Embed Size (px)
Citation preview

1 3
Exp Brain ResDOI 10.1007/s00221-013-3457-9
REsEaRch aRtIclE
Bilateral stimulation of the subthalamic nucleus has differential effects on reactive and proactive inhibition and conflict-induced slowing in Parkinson’s disease
Ignacio Obeso · Leonora Wilkinson · Maria-Cruz Rodríguez-Oroz · Jose A. Obeso · Marjan Jahanshahi
Received: 31 October 2012 / accepted: 14 February 2013 © springer-Verlag Berlin heidelberg 2013
mean cIs but produced a significant differential effect on the slowest and fastest Rts on conflict trials, further pro-longing the slowest Rts on the conflict trials relative to DBs off and to controls. these results are the first demonstration, using a single task in the same patient sample, that DBs of the stN produces differential effects on reactive and pro-active inhibition and on conflict resolution, suggesting that these effects are likely to be mediated through the impact of stN stimulation on different fronto-basal ganglia path-ways: hyperdirect, direct and indirect.
Keywords Reactive inhibition · Proactive inhibition · conflict resolution · cognitive control · Fronto-striatal circuits · Parkinson’s disease
Introduction
the ability to prevent oneself from engaging in actions that upon first consideration appeared to be the best choice but
Abstract It has been proposed that the subthalamic nucleus (stN) mediates response inhibition and conflict resolution through the fronto-basal ganglia pathways. Our aim was to compare the effects of deep brain stimulation (DBs) of the stN on reactive and proactive inhibition and conflict resolution in Parkinson’s disease using a single task. We used the conditional stop signal reaction time task that provides the stop signal reaction time (ssRt) as a measure of reactive inhibition, the response delay effect (RDE) as a measure of proactive inhibition and conflict-induced slow-ing (cIs) as a measure of conflict resolution. DBs of the stN significantly prolonged ssRt relative to stimulation off. however, while the RDE measure of proactive inhibi-tion was not significantly altered by DBs of the stN, rela-tive to healthy controls, RDE was significantly lower with DBs off but not DBs on. DBs of the stN did not alter the
Electronic supplementary material the online version of this article (doi:10.1007/s00221-013-3457-9) contains supplementary material, which is available to authorized users.
I. Obeso · l. Wilkinson · M. Jahanshahi (*) sobell Department of Motor Neuroscience and Movement Disorders, Ucl, Institute of Neurology, the National hospital for Neurology and Neurosurgery, 33 Queen square, london Wc1N 3BG, UKe-mail: [email protected]
M.-c. Rodríguez-Oroz · J. a. Obeso Department of Neurology, clínica Universitaria and Medical school of Navarra, Neuroscience centre, cIMa, University of Navarra, Pamplona, spain
M.-c. Rodríguez-Oroz · J. a. Obeso centro de Investigación Biomédica en Red sobre Enfermedades Neurodegenerativas (cIBERNED), Instituto carlos III, Ministerio de Investigación y ciencias, Madrid, spain
Present Address: I. Obeso Reward and Decision-Making Group, centre de Neurosciences cognitives, centre National pour la Recherche scientifique, lyon, France
Present Address: l. Wilkinson Brain stimulation Unit, National Institute of Neurological Disorders and stroke, Bethesda, MD 20892-1430, Usa
Present Address: M.-c. Rodríguez-Oroz Department of Neurology, University hospital Donostia and Neuroscience Unit, BioDonostia Research Institute, 20014 san sebastian, Ikerbasque, Basque Foundation for science, Bilbao, spain

Exp Brain Res
1 3
on reflection are shown to be in fact suboptimal plays a fundamental role in flexible behaviour. For instance, when crossing a busy road, the ability to stop oneself from con-tinuing to cross the road if a fast car approaches represents adaptive modification of behaviour triggered by a sud-den and unexpected stimulus, a typical example of reac-tive inhibition. Proactive inhibition involves responding with restraint to meet goals and objectives. In the above example, proactive inhibition or action restraint would be the slowing down of one’s walking pace when approaching the busy road. In everyday life, proactive inhibition often concerns the preparedness to act with restraint when faced with temptations/situations that challenge self-control, such as drinking or smoking or eating sweets. thus, proac-tive inhibition is considered essential for self-control and is the type of inhibition that most often goes awry in psychiat-ric disorders (aron 2011). at times, the necessity to decide between equally salient or valued or incompatible options can induce a conflict. It is proposed that when faced with such conflict between available options, inhibitory control is imposed on responding, to prevent hasty decisions and premature responses until an optimal decision is arrived at (Frank 2006). thus, inhibition is often also a key pro-cess in conflict resolution. these inter-related inhibitory processes, reactive and proactive inhibition and conflict resolution, are essential for executive control and to ensure adaptive behaviour (aron 2011; Frank 2006; Verbruggen and logan 2009b).
the stop signal Rt task (logan and cowan 1984) has been widely used to measure reactive inhibition, through estimation of the stop signal reaction time (ssRt), on tri-als when a stop signal is presented at variable stop signal delays (ssD) after the Go signal, instructing participants to stop the response. In this task, it is proposed that the outcome of a ‘horse race’ between the go and the stop process determines whether the participant successfully stops or fails to stop and responds on the stop trials. In the present study, we used a conditional version of the stop signal task (aron et al. 2007; De Jong et al. 1995; Obeso et al. 2011a, b), which has the advantage of concurrently including conditions involving reactive and proactive inhi-bition and conflict resolution, as reviewed below. On the Go trials of the conditional stop signal task, participants respond to green arrows pointing left or right using the index and middle finger of the dominant hand. For each participant, one of the arrows is assigned as ‘critical’ and the other as ‘non-critical’. Participants are instructed to stop their response when a stop signal (a red cross) is pre-sented following a green arrow in the ‘critical’ direction, but to ignore the stop signal and respond as usual to the Go signal when the stop signal is presented following a green arrow in the ‘non-critical’ direction. the former ‘critical’ stop trials allow estimation of the ssRt using the race
model. the latter ‘non-critical’ stop trials induce a conflict between going (as per instructions) and stopping (induced by appearance of the stop signal), which participants have to resolve, which delays the reaction time on these trials. this conflict-induced slowing can be measured as the dif-ference between the ‘non-critical’ stop and ‘non-critical’ Go trials. the difference in Rts to ‘critical’ and ‘non-crit-ical’ Go trials, the response delay effect, is the measure of proactive inhibition in this task as it reflects the partici-pants’ strategic slowing of responses on ‘critical’ but not ‘non-critical’ Go trials in anticipation that the former are followed by a stop signal.
Imaging studies in healthy participants have shown that successful motor inhibition on a stop signal reaction time task is associated with activation of the subthalamic nucleus (stN) together with frontal areas such as the pre-supplementary motor area (pre-sMa), inferior frontal cor-tex (IFc) and the anterior cingulate (aron et al. 2007; aron and Poldrack 2006; Rubia et al. 2003; li et al. 2006, 2008). Furthermore, in an fMRI study using a conditional version of the stop signal reaction time task, significant activation of the ‘braking’ network (right stN, IFc and pre-sMa) was described (aron et al. 2007), in association with both reactive inhibition in response to a stop signal and con-flict-induced slowing on trials when the stop signal was presented but had to be ignored. In imaging studies, proac-tive inhibition through adjustment of response strategies, for example, by slowing down responses to improve the likelihood of stopping, has been related to activation of the striatum (Zandbelt and Vink 2010; Vink et al. 2005). On the conditional stop signal task, responding with restraint on ‘critical’ Go trials when a stop signal could occur was asso-ciated with activation of the striatum, IFc and pre-sMa (Jahfari et al. 2010). In monkeys, recording of single- and multiple-unit activity and intracranial local field potentials during the stop signal task has demonstrated that neuronal activity in the sMa plays a role in both proactive control of motor readiness and reactive cancellation of a response in this task (chen et al. 2010).
