Embed Size (px)
Citation preview

Human Herpesvirus 6 Infection in Neoplastic andNormal Brain Tissue
Laura Cuomo,1* Pankaj Trivedi,1,2 Maria Rosaria Cardillo,2 Franco Maria Gagliardi,1,3
Aldo Vecchione,2 Riccardo Caruso,1,3 Antonella Calogero,1 Luigi Frati,1,2 Alberto Faggioni,2 andGiuseppe Ragona1,2
1Istituto Neurologico Mediterraneo Neuromed, Pozzilli, Italy2Dipartimento di Medicina Sperimentale e Patologia, Universita “La Sapienza,” Rome, Italy3Dipartimento di Scienze Neurologiche, Universita “La Sapienza,” Rome, Italy
The presence and variant distribution of humanherpesvirus 6 (HHV-6) was investigated by anested polymerase chain reaction (PCR) in 118biopsies from patients affected by nervous tis-sue tumor (115 primary tumors and 3 metasta-sis) and in 31 autopsy samples from the brainof healthy individuals. HHV-6 DNA sequenceswere detected in normal and neoplastic nervoustissue at a frequency of 32% and 37%, respec-tively. In both tissues, variant A was three timesmore frequent than the variant B. Peripheralblood lymphocytes (PBLs) derived from seventumor affected patients contained the same vari-ant as their respective brain sample, as judgedby PCR. The expression of HHV-6 encoded im-mediate early protein p41 was detected by im-munohistochemistry in neoplastic but not innormal brain. This may reflect viral reactivationfrom latency in immunocompromised patients.The seroepidemiological data indicated a fre-quency distribution of anti-HHV-6 antibodies inpatients with brain tumors similar to that foundin healthy donors. J. Med. Virol. 63:45–51,2001. © 2001 Wiley-Liss, Inc.
KEY WORDS: HHV-6; brain tumor; polymer-ase chain reaction
INTRODUCTION
Human herpesvirus 6 (HHV-6) is a ubiquitous lym-photropic herpesvirus, associated etiologically with ex-anthem subitum [Yamanishi et al., 1988]. Two variantsof HHV-6, A and B, have been isolated, which differ interms of biologic, immunologic, and molecular charac-teristics [Ablashi et al., 1993]. Variant A includes iso-lates derived from patients with lymphoproliferativedisorders [Salahuddin et al., 1986]. Variant B has beenfound mostly in children with exanthem subitum[Ablashi et al., 1988; Dewhurst et al., 1993], but it alsoincludes some isolates from adults, such as Z29 iso-
lated from a Zairian patient with AIDS [Black etal.,1989].
Currently the encephalitogenic potential of HHV-6and the association of the virus with nervous systempathology is controversial. It has been suggested thatHHV-6 invades the central nervous system (CNS) dur-ing active infection where it can persist in a latentform. HHV-6 can be reactivated in infants with roseola[Caserta et al., 1994] and in immunecompromised pa-tients, such as AIDS patients [Knox and Carrigan,1994], bone marrow transplant [Drobisky et al., 1994],and multiple sclerosis patients [Merelli et al., 1996],causing meningitis and occasionally fatal encephalitis.In a large proportion of multiple sclerosis patients,HHV-6 was also detected in active plaques [Challoneret al., 1995].
In vitro studies on the neurotropism of HHV-6 havedemonstrated that it can infect glioblastoma and neu-roblastoma cell lines [Ablashi et al., 1988; Levy et al.,1990], embryonic glia [Tedder et al., 1987], and pri-mary human fetal astrocytes [He et al., 1996]. Thesecell cultures are less permissive than B lymphocytesand viral replication less efficient.
The association of HHV-6 with various human tu-mors has been described [Josephs et al., 1988; Torelli etal., 1991; Bovenzi et al., 1993; Yadav et al., 1994]. Invitro studies also indicated that specific viral DNAfragments may possess tumorigenic potential [Raz-zaque, 1990; Razzaque et al., 1993; Kashanchi etal., 1997]. HHV-6 genomes were more frequent in nor-
Grant sponsor: Ministero della Sanita; Grant sponsor: Minis-tero dell’Universita e della Ricerca Scientifica e Tecnologica;Grant sponsor: Associazione Italiana Ricerca sul Cancro (AIRC);Grant sponsor: Istituto Superiore di Sanita; Grant sponsor: Pro-getto AIDS; Grant sponsor: Istituto Pasteur Fondazione Cenci-Bolognetti.
LC and PT have equally contributed to this work.*Correspondence to: Laura Cuomo, Istituto Neurologico Medi-
terraneo Neuromed, Localita Camerelle, 86077 Pozzilli (Isernia)Italy.
Accepted 26 January 2000
Journal of Medical Virology 63:45–51 (2001)
© 2001 WILEY-LISS, INC.

mal compared to neoplastic brain [Luppi et al., 1994,1995]. In both studies, however, the number of casesexamined was limited.
