Upload
alma-alfaro
View
353
Download
4
Embed Size (px)
Citation preview
Unidad Temtica II BiologaPor: Alma Alfaro Resumen obtenido del libro El Mundo de la clula Becker, Kleinsmith, Hardin Enzimas: los catalizadores de la vidaLas reacciones o procesos celulares son mediados por protenas catalizadoras llamadas enzimas. En algunos casos son llevados a cabo por ribozimas (ARN). Energa de activacin y el estado metaestable Muchos procesos pueden ser termodinmicamente factibles, sin embargo en cambio no es observable por la baja velocidad a la que se produce, e aqu la importancia de las enzimas. La barrera de activacin energtica debe ser superada antes de que se pueda verificar una reaccin qumica Existe una energa de activacin especfica que es la cantidad mnima de energa que los reactivos deben tener antes de que ocurra una colisin entre ellos. Esto lo logran alcanzado un estado qumico intermedio llamado estado de transicin. El estado metaestable es el resultado de la barrera de activacin La velocidad de las reacciones no catalizadas en las clulas es muy baja, las molculas que son aparentemente estables, son termodinmicamente inestables y no tienen energa suficiente para sobrepasar la energa de activacin, esto es el estado metaestable. Los catalizadores superan la barrera de la energa de activacin La energa de activacin es importante para: el mantenimiento del estado metaestable y como barrera que debe ser superada para que las reacciones tengan lugar a velocidades adecuadas. La manera de lograr que una molcula supere la energa de activacin (EA) es aumentando la proporcin de molculas energticas, esto se consigue ya sea aumentando el contenido medio de energa de todas las molculas, o reduciendo el requerimiento de energa de activacin. Una manera de incrementar el contenido de energa del sistema es suministrando calor, as se aumentar la energa cintica de las molculas (asegurando que haya un mayor nmero de molculas reactivas). El problema del aumento de temperatura es que no es compatible con organismos vivos ya que estos son isotrmicos (de temperatura constante) por lo que requieren tambin de mtodos isotrmicos. Una alternativa al incremento de la temperatura es reducir el requerimiento de la energa de activacin. Los reactivos se pueden agrupar en una
superficie que facilite que las porciones potencialmente reactivas de las molculas adyacentes se yuxtapongan, la interaccin se ver favorecida y la energa de activacin se reducir. Quien provee esta superficie reactiva son los catalizadores, los cuales aumentan la velocidad de la reaccin disminuyendo la energa de activacin. Una caracterstica primordial del catalizador es que no se modifica permanentemente, ni se consume. Sencillamente proporciona un ambiente idneo para facilitar la reaccin. Enzimas como catalizadores biolgicos Las tres propiedades bsicas de los catalizadores son: 1. Incrementa la velocidad de la reaccin disminuyendo el requerimiento de energa de activacin, permitiendo una reaccin termodinmicamente factible sin necesidad de activacin trmica. 2. Un catalizador funciona formando complejos transitorios con las molculas de sustrato, ligndolas de manera que se facilite su interaccin. 3. Un catalizador solo cambia la velocidad en que se consigue el equilibrio, no la posicin. Esto aplica a catalizadores orgnicos e inorgnicos, aunque los sistemas biolgicos rara vez usan catalizadores inorgnicos. Las molculas orgnicas son ms especficas. La mayora de las enzimas son protenas Sitio activo Uno de los conceptos ms importantes de entender de las enzimas como protenas es el sitio activo. Cada enzima contiene dentro de su estructura terciaria un grupo caracterstico de aminocidos que forman en sitio activo, en el cual ocurre el evento cataltico del cual esa enzima es responsable. Normalmente el sitio activo es un surco o bolsillo real con propiedades qumicas y estructurales que permiten la acomodacin adecuada y especfica del sustrato. El sitio activo de una enzima tpica comprende un nmero determinado de aminocidos, que normalmente no son contiguos, a lo largo de la secuencia primaria de la protena. En todo caso, se mantienen juntos, en una conformacin correcta, gracias al plegamiento tridimensional caracterstico de la cadena polipeptdica. Slo cuando la molcula alcanza su conformacin tridimensional estable se renen los aminocidos especficos para constituir el sitio activo. Algunas enzimas tambin pueden portar componentes no proteicos llamados grupos protticos los cuales normalmente son molculas orgnicas pequeas o iones metlicos que funcionan como aceptores de electrones (debido a que ninguno de los aminocidos laterales es buen aceptor de electrones), cuando estos estn presentes se localizan en el sitio activo y son indispensables para la actividad cataltica.
Especificidad enzimtica Las enzimas manifiestan un alto grado de especificad por el sustrato. La especificad es una de las propiedades ms caractersticas de los seres vivos. Los catalizadores inorgnicos son muy poco especficos ya que actan sobre una variedad de componentes con una caracterstica qumica general. No todas las enzimas son tan especficas, algunas reconocen un nmero concreto de sustratos y otras una categora amplia de sustancias, siempre que posean caractersticas estructurales comunes. Esta especificad por grupos es ms frecuente en enzimas implicadas en la sntesis y degradacin de polmeros. Diversidad enzimtica y nomenclatura Las seis clases principales de enzimas son: oxidoreductasas, transferasas, hidrolasas, liasas, isomerasas y ligasas. Oxidoreductasas: se encargan de reacciones xido-reduccin. Transferasas: se encargan de transferir grupos funcionales desde una molcula a otra. Hidrolasas: se encargan de la ruptura hidroltica de una molcula en dos. Liasas: se encargan de la eliminacin de un grupo desde, o adicin de un grupo a, una molcula, con redistribucin de electrones. Isomerasas: se encargan del desplazamiento de un grupo funcional dentro de una molcula. Ligasas: se encargan de la unin de dos molculas para formar una nica molcula. Cabe recordar el sistema EC que nombra las enzimas segn su Clase, sub clase, sub sub clase y nmero de entrada. Sensibilidad a la temperatura La velocidad de una reaccin catalizada enzimaticamente aumenta con la temperatura, hasta cierto punto, si sobrepasa ese punto la molcula inicia a desnaturalizarse. Los puentes de hidrgeno se rompen, las interacciones hidrfobas cambian, y la integridad estructural del sitio activo se interrumpe, causando la prdida de la actividad.La velocidad de reaccin de una enzima humana es mxima alrededor de los 37 C. La mayora de las enzimas de los seres homeotermos se inactivan por la temperatura, por encima de los 50-55 C. Sensibilidad al pH Las enzimas tambin son sensibles al pH, muchas slo son activas dentro de un rango de pH de 3-4 unidades. La dependencia de pH de una enzima normalmente refleja el medio ambiente en el cual la enzima est activa. Sensibilidad a otros factores Las enzimas tambin pueden ser sensibles a sustancias inhibidoras o activadoras de la enzima. La mayora de las enzimas son sensibles al medio ambiente inico, el cual es probablemente el que afecta a la conformacin
total de la enzima, debido a que los puentes de hidrgeno dependen de l. El medio ambiente inico puede afectar tambin a las uniones del sustrato porque las interacciones inicas estn a menudo involucradas en las uniones entre el sustrato y el sitio activo. La unin del sustrato, la activacin y la reaccin se producen en el sitio activo En el sitio activo de la enzima ocurre la unin, activacin y transformacin qumica del sustrato. Los catalizadores enzimpaticos procesan a una velocidad 10^7 a 10^14 ms rpido que sin enzima y los catalizadores inorgnicos trabajan de 10^3 a 10^4 veces ms rapido. La unin del sustrato El contacto inicial entre el sitio activo de la enzima y una molcula de sustrato potencial, depende de colisiones aleatorias. Una vez en la hendidura o bolsillo del sitio activo, las molculas de sustrato se unen temporalmente a la superficie de la enzima, con la orientacin apropiada para interaccionar entre s y con grupos catalticos especficos de la enzima, lo cual facilita la reaccin. La unin del sustrato suele realizarse a travs de aminocidos cargados, por medio de puentes de hidrgeno o enlaces inicos (o ambos). La suma de estas uniones (dbiles) poseen suficiente fuera para mantener a la molcula en su lugar. La unin al sustrato es fcilmente reversible. Existi la teora del modelo de la cerradura y llave el cual ajustaba el sitio activo a la molcula. Ms se provey el modelo de ajuste inducido, este modelo supone que la unin inicial de la molcula de sustrato al sitio activo deforma la enzima y el sustrato, estabilizando a las molculas de sustrato en su estado de transicin y permitiendo que los enlaces crticos sean ms susceptibles a ataque catalticos. Activacin del sustrato El papel del sitio activo no es slo reconocer y unir el sustrato apropiado sino tambin activarlo, sumergindolo en el medio qumico necesario para la catlisis. Una determinada reaccin catalizada enzimaticamente, puede involucrar a uno o ms recursos de activacin del sustrato. Los mecanismos ms comunes son los siguientes: 1. El cambio en la conformacin enzimtica inducida por la unin del inicial sustrato al sitio activo, no slo causa una mejor complementariedad y un ajuste ms estrecho enzima-sustrato, sino que tambin deforma uno o ms de sus enlaces, debilitando as dichos enlaces, hacindolos ms susceptibles al ataque cataltico. 2. La enzima tambin puede aceptar o donar protones incrementando, por lo tanto, la reactividad qumica del sustrato. 3. Como forma adicional para la activacin del sustrato, las enzimas tambin pueden aceptar o donar electrones, formando de ese modo enlaces covalentes temporales entre la enzima y su sustrato. El acontecimiento cataltico
1. El sustrato colisiona con el sitio activo, donde es mantenido porenlaces dbiles con los grupos R de los aminocidos del sitio activo. La unin del sustrato provoca un cambio conformacional en la enzima, responsable de que el sustrato sea unido ms firmemente (ajuste inducido). 2. El sustrato es convertido en productos. 3. Los productos son liberados. 4. El sitio activo est disponible para otra molcula de sustrato. (la enzima retoma su forma) Cintica enzimtica La actividad enzimtica tambin depende de la concentracin de los sustratos, de los productos y de los inhibidores presentes en la clula. Estos aspectos cuantitativos de la catlisis enzimtica son el campo de la cintica enzimtica. La mayora de enzimas siguen la cintica de Michaelis Menten La velocidad de una reaccin catalizada enzimaticamente aumenta con la concentracin de sustrato. Existe una importante relacin entre la velocidad inicial de reaccin v y la concentracin de sustrato (S) , mientras que S tiende al infinito, v tiende hacia un valor mximo que depende del nmero de molculas enzimticas y solo puede incrementarse aadiendo ms enzima. La incapacidad para aumentar la velocidad de reaccin ms all de un valor finito mximo a concentraciones de sustrato cada vez ms altas, se le llama saturacin.