Parkinson’s disease (PD) is characterized by increased and abnormally synchronized stN neuronal activity (Kuhn et al. 2009; Wichmann et al. 1994), which is consid-ered a key pathophysiological feature of the disorder and the rationale for surgical treatment with deep brain stimu-lation (DBs) of the stN or subthalamotomy (Obeso et al. 1997; hutchison et al. 1998). consistent with the imaging results implicating the stN in motor inhibition, there is evidence indicating that DBs of the stN in PD signifi-cantly alters ssRts on the standard stop signal reaction time task (Mirabella et al. 2011; Ray et al. 2009; swann et al. 2011; van den Wildenberg et al. 2006) or suppression of unwanted saccades in a memory-guided saccade para-digm (Yugeta et al. 2010). studies in the rat have shown
Exp Brain Res
1 3
that lesions of the stN are associated with increased pre-mature responses (Baunez et al. 1995), impulsivity and an inability to inhibit operant responses (Wiener et al. 2008), and a generalized impairment of stopping on a modified stop signal reaction time task (Eagle et al. 2008). Proac-tive action restraint in go/no-go Rt tasks (Ballanger et al. 2009; hershey et al. 2004, 2010) is altered with DBs of the stN. Finally, conflict resolution on paradigms such as the stroop (Jahanshahi et al. 2000; Witt et al. 2004) or a probabilistic decision-making task (Frank et al. 2007) and inhibition of incompatible responses in a simon task (Wylie et al. 2010) are also altered by stN DBs in patients with PD.
the evidence reviewed above suggests a potential effect of DBs of the stN in PD on reactive and proactive inhibi-tion and conflict resolution. the novel aspect of the present study is that we investigated the effect of DBs of the stN on reactive and proactive inhibition and conflict resolution in a single sample of PD patients using the same task. to this end, we used the conditional stop signal task (aron et al. 2007; De Jong et al. 1995; Obeso et al. 2011a, b) that allows measurement of inhibition under three different con-ditions: reactive, proactive and conflict resolution. We pre-dicted that along with the general improvement of the motor symptoms of PD, speed of movement initiation on Go trials would become faster with stN DBs. In contrast, based on the imaging and behavioural results reviewed above show-ing that inhibitory control and conflict resolution are worse in PD with stN DBs on than off (Ballanger et al. 2011; Frank et al. 2007; Jahanshahi et al. 2000; Witt et al. 2004), we predicted that:
1. ssRts as the index of reactive motor inhibition would become prolonged with DBs on versus off;
2. proactive inhibition as measured by the response delay effect (RDE) would be significantly lower with stN DBs on versus off;
3. the need for conflict resolution would result in further prolongation of Rts on conflict trials in PD with DBs on versus off.
Methods
Participants
Fifteen patients with PD (13 male and right handed, mean age 59.88, sD 6.1; range 47–70 years) took part in the study. all met UK Brain Bank diagnostic criteria for PD (hughes et al. 1992). all patients had been treated with bilateral DBs of the stN for at least 4 months (M = 28.31, sD = 20.39) and were recruited from the clinica Universidad de Nav-arra, Pamplona (spain). all patients were screened for the absence of dementia (Mini Mental Examination scores >26, MMsE, Folstein et al. 1975) and major depression (Beck Depression Inventory scores <18, Beck et al. 1961). None of the patients had major psychiatric illness, head injury, or history of alcohol or drug abuse. Post-surgical MRI con-firmed the correct placement of electrodes. all patients were treated preoperatively with levodopa in combination with a dopamine agonist, with a mean levodopa equivalent daily dose of 1,035.16 milligrams (sD = 188.9). Patients were assessed while on their usual medication.
twenty-nine healthy volunteers (15 male, 23 right handed, mean age 56.97, sD 6.9; range 45–68 years) par-ticipated in the study. None of the controls had any neuro-logical disorder, psychiatric illness, head injury, or history of alcohol or drug abuse. the study was approved by the Joint Ethics committee of the Institute of Neurology and the National hospital for Neurology and Neurosurgery and the University of Navarra clinical Investigation commit-tee. Informed consent was obtained from all participants. Information about controls and PD patients is presented in table 1 (also see table in supplementary Material).
conditional stop signal task
We employed the conditional version of the stop signal task (aron et al. 2007; De Jong et al. 1995; Obeso et al. 2011a, b), which has the advantage that it allows concurrent meas-urement of speed of initiating a response (Go Rts), inhib-iting a response (ssRt), proactive inhibition (response
Table 1 Demographic information and clinical characteristics for Parkinson’s disease patients and healthy controls
Group age (years)
Education (years)
BDI (0–63) MMsE (0–30) UPDRs-III on med, DBs on
UPDRs-III on med, DBs off
Disease duration (years)
Parkinson’s disease (n = 15)
58.93 (6.5) 13.36 (3.7) 8.20 (4.3) 29.07 (1.0) 14.69 (6.8) 25.46 (6.4) 13.47 (4.2)
healthy control (n = 29)
56.97 (6.9) 15.72 (3.5) 6.07 (5.1) 29.59 (0.6)
p 0.49 0.09 0.10 0.20 <53.88 % improvement
UPDRs-III: part III or motor section of the Unified Parkinson’s Disease Rating scale
DBS deep brain stimulation, BDI beck depression inventory, MMSE mini mental state examination

Exp Brain Res
1 3
delay effect, RDE) and speed of initiating a response under conditions of conflict (conflict-induced slowing, cIs) (Fig. 1).
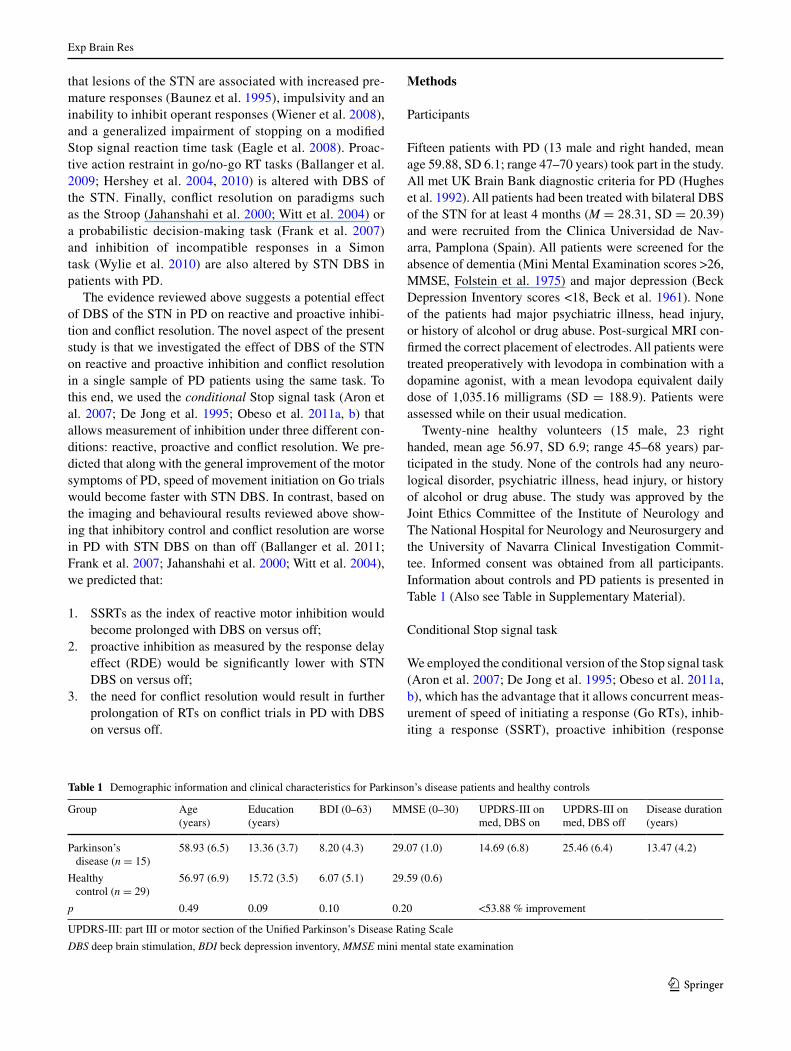
the conditional stop signal task involved a series of Go and stop trials. On Go trials (75 % of all trials), a Go signal (either a left or right pointing green arrow) was presented on the computer screen and participants had to respond as fast as possible using the index and middle fingers of their domi-nant hand to press either a left or a right key. On stop trials (25 % of all trials), a stop signal (red cross) was presented following the Go signal, after a variable stop signal delay (ssD). For each participant, either the left or right pointing arrows were designated as the ‘critical’ direction. When a stop signal was presented following a Go signal in the ‘criti-cal’ direction, participants had to stop their response to the Go signal. In contrast, if a stop signal was presented follow-ing a Go signal in the ‘non-critical’ direction, participants had to ignore the stop signal and respond to the Go signal. there were three blocks of trials, consisting of 32 stop and 96 Go trials per block (128 trials per block, 384 trials in total). In each block, the number of left and right pointing arrows was equal and for every four trials, there was 1 stop and 3 Go trials. to replicate the procedures used by aron et al. (2007) in their imaging study of the conditional stop signal task closely, durations from 0.5 to 4 s were inserted between the stop or Go trials.
the ssD value for the stop trials was sampled from one of four staircases, changing dynamically throughout the task based on the participant’s response. Initially, the four stair-cases started with ssD values of 100, 150, 200 and 250 ms, respectively. successful inhibition of a response on a stop trial made inhibition more difficult on the next stop trial by increasing the ssD by 50 ms. In contrast, if a response was not successfully inhibited, inhibition became easier by decreasing the ssD by 50 ms. staircases of four step-up and step-down algorithms were used in this way to ensure convergence to P(inhibit) of 50 % by the end of the three blocks. Each staircase (100, 150, 200 and 250 ms) moved four times within each block for stop trials of the ‘critical’ direction. ssDs for stop trials of the ‘non-critical’ direction were yoked to the ‘critical’ direction values. the staircases were independent but were randomly mixed in a block of trials.
