Embed Size (px)
Citation preview

Respuestas Fisiológicas de Siete Genotipos
de Arroz (Oryza sativa L.) a Diferentes
Temperaturas Nocturnas
Oscar Humberto Alvarado Sanabria
Universidad Nacional de Colombia
Facultad de Ciencias Agrarias, Departamento de Agronomía
Bogotá, Colombia
2016

II Respuestas Fisiológicas de Siete Genotipos de Arroz (Oryza sativa L.)
a Diferentes Temperaturas Nocturnas
Respuestas Fisiológicas de Siete Genotipos
de Arroz (Oryza sativa L.) a Diferentes
Temperaturas Nocturnas
Oscar Humberto Alvarado Sanabria
Trabajo de investigación presentado como requisito parcial para optar al título de:
Magister en Ciencias Agrarias con Énfasis en Fisiología de Cultivos
Director:
Ph.D. Hermann Restrepo Díaz
Codirector:
Msc. Gabriel Garcés Varón
Fisiología de Cultivos.
Universidad Nacional de Colombia
Facultad de Ciencias Agrarias, Departamento de Agronomía
Bogotá, Colombia
2016

Resumen y Abstract III
Resumen
La temperatura es uno de los principales factores que determina la distribución y el potencial
desarrollo de una especie. En los últimos años la temperatura promedio ha aumentado en
todo el mundo y Colombia no ha sido la excepción con un aumento de aproximadamente
2°C en los últimos 20 años. Este aumento de la temperatura perjudica el crecimiento,
desarrollo y reproducción de varias especies cultivables, dentro de ellas el arroz. Esta
especie es una de las principales fuentes de energía para la especie humana, por lo que
cualquier disminución en sus rendimientos afecta la seguridad alimentaria del mundo. La
alta temperatura nocturna (ATN) ha mostrado ser uno de los factores más perjudiciales para
el rendimiento del arroz. En el presente estudio se caracterizó la respuesta de siete genotipos
de arroz a una alta temperatura nocturna (30°C) en estado de macollamiento y durante la
maduración. Para esto se evaluaron variables de crecimiento, fotosíntesis, variables
relacionadas con estrés oxidativo, y algunos componentes de rendimiento. Los resultados
indicaron que i) la alta temperatura nocturna afecta los procesos fisiológicos (en estado
vegetativo) desde el cuarto día de exposición ii) la alta temperatura nocturna disminuye el
crecimiento vegetativo debido a una menor asimilación de CO2 y mayor respiración iii) la
alta temperatura nocturna disminuye el rendimiento por panícula del genotipo F50 y lo
aumenta en CT19021 debido a una mayor y menor producción de granos vanos,
respectivamente. Con base en los resultados obtenidos en el presente estudio, el uso de
variables fisiológicas y bioquímicas como posibles indicadores de tolerancia o
susceptibilidad a un estrés por alta temperatura podrían ser: En estado vegetativo la
asimilación de CO2, el contenido de clorofila y carotenoides, el contenido de MDA y la
respiración; en estado reproductivo el número de granos producidos y el porcentaje de
fertilidad, mientras variables como, conductancia estomática, contenido de prolina y
fluorescencia de la clorofila a varían en el tiempo o no presentan cambios al aumentar la
temperatura nocturna, por lo que no serían buenos indicadores del estrés provocado por una
alta temperatura nocturna.
Palabras clave: fotosíntesis, estrés abiótico, estrés oxidativo, fluorescencia, genotipo,
clorofila.

IV Respuestas Fisiológicas de Siete Genotipos de Arroz (Oryza sativa L.)
a Diferentes Temperaturas Nocturnas
Abstract
Temperature is one of the main factors that determine the distribution and the potential
development of a species. In recent years the average temperature has increased worldwide,
and Colombia is no exception with an increase of about 2 °C in the last 20 years. This
increase in temperature impairs growth, development and reproduction of several crop
species within them rice. This species is one of the main sources of energy for the human
species, so any decline in yields affects food security in the world. The high night
temperature has proved one of the most damaging rice yield factors. In this study the
response of seven lines of rice at high night temperature (30 °C) and tillering state during
grain filling was characterized. For this growth variables, photosynthesis, oxidative stress-
related variables, and some yield components were evaluated. The results indicated that i)
the high night temperature affects physiological processes (vegetative state) from four days
of exposure and these vary over time ii) the high night temperature decreases vegetative
growth due to lower CO2 assimilation and greater respiration iii) high night temperature
decreases production / panicle genotype F50 and CT19021 increases due to a higher
production and lower openings, respectively grains. Because of its stability over time and
that regardless of genotype tends to exhibit the same behavior, the following variables as
potential indicators of stress are suggested for high temperature CO2 assimilation, number
and percentage of grain produced vaneamiento, while variables such as, stomatal
conductance, MDA and proline content are variable in time or are unchanged by increasing
the night temperature, so they would not be good indicators of this stress.
Keywords: Photosynthesis, Abiotic stress, oxidative stress, fluorescence, genotype,
chlorophyll

Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L.) a Diferentes Temperaturas Nocturnas
V
CONTENIDO
Pág.
Resumen .............................................................................................................................. III
Abstract ............................................................................................................................... IV
Lista de figuras ................................................................................................................. VIII
Lista de tablas ...................................................................................................................... X
Lista de Símbolos y Abreviaturas ....................................................................................... XI
INTRODUCCIÓN .............................................................................................................. 13
1. MARCO TEÓRICO .................................................................................................... 15
1.1 FISIOLOGÍA DE LAS PLANTAS BAJO UNA ALTA TEMPERATURA ....15
1.2 RESPUESTA DEL CULTIVO DEL ARROZ A UNA ALTA TEMPERATURA
NOCTURNA ................................................................................................................16
1.3 LAS ALTAS TEMPERATURAS Y LA FENOLOGÍA DEL ARROZ............19
1.4 EL USO DE VARIABLES FISIOLÓGICAS EN EL FITOMEJORAMIENTO
DE ARROZ ..................................................................................................................20
1.4.1 La fotosíntesis y sus componentes ..........................................................22
1.4.2 Reproducción y Rendimiento .................................................................27
1.4.3 Peroxidación Lipídica de membranas .....................................................28
1.4.4 Osmolitos compatibles (Prolina) ............................................................29
2. OBJETIVOS ................................................................................................................ 31
2.1 Objetivo General ................................................................................................31
2.2 Objetivos Específicos ........................................................................................31
3. MATERIALES Y MÉTODOS .................................................................................... 33
3.1 RESPUESTA FISIOLÓGICA DE PLANTULAS DE ARROZ F60
SOMETIDAS A PERIODOS DE ALTA TEMPERATURA NOCTURNA. ..............33
3.1.1 Condiciones generales del experimento .................................................33
3.1.2 Tratamientos ...........................................................................................34
3.1.3 Fotosíntesis, conductancia estomática, y transpiración ..........................34
3.1.4 Uso eficiente del agua y eficiencia de carboxilación .............................35
3.1.5 Eficiencia máxima del PSII (Fv/Fm) ......................................................35
3.1.6 Contenido de clorofila a, clorofila b y carotenoides ...............................35
3.1.7 Respiración oscura de la hoja .................................................................36
3.1.8 Contenido de Prolina ..............................................................................36
3.1.9 Peroxidación Lipídica .............................................................................36
3.1.10 Análisis Estadístico.................................................................................37
3.2 RESPUESTA FISIOLÓGICA DE SEIS GENOTIPOS DE ARROZ EN
ESTADO VEGETATIVO A UNA ALTA TEMPERATURA NOCTURNA .............38

VI Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L.) a Diferentes Temperaturas Nocturnas
3.2.1 Tratamientos ...........................................................................................38
3.2.2 Condiciones generales de crecimiento ...................................................38
3.2.3 Fotosíntesis .............................................................................................39
3.2.4 Eficiencia de Carboxilación y Uso eficiente del agua ............................39
3.2.5 Respiración oscura ..................................................................................39
3.2.6 Parámetros de Fluorescencia de la clorofila a ........................................40
3.2.7 Parámetros de Crecimiento .....................................................................40
3.2.8 Pérdida de electrolitos ............................................................................40
3.2.9 Clorofila y Carotenoides .........................................................................41
3.2.10 Prolina y Malondialdehído (MDA) ........................................................41
3.2.11 Diseño experimental y análisis estadístico .............................................41
3.3 CARACTERIZACIÓN FISIOLÓGICA DE SIETE GENOTIPOS DE ARROZ
SOMETIDOS A UNA ALTA TEMPERATURA NOCTURNA ...............................42
3.3.1 Condiciones generales de crecimiento ...................................................42
3.3.2 Genotipos y tratamientos ........................................................................42
3.3.3 Componentes de rendimiento .................................................................43
3.3.4 Parámetros de Fluorescencia de la Clorofila a .......................................43
3.3.5 Contenido de Clorofila y Carotenoides ..................................................43
3.3.6 Prolina y Malondialdehído (MDA) ........................................................43
3.3.7 Análisis Estadístico.................................................................................44
4. RESULTADOS ........................................................................................................... 45
4.1 RESPUESTA FISIOLÓGICA DE PLANTULAS DE ARROZ SOMETIDAS A
PERIODOS DE ALTA TEMPERATURA NOCTURNA. ..........................................45
4.1.1 Fotosíntesis, conductancia estomática, transpiración y concentración
interna de CO2 .......................................................................................................45
4.1.2 Eficiencia de la carboxilación y uso eficiente del agua ..........................46
4.1.3 Respiración oscura y balance de carbono ...............................................47
4.1.4 Eficiencia del fotosistema II y contenido de pigmentos fotosintéticos. .49
4.1.5 Peroxidación lipídica y contenido de prolina .........................................50
4.2 RESPUESTA FISIOLÓGICA DE SEIS GENOTIPOS DE ARROZ EN
ESTADO VEGETATIVO A UNA ALTA TEMPERATURA NOCTURNA .............51
4.2.1 Fotosíntesis, Eficiencia de la Carboxilación y Uso eficiente del Agua ..51
4.2.2 Respiración .............................................................................................52
4.2.3 Variables de Crecimiento .......................................................................53
4.2.4 Fluorescencia de la clorofila a ................................................................54
4.2.5 Fuga de electrolitos, MDA, Prolina y pigmentos fotosintéticos.............55
4.3 CARACTERIZACIÓN FISIOLOGICA DE SIETE GENOTIPOS DE ARROZ
SOMETIDOS A UNA ALTA TEMPERATURA NOCTURNA ................................58

Contenido VII
4.3.1 Componentes de rendimiento (Número de granos vanos, Número de
granos llenos, Porcentaje de fertilidad y masa de granos llenos por panícula) .....58
4.3.2 Fluorescencia de la Clorofila a ...............................................................60
4.3.3 Pruebas bioquímicas (Pigmentos fotosintéticos, MDA y Prolina) .........61
5. DISCUSIÓN ................................................................................................................ 63
5.1 PRIMER EXPERIMENTO: PERIODOS DE ESTRÉS ....................................63
5.2 SEGUNDO Y TERCER EXPERIMENTO: RESPUESTA DE SIETE
GENOTIPOS DE ARROZ A UNA ALTA TEMPERATURA NOCTURNA ............65
6. MODELO TEÓRICO DEL EFECTO DE LA ALTA TEMPERATURA NOCTURNA
69
7. CONCLUSIONES Y RECOMENDACIONES .......................................................... 71
7.1 Conclusiones ......................................................................................................71
7.2 Recomendaciones ..............................................................................................71
Bibliografía ......................................................................................................................... 73

Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L.) a Diferentes Temperaturas Nocturnas
VIII
Lista de figuras
Figura 1. Efectos del fenómeno del niño (2009-2010) en los rendimientos de la zona de
Espinal (Tolima). Fuente: FEDEARROZ (2015). .............................................................. 19
Figura 2. Fotosíntesis(A), Conductancia estomática(B), transpiración(C) y Concentración
intercelular de CO2(D) de plántulas de arroz sometidas a dos temperaturas nocturnas (24 vs
30°C). Las ecuaciones representan el comportamiento cúbico en el tiempo. n.s,*, ** y ***
representa no significativo y significancia con P≤0,05, 0,01 y 0,001, respectivamente. Cada
punto representa el promedio de 12 valores. ...................................................................... 46
Figura 3. Eficiencia de carboxilación(A) y Uso eficiente del agua(B) de plántulas de arroz
sometidas a dos temperaturas nocturnas (24 vs 30°C). n.s y * representan no significativo y
significancia con P≤0,05 según la prueba de Tukey, entre temperaturas para un mismo día.
Cada barra de gráfico representa la medía de doce valores+Error estándar. ..................... 47
Figura 4. Respiración oscura de plántulas de arroz sometidas a dos temperaturas nocturnas
(24 vs 30°C). Las ecuaciones presentan un comportamiento cúbico en el tiempo. * y ***
representan significancia con P≤0,05 y 0,001, respectivamente. Cada punto representa el
promedio de 12 valores. ...................................................................................................... 48
Figura 5. Balance de CO2 de plántulas de arroz sometidas a dos temperaturas nocturnas (24
vs 30°C). n.s y * representan no significativo y significancia con P≤0,05, entre temperaturas
para un mismo día, según la prueba de Tukey. Cada barra de gráfico representa la medía de
doce valores+Error estándar. .............................................................................................. 48
Figura 6. Eficiencia máxima potencial del PSII (Fv/Fm) de plántulas de arroz sometidas a
dos temperaturas nocturnas (24 vs 30°C). n.s y * representan no significativo y significancia
con P≤0,05, entre temperaturas para un mismo día según la prueba de Tukey. Cada barra de
gráfico representa la medía de doce valores+Error estándar. ............................................ 49
Figura 7. Contenido de clorofila y carotenoides de plántulas de sometidas a dos
temperaturas nocturnas (24 vs 30°C). Líneas con la letra L presentan un comportamiento
lineal (P≤0,001). Cada punto representa el promedio de 12 valores. ................................. 50
Figura 8. MDA(A) y contenido de Prolina (B) de plántulas de arroz sometidas a dos
temperaturas nocturnas (24 vs 30°C). n.s y * representan no significativo y significancia con

Contenido IX
P≤0,05, entre temperaturas para un mismo día, según la prueba de Tukey. Cada barra del
gráfico representa la medía de doce valores+Error estándar. ............................................ 51
Figura 9. Fotosíntesis (A), Eficiencia de la carboxilación (B), CO2 intercelular (C), y Uso
eficiente del agua intrínseco (D) de hojas de plantas de seis genotipos de arroz. n.s y *
representan no significativo y significancia con P≤0,05, entre temperaturas para un mismo
día, según la prueba de Tukey. Los datos representan el promedio de 8 repeticiones±Error
estandar. .............................................................................................................................. 52
Figura 10. Respiración de hojas de plantas de seis genotipos de arroz. n.s y * representan
no significativo y significancia con P≤0,05, entre temperaturas para un mismo día, según la
prueba de Tukey. Los datos representan el promedio de 8 repeticiones±Error estándar.... 53
Figura 11. Perdida de electrolitos (A) y Producción de MDA (B) de hojas de plantas de seis
genotipos de arroz. Los datos representan el promedio de 8 repeticiones±Error. Asteriscos
significan diferencias significativas en la interacción de Temperatura (T) x Genotipo (G) a
P≤0,05 de acuerdo con el ANOVA. .................................................................................... 56
Figura 12. Efecto de dos temperaturas nocturnas (24 vs 30°C) sobre el número total de
granos (A), de granos llenos (B), de granos vanos (C) y porcentaje de fertilidad (D) en siete
genotipos de arroz. Los datos representan el promedio de 6 repeticiones±Error. Barras con
letras distintas indican diferencias significativas a P≤0,05 según la prueba de Tukey. ..... 59
Figura 13. Masa de granos llenos de panículas de arroz bajo dos temperaturas nocturnas
(A) y en siete genotipos (B). Los datos representan el promedio de 6 repeticiones±Error
estándar. Barras con letras distintas indican diferencias significativas a P≤0,05 según la
prueba de Tukey. ................................................................................................................. 60
Figura 14. Rendimiento cuántico real del fotosistema II (Y(II))(A), Disipación no
fotoquímica controlada (Y(NPQ))(B), Disipación no fotoquímica no controlada (Y(NO))(C)
y Rendimiento cuántico potencial del PSII (Fv/Fm)(D) de la hoja bandera de siete genotipos
de plantas de arroz. Los datos representan el promedio de 6 repeticiones±Error estándar.
Barras con letras distintas indican diferencias significativas a P≤0,05 según la prueba de
Tukey. ................................................................................................................................. 61
Figura 15. Modelo conceptual de los efectos y las características de tolerancia y
susceptibilidad de arroz sometido a altas temperaturas nocturnas (30°C). Los símbolos
dentro de cada recuadro indican aumento (↑), disminución (↓) o sin cambios (=) del proceso
dentro del recuadro. El origen de la flecha roja indica las posibles causas de un proceso y el
final de la flecha roja indica el proceso afectado. ............................................................... 70

X Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L.) a Diferentes Temperaturas Nocturnas
Lista de tablas
Tabla 1. Efecto de la alta temperatura nocturna en procesos fisiológicos del arroz .......... 18
Tabla 2. Rangos óptimos de temperatura diaria para el desarrollo del arroz según su etapa
fenológica. Información adaptada de Yoshida (1981). ....................................................... 19
Tabla 3. Parámetros de la fluorescencia derivados de la fluorescencia de la clorofila a ... 24
Tabla 4. Variables de crecimiento de hojas de seis genotipos de arroz expuestos a dos
diferentes temperaturas nocturnas (24 vs 30°C) ................................................................. 54
Tabla 5. Eficiencia máxima real y eficiencia máxima potencial y formas de disipación de la
energía del PSII en hojas de seis genotipos de arroz expuestos a dos diferentes temperaturas
nocturnas (24 vs 30°C) ....................................................................................................... 55
Tabla 6. Pigmentos fotosintéticos y Prolina de hojas de seis genotipos de arroz expuestos a
dos temperaturas nocturnas (24 vs 30°C) ........................................................................... 57
Tabla 7. Contenido de Pigmentos fotosínteticos (Clorofilas y Carotenoides),
Malondialdehído (MDA) y prolina en hojas banderas de siete genotipos de arroz expuestos
a dos diferentes temperaturas nocturnas (24 vs 30°C). ....................................................... 62

Lista de Símbolos y Abreviaturas XI
Lista de Símbolos y Abreviaturas
ANOVA = Análisis de varianza.
ATP = Adenosin Trifosfato.
CAR (x+c) = carotenoides.
CE = Eficiencia de carboxilación.
Chl = Clorofila.
Chl a = Clorofila a.
Chl b = Clorofila b.
Ci= CO2 subestomático.
DDE = Días de Exposición.
DE = Días después de emergencia.
E = Transpiración.
Fm = Fluorescencia Máxima de hojas adaptadas a la Oscuridad.
Fm’ = Fluorescencia Máxima de hojas adaptadas a la Luz.
Fo’ = Fluorescencia Mínima de hojas adatadas a la Luz.
Fo = Fluorescencia Mínima de hojas adatadas a la Oscuridad.
Fv/Fm = Eficiencia máxima potencial del fotosistema II.
MF= Masa Fresca.
gS = conductancia estomática.
ATN = Alta temperatura nocturna.
LHC = Complejo proteico cosechador de luz.
MDA = Malondialdehído
NAD = Nicotin Adenin dinucleotido.
NADPH = Nicotin Adenin dinucleotido reducido.
NPQ = Disipación no fotoquímica
PAR = Radiación Fotosintéticamente Activa.
Pn = tasa de asimilación de CO2
PS = Fotosistema
PSI = Fotosistema I
PSII= Fotosistema II
Ro = Respiración Oscura.
ROS = Especies Reactivas de oxígeno.
RuBisCO = Ribulosa 1,5 Bifosfato carboxilasa/oxigenasa.
RuBP = Ribulosa-1,5-bisfosfato.
TBA = Ácido Tiobarbitúrico.
WUE= Uso eficiente del agua, hallado como tasa fotosintética/tasa transpiratoria.
WUEi = Uso eficiente del agua intrínseco.

XII Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L.) a Diferentes Temperaturas Nocturnas
Y(II)= Eficiencia máxima real del fotosistema II.
Y(NO) = Eficiencia de la disipación no fotoquímica no controlada.
Y(NPQ) = Eficiencia de la disipación fotoquímica controlada

Introducción 13
INTRODUCCIÓN
El cambio climático ha generado condiciones adversas tales como periodos prolongados de
sequías, inundaciones y temperaturas extremas en varias regiones del mundo (Teixeira et
al., 2013). El cultivo del arroz es uno de los cereales con mayor importancia para la
alimentación humana en el mundo (FAO, 2002). Y en Colombia ocupó 455,000 ha
sembradas con una producción de 1,925,687 toneladas durante 2013 (FEDEARROZ, 2015).
En este sentido, periodos de altas temperaturas tanto diurnas como nocturnas han sido
registrados en los últimos años en las zonas productoras de arroz en Colombia, observándose
un impacto negativo sobre el rendimiento de este cultivo (Castilla et al., 2010; Restrepo-
Diaz y Garces-Varon, 2013; Sánchez-Reinoso et al., 2014).
El rendimiento del cultivo del arroz es fuertemente influenciado por factores ambientales
como luminosidad, disponibilidad de agua, nutrientes y la temperatura ambiental (Yoshida,
1981; Fageria, 2007). En cuanto a la temperatura ambiental, los periodos de exposición de
cultivos de arroz a altas temperaturas diurnas como nocturnas han aumentado durante los
últimos años (Restrepo-Díaz y Garcés Varón, 2013; Sánchez-Reinoso et al. 2014). En este
aspecto, periodos de estrés térmico, especialmente, altas temperaturas nocturnas pueden
afectar negativamente la fisiología y en consecuencia el rendimiento del cultivo del arroz
(Mohammed y Tarpley, 2014). En plántulas de arroz, altas temperaturas nocturnas (35-
45°C) disminuyeron la asimilación de CO2, la eficiencia del fotosistema II y la actividad de
la Ribulosa 1-5 bifosfato carboxilasa/oxidasa (RuBisCO) (Yin et al., 2010). Además de los
efectos sobre las propiedades de intercambio gaseoso de la hoja, una alta temperatura
nocturna reduce el contenido de clorofila (Kumar et al., 2011; Dong et al., 2014), afectando
la relación entre clorofila total y carotenoides (Song et al., 2013; Dong et al., 2014). Otra
respuesta fisiológica, es el incremento de la respiración nocturna expresado como perdida
de carbono (Lee y Akita, 2000; Mohammed y Tarpley, 2009b; Mohammed et al., 2013).
También, es importante mencionar que el grado de afectación negativa por una alta
temperatura nocturna sobre el rendimiento, está influenciada por el estado fenológico donde
se presente esta condición estresante (Fageria et al., 2007). Yoshida (1981) menciona que
las etapas de floración y maduración son etapas fenológicas más susceptibles a estreses
ambientales.
Una de las estrategias para afrontar los efectos negativos de las altas temperaturas sobre el
rendimiento de los cereales es la selección de genotipos tolerantes a dicha condición
ambiental (Araus et al., 2008). En este aspecto, el mejoramiento genético de cultivos

14 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
actualmente usa características fisiológicas con el propósito de aumentar la eficiencia en la
selección de genes (Reynolds et al., 2009; Jha et al., 2014). Por ejemplo, en la selección de
genotipos de trigo tolerantes a alta radiación y temperatura se han usado parámetros de la
fluorescencia de la clorofila y variables de intercambio gaseoso (Sayed, 2003; Monneveux
et al., 2003). Adicionalmente, la caracterización de genotipos tolerantes a altas temperaturas
nocturnas se tiene en cuenta variables bioquímicas como el contenido de pigmentos
fotosintéticos (clorofilas y carotenoides), la producción de osmolitos compatibles (glicina
betaína, Prolina y azucares solubles) y la integridad de la membrana celular (Bita y Gerats,
2013). En arroz, se ha reportado que los genotipos tolerantes presentan una tasa de
fotosíntesis, respiración, transporte de electrones, contenido de clorofila y rendimiento
cuántico del fotosistema II estables bajo condiciones de altas temperaturas nocturnas
(Glaubitz et al., 2014; Mohammed y Tarpley, 2014). Por lo anterior, estas variables han sido
sugeridas como herramienta para seleccionar genotipos con una posible tolerancia a
diferentes condiciones de estrés abiótico (Sayed, 2003; Araus et al., 2008).
Actualmente, la utilización de algunos parámetros fisiológicos y bioquímicos como tasa
fotosintética y producción de prolina han adquirido importancia en la caracterización de
genotipos a condiciones de estrés abiótico especialmente estrés térmico en Colombia
(Restrepo-Diaz y Garces-Varon, 2013; Sánchez-Reinoso et al. 2014). Sin embargo, los
trabajos donde se utilizan parámetros fisiológicos (fluorescencia de la clorofila a,
Peroxidación lipídica, intercambio gaseoso) siguen siendo necesarios para un mejor
conocimiento de la influencia del ambiente en la selección de genotipos con características
de tolerancia a estreses abióticos (especialmente estrés térmico) como futuros parentales en
los programas de fitomejoramiento. Por tal motivo, las hipótesis de trabajo en el presente
trabajo eran: i) El uso de variables como: fotosíntesis, crecimiento (acumulación de masa
seca y área foliar), componentes de rendimiento (tamaño de grano, número de granos,
vaneamiento) y bioquímicas (malondialdehído, contenido de prolina y contenido de
pigmentos) sirven como herramientas en la caracterización de un estrés por alta temperatura
nocturna para genotipos de arroz; ii) Los genotipos tolerantes presentan mayor crecimiento,
rendimiento, fotosíntesis, menor respiración y mayor fertilidad.