Cal es el significado de Vmax y Km? Km es la concentracin de sustrato para la cual la reaccin tiene lugar a la mitad de su velocidad mxima. Se nombra constante de Michaelis. El nmero de recambio representa la velocidad a la cual las molculas de sustrato se convierten en producto por una nica molcula de enzima, cuando sta est funcionando a su mxima velocidad. Los inhibidores enzimticos actan irreversible o reversiblemente Las enzimas estn influidas por productos, sustratos alternativos, sustratos anlogos, drogas, toxinas y una clase importante de reguladores llamados efectores alostricos. Muchas de estas sustancias tienen un efecto inhibidor en la actividad enzimtica, reduciendo (o incluso anulando) la velocidad de reaccin con el sustrato deseado. La inhibicin enzimtica desempea un papel vital como mecanismo de control de las clulas, tambin es importante en la accin de drogas y toxinas, las cuales ejercen frecuentemente sus efectos inhibiendo enzimas especficas. L
Los inhibidores pueden ser reversibles o irreversibles. Un inhibidor irreversible se une a la enzima covalentemente, causando una prdida irrevocable de la actividad cataltica. Los inhibidores irreversibles suelen ser txicos. Un inhibidor reversible se una a una enzima de forma disociable, no covalente, de manera que las formas libres y unidas del inhibidor existen en equilibrio las unas con las otras. Las dos formas ms comunes de inhibidores reversibles se clasifican como inhibidores competitivos (se une al sitio activo de la enzima y por lo tanto compite directamente con las molculas de sustrato por el mismo sitio de la enzima, reduciendo su actividad, hasta el punto de que los sitios activos de las molculas de enzima pueden tener unidas, en cualquier momento, molculas de inhibidor, en lugar de molculas del sustrato). Tambin estn los inhibidores no competitivos, este se une a la superficie de la enzima en una posicin diferente del sitio activo, por lo tanto no bloquea la unin del sustrato pero sin embargo inhibe la actividad de la enzima porque la unin del inhibidor con su sitio especfico, reduce enormemente e incluso suprime la actividad cataltica en el sitio activo. Regulacin enzimtica Algunos mecanismos reguladores son los cambios en las concentraciones de sustrato (y producto), alteraciones en el pH, y la presencia de inhibidores. La regulacin que depende directamente de las interacciones entre los sustratos y los productos con la enzima, se llama regulacin por el sustrato, aqu un incremento en la cantidad de sustrato produce un aumento en la velocidad de reaccin, por el contrario un incremento en la concentracin de producto reduce la velocidad a la cual el sustrato se convierte en producto. Tambin existen otros medios de regulacin dos de los ms importantes son: la regulacin alostrica y la modificacin covalente. Estos mecanismos permiten a las clulas conectar o desconectar a las enzimas o ajustar sus velocidades de reaccin, modulando apropiadamente las actividades de las enzimas. Las enzimas alostricas se regulan por molculas diferentes de los reactivos y productos La regulacin alostrica es el nico mecanismo importante de control, por el cual la tasa de las reacciones catalizadas enzimaticamente, se ajusta a las necesidades celulares. Retroinhibicin La regulacin deseada es posible porque el producto P es un inhibidor especfico de E1, la enzima que cataliza la primera reaccin en secuencia. Este fenmeno es llamado retroinhibicin (O inhibicin por el producto final) se representa por la flecha de lneas discontinuas que conecta el producto P con la enzima E1. Una retroinhibicin sucede cada vez que un producto metablico inhibe a una de las enzimas implicada en la va por la cual ese producto se sintetiza. La retroinhibicin es comnmente usada por la clula para asegurar que las actividades de las secuencias de reaccin se ajustan a las necesidades celulares. Regulacin alostrica
Todas las enzimas con capacidad de regulacin alostrica, pueden existir en dos estados diferentes. En una de las dos formas, la enzima tiene una afinidad alta por su(s) sustrato(s), mientras que en la otra, tiene poca o ninguna afinidad. Las enzimas con esta propiedad se llaman enzimas alostricas. Las dos formas diferentes de una enzima alostrica son fcilmente interconvertibles y estn de hecho, en equilibrio una con otra. La velocidad de reaccin es alta cuando la enzima est en la forma de alta afinidad y baja, o incluso nula, cuando la enzima est en su forma de baja afinidad. El que se favorezca la forma activa o inactiva de una enzima alostrica depende de la concentracin celular de la sustancia reguladora apropiada, llamada efector alostrico. Un efector alostrico es una pequea molcula orgnica que regula la actividad de una enzima, para lo cual no es ni el sustrato, ni el producto inmediato. Un efector alostrico influye en la actividad de la enzima unindose a una de las dos formas intercovertibles de sta, estabilizndola en ese estado. El efector se une a la enzima debido a la presencia en la superficie de est en un sitio alostrico (o regulador), distinto al sitio activo. Un efector puede ser o bien un inhibidor alostrico o un activador alostrico, dependiendo del efecto que este tenga. Los sitios activos y los sitios alostricos estn normalmente en diferentes subunidades de la protena, a las que denominamos subunidades catalticas y subunidades reguladoras. Las enzimas alostricas manifiestan interacciones cooperativas entre subunidades Muchas enzimas alostricas poseen cooperatividad lo que significa que a medida que los mltiples sitios catalticos unen molculas de sustrato, la enzima sufre cambios conformacionales, que afectan a la afinidad de los restantes sitios de unin del sustrato. Algunas enzimas muestran cooperatividad positiva, en la cual la unin de la molcula sustrato a una subunidad cataltica incrementa la afinidad de otras subunidades catalticas por el sustrato. Otras enzimas muestran cooperatividad negativa, en la cual el sustrato unido a una subunidad cataltica reduce la afinad de otros sitios catalticos con el sustrato. La cooperatividad positiva hace que la actividad cataltica de la enzima se incremente ms rpido de lo normal a medida que la concentracin del sustrato aumenta, mientras que la cooperatividad negativa significa que la actividad enzimtica aumenta ms lentamente de lo esperado. Las enzimas tambin se pueden regular por la adicin o eliminacin de grupos qumicos Muchas enzimas estn sometidas a control mediante modificaciones covalentes, vindose afectada por la adicin o eliminacin de grupos qumicos especficos. Las modificaciones comunes incluyen la adicin de grupos fosfato, grupos metilo, grupos acetilo, o derivados de nucletidos. Algunas modificaciones son reversibles y otras no. En cada caso, el efecto de la modificacin es la activacin o la inactivacin de la enzima, o por lo menos la modulacin, hacia arriba o hacia abajo, de su actividad. Fosforilacin / desfosforilacin
La adicin de grupos fosfato es una de las modificaciones ms frecuentes y mejor entendidas. La adicin de fosfato, fosforilacin, suele producirse por transferencia de un grupo fosfato desde el ATP al grupo hidroxilo de un residuo de serina, treonina o tirosina de la protena enzimtica. Las enzimas que catalizan la fosforilacin de otras enzimas (o de otras protenas) se llaman proten-quinasas. El proceso inverso, la desfosforilacin est catalizado por enzimas llamadas proten-fosfatasas e implica la eliminacin de un grupo fosfato desde la protena fosforilada. Ruptura proteoltica Un tipo diferente de activacin covalente de enzimas consiste en la eliminacin irreversible de un fragmento de la cadena polipeptdica, mediante una enzima proteoltica (que degrada protenas) adecuada. Este tipo de modificacin, llamado ruptura proteoltica, es el que tiene lugar en las enzimas proteolticas del pncreas, tripsina, quimiotripsina y carboxipeptidasa. Ribozimas: molculas de RNA con actividad enzimtica Los catalizadores constituidos por ARN son denominados ribozimas.
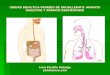
Membranas : estructura, qumica y funcinToda clula posee membrana y esta define los lmites de la clula y sus compartimientos internos. Las funciones de las membranas Funciones de la membrana: 1. Definen los lmites de la clula y limitan sus compartimentos. 2. Sirve como sitio concreto donde se realizan funciones especficas. 3. Posee protenas de transporte que facilitan y regulan el movimiento de sustancias hacia el interior y hacia el exterior de la clula y de sus compartimentos. 4. Contienen los receptores necesarios para detectar seales externas. 5. proporcionan los mecanismos para la comunicacin intercelular. 1. Las membranas permeabilidad definen lmites y sirven como barreras de
La barrera de permeabilidad de una clula en su conjunto es la membrana plasmtica ( o celular, una membrana que rodea a la clula y regula el paso de materiales hacia dentro y fuera de las clulas. Adems de esta hay varias membranas intercelulares que sirve para compartimentalizar funciones dentro de las clulas eucariotas. 2. En las membranas se sitan protenas especficas y por tanto son los lugares donde se realizan funciones especficas
3.
Las protenas de membrana regulan el transporte de solutos
-Las protenas de membrana realizan y regulan el transporte de sustancias fuera y dentro de la clula y sus organulos. -Muchas sustancias se mueven a travs de la membrana en direccin dictada por su gradiente de concentracin. -El movimiento de un in est determinado por su potencial electroqumico (gradiente de concentracin + gradiente de carga) -Este proceso de transporte no requiere energa ya que es favor del gradiente, ocurre de dos maneras diferentes: por difusin simple (agua, oxgeno, etanol molculas pequeas-) o por difusin facilitada (molculas ms grandes, ej. Azcares) en esta se mueven ayudadas por protenas transportadoras. -Alternativamente las molculas se pueden transportar en contra de su gradiente en un proceso llamado transporte activo, este requiere energa la cual es obtenida por la hidrlisis de ATP o un compuesto similar de alta energa (transporte activo directo) La energa puede ser proporcionada acoplando el transporte en contra de gradiente del soluto al transporte a favor del gradiente de los iones de sodio o de protones a travs de la misma membrana, a este proceso se le llama transporte activo indirecto. Protenas hacia la clula endocitosis Protenas fuera de la clula exocitosis 4. Las protenas de membrana detectan y transmiten seales elctricas y qumicas Transduccin de seales: consiste tanto en la deteccin de seales especficas en la superficie externa de las clulas como los mecanismos usados para transmitir tales seales a interior celular. Las clulas reciben informacin del entorno en forma de seales elctricas o qumicas. En la transduccin de seales qumicas, algunas molculas seal entran directamente en las clulas y actan en su interior. En la mayora de los casos las molculas sealizadoras que afectan a una membrana, se unen a protenas especficas de la superficie externa de la membrana plasmtica, denominadas receptores. A la unin de estas sustancias se les llama ligandos. Las seales internas generadas se denominan segundos mensajeros. 5. Las protenas de membrana median en la adhesin celular y la comunicacin clula-clula Comunicacin intercelular: es cuando una clula esta conectada a otra e intercambian componentes. Estas son proporcionadas por uniones gap (clulas animales) y plasmodesmos (clulas vegetales) Modelos de la experimental estructura de la membrana: una perspectiva
Mosaico fluido: es un modelo que sirve para describir a todas las membranas biolgicas, este imagina a la membrana como dos capas de lpidos bastante fluidas con proteinas localizadas dentro y sobre capas lipdicas y orientadas de forma especfica con respecto a las dos superficies de membrana. Overton y Langmuir: los lpidos son componentes importantes de la membrana
Los compuestos lipofdicos penetran con facilidad la membrana. Overton Los fosfolpodos son afipticos y su cabeza polar (hidroflicas) se encuentran en contacto con el agua, mientras que sus colas no polares (hidrofbicas) sobresalen del agua. Langmuir Porter y Grendel: la base de la estructura de la membrana es una bicapa lipdica Hipotetizaron la existencia de una bicapa donde las cadenas hidrocarbonadas no polares e hidrofbicas estubieran hacia el interior, fuera del medio acuoso y los grupos polares hidroflicos de cada capa estaran dirigidos hacia el exterior. Davson y Danielli: las membranas tambin contienen protenas Davson y Danielli propusieron (notese que esta teora fue desechada NO es la correcta) que las membranas biolgicas consisten en una bicapa lipdica que est recubierta en ambos lados con finas lminas de protenas (modelo de sndwich protena-lpido-protena) Luego se postulo el que las protenas hidroflicas podan atravesar la membrana y formar poros polares, esas protenas podan cambiar la permeabilidad y las propiedades de resistividad de la membrana. El interior lipidito es responsable de las propiedades hidrofbicas de la membrana y los componentes proteicos explican sus propiedades hidroflicas. Robertson: todas las membranas comparten una estructura subyacente comn La membrana tiene un aspecto trilaminar o de via de ferrocarril. Unidad de membrana: todas las membranas celulares comparten una estructura subyacente comn. Una investigacin ms detallada revel los principales defectos del modelo de Davson y Danielli Mediante diversos estudios se llego a la conclusin de que las protenas se encontraban al menos en parte, en el interior hidrofbico de la membrana. Tambin se indica que las membranas son estructuras fluidas en las que la mayora de lpidos y muchas protenas se mueven libremente. Singer y Nicholson: una membrana consiste en un mosaico de protenas dentro de una bicapa lipdica fluida (este si es cierto SI es correcto) El modelo de mosaico fluido tiene dos caractersticas principales, las cuales aparecen el nombre. El modelo imagina una membrana como un mosaico de protenas incluidas, o por lo menos unidas, de forma discontinua a una bicapa lipdica fluida. Singer- Nicholson beian las protenas como entidades globulares discretas que se asocian a la membrana basndose en su afinidad relativa por el interior hidrofbico de bicapa lipdica. Los tres tipos de protenas presentes en la membrana son: integrales, perifricas y ancladas a lpidos.
