Embed Size (px)
Citation preview

Brain Research, 224 (1981) 1-29 1 Elsevier/North-Holland Biomedical Press
Research Reports
C O N N E C T I O N S OF THE SUBTHALAMIC NUCLEUS IN THE M O N K E Y
MALCOLM B. CARPENTER, STEVEN C. CARLETON, JEFFREY T. KELLER* and PATRICIA CONTE
Department of Anatomy, School of Medicine, Uniformed Services University of the Health Sciences, Bethesda, MD 20014 (U.S.A.)
(Accepted March 12th, 1981)
Key words: subthalamic nucleus - - globus pallidus - - substantia nigra - - pedunculopontine nucleus
SUMMARY
Attempts were made to determine the afferent and efferent connections of the
subthalamic nucleus (STN) in the monkey using retrograde and anterograde axo- plasmic transport technics. Following HRP injections limited to the STN, label was transported to arrays of cells adjacent, and parallel, to the lateral medullary lamina in
the rostral two thirds of the lateral pallidal segment (LPS). Only sparse label was transported to cells of the pedunculopontine nucleus (PPN) and the locus ceruleus (LC). No enzyme was transported across the midline, or to the striatum, medial
pallidal segment (MPS), thalamus, substantia nigra (SN) or dorsal nucleus of the raphe (DNR).
HRP injected into portions of both the STN and SN produced retrograde transport of the enzyme to cells in parallel arrays in the LPS related rostrocaudally to
the injection site. Additional enzyme transport was seen in cells of the striatum, the D N R and the PPN. Only a few isolated cells were labeled in the sensorimotor cortex.
Efferent connections of the STN were studied in monkeys in which [3H]amino acids were injected hydraulically or iontophoretically into the STN. Isotope traced in
serial autoradiographs was distributed to: (1) both segments of the globus pallidus (GP) in arrays parallel to the medullary laminae, and (2) the pars reticulata of SN (SNpr). The greatest number of terminals was found in the MPS. Fibers from the rostral part of the STN descended along the dorsal border of the SN and projected ventrally to terminations in the SNpr. No isotope was transported across the midline, or to the striatum, thalamus, D N R or PPN. Isotope injected into both the STN and SN produced similar transport to the GP and transport via nigral efferent fibers to : (1)
* Postdoctoral Fellow in Anatomy on leave from the Mayfield Neurological Institute and the College of Medicine, University of Cincinnati.
0006-8993/81/0000-0000/$ 02.50 © Elsevier/North-Holland Biomedical Press

portions of the striatum, (2) specific thalamic nuclei (VAmc, VLm, DMpl), (3) deep and middle gray layers of the superior colliculus and (4) PPN. Control studies indicated that [~H]amino acids injected only into the SN were transported to PPN. HRP injected into PPN produced profuse retrograde transport in cells of the MPS and SNpr and distinct label in a few cells of the zona incerta and STN.
These data suggest that the STN receives its major subcortical input from cells of the LPS arranged in arrays which have a rostrocaudal organization. No cells of the MPS or SN project to the STN. The output of the STN is to both segments of the GP and SNpr. Major subcortical projections to PPN arise from the MPS and SNpr, but afferents also arise from other sources. The major projection of PPN is to SN.
INTRODUCTION
The deep location and small size of the subthalamic nucleus (STN) have rendered determination of its connections singularly difficult. Although degeneration studies of projections of the nucleus were complicated by interruption of fibers of passage, they provided evidence of efferent projections to both the globus pallidus (GP) and the substantia nigra (SN)8,26, 54. Autoradiographic studies indicate that subthalamopallidal fibers are topographically organized and arranged in arrays parallel to the medullary laminae 40. Retrograde transport studies provide details concerning the topographic projections of STN neurons and suggest that: (1) cells in medial and caudal regions of the nucleus project to the medial pallidal segment (M PS), (2) cells in the medial part of the middle third of the nucleus project to the rostral division of the lateral pallidal segment (LPS) and (3) cells in central portions of the rostral two-thirds of the nucleus project to the central division of the LPS 3. This investigation indicated an inverse dorsoventral topographical relationship between cells in the STN and axonal terminations in the LPS. A smaller, but substantial, projection from the STN to the SN has been described by several authors 13,~4,a5,4°,47. Most authors consider these fibers to terminate chiefly in the pars reticulata. A fluorescent retrograde double labeling study in the rat suggested that virtually all (94 ~o) cells in the STN give rise to collaterals that supply both the globus patlidus and the substantia nigra 27. Although there are suggestions of sparse projections from the STN to various thalamic nuclei, the putamen and the pedunculopontine nucleus, none of these appears to be established 4°,47.
Major inputs to the subthalamic nucleus arise from the frontal cortex and the lateral pallidal segment. The most massive input to the subthalamic nucleus (STN), derived from the lateral pallidal segment (LPS), is topographically organized with rostral parts of LPS projecting to medial regions, and central portions of the LPS supplying lateral parts of the nucleus 3,2~,~5,49. Few cells in caudal parts of the LPS appeared to project to STN and no fibers from the medial pallidal segment (MPS) terminated in the subthalamic nucleus 3,25,49. Autoradiographic studies have demon- strated corticofugal projections from the precentral motor cortex and Brodmann's areas 6, 8 and 919,28,29. Projections from the precentral motor cortex, described as somototopically organized, terminated mainly in lateral regions of STN. Less

substantial cortical efferents from premotor and prefrontal areas ended in more medial parts of the nucleus.
Relatively small projections to STN have been described from the pedunculo- pontine nucleus (PPN), the dorsal nucleus of the raphe (DNR) and the locus ceruleus ~5,43,4~. Autoradiographic studies suggest the projection from PPN to the STN may be the most substantial, but this nucleus also distributes fibers to other structures16, 43.
The purpose of the present study was to re-investigate the connections of the subthalamic nucleus in the monkey using axoplasmic transport technics to determine the pathways by which this nucleus contributes to motor function.
METHODS
In a series of squirrel monkeys (Sairniri sciureus) with weights ranging from 750 to 1000 g attempts were made to inject small volumes of horseradish peroxidase (HRP) into the STN. Coordinates were determined from dissections of formalin fixed heads with brains in situ 9. A stereotaxic approach was used in which a micropipette with an inside diameter of 15-20/~m was introduced in a vertical plane. The micropipette first was filled with mineral oil stained with a red dye. The metal plunger from a 10-#m Hamilton syringe was inserted into the shank of the micropipette for a sufficient distance to express some of the mineral oil. One or two /~1 of 5 - 1 0 ~ HRP (Worthington) was drawn into the tip of the micropipette by suction. The micropipette was attached to a calibrated stereotaxic holder, and the tip of the pipette was immersed in distilled water until used. Immediately before inserting the pipette into the brain, a small volume of HRP was ejected into a small plastic cup containing distilled water. Using a calibrated micrometer advance 0.I0-0.15 #1 of HRP were injected into the target site in a period of 10-15 min and the pipette was left in place thereafter for 10-15 rain. When the pipette was withdrawn from the brain, the patency of the tip was tested again.
After post-operative survivals of 48 h, animals were anesthetized with Nembutal and the heart was injected with 500 units of heparin and 0.5 ml of 1 ~ sodium nitrite. Animals were perfused via the left ventricle of the heart with a liter of normal saline followed by 2 1 of a fixative solution containing 1.25 ~ paraformaldehyde and 2.5 jo~ glutaraldehyde in a 0.1 M phosphate buffer (pH 7.4). Brains were immediately removed from the skull, sectioned into blocks either perpendicular to the axis of the brain stem, or in parasagittal planes, and tissue blocks were immersed in fixative overnight at 4 °C. The following day the tissue blocks were transferred to fresh 0. ! M phosphate buffer containing 30 ~ sucrose and stored at 4 °C for another 24 h.
Blocks of tissue containing the corpus striatum, diencephalon, mesencephalon and pons were cut serially at 40/~m on a freezing microtome and collected in groups of 5 or 6 sections in compartments of plastic containers filled with distilled water. Blocks of sensorimotor cortex were cut serially perpendicular to the pial surface and treated in the same manner. Selected sections were treated by the Graham and Karnovsky 1~ method to produce the reaction for the identification of HRP activity 34. Reacted