The unresolved issue of the association of HHV-6with nervous tissue was studied in 115 primary braintumors and 3 brain metastasis for the presence ofHHV-6 DNA by nested PCR. As control, 31 tissuesamples from different regions of normal brain ob-tained at necroscopy were used. All positive cases weretested further for virus variant identification. HHV-6infection was also investigated by immunohistochem-istry in brain tissue and by serology.
MATERIAL AND METHODSPrimary Tumors, Normal Tissues and Cell Lines
Fresh biopsies were obtained from 115 primary braintumors and from 3 brain metastasis. The age of thepatients ranged from 15 to 83 years. Specimens werecollected, quickly frozen in liquid nitrogen, and kept at−80°C until the time of examination. Thirty-one forma-lin-fixed, paraffin-embedded autopsy brain sampleswere taken from histological archives. The sampleswere obtained from adult immunocompetent subjectswho were free of clinical signs or records of inflamma-tory or systemic diseases associated with the centralnervous system. The histological diagnosis of the 115tumors was as follows: 31 glioblastomas, 2 neuroblas-tomas, 35 meningiomas, 2 ependimomas, 6 astrocyto-mas, and other 39 histological subtypes including cor-domas, choroid plexus papillomas, adenocarcinomas,adenomas, neurinomas, and neurilemmomas. The im-mature T cell line HSB-2 [Adams et al., 1968] wasmaintained in RPMI 1640, supplemented with 2 mMglutamine, 100 IU/ml penicillin, 100 mg/ml streptomy-cin, and 10% fetal calf serum. HHV-6 infection wasperformed by incubating 5×106 cells with 1 ml of stockvirus (titer 105 TCID50) for 2 hours at 37°C. The cellswere washed and resuspended in complete medium.
DNA Extraction
Three single (5–10 mm thick) sections were cut fromeach specimen with a single disposable blade. The firstand the third of each mirror section was retained forstandard histological staining. DNA from all sampleswas extracted by incubation of the remaining section at60°C for 4–12 hours in digestion buffer (20 mM EDTA,50 mM Tris-HCl pH 8, 0.5% SDS, and 200 mg/ml pro-teinase K). After digestion, the samples were extractedonce with phenol, once with phenol/chloroform (1:1),and once with chloroform/isoamyl alcohol (24:1) andprecipitated with ethanol. Peripheral blood lympho-cytes were isolated from heparinized blood by centrifu-gation on Ficoll-Isopaque gradients [Boyum, 1968] andDNA was extracted as described above.
Paraffin was removed from paraffin-embedded speci-mens with xylol, ethanol, and acetone. After digestionwas complete, a prewarmed saturated 5 M NaCl solu-tion was added to a final concentration of 1.5 M [Mul-lenbach et al., 1989]. The proteins were extracted by
adding 1 vol of chloroform, mixing for 30–60 minutes,and centrifuging for 10 minutes at 2,000 r.p.m. Thesupernatant was precipitated by adding 2.5 volumes ofethanol. The pellet was washed in 70% ethanol andresuspended in 100–200 ml TE.
HHV-6 DNA PCR
One microgram of each DNA samples was submittedto nested PCR where a 287 bp outer fragment and a163 bp inner fragment of HHV-6 DNA was amplified.The resulting product contained a Ban I and a Hind IIIdigestion sites present selectively in “A” and in “B” typeof HHV-6, respectively. Hind III digestion yielded twofragments of 66 bp and 97 bp; Ban I digestion produceda 90 bp and a 73 bp fragments. These enzymes wereused to identify the type of HHV-6 amplified. Primersequences and PCR conditions are essentially as de-scribed previously [Lyall and Cubie, 1995]. The prod-ucts of digestion were run in 3% agarose gel. The DNAfrom viral positive specimens was also analyzed bySouthern blot hybridization by using an 8.7 Kb HindIII fragment (pZVH14) as a probe from the HHV-6 G.S.strain [Josephs et al., 1986].
Immunohistochemistry
Three serial sections (3 mm thick) from paraffin em-bedded blocks were cut onto slides coated with 3-ami-nopropyl-ethyl-xylene (Sigma Chemical Co., St. Louis,MO) and stained by using the Dakow En Vision™working procedure to provide better sensitivity tostreptavidin-biotin method. Deparaffinized sectionswere treated with 0.3% hydrogen peroxidase (H2O2) inmethanol for 10 minutes to block endogenous peroxi-dase. To eliminate non-specific staining, the sectionswere then incubated for 20 minutes with normal serumfrom the species in which the secondary antibody wasmade. The sections were placed in a microwave ovenfor 5 minutes at 750 ww (two cycles) and incubatedsubsequently for 30 minutes with the MoAb p41/38[ABI, Columbia, MD; Cheow and Balachandran, 1991]that was directed against p41 nuclear protein presentin both variants A and B of HHV-6. They were incu-bated further with EnVision™ (Dako Immunochemi-cals, Carpinteria, CA) (1:500) for 30 minutes. Diamino-benzidine-hydrogen peroxidase (Sigma) was used as achromogen substrate and the sections were then coun-terstained with Mayer’s hematoxylin. For negativecontrols, the primary antibody was replaced by normalswine serum. HSB-2 cells infected with HHV-6 (GS)were used as positive control. The estimated visual p41intensity of immunostaining was graded on an arbi-trary four-point scale: 0 (no staining); 1+ (weak stain-ing); 2+ (moderate staining); and 3+ (strong staining).The distribution of immunostaining was evaluated asfocal (positive cells scattered or localized or less than20%) or diffuse (widespread immunostained cells andmore than 20%).