-Las protenas integrales de la membrana estn embebidas en la bicapa lipdica y se mantienen en su sitio por la afinidad de los segmentos hidrofbicos de la protena por el interior hidrofbico de la bicapa lipdica. -Las protenas perifricas, son mucho ms hidroflicas y por ello estn localizadas en la superficie de la membrana y estn ancladas de forma no covalente a los grupos polares de las cabezas de los fosfolpidos o a las partes hidroflicas de otras protenas. -Finalmente las protenas ancladas a lpidos son esencialmente hidroflicas y por ello residen en la superficie de la membrana, pero estn unidas covalentemente a molculas de lpido que estn incluidas en la bicapa. Unwin y Henderson: la mayora de las protenas de membrana contienen segmentos transmembrana Una propiedad importante de las protenas integrales es que tienen en su estructura primaria una o ms secuencias hidrofbicas que abarcan la bicapa lipdica, a esto se le llama segmentos transmembrana y anclan las protenas a la membrana y las sostienen alineadas correctamente dentro de la bicapa lipdica. Los lpidos de membrana: la parte fluida del modelo Las membranas contienen varias de las principales clases de lpidos Las principales clases de lpidos de la membrana son: fosfolpidos, glicolpidos y esteroles. Fosfolpidos: son los que se encuentran en mayor cantidad en la membrana, contienen muchas clases de fosfolpidos distintas, entre ellas los fosfoglicridos (basados en glicerol) y los esfingolpidos (basados en esfingosina). Los fosfoglicridos ms comunes son: fosfatidilcolina, fosfatidiletanolamina, fosfatidilserina y fosfatidilinositol. El esfingolpido ms comun es la esfingomielina. Glicolpidos: estos se forman al aadir a los lpidos grupos de carbohidrato. Algunos tienen como base glicerol, pero la mayora derivan de la esfingosina y por tanto se llaman glicoesfingolpidos. Ejemplos: cerebrsidos (denominados neutros ya que cada molcula tiene un azocar no cargado como grupo principal) y glanglisidos ( siempe tienen un grupo principal oligosacrido que contiene uno o ms residuos de cido silico cargados negativamente). Los cerebrsidos y ganglisidos predominan especialmente en las membranas cerebrales y clulas nerviosas. Esteroles: el principal esterol de las membranas celulares animales es el colesterol, las vegetales contienen colesterol ( poco) y grandes cantidades de fitoesteroles (campesterol, sitosterol y estigmasterol). Membranas de procariotas como bacterias y cianobacterias contienen algo similar a los esteroles denominado haponoides, que sustituye al esterol en la membrana. El esterol solamente est en la membrana plasmtica, no en membranas de organulos. La cromatografa en capa fina es una tcnica importante para el anlisis de los lpidos
La TLC es una tcnica que separa diferentes clases de lpidos basndose en sus afinidades relativas por una fase estacionaria hidroflica y una fase mvil hidrofbica La fase estacionaria suele ser una capa fina de cido silcico sobre una placa de vidrio o placa metlica, mientras que la fase mvil es una mezcla de solventes apropiados. Los cidos grasos son esenciales en la estructura y funcin de la membrana Los cidos grasos son componentes de todos los lpidos de la membrana excepto de los esteroles. Son esenciales para la estructura de la membrana debido a que sus largas colas hidrocarbonadas forman una barrera hidrofbica efectiva para la difusin de solutos polares.
Asimetra de membrana: la mayora de los lpidos se distribuyen de manera desigual entre las dos monocapas La mayora de los lpidos se distribuyen de forma desigual entre las dos monocapas. Esta asimetra de la membrana incluye diferencias tanto en las clases de lpidos presentes como en el grado de instauracin de los cidos grasos de las molculas de fosfolpidos. Movimientos que puede tener un fosfolpido en la membrana: -Difusin transversa -Rotacin -Difusin lateral *La bicapa lipdica es fluida. *Las membranas funcionan correctamente slo en estado fluido. -Temperatura de transicin -Transicin de fase *La fluidez de la membrana depende principalmente de las clases de lpidos que contiene. Hay dos propiedades especialmente importantes para determinar la fluidez: la longitud de las cadenas laterales de los cidos grasos y su grado de instauracin (es decir, el nmero de dobles enlaces presentes) *Las membranas con cadenas largas tienden a ser menos fluidas. Cadena larga saturados - (mayor temperatura de transicin) Cadena corta insaturados - (menor temperatura de transicin) Los efectos de los esteroles sobre la fluidez de la membrana Colesterol (clulas animales) Esteroles ----- Fitoesteroles (clulas vegetales) Intercalar molculas rgidas de colesterol en la membrana de una clula animal hace que a temperaturas ms elevadas la membrana sea menos fluida de lo que debera ser. Sin embargo, el colesterol tambin evita de manera eficaz que las cadenas hidrocarbonadas de los fosfolpidos encajen a medida que la temperatura va disminuyendo, reduciendo de este moedo
la tendecia de las membranas a melificarse al enfriarse. El colesterol disminuye la fluidez de la membrana a temperaturas por encima de la Tm y la aumenta a temperaturas por debajo de la Tm. Adems, una bicapa lipdica que contiene esteroles es menos permeable a los iones y a molculas pequeas que una bicapa que carece de esteroles. La mayora de los organismos pueden regular la fluidez de la membrana Los organismos procariotas y eucariotas pueden regular la fluidez de la membrana principalmente cambiando la composicin lipdica de las mismas. La capacidad de hacer esto se denomina adaptacin homeosviscosa, debido a que el efecto principal de esa regulacin es mantener la viscosidad de la membrana aproximadamente igual a pesar de las variaciones de temperatura. Las balsas lipdicas son regiones concretas de lpidos de membrana que estn implicadas en la sealizacin celular Las regiones de lpidos de membrana que secuestran protenas implicadas en la sealizacin celular se denominan microdominios o balsas lipdicas, estas se caracterizan por niveles elevados de colesterol y glicoesfingolpidos y por ello son ms espesas y menos fluidas que el resto de la membrana. Las caveolas son consideradas sub grupos de las balsas lipdicas y son pequeas invaginaciones de la membrana con forma de frasco. Protenas de membrana: la parte del modelo Los principales componentes del mosaico de la membrana son las protenas Las membranas contienen protenas perifricas y protenas ancladas a lpidos Protenas integrales de membrana Estas son protenas anfipticas que poseen una o ms regiones hidrofbicas que exhiben afinidad por el interior hidrofbico de la bicapa lipdica, por ello estn incluidas en el interior de la membrana y no se pueden extraer con facilidad. A su vez las protenas integrales pueden ser monotpicas, cuando solamente sobresalen por uno de los lados de la bicapa (muy pocas), o pueden ser protenas transmembrana (la mayora), lo que significa que cruzan la membrana y tienen regiones hidroflicas que sobresalen de la membrana por los dos lados. Las protenas transmembrana atraviesan la protena una vez (protenas de paso nico), o varias veces (protenas de paso mltiple). Algunas protenas de paso mltiple consisten en un nico polipptido, sin embargo cuando tienen dos o ms polipptidos se denominan protenas multisubunidades. La mayora de las protenas transmembrana estn ancladas a la bicapa lipdica por uno o ms segmentos transmembrana hidrofbicos, uno por cada vez que la protena atraviesa la bicapa. Protenas perifricas de la membrana integrales, protenas
Son las protenas que carecen de secuencias hidrofbicas diferenciadas y por tanto no penetran en la bicapa lipdica, en vez de ello se unen mediante fuerzas electrostticas dbiles y puentes de hidrgeno a la superficie de la membrana a travs de las porciones hidroflicas de las protenas integrales y quizs a travs de los grupos polares de las cabezas de los lpidos de la membrana. Estas se extraen ms fcilmente que las integrales y normalmente se extraen cambiando el pH o la fuerza inica. Protenas de membrana ancladas a lpidos Son las protenas cuyas cadenas polipeptdicas se localizan en una de las superficies de la bicapa lipdica, pero estn unidas covalentemente a molculas lipdicas incluidas dentro de la bicapa. Las protenas unidas a la superficie interna de la membrana se anclan por medio de formacin de enlaces covalentes con un cido graso o con un derivado isopreno denominado grupo prenil. En el caso de las protenas de membrana ancladas a cidos grasos, la protena se sintetiza en el citosol y despus se une covalentemente a un cido graso saturado incluido dentro de la bicapa. Por otro lado las protenas de membrana preniladas se sintetizan en el citosol como protenas solubles despus se les aade un grupo prenil y este se inserta en la capa lipdica de la membrana. Muchas protenas ancladas a lpidos y que se fijan a la superficie externa de la membrana se unen covalentemente a glicosilfofatidilinositol (GPI), estas son denominadas protenas de membrana ancladas GPI Las protenas se pueden separar por electroforesis en geles de poliacrilamida con SDS Un mtodo de separar las molculas para su estudio es la electroforesis la cual consiste en utilizar un campo elctrico para separar molculas cargadas elctricamente. La velocidad a la cual una molcula dada se mueve durante la electroforesis depende tanto de su carga como de su tamao. Previo a su estudio los fragmentos de membrana se solubilizan con el detergente SDS para desorganizar las uniones protena-protena y protena- lpido, las protenas se desnaturalizan y no se vuelven a plegar ya que estn cubiertas con molculas de turgente cargado negativamente. Resulta cada vez ms viable determinar tridimensional de las protenas de membrana la estructura
Las protenas integrales se han logrado estudiar por medio de la cristalografa de rayos X, la cual determina la estructura de las protenas que se puedan aislar de forma cristalina y por anlisis hidroptico aquellas que por lo menos se puedan aislar y secuenciar. La cristalografa de rayos X es utilizada para determinar la estructura tridimensional de las protenas. Por otro lado es posible inferir el nmero y posicin de los segmentos transmembrana a partir de una representacin de hidropata. Segn los valores de hidrofobicidad se calcula un ndice de hidropata, el cal es una medida numrica de la hidrofobicidad relativa de segmentos sucesivos de la cadena del polipptido. Las protenas de membrana tienen varias funciones