Marco Teórico 15
1. MARCO TEÓRICO
1.1 FISIOLOGÍA DE LAS PLANTAS BAJO UNA ALTA TEMPERATURA
La temperatura media del planeta ha aumentado los últimos años y seguirá aumentando
0,3℃ por década en comparación al lapso 1990-2010 (Lobell y Gourdji, 2012). Este
aumento de la temperatura afectará negativamente la producción de los cultivos más
importantes en la alimentación humana y animal como son: maíz, soya, trigo y arroz (Welch
et al., 2010b; Wassmann et al., 2009). La alta temperatura afectará de forma distinta a cada
región, debido a las condiciones propias de cada una, tanto climáticas como de manejo de
cultivo, por lo que son necesarias investigaciones a nivel regional.
La alta temperatura afecta el desarrollo y rendimiento de las plantas cultivables (Jha et al.,
2014; Lobell y Gourdji, 2012). Cada planta cultivable necesita unas condiciones climáticas
determinadas para desarrollarse óptimamente. Si estas condiciones no se presentan, el
desarrollo y crecimiento de las plantas disminuyen (Sage et al., 2015). La temperatura es
uno de los principales factores que afectan la supervivencia de las plantas y junto con la
disponibilidad de agua determinan la distribución de las plantas en el planeta (Lambers et al.,
2008).
La alta temperatura, tanto diurna como nocturna, afecta procesos esenciales en la fisiología
de la planta como la fotosíntesis, la respiración, la absorción de agua, el crecimiento y el
desarrollo (Mohammed y Tarpley, 2014, 2009b; Mohammed et al., 2013). A nivel de hoja,
la fotosíntesis es uno de los procesos más sensibles a la alta temperatura (Bita y Gerats,
2013; Sage y Kubien, 2007). En este sentido, tanto la parte fotoquímica como el ciclo de
Calvin son afectados (Wahid et al., 2007). En el caso de la parte fotoquímica, el fotosistema
II es la parte más sensible, ya que se pueden afectar procesos metabólicos como una la tasa
de transporte de electrones y la producción de especies reactivas de oxigeno (ROS) (Cui
et al., 2006; Yamasaki et al., 2002). Asimismo, El contenido y la distribución de los
pigmentos fotosintéticos son alterados por altas temperaturas. En este aspecto, aumenta el
contenido de carotenoides y disminuye el de clorofila con el fin de disipar el exceso de
energía y reducir las ROS (Demmig-Adams y Adams Iii, 1996; Gill y Tuteja, 2010). En el
caso del ciclo de Calvin, la actividad de la Rubisco puede ser inhibida por periodos de estrés
térmico (Wang et al., 2010; Kubien y Sage, 2008)
Otro proceso fisiológico afectado es la respiración (Wahid et al., 2007). El balance de
carbono en una planta está dado por la diferencia entre la ganancia de carbono por

16 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
fotosíntesis y la perdida de este por respiración (Turnbull et al., 2001; Sánchez-Reinoso
et al., 2014). Varios estudios han reportado cambios en respiración como respuesta a la
variación de la temperatura ambiental (Smith y Dukes, 2013). En general, la respiración se
incrementa en función de la temperatura (Lambers et al., 2008).
La interacción entre estrés térmico y el estado de desarrollo de la planta también es
importante en el rendimiento de los cultivos (Restrepo-Díaz et al., 2010). La investigación
actual ha concluido que el estado de floración es el más susceptible a las altas temperaturas,
pero las plantas pueden también sufrir alteraciones, provocados por estrés térmico, en los
demás estados fenológicos (Sánchez et al., 2014; Wassmann et al., 2009; Sage et al., 2015;
Wongpatsa et al., 2014). Una ATN puede causar aborto de flores y frutos, afectar la
viabilidad y fertilidad del polen y finalmente, la calidad del producto a cosechar (Restrepo-
Diaz et al., 2010; Sage et al., 2015). Asimismo, se ha reportado que los cultivos de cereales
sometidos a una ATN puede presentar mayor esterilidad en comparación a una alta
temperatura diurna (Welch et al., 2010a). Teniendo en cuenta los efectos del estrés térmico,
en cereales los genotipos tolerantes pueden mantener su rendimiento, fertilidad de las
espiguillas, viabilidad del polen y capacidad fotosintética (Poli et al., 2013).
Las plantas tienen respuestas bioquímicas a las altas temperaturas con el propósito de
sobrellevar esta condición estresante (Wahid et al., 2007). En plantas de cebada bajo estrés
térmico, se ha reportado un aumento en la relación Carotenoides/Clorofila, ya que una
disminución de la clorofila permite captar menos energía y evitar el calentamiento de la hoja
provocado por la reducción en la transpiración (Havaux y Tardy 1999). Otro mecanismo de
aclimatación a condiciones de alta temperatura, especialmente nocturna, es la menor
peroxidación lipídica representado en la producción de malondialdehído (MDA)
(Narayanan et al., 2016; Bahuguna et al., 2015; Zhou et al., 2012). Finalmente, las plantas
pueden presentar respuestas moleculares para tolerar episodios de estrés térmico (Jagadish
et al., 2012; Jha et al., 2014). En este sentido, genotipos tolerantes han mostrado una mayor
acumulación de proteínas de choque térmico que genotipos susceptibles (Jagadish et al.,
2010).
1.2 RESPUESTA DEL CULTIVO DEL ARROZ A UNA ALTA TEMPERATURA
NOCTURNA
La temperatura mínima (temperatura nocturna) ha presentado un mayor incremento que la
temperatura máxima diaria durante el siglo pasado (Krishnan et al., 2011). Esta condición

Marco Teórico 17
puede afectar las plantas a nivel fisiológico, bioquímico y morfológico (Krishnan et al.,
2011; Mohammed y Tarpley, 2014). En este contexto, las altas temperaturas nocturnas (35-
45°C) disminuyen la asimilación de CO2, la eficiencia del fotosistema II y la actividad de la
Ribulosa 1-5 bifosfato carboxilasa/oxidasa (RuBisCO) en arroz (Yin et al., 2010). Además
de los efectos sobre las propiedades de intercambio gaseoso de la hoja, una alta temperatura
nocturna reduce el contenido de clorofila (Kumar et al., 2011; Dong et al., 2014), afectando
la relación entre clorofila total y carotenoides (Song et al., 2013; Dong et al., 2014).
Otra respuesta fisiológica, es el incremento de la respiración nocturna expresado como
pérdida de carbono (Lee y Akita, 2000; Mohammed y Tarpley, 2009b; Mohammed et al.,
2013). Mohammed y Tarpley (2010a) han mencionado que esta pérdida de carbono es una
de las causas de la reducción del rendimiento en el cultivo del arroz. A nivel bioquímico,
altas temperaturas (40/32°C día/noche) también provocan daño en las membranas
(expresado como un mayor contenido de MDA), causando un incremento de la actividad
enzimática antioxidante (superóxido dismutasa, glutatión reductasa) y producción de
osmolitos compatibles como la prolina (Kumar et al., 2011; Xue et al., 2012). En cuanto a
los parámetros de rendimiento del cultivo, Peng et al., (2004) reportan que temperaturas
nocturnas superiores a 24 °C reducen la producción de biomasa total cerca del 20%, el
porcentaje de granos llenos y finalmente, el rendimiento de grano.
El crecimiento y desarrollo del cultivo del arroz es influenciado por la temperatura nocturna
(Shah et al., 2011; Wahid et al., 2007; Glaubitz et al., 2014). En la tabla 1 se recopila algunos
efectos de las temperaturas nocturnas sobre parámetros fisiológicos, de crecimiento y de
rendimiento en arroz.
18 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
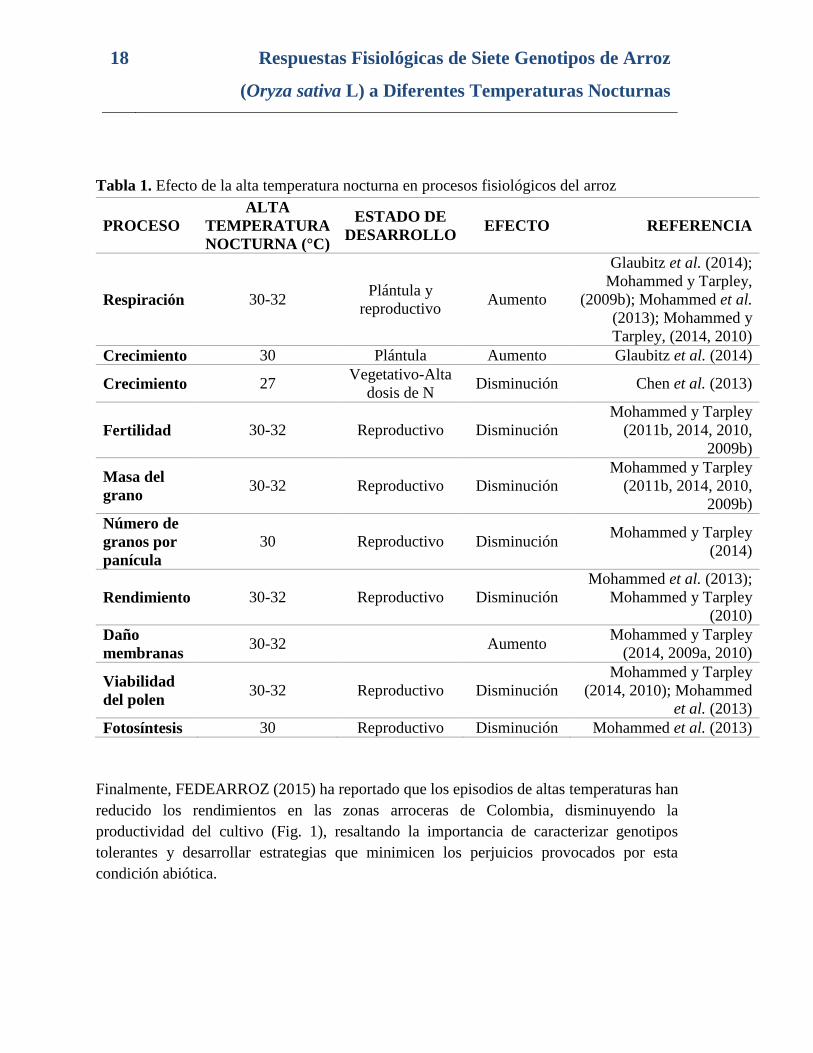
Tabla 1. Efecto de la alta temperatura nocturna en procesos fisiológicos del arroz
PROCESO
ALTA
TEMPERATURA
NOCTURNA (°C)
ESTADO DE
DESARROLLO EFECTO REFERENCIA
Respiración 30-32 Plántula y
reproductivo Aumento
Glaubitz et al. (2014);
Mohammed y Tarpley,
(2009b); Mohammed et al.
(2013); Mohammed y
Tarpley, (2014, 2010)
Crecimiento 30 Plántula Aumento Glaubitz et al. (2014)
Crecimiento 27 Vegetativo-Alta
dosis de N Disminución Chen et al. (2013)
Fertilidad 30-32 Reproductivo Disminución
Mohammed y Tarpley
(2011b, 2014, 2010,
2009b)
Masa del
grano 30-32 Reproductivo Disminución
Mohammed y Tarpley
(2011b, 2014, 2010,
2009b)
Número de
granos por
panícula
30 Reproductivo Disminución Mohammed y Tarpley
(2014)
Rendimiento 30-32 Reproductivo Disminución
Mohammed et al. (2013);
Mohammed y Tarpley
(2010)
Daño
membranas 30-32 Aumento
Mohammed y Tarpley
(2014, 2009a, 2010)
Viabilidad
del polen 30-32 Reproductivo Disminución
Mohammed y Tarpley
(2014, 2010); Mohammed
et al. (2013)
Fotosíntesis 30 Reproductivo Disminución Mohammed et al. (2013)
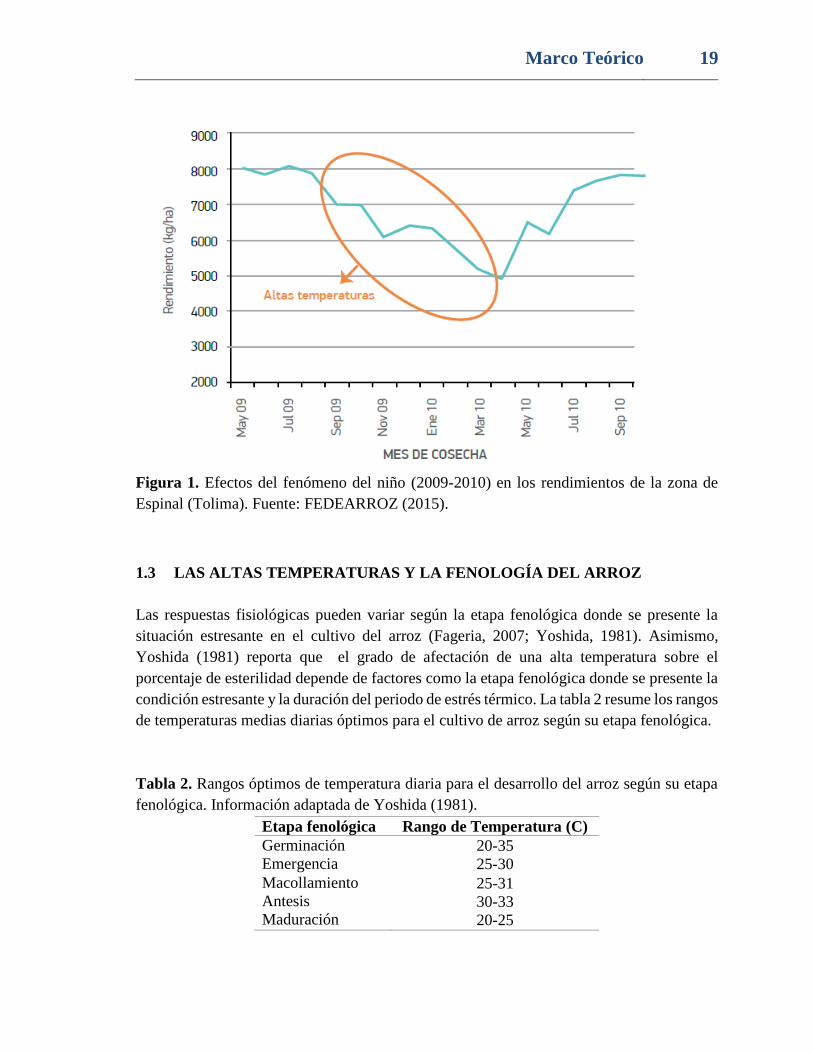
Finalmente, FEDEARROZ (2015) ha reportado que los episodios de altas temperaturas han
reducido los rendimientos en las zonas arroceras de Colombia, disminuyendo la
productividad del cultivo (Fig. 1), resaltando la importancia de caracterizar genotipos
tolerantes y desarrollar estrategias que minimicen los perjuicios provocados por esta
condición abiótica.
Marco Teórico 19
Figura 1. Efectos del fenómeno del niño (2009-2010) en los rendimientos de la zona de
Espinal (Tolima). Fuente: FEDEARROZ (2015).
1.3 LAS ALTAS TEMPERATURAS Y LA FENOLOGÍA DEL ARROZ
Las respuestas fisiológicas pueden variar según la etapa fenológica donde se presente la
situación estresante en el cultivo del arroz (Fageria, 2007; Yoshida, 1981). Asimismo,
Yoshida (1981) reporta que el grado de afectación de una alta temperatura sobre el
porcentaje de esterilidad depende de factores como la etapa fenológica donde se presente la
condición estresante y la duración del periodo de estrés térmico. La tabla 2 resume los rangos
de temperaturas medias diarias óptimos para el cultivo de arroz según su etapa fenológica.
Tabla 2. Rangos óptimos de temperatura diaria para el desarrollo del arroz según su etapa
fenológica. Información adaptada de Yoshida (1981).
Etapa fenológica Rango de Temperatura (C)
Germinación
Emergencia 20-35
25-30
Macollamiento
Antesis
Maduración
25-31
30-33
20-25

20 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
Por otro lado, la literatura existente ha reportado ampliamente la influencia de la relación
etapa fenológica, el genotipo y el estrés por altas temperaturas nocturnas sobre el
rendimiento del cultivo (Fageria, 2007; Jagadish et al., 2014). En este sentido, Mohammed
y Tarpley (2009b) no observaron ningún efecto de una alta temperatura nocturna sobre el
número de macollas, cuando las plantas se sometieron a alta temperatura nocturna desde los
20 días de emergencia hasta la cosecha, mientras Glaubitz et al. (2014) reportan que en 6 de
12 genotipos evaluados la alta temperatura nocturna reduce el macollamiento en plantas de
arroz expuestas a alta temperatura nocturna desde los 20 días después de siembra hasta los
46 días después de siembra.
En cereales, la formación de polen es uno de los estados más sensibles a la alta temperatura.
Durante esta formación, la mitosis 1 y 2 son los estados susceptibles (Barnabás et al., 2008),
siendo el primera (estado uninucleado del polen) el más susceptible a la alta temperatura.
En varias especies (incluida el arroz), la alta temperatura provoca aborto del polen en este
estado. La alta temperatura afecta el metabolismo de la microspora y puede disminuir la
síntesis de proteínas relacionadas con la síntesis de carbohidratos, lípidos, proteínas
antioxidantes y síntesis de la pared celular del polen (Sage et al., 2015). En antesis,
temperaturas nocturnas superiores a 27°C también reducen el peso del grano y disminuyen
la emergencia de la panícula (Coast et al., 2015). En la última fase de desarrollo del cultivo
(Maduración), (Krishnan et al., 2011) reportan que la alta temperatura(34°C) nocturna
reduce la masa del grano en plantas de arroz.
1.4 EL USO DE VARIABLES FISIOLÓGICAS EN EL FITOMEJORAMIENTO
DE ARROZ
Una de las estrategias para afrontar los efectos negativos de las altas temperaturas sobre el
rendimiento de los cereales es la selección de genotipos tolerantes a dicha condición
ambiental (Araus et al., 2008). En este aspecto, el mejoramiento genético de cultivos
actualmente usa características fisiológicas con el propósito de aumentar la eficiencia en la
selección de genes (Reynolds et al., 2009; Jha et al., 2014). Por ejemplo, en la selección de
genotipos de trigo tolerantes a alta radiación y temperatura se han usado los parámetros de
la fluorescencia de la clorofila y variables de intercambio gaseoso (Sayed, 2003; Monneveux
et al., 2003). Adicionalmente, la caracterización de genotipos tolerantes a altas temperaturas
nocturnas tiene en cuenta variables bioquímicas como el contenido de pigmentos
fotosintéticos (clorofilas y carotenoides), la producción de osmolitos compatibles (glicina
betaina, Prolina y azúcares solubles) y la integridad de la membrana celular (Bita y Gerats,

Marco Teórico 21
2013). En arroz, se ha reportado que los genotipos tolerantes a altas temperaturas presentan
una tasa de fotosíntesis, respiración, transporte de electrones, contenido de clorofila y
rendimiento cuántico del fotosistema II estables bajo condiciones de altas temperaturas
nocturnas (Glaubitz et al., 2014; Mohammed y Tarpley, 2014). Por lo anterior, estas
variables han sido sugeridas como herramienta o marcadores para seleccionar genotipos con
una posible tolerancia a diferentes condiciones de estrés abiótico (Sayed, 2003; Araus et al.,
2008).
Actualmente, la utilización de algunos parámetros fisiológicos y bioquímicos como la tasa
fotosintética y producción de prolina han sido utilizados para en la caracterización de
genotipos a condiciones de estrés abióticos especialmente bajo estrés de alta temperatura
diurna en Colombia (Restrepo-Diaz y Garces-Varon, 2013; Sánchez-Reinoso et al., 2014).
Teniendo en cuenta lo anterior, el comportamiento de la planta bajo condiciones de estrés
por una alta temperatura es regulado por varios procesos fisiológicos (Mittler et al., 2012).
Sin embargo, el pobre entendimiento de los complejos mecanismos de tolerancia térmica y
la interacción entre genotipo (G) vs ambiente (A) han sido uno de los cuellos de botella para
el progreso de los programas de mejoramiento genético (Jha et al., 2014). En este sentido,
los estudios con plántulas de arroz han adquirido importancia con el propósito de conocer a
profundidad los mecanismos de respuesta involucrados y así caracterizar el grado de
tolerancia o susceptibilidad de los genotipos a condición de estrés térmico durante los
últimos años (Xue et al., 2012; Sánchez-Reinoso et al., 2014). El conocimiento de la
interacción G x A mediante el estudio de procesos fisiológicos y bioquímicos involucrados
han adquirido importancia porque han permitido el uso de diferentes técnicas en el
mejoramiento clásico en Colombia con el propósito de un mejor direccionamiento y
aprovechamiento del germoplasma desarrollado hasta ahora (FEDEARROZ, 2013).
Se ha demostrado que algunos procesos fisiológicos son más sensibles a la alta temperatura
y que la evaluación de estos se puede usar para diferenciar susceptibilidad o cierta tolerancia
de los genotipos estudiados. Estas variables fisiológicas pueden aportar al mejoramiento
genético: aumentando el conocimiento de los factores que determinan el rendimiento y
definiendo características secundarias (características diferentes al rendimiento) para
seleccionar parentales (Araus et al., 2008).
A continuación, se enuncian los procesos tenidos en cuenta en este trabajo, algunas
definiciones importantes y su relación con un estrés térmico.

22 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
1.4.1 La fotosíntesis y sus componentes
Fluorescencia de la Clorofila a
La fluorescencia de la clorofila y el intercambio gaseoso pueden ser usados para fenotipar
poblaciones menores a 100 individuos, para detectar diferencias cuando otras variables no
las detectan (crecimiento o rendimiento) y para explicar la repuesta a estrés abiotico (Pask
et al., 2012). En estrés por calor, se prefiere el uso de fluorescencia de la clorofila cuando
se dispone de pocos recursos (principalmente tiempo) y cuando se estan estudiando muchos
individuos (Pask et al., 2012).
“Fluorescencia es la re-emisión de energía en forma de fotón cuando un electrón vuelve a
su estado fundamental desde un estado excitado” (Cosgrove y Borowitzka, 2010). En el caso
de la fluorescencia de la clorofila a, una molécula de la clorofila puede excitarse y alcanzar
el estado singlete después de absorber un fotón (Bolhàr-Nordenkampf y Öquist, 1993), y
volver a su estado fundamental luego de reemitir dicha energía en un haz de luz de menor
energía.
El análisis de la fluorescencia de la clorofila a es una herramienta usada para conocer el
comportamiento fotosintético de las plantas, principalmente lo relacionado con la parte
fotoquímica de la fotosíntesis (Brestic y Zivcak, 2013; Guidi y Degl’Innocenti, 2012).
Cuando la energía lumínica es captada por la hoja, se puede disipar en forma de calor, en
forma de fluorescencia o a través de la fotoquímica (Baker, 2008). Estos procesos son
competitivos, por lo que conocer el comportamiento de uno da información acerca de la
eficiencia y el desempeño de los otros dos (Murchie y Lawson, 2013).
Dependiendo del objetivo de la investigación, el análisis de la fluorescencia se puede hacer
a nivel de nanosegundos o de minutos. El primero para conocer el estado de óxido-reducción
de los primeros aceptores de electrones: la quinona A y la quinona B; y lo segundo para
determinar el uso (o disipación(quenching)) de la energía lumínica (Kalaji et al., 2014). Con
base a lo anterior, el segundo enfoque fue utilizado en el presente trabajo de investigación
El análisis de disipación de la energía usa parámetros basados en la emisión de la
fluorescencia en determinadas condiciones. Así, Fm es la fluorescencia máxima emitida
luego de someter la hoja a un haz de luz saturante (generalmente mayor a 3000 µmol m-2 s-
1), mientras Fo es la fluorescencia emitida luego de someter la hoja a un haz de luz no
saturante y con tan baja intensidad (alrededor de 0,1 µmol m-2 s-1) que no desencadene los
mecanismos de disipación de la energía (Baker, 2008). La diferencia entre estas dos

Marco Teórico 23
fluorescencias se denomina fluorescencia variable Fv, y la nomenclatura cuando se
determinan luego de un haz de luz actínica es Fm’, Fo’ y Fv’. Con estos índices y otros más
se determinan parámetros que dan información sobre el estado y la disipación de la energía
en los fotosistemas (Baker, 2008; Brestic y Zivcak, 2013)(Tabla 3):
En arroz, el estrés por calor induce un incremento en NPQ y en el estado de de-epoxidación
del ciclo de las xantofilas (Yin et al., 2010). Una disminución en la asimilación de CO2
acompañada por una no disminución de Fv/Fm inducido por estrés por calor puede
potencialmente exponer las plantas a un exceso de energía bajo condiciones lumínicas, lo
cual si no se disipa de forma segura, puede resultar en un daño del PSII debido a una sobre-
reducción de los centros de reacción (Xue et al., 2012) . Se dice que el PSII es el componente
más sensible a la alta temperatura: el daño puede ser directo o una disminución en los
mecanismos de reparación (Yin et al., 2010). Esto también es reportado por Vani et al.
(2001) quienes argumentan que la alta temperatura desorganiza los tilacoides y los
complejos donadores y aceptores de electrones en el PSII.

24 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
Tabla 3. Parámetros de la fluorescencia derivados de la fluorescencia de la clorofila a
Parámetro Formula Descripción
Fq’, ΔF’ Fm’-F’ Disipación fotoquímica de la
fluorescencia cuando los centros de
reacción del PSII abiertos.
Fv/Fm (Fm-Fo)/Fm Eficiencia máxima potencial de la
fotoquímica del PSII. Se considera que un
valor menor a 0,83 índica algún tipo de
estrés en la planta (Baker, 2008; Murchie
y Lawson, 2013)
ΦPSII, Y(II) Fq’/Fm’ Eficiencia cuántica efectiva (operativa) de
la fotoquímica del PSII a determinada
intensidad lumínica.
Fv’/Fm’ (Fm’-Fo’)/Fm’ Eficiencia máxima potencial de la
fotoquímica del PSII a determinada
intensidad lumínica.
NPQ (Fm-Fm’)/Fm’ disipación no fotoquímica
qP Fq’/Fv’ Coeficiente de la disipación fotoquímica
basado en el “puddle model” (unidades del
PSII desconectadas). Proporción de
centros de reacción oxidados o mayor
proporción del estado reducido de Qa.
qL (Fq’/Fv’)/(Fo’/F’) Coeficiente de la disipación fotoquímica
basado en el “lake model”(unidades del
PSII conectadas).
qN (Fm-Fm’)/(Fm-Fo’) Coeficiente de la disipación no
fotoquímica de la fluorescencia variable.
ΦNO, Y(NO) 1/(NPQ+1+qL(Fm/Fo-1)) Eficiencia cuántica de la disipación no
regulada de la energía en el PSII.
ΦNPQ, Y(NPQ) 1-Y(II)-Y(NO) Eficiencia cuántica de la disipación
regulada de la energía en el PSII. Asociado
con la acidificación del lumen tilacoidal y
con el estado de de-epoxidación del ciclo
de las xantofilas.
La misma tendencia es presentada por Song et al. (2013) quienes encontraron una
disminución de Fv/Fm y Y(II) asociada con un daño en la proteína D1. Además de esto, la
conductancia estomática se incrementó, así como la transpiración y la concentración
intercelular de CO2, sugiriendo que la disminución de la fotosíntesis es debida a una
limitación no estomática. Según este autor un estrés leve (temperatura diurna<40°C) no daña
el PSII. Sin embargo, a dichas temperaturas leves se dañan las proteínas encargadas de

Marco Teórico 25
regenerar dicho fotosistema. El PSII es sensible debido a que la alta temperatura aumenta la
fluidez de la membrana tilacoidal (Farooq et al., 2011).
Teniendo en cuenta lo anterior, mantener alta la tasa fotosintética y bajo NPQ bajo altas
temperaturas puede disminuir la sobre-reducción de la cadena transportadora de electrones
y la producción de H2O2 a través de la fotorespiración (Bahuguna et al., 2015). Por esto
Krishnan et al. (2011) sugieren que una alta eficiencia del fotosistema II (Fv/Fm) y Fo se
correlacionan con la tolerancia a altas temperaturas.
Asimilación de CO2
Muchas investigaciones en arroz han demostrado que la asimilación de CO2 disminuye al
aumentar la temperatura diurna o nocturna (Laza et al., 2015; Mathur et al., 2014; Cui et al.,
2006). Esto generalmente va acompañado de una mayor respiración, menor crecimiento y
menor eficiencia del fotosistema II. Sin embargo, la razón por la cual la asimilación de CO2
disminuye bajo altas temperaturas no está del todo clara.
Las dos razones más aceptadas son una disminución en el estado de activación de la Rubisco
y la baja disponibilidad de RuBP (Kubien y Sage, 2008; Scafaro et al., 2010). La
disminución del estado activo de la Rubisco está asociado con una menor actividad de la
Rubisco activasa, debido a que es más sensible que la Rubisco a la alta temperatura (Salvucci
y Crafts-Brandner, 2004). Por otro lado, la menor disponibilidad de RuBP está asociada con
la disponibilidad de ATP y NADPH (productos de la cadena transportadora de electrones)
y la actividad enzimática del ciclo de Calvin (Yamori et al., 2011).
Kubien y Sage (2008) concluyeron que el aumento del estado de activación de la Rubisco
no necesariamente aumenta la tasa de fijación de CO2, ya que al mantener una alta relación
de Rubisco activasa/Rubisco no se aumenta la tolerancia a la alta temperatura (Kubien y
Sage, 2008). Por otro lado, Wang et al. (2010) establecieron que un mayor estado de
activación de la Rubisco hace más tolerante a las plantas de arroz a una alta temperatura, y
que la isoforma grande de la Rubisco activasa es más importante bajo una alta temperatura.
Lo anterior es soportado por quienes establecieron que O. meridionalis es más tolerante que
O. sativa porque la asociación Rubisco/Rubisco activasa es más estable en O. meridionalis
sometida a altas temperaturas (Scafaro et al., 2012).
Aunque todavía no esté claro el mecanismo por el cual la asimilación de CO2 disminuya por
una alta temperatura. Se ha demostrado que una tasa de asimilación de CO2 estable bajo
condiciones estresantes puede ser un indicador de tolerancia (Ashraf y Harris, 2013; Guidi

26 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
y Degl’Innocenti, 2012). En el caso de la alta temperatura en arroz sucede lo mismo y la
resistencia de los genotipos a la alta temperatura está asociada con una tasa constante de
fotosíntesis y con la capacidad de almacenar carbohidratos en el tallo (Farooq et al., 2011;
Venkateswarlu et al., 2012).
Pigmentos Fotosintéticos
Las clorofilas son pigmentos que absorben la luz entre los 400 y 700nm con una mayor
absorción a los 480nm y entre los 650 y 700nm, la luz verde (entre los 480 y 550nm) no es
absorbida por estos pigmentos (Heldt, 2005; Taiz y Zeiger, 2006). Estos pigmentos captan
la luz y la energía adquirida la transportan a los centros de reacción de los fotosistemas
donde a través de un flujo de electrones, la luz es transformada en energía química (Heldt,
2005).
Los carotenoides son pigmentos isoprenoides con enlaces dobles que absorben la luz en la
región entre 400 y 500nm (Tanaka et al., 2008; Taiz y Zeiger, 2006). Esto pigmentos
cumplen 3 funciones: captan energía y la transfieren a la clorofila (pigmentos accesorios),
protegen el aparato fotosintético de radicales como la clorofila en estrado triplete, el oxígeno
singlete y otros ROS y participan en el ensamblaje de los LHC del PSI y en la estabilización
de las membranas tilacoidales (función estructural) (Gill y Tuteja, 2010; Havaux, 1998).
En cebada un aumento de la proporción CAR/Chl es una adaptación a alta temperatura y
radiación, provocado por una disminución de Chl, principalmente. Esto confirma la idea de
que la disminución de la clorofila es una forma de captar menos energía y evitar el
calentamiento de la hoja, provocado por la reducción en la transpiración (Havaux y Tardy,
1999).
Dentro de las características deseables en el mejoramiento genético para la tolerancia a altas
temperaturas están la producción de pigmentos fotoprotectores, estabilidad membranal,
vigor temprano y senescencia tardía (Cossani y Reynolds, 2012). Lo anterior es soportado
por la respuesta de NL-44, un genotipo tolerante a alta temperatura que mantiene su verdor
relativo (contenido de clorofila) y su capacidad fotosintética, lo que le permite sobrevivir
tanto en estado vegetativo como reproductivo (Bahuguna et al., 2015).

Marco Teórico 27
1.4.2 Reproducción y Rendimiento
El estado reproductivo es el más susceptible a las altas temperaturas, tanto diurnas como
nocturnas. La alta temperatura acelera el desarrollo de la inflorescencia, disminuye la
viabilidad del polen, el crecimiento del tubo polínico y perjudica el desarrollo de los ovarios
(Jagadish et al., 2015).
En cereales, la formación de polen es uno de los estados más sensibles a la alta temperatura.
La mitosis 1 y 2 durante la formación del grano de polen son muy sensibles a las altas
temperaturas (Barnabás et al., 2008). La primera, que corresponde al estado uninucleado del
polen es el más susceptible a la alta temperatura. En varias especies (incluida el arroz), la
alta temperatura provoca aborto del polen en este estado. La alta temperatura afecta el
metabolismo de la microspora y puede disminuir la síntesis de proteína relacionada con
síntesis de carbohidratos, lípidos, proteínas antioxidantes y síntesis de la pared celular del
polen (Sage et al., 2015).
La meiosis, proceso previo a la mitosis durante la microsporogénesis, también puede ser
afectada por la alta temperatura. Un efecto negativo de la alta temperatura en este proceso
disminuye el número de células madre de las microsporas, y por tanto el número de granos
de polen. Finalmente, aunque el polen madure, también puede ser inviable si se presentan
altas temperaturas (>30℃ por minutos u horas) o puede quedarse en las anteras debido a una
menor dehiscencia de estas últimas. Todo esto se traduce en una menor fertilidad de la
espiguilla y menor rendimiento.
La alta temperatura diurna reduce la viabilidad del polen, el crecimiento del tubo polínico,
lo que se traduce en menor fertilidad de la espiguilla. La reciente literatura muestra los
mismos efectos bajo alta temperatura nocturna (Jagadish et al., 2015). Luego de la
polinización, se puede presentar una pobre geminación del polen, (crecimiento del tubo
polínico) debido a la degradación del tejido encargado de la nutrición del tubo polínico o a
la formación de granos de polen inmaduros (Barnabás et al., 2008). Luego de la formación
del polen la alta temperatura puede evitar dehiscencia de las anteras disminuyendo la
cantidad de polen disponible para la polinización (Bokszczanin y Fragkostefanakis, 2013).
El hecho que se afecte el número de granos o el tamaño del grano depende del estado
fenológico en el que se haya presentado la alta temperatura. Si fue durante la formación de
la panícula y la antesis se afectara el número de granos, mientras si es durante el llenado del
grano se afectara el tamaño y peso del grano (Farooq et al., 2011).