46 Cuomo et al.

HHV-6 Serology
Serum samples were collected from 73 patients withbrain tumors and from 150 healthy subjects, and storedat −20°C. Sera were diluted serially and tested for thepresence of antibodies against HHV-6 by indirect im-munofluorescence assay on HSB-2 cells infected withHHV-6 (GS-strain) as viral capsid antigen source[Ragona et al., 1994]. Mock infected cells were used toevaluate nonspecific background fluorescence. Anti-body titers were expressed as the highest serum dilu-tion yielding detectable immunofluorescence on an ap-preciable number of the antigen positive cells.
RESULTSDetection of HHV-6 DNA in Normal and
Neoplastic Human Brain
To assess the sensitivity of the nested PCR, theHSB-2 cell line and peripheral cord blood lymphocyteswere infected, respectively, with HHV-6 variant A (GS)and variant B (Z29). When 90% of the cells were posi-tive for HHV-6-p41 antigen, cells were diluted stepwisewith uninfected cells from the same source and DNAwas extracted. Between 0.02 and 0.2 HHV-6 infectedcells were detected among 10,000 negative cells.
The tumor specimens (115), 3 metastasis, and 31 for-malin fixed, paraffin-embedded normal brain tissueswere examined for the presence of HHV-6 DNA by us-ing nested PCR, followed by Southern Blotting and hy-bridization with an internal oligoprobe. HHV-6 DNAsequences were detected in 43 tumor samples (37.3%),which included 14 of 31 (45%) glioblastomas, 2 of 2(100%) neuroblastomas, 9 of 35 (25%) meningiomas, 1of 2 (50%) ependimomas, 2 of 6 (33%) astrocytomas, and15 of 39 (26%) other nervous system neoplasms (TableI). Ten out of thirty-one samples from normal braintissue were found positive for HHV-6 (32.2%). Threebrain metastases were also tested and found negativefor HHV-6. The amplification products consisted of a287 bp outer amplimer and a 163 bp inner fragment.
A number of quality controls were set up to excludefalse positive results. First, the specimen sectioning,amplification, and detection procedures were per-formed in separate laboratories. Second, up to 50% ofthe total samples included in each PCR experimentconsisted of a mixture of reagent blank controls with-out DNA and samples containing DNA extracted fromHHV-6 negative cell lines. Third, each tissue specimenwas run in triplicate and when found positive in one, itwas rerun until concordance of the triplicates.
HHV-6 Variant Distribution in Human BrainTissue and in Peripheral Blood Lymphocytes
In order to identify which HHV-6 variant was pres-ent in the brain tissue, the 163 bp nested PCR am-plimer was digested with Ban I and Hind II restrictionenzymes. Ban I digests selectively HHV-6 type A DNAyielding two fragments of 90 bp and 73 bp, respectively.Type B, but not type A, variant contains an Hind IIIdigestion site that gives rise to two fragments of 66 bpand 97 bp as described by Lyall and Cubie (1995).Among the 43 HHV-6 DNA positive tumor samples, 33(76.7%) contained type A and 10 (23.2%) type B variant(Table, Fig. 1). A similar distribution was found in nor-mal brain tissues: type A was detected in 7 (70%), andtype B in 3 (30%) out of 10 HHV-6 positive specimens(Table I).
After primary infection, HHV-6 is maintained in alatent state in the peripheral blood of most healthyadults [Di Luca et al., 1996]. In order to evaluatewhether the HHV-6 variant detected in the brain wasthe same as that present in latently HHV-6-infectedPBL derived from the same donor, PBLs from 7 out ofthe 115 patients studied were tested. Six patients hadHHV-6 type A and 1 HHV-6 type B in the respectivebrain tumor. The nested PCR analysis followed by re-striction of endonuclease digestion showed that theHHV-6 variant detected in the blood was the same asthat detected in the respective brain (data not shown).
TABLE I. Presence and Variant Distribution of HHV-6 DNA in Normaland Neoplastic Brain Tissues
DiagnosisNumberof cases
PCRHHV-6+
HHV-6 VARIANTSA B
Primary tumorsGlioblastomas 31 14 13 1Neuroblastomas 2 2 1 1Meningiomas 35 9 7 2Ependimomas 2 1 1 0Astrocytomas 6 2 0 2Othersa 39 15 11 4
TOTAL 115 43 (37.3%)b 33 (76.7%)c 10 (23.2%)cMetastasis 3 0 0 0Normal brain 31 10 (32.2%) 7 (70%) 3 (30%)aOther histological types include: schwannoma, neurinoma, cordoma, choroid plexuspapilloma, adenocarcinoma, carcinoma, adenoma, and neurilemmoma.bNumbers in parenthesis express the ratio among number of HHV-6 positive cases/total number of examined cases × 100.cNumbers in parenthesis express the ratio among number of HHV-6 variant positivecases/total number of HHV-6 positive cases × 100. HHV-6 variant has been character-ized by cleavage with Hind III and Ban I restriction enzymes.