Participants were instructed to respond to the green arrows by pressing the correct response key as fast and as accurately as possible, while at the same time also look out for the appearance of the red cross and try to withhold the response when this followed a green arrow pointing in the ‘critical’ direction. Participants were specifically instructed not to let their performance on the stopping task interfere with their performance on the Go task. twenty practice tri-als were completed before the task was started.
to ensure the ‘context independence’ assumption of the Race Model, and that presentation of the stop signal does not change the Go Rt distribution, we examined the indi-vidual data of each patient block by block to detect any instances where ‘critical’ stopRespond Rts (failed stop tri-als) were longer than ‘critical’ Go Rts. to overcome this violation, it has been recommended that data from partici-pants or blocks in which ‘critical’ stopRespond Rts are longer than ‘critical’ Go Rts should be discarded (logan 2013, personal communication). In the DBs on condition, 8/16 patients showed ‘critical’ stopRespond Rt > ‘critical’ Go Rt on one block, whereas in the DBs off condition, 6/16 patients had ‘critical’ stopRespond Rt > ‘critical’ Go Rts in one block. these blocks were discarded from further analy-sis. One patient was discarded from the analysis since the violation was present across all blocks, reducing the sample size to the 15 reported in tables 1 and 2.
Using the standard Race Model (logan and cowan 1984), we estimated the stop signal reaction time (ssRt) using the ssRt integration method calculated independently for each block as recently recommended (Verbruggen et al. 2013). the integration method overcomes many problems asso-ciated with skewed Go Rt distributions or slowing which could occur in Parkinson’s disease. In this method for the calculation of ssRts, first, correct Go trials in the critical direction are rank-ordered. second, for each participant, the nth Go Rt value was obtained by multiplying the number of
Fig. 1 Examples of different types of trials on the conditional stop signal task. Every trial begins with a fixation circle. the first trial is a Go trial. the second trial in the ‘critical’ direction (where participants are asked to try to stop upon stop signal presentation) has a Go signal replaced with a stop signal after a variable stop signal delay (ssD). the last trial in the ‘non-critical’ direction (where participants are asked to ignore the stop signal and keep responding) has a Go trial followed by a stop signal that needs to be ignored and participants should respond to Go signal. three key measures can be obtained: 1 the stop signal reaction time (ssRt = ‘critical’ Go Rt-mean ssD at P inhibit 50 %); 2 response delay effect (RDE = mean ‘critical’ Go Rt-mean ‘non-critical’ Go Rt); 3 conflict-induced slowness (cIs = ‘non-critical’ stopRespond Rt-‘non-critical’ Go Rt)
Exp Brain Res
1 3
Tabl
e 2
Mea
ns a
nd s
tand
ard
devi
atio
ns o
f th
e m
easu
res
of i
nter
est
on t
he c
ondi
tiona
l st
op s
igna
l re
actio
n tim
e ta
sk f
or p
atie
nts
with
Par
kins
on’s
dis
ease
with
dee
p br
ain
stim
ulat
ion
(DB
s) o
f th
e su
btha
lam
ic n
ucle
us o
n or
off
and
for
hea
lthy
cont
rols
Mea
sure
Des
crip
tion
of m
easu
rePD
DB
s on
PD D
Bs
off
DB
s on
ver
sus
off
(w
ithin
sub
ject
s)c
ontr
ols
Gro
up c
ompa
riso
n (b
etw
een-
grou
ps)
‘Cri
tica
l’ di
rect
ion
Go
Rt
Rt
to G
o si
gnal
in ‘
criti
cal’
di
rect
ion
547.
03 (
105.
1)53
7.95
(10
9.4)
t (14)
= 0
.47,
p =
0.6
449
5.69
(89
.7)
(a)
t (43)
= 1
.71,
p =
0.0
9(b
) t (4
3) =
1.3
9, p
= 0
.17
stop
inhi
bit
% c
orre
ct in
hibi
tion
48.1
2 (7
.1)
50.9
9 (8
.8)
2 z =
−1.
62, p
= 0
.11
52.5
9 (1
0.1)
(1 a) z
= −
1.20
, p =
0.2
0(1 b)
z =
−0.
37 p
= 0
.70
stop
Res
pond
Rt
Rt
on
stop
tria
ls o
n w
hich
pa
rtic
ipan
t fai
led
to s
top
511.
69 (
97.0
)49
6.47
(79
.45)
t (14)
= 0
.74,
p =
0.4
646
5.85
(79
.6)
(a)
t (43)
= 1
.67,
p =
0.1
0(b
) t (4
3) =
1.2
9, p
= 0
.23
Go
erro
rsN
umbe
r of
om
issi
ons
6.80
(7.
6)7.
27 (
7.0)
2 z =
−1.
90, p
= 0
.06
0.62
(1.
0)(1 a)
z =
−3.
14, p
= 0
.002
(1 b) z
= −
4.79
, p <
0.0
01
stop
sig
nal d
elay
Del
ay b
etw
een
Go
and
st
op s
igna
ls12
7.36
(69
.5)
170.
87 (
101.
1)t (1
4) =
−2.
85, p
= 0
.01
209.
15 (
95.2
)(a
) t (4
3) =
−3.
29, p
< 0
.01
(b)
t (43)
= 1
.23,
p =
0.2
2
stop
sig
nal r
eact
ion
time
Mea
sure
of
inhi
bitio
n41
3.03
(13
4.2)
305.
99 (
82.8
)t (1
4) =
2.5
0, p
= 0
.02
272.
22 (
69.5
)(a
) t (4
3) =
4.6
8, p
< 0
.001
(b)
t (43)
= 1
.46,
p =
0.1
5
‘Non
-cri
tica
l’ di
rect
ion
Go
Rt
Rt
to G
o si
gnal
in
‘non
-cri
tical
’ dir
ectio
n51
4.98
(10
5.8)
516.
45 (
92.7
)t (1
4) =
−0.
09, p
= 0
.92
433.
14 (
44.3
)(a
) t (4
3) =
3.6
3, p
= 0
.001
(b)
t (43)
= 4
.05,
p <
0.0
01
stop
inhi
bit
% in
corr
ect i
nhib
ition
(f
ailu
re to
res
pond
on
‘n
on-c
ritic
al’ s
top
tria
l)
10.8
7 (7
.2)
12.8
1 (1
0.6)
2 z =
−0.
53, p
= 0
.59
3.10
(4.
4)(1 a)
z =
−3.
29, p
= 0
.001
(1 b) z
= −
2.63
, p <
0.0
1
stop
Res
pond
Rt
Rt
to ‘
non-
criti
cal’
tria
ls
with
a ‘
to b
e ig
nore
d’
stop
sig
nal
665.
40 (
205.
0)66
7.00
(20
8.8)
t (14)
= −
0.06
, p =
0.9
555
2.30
(78
.9)
(a)
t (43)
= 2
.63,
p =
0.0
1(b
) t (4
3) =
2.6
3, p
= 0
.01
Go
erro
rsN
umbe
r of
om
issi
ons
9.60
(8.
4)10
.87
(11.
6)2 z
= −
1.41
, p =
0.1
50.
45 (
0.7)
(1 a) z
= −
4.01
, p <
0.0
01(1 b)
z =
−4.
94, p
< 0
.001
con
flict
-ind
uced
slo
win
gM
easu
re o
f co
nflic
t-
indu
ced
slow
ing
150.
42 (
121.
1)15
0.54
(14
5.6)
t (14)
= 0
.01,
p =
0.9
911
9.15
(67
.6)
(a)
t (43)
= 1
.10,
p =
0.2
7(b
) t (4
3) =
0.9
8, p
= 0
.33
Oth
er v
aria
bles
Go
disc
rim
inat
ion
erro
rsD
ecis
ion
erro
rs–p
ress
ing
w
rong
res
pons
e bu
tton
24.2
0 (1
8.2)
19.4
0 (1
4.9)
2 z =
−1.
60, p
= 0
.10
0.45
(0.
6)(1 a)
z =
−5.
12, p
< 0
.001
(1 b) z
= −
5.42
, p <
0.0
01
Res
pons
e de
lay
effe
ctD
iffe
renc
e sc
ore
of ‘
criti
cal’
G
o R
t-‘n
on-c
ritic
al’ G
o R
t32
.05
(36.
5)21
.50
(60.
8)t (1
4) =
0.4
9, p
= 0
.62
62.5
5 (6
1.9)
(a)
t (43)
= −
1.01
, p =
0.0
8(b
) t (4
3) =
−2.
66, p
= 0
.02
stop
Res
pond
: th
e av
erag
e R
t f
or r
espo
nses
fol
low
ing
a st
op s
igna
l. st
op s
igna
l de
lay
(ssD
): t
he a
vera
ge s
sD,
com
pute
d fr
om f
our
stai
rcas
es (
see
“Met
hods
”) a
t th
e po
int
at w
hich
p(
inhi
bit)
= a
bout
50
%. s
top
sign
al r
eact
ion
time
(ssR
t):
com
pute
d by
sub
trac
ting
the
mea
n ss
D f
rom
the
mea
n ‘c
ritic
al’ G
o R
t f
or e
ach
part
icip
ant.