A
B
C ~

sections were mounted on slides with gelatin and lightly stained with cresyl violet. A larger number of selected sections from each brain were reacted with tetramethyl-
benzidine (TMB), mounted on slides and lightly stained with neutral red 36. Except for one animal done early in the study, the tetramethylbenzidine (TMB) reaction was the
principal one used because of its greater sensitivity. In a series of rhesus and cynomologus monkeys ranging in weight from 2.8 to 3.5
kg, attempts were made to inject iontophoretically tritiated amino acids into the STN. Tritiated proline or leucine, either alone or combined in equal parts, were concen- trated to 40/~Ci/~l and drawn into the tip, shank and distal part of the stem of the
micropipette by capillary action. A fine silver wire with the insulation stripped from the tip was inserted into the amino acid of the pipette stem. The pipette was mounted
on a calibrated carrier and advanced to the target site. The lead wires from the pipette and an indifferent electrode (attached to muscle) were connected to a high voltage
constant-current generator (Midgard Electronics, Model C-3). The driving force was
an anodal current of 0.1-0.20 /,A applied either continuously for 15 min or pulsed (7 s on/offtime) for 30 rain. At the termination of the period of iontophoresis, the polarity of the current was reversed and the pipette was withdrawn from the brain. In one
monkey attempts were made to inject cells of the subthalamic nucleus using a l-,ul
Hamilton syringe. Following post-operative survivals of 5 8 days animals were anesthetized and
perfused via the left ventricle of the heart with a liter of normal saline followed by a liter of 10 °. o neutral formalin. Brains were immediately removed, further fixed in l0 ",/, neutral formalin and cut sagitally, or transversely, to the axis of the brain stem. Blocks of tissue were washed, dehydrated, embedded in paraffin and cut serially at 15 #m.
Every tenth section was mounted. Sections through the injection site were stained with cresyl violet for evaluation. Selected sections from the pons and all more rostral levels
of the neuraxis were deparaffinized, dipped in Kodak NTB-2 nuclear emulsion, and air dried le. Dipped sections were packed in light-tight black plastic boxes with a
dehydrating agent and stored at 4 "C for exposure periods of 10-12 weeks. Exposed sections were developed in fresh Kodak D-19 developer at 15 °C for 3 rain, fixed, washed and lightly stained with cresyl violet. Selected autoradiographs were studied by bright and dark field microscopy.
RESULTS
Subthalamic nucleus afferents Attempts to determine the sources of input to the subthalamic nucleus (STN)
were made in a series of 15 squirrel monkeys by studying the retrograde transport of horseradish peroxidase (HRP) from injections of the nucleus. Because all evidence
Fig. 1. Monkey U-90. Projection drawings of parasagittal sections of the corpus striatum and diencephalon [arranged lateral (A) to medial (C)] demonstrating transport of HRP from an injection in the subthalamic nucleus (STN, B). Retrogradely labeled ceils (coarse black dots) were found in parallel arrays in the LPS (A, B), while no labeled cells were seen in the MPS (A, B, C). Regions of the STN not involved by the injection were filled with labeled cells (A, C). A few retrogradely labeled cells were found in the nucleus of the ansa lenticularis (dorsal to OT, in B, C) and in the pedunculopontine nucleus (PPN, C). See list of abbreviations and Fig. 2A-F.

6
suggests that projections to the STN are ipsilateral, injections were made bilaterally and each side was considered separately 3,4,19,z5,3°,35,42,43,49. Even though similar small volumes of HRP were injected on each side, the size of the injection sites showed considerable variation. The structure most commonly injected in addition to the STN was the rostral part of the SN. Attempts to selectively inject the STN were successful in 3 monkeys. HRP injections in 4 monkeys involved portions of both the STN and the SN on 6 sides.
Subthalamic nucleus
In monkey U-90 sagittal sections of the brain revealed a pipette track which terminated in central parts of the STN and evidence of enzyme diffusion throughout the neuropil of the nucleus (Figs. 1 and 2A). Although cells in all parts of the nucleus contained HRP granules, uptake of the marker was appreciably greater in cells in the medial half of the nucleus (Figs. t and 2B, C). No HRP diffused into the SN, although a few scattered neurons contained label in the rostral pole of the SN.
Retrograde transport of HRP to the LPS was impressive and exhibited a definite pattern (Figs. 1 and 2D, E, F). Large numbers of deeply labeled neurons were found in the rostral division of the LPS (i.e. that portion rostral to the medial medullary lamina of the pallidum) arranged in a band close to, and parallel with, the lateral medullary lamina. In rostral regions ofthe LPS the largest number of labeled cells were seen dorsolaterally; only a few clusters of labeled cells were found in medial regions of the LPS. In the central division of the LPS (i.e. that portion of the LPS flanking the lateral border of the MPS) the band of labeled cells adjacent to the lateral medullary lamina became narrower and the most heavily labeled cells were seen ventrally. Caudal regions of the LPS contained only a few labeled cells adjacent to the lateral medullary lamina. Thus, labeled neurons formed a band adjacent to the lateral medullary lamina in the rostral two-thirds of the LPS which was broad rostrally and narrow caudally. No labeled neurons were seen in the striatum, the MPS or in the thalamus, except in
the immediate vicinity of the pipette track. A cluster of labeled large cells was seen rostral to the peduncular part of the
internal capsule in the area of the nucleus of the ansa lenticularis 48 (Fig. 1). These medially located cells appeared in places to be continuous with a few labeled cells in the ventral tegmental area. One or two labeled cells were identified in the ipsilateral pedunculopontine nucleus (PPN), and the locus ceruleus (LC). No transport of HRP was seen to cells in more caudal regions of the brain stem.
In two other animals HRP injections involved small portions of the rostral third of the STN. Both injections were in the center of the nucleus. The pipette track in monkey U-81 (R) traversed and traumatized some fibers in the lenticular fasciculus. Although the HRP was confined to avery small part of the STN, cells in a much larger region of the nucleus contained HRP granules. The darkest granules were seen in cells in the medial half of the nucleus; cells in the caudal third of the nucleus and in all lateral regions contained only a few HRP granules. The injection in the STN in monkey U-87 (R) showed no evidence of diffusion into other parts of the nucleus and there was no uptake of the enzyme by cells in any region except that immediately
adjacent to the injection (Figs. 3 and 4B).

Fig. 2. Squirrel Monkey U-90. A: dark field photomicrograph of a parasagittal section demonstrating an HRP injection in the subthalamic nucleus (see Fig. 1). B: bright field photomicrograph of a parasagittal section of the subthalamic nucleus (STN) medial to the injection site showing label in virtually all cells. C: bright field photomicrograph of the STN lateral to the injection site. D: dark field photomicrograph of a parasagittal section with retrogradely labeled neurons in the lateral pallidal segment (LPS). E and F: bright field and dark field photomicrographs of labeled cells in the LPS. A-E: tetramethylbenzidine reaction, counterstained with neutral red; magnification: A, q 6; B, 25; c, I’ 100; D, x 18; E, x 100. F: diaminobenzidine reaction counterstained with cresyl violet: magnification: F, Y 63.
\
/ .' ..: , .... ~,, ~" ~,, "~ '~
/! ~ !'
J
/ i
' \
\ t
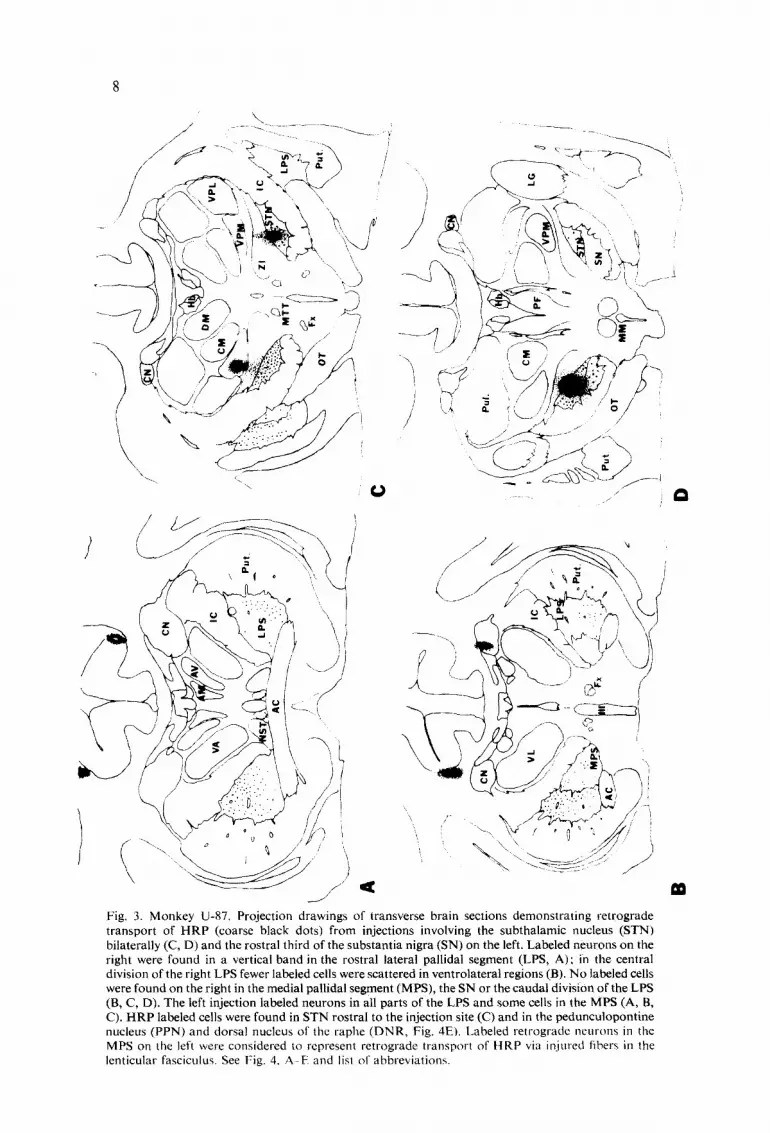
Fig. 3. Monkey U-87. Projection drawings of transverse brain sections demonstrating retrograde transport of HRP (coarse black dots) from injections involving the subthalamic nucleus (STN) bilaterally (C, D) and the rostral third of the substantia nigra (SN) on the left. Labeled neurons on the right were found in a vertical band in the rostral lateral pallidal segment (LPS, A); in the central division of the right LPS fewer labeled cells were scattered in ventrolateral regions (B). No labeled cells were found on the right in the medial pallidal segment (MPS), the SN or the caudal division of the LPS (B, C, D). The left injection labeled neurons in all parts of the LPS and some cells in the MPS (A, B, C). HRP labeled cells were found in STN rostral to the injection site (C) and in the pedunculopontine nucleus (PPN) and dorsal nucleus of the raphe (DNR, Fig. 4El. Labeled retrograde neurons in the MPS on the left were considered to represent retrograde transport of HRP via injured fibers ill the lenticular fasciculus. See Fig. 4, A E and list of abbreviations.
m