HHV-6 in Normal and Neoplastic Brain 47

Immunohistochemical Detection of HHV-6-p41Antigen in Brain Tumors
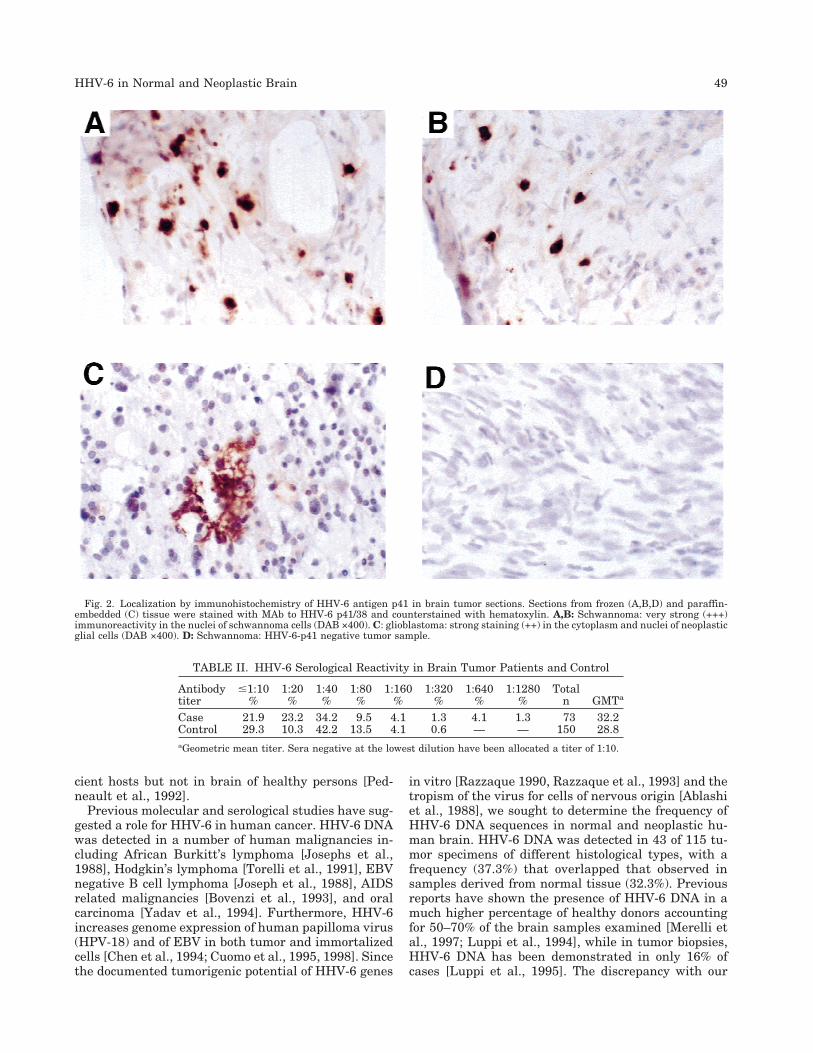
To establish whether HHV-6 was present in a repli-cative form and to identify the viral localization in thebrain, immunohistochemistry was carried out by usinga mAb against the DNA binding early-lytic protein p41.Sections from six HHV-6 positive tumor samples andfour HHV-6 negative samples were examined. The sec-tions were obtained either from paraffin-embedded orcryo-preserved samples. HHV-6 positive cells werefound in four out of six samples positive by PCR. Thep41 antigen was detected as focal or scattered stainingwith nuclear or nucleo-cytoplasmic localization in theneoplastic cells (Fig. 2). The endothelial cells were alsofound to express p41 in glioblastoma tissue sections.The inflammatory cellular infiltrates were not stainedby p41 MAb. Counterstaining with hematoxylinshowed that the sections contained only neoplastic tis-sue. None of the seven HHV-6 DNA positive controlbrain samples examined was found positive for HHV-6-p41 (not shown).
HHV-6 Antibody Detection
Immunofluorescence was employed to investigatethe antibody status to HHV-6 in 73 out of the 115 pa-tients studied compared to 150 age-matched, healthyindividuals. The antibody titer was evaluated in eachserum sample against cells infected with the HHV-6GS strain [Gentile et al., 1999]. Evidence of the pastHHV-6 infection was found in the majority of the pa-tients (78.1%) and controls (70.7%). All patients who
were HHV-6 DNA positive had detectable levels ofHHV-6 specific antibodies. The percentage distributionof HHV-6 antibodies is shown in Table II. The geomet-ric mean titers (GMT) did not differ significantly be-tween the two groups examined. However, only serafrom tumor patients had antibodies at higher dilutions.The GMT of sera taken from HHV-6 DNA positive tu-mor patients was 26.4 (antibody positive dilutionsranging from 1:20 to 1:80).