Algunas de las protenas de la membrana son enzimas , otras protenas de membrana son receptores implicados en el reconocimiento y mediacin de los efectos de seales qumicas especficas que se unen a la superficie de la clula. Las protenas de membrana tambin estn implicadas en la comunicacin intercelular. Las protenas de membrana desempean un papel principal, en la captacin y secrecin de sustancias por endositos y exocitosis; en el marcaje, clasificacin y modificacin de proteinas en el retculo endoplsmico. Finalmente las protenas poseen un papel estructural ya que estabilizan y dan forma a la membrana celular. Las protenas de membrana estn orientadas asimtricamente a travs de la bicapa lipdica Muchas protenas de membrana estn glicosiladas La mayora de membranas contienen pequeas cantidades de carbohidratos. Los glicolpidos representan una pequea porcin de los carbohidratos de membrana ya que la mayora estn en forma de glicoprotenas, protenas de membrana con cadenas de carbohidratos unidas covalmenteme a cadenas laterales de aminocidos. La adicin de una cadena lateral hidrocarbonada a una protena se denomina glicosilacin. (adicin de cadena de carbohidrato). Las glicoprotenas predominan en las membranas plasmticas, donde desempean un papel importante en el reconocimiento intercelular. Consecuentes con este papel, las glicoprotenas estn siempre situadas de tal forma que los grupos carbohidrato sobresalen de la superficie externa de la membrana celular. Esta disposicin contribuye a la asimetra de la membrana. En muchas clulas animales, los grupos carbohidrato de las glicoprotenas y de los glicolpidos sobresalen de la superficie celular y forman una cubierta de superficie denominada glicoclix. Las protenas de membrana varan en cuanto a su movilidad Las protenas de membrana son de hecho ms viables al movimiento que los lpidos, Evidencias experimentales de la movilidad proteica Un mtodo para identificar la movilidad proteica es el de aticuerpos fluorescentes donde se marcan dos tipos de protenas, seguidamente se unen y se observa el movimiento.
Transporte a travs de la membrana: saltando la barrera de permeabilidadLa naturaleza hidrfoba de las membranas las hace impermeables a la mayora de molculas y iones, manteniendo unas sustancias en el interior celular y otras en el exterior. Sin embargo las membranas no son slo barreras que restringen indiscriminadamente la entrada y saluda de sustancias, sino que poseen una permeabilidad selectiva, que permite el intercambio controlado de molculas y iones especficos.
Las clulas y el proceso de transporte Los procesos de transporte en masa que permiten el movimiento de sustancias dentro y fuera de la membrana son la endocitosis (dentro) y la exocitosis (fuera). La mayora de sustancias que atraviesan la membrana no so macromolculas, sino iones y pequeas molculas orgnicas en disolucin (solutos, mayormente Na, K, Ca2, y Cl). La mayora de molculas orgnicas de pequeo tamao que son transportadas son metabolitos sustratos, productos intermedios y finales de las rutas metablicas que se verifican en la clula o sus organulos. El transporte es la capacidad de mover selectivamente iones y molculas orgnicas a travs de una membrana, un proceso bsico en la funcin celular. Los solutos cruzan la membrana por difusin simple, difusin facilitada o transporte activo Las molculas pequeas como oxgeno, dixido de carbono y etanol, atraviesan la membrana por difusin simple, que es el movimiento de solutos a travs de la bicapa lipdica en la direccin dictada por la diferencia de concentracin del soluto entre ambos lados de la membrana (de mayor a menor). Sin embargo la mayora de solutos se mueven gracias a la presencia de protenas transportadoras (protenas integrales que reconocen sustancias con alta especificad). En algunos casos las protenas de transporte permiten la difusin facilitada, movindolos a favor del gradiente de energa, cuando las protenas permiten el transporte en contra del gradiente se denomina transporte activo. El movimiento de solutos a travs de la membrana est determinado por su gradiente de concentraciones o su potencial electroqumico El movimiento de una molcula depende de su gradiente de concentracin, de dicha molcula a ambos lados de la membrana, el movimiento ser a favor del gradiente. El movimiento de un ion, depende de su potencial electroqumico, el cual resulta de la integracin de los gradientes de concentracin y de carga de ambos lados de la membrana. Difusin simple: movimiento no asistido a favor de gradiente La forma ms sencilla por la que un soluto puede pasar de un lado a otro de la membrana es la difusin simple, movimiento neto no asistido de un soluto desde una regin donde su concentracin es mayor, a otra donde sta es menor. Este movimiento es solo para molculas pequeas y relativamente poco polares. (ej: O, CO2) La difusin mueve a los solutos hasta alcanzar el equilibrio Independientemente del punto de inicio, la difusin tiende a crear una solucin en la cual la concentracin es la misma en ambas partes, alcanzando as el equilibrio. La difusin es un movimiento que conduce siempre al equilibrio, a la vez disminuye la energa libre ya que las
molculas se mueven a favor del gradiente, por ello la difusin progresa siempre de regiones de mayor a menor energa libre. smosis en la difusin de agua a travs de una membrana con permeabilidad selectiva El agua tiende a moverse del lado donde hay menor concentracin de solutos a la parte con mayor concentracin de solutos, este movimiento en respuesta a diferencias de concentracin de solutos se llama smosis. La difusin simple est limitada a molculas pequeas no polares nicamente los solutos pequeos pueden pasar por difusin simple a travs de la bicapa lipdica, los tres factores que afectan sustancialmente la difusin de solutos son: el tamao, la polaridad y la carga (si el soluto es ion) Tamao del soluto En trminos generales, las bicapas lipdicas son ms permeables a las molculas pequeas que a las grandes. Las molculas pequeas ms relevantes para la funcin celular son el agua, el oxgeno y el dixido de carbono. La permeabilidad de la bicapa es tanto mayor cuanto menor sea la molcula. Polaridad del soluto Las bicapas lipdicas son relativamente permeables a molculas no polares y menos permeables a molculas polares. Esto ya que las no polares se disuelven ms fcilmente en la fase hidrfoba de la bicapa y as la atraviesan ms fcilmente. Permeabilidad a iones La membrana es altamente impermeable a los iones, esto es debido a su fuerte asociacin con las molculas de agua, que forman un escudo de hidratacin. La tasa de difusin simple gradiente de concentraciones es directamente proporcional al
Termodinmicamente la difusin simple es un proceso exergnico y por ello no requiere energa. Las molculas individuales difunden al azar en ambos sentidos, pero el flujo neto es siempre en la direccin de la menor energa libre, esto significa a favor de la gradiente de concentracin, por ello la tasa neta de transporte es proporcional a la diferencia de concentraciones. Esta es relevante para molculas como el etanol y el O2.
Difusin facilitada: movimiento a favor del gradiente, asistido por protenas
La mayora de sustancias en la clula son demasiado grandes o demasiado polares como para atravesar la membrana por difusin simple, estas sustancias entran y salen de la clula gracias a protenas transportadoras, que intervienen en el movimiento de solutos a travs de la membrana. Por ello la funcin de las protenas transportadoras es meramente facilitar la difusin a favor de sustancias polares o con carga, que de otra forma no podran pasar. (ej. Glucosa) Las protenas transportadoras y los canales proteicos facilitan la difusin empleando mecanismos diferentes Las protenas que facilitan el paso por difusin facilitada son protenas integrales con uno o varios segmentos transmembrana. Estas protenas se clasifican en: transportadores proteicos: tambin denominados permeasas, son las que captan una o ms molculas de soluto en un lado de la membrana, seguidamente sufren un cambio conformacional que permite la transferencia del soluto hacia el otro lado de la membrana. Tambin estn los canales proteicos, los cuales constituyen canales hidrfilos que atraviesan la membrana y permiten el paso de solutos sin necesidad de grandes cambios conformacionales. Son una especie de poros, y son relativamente grandes, estos porros estn formados por protenas transmembrana denominadas porinas, por otro lado la mayora de estos canales son muy pequeos y selectivos y como intervienen en el transporte de iones se denominan canales inicos. Las protenas transportadoras conformacionales alternan entre dos estados
El modelo de conformacin alternativa hace referencia a como una protena alterna entre los dos estados conformacionales de estar abierto primero a un lado de la membrana y luego al otro. Las protenas transportadoras son anlogas a las enzimas, por su especificad y cintica Las protenas transportadoras son denominadas a menudo como permeasas. Como en las reacciones catalizadas por enzimas, la difusin facilitada implica la unin inicial del sustrato (soluto a transportar) a u lugar especfico de la superficie de la protena (el lugar de unin del soluto). Tras la formacin de este complejo se liberar el producto (soluto transportado). Especificidad de las protenas de Transporte Las permeasas poseen especificidad, al igual que las enzimas. Cintica de la funcin de las protenas de transporte El transporte facilitado al igual que la catlisis enzimtica, exhibe una cintica de saturacin, con una velocidad mxima y una constante Km que corresponde a la concentracin de soluto necesario para alcanzar la mitad de la tasa mxima de transporte. Una ltima semejanza entre las protenas y las enzimas es la inhibicin competitiva.