Fig. 4. Squirrel monkeys U-87 (A D) and U-81 (E, F) A and B: dark field photomicrographs of HRP injections in the left and right subthalamic nucleus (see Fig. 3). C and D: bright field and dark field photomicrographs of retrogradely labeled neurons in the right (C) and left (D) lateral pallidal segments. E: dark field photomicrograph of a caudal midbrain section showing retrograde transport of HRP to cells in the peduncu[opontine nucleus (PPN) and the dorsal nucleus of the raphe. The injection in U-81 (L) like that in U-87 (L) labeled portions of both the subthalamic nucleus and the substantia nigra. F: bright field photomicrograph of neurons in PPN retrogradely labeled with HRP. A, B: diaminobenzidine reaction, counterstained with cresyl violet; magnification: A, B, ~'< 12. C-F: tetramethylbenzidine reaction, counterstained with neutral red; magnification: C, D, :< 63; E, ~: 18; F, × 63.

10
Even though only small portions of the STN were injected in these animals, HRP was transported to cells in the LPS in a pattern similar to that described for a larger injection (U-90). Labeled neurons were found in a lateral band in the rostral division of the LPS in monkey U-87 (R) but were not as numerous as in U-90 (Figs. 3 and 4D). The central division of the LPS in this animal contained only a small cluster of labeled neurons in ventral regions near the lateral medullary lamina. No labeled cells were seen in the caudal half of the LPS. The smaller injection in the rostral part of the STN in monkey U-81 (R) reproduced retrograde labeling of only small clusters of neurons in ventrolateral region of the rostral half of the LPS. In this animal a few welt-labeled cells were seen in scattered sites in the MPS, presumably due to HRP transport via injured fibers of the lenticular fasciculus. In both animals only a few retrogradely labeled cells were seen in the PPN and in the LC. No HRP was transported to cells in the striatum, thalamus, substantia nigra or the dorsal nucleus of the raphe.
Subthalamic nucleus and substantia nigra Attempts to selectively inject small volumes of HRP into the STN in 4 monkeys
resulted in concomitant involvement of portions of the STN and rostral regions of the SN (Figs. 3, 4A and 5). Six such injections were produced in 4 monkeys. The precise limits of the injection in the STN were difficult to determine, but judging from the pipette track it included at least the caudal third of the nucleus. The HRP delivered to the SN involved the rostral third of the nucleus on three sides in two animals [U-6(B) and U-87(L)] and the rostral half of the nucleus in three instances [U-81(L) and U- 92(B)]. HRP :injections in U-92 labeled similar parts of the STN and the SN on both sides (Figs. 5 and 6A). Bilateral HRP injections in U-6 involved larger portions of the STN and SN on the right. Even though these injections of the STN were partial, cells in all parts of the nucleus were heavily labeled (Figs. 5 and 6B). Unusually profuse label appeared to reflect direct diffusion of the marker and retrograde transport from terminals in the substantia nigra 13,'4,35,4°,47. These cirumstances made it difficult to
define the limits of the injection in the STN. The pattern of retrograde transport of HRP to neurons in the LPS
differed among animals of this group. In monkey U-92 the number of labeled neurons and the extent of their distribution in the LPS was greater than in any animal in this series (Figs. 5 and 6C). Well-labeled neurons were found in all parts of the LPS. In the central division of the LPS, the largest and most completely labeled cells were in dorsal and ventral regions and this pattern persisted into the caudal division of the LPS (i.e. that part of the LPS caudal to the medial medullary lamina). A similar pattern of HRP cells in the LPS was seen in monkey U-87(L), although the number of labeled cells was
less (Figs. 3 and 4C). In two monkeys [U-6(L) and U-81(L)] labeled neurons formed a band adjacent to the lateral medullary lamina in the rostral two-thirds of the LPS, as previously described. Even though the HRP injection in the right STN in monkey U-6 appeared greater than that on the left STN, retrograde transport of the marker to the LPS was less. Labeled cells in the rostral and central divisions of the right LPS were largely confined to ventral regions near the lateral medullary lamina.
t / sii'.Y . / "
C
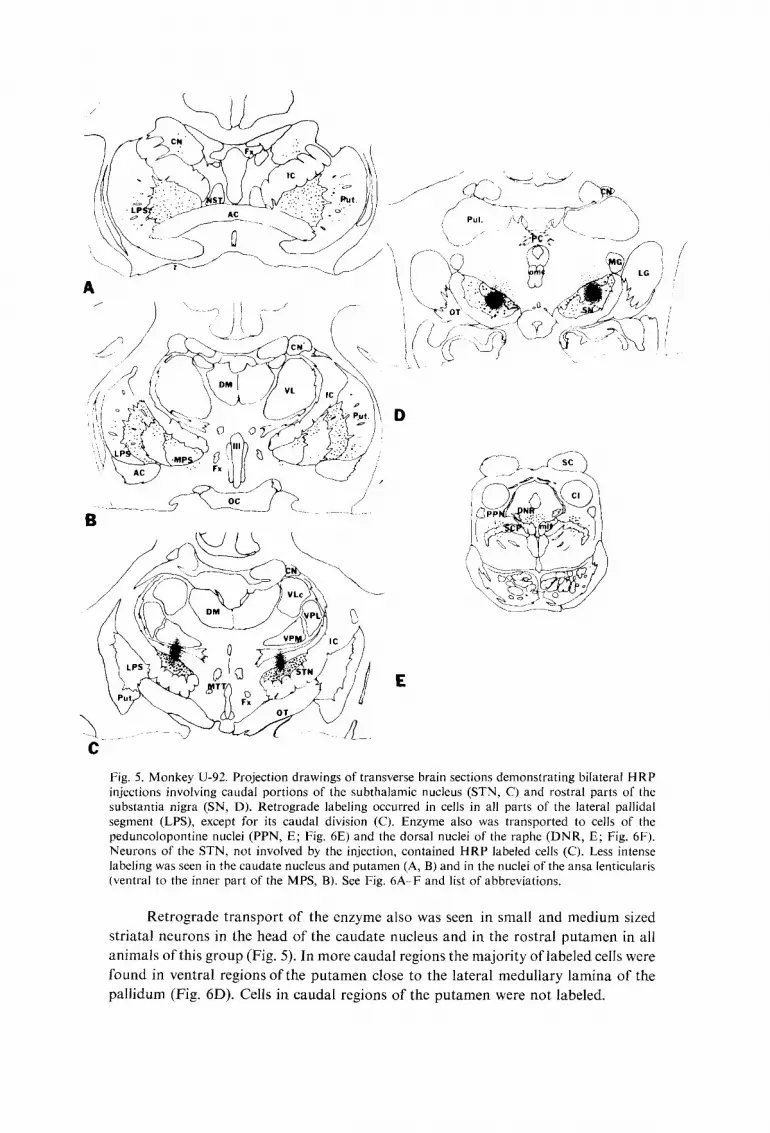
Fig. 5. Monkey U-92. Projection drawings of transverse brain sections demonstrating bilateral HRP injections involving caudal portions of the subthalamic nucleus (STN, C) and rostral parts of the substantia nigra (SN, D). Retrograde labeling occurred in cells in all parts of the lateral pallidal segment (LPS), except for its caudal division (C). Enzyme also was transported to cells of the peduncolopontine nuclei (PPN, E; Fig. 6E) and the dorsal nuclei of the raphe (DNR, E; Fig. 6F). Neurons of the STN, not involved by the injection, contained HRP labeled cells (C). Less intense labeling was seen in the caudate nucleus and putamen (A, B) arid in the nuclei of the ansa lenticu[aris (ventral to the inner part of the MPS, B). See Fig. 6A-F and list of abbreviations.
Retrograde t ranspor t of the enzyme also was seen in small and medium sized
striatal neurons in the head of the caudate nucleus and in the rostral pu tamen in all
animals of this group (Fig. 5). In more caudal regions the majori ty of labeled cells were
found in ventral regions & t h e pu tamen close to the lateral medullary lamina of the
pal l idum (Fig. 6D). Cells in caudal regions of the pu tamen were not labeled.

