Embed Size (px)
DESCRIPTION
Molekulske Osnove Bioloskih Funkcija Hormoni
Citation preview

I) Proteini kao faktori zgrušavanja krvi.
II) Imunoglobulini u imunološkoj reakciji i zaštitnim mehanizmima
ćelije i tkiva.
III) Interakcije antitelo-antigen, biohemija krvnih grupa.
Molekulske osnove bioloških funkcija.

Hemostaza (zaustavljanje krvarenja)
Hemostaza se odvija u nekoliko faza:
Ostvaruje se formiranjem krvnog ugruška (koaguluma) u zidovima oštećenih krvnih sudova, čime se sprečava gubitak krvi, a krv održava u tečnom stanju unutar vaskularnog sistema. Da bi se formirao krvni ugrušak neophodno je uključivanje većeg broja sistema i koordinisanih mehanizama. Pored toga, skup složenih, međusobno povezanih mehanizama, kao i lokalni faktori, učestvuju u održavanju ravnoteže između procesa koagulacije i antikoagulacije.
1) spazam krvnog suda 2) Formiranje privremenog trombocitnog čepa ili prstena 3) koagulacija 4) retrakcija i fibrinoliza koaguluma i urastanje vezivnog tkiva

Proteini kao faktori zgrušavanja krvi.
- Proteini iz krvne plazme - Faktori iz trombocita - endotelijalne ćelije krvnih sudova - vitamin K
Po oštećenju krvnog suda, formira se ugrušak kao posledica agregacije trombocita i formira se nerastvorna mreža fibrina koja zaustavlja i druge krvne ćelije.
Kaskada koagulacije krvi uključuje niz proteina od kojih neki imaju i enzimsku aktivnost: • Enzimi: faktori: II, VII, IX, X (zahtevaju vitamin K za svoju sintezu) • Neenzimeske proteinske faktore – označeni su rimskim brojevima (sa a u indeksu za aktivnu formu): Tkvini faktor, V i VIII, kininogen

Konačni cilj jeste da se formira mreža vlakana fibrina, koja će zaustavljati protok krvi, nagomilavanjem krvnih ćelija – što je moguće nakon što
Fibrin nastaje iz fibrinogena – cirkulišućeg proteina, pod dejstvom aktiviranog trombina.
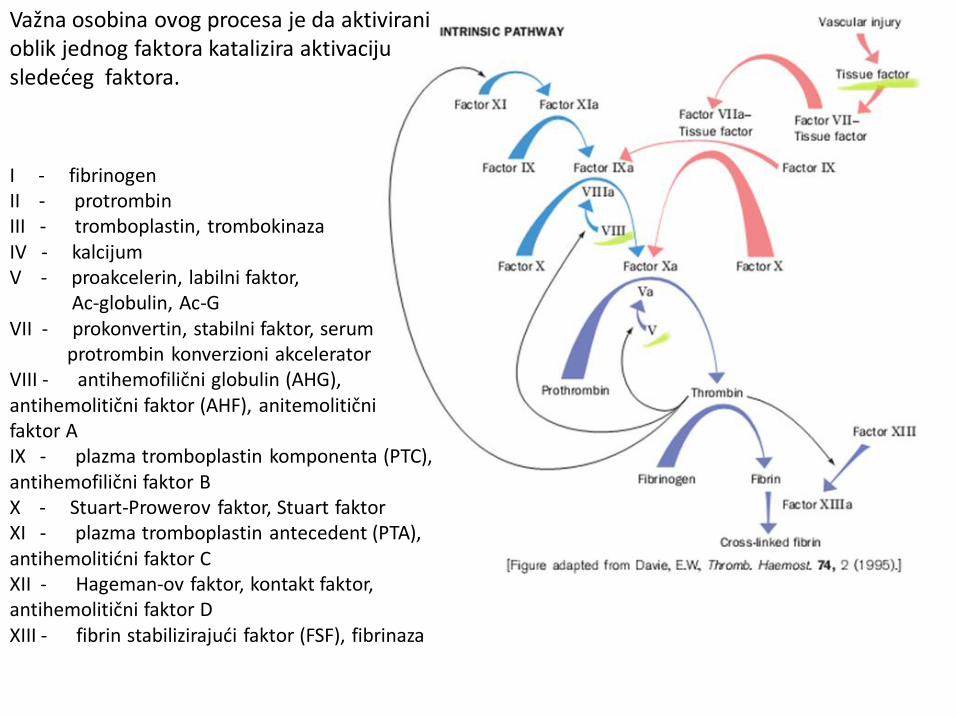
Važna osobina ovog procesa je da aktivirani oblik jednog faktora katalizira aktivaciju sledećeg faktora.
I - fibrinogen II - protrombin III - tromboplastin, trombokinaza IV - kalcijum V - proakcelerin, labilni faktor, Ac-globulin, Ac-G VII - prokonvertin, stabilni faktor, serum protrombin konverzioni akcelerator VIII - antihemofilični globulin (AHG), antihemolitični faktor (AHF), anitemolitični faktor A IX - plazma tromboplastin komponenta (PTC), antihemofilični faktor B X - Stuart-Prowerov faktor, Stuart faktor XI - plazma tromboplastin antecedent (PTA), antihemolitićni faktor C XII - Hageman-ov faktor, kontakt faktor, antihemolitični faktor D XIII - fibrin stabilizirajući faktor (FSF), fibrinaza