Las protenas transportadoras movilizan uno o dos solutos Cuando una permeasa transporta un solo soluto, el proceso se denomina uniporte, cuando se transportan dos solutos simultneamente y de manera acoplada, es decir, que no se produce el movimiento si uno de los dos solutos faltas, se llama transporte acoplado,. El transporte acoplado puede ser simporte (o contrasporte) cuando atraviesan la membrana en el mismo sentido, o antiporte cuando la atraviesan en sentidos opuestos. El transportador de glucosa del eritrocito y el intercambiador de aniones son ejemplos de protenas transportadoras El transportador de glucosa (GLUT1), es una transportador uniporte. El intercambiador de aniones del eritrocito (o intercambiador de clorurobicarbonato) es un transportador de antiporte. Los canales proteicos facilitan la difusin, hidrfilos en la membrana formando tneles
Los canales proteicos permiten a los iones desplazarse libremente a travs de la membrana. Los tres tipos de canales proteicos transmembranales son: canales inicos, porinas y acuaporinas. Canales inicos: protenas transmembrana que permiten el paso rpido de determinados iones Los canales inicos son muy selectivos, la mayora permite el paso de una sola especie inica y por ello son necesarios diferentes canales para el transporte de iones. La mayora de los canales inicos estn regulados por lo que estos se abren y cierran en respuesta a determinados estmulos. En clulas animales existen tres tipos de estmulos que controlan la apertura y cierre de los canales regulados: canales operados por voltaje, que se abren (polarizado) y cierran (despolarizado) como respuesta a cambios en el potencial de membrana; canales operados por ligando, regulados por la unin al canal de determinadas sustancias extracelulares o intracelulares y canales mecanosensibles, que responden a fuerzas mecnicas que actan en la membrana. Porinas: protenas transmembranales que permiten el paso rpido de diversos solutos Estos poros son ms grandes que los de los canales inicos, estos poros se forman por protenas denominadas porinas, que atraviesan varias veces la membrana. Los segmentos transmembranales de las porinas son barriles B (beta). El poro permite el paso de varios solutos hidrfilos, cuyo paso queda nicamente restringido por el tamao del poro de la porina en particular. Acuaporinas: protenas transmembrana que permiten el paso rpido de agua
En las clulas de algunos tejidos, existe un movimiento especfico de agua, mediado por una familia de canales proteicos, denominados acuaporinas (AQPs). Las acuaporinas no son responsables del intercambio de todo el agua que pasa a travs de cualquier membrana, ms bien facilitan la entrada y salida rpida de molculas de agua en tejidos que as lo requieren. *El sodio y el potasio mantiene la polaridad de la membrana, por ello son importantes las bombas de sodio-potasio (consumen energa).
Transporte activo: movimiento en contra de gradiente, asistido por protenas El transporte activo es aquel que hace posible el movimiento de solutos en contra del equilibrio termodinmico, por lo que requiere siempre el aporte de energa. El transporte activo realiza tres funciones principales en las clulas y sus orgnulos. Primero, hace posible la toma de sustancias nutritivas esenciales del medio circundante Segundo, permite que sean eliminadas varias sustancias, como productos de secrecin y de desecho, incluso cuando la concentracin en el exterior es mayor que en el interior. Tercero, permite que la clula mantenga constantemente un desequilibrio en las concentraciones de ciertos iones inorgnicos (K, NA, CA2+, H). El resultado final del transporte activo es un desequilibrio en estado estacionario, sin el cual la vida no sera posible. Las protenas de membrana implicadas en el transporte activo suelen denominarse bombas. A diferencia de la difusin simple o facilitada, el transporte activo suele tener direccionalidad intrinseca, por lo que generalmente solo se mueve en un sentido. Por ello se dice que el transporte activo es unidireccional o vectorial. El acoplamiento del transporte activo a una fuente de energa, puede ser directo o indirecto El transporte activo puede ser: activo directo o activo indirecto. El transporte activo directo implica la existencia de un sistema de transporte acoplado a una reaccin qumica exergnica, generalmente la hidrlisis de ATP. Las protenas de transporte que utilizan la energa liberada
por la hidrlisis de ATP se llaman ATPasas de transporte o K bombas ATPsicas. Los cuatro tipos de ATPasas de trasnporte son: tipo P (phosphorylation/ se fosforilan por ATP), tipo V (vacuola/ bombean protones a organulos como las vacuolas, lisosomas, etc), tipo F (factor / utilizan la energa de la hidrlisis del ATP) y tipo ABC ( ATP binding cassette/ forman una superfamilia de protenas). El transporte activo indirecto implica la existencia de un sistema de transporte acoplado al transporte de iones, este puede ser simporte o antiporte. El transporte activo indirecto ( o secundario) es impulsado por el movimiento de iones a favor de su gradiente electroqumico.
Metabolismo de la energa quimiotrfica: gluclisis, fermentacinRutas Metablicas Cuando consideramos todas las reacciones qumicas que ocurren dentro de la clula, hablamos de metabolismo. El metabolismo global de una clula consiste, en definitiva, en muchas rutas metablicas especficas, cada una de ellas para una operacin concreta. La vida a nivel celular se puede definir como una red integrada de reacciones metablicas cuidadosamente acopladas en la que cada una de ellas contribuye a la suma de las actividades que la clula debe llevar a cabo. Las rutas metablicas son de dos tipos generales. Las rutas en las que se sintetizan componentes celulares se denominan rutas anablicas (hacia arriba), mientras que aquellas implicadas en la degradacin de constituyentes celulares se denominan rutas catablicas. ( hacia abajo). Las rutas anablicas implican un incremento del orden (una disminucin de entropa) y son endergnicas (requieren energa). Las rutas catablicas son exergnicas (liberan energa) se disminuye el orden molecular (aumenta la entropa). Las rutas catablicas tienen dos papeles en la clula: liberar la energa libre necesaria para las funciones celulares y generar las pequeas molculas orgnicas o metabolitos, que son los ladrillos de construccin para la biosntesis. Las reacciones catablicas pueden producirse en presencia de oxgeno (aerobio) o en ausencia de oxgeno (anaerobio), se produce mayor energa en las rutas aerobias. ATP: el acoplador universal de energa
Las reacciones anablicas se encargan del crecimiento y reparacin de las clulas, las reacciones catablicas producen la energa que las reacciones anablicas necesitan. Se necesita un acoplamiento entre estos dos procesos y eso se logra gracias a ciertos tipos de molculas que mantienen la energa obtenida en reacciones exergnicas y la liberan donde es requerida. La molcula ms comnmente utilizada como intermediario energtico es el compuesto fosforilado adenosina trifosfato (ATP), este esta implicado en la mayora de transiciones energticas. El ATP tiene dos enlaces fosfoanhdrido ricos en energa El ATP es una molcula que contiene la base aromtica adenina, el azcar de 5 carbonos ribosa y una cadena de 3 grupos fosfato. Los grupos fosfato estn conectados entre s por puentes fosfoanhdridos y la ribosa por medio de un enlace fosfoster. Dependiendo de si la adenosina (adenina + ribosa) esta unida a uno, dos o tres fosfatos se formara AMP, ADP o ATP. El ATP es un buen intermediario del metabolismo energtico celular ya que la hidrlisis del tercer enlace fosfato (el ms externo) libera energa. Hidrlisis es el trmino general para las reacciones en las que un enlace molecular se rompe por reaccin con el agua. Por ello la hidrlisis de ATP (reaccin con el agua para formar ADP y fosfato inorgnico) es exergnica. La hidrlisis de ATP es altamente exergnica debido a la repulsin electrosttica y a la estabilizacin por resonancia La hidrlisis de ATP y ADP en Pi (fosfato inorgnico HPO4-2 ) es exergnica debido a la repulsin electrosttica entre los grupos fosfato adyacentes cargados negativamente y la estabilizacin de los dos productos (ADP y Pi) por resonancia. La repulsin electrosttica hace referencia a como las cargas negativas de los grupos fosfato se repelen entre s liberando energa, a la vez en la sntesis de ATP se requiere energa para vencer esta repulsin. La estabilizacin por resonancia es la segunda contribucin importante al enlace energtico de ATP. Se debe entender que la estructura de los carboxilatos y los grupos fosfato es en realidad una media de todas las estructuras que se denomina hbrido de resonancia, en el que los electrones extra estn deslocalizados sobre todos los posibles enlaces. Cuando estos electrones presentan la mxima deslocalizacin, una molcula se encuentra en su configuracin ms estable (de mnima energa) y se dice que est estabilizada por resonancia. Si uno o ms tomos de oxgeno del carboxilato o el grupo fosfato participa en un enlace covalente deja de estar disponible para la deslocalizacin electrnica de manera que el resto de electrones estn ahora deslocalizados sobre un nmero de oxgenos inferior al mximo lo que supone que la molcula se encuentra en una configuracin de ms alta energa (menos deslocalizada). El ATP es un importante intermediario del metabolismo energtico celular
El ATP ocupa una posicin intermedia en el espectro global de compuestos fosforilados ricos en energa de la clula. El fosfoenol piruvato (PEP), el difosfoglicerato o la fosfocreatina liberan ms energa, mientras que la glucosa 1-6- fosfato y el glicerol fosfato liberan menos. Un compuesto puede fosforilar a un compuesto de mayor energa libre ( un nmero menos negativo), pero uno con menor energa libre no puede fosforilar a uno con mayor. Se obtiene ATP a partir de ADP si es fosforilado por PEP, pero no puede ser fosforilado por glucosa -6 fosfato. De igual forma el ATP puede fosforilar a la glucosa, pero no el piruvato. Las reacciones que implican el movimiento de un grupo qumico de una molcula a otra, se denominan reacciones de transferencia de grupo (ej. Grupo fosfato del piruvato al ADP). Estas reacciones son muy comunes en el metabolismo celular y la transferencia del grupo fosfato es uno de lso que se hacen con mayor frecuencia. El ATP por estar en un plano medio puede ser tanto donador como aceptor de fosfato. El par ADP/ATP representa una forma reversible de conservar, transferir y liberar energa dentro de la clula. Metabolismo energtico quimitrofo El metabolismo energtico quimitrofo cosiste en las rutas y reacciones mediante las cuales las clulas catabolizan nutrientes y conservan, en forma de ATP, parte de la energa libre que se libera en ellas. Las oxidaciones biolgicas normalmente implican la eliminacin de electrones y protones y son altamente exergnicas La oxidacin es la prdida de electrones, los nutrientes como los carbohidratos, grasas o protenas son fuente de energa para la clula lo que significa que son compuestos orgnicos oxidables y que su oxidacin es altamente exergnica. La oxidacin de molculas orgnicas normalmente implica la eliminacin, no slo de electrones, sino tambin de iones hidrgeno (protones), de manera que el proceso es tambin una deshidrogenacin. La mayora de las enzimas que catalizan reacciones oxidativas en las clulas son deshidrogenadas. Ninguna de las reacciones de oxidacin puede tener lugar de forma aislada; los electrones se transfieren a otra molcula, que queda reducida en el proceso. La reduccin es lo opuesto a la oxidacin y se defina como la ganancia de electrones. En las reducciones biolgicas, como e las oxidaciones, los electrones suelen ir acompaados de protones y lar reaccin global es, por tanto, una hidrogenacin. Las oxidaciones y las reducciones tienen lugar simultneamente. Las coenzimas como el NAD+ funcionan como aceptores de electrones en las oxidaciones biolgicas En el metabolismo quimitrofo de la energa el ltimo aceptor de electrones es normalmente, aunque no siempre, el oxgeno. En la mayora de las oxidaciones biolgicas el aceptor inmediato de electrones (y donador inmediato de electrones en la mayora de reducciones) es alguna coenzima. Las coenzimas son molculas pequeas que funcionan junto con
las enzimas, como transportadores de electrones o de pequeos grupos funcionales. Las coenzimas funcionan realmente como sustratos en las reacciones en las que participan, sin embargo no son consumidas, sino recicladas. La coenzima ms comn implicada en el metabolismo energtico es la nicotinamida adenina dinucletido (NAD+), el NAD+ sirve como aceptor de un electrn por incorporacin, a su anillo aromtico, de dos electrones y un protn, generando as la forma reducida, el NADPH y un protn. La mayora de los quimitrofos consiguen la energa necesaria para oxidacin de las molculas orgnicas de los alimentos Mediante la oxidacin los quimitrofos conseguimos la mayor parte de la energa que necesitamos. Hay muchas sustancias distintas que sirven como sustratos en las oxidaciones biolgicas, para nosotros y la mayora de quimitrofos dependemos de molculas orgnicas de los alimentos como sustratos oxidables, fundamentalmente carbohidratos, grasas y protenas. La glucosa es uno de los sustratos oxidables ms importantes del metabolismo enegtico La glucosa es un azcar de seis carbonos, y es un buen candidato para la oxidacin debido a que es el azcar mayoritario en la sangre, y por lo tanto fuente principal de energa para las clulas del cuerpo. La glucosa de la sangre procede principalmente de carbohidratos ingeridos, como la sacarosa o almidn o la degradacin de glucgeno. La oxidacin de la glucosa es altamente exergnica La glucosa es una buena fuente potencial de energa porque su oxidacin es un proceso altamente exergnico. Por ejemplo la glucosa, liberar la misma energa bajo combustin o si se cataboliza por el cuerpo. Sin embargo la distincin es fundamental: una combustin incontrolada ocurre a temperaturas que son incompatibles con la vida y la mayor parte de la energa libre se libera en forma de calor. Las oxidaciones biolgicas implican reacciones catalizadas enzimaticamente, que tienen lugar sin cambios significativos de temperatura, conservndose qumicamente la energa libre, en forma de ATP.