Fig. 6. Squirrel monkey U-92. A: dark field photomicrograph showing the HRP injection in the caudal part of the subthalamic nucleus (STN) ; this injection also labeled part of the substantia nigra (see Fig. 5). B: bright field photomicrograph through the rostral third of the STN demonstrating extensively labeled neurons, C: dark field photomicrograph of retrogradely labeled neurons in the lateral pallidal segment. D: dark field photomicrograph of a single HRP labeled neuron in the putamen. E and F: dark field photomicrographs of retrogradely labeled neurons in the pedunculo- pontine nucleus (E) and the dorsal nuclei of the raphe (F). A -F: tetramethyl benzidine reaction, counterstained with neutral red; magnifications: A, ~ 12; B, 20; C, 63, D, 160: E, 25 : F, i,,~ 20.

13
HRP deposited in the SN labeled cells in both the pars compacta and pars reticulata. Cells of the pars compacta exhibited profuse uptake throughout most of their rostrocaudal extent of the nucleus. The extensive labeling of cells in the pars compacta was in part primary, in part via dendrites extending into the pars reticulata and in part probably due to retrograde transport via injured ascending axons.
HRP was transported in retrograde fashion to cells of the ipsilateral PPN and the DNR in all animals of this group (Figs. 4E, F, 5 and 6E, F). Labeling &cells in the pars compacta of PPN was profuse and consistent. Only one or two well-labeled cells were seen in each section containing the locus coeruleus. Sections through the cerebral cortex in animals in all groups with HRP injections in the STN revealed sparse retrograde transport of the enzyme to cells in the sensorimotor region.
Subthalamic nucleus q~erents Attempts to determine the efferent projections of the STN were made in a series
of rhesus and cynomologus monkeys. All injections were unilateral. Injections of the STN were suitable for detailed analysis in 2 monkeys; in a third animal portions of both the STN and SN were injected. Comparisons of the isotope transport in the third animal revealed similarities and some important differences.
Subthalamic nucleus Injected tritiated amino acids involved the rostral two-thirds of the STN in two
monkeys, although most of the isotope was in the rostral half of the nucleus. The medial part of the STN near its dorsal capsule was involved in rhesus U-41 and some isotope diffused into the lenticular fasciculus and the zona incerta (Figs. 7 and 9B). Caudal portions of the STN contained only a few silver grains and no part of the SN contained injected isotope. The injection in rhesus U-35 involved the lateral two-thirds of the STN and infiltrated the internal capsule ventral to the nucleus (Figs. 8 and 9A). Although silver grains were present in the rostral part of the SN, they were considered to represent isotope transported from the STN, since no evidence of radioactive transport was seen in the striatum, thalamus or superior colliculus.
Tritiated amino acids deposited in the rostral half of the STN were transported to both segments of the globus pallidus and to the SN, mainly in the pars reticulata. No label was transported across the midline to the opposite side at any site.
Isotope from the STN was transported to all regions of the MPS (Figs. 7 and 8). Fibers from dorsal and lateral regions of the STN arched laterally and ventrally through the internal capsule into dorsal regions of the MPS, while those from medial and ventral parts of the nucleus passed into ventral regions of the M PS. Silver grains were particularly dense in the apical region of the MPS caudally; further rostrally they were present in both sides of the accessory medullary lamina. At levels through the optic chiasm the bulk of the labeled fibers and terminals were concentrated laterally in the M PS. Groups of fibers ascending from the ventral border of the pallidum projected dorsally parallel with the accessory and medial medullary laminae. Differences in the locations of silver grains in the outer part of the MPS in these two animals could be correlated with the area of maximal primary labeling in the STN. In rhesus U-41 silver

14
\ -
i
C
J
• g .
- . ~ , . , s ~ ' ~ ' X j
-- \ j
r , - r
. " W - . _ j - z c 5 , - ~ ~ .
Fig. 7. Monkey U-41. Projection drawings of transverse autoradiographs showing transport of [ZH] amino acids (small black dots) from an injection in the dorsomedial part of the subthalamic nucleus (STN, C). Labeled axonal terminals formed well-defined vertical laminae in the rostral two-thirds of both pallidal segments (A, B). [3HI label in the caudal third of the MPS was diffuse, while no transport was evident in caudal portions of the LPS (C). Descending subthalamonigral fibers formed a distinct bundle on the dorsal surface of the SN (D), before projecting ventrally to terminations in the pars reticulata (D). See Fig. 9B, D, E, F and list of abbreviations.

15
J
/
a
Fig. 8. Monkey U-35. Projection drawings of transverse autoradiographs demonstrating anterograde transport of [:3H]amino acids from an injection in the ventral part of the subthalamic nucleus (STN, C). Terminal labeling in subtbalamopallidal fibers occurred in parallel arrays in both palIidal segments (A, B). These arrays of silver grains maintained their continuity except in caudal portions of both pallidal segments (C). Subthalamonigral fibers terminated in central portions of the pars reticulata of the SN (D). See list of abbreviations and Fig. 9A, C.

16
grains in the outer part of the MPS were found only in ventral regions, while in rhesus U-35 a larger number of silver grains were present dorsally. In the MPS labeled subthalamopaltidal fibers were found in two arrays: (1) a narrow band parallel with, and slightly medial to, the medial medullary lamina and (2) a globular array in the central region (Figs. 7, 8 and 9D).
Labeled fibers terminating in the LPS reached this segment by coursing laterally through the MPS (Fig. 9E). Fiber terminations in the LPS were confined largely to the rostral half of this segment and were not as abundant as those in the MPS. In the rostral division of the LPS two arrays of terminal fibers were seen: (1) one medial to, and parallel with, the lateral medullary lamina, and (2) a globular array located centrally (Figs. 8 and 9C). In rhesus U-41 the greatest number of silver grains were seen in ventrolateral regions of the LPS, while in the U-35 terminals were distributed equally in dorsal and ventral regions. In the central division of the LPS, arrays of terminal fibers formed a less definite pattern. Through rostral parts of this division two curved arrays, or parts of them, were arranged parallel to the lateral medullary lamina. In caudal parts of the central division of U-41 only small patches of silver grains were seen in dorsal regions (Fig. 9D). There was a suggestion of an inverse dorsoventral relationship between cells of the STN and terminations of subthalamic projections in both the MPS and the rostral LPS. In addition subthalamopallidal projections to both pallidal segments appeared to be arranged topographically in a rostrocaudal sequence.
Descending transport of isotope was to the SN. In both animals a considerable collection of silver grains was present rostrally in central parts of the SN and in a patch above its dorsal border (Figs. 7 and 8). At levels through the oculomotor rootlets, fascicles of labeled fibers descending along the dorsal border of the SN projected ventrally into the pars reticulata (Figs. 7 and 9F). This bundle continued for some distance, but became smaller and disappeared caudally. No fibers from this bundle projected dorsally into the midbrain tegmentum and no isotope was transported to the PPN, the DNR or the LC.
Subthalamic nucleus and substantia nigra In rhesus C-1377 an attempt was made to inject a small volume of tritiated
amino acids (L-leucine, L-proline and L-lysine, 100 #Ci/#l) into the STN with a Hamilton syringe via a lateral stereotaxic approach. The injection involved the caudal third of the STN and the lateral half of the rostral third of the SN (Fig. 10A). The medial half of the SN was free of silver grains at all levels. Rostral parts of the STN contained some fibers of passage, but showed no uptake from the injection. Labeled fibers emerged from the ventrolateral surface of the STN, traversed the peduncular part of the internal capsule and entered caudal portions of the globus pallidus, where they were distributed to both segments (Fig. 10B). The caudal division of the LPS contained a small collection of silver grains adjacent to the internal capsule, while extreme caudal portions of the LPS were free of silver grains. At levels through rostral portions of the STN, labeled fibers streaming into both pallidal segments were distributed mainly to dorsal regions. The majority of subthalamopallidal fibers

Fig. 9. Rhesus monkeys U-35 (A, C) and U-41 (B, D, E, F). A and B: dark field autoradiographs of /;~H]amino acids iontophoresed into the subthalamic nucleus (see Figs. 7 and 8). C: dark field autoradiograph of a transverse section through the rostral division of the lateral pallidal segment (LPS) demonstrating two parallel arrays of silver grains. D and E: dark field photomicrographs of autoradiographs through both segments of the globus pallidus; in the more caudal section (D) two parallel arrays of axonal terminals are seen in the medial pallidal segment (MPS), along with a small hand of silver grains in the dorsal part of the central division of the LPS. In E, parallel arrays of terminals are present in lateral border of the MPS and the LPS. F: dark field autoradiograph of subthalamonigral fibers descending along the dorsal border of the substantia nigra. A-F: cresyl violet; magnification: A, B, ~< 4; C, × 6; D, E, F, × 12.