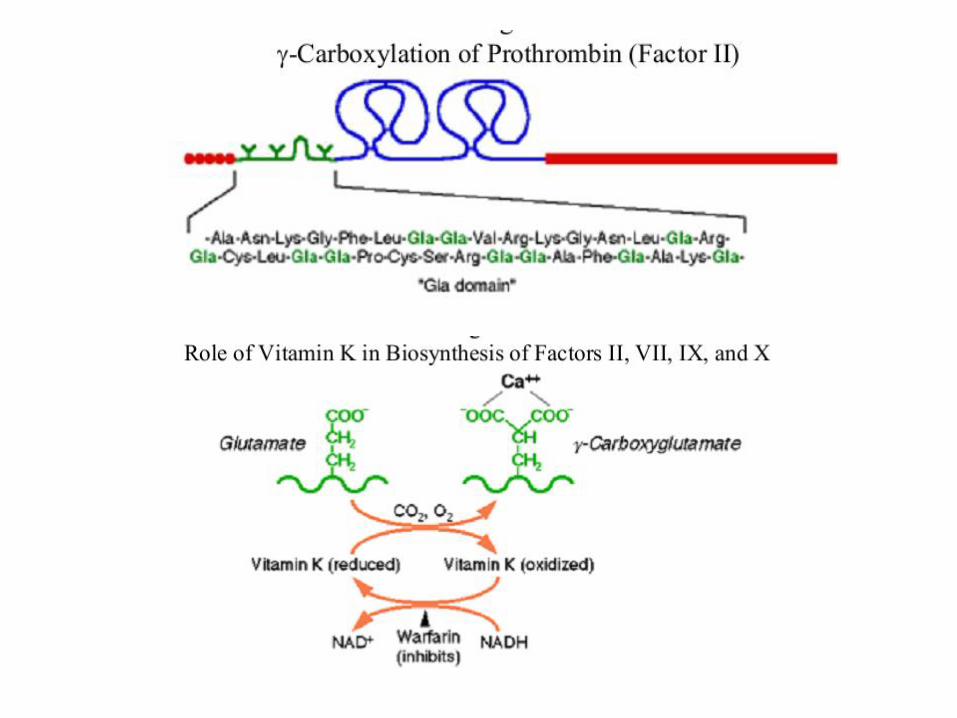
Kako se vrši prevođenje protrombina u trombin? Protrombin se, nakon što helira Ca2+ jone svojim GLA regionom, vezuje za fosfolipide na membrani krvnih pločica (trombocita). Time se protrombin dovodi u bliski kontakt sa dva proteina koji katalizuju njegovu konverziju u trombin.
Vezivanje Ca2+ jona za modifikovane ostatke glutaminske kiseline (γ-karboksi glutamat). Ovako modifikovan ostatak AK, mnogo je jači helatorski agens od ostatka glutaminske kiseline. Ova post-translaciona modifikacija protrombina, dešava se u prisustvu vitamina K.
trombin
Interakcije između komponenti „protrombinaznog” kompleksa

Faktor Xa prevodi protromin (faktor II) u trombin (faktor IIa) tako što vodi raskidanju dve peptidne veze u ovom zimogenu. Ovu reakciju ubrzava faktor Va, fosfolipidi na membrani trombocita, kao i kalcijumovi joni. Smatra se da agregacija trombocita obezbeđuje fosfolipidnu površinu na kojoj se dešava aktivacija protrombina (na mestu hemostaze). Smatra se da se aktivacija faktora X pomoću faktora IXa i njegovog kofaktora VIIIa odigrava po veoma sličnom mehanizmu .
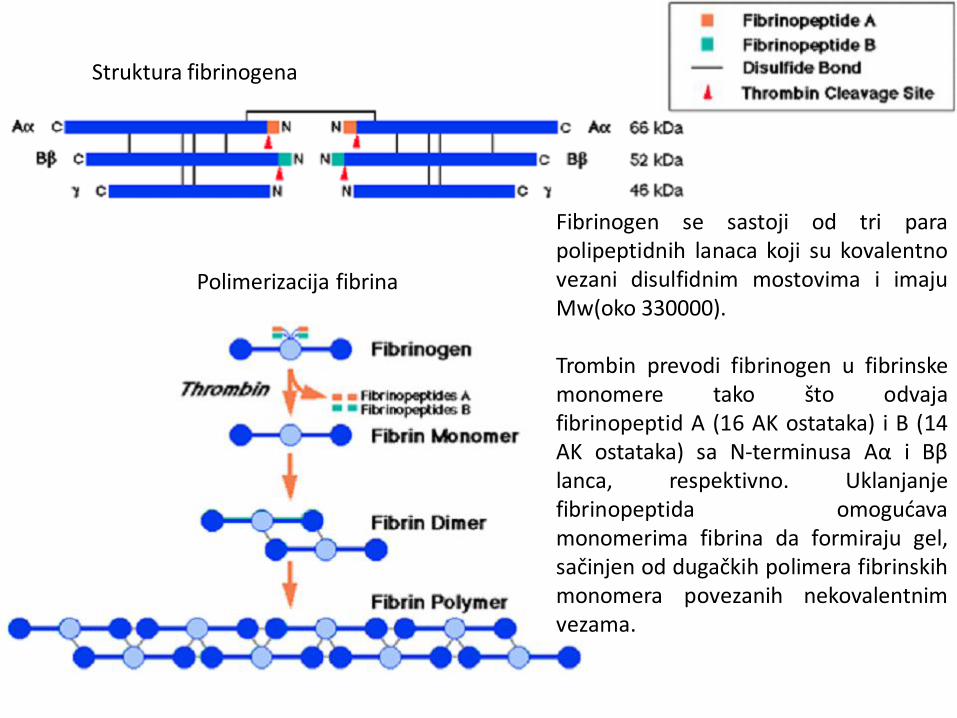
Struktura fibrinogena
Polimerizacija fibrina
Fibrinogen se sastoji od tri para polipeptidnih lanaca koji su kovalentno vezani disulfidnim mostovima i imaju Mw(oko 330000). Trombin prevodi fibrinogen u fibrinske monomere tako što odvaja fibrinopeptid A (16 AK ostataka) i B (14 AK ostataka) sa N-terminusa Aα i Bβ lanca, respektivno. Uklanjanje fibrinopeptida omogućava monomerima fibrina da formiraju gel, sačinjen od dugačkih polimera fibrinskih monomera povezanih nekovalentnim vezama.

A) Struktura zimogenih oblika proteaza uključenih u koagulaciju - Svaki od navedenih proteina se sintetiše u jetri i iz nje sekretuje u krvotok. Svaki od
ovih zimogenih formi sadrži signalni peptid koji se odvaja u toku tranzita u ER - Oko 200 ak ostataka na C-terminalnom kraju svakog od navedenih zimogena ima
homologan deo tripsinu (proteazi), i sadrži aktivni centar sa Ser, Asp i His ostacima - ovi proteini takođe sadrže zantan broj domena koji su homologi delovima drugih
proteina (EGF i fibronektin). Smatra se da su ti domeni odgovorni za specifične interakcije između proteaza i njihovih supstrata, kofaktora i/ili infibitora
Biohemija koagulacije:

Biohemija koagulacije:
Faktori II, VII, IX i X su međusobno homologi na N- terminusu. Nakon uklanjanja signalnog peptida, karboksilaza koja se nalazi u ER ili u Goldži aparatu, se vezuje za propeptidni deo svakog od ovh proteina i vodi modifikaciji 10-12 ostataka glutaminske kiseline (smeštenih u susednom regionu) – prevodeći ih u γ-karboksiglutamate. Neposredno pre sekrecije ovako modifikovanih polipeptida, uklanja se propeptidni region.