El catabolismo de la glucosa produce mucha ms energa en presencia que en ausencia de oxgeno La glucosa como sustrato oxidable asume la disponibilidad de algn tipo de aceptor de electrones (sin este no sera posible la oxidacin), el aceptor de electrones en la mayora de organismos es el oxgeno molecular. La oxidacin completa de la glucosa en dixido de carbono y agua, en presencia de oxgeno, se denomina respiracin aerobia. Algunos organismos, normalmente bacterias, pueden llevar a cabo la respiracin anaerobia, usando un aceptor final de electrones distinto del oxgeno. El proceso anaerbico para obtener energa se denomina fermentacin. En algunas clulas animales y en muchas bacterias, el producto final es el
lactato de manera que el proceso del cabolismo anaerobio de la glucosa se denomina fermentacin lctica. En la mayora de las clulas vegetales y en microorganismos como las levaduras, el proceso se denomina fermentacin alcohlica porque el producto final es el etanol, un alcohol. Basndose en sus requerimientos de oxgeno, los organismos aerbicos, anaerbicos o facultativos Los organismos clasificados segn sus requerimientos de oxgeno y su utilizacin como aceptor de electrones en el metabolismo energtico pueden ser aerobios estrictos (necesidad absoluta de oxgeno), anaerobios estrictos (no pueden usar oxgeno como aceptor bajo ninguna condicin ej. Bacterias) y finalmente los organismos facultativos (pueden desarrollarse tanto en condiciones aerobias como anaerobias ej. Moluscos y anlidos). Algunas clulas o tejidos de organismos aerobios tambin pueden, en ausencia o con escasez temporal de oxgeno, funcionar como facultativos si es necesario (ej. Clulas del msculo esqueltico). Gluclisis y fermentacin: produccin de ATP en ausencia de oxgeno La molcula de glucosa se degrada dando lugar a 2 molculas de tres tomos de carbono, cada una de las cuales es parcialmente oxidada por una secuencia de reacciones que es suficientemente exergnica para generar 2 molculas de ATP por cada molcula de glucosa fermentada. La gluclisis genera ATP El proceso de gluclisis, tambin llamado ruta glucoltica, es una secuencia de reacciones de diez pasos que convierte una molcula de glucosa en dos molculas de piruvato, un compuesto tricarbonado. La gluclisis es comn a los metabolismos aerobios y anaerobios. Un nombre alternativo es ruta Embden-Meyerhof. Visin general de la gluclisis La esencia del proceso es la separacin del azcar (glucosa) en dos molculas de tres tomos de carbono. Una de estas molculas, el gliceraldehdo-3-fosfato, es la nica molcula que se oxida en esta ruta. Algunas caractersticas importantes son la ruptura del azcar (Gly-4), la oxidacin, que genera NADH (Gly-6) y los dos pasos especficos en que esta secuencia de reacciones se acopla a la generacin de ATP (Gly-7 y Gly-10). La ruta general se considera en tres fases: preparacin y ruptura (Gly-1 a Gly-5); la secuencia oxidativa, que tambin genera ATP (Gly-6 y Gly-7); y la segunda reaccin de sntesis de ATP (Gly-8 a Gly-10) Fase 1: preparacin y ruptura El resultado neto de las tres primeras reacciones es la conversin de una molcula no fosforilada (glucosa) en una molcula doblemente fosforilada (fructosa-1, 6-bifosfato), esto se logra con la adicin de dos grupos fosfato, uno en cada carbono terminal.
Gly-1 = se fosforila el carbono 6, dando lugar a la glucosa-6-fosfato (aqu se usa un ATP que proporciona el grupo fosfato y la energa necesaria). El enlace formado por la fosforilacin de la glucosa es un enlace fosfoster y el enlace por el cual el fosfato terminal se une al ATP es un enlace fosfoanhdrido. Enzima utilizada hexoquinasa. Irreversible Gly-2 = El aldoazcar glucosa-6-fosfato se convierte en una cetoazcar, fructosa-6-fosfato, que tiene un grupo hidroxilo en el carbono 1. Enzima utilizada fosfoglucoisomerasa. Reversible Gly-3 = El grupo hidroxilo de la fructosa-6-fosfato es fosforilado, dando lugar a un azcar doblemente fosforilada, fructosa-1,6-difosfato (aqu se usa otro ATP, para la segunda fosforilacin). Enzima utilizada fosfofructoquinasa-1. Irreversible (esto debido a la diferencia de energa entre el enlace anhdrido del ATP y el enlace fosfoster de la molcula fructosa). Gly-4 = Se rompe la fructosa-1,6-difosfato dando lugar a dos triosas, llamadas dihidroxiacetona fosfato y gliceraldehdo-3-fosfato. La ruptura lo hace la enzima aldolasa. Gly-5= Estas dos triosas son interconvertibles. Ya que solo la gliceraldehdo3-fosfato se puede oxidar, se convierte a la dehidroxiacetona fosfato a gliceraldehdo-3-fosfato. Enzima utilizada triosa fosfato simerasa. Reversible Resumiendo: Glucosa + 2ATP 2-gliceraldehdo-3-fosfato + 2ADP
Fase 2: Oxidacin y sntesis de ATP Gly-6 = Se oxida el gliceraldehdo-3-fosfato a cido 3-fosfoglicerato, aqu se reduce NAD+ a NADH. Enzima utilizada gliceraldehdo-3-fosfato deshidrogenasa. Se libera una molcula de H (proton) y se dona un electrn al NAD+ produciendo NADH. Gly-7 = Se fosforila ADP con fosfato inorgnico Pi, se genera ATP (dos en total). Enzima utilizada fosfogliceroquinasa. La produccin de ATP, por la transferencia directa al ADP, de un grupo fosfato de alta energa procedente de un sustrato fosforilado como el 1,3-difosfoglicerato se denomina fosforilacin a nivel sustrato. Resumiendo: Gliceraldehdo-3-fosfato + NAD+ + ADP + Pi 3-fosfoglicerato + NADH + H + ATP (X2) Fase 3: Formacin de piruvato y generacin de ATP Gly-8 = El enlace fosfoster del grupo fosfato con el tomo de carbono se convierte en un enlace fosfoenol (proceso exergnico) este incremento de energa implica un reordenamiento de la energa interna de la molcula. El grupo fosfato de 3-fofoglicerato se traslada al tomo de carbono adyacente, formando 2-fosfoglicerato. Enzima utilizada fosfoglieromutasa. Reversible
Gly-9 = Se elimina una molcula de agua del 2-fosfoglicerato (aqu se usa la enzima enolasa) y se genera fosfoenolpiruvato (PEP), compuesto de alta energa. El PEP tiene una caracterstica fundamental de un enlace fosfato de alta energa: un grupo fosfato en un tomo de carbono que esta unido por un doble enlace a otro tomo, casi siempre carbono u oxgeno. El PEP puede encontrarse en dos formas interconvertibles (enol y ceto), estas dos se diferencian slo en la localizacin del doble enlace. Sin embargo el equilibrio favorece la forma ceto. Gly-10 = Ocurre hidrlisis en la PEP, esta transfiere su fosfato al ADP, dando lugar a otra molcula de ATP, quedando solamente como piruvato. Enzima utilizada piruvato quinasa. Irreversible. Resumiendo: 3-fosfoglicerato + ADP piruvato + ATP (X2)
Recordemos que en la primera fase se invierten dos molculas de ATP que luego son devueltas en la reaccin Gly-7, finalmente se generan dos molculas de ATP como ganancia neta en la Gly-10. Un resumen total y final sera: Glucosa + 2NAD + 2ADP + 2Pi 2H + 2ATP 2 piruvato + 2NADH +
El destino del piruvato depende de si el oxgeno se encuentra o no disponible El destino del piruvato depende de si hay o no oxgeno disponible, el presencia de oxgeno, el piruvato se oxida a otra molcula denominada acetil coenzima A, que sucesivamente, se puede oxidar completamente a CO2. As la ruta glucoltica se convierte en la primera de las componentes principales de la respiracin aerobia. Sin embargo en ausencia de oxgeno, en condiciones anaerobias, no se oxida el piruvato, no se forma acetil coenzima A, y no se genera ATP adicional. Las necesidades se ven satisfechas con el rendimiento de ATP en la ruta glucoltica, el piruvato se reduce aceptando los electrones (y protones) que se eliminan del NADH. Y como productos de la reduccin del piruvato se forma lactato o etanol y dixido de carbono. En ausencia de oxgeno, el piruvato entra en la ruta fermentativa y regenera NAD Generalmente, la ruta glucoltica acaba en la formacin del piruvato. En un proceso fermentatico, dada la necesidad de regenerar NAD, no termina aqu. En un proceso de la gluclisis, el NAD se reduce a NADH, sin embargo es necesaria la regeneracin de NAD. En precensia de oxgeno, el NADH es reoxidado por la transferencia de electrones (y protones) al oxgeno, sin condiciones anaerobias, los electrones se transfieren al piruvato redujendo su grupo carbonilo a un grupo hidroxilo. Las dos rutas de fermentacin ms
comunes utilizan el piruvato como aceptor final de electrones convirtindolo en lactato o etanol y CO2. Fermentacin del lactato Cuando el proceso anaerobio termina en la formacin de lactato, se denomina fermentacin lctica. El lactato se genera por la transferencia directa de electrones del NADH al grupo carbonilo del piruvato, reducindolo al grupo hidroxilo del lactato. En base a una molcula de glucosa, se puede representar como: 2 piruvato + 2 NADH + 2H 2 lactato + 2NAD Esta reaccin es reversible y es elaborada por la enzima lactato deshidrogenasa. La ecuacin global del metabolismo de la glucosa a lactato sera: Glucosa + 2ADP + 2Pi 2 lactato + 2ATP