Fig. 10. Rhesus monkey C-1377. A, B, C: dark field photomicrographs of autoradiographs in rhesus monkey C-1377. [3HI amino acids injected via a lateral stereotaxic approach labeled caudal parts or the subthalamic nucleus (A) and rostrolateral parts of the substantia nigra. B: demonstrates axonai transport to both pallidal segments and C shows modest transport of isotope to terminals about cells in the pedunculopontine nucleus (PPN). D: rhesus monkey C-1363 in which [3H] amino acids were injected into the left substantia nigra and the right medial patlidal segment (MPS); dark field autoradiograph of a caudal midbrain section reveals isotope in terminals of nigrotegmental (left) and pallidotegmental (right) fibers about cells of PPN. The largest number of silver grains are on the right side. E and F: squirrel monkey U-98 in which HRP was injected into the left PPN; retrograde transport of HRP was greatest in cells of the caudal MPS (E, dark field) and pars reticulata of the substantianigra(F, brightfield).A D:cresylvi° lc t ;magnif ica t i°n:A'B ' : 5 ; ( , t~:!) . ( , . f - l : : tetramethylbenzidine reaction counterstained with neutral red: magnification: E, ' , ! ~3

19
terminated in dorsal regions of both pallidal segments, but arrays of silver grains about all medullary laminae extended ventrally (Fig. 10B). Rostral pgrtions of the MPS and the LPS contained labeled fibers only in dorsolateral regions and the rostral pole of the LPS contained no silver grains. Fibers coursing through both segments of the pallidum terminated in a spotty fashion in the rostral half of the putamen and the head of the caudate nucleus. Silver grains in dense clusters were most common in medial parts of the putamen.
Isotope in thalamic nuclei conformed to the pattern consistently seen with transport from lateral portions of the SN, or the pars reticulata, in that silver grains were profuse over the ventral anterior (VAmc), the ventral lateral (VLm) and the dorsomedial (DMpl) thalamic nuclei ipsilaterally 6. In the midbrain isotope was transported caudally in the lateral part of the SN, where the bulk of the descending fibers appeared to end in the pars reticulata. Transport also was seen from the SN to the deep and middle gray layers of the ipsilateral superior colliculus ~2. Some fibers from the SN swept dorsally into the caudal midbrain tegmentum and terminated within the pars compacta of the ipsilateral pedunculopontine nucleus (Fig. 10C). No isotope was transported to the DNR or the locus ceruleus.
Controls"
In order to better evaluate descending projections from the STN and SN, autoradiographic data from two previous studies of nigral efferent pro- jections were re-examined~, 22. In three rhesus monkeys pressure injections made via a lateral stereotaxic approach involved portions of the SN selectively. Injections of [3H]amino acids involved cells in both the pars reticulata (pr) and pars compacta (pc), largely in rostrolateral (C-1363) and caudal regions (C-1315) of the SN. In rhesus C- 1374 virtually all parts of the SN were injected. In these animals anterograde transport of the label was seen in the striatum and in thalamic nuclei (i.e. VAme, VLm and DMpl). Nigrotectal projections were particularly prominent in two monkeys 22. All of these animals showed distinct labeling of terminals in the ipsilateral PPN (Fig. 10C). Silver grains over terminals in PPN were greatest dorsal to fibers of the superior cerebellar peduncle (SCP). Comparisons between transport of isotope from SN and the MPS could be made in rhesus C-1363 in which [3H]amino acids on the contralateral side were injected into the MPS. Pallidotegmental projections to PPN were much more profuse than those from the SN and terminals occupied a larger area (Fig. 10D). It was our impression that the largest concentration pallidotegmental fibers terminated in more medial regions of PPN than descending fibers from the SN.
In one squirrel monkey (U-98)HRP was successfully injected into the PPN. The injection, centered along the dorsal border of the superior cerebellar peduncle (SCP) at the level of the trochlear nucleus, caused the enzyme to diffuse throughout the PPN. Retrograde transport of HRP was most profuse in cells of the MPS where the largest number of labeled cells were seen in caudal regions (Fig. 10E). The second largest number of labeled neurons was found in the pars reticulata of SN; these cells were scattered in the lateral half of the SN (Fig. 10F). Small numbers of labeled

20
cells were identified in the zone incerta and paraventricular nucleus of th hypo- thalamus. One or two well-labeled cells were found in the rostrolateral part of the STN but none were seen in the thalamus. These data appeared consistent with observations based upon autoradiography suggesting that the PPN receives inputs from multiple subcortical sources.
DISCUSSION
These studies of the afferent and efferent connections of the subthalamic nucleus in the monkey, based upon anterograde and retrograde axoplasmic transport technics, confirm that the major connections of this nucleus are with the globus pallidus and the substantia nigra, and that connections with other brain stem nuclei are small. Although axoplasmic transport technics provide a reliable method for determining connections of the subthalamic nucleus, its small size, tens-shaped configuration, deep position and proximity to major fiber bundles made it a difficult target to inject selectively. The most difficult region of the nucleus to inject selectively was its caudal third which lies adjacent to the substantia nigra. Comparisons of the axoplasmic transport resulting from isolated injections of the subthalamic nucleus and those involving both the subthalamic nucleus and the substantia nigra provided data concerning descending brain stem projections which have been in doubt.
Globus pallidus Observations based upon retrograde transport of HRP from the subthalamic
nucleus (STN) clearly indicate that afferent fibers are derived exclusively from the lateral pallidal segment (LPS). The largest number of LPS neurons projecting to STN lie in the rostral and central divisions of that segment, but fibers also are derived from cells in the caudal division of the LPS. Cells in the rostral two-thirds of the L PS project fibers to the rostral two-thirds of the STN, and cells in the caudal division of the LPS project to the caudal third of the STN. Cells in the LPS projecting to the STN are arranged in laminar arrays parallel with the lateral medullary lamina. No clear dorsoventral topographical relationships were evident. While similar findings have been reported in the monkey, no laminar arrangement of afferent LPS neurons was described 49. In the cat McBride and Larsen 35 described a mediolateral laminar origin of pallidosubthalamic projections like that reported here. Their observations suggest- ed that cells in lateral laminae of the GP (equivalent to the LPS in the monkey) projected to lateral regions of the nucleus and that cells in medial laminae ended in medial parts of the STN. These authors reported no rostrocaudal organization of pallidosubthalamic fibers. Both degeneration and autoradiographic studies in the monkey indicate that pallidosubthalamic projections have a rostrocaudal organization in which fibers from: (1) the rostral division of the LPS project to the medial two- thirds of the STN and (2) the central division projects to the lateral third of the nucleus
excluding its rostral and caudal poles 3,4. Some retrogradely labeled cells seen in the LPS following HRP injections of
both the STN and the SN undoubtedly represented neurons projecting fibers to the

21
substantia nigra. The number of such cells probably was small t7. According to McBride and Larsen 35 HRP injections in lateral parts of the SN in the cat retrogradely labeled neurons in ventral and caudal regions of the GP (i.e. the equivalent of the LPS in the monkey), while injections in medial parts of the SN labeled neurons in ventral regions at more rostral levels. This label was considered to be transported retrograde from the pars reticulata. The large number of retrogradely labeled cells in the LPS and their pattern of distribution in these monkeys suggested that most of these cells supplied afferents to the STN.
As in previous studies there was no evidence that cells in the medial pallidal segment projected to the subthalamic nucleus3,25, 49. In those animals in which a few retrogradely labeled cells were seen in the MPS, the pipette traversed and damaged fibers of the lenticular fasciculus.
Subthalamopallidal projections have been studied by both anterograde and retrograde axoplasmic transport methods in the monkey, cat and rat a,14,27,4°,47.
Autoradiographic data in the monkey indicated the massive nature of the subthalamo- pallidal projection, suggested a topographical organization, and demonstrated that terminals end in arrays parallel to the medullary laminae. These fibers followed the course of pallidosubthalamic fibers (i.e. von Manokow's 37 second division of the ansa lenticularis) but had a reverse polarity 4°. Although profuse terminations were found in both pallidal segments, the number of arrays and the silver grain density was much greater in the medial pallidal segment (see ref. 40, their Figs. 6-13 and 15). Rostromedial parts of the STN were found to project to rostral and medial portions of the LPS (i.e. the rostral division) while lateral parts of the nucleus projected to the central division of the LPS. In the cat subthalamopallidal fibers projected to both tile entopeduncular nucleus and the globus pallidus, but did not exhibit a laminar pattern. Again the projection to the feline homologue of the medial pallidal segment (i.e. the entopeduncular nucleus) was greatest. Isotope transported to the globus pallidus (i.e. the equivalent of the LPS in the monkey) was found mainly in rostral regions (see ref. 40, their Fig. 21).
Data from the present study, based upon partial injections of the STN, indicate profuse projections to both pallidal segments in laminar arrays as described by Nauta and Cole 40. Injections of the STN involving either rostral or caudal parts of the nucleus appeared to produce the greatest transport of the isotope to MPS. However, silver grains in the MPS obviously were over both terminals and fibers in passage. Even after taking into account the fibers of passage, the number of apparent terminals was much greater in the MPS than in the LPS. Silver grains concentrated in the apical regions of the MPS at caudal levels moved laterally away from the internal capsule, and at more rostral levels were seen in arrays on both sides of the accessory medullary lamina, The locations of silver grains in the outer part of the MPS (i.e. lateral to the accessory medullary lamina) could be correlated with the location of maximal primary label in the STN. When dorsal regions of the STN were injected, anterograde transport of isotope was found mainly in ventral regions of the outer part of the MPS (U-41 j; a similar correlation was found between cells in ventral portions of the STN and fibers projecting to dorsal regions of the outer part of the M PS (U-35).