B) Proteinski (ne enzimskih) faktori koagulacije
• Faktori V i VIII su veliki proteini plazma membrane koji u sekvenci sadrže ponavljajuće domene, koji su homologi domenima ceruloplazmina (A1, A2 i A3 domeni, na slici). Ova dva faktora bivaju aktivirani pomoću trombina. Aktivirani V i VIII nemaju enzimsku aktivnost već imaju ulogu kofaktora proteaza Xa i IXa, respektivno. Faktor VIII cirkuliše u plazma membrani vezan za von Willebrand-ov faktor • Tkivni faktor je membranski integralni protein, koji je ekspresovan na površini aktiviranih monocita, endotelijalnih ćelija izloženih delovanju različitih citokina (npr. Tumor nekrozis faktor) i drugim ćelijama. Ne nalazi se u plazmi. Ovaj faktor u značajnoj meri povećava proteolitičku aktivnost enzima VIIa.

Aktivacija faktora VIII pomoću trombina:
Proteoliza, pre sekrecije


Faktor XIII - fibrin stabilizirajući faktor (FSF), fibrinaza
Transglutaminazna aktivnost XIIIa faktora
Kovalentno unakrsno povezivanje fibrinskih polimera koje je aktivirano faktorom XIIIa, značajno je za pravilno formiranje ugruška kao i proces oporavka rane u in vivo sistemima. Zimogena forma faktora XIII biva aktivirana (XIIIa) delovanjem trombina. XIIIa katalizuje transglutaminaciju - kojom se unakrsno povezju C-terminusi γ-lanaca sa susednim monomerima fibrina, ali se uočavaju i dodatna povezivanja (sve u cilju učvršćivanja ugruška). Prisustvo i aktivnost faktora XIII, može se proveriti ispitivanjem rastovrljivosti ugruška u prisustvu uree, koja ne može da raskine ovako unakrsno uspostavljene kovalente veze.

Mesto odvijanja reakcija koje vode koagulaciji. Usled enzimske aktivnosti brojnih faktora koagulacije, dolazi do pojačavanja početnog stimulusa za koagulaciju krvi. Svaki kompleks između tkivnog faktroa i faktora VIIa, može da proizvede veliki broj Xa molekula, koji pak produkuju još i već ukoličinu trombina. Ovo pojačavanje procesa koagulacije može da potiče i od pozitivne sprege: tj. Aktivacijom faktora V, VIII i XI pomoću tromibina kao i aktivacije faktora VII pomoću faktora Xa. Reakcije koje se odvijaju na površini membrana su: -Aktivacija X i IX pomoću VIIa/tkivni faktor kompleksa na površini glatkih mišićnih ćelija koje su lokalizovane ispod enotela krvnih sudova - Aktivacija X pomoću Ixa/VIIIa kompleksa na površini aktiviranih trombocita (na mestu ovrede) - aktivacija protrombina pomoću Xa/Va kompleksa na površini aktiviranih trombocita.

Trombociti - ispusti povezani zakačeni za fibrinske niti - kontrakcija (aktin i miozin) - povlačenje fibrinskih niti = retrakcija koaguluma - približavanje ivica ozleđenog krvnog suda (ubrzava zarastanje) - u zonu ozlede migriraju fibrociti (formira se novo vezivno tkivo) + epitelijalne ćelije oko ozlede proliferišu (ispunjava se ozleđena površina).
Retrakcija koaguluma + serum

Protrombin TROMBIN koag. krvi
na pov. endotel. cel.
trombomodulin
Protein C
(vit. K zavisan protein, iz jetre)
Protein S (vit. K zavisan prot.; iz
endotela, megakariocita I
hepatocita)
VIIa inakt VII Va inakt V
Inaktivacija inhibitora
tkivnog aktivatora plazminogena
Plazminogen plazmin
fibrinoliza
aktivni protein C

Klinički značaj
Dobro je poznata funkcija GAG heparina u sprečavanju koagulacije krvi.
Heparin je rasprostranjen u granulama ćelija koje oivičavaju krvne
sudove. Oslobađanje heparina iz ovih granula pri povredi i njegov ulaz u
serum dovodi do inhibicije koagulacije krvi na sledeći način: slobodan heparin se kompleksira sa antitrombinom III i aktivira ga, što dovodi do inhibicije svih serin-zavisnih proteaza u kaskadi koagulacije. Ova pojava se koristi za primenu heparina kao antikoagulansa u terapiji.

II) Imunoglobulini u imunološkoj reakciji .
• Imuni sistem ima sposobnost da razlikuje strane od sopstvenih biomolekula. Ulaz stranih molekula dovodi do imunog odgovora i sinteze antitela. Strani molekuli se nazivaju antigeni. Imuni sistem može da prepozna više miliona stranih supstanci-antigena. Imuni sistem se deli na:
• Humoralni (antitela-imunoglobulini)
• Celularni: makrofagi, limfociti (nalaze se u krvi, limfi, limfnim žlezdama i limfoidnim organima (slezina, timus, limfni čvorovi). B-limfociti (nastaju u koštanoj srži) i T-limfociti odgovorni za celularni imunitet i razvijaju se u timusu. T-ćelije imaju ključnu ulogu jer pomažu B-ćelijama u produkciji antitela.
IMUNI SISTEM


• Tokom evolucije kod kičmenjaka se razvio sistem za
odbranu od toksina i mikroorganizama, uklanjanje
starih i oštećenih ćelija. Imuni sistem prvo mora da
prepozna stranu materiju, antigen; potom proizvodi
specifične imunoglobuline (antitela) koja će vezati
antigen u antitelo-antigen kompleks (nekovalentnim
vezama). Ovaj kompleks se zatim eliminiše iz
organizma, najčešće putem fagocitoze.
ANTITELA

ANTITELA
• Brzina i sposobnost da se fiksira određena
mutacija može veoma da se razlikuje za
različite regione unutar sekvence proteina.
• Imuni sistem viših organizama zavisi od veoma
brzog stvaranja novih molekula koji će vezati i
raspoznati strane supstance koje mogu biti
štetne za organizam. Ovu funkciju obavljaju
proteini iz klase imunoglobulina, vezujući
antigen za varijabilni region.

• Antitela (Imunoglobulini) su veliki glikoproteinski molekuli, karatkerističnog oblika koji podseća na slovo Y. • Sekretuju ih plazma ćelije (vrsta B limfocita) – a funkcija im je da identifikuju i obeleže strana tela, kao što su bakterije i virusi).
Antitela prepoznaju jedinstvenu strukturu na ciljnom (stanom) telu – koji se naziva antigen.

ANTITELA
Osnove jedinice građe imunoglobulina su: -Dva velika teška lanca - dva mala, laka lanca.
Svaki teški lanac se sastoji od dva regiona: - Konstantnog i varijabilnog regiona
1. Fab region = antigen binding 2. Fc region 3. Heavy chain (blue) with one variable (VH) domain followed by a constant domain (CH1), a hinge region, and two more constant (CH2 and CH3) domains. 4. Light chain (green) with one variable (VL) and one constant (CL) domain 5. Antigen binding site (paratope) 6. Hinge regions.