Go
omis
sion
err
ors:
fai
lure
to r
espo
nd o
n a
Go
tria
l. G
o di
scri
min
atio
n er
rors
: pr
essi
ng t
he r
espo
nse
key
in t
he o
ppos
ite d
irec
tion
indi
cate
d by
the
stim
ulus
. con
flict
-ind
uced
slo
win
g (c
Is):
mea
sure
d by
sub
trac
ting
mea
n ‘n
on-c
ritic
al’
Go
Rt
s fr
om m
ean
‘non
-cri
tical
’ sto
pRes
pond
Rt
s
(a)
DB
s on
ver
sus
cont
rols
inde
pend
ent t
test
s; (
b) D
Bs
off
vers
us c
ontr
ols
inde
pend
ent t
test
s. P
aire
d t t
ests
wer
e ap
plie
d fo
r D
Bs
on v
ersu
s of
f co
mpa
riso
n1
Man
n–W
hitn
ey u
sed
for
betw
een-
grou
ps c
ompa
riso
ns2
Wilc
oxon
use
d fo
r w
ithin
-sub
ject
s co
mpa
riso
n

Exp Brain Res
1 3
Go trials in the distribution by the probability of respond-ing to the stop signal. Finally, the mean ssD is subtracted from the nth Go Rt to obtain the integrated ssRt value (Verbruggen et al. 2010). the ssD estimation was averaged from the mean values for the last six moves in each of the four staircases when the participant had converged on 50 % inhibition.
On this task, ‘critical’ Go Rts are generally slower than ‘non-critical’ Go Rts, because the necessity to stop the response with presentation of a stop signal on the former but not the latter type of Go trials results in a context-specific, strategic and proactive inhibition of responses. to quantify this ‘proactive’ inhibition/action restraint, we calculated the response delay effect (RDE), the difference in Rts between the ‘critical’ and ‘non-critical’ Go Rts. the measure of ‘conflict-induced slowing’ (cIs) was obtained by subtract-ing mean Rts for ‘non-critical’ Go trials from mean Rts for ‘non-critical’ stop trials (‘non-critical’ stopRespond Rts, Fig. 1). In addition to the analysis of the effects of DBs on mean Rt measures, we also examined how the fastest and slowest Rts were influenced by stN stimulation by ana-lysing the top and bottom Rt quartiles. this analysis was motivated by two considerations. First, such finer grained distribution analysis of Rts can be informative in clarifying which portion of the distribution, if any, is being altered by stN stimulation. For example, we have previously shown that DBs of the stN improved simple and choice Rts in PD by speeding up the 10 % slowest responses (Brown et al. 1999). second, a previous study has shown that the DBs of the stN in PD can produce opposite effects on inhibition of fast and slow responses in the simon task (Wylie et al. 2010).
Procedure
PD patients completed the conditional stop signal task twice: once with DBs ‘on’ and once with DBs ‘off’, in a counterbalanced order. the time between turning DBs off/on and starting the assessment (M = 52.81 min, sD = 29.8) was sufficiently long to ensure the clinical benefits of DBs had dissipated or returned. For 14/29 controls, the ‘criti-cal’ direction was left. For 9/15 PD patients with DBs on, the ‘critical’ direction was left and right for the remaining patients, reversed for the second performance.
statistical analysis
the analysis was completed on individual Rt measures, error scores and difference scores using sPss 16.0. For each of these measures, the analysis involved two main sets of comparisons: (1) DBs on versus DBs off, and (2) PD patients with DBs on or off relative to healthy con-trols. Given the within-subject nature of the former set of
comparisons, paired t tests were used for individual Rt and difference scores, and Wilcoxon matched pair tests for anal-ysis of errors. For the comparison of healthy controls to PD patients with DBs on or off, independent group t tests were employed for the individual Rt and Rt difference scores and Mann–Whitney U tests were used for the error data. the results involve three key sets of comparisons: DBs on versus DBs off, DBs on versus controls and DBs off versus controls. For each of these comparisons, a holm–Bonferroni correction was applied, with a p < 0.017 considered signifi-cant. For the comparison of the effect of DBs on versus off on the fastest and slowest Rt quartiles, we used a series of two-way repeated measures analyses of variance (aNOVa) with DBs (on vs. off) and quartiles (fastest vs. slowest) as the within-subject repeated measures factors. the associa-tion of variables was examined with Pearson or spearman correlation coefficients, as appropriate.
Results
the groups were matched in terms of age (PD: 59.88 (6.1), range 47–70; controls: 56.97 (6.9), range 45–68). DBs of the stN was associated with significant improvement in the motor symptoms of PD. Off medication scores on the motor section of the Unified Parkinson’s disease Rating scale (UPDRs-III; Fahn and Elton 1987) with DBs on were significantly better than that with DBs off (table 1). all patients improved substantially paralleling the results of larger series from our centre (Rodriguez-Oroz et al. 2005), which together with the post-operative MRI verification suggested correct positioning of at least one of the electrode contacts in or near the stN target.
conditional stop signal task
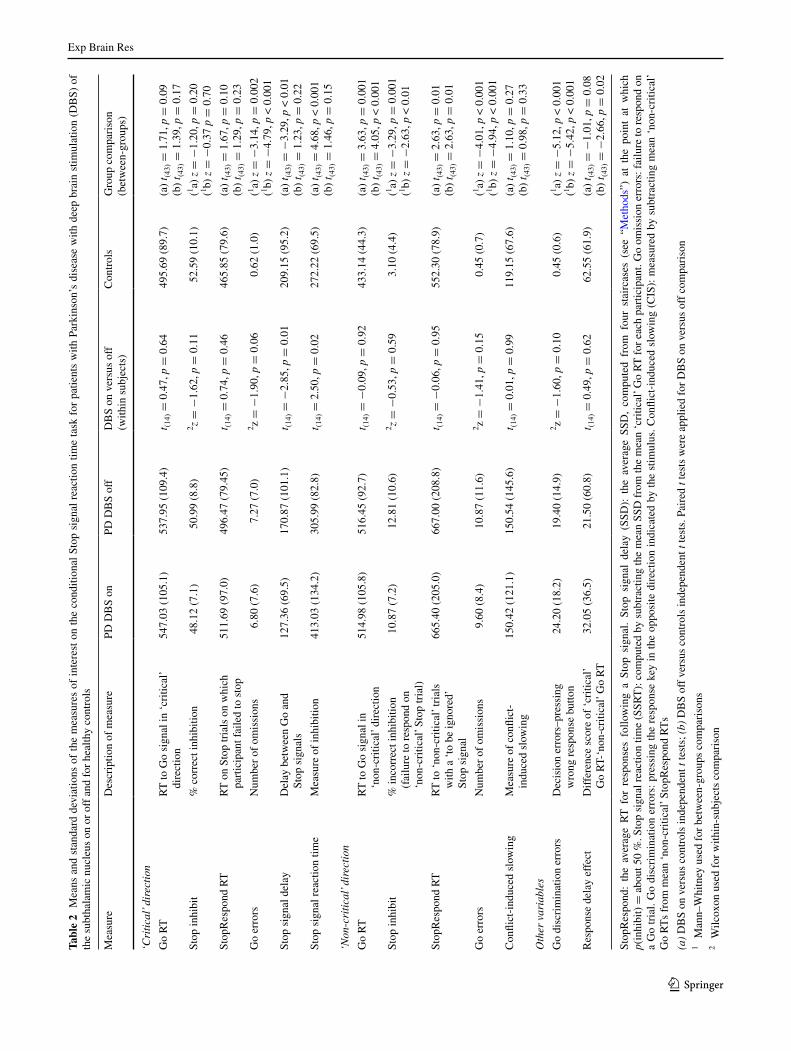
Figures 2, 3, 4, 5 and 6 and table 2 show the data relating to performance on the conditional stop signal task for ‘critical’ and ‘non-critical’ Go and stop trials for patients with DBs on and off and controls.
‘critical’ trials and ssRt
Inhibition close to 50 % of trials was achieved by PD patients with DBs on with a mean ssD (M = 127.36, sD = 69.5) which was significantly shorter (i.e. easier to inhibit) than the mean ssD with DBs off (M = 170.87, sD = 101.1; t(14) = −2.85, p = 0.01). controls achieved inhibition on about 50 % of trials with a mean ssD (M = 209.15, sD = 95.2) which was significantly longer (i.e. more difficult to inhibit) than for PD patients with DBs on (t(43) = −3.29, p = 0.01) but not with DBs off (p > 0.05).
Exp Brain Res
1 3
Importantly, as predicted, DBs of the stN significantly prolonged/worsened ssRt relative to DBs off (p = 0.02) (see table 2; Fig. 3). For the control participants, the ssRt was significantly faster (indicative of better inhibition) than PD with DBs on (p < 0.001), but the differences were not significant for DBs off (p = 0.15).
