-
7/28/2019 Jurnal Vimentin
1/4
-
7/28/2019 Jurnal Vimentin
2/4
Volume 301, number 2 FEB.5 LETTERS May 1992nucleic acid solution
(690-740 PM) were added to the vimcntin solu-tion. and emission
spectra were measured using cxcitution wav-elenyhta or280 and 295
nm. Assuming that tyrosine is not excited at295 nm, the
contribution of the tyrosine residues to the total Ruorcs-cence
wjascalculated by subtraction of the vimentin emission obtainedat
an excitation wavelength of 295 nm from that obtained at 280
nm,after normalizing the two spccrra above 380 nm (whcrc tyrosine
emis-sion is practically zero [ 141).
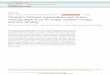
Vimentin fluorescence excitation at 280 nm
Wavelength nm)
Vimentin fluorescence excitation at 295 nm
Wavelength nm)Fig. 1. F!uorescence emission spectra or vimentin
(0.59 PM) withincreasing amounts or pdT (stock 740 PM): I, 0 yl; 2,
8,5~1; 3, 24.5yl; 4. 48.5 ~1; 5, 78.5 ~1; 6. 118.5pl; 7, 189.5 ~1.
(A) Excitationwavelength 280 nm. (B) Excitation wavelength 295 nm.
Spectra arecorrected for dilution, the starting volume is 2,000
~1.
3. RESULTSFig. IA and B show the fluorescence emissionspectra of
vimentin determined after the addition ofincreasing amounts of
poly(dT). The vimentin emissionat an excitation wavelength of 280
nm (A) shows amaximum at 310 nm, suggesting a large contribution
of
tyrosine fluorescence to the total signal. The intensityof the
blue side of the spectrum (290-320 nm) is signif-icantly decreased,
while the red side of the spectrum(360-450 nm) is hardly affected
when poly(dT) is added,Upon excitation with 295 nm light, a peak
with a max-imum at 346 nm due to tryptophan fluorescence is
ob-served, The intensity as well as the maximum emissionwavelength
are practically unaffected upon titrationwith poly(dT). When the
pure tyrosine emission is cal-culated via subtraction of the
tryptophan emission (seesection 2) one observes that the tyrosine
signal isquenched upon addition of poly(dT) (Fig. 2).
Whenrelatively large amounts of poly(dTj are added (Fig.
2),saturation occurs and the tyrosine fluorescence remainsat a
constant level, about 48% of the level in the absenceof
poly(dT).Assuming that the observed tyrosine fluorescencequenching
is caused by binding of the nucleic acid to theprotein, the
stochiometry of binding 11, which is thenumber of nucleotides
covered by each vimentin tetra-mer in the complex, can be
determined from a fluorcs-cence titration curve, in which the
quenching of tyrosinefluorescence is plotted against the ratio of
the addednucleotide to the protein tetramer concentration (Fig.3).
This is accomplished by drawing two straight lines
Calculated vimentin tyrosine fluorescence
Fig. 2. Cdlcuhilcd tyrosine contribution to the Ruoresccnce
emissionatIer excitation at 280 nm (set text). For cxplunation or
the numbersnear the spectra: see Fig. I.178
-
7/28/2019 Jurnal Vimentin
3/4
Volume 301, number 2 FEBS May 1992Titration curve for vimentin
with poly(dT)
81
[poly(dT)] [vimentin]Fig. 3, Fluorescence titration curve for
the binding ofpdT tovimcntin.The Ruorcsccnce due to tyrosine
measured at 307 nni is plotted againstthe ratio of nuclcotides and
pl*otcin ictramcrs.
representing the initial and final slopes of the curve. Theratio
at the intersection of these lines corresponds to n.For the
vimentin-poly(dT) complex, n=50 f 4 is ob-tained. Assuming
Scatchard binding to independentsites, the apporant binding
constant (Q,) can beachieved from the titration curve using the
expression[15]:K,,=W(l-W[&l]&,=apparent binding constant
(M-l), 8=ratio be-tween protein fluorescence change at a particular
ligandconcentration and fluorescence change at a saturatingligand
concentration and [P&total protein concentra-tion (M). Because
this approximation does not correctfor the accumulation of gaps
(which are smaller thanthe binding site size) between stretches of
contiguouslybound vimentin molecules, this binding constant is
a
Table IValues of the nllmber of nucleotidcs covered, 11, he
apparent bindingconstant, f& and the maximum tyrodnc quenching
due to the inter-action of vimcntin tctramers with nucleic
acids
II Max. quenching(%IpoMdT)poly(rA)poWA)
50 t- 4 2.5 ?: 0.5 x IO 48 2 244+4 5.0 2 3.0 x IO 32 ?r.4
-= no measurable tyrosinc fluorescence quenching
minimum estimate only [ 161.However, KllPp s a usefulparameter
to compare binding constants of differentpolynucleotides. For the
binding of vimentin topoly(dT), &,,=2.5 & 0.5 x 10 M-i was
calculated.Similar binding experiments as those reported
forpoly(dT) were also performed for vimentin binding topoly(dA) and
to poly(rA). The parameters n, K,,rPandthe total tyrosine quenching
of poly(dT) and poly(rA)are listed in Table 1. In case of poly(rA),
n is found tobe similar to poly(dT), while KicpP f poly(rA) is
slightlylarger. Interstingly, titration with polp(dA) did not
re-sult in a measurable fluorescence quenching (data notshown).4.
