Embed Size (px)
Citation preview

Cytotoxic T lymphocyte responses to Marek’s diseaseherpesvirus-encoded glycoproteins
Carrie J. Markowski-Grimsrud1, Karel A. Schat*
Department of Microbiology and Immunology, Unit of Avian Health, College of Veterinary Medicine,
Cornell University, Ithaca, NY 14853, USA
Received 20 June 2002; received in revised form 26 August 2002; accepted 26 August 2002
Abstract
Cell-mediated immune responses are important for protective immunity to Marek’s disease (MD), especially because MD
herpesvirus (MDV) infection is strictly cell-associated in chickens with the exception of the feather follicle epithelium. A system
previously developed using reticuloendotheliosis (REV)-transformed cell lines stably expressing individual MDV genes allows
the determination of relevant MDV proteins for the induction of cytotoxic T lymphocyte (CTL) responses. To examine the
importance of glycoproteins for the induction of CTL, the MDV genes coding for glycoproteins (g) C, D, E, H, I, K, L, and M
were stably transfected into the REV-transformed chicken cell lines RECC-CU205 (major histocompatibility complex (MHC):
B21B21) and RECC-CU91 (MHC: B19B19). All transfected cell lines were lysed by REV-sensitized, syngeneic splenocytes
obtained from MD-resistant N2a (MHC: B21B21) and MD-susceptible P2a (MHC: B19B19) chickens, indicating that the
expression of individual MDV glycoproteins did not interfere with antigen processing pathways. Only cell lines expressing gI
were recognized by CTL from both N2a and P2a MDV-infected chickens. Cell lines expressing glycoproteins gC and gK, and to
a lesser extent, gH, gL, and gM were lysed by syngeneic MDV-sensitized splenocytes from N2a birds but not P2a birds. In
contrast, gE was recognized by MDV-sensitized effector cells from the P2a line and not the N2a line. Glycoprotein D was not
recognized by either line, with the exception of one marginally significant P2a assay. These results indicate that late viral
glycoproteins are relevant for the induction of cell-mediated immunity during MDV infection.
# 2002 Elsevier Science B.V. All rights reserved.
Keywords: Cell-mediated immunity; Chickens; Cytotoxic T lymphocytes; Glycoprotein; Marek’s disease herpesvirus; Reticuloendotheliosis
virus
1. Introduction
Marek’s disease herpesvirus (MDV) is a 180 kbp
oncogenic herpesvirus of chickens. MDV was origin-
ally classified as a gammaherpesvirus due to its lym-
photropism but more recently has been reclassified as
an alphaherpesvirus based on its genomic structural
similarity to other alphaherpesviruses such as vari-
cella-zoster virus and herpes simplex virus (HSV)
(Buckmaster et al., 1988; Lupiani et al., 2001).
MDV is grouped into three serotypes: serotype 1
consists of all pathogenic virus strains, serotype 2
comprises the naturally occurring, non-oncogenic
strains in chickens, and serotype 3 includes the
Veterinary Immunology and Immunopathology 90 (2002) 133–144
* Corresponding author. Tel.: þ1-607-253-4032;
fax: þ1-607-253-3384.
E-mail address: [email protected] (K.A. Schat).1 Present address: Department of Immunology, Institute for
Cancer Research, Norwegian Radium Hospital, University of Oslo,
N-0310 Oslo, Norway.
0165-2427/02/$ – see front matter # 2002 Elsevier Science B.V. All rights reserved.
PII: S 0 1 6 5 - 2 4 2 7 ( 0 2 ) 0 0 2 2 9 - 5

non-pathogenic herpesvirus of turkeys (HVTs)
(Bulow and Biggs, 1975).
MDVis the causative agent of Marek’s disease (MD),
a lymphoproliferative disease of chickens prevalent in
poultry-producing countries worldwide (Purchase,
1985). The disease is characterized by mononuclear
infiltration of the peripheral nerves, viscera, and various
other organs, resulting in the formation of lymphoid
tumors and variable mortality (Payne, 1985). MD inci-
dence has largely been controlled by vaccination with
MDV serotypes 2 and 3, often in bi- and multivalent
combinations since the 1970s (Witter, 2001). However,
over the past decade new MDV serotype 1 strains of
increased virulence have emerged with greater immu-
nosuppressive potential (Calnek et al., 1998). It is
predicted that the protective ability of current vaccines
is limited (Kreager, 1997). A thorough understanding of
MDV immunity is urgently needed to meet the future
challenges of vaccine development.
Cell-mediated immune responses are important in
protection from herpesvirus infections in general
(Arvin, 1992; Rickinson and Moss, 1997; Riddell
et al., 1991; Schmid and Mawle, 1991). Cell-mediated
immunity is especially important in protection from
MDV infection due to the strictly cell-associated
nature of the virus (Schat and Markowski-Grimsrud,
2001).
Our laboratory has maintained a longstanding inter-
est in elucidating the importance of cell-mediated
immune responses to MDV infection. A system pre-
viously developed in the laboratory has utilized REV-
transformed cell lines stably transfected with eukar-
yotic expression vectors containing various MDV
genes of interest (Pratt et al., 1992). These cell lines
have been used as target cells in chromium release
assays (CRAs) to ascertain CTL responses to indivi-
dual MDV-encoded proteins in genetically resistant
and susceptible chicken lines with defined MHC
haplotypes.
Previous studies utilizing this system have shown
that CTL responses are generated to a number of
immediate early, early, and late MDV proteins (Uni
et al., 1994; Omar and Schat, 1996; Schat and Xing,
2000). Target cell lines expressing glycoprotein B
(gB), a late structural protein, were significantly lysed
by MDV serotype 1- or serotype 2-sensitized syn-
geneic but not allogeneic splenocytes (Omar and
Schat, 1996). The effector cells were characterized
as classical CTL expressing CD8 and TCRab but not
CD4 (Omar and Schat, 1997). These studies have been
extended to include the following glycoproteins
encoded by MDV: gC (Binns and Ross, 1989), gD
(Brunovskis and Velicer, 1995), gE (Brunovskis and
Velicer, 1995), gH (Scott et al., 1993), gI (Brunovskis
and Velicer, 1995), gK (Ren et al., 1994), gL (Yoshida
et al., 1994), and gM (Osterrieder, 1999). Preliminary
data suggested that some of the glycoproteins are
important in inducing CTL responses (Markowski-
Grimsrud et al., 2001). The complete findings for all
MDV glycoproteins are presented in this report.
2. Materials and methods
2.1. Experimental animals
Chickens were obtained from specific-pathogen-
free (SPF) departmental flocks with defined MHC
haplotypes, the N2a (MHC: B21B21) and P2a
(MHC: B19B19) lines (Weinstock and Schat, 1987).
