j���� �j� ��|�j� �� wk�������� �� j��� ����`�`j��`�� `� ���� ��
��������`������jw� ���j����`}�����|��`{� ������ k��-�� ���'�-'���`�-�� ���.�-'���
���`�. �`� ���������j�����`��� ��`���`������ �� ���� k� -�� ��� 4�-'������`� ����
����j ��`��� ����� ���`������ w4'&��j���`� �������$��������j� ���������j�����`���
���`�. ��������`� ������ k��-�� ���4�-'���`�-�� �����-'�~���`�|����`��j���`� ��
��`��� ��������� ��j�� ���j��� �� ��|������ � j ���� `� �����`��� �� ���� �������`���� ���
�����������������?E�}�
4j�� �� �� w���� ��� ���j�`� � ��� ���� k�� -'����� ��{� �� ���j �`���� �j� ��� �
� j �����`���6j������������������`���� ��������`���`�|����j������ ������`�����
�����`����� ��� ��j��j��`��� ��`� �`��j�� ��� ��� -�5� ��$� ��� '����'���+2������ ��!�)������}�
-������������`���j������`������ ��j�� ���� ���������~�`� j��� ��������`���������
�� ��������j�� j���`�j�`j��`� j��������`����j�� ���������`��-'���9j�`��`�����{�~����}�4�
��6�j�{��jk������������ ����-�5���`��j�(`�����`�����}���������������jk����j���
�`� ��������������`������� �� ���� � 5�4�� ��~�5&���{��~�5&��{��~�5&����`��~�5&����}� . ��
�����`� ��������j��`� �� -'��� `� -'��}� �� ������ �`� ������ ��� ���`����� -�������
���~-��~{�������~�`���������}�. �� ��j�`���� ��`� w��`�|�`��������� k��-'������ ���
4���� ���.�`�-'�������� ���'��/�j�� {�~�����$����`�/��`�{�~����}�
�

:%;)� <8�
�0�
<)�
=<7�.(�
;��������������������
���� ���
���
���
��7����
��� �����
�
��7��
��
��� ����
���� ���
���
��
��� ����
�� �� ��
�����������4����
� ����#��
#����������
� ������
������������ ������������������������������������������� ������������������� ������������������ ���!�� "��#� ������$����%��� ���!� �&�� ���'����������������������%������������������ �(� ���������)������$�� �!��$���*����l’expression��� �� �+� ���� �� ��$!����(� �� �'��� s’associe� �%��� ��� ��7� �� ��� �� ���� ��� ����%��� ��� ���!� ��� ��������+ ����� �� ��(� �� �� ���!��� l’activation� �� $,���� ��� ��� d’E���� �!� �*���� ���� �������$�������������������������� ���+� ���� �"��� #(���� ��!-!��.�/��*������� ����� ���+� ���� ����7�'��� �� �+� ���� � s’associe� �%��� �� ��7�� *���� )� ��� ����� ������+ �� ��(� ������.�!���+���������+ ��� �� ��� ��� ��������� �!� �&�� ��'��� ��� ��,��� �!� ,��!���� �������%���� �������������� �(� ���'��� ����%���� � ��� l’expression� �� $,���� ������������ )� ����� ���������l’ADN,�������&���������4���� ���+� ����4�"���4#(��

� ����� ����������������������� �� ����������������
PK�
�
�� �0����1 k0�-�� ���'�-'�����`�-�� ���.�-'���12�``�`� �1�00�3��j�1���`�
��20`2��`����
�w�`2����14�0�5~��j���� �� j ��2���2�1��0���j��0`��j j0���`�3���6j������j��
k1�0�`���� �� �0� 7��`j20� �� �2��00���{� ���00�`� j�� �j3��`�`���� �� wk1200���� �0�
��� ���'��'~{�'��`�'��}������� ���'~�20`� ��1 j0��`j�����w�`2� 0}��5 ����`�����`2��6j�
����`28 ���0���k1200������`2|��`���1 �6j�����|�jk���2�3j �`������`2��0�2�1`���� {�
`2��j�`���� {�1�0`�`2��j�`���� ����`� �`�� }{���~��}��0���� ��0�'�0�7�k�`�0j2� 0�-'���`�
��1�j2�0`��j 2� j2���`�|�`����`� �`�6j{�`�12�``�`����0�� ��14�014�2� �`��������`�|�`2���
�� ��12�`�����j�2�`��� �0`���/}�- ���{����0��6j�00���� �3j0�1~���`�1~��{�0��`��0�
��4��`j20��j���� �� j ��2�6j��0�6j0`2�`� 0�7��`j20���`2��0�2�1`������ ��7��� �.�+�
�.�+~�����}��0�12�`���0��� ��7��� �.�+�`�/�0�2`2�j|�`��00����0���j��12�`������ ��
7��� �'���'�~����1�j2����0`�`j2�j�����1 k��j `�12�`��6j�2�1200j2��j�1�00�3�5~�$}�
.�� ��j`� �� 14�0� 5~{� �� 12�`���� /� 0`� ��6�2�`��2��`� 2`2�j|�� 0�j0� j�� 7�2��
4�1�14�014�2� �� 6j�� j�� 12�`� �w��`2�3�2� �|�� 0� ��20� �� �� 7��� � .�+� 1�j2�
��06j2� j2� ������� `2��0��`�|�`j2� `� 2�2j`2� �0� 12�`���0� ��1 �6j�0� ���0� ��
���1��`���� `� ����� � |22�j� �3� �� �� �42���`��}� ��� 14�014�2� �`���� �� /� 1�2� 0�
���1 k0� -�� ��� '�-'����� �� �`� �``� ��`2��`���}� �� ���1 k� .�+�'�� ���`28 �
��`����`� wk1200������3��0�00�`� 0��� ���j1 ���`������ w4'&�6j���j2�� �j���14�0�
${� ` 0� 6j� �-&4� `� �� ��� ��� 4}� 'w�j`20� �� 0� �� .�+~� 0��`� �3� ��`� ����j0� 1�j2�
��4�2�`��j���32��2��0�12�`���0���4��`2��0��j���� �� j ��2}�-w0`� ���0�{��wj��1�2`{�
�� wj�6j�`���.�� �3�0�$����6j������j�`��� ����32���`�����j�-�5�1��9�1~{�`��w�j`2�1�2`{���
����� ��� .{� 6j�{� *��� -'��{�����j�`��� �� 14�014�2� �`����`�������� w��4��`������/� �1�j2�
2|j�|��2���00�3j�{��������������?H�}��
�
� �����2�3j �`������/�0`�j���|����`�72�6j�`����0� ����2����3��0�12�0`�`�6j�
�w����`�|�`�������1 �`���/�0`�0j77�0��`�1�j2�1�002� �1���`���20`2��`����`�12�``2�
�� ��� j ���2�1 �6j2�0���4'&}�5 ��w0`������1�0��`�����`�6j� ��0�3�� �0�`������1����`�
��/�0��`�2`2�j|��72�6j���`���2�3j �����0������2jk�����20{����`� ��12�0`�`}�
.�� 77`{� ���� �0� `j�j20� 12����20� `� ���� �0� `j�j20� ��`�0`�`�6j0� 12�0�`�`� j��
��07���`������`����0� ��2�3j �`�������``� |�������`���j����`�|�`�����22��`���

�

� ����� ����������������������� �� ����������������
PL�
�
.�+~����� ��0�`�� }{������}��wj���0�� `�2�`���� 72�6j���`�2`2�j|��0`� ��12`��� ��
2�3���� �42���0���6j� ~�6� 6j�� ���`��`� � 3���/� �-����� `� � }{� ~����� ��� `� � }{� ~�����
� ����`�� }{�~����}�5 ����`��2�����`������`2��6jw��1 j0���0���28 �0j2� ��2�3j �`����
�j� ��� � � j ��2{� �� 12`� �� /� 0`� ��1�2`��`� ���0� �� 12�3200���� �j� ����2� �� ��
12�0`�`�|20��0�0`��0�2�0�0`��`0��� ����0`2�`����1j�06jw �12�`�j���j3��`�`������
wk1200����`��� w��`�|�`����4/{�� j������`��`�j��3����� ���.�+~��$4�2���`�� }{���~��}�
-4�� �� 0�j2�0{� � 28 � 0j11200j2��� `j�j2���/� ���`�������`2�� 1�2� 0��� ��|� ���`����
01���7�6j� ���0� w�1�`4� �j�� 12�0`�`�6j}� - �� ����j�`� �� �� 7�2��`���� �� ���1 �0�0� ��`2��
�1�`4� �� 0� 6j�� �w�|� j�`� 1�0� �� ��2�����0� ������0��� `� � }{� ��������� 1�2� ���`2{�
w��|� ���`������ w�0� ��� ��7��� ���12�`���0�/��/�1~���1~�������j�`� ��7�2��`����
�w�������2�����0��|��j��1���`2�������1 �`{�0j33�2��`�j��2��������7���`���� �
�`2��0���77�2�`0���20��9� �`�� }{������}��
�� 1���212�0�`�j���22��2��� ��12�3200����`j��2� ����0� ��12�0`�`�
���12`���1������0� ��32������6�2�`���0�����20���j��0`����|�����0�j �3�� w��1�2`����
���``�12�`�������0� �����`����� w4����0`�0���� ��3 ����12�0`�`�6j��9���23�`�
� }{�~����}�1���6�j�j��28 ��2j��� ����0� ����`�`����`� ��2�1�2�`�����0������30���j0�0���
w4'&� �j2��`� �� 12�3200���� �j� ��� � � j ��2{� ���0�� 6j� ���0� � ��� ��4��`� ��
w�1�1`�0� �206j� 0� �0���0��� 1j|�`� 1�0� (`2� 2�1�2�0}� 1��� 0`� �3� ��`� ��1 �6j��
���0� ���� ��4��`��� ��0��0����� j ��2���2�1��0���j��0`200�����3���6j��+5${�
��������� ��'"��'� ������������ 6j�� 0`� j�� ������0�� ��`2��0�6j� 02|��`� �� 72��2� ��
12�3200�������1 �0�6j�`��``��`��� 6j���`����`� 0� -�5� �����`�� }{� �����}� 4� `�`2�
�wk�1 {�� �0`�����j�6j� w��|� ���`��������.&��4�� ��0�j2�0���� ��4�j��0��0����
���0� �� 12�0`�`{� �� 6j�� ���`� ����00�����`������`�0`�`�6j}���2� ���`2{� w��|� ���`������
��.&� ��j1 �� �� � � �� 1��� �� �`� � 14������� �w+5${� `� 2�0j `� �� w�11�2�`���� ��
`j�j20� `2�0� �3200�|0{�����`� �� �� 7�2��`���� ����`�0`�00� `� �� w�j3��`�`���� �� ��
�`� �`�� �4�� �� 0�j2�0� �-4�� `� � }{� �����}� .�7��{����0� ����� ��� ����2��� �� 12�0`�`�
�/4����%�����������'����������#������!����"������������{� w���`��`�����0� �0���0���j0�0�
1�2� ��0j2k1200������ w��`�3������j�|�2j0�$#������0� w�1�`4� �j��12�0`�`�6j�0`� �72j�`�
�� ����j �����`�|�`������1���`���/��9j2��`��`�� }{����~�}��0�`j�j20��02|�0��4��
�0� 0�j2�0� 0��`� �3� ��`� `2�0� �3200�|0� `� �� 12�3200���� �j� ����2� 0`� ���1 �`��`�
��0�0� ��� ����0`2�`���}���

�

� ����� ����������������������� �� ����������������
PM�
�
�
��� ��� ,��� ��� �!�"� ��� ����!�#���� $��%�D���&� �#� �''���("� �#����!��"�%��� #�� ������� ,�� #��
'� �"�"����"� #������#"��"��,��'#��������,��)#�!��"��,D�*'����� �����%��&�� �� !�"��"��
������������&�%���� �,�����"����D���������������D� �����,�������������>���������������
,�������������D������������������������������������ ������������������������������
���������,������������������������ ����,��� �����������������������Y������ ���� ���
Y�����,������Z�����?S=S[[>��������������������������\����������]����������������
�� ��� ��������� ,�� ������� ������� ��� �� �D���������� ,�� ��� ���������� �������������>�
������ ������� ,�� �������� ����� �D����� ,D��� ���,�� ����� �����,��� ,��� ��� �������
��������>�
� �
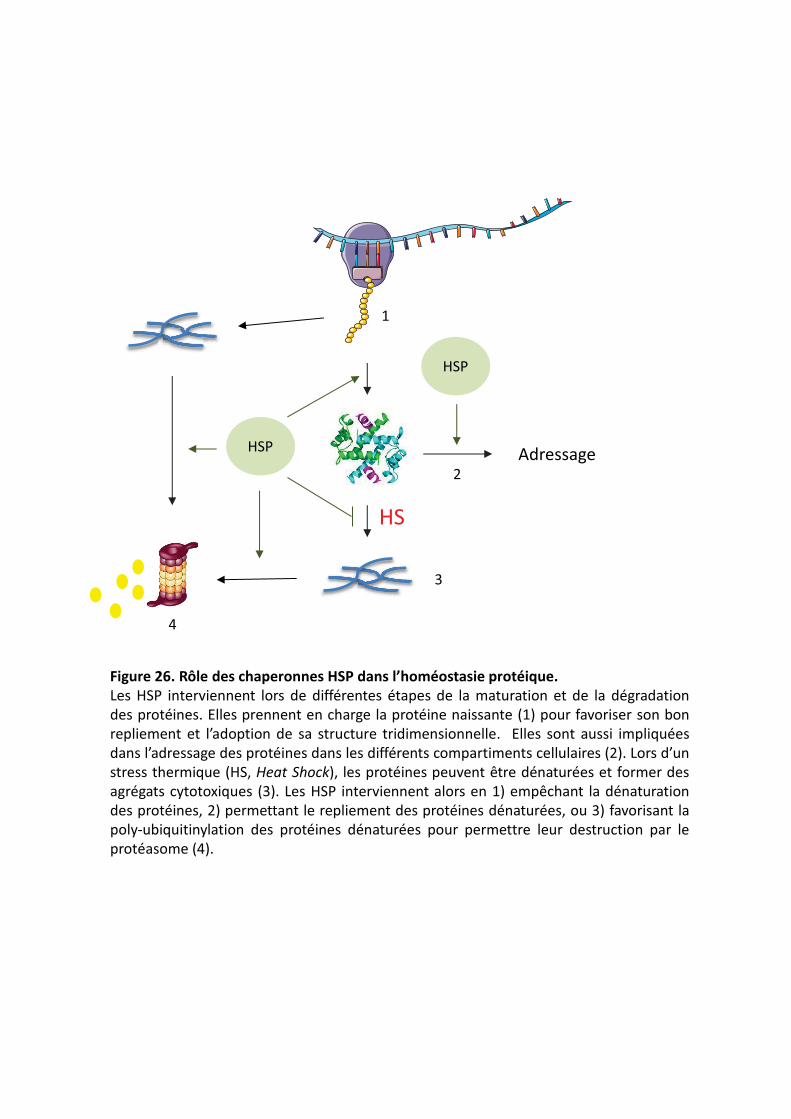
9���
���
����
4�������
��$� !�"#$�Rôle des chaperonnes HSP dans l’homéostasie protéique.��������� � ������ � �� ��������������� ������������� ��������� ������� ��������� ����������� ������������� � ��� ������ �������� �� ��� ����������������������� ��� ��������� ����� l’adoption���� �� ���������� ������� ��� ������ ������� �� ������� ������������ ��l’adressage����������� ���� ������������� ������������ ��������������� ��������d’un���������������������!�����������!� ���������� �������� ��"������ ���������������������������#����$������ �%����������� � ������ � �������� �������"�� �� ���� ������ ����������� ��!� ��������� ������������� ������������ ����� ������!����%�������� �������#&�������� #���� � ���� ������ ��� �� ������� ����� ���������� ����� ���������� � ��� ��������������'���
��
�
%�
'�

� ����� �������������������������������� ����������������!������"��������������
QO�
�
���� �������� ��������� ��� ����� �%��� ,��� ��� �������&���
����������>�
�
�
/(� )**j*� 4j+�,(� )-(`,(`� j(� .j�(`,`�� /4�(-+�(�*� �� /0-`�,({� /-j0� j(�
)-()(`0�`,-(��**�(`������������+3�+�{�/0-)4���*��1�`j0�`,-(�0`0-j|����(1�*1�)0,1`�jk�
��/0-`�,(1� �4140,{� ������ +,(2�� �(�� 5-*-j,(-77{� ��~��}� ��� 0�3j*�`,-(��� )�/0-`�-+�
)-(1`,`j� j(� ��7,� +�3j0� /-j0� *�� )**j*{� `� ��/(�� �� *�� �*�()� (`0� *�� 1�(`4�1{� *�
0/*,+(`�/0+``�(`��jk�/0-`�,(1��w��-/`0�j(�)-(7-0+�`,-(�`0,�,+(1,-((**�/0-/,)�
�� *j0��)`,|,`�{�`� *����30���`,-(}� �1� )4�/0-((1�+-*�)j*�,01� )-(1`,`j(`� j(� )*�11���
/0-`�,(1� 3-j�(`� j(� 04*� ,+/-0`�(`���(1� *� 0/*,+(`��1�/0-`�,(1� `� *j0� 0+-�*�3{�
/�0`,),/�(`��,(1,��j� ��)-(`04*�.j�*,`�����j�/0-`�-+{�(��|,`�(`�(-`�++(`� *�� 7-0+�`,-(�
�w�30�3�`1� /0-`�,.j1� )�`-`-k,.j1}� ��� /*j/�0`� �1� )4�/0-((1� /j|(`� 0+-�*0� *1�
/0-`�,(1�(-j|**+(`�1�(`4�`,1�1�/�0��1�0��)`,-(1�(��+�`,.j1���/(��(`1���*w4��{�
)-++� )w1`� *� )�1� /-j0� *1� 9$���{� 9$���� `� 9$���� ����� �!��2� ��������� �������� ?M�}�
'w�j`01{� )-++� *1� /`,`1� 9$�{� 1� 7,k(`� 1j0� *1� /0-`�,(1{� /0+``�(`� �� /0-`�30�
)0`�,(1�1-j1�j(,`�1�/(��(`�*�/0-)11j1���0+-�*�3��9�0`*�`��*}{���~~�}�.(��4-01���
)�04*�11(`,*��j�+�,(`,(���*w,(`�30,`���j�/0-`�-+{��w�j`01�7-()`,-(1��1�)4�/0-((1�
-(`� �`�� ,�(`,7,�1{� `**1� .j� *�� 0�3j*�`,-(� �� *w�)`,|,`�� �1� 0�)/`j01� �jk� 4-0+-(1�
1`�0-5�1� -j� ()-0� �� *w�11+*�3� `� �� *�� ��(�+,.j� )40-+�`,(,((}� 'jk� `�/1� ��
)4�/0-((1� 10-(`��`j�,�1���(1� )� )4�/,`0��� *�� 7�+,**��1� 9$���,(1,�.j� *1�/0-`�,(1�
)4�/0-((1��w4,1`-(1}�
�
=> ������������'���
�
���1j/07�+,**��1�9$�����w�-0���`��,�(`,7,��)-++��1�/0-`�,(1��-(`�*wk/011,-(�1`�
7-0`+(`��j3+(`��1j,`���1`011�)**j*�,0{�)-++�*1�)4-)1�`40+,.j1{��w-��**1�`,0(`�
*j0�(-+� ���+/,(3��`��*}{������}�$j,|�(`� *j0� `�,**{� *1�9$��)4�� *1�+�++,7�01�-(`��`��
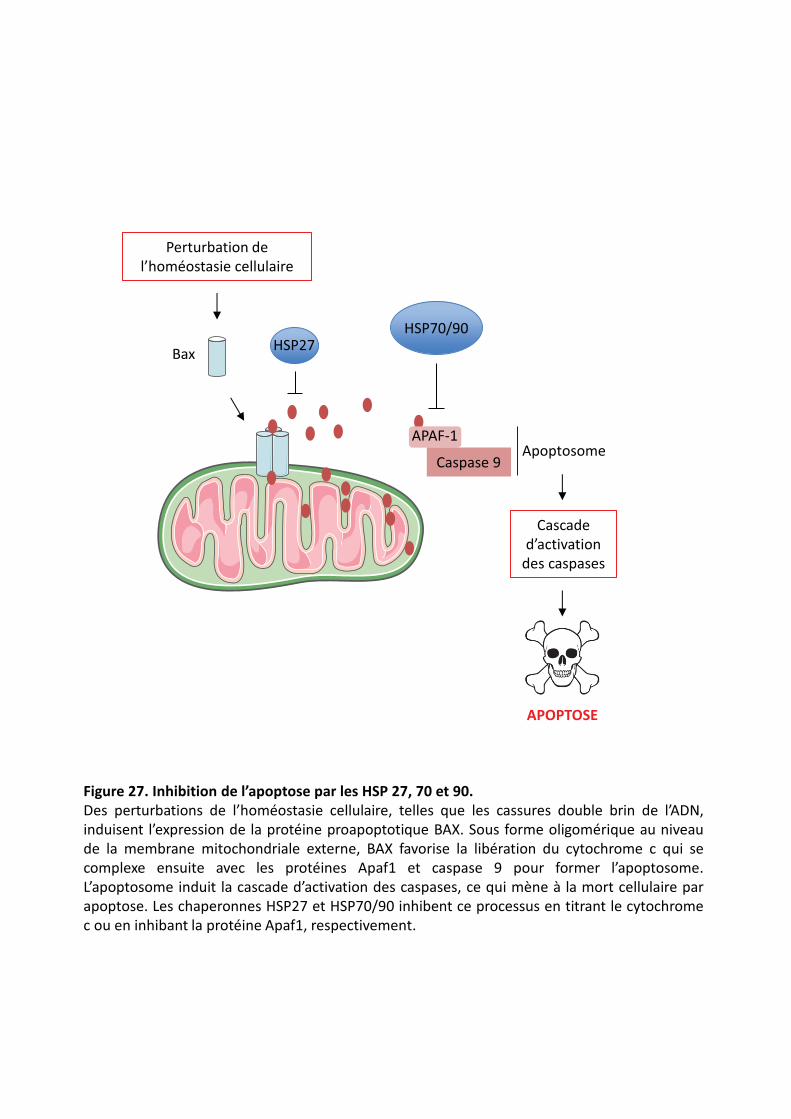
4$4�,6�
-����� �7�4�������� �
-����� �d’activation � �������� ��
�+�+�����
��������������
��� ����������l’homéostasie cellulaire�
����
�()�*+��,.��������������l’apoptose���� �������������������'�� ��� ��������� �� l’homéostasie� ��� ������ ����� � � ��� ���� ��� �� ��� ����� �� l’ADN,���� ����� l’expression��� ����������������������� ��4����� ���������� ������ �� ���!� ��� ���������������"�������� ������ �4�� ��!����� ��� ����������� � � �#���"���� �� � �� ��������� �� ��� �!�� ��� ��������� 4���$� �� ������� � �� �� ������ l’�����������L’���������� ��� ��� ����������d’activation���������������� ���%��&� ����������� ������������������'���"��������������������������"������������� ���������������#���"������� �����"�����������������4���$��������!�����

� ����� �������������������������������� ����������������!������"��������������
QG�
�
)(�))*)���+)�,j�`-�7�.,((){�)(/+�(j-�`�,((���9$���{�9$���{�9$���{�`�()�0`,`)�9$�����
~���� ���1'�{�0�-.,� (),j(()�/+� -`-/j|� (�� 9$����}� 'wj+�.�+,�-� 3*+*-�({� ()� 9$����
2�j`�0/,�)�./(*)j(�,-)�/+`�j+��)`,|,`*�4����*0+��+`{�)/+`-�,-.+`��jk�0`,`)�9$�}��
5(� )`� )/++j� ,j� ()� 9$�� -*3j(+`� (�� )`�,(,`*� /j� (�� �*3-���`,/+� �� 0-/`*,+)� �� �)`,|,`*�
/+)/)j00-)),|� /j� �� 0-/`*,+)� �� �)`,|,`*� /+)/3*+,,j� )/..� (� -*)0`j-� �)�
�+�-/3�+)�`�()�-*)0`j-)���(w.5+����++�j�`��(}{���������-)((,-�`��(}{������}�'�0(j){�
()�9$��/+`�j+�0j,))�+`�-3(�)�`/0-/`)`j-�`�)w)`�0/j-,j/,�,(�k,)`�j+�/++�)/--*(�`,/+�
+`-� (wk0-)),/+���))�0-/`*,+)�`� (�� -*),)`�+)��)�)((j()�)�+)*-j))��jk� `2*-�0,)�
�5�--,�/� `� �(}{� �����}� -� 0/`+`,(� )�`/0-/`)`j-� 7�,`� ,+`-|+,-� 0(j),j-)� .*)�+,).)�
./(*)j(�,-)}� �/j`� �w�/-�{� (j-� -3(� ��+)� (� -0(,.+`� `� (w,+`*3-,`*� �)� 0-/`*,+)� )`�
0�-`,)j(,�-.+`�,.0/-`�+`���+)�()�)((j()�)�+)*-j))}�.+�77`{�(w�)`,|,`*�.*`�/(,,j��)�
)((j()�)�+)*-j))�)`�3*+*-�(.+`�`-�)��j3.+`*�0�-�-�00/-`���)((���)((j()�)�,+){�
)�,j,�)/+�j,`���j+�)�+`2�)�0-/`*,,j�(�-3.+`��))*(*-*{�+/`�..+`�|,��(w�)`,|�`,/+��j�
)/.0(k�.�+/-~}�-�02*+/.�+�3*+�-�j+�)/,+�0(j)� ,.0/-`�+`�+�)�,j,�)/+)-+� (�
.�,+`,+��j�0-/`*/.��7,+��w*|,`-�(��7/-.�`,/+��w�3-*3�`)�0-/`*,,j)�)/+�j,)�+`���(��./-`�
�� (�� )((j(}� '� 0(j){� ()� 9$�� )/+`� )/++j)� 0/j-� ,+2,-� �� 7�8/+� �,-)`� ()� 0-/)))j)�
�0/0`/`,,j)}� 5(���*`*�./+`-*�,j� 9$����)`� )�0�(��� )� 7,k-�`��w,+2,-� (w�)`,|,`*��j�
)�`/)2-/.� ){� `� ()� 9$���� `� 9$���� ,+2,+`� ,j�+`� �� (()� 4�4+�~� ���������� ���������
���*������ ������� /�{� )� ,j,� .04)2� (w�)`,|�`,/+� �)� )�)0�))� +� �|�(� `� )`/00� �/+)� (�
0-/)))j)� ��./-`� )((j(�,-� ��-j�� `� �(}{� ������ �������� ?N�}� '�+)� (� )�)� �j� )�+)-��� (��
0-/)`�`{� ()�9$���{� ����`� ����)/+`�)j-k0-,.*)���+)� ()� `,))j)�+*/0(�),,j)�0�-�-�00/-`�
�jk� `,))j)� )�,+)���5�)+`)� �-�-�,((/��+�� 500/(,`,{� ������ +/)`-�`��(}{� �������j-�2�)2,�`��(}{�
�����}�'�0(j)�(wk0-)),/+���9$����)`�)/--*(*��j�3-�����5(�)/+�`���(��0-/3-)),/+��j�
)�+)-�|-)��)�)`��)�-*),)`�+`)��� (��)�)`-�`,/+}�/+��)�-�,)/+)���)(��)`�,j�9$����)`�
,.0(,,j*���+)�(w�00�-,`,/+��wj+��+�-/3*+/�,+�*0+��+)�+�7�|/-,)�+`�(��)j-|,�)((j(�,-�
0�-� (w,+`-.*�,�,-� �� (wk0-)),/+� `� �� (w�)`,|�`,/+� �� $`�`��� �//))2,� `� �(}{� �����}� ���
)/.,+�,)/+���`-�,`.+`)�j`,(,)*)�+�)2,.,/`2*-�0,)��|)��)�,+2,,`j-)���9$��-*�j,`�(��
0-/(,7*-�`,/+� �� (,3+*)� )((j(�,-)� 0-/)`�`,,j)� )�+)*-j))� 2j.�,+){� )/j(,3+�+`� �,+),� (�
0/`+`,(�,+`*-4`���))�./(*)j()�)/..�),(�`2*-�0j`,,j)���j�`��(}{���~���4��-j-�`��(}{�
�����}�.+7,+{�j+�*`j����.,)�+�*|,�+)�(��0-*)+)���9$������+)�(�0(�).����0�`,+`)�
�``,+`)���)�+)-)���(��0-/)`�`���)�()�)`��)�0-*)/))�������������X�������������������

��%��
'/������*�$��������/����������%�����������$���������������%���������
�/$������/��/��5����
������������������������������
�������������������������������
���������������������������
����������
��/������������/��������4 ': 65��%����������/������������/��������4 ': 65���/�:���/$�/�
'�
6�
�4� ��� '�
6�
�/�� !���
�/$�����$/��5����
Figure 28. Rôle des chaperonnes d’histone dans �’assemblage des nucléosomes. � �� 4�� �������� �� �/%������� ��� l’ADN,� ��� ��������� ': 6������������ ����/���/��� 4�/�5��������%���/���������������%����%�������%���4�.���-��:����������.������������+������7����+�-4+:��5��� ��� ����$���+� l’ADN������ .�������� �/�������� 4 ': 65���%���� ��%����$����� ��.������ �/%�������� ��� ��%������� �4!�� %������ l’intégration� ���� �������� �4: ���� �����������%�����%����+���%�����������$��������/�������%���������4%��5�����������������+������������.�������������%����������/%����������l’ADN,���../���������%��������%�����%����+��������5���d’assemblage�������/����������.������������(�������������������4��������������/$�����$/��5���9� l’intégration�������������� ': 6��%���� ��%����$����� l’4��%�� ((����.����$����� ��� ������ 4���� � � ���4�� .�� ���������� ��� ���� ������� ���� '���� ���� �� ������� ����������������� ������� ������ ���l’intégration������� ����!4"�!������������ �������l’ADN�n’ont������l’heure� ��#�������� ������ ������D’après Burgess ��������!$�%��

� ����� �������������������������������� ����������������!������"��������������
QH�
�
`�jk�&*'()j���*$4�*`�(`�+/'.�+��4�`��+}{������}�-&��/++*&�,+�)+`�+&�9$*�)/..��&�
�)`j'&� �� ,�'`� +`(�'� ��+&� +�� )�')(+/-+�&� ,'/&`�`()j� `� )/..� �&� ,/`+`(+&� /j`(+&�
�(�-+/&`()&�`�,'/+/&`()&���+w*|/+j`(/+���+��,�`./+/-(}�
�
?>�����������������,D��������
�
'�+&� +�+/��j��&� )++j+&�j)�'�/`&{� +w4'&�&`� +(*����&�,'/`*(+&� &`'j)`j'�+&��,,+*&�
.(&`/+&}��w+&.+�)/+&`(`j� +��).'/.�`(+{�)j(�,j`�4`'��(|(&*�+���.j+`(,+&�j+(`*&�
&`'j)`j'�+&{�+&�+j)+*/&/.&}�����)/j'����j�+j)+*/&/.�&`�)/+&`(`j*�����k.,+�('&���
).�)j+��&�.(&`/+&�9�4{�9��{�9��`�9�{�&j'�+)j+�&w+'/j++`�~���,�('&����&&��w4'&}�
��� ,'*&+)� �� +w.(&`/+� 9~� �� +w+`'*� `� �� +�� &/'`(� �j� +j)+*/&/.� ,'.`� �� &)++'�
+w+&.+}����,'*&+)�`�+��,/&(`(/+��&�+j)+*/&/.&�&/+`���+�.()j&��0(+���,'.``'�
+��)/.,�)`(/+�/j�+��'+�k�`(/+���+��).'/.�`(+}�-�,'/)&&j&�,'.`���'*-j+'�+w�))�&��&�
0�)`j'&� �� `'�+&)'(,`(/+� `� �� +�� .�).(+'(� `'�+&)'(,`(/+++� �� +w4'&{� `� �(+&(� )/+`'3+'�
+wk,'&&(/+�-*+()j}�-&�.*)�+(&.&���'*-j+�`(/+&{��(`&���*,(-*+*`()j&��{��*,+�+`���
,+j&(j'&� )+�&&&� �� ,'/`*(+&}� �&� ,'/`*(+&� �� ./�(0()�`(/+� �&� .(&`/+&� �*,/&+`� �&�
-'/j,.+`&� ).(.()j&� �}-}� .*`.�+&� /j� �)*`�+&�� &j'� +&� )jj&� �.(+/�`'.(+�+&� �&�
.(&`/+&}� -&� ./�(0()�`(/+&� �+`�'+`� +�� &`'j)`j'� ).'/.�`(+(++{� &/(`� ,�'� '*,j+&(/+�
*+)`'/&`�`()j��j�-'/j,.+`�|(&���|(&���+w4'&�)/..�)w&`�+�)�&���+w�)*`�+�`(/+{�&/(`�+�
,'.``�+`� +� ')'j`.+`� �w�j`'&� )/.,+k&� �� ./�(0()�`(/+&� �&� .(&`/+&� /j� ��
'./�+�-�).'/.�`(+(+}� .++&�)/+&`(`j+`��(+&(� j+� )/��)/.,+k{� +�)/���&�.(&`/+&{�
'*-(&&�+`� +w*`�`� �� +�� ).'/.�`(+}� �&� )/.,+k&� �� './�+�-� ).'/.�`(+(+� ,j|+`�
�*'/j+'� /j� +'/j+'� +w4'&� �j`/j'� �&� .(&`/+&� �� 0�8/+� 4�*��*,+��+`}� .+0(+{� +&�
).�,'/++&� �w.(&`/+&� '*-j++`� +w(+`*-'�`(/+� /j� +� '`'�(`� �&� .(&`/+&� &j'� +w4'&}� �&�
).�,'/++&��w.(&`/+&� (+`'|(+++`���,+j&(j'&�+(|�jk}�'w�/'��++&� '*-(&&+`� +��*,3`�
�&�.(&`/+&�&j'�+�'(+��w4'&�+/j|++.+`�&�+`.*`(&*�+/'&���+��,.�&�$��j�)�)+�)++j+�('�
� �!"���?O �}����&`�(+(`*{�+���(&,/+((+(`*�`�+w(.,/'`�+j)+*�('��&��(.�'&��w.(&`/+&�9��9��
+/j|++.+`�&�+`.*`(&*&��*,+�+`��&�).�,'/++&�&4$*�`�4$1~��-�.,/&�`��+}{���~��}�
��.*)�+(&.�-/j|'+�+`�+��0/'.�`(/+��&�`*`'�.�'&��w.(&`/+&�9��9��+w&`�`/j2/j'&�,�&�
(+�)/.,'(&���+w.j'��)`j++{�.�(&�(+�&`�)/++j�)j�+j'�(+`*-'�`(/+���+w4'&�+/j|++.+`�

�

� ����� �������������������������������� ����������������!������"��������������
QP�
�
0/'.*�&`�`'(j`�('���+��).�,'/++�-41�~���!��#��������#) $��������/��)j(�(+`'�-(`���
0�8/+��(')`��|)� +��,'/`*(+�*-&4����� �������*���� ��"� �����������{�j+��&�)/+&`(`j�+`�
��+��.�).(+'(�'*,+()�`(|��$.(�.�'���+��$`(++.�+{�~����}��&�.(&`/+&�,�'+`�+&�'`('*&�
�j�0j'�`���.&j'���+w�|�+)*���+��0/j').���'*,+()�`(/+�&'|+`�*-�+.+`���&/j')���
+�� 0/(&�,/j'� +�'(+�,�'+`�+�`�+*/&�+`.*`(&*{�.�(&� +&�.*)�+(&.&� (.,+()j*&�+� &/+`�,�&�
+)/'� (�+`(0(*&}� ��� ,'*&+)� �w.(&`/+&� ,�'+`�+&� `� +/j|++.+`� &�+`.*`(&*&�
)/+`'(j'�(`� �j� .�(+`(+� �wj+� .*./('� *,(-*+*`()j� �j� 0(+� �&� �(|(&(/+&� )++j+�('&�
���'-j'/+� �+�� /(+'-{� ��~��}� �&� �(.�'&� �w.(&`/+&� 9�4�9��� &/+`� ,'(&� +� ).�'-�
,'(+)(,�+.+`�,�'�+��).�,'/++�&�,~���"� ����#�����#) $���������~�}�.++�)/+`'3+�+j'�
(.,/'`�+j)+*�('��(+&(�)j�+j'�(+`*-'�`(/+���+��).'/.�`(+���0�8/+��(')`{�)/+2/(+`.+`�
�|)� +� )/.,+k� �� './�+�-� ).'/.�`(+(+� 4-1� �%�+'����'���� �!��#����� ��'�
��#�'� �����������{�/j���0�8/+�(+�(')`�+�*+(.(+�+`�+&�(+`'�)`(/+&��&,*)(0()j&�+`'�+&�
.(&`/+&�`�+w4'&�,/j'�0�)(+(`'�+��0/'.�`(/+��&�+j)+*/&/.&��4+�'�&�`��+}{���~���5`/�`��+}{�
~����� �/&�..�,�'�&`� `� �+}{� �����}� 'w�j`'&� ).�,'/++&� &/+`� (.,+()j*&� ��+&� +��
��+�.()j� �w�&&.+�-� �&� +j)+*/&/.&� (+�*,+��..+`� �� +�� '*,+()�`(/+� �� +w4'&{�
)/..� +/'&� �� +�� `'�+&)'(,`(/+� � �!"��� ?O��}� ��� ).�,'/++� 95/4� (+`'�-(`� �|)� +w4/&�
,/+�.*'�&�55��/&4,/+�55��`�,'.`�+��'*(+`*-'�`(/+��&�`*`'�.�'&�9��9���,'�&�+�,�&&�-�
�� +��.�).(+'(� `'�+&)'(,`(/++++� �5/+�'-� `� �+}{� ��~��}� 'w�j`'&� ).�,'/++&� )/..�
'�kk� (+`'|(+++`�,+j`3`���+&� +&�'*-(/+&� (+`'-*+()j&�+�'*(+`*-'�+`� +&� `*`'�.�'&�9��
9����+&�+&�'*-(/+&�'*-j+�`'()&�`�+&�'*-(/+&�`*+/.*'()j&��''�+*�`��+}{���~���5/+�'-�`�
�+}{���~��}�'.3{�)j�+`���++{�&`� (.,+()j*���+&� +w(+`*-'�`(/+��� +w.(&`/+�9���j�+(|�j��&�
'*-(/+&�.*`*'/).'/.�`()j&�*���&/+�(+`'�)`(/+��|)�+��,'/`*(+�9*~���3�,,&�`��+}{���~~�}�
.+� )� )j(� )/+)'+� +&� �(.�'&� 9�4�9��{� ,j� �w(+0/'.�`(/+&� &/+`� �(&,/+(+&� )j�+`� �jk�
).�,'/++&� (.,+()j*&���+&� +j'��*,3`� `� +j'� '`'�(`{�(+�)jw(+� &.+�)j� )� &/(`� +&�
.4.&�)j�)++&�(.,+()j*&�+/'&���+��'*,+()�`(/+���+w4'&}�*�'�+j'�'3+���+&�+w�&&.+�-�
�� +��).'/.�`(+�`� +�.�(+`(+��� +w(+`*-'(`*��j�-*+/.{� +��'*-j+�`(/+��� +wk,'&&(/+��&�
).�,'/++&��w.(&`/+&�`� +�� '*-j+�`(/+��� +j'� �)`(|(`*� &/+`�,'(./'�(�+&� �j�.�(+`(+���
+w./.*/&`�&(� )++j+�('� `� `(&&j+�('}� .+� 00`{� �� +/.'j&&� �+`*'�`(/+&� `/j).�+`� )&�
,'/`*(+&� /+`� *`*� (�+`(0(*&� )/..� *`�+`� �&&/)(*&� �� +w�,,�'(`(/+� �� ,�`./+/-(&� ).��
+w./..{� �/+`� +� )�+)'}�'�+&� +� )�+)'� �� +�� ,'/&`�`{� +�� &j'k,'&&(/+� �� -41�~� &`�
'`'/j|*���+&�+&�`j.j'&���.�j|�(&�,'/+/&`()�`���.�j`�-'����$`�(�+/�`��+}{������}�'�
,+j&{� +���(.(+j`(/+��� `&4$*{�j+� �&� (&/0/'.&��� +��).�,'/++�&4$*{�,�'� (+`'0*'+)�

�

� ����� �������������������������������� ����������������!������"��������������
QQ�
�
4/&���+&��&�)++j+&�)�+)*'j&&���,'/&`�`&�.j.�(+&��+�'/-*+/�(+�*,+��+`&�(+�j(`�
j+� �''4`� �� ,'/+(0*'�`(/+� �(+&(� )j� +w(+�j)`(/+� �� +w�,/,`/&� �4+4&|� `� �+}{� ��~~�}�
�w(&/0/'.� `&4$*�&`�k,'(.*�,'*0*'+`(++.+`���+&� +&� )++j+&�.�+(-+&{�`� )/+&`(`j�
�/+)� j+� )(+� �� )./(k� ��+&� +� `'�(`.+`� �j� )�+)'� ,j(&)j� +&� )++j+&� &�(+&� &/+`�
(+&+&(+&� �� &/+� (+.((`(/+}� 4|�+`� ./+� �''(|*� �j� +�/'�`/('{� +w*)j(,� �w�))j(+� �|�(`�
./+`'*�)j� +�� ).�,'/++��w.(&`/+&�&*�~� ��"� ���!������� /��&`� &j'k,'(.*���+&� +�
)�+)'���+��,'/&`�`�(+�*,+��..+`��j�-'����&�`j.j'&���*/`/(+-�`��+}{�������}�-``�
).�,'/++{� )j(� �� 0�(`� +w/2`� �� )� `'�|�(+� �� `.�&{� &'�� �*)'(`� ,+j&� +� �*`�(+� ��+&� +�
).�,(`'�&j(|�+`}�
� �

�

� ����� ������������������������� �����������
QR�
�
������������� ��
�
�
��� +j)+/,./&.(+� �&*�~�{� *-�+.+`� �,,+*� ���{� &+��� /j� &j.�`'(+{� �� �w�/'�� *`*�
(�+`(0(*�)/..�j+�,`(`�,./&,./,'/`*(+�+j)+*/+�('�k,'(.*�,'*0*'+`(++.+`���+&�
+&� )++j+&� +� ,'/+(0*'�`(/+� �-.�+-� `� �+}{� ~����}� .++� 0�(`� ,�'`(� �� +�� 0�.(++� �&�
+j)+*/,./&.(+&�+j)+/,+�&.(+&{�,�'.(� +&)j++&�/+�'`'/j|�&*�~{�&*��{�&*��}�-&�
,'/`*(+&� .j+`(0/+)`(/+++&� &/+`� (.,+()j*&� ��+&� �(|'&� ,'/)&&j&� )+*&� �� +w./.*/&`�&(�
)++j+�('�`�+j'��*'*-j+�`(/+�&`�(.,+()j*���+&���+/.'jk�`�,&���)�+)'&}�
�
=>���������������������� �f����������� ��
�
��� 0�.(++�&j)+*/,./&.(+�&j)+*/,+�&.(+�)/+&`(`j�j+�-'/j,���,`(`&�,'/`*(+&��/+`�
+�� 0/+)`(/+� (+(`(�+.+`� (�+`(0(*� &`� )++� �� ).�,'/++� �w.(&`/+&}� .+� 00`{� +��
+j)+*/,+�&.(+� �&*���{� ,j'(0(*� �� ,�'`('� �w/j0&� �� ����"�� ��*��{� �� *`*� +�� ,'.(�'�
).�,'/++��w.(&`/+� (�+`(0(*�-'*)���&��)�,�)(`*��� +('� +&�.(&`/+&�`��� +&��&&.+'� ���
*�����&j'�j+�./+*)j+��w4'&�+j����&4��`��+}{�~����}�5+���,�'�+��&j(`�*`*�./+`'*�j+�'3+���
&*�����+&� +���*)/+�+&�`(/+��� +��).'/.�`(+�&,'.�`()j��,'�&�0*)/+��`(/+}�*/j'�)+�{�
&*���'`('�+&�,'/`�.(+&{�)j(�'.,+�)+`�+&�.(&`/+&���+&�+&�-�.�`&�.*+&��j�)/j'&���
+��&,'.(/-+�&�`�)j(�(+�j(&+`�j+�0/'`�)/+�+&�`(/+���+��).'/.�`(+{�,/j'�,'.``'�
+w�&&.+�-� �&� .(&`/+&� 9�4�9��� �*.(+,/``� �+�� �+/{� ~����}� &*�~� �� *`*� (�+`(0(*� +�
,'.('� +(j� )/..� j+� �/+��+`� ,'/`*(+� +j)+*/+�('� +*)&&�('� �� +�� (/-+�&� �&�
'(/&/.&}�.+0(+{�&*���+w��*`*�(�+`(0(*�)j��j)/j,�,+j&�`�'�{�`�,j��w*`j�&�'+�`(|&�
�jk�0/+)`(/+&���)``�,'/`*(+�&/+`��(&,/+(+&����)4'`.j'��+��$.�)4+0/'�{�~����}�++�&�(`�
+*�+./(+&�)jw++�,�'`()(,��� +��'*-j+�`(/+��� +��(/-+�&��&�'(/&/.&�+� (+`'�-(&&�+`�
�|)� &*�~� `� )jw++� &`� )�,�+� �� +('� +&� .(&`/+&� ,/j'� ,�'`()(,'� �� +�� '*-j+�`(/+� ��
+wk,'&&(/+�-*+()j��5�����`��+}{���~���9j�+-�`��+}{��������/`/(�`��+}{������}��
�&�,'/`*(+&�&*�~{�&*��� `�&*��� &/+`� )/�*&�,�'��&� -�+&��(00*'+`&� +/)�+(&*&� ).��
+w./..�&j'� +&� '*-(/+&�).'/./&/.()j&��)��{��,�~}��`�~�)��}�~{� '&,)`(|.+`}��j'�

����������������� ����������������������������� ��� �����!�"�#�!�"�����!�"#��-��� %� ��������� ���,���� ���� ������ ����'���������� ��,�� �����%��)� .���� �������l’�������������������������������� ������������%��� ����7�������� �� �����������������4��� 4!� ��� 4%)� ��� ������ ���#&+'���������� ���� #������� ����� %����#��)� .���� ���� �����*��������l’interaction��%���d’autres����������� ������������'������������������� ����#�����%��� ����������������*�������� ���������'�'� "������� ���������#)�������!������,������������������������������������"��������������������������������#)�D’après�������0���!$�$)����
�������#$��"����������������������������%�����!�"������ ������� �'���������� ���� caractérisé�� ���� ��� ������ ��� ������� �������#��� ��l’oligomé����������� l’hétérodimé�������)�����������d’export�nuclé����� "�.�#� ,�������������pré����� ���� ������ ré���� )� ��� ������ ��������� �� ��� proté���� ��������!� �������������"4�#� �������������� ��� ���������&�7����������������������� �����������nuclé�����#���������"���#5�������ré�������������*��������l’����%������#���������������������)����-'�������������� proté���� possè�� ���������� ��������%���� ribonuclé���� ��� �������� ��� ré����� #���*����impliqué��� ���� ��� ������� ��&� ������ nuclé�*���)� .�,���� ��� ���� ���%�� ���� ��� ������������*��� spé��,�*��� �� l’��,���� �� *��� �������� !� ré����� ��+���7���� "!66� ��� !8$#�né��������� à���������������nuclé�����������proté����"���#)�D’après����������������!$$7)��
��

� ����� ������������������������� �����������
QT�
�
�/.�(+� )/j'� �.(+/�`'.(+�+� &'|�+`� �� +w/+(-/.*'(&�`(/+� `� �� +w�)`(|(`*� ).�,'/++�
�w.(&`/+� &`� +� ,+j&� )/+&'|*� �1'.+()4� `� �+}{� �����}� 4j� )/+`'�('{� +�� ,�'`(� -�`'.(+�+{�
&'|�+`� +/`�..+`� �� +w(+`'�)`(/+� �|)� �w�j`'&� ,'/`*(+&{� /j� �� +�� +(�(&/+� �jk� �)(�&�
+j)+*()j&���+&�+�)�&���&*�~{�&`�+��,+j&�|�'(�+}��&���..'&���+��0�.(++�,/&&��+`�
j+� &(-+�+� �� +/)�+(&�`(/+� +j)+*�('� (,�'`(`{� .�(&� &j+� &*�~� ,/&&��� j+� �/.�(+� ��
+/)�+(&�`(/+�+j)+*/+�('�`�j+�&(-+�+��wk,/'`�+j)+*�('����!�&��?Q�}���
&*�~�'&`�+�..'���+��0�.(++�+�,+j&�*`j�(*��j�0�(`���&/+�k,'&&(/+�j()j(`�('�`���
&/+�(.,+()�`(/+���+/.'j&&�,�`./+/-(&�)�+)*'j&&}�
�
?>�-&!� ��'�� �'&��'�&������'��� �'�� �����,�����=�
�
��� -�+� &*�~� )/.,/'`� ~�� k/+&� ).�� +w./..� `� )/�� ,/j'� �� (&/0/'.&��� &*�~}~{�
&*�~}��`�&*�~}�}����,'/`*(+�&*�~}~�&`�/`+j��,'�&�`'�+&)'(,`(/+��&�~��k/+&{��+/'&�
)j�&*�~}��&`�/`+j�-'*)���j+�*,(&&�-��+`'+�`(0���+&�+��,�'`(��w���+w4/&.{�'*&j+`�+`�
+� j+�,'/`*(+�,+j&� )/j'`� �/.4����`� �+}{� ~����}�*j��w*`j�&�,/'`+`� &j'� +w(&/0/'.�
&*�~}�}�++�&�(`�`/j`0/(&�)jw(+�+j(�.�+)j�j+�,�'`(�(+`'+��j��/.�(+�)�'/k��`'.(+�+�
�-�`'�{� &�+&�,/j'��j`�+`�+�)/++�*`'� +&�)/+&*)j+)&� 0/+)`(/++++&}� -/..� `/j`&� +&�
,'/`*(+&��� +�� 0�.(++�+j)+*/,+�&.(+�+j)+*/,./&.(+{�&*�~�,/&&���j+��/.�(+�)/j'�
�.(+/�`'.(+�+� �&�`'�� `'�&� )/+&'|*}� -��/.�(+� )/+`(+`� j+� &*)j+)��w�)(�&��.(+*&�
�)(�&{� �,,+*� 4~{� `� )j(� ,'.`� +/`�..+`� &�� +(�(&/+� �|)� +&� .(&`/+&� 9�� `� 9�}� ���
,'*&+)��&�&*)j+)&��w�)(�&��.(+*&��)(�&�4��`�4��&(`j*&�+��|�+��j��/.�(+�)/j'�
&`� (+�(&,+&�+��� +�� 0(k�`(/+��&�.(&`/+&�9�4�`�9����+4j��4(�`��+}{����~�}����,�'`(�-�
`'.(+�+���+��,'/`*(+{�'`'/j|*�j+()j.+`���+&�&*�~{�&`�)/+&`(`j*��w�)(�&��.(+*&�
�&()j&�&j(|(&��w�)(�&��.(+*&��'/.�`()j&{�`�&`� (.,+()j*�+/`�..+`���+&� +�� 0(k�`(/+�
�jk� �)(�&�+j)+*()j&� �(+&(�)j���+&� +�� 0/+)`(/+� '(/+j)+*�&()j� � �9(+-/'�+(� `� �+}{� ������
��+-�`��+}{� ~����}� -w&`�*-�+.+`���+&�)``�,�'`(�)jw&`� &(`j*� +� &(-+�+��� +/)�+(&�`(/+�
+j)+*/+�('����!�&��@S�}��
&*�~� &`� ,'*&+`� &&+`(++.+`� &/j&� 0/'.� �w.k�.�'&{� (+� )jw++� ,j(&&� k(&`'�
&/j&� 0/'.� ./+/.*'()j� �/.4���� `� �+}{� ~����}� �� ,�&&�-� �� &*�~� �&� 0/'.&�
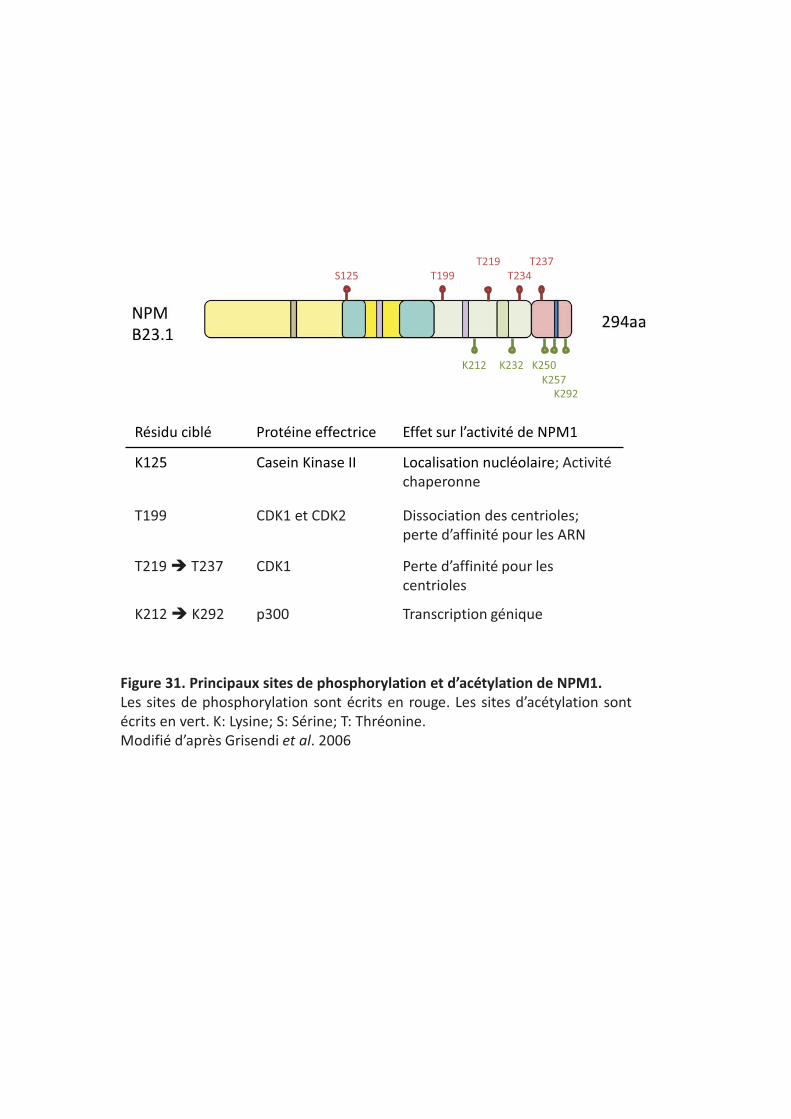
�1�6� ,122� ,�3"�,�34�,�12�
.�1�� .�3�� .�6#�.�64�
.�2��
���5 ��5#6�� 8����5���� ����5��� Effet sur l’activité de��8�1�
.1�6� 9���5��.5�����::� '���65���5���� �6��6�5��$�4��5!5����%���������
,122� 9;.1����9;.�� ;5����5��5�����������5�6��$�perte d’affinité pour les ARN�
,�12���,�34� 9;.1� Perte d’affinité pour les �����5�6���
.�1����.�2�� �3##� ,������5��5���*��5� ��
����%���(���%���������������������� ����������d’acétylation��������'����5�������%���%���6��5�����������5�������� *�9�'����5����d’acétylation���������5������!���9�.+�'��5��$��+����5��$�,+�,%����5��9���5 5��d’après�<�5���5������9��##;��

� ����� ������������������������� �����������
QK�
�
./+/.*'()j&� �jk� 0/'.&� .k�.*'()j&� 2/j� j+� '3+� (.,/'`�+`� &j'� +�� 0/+)`(/+� �� +��
,'/`*(+�,j(&)j�&/+��-'*��w/+(-/.*'(&�`(/+� (+0+j+)�&�� +/)�+(&�`(/+�+j)+*�('��)(�+�`��+}{�
�����}� .+� '*,/+&� �� &�� ,./&,./'�+�`(/+� ,�'� +�� -'3� )�)��� �j'�+`� +��.(`/&{�&*�~� 0/'.�
*-�+.+`�j+�.*`*'/�(.�'��|)�&*�~}�{�)�)j(��(.(+j�&/+��00(+(`*���+(�(&/+��|)�+w4/&{�
&j--*'�+`� j+� '3+� (+.((`j'� ��&*�~}�� &j'� +�� (/-+�&� �&� '(/&/.&� �+4j��4(� `� �+}{�
������}�5+����j&&(�*`*�.(&�+�*|(�+)��&� (+`'�)`(/+&��|)��w�j`'&�..'&��� +��0�.(++�
�&�+j)+*/,+�&.(+&�+j)+*/,./&.(+&{�)/..�&*��}�.+0(+{�(+�&`���+/`'�)j�+w�&&/)(�`(/+�
��&*�~��|)��w�j`'&�,'/`*(+&�+/+��,,�'+`*&���*`*�'�,,/'`*{�)/..�)w&`�+�)�&�,/j'�
+�'*),`j'��&��+�'/-�+&��4/�}�-``�,�'`(�&'���*|+/,,*�,+j&�+��*`�(+���+��0(+���)�
).�,(`'}�
�
@>���,���������������Z���,������� ���,�����=�
�
&*�~� &j(`� �� +/.'j&&� ./�(0()�`(/+&� ,/&`�`'��j)`(/++++&� ���!���� @=�}� ���
,./&,./'�+�`(/+�2/j�j+�'3+�)'j)(�+�,+��+`�+�)�)+�)++j+�('}���)/.,+k�)�)+(+�.�-'3��
(+�j(`�+��,./&,./'�+�`(/+���&*�~�&j'�+��`.'*/+(+�~���,+��+`�+��,.�&�5~{�)�)j(�,'.`�
&���(&&/)(�`(/+��&�)+`'(/+&�`�+w(+(`(�`(/+���+���j,+()�`(/+��j�)+`'/&/.���/4j��.��`��+}{�
���~�}� '�,+j&{�&*�~� (+.(� +w*,(&&�-��&�,'*�4/&.� +/'&)jw++�&`�,./&,./'�+*� &j'� +��
`.'*/+(+�~��� ���'�,/'�`��+}{������}����,./&,./'�+�`(/+��&� `.'*/+(+&�~��{��~�{�����`�
�����j'�+`� +��.(`/&�,�'�-'3~{�j+��j`'�..'��� +��0�.(++��&�-'3{��/+(`� +w�00(+(`*���
&*�~�,/j'� +&�4/&��+4j��4(�`��+}{������}���� +/)�+(&�`(/+�+j)+*/+�('���&*�~�*`�+`��j�
+�,�'`(�,�'�&��0/'`��00(+(`*�,/j'� +&�4/&{� (+�&`�,/&&(+�)j�)�.*)�+(&.�,�'`()(,��� +��
&/'`(� ��&*�~� �&� +j)+*/+&� `� ,'.``� �(+&(� +j'� �*&�&&.+�-}�++� ,j`� *-�+.+`�
)(`'� +�� ,./&,./'�+�`(/+� &j'� +�� &*'(+� ~��� ,�'� +�� )�&*(+� 4(+�&� 55� �-355�� )j(� '*-j+� +��
+/)�+(&�`(/+� +j)+*/+�('� �(+&(� )j� +w�)`(|(`*� ).�,'/++� �� &*�~� �&-(� �+�� ++&/+{� ������
$�+(� `� �+}{� �����}� 'w�j`'&� ./�(0()�`(/+&� ,/&`�`'��j)`(/++++&� /+`� *`*� (�+`(0(*&{�
)/..�+wj()j(`(+�+�`(/+{�+��$/�+�+�`(/+�/j�+)/'�+w�)*`�+�`(/+}����,/+�j()j(`(+�+�`(/+���
&*�~�,j`�4`'�'*�+(&*�,�'�+�)/.,+k��/-4~��4/'~�/j�,�'�+��(�(&��&�&j,,'&&j'&���
`j.j'&�,~�4/1�,~�4/1{�(+�)j���+&�)��'+('�)�&{�+wj()j(`(+�+(-�&�(.,+()j*�+�&/(`�
,�&�)/++j��5`�.�+��`��+}{��������$�`/�`��+}{������}�'�+&�+&��jk�)�&{�)+��(+�j(`�j+�,'`�
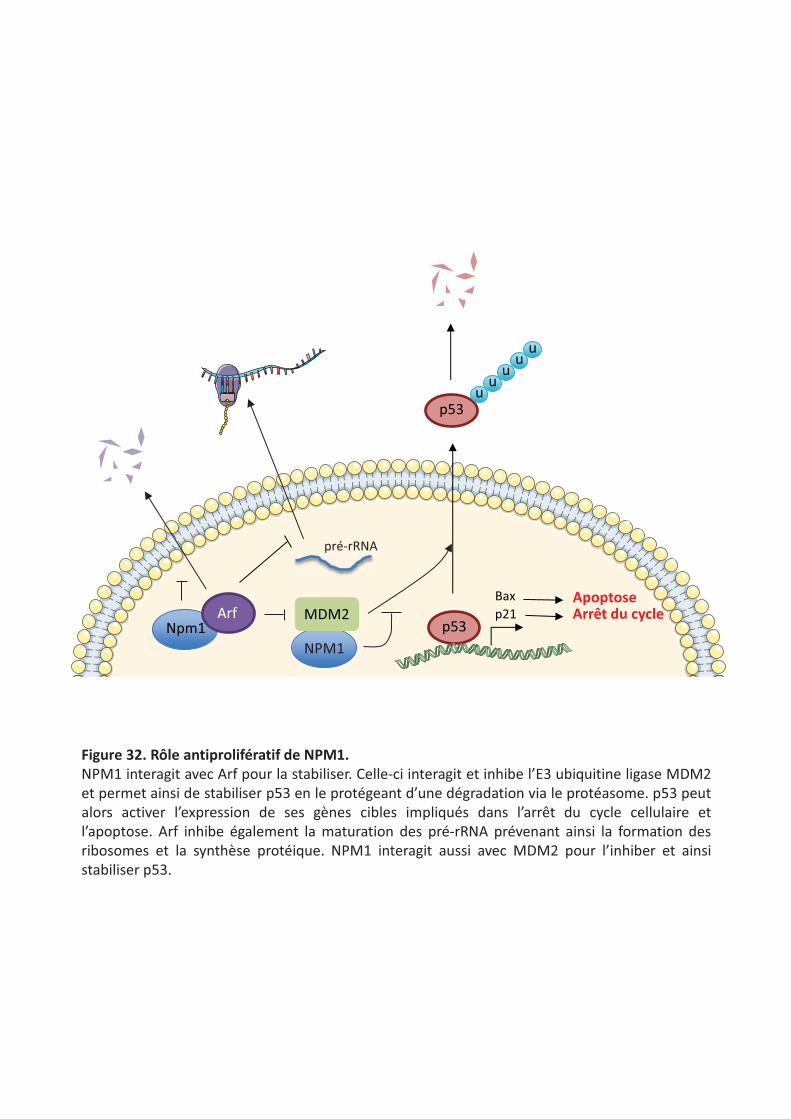
�����4���
���'��
�������������
����� �� "�������
�������� ����
�������4�
�������������������������������������������������� ���� ����4��������� ��� ���������-������������� ��������������l’E���� ���������� ���'������������ �����!� �� ������������������������ ���d’une�!��� ! ������� ��������� �������������� ����� ������� l’expression� !�� ���� �"���� ������� ����� ���� ! ��� l’arrêt� !�� �#���� ������ ���� ���l’apoptose�� 4��� ������ �� ������� � �� ��� �����!����������4������� ��� ����� � � ���� �����!������������� ��� � � �#���"��� ������ �������� ����� ���� ����� ����'�� ����� l’inhiber� ��� ������� ������������
����

� ����� ������������������������� �����������
QL�
�
$w�)`%|%`*� &`� j'&� $*-(�$�`%)'� $&� &*�~� *�(� +�� |)%&� $j� *()`*�,)-&}� ��� $/�+�+�`%)'� &`�
+w�)*`�+�`%)'�$&�&*�~�.)j&'`�(&,*&)`%|&-&'`�,j(�+��+))�+%,�`%)'�$&�+��*()`*%'&��%',%�/j&�,j(�
,��)�*�)%`*���(*-j+&(�+��`(�',)(%*`%)'�-*'%/j&{�&`�,&()'`��)($*,�*�(�+��,j%`&}�
*�(� ,&,�-j+`%*+&,�$)-�%'&,{�*�(� +�� |�(%*`*�$&� ,&,� %'`*(�)`&j(,{��%',%�/j&�*�(� +&,�-j+`%*+&,�
-)$%0%)�`%)',�*),`�`(�$j)`%)'&++&,�/jw&++&�,j%`{�&*�~�(*-j+&�$%|&(,&,�0)')`%)',�&,,&'`%&++&,�
�� +w0)-*),`�,%&� )&++j+�%(&}� -&,� *())&,,j,� *&j|&'`� 4`(&� �+`*(*,� +)(,/j&� +w&k*(&,,%)'� )j� +��
,`(j)`j(&�$&� +��*()`*%'&�&,`�-)$%0%*&� ,j%`&� ��$&,�-j`�`%)',� -*'*`%/j&,�)j�**%-*'*`%/j&,{�
))'`(%j�'`��%',%���+w�**�(%`%)'�&`�)j���+��*()-(&,,%)'�$&�')-(&jk�)�')&(,�)0&��+w0)--&}��
�
E>� � �!�,�����=�,�"!� �!���"��#!�
�
�&,��+`*(�`%)',� `)j)0�'`��j�-�'&����/� ,)'`� `(�,� 0(*/j&'`&,�$�',�$&�')-(&jk� `�*&,�$&�
)�')&(,� )0&�� +w0)--&}� 'w�)($� ))',%$*(*&� ))--&� j'�-�(/j&j(�$&� )&++j+&,� `j-)(�+&,�&`�
))--&� j'� *)`&'`%&+� *()`)�)'))-�'&{� &*�~� *&j`� *-�+&-&'`� �|)%(� $&,� 0)')`%)',� )'))�
,j**(&,,%|&,� ))--&� )w&,`� +&� )�,� $�',� +&,� )�')&(,� 0*-�`)*)1*`%/j&,}� �w*`j$&� $&�-)$�+&,�
)&++j+�%(&,� &`� -j(%',� *(*,&'`�'`� $&,� �+`*(�`%)',� $�',� +w&k*(&,,%)'� $&� &*�~� �� *&(-%,� $&�
-%&jk�$*0%'%(�,&,�-j+`%*+&,�0)')`%)',�%)+)-%/j&,�&`�+&j(,�%-*+%)�`%)',�$�',�+��`j-)(%-&'�,&}�
�
E>=>�� ���=��"$���� ��%#& ��##�'�&"�!(�'����("�!'#�!!�&"�&)#"�*(���'��!'�"���!!��#���� ��������������������
� !'��� �'#�)#"&+�*(�>�
� �
� '�������,1�������������'�������������������$��������+��
�
�w%'0%%`%)'� $&� +�� *()+%0*(�`%)'� %'$j%`&� *�(� &*�~� &,`� &'� *�(`%&� �,,))%*&� �|&)� ,�� )�*�)%`*�
$w%'`&(�)`%)'��|&)� +��*()`*%'&�*~�4/1� �*~�4/1�)0&�� +�� ,)j(%,�� ���)(#�� @?�}� -&``&�$&('%�(&�&,`�
j'&�*()`*%'&�'j)+*)+�%(&� %-*+%/j*&�$�',� +w�((4`�$j�)�)+&�)&++j+�%(&�&`� +&�$*)+&')0&-&'`�$&�
+w�*)*`),&�&'�(**)',&���j'�,`(&,,�)'))-*'%/j&�&'� %'`&(�-%,,�'`��|&)�$%00*(&'`,�*�(`&'�%(&,�
*()`*%/j&,}�'w�)(${�4/1�*&j`�%'`&(�-%(�&`�,*/j&,`(&(��'���$�',�+&�'j)+*)+&{�%'0%�'`��%',%�

�

� ����� ������������������������� �����������
QM�
�
,)'��)`%|%`*}�*���&,`��+)(,�,`�%+%,*&�&`�,)'��)`%|%`*� `(�',)(%*`%)'&++&�&,`��))(j&{�*&(-&``�'`�
+w�((4`�$j�)�)+&�)&++j+�%(&� |%�� +w&k*(&,,%)'�$j� -35�*�~�)j� +w�*)*`),&� |%�� +w&k*(&,,%)'�$&� ��k�
�$j++%|�'� &`� �+}{� ��~��� �&&(� &`� �+}{� ~����}� 4/1� *(*|%&'`� *-�+&-&'`� +�� *()+%0*(�`%)'� &'�
%'0%�'`� +��-�`j(�`%)'�$&,�4/&�*(*�(%),)-�jk���${���$�&`���$��$j-%-)`)�&`��+}{������}����
,`�%+%`*�$&�4/1�$**&'$�-(�'$&-&'`�$&�,)'� %'`&(�)`%)'��|&)�&*�~}�.'�&00&`{� +��$%-%'j`%)'�
$&�&*�~�*�(� %'`&(0*(&')&� 4/&�$%-%'j&� +�� ,`�%+%`*�$&� 4/1{��+)(,�/j&� +�� ,j(&k*(&,,%)'�$&�
&*�~�+w�j--&'`&��3j)�&`��+}{������}�'&�*+j,{�+��0)(-&�-j`�'`&�$&�4/1�%')�*�+&�$w%'`&(�-%(�
�|&)�&*�~�&,`� %',`�+&�&`�')'�0)')`%)''&++&}�-&,�0)')`%)',�)'))�,j**(&,%|&,�$&�&*�~�)'`�
*`*�%&'�*`j$%*&,�$�',�+&,�)�')&(,�0*-�`)*)1*`%/j&,}�.'�&00&`{�j'�,)j,�-()j*&�$&�+&j)*-%&,�
-�*+)1$&,��%-j&,�&,`�)�(�)`*(%,*�*�(�j'&�-j`�`%)'�$&�&*�~�(&+))�+%,�'`�+��*()`*%'&�$�',�+&�
)�`)*+�,-&� �&*�)��� �1�+%'%�&`��+}{� �����}� '�',� +&,�)&++j+&,�-�+%-'&,{�&*�)�� %'`&(�-%`��|&)�
4/1�&`�+��(&+))�+%,&�$�',�+&�)�`)*+�,-&{�%'0%�'`�+w�)`%)'�*()`&)`(%)&�$&�4(0�,j(�*����%',%�/j&�
,j(�+��%)-&'�,&�$&,�(%),)-&,���&,`&'�&`��+}{������}�'&�*+j,{�&*�)��,w0*`*()$%-*(%,&��|&)�
+&,�0)(-&,�,�j|�-&,�$&�&*�~{�/j%�'&�*&j|&'`��+)(,�*+j,�%'`&(�-%(��|&)�4/1�*)j(�*(*|&'%(�,��
$*-(�$�`%)'}��
&*�~�,`%-j+&�*-�+&-&'`� +w�)`%|%`*�$&�*��� %'$**&'$�--&'`�$&� 4/1� ���)(#�� @?�}�&*�~�,&�
0%k&�$%(&)`&-&'`���*���*)j(� +��,`�%+%,&(�&`�,`%-j+&(�,)'��)`%|%`*�`(�',)(%*`%)'&++&�,j%`&���j'�
,`(&,,�-*')`)k%/j&��-)+)-)�&`��+}{������}�'&�*+j,{�+w&k*(&,,%)'�$&�&*�~�&,`�%'$j%`&�*�(�j'&�
&k*),%`%)'��jk�(��)''&-&'`,�/}#}{�)&�/j%�*&(-&`�,)'� %'`&(�)`%)'��|&)�9'��{� +w)(`0)+)-j&�
0j-�%'�$&��'��}�-&``&� %'`&(�)`%)'�,`�%+%,&�*���&'� +w&-*4)0�'`�$&�,w�,,))%&(��|&)�9'����
�3j(2%�&`��+}{������}�-&,�$)''*&,�,)'`�(&'0)()*&,�*�(� +w),&(|�`%)'�/j&� +��,j(&k*(&,,%)'�$&�
&*�~� %'$j%`�j'&�,*'&,)&')&�$�',�$&,� 0%()+�,`&,�&k*(%-�'`�*��{��+)(,�/jw&++&� %'$j%`�j'&�
0�*&(*()+%0*(�`%)'�$�',�$&,�0%()+�,`&,�%'|�+%$*,�*)j(�*����5`�0�'��&`��+}{������}��
�
� '�������,1��4 ������������� ��������������� ������%3�������������������
� �������������������������� ����
�
&*�~�.)j&�j'�(3+&�%-*)(`�'`�$�',�+&�))'`(3+&�$&�+��(**�(�`%)'�$&�+w4'&}�-)--&�-&'`%)''*�
*(*)*$&--&'`{� +w&k*(&,,%)'�$&�&*�~�&,`� %'$j%`&��*(�,� %((�$%�`%)'�*�(�$&,� (��)''&-&'`,�
/}#}� &`� &,`� '*)&,,�%(&� �� +�� (**�(�`%)'� $&� +w4'&� ��j� &`� �+}{� �����}� '&� *+j,{� +�� *&(`&�

�$� �� ��� ��
�8�,�������
�8�,�
��',���
4�
56.��
�8�,�
��',���
4�
56.��
8'.,�
�4.�4�
�8�,� �8�,�
�4.�4� �4.�4�
���������� �� ����
���� ��������� �������������������������� �������� �������4���������������������� ���� ��������������� ����������������� ������ ����������� ����������� �� �������� ������ ��� ���� ��� �� ������� ������������ ����� �� ������ ��������� ��� ���� ���� ����!� �������� �� ��������� l’activité� �� �������� �� ��� 4���� ��� �� ���� ��������������� ������ ��������� ����������������������"�� ����������� �� ��������������"�� ��������������!�#�������� �� ������������"�� ������������������������� ������������������������������ ���������$�������$ �!��D’après��� ���� �������� %%&!�

� ����� ������������������������� �����������
RO�
�
$w&k'(&))%*+� $&� &*�~� $�+)� ,&)� 0%(*,�)`&)� &-(�*++�%(&)� $&� )*j(%)� '(*|*.j&� j+&�
�))j-j,�`%*+�$&�,��'(*`/%+&�'0*)'0*(�,/&���9�4'{�-�(.j&j(�$&)�)�))j(&)�$*j,&)�(%+)�$&�
,w4'&� �-*,*-*�&`��,}{� �����}� �&*�~�&)`�/-�,&-&+`�+/)&))�%(&��j�-�%+`%&+�$&� ,��',*0$%&}�
'�+)�$&)�)&,,j,&)�.j%&))&+`&){�&*�~�&)`�(&`(*j|/&��))*)%/&��jk�)&+`(*)*-&){�)&�.j%�%+0%&�
,&j(�$j',%)�`%*+}����'0*)'0*(�,�`%*+�$&�&*�~�)j(� ,��`0(/*+%+&�~���'�(� ,&�)*-',&k&�-�),%+&�
.�-'1��&+`(�%+&�)��$%))*)%�`%*+�$&)�)&+`(*)*-&){�)&�.j%� %+$j%`� ,��)/'�(�`%*+�$&)�)&+`(%*,&)�
&`� ,w%+%`%�`%*+� $&� ,��$j',%)�`%*+� $&)� )&+`(*)*-&)� �+2j$�� &`� �,}{� �����}� ��� (/�))*)%�`%*+� $&�
&*�~��|&)�,&)�)&+`(*)*-&)�)&�0�%`�$j(�+`�,��'0�)&���$j�)�),&�)&,,j,�%(&�&`�&)`�$/'&+$�+`&�
$&�,w�)`%|%`/�$&�$&jk�2%+�)&)�-%`*`%.j&){�*�1~�&`�&.1�4��-�*�&`��,}{�������40�+-�&`��,}{������}�
5,�&+� (/)j,`&� ,�� 0*(-�`%*+�$&�$&jk� )&+`(*)*-&)�.j%� |*+`� 0*(-&(� ,&)�$&jk�'3,&)�$j� 0j)&�j�
-%`*`%.j&){�'&(-&``�+`� ,��*++&�)/-(/-�`%*+�$&)�)0(*-*)*-&)����)����@@�}�.+��))*($��|&)�
)&)�(/)j,`�`){�,��'&(`&�$wj+��,,�,&�$&�&*�~�)0&��,��)*j(%)�&+`(�%+&�j+&�%+)`�%,%`/�-/+*-%.j&�
$j&� �� j+&� �-',%0%)�`%*+� $j� +*-(&� $&� )&+`(*)*-&)� �5(%)&+$%� &`� �,}{� �����}� -&)� )*j(%)�
$/|&,*''&+`� j+� )�+$(*-&� -�/,*$�)',�)%.j&{� )&� .j%� (&+0*()&� ,w0�'*`0�)&� $wj+� (3,&�
*+)*)j''(&))&j(�$&�&*�~����*�*�}��
�
E>?>���� ���!"�� �#$�,�����=�� %���#&��$%��,�$ �'� �%�(��� � #'�,� �
�
-*+`(�%(&-&+`��jk�)�+)&()�0/-�`*'*0/`%.j&){�&*�~�&)`�0(/.j&--&+`�)j(&k'(%-/&�$�+)�,&)�
`j-&j()� )*,%$&){�&`��j)j+&�-j`�`%*+�*j�$/,/`%*+�$j�-�+&�+w��'*j(� ,w0&j(&�/`/� (�''*(`/&}��
�w&k'(&))%*+� $&� &*�~� &)`� )*((/,/&� �� ,�� '(*,%0/(�`%*+� )&,,j,�%(&� &+� (/'*+)&� �� $&)� )`%-j,%�
-%`*-/+%.j&)��1&j&()`&%+��+$�/�+$���*{�~��~��5j%+�&`��,}{�~����}�.+�&00&`{�&*�~�&)`�$&'j%)�
,*+-`&-')�)*+)%$/(/&�)*--&� j+�-�(.j&j(�$&�)&,,j,&)�&+�'(*,%0/(�`%*+�&`�*j�)�+)/(&j)&)�
�'&(-j+*|��&`��,}{������}�4j�$&,��$&�)&)�*)&(|�`%*+){�',j)%&j()�/`j$&)�*+`�$&'j%)�-*+`(/�
,w&00&`�)�j)�,�$wj+&�)j(&k'(&))%*+�$&�&*�~�)j(�,w0*-/*)`�)%&�)&,,j,�%(&}��&)�'(%+)%'�jk�&00&`)�
%*,*-%.j&)� .j%� *+`� /`/� *)&(|/)� )j%`&� �� j+&� )j(&k'(&))%*+� $&� &*�~� ��� *����� � )*+`� j+&�
�j--&+`�`%*+�$&� ,��'(*,%0/(�`%*+�&`� ,w%+0%%`%*+�$&� ,��$%00/(&+)%�`%*+�&`�$&� ,w�'*'`*)&{� `(�%`)�
)�(�)`/(%)`%.j&)�$wj+&�`(�+)0*(-�`%*+�+/*',�)%.j&}�
�
�

�������4�46��
�������4�41��
�������4��#�� �������4�5���
��������� �������� ���������
9���� �����
����� �����
����� �����
� �������������������������� ������������ ����� ���������� �����4��� ������������������ ��������������d’un���������������������4�������� ������������������ ���������������� �!��"����������#���$�������������� ����������%��������&��$�������������� ����������%'�(��������� ���� �������� ������()�������������"���� ���l’ITS������ � �"���� �� ������������� ���"��������4�*��������4���+������+�'�
�()��
#��� #���
#���
#���

� ����� ������������������������� �����������
RG�
�
� '������������������� �����,1�������� �����)��������� ����������
�
��� ,j-&k.-&,,%/0� $&� &*�~� 1�0&� �� j0&� �))/2/-�`%/0� $j� )�)2&� )&22j2�%-&� �2/-,� 3j&� ,��
$%1%0j`%/0�.�-� %0`&-4/-&0)&�4/&�,`/..&� 2��.-/2%4/-�`%/0{�$�0,�.2j,%&j-,�1/$�2&,�$&� 2%50/&,�
$&�)&22j2&,�)�0)/-&j,&,�0j1�%0&� ��%�&`��2}{���������05�&`��2}{���~~�}�*2j,%&j-,�1/)�0%,1&,�
/0`�/`/� %$&0`%4%/,�./j-�&k.2%3j&-�)&� -62&�.-/2%4/-�`%4�$&�&*�~�)/11&�,/0� %0`&-�)`%/0��|&)�
2w4'&�./2�1/-�,&���3j%�,`%1j2&�2��-/.2%)�`%/0�$&�2w4'&����7&1j-��&`��2}{�~�����}�-&.&0$�0`{�
2wj0�$&,�-62&,�.-%0)%.�jk�$&�&*�~�$�0,�2��.-/2%4/-�`%/0�)&22j2�%-&�&,`�$j���,/0��)`%|%`/�$�0,�2��
%/5&0�,&�$&,�-%/,/1&,}�.0�&44&`{�&*�~�&,`�)�.�2&�$w%0`&-�5%-��|&)�2&,��)%$&,�0j)2/%3j&,�
&`� 0/`�11&0`� 2&,� 4/&� -%/,/1�jk� �4/&-�{� &`� )&``&� %0`&-�)`%/0� &,`� 0/)&,,�%-&� ./j-� 2��
1�`j-�`%/0�&`�2w&k./-`�)�`/.2�,1%3j&�$&,�4/&-�0/j|&22&1&0`�,�0`0/`%,/,���%0$,`-81{���~��}�
'&�.2j,{�&22&�./,,�$&�j0&��)`%|%`/�-%/0j)2/�,%3j&�&`�&22&�&,`�)�.�2&�$&�)/j.&-�2&,�.-/�4/&-�
�j�0%|&�j�$&� 2w5�$���������� �%�������)�'��������<��./j-�.&-1&``-&� 2��1�`j-�`%/0�$&,�4/&-�
�}�� &`� ��$� �$�|7j-� �0$� +2,/0{� ~����� ���!"#�� @E�}� -/04/-1/1&0`� �� )&,� /,&-|�`%/0,{� 2��
$%1%0j`%/0�$&�&*�~�.�-� %0`&-4/-&0)&�4/&�-/$j%`� 2��1�`j-�`%/0�$&,�4/&-�$�0,�j0�1/$�2&�
)&22j2�%-&�$w/,`//,�-)/1&� �5`�0�0��&`��2}{������}�&*�~�,`%1j2&�/5�2&1&0`� 2�� `-�0,)-%.`%/0�
$&,�4/&�-%/,/1�jk��%0,%�3j&�)&22&�$w�j`-&,�5�0&,�%1.2%3j/,�$�0,�2��`-�0,4/-1�`%/0�1�2%50&�
&0�)/0`-62�0`� 2w�)`%|%`/�`-�0,)-%.`%/00&22&�$j�4�)`&j-�/0)/5�0&�)�1�)���%��0$�9�00{���~����%�
&`� �2}{� �����}� '&� )&``&� 4�9/0{� &*�~� .�-`%)%.&-�%`� �� 2�� .-/2%4/-�`%/0� )/0`%0j&� $&,� )&22j2&,�
)�0)/-&j,&,�&0� -&04/-9�0`� 2��%/5&0�,&�$&,� -%/,/1&,�&`�$/0)� 2�� ,�0`0�,&�.-/`/%3j&}� 4j�
$&2��$&�,/0�-62&�,j-�)�1�){�.2j,%&j-,�/`j$&,�/0`�-�../-`/�j0�-62&�$&�&*�~�$�0,�2��-/5j2�`%/0�
$&�2��$�0�1%3j&�)0-/1�`%0%&00&�&`�2��`-�0,)-%.`%/0�5/0%3j&�./j-�.�-`%)%.&-���2w&k.-&,,%/0�/j�
�� 2��-/.-&,,%/0�$&�5�0&,�/0)/5�0&,�/j�,j..-&,,&j-,�$&�`j1&j-,{�-&,.&)`%|&1&0`}�'�0,� 2&,��
)�0)&-,�/-�jk{�&*�~�&,`��)/`�2/&�.�-� 2w0%,`/0&��)/`�2�`-�0,4/-�,&�.���{�)&�3j%�,`%1j2&�,/0�
�)`%|%`/�$&�)0�.&-/00&�$w0%,`/0&�./j-�-&1/$&2&-�2��)0-/1�`%0&�&`�)/0`-62&-�2w&k.-&,,%/0�$&�
5�0&,� %1.2%3j/,� $�0,� 2�� `-�0,4/-1�`%/0� 1�2%50&� �$0�0$%2��� &`� �2}{� �����}� &*�~� &,`�
/5�2&1&0`� )�.�2&� $w%0`&-�5%-� &`� $w%00%&-� 9.'5�~{� j0� %00%%`&j-� $j� )/1.2&k&�/&4./2� 55{�
,`%1j2�0`��%0,%�2��`-�0,)-%.`%/0�5/0%3j&��5j-j1j-`0��&`��2}{������}��
�w�)`%|%`/�.-/2%4/-�`%|&�$&�&*�~�.&j`�/5�2&1&0`�6`-&�&k.2%3j/&�.�-�,/0�%0`&-�)`%/0��|&)�$&,�
|/%&,�$&�,%50�2%,�`%/0�)&22j2�%-&�$/.&0$�0`&,�$&,�4�)`&j-,�$&�)-/%,,�0)&�/j�$/.&0$�0`&,�$&,�
%0`&-2&j7%0&,}� 4%0,%{� ,j%`&� �� 2w�)`%|�`%/0� $&� 2�� |/%&� $&� ,%50�2%,�`%/0� )41�$�4�� .�-� 2&,�

4%41'�
�7'����������
%�7��7#������7�%����l’homéostasie cellulaire�
��&����!77�
������������������� ���
�����������
4����������
� �� �����
����� �� ������ �� �����
- �� ������� �� ���������������� ����� ��������
l’ADN nucléaire�
�4�
'� ����
)�$����������
����
���
�����������������������������������������'��� ����� ��������� l’homéostasie� ������ ���� ������� ������� �������� ��� l’ADN,� ����� � � ���������������!�� l’apoptose� � � � � ���� �����!���� ��"� .����� ���������� l’expression� ��� ��������� ��� ������� ���� ������ � �� ��� ���� ��� s’������������� ��� #������ ���� ����� $� � � ��# ��� ��� � ������!�����"� � ��� �� � �������%���!������ �������������������� � � #�� ����� d’un���������� ������ ����������������������������������4� #�&�������� �� ����"�� �� �� ��������� �� �� ������ ���� �������������������$�l’activation������ �� ���� ��� ������������� �� �� ����������� ����� ������ ��������� ��� ����"� L’ensemble� ��� ���� ���������� ������� $� � � # ����� ����� ��� l’ADN������ ��� ��� $� � �������� � � �������"� -�� �������������� �� ������� � � ������!�������������� ���� ��� ����������� � ��� ��� ��% �� ���� ������� � � # ����� ����� ��� l’ADN"� D’autres,� ������'� ��'� ����� ��!������ ������������� �4��� ����������(��� ��!������ � �� �� ��������� � �� ���"� � ����� ������� ��� ������) ��� ����� (��� ��� ���� � � l’expression� ��� ��������� ���� ������� ����*����������+������������$� l’activation����������������"� -������������ ��!������ ����� �� �������%���!������������ ������������������� ������� ���� ����� ������ ����� ��������!���� �����������(�! ��� ��������s’��������������d’être� �����"�
-�� ������������������� ��������� �
�����

� ����� ������������������������� �����������
RH�
�
%0`&,2&j7%0&-{� 2�� .,/`/%0&� $�4��� .0/-.0/,�2/&� -`%1j2&� 2w&k.,&--%/0� $&� &*�~}�
//)%.,/1j&1&0`{� 2�� .,/-&0)&� $&� &*�~� &-`� 0/)&--�%,&� �� 2w�)`%|�`%/0� $&� $�4��}� .02%0{�
2w%0`&,�)`%/0�&0`,&�2&-�$&jk�.,/`/%0&-�2�)%2%`&�2��`,�0-2/)�`%/0�0j)2/�%,&�$&�$�4���./j,��)`%|&,�
-&-�3�0&-�)%2&-��/&0�&`��2}{���~��}��
&*�~�&-`�/3�2&1&0`� %1.2%1j/&�$�0-� 2��.,/2%2/,�`%/0�$&-�)&22j2&-�)�0)/,&j-&-�$&�)/2/0�&0�
%00%�0`� 2�� -/0&-)&0)&� %0$j%`&�.�,�.���&0� ,/./0-&��� j0� -`,&--�/0)/3/0%1j&� ��/03�&`��2}{�
��~��}� .22&� &-`� $&� .2j-� ,&`,/j|/&� -j,&k.,%1/&� $�0-� $&-� /)0�0`%22/0-� $&� )�0)&,� $j� )/2/0�
0j1�%0-�.�,�,�../,`���$&-�/)0�0`%22/0-�-�%0-�/j�%--j-�$w�$/0/1&-�/0%0-}��
�
�
� '�������,1����������� �� ������� �"������������������������������ ��
� ��������������������
�
�w�./.`/-&� ,&./-&� -j,� 2w�)`%|�`%/0� $wj0&� )�-)�$&� $&� )2%|�3&-� .,/`/%1j&-� �--j,/-� .�,� 2&-�
&0��1&-�$&� 2�� 2�1%22&�$&-� )�-.�-&-}� .22&�.&j`�6`,&�$/)2&0)0/&�.�,� j0� -%30�2�&k`&,0&� �|/%&�
&k`,%0-�1j&��1j%�0/)&--%`&� 2w�)`%|�`%/0�$&� ,/)&.`&j,-�1&1,�0�%,&-�/j�.�,�$&-��2`/,�`%/0-�
%0`&,0&-���2��)&22j2&��|/%&�%0`,%0-�1j&��1j%�1/%2%-&�2&-�1%`/)0/0$,%&-����!����@H�}��&-����#" ��
)/0$j%-�0`� �� 2w�./.`/-&� .�,� |/%&� %0`,%0-�1j&� .&j|&0`� 6`,&� $&-� 2/-%/0-� $&� 2w4'&{� 2��
.,/$j)`%/0� &k)&--%|&� $w&-.�)&-� /k�3/0/&-� ,/�)`%|&-� �/+${� .�����*�� �9$���� $.&)%&-�{� $&-�
$/11�3&-� %1./,`�0`-� $&� 2�� -`,j)`j,&� )&22j2�%,&� /j� &0)/,&� j0� &0|%,/00&1&0`� .�j|,&� &0�
2�)`&j,-�`,/.0%1j&-}�52-�)/0$j%-&0`��j�,&2�,3�3&�$j�)�`/)0,/1&�)�.�,�2��1%`/)0/0$,%&{�.�,�2&�
%�%-�$&�2��.,/`/%0&�.,/��./.`/`%1j&��4'���� +<���������'�'��1j%�2/,1&�j0�./,&��j�0%|&�j�$&�
2�� 1&1,�0&� &k`&,0&� $&� 2�� 1%`/)0/0$,%&� ���,`%0/j� �0$� 5,&&0{� ���~�}� �&� )�`/)0,/1&� )�
-w�--/)%&� �2/,-� �|&)� 2�� .,/)�-.�-&� �� &`� 2�� .,/`/%0&� 4.�2�~� ./j,� 2/,1&,� 2w�./.`/-/1&� &`�
$/)2&0)0&,� 2�� )�-)�$&�$w�)`%|�`%/0�$&-� )�-.�-&{�1&0�0`� 2%0�2&1&0`� �� 2�� 2,�31&0`�`%/0�$&�
2w4'&�0j)2/�%,&�&`���2��1/,`�$&�2��)&22j2&��4/j�&`��2}{�~����}��
��� -j,&k.,&--%/0�$&�&*�~� ,&02/,)&� 2�� -j,|%&� )&22j2�%,&�&0� %00%�0`� 2w�./.`/-&�$&-� )&22j2&-�
)�0)/,&j-&-�-j%`&���$%22/,&0`-�-`,&--{�&`�$/0)�)/02�,&�j0&�,/-%-`�0)&��jk�`,�%`&1&0`-�)0%1%/��
&`�,�$%/`0/,�.&j`%1j&-}�.0�&22&`{�.2j-%&j,-�/`j$&-�1/0`,&0`�1j&�2��$%1%0j`%/0�$&�&*�~�.�,�
%0`&,2/,&0)&� 4/&� -&0-%%2%-&� .2j-%&j,-� 2%30/&-� $&� )&22j2&-� )�0)/,&j-&-� )/0`,&� $%22/,&0`&-�

�

� ����� ������������������������� �����������
RP�
�
$,/3j&-�`&2-�1j&�2&�`�k/2{�2��1%`/1�)%0&�-�/j�&0)/,&�2&�)%-.2�`%0&{�&`�)/0`,&�2w%,,�$%�`%/0��|&)�
$&-�,��/00&1&0`-�/}#}���&,3-`,�20�&`��2}{��������/�&`��2}{���~���$&70�,�&`��2}{���~��}�*2j-%&j,-�
1/)�0%-1&-�1/2/)j2�%,&-�.&j|&0`�&k.2%1j&,�2��,/-%-`�0)&�%0$j%`&�.�,�&*�~}�'w�/,${�&*�~�
%0`&,�3%`�&`�-/1j&-`,&�$�0-�2&�)�`/.2�-1&�2��.,/`/%0&�.,/��./.`/`%1j&��4'{�%00%�0`��%0-%�2��
2%/,�`%/0�$j�)�`/)0,/1&�)�&`�2w�./.`/-&�.�,�|/%&�1%`/)0/0$,%�2&���/�&`��2}{���~��}�&*�~�&-`�
�j--%�)�.�2&�$w%00%&,�2w�)`%|%`/�$&�.��{�1j%�&-`�j0�,/3j2�`&j,�)2/�$&�2w�./.`/-&�&0�,/./0-&�
�jk�$/11�3&-�$&�2w4'&}�.0�,/./0-&���j0�,��/00&1&0`�/}#}{�&*�~�&-`�.0/-.0/,�2/&�.�,�2��
.,/`/%0&� 7%0�-&� 4�/}� -&``&� .0/-.0/,�2�`%/0� &0`,�%0&� -�� 2%k�`%/0� �� .��� &`� &1.6)0&� 2��
.0/-.0/,�2�`%/0� $&� )&2j%�)%� -j,� -�� -/,%0&� ~�{� &--&0`%&22&� �� -/0� �)`%|%`/� `,�0-),%.`%/00&22&�
���%3j&2�&`��2}{������}�'�0-�$&-�)/0$%`%/0-�.0�-%/2/3%1j&-{�)&``&�%00%%`%/0�$&�.���.�,�&*�~�
.&,1&`�$&�2%k&,�j0�-&j%2�$&�,��/00&1&0`���.�,`%,�$j1j&2� 2w�)`%|�`%/0�$&�.���&-`�0/)&--�%,&�
./j,�-`/..&,�2&�)�)2&�)&22j2�%,&�&`�/j�$/)2&0)0&,�2w�./.`/-&}�'�0-�j0&�)&22j2&�)�0)/,&j-&{�2��
-j,&k.,&--%/0�$&�&*�~�|���j31&0`&,�)&�-&j%2�&`�.,/|/1j&,�2w�..�,%`%/0�$wj0&�,/-%-`�0)&��jk�
,��/0-�&`�j0&� %0-`�%2%`/�3/0/1%1j&�./j|�0`� 2�|/,%-&,� 2��.,/3,&--%/0� `j1/,�2&}�.��� %0$j%`�
/3�2&1&0`�j0&��./.`/-&�-j%`&���j0�-`,&--�/0)/3/0%1j&{�)/11&�2��-j,&k.,&--%/0�$&�)��--}�
'�0-� )&-� )/0$%`%/0-{� 2�� -j,&k.,&--%/0� $&� &*�~� %00%&� 2w�)`%|%`/� $&� .��� ./j,� 2�|/,%-&,� 2��
.,/2%2/,�`%/0�%0$j%`&�.�,�)�1�)���%�&`��2}{������}�&*�~���/3�2&1&0`�/`/�1/0`,/�)/11&�/`�0`�
j0�1/$%�`&j,�&0��|�2�$&�2��|/%&�$&�-j,|%&�)&22j2�%,&�*5�1�41��$�0-�2&�0/��j}��&�`,�%`&1&0`�$&�
)&22j2&-� -j,,/0�2%&00&-� $&� ,�`� �|&)� $j� &5�� ����*�� ��(�!� ������{� 2�)`&j,� $&� ),/%--�0)&�
./j|�0`�-`%1j2&,� 2�� |/%&�*5�1�41���&0`,�%0&� 2��.0/-.0/,�2�`%/0�$&�.0/-.0/%0/-%`%$&-�./j,�
2/,1&,�$&-�*5*����!���!��������'������+�!���!����}�&*�~�-&�2%k&�-j,�)&-�*5*��&`�2&�)/1.2&k&�
&*�~�*5*��%00%&�2��2,�31&0`�`%/0�$&�2w4'&�.�,�2��-4'�������������*���'��������&`�-`/..&�
2%0�2&1&0`�2&�.,/)&--j-��./.`/`%1j&��400�&`��2}{������}��
-/11&� %2� ��/`/� ,�../,`/�$�0-� 2��.�,`%&�.,/)/$&0`&{�&*�~�&-`� j0� �)`&j,� %1./,`�0`�$&� 2��
,/.�,�`%/0� $&� 2w4'&}� '�0-� j0� )/0`&k`&� .0�-%/2/3%1j&{� )&``&� 2/0)`%/0� &-`� 0/)&--�%,&� �j�
1�%0`%&0�$&� 2w0/1//-`�-%&�)&22j2�%,&}� .0� ,&|�0)0&{�$�0-�j0�)/0`&k`&�1�2%0{�)&``&�)�.�)%`/{�
&0� )/02/,�0`� $&� 1&%22&j,&-� )�.�)%`/-� $&� ,/.�,�`%/0� �jk� )&22j2&-� `j1/,�2&-{� �j,�� j0� ,42&�
%1./,`�0`�$�0-�2��-j,|%&�)&22j2�%,&�&`�$�0-�2��,/-%-`�0)&��jk�`,�%`&1&0`-�3/0/`/k%1j&-}�
.02%0{�&*�~�.,/`�3&� 2&-�)&22j2&-�)�0)/,&j-&-�)/0`,&� 2&-� -`,&--�&0|%,/00&1&0`�jk�3/0/,/-�
.�,� 2��),/%--�0)&� `j1/,�2&}� ���*�*�{� 2��),/%--�0)&� %0%`%�2&�$wj0&� `j1&j,��|�0`� 2w�03%/3&0�-&�
),/&�j0�&0|%,/00&1&0`�0�./k%1j&�&`��..�j|,%�&0�0j`,%1&0`-�&`�&0�2�)`&j,-�$&�),/%--�0)&-{�

�

� ����� ������������������������� �����������
RQ�
�
$&jk� -%30�jk� &0|%,/00&1&0`�jk� )/00j-� ./j,� $/)2&0)0&,� 2w�./.`/-&}� ��� -j,&k.,&--%/0� $&�
&*�~�./j,,�%`�.&,1&``,&��jk�`j1&j,-�$&�)/0`%0j&,�$&�),/8`,&�$�0-�)&-�)/0$%`%/0-�.j%-1j&���
~�� 2w&k.,&--%/0� $&� &*�~� %0$j%`&� .�,� 2&� 2�)`&j,� $&� `,�0-),%.`%/0� 55�~� �$��9��� ��'"��) ��
��������&0�)/0$%`%/0�0�./k%1j&�.&,1&`�$w%00%&,� 2��.0/-.0/,�2�`%/0�$&� 2��-/,%0&�~��$&�.���
&`��%0-%�$&�2/1j&,� 2w�./.`/-&� ��%�&`��2}{� �������� ��� 2w�)`%|�`%/0�$&�/�-� -`�%2%-&� 2��.,/`/%0&�
&*�~{�&`�)&2��&-`�0/)&--�%,&�./j,� 2��-j,|%&�$&� 2%,/2�-`&-�)j2`%|/-�-�0-�-/,j1� �-0/j��0$�
-j03{����~�}�
�
�� ��� ,�� ���� ,� !��"� �#� �$$���%&� ,� ��'�D� � ,���!��$�!���� ,�����=� ���&� !��������� ��
#D$�(!��&����� ��##�#����"� $���'��� #D���� ��� ��� #D�)�&�� ,�� ��&&�� $��&!� �� ��������� #��
&�(���!� &���,� ��,������&�����>���
� �

�

� ����� ������������������������������������#������� �� ����
RR�
�
��!�� &�&�� �,����*�&�,�����$���$��
�
����������D�����,D� ������������� �,�����=���$�$ ���
�
4|�0`�1/0��,,%|/&��j�2�/,�`/%,&�2w/1j%.&�$w�))j&%2���%$&0`%2%/�2��.,/`/%0&�)0�.&,/00&�&*�~�
)/11&� /`�0`� -j,&k.,%1/� $�0-� $&-� `j1&j,-� .,/-`�`%1j&-� 0j1�%0&-{� %0$/.&0$�11&0`� $j�
-)/,&� $&� 52&�-/0� ��//`/%03� &`� �2}{� ������}� '&� .2j-{� )&``&� /`j$&� �� .&,1%-� $w/`�2%,� &*�~�
)/11&�j0�0/j|&�j�)/��)`%|�`&j,�$&�2w�)`%|%`/�`,�0-),%.`%/00&22&�$&�4/{�j0��)`&j,�1�:&j,�$&�
2��)�,)%0/3&0�-&�.,/-`�`%1j&}�-/11&�$/),%`�$�0-�2&�)0�.%`,&�.,/)/$&0`{�2��-j,&k.,&--%/0�$&�
&*�~�&-`�/-&,|/&�$�0-�$&�0/1,&j-&-�`j1&j,-�$w/,%3%0&-�0%-`/2/3%1j&-�$%22/,&0`&-}��%&0�
1j&�$&�0/1,&j-&-�/`j$&-�-&�-/%&0`�%0`/,&--/&-��jk�1/)�0%-1&-�1/2/)j2�%,&-�,&2%�0`�&*�~�
�� 2�� .,/2%2/,�`%/0� )&22j2�%,&� &`� 2�� `,�0-2/,1�`%/0� 1�2%30&� ��� *����{� %2� 0w&k%-`&� .�-� �� 2w0&j,&�
�)`j&22&�$&�$/1/0-`,�`%/0�$&��)`%|%`/�$&� `,�0-2/,1�`%/0� ��� *�*�}� 42%0�$w&k.2/,&,� 2&� ,42&�$&�
&*�~� $�0-� 2�� .0�-%/.�`0/2/3%&� .,/-`�`%1j&{� 0/j-� �|/0-� $/|&2/../� j0&� -`,�`/3%&�
.&,1&``�0`�$&�3/0/,&,�$&-�-/j,%-�`,�0-3/0%1j&-�-j,&k.,%1�0`� 2��2/,1&�0j1�%0&�$&�&*�~}�
42%0�$&�$%,%3&,�2w&k.,&--%/0�$j�`,�0-3�0&�-./)%2%1j&1&0`�$�0-�2w/.%`0/2%j1�.,/-`�`%1j&{�0/j-�
�|/0-� .2�)/� -�� -/1j&0)&� 4'&)� &0� �|�2� $j� .,/1/`&j,� 1%0%1�2� $&� 2�� .,/�-%0&� $&� ,�`� ��
��������}� -&�.,/1/`&j,{�$/0/11/� 4//�*�{� ��$/:�� .&,1%-� j0� )%2�3&� $&� 2w&k.,&--%/0� $j�
3�0&�$&�2��)02/,�1.0/0%)/2��)/`�2`,�0-2/,�-&��-4����j�0%|&�j�$&-�2/&-�.,/-`�`%1j&-��40�03�
&`��2}{� �����}�� �������� �� ����,���� ,�� ������� ��� ��� ����� ��� ,��� ��� ���� ������������ ,��
�� ����,����������������� ���������������������,��� ���������,��������������,D� �� ����
�D����� ,D� �� ����������� � ,�����=� ���� ���� ������������������������ �� $�$ >������
������� ��� ������������� ��,����� ���������=�����,��� ��!���� ���� �����,� �� ���
��������� ��������� ���D� ������� ���f��������������� �,���� ����,������������>�
����������������� ��"��D����� ��,���������������� �,�����=�,� ������� ����,�����
����������$�$ ���
�

�

� ����� ������������������������������������#������� �� ����
RT�
�
52� 0w&k%-`&� .�-� �� 2w0&j,&� �)`j&22&� $&� $/00/&-� )/0)&,0�0`� 2w/,%3%0&� $wj0&� -j,&k.,&--%/0� $&�
&*�~�$�0-� 2&�)�0)&,�$&� 2��.,/-`�`&}��w�))j1j2�`%/0�$&-�`,�0-),%`-�)/$�0`�./j,�&*�~�$�0-�
2&-�)�0)&,-�$&� 2��.,/-`�`&�0/j-���.&,1%-�$&�./-&,�$&jk�0�./`0�-&-{�0/0�&k)2j-%|&-}�'wj0&�
.�,`{�&22&�./j,,�%`�6`,&�$j&���j0&�-`�%2%-�`%/0�$&�2w4/&1�$&�&*�~�$�0-�j0�)/0`&k`&�`j1/,�2}�
'w�j`,&�.�,`{�)&``&�-j,&k.,&--%/0�./j,,�%`�6`,&� 2&�,&22&`�$wj0&��j31&0`�`%/0�$&� 2w�)`%|%`/�$&�
2�)`&j,-�$&�`,�0-),%.`%/0����0`�&*�~�./j,�3�0&�)%2&}�*2j-%&j,-�$&�)&-�2�)`&j,-�-/0`�)/00j-�
./j,�,/3j2&,�2w&k.,&--%/0�$&�&*�~{�.�,1%�2&-1j&2-�)�1�){�55��~�{�/j�&0)/,&�--~��-0�0�&`��2}{�
~����� '�03� &`� �2}{� ������ �%� &`� �2}{� �����}� *2j-� ,/)&11&0`{� $&-� �0�2�-&-� $w%11j0/�
.,/)%.%`�`%/0� $&� )0,/1�`%0&� )/j.2/-� �� j0� -/1j&09�3&� 0�j`� $/%`� $&-� 2,�31&0`-� $w4'&�
,/)j./,/-� �-05*�-&1�� /0`� .&,1%-� $&� 1/0`,&,� 2�� .,/-&0)&� $&� 1�+/� -j,� 2&� .,/1/`&j,� $&�
&*�~�$�0-� 2&-�)&22j2&-�0/.�`%1j&-�1j,%0&-� ���*�*�� �-0�|&,/jk�&`��2}{���~���}�-&-�$/00/&-�
,/|�2&0`�j0�0/j|&�j�,42&�0j)2/�%,&�$&�1�+/�$�0-�2��,/3j2�`%/0�$&�2w&k.,&--%/0�3/0%1j&}�'&�
.2j-{�2w�2`/,�`%/0�2,/1j&0`&�$&�2��|/%&�*5�1�47`�1�+/�$�0-�2��`j1/,%3&0�-&�.,/-`�`%1j&�./-&�
1�+/� )/11&� j0� )�0$%$�`� .&,`%0&0`� $�0-� 2�� ,/3j2�`%/0� $&� 2w&k.,&--%/0� $&� &*�~}� */j,�
&k.2/,&,� )&``&�.%-`&{�0/j-��|/0-� `%,/�.,/2%`�$wj0�1/$�2&�$&� -/j,%-� %0|�2%$/&-�./j,� 2&�3�0&�
*`&0� $�0-� 2w/.%`0/2%j1� .,/-`�`%1j&� ���03� &`� �2}{� �����}� -&-� �0%1�jk� $/|&2/..&0`� $&-�
�$/0/)�,)%0/1&-� .,/-&0`�0`� j0&� -%30�`j,&� 1/2/)j2�%,&� -%1%2�%,&� �� )&22&� /-&,|/&� )0&��
2w0/11&� &`� )/0-`%`j&0`� �� 2w0&j,&� �)`j&22&� 2wj0� $&-� 1&%22&j,� 1/$�2&� $&� `j1/,%3&0�-&�
.,/-`�`%1j&}�'&�.2j-{� 2w�)`%|�`%/0�)/0-`%`j`%|&�$&� 2��|/%&�*5�1�47`�$�0-�)&�1/$�2&�-j%`&��� 2��
.&,`&�$&�*`&0�.&,1&`�j0&�0�.&,�)`%|�`%/0�$&�1�+/}�� �������� ������,������ ������� ,���
���������������� � ������,�������,�������������� ���,����������������,����������������
=[� ,�� ,����� ��� ��� #$%� �� ��!��� �D�������� � ,�� ���=�� ?[� ��� ����� ��� ����Y�[�
��� ���Y�[� ����������Y�[� ��� @[� ,�� ,����� ��� �D����� ,�� ��� �� ��!��� ,�� ���=� ���
#$%�,� ��� ��� ������������>�
�
� �

�

�����
Résultats��
� �

�

� $����������%�� ���&��
RL�
�
&%#'()��=�
�
���=������������� �� ,������ ������������� �� ���������������� ,������� ��� �������
/�2%7��/j$,�{�$�,%0����1j�%,&{�-/,%00&��/j,-�-�2&`{�5�&22&��/j&�j{�-�,%22&�$&�)/j--%0&�j{�
��,`%0��400��*/2�7/|�{�-/2%0�&%�,2&%k{���j,&0`��/,&2{�-2�j$&��&�j$/%0}�
�
�����5��������-�
�
��� 0j)2//.0/-1%0&� �&*�~������ &-`� j0&� .,/`/%0&� )0�.&,/00&� $w0%-`/0&-� 1j%� .�,`%)%.&� ��
2w�--&12�3&� $&-� 0j)2//-/1&-� &`� �j� ,&1/$&2�3&� $&� 2�� )0,/1�`%0&}� '&� 0/1,&j-&-�
�2`/,�`%/0-�2/0)`%/00&22&-�$&�&*�~�/0`�/`/�,/.&,`/,%/&-�$�0-�$%22/,&0`-�`�.&-�$&�)�0)&,�&`�
2&-�`,�|�jk�$&�2w/1j%.&�$w�))j&%2�/0`�1/0`,/�1j&�)&``&�$&,0%�,&�-w&k.,%1&�2/,`&1&0`�$�0-�2&-�
`j1&j,-� .,/-`�`%1j&-� )0&�� 2w0/11&}� 4� 2w0&j,&� �)`j&22&{� %2� 0w&k%-`&� �j)j0&� .,&j|&�
&k./,%1&0`�2&�$&�2��)�.�)%`/�$&�&*�~���.,/1/j|/%,�2��`,�0-2/,1�`%/0�1�2%30&�$&-�)&22j2&-�
/.%`0/2%�2&-�$&�.,/-`�`&�&`�2w/:&)`%2�$j�`,�|�%2�|%-&���&k.2/,&,�-/0�./j|/%,�/0)/3/0%1j&�)0&��
$&-�-/j,%-�3/0/`%1j&1&0`�1/$%2%/&-}�&/-�,/-j2`�`-�1/0`,&0`�1j&�2��-j,&k.,&--%/0�)%2/&�$&�
&*�~� $�0-� 2w/.%`0/2%j1� .,/-`�`%1j&� 3,*)&� �j� .,/1/`&j,� �0$,/3/0/�$/.&0$�0`� 4//�*��
-w�))/1.�30&� $wj0&� �j31&0`�`%/0� $j� ./%$-� $&-� 2/&-� $&� 2�� .,/-`�`&� )0&�� 2&-� -/j,%-�
`,�0-3/0%1j&-�&0� )/1.�,�%-/0��|&)� 2&-� -/j,%-� -�j|�3&-}� �%&0�1jw%2�0w���%`�.�-�$&� `j1&j,-�
)0&�� 2&-� �0%1�jk� `,�0-3/0%1j&-� �� 2w*3&� �$j2`&{� j0&� �0�2�-&� 0%-`/2/3%1j&� ,/|�2&� j0�
/.�%--%--&1&0`� $&� 2w/.%`0/2%j1� .,/-`�`%1j&� �--/)%/� �� j0&� �j31&0`�`%/0� $&� 2w%0$&k�
.,/2%2/,�`%2� .�,� 1�,1j�3&� �|&)� 2w�0`%3�0&� 1%���}� -&-� 1/$%2%)�`%/0-� -w�))/1.�30&0`� $wj0&�
�j31&0`�`%/0�$&�2w&k.,&--%/0�$&�3�0&-�)%2&-�$j�2�)`&j,�$&�`,�0-),%.`%/0�.��~}�*�,1%�)&jk�)%{�
2��0�j--&�$&� 2w�))j1j2�`%/0�$&� 2��)�)2%0&�.�&-`�)/,,/2/&���j0&�2/,`&�$%1%0j`%/0�$&-�`�jk�$&�
.��7%.~{� j0� %00%%`&j,�$&� 2��.,/3,&--%/0�$j�)�)2&�)&22j2�%,&}�&/j-�.,/./-/0-� j0�1/)�0%-1&�
-&2/0� 2&1j&2�&*�~�./j,,�%`� 2&|&,� 2�� ,/.,&--%/0�&k&,)/&�.�,�/� -j,� .��~}�*�,� )/0-/1j&0`{�
2w&k.,&--%/0�$&-�3�0&-�)/$�0`�./j,� 2��-�)2%0&�.�/j�&0)/,&�$1*�� 2�|/,%-&,�%`� 2��$/3,�$�`%/0�
$&�.��7%.~�)&�1j%��/2%,�%`�2��1j%&-)&0)&�$&�2w/.%`0/2%j1�.,/-`�`%1j&}��%&0�1j&�)&-��2`/,�`%/0-�

�

� $����������%�� ���&��
RM�
�
0&�-/%&0`�.�-�-j22%-�0`&�./j,�%0%`%&,�2��`j1/,%3&0�-&����*�*�{�%2�&-`�./--%2&�1j&�2w�--/)%�`%/0�
$wj0&� -j,&k.,&--%/0� $&� &*�~� �|&)� $w�j`,&-�2/-%/0-� /0)/3/0%1j&-� .j%--&� -`%1j2&,� 2��
.,/2%2/,�`%/0� $&-� )&22j2&-�1�2%30&-� &0� 2&|�0`� 2&� 2,&%0� 1jw&k&,)&� 2�� .,/`/%0&� .��7%.~� -j,� 2��
.,/3,&--%/0�$j�)�)2&�)&22j2�%,&�&`�2��.,/2%2/,�`%/0}�.02%0{�2��.&,`%0&0)&�$&�)&-�/-&,|�`%/0-�&-`�
��1&``,&�&0�.&,-.&)`%|&��|&)� 2&-�/-&,|�`%/0-� )2%0%1j&-� )0&�� 2w0/11&�1j%� ,/|�2&0`�1j&� 2��
.&,`&� $&� .��7%.~� $�0-� 2&-� )�0)&,-� $&� .,/-`�`&� ./--�$&� j0&� |�2&j,� .,/$%)`%|&� $j� ,%-1j&�
/|/2j`%2�$&-�`j1&j,-�&`�$&�2��,/)%$%|&��.,�-�`,�%`&1&0`�)0&��2&-�.�`%&0`-��``&%0`-�$wj0�)�0)&,�
$&�2��.,/-`�`&}�
�
� �

�

��
�
���������� ����������� � �������������������� � ������������ �
����� �� �������
�����������- #��!"�#� ��!�" $"$#$!% $&'!#"�!"�%!� �"$#('&' �)$&"�*�
�
�
&&%!+� -##�$&��,�-�� .&/$!"&� #&0#&!$ ��,�-�1�� �#$!"" � 5##$(2�&$ '��,�-�� ��$!$$ � � �
+##((!" &#��,�-��#&$'!"&�1""&� #$&+#�&3���#$!"�-!&$% !4��,�-��5&#$ "'�##$ $��,�-�&"���$&#� �
- &#�#!"��,�-5"�
�
�6"!� $(!'3� �$ $%#"'� 1#� $)" �� 6"!� $(!'3� -$&!( � &(�&$�� �& ��� - � �3337�� $28-333�
�$ $%#"'2$ $$&"���$$&"� 9�
,�-&.��6#&8,:-���& ���$28-33���$ $%#"'2$ $$&"���$$&"� 9�
-("( $%��6#&��3-���& ���$28-33���$ $%#"'2$ $$&"���$$&"� 9�
�
1�#$$ "'�&��$ ((���
�
5�#$$ ("#"�&"� �'#���$&#� �- &#�#!"9��-&.�6#&8,:-2�& ����&%"#(�6"!� $(!'&!$ �� (�
�3; ���3�1� "# �-$&!( � &(�&$��1#/!������������ ��������������������������������
����������������2�������������-���� � !� �"#$%����� &�'��
���������()*��&�� +,� ��������-.%��%��+���%��/�%���$�0��&� �&� ��
����������� �()*�1-��� ���� %- +�� ��)-�%� $�+� ����/��.��� ���%� �� &�/� �+���
2$���&� $��+& ���%� &�� ��34)��3#%-�+��/� �+���++ ���&���%� &�� ����5$��$�0��&� �

�

<�
�
��������
6�'�( %) +�4 � *6+,=--<>5� 4+� � � �.� /� &� �.40�4& �+�7� (��������� �����������
����� ���������������� ��������������� ����� ����������������������������� ���
����� �� �� �������� ������ � ��� �������� ������� � ����� ��� �������������
������������������������������� ����������������������������������������������
�������� ������� ��� ��� ������ ������� ������ �� ��������� ��� ���������� �����
����������� !���"��������������������� ������!������������������ #1$$%��&� �
������������ ���������� ����"�����!��� ���������� ����������������� �� �� � ������
������� ���� ������� ������ ����� ����� ���� �����!������� ������� ��� 1$$%��"�����
����������#�-"����&������ ���� ��� ���� ����#'(&��������)"������%"������!������
(���� ��������� ����������������������������������������!������ �������������������
���*�+,� ������������� ��� !����������!������������� ������� !���� �����-������
�����)"��."!������������!��$�1����������������2��#�/0�0%&��������������������
!��-"����������� 1�� ������� ������� ��� �������������%"!��� �������� �'(������
#�/0�00%&�����������������������������������������!����������������2�������
������������3�������� ���3�!��������������������2�������������!������������������
���������������������������������������������������������������������������
��������������������2�����2*�%�������������������2%4�"����������������������
��������� ��� ���� ������ � ������� �%,����� ���������� ���� ���������� ����� �� ��
����������������������������������������������������������������������������
�����������%,����� ������� �����������������������!�������������� ������������
����������������������������������������������������������!�����������������
������������ ������ !� ������ ������� ���������� �������� ����� � �������� ���
�����������!�����������������������������������������������

)�
�
������������
����������������-��������������,����������������-��8-�������,�������'��������
����������-��������04����������9��%0�019�1���,�����������������������������������,�
-����������-����,�������'�-������������������,�������������-������01�(1����
-�����������������'������������������������������������������0�������9��
%0��19� -����� 1�(5������������-�,���� ������� �������$�������������� ��������� ����
-�� ������ ���� �������� �-��� �.� � %0� ��-�� � ������� �� �������5��������
�������������� -��-� ��� ������$���02-���'��������9��%00?19� 1� ��� -���'������������� �
������� ��� �� ���,�� �$�� -�� ��������� ���-������ ,������,� ������ �������
��,��������'������ �-�����������,��9�
����'�-�������������-�����������'����������������-�����$��������������-�����
����'�����9�����������������,�������$��-����6���������������'�������5��*�0�������
���������*�����1����������� -�������������$�����,��������-�����������������'�-��
������9������������������'�-����-��� �����,���������-����-���,�������
'� -�� ����� ������ -��,-� -�� :��� 2�� :%� ������ �-����� 0@��������� �� ��9�� %00)19� 1'���
��,����� �������� �5���� �������� 0���� �%� ���� �)1�� ���������� ������ ���� ��� �� -��
'������ '� -�� ������ ������� �5��*.�8�� -��-� ��� ���� �-��-������� ���� ���������
���������� -��$���$������ ������$$� 01������ ���� ����� ����� ��� � ����� �� ����
���� ����������� ��������� ��� ���� 2��� ������ � ��!���"� �� ���#��$� ��� ���� �"��� ���� ���
����������$���$���� �������#� ����%1���������������#� �����&��'���� ������ ������1!��$�
��� �� $��� � � � ������(����� ����#� � ���� ������� 2� ������� 2� � ������ �(���� ��)�� ��#�
��$�$�� ��� �� �� ������ ���#*��+� ����� *�� ��� ����������$� ,*�� ���#��$� ��� �� � ������
�������������1��������!�������� ����� ���������*���������������)�����*�������)-����+����
*�� ������� 2���)��!�#����#� ��� ������������ ���#��$� ��� �� � ������ �!��� #�$��#������
�.����$����� ��� ���� ����/� '������ ��� ���� �������� � ����( � ���� ����� ����������� ��� �������

7�
�
21��44�����������������1���� ������������������������������������������������
���� �������� ���� ���� ����� ������� ��� ��� ���������� ��� ���� ����� ��� ��1�
���������������� !"#�$���������������������������������������������������������
������ ��� ���� ����� ������ ���� �������� � ��%�� ������� ������ ��������� ������
�&������������'�������� (( "#��
����������%���)*��������+������- !���%��������,!-"� �������������������
�����.����������������������%����������%���%��#�$����.�����!/�+���������
��%���� �����0��� � ��������������������� ��������������������.������ �����������
��� ���� ��������%� �-����� ��� ��#�� � - "#� �)*�� ���������� �������� ������ ��������
��%���������������.�������������������������������������,+���+�� ((-"#�1������
�)*�� ���� .��� ����� ��� .�� ������� � �.���%�� .�������� ������� �������� ���23�
���������������4�����1�������������%�����.�������1� ��������������������
��� ����� ������ ��������� ������� ��� ��#�� ((5"#��)*�� ���� �� ����������%������� �
���%�������� %��������� ������ ��� �.��%��� ��������%�� �����0���� � �� ������
��.�������������%�������������+�%����6��������#�� ((7"#�1���������������%������
�����������������%������)*�������������������4�����������������������������
��� ��������� ������ ���0���� ��� ��#�� � 58� ������� ��� ��#�� � /8� ��.��� ��� ��#�� � 8�
$��+�������#��� "#�
9�� ����� ���������� ����� ������)*�� �� �������� ���� ����������� ���%������ ���
��������� ������ ������ �&:����� ��� ��#�� ((-8� &��.���� ��� ��#�� (�;"� .��� �� ������
��%����������� ����������� ����������)*�� �� ����� ���+�#�<�������� ������� ����
��������� ������� ��� ���� �������� %����� %����� �����4������� �)*�� � ����
��������� ����� ��� ������� ������� ����� �)*��%��� ��%������ ��������� ����� �������
���������.�����%����� /+�����������������������#�
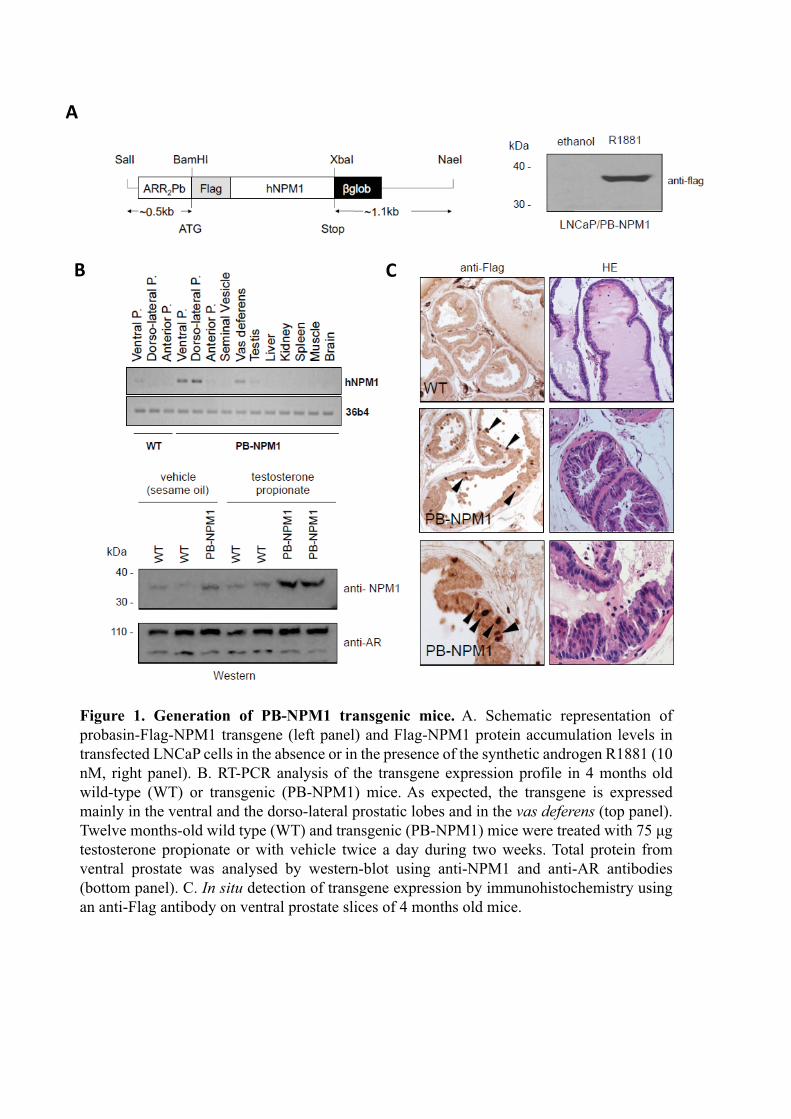
*
+ $
Figure 1. Generation of PB-NPM1 transgenic mice. A. Schematic representation ofprobasin-Flag-NPM1 transgene (left panel) and Flag-NPM1 protein accumulation levels intransfected LNCaP cells in the absence or in the presence of the synthetic androgen R1881 (10nM, right panel). B. RT-PCR analysis of the transgene expression profile in 4 months oldwild-type (WT) or transgenic (PB-NPM1) mice. As expected, the transgene is expressedmainly in the ventral and the dorso-lateral prostatic lobes and in the vas deferens (top panel).Twelve months-old wild type (WT) and transgenic (PB-NPM1) mice were treated with 75 μgtestosterone propionate or with vehicle twice a day during two weeks. Total protein fromventral prostate was analysed by western-blot using anti-NPM1 and anti-AR antibodies(bottom panel). C. In situ detection of transgene expression by immunohistochemistry usingan anti-Flag antibody on ventral prostate slices of 4 months old mice.

6�
�
�����
��������� ��������������������/����"����������$���������,� �����#�������
#$.���#��$����.0�������������$����#������������$���������;�������,� ���#���������.�
��� �$�� #������� �#��$������ (.� ��0���� ��"������� ��� �$�� �������� ��#����"�� 155�;
��(�����A�-��#������A $���������<0��===��A ����� �� �����<�/��������$�����������
�����������0���� ����������� ��� ��� �,����#������� ������ ������ ������� �� �������$� �$��
�.��$�����15���������5�11�<�1����#�����0�15�����"������ ������������������#����������
����(������>���;,� ��A �������������������$���������<�5,1�������������"������
�������� �(������� ����2;����$�� ���� ���������� ��������� �.#����������� ����.;��� (.�
��5���������$������#�������,� ��#�������<�?�#�����������$���������������#����������
�$�� #������� ��� �-;,� �� ����0����$� ��� ����#��� ��#������� ��� �$�� ������� ���� �$�� ����
��������� ��� #�"�����.� �#����� A����$�������� ��� ��<0� �==<�� A �� ��� �� �����<�
�����������.0� ,� �� #������ ��"��� ��� $��$�� ��� �$�� "������ #������� ��� �-;,� �� �����
���#���� ��� !/� ����� A �� ��� ���� �����<� >��$����0� ����.� ��.�������� ���
������������ #�#������� ������� �� ������ �������� ��� ,� �� #������ ��"��� ���.� ��� �$��
#������� �����-;,� ������0� ���������� �$�� �������� �������"��.� ��� �$�� ��������� ��
�<� >�����.0� ��� ��������� �$��� �$�� ��������� ��#������� ��� ��������� ���$��� �$��
�#��$������ ���#������� ��� �$��#������0� ��� �$����(.� �$�� 2��� ����.���� ���2�����$�����
"������#�����������!/������-;,� ��������������������(��.������������������$�������
�#���#��A �����<��
�
��������� �������� ��!"�������������������� ������! ��!� �������
�#�������� ���������������������20���������<;����$���������#$����.#�������.���<�
1�� �$���� ��� ���<� �0� �$�� ����$��� ��� "������ ���� ����;������� #������� ��(��� ����
������������.� ��������� ��� �<;����$� ���� �-;,� �� ����0� ���#���� ��� �$�������;�.#��

6
76
;6
88
9< 99::9=;
;8
;66
�38:89
>��<�
��������
�
=
:
:
9< 99::9=;
>
<
�38:86
;
=
?��
����
������
���
����
����
� ��
�28�886
> ����
���
���>�
��>����
������
��
99::9=;
�� ����*
$
*
+
� ������ ������ � ������ ������ � ������ ������
;8
88
;�
7�
�
;8
88
;�
7�
�
478
;9�
;8
;�
�
��
99::9=;
*�� ����
��
99::9=;
�� ����
��
99::9=;
���
�28�86�28�886
�28�884
��
��
?��� �=;�> � ����� � �� � � �����
99::9=;
Figure 2. NPM1 overexpression induces epithelial cellproliferation and leads to prostatic hyperplasia.A. Ventral (VP), Dorso-lateral (DLP) and Anterior (AP)prostatic lobe weight of wild type and transgenic miceaged 4 or 16 months. Bars represents ± SEM and aStudent’s T test was performed to assess statisticalsignificance (n = 4<x<7). B. HE staining of ventralprostatic lobe from 16 month old WT and PB-NPM1mice. C. Immunohistochemical staining of theproliferative marker ki67 in the ventral lobe of 16 monthold transgenic and WT mice. The ki67 index representsthe ratio between the number of ki67 positive cells and thenumber of total cells. At least 4000 cells were counted foreach condition. C. Proportion of acini displaying at leastone ki-67 postive cell (% of positive acini) and averagenumber of ki-67 positive cells per positive acinus in theventral lobe of 16 months-old WT and PB-NPM1 mice.

3�
�
�����!�������$�� � -��������$�.�����#�������������������*���� �������$�������������
��*�����������������$���������������������+����$�������� �%�����������������
��������������� ��"��������������� "����� ������ ��������������������������� ������
����� �"�� ���� ���� ��������� ������ ��� ��� ���� ���������� ��� ������� �����
�������������������� ���2�� ����� ����������������������������������������
������������������������������ ���������������������� ���������������������������
����� ������������������������������������������������ ����"����� �������"���������
����������� ���������� � ����������� ������������������������������������
����������������������������� ����������������$����������"� ���� ����� ������
�� �������� �������� ���� ����� ��� ��� ���� � �"��� �� ���������� ����� !��"�
������������� �"����� ����#�$���"��� ������� ����� ����� �����������������
�������"������� �������������� ���������%��"� �����"�������������� �������"�������
�� �������"�� ������ ��$� �� � ���� ����� � ������ ������� ��� ���� �"���� %��"�
�������"�� ������� 1������� � ������ ������� ����� ���� #�$�� �"��� ������� ��� ����
������� � ��������� ��� ����� ���������� ���������� ���� ��� ������������� ��� ����
������������
�
��������������� ���������������������� ����&�"������������"������������
#�$���"��� ������������������������������������������������%��������� �����
��������������'���� ()��������������� �*++��,�-�������� �*++"�����"��� ��������"���
�� � ���������� ��� ��������������-�#�$������ ���� ����%������� ������ ����������
������� ��� ��� ������� 2� ������� �� ����� �� ����� ������� ��� ���� ��.&� ����������
������� ����� ������� �"����� � ����� ������� ��� ������� 2�� ������ �������� ���
"����� ������ �����-�#�$������ ������ ��� ����������������1����������������
2�� �'#1� ��"���� ���� ���� ����� ��� ������ ����� ����� ����� /�� ������ � ������� ���
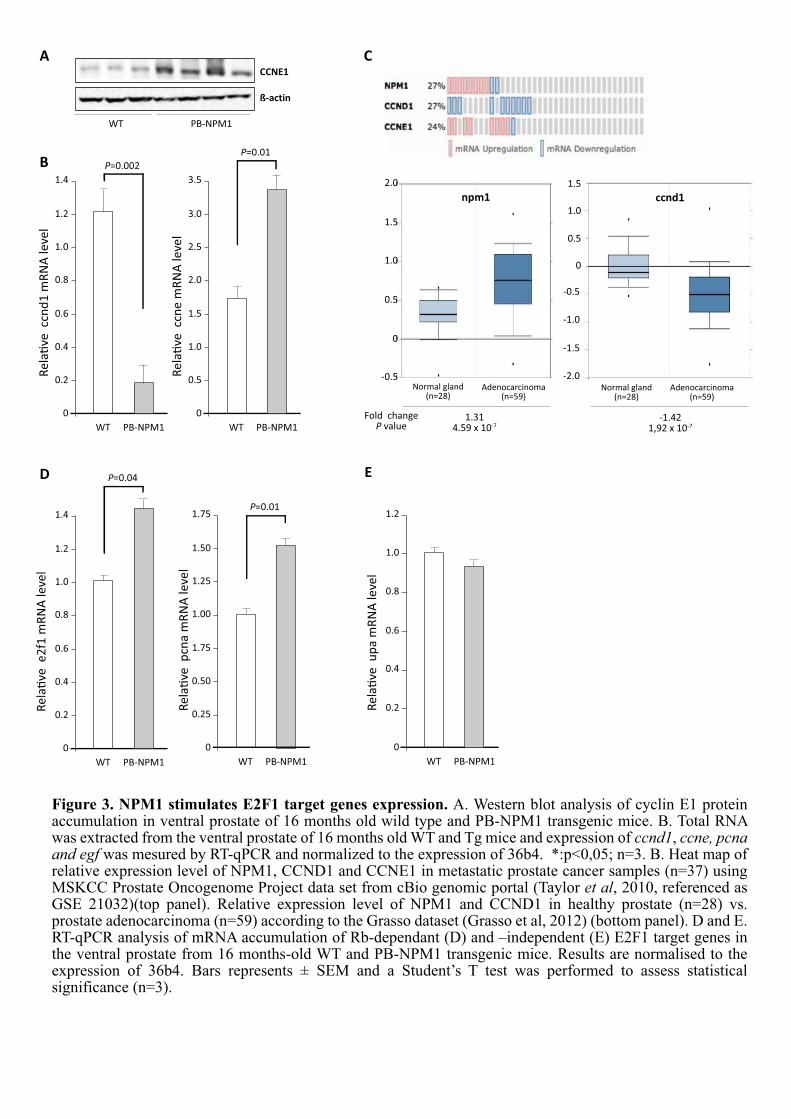
�
8:9
8:3
8:7
�� 99::9=;
8:1
4:8
�38:889
;:9
;:3
2����������
��
� ���
�
8:9
8:3
8:�
�� 99::9=;
8:1
4:8
�38:83
;:9
;:3
��������
��
�����
�
8:6
4:8
;:6
�� 99::9=;
�� 99::9=;
�������
��+��
9:8
7:6
�38:84
1:8
?:6��
����
�����
��
�����
�
8:92
8:68
;:��
�� 99�:9=�
4:88
;:92
�38:84
4:28
;:��
����
�����
��
����
�
8:9
8:3
8:�
�� 99�:9=�
8:1
4:8
�
+
�
* �
;:9
��������
����
��
? ���� �����������
�
�8:�
8:�
4:8
;:�
9:8
�
�8:�
��:8
�4:�
����
���
4��
;��
>��� ����� �<���
;�1;�?�� ; ��<>
�4����@�� ; ��<>
���� ,,!��
�������9����3�7�
?��� �����������
�������9����3�7�
Figure 3. NPM1 stimulates E2F1 target genes expression. A. Western blot analysis of cyclin E1 proteinaccumulation in ventral prostate of 16 months old wild type and PB-NPM1 transgenic mice. B. Total RNAwas extracted from the ventral prostate of 16 months old WT and Tg mice and expression of ccnd1, ccne, pcnaand egf was mesured by RT-qPCR and normalized to the expression of 36b4. *:p<0,05; n=3. B. Heat map ofrelative expression level of NPM1, CCND1 and CCNE1 in metastatic prostate cancer samples (n=37) usingMSKCC Prostate Oncogenome Project data set from cBio genomic portal (Taylor et al, 2010, referenced asGSE 21032)(top panel). Relative expression level of NPM1 and CCND1 in healthy prostate (n=28) vs.prostate adenocarcinoma (n=59) according to the Grasso dataset (Grasso et al, 2012) (bottom panel). D and E.RT-qPCR analysis of mRNA accumulation of Rb-dependant (D) and –independent (E) E2F1 target genes inthe ventral prostate from 16 months-old WT and PB-NPM1 transgenic mice. Results are normalised to theexpression of 36b4. Bars represents ± SEM and a Student’s T test was performed to assess statisticalsignificance (n=3).