DISCUSSION
A role of viruses in brain tumorigenesis has longbeen suspected but never established. Recently, the hu-man BKV and JVC polyomaviruses, which are ubiqui-tous in the human population, and the simian polyoma-virus SV40 have been considered as possible cofactorsin the pathogenesis of human brain tumors [Monini etal., 1995]. BKV sequences were detected in normal andneoplastic human brain, while JCV DNA was foundonly in normal brain tissue [Corallini et al., 1987; Dor-ries et al., 1987]. Also, SV40 Tag coding sequences werefound in ependymomas, choroid plexus papillomas, andother brain tumors but not in normal brain, which sug-gests that this virus is associated with human neo-plasm with a high frequency [Martini et al., 1996; Bar-banti-Brodano et al., 1998]. Among herpesviruses, onlyherpes simplex virus 1 and HHV-6 are able to establishlatent infection in human brain and they can be reac-tivated causing encephalitis [Fraser et al., 1981; Ca-serta et al., 1994]. Epstein-Barr virus (EBV) DNA hasbeen demonstrated in pathologic brain of immunodefi-
Fig. 1. Variant characterization of HHV-6 DNA detected in tumor brain samples by polymerase chain reaction followed by endonucleasedigestion and resolution by agarose gel electrophoresis. GS strain, prototype variant A; Z29 strain, prototype variant B; 1,3: HHV-6 variant Apositive samples; 2: HHV-6 variant B positive sample; 4,5: negative test samples from tumor brain biopsies. As negative control, the DNA ofHSB-2 cell line was used. MW, molecular weight standards, 100 bp DNA ladder.
48 Cuomo et al.
cient hosts but not in brain of healthy persons [Ped-neault et al., 1992].
Previous molecular and serological studies have sug-gested a role for HHV-6 in human cancer. HHV-6 DNAwas detected in a number of human malignancies in-cluding African Burkitt’s lymphoma [Josephs et al.,1988], Hodgkin’s lymphoma [Torelli et al., 1991], EBVnegative B cell lymphoma [Joseph et al., 1988], AIDSrelated malignancies [Bovenzi et al., 1993], and oralcarcinoma [Yadav et al., 1994]. Furthermore, HHV-6increases genome expression of human papilloma virus(HPV-18) and of EBV in both tumor and immortalizedcells [Chen et al., 1994; Cuomo et al., 1995, 1998]. Sincethe documented tumorigenic potential of HHV-6 genes
in vitro [Razzaque 1990, Razzaque et al., 1993] and thetropism of the virus for cells of nervous origin [Ablashiet al., 1988], we sought to determine the frequency ofHHV-6 DNA sequences in normal and neoplastic hu-man brain. HHV-6 DNA was detected in 43 of 115 tu-mor specimens of different histological types, with afrequency (37.3%) that overlapped that observed insamples derived from normal tissue (32.3%). Previousreports have shown the presence of HHV-6 DNA in amuch higher percentage of healthy donors accountingfor 50–70% of the brain samples examined [Merelli etal., 1997; Luppi et al., 1994], while in tumor biopsies,HHV-6 DNA has been demonstrated in only 16% ofcases [Luppi et al., 1995]. The discrepancy with our
Fig. 2. Localization by immunohistochemistry of HHV-6 antigen p41 in brain tumor sections. Sections from frozen (A,B,D) and paraffin-embedded (C) tissue were stained with MAb to HHV-6 p41/38 and counterstained with hematoxylin. A,B: Schwannoma: very strong (+++)immunoreactivity in the nuclei of schwannoma cells (DAB ×400). C: glioblastoma: strong staining (++) in the cytoplasm and nuclei of neoplasticglial cells (DAB ×400). D: Schwannoma: HHV-6-p41 negative tumor sample.
TABLE II. HHV-6 Serological Reactivity in Brain Tumor Patients and Control
Antibodytiter
#1:10%
1:20%
1:40%
1:80%
1:160%
1:320%
1:640%
1:1280%
Totaln GMTa
Case 21.9 23.2 34.2 9.5 4.1 1.3 4.1 1.3 73 32.2Control 29.3 10.3 42.2 13.5 4.1 0.6 — — 150 28.8aGeometric mean titer. Sera negative at the lowest dilution have been allocated a titer of 1:10.
HHV-6 in Normal and Neoplastic Brain 49

results could be ascribed to the larger number ofsamples examined by us.
Little is known about the distribution of HHV-6 vari-ants in normal population and the association with hu-man diseases other than exanthem subitum. The cur-rently available serological assays, including anenzyme immunoassay developed recently [Soldan etal., 1997], do not allow the determination of differencesin antibody reactivities toward the two variants, andhence the rate of infection with each variant and theidentification of a causative role in the associated dis-eases.