The departmental flocks are maintained in a filtered-
air, positive-pressure house and are free of all patho-
gens except chicken infectious anemia virus (CAV)
(Cardona et al., 2000a,b). The majority of the chicks
hatched from these breeder flocks have maternal
antibodies against CAV. Birds were hatched either
in the SPF facility and transferred to separate experi-
mental units at 1 day of age or were hatched in the
experimental units to reduce the risk of exposure to
CAV. All experiments were conducted with birds less
than 3 weeks of age before maternal antibodies have
waned, since CAV infection in birds lacking maternal
antibodies impairs the development of CTL (Mar-
kowski-Grimsrud et al., 2001). All experimental pro-
cedures were conducted in compliance with
institutional animal use protocols.
2.2. Cell cultures and cell lines
Chick kidney cell (CKC) and chicken embryo fibro-
blast (CEF) cultures were prepared from 2-week-old
SPF chicks or 10-day-old chicken embryos, respec-
tively, as previously described (Schat and Purchase,
1998). The REV-transformed chicken cell (RECC)
(Witter et al., 1979) lines CU91 (MHC: B19B19) and
CU205 (MHC: B21B21) (Schat et al., 1992) were
134 C.J. Markowski-Grimsrud, K.A. Schat / Veterinary Immunology and Immunopathology 90 (2002) 133–144

propagated in lymphocyte medium (LM) (Schat and
Purchase, 1998) containing 10% FBS (LM10) in 5%
CO2 at 41 8C. The RECC cell lines CU371 and CU368
expressing MDV gB, derived from CU91 and CU205,
respectively, have been described (Omar and Schat,
1996), and were used as positive control target cell
lines in CRA.
2.3. Viruses and inoculations
The oncogenic MDV serotype 1 strain JM-16 (Cal-
nek et al., 1984) was propagated in CKCs and was
used for all MDV inoculations. The non-defective,
low-virulence CS-strain of REV, originally obtained
from Witter (Avian Disease and Oncology Laboratory,
East Lansing, MI), was propagated in CEF as pre-
viously described (Weinstock et al., 1989). Nine- or
ten-day-old chicks were inoculated intraabdominally
with 1000–2000 focus-forming units (FFU) of JM-16,
passage 19 (JM-16/p19) or with 103.5–4.3 tissue culture
infective doses 50% (TCID50) REV-CS, or used as
uninfected controls.
2.4. Virus isolation
For isolation of JM-16 from MDV-infected chick-
ens, 250,000 splenocytes from infected birds were
inoculated onto 24–48 h CKC in 35 mm tissue culture
plates in triplicate. The presence of FFU was deter-
mined 5–7 days post-inoculation.
2.5. Plasmid construction
The plasmid constructs pcDNA3-gD, pcDNA3-gE,
and pcDNA3-gI for expression of MDV gD, gE, and gI,
respectively, were kindly provided by the late Velicer
(Michigan State University, East Lansing, MI). The
genes for MDV glycoproteins gC, gH, gK, gL, and gM
were amplified from MDV viral DNA (RB-1B (Schat
et al., 1982a) or GA-5 strain (Calnek, 1973)) by PCR
and subcloned into the pcDNA3.1/Myc-His eukaryotic
expression vector (Invitrogen, Carlsbad, CA). The pri-
mers used for the amplification of these genes are listed
in Table 1. Constructs were verified by bi-directional
sequencing at the Biotechnology Resource Center,
Cornell University. Prior to transfection each construct
was linearized using an appropriate restriction enzyme
(PvuI or ScaI) recognizing a single site in the ampicillin
resistance gene.
2.6. Target cell line generation
RECC-CU91 and CU205 were transfected with
individual plasmid constructs by electroporation as
previously described (Pratt et al., 1992). Briefly, CU91
and CU205 cells were centrifuged over Ficoll-Paque
(Amersham Pharmacia Biotech, Inc., Piscataway, NJ)
at 500 � g for 20 min 1 day prior to transfection to
remove dead cells and resuspended at 0:5 � 106 cells/
ml in LM10. On the day of electroporation the cell lines
were centrifuged again over Ficoll-Paque, resuspended
Table 1
Primers used for amplification of MDV glycoprotein genes
Gene DNA
source
Primer Sequencea RE siteb Nucleotide
location
Amplicon
length (bp)
Accession
numberc
gC GA DNA Upper 50-(GAGAAGCTT)GAAGAGACACCAAACGTAACC-30 HindIII 268–288 1554 M20001
Lower 50-(GGGCTCGAG)CATAACAATGAGATTATAATCG-30 XhoI 1800–1821
gH RB-1B DNA Upper 50-(ACACTCGAG)CTACATATCCAACATGGGTCT-30 XhoI 194–214 2452 S62555
Lower 50-(ACATCTAGA)AAGATCGTCGTACAGGCTCAAT-30 XbaI 2624–2645
gK GA DNA Upper 50-(CATCACGCGGCCGC)CACGGTAGATATTGACTCG-30 NotI 406–424 1083 U10040
Lower 50-(TTAAGTCTCGAG)AGGCAAATAGGCACGC-30 XhoI 1473–1488
gL GA DNA Upper 50-(GCGCTCGAG)TAATCGGAGCTGCTCGGTAGC-30 XhoI 212–232 606 U04994
Lower 50-(ACATCTAGA)GGCATTGGCTCGTCGGCTGCCAGA-30 XbaI 794–817
gM GA DNA Upper 50-CTACCGGCGGTCGTCGCTATG-30 N/A 24–44 1290 AF118111
Lower 50-ATCATCCCATTCGCTCTCAG-30 N/A 1294–1313
a Parentheses designate spacer or restriction enzyme (italic) sequences. Start codons designated in bold. Those not present are located
downstream of the upper primer.b Restriction enzyme site used for subcloning after PCR amplification.c GenBank accession number.
C.J. Markowski-Grimsrud, K.A. Schat / Veterinary Immunology and Immunopathology 90 (2002) 133–144 135

at 1 � 107 cells/ml in LM base without antibiotics, and
0.4 ml of cell suspension was aliquoted into 0.4 cm
cuvettes. CU91 and CU205 were mock-electroporated
or electroporated with 10 mg of each linearized con-
struct at 300 V and 500 mF using a Gene Pulser (Bio-
Rad Laboratories, Inc., Hercules, CA). After 5 min the
cells were added to 5 ml of LM10 and plated in five
wells of a 24-well-plate (Costar, Corning, Inc., Corning,
NY). At 48 h post-transfection selection with Geneticin
(GibcoBRL Life Technologies, Inc., Gaithersburg,
MD) was initiated. Different amounts of Geneticin
were administered and increased incrementally until
the mock-transfected controls were no longer viable.
Following selection of stable transformed cells 3–4
weeks later, cell lines were maintained in LM10 con-
taining 300 mg Geneticin/ml.
2.7. RT–PCR
Total RNA was extracted from cell lines using
RNAzolB (Teltest, Inc., Friendswood, TX) according
to the manufacturer’s instructions. RNA was reverse
transcribed in 20 ml reactions using the RT Core Kit
(Perkin Elmer, Inc., Boston, MA) in a PE2400 thermal
cycler using the following parameters: 42 8C for
15 min, 99 8C for 5 min, and 5 8C for 5 min. Ten
milliliters of the RT reactions were used for PCR
amplification using 100 pmol of each primer for the
respective glycoprotein (Table 2) in 50 ml reactions.