0�
�
"��������� ������� ��������������� ���-�������������������������� ��������������
���������� �����1��������������� � ��������������� �������� �� ��2������ � � ���
�������!������� � � ���������!����� �!��"������� ����#��$$%�&��'�� ������������
���� ����� � �� �������� �������� ��� ���� � �� ������ ���!� ��� ������� ���� ��������
������������(����������������$���"(������� ����#��$��&��(������������������������)�
���!� ����� ��� ������� ��� *+� ������,�� ���� � �� ������ �� ����� �� ����� �� ��� ��
� �������������������� �!� ������ �� ������!��� ������ ������ ��� ���������� "���
��&�� ��� �������#� � �� ����� �� ���� � ��� �� ���� �� � ����� � ������������ ���
������� ���� ��������2��� ���������������������������!��� ���"�������� ����#�
�$�$&�� -��������� ��� ����� �� ��� �� �� ����� � ��� ��.�/�� ���0.1�1� ����� ����#�
����!� ��2���� ������� ������� � � ������������������!������#������2�� ��� � � ��� ���
����� �� ��� ���� � �� ��� ���� "/��� � � ���#� �$$34� ��2������ � � ���#� �+++&�� ��� �� � ����
���� �����#������������ ���������������������2���� ��!� �!�������5��������������
��� �����!������������ � �� ��������� ���"������2�#����#�2���&������������������
����� � "���#� ������ ����� ���� ��� ���2����� ���� �������!�� �� � � ��&��
0 ���� �!��#����� ��� ������������ �!��������� ������������� ���6�����������
�-�����������"�����&������������� ����!!�� � �� ������� ����������������� ��2����
�� � � ������������ ����!���������� ����
�
&���������������� ��������������� ���������������1���������� ������� ���(�7-�
������ ���� ��� ��� ����!��� ������ ����80���32������� ������ ������#��������������
�������8���� � � ����9���������� �������� ���� ��������������!��� �!�������2�#�������
������� ��� �� �� � � ��� ��8��� ����� ��������� ����������� ��� ��32���� �� ��� ����)3#�
����!� �� � �� ���5�� �� ��� ��� ��� 21����5�� �� �������� ��!���� -��-2��� �� � ��
�����5�� � ��� �������� �!��� ��� "���� � � ���#� �$$)&��'�� ������� ��32���� ��� ���
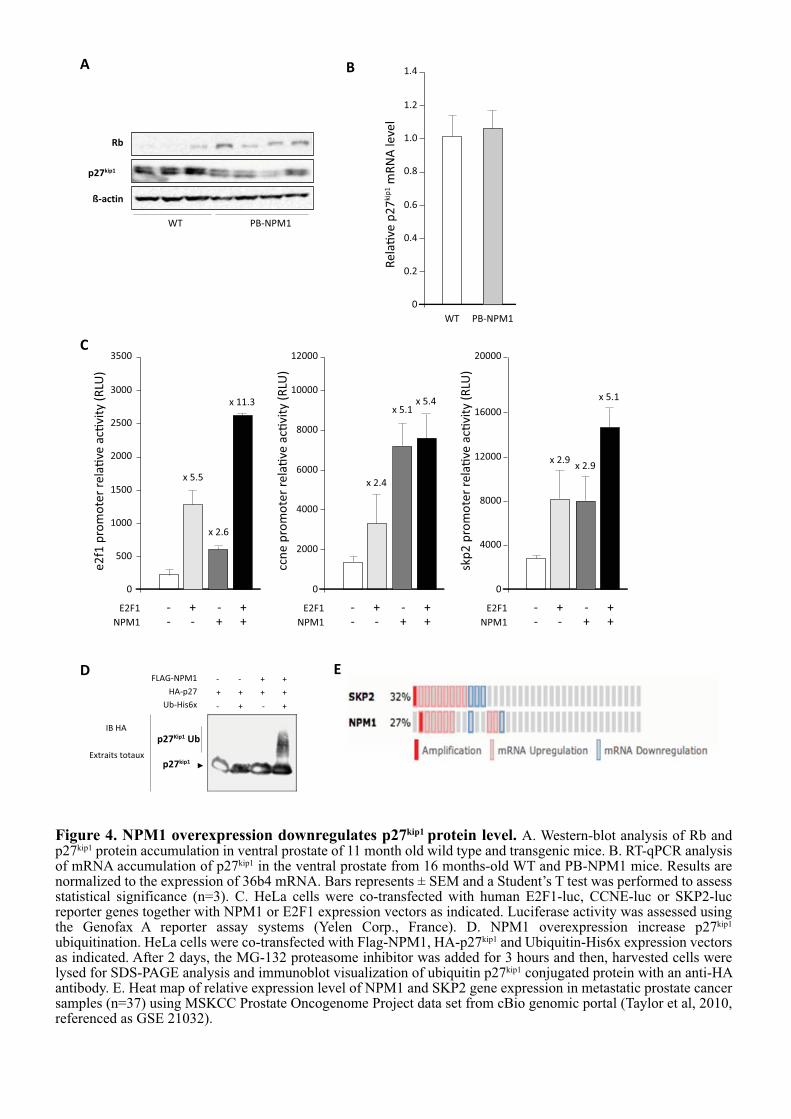
�������
%������
�������
)�
��
@�������������
��������
� �
��5���� ������
9:9=�
�
��9
��3
���
�� 9:9=�
��1
4��
;�9
;�3
: �������?
�����
������
�
�
* �
+
: : : :< : < :
< < : :
���%��� ��
�
=��
;���
���4
; �?�
; �?4
; 44?�
� ���
� ���� ��3
� ��� � ���
� ���
< : < :���� < < : :
����
7���
7���
?���
?���
���
��
����
��������������
�<7�
�
�
����
���� < : < :���� < < : :
;���
8���
;���
�����
�����
���
��
���
�����
��
�����
�<7�
�
�
;���
���� < : < :���� < < : :
;���
�����
�����
�����
���
��
���
����
��
����
�<7�
�
Figure 4. NPM1 overexpression downregulates p27kip1 protein level. A. Western-blot analysis of Rb andp27kip1 protein accumulation in ventral prostate of 11 month old wild type and transgenic mice. B. RT-qPCR analysisof mRNA accumulation of p27kip1 in the ventral prostate from 16 months-old WT and PB-NPM1 mice. Results arenormalized to the expression of 36b4 mRNA. Bars represents ± SEM and a Student’s T test was performed to assessstatistical significance (n=3). C. HeLa cells were co-transfected with human E2F1-luc, CCNE-luc or SKP2-lucreporter genes together with NPM1 or E2F1 expression vectors as indicated. Luciferase activity was assessed usingthe Genofax A reporter assay systems (Yelen Corp., France). D. NPM1 overexpression increase p27kip1
ubiquitination. HeLa cells were co-transfected with Flag-NPM1, HA-p27kip1 and Ubiquitin-His6x expression vectorsas indicated. After 2 days, the MG-132 proteasome inhibitor was added for 3 hours and then, harvested cells werelysed for SDS-PAGE analysis and immunoblot visualization of ubiquitin p27kip1 conjugated protein with an anti-HAantibody. E. Heat map of relative expression level of NPM1 and SKP2 gene expression in metastatic prostate cancersamples (n=37) using MSKCC Prostate Oncogenome Project data set from cBio genomic portal (Taylor et al, 2010,referenced as GSE 21032).

5�
�
����%�����12��2�������������� ������������������������������������������������������
���������������������������������������������������� ��!����������"����������#$1�������
���� ���� � ������ �������� ��������� ���� �-$�%�� ����� ���� �� �� & ��� ���� ����
�'����������������������� ���'����������'������������� � ���������������������� �����"�����
�����'��� �(����� � ���$�%����'���� 2�)�� ��������������� ����'��� ���� � ����*�� � ���
� ������� ������'������������ ������ ���$�%�������(��������������������������+�,��
������ ������ ��� � �� �'��� ����������� ��� � �� ���������������� ������������� ��� 2�)�"� �������� ���
-.��������������2������(����������1��� �����������/��"�������������������+�,����������� �
�'������������������������'������ �������������2�)���������������������������2�)�"�-.���
���� ������� 2 � ���������� � ��� ���� � ���� ��������������� ������������ ���������� ���$�%���
& ���� ����� � '�� ������ ���� �'�� ���� ����� � ��� $�%�� �����(������������� �� �����
�������������������������������� ���������2�����-.����������������2�)������� �����'����
�����������������������'������(������� �� ���$�%�������(����������� ��������������
'��0'�������������������&�������"���� ��������������������+1����������������(���������
������� ����� � �� +�,�� ����� ����� ��� ������������ ��� � ����$�%�� ���� +!-1'��0'�����
�(�����������������������������������'���/����������������������������"���������������������
�'�����'�+��� �'����� ����+!-1'��0'�������2'������ �������������� �'������� ����� � ��
������ ������� �'���������� ��� $&1� � ���������� ��� -�-�132� ����������� ��� ��'����
����������������'��������������������'�������������������������� ��+1�����������������
� �����2'����������'��0'���������������������� ���$�%����������(���������)���/� ��& '�"�
� ������'�������������'�� ���� ������������������ ���� �����'��0'������������������������
�����������(���������$�%���
� �

�

4�
�
����������
����� ��������� � ����������������������������� ���������� ���� ������� ������� � �
�������� ���� ����� �������� � ������� �� � �� ���� ����� �������� �� ��� ���� � �
���������������������������� � ��� ������� � � �������� �������������������
����� �� ���� !"� ����#���$%��&�� '����� ����������� � ������ ��� ������� ��������
���������� � � �(��� ����)��������� ��� ����������� �� � � �������� ���������*�� � ��
�� ������ ���� �� �� � �� �� ������ ��� �� ���� � � �������� ����������� ���������� � � ��
� ����� ���������������������������� ����!"����������$%�$+�"�������������$%�,+�
-� �� �� �����$%%.+� '�� ������ �� �����$%%/&��0����� ������������� ��� �� �����������
������� ��� � ��� ��� ���� ��� ����� ��� ������ 1�� � ��� ������ ����� �����
���������� � ����� ��� ���������� ����������� ���� ��� ��� ������� �� ��� �������
����� �� �����'� ������� ����� ����� ������ ��� ����������������������������������
�� ��� ���� �� ��� ��� ��� ������ ���������� ���������� � �������� �� ��� ��������� �
� �������������������������2 ��� ���������� ����������������������� ���������� ��
����� ����� � ��� ������ �� �-)����� ���� ���� ���� �������� �� �� �������� ��������
����� ������ �!�� ��)()&����������������������������������� ���!3� ���������$%%4&��
������ ���� ������� ���� ����� ��������� ����� �-)����(�� ��)()� ���� �� ���� ��� ��56�
������������ � ���� ��������� ��� ����� ��)()� ����������!����� ������ &��'� ��
�� ��)()������������������������� ���� ��� ��� ����� ���� ����� ���������� ���
�� ����� ���� ��� �������� � � �� � ����� � ���������� ��� ���� � �������� �� ��� �����
!"�������������$%�,&��������������������� ���� ������������������ ������������
� � ������� �������������������� �������� �� ���� 1��������� ����� ������ � �� ��������
������ ����� ���� ����������� ���� ���� ������� �� ���� ��� ����� � � �������� �� ���
��������� ��

�$�
�
�������������������� ��� ������ ���� ��� ������� ��� �������� ������ �� �� ��������
������������������������ ������������ �� ���������������� ���������� �!�����"�����
��� � ���������� �� �� �������#�������"��� ������ ����������#�� �� ���$#��
������ ���� ���$#%�������������� �����������������������#������� ����&������ � �
����� #������ �� ���� �������'�������� ����� ���� '���� �������� ��������'���
���� ��(������$#� ��� ������� ���� ����#�� ���������� ��� ����#� ��� ������ '���������
)*������ ���++,-�����"������� ���.../ ������" ����"����������������� �������������
����� �� ��$#��������������"����� � ����������� ���-%�����"������ �������'������
����#� ���� ��� ������� '����$#�)������ ������ ���++0#/ �!����� ����� ��' ���#��
���������� �� "#��������� ����&������ �� ��� ��������������12������ �� ����������
'������������������������� ��� �������� ���������������� �"�������"������������
����� ����������#���� �����$#��12�� "� ���"��� ������$1���" ����)���������
���� ���..,-�3� ����� ���++4/ �
����� ����&������ ���������������������'�������������5���#����������� ����&������ ��
���� ������� 6�� � '������� �� ��� ����� ���� ��� ���� ��� �$1��� " ���� �� �� ����
�������)������ ������ ���++0�/ ������ #������ ��"������������(��������������� �����
������ ����&��������� ������ �#��(��� � ����� ������ � � !������� ������� 6�� � '�%
������ �� ����� �� ������ �� ������ ���� ��� ���������� � �� 6�1� �������� �� )���"�%
7 #������������������...-������ ����� ���..0/ �8��"����������%������������������
���� ����� ����&������ �� �� " �� ��,����� �� ��� ����� ��� #�(����� �� ����
�� ��� "��� �������� � � 9�� ����� #������� �� �� ��� ��������� �� ��,����� �� ����
�#������� ��� � �:��� ��'��� �� �� ��� ��������� �� # �� ��������� �� ���� ����"$�1�
������ ������ #������ ���������������������� "������������� ��� �����'�������������� ���
#���� �� '�� ��� ��,����� � ��� ��� ��� ������ '��� �� ���� ������� ����������������
"�������� ���� ���"��� ������� )��������� �� �� �� �..;-� <���� �� �� �� �..;/ � ������ ����

33�
�
5673�#$��������������� ���������������������� ������������������������������������
������������������������������������������������������� � ���!�����������
������������������������������������ ���������������������������������� �������"���
������� � � ���� ����������� ��������� "����� �#����������� ���� ������ � � ���� � ���
#���� ��������� ��������� � ��� ���� ��� ��� ��� ��������� ������ ���� ��������� ����
������������ ������������������� ��
� �

�

$%�
�
����������
1&&'%���������� ���� ������ ������� ����� ��������� ���� ��� ������������� ��� ��������� ���
����������������������������������������������������� ����������-���������!"�#��
-������ ��1��� $������ ��%��� 2����&������� '�(��� ���� )����� 2�1�� ��#���� (�*��� ����������
����������������&���������������������������+�����������!��"!�,��
���� -�(��� '������� $��� ���� .������������ /�(�� 0,,#��� 1�� ���� ���&����� �0�� ��� �+���
������2������������������������������3���������������������������)������3������������
04!"05���
%������� -���(�������� ���� 1������� 2��� 1��������(��� �������� ���� 6��7��������� $��� $�� .���8��� ����
��3������ ���� ����+&��� 2��� .��������� 1��� ��� ���� 0,,4��� ��������+��� ���������+��� ���
������+����������������+������������+��������������)��2�����/��(���������049"055��
%�������/���.����'�:����-�����%���%��+��������(���������������6���������(�� 0,�,���2���+�����
�������������&���������������� ���0,,#2�;$ - �1)�0,,#�� -���� /��������������0#�!"
0�����
%���+�:��&�������/�������(�������<�� ��������������������&�����������������������������
&����������6�)1��������0��2=�����������������!!#"!9���
;��������/�����-����������/�����������(�>���)�+��'�.���%���������(�/�������;����&�����)�(��
������� 1�������:������������ 6�������� ������� 6����������� ��� ��� 1�1(6�(������
����������������95#�"95����
;���������.���>���<�:(�����&�����������������������������������.�(��������1�6���?������(�/���
/���������$����������/��� -�������� /������������ 0,�0���1��+���������� ������������� ������
����������:���������������������������)����������0!�"09!��
;���������.���(������������%�������-�������6���������6�6�� 0,,5���)��������+���������������
)������3������������9�!"4,4��

YZ�
�
/(�������1���������� ���� ����� ���YZ����������� �� � � ����������������� ������������ � ���
-����������Y����Z��
������� ������� ��� ������������������������������� �������������! ���"���#������ �� ����$���
- ���� ������! %���&� ��#������ ������� ����������[���'�(�� �� ��(����� ��������)-�Z�
��� �*1� +���� � ��� ���������������� ����,��%� �(� �� � ���� �� � � ����� ��� ������� �
���� ��� �������� � ������[-[��[.���
������/��� ���� ���YY��� *0�Y)-�Z1� 1� �����(���������� ���� ��� � ��� 2�+���� �
-�� � �����������������2 �� ��� ��-���� ���2 ���'����������Y\-��\��
�����3��������4������ ���56�������4������������7���7�� �������� ������� �������$�����8���7���
��� 9 �� 7�� �������� ���%������� �(� ���� ��������)-�Z� � ���� �� ���� ��+� �������
������:�������� ����� �� ��������( �������������,�,����0�����*�����1�������������\.�\�
\.[;��
�����9���4��� ��$���4��� ��3�5$���<�����5���9�� ������#����=������4��� ��*�����Y����2>�� ������
�(����� ��������)*0�Y����� ��� ��8������ ������������,���, � ����(����������� ��
� ��������-��� ����������-Z��
��� � �����1������4�((�������*������\���-����� ���������� ����(������� ��� ����������
������� ����� ������� �������Z�-[�Z�.���
���+ ����#���-������2�����?���� ����������5��� �������- �����������! �� �� ��0��������� ���
��� ���Y;���*0�Y���� ���� �� �� �� ������� ��8�������107��� ������ � ���������� �
���� ��� �����0����� ���� \.�\Z��
�������� ��1��� =����� 6����� -�������� 2����� ��� #� �+ � �� *���� ����;���� ��(( � ������
>�� �������(�� ����%�� �� ������%���� ��� ����� ,� �� �(�����@�%������8����A����� �
��� � �������(�������� ����� ���B� �0������ ����ZZ-�Z;;��
����������1������ ������-�6���-��������2��������#� �+ � ��*��������;+��������������
� � ������(�2+����� ��2���%��� �0������ ����� ������� ��2 ������.�Y[�.��-��

)*�
�
�+7,��7+-./�1C/�$.+0�/�$C/�21,�7/�2C/���00�7+/�1C�C/��0��,,�/�#C$C/�=�0243+/�1C/��7��0���7+/��C�
8)@@@9C�� .�4.,.7�,.+7� +%� 56F� .2� 0��4-�,��� 1� ��3D��5�7��7,� 54+254+01-�,.+7� �7��
,0.��0.���+�5-�7�%+0��,.+7C�#�7�2���8C���/�))E)9))E@C�
�+,,�,/�*C/�-�--�47,/��C/�-+--�/��C/��+7.�4/�C/���2+7/��C/���,8��8/��C/��4�.�/�=CD0C/���7���0�
7:�2,/�BC/�7.���-/�BC/������������������ ������21�������������������������������������
�������������� ��� �� ������� ����������� ���� ����������!���������� ��������� ������� 2���
���������"F�#"$%�
&����� 21� �''%��������������(����!������������������)������� �����*���� ����� -�������
�$F#�'%�
&�+�,���-��.���-��+����&��.�������/�����1������0�����1&������-��������� �''2��
23��������� ��� �������4�����5-�%� ��� ������� ���� ����������� ������������������ 6�
���4�������7$#"��
*)�,�)��� /� ���$�� �4�� 8��������� ���� ��������� ��� &�/�5&������4����5-�%�� ��
/���������������&���������1��������������6�-���4��� ��)(��������77�#77$�
��������/���4��������� 1/������� 81�� ���� ���������9� �''7�� �(����� ��!���������
��4�:����������������������������� ���&1��(��4��������4�������:��:������9������� ���
�2�F#�2%'�
;�����-��-��������9��<�0����-�������4��������<� ���$��=�������&���� ������������������
���������������������������������������������������6�&������8�������%�#�%F�
8���4����������/=��>����>��9����:�����&/������8�������� �/� ���2�� 2��!���������
/������ *����� ?��4� 8������ ��(� ���� 8���������(� ���� &���� ��� �� �������� ��� ���������
�����������������������1��������=��������1�������������=�����2�''#2��'�
84�����(���6��8,������4����.��9������88���4���4�����=��@����������98������@�������@�
���'��1���(������&�/��<�����+�������4��&���������������=�������������������������

�4�
�
1�&���������������� ��������������������������������&����&����&�-���&����������
����&�
����������������� �������������� ��!�""�#&�1����$��� ��������� ���� �� ���� ������
�����&��1�1�������%&��''&�
�������(&�&������������&�����������&��(�)���� �&�&���������������-&2&�!�**�#&��������27
��+�������$���������� ��+� �&������&������','��'�&&�
��������&-&����-�7.��������&��/�������&�&�� ����.&�&��+������� &1&��� ���������&1&������
.������� �&�&� !�**�#&� ���������� ��� ������ � ��� ��� �������� ������ ����$�� )��
����� �����&��&����&�������&������$&�����*&�%"'&�
��)��$��2&�&��������&�&��2 ����� �&�&��-��$$����� �&0&��������1&+&������1�������1& &�!�***#&�
����������� ����)���� ��� ������ ������ ������� �����2� ������ !1(� '7��,#�
���$��3������ �����4-�%&�.��1�����������*&�%"'&�
.�5�������&�� ������+&����������&�� 15�������.&��6�5�7���8&�� ���� /������� �&� !�***#&�
��������� ������ -�%&�� )���� ��� �����)������� ������ ���� ���$���������
����������1� ��������� �����������&��&�-�����&�!.�5��#������*"'�*"*&�
.���5����&�����5���8&��+������&����$�������.&������.�������&�!�**�#&���� �����������
2 �������)�������������� ������������2 ������$�����������&�������
(&�����%%���%%��&�
.������� -&�&�� ������3���&��8���������8&�� �� ������ 1&��9�����/&�� ������� -&�&�� 1������0&+&��
+����5��1&���������2&��(����-&������&�!�"�"#&����$������$������ �������$����������
�����������&��������������������&�
.��������&��+� ������&(&���-�����&��������71���3�����&��:)�����75����&��-�������������
�&�����3�������&�&��+���3���&8&����������$��������&�&�!�**&#&�������������7� �����
5���� ����)����� ��+� �� �� �� ����� ��$������ ������� ��� ������3�� ������ ������
�������������&�������(&������'���'&&�

;<�
�
04�'454=����������-���� � ������������- �� �����������*�������� � ����� ����� �� ��
� �� !"��������� � !"������������� ��# "������! �������� ���� ���$���� ������%�%&%'(��
)��!�� *���+���� ,��� - ���.�����/ �!"��������$��������� ���� ,����� ,�����������+����� -���+������0
-"������$���� ������$�*��� ������ �*������$������ 0�# � ��� � ������ � �� ��"��� �$� ��
�"�����"##� �����! � �� �������� ��������#������ ��� ������ ��� ������*�(&**%��
1��!����2�����������,���2"�#����2���)��!��3�,���$�����#�����,���� � �������,�-���-�����2�������
� �� ��� ��2�� �%((4��� -��� #*5� 6#� ������ #� ����� #���� ��� �� 0 � � �"������� ���
#��� ���������#������ ��� ���,��7����������('%&('8��
����!��,��������������������# ���*�������2��"������,���*������1���������#���� �#��9�����
#����� ���� �����!��� � ���� �#������ 0�# � ��! � � 6#� ����������"!��� !"�������
9�������! �������!�"��������������������������������2���������!�������':(4&'5%���
���"������;� � �0��������1������� ���������)��!��)���+����������)������0-"������$�������
��������1�1���*��:���*�� �!��� �#8�������9�� �� ��������������������2�"� �2�� ��
���2 ��������$������ ���� ������ ��� �������544(&54(4��
�
� �

�

� $�����������%�� ���'�
TO�
�
&������?�
�
��� �������� ���� ,�������� �������� ������������ �������=��� ����������� ������ �>�
��������j���{��� �!"��#���"j�$$"{�-���!!"��j� �-�$"`{�-���$$"��"���j �!"�j{���%$$"��j&"�j{�
-'���(�-)�|"��jk{��"�!���j$����j{���$|*�"�����!{��j�"!`����"$��!��-$�j�"��"�j���!}�
�
��"#������� ��$� �-�
�
��+��`'�!"���!� "�, ���" `��,+$�-j'"���! �$��(��� �!("�("$$j$���"�"!��'#j$�!`�$w�(`�|�`'��"�
(�&$" �(�`�+$� ,�-j" � �,+��`�!`" ���! � $"�(�!`�.$"��"� $�� �!`)* "�+��`'�-j"��j�"!(��"��j�
,'`�&�$� ,"� #$j(���-j"� "`� $�+���-j"}� �$j � �'(",,"!`{� �" � '`j�" � �!`� ��++��`'� j!� �.$"�
!j($'���"��"�, �����! �$"�(�!`�.$"��"�$w"k+�" ��!�#'!�-j"}�'"�+��� " �,j$`�+$" ���!(`��! �
`��+)�-j" {� $w)�+"��(`�|�`��!� �"� , ��� (�!�*�"� j!� �|�!`�#"� '$"(`��� �jk� ("$$j$" � "!�
+��$��'��`��!{� "`� "$$"� " `� ��'-j",,"!`� �& "�|'"� ��! � $" � `j,"j� � )j,��!" {� +��,��
$" -j"$$" ��!� �"`��j|"� $"� (�!("���"� $��+�� `�`"}� '�! � ("``"�'`j�"{�!�j �,�!`��! �#�/("�0�
$wj`�$� �`��!��"���&��&$� `" �",&���!!���" ��"� �j�� ��!|�$��' �+�j��� .�-j"�, ��� `�,j$"�
$w"k+�" ��!� �"� ��~}� �� +"�`"� �"� � .� �!�j�`� j!"� )�+"��(`�|�`��!� �"� , ��� "`�
$w�((j,j$�`��!��"�$w4�,���! ��-j"��"�$��+��`'�!"���~}�-"``"�"��"`�" `� +'(���-j"��"�, ���
+j� -j"�j!� `���`","!`�0� $����+�,�(�!"��j� $�� `��! �"(`��!��w4�� �!`"��'�"!` �����#' �(�!`�"�
, �����! ��" �("$$j$" � �j|�#" ��j� �!|�$��'" �+�j��� .��'�j�`����`","!`� $w�((j,j$�`��!�
�" �4�,���! ��-j"��" �+��`'�!" ���~}�"�(�!`�.$"��"� $w"k+�" ��!��"���~�+���, ���
w"k"�("�0��"jk�!�|"�jk}�'wj!"�+��`{�!� �"k+'��"!(" ��w�,,j!�+�'(�+�`�`��!��"�()��,�`�!"�
"`��"� `��! �"(`��!� `��! �`���"��|"(��" �#*!" � ��++��`"j� ��"� `�+"� $j(��'�� "� �!��-j"!`�-j"�
, ��� "���k"� j��$"�+��,�`"j���"���~�"`�(�!`�.$"�$w�(`�|�`'��"�("��"�!�"�}�'w�j`�"�+��`{�$��
�' `�&�$� �`��!���+��"��"� $w4�,��"���~��+�* � �!)�&�`��!��"� $�� �#!�$� �`��!�, ���+���j!�
`���`","!`�0� $�� ��+�,�(�!"� � j##*�"� j!�,'(�!� ,"��"��'#����`��!��(`�|"}� .!�"��"`{�!�j �
�|�! � �& "�|'� #�/("� j!� `���`","!`� �"� !� � ("$$j$" � �|"(� j!� �!)�&�`"j�� �"� $�� `��! (��+`��!�
��(`�!�,�(�!"�'��-j"� $w4�,��"���~�+� *�"�j!"��",��|�"� $�!#j"}�'"�+$j {� $w"��"`��"� $��
��+�,�(�!"�" `��"!|"� '�+���j!� `���`","!`�(�!1��!`�0� $w�(`�!�,�(�!"�'�("�-j�� j##*�"�-j"�
, ��� �!)�&"� $w"k+�" ��!� �wj!� #*!"� ��!`� $"� +���j�`� " `� �,+$�-j'� ��! � $�� �'#����`��!� �"�
$w4�,��"� ��~}� .!� j`�$� �!`� �" � �j�� � �!|�$��'" �+�j�� $"� j++�" "j�� �"� `j,"j��� .�

�

� $�����������%�� ���'�
TG�
�
+'(���-j","!`� ��! � $w'+�`)'$�j,� +�� `�`�-j"{� !�j � �|�! � (�!���,'� $"� (�!`�.$"� �"�
$w"k+�" ��!��"� ��~�+���, ��� ��� *�*����! � j!� (�!`"k`"�!'�+$� �-j"}� .!��!{�!�j ��|�! �
,�!`�'�+��� `��! �"(`��!��w4�� �!`"��'�"!` �-j"� $w"k+�" ��!��"� ��~�" `�!'(" ���"� 0� $��
+��$��'��`��!��" �("$$j$" �� .���}�'�! �$w"! ",&$"{�(" ��' j$`�` �,�!`�"!`�-j"�, ����'#j$"�
$w"k+�" ��!� �"� ��~� +��� �" � ,'(�!� ," � `��! (��+`��!!"$ � "`� +� `�`��! (��+`��!!"$ {� "`�
+$�("!`�("``"��"�!�*�"�(�,,"�j!�"��"(`"j��"!��|�$��"�$�� �#!�$� �`��!������4� �, ���+�j��
(�!`�.$"��$��+��$��'��`��!�("$$j$���"}�
�
�
� �

�

1
Transcriptional and posttranscriptional regulation of NPM1 gene expression by mTOR. 1
2
Short title: mTOR regulates NPM1 gene expression. 3
4
5
Rafik Boudra1,2,3, Rosyne Lagrafeuille1,2,3,†, Corinne Lours-Calet1,2,3, Cyrille de 6
Joussineau1,2,3, Gaëlle Loubeau1,2,3, Cédric Chaveroux4, Jean-Paul Saru1,2,3, Silvère Baron1,2,3, 7
Laurent Morel1,2,3 and Claude Beaudoin1,2,3* 8
9
1Université Clermont Auvergne, Université Blaise Pascal, GReD, BP 10448, F-63000 10
Clermont-Ferrand, France; 11
2CNRS, UMR6293, GReD, F-63001 Clermont-Ferrand, France; 12
3Inserm, UMR1103, GReD, F-63001 Clermont-Ferrand, France; 13
4Inserm U1052, CNRS UMR5286, Centre de Recherche en Cancérologie de Lyon, F-69000 14
Lyon, France; 15
†Current affiliation: CNRS, UMR 6023, LMGE, BP 10448, F-63000 Clermont-Ferrand, 16
France. 17
18
*Correspondance to: Claude Beaudoin; CNRS UMR6293-GReD, Campus Universitaire des 19
Cézeaux, 10 Avenue Blaise Pascal, Aubière 63178, France. Tel.: (+33)473405340; Fax.: 20
(+33)473407042; Email: [email protected] 21
22
Keywords: mTOR, NPM1, gene expression, cell proliferation, cancer. 23
Abbreviations: mTOR, mammalian target of rapamycin; NPM1/B23, nucleophosmin; MEF, 24
murine embryo fibroblast; siRNA, small interfering RNA. 25

�

2
Abstract 1
The mammalian target of rapamycin (mTOR) plays essential roles in the regulation of a wide 2
array of growth-related processes such as protein synthesis, cell sizing and metabolism in both 3
normal and pathological growing conditions. These functions of mTOR are thought to be 4
largely a consequence of its cytoplasmic activity in regulating translation rate, but 5
accumulating data now highlight supplementary role(s) for this serine/threonine kinase within 6
the nucleus. Indeed, the nuclear activities of mTOR are currently associated with the control 7
of protein biosynthetic capacity through its ability to regulate the expression of gene products 8
involved in the control of ribosomal biogenesis and proliferation. Using primary murine 9
embryo fibroblasts (MEFs), we observed that cells with overactive mTOR signaling displayed 10
higher abundance for the growth-associated NPM1 protein, in what represents a novel 11
mechanism of NPM1 gene regulation. We show that NPM1 gene expression is dependent on 12
mTOR as demonstrated by treatment of wild-type and Pten inactivated MEFs cultured with 13
rapamycin or by transient transfections of small interfering RNA directed against mTOR. In 14
accordance, the mTOR kinase localizes to the NPM1 promoter gene in vivo and it enhances 15
the activity of a NPM1-luciferase reporter gene providing an opportunity for direct control. 16
Interestingly, rapamycin did not dislodge mTOR from the NPM1 promoter but rather strongly 17
destabilized the NPM1 transcript through a posttranscriptional mechanism of mRNA decay. 18
Finally, we also show that NPM1 expression is required to promote mTOR-dependent cell 19
proliferation. Moreover, NPM1 mRNA was found up-regulated and sensitive to rapamycin in 20
a prostate-specific Pten-deleted mouse model of cancer. We therefore proposed a model 21
whereby mTOR is closely involved in the transcriptional and posttranscriptional regulation of 22
NPM1 gene expression with important implications for our understanding of how this 23
pathway operates to regulate proliferation in development and diseases including cancer. 24
25

3
Introduction 1
The mammalian target of rapamycin (mTOR) is a highly conserved serine/threonine protein 2
kinase that regulates cell growth and metabolism in response to nutrient availability, growth 3
factor and cellular stresses.1, 2 It belongs to the family of phosphatidylinositol 3-kinase 4
(PI3K)-related kinases (PIKKs) and operates in two functionally distinct cellular complexes 5
termed mTOR complex 1 (mTORC1) and 2 (mTORC2) which associate respectively the 6
regulatory-associated protein of mTOR (raptor) and the rapamycin-insensitive companion of 7
mTOR (rictor).3, 4 Once activated, mTORC1 promotes cell growth through enhanced 8
translation resulting from activation of the ribosomal S6 subunit kinase (S6K) and 9
inactivation of the eukaryotic translation initiation factor 4E (eIF4E)-binding protein (4E-10
BP1).5 This growth-promoting activity which is further strengthened by mTORC2-dependent 11
functions is now described too for its ability to activate a transcriptional program that is 12
critical for protein synthesis and cell growth. Therefore, by binding to the promoter of RNA 13
polymerase I and III transcribed genes, mTOR increases the abundance of ribosomal RNAs 14
(rRNA) and transfer RNAs (tRNA) leading to enhanced protein biosynthetic capacity and 15
increased cell growth.6 16
More recent studies have identified additional gene regulatory networks downstream of 17
mTORC1 complex that might activate specific cellular processes that are likely to contribute 18
to human physiology and disease. Using a genome-wide scale analysis, Chaveroux et al.7 19
have demonstrated that mTOR occupies many regulatory region on chromatin obtained from 20
mouse livers with a strong enrichment at the proximal promoter of the NPM1 gene. This latter 21
encodes the abundant and ubiquitously expressed nucleophosmin protein (NPM1 also known 22
as B23, numatrin and NO38) which has been originally identified as a non-ribosomal 23
nucleolar phosphoprotein found at high levels in the granular regions of the nucleus.8 Its 24
expression is found to be more abundantly expressed in proliferating cells than in resting cells 25

4
and is increased markedly and promptly in association with mitogens9, 10 or certain cellular 1
stresses.11-13 Like mTOR, NPM1 expression is altered in several different types of cancer and 2
is demonstrated to be a positive regulator of ribosome biogenesis, cell proliferation and 3
resistance to cell death. 14 To date, the regulatory mechanisms involved in the regulation of 4
the NPM1 gene expression are not understood in detail but experimental evidence have 5
demonstrated that transcription factors such as YY1 and Myc can both bind directly to the 5’ 6
region of the NPM1 gene. 15, 16 In addition to the activity of the mTOR kinase to control 7
mitochondrial oxidative activities and metabolism, 17, 18 its recruitment to the NPM1 promoter 8
region prompted us to address whether mTOR targets the NPM1 gene to modulate its 9
expression at a transcriptional level. Besides, it is currently unknown to which extent these 10
two key regulators of cell proliferation might interact to regulate common biological functions 11
with implications in diseases and cancers. 12
In this study, we demonstrate that the mRNA and the protein levels of NPM1 are elevated in 13
fibroblasts with hyperactive mTOR signaling in comparison with wild-type fibroblasts. 14
Furthermore, we show that rapamycin inhibition and RNA silencing of mTOR reduce NPM1 15
expression indicating that a mTOR-dependent transcriptional complex might be involved in 16
this control. By using a chromatin immunoprecipitation (ChIP) assay in conjunction with a 17
reporter luciferase assay, our data indicate that mTOR binds the NPM1 target gene at the 18
promoter to regulate its expression. Finally, the finding that NPM1 levels were increased in 19
prostate tumors of mutant mice with genetic inactivation of Pten raised the possibility that 20
hyperactive mTOR directly leads to elevated levels of NPM1 perhaps due to an increased 21
requirement of ribosomal synthesis in cancer cells. Therefore, our data may help to clarify the 22
mechanisms underlying the transcriptional regulation of the NPM1 gene and may shed light 23
on the ability of mTOR to control NPM1 functions in particular regarding the effect of mTOR 24
in increasing ribosomal biosynthesis during proliferation and neoplastic transformation. 25
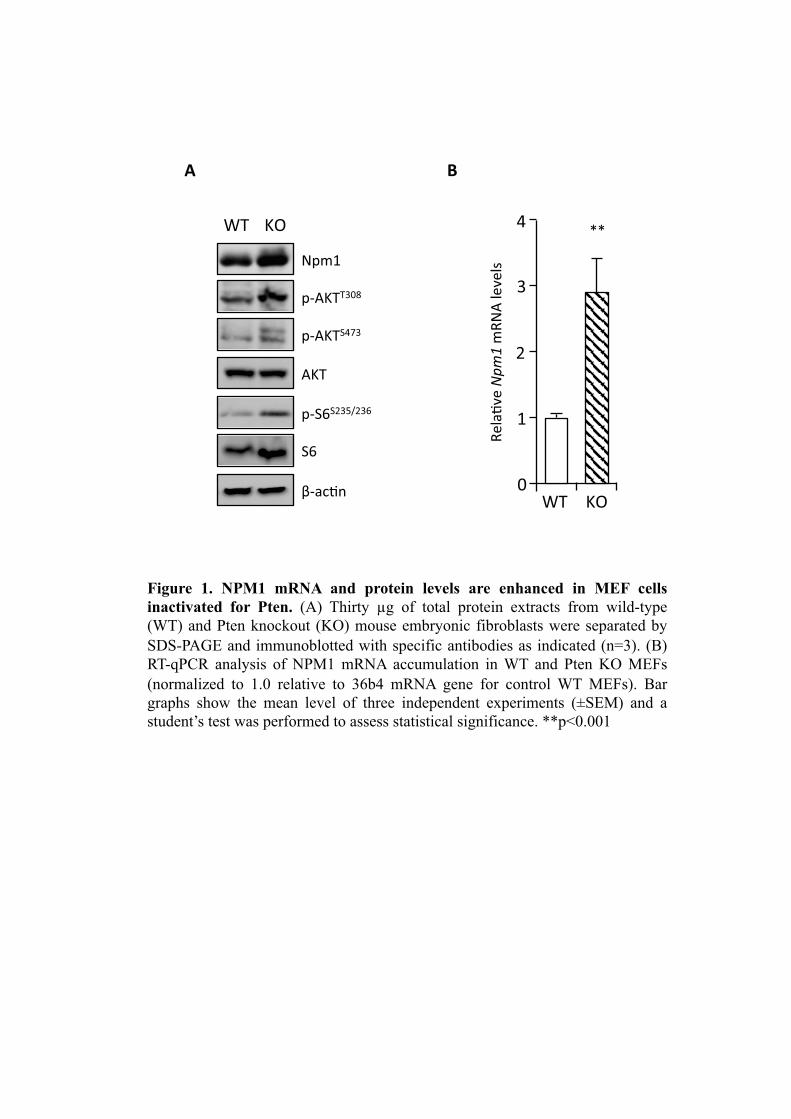
�24�������
�4������
4���
���� ��� ���
���
�����
������
��� ���
&� ��
�
��
��
��
�
���
��� �������������
������
4��������
Figure 1. NPM1 mRNA and protein levels are enhanced in MEF cells inactivated for Pten. (A) Thirty µg of total protein extracts from wild-type (WT) and Pten knockout (KO) mouse embryonic fibroblasts were separated by SDS-PAGE and immunoblotted with specific antibodies as indicated (n=3). (B) RT-qPCR analysis of NPM1 mRNA accumulation in WT and Pten KO MEFs (normalized to 1.0 relative to 36b4 mRNA gene for control WT MEFs). Bar graphs show the mean level of three independent experiments (±SEM) and a student’s test was performed to assess statistical significance. **p<0.001

5
Results 1
Pten-null fibroblasts display higher expression levels of NPM1. 2
Despite the multitude of NPM1 functions in a plethora of biological processes, the molecular 3
mechanisms of NPM1 gene activation are still poorly understood. Recent data emerging from 4
genome-wide chromatin immunoprecipitation analyses indicate that the highly conserved 5
mTOR protein kinase localized at many regions surrounding the transcriptional starting site of 6
genes with a significant enrichment at the NPM1 promoter gene.7 This suggests that mTOR 7
may regulate NPM1 expression and prompted us to address the relationship between these 8
two proteins. For this purpose, we have taken advantage of mouse embryonic fibroblasts 9
(MEFs) lacking Pten since these cells display increased Akt Thr-308 phosphorylation levels 10
due to overactive phosphatidylinositol 3-kinase (PI3K) signaling pathway. In turn, this led to 11
full activation of mTOR complexes as evidence by the feedback up-regulation of phospho-12
Akt at serine 473 and elevated phosphorylation levels of the downstream S6 ribosomal 13
(Ser235/236) target protein. Immunoblot analyses of cell lysates show that Pten-null mouse 14
embryo fibroblasts had increased levels of NPM1 protein compared with their wild-type 15
counterparts (Fig 1A). This change in protein abundance is correlated with a 2.5- to 3.0-fold 16
increase in NPM1 mRNA levels after normalization to the house keeping gene 36b4 (P < 17
0.01) by qRT-PCR assays (Fig 1B). Therefore, these data indicate that enhanced activation of 18
mTOR due to sustained Akt signaling in MEFs lacking Pten may contribute to regulate 19
positively NPM1 at a transcriptional and/or posttranscriptional level. 20
21
NPM1 expression is affected by mTOR inhibition. 22
In an attempt to define if mTOR was responsible for nucleophosmin accumulation, we treated 23
wild-type and Pten negative MEF cultures with 20 nM of the allosteric mTOR inhibitor 24
rapamycin for 24 hours and compared the amounts of NPM1 by western blotting. This long-25
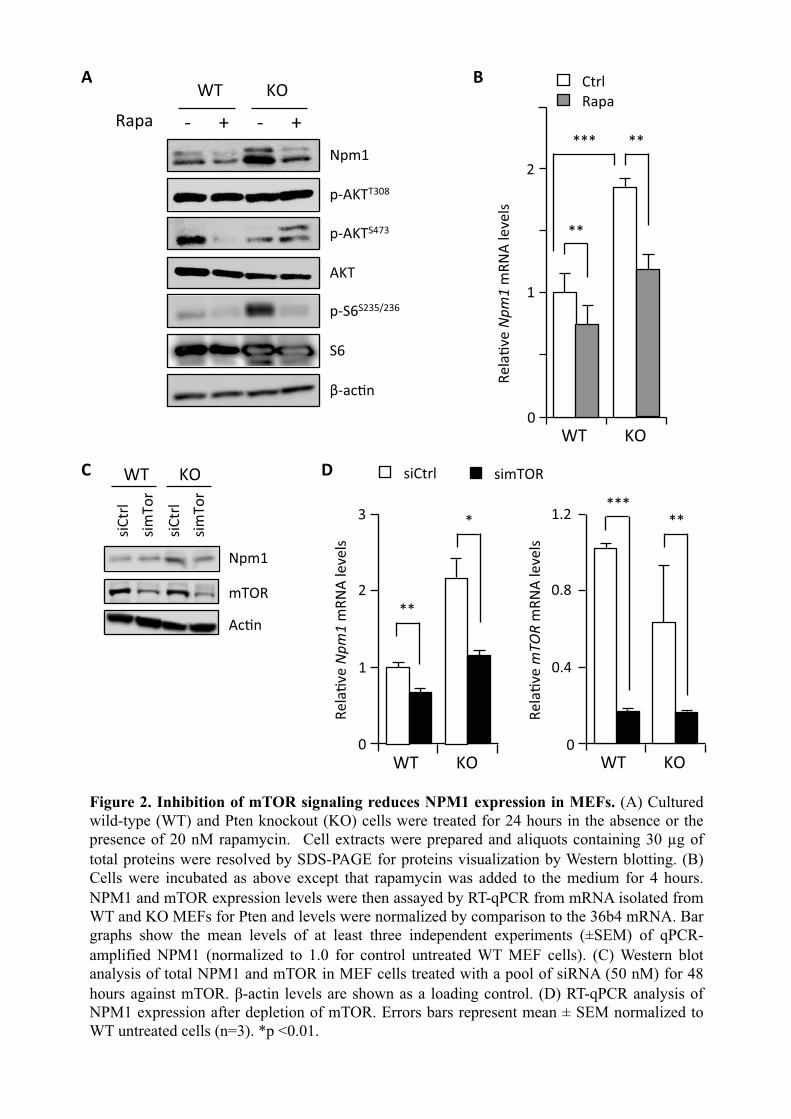
� ��
��!��"#3�"!�
�!�
$���� �
�4�������
�4������
4���
��� � �
����� � �� � ��
&� ��
��
�����
��
��� � �
�����
�� ��
4����
�������
�����
�����
����
��
��
�
�����
����
������
4��� �
���
������� ���� ��
��
!!�
��� � �
!�
"��
"��
�
�����
�����
�����
4��� �
����"��
��� � �
!!�!!!�
��
��
��
�
!!�
��� � �
�����
����
������
4��� �
���
����������
!!�!!!�
Figure 2. Inhibition of mTOR signaling reduces NPM1 expression in MEFs. (A) Cultured wild-type (WT) and Pten knockout (KO) cells were treated for 24 hours in the absence or the presence of 20 nM rapamycin. Cell extracts were prepared and aliquots containing 30 µg of total proteins were resolved by SDS-PAGE for proteins visualization by Western blotting. (B) Cells were incubated as above except that rapamycin was added to the medium for 4 hours. NPM1 and mTOR expression levels were then assayed by RT-qPCR from mRNA isolated from WT and KO MEFs for Pten and levels were normalized by comparison to the 36b4 mRNA. Bar graphs show the mean levels of at least three independent experiments (±SEM) of qPCR-amplified NPM1 (normalized to 1.0 for control untreated WT MEF cells). (C) Western blot analysis of total NPM1 and mTOR in MEF cells treated with a pool of siRNA (50 nM) for 48 hours against mTOR. β-actin levels are shown as a loading control. (D) RT-qPCR analysis of NPM1 expression after depletion of mTOR. Errors bars represent mean ± SEM normalized to WT untreated cells (n=3). *p <0.01.

6
term treatment should likely inhibit both mTOR complexes since previous studies have shown 1
that rapamycin binds to the intracellular protein FKBP12 to generate a drug-receptor complex 2
that then tethers a large fraction of the free mTOR molecules to inhibit the kinase activity of 3
mTORC1 and suppress the assembly and function of mTORC2.19 Unphosphorylated S6 was 4
used as a loading control and phosphos-S6 (Ser235/236) showed that mTORC1 signaling, 5
which stimulates phosphorylation of the ribosomal S6 protein at this site, was inhibited by 6
rapamycin. The experiment in Figure 2A clearly showed that addition of rapamycin compared 7
to vehicle slightly reduced NPM1 protein levels in wild-type MEFs as well as in Pten-deleted 8
MEFs although at a greater extent (50% inhibition, P = 0.09) that might be related to the 9
higher sensitivity to mTOR inhibition. Unlike a recent report,20 this fall in NPM1 abundance 10
is likely to occur in part at the transcriptional level since NPM1 mRNA levels are reduced by 11
25% (p < 0.01) and 40% (p < 0.001) respectively in wild-type and Pten-null MEFs after 4 h of 12
treatment with 20 nM rapamycin (Fig 2B). As shown in Figure 2A, prolonged treatment with 13
rapamycin did not lead to the loss of phospho-Akt (S473) in MEF with PTEN loss (compare 14
lanes 3 and 4) and indicates that the assembly of mTORC2 might not be completely blocked 15
in these conditions. Thus, we asked if it is possible to reduce further the amount of NPM1 16
mRNA by lowering total mTOR to decrease both mTORC1 and mTORC2 complexes. 17
Indeed, a knockdown of mTOR in Pten-deficient MEFs did no trigger a stronger decrease in 18
NPM1 mRNA levels and protein abundance although mTOR mRNA abundance is decreased 19
by more than 80% (Fig. 2C and D). Therefore, our data are consistent with a transcriptional 20
and/or posttranscriptionnal regulation of the NPM1 gene by mTOR complexes. 21
22
mTOR binds to the Npm1 promoter and modulates its transcriptional activity. 23
To confirm the above observations, we examined the DNA binding activity of mTOR in MEF 24
cultures using standard chromatin immunoprecipitation (ChIP). As shown in Figure 3, mTOR 25

&�
��
�#$%�#&�$���&�
���4%&'4&4�
��4��(��&���&���
��#��
���)�4��
���)�$���
�#'��%*+�
� ���
���)�4��
���)�$���
�#'��%*+�
)� ,� )� ,� )� ,� )� ,� )� ,� )� ,�
*�
-�
.�
%/�
�����(
��0���
��&�
(���
��
����
11�
���%��'&�
�� ��'&�
11�
2�
�����(
���34
4��'�3��
�56�%**
*7�
*�
%*�
-*�
8*������
/*�
9*�
4���%)�'&�&�$���5:47� *� *;%� *;/� *;8� *� *;8�#��-�)�'&�
����
*�
-*�
2*�
%8*�
.*�
%/*�
�����(
���34
4��'�3��
�56�%**
*7�
4���%)�'&�&�$���5:47� *� *;%� *;/� *;8� *� *;8�#��-�)�'&�
Figure 3. mTOR binding and activation of Npm1 promoter. (A) Endogenous mTOR is associated with Npm1 promoter region in a rapamycin-independent manner. ChIP assays were conducted using a mTOR antibody in WT and Pten KO MEFs in the absence or the presence of 20 nM rapamycin for 4 hours. Binding of mTOR to Npm1 promoter and Pol-III transcribed tRNAArg/Tyr were determined with PCR primer sets described in Material and Methods. Rabbit IgG were used as negative control. (B) Npm1 is a transcriptional target of mTOR. HeLa (left) and MEF (middle) cell lines were cotransfected for 48 hours with 200 ng of hNPM1-luc plasmid encoding the luciferase under the control of the human NPM1 promoter and increasing amounts of a pCMV-Flag mTOR vector (100, 200 and 500 ng). The luciferase activity measured in WT MEF cells transfected with NPM1-luc or CMV-luc was normalized to 1 and a relative fold-induced luciferase activity was then calculated for Pten KO MEFs in the same experimental conditions (right). Error bars represent ± SEM (n=3; asterisk, p<0.05; two asterisks, p<0.01; three asterisks, p<0.001).

7
immunoprecipitated DNA was analyzed with PCR primer pairs spanning the NPM1 promoter 1
region and the pol III-transcribed tRNAArg/Tyr positive control gene (see Materials and 2
Methods). In lines with previous reports, mTOR specifically targets the pol-III transcribed 3
tRNAArg/Tyr gene, but not a non-mTOR-bound genomic region (negative control) thus 4
confirming the specificity of the reaction. Examination of multiple experiments confirmed the 5
association of endogenous mTOR to the upstream region of the NPM1 gene in wild-type 6
MEFs cultured in normal growth conditions (Fig. 3A, left). This region encompasses the peak 7
found by Chaveroux et al. at -425 from the transcription starting site and seems to be specific 8
of the ChIP assay since no signal was detected with control IgG. Interestingly, Pten loss did 9
not substantially increased mTOR occupancy since we detected equivalent amount of mTOR 10
associated at this promoter (Fig 3A, right). Moreover, treatment with rapamycin did not 11
reduce the level of mTOR occupancy at the NPM1 promoter region suggesting that mTOR 12
binding is insensitive to inhibition with rapamycin. 13
The presence of mTOR at NPM1 template may allow it to regulate transcription of this gene 14
directly. This possibility was explored using the human NPM1 promoter 5’flanking region 15
from -1200 to +87 fused to the luciferase reporter gene. The resulting hNPM1-luciferase 16
construct was transiently transfected into HeLa and MEF cells either alone or in combination 17
with increasing amounts of mTOR expression vector. Following transfection, cells were 18
maintained in serum-free media and 24 h later, cells were harvested and luciferase expression 19
was measured. Ectopic expression of mTOR in HeLa and MEF cells nearly double the NPM1 20
promoter activity in a dose dependent manner in our experiment conditions (Fig. 3B, left and 21
middle). In addition, we found that the luciferase activity was strongly enhanced in Pten-null 22
MEFs (9.5-fold, p < 0.01) in comparison with wild-type MEFs (Fig 3B, right). In comparison, 23
cells were transfected with a CMV-driven luciferase expression vector (pcDNA4/T0-luc 24
provided by H Kleinert). As shown in Fig. 3B (left panel) the activity from the hNPM1-25
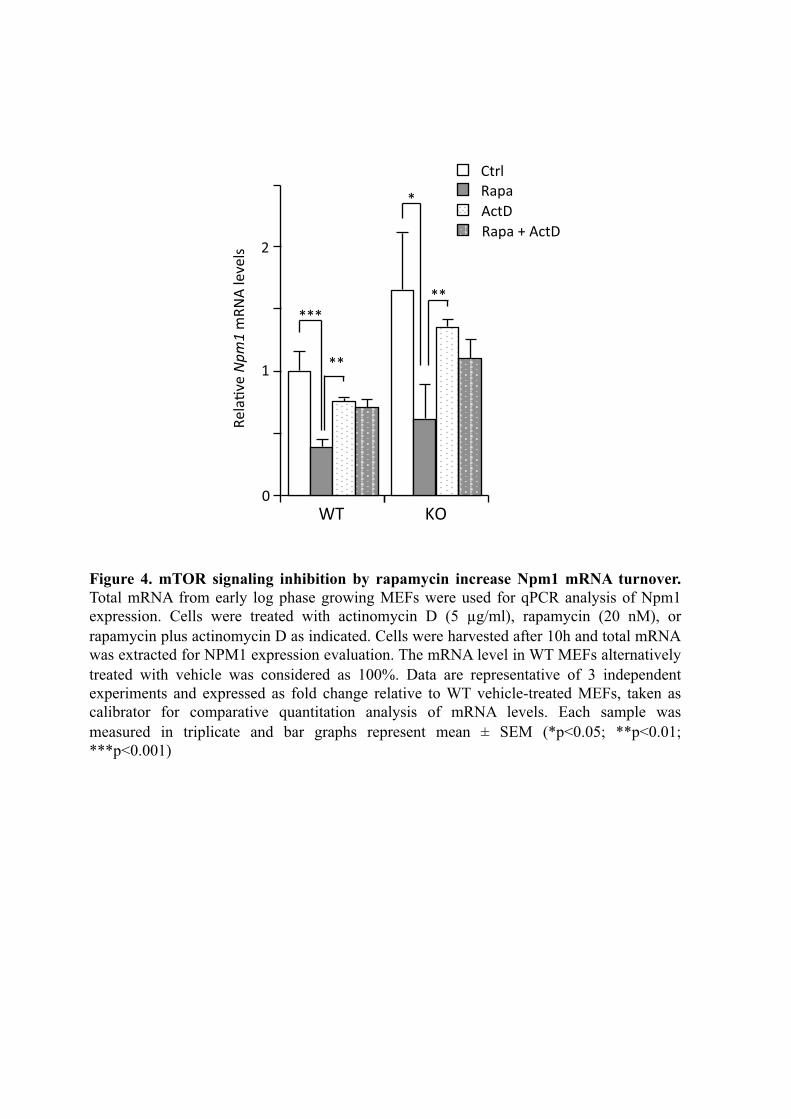
(�
��
�
!!!�
��� ���
�����
����
��� �
4���
�����
���������4����������4����
!�
!!�
!!�
Figure 4. mTOR signaling inhibition by rapamycin increase Npm1 mRNA turnover. Total mRNA from early log phase growing MEFs were used for qPCR analysis of Npm1 expression. Cells were treated with actinomycin D (5 µg/ml), rapamycin (20 nM), or rapamycin plus actinomycin D as indicated. Cells were harvested after 10h and total mRNA was extracted for NPM1 expression evaluation. The mRNA level in WT MEFs alternatively treated with vehicle was considered as 100%. Data are representative of 3 independent experiments and expressed as fold change relative to WT vehicle-treated MEFs, taken as calibrator for comparative quantitation analysis of mRNA levels. Each sample was measured in triplicate and bar graphs represent mean ± SEM (*p<0.05; **p<0.01; ***p<0.001)

8
luciferase reporter gene in Pten deleted MEFs was 4.6-fold more stimulated when compared 1
to that of the CMV-luciferase control gene (p = 0.01). This latter result is contrasting with the 2
data from our ChIP assays since we did not observed any substantial increase in the 3
association of mTOR with the endogenous Npm1 promoter in MEF cells inactivated for Pten 4
(Fig. 3A). Nevertheless, these results clearly indicate that mTOR increases the transcriptional 5
activity of the NPM1 promoter although we cannot conclude whether this association requires 6
other transcriptional co-regulators or component of the mTORC1/2 complexes. 7
8
Rapamycin regulates NPM1 steady-state mRNA levels. 9
The work described above shows that rapamycin caused a rapid significant decreased in 10
NPM1 mRNA levels in both wild-type and Pten-/- MEF cells at 4 h (Fig. 2A) suggesting that 11
inhibition of mTOR signaling might affect gene transcription and/or mRNA turnover to 12
regulate NPM1 gene expression. This latter assumption is supported by several lines of 13
evidence showing that signaling through mTOR promotes mRNA decay.21 To test this 14
possibility, the levels of NPM1 mRNA were thus quantified by RT-qPCR in MEF cells in the 15
presence of the RNA polymerase II inhibitor actinomycin D (ActD) or with rapamycin. As 16
shown in Figure 4, mTOR inactivation by rapamycin down-regulates NPM1 mRNA by 17
around 60% in both wild-type (0.41±0.04 of the control vehicle value, p < 0.001) and Pten 18
deleted MEFs (0.64±0.29 of the control vehicle value, p < 0.05). In contrast, ActD alone only 19
decreased mRNA levels from 25% in both cell lines suggesting that inhibition with rapamycin 20
strongly upregulates NPM1 mRNA turnover. Of note, addition of ActD to cells treated with 21
rapamycin prevents the effect observed with rapamycin alone. Thus the enhanced turnover of 22
NPM1 mRNA in mTOR-inhibited cells can be suppressed by inhibition of RNA polymerase 23
II transcription. This may reflect the need of a transcriptional event to recruit the general 24
cellular mRNA degradation machinery to regulate NPM1 mRNA turnover. 25
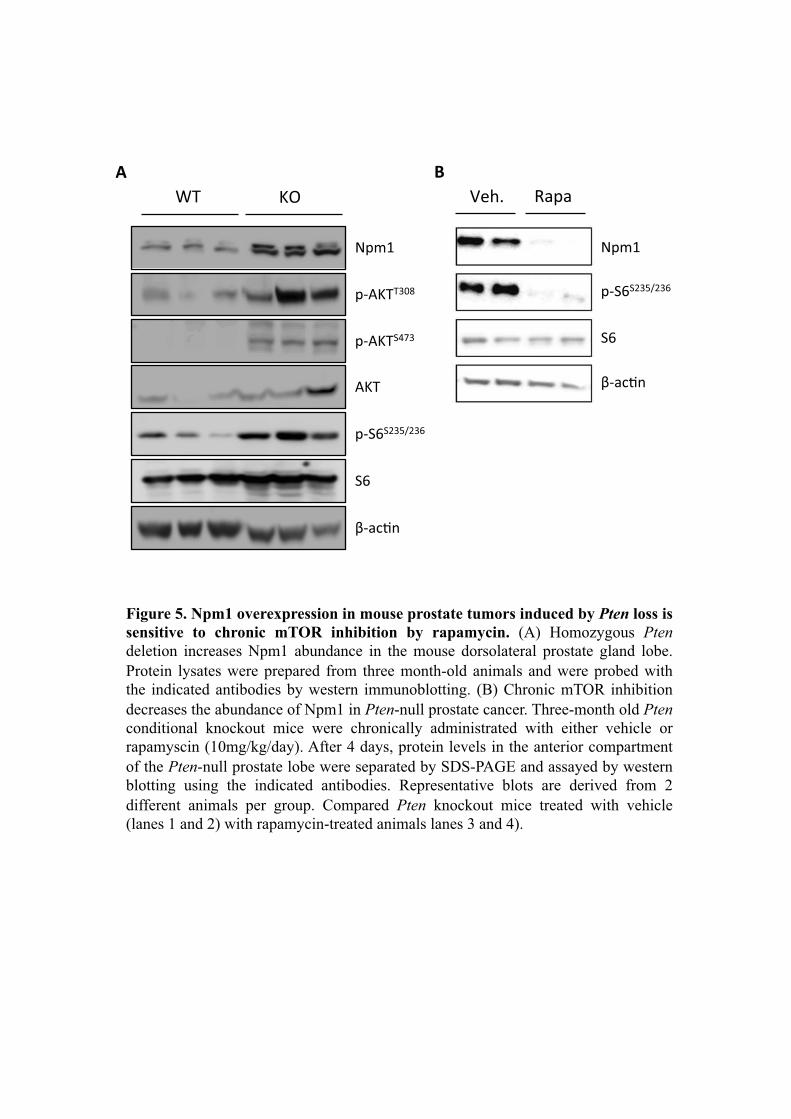
��5�"�#�"�5�
�� ��
�4�������
�4�������
4���
�5�
������
��� ���&�
�� ��
��5�"�#�"�5�
�5�
������
����� �������
Figure 5. Npm1 overexpression in mouse prostate tumors induced by Pten loss is sensitive to chronic mTOR inhibition by rapamycin. (A) Homozygous Pten deletion increases Npm1 abundance in the mouse dorsolateral prostate gland lobe. Protein lysates were prepared from three month-old animals and were probed with the indicated antibodies by western immunoblotting. (B) Chronic mTOR inhibition decreases the abundance of Npm1 in Pten-null prostate cancer. Three-month old Pten conditional knockout mice were chronically administrated with either vehicle or rapamyscin (10mg/kg/day). After 4 days, protein levels in the anterior compartment of the Pten-null prostate lobe were separated by SDS-PAGE and assayed by western blotting using the indicated antibodies. Representative blots are derived from 2 different animals per group. Compared Pten knockout mice treated with vehicle (lanes 1 and 2) with rapamycin-treated animals lanes 3 and 4).