It has been shown that variant B strain is more dif-fuse in the PBL of a healthy population [Di Luca et al.,1994]. Evidence of variant B infection has been re-ported also by Drobisky et al. [1993] as detected byPCR on virus-enriched supernatants obtained from mi-togen activated PBL of bone marrow transplanted pa-tients. Another study reported prevalence of variant Ain serum samples of transplant patients and immuno-compromised hosts [Secchiero et al., 1995]. The distri-bution of HHV-6 variants in the brain is still a matterof study. Challoner et al. [1995] reported 70% variant Bpositivity in the brain of both multiple sclerosis pa-tients and controls. In contrast, in a preliminary studyof the normal brain from 13 autopsy donors, variant Awas found to be prevalent [Luppi et al., 1994]. Simi-larly HHV-6 A is detected more frequently in the CSFafter primary infection [Hall et al., 1988].
Our results are in line with the latter studies show-ing that variant A was three times more frequent thanvariant B in normal and neoplastic brain. A confirma-tion of variant identity was obtained by PCR analysison PBLs from seven patients with tumors.
On the basis of these data, it is suggested that thetwo HHV-6 variants may differ quantitatively in theirtropism and be associated with different diseases. Theneurotropism of HHV-6 variant A has also been sug-gested in vitro by the greater efficiency of GS strain toinfect, to produce a cytopathic effect, and to replicate inprimary astrocytes compared to variant B [He et al.,1996]. Detection of HHV-6 p41 antigen, expressed se-lectively during viral replication, in neoplastic but notin normal brain, suggests that in these patients thevirus can be reactivated from the latency state as hasbeen observed in other immunocompromised hosts.Taken together, the data suggest that HHV-6 may nothave a direct role in brain tumor pathogenesis but mayact as a cofactor as it does in AIDS and in other humantumors. More in vitro transformation studies and thedevelopment of animal models will be required to de-termine whether HHV-6 is indeed involved in thepathogenesis of human tumors.
ACKNOWLEDGMENTS
PT is supported by Associazione Italiana Ricerca sulCancro. We thank Miss Sofia Iacovone and Miss Ro-sanna Bocchini for technical assistance.
REFERENCES
Ablashi DV, Lusso P, Hung CL, Salahuddin SZ, Josephs SF, Llana T,Kramarsky B, Biberfeld P, Markham PD, Gallo RC. 1988. Utili-zation of human hematopoietic cell lines for the propagation andcharacterization of HBLV (human herpesvirus 6). Int J Cancer15:787–791.
Ablashi D, Agut H, Berneman Z, Campadelli-Fiume G, Carrigan D,Ceccherini-Nelli L, Chandran B, Chou S, Collandre H, Cone R,Dambough T, Dewurst S, Di Luca D, Foa-Tomasi L, Fleckenstein,Frenkel N, Gallo R, Gompels U, Hall C, Jones M, Lawrence G,Martin M, Montagnier L, Neipel F, Nicholas J, Pellet P, RazzaqueA, Torelli G, Thompson B, Salahuddin S, Wyatt L, Yamanishi K.1993. Human herpesvirus-6 strain variants: a nomenclature. ArchVirol 129:363–366.
Adams RA, Flowers A, Davis BJ. 1968. Direct implantation and serialtransplantation of human acute lymphoblastic leukemia in ham-sters, SB-2. Cancer Res 28:1121–1125.
Barbanti-Brodano G, Martini F, De Mattei M, Lazzarin L, Corallini A,Tognon M. 1998. BK and JC human polyomaviruses and simianvirus 40:natural history of infection in humans, experimental on-cogenicity, and association with human tumors. Adv Virus Res50:69–99.
Black JB, Sanderlia KC, Goldsmith CS, Gary HE, Lopez C, Pellet PE.1989. Growth properties of human herpesvirus-6 strain Z29. JVirol Methods 26:133–145.
Bovenzi P, Mirandola P, Secchiero P, Strumia R, Cassai E, Di Luca D.1993. Human herpesvirus 6 (variant A) in Kaposi’s sarcoma. Lan-cet 341:1288–1289.
Boyum A. 1968. Isolation of mononuclear cells and monocytes fromhuman blood. Isolation of mononuclear cells by combining cen-trifugation and sedimentation. Scandinavian Journal of Clin LabInvest 21:77–89.
Caserta MT, Hall CB, Schnabel K, McIntyre K, Long C, Costanzo M;,Dewurst S, Insel R, Epstein LG. 1994. Neuroinvasion and persis-tence of human herpesvirus 6 in children. J Infect Dis 170:1586–1589.
Challoner PB, Smith KT, Parker JD, MacLeod DL, Coulter SN, RoseTM, Schultz ER, Bennet JL, Garber RL, Chang M, Schad PA,Stewart PM, Nowinski RC, Brown JP, Burmer JC. 1995. Plaque-associated expression of human herpesvirus 6 in multiple sclero-sis. Proc Natl Acad Sci USA 92:7440–7444.
Chen M, Popescu N, Woodworth C, Berneman Z, Corbellino M, LussoP, Ablashi DV, Di Paolo JA. 1994. Human herpesvirus 6 infectscervical epithelial cells and transactivates human papillomavirusgene expression. J Virol 68:1173–1178.