Klase antitela: Humani Ig se mogu podeliti u 5 klasa: (na osnovu razlike u AA sekvenci u konstantnom regionu teškog lanca – koji se označavaju sa grčkim slovima α δ ε γ i μ.

ANTITELO-ANTIGEN INTERAKCIJE
Regioni koji intereaguju nazivaju se epitop (na antigenu) i paratop (na antitelu).
-Antigeni mogu imati različite epitope, raspoređenje diskontinualno na svojoj površini. Ali glavni epitopi se nazivaju još i determinantni.
-Inerakcija je komplementarna (po sistemu ključ-brava). Interakcije Fab-epitop su slabe i ne-specifične i to: elektrostatičke sile, vodonične veze, hidrofobne interakcije, van der Waals-ove sile. -To ukazuje na to da su ove interakcije reverzibilne i da je moguće da antitelo ukršteno reaguje saa različitim antigenima i različitim afinitetima.

Na svakom vrhu ovih molekula Y-oblika nalaze se paratopi – koji su specifični za odgovarajuće epitope na antigenu. (prepoznavanje je tipa: ključ-brava). Vezivanjem stranog tela, pomoću ovog specifičnog prepoznavanja, antitelo može da obeleži mikrob ili inficiranu ćeliju – koju zatim napada imuni sistem (fagocitoza) ili samo antitelo može da neutrališe svoju metu (ako se vezivanje ujedno blokira deo mikroba koji je značajan za njegovo invazivno delovanje ili preživljavanje)

Proizvodnja antitela
Antitela se produkuju u jednoj vrsti B-limfocita (tip belih krvnih zrnaca), koje se nazivaju plazma ćelije. Antitela se javljaju u dva oblika: – rastvorni (koji se sekretuju iz ćelije) – i kružeći krvotokom tragaju za stranim telima ili mikroorganizmima. - vezani za ćelijsku membranu B ćelija (i označavaju se kao BCR (B cell receptor). Ovi molekuli (vezani za membranu) olakšavaju aktivaciju B- ćelija limfocita i kao posledica toga i diferencijaciju ovih ćelija – u fabrike antitela (plazma ćelije) ili memorijske ćelije – koje će doprinose specifičnom imunom sistemu.

Biohemija krvne grupe
• ABO su kompleksni oligosaharidi koji se nalaze u skoro svim ćelijama u telu i odreženim ekstracelularnim sekretima. Na membranama eritrocita oligosaharidi koji određuju specifičnu prirodu ABO supstanci su uglavnom glikosfingolipidi, dok su u sekretima u formi glikoproteina.

Imunološki sistem i krvne grupe
• imuni sistem - oligosaharide koje organizam sintetiše prepoznaje kao “svoje”
• strane oligosaharidne sekvence → imuni odgovor
• antigeni krvnih grupa su
oligosaharidi vezani za proteine i
ceramid
• H-antigen (kod O grupe) sadrži β-
D-Gal, β-D-GlcNAc, β-D-Gal i u
položaju 2 vezanu α-L-Fuc
• kod grupe A, za Gal se vezuje α-D-
GalNAc, a kod grupe B α-D-Gal; AB
grupa ima obe vrste antigena
• grupa A ima anti-B antitela, grupa B
– anti-A, O grupa oba tipa, a AB
grupa nema ni anti-A ni anti-B

Individua tipa A ima anti-B-antitela u plazni i aglutiniraće kako sa B, tako i sa AB-tipom.
individua sa AB tipom, nema ni anti-A- ni anti-B antitela i to je univerzalni primalac.
Individua sa O-grupom ima i anti-A i anti-B-antitela i ona je univerzalni davalac.
Objašnjenje je da organizam neće stvarati antitela za supstance koje sam sadrži. Tako
individua tipa A neće proizvoditi antitela za A-antigen ali će proizvoditi za B i AB itd.
Geni odgovori za sintezu ABO supstanci nalaze se hromozomu 9. Tu postoje tri alele:A i
B su kodominatne i O receisivni.

BIOHEMIJSKA OSNOVA SISTEMA KRVNIH GRUPA
Danas je poznat 21 sistem krvnih grupa od kojih su najpoznatiji: ABO, Rh
(Rhesis) i Mn sistem.
Krvna grupa označava sistem antigena na eritrocitima (crvenim krvnim
zrnacima) koji su genetski kontrolisani na genetskom lokusu koji ima različit
broj alela (npr. A,B, O i ABO sistem). Termin krvni tip označava antigenski
fenotip, koji se prepoznaje po korišćenju određenih antitela.
ABO sistem krvnih grupa. Landsteiner, 1900 g.
Membrane eritrocita najvećeg broja individua sadrže jednu grupu supstanci
krvi: A tip, B-tip, AR ili O-tip.

Krvne grupe antigena

HORMONI

HORMONI
Hormoni su proizvodi endokrinih žlezda koje čine HUMORALNI integracioni sistem organizma. Oni putem krvi tj.telesnih tečnosti (humora) prenose poruke ciljnim organima ili tkivima na koja deluju preko specifičnih receptora. Dele se na: 1. HIDRROSOLUBILNE: PEPTIDNI HORMONI (INSULIN, GLUKAGON, HORMONI HIPOFIZE I
PARATHORMON) I KATEHOLAMINI. DELUJU PREKO RECEPTORA NA POVRŠINI ĆELIJE,A SIGNALE PRENOSE PREKO G-PROTEINA I SISTEMA cAMP i Ca2+ jona.
2. LIPOSOLUBILNI: STEROIDNI HORMONI I HORMONI TIREOIDNE ŽLEZDE: LAKO PROLAZE KROZ PLAZMA MEMBRANU, A NJIHOVI RECEPTORI SE NALAZE U CITOSOLU ILI JEDRU. STEROIDNI HORMONI SE VEZUJU ZA ODGOVARAJUĆE RECEPTORE ŠTO DOVODI DO KONFORMACIONIH PROMENA I NJIHOVE AKTIVACIJE I POVEĆANJA NJIHOVOG AFINITETA ZA ODREĐENE SEKVENCE DNA, ČIJA JE TRANSKRIPCIJA POD KONTROLOM STEROIDNIH HORMONA.
3. ANTIHORMONI: KONKURIŠU STEROIDNIM HORMONIMA ZA RECEPTOR ILI SPREČAVAJU TRANSFER HORMON-RECEPTOR KOMPLEKSA U JEDRO. NPR. ANTI-ESTROGEN TAMOKSIFEN KORISTI SE ZA LEČENJE PECIJENATA SA ESTROGEN-ZAVISNIM TUMORIMA.