La fermentacin del lactato es principal fuente energtica para la mayora de bacterias anaerobias y culas animales en condiciones anaerbicas o de hipoxia (carencia de oxgeno). Tambin es importante comercialmente ya que se produce queso, yogurt y otros productos que dependen de la fermentacin microbiana de la lactosa. Tambin se da luego de ejercicio intenso, nuestros msculos resiven menos oxigeno del que necesita y produce fermentacin lactica, el lactato producido es llevado al hgado donde se transforma de nuevo en glucosa por medio de la gluconeognesis proceso opuesto a la fermentacin lctica. Fermentacin alcohlica En condiciones anaerobias las clulas vegetales como levaduras y microorganismos llevan a cabo la fermentacin alcohlica. En este proceso el piruvato pierde un tomo de carbono (como CO2) para dar lugar al compuesto de dos carbonos acetaldehdo. Este se reduce por NADH y da lugar al etanol. Esta secuencia de reacciones est catalizada por dos enzimas, la piruvato descarboxilasa y la alcohol deshidrogenasa. La reaccin global se puede resumir como: 2 piruvato + 2NADH + 4H 2 etanol + 2CO2 + 2NAD
Como ecuacin global de la gluclisis para la fermentacin alcohlica tenemos: Glucosa + 2ADP + 2Pi + 2H2 etanol + 2CO2 + 2AT La fermentacin libera slo una pequea fraccin de la energa libre del sustrato pero conserva esa energa eficientemente como ATP Una caracterstica fundamental de todos los procesos fermentativos es que no hay un aceptror de electrones externo y no se da una oxidacin total. Sustratos alternativos para la gluclisis
A pesar de que la glucosa es el sustrato principal para los procesos de gluclisis, tambin hay otras alternativas que pueden ser utilizadas por la clula. De aqu surge un principio general: cualquiera que sea la naturaleza qumica del sustrato alternativo, ste se convierte en un intermediario principal de la ruta catablica de la glucosa, en el menor nmero de casos posible. El glicerol y otros azcares se catabolizan tambin por la ruta glucoltica
Gluconeognesis La gluconeognesis es la formacin de glucosa a partir de precursores tri y tetracarbonados. El material inicial ms comn es el piruvato, o el lactato, en el que se convierte el piruvato en condiciones anaerbicas. Los procesos similares a los de la gluclisis, a diferencia de tres pasos, el primero, el tercero y el dcimo, esto refleja un principio del metabolismo celular: las rutas biosintticas son, rara vez el reverso de las correspondientes rutas catablicas. La gluconeognesis tiene reacciones bypass que evitan estos tres pasos difciles de revertir. Regulacin de la glucolisis y gluconeogneis La regulacin de la glucolisis y de la gluconeognesis en las clulas animales implica a uno o dos de los principales mecanismos de control, la regulacin alostrica (afecta la actividad enzimtica y es un mecanismo intracelular) y la regulacin hormonal (suele ser una reaccin intercelular) Las enzimas clave en las rutas glucoltica y gluconeogentica estn sujetas a regulacin alostrica La regulacin alostrica implica la alternancia entre dos formas de una enzima, una de las cuales catalticamente ms activa que la otra. El que la enzima est en su forma activa o inactiva depender de que el efector alostrico se una al sitio alostrico y de si el efector es activador o inhibidor. Para la gluclisis las enzimas clave son la hexiquinasa y la fosfofructoquinasa y la piruvato quinasa. Para la gluconeognesis, la fructosa-1,6-difosfato y la piruvato carboxilasa. El regulador ms importante de ambas rutasw es la fructosa-2,6-bifosfato (F2,6BP) y esta es catalizada por la fosfofructoquinasa-2 (PFK2) .
Metabolismo quimitrofo de la energa: respiracin aerobiaRespiracin celular: maximizando el rendimiento del ATP Cuando se dispone de un aceptor de electrones externo, se puede dar la oxidacin completa del sustrato, obteniendose ms ATP, esto gracias a la
respiracin celular. Se entiende por respiracin celular: el flujo de electrones en o a a travs de una membrana, desde coenzimas reducidas hasta un aceptor de electrones, normalmente acompaado de la produccin de ATP. La coenzima reducida que se genera por el catabolismo glucoltico es el NADH, tambin hay otras dos coenzimas FAD y coenzima Q tambin recogen los electrones que se liberan. El aceptor final es el xigeno, la forma reducida de este aceptor es el agua, y se conoce al proceso en conjunto como respiracin aerobia. Y tiene lugar en la mitocondria. La respiracin fermentacin aerobia produce mucha ms energa que la
La respiracin aerobia puede producir potencialmente hasta 38 molculas de ATP por molcula de glucosa en procariotas y de 36 a 38 en eucariotas. La respiracin incluye la gluclisis, el ciclo del TCA, el transporte de electrones y la sntesis de ATP Las respiracin se divide en cuatro etapas, las primeras dos refieren a procesos oxidativos mediados por coenzimas y los otros dos incluyen la reoxidacin de las coenzimas y la produccin de ATP. La etapa 1 es la ruta glucoltica, la gluclisis que tiene como resultado la oxidacin de glucosa a piruvato. El piruvato es oxidado a acetil coenzima A, el cual entra en la etapa 2, el ciclo del cido tricarboxlico (TCA), esta ruta oxida los carbonos entrantes a CO2 y conserva la energa en forma de coenzimas reducidas, compuestos ricos en energa. En la etapa 3 se da el transporte de electrones, es decir la transferencia de electrones desde coenzimas reducidas hasta el oxgeno, acoplado a un transporte activo o bombeo, de protones a travs de la membrana. (proceso exergnico) y aporta energa generando un gradiente electroqumico de protones. En la etapa 4, la energa de este gradiente se usa para impulsar la sntesis de ATP. Este modelo de sntesis de ATP dependiente de oxigeno se denomina fosforilacin oxidativa. La mitocondria: donde tiene lugar todo el proceso -Las mitocondrias se encuentran con frecuencia en los lugares en los que la necesidad de ATP es mayor. -Las mitocondrias poseen forma de salchicha, son entidades separadas y son organulos grandes y nmerosos. Las membranas externa e interna definen dos compartimientos separados La mitocondria posee una membrana interna y una membrana externa. La membrana externa no representa una barrera de permeabilidad significativa para e paso de iones y molculas pequeas, ya que dispone de unos canales proteicos transmembranosos denominados porinas, que permiten e paso de solutos. Tambin hay un espacio intermembrana. La
membrana interna si representa una barreara para la permeabilidad de la mayora de solutos, separando por tanto, los espacios intermembrana y del interior del orgnulo en dos compartimientos separados. La membrana interna posee crestas, mayor nmero de crestas, mayor cantidad de trabajo. El interior de la mitocondria esta relleno de una matriz semifluida, aqu se encuentran las enzimas involucradas en la funcin mitocondrial. Las funciones mitocondriales tienen lugar especficas o dentro de los compartimientos en membranas
La mayor parte de enzimas mitocondriales, son enzimas de la matriz. La mayora de los intermediarios en la cadena de transporte de electrones son componentenes de la membrana interna. Sobresaliendo desde la membrana interna hacia la matriz se encuentran unas esferas con forma de puo llamadas complejo F, este complejoe sta unido a un complejo Fo y forma el complejo FoF1, y se considera como una ATP sintetasa, responsable de la mayor parte de produccin de ATP en la mitocondria. En procariotas, las funciones respiratorias se localizan en la membrana plasmtica y en el citoplasma El ciclo del cido tricarboxlico: a vueltas con la oxidacin En presencia de oxgeno, el piruvato se oxida completamente a dixido de carbono y la energa liberada en el proceso se usa para promover la sntesis de ATP. La primera etapa en este proceso es ruta cclica. Un intermediario importante en esta serie de reacciones cclicas es el citrato, que contiene tres grupos carboxilo y por lo tanto es un cido tricarboxlico. Por esta razn esta ruta se denomina ciclo del cido tricarboxlico (TCA), o ciclo de Krebs. El ciclo del TCA metaboliza acetil coenzima A que consiste en un acetato unido a un transportador llamado coenzima A. El acetil CoA proviene de la descarboxilacin oxidativa del piruvato o de la ruptura oxidativa de cidos grasos. El CoA transfiere su grupo acetato a un aceptor de cuatro carbonos llamado oxalacetato, formndose de esta manera citrato. Luego el citrato se oxida y regenera el oxalacetato inicial. Cada vuelta del ciclo de Krebs implica la entrada de dos carbonos (al acetato del acetil CoA), la liberacin de dos carbonos en forma de dixido de carbono y la regeneracin de oxalacetato. Esto se dicide en cinco etapas: cuatro dentro del mismo ciclo y una reaccin que convierte piruvato en acetil CoA. Los sustratos del ciclo de Krebs son por lo tanto el acetil CoA, las coenzimas oxidadas, el ADP y el Pi y los productos son dixido de carbno, coenzimas reducidas y una molcula de ATP. El piruvato se convierte descarboxilacin oxidativa en acetil coenzima A mediante la
El paso de piruvato a acetil CoA requiere de la actividad de la piruvato deshidrogenasa (complejo multiproteico), logrando as la descarboxilacin oxidativa del piruvato. As el piruvato pasa a ser acetato liberando CO2 y reduciendo NAD en NADH, la energa libre se usa para activar el acetato y as unise al grupo coenzima A y formar el acetil CoA.