28 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
Los genotipos tolerantes tienden a mantener su rendimiento, fertilidad de las espiguillas,
viabilidad del polen y capacidad fotosintética (Poli et al., 2013). Además de estas
características, genotipos tolerantes como N22 acumula heat shock proteins HSP y cold
shock proteins CSP cuando es sometido a un estrés térmico, lo que podría explicar su
tolerancia y alta fertilidad bajo el estrés por alta temperatura (Jagadish et al., 2010).
1.4.3 Peroxidación Lipídica de membranas
Cuando se presenta una condición estresante, la capacidad fotosintética de la planta
disminuye, lo que la hace más susceptible a la fotoinhibición (disminución de la tasa
fotosintética provocada por la luz) (Gururani et al., 2015; Murata et al., 2007). Una
condición estresante generalmente disminuye la asimilación de CO2 por una menor actividad
de la Rubisco, la Rubisco activasa o por una disminución en la proteína D1 del PSII, lo que
provoca fotoinhibición (Gururani et al., 2015).
Durante este proceso los electrones procedentes de la clorofila no pueden ser disipados de
forma segura y se producen especies reactivas de oxígeno (ROS)(Gill y Tuteja, 2010). Estas
especies reactivas pueden dañar lípidos, proteínas y el ADN, lo que produce radicales libres
lipídicos y disminuye la funcionalidad de organelos celulares (Apel y Hirt, 2004).
Una de las formas de medir la peroxidación lipídica es cuantificar la producción de
malondialdehído (MDA). El MDA es el producto de la peroxidación del ácido linolénico, el
cual es uno de los componentes principales de la membrana celular. Esta peroxidación daña
la membrana, disminuyendo su selectividad y aumentando su fluidez (Gill y Tuteja, 2010).
Debido a que la peroxidación lipídica es una de las principales consecuencias de la
producción de especies reactivas de oxígeno provocado por una condición estresante, tanto
los niveles de ROS como los de TBARS (sustancias reactivas con el ácido tiobarbitúrico, un
ejemplo es el MDA ) son considerados indicadores bioquímicos de la severidad del estrés
por calor (Bahuguna et al., 2015).
Teniendo en cuenta lo anterior se considera que la tolerancia a alta temperatura nocturna
(Narayanan et al., 2016; Bahuguna et al., 2015; Zhou et al., 2012), a salinidad (Lutts et al.,
1996) y a una alta temperatura diurna(Zhou et al., 2012) se manifiesta como una menor
peroxidación lipídica (MDA). Asimismo, la pérdida de electrolitos se usa como indicador
del daño membranal ya que correlaciona con la peroxidación lipídica y en varios casos con
la senescencia y capacidad fotosintética de la planta (Mantri et al., 2012).

Marco Teórico 29
1.4.4 Osmolitos compatibles (Prolina)
La prolina es un aminoácido no estructural que generalmente se acumula en plantas cuando
se presenta una condición estresante (Ashraf y Foolad, 2007); la mayor cantidad de reportes
la asocian con salinidad, pero también se acumula cuando se presenta déficit hídrico, alta
radiación, contaminación por metales pesados, deficiencia de nutrientes, temperaturas
extremas y estrés oxidativo (Szabados y Savouré, 2010; Hare y Cress, 1997).
En condiciones de estrés se ha reportado que la prolina cumple las siguientes funciones
(Szabados y Savouré, 2010; Ashraf y Foolad, 2007; Wahid et al., 2007):
- Ajuste osmótico.
- Eliminación de radicales libres.
- Mantenimiento del balance de óxido-reducción (NADPH/NADP).
- En algunos casos promueve la transcripción de genes relacionados con la tolerancia
al estrés.
- Estabilización de estructuras sub-celulares.
Debido a lo anterior la prolina se ha usado para aliviar el estrés (Wahid et al., 2007) o como
un indicador de tolerancia a condiciones estresantes (Szabados y Savouré, 2010). Por
ejemplo Kumar et al. (2011) determinaron que bajo una alta temperatura, plántulas de arroz
y de maíz acumulan mayor cantidad de prolina como una respuesta al estrés provocado por
la alta temperatura. Otro ejemplo lo reporta Yan et al. (2012) en trigo, quienes descubrieron
que el estrés térmico daña la sección donadora y receptora de electrones del PSII y que este
efecto es aliviado por una mayor producción de prolina provocada por una aplicación previa
de sal.
Como se enuncio antes, la alta temperatura puede producir ROS, disminuir la tasa de
transporte de electrones lo que afecta la relación NADPH/NADP (Vani et al., 2001b) y
desorganiza los tilacoides (Vani et al., 2001a). Estos efectos podrían ser aliviados por la
prolina, teniendo en cuenta las funciones de este aminoácido (Szabados y Savouré, 2010).
Aunque, la acumulación de prolina este asociada con funciones que promueven la tolerancia
o alivian los efectos negativos del estrés. Mani et al. (2002) reportan que en mutantes que
sobre producen prolina o que no la degradan, los efectos no son positivos y pueden provocar
malformaciones en las plantas. Teniendo en cuenta esto, Szabados y Savouré (2010)
enuncian que las funciones positivas de la acumulación de prolina pueden estar asociadas a
los procesos de síntesis de prolina y no a la acumulación de la misma.

30 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
Con base en el anterior marco teórico se espera que la alta temperatura nocturna (30°C)
disminuya el crecimiento y rendimiento de los genotipos a evaluar y que las variables
relacionadas con fotosíntesis y componentes de rendimiento sean los mejores indicadores
de tolerancia o susceptibilidad a una alta temperatura nocturna.

Objetivos 31
2. OBJETIVOS
Con el propósito de desarrollar la hipótesis de trabajo, los siguientes objetivos fueron
planteados:
2.1 Objetivo General
Caracterizar el comportamiento fisiológico y agronómico de siete genotipos de arroz
sometidos a una alta temperatura nocturna (30°C).
2.2 Objetivos Específicos
Evaluar el efecto de cuatro diferentes periodos de dos temperaturas nocturnas (24 y
30°C) sobre la fotosíntesis y respiración de un genotipo susceptible a estrés térmico
diurno.
Caracterizar la tasa fotosintética, respiratoria, parámetros de crecimiento y de
rendimiento de siete genotipos de arroz a una alta temperatura nocturna (30°C).
Identificar la utilidad de variables fisiológicas como la fotosíntesis, parámetros de la
fluorescencia de la clorofila a y pruebas bioquímicas como herramientas en la
selección de genotipos de arroz tolerantes a condiciones de altas temperaturas
nocturnas para programas de mejoramiento genético.


Materiales y Métodos 33
3. MATERIALES Y MÉTODOS
Con el fin de cumplir los objetivos propuestos en el presenta trabajo, se desarrollaron 3
experimentos. En el primero, se evaluó el efecto de cuatro periodos de dos temperaturas
nocturnas (24 y 30°C) sobre la fotosíntesis y respiración en un genotipo susceptible a estrés
térmico diurno. En el segundo, se caracterizó la tasa fotosintética, respiratoria y parámetros
de crecimiento, de seis genotipos de arroz en fase vegetativa, a una alta temperatura nocturna
(30°C). En el tercero, se evaluó la eficiencia del PSII y parámetros de rendimiento de siete
genotipos de arroz sometidos a una alta temperatura nocturna (30°C).
A continuación, se enuncian las metodologías usadas en cada uno de los experimentos
citados:
3.1 RESPUESTA FISIOLÓGICA DE PLANTULAS DE ARROZ F60 SOMETIDAS
A PERIODOS DE ALTA TEMPERATURA NOCTURNA.
3.1.1 Condiciones generales del experimento
Se sembraron semillas del cultivar Fedearroz 60 ('F60'): es una variedad con buenos
rendimientos (mayor a 6 t ha-1) en cultivos comerciales, grano largo, buena calidad de
molino pero susceptible a altas temperaturas diurnas (Sánchez-Reinoso et al., 2014), en
materas plásticas con 450 mL de suelo. Las materas se dispusieron en un invernadero de
vidrio localizado en la Universidad Nacional de Colombia, Campus Bogotá. El invernadero
contó con un controlador de humedad relativa y resistencias calefactoras con el fin de
mantener las siguientes condiciones de crecimiento: temperatura diurna 31°C, temperatura
promedio nocturna 22°C, humedad relativa promedio de 70% y fotoperiodo natural de 12 h.
Se dispusieron dos plantas por matera las cuales fueron fertirrigadas diariamente con 50 mL
de una solución nutritiva que contenía un fertilizante completo (Wuxal®, Bayer
CropScience, Bogotá D.C., Colombia) a una dosis de 2 mL L-1 H2O, para una concentración
de nutrientes: Nitrógeno total 160 g L-1, (N amoniacal 38 g L-1, N nítrico 12 g L-1, N ureico
110 g L-1), Fósforo asimilable (P2O5) 160 g L-1, Potasio soluble (K2O) 120 g L-1, Boro (B)
10 g L-1, Cobre (Cu)* 0,21 g L-1, Hierro (Fe) 0,43 g L-1, Manganeso (Mn) 0,36 g L-1,
Molibdeno (Mo) 0,07 g L-1, Zinc (Zn)*:10 g L-1. (* Quelatados con EDTA pH en solución
al 10% 6.5, Densidad a 25°C 1.40 g cm-3).

34 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
3.1.2 Tratamientos
A los 45 días después de emergencia (DE), los tratamientos de estrés térmico fueron
establecidos y consistieron en exponer un grupo de plantas de arroz (48 plantas de las 96
que conformaban el experimento) a una temperatura nocturna de 30°C y otro grupo de
plantas (las restantes 48) a una temperatura nocturna de 24°C entre las 18:00 y 24:00 horas
a cuatro periodos de estrés (4, 8, 12 y 16 días). La temperatura nocturna de 30°C se impuso
mediante el traslado de 48 plantas desde el invernadero a una cámara de crecimiento (MLR-
351H, Sanyo, Bensenville, Illinois, USA) a las 18:00 horas. Luego, las plantas eran
retornadas al invernadero a las 06:00. En cada periodo de muestreo (4, 8, 12 y 16 días) se
tomaron 12 plantas de cada uno de los tratamientos por temperatura (24 y 30°C), para
determinar las variables de respuesta.
Las condiciones de crecimiento en la cámara eran las siguientes: un primer periodo de
temperatura nocturna de 30°C entre las 18:00-24:00 horas y un segundo periodo de 20°C
entre las 00:00 y 06:00 horas. La humedad relativa fue del 70% durante toda la noche. Las
plantas control eran aquellas que siempre estuvieron en el invernadero a lo largo del
experimento (48 plantas) con una temperatura promedio nocturna de 24°C entre las 18 y 24
horas y 20°C entre las 00:00 y 06:00. Asimismo, se seleccionó una alta temperatura nocturna
de 30°C porque es superior a las temperaturas óptimas para el crecimiento y desarrollo de
la planta de arroz (entre 23 y 25°C según Krishnan et al., (2011) y Mohammed y Tarpley,
2014a). Adicionalmente, los cuatro diferentes periodos de estrés térmico (4, 8, 12 y 16 días)
fueron establecidos siguiendo la frecuencia con que se han registrado estas condiciones
ambientales durante los últimos años en las zonas productoras de arroz en Colombia.
Finalmente, el experimento duro 61 días.
3.1.3 Fotosíntesis, conductancia estomática, y transpiración
Estas variables fueron determinadas con un medidor de fotosíntesis portátil (LSPro-SD,
ADC BioScientific Ltd.UK). Las mediciones fueron desarrolladas en la parte media de la
penúltima hoja del tallo principal de cada plántula, entre las 9am y las 12m. (Mohammed
et al., 2013; Mohammed y Tarpley, 2014). Las condiciones de la cámara de medición
fueron: flujo fotónico constante de 1200 µmol m-2 s-1, temperatura promedio de 30±2°C y
concentración de CO2 de 368±10 µmol mol-1 (Temperatura y concentración de CO2
ambientales).

Materiales y Métodos 35
3.1.4 Uso eficiente del agua y eficiencia de carboxilación
Estas dos variables fueron calculadas mediante las ecuaciones descritas por Kumar et
al.(2001) y Sikder et al. (2015)
𝑊𝑎𝑡𝑒𝑟 𝑈𝑠𝑒 𝐸𝑓𝑓𝑖𝑐𝑖𝑒𝑛𝑐𝑦 (𝑊𝑈𝐸) = 𝑃𝑛
𝐸
𝐶𝑎𝑟𝑏𝑜𝑥𝑖𝑙𝑎𝑡𝑖𝑜𝑛 𝐸𝑓𝑓𝑖𝑐𝑖𝑒𝑛𝑐𝑦 (𝐶𝐸) = 𝑃𝑛
𝐶𝑖
Donde Pn, E y Ci son la tasa fotosintética, la transpiración y la concentración de dióxido de
carbono subestomático, respectivamente.
3.1.5 Eficiencia máxima del PSII (Fv/Fm)
La relación se determinó con un fluorómetro no modulado (Handy PEA, Hansatech
Instruments, Kings Lynn, UK) en la misma hoja y a la misma hora que se realizaron las
mediciones de intercambio gaseoso. Para esto, las hojas fueron sometidas a 20 minutos de
oscuridad y luego se aplicó un pulso de luz de 0,1 µmol m-2s-1 para determinar la
fluorescencia mínima (Fo). Posteriormente, se aplicó un pulso de saturación de 3000 µmol
m-2 s-1 y se determinó la fluorescencia máxima Fm. Con estos datos se determinó Fv/Fm =
(Fm/(Fm-Fo)).
3.1.6 Contenido de clorofila a, clorofila b y carotenoides
Cincuenta miligramos de hojas frescas fueron macerados en un mortero con nitrógeno
líquido. Luego, la muestra se llevó a un volumen de 5mL con acetona al 80% y se centrifugó
a 7000g durante 10 minutos. Después, se determinó la absorbancia a longitudes de onda de
663nm, 647nm y 470nm con un espectrofotómetro BioMate 3 UV-Vis (Wisconsin, USA).
Los contenidos de clorofila a, b y carotenoides en hojas (mg L-1) se determinaron mediante
las formulas descritas por Lichtenthaler (1987) y Solarte et al. (2010)

36 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
3.1.7 Respiración oscura de la hoja
Esta variable fue determinada usando un equipo de fotosíntesis portátil LSPro-SD (ADC
BioScientific Ltd. UK.) en la parte media de la penúltima hoja de cada plántula (misma hoja
usada en fotosíntesis) entre las 21:00 y las 24:00 horas, como lo describen Mohammed y
Tarpley (2009b). En ambos tratamientos, se ajustó el flujo fotónico a 0 µmol m-2s-1 y la
temperatura de la cámara a 30°C o 24°C dependiendo del tratamiento. La respiración oscura
se asumió como la pérdida de CO2 proveniente de la hoja.
3.1.8 Contenido de Prolina
El contenido de Prolina se determinó siguiendo el protocolo de Bates et al. (1973)
modificado por Solarte et al. (2010) donde se tomaba 100 mg de tejido vegetal y se
homogenizaban en 5 mL de ácido sulfosalicílico al 3%. Posteriormente, la muestra era
centrifugaba a 4000g durante 30 minutos. Luego, en un tubo falcon se mezcló 1 mL del
sobrenadante con 1 mL de ninhidrina en medio ácido y 1 mL de ácido acético glacial. La
anterior mezcla fue agitada durante 1 minuto y en seguida sometida a baño maría a 95°C
por una hora. Después de este periodo de tiempo, la reacción fue detenida con hielo.
Finalmente, el producto de la reacción fue extraída con 3 mL de tolueno, agitando
vigorosamente los tubos con un vórtex. La absorbancia fue medida a 520 nm usando un
espectrofotómetro (Spectronic BioMate 3 UV-Vis, Thermo, Madison, WI, USA). El
contenido de Prolina se determinó usando una curva de calibración y la siguiente ecuación:
𝑚𝑖𝑐𝑟𝑜𝑚𝑜𝑙𝑒𝑠 𝑑𝑒 𝑃𝑟𝑜𝑙𝑖𝑛𝑎
𝑔𝑟𝑎𝑚𝑜𝑠 𝑑𝑒 𝑚𝑎𝑡𝑒𝑟𝑖𝑎𝑙 𝑣𝑒𝑔𝑒𝑡𝑎𝑙 𝑓𝑟𝑒𝑠𝑐𝑜=
[(
𝜇𝑔 𝑃𝑟𝑜𝑙𝑖𝑛𝑎𝑚𝐿 𝑥 𝑚𝐿 𝑇𝑜𝑙𝑢𝑒𝑛𝑜)
115.5 𝜇𝑔𝜇𝑚𝑜𝑙
]
[𝑔 𝑚𝑢𝑒𝑠𝑡𝑟𝑎
5]
3.1.9 Peroxidación Lipídica
Para la determinación de la peroxidación lipídica (representada como contenido de
Malondialdehído-MDA) se empleó el método del ácido tiobarbitúrico (TBA) descrito por
Hodges et al. (1999): Se homogenizaron 300 mg de hojas completamente desarrolladas con
nitrógeno líquido. Posteriormente, el homogenizado se mezcló con 3 mL de ácido

Materiales y Métodos 37
tricloroacético al 0,1%. La mezcla se centrifugó a 3000g por 10 min. Del sobrenadante se
extrajeron 2 mL para reaccionar en dos tubos de ensayo de la siguiente manera: (a) Con
TBA (+TBA); 1 mL de sobrenadante con 4 mL de ácido tricloroacético al 10% más ácido
tiobarbitúrico al 0,65%, (b) Sin TBA (–TBA); 1 mL de sobrenadante con 4mL de ácido
tricloroacético al 10%. Posteriormente, los tubos de ensayo fueron llevados a baño maría a
95°C durante 25 min y la reacción se detuvo con hielo. Las muestras fueron centrifugadas
nuevamente a 3000 g por 10 min y se leyeron las absorbancias a 440, 532 y 600 nm usando
un espectrofotómetro (BioMate 3 UV - Vis, WI, USA). Todas las variables fisiológicas y
bioquímicas se midieron a los 4, 8, 12 y 16 días después de iniciados los tratamientos tanto
en las plantas sometidas a 30°C como a 24°C. Para obtener la concentración de MDA se usó
la siguiente ecuación, asumiendo un coeficiente de extinción del MDA de 157 M mL-1:
𝐴 = [𝐴𝑏𝑠 532+𝑇𝐵𝐴 − 𝐴𝑏𝑠 600+𝑇𝐵𝐴 − (𝐴𝑏𝑠 532−𝑇𝐵𝐴 − 𝐴𝑏𝑠 600−𝑇𝐵𝐴)]
𝐵 = [(𝐴𝑏𝑠 440+𝑇𝐵𝐴 − 𝐴𝑏𝑠 600+𝑇𝐵𝐴) ∗ 0,0571]
𝑀𝐷𝐴 𝑒𝑞𝑢𝑖𝑣𝑎𝑙𝑒𝑛𝑡𝑒𝑠 (𝑛𝑚𝑜𝑙 𝑚𝐿−1) = (𝐴 − 𝐵157000⁄ ) ∗ 106
3.1.10 Análisis Estadístico
Los datos fueron analizados mediante un diseño factorial (temperatura nocturna (24 y 30°C)
vs tiempo de exposición (4, 8, 12 y 16 días) para un total de 8 tratamientos. Cada tratamiento
estuvo compuesto por 6 unidades experimentales y cada unidad experimental correspondió
a dos plantas. A los datos obtenidos, se les realizaron las pruebas de Kolmogorov-Smirnov
y de Levene para comprobar normalidad y homogeneidad de varianza, respectivamente.
Luego, se desarrolló un análisis de varianza ANOVA. Cuando se observaron diferencias
significativas, se realizó una prueba de Tukey o contrastes polinómicos con el programa
SPSS (v20.0, IBM Company, USA).

38 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
3.2 RESPUESTA FISIOLÓGICA DE SEIS GENOTIPOS DE ARROZ EN
ESTADO VEGETATIVO A UNA ALTA TEMPERATURA NOCTURNA
3.2.1 Tratamientos
Para el desarrollo del estudio se usó un genotipo susceptible a una alta temperatura diurna
('F60̍) (Sánchez-Reinoso et al. 2014) y cinco genotipos caracterizados por buen
comportamiento bajo condiciones de altas temperaturas tanto diurnas como nocturnas
utilizadas por la Federación Nacional de Arroceros dentro de sus programas de
fitomejoramiento (IR 1561, FLO 2764, LV447-1, CT19021, LV1401). Cuando las plantas
alcanzaron los 32 días después de emergencia (DE), estas fueron divididas en dos grupos de
16 plantas cada uno. El primer grupo siempre permaneció en el invernadero. Mientras el
segundo grupo fue ubicado en una cámara de crecimiento (MLR-351H, Sanyo, Bensenville,
Illinois, USA) a una temperatura de 30°C entre las 18:00 y 24:00 h y de 20°C entre las 00:01
y 06:00 por un periodo de 8 días. Cuando finalizaba el periodo de exposición en la cámara
de crecimiento, las plantas de arroz siempre eran retornadas a las condiciones de
invernadero. El experimento termino a los 40 DE.
3.2.2 Condiciones generales de crecimiento
El ensayo se desarrolló bajo condiciones de invernadero ubicado en la Facultad de Ciencias
Agrarias de la Universidad Nacional de Colombia, sede Bogotá. Las condiciones de
crecimiento en el invernadero fueron las siguientes: temperatura diurna 30°C, temperatura
nocturna 24°C, humedad relativa promedio de 70% y un fotoperiodo natural de 12 h. Se
dispusieron dos plantas por matera de 450 mL de los diferentes genotipos a estudiar, las
cuales fueron fertirrigadas diariamente con 80 mL de una solución nutritiva que contenía un
fertilizante completo (Wuxal®, Bayer CropScience, Bogotá D.C., Colombia) a una
concentración de 2 mL L-1 H2O, siendo la concentración del fertilizante la siguiente:
Nitrógeno total 160 g L-1, (N amoniacal 38 g L-1, N nítrico 12 g L-1, N ureico 110 g L-1),
Fósforo asimilable (P2O5) 160 g L-1, Potasio soluble (K2O) 120 g L-1, Boro (B) 10 g L-1,
Cobre (Cu)* 0,21 g L-1, Hierro (Fe)0,43 g L-1, Manganeso (Mn)0,36 g L-1, Molibdeno (Mo)
0,07 g L-1, Zinc (Zn)* 10 g L-1. (*Quelatados con EDTA pH de 6,5 en solución al 10%,
Densidad a 25°C 1,40 g cm-3).

Materiales y Métodos 39
3.2.3 Fotosíntesis
Entre los 37 y 40 DE, La penúltima hoja (de la base al ápice) del tallo principal de cada
planta fue utilizada para determinar la tasa asimilación de CO2 mediante un medidor portátil
de fotosíntesis (LI-COR 6200, Lincoln, Nebraska, USA). Durante las medidas de
fotosíntesis, las condiciones en la cámara de medición fueron las siguientes: Radiación
Fotosintéticamente Activa (PAR) mayor a 800 μmol m-2 s-1, temperatura de la hoja de 27 ±
5 °C y diferencia presión de vapor de agua del aire y la hoja 1,8 ± 0,5 kPa. La medición de
la tasa fotosintética se realizó entre las 09:00 y las 12:00 h. Finalmente, para la
determinación de la tasa de asimilación de CO2 se tomaron ocho plantas por tratamiento
(doce tratamientos: 2 temperaturas x 6 genotipos).
3.2.4 Eficiencia de Carboxilación y Uso eficiente del agua
La eficiencia de la carboxilación se determino mediante la ecuación de Sikder et al. (2015)
nombrada en el apartado 3.1.4, y el uso eficiente del agua así:
𝑊𝑎𝑡𝑒𝑟 𝑈𝑠𝑒 𝐸𝑓𝑓𝑖𝑐𝑖𝑒𝑛𝑐𝑦 (𝑊𝑈𝐸𝑖) = 𝑃𝑛
𝑔𝑆
Donde Pn corresponde a la tasa fotosintética, gS a la conductancia estomática y WUEi al
uso eficiente del agua Intrínseco.
3.2.5 Respiración oscura
Para determinar esta variable se siguió la metodología descrita por Sánchez-Reinoso et al.
(2014) que consiste en introducir las plantas en cámaras herméticas plásticas de 2 L de
capacidad. Posteriormente, sensores de Gas CO2 infrarrojo (CO2-BTA, Vernier, Beaverton,
OR, USA) fueron insertados en las cámaras y conectados a una interface portátil (LabQuest,
Vernier, Beaverton, OR, USA). Antes de introducir las plantas en las cámaras herméticas,
todas las macetas fueron cubiertas con una bolsa plástica para evitar la respiración del suelo
con el propósito de cuantificar únicamente la respiración del tejido vegetal. Luego, las
cámaras fueron colocadas en un cuarto oscuro por 300 segundos (s). Durante este periodo,
el sensor estimó cada 4 s la concentración de CO2 (mg kg-1) en la cámara, obteniéndose una
curva de tendencia lineal. Para obtener la tasa de respiración de la planta (mmol CO2 kg PF -1 h-1) se determinó la pendiente de la regresión lineal (mg kg-1/s) y se calculó sobre el peso
fresco total de la planta. Las mediciones fueron desarrolladas en las mismas fechas y el
mismo número de plantas donde se obtuvieron las medidas de fotosíntesis.

40 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
3.2.6 Parámetros de Fluorescencia de la clorofila a
Estos parámetros se calcularon en la parte media de la penúltima hoja de cada planta (las
mismas utilizadas para la estimación de la Fotosíntesis) entre las 9:00 am y las 13:00 horas
justo al finalizar el ensayo (40 DE). La fluorescencia de la clorofila fue estimada con un
fluorómetro modulado MINI-PAM (Heinz Walz, Effieltrich, Germany). Las hojas se
mantuvieron en la oscuridad por 20 minutos antes de medir Fv/Fm. La fluorescencia mínima
(Fo) fue medida con un pulso de luz modulado menor a 50 µmol m-2 s-1 y la máxima
fluorescencia fue inducida por un pulso de 0,8s con una intensidad de 3000 µmol m-2 s-1. El
rendimiento cuántico potencial del PSII (Fv/Fm) fue determinado así: Fv/Fm = (Fm-Fo)/Fm.
Para alcanzar el estado basal de la fotosíntesis (Fs’) las muestras se sometieron a una luz
actínica de 900 µmol m-2 s-1 y luego de esto se sometieron a un segundo pulso saturante
(3000 µmol m-2 s-1 por 0,8s) para obtener la máxima fluorescencia en estado adaptado a la
luz (Fm’). Se calculó Fo’ mediante la fórmula de Oxborough y Baker (1997): Fo’= 1/(1/Fo-
1/Fm+1/Fm’). Con las variables anteriores y de acuerdo con Brestic y Zivcak (2013) se
estimó: el rendimiento cuántico real del fotosistema II (ΦPSII= Fm’-F’/Fm’), Rendimiento
cuántico de la disipación de la energía no regulada (ΦNO = 1/ (NPQ+1+qL(Fm/Fo-1)),
rendimiento cuántico de la disipación de la energía regulada(ΦNPQ = 1 - ΦPSII – ΦNO),
disipación no fotoquímica (NPQ = (Fm-Fm’)/Fm’), coeficiente de disipación fotoquímica
basado en el “Lake model” (qL = ((Fm’-Fs’)/(Fm’-Fo’)*(Fo/Fs’) y coeficiente de disipación
no fotoquímica de la fluorescencia variable (qN = Fm-Fm’/Fm-Fo’).
3.2.7 Parámetros de Crecimiento
Al finalizar los tratamientos, cuatro plantas por tratamiento se sometieron a 70°C por 48h
con el fin de determinar la acumulación de masa seca por órgano (tallo+vaina foliar y lamina
foliar). Se determinó el área foliar de las láminas foliares de cada planta usando un medidor
de área foliar LI-3000C (LI-COR Lincoln, Nebraska, USA). Con las anteriores variables se
determinó masa foliar especifica (relación masa foliar/área foliar de cada planta) y la
relación hojas/tallo (masa de hojas/masa de tallo+vaina foliar por planta)
3.2.8 Pérdida de electrolitos
Esta variable fue determinada mediante el protocolo del porcentaje de fuga de electrolitos
descrito por Sanchez-Reinoso et al. (2014). Después del período de estrés a los 40 DE,
aproximadamente 300 mg de hojas frescas fueron cortados en trozos de 1 cm de longitud y

Materiales y Métodos 41
colocadas en tubos de ensayo con 30 mL de agua desionizada. Posteriormente, los tubos
fueron incubados en baño maría a 30°C por 2 h y se determinó la conductividad eléctrica
inicial (CE1). Posteriormente, las muestras fueron sometidas a 100°C en baño maría por 15
min para liberar todos los electrolitos. Finalmente, se enfrió la muestra en cama de hielo y
se determinó la conductividad eléctrica final (CE2). El porcentaje de electrolitos se calculó
así:
%𝐸𝑙𝑒𝑐𝑡𝑟𝑜𝑙𝑖𝑡𝑜𝑠 = (𝐸𝐶1
𝐸𝐶2) 𝑥 100
3.2.9 Clorofila y Carotenoides
A los 40 DE, trozos de lámina foliar de 1 cm de longitud (50 mg de peso fresco) de una hoja
completamente desarrollada fueron macerados usando nitrógeno líquido y almacenados a -
80 °C hasta que se realizaron los respectivos análisis (descrito en el apartado 3.1.6).
3.2.10 Prolina y Malondialdehído (MDA)
Después de los ocho días del periodo de estrés (40 DE), se maceraron 300 mg de hojas
completamente desarrolladas del tallo principal con nitrógeno líquido y se conservaron a
una temperatura de -80°C hasta su análisis: el cual corresponde al del apartado 3.1.8.
Para la determinación de la peroxidación lipídica (representada como contenido de MDA)
se empleó el método del ácido tiobarbitúrico (TBA) descrito por Hodges et al. (1999), el
cual también es descrito en el apartado 3.1.9.
3.2.11 Diseño experimental y análisis estadístico
Se realizó un diseño en arreglo factorial con ocho plantas por tratamiento, donde el factor
principal fueron los genotipos (F60, LV447, FLO2764, LV1401, CT19021, IR1561) y el
factor secundario fue la temperatura nocturna (24 vs 30 °C). Cuando se presentaron
diferencias significativas, se usó el test de Tukey como prueba de comparación de medias.
Los datos se analizaron usando el programa SPSS (v20.0, IBM Company, USA).