The PCR cycling parameters consisted of a hold at
94 8C for 5 min, followed by 35 cycles of 94 8C for 45 s/
56 8C for 45 s/72 8C for 45 s, and ending with a hold at
72 8C for 10 min. Control reactions lacking reverse
transcriptase were performed to ensure RNA-specific
amplification. RT–PCR products were electorphoresed
on a 1.5% agarose gel and visualized by ethidium
bromide staining using an Eagle Eye detection system
(Stratagene, La Jolla, CA).
2.8. Indirect immunofluorescence assay (IFA)
Smears of stably transfected RECC lines were
prepared on 12-well glass slides at 5 � 104 cells per
well, air-dried, and fixed in 100% acetone for 10 min.
Cells were stained for 30 min at 37 8C with rabbit
antisera specific for the various glycoproteins gC
(kindly provided by Osterrieder), gE, gH, gI, and
gL (kindly provided by Lee) followed by an FITC-
conjugated goat anti-rabbit secondary antibody (Cap-
pel/ICN Pharmaceuticals, Inc., Costa Mesa, CA).
Antisera specific for gD, gK, and gM were unavailable
for testing. Cells were also stained separately with a
REV-specific monoclonal antibody (provided by Lee),
followed by an FITC-conjugated rabbit anti-mouse
secondary antibody (Cappel/ICN). Wells were washed
twice with PBS, pH 7.2, for 10 min after each staining.
Dabco/glycerine (Beutner et al., 1987) was used to
preserve the slides. Slides were examined in a Zeiss
Axioskop2 plus microscope under both phase-contrast
and UV light at 400� and 630� magnification power.
Photographs were taken using an AxioCam digital
camera (Carl Zeiss, Inc., Thornwood, NY) and visua-
lized using Axiovision version 3.0.6 software.
2.9. Serology
Sera were obtained from all chickens at the termi-
nation of each experiment and stored at �20 8C until
Table 2
Primers used for RT–PCR confirmation of transcribed MDV glycoprotein genes
Gene Forward primer (length) Reverse primer (length) Amplicon
length (bp)
Position Accession
numbera
gC 50-TAACAGAAACGACGGGCAAGAACG-30 (24) 50-ATACTGGAATGCGGCTGAGGAGAT-30 (24) 1047 536–1582 M20001
gD 50-AACCTCCGGGCTACTCTCAATG-30 (22) 50-TCGGTCGTTTAGTTATGTATGGTG-30 (24) 841 7105–7945 L22174
gE 50-ATCCCGCGATATCCACCAGTTG-30 (22) 50-ATCGCTAGCCTCATTTTCGTCG-30 (22) 1026 9572–10597 L22174
gH 50-ACAAAGACGACGCTCAACCTACAG-30 (24) 50-TTCCGCAAATGCATTCCAGATA-30 (22) 754 634–1387 S62555
gI 50-CAATCTGCTCTTGTTGCGTTCC-30 (22) 50-GGCCTCATCAGATCTTGGTTCAC-30 (23) 834 8354–9187 L22174
gK 50-TGGACAGTAGCCTCTATGACG-30 (21) 50-CGCAAGCAAGTATAGAATGCT-30 (21) 900 554–1453 U10040
gL 50-ACGATATTCCCACGGGCATTTTTA-30 (24) 50-TGGCATTCCCCGACTATCATCTTC-30 (24) 335 429–763 U04994
gM 50-GTGCGGCATACATGGGGAGTG-30 (21) 50-CAGCAGAACGGGATTGTGTAAGTG-30 (24) 831 418–1248 AF118111
a GenBank accession number.
136 C.J. Markowski-Grimsrud, K.A. Schat / Veterinary Immunology and Immunopathology 90 (2002) 133–144

use in enzyme-linked immunoabsorbant assays
(ELISA) using the commercial Flokchek CAV Anti-
body Test kit (Idexx Laboratories, Westbrook, ME) to
ensure the presence of maternal antibodies to CAV.
Sera were diluted 1:10 and tested according to the
manufacturer’s instructions. Absorbances were read at
650 nm in a Microplate EL310 Autoreader (Bio-Tek
Instruments, Inc., Winooski, VT).
2.10. CRA
For all experiments, 6 to 12 9- or 10-day-old P2a or
N2a birds per group were used. All assays contained
an uninfected control group and an MDV-infected
group, and in most cases a third group of REV-infected
birds was included. Birds were euthanized and effec-
tor cells were prepared from spleens at 7 days post-
infection (dpi) using previously described methods
(Schat et al., 1982b; Omar and Schat, 1996). Briefly,
spleens were decapsulated and forced through a
0.60 mm pore-size nylon mesh (Tetko, Inc., Kansas
City, MO) to create single cell suspensions. Cells were
resuspended in PBS and centrifuged over Ficoll-Paque
at 500 � g for 20 min to remove red blood cells.
Lymphocytes were harvested from the interface,
washed once with PBS, and resuspended at 5�107 cells/ml in LM containing 20% FBS.
For target cell preparation, the stably transfected
RECC lines were centrifuged over Ficoll-Paque 1 day
prior to the assay and adjusted to 0.5–1:0 � 106 cells/
ml in LM10. On the day of the assay, each cell line was
labelled with 150–200 mCi of Na51CrO4 (New Eng-
land Nuclear, Boston, MA) as previously described
(Schat et al., 1982b). CRAs were performed in tripli-
cate at an effector:target cell ratio of 100:1 in 96-well
round-bottomed plates (Costar) using 5 � 104 target
cells per well. Three to six target cell lines were tested
per assay depending on the number of spleen cells
available; one complete trial, therefore, represented a
composite of several assays. Each cell line was tested
2–3 times; hence a total of 2–3 trials were performed
for N2a and P2a chickens.
Following a 4 h incubation at 38 8C, the plates were
centrifuged at 500 � g for 10 min. Half of the super-
natant (100 ml) from each well was harvested, mixed
with an equal volume of SuperMix scintillation fluid
(EG&G Wallac, Turku, Finland) in Microbeta 96-well
flexible plates and cpm were determined for each well
based on 5 min counts. The pellets were resuspended
in the remaining supernatant, and half (50 ml) from
each well was transferred to a new Microbeta plate
containing 50 ml of 2% Triton X and lysed overnight.
SuperMix (100 ml) was then added to the lysate in each
well and mixed by shaking the plates gently on a
Vortex Genie 2 (Fisher Scientific, Pittsburgh, PA) for
5–10 min, and the cpm were determined in a Trilux
1450 Microbeta (EG&G Wallac) scintillation counter.
The percent specific release (% SR) was calculated
using the following formula:
% SR ¼
cpm supernatant of sample
� cpm supernatant average control
cpm pellet þ supernatant
� cpm supernatant average control
� 100
2.11. Statistical analysis
For CRA analysis, the percent releases (% release ¼cpm supernatant=total cpm) between control and
infected groups were compared using a Student’s t
test (Snedecor and Cochran, 1989). Non-responder
chickens that tested negative for CAV antibodies were
removed from analysis to eliminate any possibility of
interference of CAV with CTL development (Mar-
kowski-Grimsrud et al., 2001).