22
Because silver grains were found in nearly all regions of the inner part of the MPS, no similar correlation could be made.
Fibers projecting to the LPS coursed through the MPS and the medullary laminae. Injections of [3H]amino acids in the rostral STN produced: (I) two parallel arrays of silver grains in rostral division of the LPS, and (2) thinner less definite arrays of silver grains in the central division of the LPS. Specific correlations between injection sites in the STN and regions of terminal transport in the LPS suggested relationships between rostral regions of both structures.
When only the caudal part of the STN was labeled, silver grains were found over terminals in the caudal regions of the LPS and relatively little isotope was transported to rostral regions. Our observations suggest that subthalamopallidal fibers projecting to the LPS in the monkey are topographically organized in a rostrocaudal sequence. Data were not sufficient to determine a topographical organization between these structures in a dorsoventral dimension but in U-41 cells in dorsal regions of STN projected preferentially upon ventral regions of the LPS. These findings tend to support previous HRP studies indicating that cells near the dorsal capsule of the STN project mainly to ventral regions of the LPS, while cells in ventral parts of the STN project to dorsal regions of the LPS 3. Although autoradiographic data are not as precise as that based upon the retrograde transport of HRP, they tend to support this inverse relationship between the STN and both segments of the gtobus pallidus.
Transport of HRP from the MPS in the monkey reportedly resulted in retrograde labeling of only a small number of cells in STN 14 which did not reflect the profuse subthalamopallidal projection to the MPS described by Nauta and Cole 4° or seen in the present study. In our own HRP study of subthalamopallidal projections in the monkey z, labeling of portions of the MPS consistently produced retrograde transport of enzyme to neurons in caudal and medial parts of the STN, a region of the nucleus which has not been demonstrated to receive projections from either the LPS or the cortex 4'19,25'42. HRP injections in divisions of the LPS in the monkey indicated that: (I) cells in the medial half of the middle third of the STN projected to the rostral division of the LPS, and (2) cells in central regions of the rostral two-thirds of the STN projected to the central division of the LPS.
Data from this and previous studies indicate that connections between the globus pallidus and the subthalamic nucleus in the primate are not totally reciprocal a, z5,49. Although the medial pallidal segment receives a profuse projection from the subthalamic nucleus, it has no return projection. Thus, there are no reciprocal connections between the MPS and the STN. The STN appears to be an important source of afferents to cells of the MPS that project to thalamic nuclei [i.e. the ventral anterior, pars principalis (VApc), the ventral lateral, pars oralis (VLo) and pars medialis (VLm), and the centromedian (CM)], the lateral habenular nucleus and the pedunculopontine nucleus (PPN)3°,42: These efferent projections from the STN arise from cells in the caudal and medial parts of the nucleus 3 and end in arrays parallel to the accessory and medial medullary laminae of the MPS.
On the basis of data from this study, pallidosubthalamic and subthalamopallidal

23
projections interconnecting the STN and LPS appear to: (1) be reciprocal, (2) arise from cells and have terminals arranged in arrays parallel to the lateral medullary lamina, and (3) be organized topographically in rostrocaudal sequence. Unfortu- nately, injections of [3H]amino acids in the STN do not provide as precise detail concerning the cells of origin of subthalamopallidal projections as the retrograde transport of HRP injected selectively into various segments of the globus pallidus '3. Data from the above-mentioned study cast doubt as to whether this apparent reciprocal relationship exist on a point for point basis. A fluorescent retrograde double-labeling study in the rat suggested that virtually all STN neurons project axon collaterals to both the globus pallidus and the substantia nigra and that both collateral systems exhibit a mediolateral topographical organization 27. These data in the rat suggest that the STN contains few, if any, interneurons, an observation that differs greatly from classic Golgi studies in the monkey 45.
Substantia nigra Both degeneration and axoplasmic transport studies indicate that the STN
projects fibers to the SN 24,26,27,35,40,47,54. Most of the evidence suggests that these
fibers terminate mainly in the pars reticulata. Physiological evidence in the rat also indicates that STN neurons supply axonal collaterals to both the GP and the SN ~:~,1~. Orthodromic and antidromic stimulations of the pathway from the STN to the SN support the view that these fibers end upon cells of the pars reticulata is. Observations in the present study based upon autoradiography suggest that subthalamonigral fibers from rostral regions of the STN descend along the dorsal border ot the SN for some distance before projecting ventrally to end largely in the pars reticulata. Fibers arising from more caudal regions of the STN may follow another trajectory. Evidence for subthalamonigral projections in the present study, based upon HRP injections, is largely indirect. In animals in which HRP was injected into caudal parts of the STN and rostral parts of the SN, virtually all STN neurons were profusely labeled. The extensive labeling of STN neurons was considered to be due in large part to retrograde transport of the enzyme by subthalamonigral fibers.
Although degeneration and autoradiographic studies have not supported the existence of a nigrosubthalamic projection 5 7,10, a recent HRP study has raised questions about such fibers 49. This study suggests that cells in both parts of the SN show retrograde transport of the marker from the STN.Although uptake of the enzyme by injured nigrostriatal axons coursing along the dorsal border of the STN was not excluded, the evidence was interpreted as suggesting the existence of a nigro- subthalamic pathway. This view received its best support from electron microscopic evidence of degenerated boutons in the STN following a lesion in the SN. In animals in this study with HRP injections confined to portions of the STN, no enzyme was transported retrograde to SN neurons at the light microscopic level. Physiological data in the rat based on a variety of forms of stimulation did not support the existence of a nigrosubthalamic pathway TM.

24
Tegmental nuclei Both anterograde and retrograde transport studies in the monkey and cat
suggest that the STN may receive relatively small projections from the PPN, the DNR and the LC t6,35,43,49. Data from the present study, based upon retrograde transport of
HRP, indicate that only a few cells of PPN project to the STN, but that a large number of PPN neurons project upon rostral portions of the SN. Large number of cells in the PPN were consistently labeled in animals in which the injections involved portions of the SN. It is difficult to explain the discrepancies between our observations and those reported in the literature without inferring that the injections were not as discretely localized to the STN as they appeared. Autoradiographic data concerning the projections of PPN in the cat suggested substantial projections to STN and SN and variable projections to the intralaminar thalamic nuclei, the hypothalamus and the central gray16,as, 39.43. Even though autoradiographic methods obviate the fiber of passage problem, other cell groups in the reticular formation may be inadvertently labeled. While cells of PPN probably project to multiple structures, including the medial pallidal segment3, ta, evidence from the present study suggests that fibers from this nucleus passing to the SN may constitute the largest bundle.
Although Nauta and Cole 4° described fibers from the STN projecting to the pars compacta of the PPN in the monkey, no similar descending pathway was seen in the present study. Injections of [all]amino acids into both the STN and the SN (C-1377), or only into SN6, 22, resulted in transport of label to terminals around cells of PPN. These findings leave little doubt that cells in the SN project to the PPN, but comparisons of nigrotegmental and pallidotegmental projections to this nucleus in the same animal (C-1363) indicate that the MPS contributes a much larger number of fibers.
Hedreen 2t first identified the nigrotegmental pathway terminating in the PPN in the rat and established its non-dopaminergic nature. The suggestion that the nigro- tegmental pathway originated from the pars reticulata has been confirmed autoradio- graphically by small selective microiontophoretic injections of [all]amino acids into the pars reticulatal; these authors also noted descending projections from the pars compacta and the ventral tegmental area that traversed the region of the PPN en route to other terminations.
In addition to reciving inputs from the SN, the PPN in the monkey receives projections from the motor cortex 2°,31 and the MPS s,25,a°,42. While projections from the motor cortex and the MPS appear to be the dominant sources of input to PPN in the monkey, this does not seem to be the case in the cat 32,a3,as. In the cat the densest projection to PPN appears to arise largely from cells of the pars reticulata of the SN as. It is of interest that the PPN receives its subcortical inputs from structures that receive striatal efferents (i.e. the MPS and the SN) and that the efferents from the nucleus largely project back upon these nuclei l,a,14,21,3s,ag. While interconnections between the PPN and the MPS may be reciprocal, the projection from PPN to the MPS appears very small a. Excellent data in the cat and rat indicate that connections between the PPN and the SN are not reciprocal in the strict sense, since nigrotegmental fibers arise from cells of the pars reticulata and ascending projections from PPN end upon