El ciclo del TCA comienza con la entrada de acetato en forma de acetil CoA En cada vuelta del ciclo del TCA, entran dos tomos de carbono en forma orgnica (como acetato) y salen dos tomos de carbono en forma inorgnica (como dixido de carbono). En la primera reaccin (TCA-1) el grupo acetato de dos carbonos de acetil CoA, se une al cpuesto de cuatro carbonos oxalacetato, para formar citrato. Esta condensacin est catalizada por la citrato sintasa. En dos reacciones de oxidacin del ciclo del TCA se forma NADH y se libera O2 TCA2 = se convierte el citrato en isocitrato, enzima aconitasa. TCA 3 y TCA 4= etapas descarboxilativas, se elimina CO2 y se reduce el nmero de carbonos de seis a cinco y de cinco a cuatro. TCA3= el isocitrato se oxida a un compuesto de seis carbonos llamado oxalusuccinano. Y se produce NADH. Inmediatamente el oxalusuccinano pasa a ser a-cetoglutarato. Se libera O2 TCA4= El a-cetoglutarato se oxida a succinil CoA, libera NADH, se libera O2. La formacin directa de GTP (o ATP) tiene lugar en una etapa del ciclo de TCA TCA5 = La energa del enlace toister del succinil CoA se usa para general una molcula de ATP (bacterias y clulas vegetales) o GTP (clulas animales) Las reacciones oxidativas finales del ciclo del TCA genran FADH2 y NADH TCA6= el succinato se oxida a fumarato. Al ser una oxidacin con menos energa, el aceptor es FAD aceptando dos protones y dos electrones, redujendose a FADH2. TCA7= el doble hlice del fumarato se hidrata para formar malato. TCA8= el grupo hidroxilo del malato se convierte en la diana de la oxidacin final, nuevamente se libera NADH, y el producto es exalacetato. En resumen: los productos del ciclo del TCA son CO2, ATP, NADH y FADH2 1. El acetato entra en el ciclo como acetil CoA y se une a una molcula aceptora de cuatro tomos de carbono para formar citrato, compuesto de seis carbonos. 2. La descarboxilacin ocurre en dos etapas del ciclo por lo que la entrada de dos carbonos en forma de acetato se compensa por la prdida de dos carbonos en forma de dixido de carbono. (TCA3 y TCA4)
3. La oxidacin tiene lugar en cuatro etapas (TCA3, 4, 6 y 8) con el NAD como aceptor en tres de ellas (3,4,8) y FAD en uno de ellos (6). 4. El ATP se genera en un solo momento, con el GTP como intermediario en las clulas animales. (5) 5. Se completa la vuelta del ciclo con la regeneracin del oxoalacetato, el aceptor original de cuatro carbonos. Hasta este punto las coenzimas formadas NADH FADH2 son compuestos de alta energa altamente exergnicos. -Varias encimas del TCA estn sujetas a regulacin alostrica. -Las protenas y los lpidos tambin pueden ser fuente de energa. -El ciclo de glioxilato convierte el acetil CoA en carbohidratos. Transporte de electrones: flujo de electrones desde las coenzimas al oxgeno Se generan dos molculas de ATP durante la gluclisis y dos durante el TCA. Se libera una gran cantidad de energa cuando las coenzimas NADH y FADH2 son reoxidadas, por la transferencia del hidrgeno al oxgeno molecular. Hasta aqu llevamos 10 NADH y 2 FADH2. El proceso de reoxidacin de las coenzimas mediante la transferencia de electrones al oxgeno se conoce como transporte de electrones, el gradiente electroqumico de protones es al mismo tiempo el resultado del transporte de electrones y el origen de la energa que impulsa la sntesis de ATP. El sistema de transporte de electrones conduce los electrones desde las coenzimas reducidas al oxgeno -El transporte de electrones implica la oxidacin altamente exergnica del NADH y de FADH2. -La transferencia de electrones se lleva a cabo en un proceso de muchos pasos, que implica una serie de transportadores de electrones oxidables reversiblemente que funcionan conjuntamente en lo que se ha llamado cadena de transporte de electrones (ETS).
El sistema de transporte de electrones consiste en cinco tipos diferentes de trasmportadores Todos menos la coenzima Q, son protenas con grupos prostticos capaces de ser reducidos y oxidados, la mayora aparecen en la membrana. -Flavoprotenas: usan como grupo prosttico FAD o FMN (mitad de FAD). -Ferrosulfoprotenas (ferrodoxinas), (Fe-S) el aceptor es el Fe, alternando entre Fe2 y Fe3. -Citocromos: el transportador es su tomo Fe. Un unico electrn, igual que arriba. -Citocromos con cobre: (Cu-Fe), ambos pueden aceptar. -Coenzima Q: unico componente no proteico de la ETS,(ubiquinona), transportadores ms abundantes ubicados en la membrana interna, acepta
electrones y protones cuando se reduce y libera electrones y protones cuando se oxida. (Bomba de electrones, recibe de un lado y libera del otro).
Base estructural de la informacin celular: DNA, cromosomas y el ncleoNaturaleza qumica del material gentico El descubrimiento del DNA por Meischer condujo a propuestas contraictorias sobre la naturaleza qumica de los genes El ADN contiene la informacin gentica, el ADN se encuentra en los cromosomas que a su vez se encuentran en el ncleo de la clula. Avery demostr que el DNA es el materia gentico de las bacterias Hershey y Chase demostraron que el DNA es el material gentico de los virus Los bacterifagos o fagos son virus que infectan bacterias. Se utilizan como sistema modelo para estudiar genes. Un fago se ancla a la pared de la clula, inyecta el ADN dentro de esta, se transcribe y traduce para luego autoensamblarse, seguidamente la clula infectada se lisa (se abren grietas) y libera las nuevas partculas fago. Este curso de acontecimientos se denomina crecimiento ltico y es caracterstico de un fago virulento, este crecimiento ltico tiene como resultado la lisis de la clula husped y la produccin de muchas partculas fago. En comparacin, un fago temperado puede producir crecimiento ltico, o integras su ADN en el cromosoma bacteriano sin causar ningn dao inmediato a la clula husped. En el estado integrado o estado lisognico, el ADN de un fago temperado se denomina profago. El profago se replica junto con el ADN bacteriano. Durante este tiempo, los genes del fago, auque potencialmente letales, para el husped, estn inactivos o reprimidos. Sin embargo bajo ciertas condiciones, el DNA del profago se escinde del cromosoma bacteriano y entra otra vez en el ciclo ltico, produciendo progenie de partculas fago y lisando la clula husped. Las reglas de Chargaff revelan que A=T y que C=G Las cuatro bases en el ADN son: adenina, guanina, citosina y timina; estas no varian con el individuo, el tejido, la edad y el estado nutricional o medio ambiente. El nmero de adeninas es igual al nmero de timinas A=T, y el nmero de guaninas igual al nmero de citosinas G=C: esto significa que el nmero de purinas es igual al nmero de pirimidinas (A+G= C+T). El significado de estas equivalencias es conocido como reglas de Chargaff. La estructura del ADN Watson y Crack descubrieron que el ADN es una doble hlice El modelo de ADN consiste en dos hebras entrelazadas, una doble hlice. En la doble hlice de Watson y Crack los esqueletos de azcar fosfato de las dos hebras estn en el exterior de la hlice, y las bases miran hacia el
interior, hacia el centro de la hlice, formando los escalones de una escalera circular a la que se parece la estructura. La hlice es dextrgira (se curva hacia arriba y hacia la derecha). Contiene diez pares de nucletidos por vuelta. Se necesita de un par purina-pirimidina. Las dos hebras se sostienen por puentes de hidrgeno entre las bases de las hebras opuestas. Adems, los puentes de hidrgeno que mantienen juntas las dos hebras de la doble hlice slo ajustan cuando se forman entre la base adenina (A) en una de las cadenas y timina (T) en la otra, o entre la base guanina (G) en una cadena y citosina (C) en la otra. Esto significa que la secuencia de bases en la cadena determina la secuencia de bases en la cadena opuesta, adems, se dice que las dos cadenas de la doble hlice de ADN son complementarias. Esto sugiere el como se replica el ADN: las dos hebras de la doble hlice de ADN se separan antes de la divisin, de manera que cada hebra funciona como un molde que dirige la sntesis de una nueva hebra de ADN, usando reglas del emparejamiento de bases. Caractersticas importantes de la doble seran que: la forma en que giran las dos hebras crea un surco mayor y un surco menor. (estos desempean un papel importante en las interacciones de ADN con varias molculas). Otra caracterstica importante es la orientacin antiparalela (esto significa que los enlaces fosfodister de las dos hebras se orientan en direcciones opuestas). La hlice dextrgira de Watson y crack se denomina B-DNA sin embargo existen formas alternativas como el A-DNA (se crea deshidratando el B-DNA) y el Z-DNA (tiene forma de zigzag levgira) El DNA puede transformarse en formas relajadas y superenrolladas Cuando el DNA gira sobre s mismo forma DNA superenrollado. Una molcula de DNA puede ir haca atrs o hacia delante entre el estado de superenrollamiento y el estado de no superenrollamiento, o estado relajado (forma circular). El superenrollamiento puede ser positivo (si se gira en la direccin en que estn entrelazadas las hebras) o negativo (si se gira en contra a como estn entrelazadas las hebras). El DNA en circular se encuentra en la naturaleza, incluyendo el de bacterias, virus y organulos eucariotas estn de manera invariable superenrolladas negativamente. El superenrollamiento se produce tambin en molculas de DNA lineal, cuando las regiones de la molcula estn ancladas a alguna estructura celular de manera que no pueda rotar libremente. El superenrollamiento influye tanto en la organizacin espacial como en el estado de energa de ADN y afecta a la capacidad de una molcula de DNA para interactuar con otras molculas. El superenrollamiento positivo implica un enrollamiento muy apretado de la doble hlice y por tanto se reducen las oportunidades para interaccionar. Por el contrario, el superenrollamiento negativo est asociado con el desenrollamiento de la doble hlice, que aumenta el acceso de sus hebras a las protenas implicadas en la replicacin o en la trascripcin del DNA. La interconversin entre la forma relajada y la forma superenrollada del ADN est catalizada por enzimas denominadas topoisomerasas. Estas se
clasifican en tipo I y tipo II. Ambos tipos catalizan la relajacin del ADN superenrollado, pero las enzimas del tipo I lo hacen introduciendo cortes transitorios en una sola hebra del ADN ( y pasa la hebra no cortada a travs de la ruptura) mientras que las enzimas del tipo II introducen rupturas transitorias en la doble hebra ( y pasa una regin no cortada de la doble hlice de DNA a travs del orificio). El tipo II requiere energa derivada de la hidrlisis de ATP. Los procariotas tienen una topoisomerasa de tipo II denominada DNA girasa que puede inducir tanto la relajacin como el superenrollamiento del DNA. Pude relajar el superenrollamiento positivo que resulta del desenrollamiento parcial de una doble hlice o puede introducir activamente vueltas negativas que promuevan la separacin de las hebras, facilitando adems el acceso a otras protenas implicadas en la replicacin del ADN. La DNA girasa requiere ATP para generar el superenrollamiento, pero no para relajar una molcula ya superenrollada. Las dos hebras de una doble hlice de DNA se pueden separar por desnaturalizacin y volver a unirse por renaturalizacin Debido a que las dos hebras de la doble hlice de DNA se mantienen unidas por enlaces no covalentes relativamente dbiles, las dos hebras pueden separarse con facilidad. La separacin de las hebras inducida de manera experimental da como resultado desnaturalizacin del DNA; el proceso inve