MEHANIZAM DELOVANJA STREROIDNIH I NESTEROIDNIH
HORMONA

INSULIN
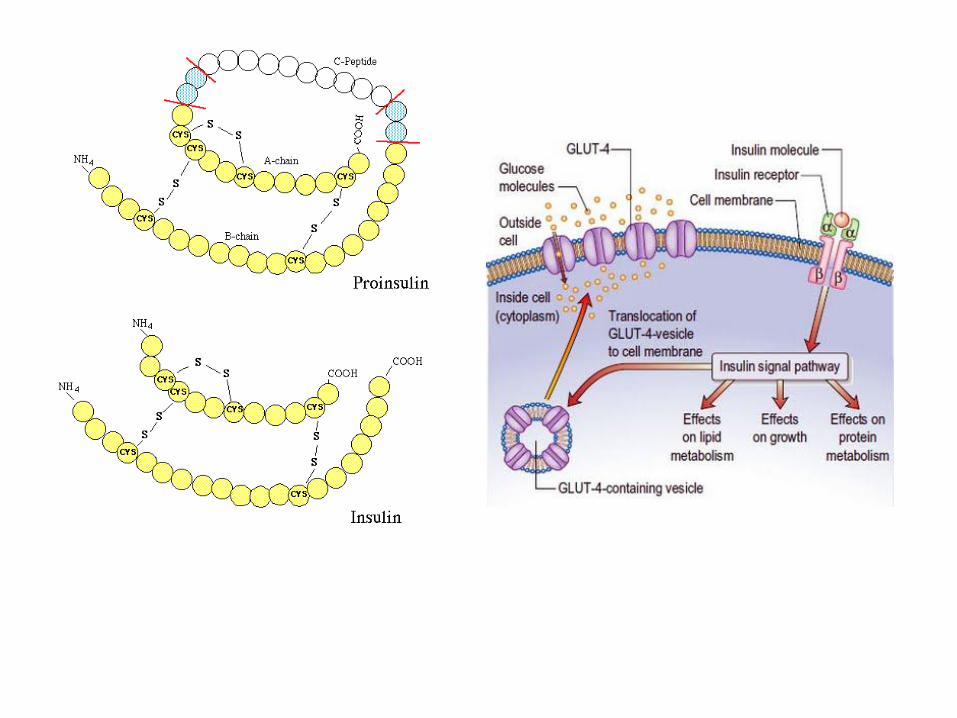
Insuln luče β-ćelije Langerhansovih ostrvaca pankreasa. Čine ga dva lanca A i B povezana sa dva disulfidna mosta i ukupno 51 aminokiselinska ostatka. U toku biosinteze nastaju neaktivni proenzimi: PREPROINSULIN I PROINSULIN. Odvajanje C-peptida nastaje aktivan hormon. Najjača stimulacija sekreceije insulina je porast glukoze u krvi. Takođe na sekreciju deluje i povećanje sadržaja aminokiselina u plazmi i pojačano lučenje gastrointestinalnih hormona (sekretina) nakon unošenja hrane. Insulin deluje preko receptora koji se nalaze u plazma membrani mišića i masnog tkiva. Receptor je tetramer izgrađem iz α i β subjedinica. β subjedinica prolazi kroz membranu, a njen domen koji je u citosolu ima aktivnost tirozin-protein-kinaze kouju aktivira insulin. α-subjedinica je smeštena ekstracelularno i vezuje hormon. Vezivanje insulina dovodi do autofosforilacije tirozinskog ostatke intracelularnog dela β subjedinice čime se aktiviraju procesi fosforilacije i defosforilacije. Insulin stimuliše ulazak glukoze u glikolizu ili njenu ugradnju u glikogen. Insulin ima anabolički efekat. Stimuliše glikolizu i glikoneogenezu, a inhibira glukoneogenezu i glikogenolizu. Takođe stimuliše sintezu troglicerida. U masnom tkivu inhibira dejstvo lipaza čime utiče na smanjenje koncentracije slobodnih masnih kiselina u krvi. Povećanjem glikolize u adipocitima stimuliše sintezu triglicerida. Povećava transport aminokiselina i proteina u mnogim tkivima.

SEKRECIJA INSULINA IZ BETA ĆELIJA PANKREASA.

GLUKAGON
Glukagon luče α-ćelije pankreasa. Jednolančani polipeptid sa 29 ak, Ima ulogu um održavanju stalne koncentracije glukoze u krvi i ANTAGONIST je insulinu. Snažan HIPERGLIKEMIJSKI FAKTOR. U JETRI STIMULIŠE GLUKONEOGENEZU I GLIKOGENOLIZU. Takođe stimuliše oksidaciju masnih kiselina i ketogenezu (sintezu ketonskih tela). Vezuje se za membranske receptore a transmisija signala odvija se preko G-proteina, adenilil ciklaze i cAMP. To dovodi do inicijacije kaskadnih rekacija fosforilacije regulatornih enzima čime se oni aktiviraju ili inhibiraju. Pripremaju organizam za vanredno delovanje. Dejstvo ostvaruju preko dve vrste receptora: α- i β-adrenergičnih receptora. Stimulacija α-receptora dovodi do kontrakcije muskularnih krvnih sudova, dok stimulacija β-receptora ima suprotne efekte. Vezivanjem adrenalina za β-receptore stimulišu se G-proteini, adenili-ciklaza i cAMP.

Mehanizam deovanja glukagona u hepatocitima jetre
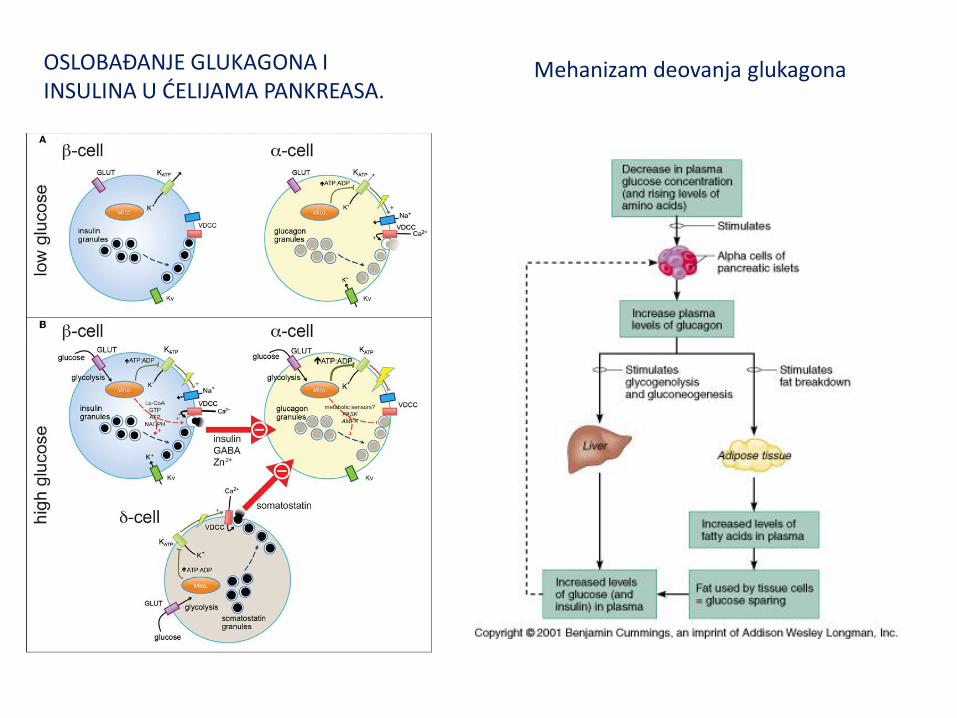
OSLOBAĐANJE GLUKAGONA I INSULINA U ĆELIJAMA PANKREASA.
Mehanizam deovanja glukagona