9
NPM1 is a downstream effector of mTOR signaling with implication in prostate cancer. 1
Our previous report that NPM1 is overexpressed in prostate cancer22, together with the fact 2
that mTOR serves a pivotal role in cancer suggest that these two proteins may be functionally 3
link to regulate events involved in prostate cancer cell growth control. To answer this, we 4
used the conditional murine Pten prostate cancer model with a constitutive activation of the 5
Akt/mTOR signaling pathway in the prostate epithelium.23 This model is greatly appropriate 6
since it recapitulates the disease progression seen in humans with the formation of high-grade 7
prostatic intraepithelial neoplasia and invasive carcinoma. Consistent with our findings in 8
MEF cells with deletion of Pten, the abundance of the NPM1 protein is increased in prostate 9
tumors of Pten-/- animals in conjunction with Akt activation and phosphorylation of the 10
downstream mTOR effector S6 ribosomal protein (Fig. 5A). Of note, chronical administration 11
for 4 days with rapamycin (10 mg/kg/day) almost completely inhibits the expression of 12
NPM1 in the dorsolateral compartment of prostate gland of Pten knockout mice (Fig 5B, 13
lanes 3-4 compared with lanes 1-2). The lack of phosphorylation of S6 protein in rapamycin-14
treated animals confirms that the mTOR pathway was efficiently blocked by our treatment 15
and let us conclude that mTOR also regulates NPM1 expression in vivo. 16
One important remaining question is to know whether NPM1 acts as a downstream mediator 17
of mTOR to promote cellular adaptation for growth. This questioning cannot be solved by 18
deleting Npm1 gene expression in the Pten knockout mice because it complete inactivation 19
leads to embryonic lethality at mid-gestation24, and no mouse model for conditional 20
inactivation of Npm1 is currently available. To circumvent this limitation, we have assessed 21
the proliferation rate of wild-type and Pten-/- MEFs, non-silenced or silenced for Npm1, using 22
the bromodeoxyuridine (BrdU) incorporation cell-based assay. As shown in Figure 6, NPM1 23
silencing efficiency was near 90% compared to control cultures transfected with a non-24
specific control unrelated GFP siRNA sequence (Fig. 6A). Interestingly, the growth 25
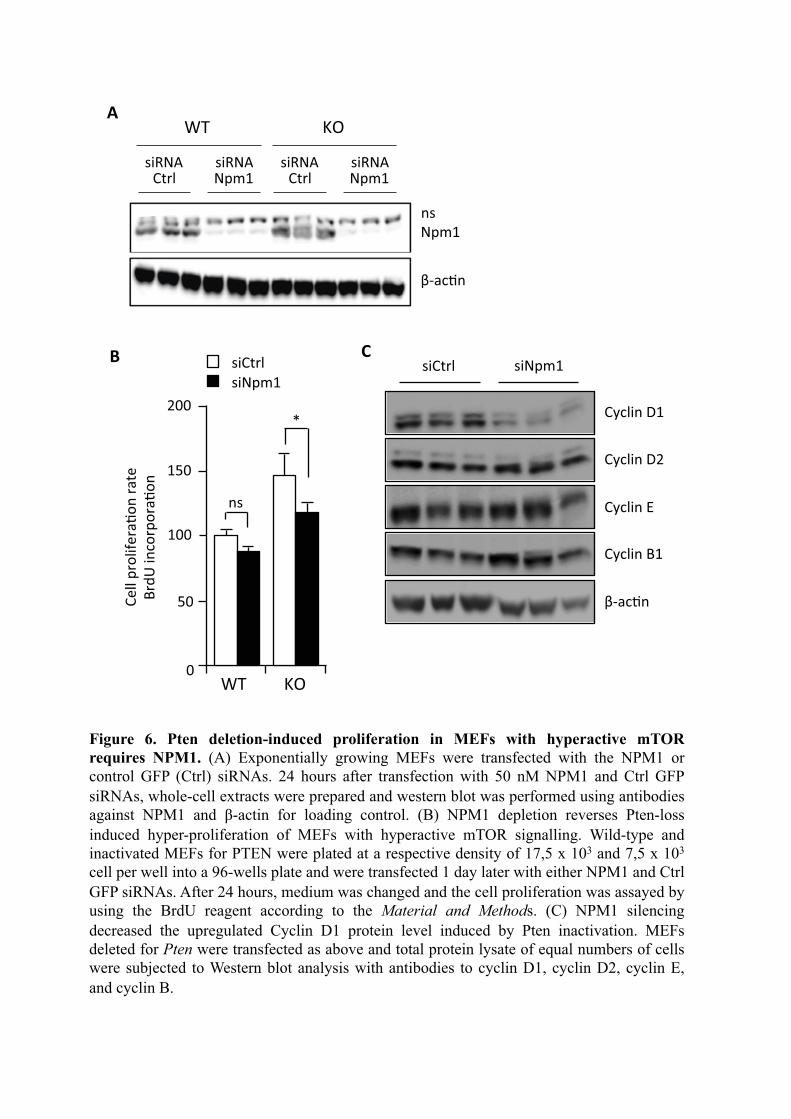
�6� ���
!$� ���
6��4�-�"��
6��4� ���
6��4�-�"��
6��4� ���
��� ���&�
�� ��
-� ����'��
-� ����'%�
-� ����.�
-� �������
6�-�"�� 6� ���
7��
�7��
��
-���� "�����"���
��"�
���
�"��
��� �" �"����
�
6�-�"��6� ���
%���
�6�
��� ���
��
����
!$� ���
Figure 6. Pten deletion-induced proliferation in MEFs with hyperactive mTOR requires NPM1. (A) Exponentially growing MEFs were transfected with the NPM1 or control GFP (Ctrl) siRNAs. 24 hours after transfection with 50 nM NPM1 and Ctrl GFP siRNAs, whole-cell extracts were prepared and western blot was performed using antibodies against NPM1 and β-actin for loading control. (B) NPM1 depletion reverses Pten-loss induced hyper-proliferation of MEFs with hyperactive mTOR signalling. Wild-type and inactivated MEFs for PTEN were plated at a respective density of 17,5 x 103 and 7,5 x 103 cell per well into a 96-wells plate and were transfected 1 day later with either NPM1 and Ctrl GFP siRNAs. After 24 hours, medium was changed and the cell proliferation was assayed by using the BrdU reagent according to the Material and Methods. (C) NPM1 silencing decreased the upregulated Cyclin D1 protein level induced by Pten inactivation. MEFs deleted for Pten were transfected as above and total protein lysate of equal numbers of cells were subjected to Western blot analysis with antibodies to cyclin D1, cyclin D2, cyclin E, and cyclin B.

10
advantage of Pten-/- MEFs is significantly reduced after depletion of Npm1 (20% inhibition, p 1
< 0.05) while NPM1 down-regulation had a moderate effect on the proliferation of control 2
wild-type MEFs (Fig. 6B). When compared to control wild-type MEFs, NPM1 silencing 3
prevents the growth enhancement induced by Pten loss. One possible explanation for this may 4
reside in the decreased expression of cyclin D1 in Pten inactivated MEFs silenced for NPM1 5
(Fig. 6C). This is an interesting finding since Pten act as general negative regulator of cyclin 6
D expression.25 Indeed, inhibiting NPM1 in MEF cells with hyperactive AKT/mTOR 7
signaling may thus contribute to counteract the increased expression of cyclin D1 in Pten 8
inactivated MEFs and to restore cell cycle regulation in these cells. Collectively, these results 9
suggest that NPM1 may function downstream of mTOR to coordinate mitogenic signals to 10
control cell growth and cell-cycle progression. 11
12

�

11
Discussion 1
Signaling through the mammalian target of rapamycin (mTOR) regulates many diverse 2
cellular processes, especially those contributing to cell growth and proliferation. In the past 3
couple of years, many important advances have been made in identifying new components of 4
the mTOR network and in further elucidating connections between known components. In the 5
present study, we provide evidence that Npm1 is a downstream signaling target of mTOR that 6
may participate in the mTOR-dependent coordination of cell growth and cell-cycle 7
progression in response to mitogenic signals. By actively shuttling between the nucleolus and 8
cytoplasm, Npm1 was reported too to take on various distinct cellular functions. Indeed, the 9
biological significance of Npm1 in ribosome biogenesis and transport, anti-apoptotic activity, 10
the regulation of centrosome duplication and the regulation of tumors suppressors have been 11
largely documented although its upstream regulatory mechanisms still represent a major gap 12
to fulfill.14, 26-28 13
Using cell lines and animal models bearing a targeted deletion of the Pten gene that leads to 14
hyperactivation of the mTOR signaling, we find that Pten-null MEFs display more Npm1 15
protein and mRNA than their wild-type counterpart. This increase is also observed in the 16
prostate gland of the conditional murine Pten cancer model indicating thus that active mTOR 17
signaling pathway in prostate cancer cells might also impinge on Npm1 expression in vivo. In 18
both situations, Npm1 enhanced expression is sensitive to rapamicyn inhibition of mTOR 19
although our in vitro data show that mTOR association with the Npm1 promoter is insensitive 20
to rapamycin. Actually, it is not clear how this kinase acts to increase the transcriptional 21
activity we observed toward the Npm1 promoter and we cannot exclude that this later may 22
serve to assemble competent transcriptional mTORC complexes onto chromatin. Of interest, 23
the mTOR enriched 5’ enhancer promoter region of Npm1 includes a functional YY1-binding 24
motif that has been already characterized.15 It is thus tempting to suggest that mTOR might 25

12
control Npm1 gene transcription through a YY1 transcriptional complex as reported for other 1
genes that adapts the mitochondrial oxidative function in response to nutrients and hormonal 2
signals.17 This should be easily assess by testing the mTOR-YY1 interaction into chromatin to 3
determine whether this interface constitute a key regulatory step to enhance Npm1 gene 4
expression when mTOR is hyperactive. Moreover, silencing YY1 as well as components of 5
the TORC complexes namely the Raptor and Rictor proteins may provide additional valuable 6
insights to unravel the mechanism of Npm1 gene transcriptional activation by mTOR 7
dependent complexes. 8
Interestingly, mTOR binds to promoters of RNA polymerase I- and III- transcribed genes and 9
these associations with ribosomal DNA (rDNA) and transfer RNA (tRNA) enhancer regions 10
are believed to be crucial to the protein synthetic capacity of the cell and its growth ability. By 11
directly controlling Npm1 gene expression, mTOR would allow the cell to connect nutrient 12
and/or hormonal signals to activate ribosome biogenesis. This is of particularly interest 13
because Npm1 was initially described as a ribosome chaperone or assembly factor that was 14
found associate with pre-ribosomal RNA processing. In support of this notion, a fraction of 15
soluble Npm1 interacts with pre-ribosomal 60S particles.29 It facilitates cleavage of ribosomal 16
RNA in vitro and acts as an endoribonuclease for the maturing rRNA transcript. 30, 31 Not 17
surprisingly, inhibition of Npm1 thus results in a decrease in cell growth as a consequence of 18
impaired processing of pre-rRNA to mature 28S rRNA and inhibition of ribosome subunit 19
export from the nucleolus to the cytoplasm.32, 33 Altogether, these observations support the 20
notion of a functional coupling between mTOR and Npm1 to coordinate the manufacturing of 21
ribosomes and our finding that rapamycin down-regulates Npm1 abundance may provide an 22
explanation to understand why mTOR signaling inhibition interferes with the processing of 23
rRNA as recently noted by Iadevaia et al.34 24

13
The regulation of Npm1 mRNA turnover we have reported in this study constitutes an 1
unexpected finding that raises important questions in regards to the specific elements within 2
the Npm1 mRNA that controls its stability and the cytoplasmic mRNA degradation machinery 3
that might be activated when mTOR pathway is inhibited. In fact, multiples mRNA were 4
demonstrated to be destabilized after treatment with rapamycin in yeast21 and our findings 5
now define Npm1 as a new potential target of the mTOR regulated mRNA decay. From our 6
complementary experiments, Npm1 mRNA destabilization is unlikely to occur through the 7
AU-rich-mediated decay machinery which has been recently described by Cammas et al.35 8
While in muscle cells the AU-rich elements (AREs) located at the 3’ untranslated region (3’-9
UTR) sequence of the Npm1 mRNA were shown to participate to the posttranscriptional 10
downregulation of the Npm1 gene expression, inhibition of mTOR by rapamycin did not 11
destabilizes a Renilla luciferase reporter gene fused to the Npm1-3’-UTR sequence (Rluc-12
NPM-3’ provided by A Cammas) in transiently transfected MEF cells (not shown). This allow 13
us to conclude that the destabilization of Npm1 mRNA may be regulated through another 14
mechanism of mRNA decay and the actinomycin D-mediated stabilization of Npm1 mRNA in 15
presence of rapamycin suggests the need of a pol II-dependent transcription event to control 16
Npm1 mRNA stability. One attracting possibility is the transcriptional repression of antisense 17
non-coding RNA (ncRNA) by mTOR dependent complexes. This hypothesis fits well with 18
our data, since inhibition of mTOR by rapamycin could relief the transcriptional repression of 19
ncRNAs and allow their complementary association with Npm1 mRNA for targeted 20
degradation. In this sense, mRNA degradation by microRNA (miRNA), small interfering 21
RNA (siRNA) and large intergenic non-coding RNA (lncRNA) all appears as plausible 22
possibilities and inhibition of pol-II transcription by actinomycin D should stabilize Npm1 23
mRNA in presence of rapamycin. This is a very interesting issue to address and further works 24

14
is required to completely understand how mTOR pathway is post-transcriptionally regulating 1
Npm1 gene expression. 2
Nevertheless, these informations combined with the results we provided here may have 3
important implications for cancer biology since mTOR is a critical effector of cell-signaling 4
pathways which is commonly deregulated in human cancers. This is particularly the case in 5
prostate cancer where several molecules within the AKT/mTOR signaling pathway are altered 6
during prostate cancer progression with potential diagnostic markers including PTEN, p-7
AKT, and mTOR.36, 37 In addition, mTOR signaling molecules have been specially proposed 8
as a central player in prostate cancer because of their influence on cell growth capacities.38 9
The general belief is that activated mTOR may provide certain tumors cells with a growth 10
advantage by promoting protein synthesis, which is the best-described function of mTOR. By 11
regulating Npm1 gene expression through transcriptional and posttranscriptional mechanisms, 12
increased mTOR activity may lead to the enhance Npm1 expression that has been already 13
reported by us and others in human prostate cancer.22, 39 It is currently unknown whether 14
deregulated mTOR and Npm1 are associated in clinical samples of prostate cancer but it is 15
tempting to speculate that mTOR might stabilize Npm1 mRNA level to stimulate cell growth 16
by regulating protein synthesis and cell cycle progression. The increased abundance of Npm1 17
in Pten knockout mouse prostate cancer model is in good agreement with this assumption and 18
the possibility to restore cell cycle progression in Pten inactivated MEF cells silenced for 19
Npm1 further strengthen its functional role as a downstream effector of mTOR signaling. 20
Collectively, our results suggest that mTOR is closely involved in the regulation of Npm1 21
gene expression with potential implication for prostate cancer therapies that may benefit from 22
a better understanding of the mTOR signaling pathway to identify new molecule targets to 23
inhibit in prostate cancer progression and the evolving treatment landscape of resistant 24
prostate cancer. 25

�

15
Materials and Methods 1
Chemicals and biochemicals. 2
Life Technologies (Cergy Pontoise, France) supplies the Dulbecco’s modified Eagle medium 3
(DMEM, cat#41965), glutamine (cat#25030), antibiotics (Pen/Strep, cat#15140), DPBS 10X 4
(cat#14200). Fetal bovine serum was obtained from Biowest (cat#51810). Rapamycin and the 5
non-ionic emulsifier Cremephor EL were respectively purchased from LC Laboratories 6
(#catR5000) and Sigma-Aldrich (#catC5135). Goat polyclonal anti-NPM1 (C-19, sc-6013), 7
rabbit polyclonal anti-cyclin D1 (M20, sc-718), rabbit polyclonal anti-cyclin E (M-20, sc-8
481), rabbit polyclonal anti-cyclin A (C-19, sc-596) and rabbit polyclonal anti-cyclin B1 (H-9
433, sc-752) were purchased from Santa Cruz Biotechnology; rabbit polyclonal anti-phospho-10
AKT Ser473 (ab81283) from AbCam, rabbit monoclonal anti-phospho-AKT Thr308 11
(cat#2965), rabbit monoclonal anti-AKT (cat#9272), rabbit monoclonal anti-S6 (cat#2217), 12
rabbit polyclonal anti-phospo-S6 Ser235/236 (cat#2211), rabbit polyclonal anti-mTOR 13
(cat#2972) from cell signaling; and rabbit polyclonal anti-βactin from Sigma-Aldrich 14
(cat#A2066). The Npm1-luciferase reporter gene was obtained by cloning the PCR amplified 15
DNA fragment from the -1200 to +86 promoter region of the human NPM1 gene (Forward 16
primer, 5’-GATCGGTACCGTGACACAGGAGCTCTCAGAAAGG-3’; Reverse primer 5’-17
GATCCTCGAGACTTAGGTAGGAGAGAAGGCGGAC-3’) directly into the KpnI/XhoI 18
sites of the pGL3-basic luciferase reporter vector (Promega, Charbonnière Les Bains, France). 19
20
Cell cultures and transfections. 21
Mouse Embryonic Fibroblast cells were cultured in 10 cm plates in DMEM supplemented 22
with 10% fetal calf serum at 37°C in a humidified atmosphere containing 5% CO2. HeLa cells 23
were maintained in DMEM supplemented with 10% fetal calf serum, as above. For transient 24
transfections, 3 x 105 HeLa cells per well were plated in 6 well-plates and the next day, 25

16
transient transfections were performed using the Metafecten procedure (Biontex Laboratories 1
GmbH, Germany). For wild-type and Pten knockout MEFs, 2 x 105 and 1 x 105 cells/well 2
were plated respectively and cells were transfected the next day according to the same 3
procedure. After 12 h, the medium was replaced with fresh complete medium and 2 days 4
later, cells were lysed and the luciferase activity was measured with the assay system from 5
Yelen. For depletion of Npm1 and mTOR, we used the siGenome mouse Npm1 (cat#18148) 6
and mouse mTOR (cat#56717) siRNA- SMART pools from Dharmacon (sequences available 7
upon request). 8
9
Animals and treatments. 10
Mice were housed individually (24°C; light on 07.00-19.00h) and allowed free access to 11
water and diet (A03 standard diet cat#U8200G10R from SAFE, Augy, France). When stated, 12
animals received daily i.p. injections of either vehicle for the control or Sirolimus at a dose of 13
10mg/kg per day for 4 days. Mice were killed and tissues were freeze-clamped and stored at -14
80°C for further analyses. All animal care and experimental procedures were in accordance 15
with the French guidelines for animal experimentation and were ethically approved by the 16
local committee on Animal Experiments. 17
18
RNA extraction and RT-qPCR analysis. 19
Total RNA was isolated from MEF cells using the RNAzol method according to the 20
manufacturer’s instructions (Quantum, Montreuil-sous-bois, France). For cDNA synthesis, 21
five hundred nanograms mRNA were reverse transcribed for 1 h at 42°C with 5pmoles of 22
random hexamer primers, 200 units reverse transcriptase (MuMLV RT, Promega), 2mM 23
dNTPs and 20 units RNAsin (Promega) as described. 40 A half microliter of one-tenth dilution 24
of the cDNA was used in each quantitative PCR reaction. These were conducted with the 25

17
mouse Npm1 sequence specific primers (Forward: 5’-GCAGGGGCAAAAGATGAGT-3’; 1
Reverse: 5’-CAAAGCCCCCTAGGGAAAC). Each reaction was performed in triplicate in a 2
final volume of 15µl with 0.75µl of the appropriate 20X probe mix and 7.5µl of PCR 3
Mastermix (Precision-iC., PrimerDesign.co.uk). Relative mRNA accumulation was 4
determined by the ΔΔCt method and 36b4 (Forward: 5’-GTCACTGTGCCAGCTCAGAA-3’, 5
Reverse: 5’-TCAATGGTGCCTCTGGAGAT-3’) was used for quantative normalization of 6
cDNA in each cell-derived sample. Statistical analysis was performed with Student's t-test. 7
8
Preparation of cell extracts and Western Blot analysis. 9
Mouse tissues were harvested from three-month old wild-type and PTENLoxP/LoxP ; PB-Cre4+ 10
mice and immediately homogenized in ice cold NaCl buffer [0.42M NaCl, 20mM Hepes (pH 11
8.0), 1.5mM MgCl2, 0.2mM EDTA, 25% glycerol, 0.1% NP40, 1mM PMSF, 1μg/ml 12
apoprotinin, 1μg/ml leupeptin]. To make the whole-cell lysates, MEF cells were washed with 13
1X PBS and resuspended in ice cold NaCl buffer. After brief sonication on ice and 14
clarification for 30 min at 15,000 x g, 40µg of proteins were resolved through SDS-15
polyacrylamide gels and transferred to nitrocellulose membranes. Membranes were blocked 16
with milk buffer [5% milk in Tris-buffered saline (TBS)-Tween 20] for 1 h and incubated 17
with the indicated primary antibody in milk buffer overnight at 4°C. The membranes were 18
washed three times with TBS-Tween 20 and then incubated 90 min with horseradish 19
peroxidase-conjugated donkey anti-rabbit or goat anti-rabbit immunoglobulin G secondary 20
antibody (P.A.R.I.S biotech, Compiègne, France), followed by enhanced chemiluminescence 21
(Santa Cruz Biotechnology) according to manufacturer's instruction. 22
23
24
25

18
Cell proliferation assay. 1
Cell proliferation was determined using the cell-based BrdU (Bromode oxyuridine) 2
proliferation assay. Briefly, MEF cells were seeded in 96-wells plate and transfected with 50 3
nM Npm1 or control GFP siRNAs. The next days, medium was changed and cells were 4
cultured in 200 µl of complete medium. After 48 hours, the BrdU labelling solution was 5
added at a final concentration of 10 mM for 2.5 hours and the absorbance was measured at 6
655 nm following fixation after an additional incubation of 1 hour with the anti BrdU-POD 7
solution according to the Cell Proliferation ELISA BrdU colorimetric kit from Roche Applied 8
Science. 9
10
Chromatin immunoprecipitation. 11
Chromatin immunoprecipitation assays were performed according to the Upstate protocol. 12
Sheared DNA fragments were immunoprecipitated with an anti-mTOR (Abcam, ab32028) 13
and a mock rabbit IgG (Diagenode) was used as a negative control. Sequences of primers 14
used for mouse Npm1 promoter PCR were: Forward 5’-GCAGGGGCAAAAGATGAGT-3’ 15
and Reverse 5’-CAAAGCCCCCTAGGGAAAC). Primer sets for mouse tRNAArg/Tyr (chr3 16
segment 19528000 to 19528500: Forward 5’- CTTGCCGCTCCCTTCGATAG-3’; Reverse 17
5’-AGAGGAGGAGGAGCGCACCTG-3’) and a negative control non-mTOR-bound 18
genomic region (Forward 5’-TTGGCATTGATATTGGGGGTGGGAGCAACT-3’; Reverse 19
5’ GACTTCTTACTTTGACGCTTTCCTCCA-3’) were used as described.7 PCR conditions 20
for all primers were as follows: 94°C for 3 min, followed by 32-35 cycles at 94°C for 30s, 55-21
58°C for 45s and 72°C for 45s. 22
23
24
25

19
Statistical analyses 1
All experiments were repeated at least three times and results were expressed as mean ± SEM. 2
Statistical differences between groups were determined by Student’s t test and the criterion 3
for statistical significance was P<0,05. For western blotting results, the band intensities were 4
measured by using the ImageJ and normalized with β-actin. 5
6
7

20
Acknowledgments 1
This research was supported by grants awarded by the Centre National de la Recherche 2
Scientifique (CNRS), the Université Blaise Pascal (UBP), the Association pour la Recherche 3
sur les Tumeurs de la Prostate (ARTP) and the Association pour la Recherche sur le Cancer 4
(ARC). RB is holder of a studentship from the French Foundation for Medical Research 5
(FRM). 6

�

21
References 1
1. Aramburu J, Ortells MC, Tejedor S, Buxade M, Lopez-Rodriguez C. Transcriptional 2 regulation of the stress response by mTOR. Sci Signal 2014; 7:re2. 3
2. Laplante M, Sabatini DM. mTOR signaling in growth control and disease. Cell 2012; 4 149:274-93. 5
3. Hara K, Maruki Y, Long X, Yoshino K, Oshiro N, Hidayat S, Tokunaga C, Avruch J, 6 Yonezawa K. Raptor, a binding partner of target of rapamycin (TOR), mediates TOR action. 7 Cell 2002; 110:177-89. 8
4. Sarbassov DD, Ali SM, Kim DH, Guertin DA, Latek RR, Erdjument-Bromage H, 9 Tempst P, Sabatini DM. Rictor, a novel binding partner of mTOR, defines a rapamycin-10 insensitive and raptor-independent pathway that regulates the cytoskeleton. Curr Biol 2004; 11 14:1296-302. 12
5. Ma XM, Blenis J. Molecular mechanisms of mTOR-mediated translational control. 13 Nat Rev Mol Cell Biol 2009; 10:307-18. 14
6. Tsang CK, Liu H, Zheng XF. mTOR binds to the promoters of RNA polymerase I- 15 and III-transcribed genes. Cell Cycle 2010; 9:953-7. 16
7. Chaveroux C, Eichner LJ, Dufour CR, Shatnawi A, Khoutorsky A, Bourque G, 17 Sonenberg N, Giguere V. Molecular and genetic crosstalks between mTOR and ERRalpha are 18 key determinants of rapamycin-induced nonalcoholic fatty liver. Cell Metab 2013; 17:586-98. 19
8. Feuerstein N, Mond JJ. "Numatrin," a nuclear matrix protein associated with induction 20 of proliferation in B lymphocytes. J Biol Chem 1987; 262:11389-97. 21
9. Chan WY, Liu QR, Borjigin J, Busch H, Rennert OM, Tease LA, Chan PK. 22 Characterization of the cDNA encoding human nucleophosmin and studies of its role in 23 normal and abnormal growth. Biochemistry 1989; 28:1033-9. 24
10. Feuerstein N, Chan PK, Mond JJ. Identification of numatrin, the nuclear matrix 25 protein associated with induction of mitogenesis, as the nucleolar protein B23. Implication for 26 the role of the nucleolus in early transduction of mitogenic signals. J Biol Chem 1988; 27 263:10608-12. 28
11. Li J, Zhang X, Sejas DP, Bagby GC, Pang Q. Hypoxia-induced nucleophosmin 29 protects cell death through inhibition of p53. J Biol Chem 2004; 279:41275-9. 30
12. Wu MH, Chang JH, Chou CC, Yung BY. Involvement of nucleophosmin/B23 in the 31 response of HeLa cells to UV irradiation. Int J Cancer 2002; 97:297-305. 32
13. Wu MH, Yung BY. UV stimulation of nucleophosmin/B23 expression is an 33 immediate-early gene response induced by damaged DNA. J Biol Chem 2002; 277:48234-40. 34

22
14. Grisendi S, Mecucci C, Falini B, Pandolfi PP. Nucleophosmin and cancer. Nat Rev 1 Cancer 2006; 6:493-505. 2
15. Chan PK, Chan FY, Morris SW, Xie Z. Isolation and characterization of the human 3 nucleophosmin/B23 (NPM) gene: identification of the YY1 binding site at the 5' enhancer 4 region. Nucleic Acids Res 1997; 25:1225-32. 5
16. Zeller KI, Haggerty TJ, Barrett JF, Guo Q, Wonsey DR, Dang CV. Characterization of 6 nucleophosmin (B23) as a Myc target by scanning chromatin immunoprecipitation. J Biol 7 Chem 2001; 276:48285-91. 8
17. Cunningham JT, Rodgers JT, Arlow DH, Vazquez F, Mootha VK, Puigserver P. 9 mTOR controls mitochondrial oxidative function through a YY1-PGC-1alpha transcriptional 10 complex. Nature 2007; 450:736-40. 11
18. Duvel K, Yecies JL, Menon S, Raman P, Lipovsky AI, Souza AL, Triantafellow E, 12 Ma Q, Gorski R, Cleaver S, et al. Activation of a metabolic gene regulatory network 13 downstream of mTOR complex 1. Mol Cell 2010; 39:171-83. 14
19. Sarbassov DD, Ali SM, Sengupta S, Sheen JH, Hsu PP, Bagley AF, Markhard AL, 15 Sabatini DM. Prolonged rapamycin treatment inhibits mTORC2 assembly and Akt/PKB. Mol 16 Cell 2006; 22:159-68. 17
20. Pelletier CL, Maggi LB, Jr., Brady SN, Scheidenhelm DK, Gutmann DH, Weber JD. 18 TSC1 sets the rate of ribosome export and protein synthesis through nucleophosmin 19 translation. Cancer Res 2007; 67:1609-17. 20
21. Albig AR, Decker CJ. The target of rapamycin signaling pathway regulates mRNA 21 turnover in the yeast Saccharomyces cerevisiae. Mol Biol Cell 2001; 12:3428-38. 22
22. Leotoing L, Meunier L, Manin M, Mauduit C, Decaussin M, Verrijdt G, Claessens F, 23 Benahmed M, Veyssiere G, Morel L, et al. Influence of nucleophosmin/B23 on DNA binding 24 and transcriptional activity of the androgen receptor in prostate cancer cell. Oncogene 2008; 25 27:2858-67. 26
23. Wang S, Gao J, Lei Q, Rozengurt N, Pritchard C, Jiao J, Thomas GV, Li G, Roy-27 Burman P, Nelson PS, et al. Prostate-specific deletion of the murine Pten tumor suppressor 28 gene leads to metastatic prostate cancer. Cancer Cell 2003; 4:209-21. 29
24. Grisendi S, Bernardi R, Rossi M, Cheng K, Khandker L, Manova K, Pandolfi PP. Role 30 of nucleophosmin in embryonic development and tumorigenesis. Nature 2005; 437:147-53. 31
25. Diao L, Chen YG. PTEN, a general negative regulator of cyclin D expression. Cell 32 Res 2007; 17:291-2. 33
26. Lim MJ, Wang XW. Nucleophosmin and human cancer. Cancer Detect Prev 2006; 34 30:481-90. 35

23
27. Lindstrom MS. NPM1/B23: A Multifunctional Chaperone in Ribosome Biogenesis 1 and Chromatin Remodeling. Biochem Res Int 2011; 2011:195209. 2
28. Yung BY. Oncogenic role of nucleophosmin/B23. Chang Gung Med J 2007; 30:285-3 93. 4
29. Yu Y, Maggi LB, Jr., Brady SN, Apicelli AJ, Dai MS, Lu H, Weber JD. 5 Nucleophosmin is essential for ribosomal protein L5 nuclear export. Mol Cell Biol 2006; 6 26:3798-809. 7
30. Savkur RS, Olson MO. Preferential cleavage in pre-ribosomal RNA byprotein B23 8 endoribonuclease. Nucleic Acids Res 1998; 26:4508-15. 9
31. Szebeni A, Herrera JE, Olson MO. Interaction of nucleolar protein B23 with peptides 10 related to nuclear localization signals. Biochemistry 1995; 34:8037-42. 11
32. Itahana K, Bhat KP, Jin A, Itahana Y, Hawke D, Kobayashi R, Zhang Y. Tumor 12 suppressor ARF degrades B23, a nucleolar protein involved in ribosome biogenesis and cell 13 proliferation. Mol Cell 2003; 12:1151-64. 14
33. Maggi LB, Jr., Kuchenruether M, Dadey DY, Schwope RM, Grisendi S, Townsend 15 RR, Pandolfi PP, Weber JD. Nucleophosmin serves as a rate-limiting nuclear export 16 chaperone for the Mammalian ribosome. Mol Cell Biol 2008; 28:7050-65. 17
34. Iadevaia V, Zhang Z, Jan E, Proud CG. mTOR signaling regulates the processing of 18 pre-rRNA in human cells. Nucleic Acids Res 2012; 40:2527-39. 19
35. Cammas A, Sanchez BJ, Lian XJ, Dormoy-Raclet V, van der Giessen K, Lopez de 20 Silanes I, Ma J, Wilusz C, Richardson J, Gorospe M, et al. Destabilization of nucleophosmin 21 mRNA by the HuR/KSRP complex is required for muscle fibre formation. Nat Commun 22 2014; 5:4190. 23
36. Cairns P, Okami K, Halachmi S, Halachmi N, Esteller M, Herman JG, Jen J, Isaacs 24 WB, Bova GS, Sidransky D. Frequent inactivation of PTEN/MMAC1 in primary prostate 25 cancer. Cancer Res 1997; 57:4997-5000. 26
37. Majumder PK, Yeh JJ, George DJ, Febbo PG, Kum J, Xue Q, Bikoff R, Ma H, 27 Kantoff PW, Golub TR, et al. Prostate intraepithelial neoplasia induced by prostate restricted 28 Akt activation: the MPAKT model. Proc Natl Acad Sci U S A 2003; 100:7841-6. 29
38. Kremer CL, Klein RR, Mendelson J, Browne W, Samadzedeh LK, Vanpatten K, 30 Highstrom L, Pestano GA, Nagle RB. Expression of mTOR signaling pathway markers in 31 prostate cancer progression. Prostate 2006; 66:1203-12. 32
39. Subong EN, Shue MJ, Epstein JI, Briggman JV, Chan PK, Partin AW. Monoclonal 33 antibody to prostate cancer nuclear matrix protein (PRO:4-216) recognizes 34 nucleophosmin/B23. Prostate 1999; 39:298-304. 35

24
40. Berthon A, Sahut-Barnola I, Lambert-Langlais S, de Joussineau C, Damon-1 Soubeyrand C, Louiset E, Taketo MM, Tissier F, Bertherat J, Lefrancois-Martinez AM, et al. 2 Constitutive beta-catenin activation induces adrenal hyperplasia and promotes adrenal cancer 3 development. Hum Mol Genet; 19:1561-76. 4
41. Herrera JE, Savkur R, Olson MO. The ribonuclease activity of nucleolar protein B23. 5 Nucleic Acids Res 1995; 23:3974-9. 6
7
8

�

�

�
����
Discussion et �Perspectives�
� �

�

� ��������������� �����
<=�
�
�
4)�#&���#��''�)���(� ���'�&��'�*� +$(�����)��&���#&'�$(�� ��#(� ����)��#��,���-.��)&�
)('�/�'���� ��#)� �)� ��#��')� ����'�)&�&�� �(���#� �&�$(+� �� %�(�� �� '� �� ��� ��0��&�)�&�('�
&'�#)�'��&��##� ��(� ��'���&�('���)��#�'�"+#�)�,��&��#"��&�� <*�=11D�.<�'�#)������#&�/&�*�
��#� &'�)�� � ��� &�+)�� �� ��#)�)&� � �.� ����'�#�'�� �&� ��'��&'�)�'� � �)� ���#�)��)� $(��
��#�(�)�#&� � �� )('�/�'�))��#� ��� ���-� �')� ��� �� &(��'�"�#+)�� �'�)&�&�$(�*� �&� ��.�
��'��&'�)�'� �)��**�&)���#�&2��$(�)��&��� �( ��'�)��+(#��)('�/�'�))��#�������-�����<<�
'+(#����'&*�#�)���##�)���#&'�#&�$(�� +�/�'�))��#����'(��������-���#)� ��#&�/&���&�#030�
�)&��4� � ��)&��( �&��#���� +�/�'�))��#��(�"+#����#)��$(+ � ��)&��� �)�&��#����)�#�43�����'�
�23<� '+�(&'����'&*�#�()��)�#)� ��#&'�$(+(#�� )('�/�'�))��#� ��� ���-� ��#)� +��&� �(��
�'�)&�&�$(�� � ���� �+#�� � (#�� �("��#&�&��#� ��� �� �'� �*'�&��#� �&� � +����'�&��#� �+(#��
�2��'� �)��*��))���)� ���)��'+" ���#&)���� +�/�'�))��#�����'�&�#�)�'"( �&'���)��(��2� ��
�� ( ��'�<����#�$(�����&'�)�� �����'&����)���##�)��'�"�#� �)�$(�#&��(�'� ��������-���#)� ��
��2)�� �"��� �'�)&�&�$(�� � ���*� � � )�( +)�� ��� #���'�()�)� $(�)&��#)� ��#��'#�#&� �)�
���#�)��)��� �( ��'�)��'��)� ��� �$()���#)� �)���#��+#�)���2)����&�� �"�$(�)� �$(��
#�()��)�#)���)�'))<�
�
�
���!����"�#�� ��$������$�%�$����&��$���"�'��!(�)&�")��*��
�
�23��)&�(#��)'�#�0&�'�#�#����#�)������'&�#�#&� � ��*��� ����)��677�,������������� �
������ ������� 7�#�)�.<� . �� ����'&��#&� � =� ���� �/�)*� �23--� �&� �23-=*� ��#&� ��
�����)�&��#*� ����������'"( �&��#���#)��$(�� �)���� �)��� ( ��'�)���**+'�#&�,��#"(�&���&�� <*�
=1-1.<� 4(� �� � ��� )�#� '� �� ��#)� �� �2&�� �)��� ��#��'#�#&� �� ��#&'� �� ��� �� )2#&�+)��
�'�&�$(�*� �&� ��)� �&��� �)��)� " (����$(�)� �&� �����$(�)*� �23� �/�'��� �())�� ��)� ��&��#)�
#(� ��'�)� ��#&� �� ��#&'� �� ��� +�/�'�))��#� "#�$(�,��('� '�)(��)��'� ��� �#&���#������&�#�*�
=1->.<� -�� ��#&'� ����(&� )+�/�'��'� ��� ��#�+'�� �#��'��&�*� )�#)���'&�����&��#� ��� �23� �(#�
���� �/�� ��'���&�#��#<� 4�#)�*� �����)���'2 �&��#� ��� �&�&>� )('� �� )'�#�� B=B���'� �23--�
��'��&�)��&'�#) ���&��#�#(� ��'���&�)�#���&�)�&��#�������*��&�('����&'�#)�'��&��#�,7����&�
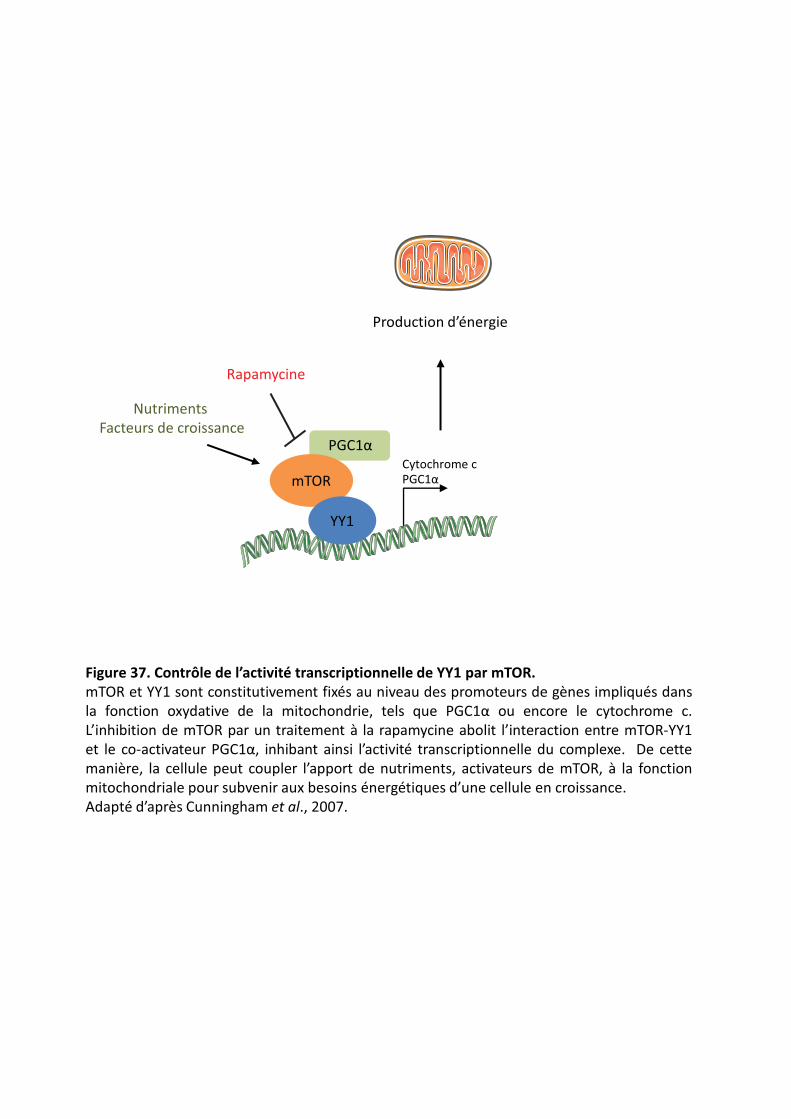
!����
(<?*α�
����!� �+��
��*�
?��� ���!�� �(<?*α�
Figure 37. Contrôle de l’activité ������������ �������������������!���������*���+�� �+����"����!�+���������"�+����"��������!���"��������+����!�#�$"�����+��#�� ��+ ���+� ���������� ��� #�� !��� ��+����&� ��#�� $"�� (<?*α� �"� �+ ���� #�� ��� ���!�� '�L’inhibition����!��������"+�������!�+���� #������!� �+��� �#��� l’interaction��+����!���(��*���� #�� �(� ������"��(<?*α&� �+�� �+����+��� l’activité� ���+� ������++�##���"� �!�#���'� �$�� �����!�+����&� #�� �##"#����"�� �"�#��� l’apport����+"���!�+��&�� ������"������ !���&��� #�� ��+ ���+�!��� ��+����#����"���" ��+����"�� ����+���+������$"���d’une� �##"#���+� ������+ �'�Adapté d’après Cunningham �����'&�8,,9'��
�"���!�+����%� ��"������ ������+ ��
Production d’énergie�

� ��������������� �����
<;�
�
��5:�@AA£-��)�)�����*.���5:�@AAAB5�����-C�%*/.�����*�*&.��)&.+'�*+��6*7%+*,,�)&� *�!��.*/+,�
*� .+�&,�+�%.�)&�*&� ")/�&.� ,/+� ��� .+� /�.�)&� *� �*/+,� 4���5� -6*,.� &).���*&.� �*� ��,� %)/+�
��4�8� *.� �#C;� 9�*� ���(/*+� *.� ��5:� @AAD-� ':;*�� *.� ��5:� @ACAB5� �*� �)&.+'�*� *� ����� ,/+�
�6*7%+*,,�)&� *,���&*,�%*/.�����*�*&.�,6*7*+�*+� *�!�$)&� �+*�.*5�����%��(/*�,���)����,�.�)&�
�� ��� � +)��.�&*� ,/+� *,� %+)�).*/+,� *���&*,� �%*& �&.,� *� �64��� %)�0��+�,*� :� �)/� �
9-/&&�&� ��� *.� ��5:� @AAD-� �,�&�� *.� ��5:� @ACAB5� &*� �./ *� +��*&.*� �� �)&.+�:� %�+� /&*�
�%%+)� *� 6���/&)%+���%�.�.�)&� *�� +)��.�&*��)/%��*���/&�,�(/*&$��*� �/.� ���.�9- �<
,*(B:�/&� *&+�� �,,*�*&.� *������ �/� &�;*�/� *� ��� ,�(/*&�*� %+)�).+��*� *����C:� �� E@F�
%��+*,� *���,*,�*&���)&.� /�,�.*� 6�&�.��.�)&� *� ��� .+�&,�+�%.�)&� 9- �;*+)/7�*.���5:�@ACG�B5�
�)/,��;)&,��)&!�+����*,� )&&�*,�)�.*&/*,���%�+.�+� *�!)�*,� *�,)/+�,� �&,�&).+*��) ��*� *�
!��+)���,.*,� *��+0)&&��+*,��/+�&,� *.� ��)&.+�� (/*� ��� ,��&���,�.�)&� �G�=4�.:� ��������:�
�)&.+'�*��6*7%+*,,�)&� *����C5�-*,� )&&�*,�%���*&.����&.*&�&.����C��)��*�/&�%).*&.�*��
*!!*�.*/+� *&� �;��� *� ��� ;)�*� *.� ,)/��;*&.� %�/,�*/+,� (/*,.�)&,� ,/+� �*,� ����&�,�*,�
�)���/���+*,���%��(/�,5�
�
���������� ���� ���� ����������������������� �����+������
�),� )&&�*,� ��)&.+*&.�����+*�*&.�(/*��6*7%+*,,�)&� *����C�*,.� �%*& �&.*� *��6��.�;�.��
*�����5� �*,�*7%�+�*&�*,��*&�*,��;*�� �*� %+)�).*/+� *����C� ��)&��*&���)&.� /���&*�
�) �&.� %)/+� ��� �/��!�+�,*� ��&,�� (/*� &),� +�,/�.�.,� *� - �� ,/���+*&.� !)+.*�*&.�/&� +'�*� *�
������/� &�;*�/�� +)��.�&�*&:� �/�� %*+�*..�&.���&,�� *��)&.+'�*+� �6*7%+*,,�)&� *����C� *�
��&��+*� �+*�.*5� &�.+��.*�*&.������+�%��0��&*�+� /�.�� *�!�$)&���%)+.�&.*��6*7%+*,,�)&� *�
���C����,� ��� &6��.�+*� �*%*& �&.� %�,� ��� ��%���.�� *� ����,)&� *�������/� %+)�).*/+5� ��*,.�
)&��%),,���*�(/*������,)�.�!�7�� *���&��+*��)&,.�./.�;*������� +)��.�&*�*.�(/*�,)&���.�;�.��
��&�,*� %*+�*..*� *� �) /�*+� �*� +*�+/.*�*&.� *.=)/� �6��.�;�.�� *� �)<��.�;�.*/+,�
.+�&,�+�%.�)&&*�,5�-6*,.��*���,�,/+��*,�%+)�).*/+,� *���&*,�+��/��&.��*,�!)&�.�)&,�)70 �.�;*,�
��.)� )& +���*,:� .*�,�(/*� �*��0.)� +)�*�-�)/���-C<;� 9-/&&�&� ���*.���5:�@AADB5� �'�&,��*�
�) ��*:������ ,*� !�7*� �/� !��.*/+� *� .+�&,�+�%.�)&� ��C� �/� &�;*�/� *� ��� � +)��.�&*� *.� /&�
.+��.*�*&.��� ��� +�%��0��&*���)(/*� �*� +*�+/.*�*&.� /��)<��.�;�.*/+���-C<;:� �& ���&.���&,��
�6��.�;�.�� .+�&,�+�%.�)&&*��*� /� �)�%�*7*� 9�������B5� �6�./ *� /� %+)�).*/+� *����C�� *>�
�6 )��*�+�;��*����%+�,*&�*� 6/&�,�.*� *�!�7�.�)&�%)/+���C�*&.+*�GDC�*.�GEE�%��+*,� *���,*,�

�

� ��������������� �����
<A�
�
����������� 3����13�3�3��3������������ ��3��3��2�4��������-3�<==B4-��������3����3����� ����3�
��� 3������03 ��3���������� �����(������ 3�3������������.���2567843����3��� ������� 3 ���
���� ��� �����3 ��� 4�(������� �1� ��� 3�� ��� ��"<� 3�� 3�3��3��� �� ����3� ��(�� ����
����3�44�������������������%��4�����-������� 3�����4:�5 �������3 �������4�(��� ������-����
79</� �1�� �� � ����3 � �13 ���� ��� ������� � �� �� � � ���� ��� 03 ��3�� ��� ����� ��� ���
�4����3��-�:��� �������� � �����������35�3���� ��03 ���� ����14�����3 �����$�������0�������
������� ��3��3���������<-�����(��303���������4%��4� �3��� ��� ���� � ��������3 ���
�����4:���3�34����������������� ��(3���3�3���� ��������3��������<����� 3����0�������4��3�
�03�� ��� (3�� 3� ����� ������� ��� ������������ ��� ����� ��� �������� ��� ��"<-� :�� ���3��
�4��������3����� ��������3�3������1� ��� 3�����������03�����������3���� 3�����35�3�3�0����
��� ��� �����������������<� ��� ������������� ���� �-���03�3� �� � ��� � ���� ����3 ���
���� �� ��� 3����5����3�3���3�� ������3����� ��� �4����3��� 2�4:�5���4:�4� �03�� ���
��03����������� ������1�������� ������5��<� �������������������"<-���������3��� �
����3�3���� � ��� � ��� ����3��� ���� � �� � 0��4���� ��14��� � �3� � �� ��<-���� 5�3� �����
����3�3�� � ���� ������ 03 ��� �1�3����1������3��� ��3�34�� ������ ����-� ��� ��� �������� ���
�������������3��������"<���� � �1������ 03�������������������03����������4%��4� �-�
�� � ����3� � ���� ��� �3��� ����3 ��3� �� ����3�� �1� ���3� � ������3�� 3� ��� �
3��������3�3���3� 3� �1�����3�34��� ������ �������� �1������ ��������<-� �13����303���3�����
�������3�� � ����� � �� ���� � ����3�� 3� ���� ���� �����4�� ��� ���������3����� �� �3�
�������� ���� �34��������������0����������� ��� �������������� ����4����3���� ���������
��������3�43 3�3���������-��
:������������4��������3��3 ��� � ������������3������� �3��������������� �3������<���:��
�����73� ������ �� ��� ������� �3(�����4����3�3��-���������3� �� � �� ����� ���� � ���
����3��� ���� � �1���3(3������ �������������"<� �� �1�3��� �1��� 4���� ���������� ���30��� ��
���� ��(3���3�3���� �1� ��� 3�����������2�����<4����3����2�����74����� 3����0�������
4��-� �� � ����� � ������� ����� �������� � ��� ����% ���� ��� ������������ ��� ����� �� ���
�4����3������ ��� ���� ����3�3� -� :���1� �������������� �� ������������� 3�� 03 ���� ���
�4����3��� ��� �3�� �1��� ����� �� ������ ���� �����<� �� �����7-� ��� � ��� �� 5��3� �� �
����� ������� ����� ���������� ���������� ���������3������1����% �� ���������3������ �
3���������� � ������3�� � ��� ����� �����3���� ��� � �� � �3���� �� 3 ���� �� � ����3��� �
�����3�� -�

��"0�
#41#�)�
�������)�
β1���3���
��.�1>1�
4� ��
������������������������������������������������������l’expression����������'��� 03������������$��%����3���������%�3����%*�4��� +��2��%� 3�*��3����#�%����.�� +��.�1>12������������3����#�������;4��*������l’éthanol�+*�43�%��2��%��*���033��"�������3��?���������%�����������%3������� �%�����#%3�� ����4������ ����#����3�������������6���3���#�%�����������%����#���!��1�-��+42����#���&������1�����+�2?�L’expression������"0�+42��������$��3����#�����##����,�l’expression�������β1���3��?�
3
345
0
045
6
645
7
�4 ���3� �4 ���3�
�� ��.�1>1
��"
0�$��
4������3*����
*���
���3�� 1� 8�

� ��������������� �����
FF�
�
���� ��� ���� �������� ! ����� ������ ���� �� ��� �� ��� ���� ���������"�!�
��� +�%�"0��'*� ���.� *'� ,*� ��7�'.� ,/+� �,�""/'#% ���'*� �'.+��*��/���+*� #���<@9� �*� �#"%�*7*�
#���<@(+�%�"0��'*� �'.*+���.� *',/�.*� �;*�� "���� �+�����,�'.� ,#'� �'.*+��.�#'� �;*�� �*,�
%+#.��'*,� /� �#"%�*7*�"���-<-� #'.���%.#+9� -*��� �#' /�.���#+,��� ��� %*+.*� ,�'.��+�.�� *�
"���-<� *.� #'�� �� �,�' ���.�#'� *� ,#'� ��.�;�.�� ��'�,*� .��%� *.� ��9-� @=<=:9� -*%*' �'.-� /'�
.+��.*"*'.����:��� ���+�%�"0��'*�',��.�+*�%�,� �,��.�;�.����.��0.�(/*� /��#"%�*7*�"���-@9����
+�%�"0��'*�',���.� #'��%�,� *���$#'� �+*�.*�,/+��*�,�.*���.��0.�(/*� *�"���-�*.����*,.�%#,,���*�
(/,�/�,*�'� *�,#'��#"%�*7*�� +#"�.�'�*'-��,��.�;�.����'�,*� *�"����,#�.�"��'.*'/*9�4��'� *�
+�%#' +*����*..*�(/*,.�#'-�'#/,��;#',�.+��.��'#,��*��/�*,��;*�� *� ����#+�'�<-�/'*�"#���/�*�
(/��,*���7*� �',��*�,�.*���.��0.�(/*� *�"����*"%$� �'.���',�������7�.�#'� *��,4���*.��' ���'.�
#'��,#'���.�;�.����'�,*�.� #+**'�*.���9-�@==?:9��#,�%+*"��+*,�*7%�+�*'�*,�"#'.+*'.�%*/�#/�
%�,� ,*��*.� *� ��� �#+�'� <� ,/+� �,*7%+*,,�#'� *����<-� ,/���+�'.� #'�� (/*� ��� +��/��.�#'� *�
�,*7%+*,,�#'� *� ���<� '*� +*(/�*+.� %�,� �,��.�;�.�� *'>0"�.�(/*� *� "���� .����� #$:9� -*,�
#''�*,� %+���"�'��+*,�,#'.��*%*' �'.��� %+*' +*��;*��%+���/.�#'��.�'.� #''�� ��� �����/�.��
,$.+*��,,/+�� *� ���,%�������.�� ,��.�#'� *��*.� �' ���.*/+�% �+"��#�#��(/*9��#/+��#'.#/+'*+�
�*..*� �����/�.�-�'#/,�%+#%#,#',� *�.+�',�*�.*+�/'*��#+"*� #"�'�'.*('���.�;*� *�"���-�(/��
�/+��.� %*+ /� ,*/�*"*'.� ,#'� ��.�;�.�� ��'�,*-� �',� '#.+*� "# ��*� �*��/���+*9� -*..*� �#+"*�
*'>0"�.�(/*"*'.��'��.�;*� *�"����."�����'-�����������:��+��*������"/.�.�#'��/�,*�'� /�
,�.*���.��0.�(/*� *� �,�,%�+���'*�*'�%#,�.�#'�@@A@�.�4("����4,'@@A@�0,:��� �����.�� ��+�.*�
*.� *,.� #'�� �,%#'���*� .�#'�� *.� ��9-� @==B:-� *.� �,�'��0,*� *� ,#'� �"%��.� ,/+� �,*7%+*,,�#'� *�
���<� %#/++��.� %*+"*..+*� ��#+,� *� �#'��+"*+� #/� '#'� �*..*� 0%#. �,*9� �#/,� %+#%#,#',�
����*"*'.� *��#(.+�',�*�.*+��*�%��,"� *��# �'.�%#/+�"�����'��;*��'#.+*�%+#"#.*/+����<�
��#'�� *'� �"#'.� /� ��'*� *� ��� �/����+�,*� %#/+�"*,/+*+� �,��.�;�.�� /� %+#"#.*/+� �',� �*,�
�#' �.�#',9� ��� �,��.�;�.�� ��.��0.�(/*� *� "���� *,.� '#'(*,,*'.�*��*� �� ��� +��/��.�#'� *�
�,*7%+*,,�#'� *����<-� ���*,.���#+,� %+#����*�(/*�"����,*+;*�,*/�*"*'.� *�%��.*(�#+"*� *�
+*�+/.*"*'.�%#/+��*,� ����+*'.,��#(��.�;�.*/+,�.+�',�+�%.�#''*�,�'��*,,��+*,����,��.�;�.�#'� *�
���.+�',�+�%.�#'9�'�',��*���,-����,*+��.��"%#+.�'.��%+�,��;#�+�� *'.������*,���.*/+,�%+�,*'.,-� *�
��+��.�+�,*+��*,� #"��'*,� ,�'.*+��.�#',� *�"�����;*���*,�*��*�.*/+,9�-*..*� *+'��+*�%�+.�*�
,*+��+����,�*�*'�/.���,�'.��#""*��%%�.�/'*�%+#.��'*�+*�#"��'�'.*�����.�� � ������������
�����������:��/,�#''�*��/7� ����+*'.,� #"��'*,� *�"����(/*��,#'��'�/�*+���;*�� *,�*7.+��.,�
%+#.��(/*,�'/�����+*,9� �,/.���,�.�#'� *�����*,� *���/.�. �#'(���+#,*� %*+"*..+�� *� %+���%�.*+�

4�.� 3’UTR�;’UTR� 444444444�
�/��
��-�
��=� 4���
%>D�
��-� -'�-� -��=� %>D�
���� ���-� 4 �-� �����
��=4�
���-�
-'�-�
4�.� 3’UTR�;’UTR� 444444444����-�
����� �4���.���-;�
�/��
&�
��
����������������� �����&������� ����������&�������%&�+��&<�-&�0>%&�'&�'!�)�'�0�"��'&*�4����'!%&�'�'&����!�1�0�"��'&�%)"0!��&*�'!*0�����*�0)��&*�22&)04�"/�*0�����*�0)��&*�22�"�&04�*/)��&*�*!#/&��&*�4�.�2�������������4�'��*����)!��"��>’UTR�'&*�4���<��&*�%)"0!��&*� '!*0�����*�0)��&*� �"��&� ����� )&�)/0&�0� �� �&/)� 0"/)� '&*� '!�'!�>��*&*� &0� '&*�&1")��"�/��!�*&*�#/��%&)�&00&�0� ���'��&*0�"��'&� l’ARNm�����!<����!�1�0�"��'&��/��&*0�*0�����*�0)��&�puisqu’elle� &�%�� &� ��� !�1�0�"�� &0� l’activité� '&*� %)"0!��&*� '!*0�����*�0)��&*<� �"/0&*� �&*� %)"0!��&*�*"�0� )!�/�!&*� %�)� l’action� '&� %)"0!��&*� ����*&*� &03"/� % "*% �0�*&*� '!%&�'��0&*� '&*� 2"�&*� '&�*������*�0�"��'&*�!��0&/)*�'&��)"�**���&�"/�'/��>��&��&��/���)&*<����'��*��&���*�'&�l’ARNm�'&����-3��/��&0������!")�&�0�/���"�%�&1&�'!*0�����*�0&/)�#/��)&�)/0&� �&*�)��"�/��!�*&*� ��4���&0�.���-;<����'!�)�'�0�"��'&� l’ARNm�'&����-�%�)��&��!����*�&�&*0�&**&�0�&��&��/��"��'!)"/�&�&�0�'&� ����>"�&�4*&<�4'�%0!�d’après��� "&��&)��+���#/�03�=1-=<��

� ��������������� �����
F<�
�
�*,�$(%-�*.*,�!(/%�,�-?�,� *�5�/�!�*/�-�/�0*,=*/ 6��(=����-/�,* $*� *,�*!!*$=*?/,)�-*��� (?,�
-*/%*==/�� *� �=*/%� */�1?*�>,@� (%�� *>,@� 2� =*/�$=�( �,( =�/*1?�,�-(?/� ���!(/%�=�( � ?�
$(%-�*.*�$ /(%�=� �* �-�/�%���)�4��( ��=*/%*-�$*,� ( �*,�-(?//��* =�(?5/�/� *� (?5*��*,�
-*/,-*$=�5*,� = �/�-*?=�1?*,� * � -*/%*==� =� *� *,,� */� *,� %(��$?�*,� � ��� =� �*�
/*$/?=*%* =� *�-�/=* ��/*,� *�%������ ���$ /(%�=� *�-(?/�,=(--*/� �2*.-/*,,�( � *����A-�
�5�=� =��� ,���*,� (%�/*?.�*!!*=,�,*$( ��/*,�� �,�/���*,� �$�* $ �,�-�/�? �=/��=*%* =������
/�-�%7$� *�>- � -�BCCD8���� (=�*=���)-�BCCC@)�
�
'���� �������'� ��$��$�� ���$� ���$,&�����'�� ��'�������� �
���/�-�%7$� *�* =/�� *�? *�$ ?=*� *�CCD� ?�,=($�� 24��%�$( � =�-(?/����A� �,�D� *?/*,�
*�=/��=*%* =)�'*�-�?,-��2� ���=�( � *����=/� ,$/�-=�( �-* � =�AB ����2�� *� 2? �=/��=*%* =����
�2�$=� (%7$� *�'� 2�!!*$=*�-�,����1?� =�=�� 24��%�$( � =�-(?/����A-�%( =/� =�1?*�$*�?�6$��
*,=� =/�,� ,=���*� � ,� (=/*� %( ��*)� -*,� ( �*,� ,?���/* =� ( $� 1?*-� �?� *��� 2? *�
/��?��=�( � *��2*.-/*,,�( -����/�-�%7$� *��(?*�,?/����,=�����=�� *,�4��%����A�* �* =/�� � =�
�*?/� ��/� �=�( )���*.�,=*� *?.�=7-*,� *� ��/� �=�( � *,�4��%�3����-/*%��/*�$( ,�,=*�* �? �
$( =/*�*61?���=�� *,� 4��%�-(?/��5�=*/� ���-/( ?$=�( � *�-/(=�� *,�$7=(=(.�1?*,� =/( 1?�*,�
*=9(?� ( =����!( $=�( �,*/��=�-*/ ?*�(?�$ � ��*�>��'-�� ��� �������������������@8����
,*$( *� 5�,*� �� �%� ?*/� ��� *%�65�*� *,� 4��%� �!� � *� /��?�*/� ��� 1?� =�=�� *� -/(=�� *,�
!( $=�( *��*,� -/( ?�=*,� >-(?/� /*5?*-� 5(�/� �$ (* �*/�� � � ��1?�=-� BCAB@)� -*��*6$�� *,=�
,(?5* =� �-* � =*� *����-/�,* $*� *�,�1?* $*,�/�$ *,�* �4��>4�.-�������� �����@� � ,�
�*,�/���( ,� ( 6=/� ?�=*,� *��24��%�>���-�������� ���������@�>�������"&�@)��-*,�,�1?* $*,�
-*/%*==* =� *� /*$/?=*/� *,� -/(=�� *,� =*��*,� 1?*� ���� >���������� �@-� ���A� >���������
�����������������@-������ >����� ���� �������� ������ �����@�(?�* $(/*� 4��A� >��� ����
� ������!������������A@�1?���� �*?/� =(?/� /*$/?=* =� *,�-/(=�� *,����$=�5�=�� /��( ?$���,*,�
-(?/� ��/� */��24��%)����-/(=�� *��?��>�������������@�*,=�����*%* =�$( ?*�-(?/�!�.*/�
$*,� ,�1?* $*,� %��,� ,( � *!!*=� ,?/� �24��%� *,=� �� �/��*%* =� ,=�����,�=*?/)� ��� !�.�=�( � *�
�2* ,*%��*� *�$*,�-/(=�� *,�*,=�/��?��*�-�/��*?/� *�/�� *�- (,- (/7��=�( -�!/?�=� *��2�$=�5�=��
*�-/(=�� *,� �� �,*,� *=�- (,- �=�,*,� �-* � =*,� (=�%%* =� *,� 5(�*,� *� ,�� ���,�=�( �
�-* � =*,� *,� !�$=*?/,� *� $/(�,,� $*)� -*�%�$� �,%*� ,(- �,=�1?�� -*/%*=� *� $( =/*�*/�
-�?,� !� *%* =� ��� -/( ?$=�( � *� -/(=�� *,� * � !( $=�( � *� �2* 5�/( *%* =� $*��?���/*)�� *�

!����
��
�����
4��
� �����������4�������4���
4���������
Figure 40. Modèle de stabilisation de l’ARNm de NPM1 par ����������� ��� ������������� ������������ �� ��������� ��� � � �������� ����� l’expression� d’un�! ���� �� ���� �� "������ � ���#� � � �� ��� l’ARNm��������$� % ��� ���� � � � ������ &��� ������� l’ARNm��������� �����#� ��� ����� '� ��� � ������ '� � � � � �(������ ���� � ���l’hypothèse�������! ����� �����4���� �� � �������4� �������4�$�) ������ � �� &��l’ARNm�������������� ��'�� ���#� � � ��� ������� �������� ����� ���$�L’ �� �(����������"� �� �� l’activité�����4���� �(��� ��������� ��*�" �� ����� ����+������ ��#��������� �����l’effet����� �� � �(���$�
� � �

� ��������������� �����
FB�
�
&+:4�� ,&'�,+�� �� (�,%�-� 4-�4�,+����,� :,� %&'�,�-%�� (�,� ��.:��� �-4� %� 4�� ��.� �-+�
4&-+�"���-&�(�,� �-�'+�/,�4:�'/%(��0��5:,1�� ���+�4�� ��� ,�"/,:'�&�-���4� � 2-�%%�-��+���34�
56.78�2��������83�-��%&'�,�-%��,�.:��,+� ���(,&-�,'��4-:,��-&.:�,'��,�' ���,�:,�'����2�9
,�' 8�4�,-� ��� ,&��/,�:-�,/,� +,�4:�+��4�� ��.� 2:-���4���"���� �����������84�(�,%�++�,+� ���
��0�+�/,�4:�'/%(��0��5:,1�� �3��/:,�+�-+�,��-�%(/,+�,'��4��'�++��,&��/,�4�,-�,/+,��%/4���4�
,/:-��;/,-�+,�,-��'+&�:,�(��-%�4��'/,+�,�,+��-4' '�4��������� �������������'/:(�&��;�'����
,&��/,� :-���� 4�� ��.� %:+&�� /:� ,/,� (/:,� '�++�� -&.:�,'�� �9,�' � 4�,-� ,/-��.�3� /:-�
,-�;/,-�(�-�/"-�,;&�4��%/4���'�+�/,�4���-�'+�;�+&��:'��&,�-���+�4�����.:�,+�+&�4-4� %�'/4�,+�
(/:,� ������� ������������� -:�+���� :,� +,��+�%�,+��� ��� ,�(�%<'�,�3� -�� ,&-:�+�+� -:���,��4/,'�
.:�����,&�:��+�/,�4���-�0(,�--�/,�4�� ��.�(�,�%����,��4&(�,4�(�-�4:�%&'�,�-%��,�((/,+&�
(�,�-�%�-����� �3��
'��%�,��,���,+&,�--�,+�4��-����+�4�����,�(�%<'�,���-+�,�,;�,-&�(�,�:,�+,��+�%�,+�'/,�/�,+���
�-�'+�,/%<'�,��'3�-�� ,&-:�+�+�-:���,��4/,'�.:�� ���4&�,�4�+�/,��'+�;��4�� �-4� %�4�� ��.�
�,4:�+��(�,� :,� +,��+�%�,+��� ��� ,�(�%<'�,���-+� +,�":+��,��4�� ��� +,�,-',�(+�/,��&,�.:�3� /:-�
(,/(/-/,-�4/,'�:,�%/4����4�,-���.:���%�����-+��%(��.:&4�4��%�,��,��4�,�'+��/:��,4�,�'+�4�
4�,-� ���,&(,�--�/,�4-:,���,��4/,+� ���(,/4:�+�2��'+�:,��8��-+�,&'�--��,���� ���4&�,�4�+�/,�4��
�-4� %� 4�� ��.� 2������� ��83� �,� +,��+�%�,+� �� ��� ,�(�%<'�,�4� ;��� �-�, �"�+�/,� 4�� %���4�
��;�,���'�++��,&(,�--�/,��+�(�,%�++,��+���,-���-�'':%:��+�/,�4��'����'+�:,����+����4&�,�4�+�/,�
4�� �-4� %3� �,� +,��+�%�,+� '/,�/�,+� �;�'� �-�'+�,/%<'�,�� '� %�-.:�,��+� ��/,-� �-����+� 4�� ���
,�(�%<'�,�� �,� �,+�,4�-�,+� +/:+� +,�,-',�(+�/,3� �,� %&'�,�-%�� 4�� +,�,-',�(+�/,9+,�4:'+�/,�
(/:,� ��� (,/4:'+�/,� 4�� '�� ��'+�:,� �� (�,��+� (�:� (,/"�"��� '/%(+�� +�,:� 4�� ��� ,�(�4�+&� 4��
4&�,�4�+�/,�4���-4� %� ��.�(�,����,�(�%<'�,�3� /:-�(�,-/,-�4/,'�.:��'����'+�:,��-+�4��
,�+:,��,�"/,:'�&�.:�3�����-+�'/,,:�4�(:�-��/,�+�%(-�.:��4�-�(�+�+-�4� �,/,9'/4�,+-�2-�9�/:�
%�� 44� #�� � ������������� �+� ��������8� -/,+� �%(��.:&-� 4�,-� ��� 4&�,�4�+�/,� 4�-� 4� %�
25/:-���<� �,4� �/���,;�<4�566=83���:-� ,&'�%%�,+4� ��� ��&+&�%/,+,&�.:�� ��-� �/,�-� 4� �,/,9
'/4�,+-� 2��,'� 44� ����� ����������� ���� ������� � 48� -/,+� &����%�,+� �%(��.:&-� 4�,-� ���
4&�,�4�+�/,�4�-�4� %��,��:�4�,+�4�-�'/%(��0�-�,�"/,:'�&/(,/+&�.:�-�����'+�:,-�-:,���:,-�
'�"��-� 2�/,�� �,4���.:�+4�56..83� 4��,�4-�4�,+����,�4�-� '�,4�4�+-�,/:-�(,/(/-/,-�(�:-��:,-�
�((,/' �-3����(�:-� -�%(���'/,-�-+���� ,�' �,' �,� ��������4-&;�,+:��-�'�,4�4�+-��,� :+���-�,+�
4�-�/:+��-�4��(,&4�'+�/,�"�/�,�/,%�+�.:��4�-(/,�"��-�-:,��,+�,,�+4�+��-�.:������,4��2��+����+�
��34� 566>83� 4��,� 4�� ;���4�,� ��� (�,+�,�,'�� "�/�/��.:�� 4�-� '�,4�4�+-� -&��'+�/,,&-4� ,/:-�
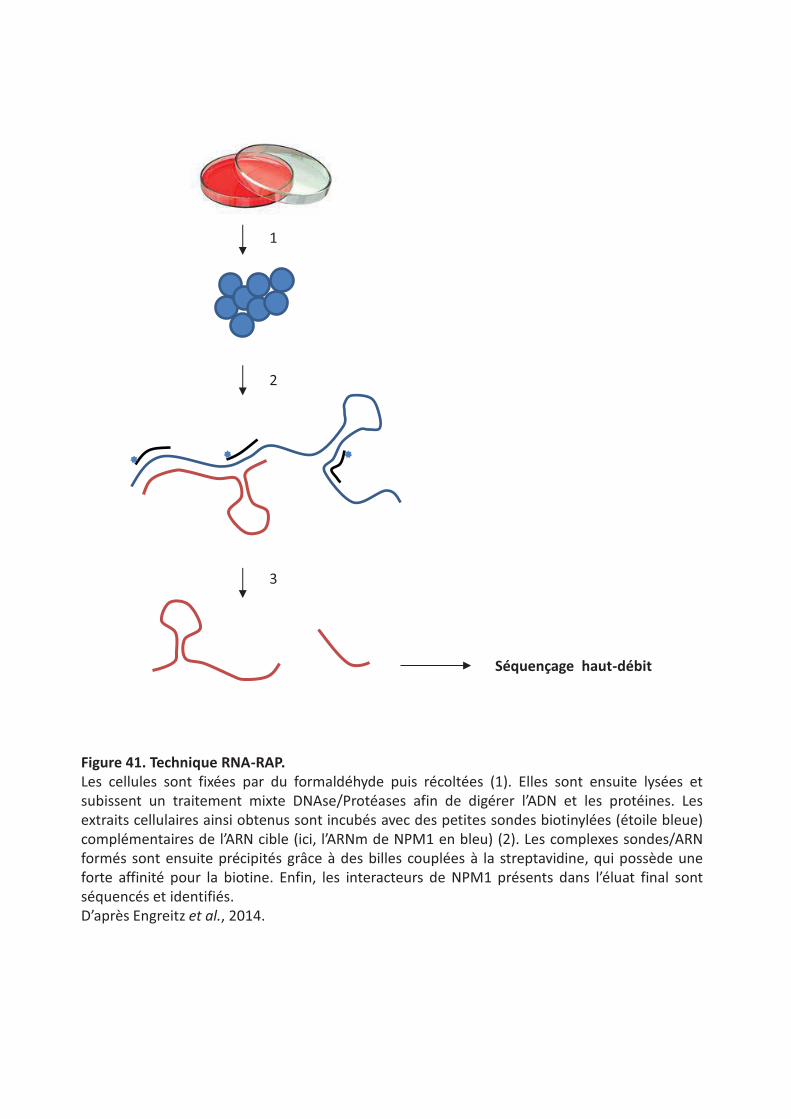
��
?�
5�
� ���������������� ����
����������������������&��&������� ����#���� �/$%� ������� ��&� �#� �/& ��������� �#��� &��/�%���� )�*'� %����� �/$%� �$�#�%�� ������� �%��#�����$%� #$� %&��%� �$%� ��%�� ��4��+(&/%������ ���$� ��� �� �&�&� l’ADN� �%� ���� �&/%��$��'� ������%&��%������#���&�����$���/�%�$#���/$%��$�#���������������%�%����/$������/%�$������)�%/�������#�*��/ ��� �$%��&������ l’ARN�������)���,� l’ARNm�����(����$����#*�)?*'������/ ��������/$���+4����/& ����/$%��$�#�%���&�����%��� &������������������/#�������� ����%&��%�����$�,��#���/������#$���/&%�� ����$�%���/#&� �����/%�$�'�%$��$,� ���� �$%�&��%�#&�� ����(����&���$%�� ��$�� l’��#�%� ��$��� �/$%����#�$�����%����$%�����'��D’après�%$ &��%������ �,�?-�0'�

� ��������������� �����
FG�
�
��!���'��)�"�('���&�(�&#��'5���,�"?(&�"�)�&���������&�&)�4�������"#���&��'�)����)���)���""("�)�
&'��&#�)��(�����,�"��'����)����*��&���)('�'��)�"?�*�'�))����������-���'���$�������)���)�
�����&���)@� #?��!���&���� �(� ����4� ���&� "?�*�'�))���� �)&� '#�("#�� ��'� ����� ��('��&� �"�')�
'��(�')�'�"?����&����"��'����)�����)('�"���#�'���&�������"?4�������-@�#��)����������'��!��
���)�)&�� ,� ���"))�'� ��� ������ �"���"�� "?�����&� �?(�� &'��&����&� ,� "�� '����)����� )('�
"?�*�'�))���� ��)� ��&�&)� 4��� ���;�����&)� �'5��� ,� (��� �(��� ����4� ��)�����"�� ���)� "��
�����'��@���()�)#"��&�����'��)���)(�&����(*����&�"?�*�'�))�����(����&�*��&�$(���'#)��&��
(��� ����"#���&�'�&#� ��� )#$(����� �(��� "?4���� ��� ���-@� #�� (�"���&���� ��)� �������&)�
��(''��)�����'����� "���.���������$(����('���(*� ����&���#)���'� "?���'��!���������'��&�$(�@�
���� ��'�� '�� ���'��!�� ���)�)&�'��&� ��� "?����&�����&���� ��)� "�����4� $(�� ��&�'���))��&� �(���
"?4����������-�)(�&��,�(��&'��&����&�,� "��'����)����@���"����(''��.&'��'#�"�)#��'5���,� "��
&��!��$(�� ��� �('�����&���� �?4��� ��'� ��)� )����)� ��&�)��)� 1��4;�4�*� ��4<��4� 4�&�)��)�
�('�����&���6� '#������&��#�'�&�)� 1���'��&+� �&� �"@*�=1-76� 1������ ��6@�������� '�� )(�����&�*�
��&&�� ���'��!�� ���)�)&�� ,� ��*�'� "�)� ��""("�)� �(��� �(� ��'��"�#!)��� ��('� �'#)�'(�'� "�)�
��&�'��&���)�4��<4��@�#�)� ��""("�)�)��&���)(�&�� "))#�)��(�)� "�)��*&'��&)�)��&� &'��&#)����)� "��
�(&�������#'�'� "?4������)��$(�� "�)��'�&#���)@�#?4����)&��"�')� �'�����&#��&� ���(�#��(�����)�
)����)����&��)"#�)�����"#���&��'�)������&'��4������"�@�#?���(���'#����&�&�����)&�'#�"�)#��
�'5��� ,� ��)� ��""�)� ��� )&'��&�(������ �&� "?#"(5&� )�(��)� ,� (�� )#$(������� !�(&� �#��&@� #�)�
��&�'��&���)�$(��)�'��&�"�)��"()���'&����&�)�)�'��&�(#'���#�)��(���)���'���)���'���;$������' )�
�'#����&�&�������"?4����������-�)�"���"���.����'�&���"�@�#?���'��!����(�')�����)�)&��&�,�
�'#����&�'� "�� �������&� �&�(#'����'� "���'#)�������� "?4����������-���'��&&'����� �����'��'�
�#����&�(����&� "�)� ��&�'��&���)@������*� ��()���(''��)�(�"���'� "?����'&����� ����&�����""�����
��)� ��&�'��&���)� ��� �����(��&� "?�*�'�))���� ��� �!��(�� ��)� �������&)� �'5��� ,� ��)� 4���
��&�'�#'��&)*��(����"?�(����&��&��'5���,���)�(��&�(')��?�*�'�))���@�
�
����� �������� ��������������������+��������� ������������
#?!)��'��&�(�&���� ��� "�� (���� �:A�<4$&<����� �)&� "?(��� ��)� �"&#'�&���)� "�)� �"()� �'#$(��&�)�
��)�'(#�)� ���)� "�� �'��'�))���� �(� �����'� ��� "�� �'�)&�&�� �(�)$(?�""�� �)&� �'#)��&�� ���)� "��
$(�)�;&�&�"�&#���)������')��#&�)&�&�$(�)�1��('������&��"@*�=11=B���)"�'��&��"@*�=1-1�6@���('�
��&&��'��)��*�"?(&�"�)�&�������'����)������&����)�)����"��(�)�1'���"��(�)6������(�)�"���&���)�

����&��� ������������ �
�
���!���" #�$�������������������������!��!����������������������������������������������� ���#���!%����!��"�������!�!���"���6�����#%�'#��*!�����!#�%�%'%�(���#%���%�(���������2��� 38/���(�"!���� ���� %'��'��� 2% %�� ��� *"����� �"�#���/� �'� #�(��'� ��� "�� ""�#��� ������!���9���'�(�"�#%���� "����!�%�%�����&� "����!�!���"�=����� %'��'����!#%�����'�����#�(��%�"�������l’expression� ��� "�� �#�=� ���� ��""'"��� ��� ���� %'��'��� �'��6�����#%� ����9� l’!�%�!"!"'����!�!���"��#�������3=��4��>��#%����"�#%����l’ADN=�"����%�������'���"���#!%�'6=���!���"��>���!%��#�����"��'�����#��l’adhésion���""'"������'�#�(��'��������%��"�'�=��""�����%�������'���"���!'�%!'��������""'"������%��"��"�����**���#�����=�

� ��������������� �����
<A�
�
,.,�.�,.,�����%��,����!�!(/��%"/+����.+�!.����.���,�-3�-:�,��,��,�,�!���,/+����,/+4!��"�"�����
��,� %�.!��.,� 5�!..!�"� ���� 4+�,.+"�":� A@BC?0� -�.� ,����� %�/.� ,6�;%�!(/�+� !?� %�+� ��� "+�����
4�+!,.,��6�����.�/+,�������4"!�:��.�������(/�����,%,�!�!�!.,���,�+�%��""/�,��.�!!?�%�+��6��"�!.!"��
�/�+,.+"�"�.+-����,"�.!���;�+�,�%�+��123-B�,/+��123-A�,/!.���/�.+�!.����.:�+,,/�.��.����
/��� ��.!4�.!"�� ���+/�� ��� 4�.� 5'!����� �.� ��0:� A@@=?0� 6�� �,.� �"��� !�%"+.��.� �6!,"��+� �.� ���
��+��.,+!,�+� ��,������.�/+,�����4������ ���4"!���6C774�.7�123���!���6�"!+�,%,�!�!(/����.�,/+�
��,� �!���,� %".��.!����,0� �",� +,,/�.�.,��"�.+��.� (/�� ��� ,/+�;%+�,,!"�� ��� ���B� !��/!.�� %�+�
�123� �,.� �,,��.!����� %"/+� ��!�.��!+� ��,� ����/��,� ���,� /�� ,.�.� %+"�!�,+�.!�0� '�� ���!$+��
!�.,+�,,��.�:�����!�!�/.!"��������B�%�+�!�.�+�,+�����43��+,�/!.����.�/;��6!��"+%"+�.!"�����
�+����.� �6���/�/��.!"����� ����1��!���'B����,���,�����/��,��.��<7<���!,�%�,����,� ��,�����/��,�
,�/4�"�,0� -�,� �"��,�,� �"�.+��.� �"��� /��� �,%�������� ��,� ����/��,� 4!,<�<4!,� ��� ���B�
�"+,(/���.����,.��/.,:��.������!.��"���/����!���������"!;�%"/+�/���!���"��.�,+�%�/.!(/�0��!���
(/�� �"/,� �1"�,� �"��!+�,� ��� �!��� ��.+�� �6�1%�+��.!4�.!"�� ����123� �.� ��� ,/+�;%+�,,!"�� ���
���B����,���,�%+",.�.�,����,"/+!,�72�%"/+��.��:��6��,��������,"/+!,��4���/�����$�����";,����
���B� +���� �!��!�!��� ��� �,�"�,.+�.!"�� �6/��� 8����B<�,%��������2� ��� ��0� 4�!�� ���
�"�."/+��+� ��� %+"��$��:� �"/,� �,4��"%%"�,� ��./�������.� /�� �"�$��� ��� �+","%�!���
�;%+!���.� /�� 43�� !�.�+�,+��.� �!+!",� �"�.+�� ���%:� �6�"�"�""/�� ��� ���B� 53�4!<���%?�
5�!��,.+'�:�A@BB?0� ��,� +,,/�.�.,� %+,�!�!��!+�,��"�.+��.� (/�� ��� ,/+�;%+�,,!"�� �6/��� �"+���
�/.��.���"�,.!./.!4����.���.!4�����3�,�53�, BA?���.+�!�������"+��.!"�����./��/+,��/��!4��/�
��� ��� "������ ����,,"!+�:� ,(/!4����.� ��� ��� %+",.�.�� ���.� ��� �+","%�!��0� '�� ���!$+��
!�.,+�,,��.�:� ��,� ����/��,� ./�"+���,� ,/+�;%+!���.� ����� � 5$�$�,�� ��?0� ��,� .+�4�/;� ��� �"/+,�
4!,��.� ��!�.����.� �� �,�������+� ��� ./�"+!"��$,�� !��/!.�� %�+� �6��.!4�.!"�� ��� ��� 4"!��
�6C774�.7�123� ��� ,/+�;%+!���.� /��� �"+��� �1+!,."1�,�� �.� �"��� �"�,.!./.!4����.� ��.!4��
�64�.� %"/+� ,/!4+�� �6�;%+�,,!"�� ��� ���%� ���,� ��� �"�.�;.�0� .�� /.!�!,��.� ��� �"�$��:� �"/,�
%"/++!"�,�+,�/!+���6�;%+�,,!"����������"+��������,��"�,.+/�.!"�,�3�4!��,#���!,%"�!���,�%"/+�
4"!+�,!������!���/��,/+����./�"+!"��$,��!��/!.��%�+�4�.0��
.��!�:�."/.����.+�4�!����4+���.+���"�%�,.,�%�+���,�����1,�,���!�!(/�,:�(/!��"/,�%�+��..+"�.�
����,.�+�!��+�,6!���;!,.��/���!�����.+���123:����B��.��6�"+�,,!4!.,��/������+�������%+",.�.��
���,� ��,� ,����.!��"�,� ./�"+�/;� �/��!�,0� �"/,� /.!�!,�+"�,� %"/+� ����� ��,� ,����.!��"�,�
./�"+�/;�"+��,,��.�.+!,,�,��"�� �6�"+�,,!4!.,��/������+��.�,��+,,!,.������/;�.+�!.����.,:�(/!�
,"�.� �!,%"�!���,� ���,� ��� �"���+��� ,"/,� �"+��� ��� 1�4� 5������� ����������?0� �"/,�

��/�
4��
���
� �!���"��������#��#�� ����
����$�����#�#"(��
���
���
���
��)��
/�
��)��
/�
��)��
/�
������� �����������������������&������������������������������� �'�#� #�� �#� #"��" �#� *��3� ��� � "�# ������#� *��)��/3� ��� 0� (��#� ���� ���� ��������#������������(����!���"���/4���� #�"����/49�( +� ����! �!���"��������#��#�� �������"����$����$�(���� �#!���"���d’huile����#�#"(��*������$�3%�-����"����: ����! ������#��+� "���#��� !" �� � ��#� $���#� ! �#�"�����#� ���� "�+� ��� ��"���� #�� �#� ���� ��#����� ���� "�"$(#�#� !" ���#�� �)�$���"������#�"����� !#��� � �#����� ����/����4�%�

� ��������������� �����
<=�
�
��,0&�,"�%�&�%��4�",�&�� ���%���'!&",(%��!,!",%�&"�',�����;=�(�"%(�"57���;�60&!��%'����
��,0&�&,�����<�&'!)!',��������&��%%!0&����'�&'!�!%,����%���%�,'&��%�&�!�!0&�%1=��'��'��>�'�%�
(,��!$,�%� �"��,�%� "�'��&�%� ",�&�� � &��� &"����",�'!"�� �)�&� ��� &��',�� ��� �&''�� &"�',�� ���
&��&�,� ����� ������� �� -����� �������� ��� � ����������� ��� ����������� ��� ��� � ����
������ �������������� ����������������� ���� ����������������������
�
�+�����������������������������,�������
�
���� ���� � �� �� ����� � ��� ��� �� ���� ������� ������������� ����������� � ����� ����
���� !��������� � ������������������ �� �������������� �� ������������������� � ��� �
�� ������ �����.���""� #�������������������� ����!$�������������� � ������������������ �
������������������������������������������������ � ��������� ������ ��������� ������
�!�%� ��� ������� ������������� �� � ������� � ������ �#� � � ���� �� � �������� � ��� �����
����"��� �"�������������� ���� !������'�������� ����������������#� ������������������
������&��� ����� ��� ������� ��������������"�� ����� ����� �� ������ ������� ������
�$��� �������� � � � ���� � ��� ���� �� ���������� �� �$ ���""������� ����� -��� �������
��� ��� ������������ ��������������������� �������� ��""� ����� ������!��� ��������#�
������������� �������"�� ��������������"���������������������������&��������� �� ������������
�� ���� ��� �����������������'��� �������� ����������������������������������������
���� !��������� � ������������������ ������������������� ���������%�"�� ������� ��� �������
���������� � ������������ �������� �� �����&��� �� � ������ � �� ��� ��� �� �� 4(#� ���
����������� ��������) �������"���������""�� ��������� ������� ���� ��������� �����"�� ��
������ �� �� ������������������ ���������������������������������������* ��������"����
��""����� ������� ���������!�������*�� �� ���� ��������� ��+,���-.������������ ����
�� �� ������ ���� ������� ����� � �+� *������ ������������� �� �����&��� ����� ������ � �����
���������� � ������ ��� /������� �!0� ���� ������ � ���� ������� �$�� � ��1�� ��� ��� ���� �
��� �2����
�
�

�� ��
-���%�����%�4��
-���%����������
-���%������%��:������3���
-���%�����%$3�����
� ���- ���������� ���� ������ ����
Figure 44. Principaux marqueurs de différenciation dans l’épithélium prostatique.�����300�����3��3�����������%��� ��%�4���#������3!%����������%�����#3�4��3������������&��%$3��������� ��%��:������3���� 3$#�3!%�� ��� #����4�� #��� #�%�3�%��� ������� 3����$��3�3���;� �������� ���#������%�&� l’acquisition� �%� ��� #����� ��� l’expression� ��� $��!%�%��� ����� !%�� ��%'� ����3��� �3:����%��#��$���d’identifier��������������300�����3��3���d’une�����%��;�7��#��&�=332;�
3
3&3
4
4&3
=
=&3
>
>&3
51�6377 6377��:���4�
7�6�$8�
4������3*
����
*���
83;33�
51� �4�-8��4���4�
Figure 45. NPM1 induit l’expression de l’EGF.����� 48�� ����%'� �'���3��� �� ������ #������3!%��� *�����%'� ��� ��%�3�� ��%*�4��� 8519� ��������4��3!%��� 8��:���49����7�$�3��������������-����#���81:!�-8;�L’expression����l’EGF��������$��3����#�����##���� �l’expression����>4�7;��.�4�#�%���4�!%��4����-#���;�����48������%'��'���3�� ��� ����%���� ��-��� �'#�3$����%�� 48�� 3����0������ �3�34�� ������� ���4� 8�4���49��%�%��� ��!%����� ���� �#��303!%�� 8�4�-89� ���� ���� ����-���� #��� 81:!�-8;� ���� ���%������ ���l’expression����l’EGF���������$��3����#�����##���� � l’expression�������4;��.�>��'#��3������3���#��������;�����34�303���3*3�������������%�����*���%�������1;����
4� ��

� ��������������� �����
<:�
�
����������������������,�&�������,����&�����������������,�������
�� �!!�,*'$��+�#'� *-� $*��:�*-� �(�+ �����*-�(,#-+�+�.:*-� *+� ��� ,��:��+�#'� :� $9$�*� $*��:���,*�
-#'+� *-� (,#$*--:-� �'+�"*"*'+� ���-/� � 4:5 *��� *� �**0(,*--�#'� *� 4�-� .:�� �++*-+*� *� ���
�!!�,*'$��+�#'� !#'$+�#''*��*-� �*-�$*��:�*-��(�+ �����*-�"�+:,*-��$$:":�*'+� �*�-���(:;��(</�-*�
"�,.:*:,� *� �!!�,*'$��+�#'� +*,"�'��*� -*� !�0*�*+� �' ��*� ��� -'�:-����--�'+���'-�� $#""*� :'�
(:�--�'+�!,*�'��� ���(,#�,*--�#'� :�$9$�*�$*��:���,*�*+�$#""*�:'���,�'+� *� ���.:�*-$*'$*� *-�
$*��:�*-��(�+ �����*-�:'*���,>#�*+���/-�<££=8��:����*+���/-�:>>?@/�'*�"�'��,*��'+�,*--�'+*-����
-:,*0(,*--�#'� *���<� �'-����(,#-+�+*� *�'#-��'�"�:0�*-+�$#,,���*���:'*�(*,+*� *�(:;��(<�
*+���:'*� �"�':+�#'� *��**0(,*--�#'� *���'*-�$���*-� *�4�/�'*�(�:--� *-�,�-:�+�+-�*'�$#:,-� *�
(:���$�+�#'-�"#'+,*'+�.:*� *-�$*��:�*-��(�+ �����*-� *�$�'��� �!�,*'+�(*, *'+� �**0(,*--�#'�
*���<��:�!:,�*+���"*-:,*� *���� �!!�,*'$��+�#'�:�#+#�'�������-�-#:"�-@/�-*++*� �"�':+�#'�
*� ��<� *-+� �--#$��*� �� �*�$$:":��+�#'� *� (:;��(<-� $*� .:�� ,*'!#,$*� �*� ��*'� *'+,*� $*-� *:0�
(,#+��'*-/�'*�:+,*-��+: *-�#'+�$#,,�����*�'�;*�:� **0(,*--�#'� *���<�*+���� �!!�,*'$��+�#'�
$*��:���,*/� �� (,�-*'$*� *� ��<� *-+� *'� *!!*+� '�$*--��,*� (#:,� �*� "��'+�*'� :� -+�+:+�
�' �!!�,*'$��� *-�$*��:�*-�-#:$ *-�":-$:���,*--� *�$*��:�*-�$�'$�,*:-*-� �"�+#(#��+�.:*-�#:�
*'$#,*� *-� $*��:�*-� -#:$ *-� *"�,9#''��,*-� :. --� ."�,9#''�$� +*"� -*��-@� :���:-:� *+� ��/-�
:><<8� -�""�-�*+���/-�:><A8�5*,�*,�*+���/-�:>>;8� �# �'--#'��' � �"#'--#'-�:><>@/� ��� -*,��+�
#'$��'+�,*--�'+� *� �+*,"�'*,��*�"(�$+� *����-:,*0(,*--�#'� *���<�-:,���� �!!�,*'$��+�#'�
*-�$*��:�*-��(�+ �����*-�(,#-+�+�.:*-/��#:,�$*��-�'#:-�(,#(#-#'-� *�'��9-*,�� ������*�(,#!��� *�
�!!�,*'$��+�#'� *-�$*��:�*-�*0(,�"�'+��*�+,�'-��'*�(�,��"":'#!�:#,*-$*'$*�:$*��:�*-�����¥@/�
#:-� ,����-*,#'-� :'� $#5"�,.:��*� *'� :+���-�'+� :'*� $#"��'��-#'� *�'+�$#,(-� !�:#,*-$*'+-�
�,���-� $#'+,*� �*�(�+#(*� !���� *+� $#'+,*� *-� "�,.:*:,-� *-� �!!�,*'+*-� �+�(*-� *� ���
�!!�,*'$��+�#'� *��*�(�+ ���:"�(,#-+�+�.:*�:� ������@/�.'��+����--�'+�:'�(,#!���(,�$�-� *�$*-�
"�,.:*:,-� �'-� �*-� $*��:�*-� ����¥-� '#:-� (#:,,�#'-� �+*,"�'*,� .:*��*� �+�(*� *� ���
�!!�,*'$��+�#'��(�+ �����*�*-+���#.:�*�(�,� :'*� -:,*0(,*--�#'� *� ��</� ���'**-+�$*(*' �'+�
(�-�*0$�:�.:*�$*++*� *,'��,*� �' :�-*�:'*� � �!!�,*'$��+�#'� *-�$*��:�*-�.:�� ���-:,*0(,�"*'+/�
�#:,�+*-+*,�$*++*� 9(#+ �-*-�'#:-�(#:,,�#'-�:+���-*,����"�+ # *� *����'��*�$*��:���,*��,�$*���
*-�(,#+��'*-�!�:#,*-$*'+*-/��#:,�$*������!�: ,��+���'�,*,� *-�-#:,�-�(#--� �'+��*���'*� *����
,*$#"��'�-*�-�.�:����������� ���� @�-#:-� �*�$#'+,+�*� :�(,#"#+*:,� *:'�"�,.:*:,� *�
�!!�,*'$��+�#'�+*,"�'��*�$#""*�(:;��(<-���'-��.:*����$#'-+,:$+�#'�"B"���:�'�;*�:� :��#$:-�
�#-��:C-�$#'':�(#:,�-#'�*0(,*--�#'�:��.:�+��,*�: #,��'#-�<£££@/�-*++*�$#'-+,:$+�#'�$#'-�-+*�

Figure 46. Activation des récepteurs de l’EGF dans un contexte sain et néoplasique.�' ���%���#��*���%$�� ��4� �������#��%������ l’EGF�������%������%�� ���� ,����������������!%���������������%���������������=�L’EGF�������� %���(� %� #� �����#�%������# ������ �������������������#��%������ l’EGF������#�������qu’aux�! ��%����������� ���#��(�� ����%�����$ =�' ���%������%����#� ��!%�4�� �#��������l’intégrité����l’épithélium,���� $$�����0��'�� �#�����������,����������������%� ����4� %�������l’EGF�d’origine��#��*��� ���'� ��(����������#��%������! ���� %������=�
'�-���16�
��.�#232�
�"2���
�7��.�#232�
;
;4;(
;4�
;4�(
;4�
;4�(
;4�
;4�(
;4-��<�;4���
�����������������������������������������������������������������)����������� ��������)���� �#��������%�$ �!%�%�����#����!�� �������16� ����� � �%����# �� �$$%��*����*�$����%�������%#��������"������� %62� ��� %6�#���� ��!%��������%��������#232������2����7����#232=�L’index� #����!�� ��!� ���� �%��� ��� � ##��� ��� ��� ��$"��� ��� ���%���� �$$%���� ��(��� %���$"���������%������� ���=�4%�$�����-;;;����%�������������$#�����#�%��* !%������(��%�8�<����%�������* !%��#����%#�==� �
'���
6�#����!�� ��!
�
��.�
#232�
�"2�
���7
���
.�#232�

� ��������������� �����
<=�
�
�,�?,�$'�4�$/4�,=�-/?/����-//=&�,���/%�=/�5-//=&�,�� �?//�,$�,=��//?!�?�,?�.��4*?,�$/4/,�
�=/-/��*4'�$�4���/%�=/��,=��,$�4/&�-�/�4�,�,�=�,� �/.4��=��,=�,?�.��4���*4'�$�$/4�,=�-/?/����
����5������ ���������������?/�4�,,�4�4�,,���,��$���?��,�//����-//%/=�?/�4��-:C��-<��,=��$=�.&4�
���-�.��,=��.-/�%&���=�������,4?�=��*�.$�,�/,�4���*4'�$�4���/%�=/���,,��1?��4?�$/4/,�,=/-��?�
,�.��?�4?� �/$?,��/,�:D/����$���?���,�/����//,�%�/1?&��4�� �$/,� �//&.�/,������,�.�/=�!/�$����
�*�.-/�,,�/,�4��������/�4?�$/,=/��/�4���,�$���?��,�,*�7�,=�-�,��==��,=���,�,=�4�,�=�/%�,�?.�4��
��� 4� &/�,$��=�/,� �--�/��=//,=� �,� //?!�/� 4-/�,� �./�/� $//�,&� $�,� ,/?/�,� �.�$� ,/,� ,/?/�,�
4��:��<���<4�,/?,��,��7,�//,,����$/?��?/�4�,�$���?��,��.-/�%�,=�,/=/��=/�,,!�,�/��������,�
,/,=�.�/=�,4�$*�,=�1?�����<��,4?�=����4&4� &/�,$��=�/,�4�,�$���?��,�&-�=�&�����,/��������,�,/,=�
//?!�,4�$*�,=�1?�����<���/1?��?,��&=�-��4�����4� &/�,$��=�/,/�
�
���������������������������������������� ������������������
�/,�4/,,&�,�%/,=/�,=�1?�����,?/�.-/�,,�/,�4�����<�4�,,����-//,=�=���,4?�=��*�.-/�,,�/,�4?�
�$=�?/�4��$//�,,�,$��.���5�������!?/�-��/&,?�=�=��,=���%�==/���,�-�/��������.�$�,/,�=/�.�?.�
�,=&/��?/,� ,?/� ��,� $���?��,� �?%��,�,� 4�� $�,$�/� -//,=�=�1?�� ��-��4� 4�,,� ��,1?����,� ���
4�%�,?=�/,�4�����<�-�/��,=�/ &/�,$��4���/&4?�=�4�� �$/,�,�!,� �$�=�.���*�.-/�,,�/,�4���*.���
5�/?���?� �=� ��/4� :><A?/� '�,,� ��,� $���?��,� ��-��4� $�==�� 4�%�,?=�/,� �,=� �,,/$�&� �� ?,�� -�/=��
4*�$=�.�=�/,�4�����./���4�,��4���%�,?/&���,��,��7,�,=��*&=�=�4��-�/,-�//7��=�/,�4���*.����
�=� 4�� .��/� '��%�,��/�� ,?/-/�,�,=�4� �*�$=�.�=�/,� 4�,��4��� ,*�,=� -�,� /�-//4?�=�� 4�,,� ��,�
-//,=�=�,� 4�� ,/,� ,/?/�,� =/�,,!&,�1?�,/� �,�� �.-��$�=�/,� -��?,����� �,=� 1?�4� �,� $/,4�=�/,,�
-�7,�/�/!�1?�,4� ��,� ,&$/&=�/,,� &-�=�&�����,4� 1?�� /,=� ���?� �?� -*��� �-�$��� 4�,� $���?��,4� ,��
-�?.�,=��==��,4/����,�/&$�-=�?/,��?.� �$=�?/,�4��$//�,,�,$��,�=?&,��?�,�.��?���,���,/?,�4�,�
�/,$=�/,,� ,�//&�,� &=�,$��,� 5�/,=�,/� �,4� '��%!/>4� :>>A�?� 5������ � ?/� '�,,�?,� $/,=�.=��
&-�=�&�����,/,<%���,4��� �*�%�!��4��,/=/��%/4���4�$�==��4�,-/,�=�/,�,*�?=//�,�� �*�$=�.�=�/,�4��
$��=7-��4��/&$�-=�?/,�1?��-�/���,� �$=�?/,�4��$//�,,�,$��,&$/&=&,�-�/���,�$���?��,�,=//%���,/���
,�/��=� 4/,$� �,=&/�,,�,=� 4�� $//�,�/� $�,� ,/?/�,� �.�$� ?,� %/4���� 4&.��/--�,=� 4�,� �&,�/,,�
-//,=�=�1?�,� ,&/-��,�1?�,� $/%%�� ��,� ,/?/�,� ��� -/?/� ��.�� 5�=�,-$<7<?/� .,� 4&,//!�,�,�,=�
�*&-�=�&��?%4�$����-�/%�==/��=�-�?=��=/��4��/�,4/����,�/&$�-=�?/,�.����4�,-/,����,�-/?/��*.���
4*//�!�,��&-�=�&�����-/?/�%�==/���,�-��$��?,���/?$����?=/$/�,��4*�$=�.�=�/,�4���*.���/�4�,,�4�
��,�$���?��,�&-�=�&�����,�%���!,�,�4�.��,4/���,=��?=/<,? �,�,=���,� �$=�?/�4��$//�,,�,$�4�$��

�

� ��������������� �����
<;�
�
(/��$)&,.�./*�/&*�$�+�$.�+�,.�(/*� ��%)+.�&.*� *,�$*��/�*,�$�&$�+*/,*,�*&���&�+���25�&� �&�
�& ��*�&�*+�4� D6..�83� ��� *,.� ����*�*&.� %),,���*� (/*� $*..*� ,)/+$*� -.��� ,/%%���*&.��+*�
4�*&&*� ,-�&���.+*+� �&,� �*� ,.+)��� %)/+� �)/*+� �*� +'�*� *� $ *�)9�..+�$.�&.� ��4)+�,�&.� ��&,��
�-�&4�,�)&� �)$��*�*.� ��� �)+��.�)&� *���.�,.�,*,�$)��*� �$+�.�%+�$� *��*&.� 2����&�*.���34�
.==A83� -*,� 1%). �,*,� ,)&.� �$./*��*�*&.� *&� $)/+,� *� 4�+���$�.�)&� %/�,(/*� &)/,� �4)&,�
�4*�)%%�� *.� $)��*&$�� �� �&��1,*+� �*� % �&).1%*� *� ,)/+�,� 4��D��9 %�.1�.*&%$9193� -*,�
,)/+�,�)&.� *,�./�*/+,�%+),.�.�(/*,�(/��)&.�.*& �&$*���$.+*�%�/,��+),,*,�*.���%+�,*&.*+�/&�
�& *;�%+)����+�.���%�/,���*4��(/*�$ *.��*,�,)/+�,��.*&%$919�2$�$�,���83�-*%*& �&.4�$*,��&���/;�
&*�%+�,*&.*&.�%�,� *���.�,.�,*,���.D��)�,�*.�&)/,�&-�4)&,�%�,�)�,*+4�� -�& /$.�)&� *� ���
4)�*� *,��4��3� -*,� )�,*+4�.�)&,� ,)&.� %)/+� �-�&,.�&.� �� %)& �+*+� �4*$� �*� �����*� &)��+*�
-�&���/;� (/*� &)/,� �4)&,� %/� �&��1,*+3� '*,� ,)/+�,� ,)&.� �$./*��*�*&.� *&� $)/+,� *�
4�*����,,*�*&.� *.� 4�*& +)&.� %+)$ ��&*�*&.� �$$+)�.+*� &),� $) )+.*,� %)/+� �/��*&.*+� ���
%/�,,�&$*�,.�.�,.�(/*� *�&).+*�.+�4���3�
&*��/.+*�%),,�����.�� (/�&.��/� +'�*� -/&*�,/+*;%+*,,�)&� *� �-.��� �&,� �*,�% �&).1%*,�(/*�
&)/,��4)&,�)�,*+4�,�*,.�(/-*��*��/+��.�/&� ��%�$.� /+�&.� �*� �4*�)%%*�*&.�%),.9&�.��� *����
%+),.�.*3� .&� *��*.4� $)��*� �*&.�)&&�� �&,� �*� $ �%�.+*� � � � � *� �-�&.+) /$.�)&�
�����)�+�% �(/*4��-.���*;*+$*�/&�+'�*�%+)����+�.���,/+��-�%�. ���/��%+),.�.�(/*��)+,� *����% ,*�
*4*�)%%*�*&.��*� *��)/+�*)&&*�*&.3���*&�(/*�4��&*�,)�.�%�,�*;%+���� �&,��-�%�. ���/����
$*� ,.� *9��4� ��� *,.� $)&&/� (/*� �*� %+)�).*/+� 4��D��� *,.� �$.��� �,� ��� &��,,�&$*� �&,� �*,�
�)/+�*)&,��%�. ����/;���&,��(/*� �&,� �*,�$*��/�*,�,.+)���*,� 2��+�,)..)��& ��*..�*+4�D6.:83�
)/,�%)/++�)&,�&)/,�*&��,,/+*+�*&�%+��*4�&.� *,�,�&/,�/+)9��&�.�/;� *�,)/+�,����*,� *�.���
.B� �)/+,3��-*;%+*,,�)&� *�&).+*�.+�&,��&*�*.�,�� �)$���,�.�)&�%)/++��*&.���)+,�$.+*��&��1,�*,�
%�+� 1�+� �.�)&���������*&�/.���,�&.�/&*�,)& *�4� �$)�%���*&.��+*� *�����)+�*� /���&*� *�
��.3���� �*� .+�&,��&*�*,.���*&�*;%+������$*�,.� *� *� �4*�)%%*�*&.4�&)/,�%*&,)&,�(/-���
%)/++��.� �& /�+*� �-*;%+*,,�)&� *� �-.���*.���&,����4)+�,*+� �-�%%�+�.�)&� -/&�%�/,��+�& �&)��+*�
*��)/+�*)&,��%�. ����/;3�4� �-��*�� /�.*4�$*���%)/++��.�,*�.+� /�+*�%�+�/&*� *&,���$�.�)&� /�
+�,*�/� *,� �$�&�� (/�� *;%��(/*+��.� �*� %)� ,� ,/%�+�*/+� *,� %+),.�.*,� %+)4*&�&.� *,� ,)/+�,�
4��D��9 ��.�%�+�+�%%)+.��/;�,)/+�,�,�/4��*,3��
�
�

Gène de l’EGF humain�
Gène de l’EGF murin�
Gène de l’EGF de rat�
���������%������������������������������������������������� ������������du gène de l’EGF de l’homme, de la souris et du rat.�E�� ����� ����� ��� ����� � �� ������ �� �� ���� ������ �������� �������� �� �� � � ����� �� ������� ��� ����� NFκB�� ���� �� ���� � �� ���� ��������� d’analyses� ��� ��������� ������ ����������������� ������� !!"""���������� �������!#��

� ��������������� �����
<A�
�
��������������������!������������������������������� ��
���$%� �&��� �� �'���(��%� (�)�*��()� �&��� '�� �+� ����*�+� �� �� ���(�� ��+%� �,��+%� -�'*�
+�((�(�*�+����))��.����)���������*����-�+�(/'�0�-���*�(�*��+)���,���(+���&����&�����
���-�1���*�23��+����/'(�-�*��)�+�����)(&�)(����)�%������+���%���������&�(���+��4���
4��*� �)� �0%� 255670� � �+)� -�++(��� /'�� ���� ��))�� ��� -���� '��� ��'��� 8��-(�(��)(���
�')��*(���(�-(/'��)�8�'����)�)(�����8�1-*�++(�����8.��0�.������)%�'������9+��+'��(��)��
'� -*���)�'*� -*�1(����� 8.�����'+��� -�*�(+�����))*������&(����� '�� +()���� �(1�)(���
-�'*������)�'*���)*��+�*(-)(�����,:���.�;�8.����%���+�'*(+��(�+(�/'����*�)%�+'���*��)�/'��
��'(,�(�+�()�����)(�����4������"#70��*�(����)�����)*��/'����&�(���+��4�����)(&����:����+�
�+���'�+��-().�(��+�-*�+)�)(/'�+� 4�(���)��0%�255270� �+�*�()����� (�)�*�++��)�8���9+�*�
8(�-��)� 8'��� (�(�')(��� �� � ����� +'*� 8��)(&()�� 8'��� ���+)*'�)(��� ���)����)� 84'���
����)�-�'*� �� '�(��*�+�����������&��'�-*���)�'*��� 8.��0��(� ��*��)(�����)*��������)�
8�1-*�++(����� 8.����+)� ����(*���� -�*� ��))���--*��.�%���'+� -�'**(��+���+'*�*� -�*� -.��
8��*(�.(++����)� �� ��:�� +'*� �� -*���)�'*� �� 8.��� ��� ����)(��� �� �� -*�+����� �'� ��
8��+����� �� ���$0� �(� ��� ���)�'*� �8�+)� -�+� (�-(/'�%� '��� ���9+�� �(�(���*��)(/'�� -'+�
-�'++���'�-*���)�'*����')((+��)�'�� ��(�(��)��/'��������)(1<���'+�*�&&�*�()�8�')*�+�
���(�)+�-�)��)(�+0�
�
���������%�����������������������!�������������������������������������
4'� &'� �� ��+� *�+')�)+� 8'��� �+� .9-�).&+�+� /'�� ��'+� -�'*+'(&��+� �+)� /'�� ���$%� ���
(�)�*��(++��)��)���� (�.(���)���%� &&�� �� *�-*�++(��� +'*� �� ���)�'*��� )*��+�*(-)(��� .2�$0� -��
�*�(�*�-�'**�()���*+�+)(�'�*�8�1-*�++(��������9�(���.�-�'*�-�*��))*����-*��*�++(���'�
�9�����'�(*���)��(�+(����'(*����'���-*�(��*�)(��0�4&��)���+�)*�&�'1%��+��/'(-�+��&�(��)�
�������)*��/'����+'*�1-*�++(��������$��'�+��+'��9�)(���+'*���*�+('�9+(������-�+()(���
2=>��)�(��)� *�+-��+������ +��� (�)�*��)(����)��� �� +�/'�+)*�)(����������+� ���'����%�
(��*��)� ��(�+(� 8��)(&()�� )*��+�*(-)(�������.2�$� 4�('��)��0%�255?@������'*���)��0%�$666�70��
��'+��&��+��++�9��������(*��*�8(�)�*��)(�����3���$�-�*�(��'��-*��(-()�)(����(�+(�/'����
+�/'�+)*�)(����������+����'�����-�*�(��'���'�*�+��������-�*)(*��+�-*�+)�)�+�����+�
��(��'10� ��.�'*�'+����)%� ��+� )��)�)(&�+� +��)� *�+)��+� (��*'�)'�'+�+%� -*��������)� ��
��'+�� �� �� ��(��� ���(�()�� �� 8��)(��*-+���� ')((+�0� .�� �))����)� �� +�� -*��'*�*� '�� �')*��

�

� ��������������� �����
<F�
�
�'.�$#+%,�*.� *�"*..+*��/�%#�'.�,*,�$#' �.�#',� ,/.���,�.�#'�*A%�+�"*'.��*,:�'#/,��4#',�.#/.�
*�"$"*�"#'.+�� (/*� ��� ,/+*A%+*,,�#'� *����C� �' /�,��.� �,�$.�4�.�#'� *� ��'*,� $���*,� *�
.@�C� %�(�B �%*' �'.,�0� 5.@�C:� (�:� $C$��'*� .� *.� �$'�?:� ��#+,� (/*� �*� ��'*� $# �'.� %#/+�
�, +#��'�,*� ���,"�'#�*'� 4$.�4�.#+:� (/�� �/�� *,.� %�(�B�' �%*' �'.�0:� �.��.� �',*',���*9� '*�
"�'��+*� �'.�+*,,�'.*:� $*..*� �/�"*'.�.�#'� *� �,*A%+*,,�#'� *,� ��'*,� $���*,� *� .@�C� *,.�
�,,#$��*���/'*��#+.*� �"�'/.�#'� *��,*A%+*,,�#'� *����-C$��'*�'C9�-*�%�$C$��'�,9�.$ �0��� ����
�.�� �$+�.� �',� �*� "# ��*� *� ,#/+�,� �(4��� 5���������� ����� ������ ��� ���� ������
� ������?��5�� �,#'�*.���9:�@AAD�?9�- *>�$*,�,#/+�,:����,/+*A%+*,,�#'� *��,�'.���'*��� /�4�+/,�
��DA� �'.*+���.� *.� �' ��*� �*,� ,/%%+*,,*/+,� *� ./"*/+,� (�� *.� %EF:� "*'�'.� ��',�� �� /'*�
%+#����+�.�#'��'�+$ �(/*� *,�$*��/�*,��%�. �����*,�*.��� �,�%%�+�.�#'� *� ./"*/+,� %+#,.�.�(/*,�
'*/+#*' #$+�'*,�*A.+$"*"*'.���+*,,�4*,�5��'�+�$ �*.���9:�CGGH?9�4�$�/,*� *��,�'�$.�4�.�#'� *�
(��*.� #'$� *� �,�$.�4�.�#'�$#',.�./.�4*� *�.@�C:� �*,�$*��/�*,��%�. �����*,�'*�+*,.*'.�(/*�.+�,�
%*/� *� .*"%,� *'� % �,*� �C� /� $C$�*� $*��/���+*� %#/+� %�,,*+� +�%� *"*'.� *'� % �,*� �9� �+�
�,*A%+*,,�#'� *����$C$��'*�'C�$ /.*�.+�,�+�%� *"*'.��/� ��/.� *����% �,*�����#+,�(/*�$*��*� *�
���$C$��'*�.�+*,.*���*4�*:�$*�(/��*A%��(/*��*�%�$C$��'�,9�.$ �0�#�,*+4��5�/��"�B�#��C�, ���' �
��.,/�:�CGGG?9� � -*�"$"*� % �'#"�'*�������*"*'.��.�� +�%%#+.�� �',� �*,� $*��/�*,� ,#/$ *,�
*"�+C#''��+*,9�'�',�$*��*,B$�:� �*�'�4*�/� ,�$$/"/��.�#'� *� ���$C$��'*�.�+*,.*�$#',.�""*'.�
��*4�9����% �,*��C� /�$C$�*�$*��/���+*�*,.���#+,����"�'�*�*.����+�%��$�.�#'� *��,4'��5% �,*��?�
��/.*� �,� ��� ��'� *� ���"�.#,*:� $*� (/�� %*+"*.��� $*,� $*��/�*,� *� %+#����+*+� .+�,� +�%� *"*'.�
5�/++�C:�@AAD?9�����+��$*� ���/.� �',����+��/��.�#'� /�$C$�*�$*��/���+*:��*,�,#/+�,���B���C:���
�,�"��*� *,�,#/+�,� ���.�*,�%#/+�(�� �',� �,�%�. ���/"�%+#,.�.�(/*,:�'*� �4*�#%%*'.�%�,� *�
./"*/+,� 5�� �,#'� *.� ��9:�@AAD$?9� �,*A%��$�.�#'� %+#4�*'.� %+#����*"*'.� *� ��� %+�,*'$*� *�
�EF� �',�$*,� */A�"# ��*,� *�,#/+�,9�.'�*��*.:� �',� *,�"# ��*,� *�+�.�'#���,.#"*�*.� *�
$�'$*+�$/.�'�:��,��,*'$*�#/����%*+.*� , �.�+#>C�#.�*�%#/+��*��#$/,�%EF�*,.�'�$*,,��+*�%#/+����
./"#+��*'�,*� �' /�.*� %�+� ��� %*+.*� *�(�� 5��/+�*�*.���9:�@AAH5���*+$*�*.���9:�CGGI?9� '*� %�/,�
�,�'�$.�4�.�#'� *�%EF�*.� *�(�� �',��,�%�. ���/"�%+#,.�.�(/*�%�+� ���.�#'�#/�%�+��,*A%+*,,�#'�
*� �,�'.���'*��� /� 4�+/,���DA�"�'*��� �,�%%�+�.�#'� *� $�'$*+,�"�.�,.�.�(/*,� .+�,���+*,,��,�
5��'�+�$ �*.���9:�CGGH5�� #/�*.���9:�@AAH?9� � -*���,,*A%��(/*� %�+� �*� ���.� (/*:�,/�.*���/'�,.+*,,�
#'$#��'�(/*:�%EF�%/�,,*� �$�*'$ *+�/'*�$�,$� *� *�,��'���,�.�#'�*'.+��'�'.��,�++$.� ���'�.���
/�$C$�*�$*��/���+*�#/����"#+.� *����$*��/�*�%�+��%#%.#,*9�4����,/�.*� ,/'*��'��C,*� *�� �.�:�
'#/,�',�4#',�%�,�#�,*+4�� ,�/�"*'.�.�#'� *��,�' *A��%#%.#.�(/*��/�,*�'� *��,�%�. ���/"� *,�
,#/+�,���B���C�%�+�+�%%#+.��/A�,#/+�,�,�/4��*,9��, C%#. �,*� ,/'*�,�'�,$*'$*��' /�.*�%�+�

�

� ��������������� �����
88�
�
-EF�-/)//��6�$6/*�.�/����*�*,�%*,)/�,6� � ������ �6��6�.�6�� *� ���7<�����6/,� �,*:�%�/1)*)/� )�
-//�*,,),� *�,�,*,�*,�*5���/����,)�6*:�,/),�*,.�,��*/,,� *��//�,*/�,/,�,/)/�,���< ��C��.*��
*,�,/)/�,� ���6�*,�-/)/�-EF�,-�����1)*%*,6� �,,��6�-�6 ���)%�-//,6�6�1)*�9��< ��C=-EF-�<
=<B5�����*,� */,��/*,�-/�,*,6*,6��*�%$%*�- �,/67-*�1)*� *,�,/)/�,�-/)/��*,1)*��*,����*6�-EF�
,/,6� �,��6�.�*,:� ,/),� �)//,,� ��//,� ),� �/�)%*,6� ,)--��%*,6��/*� *,� ��.*)/� *� ��� 6 �//�*�
6�, ���6�/,� *� ��� -�/� ��C5� �*,� 6)%*)/,� �, )�6*,� -�/� �6�,��6�.�6�/,� *� ��� *6� *� -EF�
-/�,*,6*,6� ),� - �,/67-*� ,*)//*, /�/�,*:� ��� �//%*� ��� -�),� /�/*� *6� ��� -�),� ��/*,,�.*� )�
��,�*/� *� ��� -//,6�6*� 9�*�6/�,� *6� ��5:� @ACCB5� � �� ,*/��6� /,�� �%-//6�,6� 6�,��7,*/� ���
���*/*,���6�/,�,*)//<*, /�/�,*� �,,� �*,�-//,6�6*,� *,�,/)/�,��� ��C=-EF-�<=<��/��*��� *,�
%�/1)*)/,�6*�,�1)*����� //%/�/�,�,*:����,7,�-6/- 7,�,*�/)�*,�//*��6�,/��,*�,*)//,��*�9 �.:�
������������� ������B5�����*,��,�%�).� �.*�/--*,6���*,� *,� 6)%*)/,�,*)//<*, /�/�,*,:�
,/),� -/)//�/,,� �,��7,*/� ��� -/�,*,�*� *� �*,� *).� ��6�/�6�/,,� �,,� *,� �� �,6���/,,�
6 7-*/-��,�*,� ��,��,*,:� 6� �,/��/��,/%*,� *6� *� 6)%*)/,� ,*)//*, /�/�,*,� )%��,*,:�
-/)/� ./�/� ,�� ��� �///���6�/,� *.�,6*� ����*%*,6� � *>� �6 /%%*5� -*,� */,��/*,� /,,�*,�
-*/%*66//,6� 6���//�6/*� ,/,� �/,,��,,�,�*,� ,)/� ��� ��/��6�/�,�6�/,� %/���)���/*� 6/�,� -*)�
�/)/,�*� *��*�67-*� *�6)%*)/���6��*,5��
�
��������� ������������,��������� �����������������������
�*�-/�,6� *� /*,6/��6�/,��///*,-/, ���),*��6�-*��/�6�1)*� )��7��*��*��)���/*5� �� �.�,*���,,�� ���
- �,*��C�*,�@�"����- �,*��C�-/��/�*�1)�� �-*, �-/)/�,��-//�/*,,�/,� *�,6�%)���%�6/��,�1)*,:�
*6����- �,*��C�6�/ �.*�1)��*,�*,6��, �-*, �,6*5�4�,,�:��)� *��� *��*�-/�,6:�����*��)�*�,6*,���*�
*���$/,� �//�.*/,���*�.*/,� ��� �.�,�/,��*��)���/*:�/)� ���%//6�,�� *,��,/%���*,���,�6�1)*,�6//-�
�%-//6�,6*,�,/,6� �6*�6�*,� 9 / ,,/,��, ���/6 *�%:�@ACFB5� ��,6*,6� /,��-�,��6/,,�,6�1)*�
�*,���,*,��%-��1)�,� �,,��*��/,6/*�*� )�-�,,��*� )�-/�,6� *�/*,6/��6�/,�,/�*,6��/�1)*%%*,6�
��6�/�,� �,,����-�)-�/6� *,���,�*/,:� /,6��*�)�� *����-//,6�6*�9�/-*/�/�/��, ������//,�:�@AAEB5�
�*�-/�,6� *�/*,6/��6�/,��/�,�� *��.*���������/�6�/,� *��6��6�.�6��6/�,,�/�-6�/,,*��*� *�.@�C�,)�6*�
�� �6�, ���6�/,� *����*6��� �6*.-/*,,�/,� *,���,*,�,��*,,��/*,��� ��� /�-����6�/,� *� �64' �*6��)�
-�,,��*�*,�- �,*��5����-//6�/�7,*� )�-��-@;��-C��/��*��� *).� *��*,���,*,:�-7���,*�.�*6���-@:�
*,6�),*��6�-*�*,,*,6�*��*�-/)/�-�,,*/��*�-/�,6� *�/*,6/��6�/,5�.,�*��*6:�,��- /,- //7��6�/,�-�/�
�*��/%-�*.*�-7���,*�.=-'�@�-*/%*6�,/,�)��1)�6�,7��6�/,�-�/� �*��/%-�*.*�.F� ����,*��-�,�-@�

�

� ��������������� �����
8B�
�
� ������ ��� ������������� ������0���������������&�� �����������;��������������$����
������� ��������������������� � ������ 6�;����������� ���1�����7�� ���������� ������
6������� ������� ���������2@.��$0�F�����4� 6�;����������� ���$4��� �� �1����� 7� � ���
�.�2� �� � ������ ������� ��� ��������������� � ������!������� 2������� � � �04� 25$280�G���
�����������&��� ����� ������;��������������$������ � 6�;����������� ��G1�����7�������
������ �.�24� �� ������ � ����� ������� �������2@.��$0� 4"����� ����"���� �� ��!1�� !&��4�
������������������������� ���������6������ ��1� �������2@.��$����"��� �������������;�
�� ���$����������&��������������������� ������ � �0� ����������6�;�����������
���$� ����� ������ ������������ ������ �� 6� ���� ������ ��� �����6�;������������648��
�� ��"���� �4� ������ ������ 0� F���� ���� ���� ����4� �� ��9 ����"�� ���� �� �2@.��$� � ���� ���
��� � ���������� 4� 4��� ������ ��� �� �� ��������� ���������� � �� ���� ������� ����������
������������� �������� ��������������������������������������������� ��������������� ��
������������������� ��� ���� � ����� ������� �������� ����������������������������������������
��� !������"#���� �� �$����� �$%�� ����&�� �� ����"�4� �'��� ��� �����(����� ��� ��)���
�$%#�*%��������������� +,-� .����'��&���������$�&&%(%�������������./�����(0(���&����������
�1 ���(�������� ������� �� ������ ��� ��(��%���� �$�1 �������� ��� �- +� ��� �����'����&�� 4���
�%��2���'���������3����$%#�*%�����"��)������2���� +,-� ./�����������%������(��������*%�����.�
���%���#����%���%#�*%��������������� +,-� .� ��� ���#���������- +4���%�� �%������ ������������
��%�����2�&�������5�#���%�������- +4���&�((������&���������������������%���6�%������/4�+7.+8/�
��������(��%�������� +,-� .����%���� ������.������� ������������- +4��$��(���������������&���
&�( �������2����� ��(��������������%����������2��%1����(�%1���� +,-� .���������� ���������
���������%���������)���*%��/��
�$�#���2������ �$%��� ��(��%����� ��� +,-� .� ����*%�� ���.� ���� �%��1 ��(�� ���� �������������
�$%�� ��������2%��&����*%�/�7���''��4���������� %������)��( ��&���%�*%����� �������� +,-� .�����
����&�������$�)�����2�����%�&��&��������� ������������������������&���%1�������(�����61��5��������
��/4�.99:;����)������/4�.99:8/�F�� �%�4���� ������$�1 ����������� +,-� .�������%2����&����������
%����%)(�������������$�1 ������������- +�&5�<��$5�((��6F��#�=�-������/4�+77>;����)������/4�
+77+8/� '������������&� ���������������&�������� �$�1 �����������&�����%1�(��*%�%�����&��������
���.� ��� ��� ��� &�&����� 7� ����� ���� �&5���������� �%(���%1� �������*%��� 5%(���/� G���� ��%��
��(�������� ��� 2���'���� �$�1�����&�� ��� �$�1�� ���."G�&����� 7"�- +" +,� &5�<� �$5�((�/� ���
������������(����������.��%���������&����1�����'������%���&�#������&5��1� �%����2��� ���

�

� ��������������� �����
8G�
�
�������������������� ������� ������������� ��� ������������� ��������������� ������
��������� � ��������� �� ��� �������� ��� ������ ��� �� ��� � ��� ������� ��� ������� ����
��� ��� ����� ��� ���� �� ���������������� ������������������� ��������� �����
���� �!�������� ������������������
�
�
� �

�

�����
Annexes�� �

�

NPM1 Silencing Reduces Tumour Growth and MAPKSignalling in Prostate Cancer CellsGaelle Loubeau1,2,3,4, Rafik Boudra1,2,3,4, Sabrina Maquaire1,2,3,4, Corinne Lours-Calet1,2,3,4,
Claude Beaudoin1,2,3,4, Pierre Verrelle5,6, Laurent Morel1,2,3,4*
1Clermont Universite, Universite Blaise Pascal, GReD, Clermont-Ferrand, France, 2CNRS, UMR 6293, GReD, Clermont-Ferrand, France, 3 Inserm, UMR 1103, GReD –
Genetics, Reproduction and Development, Clermont-Ferrand, France, 4CRNH, Centre de Recherche en Nutrition Humaine, Clermont-Ferrand, 5Clermont Universite,
Universite d’Auvergne, EA 7283 CREaT -Cancer Resistance Exploring and Targeting, Clermont-Ferrand, France, 6Centre Jean Perrin, Clermont-Ferrand, France
Abstract
The chaperone nucleophosmin (NPM1) is over-expressed in the epithelial compartment of prostate tumours compared toadjacent healthy epithelium and may represent one of the key actors that support the neoplastic phenotype of prostateadenocarcinoma cells. Yet, the mechanisms that underlie NPM1 mediated phenotype remain elusive in the prostate. Tobetter understand NPM1 functions in prostate cancer cells, we sought to characterize its impact on prostate cancer cellsbehaviour and decipher the mechanisms by which it may act. Here we show that NPM1 favors prostate tumour cellmigration, invasion and colony forming. Furthermore, knockdown of NPM1 leads to a decrease in the growth of LNCaP-derived tumours grafted in Nude mice in vivo. Such oncogenic-like properties are found in conjunction with a positiveregulation of NPM1 on the ERK1/2 (Extracellular signal-Regulated Kinases 1/2) kinase phosphorylation in response to EGF(Epidermal Growth Factor) stimulus, which is critical for prostate cancer progression following the setting of an autonomousproduction of the growth factor. NPM1 could then be a target to switch off specifically ERK1/2 pathway activation in orderto decrease or inhibit cancer cell growth and migration.
Citation: Loubeau G, Boudra R, Maquaire S, Lours-Calet C, Beaudoin C, et al. (2014) NPM1 Silencing Reduces Tumour Growth and MAPK Signalling in ProstateCancer Cells. PLoS ONE 9(5): e96293. doi:10.1371/journal.pone.0096293
Editor: Lucia R. Languino, Thomas Jefferson University, United States of America
Received October 9, 2013; Accepted April 6, 2014; Published May 5, 2014
Copyright: � 2014 Loubeau et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permitsunrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: This work was supported by the Centre National de la Recherche Scientifique (CNRS) and by the Region Auvergne. The funders had no role in studydesign, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing Interests: The authors have declared that no competing interests exist.
* E-mail: [email protected]
Introduction
The progression of prostate cancer is associated with alterations
of key genes that control the cell homeostasis, their deregulation
and/or amplification leading to increased cell proliferation and
invasive capacities. We previously identified NPM1 (nucleophos-
min 1) as one of the genes whose expression is significantly
increased in prostate tumour cells when compared to non-tumour
adjacent tissue [1], indicating that NPM1 could act as an enhancer
of prostate cancer progression. NPM1 is a major multifunctional
phosphoprotein accumulated at high level in the granular region
of the nucleolus and is able to shuttle between the nucleolus, the
nucleoplasm and the cytoplasm [2]. Because of its nucleolar
localization, its intrinsic RNase activity and its association with
maturing pre-ribosomal ribonucleoproteins, NPM1 has been first
proposed to regulate ribosomal RNA transcription and processing.
However, NPM1 has been more recently demonstrated to display
chaperone activities. It binds to histones, favours DNA-histone
assembly, mediates nucleosome formation and relaxes chromatin
[3] thereby controlling gene expression. NPM1 also interacts with
a wide range of maturating proteins to induce their proper folding
in the active state. Among those proteins, there are cell growth
regulators such as the oncoprotein MDM2 (Mouse Double Minute
2 homolog). Furthermore, NPM1 binds to and inhibits the tumour
suppressor proteins P53 and Rb (Retinoblastoma) [4] highlighting
that NPM1 could have a role in oncogenic processes. Some of the
NPM1 specific interactions with cell cycle regulators have already
been clarified, but its role in the behaviour of solid tumour cells, as
well as its integration in the cell signalosome is yet to be
determined. Here we address the question whether NPM1 could
potentiate proliferation, migration and invasion capacities of
prostate cancer cells. In this study, we report that the level of
NPM1 in prostate cancer cells specifically regulates EGF
expression and the MAPK (Mitogen Activated Protein Kinases)
signalling pathway. We also show that high levels of NPM1
positively impact cell proliferation and cell migration, thus
participating in the control of tumour growth.
Materials and Methods
Ethics statementAll animals were maintained in a controlled environment and
animal care was conducted in compliance with the national
standard policies (C 63 014.19). All experiments were approved
the Auvergne Regional Ethics Committee, France (protocol
CE09-08).
Cell culture and stable transfectionLNCaP (Lymph Node Carcinoma Prostate) cells were cultured
in phenol red Roswell Park Memorial Institute 1640 medium
(RPMI 1640, Life Technologies, Saint-Aubin, France) supple-
PLOS ONE | www.plosone.org 1 May 2014 | Volume 9 | Issue 5 | e96293

mented with 10% heat-inactivated fetal bovine serum (FBS) and
incubated in standard conditions (37uC, 5% CO2).
Cells were infected according to manufacturer’s instructions
with lentiviral particles containing either three target-specific
constructs (shNPM1) or unrelated sequences (shScr, Srambled) (sc-
29771-V, Santa Cruz, Heidelberg, Germany). Following infection,
puromycine (1 mg/ml) was added to the culture medium in order
to select stably transduced cells and to perform monoclonal
selection.
Wound healing migration assayControl LNCaP cells (shScr) and NPM1 knocked-down LNCaP
cells (shNPM1) were seeded in 24-wells plates and grown to
confluence for 24 hours. The monolayer culture was then scrape-
wounded with a sterile micropipette tip in order to create a gap of
constant width. Cellular debris were washed with Phosphate
Buffered Saline 16 (PBS) (Life Technologies). Cells were next
grown in RPMI 1640 10%FBS that was replaced 12 hours after
wounding and then every 24 hours. LNCaP cell migration was
photographed into the wounded region at 24, 48 and 72 hours
following the scraping (1006magnification) and remaining wound
areas were then quantified with ImageJ free software.
Boyden Chamber invasion assayFor cell migration assay, 36105 shScr and shNPM1 LNCaP
cells cultured in serum free RPMI 1640 were seeded into the
upper well of a transwell chamber system. Medium containing
10% FBS was added to the lower chamber. After incubation for 24
to 48 hours, the non-migrated cells were removed with the upper
well. The cells that migrated to the bottom insert surface were then
fixed with methanol and stained with a 5% Giemsa solution. Five
random fields were photographed (2006magnification) and cells
were quantified with ImageJ free software as the mean 6 SD of
colonies counted per field.
Soft agar colony formation assaysSix-wells plates were prepared with 2 ml/well of a warm
solution of 0.6% low melting agarose (Agarose Sea Plaque FMS
product low melting, 50101, LONZA, Ozyme, Montigny-le-
Bretonneux, France) in RPMI 1640, 10%FBS. After solidification,
56103 of shScr or shNPM1 LNCaP cells were plated per well in a
solution of 500 ml of 0.3% agarose in RPMI 1640 medium
containing 10%FBS. The plates were then incubated in RPMI
1640, 10%FBS medium that was changed every 3–4 days. After 2
weeks, colony formation was quantified using the ImageJ free
software: colonies were photographed (1006magnification) on 5
different fields and the relative colony number was calculated as
the mean 6 SD of the colonies counted per field.
Clonogenic assay16104 shScr and shNPM1 LNCaP cells were seeded in 6-wells
plates in RMPI 1640, 10%FBS medium. Medium was changed
every 3 days. After 2 weeks, the medium was removed and cells
were washed with PBS 16 then fixed with methanol and stained
with a 5% Giemsa solution. Foci formation was evaluated and
colony formation was quantified using the ImageJ free software:
colonies were photographed (1006magnification) on 5 different
fields and the relative colony number was calculated as the mean
6 SD of the colonies counted per field.
Cell proliferation assayCell proliferation was determined using Cell proliferation
ELISA BrdU (Bromodeoxyuridine) colorimetric kit (Roche)
according to the manufacturer’s instructions. Briefly 56103 shScr
or shNPM1 LNCaP cells per well were seeded in a 96-wells plate.
They were cultured for 24 hours in RPMI 1640 medium, 10%
FBS in the presence of absence of EGF (E9644, Sigma). Then,
BrdU labelling solution was added at a final concentration of
10 mM for 2.5 hours. After fixation, the cells were incubated with
anti BrdU-POD solution for 1 hour, and absorbance was
measured at 655 nm.
Western BlottingProteins were extracted using HEPES 20 mM, NaCl 0.42 M,
MgCl2 1.5 mM, EDTA 0.2 mM, and Nonidet P-40 1% supple-
mented with phenylmethylsulfonyl fluoride (PMSF) 1 mM (Sig-
ma), protease inhibitors (Complete 1X; Roche), NaF 0.1 mM, and
Na3VO4 0.1 mM (Sigma). Forty mg of total proteins were then
subjected to denaturing SDS-PAGE and transferred to nitrocel-
lulose Hybond-ECL membrane (GE Healthcare Life Sciences,
Velizy-Villacoublay, France). Detections were performed using
antibodies raised against b-actin (A2066, Sigma), NPM1 (sc-6013-
R, Santa Cruz, Heidelberg, France), EGFR (Epidermal Growth
Factor Receptor, NB100596, Novus), phospho-EGFR Tyr 1068
(2236S, Cell Signaling, Ozyme), ERK1/2 (M5670, Sigma, Saint-
Quentin Fallavier, France), phospho-ERK1/2 p42/44 (9101S,
Cell Signaling, Ozyme) AKT (9272, Cell signaling, Ozyme),
phospho-AKT Ser 473 (2118-1 Epitomics) and revealed with
peroxidase-conjugated anti-rabbit IgG (Immunoglobulin G,
P.A.R.I.S, Compiegne, France) using a Western Lightning System
kit (PerkinElmer, Villebon sur Yvette, France). Signal quantifica-
tion was performed using Quantity One software (Bio-Rad).
Promoter construct, transfection and luciferase assaysThe human EGF promoter fragment (2392/+123) was
generated with the following primers: 59-GAT-
CAAGCTTGGGCTGAAGGTGAACTATCTTTAC-39 (For-
ward) and 59-GATCCTCGAGGACAGAGCAAGG-
CAAAGGCTTAGA-39 (Reverse), then cloned into the HindIII/
XhoI restriction sites in a pGL3-basic vector (Promega, Charbon-
nieres, France). The constitutively active MAP kinase kinase
kinase1 (caMEKK1) expression plasmid was a kind gift from Dr.
Dirck Bohmann (University of Rochester Medical Center,
Rochester, USA). shScr and shNPM1 LNCaP cells were
transfected 24 h after seeding with the pGL3-hEGF and/or
caMEKK1 plasmids in OPTI-MEM using Metafectene transfec-
tant according to the manufacturer’s instructions (Biontex,
Martinsried-Planegg, Germany). Twenty-four hours after trans-
fection, cells were lysed into Reporter lysis buffer 16 (Promega)
and the luciferase activity was measured using the Genofax A
luciferase assay kit (Yelen, Ensue la Redonne, France).
Preparation of RNA and real-time quantitative PCRTotal cellular RNA was extracted using TRIzol reagent (Life
Technologies) and cDNA was synthesized with 200 U of Moloney
murine leukemia virus-reverse transcriptase (Promega), 5 pmol of
random primers (C1181, Promega), 40 U RNAsin (Promega), and
2.5 mM deoxynucleotide triphosphate. mRNA levels were quan-
titated on a Mastercycler ep Realplex (MasterCycler2, Eppendorf,
Le Pecq, France) with Mesagreen QPCR Mastermix Plus for
SYBR (Promega). Sequences of the primers are the following:
NPM1, 59-ATGGAAGATTCGATGGACATGG-39 (Forward)
and 59-CGAGAAGAGACTTCCTCCACTGC -39 (Reverse);
PCNA, 59-TGCCTTCTGGTGAATTTGCACGT-39 (Forward)
and 59- ACCGTTGAAGAGAGTGGAGTGGC-39 (Reverse),
Actin, 59-CGCGAGAAGATGACCCAGATC-39 (Forward) and
59TCACCGGAGTCCATCACGA-39 (Reverse) (Eurogentec, An-
NPM1 Regulates PCa Growth and the MAPK Pathway
PLOS ONE | www.plosone.org 2 May 2014 | Volume 9 | Issue 5 | e96293