Cheow KC, Balachandran N. 1991. Identification, characterization,and sequence analysis of a cDNA encoding a phosphoprotein ofhuman herpesvirus 6. J Virol 65:2884–2894.
Corallini A, Pagnani M, Viadana P, Silini E, Mottes M, Milanesi G,Gerna G, Vettor R, Trapella G, Silvani V, Gaist G, Barbanti-Brodano G. 1987. Association of BK virus with human brain tu-mors and tumors of pancreatic islets. Int J Cancer 39:60–67.
Cuomo L, Angeloni A, Zompetta C, Cirone M, Calogero A, Frati L,Ragona G, Faggioni A. 1995. Human herpesvirus 6, variant A, butnot variant B, infects EBV-positive B-lymphoid cells, activatingthe latent EBV genome through a BZLF-1 dependent mechanism.AIDS Res Hum Retroviruses 11:1241–1245.
Cuomo L, Trivedi P, de Grazia U, Calogero A, D’Onofrio M, Yang W,Frati L, Faggioni A, Rymo L, Ragona G. 1998. Upregulation ofEpstein-Barr virus-encoded latent membrane protein by humanherpesvirus 6 superinfection of EBV-carrying Burkitt lymphomacells. J Med Virol 55:219–226.
Dewhurst S, Dollard SC, Pellet PE, Dambaugh TR. 1993. Identifica-tion of a lytic-phase origin of DNA replication in human herpes-virus 6B strain Z29. J Virol 67:7680–7683.
Di Luca D., Dolcetti R, Mirandola P, De Re V, Secchiero P, Carbone A,Boiocchi M, Cassai E. 1994. Human herpesvirus 6: a survey ofpresence and variant distribution in normal peripheral lympho-cytes and lymphoproliferative disorders. J Infect Dis 170:211–215.
Di Luca D., Mirandola P, Ravaioli T, Bigoni B, Cassai E. 1996. Dis-tribution of HHV-6 variants in human tissues. Infect Agents Dis5:203–214.
Dorries K, Loeber G, Meixensberger J. 1987. Association of polyoma-virus JC, SV40, and BK with human brain tumors. Virology 160:268–270.
Drobyski WR, Eberle M, Mayewski D, Baxter-Lowe LA. 1993. Preva-
50 Cuomo et al.

lence of human herpesvirus 6 variant A and B infections in bonemarrow transplant recipients as determined by polymerase chainreaction and sequence-specific oligonuceotide probe hybridization.J Clin Microbiol 31:151–20
Drobyski WR, Knox KK, Majewski D, Carrigan DR. 1994. Fatal en-cephalitis due to variant B human herpesvirus-6 infection in abone marrow-transplant recipient. N Engl J Med 330:1356–1360.
Fraser NW, Lawrence WC, Wroblewska Z, Gilden DH, Koprowski H.1981. Herpes simplex type 1 DNA in human brain tissue. ProcNatl Acad Sci USA 78:6461–6465.
Gentile G, Mele A, Ragona G, Faggioni A, Zompetta C, Tosti ME,Visani G, Castelli G, Pulsoni A, Monarca B, Martino P, Mandelli F.1999. Human herpes virus 6 seroprevalence and leukaemias: acase-control study. GINEMA. Br J Cancer 80:1103–1106.
Hall CB, Caserta MT, Schnabel KC, Long C, Epstein LG, Insel RA,Dewhurst S. 1998. Persistence of human herpesvirus 6 accordingto site and variant: possible greater neurotropism of variant A.Clin Infect Dis 26:132–137.
He J, McCarthy M, Zhou Y, Chandran B, Wood C. 1996. Infection ofprimary human fetal astrocytes by human herpesvirus 6. J Virol70:1296–1300.
Josephs SF, Salahuddin SZ, Ablashi DV, Schachter F, Wong Staal F,Gallo RC. 1986. Genomic analysis of the human B-lymphotropicvirus (HBLV). Science 234:601–603.
Josephs SF, Buchbinder A, Streicher HZ, Ablashi DV, Salahuddin SZ,Guo HG, Wong-Staal F, Cossman J, Faffeld M, Sundeen J, LevineP, Biggar R, Krueger GRF, Fox RI, Gallo RC. 1988. Detection ofhuman B-lymphotropic virus (human herpesvirus 6) sequences inB cell lymphoma tissues. Leukemia 2:132–135.
Kashanchi D, Araujo J, Doniger J, Muralidhar S, Hoch R, Khleif S,Mendelson E, Thompson J, Azumi N, Brady JN, Luppi M, TorelliG, Rosenthal LJ. 1997. Human herpesvirus 6 (HHV-6) ORF-1transactivating gene exhibits malignant transforming activity andits protein binds to p53. Oncogene 14:359–367.
Knox KK, Carrigan DR. 1994. Disseminated active HHV-6 infectionsin patients with AIDS. Lancet 343:577–578.
Levy JA, Ferro F, Lennette ET, Oshiro L, Poulin L. 1990. Character-ization of a new strain of HHV-6 (HHV-6 SF) recovered from thesaliva of HIV-infected individual. Virology 178:113–121.