KATEHOLAMINI
Pripadaju klasi biogenih amina: 1. ADRENALIN (EPINERFIN) 2. NORADRENALIN (NOREPINERFIN) 3. DOPAMIN Adrenalin i noradrenalin se sintetišu u srži nadbubrega (iz Tyr kao prekursora). Razgrađuju se oksidativnom deaminacijom pod dejstvom monoaminooksidaza (MAO) i O-metilacijom delovanjem katehol-O-metiltransferaza (COMT). Delovanje: U mozgu kateholamini deluju kao neurotransmiteri, dok izvan nervnog sistema adrenalin i nor-adrenalin deluju kao regulatori metabolizma. Oslobađaju se iz depoa u srži nadbubrega kao odgovor na stres, napor, hladnoću, i nizak nivo glukoze u krvi. Povećavaju razgradnju triglicerida, pojačavaju rad srca i podižu krvni pritisak.

RECEPTORI ADRENALINA I NORADRENALINA RECEPTORI DOPAMINA

HORMONI TIREOIDNE ŽLEZDE
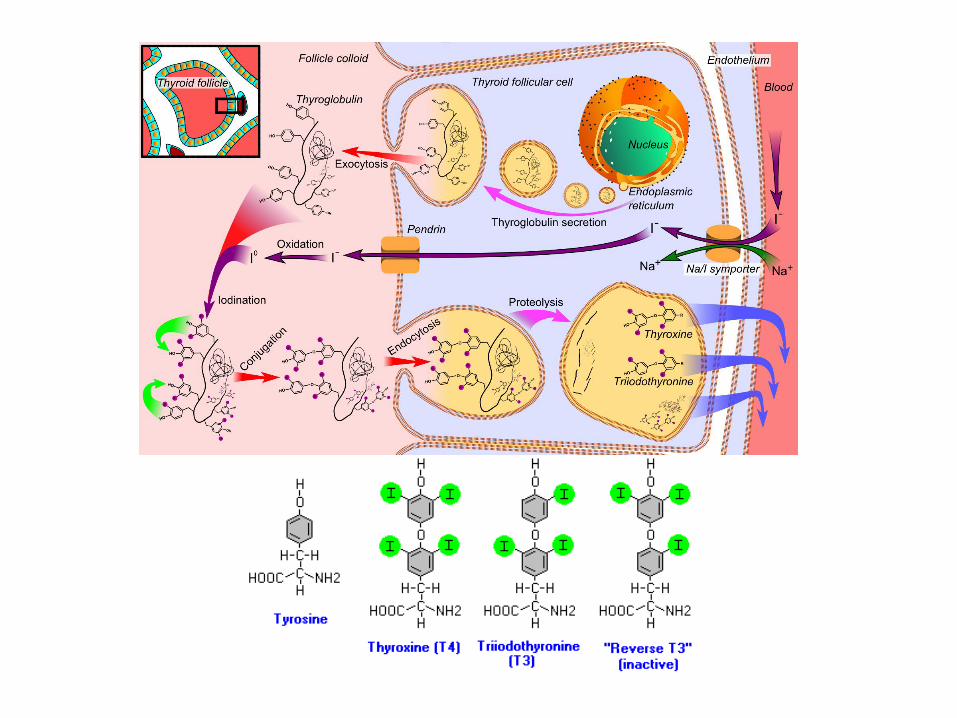
Glavni hormoni tireoidne žlezde su: 1. TIROKSIN (T4) 2. TRIJOD-TIRONIN (T3). Neophodni su za rast i razvoj. Njihov nedostatak u prenatalnom periodu izaziva KRETENIZAM. Glavni si anabolički agensi u toku rasta zajedno sa hormonom rasta. Utiču na povečanje bazalnog metabolizma, stimulacijom Na/K-ATP-aze. Sintetišu se u TIREOCITIMA folikula tireoidne žlezde iz proteina TIREOGLOBULINA koji sadrži brojne tirozil-grupe od kojih jodiranjem nastaju T3 i T4. Tireociti imaju specifičnu sposobnost da usvajaju i oksidišu jone joda, koji se inkorporiraju u tirozil ostatke tireoglobulina. Ovi hormoni se metabolišu dejodinacijom (jetra) i dezaminacijom (bubrezi) i ekskrecijom preko fecesa i urina. Smanjena aktivnost kod odraslih dovodi do pojave MIKSEDEMA kga karakteriše usporeni metabolizam i nagomilavanje mukopolisaharida. Hiperaktivnost dovodi do BAZEDOVLJEVE BOLESTI i TIREOTOKSIKOZE.