‘Non-critical’ trials and cIs
PD patients with DBs on and off responded significantly slower on ‘non-critical’ Go trials than controls (p = 0.001; table 2). similarly, PD patients had significantly slower ‘non-critical’ stopRespond Rts than controls with both DBs on and off (p = 0.01, table 2). stimulation condition did not influence ‘non-critical’ Go or stopRespond Rts either (p > 0.05). the mean cIs difference score—the measure of conflict-induced slowing calculated by subtracting mean
Fig. 2 Mean Rts in millisec-onds in the ‘critical’ direction for the Go and stopRespond (trials with a stop signal on which participants failed to stop and responded) and ‘non-criti-cal’ Go and stopRespond trials (trials with a to be ignored stop signal) for healthy controls and patients with Parkinson’s dis-ease with deep brain stimulation of the subthalamic nucleus on (PD DBs on) or off (PD DBs off). Error bars sE of the mean
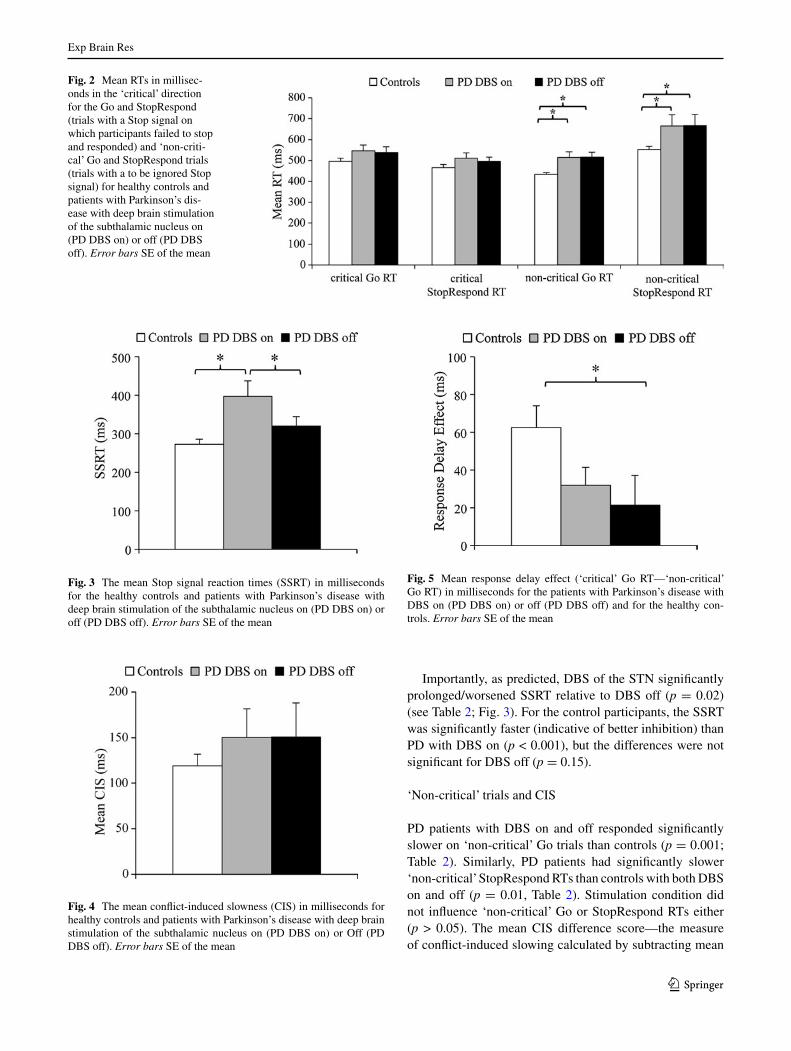
Fig. 3 the mean stop signal reaction times (ssRt) in milliseconds for the healthy controls and patients with Parkinson’s disease with deep brain stimulation of the subthalamic nucleus on (PD DBs on) or off (PD DBs off). Error bars sE of the mean
Fig. 4 the mean conflict-induced slowness (cIs) in milliseconds for healthy controls and patients with Parkinson’s disease with deep brain stimulation of the subthalamic nucleus on (PD DBs on) or Off (PD DBs off). Error bars sE of the mean
Fig. 5 Mean response delay effect (‘critical’ Go Rt—‘non-critical’ Go Rt) in milliseconds for the patients with Parkinson’s disease with DBs on (PD DBs on) or off (PD DBs off) and for the healthy con-trols. Error bars sE of the mean
Exp Brain Res
1 3
‘non-critical’ Go Rts from mean ‘non-critical’ stopRespond Rts—was comparable for controls and for PD patients with DBs on or off (p > 0.05). Most notable was the finding that there was no difference in mean cIs for PD patients with DBs on and off (p > 0.05) (see table 2; Fig. 4).
Effect of stN stimulation on proactive inhibition/action restraint
as expected, for the controls, ‘non-critical’ Go Rts were significantly faster than the ‘critical’ Go Rts (t(28) = 5.43, p < 0.001). With DBs on, PD patients also had signifi-cantly faster ‘non-critical’ Go Rts than ‘critical’ Go Rts (t(14) = 3.39, p = 0.004), indicating operation of proac-tive action restraint during ‘critical’ trials, whereas with DBs off, this difference was not significant (t(14) = 1.36, p = 0.19). In fact, for the response delay effect (RDE, table 2; Fig. 5), the difference score between ‘critical’ and ‘non-critical’ Go Rts, which is a measure of proactive action restraint, was significantly greater for the controls than PD patients with DBs off (t(43) = −2.66, p = 0.02). For the comparison of controls versus PD with DBs on, the RDE difference was not significant (t(43) = −1.01, p = 0.08). the RDE was higher, indicative of greater proac-tive action restraint, with DBs on (M = 32.05, sD = 36.5) than off (M = 21.50, sD = 60.8), but the difference was not significant (t(14) = 0.49, p = 0.62). In addition, while the magnitude of the RDE was significantly different from zero for the controls (t(28) = 5.44 p < 0.001) and PD patients with stN DBs on (t(14) = 3.39, p < 0.01), indicating pres-ence of notable proactive inhibition, this was not the case for the patients with DBs off (t(14) = 1.36, p = 0.19).
‘critical’ and ‘non-critical’ Go and stopRespond Rts
contrary to our prediction, despite significant improve-ment of the motor symptoms with stN DBs, there was
no difference in ‘critical’ or ‘non-critical’ Go Rts for PD patients with DBs on and off (see table 2; Fig. 5). PD patients with DBs on and off responded more slowly on ‘critical’ and ‘non-critical’ Go trials than controls, but the differences were only significant for the ‘non-critical’ Go trials (p < 0.001), but not the ‘critical’ Go trials (table 2). Furthermore, ‘critical’ stopRespond Rts (stop trials on which participants failed to inhibit and responded) were slower for PD patients with both DBs on and off rela-tive to controls, but the differences were not significant (p > 0.05). there was no difference in ‘critical’ stopRe-spond Rts for PD patients with DBs on and off (see table 2). the ‘non-critical’ stopRespond Rts (trials with ‘to be ignored’ stop signal) for the patients were signifi-cantly slower than those of controls both with DBs on and off (p < 0.01), but the differences with or without stN stimulation were not significant (p > 0.05).
analysis of the error data in table 2 is presented in the supplementary Material.
Effect of stN stimulation on the fastest and slowest Rts
We also investigated the effect of stN stimulation on the two tail ends of the distribution for every Rt measure (‘criti-cal’ Go Rt, ‘critical’ stopRespond Rt, ‘non-critical’ GoRt, ‘non-critical’ stopRespond Rt) and difference scores (ssRt, cIs and RDE). significant results were obtained for the ‘non-critical’ stopRespond Rt and the cIs.