42 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
3.3 CARACTERIZACIÓN FISIOLÓGICA DE SIETE GENOTIPOS DE ARROZ
SOMETIDOS A UNA ALTA TEMPERATURA NOCTURNA
3.3.1 Condiciones generales de crecimiento
El ensayo fue desarrollado bajo condiciones de casa de mallas en el Centro de Investigación
Las Lagunas de la Federación Nacional de Arroceros (FEDEARROZ) ubicado en el
municipio de Saldaña (Colombia) (3º54’46’’ N; 74º59’7’’ W) entre Septiembre de 2014 y
Enero de 2015. En bandejas plásticas de 9L de capacidad se sembraron seis plantas por
genotipo. En cada bandeja se usó suelo franco arenoso como sustrato y cada planta fue
fertilizada con 221mg de N (157 Kg ha-1), 15mg de P (54 Kg ha-1), 215mg de K (142 Kg ha-
1), 71mg de S (13 Kg ha-1) y 23 mg de Zn (12 Kg ha-1). Las condiciones ambientales a lo
largo del experimento fueron las siguientes: temperatura diurna promedio de 33°C,
temperatura nocturna promedio de 23.5°C, Humedad relativa del 77% y un fotoperiodo
natural de 12 horas. Los anteriores valores fueron registrados por una estación climática
(Davis Vantage Pro 2 Plus, NSW, AUS) ubicada en la zona de experimento.
3.3.2 Genotipos y tratamientos
Para el desarrollo del presente estudio, se usaron dos variedades comerciales de arroz
susceptibles a condiciones de estrés térmico (Fedearroz 50 y 60) (Restrepo-Diaz y Garces-
Varon, 2013; Sánchez-Reinoso et al., 2014). Adicionalmente, se seleccionaron cinco líneas
utilizadas por la Federación Nacional de Arroceros dentro de sus programas de
fitomejoramiento (IR 1561, FLO 2764, LV447-1, CT19021, LV1401). Estas líneas fueron
seleccionadas porque han mostrado un buen comportamiento bajo condiciones de altas
temperaturas tanto diurnas como nocturnas. Los tratamientos correspondieron en separar
cada material genético en dos grupos de seis plantas cada uno. Un primer grupo (Plantas
Control) siempre estuvo situado en la casa de mallas a una temperatura nocturna ambiental
de ̴24°C. El segundo grupo de plantas de arroz fue ubicado en una cámara de crecimiento
(KBW-400, Binder, Germany) a una temperatura de 30°C entre las 18:00 y 24:00h por un
periodo de 8 días. Al noveno día las plantas fueron retornadas a la casa de mallas y se
estimaron las variables de respuesta fisiológica y bioquímica. Asimismo, la exposición de
las plantas a 30°C fue desarrollada cuando estas estaban en el estado fenológico grano
lechoso. Debido a las diferencias en el desarrollo fenológico de los genotipos evaluados, las
plantas fueron ubicadas en diferentes fechas en la cámara de crecimiento: IR 1561 y F50 el

Materiales y Métodos 43
23 de diciembre de 2014, LV447-1 y FLO 2764 el 30 de Diciembre de 2014, LV1401 y
CT19021 el 6 de enero de 2015 y F60 el 15 de enero de 2015.
3.3.3 Componentes de rendimiento
Al final del ciclo de cada genotipo se tomaron las panículas de cada planta y se determinaron
por panícula los siguientes componentes: masa de granos llenos, número total de granos,
número de granos vanos, número de granos llenos (presionando entre los dedos se determinó
completamente y parcialmente llenos), porcentaje de fertilidad (expresado como la relación
entre granos llenos y número total de sitios reproductivos) y masa promedio de grano.
3.3.4 Parámetros de Fluorescencia de la Clorofila a
Esta variable se determinó en la parte media de la hoja bandera de cada planta en ambos
tratamientos de temperaturas nocturnas entre las 9:00am y la 1:00pm, al día siguiente de
finalizar el periodo térmico de las plantas expuestas a 30°C en cada genotipo. Los
parámetros de fluorescencia de la clorofila a fueron estimados como se describe en el
apartado 3.2.6
3.3.5 Contenido de Clorofila y Carotenoides
Luego de la determinación de los parámetros de fluorescencia de la clorofila a, las hojas
bandera de cada panícula fueron cortadas y almacenadas bajo refrigeración un máximo de
12 horas. Posteriormente, las muestras fueron maceradas usando nitrógeno líquido y
almacenadas a -80 °C hasta que se realizaron las respectivas determinaciones (descrito en el
apartado 3.1.6).
3.3.6 Prolina y Malondialdehído (MDA)
Para el análisis de estas dos variables se usaron muestras de las hojas bandera del tallo
principal almacenadas a una temperatura de -80°C hasta su análisis: el cual corresponde a
los apartados 3.1.8 y 3.1.9

44 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
3.3.7 Análisis Estadístico
Para el análisis estadístico se usó un diseño factorial, donde el primer factor correspondió a
la temperatura nocturna (24 vs 30°C) y el segundo factor al genotipo (F50, F60, IR 1561,
FLO 2764, LV447-1, CT19021 y LV1401), para un total de 14 tratamientos (cada uno con
6 unidades experimentales representada por una planta). A los datos obtenidos se les realizo
la prueba de normalidad de Kruskal-Wallis y de homogeneidad de varianzas de Levene
previamente al análisis de varianza (ANOVA). Cuando se observaron diferencias se
realizaron la prueba de comparación de medias de Tukey. El análisis de datos fue
desarrollado con el programa estadístico SPSS (v20.0, IBM Company, USA).