PARATIREOIDNI HORMON (PTH) I D-VITAMIN
Paratireoidni hormon i vitamin D imaju ključnu ulogu u metabolizmu kalcijuma i fosfora. U organizmu postoji stalna izmena kalcijuma između plazme i kostiju, creva i bubrega pri čemu je koncentracija kalcijuma Ca2+ jona konstantna (2.25-2.75mM/L). Polovina kalcijuma plazme je u jonskom obliku (Ca2+ jona) , a druga polovina vezana za albumine plazme. Hipersekrecija parathormona (PTH) i trovanje D-vitaminom, kao i neke maligne bolesti dovode do hiperkalcemije, dok nedovoljna količina PTH i vitamina D dovodi do hipokalcemije. Nizak nivo kalcijuma u plazmi dovodi do parestezija (spontanih muskularnih kontrakcija, razdražljivosti i grčeva). Ovi hormoni takođe utiču na nivo fosfata i fosfora (88% se nalazi u kostima). PTH igra važnu ulogu u reapsorpciji fosfata od strane proksimalnih tubula bubrega. PTH HORMON PARATIREOIDEE. Polipetid sadrži 64 aminokiselinska ostatka. Sekrecija je uslovljena koncentracijom kalcijuma u plazmi, i najmanji pad stimuliše lučenje PTH. Deluje preko cAMP na kosti i bubrege: U bubrezima deluje na tri načina: 1. Povećava reapsorpciju kalcijuma i magnezijuma 2. Nephodan je za sintezu kalcitriola (1,25.dihidroksiholekalciferol) 3. Inhibira reapsorpciju fosfata u proksimalnim tubulima Povećava resorpciju kostiju stimulišući osteoklaste i osteocite. Hiperparatireoidizam izaziva hiperkalcemiju. Hipoparatireoidizam dovodi do tretanije zbog
sniženja koncentracije kalcijuma u plazmi.
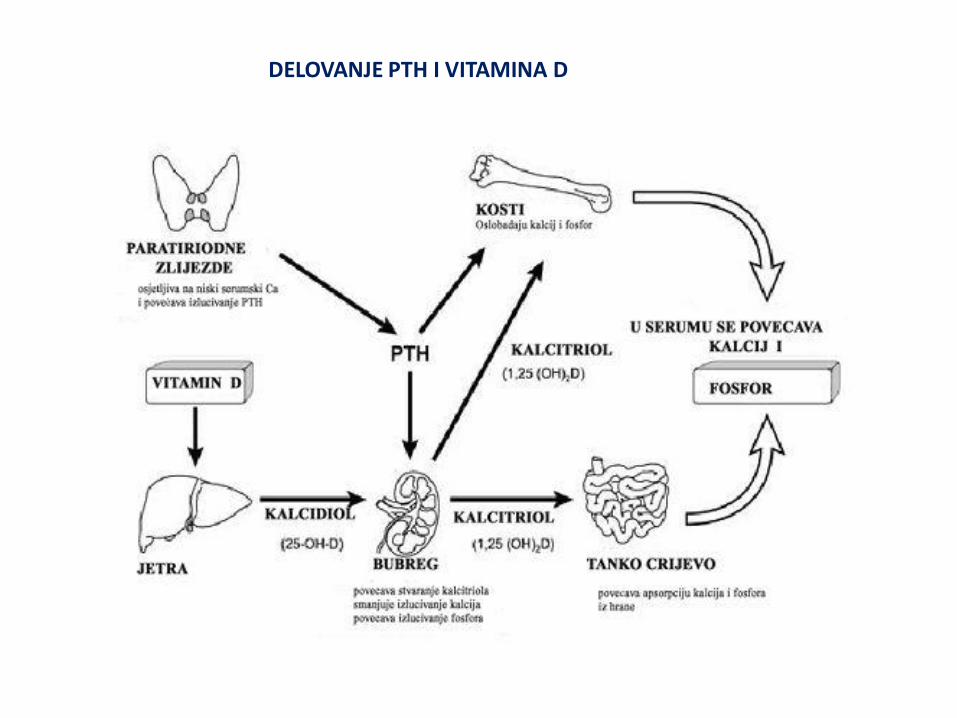
DELOVANJE PTH I VITAMINA D

HORMONI KORE NADBUBREGA-KORTIKOSTEROIDI
Kortikosteroidi se sintetišu putem mevalonske kiseline iz aceti-CoA. Prekursor u njihovoj sintezi je PREGNENOLON iz koga nastaju: KORTIZON, KORTIKOSTERON i ALDOSTERON. Sinteza kortikosdteroida je pod kontrolom hipotalamusa i hipofize koja luči ADRENOKOTRIKOTROPNI HORMON (ACTH). Sinteza se pojačava u stresnim stanjima i naporima. Sintezu ALDOSTERONA reguliše sistem RENIN-ANGIOTENZIN i ACTH. Mehanzam delovanja: Oni lako prolaze kroz plazma membranu i deluju u jedru. Primarnu efekat je EKSPRESIJA GENA i on se ispoljava tek nakon nekoliko sati posle lučenja hormona. U prvom koraku vezuje se za receptor. Kompleks hormon-receptor migrira u jedro i vezuje se za regulatorne sekvence DNA (specifične enhensere). Transport u krvi odvija se pomoću specifičnih proteina_TRANSKORTIN. Prema metaboličkom delovanju dele se na: 1. GLUKOKORTIKOIDE (KORTIZOL I KORTIKOSTERON): DELUJU NA METABOLIZAM
GLUKOZE I PROTEINA. Stimulišu glukoneogenezu. Stimulišu razgradnju proteina u mišićima, limfnim žlezdama i kostima. U limfama deluju imunosupresivno, smanjuju sintezu antitela i deluju antiinflamatorno
2. MINERALOKORTKOIDE (ALDOSTERON): Deluju na retenciju natrijuma i izlučivanje kalijuma u bubrežnim tubulima.


HORMONI HIPOFIZE
Hipofiza kontroliše rad većine endokrinih žlezda i predstavlja dominantni faktor humoralne regulacije organizma. Lućenje hormona hipofize je pod kontrolom HIPOTALAMUSA koji proizvodi RELAISING FAKTORE ili LIBERINE. Na taj način je hipofiza ključni posrednik između humoralnog i nervnog integracionog sistema organizma. Preko mehanizma povratne sprge na lučenje hormona hipofize utiču hormoni drugih emdokrinih žlezda, naročito gonada, tireoidee i kore nadbubrega. HORMONI ADENOHIPOFIZE: 1. ADRENOKORTIKOTROPNI HORMON (ACTH): fiziološki efekti slični glukoortikoidima 2. HORMON RASTA ili SOMATOTROPIN. Podstiče rast celog organizma a hipersekrecija u
detinjstvu dovodi do GIGANTIZMA a nedovoljno lučenje do PATULJASTOG RASTA. Stimuliše sintezu proteina, posbno kolagena. Antagonist insulina. Hipšersekrecija može dovesti i do AGROMEGALIJE (prekomerni rast isturenih delova tela).
3. TIREOTROPNI HORMON (TSH). Stimuliše sintezu i izlučivanje hormona tireoidee 4. GONADOTROPNI HORMONI: FOLIKULOSTIMULIŠUĆI HORMON (FSH): LUTEINIZIRAJUĆI
HORMON (LH) I HORINGONADOTROPINI (HCG). Deluju na razvoj folikula u ovarijumu i testisima, utiču na sekreciju estrogena i progesterona, stimulišu produkciju ovih hormona u trudnoći.
5. PROLAKTIN: dovodi do rasta mlečnih žlezda i stimuliše laktaciju i produkciju mleka kod dojilja.