Luppi M, Barozzi P, Maiorana A, Marasca R, Torelli G. 1994. Humanherpesvirus 6 infection in normal human brain tissue. Journal ofInfectious Diseases 169:943–944.
Luppi M, Barozzi P, Maiorana A, Marasca R, Trovato R, Fano R,Ceccherini-Nelli L, Torelli G. 1995. Human Herpesvirus-6: a sur-vey of presence and distribution of genomic sequences in normalbrain and neuroglial tumors. Journal of Medical Virology 47:105–111.
Lyall EGH and Cubie HA. 1995. Human herpesvirus-6 DNA in thesaliva of paediatric oncology patients and controls. Journal ofMedical Virology 47:317–322.
Martini F, Iaccheri L, Lazzarin L, Carinci P, Corallini A, Gerosa M,Iuzzolino P, Barbanti-Brodano G, Tognon M. 1996. SV40 earlyregion and large T antigen in human brain tumors, peripheralblood cells and sperm fluids from healthy individuals. Cancer Re-search 56:4820–4825.
Merelli E, Sola P, Barozzi P, Torelli G. 1996. An encephalitic episodein a multiple sclerosis patient with human herpesvirus 6 latentinfection. Journal of Neurological Science 137:42–46.
Merelli E, Bedin R, Sola P, Barozzi P, Mancardi GL, Ficarra G,Franchini G. 1997. Human herpesvirus 6 and human herpesvirus8 DNA sequences in brains of multiple sclerosis patients, normaladults and children. Journal of Neurology 244:450–454.
Monini P, De Lellis L, Barbanti-Brodano G. 1995. Association of BKand JC human polyomaviruses and SV40 with human tumors. In:Barbanti-Brodano G, Bendinelli M, Friedman H, editors. DNA Tu-mor Viruses. Oncogenic Mechanism. New York: Plenum Press. p51–73.
Mullenbach R, Lagoda PJ, Welter C. 1989. An efficient salt chloroformextraction of DNA from blood and tissues. Trends Genetics 5:391.
Pedneault L, Katz BZ, Miller G. 1992. Detection of Epstein-Barr virusin the brain by polymerase chain reaction. Annals of Neurology32:184–192.
Ragona G, Calogero A, Cirone M, Cuomo L, Gonnella R, Zompetta C,Gentile G, Martino P, Menichella D, Frati L, Faggioni A. 1994.HHV-6 infection in Italy: characterization of an endemic isolateand seroepidemiologic analysis. Clinical and Diagnostic Virology1:261–270.
Razzaque A. 1990. Oncogene potential of human herpesvirus-6 DNA.Oncogene 5:1365–1370.
Razzaque A, Williams O, Wang J, Rhim JS. 1993. Neoplastic trans-formation of immortalized human epidermal keratinocytes by twoHHV-6 DNA clones. Virology 195:113–120.
Salahuddin S, Ablashi DV, Markham PD, Josephs SF, SturzeneggerS, Kaplan M, Halligan G, Biberfeld P, Wong-Staal, Kramarsky B,Gallo RC. 1986. Isolation of a new virus HBLV in patients withlymphoproliferative disorders. Science 234:596–601.
Secchiero P, Carrigan DR, Asano Y, Benedetti L, Crowley W, Koma-roff AL, Gallo RC, Lusso P. 1995. Detection of human herpesvirus6 in plasma of children with primary infection and immunosup-pressed patients by polymerase chain reaction. Journal of Infec-tious Diseases 171:273–280.
Soldan SS, Berti R, Salem N, Secchiero P, Flamand L, Calabresi PA,Brennan MB, Maloni HW, McFarland HF, Lin HC, Patnaik M,Jacobsen S. 1997. Association of human herpesvirus 6 (HHV-6)with multiple sclerosis: Increased IgM response to HHV-6 earlyantigen and detection of serum HHV-6 DNA. Nature Medicine3:1394–1397.
Tedder RS, Briggs M, Cameron CH, Honess R, Robertson D, WhittleH. 1987. A novel lymphotropic herpesvirus. Lancet 2:390–392.
Torelli G, Marasca R, Luppi M, Selleri L, Ferrari S, Narni F, MarianoRT, Federico M, Ceccherini-Nelli L, Bendinelli M, Montagnani G,Montorsi M, Artusi T. 1991. Human herpesvirus-6 in human lym-phomas: identification of specific sequences in Hodgkin’s lympho-mas by polymerase chain reaction. Blood 77:2251–2258.
Yadav M, Chandrashekran A, Vasudevan DM, Ablashi DV. 1994. Fre-quent detection of human herpesvirus 6 in oral carcinoma. Journalof National Cancer Institute 7:1792–1794.
Yamanishi K, Okuno T, Shiraki K, Takahashi M, Kondo T, Asano Y,Kurata T. 1988. Identification of human herpesvirus 6 as a casualagent for exanthem subitum. Lancet 1:1065–1067.
HHV-6 in Normal and Neoplastic Brain 51





