3. Results
3.1. Development of REV-transformed cell lines
expressing MDV glycoprotein genes
REV-transformed cell lines stably transfected with
MDV glycoprotein constructs were tested for tran-
scription and expression of individual MDV genes by
RT–PCR and IFA analysis, respectively. All cell lines
tested positive for the appropriate transcripts of the
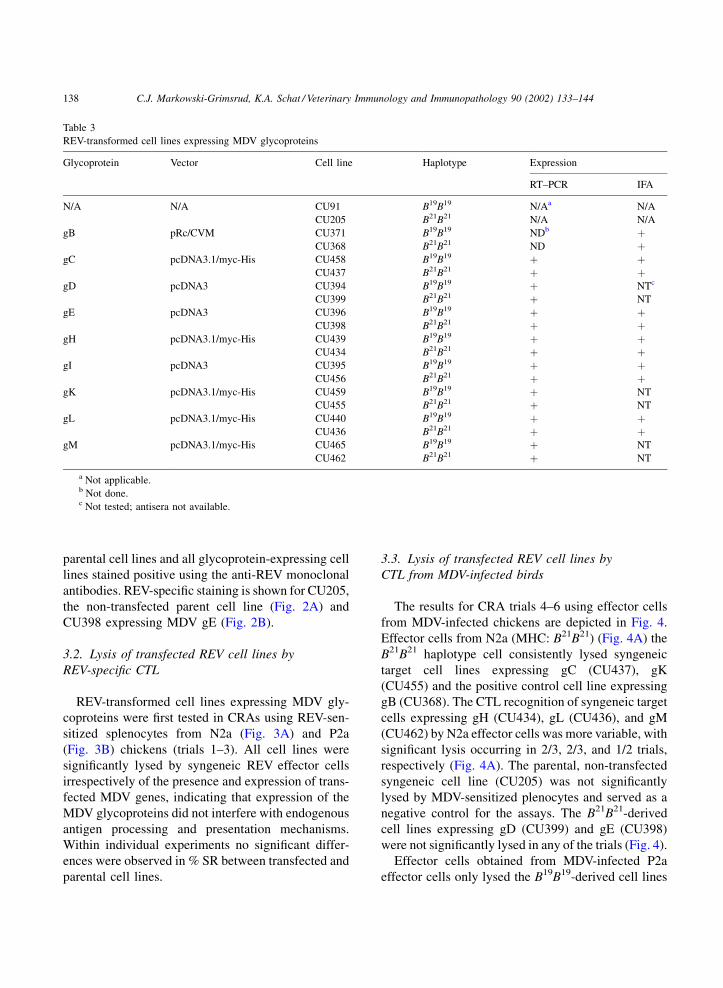
transfected MDV genes (Table 3 and Fig. 1).
The IFA results are summarized in Table 3. The
specific fluorescence for selected cell lines expressing
gC, gE, gH, gI, and gL is shown in Fig. 2. While there
was a faint non-specific staining pattern present in the
parent cell lines for all glycoprotein antisera (Fig. 2A),
the glycoprotein-expressing cell lines exhibited a
brighter, granular staining throughout the cytoplasm
of virtually all cells (Fig. 2B). As expected, both the
C.J. Markowski-Grimsrud, K.A. Schat / Veterinary Immunology and Immunopathology 90 (2002) 133–144 137
parental cell lines and all glycoprotein-expressing cell
lines stained positive using the anti-REV monoclonal
antibodies. REV-specific staining is shown for CU205,
the non-transfected parent cell line (Fig. 2A) and
CU398 expressing MDV gE (Fig. 2B).
3.2. Lysis of transfected REV cell lines by
REV-specific CTL
REV-transformed cell lines expressing MDV gly-
coproteins were first tested in CRAs using REV-sen-
sitized splenocytes from N2a (Fig. 3A) and P2a
(Fig. 3B) chickens (trials 1–3). All cell lines were
significantly lysed by syngeneic REV effector cells
irrespectively of the presence and expression of trans-
fected MDV genes, indicating that expression of the
MDV glycoproteins did not interfere with endogenous
antigen processing and presentation mechanisms.
Within individual experiments no significant differ-
ences were observed in % SR between transfected and
parental cell lines.
3.3. Lysis of transfected REV cell lines by
CTL from MDV-infected birds
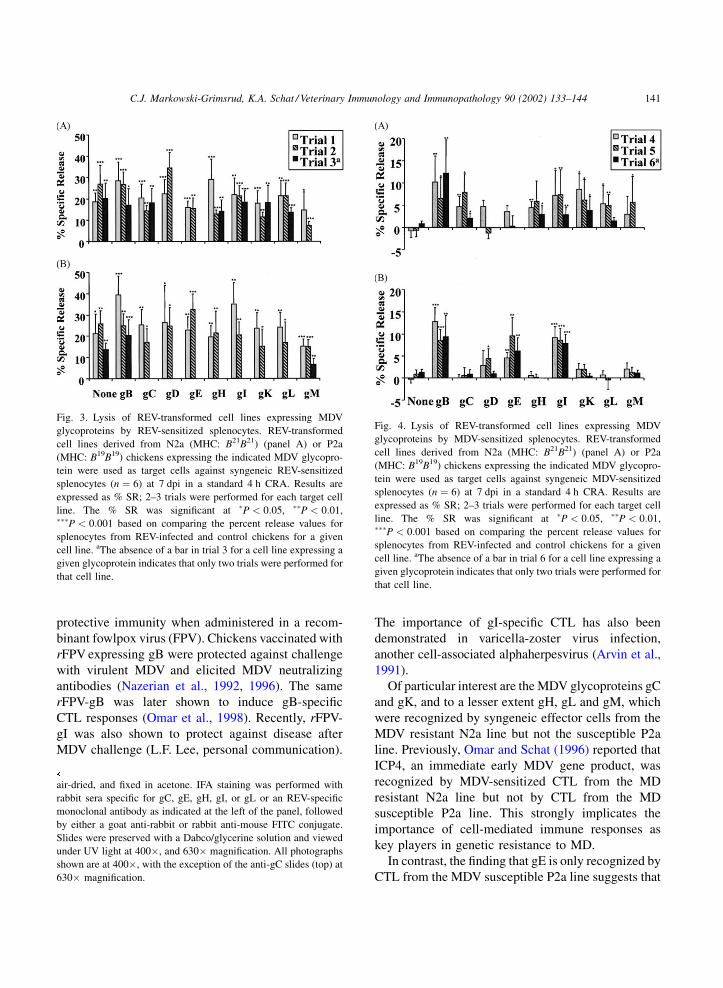
The results for CRA trials 4–6 using effector cells
from MDV-infected chickens are depicted in Fig. 4.