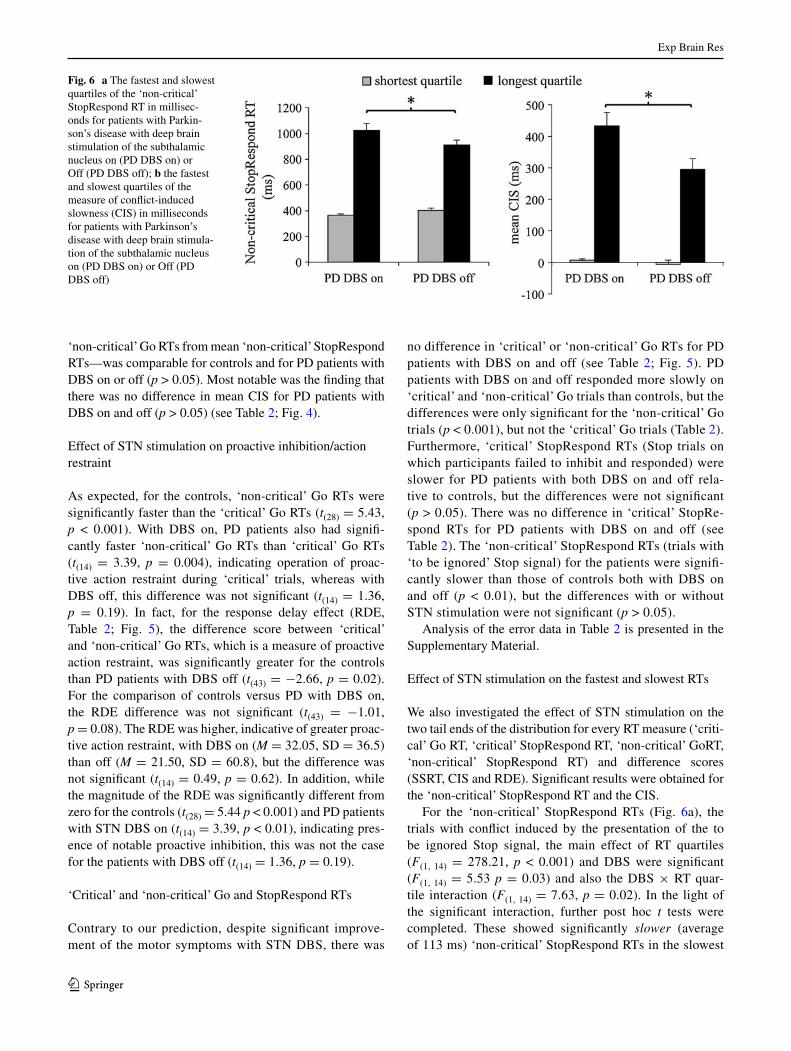
For the ‘non-critical’ stopRespond Rts (Fig. 6a), the trials with conflict induced by the presentation of the to be ignored stop signal, the main effect of Rt quartiles (F(1, 14) = 278.21, p < 0.001) and DBs were significant (F(1, 14) = 5.53 p = 0.03) and also the DBs × Rt quar-tile interaction (F(1, 14) = 7.63, p = 0.02). In the light of the significant interaction, further post hoc t tests were completed. these showed significantly slower (average of 113 ms) ‘non-critical’ stopRespond Rts in the slowest
Fig. 6 a the fastest and slowest quartiles of the ‘non-critical’ stopRespond Rt in millisec-onds for patients with Parkin-son’s disease with deep brain stimulation of the subthalamic nucleus on (PD DBs on) or Off (PD DBs off); b the fastest and slowest quartiles of the measure of conflict-induced slowness (cIs) in milliseconds for patients with Parkinson’s disease with deep brain stimula-tion of the subthalamic nucleus on (PD DBs on) or Off (PD DBs off)

Exp Brain Res
1 3
quartile for DBs on (M = 1,025.53; sD = 236.3) com-pared to DBs off (M = 912.96; sD = 145.0; t(14) = 2.04, p = 0.03). In contrast, for the fastest quartile, ‘non-crit-ical’ stopRespond Rts were faster (average of 36 ms) with DBs on (M = 364.65; sD = 52.0) than DBs off (M = 400.66; sD = 82.2), but the difference was not sig-nificant (t(14) = −1.67, p = 0.16).
For the cIs measure, the aNOVa showed significant main effects of Rt quartile (F(1,14) = 91.02, p < 0.001) and DBs (F(1, 14) = 7.98, p = 0.01), but no significant DBs × Rt quartile interaction (F(1, 14) = 1.88, p = 0.19).
correlational analysis
We examined the association between DBs on versus off measures of ssRt, cIs, RDE and errors with the clinical and demographic characteristics of the PD patients (i.e. age, duration of illness, UPDRs-III, levodopa dose equivalents and MMsE) and with DBs parameters. No correlations showed notable or significant results.
DBs parameters (frequency, voltage or pulse width) did not show any significant correlations with any Rt measures or difference scores. however, the number of discrimina-tion errors both with DBs on (r = 0.59, p = 0.010) and off (r = 0.66, p = 0.007) was significantly and positively correlated with pulse width, indicative of a lasting effect of DBs on response selection accuracy even after the stimula-tors were switched off.
Discussion
the patients with PD clearly benefited from DBs surgery as shown by a significant improvement of UPDRs-III scores with DBs on compared to off, indicating marked improve-ment of the motor symptoms with stN stimulation. Despite this improvement of the motor symptoms, DBs of the stN significantly slowed ssRt compared to DBs off (in 13/15; 86 % of the sample) and relative to controls. In addition, with DBs switched on, patients only achieved successful inhibition with a significantly shorter (‘easier’) mean stop signal delay (ssD) relative to both DBs off and controls. In contrast, the response delay effect (RDE), the index of proactive inhibition, was significantly worse than controls only with DBs off, but not with DBs on. turning DBs on did not have a significant effect on mean cIs. however, on the trials with conflict, with stN stimulation on, the slowest ‘non-critical’ stopRespond Rts became differentially and significantly slower (by an average of 109 ms) relative to DBs off. Our results suggest differential effects of DBs of the stN on reactive and proactive inhibition and conflict-induced slowing.
DBs of the stN slowed down reactive motor inhibition in PD
DBs of the stN prolonged/slowed down ssRts in PD rela-tive to when the stimulators were off. Our finding that ssRts with DBs on were significantly longer than with DBs off indicate that with stN stimulation, PD patients could only inhibit the response with shorter ssDs (inhibition easier) when the Go response was not triggered and was being planned rather than close to execution. In their fMRI study with healthy controls, aron and Poldrack (2006) found that stN activation on successful stop trials was greater with longer ssDs, indicating that the stN was more engaged in successful inhibition in trials when the go process was closer to execution. this suggests a greater role for the stN in late phases of inhibition, when the Go response has already been triggered and is close to execution, which requires fast inhi-bition. this means that with stN stimulation, PD patients cannot engage in fast inhibition of their responses when a stop signal is presented. this is consistent with the finding that in PD, fast premature response captured by the incon-gruent stimulus was increased with stN stimulation in the simon interference task, suggesting impairment of fast inhi-bition (Wylie et al. 2010).
In PD, stN DBs ‘on’ versus ‘off’ methodology has been adopted in four previous studies with the standard version of the stop signal Rt task, the results of which are inconsist-ent: three reporting that bilateral DBs of the stN improved ssRts in PD (Mirabella et al. 2011; swann et al. 2011; van den Wildenberg et al. 2006), whereas the other found that unilateral stN stimulation had no overall effect on mean ssRts but slowed ssRts in the contralateral hand only for those PD patients with baseline (DBs off) ssRts similar to controls (Ray et al. 2009). Our results with bilateral DBs of the stN on the conditional stop signal task are consist-ent with those of Ray et al. (2009) on the standard version of this task. the inconsistent results of stN DBs in PD on ssRt across these studies with the stop signal task may be related to a number of factors. First, the above studies use the mean ssRt estimation, which can be problematic in patient populations with skewed and slowed Rt distribu-tions (Verbruggen et al. 2013). second, baseline effects are important in determining the direction of stN DBs-induced effects on ssRt (Ray et al. 2009). Finally, the proportion of stop signals and prepotency of the Go response, whether DBs was unilateral (Ray et al. 2009) or bilateral (Mirabella et al. 2011; swann et al. 2011; van den Wildenberg et al. 2006), whether stop signal task performance was uniman-ual (present study; Mirabella et al. 2011; Ray et al. 2009) or bimanual (swann et al. 2011; van den Wildenberg et al. 2006), the type of movement performed (reaching, Mira-bella et al. 2011 or manual key press, present study; Ray et

Exp Brain Res
1 3
al. 2009; van den Wildenberg et al. 2006), whether patients were assessed on (present study; Ray et al. 2009; swann et al. 2011; van den Wildenberg et al. 2006) or off (Mirabella et al. 2011) medication are important methodological differ-ences across studies that would affect the results.
stN stimulation and conflict resolution in PD
as expected, the ‘non-critical’ stopRespond Rts were the slowest Rts in this task for the healthy controls as well as the PD patients with both DBs on and off, which suggests that this conflict was operational for all participants. also as expected, in PD, the ‘non-critical’ stopRespond Rts were significantly slower than those of healthy controls with both DBs on and off. the differential effect of stN DBs on the slowest (which became significantly slower with DBs on than off) and fastest Rts (non-significantly faster with DBs on than off) to the ‘non-critical’ conflict trials is intriguing. this may reflect adoption of two alternative strategies on the conflict trials. On some trials, the participants’ respond-ing could have been guided by the appropriate rule derived from the experimental instructions that ‘with an arrow in the “non-critical” direction, ignore the stop signal and always respond’. this would have resulted in fast Rts on these con-flict trials, which were further speeded up by an average of 36 ms with stN DBs on. Other trials may have entailed adoption of a second general strategy of ‘stop the response when a stop signal is presented’ regardless of whether this occurred on a ‘critical’ or ‘non-critical’ trial. adoption of such a general stopping strategy would have required reacti-vation of the response on the ‘non-critical’ stop trials, which would have further prolonged the Rts already slowed by presentation of a stop signal. these slowest ‘non-critical’ stopRespond Rts were significantly prolonged by stN stimulation in PD, suggesting that on average it took a fur-ther 113 ms to reactivate the inhibited response when the stimulators were on compared to DBs off. this implies that stN stimulation significantly affected the speed and effi-ciency of action reprogramming on the conflict trials.
the effect of stN DBs in PD on conflict resolution is not consistent across existing studies (Frank et al. 2007; Wylie et al. 2010; cavanagh et al. 2011). the main reason for this disparity is that the nature of the conflict faced by partici-pants differs across experimental tasks, as a result of which the optimal strategies for conflict resolution also vary. For example, in the ‘win–win’ trials of the probabilistic deci-sion-making task used by Frank and colleagues (Frank et al. 2007), optimal conflict resolution as shown by controls and PD patients with DBs off was to slow down Rts to take time to reflect before responding. PD patients with DBs on did not show this optimal pattern and were impulsive. On our conditional stop signal task, while DBs was on, the magnitude of conflict-induced slowing was excessive and
significantly greater for the slowest quartiles compared to DBs off. thus, the precise nature of the conflict and the optimal strategies for dealing with it influence the direction of the impact of stN DBs on conflict resolution in PD.