Resultados 45
4. RESULTADOS
4.1 RESPUESTA FISIOLÓGICA DE PLANTULAS DE ARROZ SOMETIDAS A
PERIODOS DE ALTA TEMPERATURA NOCTURNA.
4.1.1 Fotosíntesis, conductancia estomática, transpiración y concentración interna de
CO2
Según el análisis polinomico la tasa fotosintética (Pn) de plantas control (24°C) no se ajustó
significativamente a una ecuación lineal, cuadrática ni cúbica (Fig. 2A). Sin embargo, se
observó un comportamiento cúbico (P≤0,01) en plantas sometidas a altas temperaturas
nocturnas (ATN), presentándose una disminución drástica de la tasa de asimilación de
carbono en plantas de arroz sometidas a ATN durante 16 días. En general, la Pn disminuyó
alrededor de un 12% en plantas de arroz sometidas a temperaturas nocturnas de 30°C en
comparación al control (24°C) (Fig. 2A). En relación a la conductancia estomática (gS),
también se presentó un comportamiento cúbico en plantas sometidas a ATN, con una
disminución a los 8 y 16 días de exposición (DDE), en comparación al control (24°C) (Fig.
2B). Con respecto a la transpiración (E), esta variable también mostró un comportamiento
cúbico en ambas temperaturas, con una mayor transpiración a los 8 y 12 días en plantas
sometidas a 30°C. Sin embargo, se observó una disminución de la tasa de transpiración a los
16 DDE en las dos condiciones de temperaturas nocturnas (Fig. 2C). Por otro lado, la
concentración interna de CO2 (Ci) también fue mayor en plantas sometidas a 30 °C a los 4,
8 y 12 DDE y presentó una respuesta cúbica en ambas temperaturas (P≤0,01). Sin embargo,
estas diferencias no se mantuvieron al final del experimento (16 DDE) (Fig. 2D).
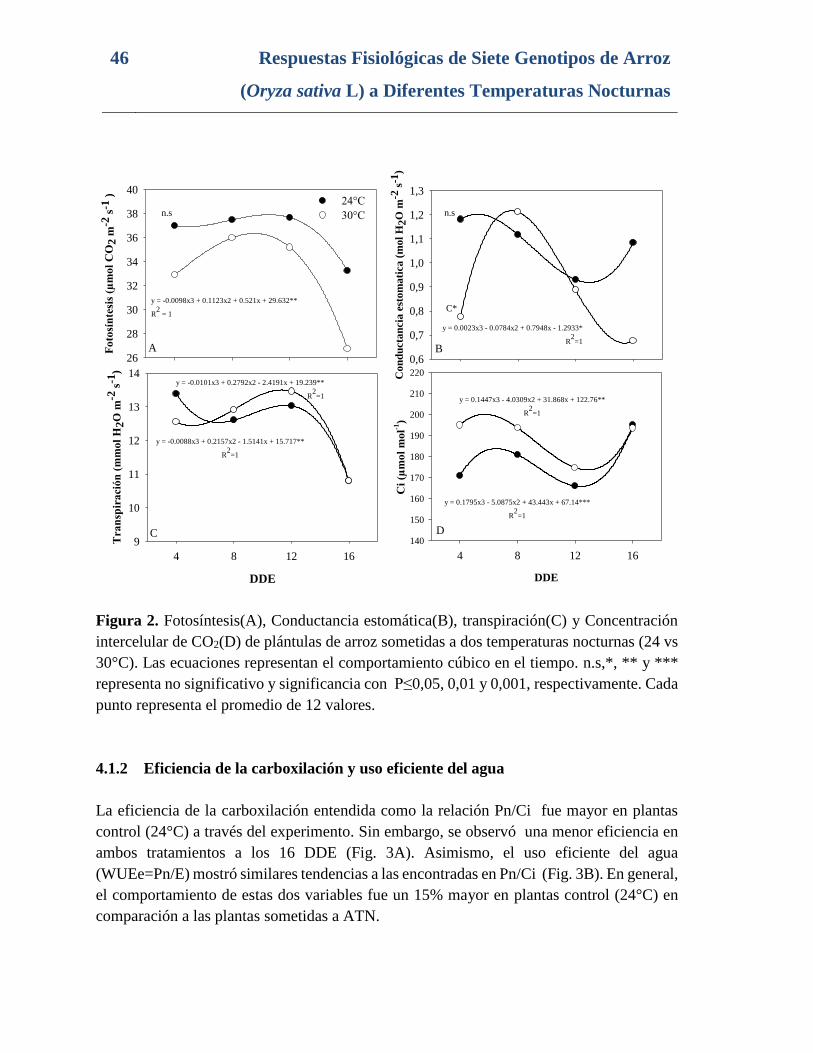
46 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
DDE
4 8 12 16
Tra
nsp
ira
ció
n (
mm
ol
H2O
m-2
s-1
)
9
10
11
12
13
14 Con
du
cta
ncia
est
om
ati
ca (
mol
H2O
m-2
s-1
)
0,6
0,7
0,8
0,9
1,0
1,1
1,2
1,3
Fo
tosí
nte
sis
(µm
ol
CO
2 m
-2 s
-1 )
26
28
30
32
34
36
38
4024°C
30°C
A B
C*
n.s
DDE
4 8 12 16
Ci
(µm
ol
mo
l-1)
140
150
160
170
180
190
200
210
220
DC
y = 0.0023x3 - 0.0784x2 + 0.7948x - 1.2933*
R2
=1
y = -0.0098x3 + 0.1123x2 + 0.521x + 29.632**
R2
= 1
y = -0.0101x3 + 0.2792x2 - 2.4191x + 19.239**
R2
=1
y = -0.0088x3 + 0.2157x2 - 1.5141x + 15.717**
R2
=1
y = 0.1447x3 - 4.0309x2 + 31.868x + 122.76**
R2
=1
y = 0.1795x3 - 5.0875x2 + 43.443x + 67.14***
R2
=1
n.s
Figura 2. Fotosíntesis(A), Conductancia estomática(B), transpiración(C) y Concentración
intercelular de CO2(D) de plántulas de arroz sometidas a dos temperaturas nocturnas (24 vs
30°C). Las ecuaciones representan el comportamiento cúbico en el tiempo. n.s,*, ** y ***
representa no significativo y significancia con P≤0,05, 0,01 y 0,001, respectivamente. Cada
punto representa el promedio de 12 valores.
4.1.2 Eficiencia de la carboxilación y uso eficiente del agua
La eficiencia de la carboxilación entendida como la relación Pn/Ci fue mayor en plantas
control (24°C) a través del experimento. Sin embargo, se observó una menor eficiencia en
ambos tratamientos a los 16 DDE (Fig. 3A). Asimismo, el uso eficiente del agua
(WUEe=Pn/E) mostró similares tendencias a las encontradas en Pn/Ci (Fig. 3B). En general,
el comportamiento de estas dos variables fue un 15% mayor en plantas control (24°C) en
comparación a las plantas sometidas a ATN.
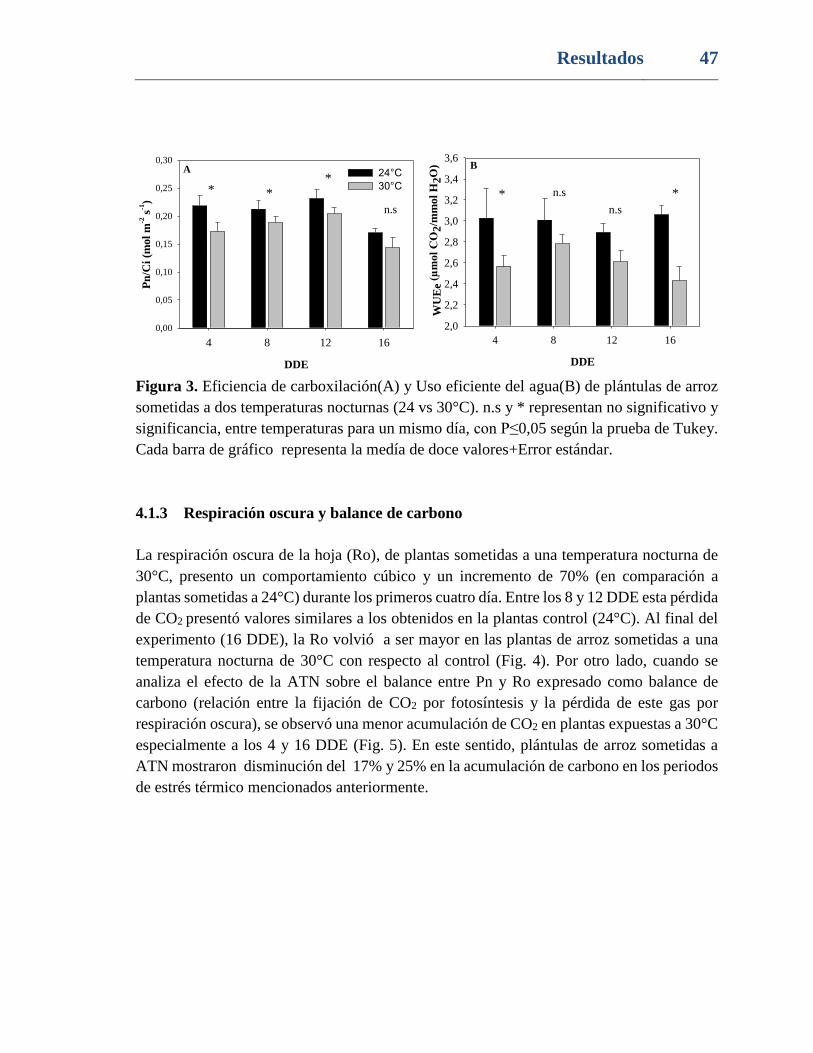
Resultados 47
DDE
4 8 12 16
WU
Ee
(µm
ol
CO
2/m
mol
H2O
)
2,0
2,2
2,4
2,6
2,8
3,0
3,2
3,4
3,6
DDE
4 8 12 16
Pn
/Ci
(mol
m-2
s-1
)
0,00
0,05
0,10
0,15
0,20
0,25
0,30
24°C
30°C * **
**
C*
A B
n.s
n.s
n.s
Figura 3. Eficiencia de carboxilación(A) y Uso eficiente del agua(B) de plántulas de arroz
sometidas a dos temperaturas nocturnas (24 vs 30°C). n.s y * representan no significativo y
significancia, entre temperaturas para un mismo día, con P≤0,05 según la prueba de Tukey.
Cada barra de gráfico representa la medía de doce valores+Error estándar.
4.1.3 Respiración oscura y balance de carbono
La respiración oscura de la hoja (Ro), de plantas sometidas a una temperatura nocturna de
30°C, presento un comportamiento cúbico y un incremento de 70% (en comparación a
plantas sometidas a 24°C) durante los primeros cuatro día. Entre los 8 y 12 DDE esta pérdida
de CO2 presentó valores similares a los obtenidos en la plantas control (24°C). Al final del
experimento (16 DDE), la Ro volvió a ser mayor en las plantas de arroz sometidas a una
temperatura nocturna de 30°C con respecto al control (Fig. 4). Por otro lado, cuando se
analiza el efecto de la ATN sobre el balance entre Pn y Ro expresado como balance de
carbono (relación entre la fijación de CO2 por fotosíntesis y la pérdida de este gas por
respiración oscura), se observó una menor acumulación de CO2 en plantas expuestas a 30°C
especialmente a los 4 y 16 DDE (Fig. 5). En este sentido, plántulas de arroz sometidas a
ATN mostraron disminución del 17% y 25% en la acumulación de carbono en los periodos
de estrés térmico mencionados anteriormente.
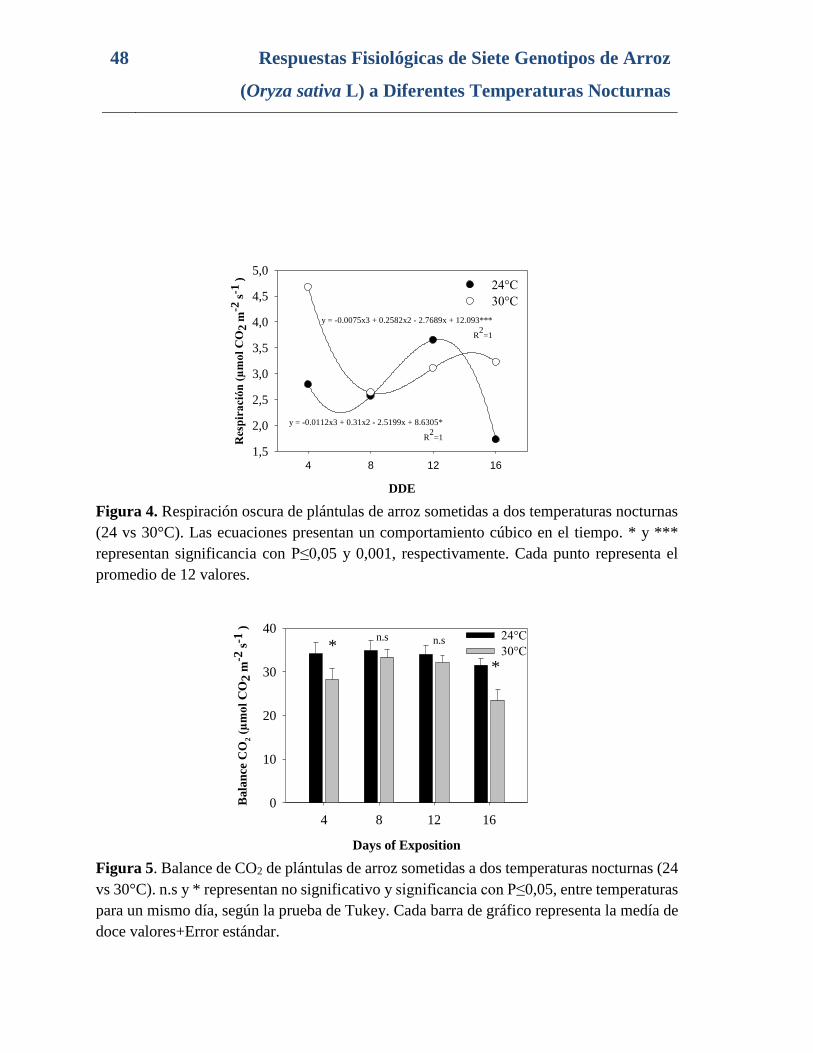
48 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
DDE
4 8 12 16
Resp
ira
ció
n (
µm
ol
CO
2 m
-2 s
-1 )
1,5
2,0
2,5
3,0
3,5
4,0
4,5
5,024°C
30°C
y = -0.0075x3 + 0.2582x2 - 2.7689x + 12.093***
R2
=1
y = -0.0112x3 + 0.31x2 - 2.5199x + 8.6305*
R2
=1
Figura 4. Respiración oscura de plántulas de arroz sometidas a dos temperaturas nocturnas
(24 vs 30°C). Las ecuaciones presentan un comportamiento cúbico en el tiempo. * y ***
representan significancia con P≤0,05 y 0,001, respectivamente. Cada punto representa el
promedio de 12 valores.
Days of Exposition
4 8 12 16
Ba
lan
ce C
O2 (
µm
ol
CO
2 m
-2 s
-1 )
0
10
20
30
4024°C
30°C *
*
n.s n.s
Figura 5. Balance de CO2 de plántulas de arroz sometidas a dos temperaturas nocturnas (24
vs 30°C). n.s y * representan no significativo y significancia con P≤0,05, entre temperaturas
para un mismo día, según la prueba de Tukey. Cada barra de gráfico representa la medía de
doce valores+Error estándar.
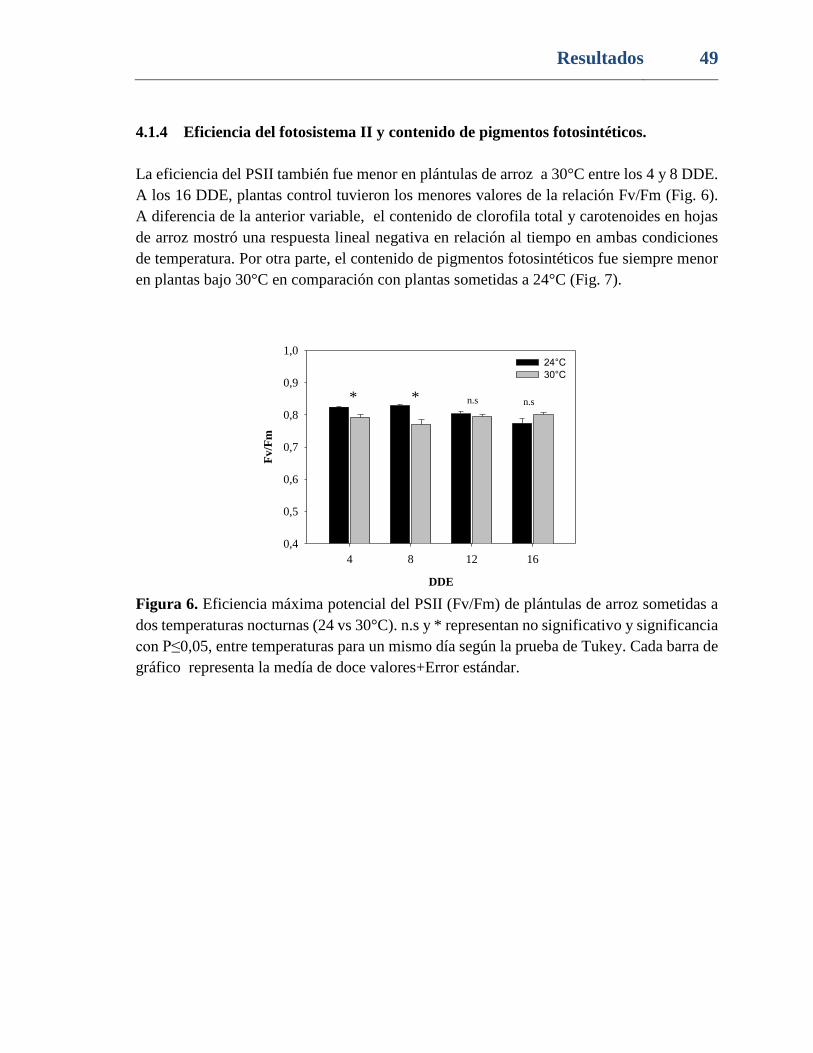
Resultados 49
4.1.4 Eficiencia del fotosistema II y contenido de pigmentos fotosintéticos.
La eficiencia del PSII también fue menor en plántulas de arroz a 30°C entre los 4 y 8 DDE.
A los 16 DDE, plantas control tuvieron los menores valores de la relación Fv/Fm (Fig. 6).
A diferencia de la anterior variable, el contenido de clorofila total y carotenoides en hojas
de arroz mostró una respuesta lineal negativa en relación al tiempo en ambas condiciones
de temperatura. Por otra parte, el contenido de pigmentos fotosintéticos fue siempre menor
en plantas bajo 30°C en comparación con plantas sometidas a 24°C (Fig. 7).
DDE
4 8 12 16
Fv
/Fm
0,4
0,5
0,6
0,7
0,8
0,9
1,024°C
30°C
n.s n.s* *
Figura 6. Eficiencia máxima potencial del PSII (Fv/Fm) de plántulas de arroz sometidas a
dos temperaturas nocturnas (24 vs 30°C). n.s y * representan no significativo y significancia
con P≤0,05, entre temperaturas para un mismo día según la prueba de Tukey. Cada barra de
gráfico representa la medía de doce valores+Error estándar.
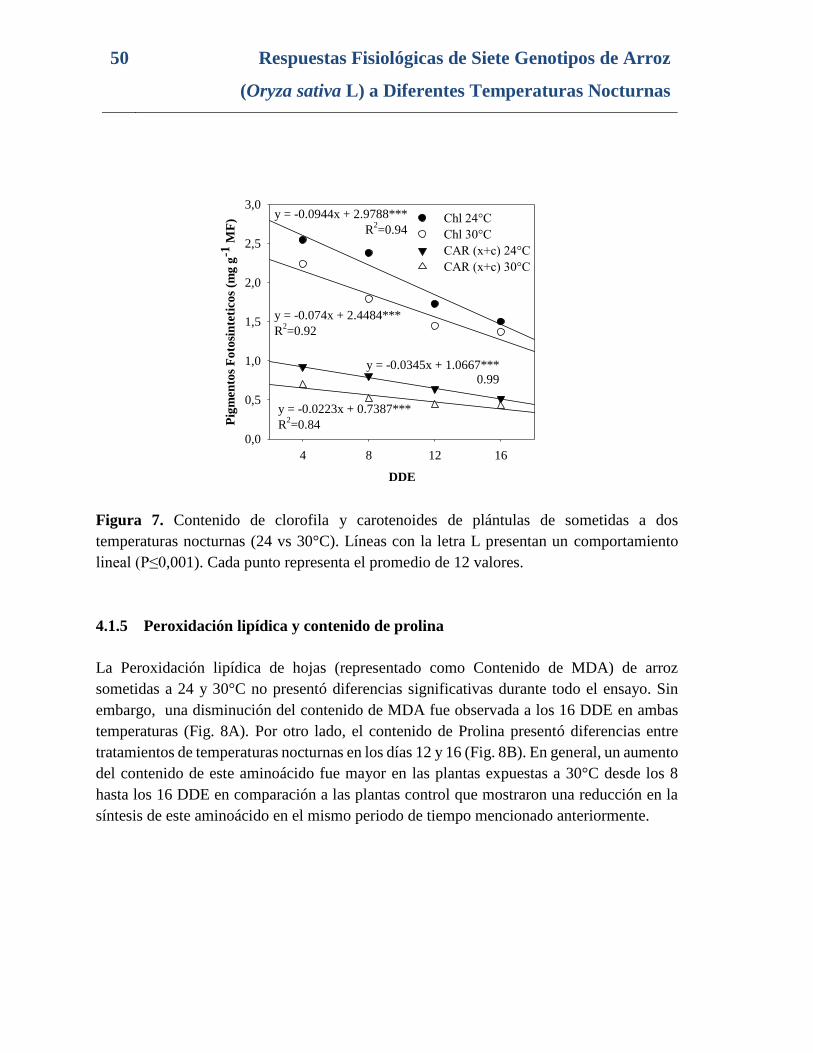
50 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
DDE
4 8 12 16
Pig
men
tos
Fo
tosi
nte
tico
s (m
g g
-1 M
F)
0,0
0,5
1,0
1,5
2,0
2,5
3,0Chl 24°C
Chl 30°C
CAR (x+c) 24°C
CAR (x+c) 30°C
y = -0.0944x + 2.9788***
R2=0.94
y = -0.074x + 2.4484***
R2=0.92
y = -0.0345x + 1.0667***0.99
y = -0.0223x + 0.7387***
R2=0.84
Figura 7. Contenido de clorofila y carotenoides de plántulas de sometidas a dos
temperaturas nocturnas (24 vs 30°C). Líneas con la letra L presentan un comportamiento
lineal (P≤0,001). Cada punto representa el promedio de 12 valores.
4.1.5 Peroxidación lipídica y contenido de prolina
La Peroxidación lipídica de hojas (representado como Contenido de MDA) de arroz
sometidas a 24 y 30°C no presentó diferencias significativas durante todo el ensayo. Sin
embargo, una disminución del contenido de MDA fue observada a los 16 DDE en ambas
temperaturas (Fig. 8A). Por otro lado, el contenido de Prolina presentó diferencias entre
tratamientos de temperaturas nocturnas en los días 12 y 16 (Fig. 8B). En general, un aumento
del contenido de este aminoácido fue mayor en las plantas expuestas a 30°C desde los 8
hasta los 16 DDE en comparación a las plantas control que mostraron una reducción en la
síntesis de este aminoácido en el mismo periodo de tiempo mencionado anteriormente.
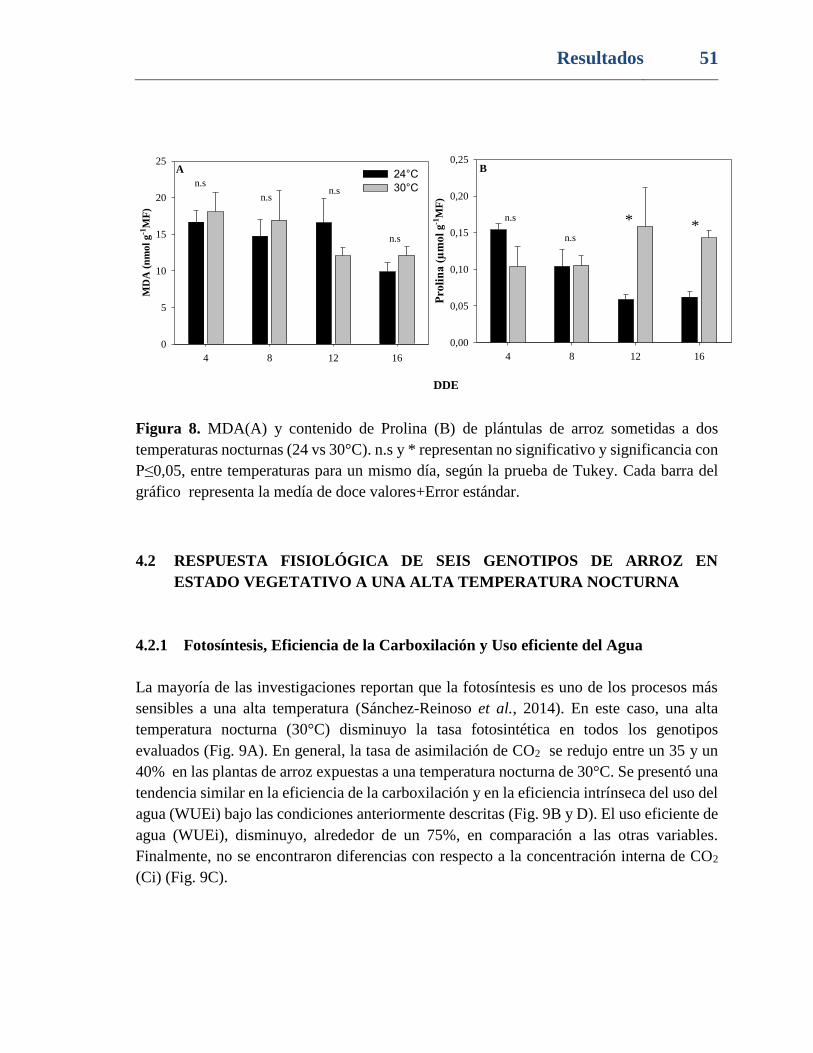
Resultados 51
DDE
4 8 12 16
MD
A (
nm
ol
g-1
MF
)
0
5
10
15
20
25
24°C
30°C
4 8 12 16
Pro
lin
a (
µm
ol
g-1
MF
)
0,00
0,05
0,10
0,15
0,20
0,25A B
* *
n.s
n.sn.s
n.s
n.s
n.s
Figura 8. MDA(A) y contenido de Prolina (B) de plántulas de arroz sometidas a dos
temperaturas nocturnas (24 vs 30°C). n.s y * representan no significativo y significancia con
P≤0,05, entre temperaturas para un mismo día, según la prueba de Tukey. Cada barra del
gráfico representa la medía de doce valores+Error estándar.
4.2 RESPUESTA FISIOLÓGICA DE SEIS GENOTIPOS DE ARROZ EN
ESTADO VEGETATIVO A UNA ALTA TEMPERATURA NOCTURNA
4.2.1 Fotosíntesis, Eficiencia de la Carboxilación y Uso eficiente del Agua
La mayoría de las investigaciones reportan que la fotosíntesis es uno de los procesos más
sensibles a una alta temperatura (Sánchez-Reinoso et al., 2014). En este caso, una alta
temperatura nocturna (30°C) disminuyo la tasa fotosintética en todos los genotipos
evaluados (Fig. 9A). En general, la tasa de asimilación de CO2 se redujo entre un 35 y un
40% en las plantas de arroz expuestas a una temperatura nocturna de 30°C. Se presentó una
tendencia similar en la eficiencia de la carboxilación y en la eficiencia intrínseca del uso del
agua (WUEi) bajo las condiciones anteriormente descritas (Fig. 9B y D). El uso eficiente de
agua (WUEi), disminuyo, alrededor de un 75%, en comparación a las otras variables.
Finalmente, no se encontraron diferencias con respecto a la concentración interna de CO2
(Ci) (Fig. 9C).
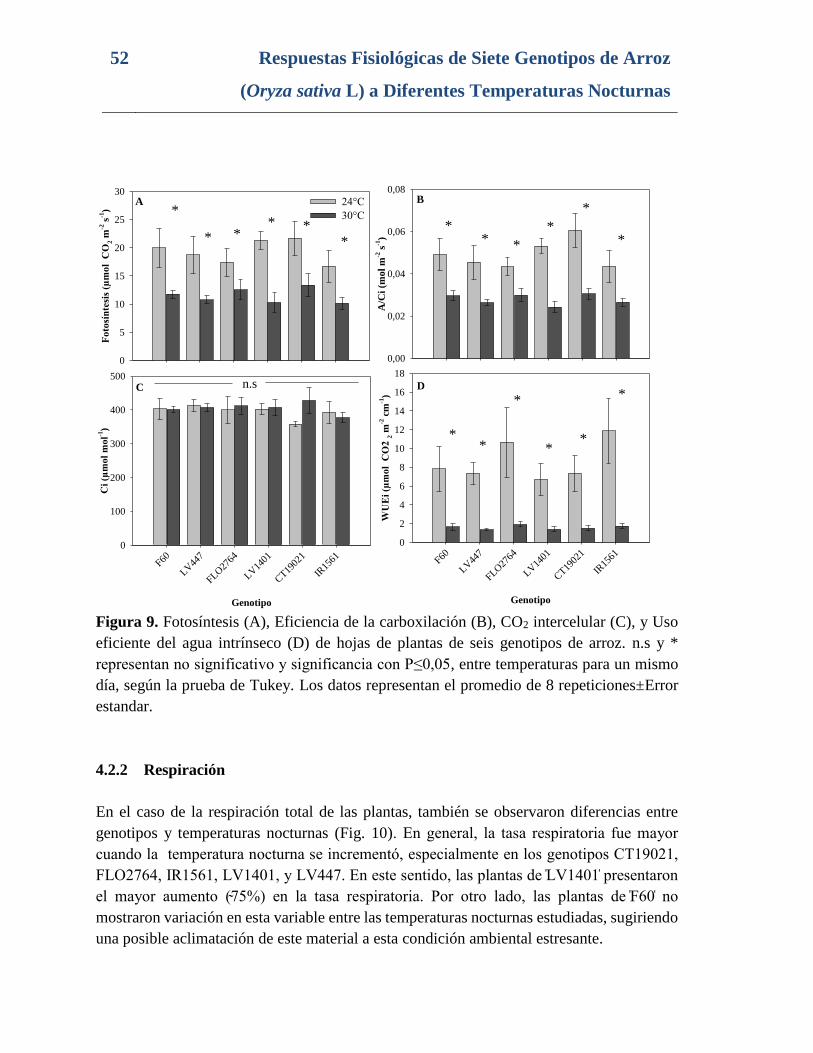
52 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
Fo
tosí
nte
sis
(µm
ol
CO
2 m
-2 s
-1)
0
5
10
15
20
25
3024°C
30°C
Genotipo
F60
LV44
7
FLO27
64
LV14
01
CT19
021
IR15
61
Ci
(µm
ol
mo
l-1)
0
100
200
300
400
500A
/Ci
(mo
l m
-2 s
-1)
0,00
0,02
0,04
0,06
0,08
Genotipo
F60
LV44
7
FLO27
64
LV14
01
CT19
021
IR15
61
WU
Ei
(µm
ol
CO
2 2
m-2
cm
-1)
0
2
4
6
8
10
12
14
16
18
A B
C D
*
** *
***
* *
*
*
*
*
*
**
*
*
n.s
Figura 9. Fotosíntesis (A), Eficiencia de la carboxilación (B), CO2 intercelular (C), y Uso
eficiente del agua intrínseco (D) de hojas de plantas de seis genotipos de arroz. n.s y *
representan no significativo y significancia con P≤0,05, entre temperaturas para un mismo
día, según la prueba de Tukey. Los datos representan el promedio de 8 repeticiones±Error
estandar.
4.2.2 Respiración
En el caso de la respiración total de las plantas, también se observaron diferencias entre
genotipos y temperaturas nocturnas (Fig. 10). En general, la tasa respiratoria fue mayor
cuando la temperatura nocturna se incrementó, especialmente en los genotipos CT19021,
FLO2764, IR1561, LV1401, y LV447. En este sentido, las plantas de ̍LV1401̍ presentaron
el mayor aumento (̴75%) en la tasa respiratoria. Por otro lado, las plantas de ̍F60̍ no
mostraron variación en esta variable entre las temperaturas nocturnas estudiadas, sugiriendo
una posible aclimatación de este material a esta condición ambiental estresante.
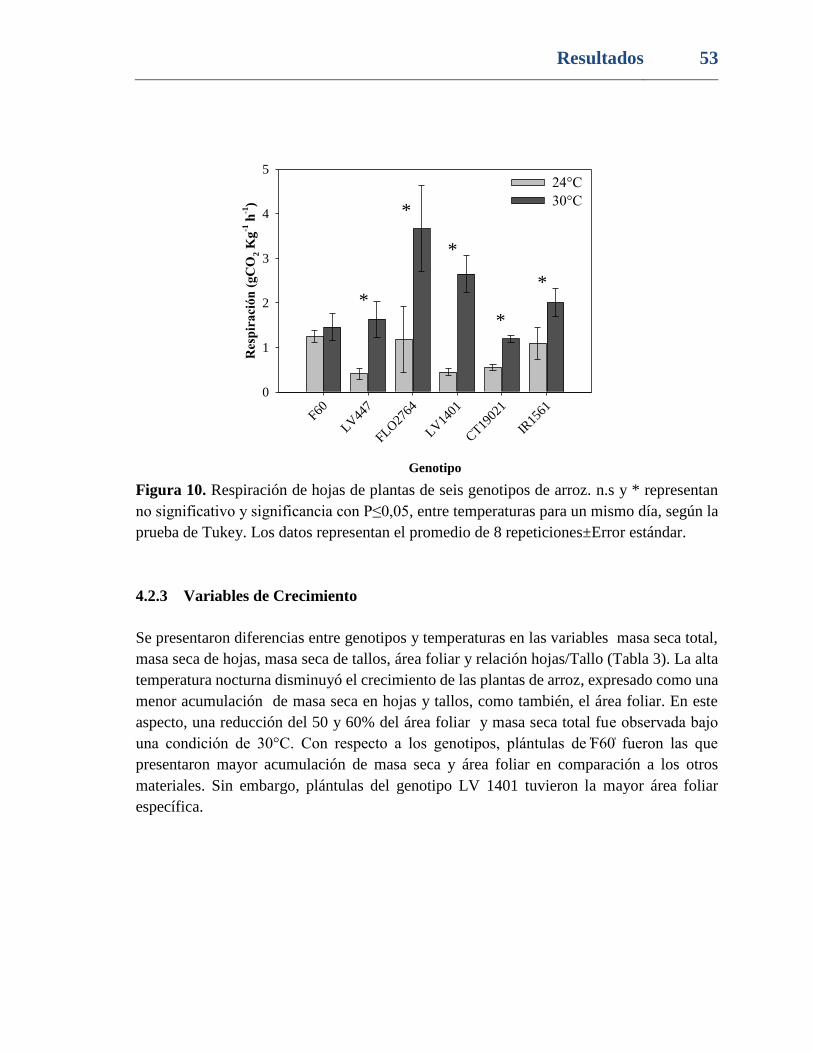
Resultados 53
Genotipo
F60
LV44
7
FLO27
64
LV14
01
CT19
021
IR15
61
Res
pir
aci
ón
(g
CO
2 K
g-1
h-1
)
0
1
2
3
4
524°C
30°C
*
*
*
*
*
Figura 10. Respiración de hojas de plantas de seis genotipos de arroz. n.s y * representan
no significativo y significancia con P≤0,05, entre temperaturas para un mismo día, según la
prueba de Tukey. Los datos representan el promedio de 8 repeticiones±Error estándar.
4.2.3 Variables de Crecimiento
Se presentaron diferencias entre genotipos y temperaturas en las variables masa seca total,
masa seca de hojas, masa seca de tallos, área foliar y relación hojas/Tallo (Tabla 3). La alta
temperatura nocturna disminuyó el crecimiento de las plantas de arroz, expresado como una
menor acumulación de masa seca en hojas y tallos, como también, el área foliar. En este
aspecto, una reducción del 50 y 60% del área foliar y masa seca total fue observada bajo
una condición de 30°C. Con respecto a los genotipos, plántulas de ̍F60̍ fueron las que
presentaron mayor acumulación de masa seca y área foliar en comparación a los otros
materiales. Sin embargo, plántulas del genotipo LV 1401 tuvieron la mayor área foliar
específica.
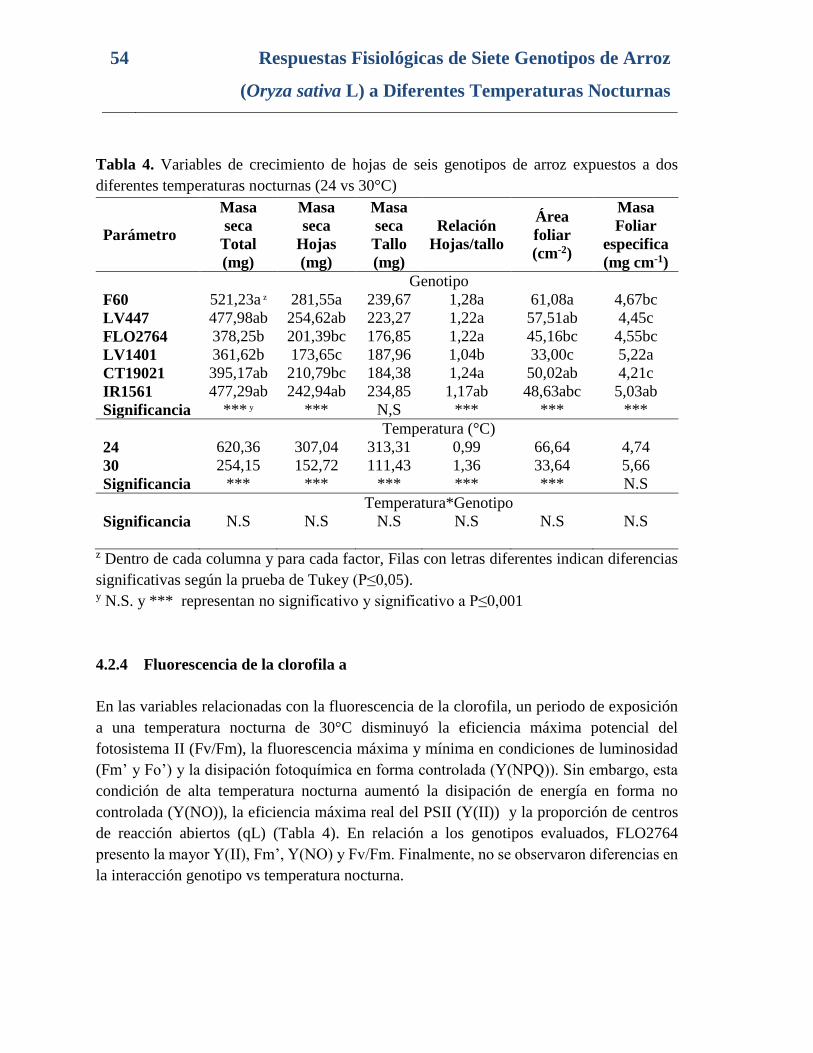
54 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
Tabla 4. Variables de crecimiento de hojas de seis genotipos de arroz expuestos a dos
diferentes temperaturas nocturnas (24 vs 30°C)
Parámetro
Masa
seca
Total
(mg)
Masa
seca
Hojas
(mg)
Masa
seca
Tallo
(mg)
Relación
Hojas/tallo
Área
foliar
(cm-2)
Masa
Foliar
especifica
(mg cm-1)
Genotipo
F60 521,23a z 281,55a 239,67 1,28a 61,08a 4,67bc
LV447 477,98ab 254,62ab 223,27 1,22a 57,51ab 4,45c
FLO2764 378,25b 201,39bc 176,85 1,22a 45,16bc 4,55bc
LV1401 361,62b 173,65c 187,96 1,04b 33,00c 5,22a
CT19021 395,17ab 210,79bc 184,38 1,24a 50,02ab 4,21c
IR1561 477,29ab 242,94ab 234,85 1,17ab 48,63abc 5,03ab
Significancia *** y *** N,S *** *** ***
Temperatura (°C)
24 620,36 307,04 313,31 0,99 66,64 4,74
30 254,15 152,72 111,43 1,36 33,64 5,66
Significancia *** *** *** *** *** N.S
Temperatura*Genotipo
Significancia N.S N.S N.S N.S N.S N.S
z Dentro de cada columna y para cada factor, Filas con letras diferentes indican diferencias
significativas según la prueba de Tukey (P≤0,05). y N.S. y *** representan no significativo y significativo a P≤0,001
4.2.4 Fluorescencia de la clorofila a
En las variables relacionadas con la fluorescencia de la clorofila, un periodo de exposición
a una temperatura nocturna de 30°C disminuyó la eficiencia máxima potencial del
fotosistema II (Fv/Fm), la fluorescencia máxima y mínima en condiciones de luminosidad
(Fm’ y Fo’) y la disipación fotoquímica en forma controlada (Y(NPQ)). Sin embargo, esta
condición de alta temperatura nocturna aumentó la disipación de energía en forma no
controlada (Y(NO)), la eficiencia máxima real del PSII (Y(II)) y la proporción de centros
de reacción abiertos (qL) (Tabla 4). En relación a los genotipos evaluados, FLO2764
presento la mayor Y(II), Fm’, Y(NO) y Fv/Fm. Finalmente, no se observaron diferencias en
la interacción genotipo vs temperatura nocturna.
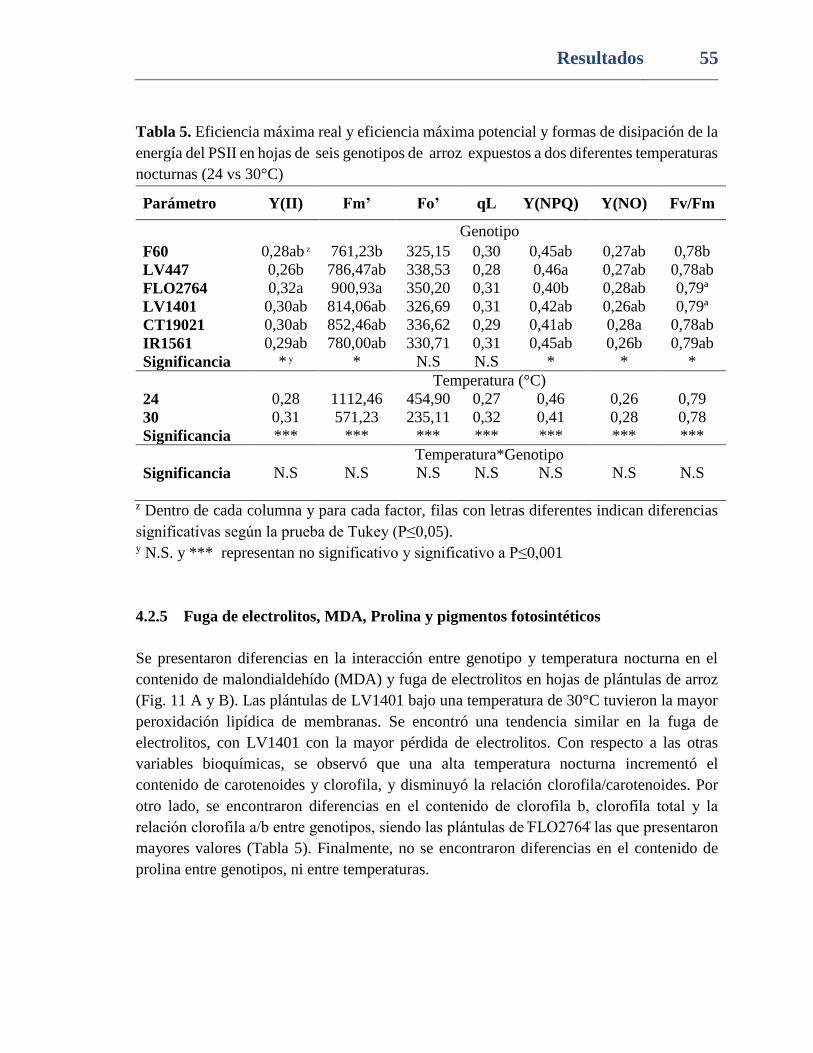
Resultados 55
Tabla 5. Eficiencia máxima real y eficiencia máxima potencial y formas de disipación de la
energía del PSII en hojas de seis genotipos de arroz expuestos a dos diferentes temperaturas
nocturnas (24 vs 30°C)
Parámetro Y(II) Fm’ Fo’ qL Y(NPQ) Y(NO) Fv/Fm
Genotipo
F60 0,28ab z 761,23b 325,15 0,30 0,45ab 0,27ab 0,78b
LV447 0,26b 786,47ab 338,53 0,28 0,46a 0,27ab 0,78ab
FLO2764 0,32a 900,93a 350,20 0,31 0,40b 0,28ab 0,79ª
LV1401 0,30ab 814,06ab 326,69 0,31 0,42ab 0,26ab 0,79ª
CT19021 0,30ab 852,46ab 336,62 0,29 0,41ab 0,28a 0,78ab
IR1561 0,29ab 780,00ab 330,71 0,31 0,45ab 0,26b 0,79ab
Significancia * y * N.S N.S * * *
Temperatura (°C)
24 0,28 1112,46 454,90 0,27 0,46 0,26 0,79
30 0,31 571,23 235,11 0,32 0,41 0,28 0,78
Significancia *** *** *** *** *** *** ***
Temperatura*Genotipo
Significancia N.S N.S N.S N.S N.S N.S N.S
z Dentro de cada columna y para cada factor, filas con letras diferentes indican diferencias
significativas según la prueba de Tukey (P≤0,05). y N.S. y *** representan no significativo y significativo a P≤0,001
4.2.5 Fuga de electrolitos, MDA, Prolina y pigmentos fotosintéticos
Se presentaron diferencias en la interacción entre genotipo y temperatura nocturna en el
contenido de malondialdehído (MDA) y fuga de electrolitos en hojas de plántulas de arroz
(Fig. 11 A y B). Las plántulas de LV1401 bajo una temperatura de 30°C tuvieron la mayor
peroxidación lipídica de membranas. Se encontró una tendencia similar en la fuga de
electrolitos, con LV1401 con la mayor pérdida de electrolitos. Con respecto a las otras
variables bioquímicas, se observó que una alta temperatura nocturna incrementó el
contenido de carotenoides y clorofila, y disminuyó la relación clorofila/carotenoides. Por
otro lado, se encontraron diferencias en el contenido de clorofila b, clorofila total y la
relación clorofila a/b entre genotipos, siendo las plántulas de ̍FLO2764̍ las que presentaron
mayores valores (Tabla 5). Finalmente, no se encontraron diferencias en el contenido de
prolina entre genotipos, ni entre temperaturas.
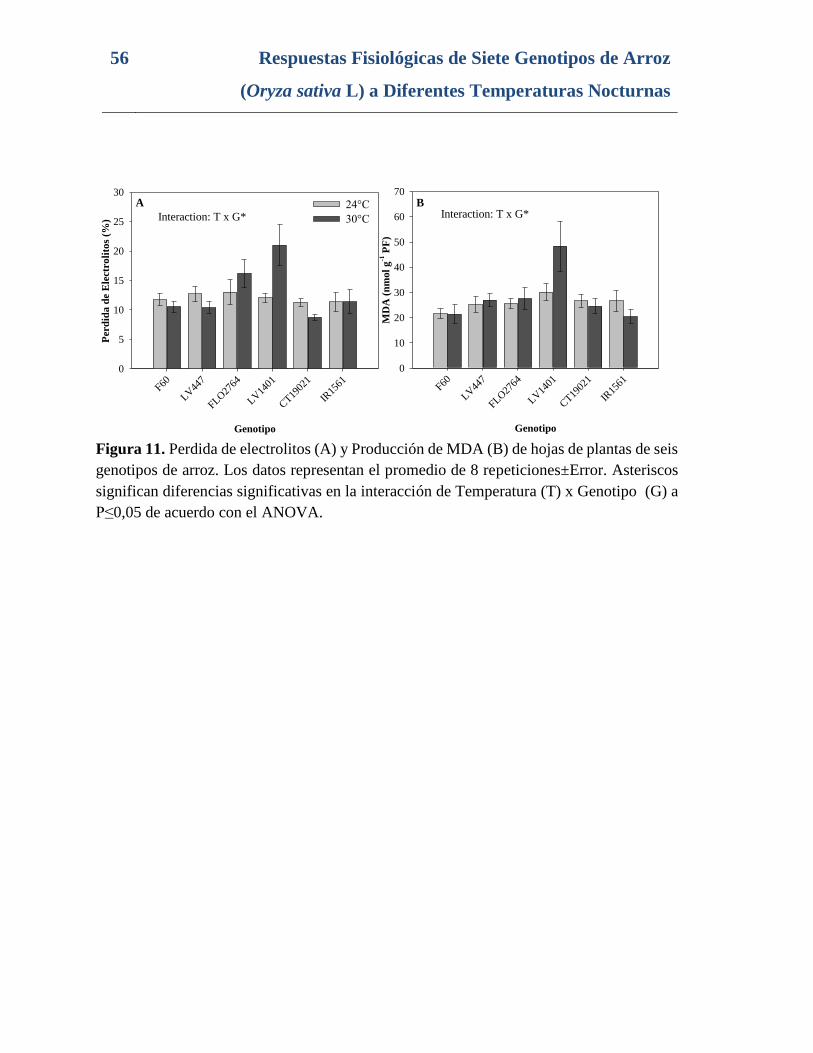
56 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
Genotipo
F60
LV44
7
FLO27
64
LV14
01
CT19
021
IR15
61
Per
did
a d
e E
lect
roli
tos
(%)
0
5
10
15
20
25
30
24°C
30°C
Genotipo
F60
LV44
7
FLO27
64
LV14
01
CT19
021
IR15
61M
DA
(n
mo
l g
-1 P
F)
0
10
20
30
40
50
60
70A B
Interaction: T x G* Interaction: T x G*
Figura 11. Perdida de electrolitos (A) y Producción de MDA (B) de hojas de plantas de seis
genotipos de arroz. Los datos representan el promedio de 8 repeticiones±Error. Asteriscos
significan diferencias significativas en la interacción de Temperatura (T) x Genotipo (G) a
P≤0,05 de acuerdo con el ANOVA.
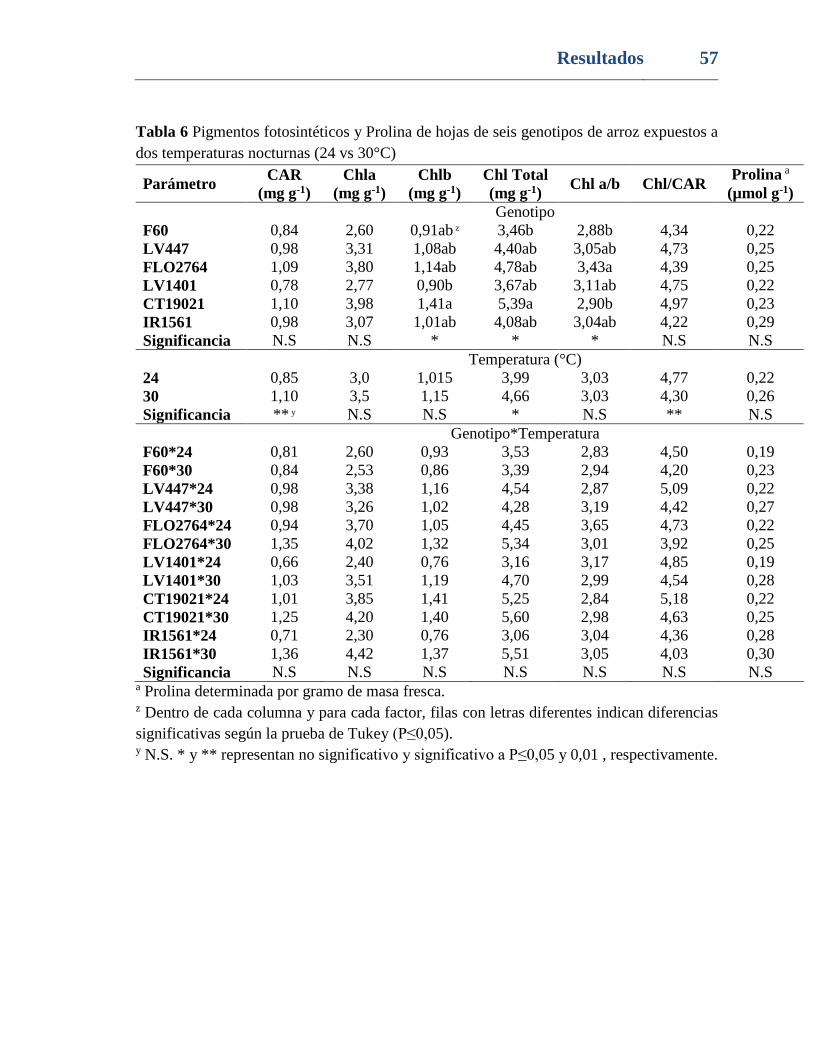
Resultados 57
Tabla 6 Pigmentos fotosintéticos y Prolina de hojas de seis genotipos de arroz expuestos a
dos temperaturas nocturnas (24 vs 30°C)
Parámetro CAR
(mg g-1)
Chla
(mg g-1)
Chlb
(mg g-1)
Chl Total
(mg g-1) Chl a/b Chl/CAR
Prolina a
(µmol g-1)
Genotipo
F60 0,84 2,60 0,91ab z 3,46b 2,88b 4,34 0,22
LV447 0,98 3,31 1,08ab 4,40ab 3,05ab 4,73 0,25
FLO2764 1,09 3,80 1,14ab 4,78ab 3,43a 4,39 0,25
LV1401 0,78 2,77 0,90b 3,67ab 3,11ab 4,75 0,22
CT19021 1,10 3,98 1,41a 5,39a 2,90b 4,97 0,23
IR1561 0,98 3,07 1,01ab 4,08ab 3,04ab 4,22 0,29
Significancia N.S N.S * * * N.S N.S
Temperatura (°C)
24 0,85 3,0 1,015 3,99 3,03 4,77 0,22
30 1,10 3,5 1,15 4,66 3,03 4,30 0,26
Significancia ** y N.S N.S * N.S ** N.S
Genotipo*Temperatura
F60*24 0,81 2,60 0,93 3,53 2,83 4,50 0,19
F60*30 0,84 2,53 0,86 3,39 2,94 4,20 0,23
LV447*24 0,98 3,38 1,16 4,54 2,87 5,09 0,22
LV447*30 0,98 3,26 1,02 4,28 3,19 4,42 0,27
FLO2764*24 0,94 3,70 1,05 4,45 3,65 4,73 0,22
FLO2764*30 1,35 4,02 1,32 5,34 3,01 3,92 0,25
LV1401*24 0,66 2,40 0,76 3,16 3,17 4,85 0,19
LV1401*30 1,03 3,51 1,19 4,70 2,99 4,54 0,28
CT19021*24 1,01 3,85 1,41 5,25 2,84 5,18 0,22
CT19021*30 1,25 4,20 1,40 5,60 2,98 4,63 0,25
IR1561*24 0,71 2,30 0,76 3,06 3,04 4,36 0,28
IR1561*30 1,36 4,42 1,37 5,51 3,05 4,03 0,30
Significancia N.S N.S N.S N.S N.S N.S N.S a Prolina determinada por gramo de masa fresca.
z Dentro de cada columna y para cada factor, filas con letras diferentes indican diferencias
significativas según la prueba de Tukey (P≤0,05). y N.S. * y ** representan no significativo y significativo a P≤0,05 y 0,01 , respectivamente.