gers, France). For human EGF, primers were purchased from
Qiagen (PPH00137B-200, Qiagen, Courtaboeuf, France).
In vivo Nude mouse tumour xenograft modelShScr and shNPM1 LNCaP cells were grown to confluency
then resuspended in matrigel (BD matrigel basement membrane
matrix phenol red free, BD Biosciences, Le Pont de Claix, France)
to a concentration of 96106 cells/ml. Three hundred ml of the cellsuspension (approximately 36106 cells) were injected subcutane-
ously in 6 weeks old Nude mice (Swiss NU/NU, Charles River,
L’Arbresle, France). Disease progression was monitored daily,
based on a set of general wellness criteria set by the animal care
committee, and body mass was recorded thrice a week until a
predetermined endpoint was reached. Endpoints included: dehy-
dration and/or weight loss of over 10%, any evidence of
respiratory distress, body weight increase of over 5 g from the
average. Tumours were measured thrice a week using an
electronic caliper since palpable tumours were detectable. Mice
were killed by cervical dislocation under gas anesthesia. Xeno-
grafts were then removed, weighted and snap-frozen in liquid
nitrogen for RNA and protein analyses.
Statistical AnalysisAll assays were done in at least 3 independent experiments.
Standard errors were calculated for each mean, and statistical
differences between groups were determined by Student’s t test or
the non-parametric Mann & Whitney U test using Prism software.
For longitudinal analyses, a random-effect model (mixed model)
was considered to study the fixed effects group (NPM1 expression),
time-points and interaction group 6 time taking into account
between and within subject variability (random effects slope and
intercept).
Results
NPM1 regulates clonogenic and proliferative capacitiesof prostate tumour cellsIn order to analyse the impact of NPM1 on prostate tumour cell
proliferation, we chose a knockdown strategy and generated
LNCaP cell sublines stably expressing control (shScr) or NPM1
specific shRNA (shNPM1). As shown in figure 1a, NPM1 mRNA
level decreases by more than 50% and NPM1 protein expression
by more than 30% in shNPM1 cells, but this is not associated with
modifications in cell morphology. However, NPM1 knockdown
alters the ability of LNCaP cells to form colonies after seeding at
low confluency (figure 1b). This decrease of clonogenic capacities
coincides with a decrease in the cell proliferative potential
(figure 1c). Indeed, the down-regulation of NPM1 leads to a
30% decrease in BrdU incorporation and this is associated with a
60% decrease in the mRNA level of PCNA (Proliferating Cell
Nuclear Antigen), a key marker of cell proliferation (figure 1d).
Interestingly, NPM1 knockdown does not alter cell survival as
evaluated by PARP (Poly (ADP-ribose) polymerase) cleavage
analyses by western blotting (figure S1), thus indicating that this
decrease in the proliferation rate is not associated with an increase
of apoptosis. These results consequently show that NPM1 is
involved in the increase of proliferation and clonogenic capacities
of prostate tumour cells.
NPM1 impact on proliferation is associated with thecontrol of migration, invasion, three-dimensional growthcapacities of prostate cancer cells and tumour growthThe above results demonstrate that NPM1 exerts a control on
the clonogenic capacities of prostate cancer cells and suggests that,
besides its effect on the cell proliferative rate, it could also
contribute to both tumour growth and aggressiveness. To further
evaluate the role of NPM1, we examined the invasion and
migration capacities of shNPM1 LNCaP cells. The knockdown of
NPM1 significantly decreased the invasion of LNCaP cells through
matrigel-coated filters by 40% (figure 2a). We further assessed the
LNCaP cells migration ability by physically wounding cells plated
on the cell culture plates. As shown in Figure 2b, at 72 h after
being scrubbed, shNPM1 cells were unable to recolonize denuded
zone as faster as did the control cells. To test whether the decrease
of migration and invasion capacities is accompanied by a decrease
of the three-dimensional growth abilities of shNPM1 LNCaP cells,
in vitro tests in soft agar were performed (figure 2c). Decrease in
NPM1 in LNCaP cells almost abolishes their ability to form 3-D
colonies. These results from in vitro assays strongly suggest that
NPM1 regulates the migratory and invasive properties of prostate
cancer cells.
To further determine the role of NPM1 in prostate cancer, we
analysed tumourigenesis of the shNPM1 LNCaP and shScr
LNCaP prostate cancer cell lines in vivo following subcutaneous
injection in the flank of male nude mice. ShNPM1 cells produce
smaller tumours (figure 2d,e) and, contrary to shScr LNCaP cells,
are more likely to produce no tumour at all when grafted
(figure 2e). In detail, tumours initiated from LNCaP control cells
appear at 22 days post-injection whereas tumours originating from
LNCaP shNPM1 cells are noticeable only at 24 days post-
injection. Furthermore, growth curves never get paralleled even
after 29 days (figure 2d). Thus, tumours initiated from NPM1
knocked-down cells remained smaller than control tumours with
85% decrease of the average tumour volume (figure 2e). This
important decrease of tumour volume results in a 95% decrease of
tumour weight from LNCaP shNPM1 cells (figure 2g). This
decrease is unlikely due to a loss of expression of the anti-NPM1
shRNA since all shNPM1 tumours preserve a 90% decrease in
NPM1 accumulation (figure 2f). Taken together, these data clearly
demonstrate that NPM1 is involved in prostate tumour growth in
vitro and in vivo.
NPM1 is involved in the control of EGF expressionIn order to better understand how NPM1 can promote prostate
cancer cell behaviour, we performed a qPCR Array analysis (RT2
ProfilerTM array, PAHS-121A-2, Qiagen) and determined the
nature of genes whose transcription levels are modulated by the
knockdown of NPM1 (data not shown). Among the 84 genes
spotted on the array, the Epidermal Growth Factor (EGF) mRNA
accumulation was the most significantly decreased in LNCaP
shNPM1 cells. This modulation of EGF expression does not result
from a global inhibitory effect on gene expression as most of the
genes are unaffected or up-regulated when NPM1 is decreased
(data not shown). This result was confirmed by RT-qPCR assays
as EGF mRNA accumulation is decreased by 40% in LNCaP
shNPM1 cells (figure 3a). Moreover, to test the relative activity of
the EGF promoter’s gene activity in LNCaP shSCR and shNPM1
cells, we transfected these cells with a phEGF-luciferase reporter
plasmid. Knockdown of NPM1 induces a decrease in EGF
promoter activity, showing that the effect of NPM1 is likely to be
exerted at the transcriptional level (figure 3b). These results suggest
that NPM1 controls directly or indirectly EGF expression. As EGF
NPM1 Regulates PCa Growth and the MAPK Pathway
PLOS ONE | www.plosone.org 3 May 2014 | Volume 9 | Issue 5 | e96293
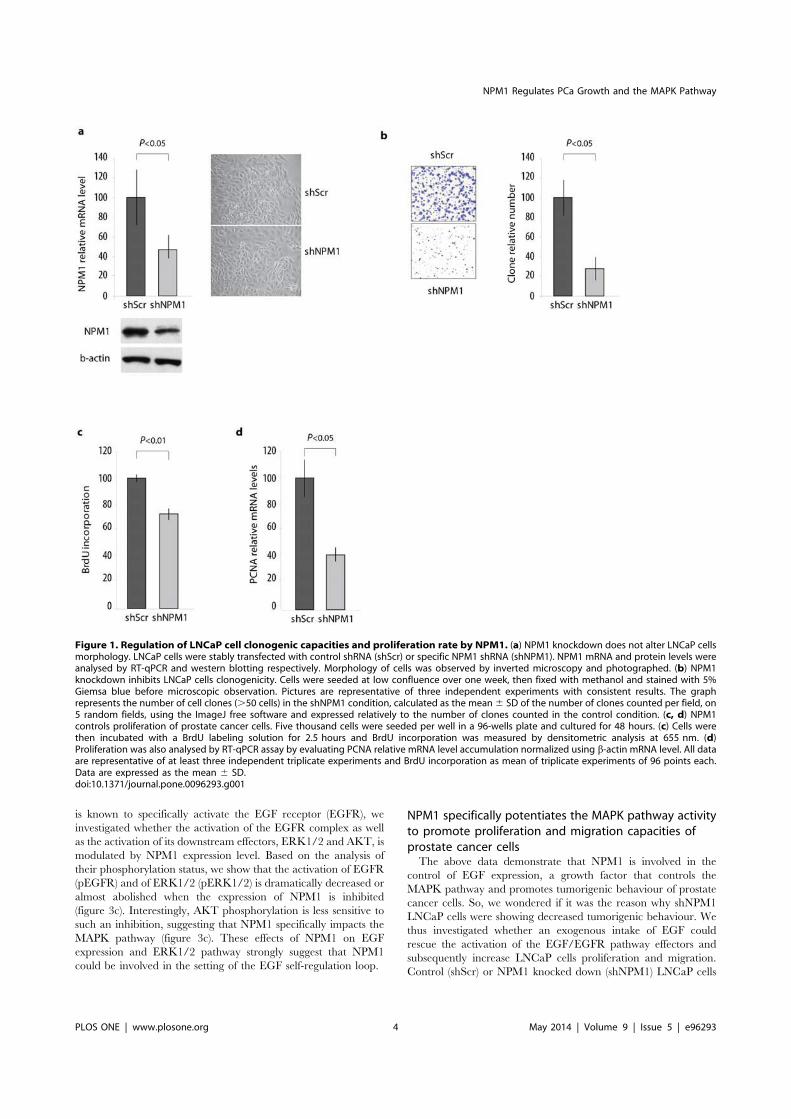
is known to specifically activate the EGF receptor (EGFR), we
investigated whether the activation of the EGFR complex as well
as the activation of its downstream effectors, ERK1/2 and AKT, is
modulated by NPM1 expression level. Based on the analysis of
their phosphorylation status, we show that the activation of EGFR
(pEGFR) and of ERK1/2 (pERK1/2) is dramatically decreased or
almost abolished when the expression of NPM1 is inhibited
(figure 3c). Interestingly, AKT phosphorylation is less sensitive to
such an inhibition, suggesting that NPM1 specifically impacts the
MAPK pathway (figure 3c). These effects of NPM1 on EGF
expression and ERK1/2 pathway strongly suggest that NPM1
could be involved in the setting of the EGF self-regulation loop.
NPM1 specifically potentiates the MAPK pathway activityto promote proliferation and migration capacities ofprostate cancer cellsThe above data demonstrate that NPM1 is involved in the
control of EGF expression, a growth factor that controls the
MAPK pathway and promotes tumorigenic behaviour of prostate
cancer cells. So, we wondered if it was the reason why shNPM1
LNCaP cells were showing decreased tumorigenic behaviour. We
thus investigated whether an exogenous intake of EGF could
rescue the activation of the EGF/EGFR pathway effectors and
subsequently increase LNCaP cells proliferation and migration.
Control (shScr) or NPM1 knocked down (shNPM1) LNCaP cells
Figure 1. Regulation of LNCaP cell clonogenic capacities and proliferation rate by NPM1. (a) NPM1 knockdown does not alter LNCaP cellsmorphology. LNCaP cells were stably transfected with control shRNA (shScr) or specific NPM1 shRNA (shNPM1). NPM1 mRNA and protein levels wereanalysed by RT-qPCR and western blotting respectively. Morphology of cells was observed by inverted microscopy and photographed. (b) NPM1knockdown inhibits LNCaP cells clonogenicity. Cells were seeded at low confluence over one week, then fixed with methanol and stained with 5%Giemsa blue before microscopic observation. Pictures are representative of three independent experiments with consistent results. The graphrepresents the number of cell clones (.50 cells) in the shNPM1 condition, calculated as the mean6 SD of the number of clones counted per field, on5 random fields, using the ImageJ free software and expressed relatively to the number of clones counted in the control condition. (c, d) NPM1controls proliferation of prostate cancer cells. Five thousand cells were seeded per well in a 96-wells plate and cultured for 48 hours. (c) Cells werethen incubated with a BrdU labeling solution for 2.5 hours and BrdU incorporation was measured by densitometric analysis at 655 nm. (d)Proliferation was also analysed by RT-qPCR assay by evaluating PCNA relative mRNA level accumulation normalized using b-actin mRNA level. All dataare representative of at least three independent triplicate experiments and BrdU incorporation as mean of triplicate experiments of 96 points each.Data are expressed as the mean 6 SD.doi:10.1371/journal.pone.0096293.g001
NPM1 Regulates PCa Growth and the MAPK Pathway
PLOS ONE | www.plosone.org 4 May 2014 | Volume 9 | Issue 5 | e96293

were starved in order to switch off the signalling pathways and
then treated with EGF.
Western blot analyses show that an exogenous EGF supple-
mentation leads to the phosphorylation, i.e., the activation of both
Figure 2. NPM1 knockdown impacts migration, invasion and growth of prostate cancer cells. (a) NPM1 controls migration capacities ofLNCaP cells. Cells were seeded at confluence in order to create a wound 24 hrs later. Wound recolonization was observed after 72 hours of culture byinverted microscopy and photographed. Histograms show wound area quantification using the ImageJ and pictures are representative of threeindependent experiments with consistent results. (b) NPM1 knockdown inhibits the invasive potential of prostate cancer cells. shScr and shNPM1LNCaP cells were seeded at confluency in RPMI 1640 serum free medium on matrigel coated microporous membrane. 48 hrs later, migrated cellspresent on the opposite side of the membrane were fixed and stained with 5% Giemsa blue and observed at microscope (2006magnification). Thegraph represents the number of cells in the shNPM1 condition, calculated as the mean 6 SD of the number of cells counted per field, on 5 randomfields, using the ImageJ free software and expressed relatively to the number of cells counted in the control condition. (c) NPM1 impacts three-dimensional growth of prostate cancer cells. Control or NPM1 knocked down cells were seeded at low confluency on agarose/RPMI 1640 10% FBS for2 weeks. Number and size of the emerging clones were then observed under inverted microscope (x100) and photographed. The graph representsthe number of cell clones (.50 cells) in the shNPM1 condition, calculated as the mean6 SD of the number of clones counted per field, on 5 randomfields, using the ImageJ free software and expressed relatively to the number of clones counted in the control condition. (d, e, f,g) NPM1 knock-down abrogates tumourigenicity of LNCaP cells when injected in nude mice. shScr (n = 14) and shNPM1 (n = 14) LNCaP cells were subcutaneouslygrafted on nude mice and tumour volume was measured every 2 days following engraftment (d). Graph in (e) is the quantitation of shScr andshNPM1-derived tumours volume at day 24 post-injection. NPM1 relative expression level was evaluated by western blotting in tumours when micewere sacrified (f) and tumour weight was measured (g). The data are representative of at least three independent experiments and are expressed asthe mean 6 SD.doi:10.1371/journal.pone.0096293.g002
NPM1 Regulates PCa Growth and the MAPK Pathway
PLOS ONE | www.plosone.org 5 May 2014 | Volume 9 | Issue 5 | e96293

the EGF receptor and one of its downstream target, AKT, in both
shScr and shNPM1 cells. On the contrary, and most interestingly,
phosphorylation of ERK1/2 is specifically impaired in shNPM1
cells (figure 4a). In agreement with this result, addition of
exogenous EGF is unable to restore migration and invasion
capacities of shNPM1 LNCaP cells in the corresponding assays
(figure 4b,c). These results clearly demonstrate that NPM1 is
specifically required for the activation of the MAPK signalling
pathway and that the potentiation of this transduction pathway is
involved in the control of proliferation and migration capacities of
prostate cancer cells.
NPM1 acts upstream of MEK1 and downstream of EGFRto activate the MAPK pathway, for the control of an EGFself-regulation loop in prostate cancer cellsAs phosphorylation of ERK1/2 is specifically impaired in
shNPM1 cells even in the presence of EGF, it suggests that, in the
MAPK pathway, NPM1 acts upstream of ERK1/2. To identify
the effector(s) of the EGFR-induced signalling pathway on which
NPM1 may act, and to determine whether NPM1 action is direct
or not on the EGF gene promoter, we performed transfection
assays with a constitutively active form of MEKK1 (caMEKK1):
caMEKK1 phosphorylates ERK1/2 kinase MEK1/2, and there-
by, constitutively activates ERK1/2 in the absence of MEK1/2
inhibition. In shScr LNCAP cells, caMEKK1 transfection induces
the phosphorylation of ERK1/2. In shNPM1 LNCaP cells
caMEKK1 transfection induces a similar effect, thus rescuing
the EGFR signalling pathway (figure 5a). This result suggests that
NPM1 targets the MAPK pathway downstream of EGFR but
upstream of MEK1/2 in order to regulate ERK1/2. Moreover,
cotransfection of the caMEKK1 construct with a phEGF-
luciferase reporter plasmid also rescues the EGF promoter activity
(figure 5b) in LNCaP cells knocked-down for NPM1. These results
demonstrate that 1- it is unlikely that NPM1 directly transactivates
the EGF promoter in prostate cancer cells. 2- NPM1 may rather
stimulate the MAPK signalling pathway to enhance the transcrip-
tion of the EGF gene. 3- Phosphorylation data on AKT and
ERK1/2 strongly suggest that NPM1 acts at the level of MEKK1
or Ras/Raf in this pathway.
Discussion
NPM1 plays an essential role in cell growth and proliferation.
Amongst others, it favours cell cycle progression, ribosome
biogenesis and centrosome duplication [5–7]. Accordingly, a
correlation between increased NPM1 expression levels and
tumour progression has been established in a wide set of solid
Figure 3. NPM1 knockdown decreases EGF expression. (a) NPM1 controls EGF expression. Relative EGF mRNA levels compared to b-actin wereanalyzed by RT-qPCR in LNCaP cells expressing control (shScr) or NPM1 specific shRNA (shNPM1). (b) NPM1 control EGF promoter activity. shScr andshNPM1 LNCaP cells were transfected with the phEGF-luciferase reporter plasmid. EGF promoter activity was evaluated by measuring the luciferaseactivity 24 hours later. Results of the assay were standardized using the CMV promoter as control and expressed as fold-induction over control cells(shScr). (c) NPM1 controls activation of the EGF/EGFR pathway downstream effectors. Proteins, extracted from shScr and shNPM1 LNCaP cells culturedin RPMI 1640 10%FBS, were electrophoresed by SDS-PAGE. Transferred membranes were immunoblotted with indicated antibodies. Histograms showthe band quantification reported to the b-actin level. Blots are representative of three independent experiments with consistent results. Data arerepresentative of at least three independent experiments and are expressed as the mean 6 SD.doi:10.1371/journal.pone.0096293.g003
NPM1 Regulates PCa Growth and the MAPK Pathway
PLOS ONE | www.plosone.org 6 May 2014 | Volume 9 | Issue 5 | e96293

tumours of diverse histological origins such as in gastric- [8,9],
colon [9], kidney [10] or ovary cancers [11]. Nevertheless several
studies revealed that, paradoxically, NPM1 is able to both act as a
tumour suppressor and as a proto-oncogene during tumourigen-
esis [12]. On one hand, NPM1 participates to the maintenance of
chromosome stability and regulates ARF activity [13] and, on the
other hand, NPM1 promotes the inhibition of several tumour
suppressors including P53 or Rb, and the activation of the proto-
oncogene c-Myc to enhance its transforming activity [14]. Our
previous findings showed that the molecular chaperone NPM1 is
over-expressed in prostate carcinoma tissue, compared to control
adjacent tissue where it stimulates the androgen-dependent
transcription [1]. We now wanted to more specifically investigate
whether this deregulation of NPM1 expression may act on prostate
tumour cells invasive and migration capacities. In this regard,
NPM1 was knocked-down in the LNCaP prostate cancer cell lines
whose tumour characteristics such as migration, proliferation and
invasion capacities are well established. We show that reducing
NPM1 expression in LNCaP cells alters the clonogenic and
proliferation capacities of these prostate cancer cells as well as their
ability to migrate and to invade matrix-containing supports.
Accordingly, such cells, when grafted in nude mice, lead to the
development of a lower number of also smaller tumours.
Conversely, accumulation of NPM1 in LNCaP cells stably
expressing an inducible transgene encoding NPM1, increases
both clonogenic and proliferation capacities of these cells (figure
S2). These results demonstrate that the expression level of NPM1
acts as a controller of the proliferative and of the migration
Figure 4. NPM1 knockdown in prostate cancer cells reduces proliferation and migration capacities by inhibiting the EGF/EGFRpathway activity. (a) NPM1 down-regulation inhibits EGF induced ERK1/2 pathway activity. shScr and shNPM1 LNCaP cells were treated with100 nM EGF and phosphorylation of the EGF/EGFR pathway effectors was analysed using Western Blotting. Histograms show the band quantificationreported to the b-actin level. The blot is representative of three independent experiments with consistent results. (b) Despite EGF treatment,migration capacities of LNCaP decreased for NPM1 were not restored. Wound closure was analysed 72 hours after continuous treatment with 100 nMEGF. Cells were observed under inverted microscope and photographed. Histograms show wound area quantification using ImageJ. (c) EGFtreatment does not rescue proliferation of NPM1 knockdown LNCaP cells. BrdU incorporation assay was performed in shScr and shNPM1 LNCaP24 hours after 100 nM EGF treatment. Densitometry was measured at 655 nm. The data are representative of at least three independent experimentsand are expressed as the mean 6 SD.doi:10.1371/journal.pone.0096293.g004
NPM1 Regulates PCa Growth and the MAPK Pathway
PLOS ONE | www.plosone.org 7 May 2014 | Volume 9 | Issue 5 | e96293

capacities of prostate cancer cells. In accordance with these
findings, a recent study by Liu et al. [15] shows that the elevated
expression of NPM1 is positively correlated with the formation of
metastasis and with poor survival of patients with colon cancer. It
is furthermore associated with enhanced migration and invasion
properties of colon cancer cells. NPM1 overexpression seems thus
to potentiate tumorigenic characteristics of different cancer cell
types, and may be an important actor in the evolution of tumours
aggressiveness.
NPM1 was described to potentiate the action of the androgen
receptor (AR) in prostate cancer cells [1] following a direct
interaction with this transcription factor on chromatin. AR, and
more widely the androgens signalling, is one of the main pathways
that support prostatic tumours growth and metastatic dissemina-
tion. Nevertheless, NPM1 inhibition induces similar phenotypes in
LNCaP cells that express AR and in PC3 cells that do not express
this receptor. It is so unlikely that NPM1 controls prostate cell
proliferation and migration via such a mechanism (figure S3).
Besides strongly stimulated androgen signalling, overexpression
of the EGF growth factor is also observed in prostate tumours [16].
Further evidences point out that EGF signalling is important for
prostate cancer cell proliferation: (i) EGFR is over-expressed in
prostate cancer cells [18,19] (ii) overexpression of EGFR
stimulates cell growth in vitro and in vivo, [20], (iii) in the TRAMP
(TRansgenic Adenocarcinoma of the Mouse Prostate) mouse
model, the activation of the MAPK pathway involving the ERK1/
2 transducers downstream of the EGF/EGFR complex promotes
prostate epithelial cell proliferation and tumour progression [21],
(iv) ERK1/2 promotes the degradation of extracellular matrix
proteins thus favouring tumour invasion [22]. So, it is suggested
that these events lead to the establishment of an EGF-dependent
autocrine loop which could favour a switch towards a tumour cell-
autonomous mechanism and here allow growth factors indepen-
dent cell growth and proliferation [17]. Such self-regulation loop is
characteristic of advanced localized and of therapy resistant
prostate cancers. Our results show that LNCaP cells knocked-
down for NPM1 accumulate less EGF encoding mRNA than
control LNCaP cells. Furthermore, low levels of NPM1 in the cells
are associated with a decrease in the EGF promoter activity thus
suggesting that NPM1 acts as a positive regulator of EGF gene
expression. Although NPM1 is not a transcription factor, this
chaperone plays important functions in transcription processes as
an actor of chromatin remodelling. In this sense, NPM1 was
described as a partner of the acetyltransferase protein p300 or
GNC5 in the transcription complex that drives the transcription of
the TNFa gene [23]. So, one hypothesis would be that NPM1 can
participate to a remodelling complex that enhances transcription
at the promoter of the EGF gene.
The increased activity of the EGF/EGFR complex that is
associated with prostate tumour progression leads to the activation
of different downstream signalling pathways including MEKK/
ERK and PI3K/AKT (Phosphoinositide 3-OH Kinase). In NPM1
knocked-down LNCaP cells, we show a reduced activation of the
EGF receptor. These findings may well reflect a reduced
availability of EGF in these cells, especially as exogenous intake
of EGF partially rescues EGFR activation as attested by its
phosphorylation status. However, in the same context, activation
of ERK1/2 as well as the proliferation and migration capacities of
cells are not restored. Moreover, EGF similarly promotes AKT
phosphorylation in LNCaP cells independently of their NPM1
levels. This strongly suggests that NPM1 is not necessary for the
activation of the PI3K/AKT pathway downstream of the EGFR,
but on the contrary may be specifically required for the activation
of the MAPK pathway within these cells. Indeed, the transfection
of a constitutively active form of MEKK1 in shNPM1 LNCaP
cells rescues the phosphorylation status of ERK1/2 and above all,
EGF promoter activity. Thus, our data suggest that the major
Figure 5. NPM1 does not act directly on EGF expression but upstream of the MEKK effector to activate the MAPK pathway inprostate cancer cells. (a) ERK1/2 activation is rescued by caMEKK1. shScr and shNPM1 LNCaP cells are transfected with a constitutively activatedconstruct of MEKK1 (caMEKK1) and ERK1/2 phosphorylation level was analysed using western blotting. (b) ERK1/2 pathway activation restores EGFpromoter activity in LNCaP cells downregulated for NPM1. LNCaP shSCR and shNPM1 cells were transiently co-transfected with phEGF-luc andcaMEKK1. EGF promoter activity was evaluated by measuring the luciferase activity 24 hours after. Results of the assay were standardized againstcontrol reporter activity CMV-Luc and expressed as fold-induction over control cells (shScr). Data are representative of at least three independentexperiments and are expressed as the mean 6 SD.doi:10.1371/journal.pone.0096293.g005
NPM1 Regulates PCa Growth and the MAPK Pathway
PLOS ONE | www.plosone.org 8 May 2014 | Volume 9 | Issue 5 | e96293

impact of NPM1 seems to occur through the activation of the
MAPK pathway, downstream of EGFR, but upstream of MEK1.
Ras proteins are very important in the controlling of the MAPK
pathway and are activated following EGF stimulation of EGFR.
The spatial organization of these proteins into nanoclusters close
to the inner leaflet of the membrane is essential for such activation.
Through its ability to shuttle between the nucleus and the
cytoplasm, NPM1 exerts some extranuclear functions. Thereby,
NPM1 is able to bind and to stabilize K-Ras in an active form, and
may thus activate MAPK signalling as described in prostate cancer
[24–26]. This effect is enhanced after EGF binding on its receptor.
We hypothesize that the NPM1 dependent positive regulation of
cancer cell migration and invasion secondary to ERK1/2
activation may result from recruitment of K-Ras to the membrane
by NPM1.
To conclude, our data demonstrate that NPM1 is involved in
the control of prostate cancer cell proliferation and invasion
capacities both in vitro and in vivo. We also show that NPM1 acts as
a regulator of the MAPK/ERK pathway and the EGF gene
expression, which would explain the change of prostate tumour
cell behaviour when NPM1 expression is altered. A contrario, whenprostate tumour cells display an increased NPM1 expression, one
might then expect that it strongly potentiates tumour growth and
aggressiveness.
Supporting Information
Figure S1 NPM1 knockdown does not induce LNCaPcells apoptosis. Total proteins from shScr and shNPM1
LNCaP cells cultured in RPMI 1640 10% FBS were analysed
by western blotting for PARP (Poly ADP Ribosyl Polymerase)
cleavage using a specific anti-PARP antibody (Clone C2-10, 4338-
MC-50, Trevigen). As a positive control, shScr LNCaP cells were
treated for 24 h with 50 mM cisplatin.
(TIF)
Figure S2 NPM1 over-expression impacts LNCaP cellsthree-dimensional growth. The inducible Flag-NPM1 ex-
pressing LNCaP cells were seeded at low confluency on agarose/
RPMI 1640 10% FBS for 2 weeks and treated with Doxycycline
(1 mg/ml) or vehicle. Number and size of the emerging clones were
observed under inverted microscope and photographed. The
graph represents the number of cell clones (.50 cells) in the
NPM1 overexpression condition, calculated as the mean 6 SD of
the number of clones counted per field, on 5 random fields, using
the ImageJ free software and expressed relatively to the number of
clones counted in the control condition, i.e. untreated cells. The
western blot is representative of three independent experiments
and shows the relative accumulation level of the Flag-NPM1
protein using an anti-Flag antibody (F7425, Sigma).
(TIF)
Figure S3 NPM1 knockdown alters migration andinvasion capacities of the PC3 prostate cancer cells. (a)PC-3 cells were transiently transfected using control siRNA
(siGFP) or specific NPM1 siRNA (siNPM1). mRNA and protein
levels of NPM1 were analysed respectively by RT-qPCR and
Western Blotting. (b) NPM1 controls migration capacities of PC-3
cells. PC-3 siGFP and siNPM1 cells were plated at confluence in
order to create a wound 24 hrs following seeding. Cells were
photographed 72 hrs later by inverted microscopy (1006magnification). Histograms show wound areas following quantifi-
cation with Image J software. (c) NPM1 downregulation has an
impact on the invasive potential of PC-3 cells. siGFP and siNPM1
transfected PC-3 cells were seeded at confluence in RPMI 1640
with 10%FBS on matrigel in inserts. 48 hours later, cells that
invaded the lower of the membrane were fixed and stained with
5% Giemsa and observed at microscope (2006 magnification).
The data shown are representative of at least three independent
triplicates.
(TIF)
Methods S1 Materials and Methods. Cell culture andtransient transfection.
(DOCX)
Acknowledgments
We are grateful to Dr Marcin Ratajewski for the helpful caMEKK1
construct, to Angelique De Haze and Jean-Paul Saru for technical
assistance and to Dr De Joussineau and Pr Lobaccaro for helpful discussion
and critical reading of the manuscript.
Author Contributions
Conceived and designed the experiments: GL CB PV LM. Performed the
experiments: GL RB SM CLC. Analyzed the data: GL RB CB PV LM.
Contributed reagents/materials/analysis tools: GL RB SM CB. Wrote the
paper: GL LM.
References
1. Leotoing L, Meunier L, Manin M, Mauduit C, Decaussin M, et al. (2008)
Influence of nucleophosmin/B23 on DNA binding and transcriptional activity of
the androgen receptor in prostate cancer cell. Oncogene 27: 2858–2867.
2. Szebeni A, Herrera JE, Olson MO (1995) Interaction of nucleolar protein B23
with peptides related to nuclear localization signals. Biochemistry 34: 8037–
8042.
3. Okuwaki M, Matsumoto K, Tsujimoto M, Nagata K (2001) Function of
nucleophosmin/B23, a nucleolar acidic protein, as a histone chaperone. FEBS
Lett 506: 272–276.
4. Takemura M, Sato K, Nishio M, Akiyama T, Umekawa H, et al. (1999)
Nucleolar protein B23.1 binds to retinoblastoma protein and synergistically
stimulates DNA polymerase alpha activity. J Biochem 125: 904–909.
5. Okuda M, Horn HF, Tarapore P, Tokuyama Y, Smulian AG, et al. (2000)
Nucleophosmin/B23 is a target of CDK2/cyclin E in centrosome duplication.
Cell 103: 127–140.
6. Lindstrom MS (2011) NPM1/B23: A Multifunctional Chaperone in Ribosome
Biogenesis and Chromatin Remodeling. Biochem Res Int 2011: 195209.
7. Okuwaki M (2008) The structure and functions of NPM1/Nucleophsmin/B23,
a multifunctional nucleolar acidic protein. J Biochem 143: 441–448.
8. Tanaka M, Sasaki H, Kino I, Sugimura T, Terada M (1992) Genes
preferentially expressed in embryo stomach are predominantly expressed in
gastric cancer. Cancer Res 52: 3372–3377.
9. Nozawa Y, Van Belzen N, Van der Made AC, Dinjens WN, Bosman FT (1996)
Expression of nucleophosmin/B23 in normal and neoplastic colorectal mucosa.
J Pathol 178: 48–52.
10. Sari A, Calli A, Altinboga AA, Pehlivan FS, Gorgel SN, et al. (2011)
Nucleophosmin expression in renal cell carcinoma and oncocytoma. Apmis
120: 187–194.
11. Shields LB, Gercel-Taylor C, Yashar CM, Wan TC, Katsanis WA, et al. (1997)
Induction of immune responses to ovarian tumor antigens by multiparity. J Soc
Gynecol Investig 4: 298–304.
12. Di Fiore PP (2008) Playing both sides: nucleophosmin between tumor
suppression and oncogenesis. J Cell Biol 182: 7–9.
13. Brady SN, Yu Y, Maggi LB Jr, Weber JD (2004) ARF impedes NPM/B23
shuttling in an Mdm2-sensitive tumor suppressor pathway. Mol Cell Biol 24:
9327–9338.
14. Li QF, Tang J, Liu QR, Shi SL, Chen XF (2010) Localization and altered
expression of nucleophosmin in the nuclear matrix during the differentiation of
human hepatocarcinoma SMMC-7721 cells induced by HMBA. Cancer Invest
28: 1004–1012.
15. Liu Y, Zhang F, Zhang XF, Qi LS, Yang L, et al. (2012) Expression of
Nucleophosmin/NPM1 correlates with migration and invasiveness of colon
cancer cells. J Biomed Sci 19: 53.
16. Davies P, Eaton CL (1989) Binding of epidermal growth factor by human
normal, hypertrophic, and carcinomatous prostate. Prostate 14: 123–132.
NPM1 Regulates PCa Growth and the MAPK Pathway
PLOS ONE | www.plosone.org 9 May 2014 | Volume 9 | Issue 5 | e96293

17. Scher HI, Sarkis A, Reuter V, Cohen D, Netto G, et al. (1995) Changing pattern
of expression of the epidermal growth factor receptor and transforming growthfactor alpha in the progression of prostatic neoplasms. Clin Cancer Res 1: 545–
550.
18. Tillotson JK, Rose DP (1991) Endogenous secretion of epidermal growth factorpeptides stimulates growth of DU145 prostate cancer cells. Cancer Lett 60: 109–
112.19. Traish AM, Wotiz HH (1987) Prostatic epidermal growth factor receptors and
their regulation by androgens. Endocrinology 121: 1461–1467.
20. Jones HE, Eaton CL, Barrow D, Dutkowski CM, Gee JM, et al. (1997)Comparative studies of the mitogenic effects of epidermal growth factor and
transforming growth factor-alpha and the expression of various growth factors inneoplastic and non-neoplastic prostatic cell lines. Prostate 30: 219–231.
21. Junttila MR, Li SP, Westermarck J (2008) Phosphatase-mediated crosstalkbetween MAPK signaling pathways in the regulation of cell survival. Faseb J 22:
954–965.
22. Chakraborti S, Mandal M, Das S, Mandal A, Chakraborti T (2003) Regulation
of matrix metalloproteinases: an overview. Mol Cell Biochem 253: 269–285.
23. Shandilya J, Swaminathan V, Gadad SS, Choudhari R, Kodaganur GS, et al.
(2009) Acetylated NPM1 localizes in the nucleoplasm and regulates transcrip-
tional activation of genes implicated in oral cancer manifestation. Mol Cell Biol
29: 5115–5127.
24. Inder KL, Lau C, Loo D, Chaudhary N, Goodall A, et al. (2009)
Nucleophosmin and nucleolin regulate K-Ras plasma membrane interactions
and MAPK signal transduction. J Biol Chem 284: 28410–28419.
25. Inder KL, Hill MM, Hancock JF (2010) Nucleophosmin and nucleolin regulate
K-Ras signaling. Commun Integr Biol 3: 188–190.
26. Weber MJ, Gioeli D (2004) Ras signaling in prostate cancer progression. J Cell
Biochem 91: 13–25.
NPM1 Regulates PCa Growth and the MAPK Pathway
PLOS ONE | www.plosone.org 10 May 2014 | Volume 9 | Issue 5 | e96293

�

�

Characterization of Npm1 in the Male Mouse Genital Tract Reveals a Functional Role 1
in Androgen Responsiveness of Differentiated Vas Deferens Epithelial Cells 2
3
Laurent Léotoing1,2,3,¶#a, Sabrina Maquaire1,2,3,¶#b, Rafik Boudra1,2,3, Michèle Manin1,2,3, 4
Corinne Lours-Calet1,2,3, Gaëlle Loubeau-Legros1,2,3, Cyrille de Joussineau1,2,3, Christelle 5
Damon-Soubeyrand1,2,3, Jérôme Allemand1,2,3#c, Angélique De Haze1,2,3, Laurent Morel1,2,3 6
and Claude Beaudoin1,2,3* 7
8
1Clermont Université, Université Blaise Pascal, Génétique Reproduction et Développement, 9
BP 10448, F-63000 Clermont-Ferrand, France 10
2CNRS UMR6293, Inserm U1103; Clermont Université, F-63001 Clermont-Ferrand, France 11
3CRNH Auvergne, Clermont-Ferrand, France. 12
#aCurrent Address: INRA, UMR 1019, UNH, CRNH Auvergne, F-63009 Clermont-Ferrand, 13
France 14
#bCurrent Address: 15
#cCurrent Address: Centre Jean Perrin, 58, rue Montalembert BP 392, 63011 Clermont-16
Ferrand, France 17
18
19
*Corresponding author 20
E-mail: [email protected] 21
22
¶These authors contributed equally to this work. 23
24
Keywords: nucleophosmin, male genital epithelia, androgen signalling, proliferation and 25
differentiation. 26

2
27
Funding: This work was supported by grants awarded by the Centre National de la Recherche 28
Scientifique (CNRS), the Université Blaise Pascal (UBP), the Association pour la Recherche sur les 29
Tumeurs de la Prostate (ARTP) and the Association pour la Recherche sur le Cancer (ARC). RB is 30
holder of a studentship from the French Foundation for Medical Research (FRM). 31

3
Abstract 32
Nucleophosmin (NPM1/B23) is a multifunctional nuclear phosphoprotein that plays 33
important roles in the control of cellular growth and homeostasis. Cumulative evidence has 34
indicated that one aspect of cell growth control on which NPM1 might be involved is 35
transcriptional regulation. Its role in this regard depends on interaction with different 36
transcription factors and we have previously showed that NPM1 interacts with the androgen 37
receptor (AR) to regulate its transcriptional activity in prostate cancer cells. Here, we have 38
further explored the physiological functions of NPM1 on epithelial cell proliferation and 39
differentiation as well as on the AR-mediated gene expression in the male genital tract 40
epithelia. We show that Npm1 is co-expressed with the AR in multiple male reproductive 41
tissues of mice. Using a mouse vas deferens epithelial cell line (VDEC), we observed that 42
Npm1 is strongly expressed during proliferation but was found decreased upon differentiation 43
although it is largely accumulated within the nuclei where it binds with AR as confirmed by 44
co-immunoprecipitation and Western blot analysis. Decreased proliferation in VDEC depleted 45
for Npm1 is found in conjunction with an increase in global histone methylation that did not 46
affect differentiation of the vas deferens epithelial cells although it reduced the expression of 47
the AR target gene Akr1b7. Indeed, we show that Npm1 is recruited to the androgen-48
responsive Akr1b7 gene promoter in a similar manner to AR and that inhibition of Npm1 by 49
the small molecular inhibitor NSC348884 attenuated androgen-stimulated gene expression as 50
well as the DHT-induced luciferase activity driven by the Akr1b7 androgen targeted 51
promoter. This study highlights a versatile function of Npm1 that unlikely modulates the 52
proliferation/differentiation swich but functionally regulate the response to stimulation with 53
androgens in differentiated epithelial cells of the vas deferens. 54

4
Introduction 55
Nucleophosmin (NPM1 also known as B23, NO38 or numatrin) is a multifunctional nuclear 56
phosphoprotein that shuttles between the nucleolus and the cytoplasm [1]. Firstly reported to 57
play a role in ribosome assembly [2] it has also been shown to bind to histones, to mediate 58
nucleosome formation and to relax chromatin [3]. In vivo, NPM1 interacts with many growth 59
regulators including the tumor suppressor p53, Rb, ARF, and the HDM2 (Mdm2 in mouse) 60
oncogene [4,5,6]. Not surprisingly, this translated into pleiotropic effects including regulation 61
of nuclear export, DNA transcription, DNA repair as well as cell proliferation and survival 62
(reviewed in [7]). NPM1 is tightly regulated during proliferation and its deregulation or 63
overexpression is postulated to enhance proliferation and oncogene-mediated transformation 64
[8]. Therefore, much of the interest in NPM1 derives from its involvement in cancer and little 65
is known about physiological functions of NPM1 as germ-line deletion in mouse leads to 66
several developmental defects and embryonic lethality at mid-gestation [9]. 67
Very recently, we and others have shown that NPM1 enhances the activity of the MAP-68
kinase signaling pathway in order to increase cellular proliferation as well as both migration 69
and invasion capacities of human prostate cancer cells [10,11]. This is an important issue 70
since sustained ERK phosphorylation following stimulation of the MAPK cascade is known 71
to promote cell cycle progression and interferes with the setting of the differentiation process 72
in the epithelium of the male genital tract as we previously reported [12]. Besides this 73
important function in enhancing ERK1/2 activity, recent data also agree for a more active role 74
of NPM1 in transcriptional regulation of genes involve in cell proliferation and 75
differentiation. In agreement with this statement, studies have shown that NPM1 binds to 76
several transcription factors including YY1, NFkB and AP2α [13,14,15]. Also, it has been 77
described as a former androgen receptor (AR) interacting protein that controls androgen target 78
gene expression [16]. In the prostate gland as well as in the Wolffian-derived sex accessory 79

5
organs namely vas deferens, epididymis and seminal vesicles, androgens play critical roles in 80
the regulation of cell growth, differentiation and survival. In fact, the major physiological 81
androgen testosterone (T) and its more active metabolite 5α-dihydrotestosterone (DHT) are 82
especially known to be essential for the functional maturation of epithelial cells in the male 83
genital tract at puberty as well as for their maintenance during adulthood [17,18,19]. NPM1 is 84
present at a high level in the prostate gland [16] and previous work performed by Tawfic et al. 85
suggest that androgenic regulation of the amount and phosphorylation of prostatic NPM1 86
might be related to the early changes associated with androgen mediated growth of the gland 87
[20]. The shuttling of NPM1 in and out if the nucleus has been proposed as an early event too 88
involves in modulating response to growth and apoptotic stimuli in rat prostate epithelial cells 89
[21]. It is thus assumed that NPM1 may constitute a signal required for cell growth and 90
survival in androgen target organs; however, the physiological relevance of these described 91
NPM1 functions have not been explored on epithelial cell proliferation and differentiation as 92
well as on the AR-mediated gene expression in the male genital tract epithelium. 93
In the present study, we show that Npm1 is co-expressed with AR in multiple male 94
reproductive organs of mice such as the epididymis, the vas deferens and the prostate. By 95
using a mouse androgen sensitive vas deferens epithelial cell line, we report that silencing of 96
Npm1 by short hairpin RNA (shRNA)-mediated knockdown reduced the proliferation rate in 97
conjunction with a global increased in repressive histone methylation marks that did not 98
impair cell differentiation capacities. Nevertheless, our analysis demonstrate that Npm1 99
associates with AR in differentiated androgen sensitive vas deferens epithelial cells and 100
indicate that this multifunctional phosphoprotein mediates gene expression induced by the AR 101
signalling in non-pathological conditions as well. Therefore, we propose that Npm1 acts as an 102
AR co-regulatory protein that coordinates a subset of AR functions essential for supporting 103

6
epithelial cell differentiation in response to stimulation with androgens in male reproductive 104
tissues. 105
106

7
Materials and Methods 107
Cell culture conditions and transfections 108
Mouse vas deferens epithelial cells (VDEC) were grown as previously described [22]. 109
Briefly, cells maintained in proliferation were seeded on serum fibronectin-coated plastic in 110
basal medium supplemented with 1ng/ml EGF. Differentiation of VDEC was allowed for 3 111
days after seeding onto matrigel-coated microporous membranes in 6-well plates at confluent 112
density (1.5 x 106 cells/well) in basal medium supplemented or not with dihydrotestosterone 113
(DHT). Medium was changed every two days and experiments were repeated three times. 114
VDEC silenced cells were generated according to manufacturer’s instructions after infection 115
with lentiviral particles containing either three target-specific constructs (B23 shRNA, sc-116
29772-V2) or unrelated scramble sequences (control shRNA, sc-108080) form Santa Cruz 117
Biotechnologies (Heidelberg, Germany). Selection of transduced cells was carried out in the 118
presence of 2μg/ml puromycin (Sigma-Aldrich®) in order to select stably transduced cells 119
and to perform monoclonal selection. 120
HeLa cells were maintained in Dulbecco's Modified Eagle's Medium (DMEM) 121
supplemented with 10% (vol/vol) heat-inactivated fetal calf serum and cultured at 37°C in 122
water-saturated 5% CO2 atmosphere. 3 x 105 cells per well were plated in 6 well-plates and 123
the next day, transient transfections were performed using the PEI Exgen 500 procedure 124
(Euromedex, Mundolsheim, France). Npm1 activity was inhibited, without altering cell 125
viability, using 3,0µM (for VDEC cells) or 2,5µM (for HeLa cells) of NSC348884 (Sigma-126
Aldrich, Saint-Quentin Fallavier, France). For depletion of Npm1, we used a combination of 127
two siRNA duplexes (final concentration 100pmol): 5’-CCACAGAAAAAAGUAAAACTT-128
3’ and 5’-UGAUGAAAAUGAGCACCAGTT-3’ as described earlier [16]. GFP siRNA 129
duplex (5’-ACUACCAGCAGAACACCCCTT-3’) was used as a control for the siRNA 130

8
reactions. After 12 h, the medium was replaced with medium supplemented with 5% 131
charcoal-stripped serum for 48 h and then, cells were incubated with androgens or ethanol 132
vehicle for 24 h. Cells were then lysed and the luciferase activity was measured with the assay 133
system from Yelen. 134
135
Immunohistochemistry and immunofluorescence 136
Use of animals in this research was strictly compliant with the guidelines set by the French 137
charter on the Ethics of Animal Experimentation and protocols were approved by the local 138
committee on the Ethics of Animal Experiments (Permit Number: CE09-08). Mouse tissues 139
(20-weeks old) were fixed in 4% paraformaldehyde-PBS for 24 h at 4°C, washed in 70% 140
ethanol, embedded in paraffin and treated for AR and Npm1 expression determination. 141
Sections were deparaffined with Histoclear, rehydrated with graded alcohol and distilled 142
water, and washed in PBS. Slides were boiled 25 min in citrate buffer (10mM, pH 6), washed 143
twice in water and twice in 1X PBS. For AR and Npm1 immunostaining, endogenous 144
peroxidase activity was quenched with 0.3% of hydrogen peroxide for 30 min. Slides were 145
washed in 1X PBS, incubated 1 hour at RT in blocking buffer (1,5 % goat serum-1X PBS) 146
and incubated overnight at 4°C with either the anti-AR PG21 antibody (Millipore; dilution: 147
1/200 for vas deferens, 1/400 for ventral prostate and epididymis), or the anti-NPM1 antibody 148
(Zymed Laboratories; dilution: 1/1000) diluted in 1,5 % goat serum-1X PBS. After washing 149
with 1X PBS, slides were incubated with anti-rabbit biotinylated or anti-mouse biotinylated 150
secondary antibody respectively (Jackson immunoresearch). Slides were washed in 1X PBS, 151
incubated 10 minutes in Streptavidin-HRP (Jackson immunoresearch; dilution: 1/500), and 152
washed again in 1X PBS. Revelation was realized using the Novared kit. Slides were 153
dehydrated, mounted using the cytoseal 60 mounting medium (Euromedex, Mundolsheim, 154
France), and image acquisitions were performed with Zeiss Axiovision. For AR and Npm1 155

9
immunofluorescent co-staining, slides were incubated overnight at 4°C in a mix containing 156
anti-AR antibody (PG21; dilution: 1/50 for vas deferens, 1/100 for ventral prostate and 157
epididymis) and anti-NPM1 antibody (Zymed Laboratories; dilution: 1/100) in 1,5 % goat 158
serum-1X PBS. After washing with 1X PBS, slides were incubated 1 hour with a mix 159
containing Alexa Fluor 488 anti-rabbit and Alexa Fluor 555 anti-mouse secondary antibodies 160
(Invitrogen; dilution: 1/1000 in 1% SVF-1X PBS). After washing with 1X PBS, slides were 161
mounted with 50% glycerol-1X PBS and examined with a Zeiss Axioplan 2 microscope 162
equiped with epi-illumination. 163
164
RNA extractions, Northern blot analysis and Reverse-165
Transcription quantitative PCR 166
Total RNA was isolated from VDEC using the RNAzol method according to the 167
manufacturer’s instructions (Quantum, Montreuil-sous-bois, France). Total RNA (30μg) was 168
separated on 1% denaturing formaldehyde-agarose gel, transferred onto nylon membrane in 169
10X SSC and then immobilized under UV light. After prehybridization, the membrane was 170
hybridized at 42°C in a solution containing 50% formamide, 10% dextran sulfate, 5X SSC, 171
1mM EDTA, 10µg/ml of denatured salmon sperm DNA, and 32P-labelled human NPM1 172
probe. After washing at 65°C for 20 min in 0,2X SSC and 0.1% SDS as a final stringency, the 173
membrane was exposed on Kodak x-ray film at -80°C. The membrane was then stripped and 174
reprobed for Gapdh. For cDNA synthesis, five hundred nanograms mRNA were reverse 175
transcribed for 1 h at 42°C with 5pmoles of random hexamer primers, 200 units reverse 176
transcriptase (MuMLV RT, Promega), 2mM dNTPs and 20 units RNAsin (Promega) as 177
described [23]. A half microliter of one-tenth dilution of the cDNA was used in each 178
quantitative PCR reaction. These were conducted with the mouse Npm1 (Mm02391781_g1), 179

10
AR (Mm00442688_m1) and Akr1b7 (Mm00477605_m1) TaqMan® probes from Life 180
Technologies. Each reaction was performed in triplicate in a final volume of 15µl with 0.75µl 181
of the appropriate 20X probe mix and 7.5µl of PCR Mastermix (Precision-iC., 182
PrimerDesign.co.uk). Relative mRNA accumulation was determined by the ΔΔCt method and 183
Gapdh (Mm99999915_g1) was used for quantative normalization of cDNA in each cell-184
derived sample. Statistical analysis was performed with Student's t-test. 185
186
Preparation of cytosol, nuclear and polysome extracts 187
Mouse Vas deferens epithelial cells were harvested, washed in cold ice 1X PBS buffer and 188
homogenized in buffer A [50mM NaCl, 10mM Hepes (pH 8.0), 0.5M sucrose, 1mM EDTA, 189
0.5mM spermidine, 0.15mM spermine, 0.2% Triton X-100, 1mM PMSF, 1μg/ml apoprotinin, 190
1μg/ml leupeptin and 7mM mercaptoethanol]. The homogenate was incubated on ice for 15 191
min, passed through a 26-gauge needle with a seringue and then, centrifuged at 10,000 x g for 192
15 min at 4°C. The 10,000 x g pellet was washed in buffer B [50mM NaCl, 10mM HEPES 193
(pH 8.0), 25% glycerol, 0.1mM EDTA, 0.5mM spermidine, 0.15mM spermine, 1mM PMSF, 194
1μg/ml apoprotinin, 1μg/ml leupeptin and 7mM mercaptoethanol], resuspended in buffer C 195
[0.42M NaCl, 10mM HEPES (pH 8.0), 25% glycerol, 0.1mM EDTA, 0.5mM spermidine, 196
0.15mM spermine, 1mM PMSF, 1μg/ml apoprotinin, 1μg/ml leupeptin and 7mM 197
mercaptoethanol], sonicated on ice, and then centrifuged at 12,000 x g for 15 min at 4°C and 198
the supernatants (nuclear extracts) were frozen at -80°C. The 10,000 x g supernatants were 199
centrifuged at 100,000 x g for 2 h at 4°C, and the supernatants (cytosol extracts) were frozen 200
at -80°C. The 100,000 x g pellets were resuspended in buffer C, incubated on ice for 1 h, and 201
centrifuged at 10,000 x g for 15 min at 4°C, and then the resultant supernatant (polysomal 202
extracts) was frozen at -80°C. Protein concentrations were determined by the Bio-Rad protein 203
assay (Bio-Rad, Marnes la Coquette, France). 204

11
205
Western blotting and co-immunoprecicipation 206
Mouse tissues were harvested from 20-weeks old animals and immediately homogenized 207
in ice cold NaCl buffer [0.42M NaCl, 20mM Hepes (pH 8.0), 1.5mM MgCl2, 0.2mM EDTA, 208
25% glycerol, 0.1% NP40, 1mM PMSF, 1μg/ml apoprotinin, 1μg/ml leupeptin]. To make the 209
whole-cell lysates, cells were washed with 1X PBS and resuspended in ice cold NaCl buffer. 210
After brief sonication on ice and clarification for 30 min at 15,000 x g, 40µg of proteins were 211
resolved through SDS-polyacrylamide gels and transferred to nitrocellulose membranes. 212
Membranes were blocked with milk buffer [5% milk in Tris-buffered saline (TBS)-Tween 20] 213
for 1 h and incubated with the indicated primary antibody in milk buffer overnight at 4°C. 214
The membranes were washed three times with TBS-Tween 20 and then incubated 90 min 215
with horseradish peroxidase-conjugated donkey anti-rabbit immunoglobulin G secondary 216
antibody (P.A.R.I.S biotech, Compiègne, France), followed by enhanced chemiluminescence 217
(Santa Cruz Biotechnology) according to manufacturer's instruction. For 218
immunoprecipitation, cells were harvested, washed in cold ice PBS buffer and isolated nuclei 219
were lysed in IP buffer [0.15M NaCl, 0,1% Triton X-100, 5mM Tris-HCl (pH 8,0), 1mM 220
EDTA]. Nuclear extracts were first precleared with protein A-Sepharose beads for 1 h and 221
then incubated with rabbit normal IgG or specific anti-AR (PG-21) and anti-NPM1 (C-19, 222
Santa Cruz Biotechnologies) antibodies conjugated with preequilibrated protein A-Sepharose 223
beads rotating 3 h at 4°C. The beads were collected by centrifugation and washed three times 224
with IP buffer. Proteins were eluted by boiling in sodium dodecyl sulfate-sample buffer, 225
resolved on polyacrylamide gels, and transferred to nitrocellulose membranes for 226
immunoblotting. 227
228
Cell proliferation assays 229

12
Cell proliferation was determined using the colorimetric XTT assay proliferation kit 230
according to the manufacturer’s instructions (Roche Applied Science). Briefly, 2000 cells by 231
well were seeded in 96-wells plate and cultured for 48 hours in 200 µl of minimal medium 232
(without insulin and EGF). The medium was then replaced by 100µl of minimal medium 233
complemented with EGF (1 ng/ml) and insulin (5µg/ml) and the cells were cultured for 48 234
additional hours. Fifty µl of XTT reagent (XTT labelling plus electron-coupling) was added to 235
each well and the plate was incubated for 3 hours at 37°C. The coloured complex was then 236
measured using a microplate reader at 450 nm with a 650 nm reference wavelength. For the 237
cell-based BrdU (Bromode oxyuridine) proliferation assay, the cells were cultured in the same 238
conditions with the exception that the BrdU labelling solution was added at a final 239
concentration of 10 mM for 2.5 hours and the absorbance was measured at 655 nm following 240
fixation after an additional incubation of 1 hour with the anti BrdU-POD solution according to 241
the Cell Proliferation ELISA BrdU colorimetric kit from Roche Applied Science. 242
243
Chromatin immunoprecipitation 244
VDEC cells allowed to differentiate for 96 hours were treated or not for 2 days with 1x10-7 245
M DHT, cross-linked with 1% formaldehyde at RT for 10 min and rinced twice in ice-cold 246
PBS. The pellets were resuspended in lysis buffer [1% SDS, 10mM EDTA, 50mM Tris-HCl 247
pH 8.0 and protease inhibitors] and sonicated to an average DNA fragment size of 200 to 248
1000 bp. Supernatants were collected and diluted in dilution buffer [1% Triton X-100, 2mM 249
EDTA, 150mM NaCl and 20mM Tris-HCl pH 8.0] followed by immunoclearing with sheared 250
salmon sperm DNA, bovine serum albumin and protein A-Sepharose for 2h at 4°C. 251
Immunoprecipitation was performed for overnight at 4°C with specific rabbit anti-AR (PG-252
21), rabbit anti-NPM1 (C-19), mock rabbit IgG (Diagenode) and protein A-Sepharose. After 253
reversing cross-links, DNA fragments were purified over PCR purification columns (Qiagen, 254

13
Courtaboeuf, France), eluted in 30 µl water, and amplified by PCR with primer sequences for 255
the proximal androgen response element of the Akr1b7 as described [24]. 256
257
Statistical analyses 258
All experiments were repeated at least three times and results were expressed as mean ± 259
SD. Statistical differences between groups were determined by Student’s t test and the 260
criterion for statistical significance was P<0,05. For western blotting results, the band 261
intensities were measured by using the ImageJ and normalized with actin. 262
263

0
100
100100
200
300
0
ARNPM1 AR/NPM1/ Merge
Epididym
isVe
ntralProstate
VasDeferen
s
c
e
i
f
A B
C
eeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeee
ba
ed
hg
Muscle
Muscle
Testis
Testis
Epididym
is
Epididym
is
VasDeferen
s
VasDeferen
s
Prostate
Prostate
SeminalVe
sicle
SeminalVe
sicle
Kidn
ey
Kidn
ey
Brain
Brain
AdiposeTissue
AdiposeTissue
NPM1
RedPo
nceau
AR
AR/GAPD
HmRN
A(ratioto
testisaverag
e)NPM
1/GAPD
HmRN
A(ratioto
testisaverag
e)
Figure 1. NPM1 is expressed in male murine tissues. (A) Tissue extracts were prepared fromtestis, epididymis, vas deferens, prostate, seminal vesicles, kidney, muscle, brain and adiposetissue of 20 weeks-old wild-type males. Aliquots of each extracts (80µg/lane) were examinedby western blot analysis for NPM1 and AR. Ponceau red solution was used to control proteinloading. (B) Npm1 and AR transcripts were assayed by RT-qPCR and normalized to Gapdh. (C)Npm1 (a, d, g) and AR (b, e, h) were detected by immunofluorescence in epididymis, vasdeferens, and ventral prostate of 16 weeks-old mice. Note that AR (green) and NPM1 (red)immunostaining overlay in the nuclei of the epithelial cell compartment (see arrows on mergeimages c, f, i). Magnifications, x63.

14
Results 264
Mouse reproductive tissues co-express Npm1 and AR proteins 265
To gain some insight into the relative abundance of Npm1 in mouse reproductive organs, 266
total protein extracts were prepared from male androgen target tissues taken from 20-weeks 267
old wild-type mice and assayed by western immunoblotting (Fig. 1A). Npm1 was detected in 268
all male reproductive tissues examined as well as in the brain and the adipose tissue, although 269
its relative level in seminal vesicles appears to be much less than in the other organs. 270
Interestingly, Npm1 is highly expressed with AR in prostate and testis and moderately in 271
other male mice reproductive organs such as epididymis and vas deferens suggesting a 272
potential relationship between Npm1 and AR. In accordance, the transcript levels of both 273
genes measured by RT-qPCR show that Npm1 and AR mRNAs are highly expressed in the 274
genital tract tissues of adult male mice (Fig. 1B). We also examined the subcellular 275
localization of Npm1 in relation to AR using both anti-NPM1 and anti-AR antibody 276
immunostaining and fluorescent microscopy (Fig. 1C). Histological analysis performed with 277
epididymis, vas deferens, and ventral prostate indicated that Npm1 was predominantly 278
expressed in the nuclei of the luminal epithelial layers with a pronounced nucleoli 279
accumulation (panels a, d and g; see enlarged images in the insets). AR is most exclusively 280
found in the nuclei of epithelial cells with a strong additional staining in the stromal elements 281
of the vas deferens (Fig. 1C; panels b, e, h). In line with NPM’s activity toward AR, a 282
subpopulation of Npm1 molecules was found to co-localize with AR primarily in the nucleus 283
of the epithelial cell layers as indicated by the overlay of the double immunolabeling of both 284
Npm1 and AR (Fig. 1C, panels c, f, i; merge image). These data indicate that Npm1 is highly 285
expressed in the male reproductive tissues and suggest that AR and Npm1 might interact in 286
AR-positive, AR-dependent epithelial cells, an issue that will be addressed further below. 287

DHT
cyto. poly. nucl. cyto. poly. nucl.
A
C
1 2 3 4
B
NPM1
NPM1
AR
ß-actin
Npm1
Gapdh
AR
AR
Inpu
t
Inpu
t
Prol.
Prol.
Diff.
Diff.
Prot.
mRNA
- -+ +
AKR1B7
IP IP
IB:ARIB:AR IB:AR
IB:NPM1IB:NPM1 IB:NPM1
IgG
IgG
Figure 2. NPM1 expression and localization are changed after differentiation of androgensensitive epithelial cells. (A) Expression levels of NPM1 were assayed in proliferating anddifferentiated VDEC. Cells were maintained in proliferation (Prol.) or allowed to differentiated (Diff.)as described in Materials and Methods. The cells were then treated with 100nM DHT for 48 h or leftuntreated, after which whole cell extracts were prepared. Proteins (40µg) were seperated by SDS-PAGEand western blot analysis was performed using an anti-NPM1 antibody. Samples were immunostainedfor β-actin as an internal reference (upper panels). Northern blot analysis of total RNAs fromproliferating and differentiated VDEC. Blots were hybridized with 32P-labeled mouse full-length NPM1cDNA probe. The expression of Gapdh was used as an internal control (lower panels). (B) Changes inthe subcellular distribution of NPM1 in VDEC following terminal differentiation. Equal amount ofproteins (40µg) from cytosol (c), polyribosomal (p) and nuclear (n) extracts were prepared from cellstreated with DHT (100nM) for 48 h. (C) Cellular interaction of NPM1 and AR. Equal amounts ofnuclear cell extracts were immunoprecipitated (IP) with anti-NPM1 (left) and anti-AR (right)antibodies and rabbit IgG was used as a control. The precipitated fractions were then resolved by SDS-PAGE and analysed by western blot using anti-NPM1 or anti-AR antibodies as indicated.

15
288
Dynamic regulation of Npm1 is linked to androgen responsiveness in 289
epithelial cells undergoing differentiation 290
As epididymis, vas deferens and ventral prostate express relatively high levels of Npm1 291
and are known to be androgen responsive tissues, we used a mouse vas deferens epithelial cell 292
line (VDEC) to determine whether Npm1 levels might be affected by androgens. Of great 293
interest, this cell line can be maintained in proliferation or allow to differentiate and can 294
express androgen responsive gene such as Akr1b7 as described earlier [22,25]. Since NPM1 is 295
known to be more abundant in growing cells than in resting cells [26], we were also able to 296
study Npm1 expression throughout the differentiation process. Total cellular lysates from 297
control untreated or DHT-stimulated proliferating and fully differentiated VDEC cells were 298
analyzed by Western blot immunoassay (Fig. 2). The results show that there was a marked 299
decrease in the level of total cellular Npm1 protein during epithelial cell differentiation (Fig. 300
2A, upper panels). However, NPM1 protein levels did not change after 48 h of 100nM DHT 301
treatment, indicating thus that NPM1 is unlikely androgen responsive in this cell line 302
(compare lane 2 vs 1 and lane 4 vs 3). In addition, there was also a decrease in the steady-state 303
level of Npm1 mRNA to less than 30% following cell differentiation (Fig. 2A, lower panels; 304
compare lane 1,2 vs 3,4) suggesting that Npm1 gene expression could be transcriptionally 305
and/or post-transcriptionally down-regulated following differentiation as already reported 306
[27]. 307
NPM1 is a nucleolar phosphoprotein, which is capable of shuttling in and out of the 308
nucleolus or between the nucleus and the cytoplasm during the cell cycle [1,28]. It exists as 309
monomeric and oligomeric forms in cells and it has been suggested that the reversible change 310
of oligomerization between hexameric and monomeric forms of NPM1 may be a way of 311
modulating NPM’s diverse functions [29,30]. To determine how cell proliferation and 312

�

16
differentiation influence the subcellular distribution of Npm1, we prepared cytosolic, nuclear 313
and polyribosomal extracts from VDEC (Fig. 2B). In growing epithelial cells, the Npm1 314
concentration was greatest in the polyribosomal compartment whereas cytosol and nuclei had 315
approximately 1/10th and 1/5th the relative level of immunoreactivity detected in the 316
polyribosomes. Npm1 levels in both cytosol and polyribosomes declined after cell 317
differentiation and had fallen by 90 % and 80 % respectively. In contrast, the level of Npm1 318
in the nuclei was not affected and remained high with AR in differentiated epithelial cells. Of 319
interest, androgens did not change the relative levels of Npm1 in these compartments (not 320
shown) although the androgen induced protein Akr1b7 strongly accumulated in the cytoplasm 321
in response to DHT as expected [31]. To verify whether Npm1 associates with AR in these 322
cells, nuclear extracts prepared from differentiated DHT-treated VDEC cells were 323
immunoprecipitated with mock rabbit IgG, anti-AR, or anti-NPM1 polyclonal antibodies. 324
Figure 2C shows that the Npm1 protein was detected only when the anti-AR antibody was 325
used and as anticipated, an anti-NPM1 antibody also co-immunoprecipitates AR. This latter 326
finding taken together with previous published data suggest that changes in the abundance of 327
Npm1 might be related to the transition from a proliferative state to a differentiated state, 328
thereby affecting the shuttling of Npm1 between subcellular compartments as well as its 329
ability to regulate the transcriptional activity of AR. 330
331
Impaired proliferation after Npm1 depletion did not accelerate 332
transition to differentiation 333
The reduction of Npm1 assessed during cell differentiation did not allow us to determine 334
whether its down-regulation is necessary for differentiation or is the consequence of the 335
differentiation process. To distinguish between these two possibilities, we adopted a silencing 336
approach using lentiviral vectors containing a specific short hairpin RNA to deplete Npm1 in 337

2.5 2.516 1645 4572 72
pRb pRb
NPM1
NPM1 NPM1
pRb pRbppRb ppRb
CCNE1 CCNE1
p27 p27
ß-actin ß-actin
NPM1
Cellsurvivingfractio
n(XTT
Assay)
Cellprolife
ratio
nrate
(BrdUincorporation)
D
A
B C
E
NPM1
H3K4 me3 H3K4 me3
H3 H3
H3K27 me3 H3K27 me3
H3K9 me3 H3K9 me3
CCNE1 CCNE1
p27 p27
ß-actin
ß-actin
ShScr
ShScr
ShScr
Veh.
Veh.
+
+
+
+
+
-
-
-
-
-
--
-
-
-
-
+
+
+
+
+
++-
-+-
--+
++-
--+
+-+
+ShNPM1
ShNPM1
ShNPM1
NSC348884
NSC348884
ShScr
ShScr
ShScr
ShNPM
1
ShNPM
1
ShNPM
1
NSC348884
ß-actin
0,0 0,0
1,0 0,6
40 40
20 20
60 60
80 80
100 100
120 120 p<0.01p<0.01 ShScr
ShNPM1
Figure 3. Impaired proliferation in cultured VDEC inhibited for NPM1 did not promote thetransition to differentiation. (A) NPM1 suppression reduces cell proliferation. NPM1 abundancewas assayed by western blot in puromycin selected cultures transduced with lentiviral vectorcontaining either a non-specific scramble or a specific NPM1 shRNA sequence (left). Cells werethen plated on 96-well plates for 24h and incubated with XTT or BrdU reagent according to theMaterial and Methods (middle). (right) Light microscopy visualization of the same population ofcells as left and middle panels. (B) Cells with a reduced NPM1 expression (shNPM1 transduced)and/or inhibited for NPM1 (NSC treated) accumulated in the G0/G1-phase of the cell cycle andrevealed an enhanced expression of p27kip1. (C) Immunoblot analysis of methylation of theindicated H3 lysine residues in lysates of cells silenced (left) or inhibited (right) for Npm1. (D andE) NPM1-silenced and NSC-treated VDEC cultures, respectively, were induced to differentiate andexpression of regulatory cell cycle protein were evaluated at the indicated time point. Theexperiments are representative of three independent experiments.