Effector cells from N2a (MHC: B21B21) (Fig. 4A) the
B21B21 haplotype cell consistently lysed syngeneic
target cell lines expressing gC (CU437), gK
(CU455) and the positive control cell line expressing
gB (CU368). The CTL recognition of syngeneic target
cells expressing gH (CU434), gL (CU436), and gM
(CU462) by N2a effector cells was more variable, with
significant lysis occurring in 2/3, 2/3, and 1/2 trials,
respectively (Fig. 4A). The parental, non-transfected
syngeneic cell line (CU205) was not significantly
lysed by MDV-sensitized plenocytes and served as a
negative control for the assays. The B21B21-derived
cell lines expressing gD (CU399) and gE (CU398)
were not significantly lysed in any of the trials (Fig. 4).
Effector cells obtained from MDV-infected P2a
effector cells only lysed the B19B19-derived cell lines
Table 3
REV-transformed cell lines expressing MDV glycoproteins
Glycoprotein Vector Cell line Haplotype Expression
RT–PCR IFA
N/A N/A CU91 B19B19 N/Aa N/A
CU205 B21B21 N/A N/A
gB pRc/CVM CU371 B19B19 NDb þCU368 B21B21 ND þ
gC pcDNA3.1/myc-His CU458 B19B19 þ þCU437 B21B21 þ þ
gD pcDNA3 CU394 B19B19 þ NTc
CU399 B21B21 þ NT
gE pcDNA3 CU396 B19B19 þ þCU398 B21B21 þ þ
gH pcDNA3.1/myc-His CU439 B19B19 þ þCU434 B21B21 þ þ
gI pcDNA3 CU395 B19B19 þ þCU456 B21B21 þ þ
gK pcDNA3.1/myc-His CU459 B19B19 þ NT
CU455 B21B21 þ NT
gL pcDNA3.1/myc-His CU440 B19B19 þ þCU436 B21B21 þ þ
gM pcDNA3.1/myc-His CU465 B19B19 þ NT
CU462 B21B21 þ NT
a Not applicable.b Not done.c Not tested; antisera not available.
138 C.J. Markowski-Grimsrud, K.A. Schat / Veterinary Immunology and Immunopathology 90 (2002) 133–144

expressing gI (CU456), gE (CU396), and the positive
control cell line expressing gB (CU371) (Fig. 4B). The
B19B19 cell lines expressing gC (CU458), gH
(CU439), gK (CU459), gL (CU440), and gM
(CU465) were not significantly lysed in any of the
trials using P2a-derived effector cells (Fig. 4B).
Finally, the B19B19 cell line expressing gD (CU394)
was not lysed by MDV-sensitized P2a effector cells,
with one exception in trial 2 with a marginally sig-
nificant % SR (Fig. 4B).
3.4. Virus isolation
MDV was reisolated from the MDV-infected chick-
ens in all experiments ensuring that negative CRA
results were not caused by the absence of infection
(data not shown).
3.5. CAV serology of infected birds
Serology analysis performed on all MDV- and REV-
infected birds failed to show a correlation between
CTL response and CAV maternal antibody status,
indicating a lack of CAV-induced CTL impairment.
4. Discussion
The generation of CTL responses to late structural
proteins might serve as an advantage to most herpes-
viruses that exist in cell-free form throughout the host,
as cell lysis at a late stage of virion packaging would
aid the release of virus and perpetuate the infectious
cycle. However, since MDV is strictly cell-associated
with the exception of the feather follicle epithelium
(Calnek et al., 1970), release of infectious virions from
CTL-lysed cells is probably minimal. The function-
ality of MDV-specific antibodies is minimal in this
context of intracellular virions. Therefore, CTL
responses are of paramount importance to the adaptive
immune response during MDV infection.
The findings presented in this study indicate the
importance of several late glycoproteins in inducing
CTL responses in MDV-infected birds. Two glyco-
proteins have now been identified that are recognized
by CTL from N2a and P2a chicken strains, gB (Omar
and Schat, 1996) and gI in this report. Interestingly,
these two glycoproteins have been shown to induce
Fig. 1. RT–PCR analysis of MDV glycoprotein gene transcription in
stably transfected REV-transformed cell lines derived from the CU91
(MHC: B19B19) and CU205 (MHC: B21B21) parental lines. Total RNA
was extracted from approximately 107 cells from each cell line, and
aliquots of each were treated with DNAse to remove contaminating
DNA. RT–PCR was performed using each corresponding primer set
as described in the materials and methods. The black arrowheads
indicate the bands of expected length for each individual glycoprotein
gene (see Table 2 for details). The three negative control lanes
corresponding to CU91, CU205, and water were performed with gC
primer pairs in this figure and are representative of results obtained
with any of the glycoprotein primer sets. This figure is a composite of
several gels aligned using the DNA standards.
C.J. Markowski-Grimsrud, K.A. Schat / Veterinary Immunology and Immunopathology 90 (2002) 133–144 139
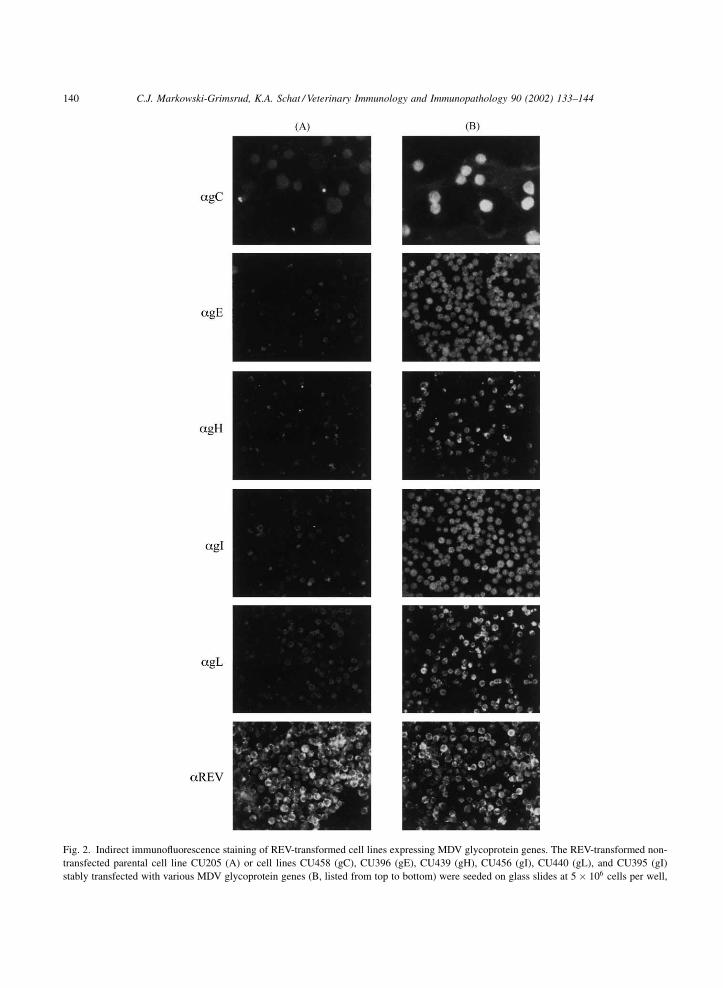
Fig. 2. Indirect immunofluorescence staining of REV-transformed cell lines expressing MDV glycoprotein genes. The REV-transformed non-
transfected parental cell line CU205 (A) or cell lines CU458 (gC), CU396 (gE), CU439 (gH), CU456 (gI), CU440 (gL), and CU395 (gI)
stably transfected with various MDV glycoprotein genes (B, listed from top to bottom) were seeded on glass slides at 5 � 106 cells per well,
140 C.J. Markowski-Grimsrud, K.A. Schat / Veterinary Immunology and Immunopathology 90 (2002) 133–144
protective immunity when administered in a recom-
binant fowlpox virus (FPV). Chickens vaccinated with
rFPV expressing gB were protected against challenge
with virulent MDV and elicited MDV neutralizing
antibodies (Nazerian et al., 1992, 1996). The same
rFPV-gB was later shown to induce gB-specific
CTL responses (Omar et al., 1998). Recently, rFPV-
gI was also shown to protect against disease after
MDV challenge (L.F. Lee, personal communication).