58 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
4.3 CARACTERIZACIÓN FISIOLOGICA DE SIETE GENOTIPOS DE ARROZ
SOMETIDOS A UNA ALTA TEMPERATURA NOCTURNA
4.3.1 Componentes de rendimiento (Número de granos vanos, Número de granos
llenos, Porcentaje de fertilidad y masa de granos llenos por panícula)
En las variables N° total de granos, N° de granos llenos, N° de granos vanos y porcentaje de
fertilidad (Fig. 12) se encontraron diferencias significativas (P≤0,05) entre la interacción
Genotipo-Temperatura, mientras no hubo diferencias en la masa promedio del grano. Plantas
de 'F50' produjeron mayor número total de granos en comparación a los otros genotipos
sometidos a una temperatura nocturna de 30°C (Fig. 12A). En relación al número de granos
llenos, los genotipos F50, F60 y LV1401 tuvieron el mayor valor en ambas condiciones de
temperatura (24 y 30°C) (Fig. 12B). La mayoría de genotipos evaluados no presentaron
diferencias sobre el número de granos vanos, excepto plantas de arroz de CT19021 y 'F50'
que presentaron el mayor número de granos vanos a 30 y 24°C, respectivamente. Los
anteriores resultados son corroborados mediante el porcentaje de fertilidad, ya que esta
variable fue menor en plantas de 'F50' y 'CT19021' en las condiciones mencionadas (Fig.
12D).
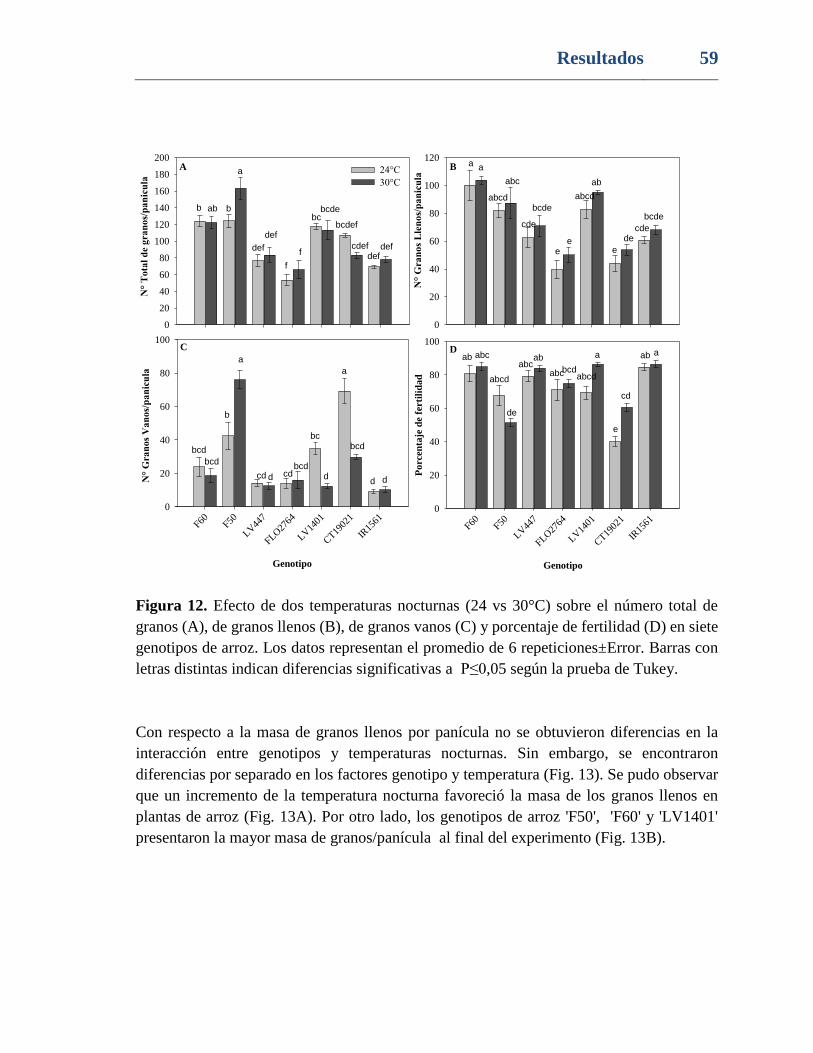
Resultados 59
Genotipo
F60 F50
LV44
7
FLO27
64
LV14
01
CT19
021
IR15
61
N°
Gra
no
s V
an
os/
pa
nic
ula
0
20
40
60
80
100
N°
Gra
no
s L
len
os/
pa
nic
ula
0
20
40
60
80
100
120
N°
Tota
l d
e g
ra
no
s/p
an
icu
la
0
20
40
60
80
100
120
140
160
180
200
24°C
30°C
A B
C
abcd
ab
a a
abcd
abc
bcdebcde
cde cdede
ee
e
a
a
bcd
bcd
b
bc
bcd
bcd
cd cdd ddd
abb
cdef
bcdef
bcdebc
f
def
a
def
b
defdef
f
Genotipo
F60 F50
LV44
7
FLO27
64
LV14
01
CT19
021
IR15
61
Po
rcen
taje
de f
erti
lid
ad
0
20
40
60
80
100D
ab a
abcd
abcabc
aab abc ab
de
bcdabcd
e
cd
Figura 12. Efecto de dos temperaturas nocturnas (24 vs 30°C) sobre el número total de
granos (A), de granos llenos (B), de granos vanos (C) y porcentaje de fertilidad (D) en siete
genotipos de arroz. Los datos representan el promedio de 6 repeticiones±Error. Barras con
letras distintas indican diferencias significativas a P≤0,05 según la prueba de Tukey.
Con respecto a la masa de granos llenos por panícula no se obtuvieron diferencias en la
interacción entre genotipos y temperaturas nocturnas. Sin embargo, se encontraron
diferencias por separado en los factores genotipo y temperatura (Fig. 13). Se pudo observar
que un incremento de la temperatura nocturna favoreció la masa de los granos llenos en
plantas de arroz (Fig. 13A). Por otro lado, los genotipos de arroz 'F50', 'F60' y 'LV1401'
presentaron la mayor masa de granos/panícula al final del experimento (Fig. 13B).
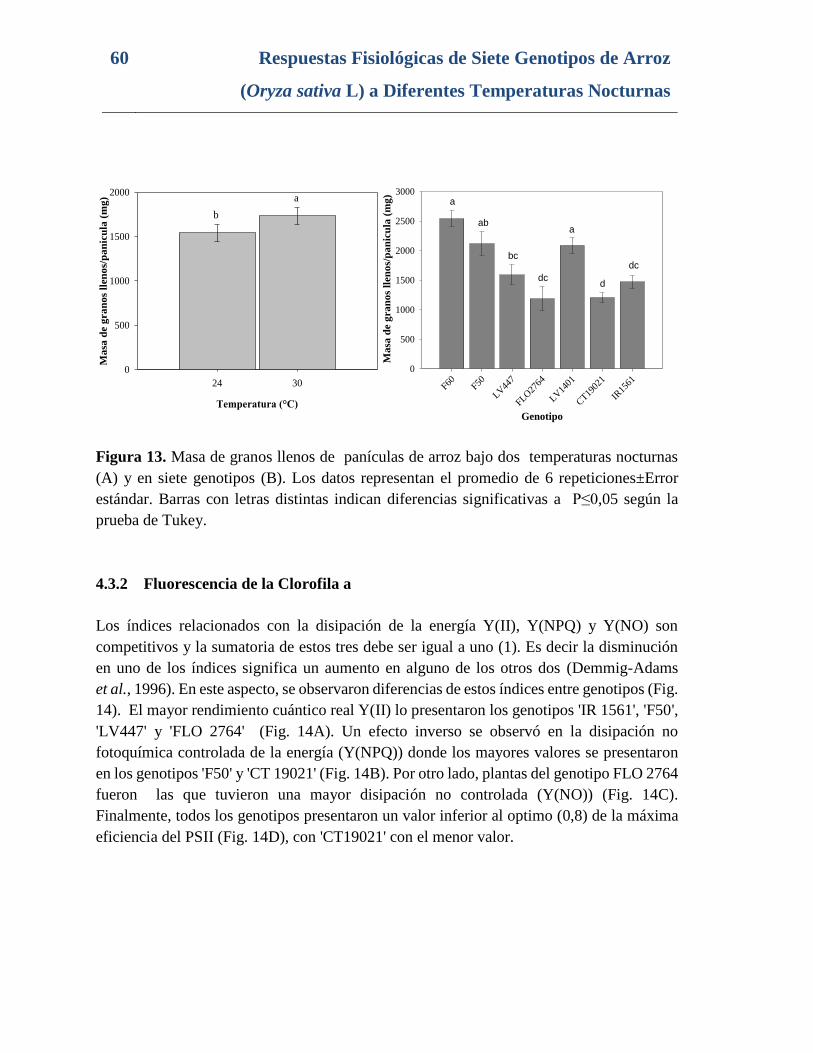
60 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
Genotipo
F60 F50
LV44
7
FLO27
64
LV14
01
CT19
021
IR15
61M
asa
de g
ra
no
s ll
en
os/
pa
nic
ula
(m
g)
0
500
1000
1500
2000
2500
3000
Temperatura (°C)
24 30
Ma
sa d
e g
ra
no
s ll
en
os/
pa
nic
ula
(m
g)
0
500
1000
1500
2000a
ab
bc
dc
a
d
dc
a
b
Figura 13. Masa de granos llenos de panículas de arroz bajo dos temperaturas nocturnas
(A) y en siete genotipos (B). Los datos representan el promedio de 6 repeticiones±Error
estándar. Barras con letras distintas indican diferencias significativas a P≤0,05 según la
prueba de Tukey.
4.3.2 Fluorescencia de la Clorofila a
Los índices relacionados con la disipación de la energía Y(II), Y(NPQ) y Y(NO) son
competitivos y la sumatoria de estos tres debe ser igual a uno (1). Es decir la disminución
en uno de los índices significa un aumento en alguno de los otros dos (Demmig-Adams
et al., 1996). En este aspecto, se observaron diferencias de estos índices entre genotipos (Fig.
14). El mayor rendimiento cuántico real Y(II) lo presentaron los genotipos 'IR 1561', 'F50',
'LV447' y 'FLO 2764' (Fig. 14A). Un efecto inverso se observó en la disipación no
fotoquímica controlada de la energía (Y(NPQ)) donde los mayores valores se presentaron
en los genotipos 'F50' y 'CT 19021' (Fig. 14B). Por otro lado, plantas del genotipo FLO 2764
fueron las que tuvieron una mayor disipación no controlada (Y(NO)) (Fig. 14C).
Finalmente, todos los genotipos presentaron un valor inferior al optimo (0,8) de la máxima
eficiencia del PSII (Fig. 14D), con 'CT19021' con el menor valor.
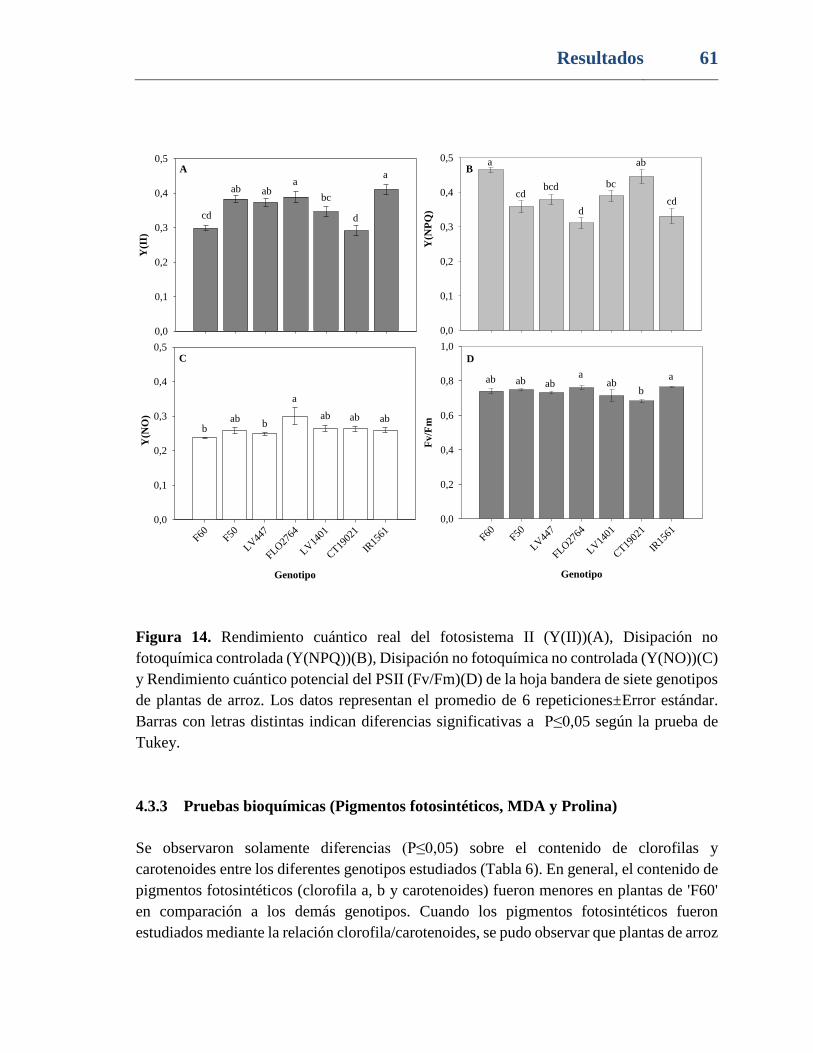
Resultados 61
Genotipo
F60 F50
LV44
7
FLO27
64
LV14
01
CT19
021
IR15
61
Y(N
O)
0,0
0,1
0,2
0,3
0,4
0,5
Genotipo
F60 F50
LV44
7
FLO27
64
LV14
01
CT19
021
IR15
61
Fv
/Fm
0,0
0,2
0,4
0,6
0,8
1,0
Y(N
PQ
)
0,0
0,1
0,2
0,3
0,4
0,5
Y(I
I)
0,0
0,1
0,2
0,3
0,4
0,5A B
C D
a
ab aba
bc
cd d
a
bb
abab ab ab
aa
bab ab abab
a ab
bcbcd
cdcd
d
Figura 14. Rendimiento cuántico real del fotosistema II (Y(II))(A), Disipación no
fotoquímica controlada (Y(NPQ))(B), Disipación no fotoquímica no controlada (Y(NO))(C)
y Rendimiento cuántico potencial del PSII (Fv/Fm)(D) de la hoja bandera de siete genotipos
de plantas de arroz. Los datos representan el promedio de 6 repeticiones±Error estándar.
Barras con letras distintas indican diferencias significativas a P≤0,05 según la prueba de
Tukey.
4.3.3 Pruebas bioquímicas (Pigmentos fotosintéticos, MDA y Prolina)
Se observaron solamente diferencias (P≤0,05) sobre el contenido de clorofilas y
carotenoides entre los diferentes genotipos estudiados (Tabla 6). En general, el contenido de
pigmentos fotosintéticos (clorofila a, b y carotenoides) fueron menores en plantas de 'F60'
en comparación a los demás genotipos. Cuando los pigmentos fotosintéticos fueron
estudiados mediante la relación clorofila/carotenoides, se pudo observar que plantas de arroz
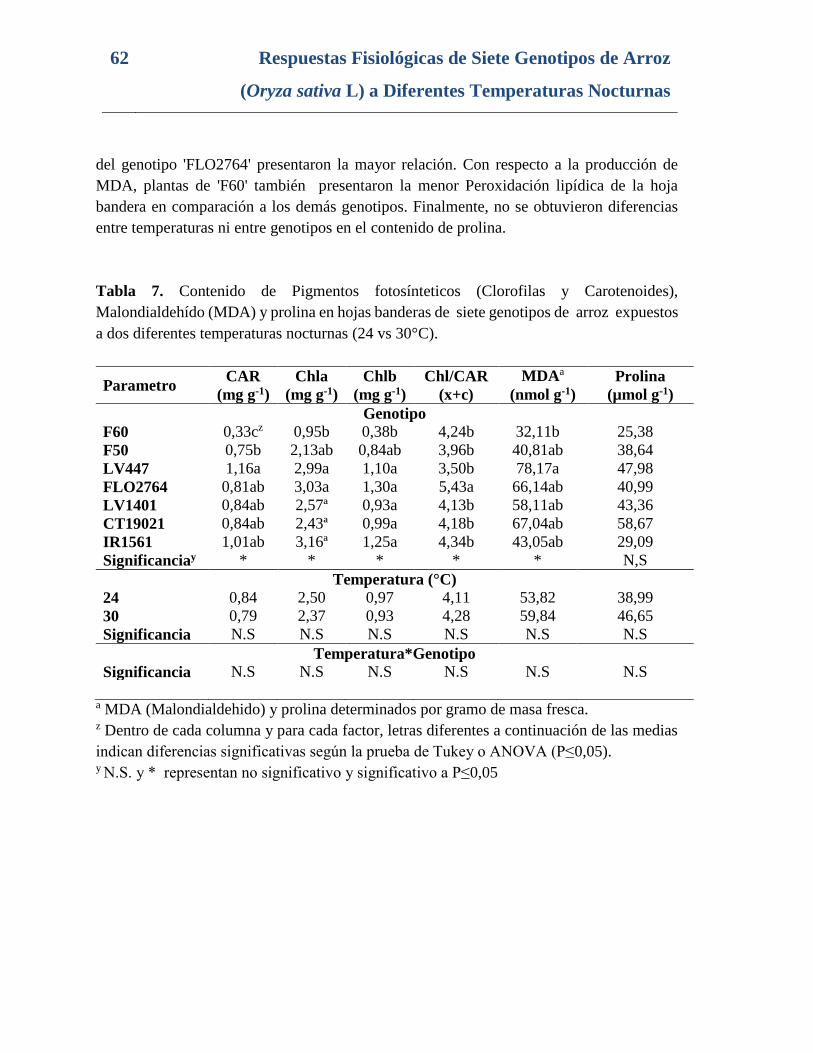
62 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
del genotipo 'FLO2764' presentaron la mayor relación. Con respecto a la producción de
MDA, plantas de 'F60' también presentaron la menor Peroxidación lipídica de la hoja
bandera en comparación a los demás genotipos. Finalmente, no se obtuvieron diferencias
entre temperaturas ni entre genotipos en el contenido de prolina.
Tabla 7. Contenido de Pigmentos fotosínteticos (Clorofilas y Carotenoides),
Malondialdehído (MDA) y prolina en hojas banderas de siete genotipos de arroz expuestos
a dos diferentes temperaturas nocturnas (24 vs 30°C).
Parametro CAR
(mg g-1)
Chla
(mg g-1)
Chlb
(mg g-1)
Chl/CAR
(x+c)
MDAa
(nmol g-1)
Prolina
(µmol g-1)
Genotipo
F60 0,33cz 0,95b 0,38b 4,24b 32,11b 25,38
F50 0,75b 2,13ab 0,84ab 3,96b 40,81ab 38,64
LV447 1,16a 2,99a 1,10a 3,50b 78,17a 47,98
FLO2764 0,81ab 3,03a 1,30a 5,43a 66,14ab 40,99
LV1401 0,84ab 2,57ª 0,93a 4,13b 58,11ab 43,36
CT19021 0,84ab 2,43ª 0,99a 4,18b 67,04ab 58,67
IR1561 1,01ab 3,16ª 1,25a 4,34b 43,05ab 29,09
Significanciay * * * * * N,S
Temperatura (°C)
24 0,84 2,50 0,97 4,11 53,82 38,99
30 0,79 2,37 0,93 4,28 59,84 46,65
Significancia N.S N.S N.S N.S N.S N.S
Temperatura*Genotipo
Significancia N.S N.S N.S N.S N.S N.S
a MDA (Malondialdehido) y prolina determinados por gramo de masa fresca. z Dentro de cada columna y para cada factor, letras diferentes a continuación de las medias
indican diferencias significativas según la prueba de Tukey o ANOVA (P≤0,05). y N.S. y * representan no significativo y significativo a P≤0,05

Discusión 63
5. DISCUSIÓN
5.1 PRIMER EXPERIMENTO: PERIODOS DE ESTRÉS
La fotosíntesis (Pn) de las plantas es uno de los procesos fisiológicos más sensibles a las
altas temperaturas, tanto diurnas como nocturnas (Ashraf y Harris, 2013). En el presente
estudio, la tasa fotosintética disminuyó alrededor de un 15% por una alta temperatura
nocturna (30°C) a lo largo del experimento (Fig. 2). Mohammed et al. (2013) obtuvieron
resultados similares, donde temperaturas nocturnas de 30°C causaron una reducción
del ̴10% sobre la Pn de arroz. Esta reducción de la Pn, causada por ATN, puede deberse a
alteraciones tanto en la fase lumínica como en la fase de carboxilación del proceso
fotosintético. Con respecto a la fase lumínica, se ha reportado que periodos largos de altas
temperaturas nocturnas disminuyen el contenido de pigmentos fotosintéticos y la eficiencia
del PSII en arroz. En este ensayo, La alta temperatura nocturna provocó un daño leve en el
PSII lo que pudo producir una fotoinhibición, teniendo en cuenta que la relación Fv/Fm
disminuyo en los primeros 8 días (Fig. 6) y es la medida más usada para medir fotoinhibición
(Goh et al., 2012).
Asimismo, una menor asimilación de CO2 puede deberse al hecho que ATN causa
limitaciones asociadas al ciclo de Calvin (Song et al., 2013; Mohammed y Tarpley, 2014).
En este aspecto, se observó una menor eficiencia de carboxilación y una acumulación de
CO2 intercelular (Fig. 2D y 3). Durante todo el ensayo se presentó una menor tasa de
carboxilación (Fig. 3) y hacia el día 12 está pudo ser la principal causa de la disminución de
la tasa fotosintética, teniendo en cuenta que la relación Fv/Fm no disminuyo luego del día
12 en plantas sometidas a 30°C.
La menor eficiencia de carboxilación significa una menor capacidad de la Rubisco para fijar
CO2, provocando una mayor acumulación intercelular de este gas (Kiran et al., 2013). Esta
menor eficiencia de carboxilación pudo ser causada por la disminución de pigmentos
fotosintéticos y una menor actividad de la Rubisco activasa (Yamori et al., 2012; Yin et al.,
2010), ya que se ha reportado que la Rubisco activasa es más susceptible a la alta
temperatura que la Rubisco.
Un mecanismo que soporta la idea de que el principal factor que limita la fotosíntesis durante
una alta temperatura es un daño en el PSII y en la menor actividad de la Rubisco es una
reorganización en la membrana tilacoidal que hace que mayor número de unidades del
LHCII se asocien al PSI y no al PSII (Marutani et al., 2014; Pastenes y Horton, 1996;

64 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
Allakhverdiev et al., 2008), aumentando el flujo cíclico de electrones y disminuyendo la
tensión en el PSII. Esto ocurre mediante la fosforilación de las antenas LHCII lo cual
provoca que estos complejos se asocien al PSI y no al PSII (Shikanai, 2014; Goldschmidt-
Clermont y Bassi, 2015; Minagawa, 2011).
El hecho de que no haya una relación positiva entre el contenido de CO2 intercelular y la
conductancia estomática (Fig. 2) soporta la hipótesis de que la disminución de la tasa
fotosintética no se debe a un cierre estomático sino a una limitación no estomática: menor
eficiencia de la Carboxilación de la Rubisco provocada por un menor estado de activación
y daño en el PSII. lo cual también es reportado por (Kubien y Sage, 2008) en plantas de
tabaco sometidas a altas temperaturas.
Trabajos desarrollados por Mohammed y Tarpley, (2009b) mostraron que periodos largos
de ATN causaron una disminución de pigmentos fotosintéticos y relación Fv/Fm
repercutiendo negativamente en la Pn en plantas de arroz. Similares observaciones fueron
también obtenidas en el presente estudio donde una reducción del contenido de clorofilas y
carotenoides y una menor relación Fv/Fm fueron encontradas en plantas de arroz expuestas
a 30°C (Fig. 6). Lo anterior junto con la menor eficiencia de carboxilación pudo provocar la
menor capacidad de la Rubisco para fijar CO2, y en consecuencia se produjo una mayor
acumulación intercelular de CO2 (Kiran et al., 2013).
Con respecto a la Ro, está mostró un incremento alrededor del 50% en plántulas de arroz
sometidas a 30°C en comparación al control (24°C) principalmente a los 4 y 16 DDE (Fig.
4). Esta pérdida de carbono en forma de respiración también ha sido reportada por
Mohammed y Tarpley (2010b) y Zhang et al. (2013) donde muestran que temperaturas
nocturnas entre 30 y 32°C causaron un aumento aproximadamente del 50% de la Ro en
plantas de arroz. Este incremento de la respiración se puede deber a factores como un
aumento de la actividad de la Oxidasa alternativa con el fin de mantener el flujo de electrones
en la mitocondria y evitar estrés oxidativo (Plaxton y Podestá, 2006), sintetizar más ATP
para producir NAD+ (el cual es afectado negativamente bajo una alta temperatura) (Wahid
et al., 2007) y aumentar la síntesis y regeneración de proteínas (Lambers et al., 2008). Los
procesos descritos anteriormente tienen el propósito de reducir los efectos negativos
producidos por ATN (Plaxton y Podestá, 2006).
Por otro lado, el contenido de MDA no mostró diferencias entre las dos temperaturas
nocturnas (Fig. 8A). Sin embargo, estos resultados contrastan con otros estudios donde la
Peroxidación lipídica fue mayor cuando se incrementó la temperatura nocturna (Kumar
et al., 2011; Xue et al., 2012). Lo anterior permite inferir que una temperatura nocturna de