25
cells of the pars compactal,3s, 39. The functional nature of this mesencephalic loop circuit remains unknown, but conceivably these connections might modulate or integrate the activities of nigral and pallidal neurons.
Retrograde transport studies indicated that no cells of the DNR project to the STN. HRP injections involving the SN consistently resulted in labeling of cells in the ipsilateral DNR. This observation has been well documented by prior studies which suggest that most fibers from this nucleus end upon cells in the pars compactaZ, 11,24, 5:3. It is of interest that cells of the pars compacta of SN project caudally to the DN R, establishing a closed loop circuit 1. Anterograde transport methods have not demon- strated any projection from the STN to the DNR in this or other studies 4°.
No definite conclusion can be drawn concerning possible projections from the LC to the STN. A few sparsely labeled cells in the LC were seen in all animals in which 1~1 RP was injected in the region of the STN. It was our impression that label in cells of the LC probably resulted from uptake by ascending fibers traversing regions adjacent to the STN. This possibility also was considered in another study in the monkey, but dismissed because HRP injections dorsal to the STN labeled fewer cells in the LC 4.~. These authors suggested the labeled cells in the LC might be those few which do not contain cateeholamines z3. Anterograde transport of isotope revealed no evidence of a projection from the STN to the LC in this or other studies in the monkey 4°. Autoradiographic data in the rat indicate that neither the pars compacta nor the pars reticulata of the SN project to LC 1.
Thalamus, striatum and cerebral cortex
Data from this study do not indicate that the STN receives or projects fibers to the thalamus or striatum. Although it has been reported that striatal stimulation in the awake monkey produces a monosynaptic short latency spike discharge in the STN followed by a long lasting depression of spontaneous activity 44, no retrograde transport of HRP from the STN to striatal neurons was found in the present study. These findings confirm earlier observations in both the monkey and cat 49. Nauta and Cole 4° raised the question of a possible projection from the STN to the putamen because of observations in one animal. Massive HRP injections of the striatum in the rat, cat and monkey have not produced retrograde transport of the enzyme to cells of the STN 41,51,52. Autoradiographic findings suggested that a few labeled axons
from STN may project to the ventral lateral and ventral anterior thalamic nuclei 40. No evidence of a subthalamo-thalamic projection has been reported in previous degenera- tion s or retrograde transport studies 49, or in the present investigation.
Attempts to confirm and further define the origins of corticosubthalamic projections, demonstrated by autoradiographic methodslg,28, zg, by using retrograde transport technics have been disappointing. Although degenerated corticosubthalamic fibers have been demonstrated electron microscopically 50, only a small number of cortical neurons have been labeled in retrograde fashion after HRP injections of the STN a'%49. The number of location of labeled cortical neurons appeared to vary without apparent relationship to the injected HRP. McBride and Larsen 35 attributed failure to label cortical neurons to the thinnest of terminal collaterals and the

26
insensitivity of DAB as a chromagen. Findings in the present study were equally unrewarding with respect to the labeling of corticosubthalamic neurons even though the more sensitive TMB was used as the chromagen. Failure to retrogradely label cortical neurons may be due to inability to inject sufficiently high concentrations of HRP into lateral regions of the STN, the area that receives the largest number of corticosubthalamic fibers 19.
The subthalamic nucleus receives projections from cells of the lateral pallidal segment arranged in arrays parallel to the medullary lamina which have a rostrocaudal organization. No cells of the medial pallidal segment or of the substantia nigra project directly to the subthalamic nucleus. The STN may receive a very small number ot" fibers from the pedunculopontine nucleus which receives inputs from the motor cortex, the medial pallidal segment and the pars reticulata of the substantia nigra. Based upon these data, it is difficult to visualize the output of the peduncutopontine nucleus as an important relay by which impulses from cells of medial pallidal segment or the pars reticulata of the substantia nigra could influence the activities of STN neurons. The subthalamic nucleus does not receive afferents from thalamic nuclei, the striatum or the dorsal nucleus of the raphe. No conclusions can be drawn on the basis of retrograde transport studies concerning projections of the cerebral cortex or the locus ceru|eus upon the subthalamic nucleus, although the latter appears doubtful.
Projections of the subthalamic nucleus are massive to both segments of the globus pallidus and the pars reticulata of the substantia nigra. Subthalamopallidal fibers terminate in laminar arrays parallel to the medullary laminae and are most numerous in the medial pallidal segment, the segment that gives rise to the major efferent system of the corpus striatum. Subthalamopallidal projections to both segments of the globus pallidus are arranged topographically in a rostrocaudal sequence and appear organized in an inverse dorsoventral relationship. Although interconnections between the subthalamic nucleus and the lateral patlidal segment appear reciprocal, other data suggest this relationship does not exist on a point for point basis a. Subthalamonigral fibers arise from cells in all parts of the STN and
project mainly to cells of the pars reticulata. The medial pallidal segment and the pars reticulata of the SN appear to
represent the two main sources of descending subcortical input to the pedunculo- pontine nucleus. In the monkey the largest number of afferents to this nucleus arise from the medial pallidal segment. While the pedunculopontine nucleus has projec- tions, among possible others, to the medial pallidal segment and the substantia nigra,
those to the substantia nigra are most profuse.
ABBREVIATIONS CN caudate nucleus DM dorsomedial nucleus
AC anterior commissure DMpl dorsomedial nucleus, pars laminaris AD anterodorsal nucleus DNR dorsal nucleus of the raphe AM anteromedial nucleus Fx fornix AV anteroventral nucleus G P globus pallidus CC crus cerebri Hb habenula CI inferior colliculus IC internal capsule CM centromedian nucleus LC locus ceruleus

27
LD lateral dorsal nucleus Put. putamen LG lateral geniculate RN red nucleus LP lateral posterior nucleus SC superior colliculus LPS lateral pallidal segment SCP superior cerebellar peduncle MG medial geniculate SN substantia nigra MLF median longitudinal fasciculus STN subthalamic nucleus MM mammillary body VA ventral anterior nucleus MPS medial pallidal segment VAmc ventral anterior nucleus, pars magno- MTT mammillothalamic tract cellularis N.]II 3rd nerve VApc ventral anterior nucleus, pars prin- NST nucleus of the stria terminalis cipalis OC optic chiasm VL ventral lateral nucleus OT optic tract VLc ventral lateral nucleus, pars caudalis OMC oculomotor nuclear complex VLm ventral lateral nucleus, pars medialis PC posterior commissure VLo ventral lateral nucleus, pars oralis pc pars compacta VPL ventral posterolateral nucleus PF parafascicular nucleus VPM ventral posteromedial nucleus PPN pedunculopontine nucleus ZI zona increta pr pars reticulata Ill 3rd ventricle Pul. pulvinar
ACKNOWLEDGEMENTS
The authors acknowledge with sincere thanks the contr ibut ions o f Mr. An ton io
B. Pereira and Mr. Thomas T. Nevin who prepared the histological material and Mrs.
Lillian Magruder who typed the manuscript .
This work was supported by research grantNS-14802 from the Nat iona l Insti tute
of Neurological and Communica t ive Disorders and Stroke o f the Nat iona l Institutes
o f Health, Bethesda, Maryland, and research grant 811 C07005 from the Depar tmen t
of Defense, Uni formed Services Universi ty o f the Heal th Sciences.
REFERENCES
I Beckstead, R. M., Domesick, V. B. and Nauta, W. J. H., Efferent connections of the substantia nigra and ventral tegmental area in the rat, Bra& Research, 175 (1979) 191-217.
2 Bunney, B. S. and Aghajanian, G. K., The precise localization of nigral afferents in the rat as determined by a retrograde tracing technique, Brain Research, 117 (1976) 423 435.
3 Carpenter, M. B., Batton, R. R., Carleton, S. C. and Keller, J. T., Interconnections and organization of pallidal and subthalamic nucleus neurons in the monkey, J. comp. Neurol., 197 (1981) 579-603.
4 Carpenter, M. B., Fraser, R. A. R. and Shriver, J., The organization of the pallidosubthalamic fibers in the monkey, Brain Research, 11 (1968) 522 559.
5 Carpenter, M. B. and McMasters, R. E., Lesions of the substantia nigra in the rhesus monkey. Efferent fiber degeneration and behavioral observations, Amer. J. Anat., 114 (1964) 293-320.
6 Carpenter, M. B., Nakano, K. and Kim, R., Nigrothalamic projections in the monkey demon- strated by autoradiographic technics, J. comp. Neurol., 165 (1976) 401-416.
7 Carpenter, M. B. and Peter, P., Nigrostriatal and nigrothalamic fibers in the rhesus monkey, J. comp, Neurol., 144 (1972) 93-116.
8 Carpenter, M. B. and Strominger, N, L., Efferent fiber projections of the subthalamic nucleus in the rhesus monkey. A comparison of the efferent projections of the subthalamic nucleus, substantia nigra and globus pallidus, Amer, J. Anat., 121 (1967) 41-72.
9 Carpenter, M. B. and Whittier, J. R., Study of method for producing experimental lesions of the central nervous system with special reference to stereotaxic techniques, J. comp. Neurol., 97 (1952) 73-132.