The importance of gI-specific CTL has also been
demonstrated in varicella-zoster virus infection,
another cell-associated alphaherpesvirus (Arvin et al.,
1991).
Of particular interest are the MDV glycoproteins gC
and gK, and to a lesser extent gH, gL and gM, which
were recognized by syngeneic effector cells from the
MDV resistant N2a line but not the susceptible P2a
line. Previously, Omar and Schat (1996) reported that
ICP4, an immediate early MDV gene product, was
recognized by MDV-sensitized CTL from the MD
resistant N2a line but not by CTL from the MD
susceptible P2a line. This strongly implicates the
importance of cell-mediated immune responses as
key players in genetic resistance to MD.
In contrast, the finding that gE is only recognized by
CTL from the MDV susceptible P2a line suggests that
air-dried, and fixed in acetone. IFA staining was performed with
rabbit sera specific for gC, gE, gH, gI, or gL or an REV-specific
monoclonal antibody as indicated at the left of the panel, followed
by either a goat anti-rabbit or rabbit anti-mouse FITC conjugate.
Slides were preserved with a Dabco/glycerine solution and viewed
under UV light at 400�, and 630� magnification. All photographs
shown are at 400�, with the exception of the anti-gC slides (top) at
630� magnification.
Fig. 3. Lysis of REV-transformed cell lines expressing MDV
glycoproteins by REV-sensitized splenocytes. REV-transformed
cell lines derived from N2a (MHC: B21B21) (panel A) or P2a
(MHC: B19B19) chickens expressing the indicated MDV glycopro-
tein were used as target cells against syngeneic REV-sensitized
splenocytes (n ¼ 6) at 7 dpi in a standard 4 h CRA. Results are
expressed as % SR; 2–3 trials were performed for each target cell
line. The % SR was significant at �P < 0:05, ��P < 0:01,���P < 0:001 based on comparing the percent release values for
splenocytes from REV-infected and control chickens for a given
cell line. aThe absence of a bar in trial 3 for a cell line expressing a
given glycoprotein indicates that only two trials were performed for
that cell line.
Fig. 4. Lysis of REV-transformed cell lines expressing MDV
glycoproteins by MDV-sensitized splenocytes. REV-transformed
cell lines derived from N2a (MHC: B21B21) (panel A) or P2a
(MHC: B19B19) chickens expressing the indicated MDV glycopro-
tein were used as target cells against syngeneic MDV-sensitized
splenocytes (n ¼ 6) at 7 dpi in a standard 4 h CRA. Results are
expressed as % SR; 2–3 trials were performed for each target cell
line. The % SR was significant at �P < 0:05, ��P < 0:01,���P < 0:001 based on comparing the percent release values for
splenocytes from REV-infected and control chickens for a given
cell line. aThe absence of a bar in trial 6 for a cell line expressing a
given glycoprotein indicates that only two trials were performed for
that cell line.
C.J. Markowski-Grimsrud, K.A. Schat / Veterinary Immunology and Immunopathology 90 (2002) 133–144 141

CTL responses to gE have an insignificant role in
protective immunity. The fact that gD did not appear
to play a significant role in CTL induction was not
surprising, since gD is poorly expressed during MDV
infection and was recently reported to be undetectable
in cell culture (Tan et al., 2001). The minor relevance of
gD for cell-mediated immunity and the fact that it is a
non-essential gene for in vivo infectivity (Parcells et al.,
1994) suggests that this gene is a candidate locus for the
development of recombinant MDV vaccines expressing
genes for other poultry pathogens such as Newcastle
disease virus, infectious bursal disease virus, and others
(Hirai and Sakaguchi, 2001).
The MDV-specific % SR are generally low, albeit
significant, typically ranging from 5 to 15% (Uni et al.,
1994; Omar and Schat, 1996) compared with HSV
studies, where secondary stimulation of splenocytes is
often used to demonstrate CTL activity (Borysiewicz
and Sissons, 1994). The same cell lines were lysed to a
greater extent by REV-sensitized splenocytes (10–
35%), which is also consistent with previous findings
(Weinstock et al., 1989; Pratt et al., 1992; Omar and
Schat, 1996). The differences in % SR by REV- versus
MDV-sensitized splenocytes can be caused by the fact
that based on IFA studies all cells within a cell line
strongly express REV proteins, but not all cells
express the transfected MDV gene to the same level.
It is also plausible that more than one REV protein is
processed and presented to REV-specific CTL or that
an immunodominant epitope is present, which might
account for the higher degree of REV-specific lysis.
One notable difference between this study and
previous studies (Pratt et al., 1992; Uni et al., 1994;
Omar and Schat, 1996; Schat and Xing, 2000) is that in
this study 9- to 10-day-old birds were used in all
assays as opposed to 4–5-week-old birds in the pre-
vious studies. This change was needed because: (a) the
SPF flocks had become infected with CIAV (Cardona
et al., 2000a,b), (b) Markowski-Grimsrud et al. (2001)
have shown that MDV- and REV-specific CTL devel-
opment are impaired during concurrent CAV infection
in birds lacking virus-neutralizing maternal antibo-
dies, and (c) the possibility of CAV infection in birds
lacking maternal antibodies could not be excluded.
However, the finding that the serology data in this
study did not correlate with relative CTL-mediated
lysis strongly suggests that CAV infection was not
present in the birds hatched in the experimental units.
While the use of younger birds possessing CAV
maternal antibodies was essential for the continuation
of this study as a precautionary measure, a lower level
of immunocompetence of the younger birds may
account for the relatively low and sometimes variable
results in the MDV- and REV-specific CTL assays.
The REV CTL assays serve as an important control
in two respects. It is well known that many proteins
encoded by herpesviruses are involved in the subver-
sion of the immune system (Ploegh, 1998). In this
respect the CRAs using transfected target cells and
REV-sensitized effector cells ensured that a given
MDV protein did not interfere with the antigen pro-
cessing and presentation of the MHC class I context.
Secondly, they served as a positive control for the
assay itself, especially in light of the potential CAV
problems discussed above.