Discusión 65
30°C no causa un mayor daño de membranas a nivel celular en este cultivar de arroz
colombiano a pesar que la fotosíntesis y respiración sufran modificaciones. Esto es
reportado como una característica de tolerancia a altas temperaturas (Narayanan et al., 2016;
Bahuguna et al., 2015; Zhou et al., 2012), sin embargo es necesario comparar con otros
genotipos para corroborar dicha hipótesis.
Asimismo, otra respuesta bioquímica bajo condiciones de estrés térmico es la producción
de prolina (Sánchez-Reinoso et al., 2014). En nuestro estudio, la producción de prolina
mostró diferencias entre temperaturas nocturnas, observándose un aumento de este osmolito
en plántulas de arroz a 30°C a través del experimento (Fig. 8B). La mayor concentración de
prolina producida en plantas bajo un estrés térmico es reportado por otros autores como
Kumar et al. (2011): una temperatura superior a 35/30°C día/noche provoca un aumento en
plántulas de arroz y teniendo en cuenta que la alta temperatura redujo la eficiencia de PSII
(Fig. 6) y que (Yan et al., 2012; Vani et al., 2001b) reportan que esto se debe a un daño en
la sección donadora y aceptora de electrones. En estas condiciones, la prolina puede actuar
como un donador de electrones del PSII y como antioxidante evitando daños en el PSII (De
Ronde et al., 2004). Por lo anterior se puede inferir que la acumulación de prolina en
plántulas de arroz de 'F60' es una estrategia de aclimatación a la alta temperatura nocturna.
5.2 SEGUNDO Y TERCER EXPERIMENTO: RESPUESTA DE SIETE
GENOTIPOS DE ARROZ A UNA ALTA TEMPERATURA NOCTURNA
El uso de genotipos tolerantes a altas temperaturas es una de las herramientas más efectivas
para mantener la estabilidad y altas producciones en el cultivo del arroz con el propósito de
mitigar los efectos negativos del cambio climático (Horie et al., 1996) Por tal motivo, es
importante identificar métodos o técnicas para determinar la tolerancia de genotipos (Zhang
et al., 2013). En consecuencia, varios autores han reportado que técnicas como los
parámetros de la fluorescencia de la clorofila (Sayed 2003; Šebela et al. 2015), la tasa de
respiratoria (Cheng et al., 2009; Mohammed et al., 2013), la acumulación de biomasa y
rendimiento (Zhang et al., 2013), pruebas bioquímicas (Sánchez-Reinoso et al. 2014) y la
fotosíntesis (Rodríguez et al., 2005) son importantes para conocer el grado de tolerancia o
susceptibilidad a condiciones de estrés abiótico. En este aspecto, en el presente estudio, se
observaron diferencias en algunas variables de crecimiento debido a la alta temperatura
nocturna (Tabla 3). La biomasa total de plantas de arroz sometidas a 30°C disminuyó
alrededor de un 60% con respecto a las plantas de 24°C de temperatura nocturna. Zhang
et al. (2013) también encontraron que una alta temperatura nocturna (ATN) reduce la
acumulación de biomasa debido a una menor tasa del crecimiento del cultivo. En general, la
tasa de producción de materia seca del arroz está condicionada por el balance entre

66 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
fotosíntesis y respiración nocturna (Cheng et al., 2009; Mohammed et al., 2013). Una ATN
reduce la tasa fotosintética, incrementa la respiración y puede afectar negativamente la
estabilidad de las membranas, causando una disminución en la biomasa (Peng et al., 2004).
En este sentido, los resultados obtenidos en el presente experimento pueden soportar las
anteriores afirmaciones donde una ATN disminuyo la fotosíntesis e incremento la
respiración (Fig. 9 y 10), disminuyendo la acumulación de biomasa de la planta. Por otro
lado, las diferencias en crecimiento entre genotipos puede deberse principalmente a
diferencias varietales, ya que el ANOVA no mostro un interacción entre genotipos y
temperatura nocturna (Zhang et al., 2013; Garcés-Varon y Restrepo-Díaz, 2015).
Con respecto a los parámetros de Fluorescencia de la clorofila a, una ATN (30°C)
disminuyó la eficiencia máxima potencial del PSII (Fv/Fm) cuando las hojas estaban en
estado vegetativo (Tabla 4). La relación Fv/Fm indica la máxima proporción de la energía
que la planta es capaz de usar en la fotosíntesis y generalmente disminuye bajo una
condición de estrés (Baker, 2008). Esta reducción es causada generalmente por un daño en
la proteína del PSII: D1 o en el sistema de reparación de esta proteína (Gururani et al.,
2015). Asimismo, un mayor qL fue observado en hojas de arroz sometidas a una ATN en
fase vegetativa, lo que significa una mayor capacidad de la planta para usar los productos
finales de la fotosíntesis (Lu y Zhang, 1999; Marutani et al., 2014). El aumento de este
índice al aumentar la temperatura nocturna puede ser una forma de aclimatación de la planta,
sugiriendo que las hojas desarrollan mecanismos para consumir los productos de la
fotoquímica (por ejemplo fotorespiración) con el fin de mantener el flujo de electrones y
evitar la fotoinhibición (Ort y Baker, 2002; D’Ambrosio et al., 2006). Por otro lado, la ATN
no provocó cambios en los parámetros de la fluorescencia de la clorofila a en la fase de
maduración. En este sentido estudios desarrollados por Mohammed y Tarpley (2014) y
Glaubitz et al. (2014) tampoco encontraron diferencias sobre estos índices cuando se
expusieron diferentes genotipos de arroz a altas temperaturas nocturnas (≥ 28°C).
Aunque no se encontraron diferencias entre temperaturas nocturnas, si se observaron entre
las hojas bandera de los genotipos en estado de maduración. Es importante señalar que el
índice Y(NO), el cual se ha definido como una forma no controlada y perjudicial de disipar
la energía (Brestic y Zivcak, 2013), mostró un incremento en el genotipo 'FLO
2764'(Y(NO)= 0,3) . Por lo anterior, Y(NO)estaría indicando que el genotipo 'FLO 2764'
pudo haber presentado ciertos daños en el PSII en las condiciones de la zona arrocera donde
se desarrolló la presente investigación en comparación a 'LV447' y 'F60' (Y(NO)= 0,25 y
0,24), respectivamente. La relación Fv/Fm de todos los genotipos fue menor al valor óptimo
reportado en la literatura (~0,8 (Murchie y Lawson, 2013; Baker, 2008)) en el experimento
de la fase de maduración. Sin embargo, un menor valor entre 0,7 y 0,8 no necesariamente

Discusión 67
significa un daño en el PSII debido a una condición estresante, sino puede ser propio del
genotipo o producto de la senescencia de la hoja (Lim et al., 2007; Panda y Sarkar, 2013).
La principal función de las clorofilas y carotenoides es recolectar y trasmitir la energía
lumínica para luego ser transformada en energía química (Taiz y Zeiger, 2006). Igualmente,
la concentración de estos pigmentos generalmente disminuye bajo una condición estresante
(Mohammed y Tarpley, 2009a) o durante la senescencia de la hoja (Hörtensteiner, 2006) .
El contenido de pigmentos fotosintéticos como carotenoides, clorofila total y clorofila b
aumentó en plantas sometidas a una ATN en el segundo experimento (Fase vegetativa)
(Tabla 3). El aumento del contenido de clorofilas es contrario a lo reportado en plantas de
arroz sometidas a altas temperaturas nocturnas Mohammed y Tarpley, (2009b). Sin
embargo, es importante analizar que la relación clorofila:carotenoides disminuyó a una
temperatura de 30°C. Según Camejo et al. (2005) y Wahid et al. (2007), una reducción de
esta relación es una respuesta de tolerancia por parte de la planta al estrés térmico.
Asimismo, la acumulación de carotenoides puede ser otro mecanismo de la planta para
reducir el estrés producido por la alta temperatura, ya que los carotenoides pueden disipar
la energía en forma de calor a través del ciclo de las xantofilas o actuar como antioxidantes
(Jahns y Holzwarth, 2012). Sin embargo, estas tendencias no se mantuvieron en la fase de
maduración entre temperatura y solo se presentaron diferencias entre los diferentes
genotipos evaluados. Dong et al. (2014) encontraron que la concentración de clorofila a y b
disminuyo cuando sometieron dos materiales de arroz a una alta temperatura nocturna
(28°C) y concluyeron que las diferencias pueden depender del momento en el que se
desarrolla la evaluación. En nuestro caso, las diferencias entre genotipos y la no
significancia entre temperaturas puede deberse al hecho que la evaluación de esta variable
fue desarrollada en estado avanzados de la etapa de maduración donde se presenta una
mayor senescencia de las hojas (Restrepo-Diaz y Garces-Varon, 2013).
En cuanto a los marcadores bioquímicos, resultados contrastantes fueron observados en la
producción de MDA entre los experimentos dos y tres. La producción de MDA fue mayor
en el genotipo LV 1401 cuando fue expuesto a una ATN en la fase vegetativa. Por otro lado,
las temperaturas no provocaron un aumento de la Peroxidación lipídica ni la producción de
prolina (osmolito involucrado en la homeostasis celular) en la fase de maduración. Esta
diferencia entre los resultados observados entre fases de desarrollo es debido también a la
senescencia, ya que la peroxidacion lipídica esta influenciada por la edad de la hoja (Zapata
et al., 2005). Los anteriores resultados permiten sugerir que la peróxidación lipídica y el
aumento de la permeabilidad de las membranas puede ser usadas para seleccionar genotipos
susceptibles a altas temperaturas en estado vegetativo del cultivo del arroz.

68 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
Una temperatura nocturna de 30°C disminuyo el porcentaje de fertilidad en plantas de 'F50'
alrededor de un 30% aproximadamente en comparación a los genotipos ̍F60̍, 'LV1401' y
'IR1561'(Fig. 12). Una posible explicación a las diferentes respuestas entre los genotipos
estudiados en este trabajo puede ser que las variedades presenten diferentes mecanismos de
aclimatación bajo esta condición de estrés (Jagadish et al., 2010). En este sentido,
Mohammed y Tarpley (2009a) argumentan que un alto número de espiguillas estériles puede
ser causado por un incremento de la respiración de mantenimiento en la etapa de
maduración, mientras que Shi et al. (2013) lo asocian con una reducción en la translocación
de nitrógeno y carbohidratos no estructurales desde las hojas y tallos hacía las panículas,
como también, una menor expresión de proteínas de choque térmico (del tipo FKBP),
proteínas de señalización de calcio (calmodulinas y quinasas) y mecanismos de reparación
o modificación de ADN y proteínas (especialmente en la etapa de maduración).
Teniendo en cuenta que la producción de todos los genotipos está asociada con el número
de granos llenos (Fig. 12 y 13) y que la masa promedio de grano no fue afectada por la alta
temperatura, se puede inferir que la disminución en la fertilidad del genotipo 'F50' y el
aumento en 'CT19021' está asociada con el proceso de polinización y viabilidad del polen y
no con el proceso de llenado del grano. Esto lo confirma Jagadish et al. (2015), quien afirma
que la alta temperatura disminuye la viabilidad del polen, el crecimiento del tubo polínico
y perjudica el desarrollo de los ovarios y que los genotipos que no presentan estas
características son tolerantes a altas temperaturas.
Con respecto a otro componente de rendimiento como es la masa de los granos llenos, las
variedades comerciales ('F50' y 'F60') y el genotipo LV1401 fueron los que mayor masa de
granos presentaron (Fig. 13.). Teniendo en cuenta lo anterior y que no se presentaron
diferencias en la masa del grano (datos no mostrados) los mayores rendimientos en estos
genotipos se debe a un mayor número de granos/panícula producidos, como también lo
enuncia Mohammed y Tarpley (2011).

Modelo Teórico 69
6. MODELO TEÓRICO DEL EFECTO DE LA
ALTA TEMPERATURA NOCTURNA
Con base a los resultados obtenidos y como una forma de conclusión general del presente
estudio de investigación, se generó un modelo o esquema conceptual del efecto de una alta
temperatura nocturna sobre la fisiología de una planta de arroz. Este modelo empieza
indicando que una alta temperatura nocturna (30°C) disminuyo el contenido de pigmentos
(Fig. 7 y Tabla 5), la capacidad del PSII y probablemente afecto negativamente la actividad
de la Rubisco activasa. Estos eventos provocan una menor asimilación de CO2 en la hoja
(Fig. 2 y 9). En cuanto a la respiración, la alta temperatura puede disminuir la vida media de
moléculas como proteínas y lípidos, lo que aumento la tasa metabólica y la necesidad de
regenerar dichas moléculas, lo cual requiere energía y dicha energía es suministrada por la
respiración. Esto provocó una mayor tasa respiratoria en la mayoría de los genotipos (Fig.
10). En este sentido, las alteraciones en la fotosíntesis y la respiración pueden causar a una
menor acumulación de carbohidratos que son usados en el crecimiento, lo cual también se
observó en el segundo experimento donde se evaluaron 6 genotipos y todos presentaron una
disminución en su crecimiento expresado como menor acumulación de masa seca y área
foliar (Tabla 3). Asimismo, la temperatura disminuyó la asimilación de CO2, sin embargo;
es poco probable que se deba a daños en el PSII, ya que no hubo una correlación entre Fv/Fm
Y(II) en general.
Por otro lado, la eficiencia de la carboxilación presentó un comportamiento similar a la
asimilación de CO2: constante en el tiempo y menor en plantas de F60 sometidas a 30°C
(Fig. 2A, 2B y 3), y menor en los 6 genotipos, en estado vegetativo, sometidos a una ATN
(Fig. 9A y 9B). Teniendo en cuenta lo anterior y que tampoco se encontró una relación
directa entre conductancia estomática y asimilación de CO2 se podría inferir que la menor
asimilación de CO2 se debe a una limitación en la capacidad de carboxilación de la RUbisCO
o en su activación y no a una disminución en la tasa de transporte de electrones o a un cierre
estomático.
Teniendo en cuenta las variables de fluorescencia y la asimilación de CO2 en los dos
primeros experimentos, se puede inferir que la distribución de la energía lumínica es la
siguiente: la energía destinada a la fotorespiración, reacción de Mehler o al flujo cíclico de
electrones aumenta (es necesario realizar otras investigaciones para determinar cuál de estos
procesos es el de mayor demanda de electrones en estas condiciones), mientras la destinada
a la fotosíntesis disminuye. Esto se comprueba con un aumento en la eficiencia operativa

70 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
del PSII (Y(II))(Tabla 4) acompañado por una disminución en la asimilación de CO2 (Fig.
9), lo que quiere decir que aumenta la energía lumínica usada en el transporte de electrones,
pero que la energía química producto de esta transporte de electrones no se está usando en
la asimilación de CO2 sino probablemente en la fotorespiración (Foyer et al., 2012; Murchie
y Lawson, 2013). Lo mismo sucede con la disipación en forma de calor (Y (NPQ)) que
disminuye con un aumento de la temperatura. Mientras aumenta la forma de disipación no
controlada, lo que podría indicar junto con lo nombrado anteriormente que el principal
mecanismo de disipación de la energía es la fotorespiración y que cuando no se puede disipar
de esta forma, dicha energía se disipa de forma no controlada, lo que podría ocasionar un
daño del aparato fotosintético.
En el caso del estado de maduración, no se presentaron los efectos provocados por las altas
temperaturas sobre las hojas en las plantas en estado vegetativo (Fig. 14 y Tabla 5). Sin
embargo, las variables reproductivas mostraron que el genotipo F50 es más susceptible a la
alta temperatura nocturna que los demás genotipos y los genotipos LV1401 y CT19021
presentan mayor tolerancia a la alta temperatura nocturna (Fig. 12). Finalmente, la Figura
15 resume el modelo teórico expuesto anteriormente:
Figura 15. Modelo conceptual de los efectos y las características de tolerancia y
susceptibilidad de arroz sometido a altas temperaturas nocturnas (30°C). Los símbolos
dentro de cada recuadro indican aumento (↑), disminución (↓) o sin cambios (=) del proceso
dentro del recuadro. El origen de la flecha roja indica las posibles causas de un proceso y el
final de la flecha roja indica el proceso afectado.

Conclusiones y Recomendaciones 71
7. CONCLUSIONES Y RECOMENDACIONES
7.1 Conclusiones
Periodos superiores a cuatro días de exposición a una ATN pueden causar
alteraciones en el balance de carbono en estado vegetativo y una disminución en los
componentes de rendimiento de la panícula.
El uso de variables fisiológicas como fotosíntesis, respiración, crecimiento de
órganos y contenidos de pigmentos fotosintéticos pueden ser consideradas como
marcadores de tolerancia o susceptibilidad en programas de mejoramiento genético
de arroz a una condición de ATN, especialmente, en etapas de desarrollo iniciales.
Los parámetros asociados a la fluorescencia de la clorofila, en general, no mostraron
utilidad para caracterizar tolerancia o susceptibilidad de genotipos a una ATN en una
etapa de maduración.
La determinación de MDA y fuga de electrolitos permite cuantificar susceptibilidad
de genotipos en etapas iniciales de la planta.
Los componentes de rendimiento de la panícula (fertilidad de la panícula y número
de granos vanos) mostraron ser un componente importante para la caracterización de
genotipos sometidos a un estrés por ATN en fase de maduración.
7.2 Recomendaciones
Evaluar el efecto de periodos de tiempo, de alta temperatura nocturna igual o
menores a dos días sobre la fisiología de plantas de arroz en estado vegetativo.
En una condición de alta temperatura nocturna, evaluar variables relacionadas con
la viabilidad del polen, tiempo de floración, características de las anteras y del
estigma de las líneas F60, F50 y CT19021, con el fin de conocer los mecanismos que
las hacen tolerantes y susceptibles a esta condición.
Determinar el efecto de la alta temperatura nocturna sobre la fotorespiración, además
de la fotosíntesis y variables de la fluorescencia, para tener un mayor entendimiento
sobre como la planta disipa la energía lumínica.

72 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
Evaluar el efecto de la alta temperatura nocturna sobre variables relacionadas con
fotosíntesis y con estrés oxidativo en otros estados de desarrollo de la planta como,
floración y embuchamiento.

Bibliografía 73
Bibliografía
Allakhverdiev, Suleyman I., Kreslavski, Vladimir D., Klimov, Vyacheslav V., Los, Dmitry
A., Carpentier, Robert y Mohanty, Prasanna¨Heat stress: An overview of molecular
responses in photosynthesis¨, [en línea] Photosynthesis Research, ISSN-01668595, DOI-
10.1007/s11120-008-9331-0, 98(1–3): 541–550, 2008.
Apel, Klaus y Hirt, Heribert¨REACTIVE OXYGEN SPECIES: Metabolism, Oxidative
Stress, and Signal Transduction¨, [en línea] Annual Review of Plant Biology, ISSN-1543-
5008, DOI-10.1146/annurev.arplant.55.031903.141701, 55(1): 373–399, 2004.
Araus, José Luis., Slafer, Gustavo a., Royo, Conxita y Serret, M Dolores¨Breeding for Yield
Potential and Stress Adaptation in Cereals¨, [en línea] Critical Reviews in Plant Sciences,
ISSN-0735-2689, DOI-10.1080/07352680802467736, 27(6): 377–412, 18 de noviembre de
2008.
Ashraf, M y Foolad, MR¨Roles of glycine betaine and proline in improving plant abiotic
stress resistance¨, [en línea] Environmental and Experimental Botany, ISSN-00988472,
DOI-10.1016/j.envexpbot.2005.12.006, 59(2): 206–216, 2007.
Ashraf, M y Harris, PJC¨Photosynthesis under stressful environments: An overview¨,
Photosynthetica, 51(2): 163–190, 2013.
Bahuguna, Rajeev N., Jha, Jyoti., Pal, Madan., Shah, Divya., Lawas, Lovely MF., Khetarpal,
Sangeetha y Jagadish, Krishna S V¨Physiological and biochemical characterization of
NERICA-L-44: a novel source of heat tolerance at the vegetative and reproductive stages in
rice¨, [en línea] Physiologia Plantarum, ISSN-1399-3054, DOI-10.1111/ppl.12299, 154(4):
543–559, 1 de agosto de 2015.
Baker, NR¨Chlorophyll fluorescence: A probe of photosynthesis in vivo¨, Annu. Rev. Plant
Biol., 59: 89–113, 2008.
Barnabás, Beáta., Jäger, Katalin y Fehér, Attila¨The effect of drought and heat stress on
reproductive processes in cereals.¨, [en línea] Plant, cell & environment, ISSN-0140-7791,
DOI-10.1111/j.1365-3040.2007.01727.x, 31(1): 11–38, enero de 2008.
Bates, LS., Waldren, RP y Teare, ID¨Rapid determination of free proline for water-stress
studies¨, Plant and Soil, 39(1): 205–207, 1973.
Bita, Craita y Gerats, Tom Plant tolerance to high temperature in a changing environment:
scientific fundamentals and production of heat stress tolerant crops, 2013,
Bokszczanin, Kamila L y Fragkostefanakis, Sotirios¨Perspectives on deciphering
mechanisms underlying plant heat stress response and thermotolerance.¨, [en línea]

74 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
Frontiers in plant science, ISSN-1664-462X, DOI-10.3389/fpls.2013.00315, 4(August):
315, 2013.
Bolhàr-Nordenkampf, HR y Öquist, G ̈ Chlorophyll fluorescence as a tool in photosynthesis
research¨, Photosynthesis and production in a changing environment, pp. 193–206, Ed.
Springer, 1993.
Brestic, Marian y Zivcak, Marek ¨PSII Fluorescence Techniques for Measurement of
Drought and High Temperature Stress Signal in Crop Plants: Protocols and Applications¨,
[en línea] edit. Gyana Ranjan Rout y Anath Bandhu Das, Molecular Stress Physiology of
Plants, ISBN-978-81-322-0806-8, pp. 87–131, Ed. Springer India, 2013.
Camejo, Daymi., Rodríguez, Pedro., Angeles Morales, Ma., Miguel Dell’Amico, José.,
Torrecillas, Arturo y Alarcón, Juan José¨High temperature effects on photosynthetic activity
of two tomato cultivars with different heat susceptibility¨, [en línea] Journal of Plant
Physiology, DOI-http://dx.doi.org/10.1016/j.jplph.2004.07.014, 162(3): 281–289, 2005.
Castilla, LA., Sierra, J., Pineda, D., Echeverry, J., Garces, G., Perafan, R., Ospina, JJ y
Flores, E Analisis de la producción arrocera e el Tolima, edit. FEDEARROZ, 2010,
Chen, S., Zhang, X., Zhao, X., Wang, D., Xu, C y Ji, C¨Response of rice nitrogen physiology
to high nighttime temperature during vegetative stage¨, The Scientific World Journal, 2013,
2013.
Cheng, Weiguo., Sakai, Hidemitsu., Yagi, Kazuyuki y Hasegawa, Toshihiro¨Interactions of
elevated [CO2] and night temperature on rice growth and yield¨, [en línea] Agricultural and
Forest Meteorology, ISSN-01681923, DOI-10.1016/j.agrformet.2008.07.006, 149(1): 51–
58, enero de 2009.
Coast, Onoriode., Ellis, Richard H., Murdoch, Alistair J., Quiñones, Cherryl y Jagadish,
Krishna S V¨High night temperature induces contrasting responses for spikelet fertility,
spikelet tissue temperature, flowering characteristics and grain quality in rice¨, Functional
Plant Biology, 42(2): 149–161, 2015.
Cosgrove, Jeff y Borowitzka, Michael A ¨Chlorophyll fluorescence terminology: an
introduction¨, Chlorophyll a fluorescence in aquatic sciences: methods and applications,
ISBN-9048192676, pp. 1–17, Ed. Springer, 2010.
Cossani, C Mariano y Reynolds, Matthew P¨Physiological Traits for Improving Heat
Tolerance in Wheat¨, [en línea] Plant Physiology, DOI-10.1104/pp.112.207753, 160(4):
1710–1718, 2012.
Cui, Langjun., Li, Jianlong., Fan, Yamin., Xu, Sheng y Zhang, Zhen¨High temperature
effects on photosynthesis, PSII functionality and antioxidant activity of two Festuca
arundinacea cultivars with different heat susceptibility¨, Botanical Studies, ISSN-

Bibliografía 75
1817406X, 47(1): 61–69, 2006.
D’Ambrosio, Nicola., Arena, Carmen y De Santo, Amalia Virzo¨Temperature response of
photosynthesis, excitation energy dissipation and alternative electron sinks to carbon
assimilation in Beta vulgaris L.¨, [en línea] Environmental and Experimental Botany, ISSN-
00988472, DOI-10.1016/j.envexpbot.2004.11.006, 55(3): 248–257, 2006.
De Ronde, JA., Cress, WA., Kr??ger, GHJ., Strasser, RJ y Van Staden, J¨Photosynthetic
response of transgenic soybean plants, containing an Arabidopsis P5CR gene, during heat
and drought stress¨, [en línea] Journal of Plant Physiology, ISSN-01761617, DOI-
10.1016/j.jplph.2004.01.014, 161(11): 1211–1224, 2004.
Demmig-Adams, B y Adams Iii, WW¨The role of xanthophyll cycle carotenoids in the
protection of photosynthesis¨, Trends in Plant Science, 1(1): 21–26, 1996.
Demmig-Adams, Barbara., Adams III, William W., Barker, David H., Logan, Barry A.,
Bowling, David R y Verhoeven, Amy S¨Using chlorophyll fluorescence to assess the
fraction of absorbed light allocated to thermal dissipation of excess excitation¨, [en línea]
Physiologia Plantarum, ISSN-00319317, DOI-10.1034/j.1399-3054.1996.980206.x, 98(2):
253–264, 9 de octubre de 1996.
Dong, Wenjun., Chen, Jin., Wang, Lili., Tian, Yunlu., Zhang, Bin., Lai, Yongcai., Meng,
Ying., Qian, Chunrong y Guo, Jia¨Impacts of nighttime post-anthesis warming on rice
productivity and grain quality in East China¨, [en línea] The Crop Journal, ISSN-22145141,
DOI-10.1016/j.cj.2013.11.002, 2(1): 63–69, febrero de 2014.
Fageria, NK¨Yield Physiology of Rice¨, [en línea] Journal of Plant Nutrition, ISSN-0190-
4167, DOI-10.1080/15226510701374831, 30(6): 843–879, 9 de mayo de 2007.
Farooq, Muhammad., Bramley, Helen., Palta, Jairo A y Siddique, Kadambot HM¨Heat
Stress in Wheat during Reproductive and Grain-Filling Phases¨, [en línea] Critical Reviews
in Plant Sciences, DOI-10.1080/07352689.2011.615687, 30(6): 491–507, 2011.
FEDEARROZ Manejo integrado del cultivo de Arroz, Ed. Produmedio, pp. 566, Colombia,
2013.
FEDEARROZ Producción y Área cultivada de Arroz paddy en 2013, Area, Produccion y
Rendimiento de Arroz Paddy, [en línea] 2015, Disponible en:
http://www.fedearroz.com.co/new/apr_public.php [Consulta: 18 de junio de 2015].
Foyer, Christine H., Neukermans, Jenny., Queval, Guillaume., Noctor, Graham y
Harbinson, Jeremy¨Photosynthetic control of electron transport and the regulation of gene
expression¨, [en línea] Journal of Experimental Botany, ISSN-00220957, DOI-
10.1093/jxb/ers013, 63(4): 1637–1661, 2012.
Garcés-Varon, Gabriel y Restrepo-Díaz, Hermann¨Growth and yield of rice cultivars sowed

76 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
on different dates under tropical conditions¨, [en línea] Ciencia e Investigación Agraria;
Vol 42, No 2 (2015), DOI-10.4067/rcia.v42i2.1511, , 2015.
Gill, SS y Tuteja, N¨Reactive oxygen species and antioxidant machinery in abiotic stress
tolerance in crop plants¨, Plant Physiology and Biochemistry, 48(12): 909–930, 2010.
Glaubitz, Ulrike., Li, Xia., Köhl, Karin I., Van Dongen, Joost T., Hincha, Dirk K., Zuther,
Ellen., Van Dongen A, Joost T., Hincha, Dirk K y Zuther, Ellen¨Differential physiological
responses of different rice (Oryza sativa) cultivars to elevated night temperature during
vegetative growth¨, [en línea] Functional Plant Biology, ISSN-14454408, DOI-
10.1071/FP13132, 41: 437–448, 2014.
Goh, Chang Hyo., Ko, Suk Min., Koh, Seokchan., Kim, Young Joo y Bae, Hyeun
Jong¨Photosynthesis and Environments: Photoinhibition and Repair Mechanisms in Plants¨,
[en línea] Journal of Plant Biology, ISSN-12269239, DOI-10.1007/s12374-011-9195-2,
55(2): 93–101, 2012.
Goldschmidt-Clermont, Michel y Bassi, Roberto¨Sharing light between two photosystems:
mechanism of state transitions.¨, [en línea] Current opinion in plant biology, ISSN-1879-
0356, DOI-10.1016/j.pbi.2015.04.009, 25: 71–78, 2015.
Guidi, Lucia y Degl’Innocenti, Elena ¨Chlorophyll a Fluorescence in Abiotic Stress¨, [en
línea] edit. B Venkateswarlu, Arun K Shanker, Chitra Shanker, y M Maheswari, Crop
Stress and its Management: Perspectives and Strategies SE - 10, ISBN-978-94-007-2219-
4, pp. 359–398, Ed. Springer Netherlands, 2012.
Gururani, Mayank Anand., Venkatesh, Jelli y Tran, Lam Son Phan¨Regulation of
Photosynthesis during Abiotic Stress-Induced Photoinhibition.¨, [en línea] Molecular plant,
ISSN-1752-9867, DOI-10.1016/j.molp.2015.05.005, 8(9): 1304–20, 7 de septiembre de
2015.
Hare, PD y Cress, WA¨Metabolic implications of stress-induced proline accumulation in
plants¨, [en línea] Plant Growth Regulation, DOI-10.1023/a:1005703923347, 21(2): 79–
102, 1997.
Havaux, Michel¨Carotenoids as membrane stabilizers in chloroplasts¨, [en línea] Trends in
Plant Science, ISSN-13601385, DOI-10.1016/S1360-1385(98)01200-X, 3(4): 147–151,
abril de 1998.
Havaux, Michel y Tardy, Florence¨Loss of chlorophyll with limited reduction of
photosynthesis as an adaptive response of Syrian barley landraces to high-light and heat
stress¨, [en línea] Australian Journal of Plant Physiology, ISSN-0310-7841, DOI-
10.1071/PP99046, 26: 569, 1999.
Heldt, HW¨The use of energy from sunlight by photosynthesis is the basis of life on earth¨,