DISCUSSION
The specific quenching of tyrosine fluorescence in thetitration
experiments with poly(dT) and poly(rA) is in-terpreted as an
interaction between tyrosyl rings of theprotein and the
heterocyclic bases of the nucleic acids.Since it has been shown
that the N-terminus of vimentinis essential for its interaction
with nucleic acids [6], it isobvious that some or all tyrosine
residues located in thisregion are involved in this process. The
interactioncould be caused by intercalation of aromatic residues
ofamino acids between nucleotide bases, as has beenshown for other
ssDNA binding proteins such as gene5 protein of fd bacteriophage
and phage T4gp32 [9,17].Crosslinking experiments of vimentin with
tetrani-tromethane (Traub, unpublished results), which
showedprevention from nitration of the tyrosyl residues byrRNA or
fd DNA also indicate a shielding of tyrosinedue to the nucleic
acid. Shape and emission maximumof the tryptophan spectrum remain
co;tstant upon theaddition of saturating amounts of nucleic acids,
indicat-ing that no conformational changes in the tryptophanregion
occur and that the tryptophan residue, which islocated in the
central rod domain [8], does not play arole in the nucleic acid
binding process either.The high affinity for poly(dT) and the
absence ofbinding of poly(dA) are in reasonable agreement withthe
results of investigations via a quantitative filterbinding assay
171. The advantage of the fluorescencetechnique is that only
spec!.ficbinding in which tyrosineresidues are involved, is
detected. Moreover, the bind-ing site size /I and the degree of
fluorescence quenching,which yields information about the binding
process, canbe determined. In this study, it is shown that the
bindingsite size for poly(dT) and poly(rA) is approximately
thesame, suggesting the involvement of the same numberof tyrosine
residues in the binding process. The appar-ent binding constant is
somewhat larger for poly(rA).At saturating amounts of poly(dT),
however, the tyro-sine quenching is significantly larger than at
saturatingconditions of poly(rA). This could be due to two
differ-ent modes of these nucleic acids to the protein. Thestriking
difference in binding properties of poly(rA) and
179
-
7/28/2019 Jurnal Vimentin
4/4
Volume 302. number 2 FEBS LETTERS May 1992poly(dA) suggests that
not only base composition butbackbone properties of the nucleic
acids as well play arole in the process. The physiological meaning
of nu-cleic acid-vimentin interaction, if existent, is poorly
un-derstood. An active role for IF subunit proteins in mod-ulating
DNA replication, recombination, and repair, aswell as in gene
expression, has been postulated [18].Vimentin has been identified
as a DNA attachment prc;-tein within nuclei of Chinese Hamster
Ovary cells [19].However, more experiments are needed to fully
charac-terize the nucleic acid binding properties of vimentinunder
several conditions. The present study, whichshows a strong specific
interaction between vimentinand some types of ssDNA and RNA, may be
helpful inthe search for the (possible) physiological role of
thebinding.A~k,to,~,/~~~~nr~ol~s.e thank the Department of Plant
Physiology ofthe Free University Amsterdam for the use of the
Aminco fluorome-ter. This work was supported by the Netherlands
Oryanizition ofScientific Research (NWO), in part via the
Foundation of Biophysicsand Biology,REFERENCES[I] Stcinert. P.M.
and Roop, D,R. (1988) Annu. Rev. Biochem. 57.593-625.[2]
Quax-Jcukcn, Y.E,F.M.. Qunx, W.J, and Bloemcndal, H. (1962)Proc.
Natl. Acad. Sci. USA BO,3548-3552.
[3] Ip, W., Hartzer, M.K., Pang, S. and Robson, R.M. (1985) J.
Mol.Biol. 183, 365-375,[4] Potschka, M. (1986) Biophys. J. 49,
129-130,[5] Truub. P., Nelson, W.J., Kuhn, S. and Vorgias, C.E.
(1983) J.Biol. Chem. 258. 14561466.[6] Shoeman, R,L.. Wddk?. S.,
Schcrbarth, A. and Traub, P. (1988)J. Biol. Chem. 263,
18744-18749.[7] Shoeman, R,L, and Traub, P. (1990) J. Biol. Chem.
265, 90%9061.[8] Capetanuki. Y,G,, Kuisk. I., Rothblum, K.N. and
Starnes, S.(1990) Oncogenc 5, 645-655.P-J1
[lOI11 I;tsi11411151[I611171iI81
Prutorius, H,T.. Klein. M. and Day, L.A. (1975) J. Biol.
Chcm.250. 9262-9269.Nelson, W-J,. Vorgius. C.E. and Traub, P.
(1982) Biochem. Bio-phys. Rcs. Commun. 106. 1141-l 147.Van
Amerongcn, H,, van Grondclle, R. and van dcr Vlict, PC.(1987)
Biochemistry 26. 4646-4651.Peterson, G.L. (1917)Anal. Biochem. 83.
346-356.Bradford, M.M. (1976) Anal. Biochcm. 72. 248-254.Lakowicz,
J.R. (1983) Principles of Fluorescence Spectroscopy,Plenum Press,
New York.Kelly. R.C., Jcnscn, D.E. and von Hippel, P,N. (1976) 1.
Biol,Chem. 251. 7240-7250.McGhce. J.D. and von Hippel. P.H. (1974)
J. Mol. Biol. 86,469-489.Chase, J.W. and Williams, K.R. (19L6)Annu.
Rev. Biochem. 55,103-136.Truub, P., Plagcns, U,. Kilhn, S. and
Pcridcs, G. (1987) Forlschr,Zool. 34. 175-287.[I91 Cress, A.E. und
Kurath, K.M. (1988) J. Biol. Chcm. 263. 19678-19683.
180