Whether the MDV-specific CTL confer protection
from disease remains to be elucidated by passive
transfer assays. Previous studies using passively trans-
ferred primary splenocytes from chickens infected
with an rFPV expressing the MDV gB gene failed
to demonstrate a reduction in viral replication upon
MDV challenge (Omar, 1997). The negative results
were likely due to the limited gB-specific CTL sub-
population(s) present in the primary splenocyte pool.
These experiments have been hindered by a lack of
means to amplify antigen-specific CTL in vitro in the
absence of MDV virus that can be present in the
lymphocytes after in vivo infection.
Although MD has been largely been curtailed by
vaccination and in fact represents the first successful
vaccine against a tumor-inducing virus in any species,
the mechanisms of vaccinal immunity have remained
elusive. Although JM-16/19, a low-passage serotype-1
MDV strain was used for all inoculations in the
experiments presented here, previous studies in the
laboratory indicated that both JM-16 and the serotype-
2 vaccine strain SB-1 induce MDV-specific CTL to
comparable levels (Omar and Schat, 1996). It is of
increasing importance to identify the antigens relevant
to vaccinal immunity. MD outbreaks in vaccinated
flocks have historically occurred at 10-year intervals
as MDV strains of increased virulence emerge (Witter,
1997). Since there are currently no new vaccine strains
available for commercial use, the development of
recombinant vaccines is of major importance to con-
trol disease in the near future.
142 C.J. Markowski-Grimsrud, K.A. Schat / Veterinary Immunology and Immunopathology 90 (2002) 133–144

Acknowledgements
This work was supported in part by the Cooperative
State Research, Education, and Extension Service, US
Department of Agriculture, under agreements #96-
38420-3061 and #98-35204-6425 and grants #298
and #426 from the US Poultry and Egg Association.
C.J.M. was supported by a USDA Biotechnology
Training Fellowship and a dissertation fellowship
from the American Association of University Women.
The authors are grateful to the late Dr. Leland F.
Velicer for kindly providing several plasmid con-
structs containing MDV glycoproteins, and Drs. Lucy
Lee and Nicolas Osterrieder for generously providing
the glycoprotein-specific antibodies. We also thank
Dr. Keith W. Jarosinski and Priscilla O’Connell for
technical assistance with several of the CRAs. Special
thanks are due to Dr. Ole M. Grimsrud for help with
preparing the figures and tables.
References
Arvin, A.M., 1992. Cell-mediated immunity to varicella-zoster
virus. J. Infect. Dis. 166, S35–41.
Arvin, A.M., Sharp, M., Smith, S., Koropchak, C.M., Diaz, P.S.,
Kinchington, P., Ruyechan, W., Hay, J., 1991. Equivalent
recognition of a varicella-zoster virus immediate early
protein (IE62) and glycoprotein I by cytotoxic T lymphocytes
of either CD4þ or CD8þ phenotype. J. Immunol. 146, 257–
264.
Beutner, E.H., Chorzelski, T.P., Kumar, V., 1987. Immunopathol-
ogy of the Skin, 3rd ed. Wiley, New York.
Binns, M.M., Ross, N.L.J., 1989. Nucleotide sequence of the
Marek’s disease virus (MDV) RB-1B A antigen and the
identification of the MDV A antigen as the herpes simplex
virus-2 glycoprotein C homologue. Virus Res. 12, 371–382.
Borysiewicz, L.K., Sissons, J.G.P., 1994. Cytotoxic T cells and
human herpes simplex virus. Curr. Top. Microbiol. Immunol.
189, 123–150.
Brunovskis, P., Velicer, L.F., 1995. The Marek’s disease virus
(MDV) unique short region: alphaherpesvirus-homologous,
fowlpox virus-homologous, and MDV-specific genes. Virology
206, 324–338.
Buckmaster, A.E., Scott, S.D., Sanderson, M.J., Boursnell, M.E.,
Ross, N.L., Binns, M.M., 1988. Gene sequence and mapping
data from Marek’s disease virus and herpesvirus of turkeys:
implications for herpesvirus classification. J. Gen. Virol. 69,
2033–2042.
Bulow, V.V., Biggs, P.M., 1975. Differentiation between strains of
Marek’s disease virus and turkey herpesvirus by immunofluor-
escence assays. Avian Pathol. 4, 133–146.
Calnek, B.W., 1973. Influence of age at exposure on the
pathogenesis of Marek’s disease. J. Natl. Cancer Inst. 51,
929–939.
Calnek, B.W., Adldinger, H.K., Kahn, D.E., 1970. Feather follicle
epithelium: a source of enveloped and infectious cell-free
herpesvirus from Marek’s disease. Avian Dis. 14, 219–233.
Calnek, B.W., Schat, K.A., Ross, L.J., Shek, W.R., Chen, C.L.,
1984. Further characterization of Marek’s disease virus-
infected lymphocytes. I. In vivo infection. Int. J. Cancer 33,
389–398.
Calnek, B.W., Harris, R.W., Buscaglia, C., Schat, K.A., Lucio, B.,
1998. Relationship between the immunosuppressive potential
and the pathotype of Marek’s disease virus isolates. Avian Dis.
42, 124–132.
Cardona, C., Lucio, B., O’Connell, P., Jagne, J., Schat, K.A.,
2000a. Humoral immune responses to chicken infectious
anemia virus in three strains of chickens in a closed flock.
Avian Dis. 44, 661–667.
Cardona, C., Oswald, W.B., Schat, K.A., 2000b. Distribution of
chicken anaemia virus in the reproductive tissues of specific-
pathogen-free chickens. J. Gen. Virol. 81, 2067–2075.
Hirai, K., Sakaguchi, M., 2001. Polyvalent recombinant Marek’s
disease virus vaccine against poultry diseases. Curr.
Top. Microbiol. Immunol. 225, 261–287.
Kreager, K., 1997. Marek’s disease: clinical aspects and current
field problems in layer chickens. In: Fadly, A.M., Schat, K.A.,
Spencer, J.L. (Eds.), Diagnosis and Control of Neoplastic
Diseases of Poultry. American Association of Avian Patholo-
gists, Kennett Square, PA, pp. 23–26.
Lupiani, B., Lee, L.F., Reddy, S.M., 2001. Protein-coding content
of the sequence of Marek’s disease virus. Curr. Top. Microbiol.
Immunol. 255, 159–190.
Markowski-Grimsrud, C.J., O’Connell, P., Schat, K.A., 2001.
Syngeneic cell-mediated immune responses to Marek’s disease
virus-encoded glycoproteins. In: Schat, K.A., Morgan, R.M.,
Parcells, M.S., Spencer, J.L. (Eds.), Current Progress on
Marek’s Disease Research. American Association of Avian
Pathologists, Kennett Square, PA, pp. 127–131.
Nazerian, K., Lee, L.F., Yanagida, N., Ogawa, R., 1992. Protection
against Marek’s disease by a fowlpox virus recombinant
expressing the glycoprotein B of Marek’s disease virus. J.
Virol. 66, 1409–1413.
Nazerian, K., Witter, R.L., Lee, L.F., Yanagida, N., 1996.