Bibliografía 77
Plant Biochemistry, 3: 56–58, 2005.
Hodges, D Mark., DeLong, John M., Forney, Charles F y Prange, Robert K¨Improving the
thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant
tissues containing anthocyanin and other interfering compounds¨, [en línea] Planta, DOI-
10.1007/s004250050524, 207(4): 604–611, 1999.
Horie, Takeshi., Matsui, Tsutomu., Nakagawa, Hiroshi y Omasa, Kenji ¨Effects of Elevated
CO2 and Global Climate Change on Rice Yield in Japan¨, [en línea] Climate Change and
Plants in East Asia SE - 4, ISBN-978-4-431-66901-2, pp. 39–56, Ed. Springer Japan, 1996.
Hörtensteiner, S¨Chlorophyll degradation during senescence.¨, [en línea] Annual review of
plant biology, ISSN-1543-5008, DOI-10.1146/annurev.arplant.57.032905.105212, 57: 55–
77, 2 de enero de 2006.
Jagadish, SVK., Murty, MVR y Quick, WP¨Rice responses to rising temperatures –
challenges, perspectives and future directions¨, [en línea] Plant, Cell & Environment, ISSN-
1365-3040, DOI-10.1111/pce.12430, 38(9): 1686–1698, 1 de septiembre de 2015.
Jagadish, SVK., Muthurajan, R., Oane, R., Wheeler, TR., Heuer, S., Bennett, J y Craufurd,
PQ¨Physiological and proteomic approaches to address heat tolerance during anthesis in rice
(Oryza sativa L.)¨, [en línea] Journal of Experimental Botany, DOI-10.1093/jxb/erp289,
61(1): 143–156, 2010.
Jagadish, SVK., Septiningsih, EM., Kohli, A., Thomson, MJ., Ye, C., Redoña, E., Kumar,
A., Gregorio, GB., Wassmann, R., Ismail, AM y Singh, RK¨Genetic Advances in Adapting
Rice to a Rapidly Changing Climate¨, [en línea] Journal of Agronomy and Crop Science,
DOI-10.1111/j.1439-037X.2012.00525.x, 198(5): 360–373, 2012.
Jagadish, Krishna S V., Craufurd, Peter., Shi, Wanju y Oane, Rowena¨A phenotypic marker
for quantifying heat stress impact during microsporogenesis in rice (Oryza sativa L.)¨, [en
línea] Functional Plant Biology, ISSN-14454408, DOI-10.1071/FP13086, 41(Hedhly
2011): 48–55, 2014.
Jahns, Peter y Holzwarth, Alfred R The role of the xanthophyll cycle and of lutein in
photoprotection of photosystem II, [en línea] 2012,
Jha, Uday Chand., Bohra, Abhishek y Singh, Narendra Pratap¨Heat stress in crop plants: its
nature, impacts and integrated breeding strategies to improve heat tolerance¨, [en línea]
Plant Breeding, ISSN-1439-0523, DOI-10.1111/pbr.12217, 133(6): 679–701, 1 de
diciembre de 2014.
Kalaji, Hazem M., Schansker, Gert., Ladle, Richard J., Goltsev, Vasilij., Bosa, Karolina.,
Allakhverdiev, Suleyman I., Brestic, Marian., Bussotti, Filippo., Calatayud, Angeles.,
Dąbrowski, Piotr., Elsheery, Nabil I., Ferroni, Lorenzo., Guidi, Lucia., Hogewoning, Sander
W., Jajoo, Anjana., Misra, Amarendra N., Nebauer, Sergio G., Pancaldi, Simonetta., Penella,

78 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
Consuelo., Poli, DorothyBelle., Pollastrini, Martina., Romanowska-Duda, Zdzislawa B.,
Rutkowska, Beata., Serôdio, João., Suresh, Kancherla., Szulc, Wiesław., Tambussi,
Eduardo., Yanniccari, Marcos y Zivcak, Marek¨Frequently asked questions about in vivo
chlorophyll fluorescence: practical issues.¨, [en línea] Photosynthesis research, ISSN-1573-
5079, DOI-10.1007/s11120-014-0024-6, 122(2): 121–58, diciembre de 2014.
Kiran, T V., Rao, Y V., Subrahmanyam, D., Rani, NS., Bhadana, VP., Rao, PR y Voleti,
SR¨Variation in leaf photosynthetic characteristics in wild rice species¨, [en línea]
Photosynthetica, ISSN-0300-3604, DOI-10.1007/s11099-013-0032-3, 51(3): 350–358,
2013.
Krishnan, P., Ramakrishnan, B., Reddy, KR y Reddy, VR High-Temperature Effects on Rice
Growth, Yield, and Grain Quality, 2011,
Kubien, David S y Sage, Rowan F¨The temperature response of photosynthesis in tobacco
with reduced amounts of Rubisco¨, [en línea] Plant, Cell and Environment, ISSN-
01407791, DOI-10.1111/j.1365-3040.2008.01778.x, 31(4): 407–418, 2008.
Kumar, B., Pandey, DM., Goswami, CL y Jain, S¨Effect of Growth Regulators on
Photosynthesis, Transpiration and Related Parameters in Water Stressed Cotton¨, [en línea]
Biologia Plantarum, ISSN-0006-3134, DOI-10.1023/A:1012408624665, 44(3): 475–478,
2001.
Kumar, Sanjeev., Gupta, Deepti y Nayyar, Harsh¨Comparative response of maize and rice
genotypes to heat stress: status of oxidative stress and antioxidants¨, [en línea] Acta
Physiologiae Plantarum, ISSN-0137-5881, DOI-10.1007/s11738-011-0806-9, 34(1): 75–
86, 17 de julio de 2011.
Lambers, Hans., Chapin III, F Stuart y Pons, ThijsL ¨Respiration¨, [en línea] Plant
Physiological Ecology, ISBN-978-0-387-78340-6, pp. 101–150, Ed. Springer New York,
2008.
Laza, Ma Rebecca C., Sakai, Hidemitsu., Cheng, Weiguo., Tokida, Takeshi., Peng,
Shaobing y Hasegawa, Toshihiro¨Differential response of rice plants to high night
temperatures imposed at varying developmental phases¨, [en línea] Agricultural and Forest
Meteorology, ISSN-01681923, DOI-10.1016/j.agrformet.2015.04.029, 209–210: 69–77,
septiembre de 2015.
Lee, KH y Akita, S¨Factors causing the variation in the temperature coefficient of dark
respiration in rice (Oryza sativa L.)¨, Plant Production Science, 3(1): 38–42, 2000.
Lim, Pyung Ok., Kim, Hyo Jung y Nam, Hong Gil¨Leaf senescence.¨, [en línea] Annual
review of plant biology, ISSN-1543-5008, DOI-
10.1146/annurev.arplant.57.032905.105316, 58: 115–36, enero de 2007.

Bibliografía 79
Lobell, David B y Gourdji, Sharon M¨The Influence of Climate Change on Global Crop
Productivity 1¨, [en línea] Plant Physiology, ISSN-0032-0889, DOI-
10.1104/pp.112.208298, 160(December): 1686–1697, 2012.
Lu, C y Zhang, J¨Effects of water stress on photosystem II photochemistry and its
thermostability in wheat plants¨, [en línea] Journal of Experimental Botany, ISSN-0022-
0957, DOI-10.1093/jxb/50.336.1199, 50(336): 1199–1206, 1999.
Lutts, S., KINET, JM y BOUHARMONT, J¨NaCl-induced Senescence in Leaves of Rice
(Oryza sativa L.) Cultivars Differing in Salinity Resistance¨, [en línea] Annals of Botany,
DOI-10.1006/anbo.1996.0134, 78(3): 389–398, 1 de septiembre de 1996.
Mani, Srikrishnan., Van De Cotte, Brigitte., Van Montagu, Marc y Verbruggen,
Nathalie¨Altered levels of proline dehydrogenase cause hypersensitivity to proline and its
analogs in Arabidopsis.¨, [en línea] Plant physiology, ISSN-0032-0889, DOI-
10.1104/pp.010572, 128(1): 73–83, 2002.
Mantri, Nitin., Patade, Vikas., Penna, Suprasanna., Ford, Rebecca y Pang, Edwin ¨Abiotic
Stress Responses in Plants: Present and Future¨, [en línea] edit. Parvaiz Ahmad y M N V
Prasad, Abiotic Stress Responses in Plants SE - 1, ISBN-978-1-4614-0633-4, pp. 1–19, Ed.
Springer New York, 2012.
Marutani, Yoko., Yamauchi, Yasuo., Miyoshi, Akihito., Inoue1, Kanako., Ikeda, Ken Ichi.,
Mizutani, Masaharu y Sugimoto, Yukihiro¨Regulation of Photochemical Energy Transfer
Accompanied by Structural Changes in Thylakoid Membranes of Heat-Stressed Wheat¨, [en
línea] International Journal of Molecular Sciences, ISSN-14220067, DOI-
10.3390/ijms151223042, 15(12): 23042–23058, 2014.
Mathur, Sonal., Agrawal, Divya y Jajoo, Anjana¨Photosynthesis: Response to high
temperature stress¨, [en línea] Journal of Photochemistry and Photobiology B: Biology,
ISSN-18732682, DOI-10.1016/j.jphotobiol.2014.01.010, 137: 116–126, 2014.
Minagawa, Jun¨State transitions-the molecular remodeling of photosynthetic
supercomplexes that controls energy flow in the chloroplast¨, [en línea] Biochimica et
Biophysica Acta - Bioenergetics, ISSN-00052728, DOI-10.1016/j.bbabio.2010.11.005,
1807(8): 897–905, 2011.
Mittler, Ron., Finka, Andrija y Goloubinoff, Pierre¨How do plants feel the heat?¨, [en línea]
Trends in Biochemical Sciences, DOI-http://dx.doi.org/10.1016/j.tibs.2011.11.007, 37(3):
118–125, 2012.
Mohammed, Abdul Razack y Tarpley, Lee¨Effects of high night temperature and spikelet
position on yield-related parameters of rice (Oryza sativa L.) plants¨, [en línea] European
Journal of Agronomy, ISSN-11610301, DOI-10.1016/j.eja.2009.11.006, 33(2): 117–123,
agosto de 2010.

80 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
Mohammed, Abdul Razack y Tarpley, Lee¨Differential response of two important Southern
US rice (Oryza sativa L.) cultivars to high night temperature¨, Australian Journal of Crop
Science, 8(2): 191–199, 2014.
Mohammed, AR y Tarpley, L¨High nighttime temperatures affect rice productivity through
altered pollen germination and spikelet fertility¨, [en línea] Agricultural and Forest
Meteorology, DOI-http://dx.doi.org/10.1016/j.agrformet.2008.12.003, 149(6–7): 999–
1008, 2009a.
Mohammed, AR y Tarpley, L¨Impact of high nighttime temperature on respiration,
membrane stability, antioxidant capacity, and yield of rice plants¨, Crop Science, 49(1):
313–322, 2009b.
Mohammed, AR y Tarpley, L¨Effects of Night Temperature, Spikelet Position and Salicylic
Acid on Yield and Yield-Related Parameters of Rice (Oryza sativa L.) Plants¨, Journal of
Agronomy and Crop Science, 197(1): 40–49, 2011a.
Mohammed, AR y Tarpley, L¨High night temperature and plant growth regulator effects on
spikelet sterility, grain characteristics and yield of rice (Oryza sativa L.) plants¨, [en línea]
Canadian Journal of Plant Science, ISSN-0008-4220, DOI-10.4141/CJPS10038, 91(2):
283–291, 1 de enero de 2011b.
Mohammed, R., Cothren, JT y Tarpley, L¨High Night Temperature and Abscisic Acid
Affect Rice Productivity through Altered Photosynthesis, Respiration and Spikelet
Fertility¨, [en línea] Crop Sci., DOI-10.2135/cropsci2013.01.0060, 53(6): 2603–2612,
2013.
Monneveux, Philippe., Pastenes, Claudio y Reynolds, Matthew P¨Limitations to
photosynthesis under light and heat stress in three high-yielding wheat genotypes.¨, [en
línea] Journal of plant physiology, ISSN-0176-1617, DOI-10.1078/0176-1617-00772,
160(6): 657–66, junio de 2003.
Murata, Norio., Takahashi, Shunichi., Nishiyama, Yoshitaka y Allakhverdiev, Suleyman
I¨Photoinhibition of photosystem II under environmental stress.¨, [en línea] Biochimica et
biophysica acta, ISSN-0006-3002, DOI-10.1016/j.bbabio.2006.11.019, 1767(6): 414–21,
junio de 2007.
Murchie, EH y Lawson, T Chlorophyll fluorescence analysis: A guide to good practice and
understanding some new applications, 2013,
Narayanan, Sruthi., Tamura, Pamela J., Roth, Mary R., Prasad, PV Vara y Welti,
Ruth¨Wheat leaf lipids during heat stress: I. High day and night temperatures result in major
lipid alterations¨, [en línea] Plant, Cell & Environment, ISSN-01407791, DOI-
10.1111/pce.12649, (39): 787–803, 2016.

Bibliografía 81
Ort, Donald R y Baker, Neil R¨A photoprotective role for O2 as an alternative electron sink
in photosynthesis?¨, [en línea] Current Opinion in Plant Biology, ISSN-13695266, DOI-
10.1016/S1369-5266(02)00259-5, 5(3): 193–198, 2002.
Oxborough, Kevin y Baker, NeilR¨Resolving chlorophyll a fluorescence images of
photosynthetic efficiency into photochemical and non-photochemical components –
calculation of qP and Fv-/Fm-; without measuring Fo-;¨, [en línea] Photosynthesis
Research, ISSN-0166-8595, DOI-10.1023/A:1005936823310, 54(2): 135–142, 1997.
Panda, Debabrata y Sarkar, Ramani Kumar¨Natural leaf senescence: probed by chlorophyll
fluorescence, CO(2) photosynthetic rate and antioxidant enzyme activities during grain
filling in different rice cultivars¨, [en línea] Physiology and molecular biology of plants : an
international journal of functional plant biology, ISSN-0971-5894, DOI-10.1007/s12298-
012-0142-6, 19(1): 43–51, 27 de enero de 2013.
Pask, Alistair., Pietragalla, Julian y Mullan, Debra Physiological Breeding II: A Field Guide
to Wheat Phenotyping., [en línea] pp. 140, ISBN-9789706481825, 2012.
Pastenes, Claudio y Horton, Peter¨Effect of High Temperature on Photosynthesis in Beansl¨,
[en línea] Plant Physiology, DOI-10.1007/s11099-008-0100-2, 112(112): 1253–1260,
1996.
Peng, Shaobing., Huang, Jianliang., Sheehy, John E., Laza, Rebecca C., Visperas, Romeo
M., Zhong, Xuhua., Centeno, Grace S., Khush, Gurdev S y Cassman, Kenneth G¨Rice yields
decline with higher night temperature from global warming¨, [en línea] Proceedings of the
National Academy of Sciences of the United States of America, DOI-
10.1073/pnas.0403720101, 101(27): 9971–9975, 2004.
Plaxton, William C y Podestá, Florencio E¨The Functional Organization and Control of
Plant Respiration¨, [en línea] Critical Reviews in Plant Sciences, ISSN-0735-2689, DOI-
10.1080/07352680600563876, 25(September 2014): 159–198, 2006.
Poli, Yugandhar., Basava, Ramana Kumari., Panigrahy, Madhusmita., Vinukonda, Vishnu
Prasanth., Dokula, Nageswara Rao., Voleti, Sitapathi Rao., Desiraju, Subrahmanyam y
Neelamraju, Sarla¨Characterization of a Nagina22 rice mutant for heat tolerance and
mapping of yield traits.¨, [en línea] Rice (New York, N.Y.), ISSN-1939-8425, DOI-
10.1186/1939-8433-6-36, 6(1): 36, enero de 2013.
Restrepo-Diaz, Hermann y Garces-Varon, Gabriel Response of rice plants to heat stress
during initiation of panicle primordia or grain-filling phases, 2013,
Reynolds, M., Manes, Y., Izanloo, A y Langridge, P¨Phenotyping approaches for
physiological breeding and gene discovery in wheat¨, [en línea] Annals of Applied Biology,
ISSN-1744-7348, DOI-10.1111/j.1744-7348.2009.00351.x, 155(3): 309–320, 1 de
diciembre de 2009.

82 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
Rodríguez, Mario Gutiérrez., Reynolds, Matthew Paul., Estrada, José Alberto Escalante y
Larqué-Saavedra, Alfonso¨Algunas consideraciones en la relación entre fotosíntesis y el
rendimiento de grano en trigo¨, CIENCIA ergo-sum, ISSN-1405-0269, 12(2): 149–154,
2005.
Sage, RF y Kubien, DS¨The temperature response of C3 and C4 photosynthesis¨, Plant, Cell
and Environment, 30(9): 1086–1106, 2007.
Sage, Tammy L., Bagha, Shaheen., Lundsgaard-Nielsen, Vanessa., Branch, Haley A.,
Sultmanis, Stefanie y Sage, Rowan F¨The effect of high temperature stress on male and
female reproduction in plants¨, [en línea] Field Crops Research, ISSN-03784290, DOI-
10.1016/j.fcr.2015.06.011, 182: 30–42, octubre de 2015.
Salvucci, ME y Crafts-Brandner, SJ¨Inhibition of photosynthesis by heat stress: The
activation state of Rubisco as a limiting factor in photosynthesis¨, Physiologia Plantarum,
120(2): 179–186, 2004.
Sánchez, Berta., Rasmussen, Anton y Porter, John R¨Temperatures and the growth and
development of maize and rice: A review¨, [en línea] Global Change Biology, ISSN-
13541013, DOI-10.1111/gcb.12389, 20: 408–417, 2014.
Sánchez-Reinoso, Alefsi David., Garcés-Varón, Gabriel y Restrepo-Díaz,
Hermann¨Biochemical and physiological characterization of three rice cultivars under
different daytime temperature conditions¨, [en línea] Chilean journal of agricultural
research, ISSN-0718-5839, DOI-10.4067/S0718-58392014000400001, 74(4): 373–379,
diciembre de 2014.
Sayed, OH¨Chlorophyll Fluorescence as a Tool in Cereal Crop Research¨, [en línea]
Photosynthetica, DOI-10.1023/B:PHOT.0000015454.36367.e2, 41(3): 321–330, 2003.
Scafaro, Andrew P., Yamori, Wataru., Carmo-Silva, A Elizabete., Salvucci, Michael E., Von
Caemmerer, Susanne y Atwell, Brian J¨Rubisco activity is associated with photosynthetic
thermotolerance in a wild rice ( Oryza meridionalis)¨, [en línea] Physiologia Plantarum,
DOI-10.1111/j.1399-3054.2012.01597.x, 146(1): 99–109, 2012.
Scafaro, AP., Haynes, PA y Atwell, BJ¨Physiological and molecular changes in Oryza
meridionalis Ng., a heat-tolerant species of wild rice¨, Journal of Experimental Botany,
61(1): 191–202, 2010.
Shah, F., Huang, J., Cui, K., Nie, L., Shah, T., Wu, W., Wang, K., Khan, ZH., Zhu, L y
Chen, C¨Physiological and biochemical changes in rice associated with high night
temperature stress and their amelioration by exogenous application of ascorbic acid (vitamin
C)¨, Australian Journal of Crop Science, 5(13): 1810–1816, 2011.
Shi, Wanju., Muthurajan, Raveendran., Rahman, Hifzur., Selvam, Jagadeesh., Peng,

Bibliografía 83
Shaobing., Zou, Yinbin y Jagadish, Krishna S V¨Source–sink dynamics and proteomic
reprogramming under elevated night temperature and their impact on rice yield and grain
quality¨, [en línea] New Phytologist, ISSN-1469-8137, DOI-10.1111/nph.12088, 197(3):
825–837, 1 de febrero de 2013.
Shikanai, Toshiharu¨Central role of cyclic electron transport around photosystem I in the
regulation of photosynthesis¨, [en línea] Current Opinion in Biotechnology, ISSN-
09581669, DOI-10.1016/j.copbio.2013.08.012, 26(Figure 1): 25–30, 2014.
Sikder, S., Foulkes, J., West, H., De Silva, J., Gaju, O., Greenland, A y Howell, P¨Evaluation
of photosynthetic potential of wheat genotypes under drought condition¨, [en línea]
Photosynthetica, ISSN-0300-3604, DOI-10.1007/s11099-015-0082-9, 53(1): 47–54, 2015.
Smith, Nicholas G y Dukes, Jeffrey S¨Plant respiration and photosynthesis in global-scale
models: Incorporating acclimation to temperature and CO2¨, [en línea] Global Change
Biology, ISSN-13541013, DOI-10.1111/j.1365-2486.2012.02797.x, 19(1): 45–63, 2013.
Solarte, María Elena., Moreno, Leonard y Melgarejo, Luz Marina ¨FOTOSÍNTESIS Y
PIGMENTOS VEGETALES¨, edit. Luz Marina. Melgarejo, Experimentos en Fisiología
Vegetal, Primera ed ed. , pp. 279, Ed. Universidad Nacional de Colombia, 2010.
Song, Lili., Yue, Lulu., Zhao, Huaqiang y Hou, Meifang¨Protection effect of nitric oxide on
photosynthesis in rice under heat stress¨, [en línea] Acta Physiologiae Plantarum, DOI-
10.1007/s11738-013-1365-z, 35(12): 3323–3333, 2013.
Szabados, L y Savouré, A¨Proline: a multifunctional amino acid¨, Trends in Plant Science,
15(2): 89–97, 2010.
Taiz, Lincon y Zeiger, Eduardo Plant physiology, Ed. Sinaver Associates, Inc, Book. vol.
Fourth, pp. 700, 2006.
Tanaka, Yoshikazu., Sasaki, Nobuhiro y Ohmiya, Akemi¨Biosynthesis of plant pigments:
Anthocyanins, betalains and carotenoids¨, [en línea] Plant Journal, ISSN-09607412, DOI-
10.1111/j.1365-313X.2008.03447.x, 54(4): 733–749, 2008.
Teixeira, Edmar I., Fischer, Guenther., Van Velthuizen, Harrij., Walter, Christof y Ewert,
Frank¨Global hot-spots of heat stress on agricultural crops due to climate change¨, [en línea]
Agricultural and Forest Meteorology, ISSN-01681923, DOI-
10.1016/j.agrformet.2011.09.002, 170: 206–215, marzo de 2013.
Turnbull, MH., Whitehead, D., Tissue, DT., Schuster, WS., Brown, KJ y Griffin,
KL¨Responses of leaf respiration to temperature and leaf characteristics in three deciduous
tree species vary with site water availability.¨, [en línea] Tree physiology, ISSN-0829-318X,
DOI-10.1093/treephys/21.9.571, 21(9): 571–8, 2001.
Vani, B., Pardha Saradhi, P y Mohanty, Prasanna¨Alteration in chloroplast structure and

84 Respuestas Fisiológicas de Siete Genotipos de Arroz
(Oryza sativa L) a Diferentes Temperaturas Nocturnas
thylakoid membrane composition due to in vivo heat treatment of rice seedlings: correlation
with the functional changes¨, [en línea] Journal of Plant Physiology, DOI-
http://dx.doi.org/10.1078/0176-1617-00260, 158(5): 583–592, 2001a.
Vani, B., Saradhi, PP y Mohanty, P¨Characterization of high temperature induced stress
impairments in thylakoids of rice seedlings¨, (April), 2001b.
Venkateswarlu, B., Shanker, Arun K., Shanker, Chitra y Maheswari, M ¨Heat Stress in Rice
– Physiological Mechanisms and Adaptation Strategies¨, [en línea] Crop Stress and its
Management: Perspectives and Strategies, ISBN-9789400722200, vol. 9789400722, pp.
193–224, 2012.
Wahid, A., Gelani, S., Ashraf, M y Foolad, MR¨Heat tolerance in plants: An overview¨,
Environmental and Experimental Botany, 61(3): 199–223, 2007.
Wang, Dun., Li, Xiao Fei., Zhou, Zheng Jian., Feng, Xu Ping., Yang, Wan Jun y Jiang, De
An¨Two Rubisco activase isoforms may play different roles in photosynthetic heat
acclimation in the rice plant¨, [en línea] Physiologia Plantarum, ISSN-00319317, DOI-
10.1111/j.1399-3054.2009.01344.x, 139(1): 55–67, 2010.
Wassmann, R., Jagadish, SVK., Heuer, S., Ismail, A., Redona, E., Serraj, R., Singh, RK.,
Howell, G., Pathak, H y Sumfleth, K ¨CLIMATE CHANGE AFFECTING RICE
PRODUCTION: THE PHYSIOLOGICAL AND AGRONOMIC BASIS FOR POSSIBLE
ADAPTATION STRATEGIES¨, [en línea] edit. D L Sparks, Advances in Agronomy, Vol
101, ISBN-0065-2113978-0-12-374817-1, vol. 101, pp. 59–122, Ed. Elsevier Academic
Press Inc, San Diego, 2009.
Welch, Jarrod R., Vincent, Jeffrey R., Auffhammer, Maximilian., Moya, Piedad F.,
Dobermann, Achim y Dawe, David¨Rice yields in tropical/subtropical Asia exhibit large but
opposing sensitivities to minimum and maximum temperatures.¨, [en línea] Proceedings of
the National Academy of Sciences of the United States of America, ISSN-1091-6490, DOI-
10.1073/pnas.1001222107, 107(33): 14562–7, 17 de agosto de 2010a.
Welch, JR., Vincent, JR., Auffhammer, M., Moya, PF., Dobermann, A y Dawe, D¨Rice
yields in tropical/subtropical Asia exhibit large but opposing sensitivities to minimum and
maximum temperatures¨, Proceedings of the National Academy of Sciences of the United
States of America, 107(33): 14562–14567, 2010b.
Wongpatsa, Usanee., Kaveeta, Lily., Sriwongchai, Tanee y Khamsuk, Ornusa¨Effects of
temperature on male sterility of two inbred lines of hybrid rice¨, Kasetsart Journal - Natural
Science, ISSN-00755192, 48(4): 525–533, 2014.
Xue, DW., Jiang, H., Hu, J., Zhang, XQ., Guo, LB., Zeng, DL., Dong, GJ., Sun, GC y Qian,
Q¨Characterization of physiological response and identification of associated genes under

Bibliografía 85
heat stress in rice seedlings¨, Plant Physiology and Biochemistry, 61: 46–53, 2012.
Yamasaki, Takenobu., Yamakawa, Tomokazu., Yamane, Yoshihiro., Koike, Hiroyuki y
Satoh, Kazuhiko¨Temperature Acclimation of Photosynthesis and Related Changes in
Photosystem II Electron Transport in¨, [en línea] Plant Physiology, ISSN-0032-0889, DOI-
10.1104/pp.010919.1, 128(March): 1087–1097, 2002.
Yamori, Wataru., Masumoto, Chisato., Fukayama, Hiroshi y Makino, Amane¨Rubisco
activase is a key regulator of non-steady-state photosynthesis at any leaf temperature and,
to a lesser extent, of steady-state photosynthesis at high temperature¨, [en línea] Plant
Journal, ISSN-09607412, DOI-10.1111/j.1365-313X.2012.05041.x, 71(6): 871–880, 2012.
Yamori, Wataru., Takahashi, Shunichi., Makino, Amane., Price, G Dean., Badger, Murray
R y Von Caemmerer, Susanne¨The roles of ATP synthase and the cytochrome b6/f
complexes in limiting chloroplast electron transport and determining photosynthetic
capacity.¨, [en línea] Plant physiology, ISSN-0032-0889, DOI-10.1104/pp.110.168435,
155(2): 956–962, 2011.
Yan, K., Chen, P., Shao, H., Zhao, S., Zhang, L., Xu, G., Sun, J., Zhang, L., Xu, G y Sun,
J¨Responses of Photosynthesis and Photosystem II to Higher Temperature and Salt Stress in
Sorghum¨, [en línea] Journal of Agronomy and Crop Science, ISSN-09312250, DOI-
10.1111/j.1439-037X.2011.00498.x, 198(3): 218–225, 2012.
Yin, Y., Li, S., Liao, W., Lu, Q., Wen, X y Lu, C¨Photosystem II photochemistry,
photoinhibition, and the xanthophyll cycle in heat-stressed rice leaves¨, Journal of Plant
Physiology, 167(12): 959–966, 2010.
Yoshida, Shouichi Fundamentals of rice crop science, Ed. Int. Rice Res. Inst., BOOK.
ISBN-9711040522, 1981.
Yoshida, Shouichi Fundamentals of Rice Yield,
Zapata, JM., Guéra, A., Esteban-Carrasco, A., Martín, M y Sabater, B¨Chloroplasts regulate
leaf senescence: delayed senescence in transgenic ndhF-defective tobacco.¨, [en línea] Cell
death and differentiation, ISSN-1350-9047, DOI-10.1038/sj.cdd.4401657, 12(10): 1277–
84, 2005.
Zhang, Y., Tang, Q., Peng, S., Zou, Y., Chen, S., Shi, W., Qin, J y Laza, MRC¨Effects of
high night temperature on yield and agronomic traits of irrigated rice under field chamber
system condition¨, Australian Journal of Crop Science, 7(1): 7–13, 2013.
Zhou, Wei-hi., Xue, Da-wei y Zhang, Guo-ping-¨Identification and physiological
characterization of thermo-tolerant rice genotypes¨, Journal of Zhejiang University, 38(1):
1–9, 2012.





