Effect of DBs of the stN on proactive action restraint in PD
Proactive response strategy adjustments become evi-dent when stop signals are expected as a result of which response thresholds are raised to slow down Go Rts and increase the likelihood of stopping (Jahfari et al. 2010; aron et al. 2007; Verbruggen and logan 2009a, b; chikazoe et al. 2009). In our task, ‘anticipatory’ or ‘strategic’ slowing of Go Rts in the context of ‘critical’ Go trials where a propor-tion of the responses would have to be subsequently stopped represents proactive inhibition or action restraint. With DBs turned off, patients could not reach the desired level of pro-active adjustment compared to controls. Given the between-groups nature of this difference and since no significant change of RDE with DBs was observed, the effect of stN DBs on proactive inhibition must be interpreted with cau-tion. the RDE measure of proactive inhibition is likely to mainly reflect adoption of a ‘waiting strategy’ and respond-ing with restraint, which reflect modulation of the speed of the Go response on the ‘critical’ Go trials in anticipation of occurrence of a stop signal. thus, the RDE may be linked to alteration/raising of the response threshold on ‘critical’ versus ‘non-critical’ Go trials, but it could also partly reflect a ‘fine tuning of preparedness to inhibit’.
Differential effects of stN stimulation on reactive and proactive inhibition and conflict resolution are likely mediated through different frontal-basal ganglia pathways
We propose that stN stimulation produced different effects on reactive and proactive inhibition and conflict resolu-tion in our PD patients because these effects are mediated through stN stimulation influencing different fronto-basal ganglia pathways: the hyperdirect, indirect and direct path-ways. this proposal is consistent with emerging evidence differentiating the roles of the stN and striatum in inhibi-tory control, with the stN considered to be particularly per-tinent to ‘hold your horses’ and global reactive inhibition (Frank 2006), whereas proactive and selective inhibitory control may be achieved via the striatum and its direct and indirect connections with the cortex (for review see Zand-belt and Vink 2010; aron 2011). this proposal is consist-ent with imaging (e.g. Jahfari et al. 2011) and tMs and tractography (Neubert et al. 2010) evidence suggesting that response inhibition can be implemented through multiple routes, for example, the hyperdirect and indirect pathways acting together or through cortico-cortical and subcortical

Exp Brain Res
1 3
pathways. however, our interpretation requires examination in an imaging and DBs on versus off or combined tMs and imaging studies. Based on available imaging results with the conditional stop signal task (aron et al. 2007), it is likely that the prolongation of the ssRt observed by us on this task is mediated by the hyperdirect pathways between the pre-sMa and IFc and the stN, which requires direct verifica-tion in future studies. In relation to clinical relevance, future studies also need to investigate if and how these deficits in cognitive and executive control on a range of experimental tasks such as the current conditional stop signal task, the stroop (Jahanshahi et al. 2000), go/no-go Rts (Ballanger et al. 2009; hershey et al. 2004), fast-paced random number generation (thobois et al. 2007) or probabilistic decision-making under conflict (cavanagh et al. 2011; Frank et al. 2007) relate to the development of new impulse control disorders (lim et al. 2009) and increased suicide (Voon et al. 2006) documented following stN DBs surgery in some patients with PD.
Acknowledgments We are grateful to all the participants. this research was supported by a PhD studentship from Fundación caja Madrid, spain (IO), and a Post-doctoral career Development Fellow-ship from Parkinson’s UK (lW).
References
aron aR (2011) From reactive to proactive and selective control: devel-oping a richer model for stopping inappropriate responses. Biol Psychiatry 69(12):e55–e68. doi:10.1016/j.biopsych.2010.07.024
aron aR, Poldrack Ra (2006) cortical and subcortical contributions to stop signal response inhibition: role of the subthalamic nucleus. J Neurosci 26(9):2424–2433
aron aR, Behrens tE, smith s, Frank MJ, Poldrack Ra (2007) tri-angulating a cognitive control network using diffusion-weighted magnetic Resonance imaging (MRI) and functional MRI. J Neu-rosci 27(14):3743–3752
Ballanger B, van Eimeren t, Moro E, lozano aM, hamani c, Boulinguez P, Pellecchia G, houle s, Poon YY, lang aE, strafella aP (2009) stimulation of the subthalamic nucleus and impulsiv-ity: release your horses. ann Neurol 66(6):817–824. doi:10.1002/ana.21795
Baunez c, Nieoullon a, amalric M (1995) In a rat model of Parkin-sonism, lesions of the subthalamic nucleus reverse increases of reaction time but induce a dramatic premature responding deficit. J Neurosci 15(10):6531–6541
Beck at, Ward ch, Mendelson M, Mock J, Erbaugh J (1961) an inven-tory for measuring depression. arch Gen Psychiatry 4:561–571
Brown RG, Dowsey Pl, Brown P, Jahanshahi M, Pollak P, Benabid al, Rodriguez-Oroz Mc, Obeso J, Rothwell Jc (1999) Impact of deep brain stimulation on upper limb akinesia in Parkinson’s disease. ann Neurol 45(4):473–488
cavanagh JF, Wiecki tV, cohen MX, Figueroa cM, samanta J, sherman sJ, Frank MJ (2011) subthalamic nucleus stimulation reverses mediofrontal influence over decision threshold. Nat Neu-rosci. doi:10.1038/nn.2925
chen X, scangos KW, stuphorn V (2010) supplementary motor area exerts proactive and reactive control of arm movements. J Neurosci 30(44):14657–14675. doi:10.1523/JNEUROscI.2669-10.2010
chikazoe J, Jimura K, hirose s, Yamashita K, Miyashita Y, Konishi s (2009) Preparation to inhibit a response complements response inhibition during performance of a stop-signal task. J Neu-rosci 29(50):15870–15877. doi:10.1523/JNEUROscI.3645- 09.2009
De Jong R, coles MG, logan GD (1995) strategies and mechanisms in nonselective and selective inhibitory motor control. J Exp Psy-chol hum Percept Perform 21(3):498–511
Eagle DM, Baunez c, hutcheson DM, lehmann O, shah aP, Rob-bins tW (2008) stop-signal reaction-time task performance: role of prefrontal cortex and subthalamic nucleus. cereb cortex 18(1):178–188
Fahn s, Elton Rl, Members of the UPDRs Development committee (1987) Unified Parkinson’s disease rating scale. In: Fahn s, Mars-den cD, Goldstein M, clane DB (eds) Recent developments in Parkinson’s disease. Macmillan healthcare Information, Florham Park, pp 153–163
Folstein MF, Folstein sE, Mchugh PR (1975) “Mini-mental state”. a practical method for grading the cognitive state of patients for the clinician. J Psychiatr Res 12(3):189–198
Frank MJ (2006) hold your horses: a dynamic computational role for the subthalamic nucleus in decision making. Neural Netw 19(8):1120–1136
Frank MJ, samanta J, Moustafa aa, sherman sJ (2007) hold your horses: impulsivity, deep brain stimulation, and medication in Parkinsonism. science 318(5854):1309–1312. doi:10.1126/science.1146157
hershey t, Revilla FJ, Wernle a, Gibson Ps, Dowling Jl, Perlmutter Js (2004) stimulation of stN impairs aspects of cognitive control in PD. Neurology 62(7):1110–1114
hershey t, campbell Mc, Videen tO, lugar hM, Weaver PM, hartlein J, Karimi M, tabbal sD, Perlmutter Js (2010) Mapping Go-No-Go performance within the subthalamic nucleus region. Brain. doi:10.1093/brain/awq256
hughes aJ, Daniel sE, Kilford l, lees aJ (1992) accuracy of clini-cal diagnosis of idiopathic Parkinson’s disease: a clinico-patho-logical study of 100 cases. J Neurol Neurosurg Psychiatry 55(3): 181–184