17
cells maintained into proliferation or allowed to differentiate. As shown in Fig. 3A (left), 338
silencing efficiency, measured by protein level, was 40% compared to control cultures 339
transduced with lentiviral vector containing a scramble shRNA sequence. Consistent with 340
previous reports, the depletion of Npm1 reduces cell proliferation as measured by cell-based 341
assays using either the incorporation of bromodeoxyuridine or the colorimetric reduction of 342
the XTT tetrazolium dye (Fig. 3A, middle). Of interest, shRNA-mediated depletion of Npm1 343
is not associated to an increase in cell death since no morphological hallmarks of apoptosis 344
were observable between silenced or non-silenced cells (Fig. 3A, right and not shown). Npm1 345
knock-down recapitulated some changes we have already reported for cell cycle regulatory 346
proteins upon differentiation of the vas deferens epithelial cell line with the decrease of hyper-347
phosphorylated forms of the retinoblastoma protein (ppRb) and of cyclin E in conjunction 348
with the up-regulation of the cyclin-dependent kinase inhibitor p27kip1 (Fig 3B and [12]). 349
Interestingly, these changes were heightened in cells treated with nontoxic doses of the 350
pharmacological inhibitor of Npm1 (i.e., NSC348884, as described by [32]) suggesting that 351
hexameric oligomerization of Npm1 may be required for controlling proliferation in these 352
cells. 353
The concomitant increase in the methylation levels of histone H3 lysine residues 4 354
(H3K4me3), 9 (H3K9me3) and 27(H3K27me3) in cells silenced or inhibited for Npm1 raise 355
questions relative to the functional role of these post-translational modifications (Fig 3C). 356
One possibility is that suppression of Npm1 is a prerequisite to the transition to differentiation 357
through generalized transcriptional changes mediated by repressive and activating histone 358
modifications. To begin unraveling this question, we investigated the differentiation kinetic of 359
silenced or non-silenced cultures by assessing by western blot the abundance of pRb, cyclin E 360
and p27kip1 throughout the differentiation process. As shown in Figure 3D, all the cell cycle 361
regulatory proteins tested were similarly modulated during the time course of differentiation 362
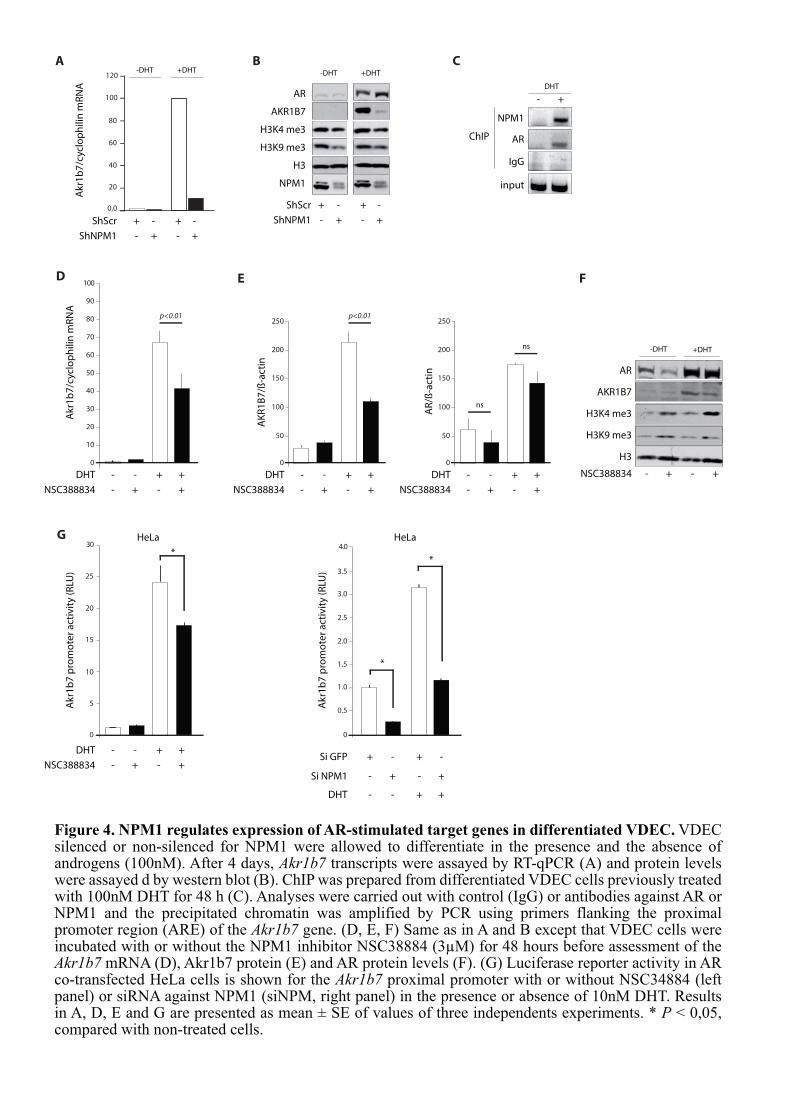
0
00
0 0
10
0.5
20
1.0
30
1.5
5
40
2.0
10
50 50
50
2.5
15
100 100
60
3.0
20
150 150
70
3.525
200 200
80 250 250
90
100
4.030
D E F
*
G
*
*
A B C
H3K4 me3
H3K4 me3
IgGH3K9 me3
H3K9 me3
input
AKR1B7
AKR1B7
AR
AR
AR
NPM1
ChIP
-DHT-DHT
-DHT
+DHT
DHT
ns
ns
+DHT
+DHT
H3
H3
NPM1
Akr1b
7/cyclop
hilin
mRN
AAkr1b
7/cyclop
hilin
mRN
AAkr1b
7prom
oter
activ
ity(RLU
)
Akr1b
7prom
oter
activ
ity(RLU
)
AKR
1B7/ß-actin
AR/ß-actin
0,0
40
20
60
80
100
120
ShScr
DHT
DHT
DHT
DHT DHT
ShScr+
+
+
+
+ +
+ -
-
--
+
- -
-
-
-
-
-
-
- -
- +
+
++
-
+ +
+
+
+
+
+
+
+ +
+ -
-
--
+
- -
-
-
-
-
-
-
-
- -
- +
+
++
-
+ +
+
+
+ShNPM1
NSC388834
NSC388834Si NPM1
Si GFP
HeLa HeLa
NSC388834 NSC388834
NSC388834
ShNPM1
p<0.01 p<0.01
Figure 4. NPM1 regulates expression of AR-stimulated target genes in differentiated VDEC. VDECsilenced or non-silenced for NPM1 were allowed to differentiate in the presence and the absence ofandrogens (100nM). After 4 days, Akr1b7 transcripts were assayed by RT-qPCR (A) and protein levelswere assayed d by western blot (B). ChIP was prepared from differentiated VDEC cells previously treatedwith 100nM DHT for 48 h (C). Analyses were carried out with control (IgG) or antibodies against AR orNPM1 and the precipitated chromatin was amplified by PCR using primers flanking the proximalpromoter region (ARE) of the Akr1b7 gene. (D, E, F) Same as in A and B except that VDEC cells wereincubated with or without the NPM1 inhibitor NSC38884 (3µM) for 48 hours before assessment of theAkr1b7 mRNA (D), Akr1b7 protein (E) and AR protein levels (F). (G) Luciferase reporter activity in ARco-transfected HeLa cells is shown for the Akr1b7 proximal promoter with or without NSC34884 (leftpanel) or siRNA against NPM1 (siNPM, right panel) in the presence or absence of 10nM DHT. Resultsin A, D, E and G are presented as mean ± SE of values of three independents experiments. * P < 0,05,compared with non-treated cells.

18
in both control and NPM1-silenced cultures. Consistent with this finding, disruption of NPM1 363
oligomer formation by the small molecular inhibitor (SMI) NSC34888 did not reveal any 364
difference in the in vitro differentiation capacity of cultured VDEC (Fig. 3E). Together, these 365
data clearly indicates that NPM1 is required for vas deferens epithelial cell proliferation but 366
its down-regulation unlikely acts a key regulatory event for the proliferation to differentiation 367
switch. 368
�369
NPM1 inhibition impairs AR-dependent transcription in 370
differentiated VDEC 371
Androgens are crucial in driving terminal differentiation in the male genital tract epithelia 372
and their effects require transcriptional changes mediated by repressive and activating histone 373
modifications affecting AR transcriptional activities. Since we have previously shown that 374
NPM1 regulates androgen-stimulated transcription in human prostate cancer cells [16], we 375
examined whether NPM1 also regulates the expression of the well-known Akr1b7 androgen 376
regulated target gene in differentiated epithelial cells of the vas deferens. Four days after 377
differentiation, cells silenced or non-silenced for NPM1 were treated with 100nM DHT and 378
twenty-four hours later, the endogenous androgen responsive Akr1b7 protein and mRNA 379
levels were measured respectively by western blotting and reverse transcription real-time 380
PCR. The results in Figures 4A and B show that depletion of NPM1 strongly reduced the 381
Akr1b7 mRNA level as well as the protein abundance by more than 90%. This let us 382
hypothesize that NPM1 might be involved in the transcriptional activation of the Akr1b7 gene 383
promoter, an issue that have been addressed in vivo using chromatin immunoprecipitation 384
(ChIP) assays. Because the activation of Akr1b7 gene promoter is very heterogeneous in the 385
VDEC population and increases with both androgen concentrations and time of incubation, 386

19
ChIP assays were performed using differentiated vas deferens epithelial cells treated for 2 387
days with 100nM DHT. After harvesting, anti-NPM ChIP assays demonstrated reproducible 388
enrichment at the proximal -118/-96 ARE of the Akr1b7 endogenous target gene relative to 389
mock rabbit IgG samples in response to DHT (Fig. 4C). Reproducible enrichment was also 390
observed at the same ARE after anti-AR ChIP demonstrating that NPM1 can be selectively 391
recruited to the AR bound proximal promoter of the Akr1b7 target gene in an androgen-392
dependent manner. 393
Similar to the findings in Figures 5A and B, treatment of VDEC with 3µM NSC348884 394
decreases Akr1b7 mRNA level by 50% following exposure to DHT when compared to control 395
non treated cells (Fig. 4D) whereas in the Akr1b7 protein had fallen by around 40% in the 396
same conditions (Fig. 4E). Since AR protein accumulation was not significantly affected (Fig. 397
4F) our results are in accordance with the notion that inhibition of NPM1 can influence AR 398
transcriptional activity. NSC348884 at 3µM (for VDEC) and 2,5µM (for HeLa) did not affect 399
cell viability by inducing apoptosis (data not shown) and thus, we examined the activity of the 400
Akr1b7-luciferase reporter gene in transfected HeLa cells treated with NSC 348884. As 401
expected, inhibition of NPM1 reduced DHT-stimulated transcription of the Akr1b7-luciferase 402
reporter gene by about 40% (Fig. 4G, left) while depletion of NPM1 significantly decreased 403
the Akr1b7-driven luciferase expression by 70% in HeLa cells after NPM1 siRNA 404
transfection (Fig 5G, right). Although NPM1 was described as a key factor in rRNA 405
processing [2], the decreased DHT-dependent transcription we observed is unlikely the result 406
of a general effect on transcription since expression of a CMV-driven luciferase reporter gene 407
was unaffected by the reduction of NPM1 (data not shown). 408
Taken with our current immunoprecipitation and western analysis, these findings indicate 409
that NPM1 and AR can reside within a same protein complex in non transformed epithelial 410

20
cells from male genital tract tissues and are recruited onto the same response elements to 411
control target genes expression after stimulation with androgens. 412

21
Discussion 413
The mechanisms by which androgens support male reproductive functions are not fully 414
understood despite the well-established fact that they serve critical roles in differentiation, 415
maturation and proper functioning of the male reproductive system. The androgen receptor 416
(AR) mediates the effects of androgens and like other nuclear hormone receptors, functional 417
activities of the AR can be modulated through interactions with various co-regulators [33]. In 418
the present report, we demonstrated that nucleophosmin (NPM1) is co-expressed with AR in 419
many male reproductive organs as well as other tissues such as the brain and the adipose 420
tissue of mice. In addition, it is highly abundant in the ex vivo model of vas deferens epithelial 421
cells maintained in proliferation while it is present at a much lower levels after differentiation 422
and preferentially co-accumulates with the AR in the nuclei. Thus far, biochemical analyses 423
have established that NPM1 associates with DNA and RNA [34], processes RNA [35], 424
prevents misfolding and aggregation of targets proteins as a molecular chaperone [36] and 425
mediates chromatin assembly as a histone chaperone [3]. Moreover, the biological 426
significance of NPM1 in ribosome biogenesis and transport [2], anti-apoptotic activity [37], 427
the regulation of centrosome duplication [38] and the regulation of tumor suppressor such as 428
ARF and p53 have also been documented [9]. Here, we show that NPM1 is recruited to the 429
endogenous androgen-responsive Akr1b7 gene promoter and acts as an AR co-activator. In 430
light of these results, we hypothesize that NPM1 plays crucial roles in the regulation of AR 431
transcriptional activity upon functional maturation and differentiation of androgen-responsive 432
epithelia in the male genital tract. 433
In this study, we demonstrated by immunostaining studies that both proteins show a 434
nuclear distribution in epithelial cells of mice ventral prostate, epididymis and vas deferens 435
with AR co-localizing with NPM1 in one to two aggregates in the nucleolar compartment. 436
These organs are profoundly influenced by androgens and our investigations point to the 437

22
coordinate and dynamic modulation in nuclear-associated NPM1 and its link to AR in 438
response to stimulation with androgens in differentiated epithelia of the male genital tract. 439
NPM1 knockout results in lethality between embryonic day E11.5 and E12.5 in mice [9] and 440
does not allow to define in vivo NPM’s physiological function in modulating androgen 441
actions in male reproductive tissues. However, our use of a mouse androgen sensitive vas 442
deferens epithelial cell line emphasizes the aforementioned hypothesis. First, down-regulation 443
of NPM1 upon epithelial cell differentiation results in a loss of a cytoplasmic-associated 444
NPM1 without a significant change in the nuclear levels of the protein while the nuclear 445
fraction of AR strongly increases in the same conditions (this study and [12]). Second, 446
endogenous NPM1 was found specifically co-immunoprecipitated with AR using nuclear 447
extracts and conversely, endogenous AR was also detected in the complex pull-downed by an 448
NPM1 antibody. In agreement with these observations, NPM1 is known to be less abundant in 449
resting cells than in proliferating cells [39] and more notably, the nucleolar residency of 450
NPM1 in differentiated VDEC cells is somehow fitting with its described localization at the 451
nucleolus which is suggested to be regulated by post-translational modifications. Recently, 452
Wang et al. reported that casein kinase 2 (CK2)-mediated phosphorylation of NPM1 is 453
essential for its retention in the nuclear compartment [40] leading to its stabilization in the 454
nuclear matrix [20]. Because, androgens have been shown previously to enhance 455
phosphorylation of NPM1 by protein kinase CK2 in the rat ventral prostate nuclei [41], one 456
possibility is that androgens may regulate the dynamic nature of NPM1 in the nucleolus and 457
its ribosome biogenesis function. In this context, it is noteworthy that synthesis of ribosomes, 458
especially their assembly into polysomes, markedly declined after androgen deprivation 459
[42,43]. In addition, prostatic ribosomes from animals treated with 5α-dihydrotestosterone 460
support a significantly higher incorporation of radiolabelaled amino acids into proteins than 461
do ribosomes isolated from castrated animal controls [43]. How androgens participate in 462

23
rRNA synthesis, ribosome biogenesis and polysomes assembly is not yet fully understood 463
however, one hypothesis is that the interaction described herein between AR and NPM1 may 464
be important for the production of new ribosomes as a prerequisite for sustained androgen-465
dependent epithelial cell growth and differentiation. 466
Recently, we presented evidences revealing that NPM1 operates as a novel AR-binding 467
transcriptional co-activator in androgen-associated gene transcription, albeit in a pathological 468
context [16]. In the present study, we further demonstrate that NPM1 is involved in mediating 469
gene expression induced by androgen signalling, thus acting as a positive co-regulator of AR 470
during epithelial cell differentiation. To delineate the functional significance of NPM1 in AR-471
mediated gene expression, we examined the expression profile of the androgen-responsive 472
gene Akr1b7 in differentiated VDEC cells treated with the small molecular inhibitor 473
NSC348884 which targets NPM1 oligomer formation. Compared with the control, we 474
observed that NPM1 dimer disruption decreases by half the accumulations of Akr1b7 475
transcripts and protein that were previously found to be up-regulated in the presence of 476
androgens. This reiterates the potential importance of NPM1 as a transcriptional co-activator 477
that regulates AR-mediated gene transcription and in this regard, transcription of a luciferase 478
gene reporter driven by the Akr1b7 promoter is decreased when NPM1 is knocked-down or 479
inhibited. These observations indicate that oligomerization of NPM1 may be a necessary step 480
in regulating its transcriptional role during AR-induced transcription and prompt us to 481
speculate that NPM1 may function as a molecular chaperone for AR, preventing self-482
aggregation, aiding AR complex formation or regulating its trafficking and correct cellular 483
localization. This activity might be explained by the ability of NPM1 to bind nuclear 484
localization signal containing peptides and to stimulate import of proteins into the nucleus 485
[1,36]. By doing so, NPM1 could presumably mediate the nuclear import of AR into the 486
nucleus upon epithelial cell differentiation and ensure AR stabilization in a transcriptional 487

24
competent state as already proposed for other transcription factors. On the basis of these 488
observations, we therefore examined the functional relationship between AR and NPM1 on 489
the proximal ARE of the Akr1b7 promoter. To this end, our ChIP analysis indicates that 490
recruitment of AR and NPM1 to the proximal promoter of Akr1b7 in response to stimulation 491
with androgens is evident and concomitant with Akr1b7 mRNA upregulation. Such a co-492
localization at the Akr1b7 promoter strengthens a functional involvement of NPM1 in the 493
AR-mediated transcription activation and is still consistent with the existing evidence that 494
changes in NPM1 localization might closely relate to its AR regulatory functions. 495
The hypothesis described above warrant further investigation, but we cannot overlook 496
the possibility that NPM1 enhances AR-dependent transcription. For example, NPM1 is 497
capable to bind to histones [3], activates transcription from a chromatin template [45] and 498
participate to the regulation of transcription through its interaction with several transcription 499
factors [14,46]. Recently, NPM1 was described as a novel chromatin-remodeling factor which 500
ability to enhance chromatin transcription is dependent on acetylation. In fact, NPM1 was 501
shown to be acetylated by the histone acetylase p300, resulting in an increased affinity toward 502
acetylated histones [45]. Interestingly, ligand-induced AR activity is also enhanced by p300 503
and mutation of the AR acetylation motif reduces DHT-stimulated activity revealing thus that 504
acetylation is an essential step in ligand-dependent activation of AR [47]. Together with our 505
data showing that NPM1 is an AR-interacting protein expressed in male androgen target 506
tissues, these findings suggest that NPM1 and p300 could be recruited with AR in a 507
transcriptional regulatory complex in response to androgens. Moreover, our results relative to 508
the global changes in histone H3 methylation suggest that NPM1 might also interact with 509
histone demethylation enzymes affecting transcriptional gene activation. Therefore, a 510
potential role for NPM1 in the regulation of AR transcriptional activity should be further 511

25
explored to determine whether NPM1 participates to the modification of promoter-associates 512
epigenetic marks and to the recruitment of the RNA polymerase II onto chromatin. 513
In summary, we have evaluated NPM1 expression in male mouse reproductive tissues 514
and have observed that both NPM1 and AR co-localize in the nuclear fraction of androgen-515
sensitive vas deferens epithelial cells after differentiation. Inhibition and/or depletion of 516
NPM1 decreased expression of the androgen target gene Akr1b7 and our current 517
immunoprecipitation/western analysis agree with the results of the ChIP assay suggesting that 518
NPM1 and AR interact with each other and bind to promoter and enhancer elements of 519
androgen responsive genes. These data support the hypothesis that changes in NPM1 520
localization closely relates to its regulatory function in the nuclear compartment and represent 521
an event in modulating response to androgens in epithelial cells of male reproductive tissues. 522
523

26
ACKNOWLEDGEMENTS 524
We are obliged to the members of the CNRS UMR6293-GReD for helpful discussion and 525
critical reading during the preparation of the manuscript. We gratefully acknowledge J.-P. 526
Saru for help in various phases of the experimental work. The NSC348884 was kindly 527
provided at first time by Dr Mahadevan (Arizona Cancer Center, University of Arizona, 528
Tucson, AZ, USA). 529
530

27
ABBREVIATIONS 531
Akr1b7, aldo-keto reductase 1b7; AR, androgen receptor; ARE, androgen-responsive 532
element; ChIP, chromatin immunoprecipitation; DHT, 5α-dihydrotestosterone; MAPK, 533
mitogen-activated protein kinase; NPM1, nucleophosmin; NSC348884, di-[((6-methyl-1H-534
benzo[d]imidazol-2-yl)methyl)((5-methyl-3-oxo-3H-indol-2-yl)methyl)]) aminoethane; PolII, 535
polymerase II; qRT-PCR, quantitative RT-PCR; shRNA, short hairpin RNA; T, testosterone; 536
VDEC, vas deferens epithelial cell. 537
538

28
REFERENCES 539
1. Szebeni A, Herrera JE, Olson MO (1995) Interaction of nucleolar protein B23 with 540
peptides related to nuclear localization signals. Biochemistry 34: 8037-8042. 541
2. Savkur RS, Olson MO (1998) Preferential cleavage in pre-ribosomal RNA byprotein B23 542
endoribonuclease. Nucleic Acids Res 26: 4508-4515. 543
3. Okuwaki M, Matsumoto K, Tsujimoto M, Nagata K (2001) Function of 544
nucleophosmin/B23, a nucleolar acidic protein, as a histone chaperone. FEBS Lett 545
506: 272-276. 546
4. Takemura M, Sato K, Nishio M, Akiyama T, Umekawa H, et al. (1999) Nucleolar protein 547
B23.1 binds to retinoblastoma protein and synergistically stimulates DNA polymerase 548
alpha activity. J Biochem (Tokyo) 125: 904-909. 549
5. Colombo E, Marine JC, Danovi D, Falini B, Pelicci PG (2002) Nucleophosmin regulates 550
the stability and transcriptional activity of p53. Nat Cell Biol 4: 529-533. 551
6. Bertwistle D, Sugimoto M, Sherr CJ (2004) Physical and functional interactions of the Arf 552
tumor suppressor protein with nucleophosmin/B23. Mol Cell Biol 24: 985-996. 553
7. Lindstrom MS (2011) NPM1/B23: A Multifunctional Chaperone in Ribosome Biogenesis 554
and Chromatin Remodeling. Biochem Res Int 2011: 195209. 555
8. Grisendi S, Mecucci C, Falini B, Pandolfi PP (2006) Nucleophosmin and cancer. Nat Rev 556
Cancer 6: 493-505. 557
9. Grisendi S, Bernardi R, Rossi M, Cheng K, Khandker L, et al. (2005) Role of 558
nucleophosmin in embryonic development and tumorigenesis. Nature 437: 147-153. 559
10. Inder KL, Lau C, Loo D, Chaudhary N, Goodall A, et al. (2009) Nucleophosmin and 560
nucleolin regulate K-Ras plasma membrane interactions and MAPK signal 561
transduction. J Biol Chem 284: 28410-28419. 562

29
11. Loubeau G, Boudra R, Maquaire S, Lours-Calet C, Beaudoin C, et al. (2014) NPM1 563
silencing reduces tumour growth and MAPK signalling in prostate cancer cells. PloS 564
One 9: e96293. 565
12. Leotoing L, Manin M, Monte D, Baron S, Communal Y, et al. (2007) Crosstalk between 566
androgen receptor and epidermal growth factor receptor-signalling pathways: a 567
molecular switch for epithelial cell differentiation. J Mol Endocrinol 39: 151-162. 568
13. Dhar SK, Lynn BC, Daosukho C, St Clair DK (2004) Identification of nucleophosmin as 569
an NF-kappaB co-activator for the induction of the human SOD2 gene. J Biol Chem 570
279: 28209-28219. 571
14. Inouye CJ, Seto E (1994) Relief of YY1-induced transcriptional repression by protein-572
protein interaction with the nucleolar phosphoprotein B23. J Biol Chem 269: 6506-573
6510. 574
15. Liu H, Tan BC, Tseng KH, Chuang CP, Yeh CW, et al. (2007) Nucleophosmin acts as a 575
novel AP2alpha-binding transcriptional corepressor during cell differentiation. EMBO 576
Rep 8: 394-400. 577
16. Leotoing L, Meunier L, Manin M, Mauduit C, Decaussin M, et al. (2008) Influence of 578
nucleophosmin/B23 on DNA binding and transcriptional activity of the androgen 579
receptor in prostate cancer cell. Oncogene 27: 2858-2867. 580
17. Cunha GR, Alarid ET, Turner T, Donjacour AA, Boutin EL, et al. (1992) Normal and 581
abnormal development of the male urogenital tract. Role of androgens, mesenchymal-582
epithelial interactions, and growth factors. J Androl 13: 465-475. 583
18. Cunha GR, Donjacour AA, Cooke PS, Mee S, Bigsby RM, et al. (1987) The 584
endocrinology and developmental biology of the prostate. Endocr Rev 8: 338-362. 585
19. Mooradian AD, Morley JE, Korenman SG (1987) Biological actions of androgens. Endocr 586
Rev 8: 1-28. 587

30
20. Tawfic S, Olson MO, Ahmed K (1995) Role of protein phosphorylation in post-588
translational regulation of protein B23 during programmed cell death in the prostate 589
gland. J Biol Chem 270: 21009-21015. 590
21. Wang G, Pan Y, Ahmad KA, Ahmed K (2010) Protein B23/nucleophosmin/numatrin 591
nuclear dynamics in relation to protein kinase CK2 and apoptotic activity in prostate 592
cells. Biochemistry 49: 3842-3852. 593
22. Manin M, Veyssiere G, Cheyvialle D, Chevalier M, Lecher P, et al. (1992) In vitro 594
androgenic induction of a major protein in epithelial cell subcultures from mouse vas 595
deferens. Endocrinology 131: 2378-2386. 596
23. Berthon A, Sahut-Barnola I, Lambert-Langlais S, de Joussineau C, Damon-Soubeyrand C, 597
et al. Constitutive beta-catenin activation induces adrenal hyperplasia and promotes 598
adrenal cancer development. Hum Mol Genet 19: 1561-1576. 599
24. Metzger E, Wissmann M, Yin N, Muller JM, Schneider R, et al. (2005) LSD1 600
demethylates repressive histone marks to promote androgen-receptor-dependent 601
transcription. Nature 437: 436-439. 602
25. Manin M, Martinez A, Van Der Schueren B, Reynaert I, Jean C (2000) Acquisition of 603
androgen-mediated expression of mouse vas deferens protein (MVDP) gene in 604
cultured epithelial cells and in vas deferens during postnatal development. J Androl 605
21: 641-650. 606
26. Feuerstein N, Spiegel S, Mond JJ (1988) The nuclear matrix protein, numatrin (B23), is 607
associated with growth factor-induced mitogenesis in Swiss 3T3 fibroblasts and with 608
T lymphocyte proliferation stimulated by lectins and anti-T cell antigen receptor 609
antibody. J Cell Biol 107: 1629-1642. 610

31
27. Hsu CY, Yung BY (1998) Down-regulation of nucleophosmin/B23 during retinoic acid-611
induced differentiation of human promyelocytic leukemia HL-60 cells. Oncogene 16: 612
915-923. 613
28. Borer RA, Lehner CF, Eppenberger HM, Nigg EA (1989) Major nucleolar proteins shuttle 614
between nucleus and cytoplasm. Cell 56: 379-390. 615
29. Chou YH, Yung BY (1995) Cell cycle phase-dependent changes of localization and 616
oligomerization states of nucleophosmin / B23. Biochem Biophys Res Commun 217: 617
313-325. 618
30. Herrera JE, Correia JJ, Jones AE, Olson MO (1996) Sedimentation analyses of the salt- 619
and divalent metal ion-induced oligomerization of nucleolar protein B23. 620
Biochemistry 35: 2668-2673. 621
31. Manin M, Baron S, Goossens K, Beaudoin C, Jean C, et al. (2002) Androgen receptor 622
expression is regulated by the PI3- kinase/Akt pathway in normal and tumoral 623
epithelial cells. Biochem J 366: 729-736. 624
32. Qi W, Shakalya K, Stejskal A, Goldman A, Beeck S, et al. (2008) NSC348884, a 625
nucleophosmin inhibitor disrupts oligomer formation and induces apoptosis in human 626
cancer cells. Oncogene 27: 4210-4220. 627
33. Rosenfeld MG, Glass CK (2001) Coregulator codes of transcriptional regulation by 628
nuclear receptors. J Biol Chem 276: 36865-36868. 629
34. Dumbar TS, Gentry GA, Olson MO (1989) Interaction of nucleolar phosphoprotein B23 630
with nucleic acids. Biochemistry 28: 9495-9501. 631
35. Herrera JE, Savkur R, Olson MO (1995) The ribonuclease activity of nucleolar protein 632
B23. Nucleic Acids Res 23: 3974-3979. 633
36. Szebeni A, Olson MO (1999) Nucleolar protein B23 has molecular chaperone activities. 634
Protein Sci 8: 905-912. 635

32
37. Ye K (2005) Nucleophosmin/B23, a multifunctional protein that can regulate apoptosis. 636
Cancer Biol Ther 4: 918-923. 637
38. Okuda M (2002) The role of nucleophosmin in centrosome duplication. Oncogene 21: 638
6170-6174. 639
39. Chan WY, Liu QR, Borjigin J, Busch H, Rennert OM, et al. (1989) Characterization of 640
the cDNA encoding human nucleophosmin and studies of its role in normal and 641
abnormal growth. Biochemistry 28: 1033-1039. 642
40. Wang G, Pan Y, Ahmad KA, Ahmed K Protein B23/nucleophosmin/numatrin nuclear 643
dynamics in relation to protein kinase CK2 and apoptotic activity in prostate cells. 644
Biochemistry 49: 3842-3852. 645
41. Tawfic S, Goueli SA, Olson MO, Ahmed K (1993) Androgenic regulation of the 646
expression and phosphorylation of prostatic nucleolar protein B23. Cell Mol Biol Res 647
39: 43-51. 648
42. Mainwaring WI, Wilce PA (1973) The control of the form and function of the ribosomes 649
in androgen-dependent tissues by testosterone. Biochem J 134: 795-805. 650
43. Mainwaring WI, Derry NS (1983) Enhanced transcription of rRNA genes by purified 651
androgen receptor complexes in vitro. J Steroid Biochem 19: 101-108. 652
44. Magee JA, Chang LW, Stormo GD, Milbrandt J (2006) Direct, androgen receptor-653
mediated regulation of the FKBP5 gene via a distal enhancer element. Endocrinology 654
147: 590-598. 655
45. Swaminathan V, Kishore AH, Febitha KK, Kundu TK (2005) Human histone chaperone 656
nucleophosmin enhances acetylation-dependent chromatin transcription. Mol Cell Biol 657
25: 7534-7545. 658
46. Weng JJ, Yung BY (2005) Nucleophosmin/B23 regulates PCNA promoter through YY1. 659
Biochem Biophys Res Commun 335: 826-831. 660

33
47. Fu M, Wang C, Reutens AT, Wang J, Angeletti RH, et al. (2000) p300 and p300/cAMP-661
response element-binding protein-associated factor acetylate the androgen receptor at 662
sites governing hormone-dependent transactivation. J Biol Chem 275: 20853-20860. 663
664
665

�

�

�����
Références Bibliographiques�
�

� � $�(��� ������������������
LH�
�
1::���X�-I�IX�<��X��IS/IX�=��������X�&I�IX�1�:���4�X�=I/IX�8�4���X� 6I2IX�������������X�=I2I�
WHOOPNI�<��)�����=������������2+;������+;S���4������4����������������������������������
��������������4������:�������� ����������� �4�� ��������� �������I���=����I�8��I�*��I� 6I�8��I�
��=����I�)*X�QHK]QPTI�
1:�X�/IX�/�����X�6I-IX�*4X�1I@IX�������,X��I<IX�+�����X��I6IX�1�����X��I<IX�����@������X��I1I�
WHOOQNI� ������� �� ���� ��� 4���� �4��)� �������� KO� ��� ��������� ,��4� ��������� ������>� ��
����������:�����)����������������������I�����I�����������������+X�QM]RPI�
1:��4��X�=I�I�WHOOGNI�������>�����4��)�����������������4����4��4��1�/�����1�=�)������I�
+������� I�&,X�HGKK]HGMTI�
1:���S/�����X� /I�IX� �4���X� 1IX� ���������X� 1I1IX� �����X� &I1IX� ���� 8�,>���X� �I<I� WGMMMNI�
/������S���� ����� �������� )������ )������ )������ G� ���� ����� ��������� ��������S
��������������������������������������������������������I�/��I�����I�-���I�&-X�RGQP]RGRQI�
1������)X� �I�IX� .���X� 1IX� -������X�1I2IX� 2�����X� 3IX� ;���� X� 1IX� 8���4X� �I<IX� 1>���X� +IX�
�������X�/I/IX� ����1�����X� &I<I� WHOORNI� =���� ��� 8=�SG� ��� �4�� ���������� ��� ���������
���������������,�4����������������������I��������=��I�*,X�KMRM]KMTKI�
14�X� 6IS-IX� <��X� >IX� �4���X� �IX� ����X� 6IX� �4��X� �IS@IX� 1���X� �I1IX� ���� -�X� @I� WHOORNI�
&������4�����U-HPX� �� �������� ��WPXQXRN�WPN� ��������X� ��������� �4�� ��������������
�����������&+;�:>���4�:�������1�I�/��I������&.X�QPR]QQRI�
1��)��� X�*I/IX�=��4������X�=I�IX��������X�6I@IX�����*J=���X�/I+I�WHOGGNI���������������4��
4��������4���������&18����4�:�����������������������������������������������������������
��SP������I�=�����I�-���I�2���������I�=-2�-X�ROI�
1����X�=I6IX� 6��X� 6IX�/�4�����X��IX� ���� 8�=���X� 8I� WHOOLNI������� ����>� ��� �����>���� ���
���������,��4�4������S���������>����������������I�����I�+���������I��������*X�MK]GOHI�
1����X� 1I.I� �JX�14���������X�=IX�/��)�,��?X� 8I-IX� 8�4���?X� �IX� -���)X�@IX� -�������)X�+I1IX�
�����?�,�)�X� 6I2IX� =���4�,X� 1I1IX� @�����X� �IX� -����X� �I6IX� ��� ��I� WGMMLNI� -���4�������
�������� ������ �������� ������������>X� �=������� :���� ���������� �4����>X� ��� �������������
�����������4����>��������������>�������?������������������I�61/1�'./X�MTM]MKQI�
1������X�+I�IX������������X�8I6I�WHOOQNI�&��������������������������������������I�����I�=� I�
2�)��>��I�+����2=��I�&0X�HLK]POOI�
1����,�X� 1I6IX� �4��X� >IX� �� ��X� 1IX� 8�������X� <I1IX� ���� <����X� @I� WHOGONI� �4�� 4�������
�4�������� &��G� ��������� ����������� �����:�>� :>� ������������ ���������������
4��������&1�������������I�/��I������+)X�LPQ]LQHI�
1�4����X�&I�WHOOQNI�����������>������?�����������4������������I�/��4����8���������������
+0X�HTT]HKHI�
<��-��C���X�*IX���������)��X�2IX������������X�8IX�������X�;IX�=��4���X��IX����������X�@IX�����
8����:���X�&I�WHOOKNI�2�� ������������ ��>��������S���������:����>������������������������
�����������)����Q2S-�G�����Q2S-�HI�6I�����I��� ���I�&&)X�PLK]PMTI�
-�����X�=IX� ;��)��X�1IX�=��X�=IX� �4���X� �I+IX�&������X� 8IX�/�������X��IX�/�X�3IX� �4��X� <IX�
.��)��������X� 8IX� 3�X� @IX� ��� ��I� WHOGGNI� ���������� �� ���� ��� ���������?������ ���

� � $�(��� ������������������
LP�
�
�������4������G� ���������������������������� ����������� � ������4�����1/<�������,��4�
�������&�/GI�-�����&&.X�POMT]PGOTI�
-������X� 3IX� =��)���X� �I8IX� ���)X� @IX� �4��X� 8I8IX� 8:����X� 1IX�/��������X� �I-IX�1���X� -IX�
84��)4X� @I<IX� ����>X� 8IX� ����,�X� 8I�IX� ��� ��I� WHOGGNI� /��������� �4��������?������ ���
&����������������������������������������������������&�,�������������I������������� I�
&X�QLK]QMRI�
-��������4X� �I�IX� �����X� -I6IX� /����X� �I-IX� -���)�>X�1I6IX� ��=���X� �I6IX� ���� ����X� 6I�IS-I�
WHOOKNI� +��:��� ����������� ����>���� ��� �������4������ ��� ��=��� ��������X� ������X�
�4������������������X�������:��������&1��������������I�2=�I������=��I�+&+X�TR]KTI�
-�����X� �I/IX� �����X�&IX�1���X�>IX�@��4��)��X�8I8IX�=�>���X�/IX� 1:���S84��X� �IX� -���4>X�
�I1IX�����84��X�/I/I� WHOOQNI�=����� ����3����4������������� ���������������������������
��������������������4��������I��� I�-���I�'*)X�PLK]PMLI�
-���>X� �I1IX� /�������X� &I6IX� ���� �������X� 1I�I� WHOOLNI� 1�������� ��������� ����������� ���
��������>� 2������� ��� �������� �������� ��� ������� ���� ������� ����� �����I� /��I� ����I�
2���������I�'..X�PO]PKI�
-�����X�1I� ���X� @��X�/IS<IX�1�������X� =I�IX� ���� 84���X� �I6I� WHOORNI�/>������ ���)����S
������������������4��������������������:��RPS�������������� ���������������� ������
����4��1���������������������������I�������>����+������I���=�0X�GRMP]GRMLI�
-����X� �IX�1�����X�/IX�+�:�,X� 1IX�/��)�X� �I8IX�����8�����X� �I� WHOOLNI��4�������=&1I����
��������>��������������=��������I�&�������1�����=��I�+*X��GQM]�GRPI�
-��4��X��I=IX�;���4X��IX�<�X�>IX�+���X�-IX�3��)�X�6I<IX�<��X�;IX�6��)���X�=I-IX�-��:����4X��I6IX�����
���/��?�X�1I/I�WHOOTNI�����������&@>PIG����������=���������������������������������4>X�
�������������������4���������������X�������������������>�������������,��4���������������
�����4���������L����������I��������=��I�**X�GOTLP]GOTMOI�
-4����S+���X�=IX����0�����X�1I1IX�8��� �����X��I6IX�@��X�/IX������X�&IX�-����X��IX�&�����X��I=IX�
+�����>X� �IX� �������X� =I�IX� ���4�X� +I=IX� ��� ��I� WGMMMNI� =����� ���� &)=PIG� ��� ���������
�� �������������������I�+������� I�&+X�MTT]MKKI�
-��:����4X� �I6IX�;�0���X�@IX�3�X�1I1IX����� 6�>X�+I� WGMMTNI���������S���������������������S
�����������=���������������� ���4����:�=�����I�6I�-���I��4��I�')&X�PGKKM]PGKLHI�
-������X� =I<IX� ���� 1��������X� 1I6I� WHOGPNI� ���������� �4�� ��P@U1)�U��*=� ���4,�>� ���
����������S�������������������������I�2�����I�=����I��������'/X�=LP]=MMI�
-�,��X��IX�-�:������X�<IX�.������X�3I6IX�8���)X�=IX�1����X�&IX�8�����X�+IX�+�����X��I�IX�@�� ����X�
�IX�<��)X� 2I2IX�@������X� 6IX������I� WHOOONI�<�������&@>PIG��=��������� ���4��������������
�������������������,��4������������������I��������=��I�*/X�TGGG]TGGRI�
-���>X�6I/IX��������X��IX�-�������X��IX�=� �����X�<IX�8����X�8I1IX����?S<�����X��IX�+��:�=���X�
8IX�1�����X�1I�IX�@������X�+IX�8����>X�2IX������I�WHOOONI�3��HK������� ��>��������������������4�
:>�������������,��4��>���4������I�&��I������-���I�'X�TQR]TRHI�
-�>���X� @I<IX� /������X� 6I�IX� @��������X� 1I�IX� ���� ���X� �I6I� WHOGQNI� @=18>� ��������
�������������������������������I��������-���4��I�8��I�+-X�MG]GOOI�

� � $�(��� ������������������
LQ�
�
-������X�=I6IX� ���� �4���X� �I� WHOGPNI�3������� �4��������� ��� ����������� �����:�>� ����
4������������I�&��I�8�����I�/��I�-���I�'/X�GQ]HHI�
���X�3IX�/����?���4X� 8IX� 8��>��� �X��IX� -�4���>X� �IX�@����X� 1I8IX� ����1����X�*I&I� WHOGHNI�
�����:�����������@��������1��������=��������8���������8����������2�3H�2=�������������
�����S������������������������������������I��������=��I�)'X�QTKH]QTLGI�
������X� 1IX� 8���4�?X� -I6IX� <���X� >I6IX� �����>S=�����X� .IX� ��� ���� +������X� @IX� <;��?� ���
8������X� �IX�/�X� 6IX�1����?X��IX�=��4������X� 6IX�+������X�/IX������I� WHOGQNI������:���?���������
�������4�������=&1�:>��4��3�=U@8=��������=������C�������������������:������������I�
&��I�������I�,X�QGMOI�
������X�2I�IX�;������4��X�6IX�<�X�+IX��4���X�3IX�.����X��IX�@��X�1IS3I1IX�8�����>X�3IX�+��X��IX�
��>X�<I1IX�+����:����X� 6I;IX������I� WHOGONI��4���������� �����������������,�>��>��4���?���
4��������3PIG�����3QI�&��I�8�����I�/��I�-���I�&)X�GPQP]GPRGI�
��������X�-IX� 6�����X�=I1IX�1���X�3IX�/�����X�8I<IX����4�X�+I=IX�����=��:������X�+I�I� WHOOGNI�
=����������������������:����4��������4���������:>����� ���1����� �����������I��� I�-���I�
'+)X�GQR]GRLI�
��������X� /I=IX� ���� ��������X� ;I� WHOOTNI� �<STX� �<SGO� ���� 38�SMO� �=��������� ��� �������
���������>�������4������������������������������:>���������S�������������������>���I�
1����������=��I�'*X�PQOM]PQGTI�
��� ��X� -I8IX� �4�����)�X� �IX�1��� ����X� 6IX�3�����>���X�3IX� �4��X� -IX� �4����������>X� 8IX�
1����X�.I@IX�<�X��IX�@����4��X�6IX�8�4��X�3IX������I�WHOGGNI�=����������;���:��)�=�������������
��P@�����1��������=��������8��������� �����2&S������������������� ������I� ������� �����
&-X�RKR]RLTI�
�4��X�8I�WHOOQNI������������4���������������������������>����W��*=N>�����,��������4�
������������������I�-�I�6I��������-&X�GQHO]GQHQI�
�4��X��I@IX��4��X�;I-IX�/�����X�8I1IX�����>��X��I� WGMMKNI� ���������������4��������?���������
�4��4������������4�����U-HP�W&�/N�����>��������������������4��--G�:����������������4��
RJ���4������������I�&�������1�����=��I�',X�GHHR]GHPHI�
�4���X�6I3IX����:��X��I8IX�����*����X�/I*I�WGMLLNI���&1�������������������>�����������
��� ������������ -HPX��������������������� �����������4��4�>� ������ ��� ��C������I� 6I� -���I�
�4��I�'*+X�GHLHQ]GHLHKI�
�4���X�@IS3IX�<�X�=IX�@���X� -IX�<����X�-IX�=��4�:���X� �I+IX�<��X� 6IX�.�������X�=IX�&�����X��I8IX�
@����X� �IX� +��X� >IX� ��� ��I� WHOGPNI� 1� ����S��S��������� ��������� ��� �3�� �>��4����� ���
����������S�������������������������I������&,0X�GOKQ]GOLQI�
�4���X�1I-IX�-���4X�<IX�1���4��X��IX�+���X�<I�IX����������X�+I8I�WGMMMNI�&�����������������
�=���������������4������������������,�4�������S:���������������>���������4���� ��������
���� ��������� ���� :���)�� �4�� ���������� �HGW���GU,��GN� �=��������� ����������� ,��4�
����4���������������������I�2�����������>�&0/X�HLOG]HLGPI�
�4� ����=X� �IX� 2��4���X� <I6IX� ������X� �I=IX� 84����,�X� 1IX� @4������)>X� 1IX� -���C��X� +IX�
8����:���X�&IX�����+���-��X�.I�WHOGP�NI�/�����������������������������)��:��,������*=�
���� 2==^� ���� )�>� ������������� ��� �����>���S�������� �������4����� ����>� �� ��I� �����
/���:I�&)X�RLT]RMLI�

� � $�(��� ������������������
LR�
�
�4� ����=X� �IX� 2��4���X� <I6IX� ������X� �I=IX� 84����,�X� 1IX� @4������)>X� 1IX� -���C��X� +IX�
8����:���X�&IX�����+���-��X�.I�WHOGP:NI�/�����������������������������)��:��,������*=�
���� 2==^� ���� )�>� ������������� ��� �����>���S�������� �������4����� ����>� �� ��I� �����
/���:I�&)X�RLT]RMLI�
�4��X��IX��������X�<I�IX�84�����X��IX�<��X�3IS@IX������X��I1IX�&�)�X�/IX�@����4��X� 6I1IX�8�4��X�
3I�IX� <��,��X� �IX� +�����X� 1IX� ��� ��I� WHOORNI� �������� ����� ��� �RPS���������� ���������
���������������������������������S�����������������������I�&������0+*X�KHR]KPOI�
�4����X��IX�<���������X�-IX�<��X�2IX�-����X�@I2IX�������X�6I<IX�1����4��X�3IX�����-���4>X��I1I�
WGMMTNI� �>������� ���� ������� �� �=���� ����������� �������� ���)���� 8����� 4����4��� �����
��������I�&������+.+X�QOK]QGPI�
�4��X� �I�IX� ���� -���X� -I-I� WHOOGNI� ���������� ���:����>� ��� �������4�����U-HP� ��� ����S
������������������������������������������ �����I�/��I��4�������I�,-X�PL]QRI�
���������X�;IX�.����0��X�+IX�8�4�����)���X�2IX�3������X�1IX��������X�-IX�.��4�� ��X�+IX�����
=��:����X�1I�WHOOGNI�8������ ���&1�:�������:>��4����������������������������4������
����4������S������������������������I�6I�8�������-���4��I�/��I�-���I�)*X�HP]POI�
�����:�X�2IX�/�����X�6IS�IX����� �X��IX�;�����X�-IX������������X��I+I� WHOOHNI�&������4������
�����������4�����:����>������������������������� ��>�����RPI�&��I������-���I�0X�RHM]RPPI�
���)X� �IX�.�?���X� �I/IX� 1�������X� 8I3IX�84�,X� 1IX�-�X�/IX���������X�=I2IX� ���� -��4���X�1I�
WHOOKNI�&�����������C�������������������:������������������������:�����������4��������
��������I��� I�-���I�+&'X�HGK]HPOI�
���)�X� �I8IX� -����X� �IX� ���� ���4�X� +I=I� WGMMGNI� 1�������� =�������� 2=��������� ���
�� ��������/����=��������� ��*�����_I�2�����������>�&'.X�HLTK]HLKPI�
�����>X�@I1IX�1��?��X� 6I�IX�/���0 ��X� 8I�IX�/����)�X� 6I1IX� 8��������X��I�IX� ����1�0��X�@I6I�
WGMMTNI�������������������������������������GPC�������������������I��������=��I�,*X�GGQH]GGQRI�
�����X��IX�3�:���4X�1IX���������X�/I.IX�=����>�X��IX��������X� 6IX�3�������X�1IX�-�����4X�+IX�
����@���)��X�3I� WGMMQ�NI�1��������������������� ������ ������������� ����������� ������:>�
�������S��)�� ���,�4� ������S�X� )��������>��� ���,�4� ������X� ���� ���������� ���,�4� ������I�
�������=��I�,0X�RQKQ]RQKLI�
�����X��IX�3�:���4X�1IX���������X�/I.IX�=����>�X��IX��������X� 6IX�3�������X�1IX�-�����4X�+IX�
����@���)��X�3I� WGMMQ:NI�1��������������������� ������ ������������� ����������� ������:>�
�������S��)�� ���,�4� ������S�X� )��������>��� ���,�4� ������X� ���� ���������� ���,�4� ������I�
�������=��I�,0X�RQKQ]RQKLI�
�����X��IX�3�:���4X�1IX�-�����4X�+IX�����@���)��X�3I�WHOOONI�1����������������SS����������
������4�����������������������������������I�����I�=��I�'.X�HGG]HGMI�
�����X��IX�@���)��X�3IX�-�����4X�+IX�����3�:���4X�1I�WHOOHNI�1������������������������������
������I�2�����I�=����I��������-X�GRR]GKOI�
���4�X�+I=I� WGMKPNI� �4�� ����� ��� ���������� ��� �4�� ����4����S������4>���� �������������
�� �� ��������������������4��������������:�>���������I�1���I�=��I�&),X�LK]MTI�

� � $�(��� ������������������
LT�
�
���4�X� +I=IX� ���� ���0�����X� 1I1I� WGMLMNI� /�����4>���S����4������ ������������� ��� �4��
���,�4������� �������������4����������I�������������I�=��I�0*X�GRM]GKRI�
���4�X� +I=IX� �4���X� <I1IX� 84�����X� 6I/IX� �����4�X� *IX� ���� ;�0��X� 3I� WGMLPNI� 3������S
�������� ����4��������� ���� ���,�4>� ����� ��� ������4>���S����4������ ������������I�
=����������I�3���I�=��I�+-X�RRM]RMLI�
�������4��X�6I�IX�=������X�6I�IX�1���,X��I3IX�.�?C��?X�;IX�/���4�X�.I@IX������������ ��X��I�
WHOOKNI� ��*=� ��������� �����4�������� �=����� �� ��������� �4����4� �� --GS�+�SG���4��
����������������������=I�&������0,/X�KPT]KQOI�
����X��I.IX�*J�������X�@I1IX�������X�@I�IX�&��>��X��IX�*��4��X�=I�IX�����<�X�;I� WHOOTNI��4���S
/>�����������������,��)I�8����I��������-���I�&*X�HRP]HTQI�
��/��?�X� 1I/IX� &�����X�1I+IX� ������X�1I-IX� ���� 2������X� 6I�I� WHOOPNI� ���4��������� ����
������������������������������������I��4��<������+*&X�MRR]MTQI�
������� �X�&I&IX�-��>�4� �X��I�IX�1������)�X�2I+IX�84��)� �X�1I�IX����� �X�1I&IX�+��0���X�
2I+IX� �����)X� *I1IX� ���������X� *I.IX� ���� /����4��)�X� *I8I� WHOOHNI� 1� ��0��� ����������
�������� -HP� ��� �����)��� ��� �������������� ���� ��>� ��� 4����� �����4����� �>��4��>���I�
�������I�<���I�.+X�TK]KHI�
��8����X�<I/IX�����������X�/I�WHOOGNI�=��������������4��+G����8������������:>��4���:�C������
���4,�>I�;2-8�<���I�0-/X�GKM]GLMI�
�9�?S;�����X�<IX�/�����X�6I;IX�+���?���?X�=IX�.�����X�3IX�.���������X�;IX�1� ���?S1��@�����X�3IX�
���� �9�?S;�����X� <I� WHOOTNI� 1����� ����� ���� �������S������>���� ����� ��������I� 3�����I�
3�������4��I�'&X�MMR]GOHKI�
��::��X� �I�IX� 1����X� 6I/IX� ���� /������X� -I�I� WHOOMNI� �4��������?������ ��� =������
�4���4��>�������8�����=� ������������=���������������*=�������=�H�:>�8T@GI�/��I�����I�
-���I�'-X�RTRK]RTKOI�
����X��IX�1�X��IS6IX��4�X�+I�IX�>���X�-IX�3�X��IX��4���X�6IX�����>X�8I=IX�<�:���X�2I8IX�1�X�>IX�<��X�=IX�
��� ��I� WHOGGNI� 8/1�QS���������� :������� ����������� ��������� ������� ���,�4� ����
����������������������I�&������0)/X�HTM]HKPI�
���0�����X�1I1IX��4�����X�1I1IX��������4�X�+I=I�WHOOPNI�;+;SGO����>�����������������������
�4�����,�4�����4����������������I��� I�-���I�'*&X�PM]RQI�
��������X� &I.IX� ����4������X� 1IX� +����� ������X� �IX� ���:���X� &I3IX� 84�����X� &I2IX� ����
������X� /I� WHOOTNI� 8T@GS� ���� :����=��S��������� ������������ ��� ����Q� ���������
��������������������������������,�4I�8�������+&0X�QTK]QKGI�
����?X� �IX� *�����4��X� @IX� �����=X� 1IX� 84���:X�/IX� ���� 3����4�X� 1I� WHOGONI� �4�� ����4S
����������� �������� �1>>� ��� �� �� ��� 4������� �4�������� �� �� ��� ��� �4�� �����������S
��������������������������3PIPI�+������� I�'0X�GHRP]GHTRI�
���:�0�)X�/IX�/������X� 6IX�����0�X�8IX�/��?��X�@IX�1���?���)X�=IX�<� �����X�-IX���������4S
6��C�����X�1IX�����)>X��IX�;������X� 6IX�����?S8����X�=IX������I� WHOOPNI�1��������=������������
�HK�����8)�H��������������������������������1������S1����������������I�����I��������=��I�
*��I�6I�1�I�1����I��������=��I�-X�HTGP]HTGMI�

� � $�(��� ������������������
LK�
�
��:�� �)�X� 1IX�@��X� 8IX� 8�������X�=I6IX�1��)��X� 6I=IX�/����X� 8IS/IX� +���9�S2�4� ���9�X� �IX�
8�4���?X� �I+IX� ���� =���>X� .I1I� WHOOMNI� �4�� ����� ��� ��2&U1)�U��P@� ���������� ��� �4��
���������������� ��:����>������������������������S��)������������������I�����I�&���I�1���I�
8��I��I�8I�1I�&/*X�HTL]HKPI�
�@ ��X�@IX�-�����X� 6I<IX�/����X�8IX�=����X��IX�<��� �)>X�1I�IX�8��?�X�1I<IX�������������,X�2IX�
/�X�;IX�+���)�X�=IX� ���� ��X�8IX� ��� ��I� WHOGONI� 1��� ��������� ��/���:�����+����=��������>�
&��,��)���,�������������*=�������=�GI�/��I������+-X�GKG]GLPI�
2���������X� @I�IX� ���� ������4�X� /I=I� WHOOPNI� 3�=:GP� ��� ��C������ ���� �������
����������������������������>��������������4�� ���������������I��� ���������&+/X�HOTG]HOTMI�
2�,����X�8IX�����:���X��IX�;��4�X��IX�84����>X�6IX�+�������X��IX���S�����X�=IX�������X�1IX�;�����X�
�IX� ����)X� 6IX� 64� ��X�8IX������I� WHOORNI�2=�������������>������������������>������������>�
�����������&1��������4��4���4���3*>-GP���������)������4��������������������I� -�I� 6I�
�������-'X�PKT]PLGI�
2������?X� 6I/IX�8���)���X�@IX�/������X��IX�84��4)��X�1I1IX�8��)�X��IX�=������X��IX�+�������X�
8I=IX� �4�,X� 1I-IX� +������X�/IX� ���� <�����X� 2I8I� WHOGQNI� =&1S=&1� ������������� 2��:���
8���������������������&���������=&1�����&����������S�=&1�������4��������8����I������
&,-X�GLL]GMMI�
;�����X� -IX�/������X� �IX� ������X� 2IX� 1�����>X�/IX� =�����X� =IX� ���C�������X� <IX� <�� 8���?�X� =IX�
�� ����X��IX������:�X�2IX�8�������X�1IX������I�WHOORNI��>����������&������4���������1�����
/>����������<��)�����,��4���&������@��>��>��I�&I�2���I�6I�/��I�+,'X�HRQ]HTTI�
;���X� -IX� ;����X� 1I2IX� =�:���X� �I/IX� ���� ������X� 1I6I� WGMMTNI� 3��MO� ���������� ���������
���������4�������:��������������>���� � �I�6I�-���I��4��I�')&X�HLTMK]HLKOHI�
;������X� -I6IX� ���� ;������X� �I� WHOOGNI� �4�� �� ��������� ��� ��������S������������
���������������I�&��I�=� I��������&X�PQ]QRI�
;���������X�&IX�����=����??�X��I1I�WGMMGNI���� � ��������� ������4���4��>������������������
��������X���������>�����������������������������X�����������S�����������&�3�P�P�3�=������I�
2=�I������=��I�&-0X�HLM]HMTI�
;�����X��I�IX�8�����X�8IX�����X��IX�3����,X�2IX�����-�����X�6I�WHOOHNI�/����������������?�����
�����������:>���*=�����������,����������������8T@G�����Q2-�GU��;Q2I�+������� I�&*X�GQKH]GQLKI�
;��)�X� 1IX� ����+����:�����X��I� WHOGPNI���������������� �����4����� ����� ��������� �������
���4������� ��� �4�������S��������� �������� 4����������I� ����� 8������ �4��������� &.X�RMG]TORI�
;�����X� �I8IX� ������X� 1I=IX� 1�:�������X�<IX�;��4��X�+IX�/>����X�3IX� ����)X� 6IX� 1�����X�+IX� ��S
-���X�6IX�8�������X��IX�=�����X�.I2IX������I�WHOOMNI�3��SHK��=�������������������������������
��������������������������������������������������������2�8S������������������I�-�I�6I�
�������&/&X�GGPK]GGQQI�
;��������X� 8I3IX�/��)��X� �IX�+����X�*I�IX� ���������X� �I�IX� ���4�X�+I=IX�3������X� �IX� ����
�4�����X� 1I1I� WHOOPNI� 8����� 4����4��� ���������� ���������� ���,�4� ���� ����4������
���������������I��� I�-���I�'*0X�PRH]PTHI�

� � $�(��� ������������������
LL�
�
;��4���)X� <I6IX� 2��9�S<;��?X� 6I/IX� ���� 1���;X� 6I� WHOOKNI� &�,� �����4��� ����� �4��
�������4�����U�������������������>�������������4��������I�-��2���>��&�,��=� I�/��I�
����I��� I�-���I�'-X�QM]RMI�
;���?��4�X� 8IX� @��?������X�/IX�3�������X�/I6IX�/@����X�/IX� 2�����X� =IX� +�>��X�3IS6IX� ����
8�4��?X�1I1I� WHOOTNI� ������������ ��,�S����������� ��� 8�=-G� ���� 8�=-H� ��� ���������
���������I�2�����I�=����I��������&+X�LPM]LQMI�
;�)���S@�:�>��4�X�6IX�����/�����X�-I�WGMMMNI��>������G���4�:������������������������4����4�
:������������&1�������)HI�2=�I������=��I�'0*X�PPL]PQKI�
+����X�8I8IX�84�����>�X�6IX�@��4���X�1I3IX�����@����X��I@I�WHOGONI�&�/PX������:�������4��
�������4�����U�������������� �����>X� ��4������ ���� ����S���������� �������������I�
-���4������>�W/���IN�0-X�GPRR]GPRKI�
+������X��IX�-�����X�/IX��������X��IX��������X�-IX�8�4����X�2IX�����@������X�+I�WHOOTNI�3����
�4��)� ��������� HK� ���� KO>� ����S���������� ��������� ,��4� ������������ ����������I� �����
�>����+������I���=�,X�HRMH]HTOGI�
+������4X� 6I=IX� -������X� =I6IX� /�����X� =I1IX� -�>��X� -I;IX� ��/�>�X� ;I6IX� ;�������X� /I6IX�
1�����������X�=IX�=����X�6I/IX�����+����:���X�&I/I�WGMMTNI�/�����������������������������
�������������/����I��������=��I�,*X�QOMT]QGOHI�
+�����X� �IX� ���� ����4��X� -I/I� WHOGHNI� ����S��������������������������� ��� �4�� ���������
��������I�/��I�����I�2���������I�+,'X�KO]KLI�
+�����X��IX�/������X�6I1IX��������X�+I=IX�;�������X�3I;IX�����1�:��X�/I6I�WGMMMNI�1��� ���������
�������S���� �������������)������ �����������,��4���������� ������������������I� �������
=��I�,-X�HKM]HLQI�
+�� �������X�2IX�8�������X�/I6IX�@���4� ��X�@IX�-��,�X�/IX���4�X��IX�-����)>X�1IX��������X� 6IX�
3����)���X��I3IX�����@������X��I1I�WGMMKNI��4���1+��������,��4����4��������������������
����������������������4���������������������I�����I�&���I�1���I�8��I��I�8I�1I�-0X�PPHO]PPHPI�
+������X� �I;IX� ���� /��������X� +I�I� WGMKQNI� ����������� ��� ���������� ���� ����������
���������������:>����:�����4����������������������������������������I�6I�����I�&&&X�RL]TQI�
+������������X� .I6IX� <����X�3I-IX� ��������X� 1I8IX�&���X� �I2IX� ���� =�:���X� �I&I� WHOOGNI�
2=������������=1��PX�����������4������������������S���� ����� ������������������I�-�I� 6I�
�������.,X�GMHL]GMPTI�
+���:���X� 1I�IX� -����?>��)�X� <I1IX� &�4X� @IS/IX� <�,��X� �I1IX� 2��������X� 8I6IX� 8������X� 8IX�
��,���X�8IX�<�,X�/IX�+��X�>IX�<�X�>IX������I� WHOGONI���������� ����������������4������� �������
3PIP�������?���������������������������������I������&0/X�TKL]TMGI�
+���X��IX�����/�C���X�<I2I�WHOGGNI����=&1����������� ����8�1�GS����������=&1�����>�
:>������=����,��4�PJ���=�� ���1�����������I�&������0)/X�HLQ]HLLI�
+�����X��I8IX�1�X�-IS/IX�=�:�����X��I=IX����X�>IX��4�����)����X�8I/IX�@4��X�1I�IX�;����X�/I6IX�
6���X� >IX� <������X� =I6IX� -������X� 6I�IX� ��� ��I� WHOGH�NI� �4������������� ���������� ��� ���4���
����������S�������������������������I�&������0.)X�HPM]HQPI�

� � $�(��� ������������������
LM�
�
+�����X��I8IX�1�X�-IS/IX�=�:�����X��I=IX����X�>IX��4�����)����X�8I/IX�@4��X�1I�IX�;����X�/I6IX�
6���X� >IX� <������X� =I6IX� -������X� 6I�IX� ��� ��I� WHOGH:NI� �4������������� ���������� ��� ���4���
����������S�������������������������I�&������0.)X�HPM]HQPI�
+�����>X��I1IX�3�X�-IX�6�4����X�=I�IX�;���X�*I3IX�/�4���X�6I<IX�;����4X�;I8IX�����1�����X�2I/I�
WHOOGNI�1����4���������������������������S������������������������������������������
�������������� �������4����>I��������=��I�*&X�QPGR]QPGMI�
+�����>X��I1IX�;��X�>IX��������X�<I1IX�3�X�-IX�-���X�3I/IX�;����4X�;I8IX�����1�����X�2I/I�WHOOQNI�
2������������,�4� ������� ���������� ������ ��������� �4����������� ��������� ��� ����������
���������������I�6I�-���I��4��I�')-X�KGGM]KGPOI�
+�������X�8IX�-�������X�=IX�=����X�/IX��4���X�@IX�@4���)��X�<IX�/��� �X�@IX�������������X��I�I�
WHOORNI�=��������������4������ ��� ��:�>������� ��������� ���� �������������I�&������
0+)X�GQK]GRPI�
+���4���X� �I-IX�@��X�8I-IX�;������X��IX�/�)����)� �X�3I�IX�����1�����X��I�I� WHOORNI�-/�K�
��4�:���� :����4���� ����4��������� ��� �4�� ��������� ������ ���� ����������� ,��4� &���4�
���������I��� I�-���I�'..X�PPQ]PQKI�
+�:��X� 1I&IX� &0�����X� 6I/IX� -�������X� +I+IX� ����/�����X� 6I<I� WGMMMNI� +���� �=��������� ���
��������������4�������>�4����������I�+��������,-X�GTL]GKKI�
+������X��I1IX�8�� ���X��I/IX��4�����X��I�IX�-����X�1I1IX�@�����>X�&I-IX�/�����X�6IX�-��,�X�/IX�
;��?������X�@I6IX�����8�:�����X��I/I� WHOOTNI�1:������� ���/������� �4����*=�������������
������X�������X�����<8�L�=� ������4�����*=�H����=�C����������8������������1)�S;*>*�����
�@�^X�:���&���8T@GI��� I������&&X�LRM]LKGI�
+��X�<IX�����������X�<IX�����;��4�X�2I�WGMMTNI�@��������>������,�4�������������C����������
4������ ���������:�����������,�����4������I�+������� I�&/X�GTR]GKRI�
+��X��IX�-���X�>IX� 8��X�;IX� 6����X�=IX�<���X� �I2IX� �4��X�3IX� �4��X�3IX�@���X�>IX�/������X� 6IX�
������X� �I+IX� ��� ��I� WHOOMNI� 1� �� ��� ��������� ��������� ������� ������� ��� ��S����������
������������������������������������������������������������������S�������������,�4I�
�������=��I�*-X�HPOR]HPGPI�
+�������4>X�/IX����X��I3IX�&�X�=IX�������X�<IX�<��X� 6IX�<��X� 6IX���>X�1IX��4���X�=IX�<�X�;IX�<��X�
�I/IX� ��� ��I� WHOOLNI� &������4������ ���������� ,��4� 32>�/G� ���� ���������� =&1�
���>�����������������������I�6I�/��I�-���I�+).X�POH]PGKI�
3��)����X�@IX�+������������X�.I6IX�/�4��X��I-IX�<����X� �I=IX�-���>X�/I2IX� ���)X�8IX�<����X�
3I-IX�&���X� �I2IX� ���� =�:���X� �I&I� WHOOPNI� 2=��������� ��� ���TOX� ��� ��������� ���������
������ ����X����������������������������������� ��������I�*��������''X�HQTT]HQKKI�
3�������X� 3IX� �����X� 8IX� -�������X� �IX� ���� &��4���X� 2I� WHOOHNI� 8�����>G� ���� 8�����>H�
��� ���� �� �����������4������ ���� �4��=��U/1�@� ����������� ���4,�>I�&��I� ����� -���I� 0X�LRO]LRLI�
3���4��X��IX�����1���:���X�=I1I�WHOGG�NI�3������)�����������>��4����=������������I������
&00X�TQT]TKQI�
3���4��X��IX�����1���:���X�=I1I�WHOGG:NI�3������)�����������>��4����=������������I������
&00X�TQT]TKQI�

� � $�(��� ������������������
MO�
�
3�����X�@I1IX�����=����X��I/I�WHOOGNI����������������������4������S���������>����������
������>���������������������������4��I�������*&X�HGKK]HGMHI�
3����X� ;I�IX� -���4��X� 1IX� ����3�>��S3����X�/I� WHOGGNI�/��������� �4��������� ��� ��������
������������������������I�&������0),X�PHQ]PPHI�
3�����:���X�3I-IX�8�����4���X��I1IX�+��������X�6I�IX�1��4����X��I/IX�-�?���X�+I8IX�/���X�6I1IX�
���� 8�� ���� �X� 8I� WGMMRNI� 1���������� ��� �4�� ������ 8���������� +���� �RP� ��� �� 3��4�
;�����������3�������=��������>����������������I�6I�����I�&,0X�QGQ]QHGI�
3�����X� -IX�;�������?X��IX�;��)��X�1IX�=��X�=IX� ���� -4����X�@I� WHOOKNI������������������
�������4������W&�/NG��������������������������������������������������� � ������4�����
1/<������I��������=��I�*)X�QOLQ]QOLQI�
3���X�=IX�8���X�-IX��������X�=I�IX�����.����>)�X��I�WHOORNI�3�������������������� ��������
����������:>� ���������=:���������� ��������������������������������������I��������=��I�*,X�GOHQP]GOHRQI�
3��������X�@IX� 8?�:���X� 1IX� ����*����X�/I*I� WHOOONI�/������� �4�� ����������������������
������������������-HPI�6I�-���I��4��I�'),X�HQQRG]HQQRKI�
3������X�/I�IX�1����� �X� �IX��4���X�8IX�<��X�<I6IX�.��4�� ��X�/I�IX��4��X�2IX�-��)X�8I�IX�����
3�����:���X� 1I&I� WHOORNI� �4�� 1�������� =�������� =�������� &������� =��������
��=���������W&S��=N�����4��������������/������������ �������&���������������=� ���������
&� ��� /��������� /��4������ ���� 1�������� =�������� 1����������I� 6I� -���I� �4��I� './X�TRGG]TRGMI�
3���X�3IX�@��X��IX�6���X�/IS3IX�2:��X�6I&IX�@��4X�/I*IX�+������X��I1IX��4���X�8IX�<�X�<IX����X��IS>IX�
3�X� �IX� ��� ��I� WHOOQNI� 1:������� �=��������� ��� �1=/GX� �� ���������������� ������ ����� ���
��������� ��������X� ��� �4�� �� ��������� ��� ��������� ���������� ���� ��������S
������������������I��������&/&X�LP]LMI�
3������>X�6IX����������� �>X��I�WHOOMNI��4��/��>����4,�>�����=&1������������I������&+*X�KTP]KKTI�
3�X�=IX�����X��I1IX�1��X�8IX���4��,��X�8IX�.�����X�=I1IX�3���4��>�X�2IX�3��X�/IX�������X�1I1IX�
.�������X�=I<IX�������X�1I-IX������I�WHOOMNI�<�����S������������������������������ ��������
���� ��� ����� ��������� ��� ��>����� �=���� ������>� 4������S���������>� ��������� ������I�
�������=��I�*-X�GT]HHI�
3�X�-IS�IX�-�4X�8IX�-�4X�8IS�IX�8������X�2I=IX�3����X�6IX�<�X��IX�3��X��IS<IX�����X�3IS6IX�<��X�3IS@IX�
1���X� <IX� ��� ��I� WHOOQNI� ;���������� ������� ���������� ����>���� ��� ��������� ���������
������������1=1KO�����������������������=���������������������������I�6I�-���I��4��I�')-X�PPQPL]PPQQTI�
3����X�6IX���::��X��I�IX�/����?�)�X�/IX�����/������X�-I�I�WHOOLNI��4���8�GS�8�H�������=�
�����C��������������������� �����������*=�������=�HI�/��I�����I�-���I�'.X�QGOQ]QGGRI�
3����X�<IX���X�-IX�1���X�8IX�-���4X�<IX����������X�+I8I�WHOOR�NI��4����������;��GO��������������
:����4��������4�������������������=������������ �4�� ������������������>� ��:�S���������
������������:>�������������������I��� I�-���I�').X�PMT]QGQI�

� � $�(��� ������������������
MG�
�
3����X� <IX� ��X� -IX�3�X�1I-IX� -���4X� <IX� <�����S������X� �IX� -������4�X��IX� ���� �����X�+I8I�
WHOOMNI��4����������1��R���������������������� ��������I��� I�-���I�+'.X�GLL]GMMI�
3����X�&IX�&���X�8IX�8?�:���X�1IX�����*����X�/I*I6I� WHOOR:NI���������&�/P� ����������,��4�
�4�� ���������������� ���������� �������� -HPU�������4������ ���� ��4�:���� ��:������
:���������I�6I�-���I��4��I�'./X�RQMT]RROHI�
3�++�&8��X�8�2.2&8�=2X�6�IX�����3*�+28��.�WGMQGNI�8��������������������������>���I��4��
��������������������������� ��������������������4����������������I�1��4I�8���I�0+X�HOM]HHPI�
3��,��?X� 1I1IX� ;�����X� -I1IX� 1������X� 6I�IX� +����:���X� &I/IX� ���� @,��X� 2I�I� WHOOGNI� �4��
�=1/��������������������������������������I�����I�������I��������I�2�I�6�4��2���������
1�� ������'/X������HOIRI�
�����X�@I<IX�<��X��IX�<��X��IX��4���4��>X�&IX�+������X�1IX�/�����X�8IX�6����X�1IX� �������3�� ��X�
�IX�������X�=I+IX�3���X�/I/IX������I� WHOOMNI�&������4����������&���������=��������@S=���
������� /��:����� ������������� ���� /1�@� 8������ ������������I� 6I� -���I� �4��I� '.0X�HLQGO]HLQGMI�
���4��X��I1IX�&�)���X�-IX�����8����X��I�WHOGGNI�/��4������������������������3����4���
�������������������4������?��I�&��I�=� I�+����I�&'X�PMP]QOTI�
���4���X�@IX�-4��X�@I�IX�6��X�1IX����4���X�-IX�3�,)�X��IX�@�:�>��4�X�=IX������4���X�-I�WHOOP�NI�
������ 8���������� 1=;� ��������� -HPX� �� &��������� �������� �� �� ��� ��� =�:������
-��������������������������������I�/��I������&'X�GGRG]GGTQI�
���4���X�@IX�-4��X�@I�IX�6��X�1IX����4���X�-IX�3�,)�X��IX�@�:�>��4�X�=IX������4���X�-I�WHOOP:NI�
������ 8���������� 1=;� ��������� -HPX� �� &��������� �������� �� �� ��� ��� =�:������
-��������������������������������I�/��I������&'X�GGRG]GGTQI�
���X��IX� -�����X�/IX���?��X�/I6IX�@�:�>��4�X�=IX� ����@�������X� 6I�I� WGMMKNI� 1�;X� ��� �81�S
���������������1��S�����?�����4�������������:�>���������������� ������I������-/X� GQR]GRRI�
6������X� 2IX�<��,��4X�=IX�8�4����X� 1IX�<��X�8IX�=@���X�/I1IX�3���X� 1IX�����3���X�/I&I� WHOOQNI�
/����������*=�������=�H�����������4���������>���)�������������������>������������� �I�
&��I������-���I�*X�GGHH]GGHLI�
6������X� +IX� ��� ���� @�����X� 3I1I+I/IX� �������X� 6IX� ���� -���)����X� 1I*I� WGMMR�NI�
��������������� ����,���������������� 1��� ������������ ��� �4��&S��������� ������� ��� �4��
3�����1��������=�������I�6I�-���I��4��I�')/X�KPQG]KPQTI�
6������X�+IX� �������@�����X�3I1IX��������X�6IX�����-���)����X�1I*I�WGMMR:NI����������������
����,�� ������������������ ������������ ��� �4��&S������������������� �4��4��������������
��������I�6I�-���I��4��I�')/X�KPQG]KPQTI�
6���X�-IX�+��X��IX� 8��X� 6IX� 84��X�;IX�;���X�;IX� 6���X�-IX� ����-���X� �I� WHOOMNI�=&1� ���������
������������ ,��4� �������4������ ���������?������ ������� ���������� ��� ������� �����I�
*��������'.X�QHOG]QHGGI�

� � $�(��� ������������������
MH�
�
6�4������X� 3IX� ���� 8��������X� 8I� WHOGONI� ����� �������������� �������X� *��QX� 8�=H� ����
&����X����� ������>������������=���,��4��������4������W&��GN��������������:�>�����
�����W28N������������������������I�1�����'X�LGR]LHHI�
6�4����X�1IX�����8)��4���X� 6I/I� WHOGPNI�8��������� �4�� �����������������I� ����I�*���I� �����
-���I�',X�KGK]KHPI�
6���X��IX�@��X�=IS8IX��4���X�3IS6IX�<��X�8IS6IX�����6���X�/IS3I�WHOOQNI�3*>-GP������������,�4�
������������ ��� ��������� ������� ������ ��� �� ���������� ��� 4������S���� ����� ���������
������������������I��������=��I�*0X�MGLR]MGMHI�
@�������X�3I3IX�3������X�6IX�.��X�/I6IX�@�:���X�3IX�������>X�=I/IX�-������X�2I1IX��4���4��X�
/I2IX��4��X�-IX�����3��4��,��X�<I2I�WHOOMNI�+���������������4�������������������4��4�����
4�����4��)���������I������8�������4���������&0X�GOR]GGGI�
@�����X�;IX�1�������X��IX�/��4�,X�.IX�-�X�6IX��4���X�<IX�@4��������X�/I8IX��4�����>��X�1I/IX�
<����X�@IX�2�4����X�8IX�8�4������X�=IX������I�WHOGGNI��4���2@������������������8�W ��N��4������
�������������4������4����������������>I�+������� I�',X�TKP]TKLI�
@�����X�8I�WHOOLNI�2=���������4��������������4�������������������������������������������
����I�8���������=� I�0X�GMP]HOGI�
@��X� -IS-IX� +�>���X� =I-IX� 8���X� @IX� ������4���X� 1IX� ���� 6���X� /I� WHOOHNI� ���������� ��
���� ������ ���&;S)����-� ���@�S���S������������ ��������� ����4������ �����I�*�������� '&X�QQMO]QQMKI�
@��X� 6IS3IX� -���X� /IS8IX� ���� �4��X� 6I� WHOOMNI� 8������ ����������� ���� ���� ����� ���
�������������� P� W8�1�PN���������� ������ ����� ��4�:������ ��� �������� ���������� �4����4�
�������KHK��4���4��>������I�6I�-���I��4��I�'.0X�PRQHR]PRQPHI�
@���X� �I6IX� +�����X� <IX� ���� <�����X� 2I� WHOOLNI� 3����4��� ����������� ��� ����������
�� ��������������������I�6I�2���������I�&-.X�QPM]QROI�
@��?������X��I�WHOOKNI��4���������������������������4������������I������&'.X�TMP]KORI�
@����:���X� 6I�IX�/���)X� 8I&IX� ���4���X� �I6IX� -������X� =I+IX� ���>��X� �I1IX� @����:���X� 8IX� ����
+4��4X��I/I� WHOOQNI��4���4��>���������� 1)�� W8��QKPN� �������=��������������������������
�����������������������������������I��������=��I�*0X�RHPH]RHPTI�
@�X�8IX�<������X�2IX�1�����>�X�=IX�=���)���4���X�8IX�2����X�<IX���������X�=I�WHOGQNI���4�:���������
4��MO� ��������� ������=��� �4����>� ��� ��������� ���������� ��������� ������I� ���8� *��� -X��GOPTLOI�
@��X�/IS<IX�-�����X�1I����X�-���,�����X��IX�=������X�/I;IX�����84���X��I6I�WHOOQNI�&S���������
���>�:�C�������������������������������4��1�������������������I�+������� I�&.X�GLTH]GLKQI�
@���4��4�X��IX�/�>�)�X�3IX�3���X� �IX� ���� ;�0���,�X�/I� WHOOKNI� 2=��������� �����0���4����
�4��)� ��������� ��� ��������� ������>� ������������ ,��4� ����������4��������� ��������� ���
����������������������������������������>I�6I�����I�&))X�KRK]KTGI�

� � $�(��� ������������������
MP�
�
@��)�X� 8IX���������X�@IX�<������X�<IX�@� �4��0�X��I/IX�*0���X��I/IX�/��)X� �IX� ����<��4�X�/I�
WHOOQNI�&��������� ��������&�/� ����������,��4�3�/H� ���� ��������� ������ �����������
���������RP������3�/HS��������������������I�������������,X�QTR]QKRI�
<���X� /I<IX� �������)X� �I1IX� -������X� �I3IX� <��X� 6IX� �������X� 6I;IX� 3�:���X� �I/IX� ����
-��4���X�1I�WHOOGNI�/�����4>�����������:��������4�����������������Q������������������
:�����������:����4��������4�������������4���� ����������������I��� I�-���I�'+'X�POG]PGQI�
<������X� �IX� ��� �4����X� 1IX�/�����X� 8IX� �������X� �IX� ���� +������X� �I� WHOOKNI� 1���������
��������������������������>���������4�����4��)����������38�MOX�38�KO�����38�HKI�������&X�RP]TOI�
<�������X�/IX�����8�:�����X��I/I� WHOGPNI�=���������������*=�G����� ���� ���������������
�=��������������������I�6I������8��I�&'*X�GKGP]GKGMI�
<���4�)X� /IX� -��4���X� /IX� 8�������X� @IS�IX� ���� 3���������X� 1I� WHOGGNI� ���:����>� ���
&��=U8/=��������������������������������������������������� ��>�������������������������
�����I����I�6I�/��I�/��I�'.X�TQR]TRGI�
<��)�>X�=I1IX�3����X�-I/IX�/����X�1I�IX�����;���4X�6I�I�WGMKLNI�&��������������������:����
:>�������������������,4��4�:�����4�����������������������4�������&1I�&������'),X�QGT]QHOI�
<�����?)�X��IX�����/�?���X��I�WGMKKNI���������������4���������������������:>��������������
�������������I�6I�2���������I�)0X�QK]RRI�
<�����X�&I1IX����� ��X�8I<IX�84�4X��IS8IX��4���X�6IX�/����X�&IX�;�����X��IX���������X�1IX�<��X�8IX�
=����X�-IX�/�4��X�1IX������I� WHOOTNI� ������ ��������� �4���RP����4,�>� ���������:�������I�
&������000X�TG]TTI�
<����X�@I+IX�1���X� -IS2IX� 6�4����X� <IX� ����+��X�1IS;I� WHOOLNI�+���������� ��� �� ���������
�����������������������������I�&������0,*X�LOQ]LOLI�
<?������X�<IX�/������X�<IX�/����X�/IX�/������X��IX����������X�/IX�.����0��X�+IX����������X�;IX�
-���4���X�/IX�.�>���-��X�+IX�/����X�<IX������I� WHOOL�NI� ��������������������4�����U-HP�
����&1�:������������������������������� ��>�����4���������������������������������������
����I�*��������')X�HLRL]HLTKI�
<?������X�<IX�/������X�<IX�/����X�/IX�/������X��IX����������X�/IX�.����0��X�+IX����������X�;IX�
-���4���X�/IX�.�>���-��X�+IX�/����X�<IX������I� WHOOL:NI� ��������������������4�����U-HP�
����&1�:������������������������������� ��>�����4���������������������������������������
����I�*��������')X�HLRL]HLTKI�
<�X��IX�����3���X�8I=I�WHOGPNI�&������4������������������������S/>������������������?������
�����S/>�S�����������&1��������������I�*��������+'X�GMLL]GMMQI�
<�X��IX�<������X��IX�;������X�1IX�<��������X�6IX��4����X��IX�1����:���X��IX�8�������X�-IX�<��X�-IX�
2)���X� �IX� 1���X� +IX� ��� ��I� WGMMLNI� ��������������� ��� �,�� ��������� �������� �������� ���
�4���������GP�������������������I�*��������&*X�QLG]QLKI�

� � $�(��� ������������������
MQ�
�
<�X� 6IX� �4���X� >IX� 8�0��X� �I�IX� -��:>X� +I�IX� ���� ����X� ;I� WHOOQNI� 3>��=��S��������
�������4������������������������4��4����4���4�:����������RPI�6I�-���I��4��I�')-X�QGHKR]QGHKMI�
<�X�6IX�;�X�6IX������?��X��IX�-���X�3IS+IX�����1���X�6I�WHOOTNI�1����������4�������S���������
W&N��������:�=>�S���������W�N����������������:��������������������������������4�������I�
/��I�2���������I�-�����I�/��'/X�KKT]KLRI�
<�X� 6IX�8�0��X��I�IX�-����X�8IX� �4��X��I6IX���������X�;I� WHOOKNI�&������4�����������������
��������S�������� ���������� ���� ����������� ���� ��4������ ���������� ������������ ���
������,��4��������������:����>I����������������'.X�GGTP]GGKOI�
<�X��IX�-����X��IX�����3���X�8I=I�WHOOLNI�&������4�����������������������>�,��4��S/>������
����������S/>�S��������4>�����������������������������������I�����I�&���I�1���I�8��I��I�8I�
1I�&/,X�GLKMQ]GLKMMI�
<��X�-IX�<��X�+IX��4���X�-IX�3�X�-IS�IX�-�X�@IX�<��X��IX�/�@��4��X�@IX�>���X� 6I1IX�*����?X��I/IX�
84��X�/I/IX������I�WHOOKNI�;�:��:��������,�4�����������������H��>�������)�����������C������
������������������4��������������4����C�������������������������������������>�����������
�������4����������I��� I����:I�2���I�&+0X�KHP]KPQI�
<������>�X� /I8I� WHOGONI� &�/GU-HP>� 1� /��������������� �4�������� ��� =�:������
-���������������4��������=���������I�-���4��I�=��I����I�'/&&X��GMRHOMI�
<������>�X� /I8I� WHOGGNI� &�/GU-HP>� 1� /��������������� �4�������� ��� =�:������
-���������������4��������=���������I�-���4��I�=��I����I�'/&&X�GMRHOMI�
<��0�X�/I6IX�8� �������X�@I6IX�8����?)�X�*I=IX��������X��I<I6IX�.�������X�=I<IX�����.���)����X��I�
WHOOGNI� 1������������� ���� * ���=��������� ��� 1�������� =�������� +���� ��� 3������S
=��������>����������������I��������=��I�*&X�PRRO]PRRRI�
<��X�>IX�<��X��IX�6���X�8IS1IX�/�X��IX�84������X�@IX�@���X�8IX�����X�8IX��4��X�6IX�;�)���,�X�@IX�
���� -�X� @I� WHOOKNI� 8���>������� ��� �������4�����U-HP� ���������� ���� ��:���������
������?�����X������������������������������������� � ��I�����I�&���I�1���I�8��I�&/0X� MTKM]MTLQI�
<�X� 8IS6IX� ;��X� <IS�IX�����X� -IS;IX� <��X�@IS-IX�3����X�3IS<IX�1���X� �IS3IX� <��X�3IX� �4��X� �IS�IX�
3����X�8IS;IX��4���X��IS6IX������I�WHOGPNI�1��� �������������������������4������,��4�-�<HS
�����������>������������������������4�� ���������������������� ��>� ���4�����4��������
�����I�3������I�-�����I�/��,)X�GLMP]GMORI�
<���X� >IX� <��X� -IX� *���?S.���X� 8IX� -���?�,�X� @IX� ���� 1 ���4X� 6I� WHOORNI� =4�:� -����� ����
=����������4����*=�@�����I�����I�-���I�&,X�KOH]KGPI�
<��������X�1IX��������������X�&I�WHOOMNI�-���������������)�����������������������������
���������������I��������&&,X�PORL]POTKI�
<����X� �I<IX� +����X� -IX� 8��������X� 8IX� 2����X� �IX� <��X� 1IX� >�X� 6IX� 3��)�X� 6I<IX� ���)X� -I3IX�
3���4��>�X�2IX�������X�1I1IX������I�WHOGGNI���2&��������������:>���������������>�����>����
���������� ���� ����������� ���������� ���� �� 4��4� ���)� ��������� ��4���� ��� ��������� �������
��������I�����I��������=��I�*��I�6I�1�I�1����I��������=��I�&)X�TRTP]TRKPI�

� � $�(��� ������������������
MR�
�
<��:���X�+IX�-�����X�=IX�/�C�����X�8IX�<����S�����X��IX�-�������X��IX�.�������X��IX�����/����X�
<I� WHOGQNI� &�/G� ���������� �������� ������� ���,�4� ����/1�@� ����������� ��� ���������
������������I����8�*���-X��MTHMPI�
<�,�X�8I1IX�������X�2IX�����2 ��X�+I� WHOOQNI� ���������� ������������������I�&������0+'X�POK]PGRI�
<�X�1IX� <��X� -IX� @��X�/IX� ����/�@��4��X�1I<I� WGMMMNI� ;�:��:����� ���,�4� ������SGOI� 1�
����������������� �������� �������4���������������������� �����������I� 6I� -���I� �4��I�')0X�GHLHK]GHLPQI�
/��1��4��X� �I1IX� ���� 84��)������X�+I/I� WGMMKNI�&��P>� ���� ��X�,����>� �=�������� �����
��������� �� �������� �������� ��� �4�� ���������� �4��������� �������������� ����
�������4�����I�+��������0'X�GPK]GQOI�
/�������X�<I1IX�8��4������X�-I1IX�-������X�=I6IX�����+����:���X�&I/I�WHOOQ�NI�������������
������������=:������������>����������������������I��������=��I�*0X�TOGL]TOHRI�
/�������X� <I1IX� 3���X� 1I6IX� -������X� =I/IX� ���� +����:���X� &I/I� WHOOQ:NI� �������������
�=������������������>�������������>���������������� ������� ������A�>������,���4B��������
������������������������������I��4�����������,.X�PPR]PQQI�
/�������X�<I1IX�8��4������X�-I1IX�-������X�=I6IX�����+����:���X�&I/I�WHOOQ�NI�������������
������������=:��������2���>�8��������������������I��������=��I�*0X�TOGL]TOHRI�
/������X��I1IX�6����X�<IX��4�)�� ���>X��IX�-���X��IX������������X�;I�WHOOQNI�&������4������
8�������4���4���������RP�=������������.�=��������I�/��I�����I�-���I�'0X�PKOP]PKGGI�
/�0�����X� �I@IX� ;�::�X� �I+IX� -�)���X� =IX� -�����X� =IX� >��X� ;IX�/�/�4��X� <I/IX�/�����X� 6IX�
-���������X� 6IX�/��������X��I6IX�+���:X��I=IX������I� WHOOQNI���*=� ��4�:�������� ������1)�S
����������������������������4�����������������4����4�����������������������������3�;SGS
�������������4,�>�I�&��I�/��I�&/X�RMQ]TOGI�
/�0�����X� 8IX� <��X�-IX� ;���X�*I3IX�/�4���X� 6I<IX� ����14���X�-I2I� WHOOTNI� �� �� ������ ���
������������4>������������� �1=/G� ��������������������� �����������������������������
����� ��:����>I��4�����������**X�GHMH]GPOGI�
/��������X� =IX� ���� =���:���X� �I� WHOGONI� �4�������� ���������� ���� �4�� ��4��������� ���
����������������������I�&��I�=� I�+����I�&&X�HLR]HMTI�
/�������X�6I�IX�����+����X��I=I�WHOOGNI�-���)�����4�������4��������:������I�&��I�=� I�/��I�
�����-���I�'X�TP]TKI�
���/��?�X� 1I/IX�/��)��X� 1I@IX� 2������X� 6I�IX� ���� �����>X� �I8I� WGMMLNI� ��������� 8���� �����
������������I�1�I�6I����4��I�&,+X�MGG]MGMI�
/������?X�6I�WHOOQNI�+G�����S�>����������������������I�&������0+'X�HML]POTI�
/������X�6IX�2��4���X�6I/IX������������X�/I/I�WGMMKNI�1����������������4���������GPC����
4�����������������������I�����I��������=��I�*��I�6I�1�I�1����I��������=��I�+X�GLTK]GLKHI�
/������4���X� �I@IX�.� ����X� �IX�@,��X�1IX�����X��IX�1��� ����X� 6IX�����8�,>���X��I<I� WHOOQNI�
32=HU����)�����S����������������������������������������������������4����4���������
����&1�:��������������:����>I�������������*X�RGK]RHKI�

� � $�(��� ������������������
MT�
�
/�����X� +I6IX�/�����X�3I<IX� ��� -�)4� ��X� 1IX� <��:���X� 6I=IX�1���4���X� �I&IX� 8�4������X�*IX�
<����X� /I8IX� ���� &������X� 8I@I� WHOOPNI� 1:������� 3*>�� 2=��������� 1����������� �4��
/����������4����>������3�������������I��������=��I�*+X�RLKM]RLLLI�
/������X�>IX������0����?X�/I-I1I�WHOOQ�NI�2������������,�4�������X����������4��������
�4���������������������������������������I�;2-8�<���I�,)&X�G]LI�
/������X�>IX������0����?X�/I-I1I�WHOOQ:NI�2������������,�4�������X����������4��������
�4���������������������������������������I�;2-8�<���I�,)&X�G]LI�
/��������>X�=I-IX�/�����4��X�2I1IX�.�������X�=IX�3���X��I<IX�@��4���X��I;IX�3�����X��I8IX�����X�
<I�IX� ���� &�����X� �I8I� WHOOLNI� /����������� ��� ������������� ���������� ��� �����������
��������� ������>� �����4������ ���� ����������S���������� ������ ���,�4I� ������� =��I� *.X�QQQK]QQRQI�
/������������X� &IX� 2,���X� �I8IX� ���� ���:�����X� <I;I� WHOOHNI� 1� ����� ���� �����������
�����:�>���������G�����4�����������������������4��������3H1�����3H-I�2/-*�6I�'&X�TRHK]TRPLI�
/����X�&IX�8�?�)�X�@IS�IX�3�����X�=IX� 6�4����X��IX�*�����X�@IX�@�)��4�X�-IX�����-��4�?���X�@I�
WHOOLNI� ��������������� ���� �4��������?������ ��� �������������� P� ��� �� 4������S:�������
�������������:�>���������������I��� I�+��,�4�������I�,/X�POK]PHOI�
/�)��X�/IX� ����X�;IX�3���>X�/I�IX�@�>�)�,�X�3IX���������X�=I2IX� ���� ���)�X��I8I� WHOORNI�
1������� ���������� ����4������ �������������� ���� ���������X� ���������� �� ��������X� ����
������������������������������)�����>����S����������)��������4�:�����I�-���I�=�����I�)+X�MRG]MRLI�
/�)4��0��X�=IX� -�������X� 6I/I8IX�@���4��X�&I8IX������,���X�/I1IX� ���� 2�,����X� 6I� WHOORNI�
=��SG� �=�����������>� ���������� ������������ ��� ��������� ��������� �� ��������� ������I�
�4�����������*0X�GOG]GOKI�
/��4������X��I6IX�����X�<I/IX�<�X�-IX����X�3IX�/����X�1IX�1���X�8IX���������X�8IX�+����,�>X��I�IX�
3����X� 6IX� +���:��X� �I+IX� ��� ��I� WHOGGNI� ����� ����������� ����� ��� ��2&� ��� �����������
����������S�����������������������������,�4I�������������&-X�KMH]LOQI�
/��4������X��I6IX�@�:�>��4�X�&IX�=�������X�/IX��4�X�1IX�����X�<I/IX�3����X�6IX�+��� �X�/IX�����
1�X� 3I� WHOGHNI� ����� ����� ���� =18U/1�@� ���� ������ ���������� ��� �������� 2/�� ����
����������� ���������� �������������������������U����������������I��������=��I�)'X�GLKL]GLLMI�
/����>X�1I1I�WHOOQNI�=��>�������4��������>���>��>������=� ������I������&&*X�HHG]HPQI�
&���X�8I8IX�����*����X�/I*I6I�WHOOTNI�2����������������4�����������������4���4��>����������
�4����:����>�����������������������������������-HPI�6I������8��I�&&-X�PTKT]PTLRI�
*)���X�/IX�3���X�3I;IX���������X��IX���)�>���X�-IX�8������X�1I+IX��4��X��I@IX�@������X�2I8IX�
3������X� �I1IX�8�>���X� 6I�IX� -� �X�@I2IX������I� WHOOONI�&������4�����U-HP� ����� ����������
��@HU�>�����2��������������������������I������&/+X�GHK]GQOI�
*)�,�)�X� /IX� /��������X� @IX� ���0�����X� /IX� ���� &�����X� @I� WHOOGNI� ;�������� ���
�������4�����U-HPX���������������������������X������4��������4�������I�;2-8�<���I�,/*X�HKH]HKTI�

� � $�(��� ������������������
MK�
�
*)�,�)�X� /IX� ���0�����X� /IX� ���� &�����X� @I� WHOOH�NI� �4�� =&1� :������� ���� ��>� ��� ��
��:������:����������������X��������4�����U-HPX��������������:>��4���4��>�������,��4�
�� ����� �>���S���������� )������ ���� :>� ������������,��4� ���� ��:�>��I�/��I� -���I� ����� &+X�HOGT]HOPOI�
*)�,�)�X� /IX� ���0�����X� /IX� ���� &�����X� @I� WHOOH:NI� �4�� =&1� :������� ���� ��>� ��� ��
��:������:����������������X��������4�����U-HPX��������������:>��4���4��>�������,��4�
�� ����� �>���S���������� )������ ���� :>� ������������,��4� ���� ��:�>��I�/��I� -���I� ����� &+X�HOGT]HOPOI�
����������X� 1IX� -�����X�/IX� 8�4����X� 2IX� ���X� 2IX� �������X� �IX�3������X� 1IX�&�)�>���X�@IX�
&�)�>���X�@I�IX�@4��4:��X�8IX�8����>X� 2IX������I� WHOOTNI�38�HK� �� �����:�C���������������
����������������������������HK@��G�����4�����8S�4������S����>������������������I�;182-�
6I�*��I���:�I�;��I�1�I�8��I�2=�I�-���I�'/X�GGKM]GGLGI�
���������X�/IX�����/��?���X��I�WHOGPNI�+���������>�������������������������������������
������I�/��I�*����I�)X�GMO]HORI�
�����X�=IX�+��X�/IX�14���X� �IX�;������X� 6IX�8���4X�<I-IX�=��X��I8IX�/�@��X�1I-IX�8�>,���4�X�/IX�
-��������X�=I6IX�2�,����X�6IX������I�WHOGPNI�8�����>HX���2&X�������H1����������������������
���������������������������I�6I�����I��� ���I�&'+X�GGRK]GGKRI�
�������X�3I-IX��4����X��I6IX���������)�X�1I=I�WHOOMNI�@S��������1��������������>�����?�����
��������������������������������������4�������I��������=��I�*-X�MQ]GOGI�
�4������X� 1IX� ���� <���X� +I3I� WGMMHNI� &������������� ��������� ������ �4�������� ���
>������������=������I������*-X�KRM]KTKI�
������X�1I/IX�+�����?S�����X��I-IX�8�4������S-��������X�=IX�/������?X�<I1IX������X��I6IX�����
6�4����X��I+I�WGMMLNI�����������2H;G����� ��>����������)������������������4�����?>�����
���������?>����������RPI�����I�&���I�1���I�8��I�-,X�LLRL]LLTPI�
�������)X� �I1IX� ��:����X� �IX� ���� -��4���X� 1I� WGMMKNI� /���� ��������>� ��=� ������
����4��������������������:>���������������������3�=�SGPI��� I��>�I�*��I���:�I�1�I�1����I�
1���I�'/.X�QRQ]QTRI�
�������)X��I1IX�-������X��I3IX��������X�6I;IX�-�)X��I/IX�����-��4���X�1I�WGMMM�NI����������
�� ��������� ��C������ 8����� 4����4��� �=�������� :>� �4�� ����������� ������ ����4�����I�
�� I�-���I�'/-X�HL]PMI�
�������)X��I1IX��������X�6I;IX�����-��4���X�1I�WGMMM:NI�3�=�SGP�����������������������
����:��������������� ���������������������� ��������I�6I�����I�&*&X�GTRR]GTTGI�
��������X�+IX������X�1IS1IX�8����X�8IX�1:���X�1I=IX�/�4����X�1IX�1�� �X�3I1IX�14����X�1IX�����
8������X�3I=I�WHOOMNI�=���������2&�+�������������������������������������I�����I�6I�0X�MR]GOOI�
��������X�6IX���������?��X�+I1I�WHOOTNI�/����������4����������4����4�����4��������>�������
�4�������������������I��������<���I�'+&X�GH]GMI�
�����X� +I8IX� ���� ���?X� *I� WHOOLNI�/��������� ���������� ���4,�>�� �4��� ��������� ���������
�������� ��������I�����������������)*X�TQG]TRMI�

� � $�(��� ������������������
ML�
�
��X� -IX�3����X� <IX� ���� �����X� +I8I� WHOOQNI� 8����� 4����4��S����4��� +��� ���������� ��� �4��
�� ��������������������������>���:�S���������������������:>����������������������������
����������,�4�����:����4���I��� I�-���I�')+X�HRK]HKRI�
��X� -IX� 3����X� <IX� -���4X� <IX� ���� �����X� +I8I� WHOOKNI� 1�������� ����������� ��� ���������
����4����������>� ����� �=��������>� ;��GOS���������� ���� S������������ ���4,�>�I�
2�����������>�&0.X�GTMK]GKOTI�
;��X�6I�WHOOTNI�2����������>��������4����>��4��>I�&������00&X�GQP]GQRI�
=�0��X� =IX� .����������X� =IX� @����X� 8IX� <>��4X� 6IX� �4������X� =IX� ���� �0�)��,X� �I� WGMMTNI�
2������������,�4� ������� W2+;N�����������4������������������4�������������� ������
����� ����X� ���� 2+;� ������������ �� ��������� ���� �������� ��� ������ �������������� ��� �4��
�����������>��4�������������������>�:���I��4�����������'.X�G]MI�
=��X� �IX� 1����X� 6I<IX� ���X� 6I6IX� 3������X� �IX� -���)���X� @IX� ���� ��� +�- �X� 6I� WHOGQNI�
�4���4��>������8�1�P��4>������>�����������,��4�&�/���������������������>���4����������
�=������������������I�*�������I�
=�:?>)X�@IX�*��X�3IX�-��4����X�+IX�-�����X�<I/IX������>X�1I�IX�/�����X�1I@IX�=����X�&IX�����
������X�1I6I�WHOOKNI���������������4������S�����������������4�������S��������������
�4��<KOG3��������������4�������������������I�/��I�����I�2���������I�'*.X�TK]KQI�
=���4�X��IX�-������X�2IX�2�������X�8IX�;�?��X�<IX�.�������X�=I<IX�&�����X��IX�����+��� �X�/I�WHOORNI�
����������3��HK�����������������:���������������������������S������������������������
��� ��������� ������� ��� ������� ������������ ���� ���� ������ ��� �������������� PS���������
������������������������I��������=��I�*,X�GGOLP]GGOMPI�
��� =�>��X�/I2IX� ��� ���������X�1I1IX� ��� .��X� �IX� 3����������X� 1I-IX� ���� �������X� 6I�
WHOGHNI�8���,����������������������������?�����I�6I������8��I�&',X�GMKO]GMKMI�
=�������X�/I1IX� ����1�X� 3I� WHOGPNI� ��2&� ��� ��������� ������I� ��� ��������� ������X� �I6I�
�������X���I�W8��������&�,�-��)NX���I�LK]GPKI�
8�)�X� 1IX� +�X� 3IX� -�X� �IX� 6�X� �IX� ���� 6�X� �I� WGMMPNI� �4�� ���C����>� ��� ���������� ����
���������4��������������������4��������������>������������������I�6I�����I�&,/X�PKM]PLRI�
8�� ���X�/IX�;����X� 1IX�=����X�&IX� -�����X� �IX�����������X�/IX� 1�����X�+I��JX�*��4��������X�+IX�
���4�����X�1IX�����������X�2I�WHOORNI�-���������������� ��������������������I�=���������GP�
�����������������������>��������4������������I�6I�2=�I�����I��������=��I��=�'0X�HOP]HOKI�
8��:���� X� �I�IX� 1��X� 8I/IX� ���� 8�:�����X� �I/I� WHOORNI� +��,���� ������ ���� �4�� ��*=�
���4,�>I�����I�*���I������-���I�&)X�RMT]TOPI�
8��:���� X��I�IX�1��X�8I/IX�8�������X�8IX�84���X�6IS3IX�3��X��I�IX�-����>X�1I;IX�/��)4���X�1I<IX�
����8�:�����X��I/I�WHOOTNI�����������=����>����������������4�:������*=�H�1����:�>�
����1)�U�@-I�/��I������''X�GRM]GTLI�
8���X�@IX�3�>���X�=IX�1�X�1IX�&��4�)�,�X��IX�&��4�)�,�X�3IX�*)���X�-IX�*����X�3IX�;�)���X�/IX�
����*4��X��I�WHOOQNI�&������4�����U-HP������������������:������������4��-=�1GS-1=�G�
�:�C������������I�6I�-���I��4��I�')-X�POMGM]POMHHI�

� � $�(��� ������������������
MM�
�
8� )��X� =I8IX� ���� *����X� /I*I� WGMMLNI� ������������� ���� ���� ��� ���S��:������� =&1�
:>��������-HP�������:���������I�&�������1�����=��I�'*X�QROL]QRGRI�
8�4���:���X��I=IX�����/�C���X�<I2I�WHOGHNI�=��������������>�����������=&1�����>I�&��I�
=� I�+����I�&+X�HQT]HRMI�
8�4��?���X� 6I<IX� ���� /�����X� +I=I� WHOGHNI� 8�����>� ������ ��������� ��� ������������ ���
����������������������I�����I�&���I�1���I�8��I��I�8I�1I�&/-X�HOOHP]HOOHLI�
8�)4��X�@I=IX�-������X�/IX�.��)����,����X�1IX�8���X�8IX�����4���X�&I=IX�����)�X��I1IX�3���X�
8I=IX�+���X�<IX�-�����X�=IX�1::��X��IX������I�WHOGQNI�����������&������4������G�=����������
��=��������8������>�����=���������8������?�����I����I�6I�=�����I�*����I�.-X�GGOT]GGGQI�
8�������X�8IX���������X��I=IX�����8�:�����X��I/I�WHOGONI�=��������������4����*=�������=�
G����4,�>�:>����������X����,�4��������X�����������I�/��I������0/X�PGO]PHHI�
84�����>�X�6IX�8,������4��X�.IX�+����X�8I8IX��4���4���X�=IX�@��������X�+I8IX�����@����X��I@I�
WHOOMNI� 1���>������ &�/G� ������?��� ��� �4�� ������������ ���� ���������� ����������������
���� ������ ��� ������ ����������� ��� ����� ��������������������I�/��I� ����I� -���I� '-X� RGGR]RGHKI�
84����X� 1IX� -��,X�1IS8IX� 2����X� 1IX� �������X� �IX� �����X� &IX� �4���� ��X� �IX�/�������>X� �IX�
�4���X� >IX� �������)X� �I2I8IX�1��)��,��?X� 1I@IX� ��� ��I� WHOGONI� �4�� ������:�������� ������
�������������������������������������������4��������������������������������I�6I�����I�
�� ���I�&'/X�QQKL]QQMHI�
84��X� /I/IX� ���� 1:���S84��X� �I� WHOGONI� /��������� ��������� ��� ��������� ������>� ��,�
�������������������4��������I�+������� I�'0X�GMTK]HOOOI�
84�:�4���X�@IX�����8�������X�-I�WGMMMNI�=����������S�������������)��������&1�:>���&1�
�������������1;SGS����������4�������������4�������I������-*X�RKR]RLRI�
84����X� /IX� -�)���?�X� 1IX� /���:��4�X� �IX� ����X� -IX� ���)��4�X� 6IX� 2��X� /IX� ��4����X� �IX�
;�0�����X�&IX�����&����X�8I�WHOGONI����TO���������������������������������������������:>�
�����������������������������������������4���������I��4�����������)/X�RQO]RRQI�
8���������X�8IX�1�������>X��IX����)�X�6IX������X�-IX�<��X��IX�+����,�>X�<IX�-���X�1IX�/��������X�=IX�
/�@���X�;IX�����<���X�/I�WHOOONI��TP���������������:�������������)������������C����������
����������� ��������I�1�I�6I����4��I�&,)X�GKTM]GKKRI�
8���������X�8IX������X�/I/IX�<�������X�/IX�3�����X�6I1IX�+�����?��X��IX��4��X�8IX�/�0�����X��IX�
/�@���X�;IX�@������X��I1IX�8������X�1I=IX������I� WHOORNI��TP��������������������������4��
���������������������I�����I�&���I�1���I�8��I��I�8I�1I�&/'X�GGPRR]GGPTOI�
8������X� �I@IX� ����1�����X� 6I�I� WGMKQNI� ������������� ;��������� ����/���:������ �������
/����8�=�������������������� ��� �4��3����� 2�:�>�I� 6I� ����I� 2���������I�/���:I�+.X� GGP]GHRI�
8������X�&IX�����&�����0X�=I�WGMMRNI�8��������������I�-��2���>��&�,��=� I�/��I�����I��� I�
-���I�&)X�QGR]QHHI�

� � $�(��� ������������������
GOO�
�
8)0>�4X��I3I2IX�������������X�*IS+I�WHOOTNI����������������������������������������������������
4�����������������������������������:��������������������������������������I����I�6I�*����I�
'.X�HGK]HHMI�
8������X��I�WGMMMNI�+�������?���������=���������,��4��4��=*81HT��������������������I�&��I�
+����I�'&X�KO]KGI�
8���)�X��I1IX�����+������X��I1I�WHOGONI�������������*=>�����������������*=�������=�H�
��4�:�����������������4����>I�*��������'-X�PKPP]PKQQI�
8�� �)� X�/IX� ����;��4��X� 1I+I� WHOOKNI� 2���������� ����������� ��� ����S����� �������>I�&��I�
=� I�+����I�.X�HTP]HKGI�
8���:���X� 8IX�/������X�/IX�/������X� ;I�IX� @���������X� 1IX� 8�� �����X� +IX� ��� -��������X�/IX�
�4�����X��IX�1������X�.IX����?����X��IX� ������X�+IX������I� WHOOMNI�* ���=�������������4��������
�����:�>� ������SG� W�1;SGN��TO� ����������� ����� �� �����:�4� ������������������ ������I�
3�������4����>�,0X�RLO]RLMI�
8���:����4X� /IX� <�� X� �IX� @,��X� �I1IX� -�:��>X� +I6IX� ���� -��)X� 8I�I� WHOOGNI� ����������
���������4�������������������������=����������������������������������������������������
����4�����I�����I�&���I�1���I�8��I��I�8I�1I�-.X�GOLHP]GOLHLI�
8���:����4X�/IX�-�:��>X�+I6IX�=���X�@IX�+���:X��I=IX�=�:��X�/I1IX��������X��I/IX�;�::�X��I+IX�
���� -��)X�8I�I� WHOOTNI� ���������� �=������������������ ��� ������� �������� ���������� ���
������������������������S���������������������������I��������=��I�**X�HLGR]HLHRI�
8�������X�/IX� @��X�/IS<IX� =������X�/I;IX� ���� 84���X� �I6I� WHOOPNI� &��������� 1��� ������
8������������4�:����=�:�������=&1�����������I�/��I������&&X�QGR]QHQI�
8�������X�-IX�;�����X�-I1IX�3��X�-I@IX�<����4��?X�6I3IX�=�:��X�6I8IX�;���4X��I1IX�1�������X�8I1IX�
3�>��4�X�&IX�@�,�����X�6IX��������4�X�+I=I�WGMMTNI�@��������>������,�4��������W@+;N�����
�������������������������4���������:����4��������4�������������4������ ���������������I�
���I�6I��� I�-���I�0/X�MQG]MRGI�
8���� ��X�@I�IX�+������S-�4�X��I<IX�3���>X�=I2IX�;���)��X�6IS<IX�����2�������X�6I/I�WHOGHNI��4��
�RP���������:����I�-���4��I�-���4>�I�1����--1�S�=� I��������&.',X�HHM]HQQI�
8?�:���X�1IX�3��������X�@IX�&���X�8IX�����*����X�/I*I6I� WHOOPNI�=���������������)�������@H�
�4���4��>������� ��� �4������������ �4�������� ���� ��>� ���������������������:HPI� 6I� -���I�
�4��I�').X�MGOK]MGGRI�
��)���X�3IX�<�����?)�X� �IX�����/�?���X��I� WGMLTNI�1���>�����������������:��� ����������:>�
:����������������������������4����������������������������I�6I�2���������I�&&/X�QTK]QKOI�
��)�����X�/IX�8���X�@IX�&��4��X�/IX� 1)�>���X��IX����)�,�X�3IX�����-��4���X�8I� WGMMM�NI�
&�����������������-HPIG�:��������������:���������������������>������������>������������
�&1����>����������4������ ��>I�6I�-���4��I�W��)>�N�&',X�MOQ]MOMI�
��)�����X�/IX�8���X�@IX�&��4��X�/IX� 1)�>���X��IX����)�,�X�3IX�����-��4���X�8I� WGMMM:NI�
&�����������������-HPIG�:��������������:���������������������>������������>������������
�&1����>����������4������ ��>I�6I�-���4��I�W��)>�N�&',X�MOQ]MOMI�

� � $�(��� ������������������
GOG�
�
���X��IX�������X�>I�WHOGHNI�@=18�������������������������������������������������I�1�����6I�
+������������I�16+�&.X�RGKG]RGLOI�
���X�6IX�84�����X�-IX�3����X�@I+IX�+�����>X��I1IX�����X��I-IX�8��X�/IX�+�������)X��I3IX���.����
14���X�=I1IX�������,X��I+IX�3�����X�8I2IX������I� WGMMKNI���4>����������������������� �����
������� ��������� ���������� �=�������� ��� �4�� ��������S���������� 4����� ���������
�������=����������1=HH�����<&���������I�/��I�2���������I�-�����I�/��&&X�QRO]QRMI�
����� �0)��X�@IX�@���)��X�<I/IX�����������X�@I6I�WHOOQNI��>���������������������������������
��������������:���I�6I�����I�-���4��I�-&X�KOT]KGKI�
������X�/IS2IX�-�:��>X�+I6IX�84�����X��I�IX�;����?X�/I2IX�8������X�1I2IX�*����X�+I@IX�@���X�3I&IX�
����-��)X�8I�I�WGMMRNI�/������������4��1�������S=��������+�������/����������1�������S
���������������������������I�&I�2���I�6I�/��I�++'X�GPMP]GPMLI�
������X�/I2IX� -�:��>X�+I6IX�@�X�-I6IX�8����X� 2I6IX������X�/IX�=�0��4)����X� -IX����� -��)X�8I�I�
WGMMMNI� 8��������� ���� ��������� ������������������� ��� ��������� �������� �������� ,��4�
�������������������I��������=��I�,-X�HRGG]HRGRI�
��������X� �IX� 84������X� @IX� 8�?�)�X� 3IX� ��)�>���X� -IX� @��X� 8IS3IX� /�>���X� 1IX� ����
;�)���,�X� @I� WHOOTNI� �4�GMM� �4���4��>������� �������� �������4������ ��� ��������
����)���������������������S�=&1�����������I�;2-8�<���I�,./X�PMM]QOMI�
��>���X� -I8IX� 8�4���?X�&IX�3�����>���X�3IX�+������X� 1IX�>���X�-IX� ��� ��X� -I8IX� 1����X�.I@IX�
@���4�)X��IX�������X�2IX�=� �X�-IX������I� WHOGO�NI� ��������� �����������������������4�����
���������������I�������������&.X�GG]HHI�
��>���X� -I8IX� 8�4���?X�&IX�3�����>���X�3IX�+������X� 1IX�>���X�-IX� ��� ��X� -I8IX� 1����X�.I@IX�
@���4�)X��IX�������X�2IX�=� �X�-IX������I�WHOGO:NI���������� ��+��������������������3�����
���������������I�������������&.X�GG]HHI�
���������X�1I6IX�������X�.IX����4����X�=IX�=��4�������X��IX�-?����4�X��IX���>��X��IX�+�����4�X�
*IX� -�����X� /IX� ;��4���X� 2IX� 8������X� ;IX� ��� ��I� WHOGPNI� �4���� H� ������ ��� ������S������
� ��������� ��� �4����4����>S��� �� ���������,��4� ����������S���������� ��������� �������
W81@@�OLUOLNI�2��I�����I�*0X�GRO]GRLI�
�4�����X� 1I1I� WHOOGNI� =���� ��� ���������� ���� ��:��:����� ���,�4� �������� ��� ����������
�� ��������I�=������������&'&X�GLK]GMRI�
�4�����X� �I�IX�@���X� 8I1IX� �4���X� 6I1IX� <��X�;IX� �4���X� 6IX�+��X� -IX�=���4����X� <I6IX� 8��X��IX�
8�:�����X� �I/IX� ���� +��>X� &I8I� WHOOMNI� 1�� 1��S��������� �� ���������� ������� ���
�����>������4�:������� ����������>���S�������������������������*=�GI�6I�-���I��4��I�'.0X�LOHP]LOPHI�
�����>X� 1I�IX� -��4����X� +IX� 3��)�>X� �I2IX� ���� -�����X� 6I/I� WGMMTNI� /��������� ��� �4��
��������� �����������������������������,��4����������������4�������������� ������� ���
���������������������I�����I��������=��I�*��I�6I�1�I�1����I��������=��I�'X�HKK]HLRI�
�9�@�X� 6I� WHOGQNI��4�� ��������� ���� ��������@=18� �������������� ������S�����S����� �����
������I�����I�*���I�*����I�'*X�GPL]GQQI�

� � $�(��� ������������������
GOH�
�
��)�>���X�-IX�3���X�3I;IX�@�,�����X�@IX���������X��IX�����;�)���,�X�@I�WHOOGNI�8��������
�4���4��>�����������������4����������4�WGMMN�:>��>����S����������)������HS�>�����2�
��������������������������������������I�6I�-���I��4��I�')*X�HGRHM]HGRPKI�
�������X� 8I1IX�/�4��X�=IX�=4����X� �I=IX� ���X�>IX�1���X� <IX� �4�����)����X� 8I/IX�@��>���S
8�������X� 8IX�1��X� 6I�IX� =�:��X� /I1IX� ������X� @I6IX� ��� ��I� WHOOKNI� ��������� ������������
�����������������������������������������������I�&��I�+����I�+-X�QG]RGI�
���������X� �I�IX� ;��������X� 8I3IX� +����X� *I�IX� ���� �4�����X� 1I1I� WHOOQNI� �������������
����������� ����������������,�4� ������S:���G��������������������������� ��� �4���� ��������
��������I�2�����������>�&0,X�QHMH]QPOOI�
�>�����X�&IX� 6�����X� <I.IX�1��X� 6I+IX� 8>������X� ;I-IX� �0��4���X� 6I�IX� ����&�=>X� 2I� WHOOGNI�
�4������ ���������� ,��4� ���������� ���,�4� ������� �������� ���,�4� ��� �4�� ���� �������
��������I�8����I�6I�����I�&��4���I�+,X�PPM]PQQI�
�����X� �I@IX� <��X� 3IX� ���� �4���X� >I;I8I� WHOGONI���*=� :����� ��� �4�� ���������� ��� =&1�
���>��������S��������S��������:��������I�������>����-X�MRP]MRKI�
���4����X�6IX�@������X�<I=IX���-���X�+IX�/��� �S����?���X��IX��:������,�)�X��IX�-4�����4��>�X�
&IX� ���?� ����X�+I�IX�@���?X� <I3IX� ���� 8����������X� 6I/I� WGMMLNI� <���� ��� �>����S����������
)������ ��4�:����� �HK@��G� ��� �� �� ��� ����������� ������� ��� ������?��� 4����� ���������
��������������I��������=��I�,.X�RQH]RQLI�
�>���X� =I@IX� <� �� �)>X� -IX� 14�X� 8I�IX� 8���X� �I8IX� �4�����0��X� -IX� ���� =�>X� 1I@I� WHOOONI�
�>������� ��� �������������� �� ������ ���� �������>���������� ���>������ ��� �4�� ������S
���� ���������������������������� ���������I�/��I�2���������I�-�����I�/��&0X�GGTH]GGKQI�
���)�,�X�3IX��4���X�6I3IX��������X�6I6IX�1���X��IX�1��������X��I�IX�����*����X�/I*I�WGMMPNI�
&�����������������-HP>�:����������=��������X�������������X����������?������������������>�
���������������,����������I�����I�/��I�-���I�=��I�+-X�TPR]TQRI�
.�����X� 8IX� ;�� ��X� 8IX� 1������X� �IX� ����=�>����X� 2I� WHOORNI���*=S��������� �4����>� ���
�������,��4� �����>�������� ��� ��I� 1��I�*����I�*��I� 6I� 2��I� 8��I�/��I�*����I� 28/*� &*X�RHR]RPKI�
.���)����X��IX�3>>�����X�2IX�@�� ����X��IX�������X�/IX�@���?���X�=IX�����:���X��IX��������X�1IX�
�������X��IX������X�6IX�����@����������X�*IS�I�WGMMRNI���� � ��������������������4�����������
���������������������������������4��������������������I�&��I�+����I�-X�QOG]QOTI�
1�������>X� �IX� 1����X� -IX� �������X� &IX� ��� <� ��X� 6IX� ���� �������� �X� .I� WHOOHNI�
* ���=�������������4��4����:�=������3*>�L����4��������������������������������,��4�
�����������������������������I��4�����������,/X�GTH]GTMI�
1���X� -IS2IX� 84��X� 6IX� =���X� 8IX� @������X�3IX� ��� 8�� ���X� ;I6IX� ���� +��X�1IS;I� WHOOP�NI�
��4�:�������������4�������������:����4���� ����4�����������:>�������4����4��� ��� ���������>�
���������:>��������������I�6I�-���I��4��I�').X�GLROT]GLRGPI�
1���X� �IX� -������X� 1IX� 8?�:���X� 1IX� ����*����X�/I*I� WGMMQNI��4�� �������� ����� :�������
���� ��>����������������������-HPIG�������������������:�=>�S������������I�6I�-���I��4��I�'*-X�POMMQ]POMMLI�

� � $�(��� ������������������
GOP�
�
1���X� ;IS;IX� �4���X� �IS-IX� >���X� 6IS-IX� -�X� �IX� <�X� <IS�IX� 3����X� -IX� ���� -��X� 6IS�I� WHOGGNI�
@���)��,������������4��������������8S�4��������������3��+H������I��4��I�6I��������+/X�LRP]LTOI�
1���X� 8IX�+��X� 6IX�<��X�;IX�=�?������X�&IX������4���X� �IX� 6���X� 6IX��4����X�+I.IX�<�X�+IX�=�>S
-�����X� �IX�&�����X� �I8IX� ��� ��I� WHOOP:NI� ��������S��������� ��������� ��� �4��������� �����
���������������������������������������������������������I�������������0X�HOM]HHGI�
1���X� -IX�3�>,���X� 8IX� ���X�/IX� �4�>��X� @IX� ���� ���4�X� +I� WHOOGNI� ����� ����������������
������������4����������I�������I�=��I�-���I��� ���I�*.X�HKO]HKMI�
1�:��X�/I6IX�����+�����X��I� WHOOQNI�=������������� ������������������������������I� 6I� ����I�
-���4��I�-&X�GP]HRI�
1�:��X� 6I�IX���>���X�<I6IX�=������X�/I;IX�84���X��I6IX�����-��S8���X��I� WGMMMNI�&���������1���
��C��������/��H��������� ������RPI�&��I������-���I�&X�HO]HTI�
������1�0������X��I6IX���::��)X�3I6IX� ���=�>��X�/I2IX��������X�6IX�����6������X�+I�WHOGHNI�
1�������� ��������� ������������>� ������������ ��� �4�� ������ �����:������� ���� �I�/��I�
����I�2���������I�+,'X�RK]TMI�
1�����X� 6I�IX�����<�����?)�X� �I� WGMKGNI���4>���������������� ���������� ��� ������ �����������
�4����::����������I�2�����������>�.-X�TRM]TTLI�
1����)���X� �IX� �������X� �I�IX�+�����X�;I<IX�����8�:��X�<I3I� WHOOHNI��&/� ��������� ������
����������������� ������I��������-0X�HRGG]HRGTI�
1��?��X��IS�IX�@�4X�<I;IX��������)���X�&I�I�WHOGONI�=��������������>������G�������=��������I�
-���4��I�8��I������I�+.X�HGKI�
1���X�6I�I�IX�3����X�/I=IX�=�4���X�/IX�-�X�1I�IX��4��X�8I@IX�8�4������X��I;IX�@�����)�X�3I;IX�
<��X�3I6IX�*,��X��IX��������X��I�I�WHOGPNI�&������4������GX�����������������������������
�������� ��� �4�� �����X� ��4�:���� �RPS��������� ��������� ����������I� ���I� 6I� ������� 6I� ���I�
�������&++X�GRTK]GRKKI�
1�X�<IX�+�����>��X�1I.IX�<�X�-IX�3��X�-IX�������X�/IX����������?�X��I6I�WHOGHNI�8��������������
�����������4�:���������8)�HS����������HK������������I��4��I�-���I�&-X�GRGR]GRHQI�
1�X� /I3IX� �4���X� 6I3IX� �4��X� �I�IX� ���� -���X� -I-I/I� WHOOHNI� �� �� ������ ���
�������4�����U-HP�����4��������������3�<������������.������������I����I�6I��������6I����I�
�������-)X�HMK]PORI�
>�X�6IX��4���X�8I<IX�������X�8I�IX�1���>X�@I2IX�1�)����X�;IX�8��X�6IX�@����X�1I@IX�<�X�+IX��������X�
<I�IX�@��X�8IS�IX������I�WHOGONI���4��������������� ������������������������������ ��:�������
������������������������I�����I�&���I�1���I�8��I�&/)X�HGPT]HGQOI�
-������4�X� �I�IX� -�����>X� 1IX�/�/�4��X� 1I�IX� ���� 6����X� 8I� WGMMMNI� 1�1��R�� ���4,�>�
����������������,�4����������������������������4�� ����:�������:�>�I��� ���������&'*X�GHGG]GHHPI�
-��X�+IX�;�)�:���X�-IX�&�)�����������X�8IX�1���X�;IX�����/�@��4��X�1I<I�WGMMHNI�3������S
:�������)��������>������,�4������������������������������S��S����4�����S���������������I�
/��I�2���������I�-�����I�/��*X�HGHP]HGHLI�

� � $�(��� ������������������
GOQ�
�
-���X�+IX�1>���X�+IX����/��?�X�1IX�����X�1IX�;���� X�1IX�14�����X��I/IX��4������X��I�IX�����
3�����X� 6I1I� WHOOHNI� 2�� ����� 8)�H� �������� �=��������� ��� 4����� ��������� ������>�
������������,��4� �������� �4���>����S����������)������ ��4�:������HK�������2&�����,��4�
������������������S�������� � ��I�����I��������=��I�*��I�6I�1�I�1����I��������=��I�.X�PQGM]PQHTI�
-���X�=I/IX�&����4X�6IX�/���4>X�/IX�1���X�3I6IX��4��������X�6IX���@������X�6I-IX�<���X�/IX�����
=�����X�=I2I� WGMMLNI�<�,��HK��=������������������������������S�������� � ��� ������������
,��4����������������I�6I�����I�&,-X�MQG]MQRI�
-��X�6IX�;�X��IX�����X�>IX�+��X��IX�������)�X�1IX��4��X�-IX�14���X�@IX�<���X�6IX����X��IX�����-��X�>I�
WHOOQNI�&�)H1�)���������������� �4�� ������?������������������ �������������� ����������I�
;2-8�<���I�,),X�GGH]GGLI�
-�4X� 8IX� ���� �4���X� �I� WGMMTNI� �������� ���� �4��������?������ ��� �� ��������� ������ ����X�
1=1KOX������4�����������������������4�������������������I�����I�&���I�1���I�8��I��I�8I�1I�
-+X�RRGK]RRHGI�
-��X� �I@IX�/�����X� @IX�1��?X� �IX� 8�:�����X� �I/IX� ���� @���X� 8I1I� WHOGONI� 8��������� ��� �4��
3�������*=� ������=� �� ���� ���� ������������� ����=����>���� ��4�:�����I�/��I� ����� +.X�KTL]KKQI�
-�)�����X�@IX�1�)���)�X�8IX�1 ���4X� 6IX�����=�� ��X�8I1I� WHOOONI�8�������4���4��>�������
������=��������� ���������8�1�P���������&�;����������� ������������:>��4�������>����
���������*=I�����I�-���I��-�&/X�QK]ROI�
-��X�6IS-IX�<��X�-I6IX��4��X�3IS@IX�3���X�8I1IX�6���X�3I8IX�@��X��IX��4��X�@IS3IX��4��X�@IS�IX�����
-���X� 3IS+I� WHOGPNI� �@HS&��=� ���������� �������� ��������� ��������� �������������I�
*����������0X�MKH]MLPI�
-�����X��IX�<��X��IS3IX�*��X�8I3IX�1���X�2I8I/IX�<��X�6IX�<�X��I<IX�<����X�3I;IX�;���X��I1IX�����
+�>X� +I=I� WHOOHNI� 8�����>H� ��4�:���� �4�� =��U/1�� )������ ���4,�>� :>� ��4�:������ �4��
���� ���������=��I�6I�-���I��4��I�'))X�PGMR]PHOGI�
������X�&IX�+�:��X�.I<IX������X� 6I1IX�������,���X�8IX�����84�����X�/I-I� WHOOTNI�����������
4���� �4��)� ��������� ��� �������?�� ������� ������ ��� ����������� ���� 3��MO� ��4�:�����I�
�������=��I�**X�GKLP]GKMGI�
����X�-IX�*���)��X�@IX�3�� ��4X�<I+IX�8��4������X�=I<IX�����1�������X�2I�I�WHOORNI�<>��4�����
������������>������>��4�����������������������������������I��4�����������*,X�HHH]HPOI�
������:���X�1IX�<������X�*IX�����1����X�@I+I�WGMMRNI�14�������4�������������������V�����I�
*���I������-���I�)X�LPR]LQHI�
�4���X� 3IX� 84�X� >IX� ������X� 3IX� 3������X� /IX� ���X� 1IX� ���� �����4X� 8I� WHOOQNI�
-HPU�������4������ ������� Q� �4���4��>������������������������ ���������� ��� ����S��)��
)������GI�6I�-���I��4��I�')-X�PRKHT]PRKPQI�
�4���X�6IX��4����X��I�IX�@�����X�8IX�����/�����)X�=I6I�WHOOONI�1��������������������:�����
�����������������4��4��� ���������������S���������������=����������4����4������������:>�
��������������������������������� ������������ � �I�2�����������>�&0&X�QTML]QKGOI�

� � $�(��� ������������������
GOR�
�
�4��X�>I-IX�/����>X��I6IX�@���4���X�1I.IX�8,���X�8IX�&� ���X�&I/IX����4�X��I/IX�����;������X�
�I� WHOOONI� +��������������� ���� �������� ��������S������������ ���,�4� ��� ���������
��������������4����4����������������������������I�&��I�/��I�*X�KOP]KOTI�
�4��X�+IX�3��4�����X�-IX�@,�)X� �IX�����X� 8I-IX� ��������X�/IS6I� WHOOPNI�=���� ��� �4�� ��������
��������������� �����8=�SP������������,�4I�/��I�����I�-���I�'+X�KKQH]KKRRI�
�4��X��IX�;���)��S&�)����X�1IX������>X��I�IX�1���X�1IX�+������4X��I1IX�=�>S-�����X��IX�����
&�)����X�1I-I�WHOOTNI�8>����>�����RP�����=:����������>������������������/�����/���������
/�������������������������I��������=��I�**X�KLLM]KLMLI�
�4��X��I>IX�8��X�/IX�8�������X�6I1IX�<���X�/I.IX�����1�����X�2I/I�WGMMQNI�1�������S����������
:��������������������������������������4��4����������������������I�=�C��������������4��
�&1S:������� ������� ���� ����������� :>� &3HS��������� ���� ���:�=>�S���������
��C������I�6I�-���I��4��I�'*-X�GPGGR]GPGHPI�
�4��X� �I>IX�@���������X� 6I1IX� ����1�����X� 2I/I� WGMMRNI� ��������������� ��� �4���� �������S
����������4���4��>�����������������4��4����������������������I�/��I�2���������I�-�����I�
/��-X�TOR]TGRI�
���������X� 6IX� 1���4�X� 1I�IX� ���������S��)�X� 6IX� &������X� <IX� ���� +���������X� 6I1I� WGMMRNI�
/���������� ��� �&1S:������� ����������>� ,��4��� �4�� �������� ��������� �����>� :>�
��:������������������������������������������I����������'&X�RK]TKI�
���?����X�.IX�8����)�X� �IX�*�������X�1IX� ����3���X�/I&I� WHOGGNI� 1��� �����������*=�H�:>�
������������,��4��4����:�����I������&00X�KRK]KTLI�
���X�3IX�<�X�-IX�<��X�>IX�����1���X�>I�WGMMMNI�1��1�1;SGI�>���4������������������������=�
����������������������������4������� ���������������SMI�6I�-���I��4��I�')0X�GGRQM]GGRRTI�
��
�





