28
10 Cole, M., Nauta, W. J. H. and Mehler, W. R., The ascending efferent projections of the substantia nigra, Trans. Amer. neurol. Ass., 89 (1964) 74-78.
11 Conrad, L. C., Leonard, C. M. and Pfaff, D. W., Connections of the median and dorsal raphe nuclei in the rat. An autoradiographic and degeneration study, J. comp. Neurol., 156 (1974) 179-206.
12 Cowan, W. M., Gottlieb, S. 1., Hendrickson, A. E., Price, J. k. and Woolsey, T. A., The autoradio- graphic demonstration of axonal connections in the central nervous system, Brain Research, 37 (1972) 21-51.
13 Deniau, J. M., Hammond, C., Chevalier, G. and Feger, J., Evidence for branched subthalamic nucleus projections to substantia nigra, entopeduncular nucleus and globus pallidus, Neurasci. Lett., 9 (1978) 117 121.
14 DeVito, J. L., Anderson, M. E. and Walsh, K. E., A horseradish peroxidase study of afferent connections of the globus pallidus in Macaca mulatta, Exp. Brain Res., 38 (1980) 65 73.
15 Graham, R. C., Jr. and Karnovsky, M. J., The early stages of absorption of injected horseradish peroxidase in the proximal tubules of mouse kidney: ultrastructural cytocbemistry by a new tech- nique, J. Histochem. Cytochem., 14 (1966) 291-302.
16 Graybiel, A. M., Direct and indirect preoculomotor pathways of the brain stem: an autoradio- graphic study of the pontine reticular formation in the cat, J. comp. Neurol., 175 (1977) 37-78.
17 Grofov/t, I., The identification of striatal and pallidal neurons projecting to substantia nigra. An experimental study by means of retrograde axonal transport of hroseradish peroxidase, Brain Research, 91 (1975) 28f~291.
18 Hammond, C., Deniau, J. M., Rizk, A. and Feger, J., Electrophysiological demonstration of an excitatory subthalamonigral pathway in the rat, Brain Research, 151 (1978) 235 244.
19 Hartmann-von Monakow, K., Akert, K. and KiJnzle, H., Projections of the precentral motor cortex and other cortical areas of the frontal lobe to the subthalamic nucleus in the monkey Exp. Brain Res., 33 (1978) 395-403.
20 Hartmann-von Monakow, K., Akert, K. and Kiinzle, H., Projections of precentral and premotor cortex to the red nucleus and other midbrain areas in Macaca fascieularis, Exp. Brain Res., 34 (1979) 91-105.
21 Hedreen, J. C., Separate demonstration of dopaminergic and non-dopaminergic projections of the substantia nigra in the cat, Anat. Rec., 169 (1971) 338 (Abstract).
22 Jayaraman, A., Batton, R. R. and Carpenter, M. B., Nigrotectal projections in the monkey: an autoradiographic study, Brain Research, 135 (1977) 147--152.
23 Jones, B. E. and Moore, R. Y., Catecholamine-containing neurons of the nucleus locus coeruleus in the cat, J. comp. Neurol., 157 (1977) 43-52.
24 Kanazawa, I., Marshall, G. R. and Kelly, J. S., Afferents to the rat substantia nigra studied with horseradish peroxidase, with special reference to fibres from the subthalamic nucleus, Brain Research, 115 (1976) 485M-91.
25 Kim, R., Nakano, K., Jayaraman, A. and Carpenter, M. B., Projections of the gtobus pallidus and adjacent structures: an autoradiographic study in the monkey, J. eomp, Neurol., 169 (1976) 263-289.
26 Knook, H. L., The Fibre-Connections of the Forebrain, Van Gorcum, Assen, 1965. 27 Kooy, D. van der and Hattori, T., Single subthalamic nucleus neurons project to both the globus
pallidus and substantia nigra in rat, J. comp. Neurol., 192 (1980) 751-768. 28 K/Jnzle, H., An autoradiographic analysis of the efferent connections from pretnotor and
adjacent prefrontal regions (areas 6 and 9) in Macaeafascieularis, Brain, Behav. Evol., 15 (1978) 135-234.
29 K~nzle, H. and Akert, K., Efferent connections of cortical area 8 (frontal eye field) in Macaca Jascicularis. A reinvestigation using the autoradiographic technique, J. comp. Neurol., 173 (1977) 147- 164.
30 Kuo, J. S. and Carpenter, M. B., Organization of pallidothalamic projections m the rhesus monkey, J. comp. Neurol., 151 (1973) 201-236.
31 Kuypers, H. G. J. M. and Lawrence, D. G., Cortical projections to the red nucleus and the brain stem in the rhesus monkey, Brain Research, 4 (1967) 151-188.
32 Larsen, K. D. and McBride, R. L., The organization of feline endopeduncular nucleus projections: anatomical studies, J. comp. Neurol., 184 (1979) 293-308.
33 Larsen, K. D. and Sutin, J., Output organization of the feline entopeduncular and subthalamic nuclei, Brain Research, 157 (1978)21-31.
34 LaVail, J. H., Retrograde cell degeneration and retrograde transport techniques. In W. M. Cowan

29
and M. Cu6nod (Eds.), The Use of Axonal Transport fin" Studies of Neuronal Connectivity, Elsevier, Amsterdam, 1975, pp. 217 247.
35 McBride, R. L. and Larsen, K. D., Projections of the feline globus pal[idus, Brain Research, 189 (1980) 3-14.
36 Mesulam, M. M., Tetramethyl benzidine for horseradish peroxidase neurohistochemistry: a non- carcinogenic blue reaction product with superior sensitivity for visualizing neural afferents and efferents, J. Histochem. Cytochem., 26 (1978) 106 117.
37 Monakow, C. yon, Experimentelle und patho[ogischanatomische Unterschungen fiber die Haubenregion, Schhfigel und die Regio subthalamica, Arch. Psychiat. Nervenk., 27 (1895) 1 129.
38 Moon Edley, S. L., A Neuroanatomical Study of the Nucleus Tegmenti Pedunculopontinus #1 the Cat, Doctoral Dissertation, Mass. Institute of Technology, Cambridge, MA 1979.
39 Moon Edley, S. and Graybiel, A. M., Connections of the nucleus tegmenti pedunculopontinis, pars compacta (TPc) in cat, Anat. Rec., 196 (1980) 129A (Abstract).
40 Nauta, H. J. W. and Cole, M., Efferent projections of the subthalamic nucleus: an autoradio- grapic study in monkey and cat, J. comp. Neurol., 180 (1978) 1 16.
41 Nauta, H. J. W., Prilz, M. B. and Lasek, R.L., Afferents to the rat caudoputamen studied with horseradish peroxidase. An evaluation of a retrograde neuroanatomical research method, Brain Research, 67 (1974) 219-238.
42 Nauta, W. J. H, and Mehler, W. R., Projections of the lentiform nucleus in the monkey, Brain Research, 1 (1966)3 42.
43 Nomura, S., Mizuno, N. and Sugimato, T., Direct projections from the pedunculopontine teg- mental nucleus to the subthalamic nucleus in the cat, Brain Research, 196 (1980) 223 227.
44 Ohye, C., LeGuyader, C. and Feger, J., Responses of subthalamic and pal[idal neurons to striatal stimulation: an extracellular study on awake monkeys, Brain Research, 111 (1976) 241-252.
45 Rafols, J. A. and Fox, C. A., The neurons in the primate subthalamic nucleus: a Golgi and electron microscopic study, J. comp. Neurol., 168 (1976) 75 112.
46 Ranson, S. W. and Ranson, S. W. Jr., Efferent fibers of the corpus striatum, Res. Publ. Ass. nerv. ment. Dis., 21 (1942) 69-76.
47 Ricardo, J. A., Efferent connections of the subthalamic region in the rat. I. The subthalamic nucleus of Luys, Brain Research, 202 (1980) 257 271.
48 Riley, H. A., An Atlas of the Basal Ganglia, Brain Stem, and Spinal Cord Based on Myelin- stained Material, Williams and Wilkins, Baltimore, 1943.
49 Rinvik, E., Grofov~, I., Hammond, C., Feger, J. and Deniau, J. M., A study of the afferent con- nections to ithe subthalamic nucleus in the monkey and the cat using the HRP technique. In L. J. Poirier, T. L. Sourkes and P. J. B6dard (Eds.), Advances in Neurology, Vol. 24, Raven Press, New York, 1979, pp. 53 70.
50 Romansky, K. V., Usunoff, K. G., Ivanov, D. P. and Galabov, G. P., Corticosubthalamic pro- jection in the cat: an electron microscopic study, Brain Research, 163 (1979) 319-322.
51 Szabo, J., Organization of the ascending striatal afferents in monkeys, J. comp. Neurol., 189 (1980) 307-321.
52 Szabo, J., Distribution of striatal afferents from the mesencephalon in the cat, Brain Research, 188 (1980) 3 21.
53 Taber Pierce, E., Foote, W. E. and Hobson, J. A., The efferent connection of the nucleus raphe dorsalis, Brain Research, 107 (1976) 137-144.
54 Whittier, J. R. and Mettler, F. A., Studies on the subthalamus of the rhesus monkey, lI. Hyper- kinesia and other physiological effects of subthalamic lesions with special reference to the subthalamic nucleus of Luys, J. comp. Neurol., 90 (1949) 319-372.





