hutchison WD, allan RJ, Opitz h, levy R, Dostrovsky JO, lang aE, lozano aM (1998) Neurophysiological identification of the sub-thalamic nucleus in surgery for Parkinson’s disease. ann Neurol 44(4):622–628. doi:10.1002/ana.410440407
Jahanshahi M, ardouin cM, Brown RG, Rothwell Jc, Obeso J, alba-nese a, Rodriguez-Oroz Mc, Moro E, Benabid al, Pollak P, limousin-Dowsey P (2000) the impact of deep brain stimula-tion on executive function in Parkinson’s disease. Brain 123(Pt 6):1142–1154
Jahfari s, stinear cM, claffey M, Verbruggen F, aron aR (2010) Responding with restraint: what are the neurocognitive mech-anisms? J cogn Neurosci 22(7):1479–1492. doi:10.1162/jocn.2009.21307
Jahfari s, Waldorp l, van den Wildenberg WP, scholte hs, Ridderink-hof KR, Forstmann BU (2011) Effective connectivity reveals important roles for both the hyperdirect (fronto-subthalamic) and the indirect (fronto-striatal-pallidal) fronto-basal ganglia path-ways during response inhibition. J Neurosci 31(18):6891–6899. doi:10.1523/JNEUROscI.5253-10.2011
Kuhn aa, tsui a, aziz t, Ray N, Brucke c, Kupsch a, schneider Gh, Brown P (2009) Pathological synchronisation in the sub-thalamic nucleus of patients with Parkinson’s disease relates to both bradykinesia and rigidity. Exp Neurol 215(2):380–387. doi:10.1016/j.expneurol.2008.11.008
li cs, huang c, constable Rt, sinha R (2006) Imaging response inhi-bition in a stop-signal task: neural correlates independent of signal monitoring and post-response processing. J Neurosci 26(1):186–192. doi:10.1523/JNEUROscI.3741-05.2006

Exp Brain Res
1 3
li cs, Yan P, sinha R, lee tW (2008) subcortical processes of motor response inhibition during a stop signal task. Neuroimage 41(4):1352–1363. doi:10.1016/j.neuroimage.2008.04.023
lim sY, O’sullivan ss, Kotschet K, Gallagher Da, lacey c, law-rence aD, lees aJ, O’sullivan DJ, Peppard RF, Rodrigues JP, schrag a, silberstein P, tisch s, Evans ah (2009) Dopamine dysregulation syndrome, impulse control disorders and punding after deep brain stimulation surgery for Parkinson’s disease. J clin Neurosci 16(9):1148–1152. doi:10.1016/j.jocn.2008.12.010
logan GD, cowan WB (1984) On the ability to inhibit thought and action: a theory of an act of control. Psychol Rev 91(3):295–327
Mirabella G, Iaconelli s, Romanelli P, Modugno N, lena F, Manfredi M, cantore G (2011) Deep brain stimulation of subthalamic nuclei affects arm response inhibition in Parkinson’s patients. cereb cor-tex. doi:10.1093/cercor/bhr187
Neubert FX, Mars RB, Buch ER, Olivier E, Rushworth MF (2010) cortical and subcortical interactions during action reprogramming and their related white matter pathways. Proc Natl acad sci Usa 107(30):13240–13245. doi:10.1073/pnas.1000674107
Obeso Ja, Guridi J, De long M (1997) surgery for Parkinson’s dis-ease. J Neurol Neurosurg Psychiatry 62:2–8
Obeso I, Wilkinson l, casabona E, Bringas Ml, alvarez M, alvarez l, Pavon N, Rodriguez-Oroz Mc, Macias R, Obeso Ja, Jahan-shahi M (2011a) Deficits in inhibitory control and conflict reso-lution on cognitive and motor tasks in Parkinson’s disease. Exp Brain Res 212(3):371–384. doi:10.1007/s00221-011-2736-6
Obeso I, Wilkinson l, Jahanshahi M (2011b) levodopa medication does not influence motor inhibition or conflict resolution in a con-ditional stop-signal task in Parkinson’s disease. Exp Brain Res 213(4):435–445. doi:10.1007/s00221-011-2793-x
Ray NJ, Jenkinson N, Brittain J, holland P, Joint c, Nandi D, Bain PG, Yousif N, Green a, stein Js, aziz tZ (2009) the role of the subthalamic nucleus in response inhibition: evidence from deep brain stimulation for Parkinson’s disease. Neuropsychologia 47(13):2828–2834. doi:10.1016/j.neuropsychologia.2009.06.011
Rodriguez-Oroz Mc, Obeso Ja, lang aE, houeto J-l, Pollak P, Rehn-crona s, Kulisevsky J, albanese a, Volkmann J, hariz MI, Quinn NP, speelman JD, Guridi J, Zamarbide I, Gironell a, Molet J, Pas-cual-sedano B, Pidoux B, Bonnet aM, agid Y, Xie J, Benabid al, lozano aM, saint-cyr J, Romito l, contarino MF, scerrati M, Fraix V, Van Blercom N (2005) Bilateral deep brain stimulation in Parkinson’s disease: a multicentre study with 4 years follow-up. Brain 128:2240–2249
Rubia K, smith aB, Brammer MJ, taylor E (2003) Right inferior prefrontal cortex mediates response inhibition while mesial pre-frontal cortex is responsible for error detection. Neuroimage 20(1):351–358
swann N, Poizner h, houser M, Gould s, Greenhouse I, cai W, strunk J, George J, aron aR (2011) Deep brain stimulation of the sub-thalamic nucleus alters the cortical profile of response inhibition in the Beta frequency band: a scalp EEG study in Parkinson’s disease. J Neurosci 31(15):5721–5729. doi:10.1523/JNEUROscI.6135-10.2011
thobois s, hotton GR, Pinto s, Wilkinson l, limousin-Dowsey P, Brooks DJ, Jahanshahi M (2007) stN stimulation alters
pallidal-frontal coupling during response selection under compe-tition. J cereb Blood Flow Metab 27(6):1173–1184. doi:10.1038/sj.jcbfm.9600425
van den Wildenberg WP, van Boxtel GJ, van der Molen MW, Bosch Da, speelman JD, Brunia ch (2006) stimulation of the sub-thalamic region facilitates the selection and inhibition of motor responses in Parkinson’s disease. J cogn Neurosci 18(4):626–636
Verbruggen F, logan GD (2009a) automaticity of cognitive control: goal priming in response-inhibition paradigms. J Exp Psychol learn Mem cogn 35(5):1381–1388. doi:10.1037/a0016645
Verbruggen F, logan GD (2009b) Proactive adjustments of response strategies in the stop-signal paradigm. J Exp Psychol hum Percept Perform 35(3):835–854. doi:10.1037/a0012726
Verbruggen F, aron aR, stevens Ma, chambers cD (2010) theta burst stimulation dissociates attention and action updating in human inferior frontal cortex. Proc Natl acad sci Usa 107(31):13966–13971. doi:10.1073/pnas.1001957107
Verbruggen F, chambers cD, logan GD (2013) Fictitious inhibitory differences: how skewness and slowing distort the estimation of stopping latencies. Psychol sci [Epub ahead of print]
Vink M, Kahn Rs, Raemaekers M, van den heuvel M, Boersma M, Ramsey NF (2005) Function of striatum beyond inhibition and execution of motor responses. hum Brain Mapp 25(3):336–344. doi:10.1002/hbm.20111
Voon V, Kubu c, Krack P, houeto Jl, troster aI (2006) Deep brain stimulation: neuropsychological and neuropsychiatric issues. Mov Disord 21(suppl 14):s305–s327. doi:10.1002/mds.20963
Wichmann t, Bergman h, Delong MR (1994) the primate subtha-lamic nucleus. III. changes in motor behavior and neuronal activ-ity in the internal pallidum induced by subthalamic inactivation in the MPtP model of parkinsonism. J Neurophysiol 72(2):521–530
Wiener M, Magaro cM, Matell Ms (2008) accurate timing but increased impulsivity following excitotoxic lesions of the sub-thalamic nucleus. Neurosci lett 440(2):176–180. doi:10.1016/ j.neulet.2008.05.071
Witt K, Pulkowski U, herzog J, lorenz D, hamel W, Deuschl G, Krack P (2004) Deep brain stimulation of the subthalamic nucleus improves cognitive flexibility but impairs response inhibition in Parkinson disease. arch Neurol 61(5):697–700. doi:10.1001/archneur.61.5.697
Wylie sa, Ridderinkhof KR, Elias WJ, Frysinger Rc, Bashore tR, Downs KE, van Wouwe Nc, van den Wildenberg WP (2010) sub-thalamic nucleus stimulation influences expression and suppres-sion of impulsive behaviour in Parkinson’s disease. Brain 133(Pt 12):3611–3624. doi:10.1093/brain/awq239
Yugeta a, terao Y, Fukuda h, hikosaka O, Yokochi F, Okiyama R, taniguchi M, takahashi h, hamada I, hanajima R, Ugawa Y (2010) Effects of stN stimulation on the initiation and inhibition of saccade in Parkinson disease. Neurology 74(9):743–748. doi:10.1212/WNl.0b013e3181d31e0b
Zandbelt BB, Vink M (2010) On the role of the striatum in response inhibition. Plos ONE 5(11):e13848. doi:10.1371/journal.pone.0013848





