HIPOTALAMUS KONTROLIŠE RAD MNOGIH ENDOKRINIH ŽLEZDA
HIPOFIZA
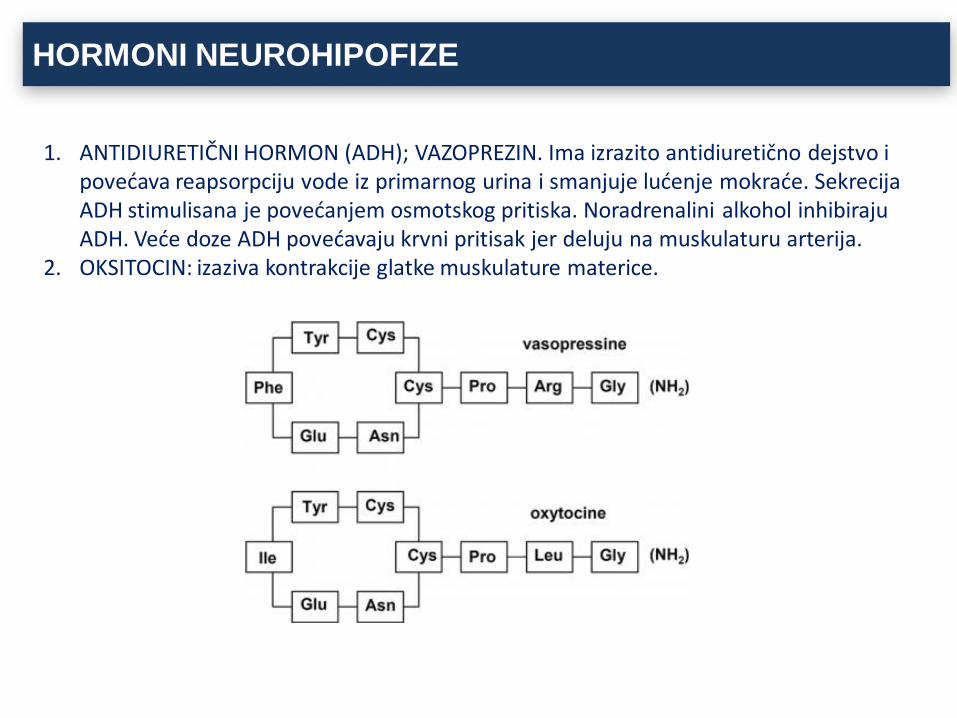
HORMONI NEUROHIPOFIZE
1. ANTIDIURETIČNI HORMON (ADH); VAZOPREZIN. Ima izrazito antidiuretično dejstvo i povećava reapsorpciju vode iz primarnog urina i smanjuje lućenje mokraće. Sekrecija ADH stimulisana je povećanjem osmotskog pritiska. Noradrenalini alkohol inhibiraju ADH. Veće doze ADH povećavaju krvni pritisak jer deluju na muskulaturu arterija.
2. OKSITOCIN: izaziva kontrakcije glatke muskulature materice.

MELATONIN
Melatonin luči PINEALNA ŽLEZDA ili EPIFIZA kao i neki peroferni nervi. Nastaje iz serotonina: Lučenje je pod kontrolom dan-noć. Deluje na melanocite kože žabe (pod njegovim dejstvom postaju svetliji) i po tome je dobio naziv melatonin. Inhibiora dejstvo ACTH i deluje kao fotosenzitivni organ. Fiziološka uloga melatonina povezana je sa regulacijom dnevnog (noćnog) ritma, utiče na spavanje, krvni pritisak, razvoj u pubertetu, adaptaciju na promenu uslova života itd. Značajan je endogeni antioksidant.
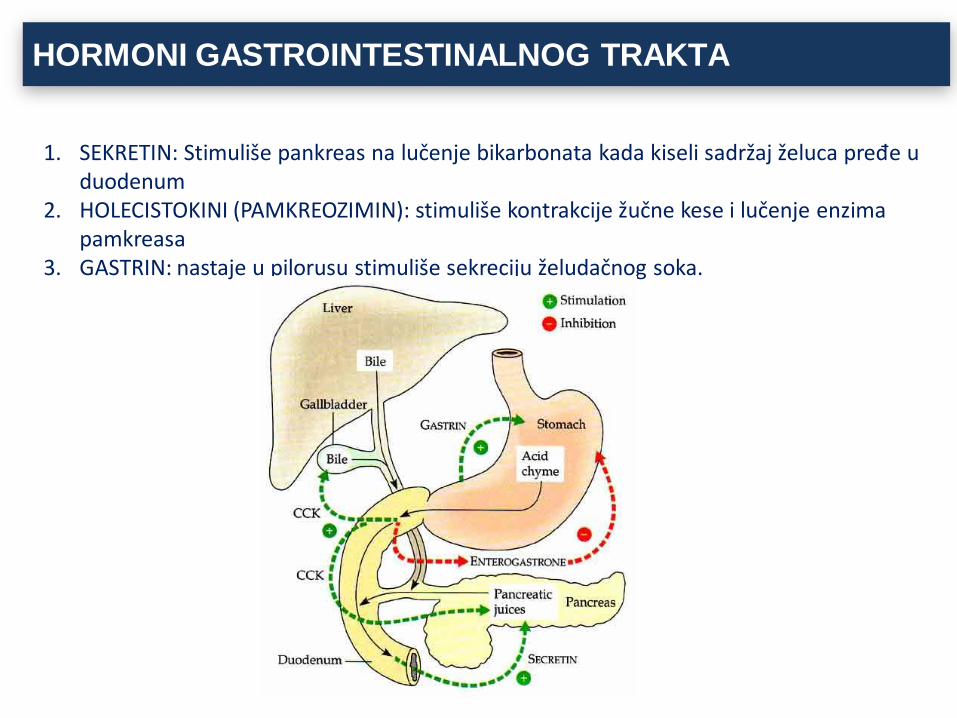
HORMONI GASTROINTESTINALNOG TRAKTA
1. SEKRETIN: Stimuliše pankreas na lučenje bikarbonata kada kiseli sadržaj želuca pređe u duodenum
2. HOLECISTOKINI (PAMKREOZIMIN): stimuliše kontrakcije žučne kese i lučenje enzima pamkreasa
3. GASTRIN: nastaje u pilorusu stimuliše sekreciju želudačnog soka.

PROSATGLANDINI
METABOLITI ARAHIDONSKE KISELINE: 1. PROSTAGLANDIN E2 I F2a
2. TROMBOKSAN A2
3. PROSTACIKLIN Deliju uglavnom lokalno na: promenu krvnog pritiska, sekreciju vode i natrijuma u bubrezima i mišićima, agregaciju trombocita, ekskreciju žučnih kiselina, lučenje insulina. Imaju inflamatorno delovanje. Tromboksan dovodi do vazokonstrikcije koronarnih krvnih sudova i agregacije trombocita. Prostaciklin je koronarni vazodilatator i inhibira agregaciju trombocita.
















![Predavanja - Hormoni [Ppt]](https://img.dokumen.tips/doc/110x75/55117ba94a7959c4028b4698/predavanja-hormoni-ppt.jpg)