Protection and synergism by recombinant fowl pox vaccines
expressing genes from Marek’s disease virus. Avian Dis. 40,
368–376.
Omar, A.R., 1997. Cytotoxic T lymphocyte responses against
Marek’s disease herpesvirus. Ph.D. Thesis. Cornell University,
Ithaca, NY.
Omar, A.R., Schat, K.A., 1996. Syngeneic Marek’s disease virus
(MDV)-specific cell-mediated immune responses against im-
mediate early, late, and unique MDV proteins. Virology 222,
87–99.
Omar, A.R., Schat, K.A., 1997. Characterization of Marek’s
disease herpesvirus (MDV)-specific cytotoxic T lymphocytes
in chickens inoculated with a nononcogenic vaccine strain of
MDV. Immunology 90, 579–585.
C.J. Markowski-Grimsrud, K.A. Schat / Veterinary Immunology and Immunopathology 90 (2002) 133–144 143

Omar, A.R., Schat, K.A., Lee, L.F., Hunt, H.D., 1998. Cytotoxic T
lymphocyte response in chickens immunized with a recombi-
nant fowlpox virus expressing Marek’s disease herpesvirus
glycoprotein B. Vet. Immunol. Immunopathol. 62, 73–82.
Osterrieder, N., 1999. Sequence and initial characterization of the
U(L)10 (glycoprotein M) and U(L)11 homologous genes of
serotype 1 Marek’s disease virus arch. Virology 144, 1853–1863.
Parcells, M.S., Anderson, A.S., Morgan, R.W., 1994. Characteriza-
tion of a Marek’s disease virus mutant containing a lacZ
insertion in the US6 (gD) homologue gene. Virus Gen. 9, 5–13.
Payne, L.N., 1985. Pathology. In: Payne, L.N. (Ed.), Marek’s
Disease. Martinus Nijhoff, Boston, pp. 43–75.
Ploegh, H.L., 1998. Viral strategies of immune evasion. Science
280, 248–253.
Pratt, W.D., Morgan, R., Schat, K.A., 1992. Cell-mediated cytolysis
of lymphoblastoid cells expressing Marek’s disease virus-specific
phosphorylated polypeptides. Vet. Microbiol. 33, 93–99.
Purchase, H.G., 1985. Clinical disease and its economic impact. In:
Payne, L.N. (Ed), Marek’s Disease. Martinus Nijhoff, Boston,
pp. 17–24.
Ren, D., Lee, L.F., Coussens, P.M., 1994. Identification and
characterization of Marek’s disease virus genes homologous to
ICP27 and glycoprotein K of herpes simplex virus-1. Virology
204, 242–250.
Rickinson, A.B., Moss, D.J., 1997. Human cytotoxic T lymphocyte
responses to Epstein–Barr virus infection. Ann. Rev. Immunol.
15, 405–431.
Riddell, S.R., Reusser, P., Greenberg, P.D., 1991. Cytotoxic T cells
specific for cytomegalovirus: a potential therapy for immuno-
compromised patients. Rev. Infect. Dis. 13, S966–973.
Schat, K.A., Markowski-Grimsrud, C.J., 2001. Immune responses
to Marek’s disease virus. Curr. Top. Microbiol. Immunol. 225,
91–120.
Schat, K.A., Purchase, H.G., 1998. Cell-culture methods. In:
Swayne, D.E., Glisson, J.R., Jackwood, M.W., Pearson, J.E.,
Reed, W.M., (Eds.), A Laboratory Manual for the Isolation and
Identification of Avian Pathogens, 4th ed. American Associa-
tion of Avian Pathologists, Kennett Square, PA, pp. 223–234.
Schat, K.A., Xing, Z., 2000. Specific and nonspecific immune
responses to Marek’s disease virus. Dev. Comp. Immunol. 24,
201–221.
Schat, K.A., Calnek, B.W., Fabricant, J., 1982a. Characterisation of
two highly oncogenic strains of Marek’s disease virus. Avian
Pathol. 11, 593–605.
Schat, K.A., Shek, W.R., Calnek, B.W., Abplanalp, H., 1982b.
Syngeneic and allogeneic cell-mediated cytotoxicity against
Marek’s disease lymphoblastoid tumor cell lines. Int. J. Cancer
29, 187–194.
Schat, K.A., Pratt, W.D., Morgan, R., Weinstock, D., Calnek,
B.W., 1992. Stable transfection of reticuloendotheliosis virus-
transformed lymphoblastoid cell lines. Avian Dis. 36, 432–
439.
Schmid, D.S., Mawle, A.C., 1991. T cell responses to herpes
simplex viruses in humans. Rev. Infect. Dis. 13, S946–949.
Scott, S.D., Smith, G.D., Ross, N.L.J., Binns, M.M., 1993.
Identification and sequence analysis of the homologues of the
herpes simplex virus type 1 glycoprotein H in Marek’s disease
virus and the herpesvirus of turkeys. J. Gen. Virol. 74, 1185–
1190.
Snedecor, G.W., Cochran, W.G., 1989. Statistical Methods, 8th ed.
Iowa State University Press, Ames, IA.
Tan, X., Brunovskis, P., Velicer, L.F., 2001. Transcriptional analysis
of Marek’s disease virus glycoprotein D, I, and E genes: gD
expression is undetectable in cell culture. J. Virol. 75, 2067–
2075.
Uni, Z., Pratt, W.D., Miller, M.M., O’Connell, P.H., Schat, K.A.,
1994. Syngeneic lysis of reticuloendotheliosis virus-trans-
formed cell lines transfected with Marek’s disease virus genes
by virus-specific cytotoxic T cells. Vet. Immunol. Immuno-
pathol. 44, 57–69.
Weinstock, D., Schat, K.A., 1987. Virus specific syngeneic killing
of reticuloendotheliosis virus transformed cell line target cells
by spleen cells. In: Weber, W.T., Ewert, D.L.(Eds.), Avian
Immunology. Alan R. Liss, New York, pp. 253–263.
Weinstock, D., Schat, K.A., Calnek, B.W., 1989. Cytotoxic T
lymphocytes in reticuloendotheliosis virus-infected chickens.
Eur. J. Immunol. 19, 267–272.
Witter, R.L., 1997. Increased virulence of Marek’s disease virus
field isolates. Avian Dis. 41, 149–163.
Witter, R.L., 2001. Protective efficacy of Marek’s disease vaccines.
Curr. Top. Microbiol. Immunol. 255, 57–90.
Witter, R.L., Calnek, B.W., Kato, S., Powell, P.C., 1979. A
proposed method for designating avian cell lines and
transplantable tumours. Avian Path. 8, 487–498.
Yoshida, S., Lee, L.F., Yanagida, N., Nazerian, K., 1994.
Identification and characterization of a Marek’s disease virus
gene homologous to glycoprotein L of herpes simplex virus.
Virology 204, 414–419.
144 C.J. Markowski-Grimsrud, K.A. Schat / Veterinary Immunology and Immunopathology 90 (2002) 133–144





























