Embed Size (px)
Citation preview

Vimentin fibers orient traction stressNancy Costigliolaa,b, Liya Dinga,c, Christoph J. Burckhardta,c, Sangyoon J. Hana,c, Edgar Gutierrezd, Andressa Motaa,Alex Groismand, Timothy J. Mitchisonb, and Gaudenz Danusera,c,1
aDepartment of Cell Biology, Harvard Medical School, Boston, MA 02115; bDepartment of Systems Biology, Harvard Medical School, Boston, MA 02115;cDepartment of Cell Biology, University of Texas Southwestern Medical Center at Dallas, Dallas, TX 75390; and dDepartment of Physics, University ofCalifornia, San Diego, La Jolla, CA 92093
Edited by Jennifer Lippincott-Schwartz, Howard Hughes Medical Institute, Janelia Research Campus, Ashburn, VA, and approved March 16, 2017 (received forreview September 1, 2016)
The intermediate filament vimentin is required for cells to transitionfrom the epithelial state to the mesenchymal state and migrate assingle cells; however, little is known about the specific role ofvimentin in the regulation of mesenchymal migration. Vimentin isknown to have a significantly greater ability to resist stress withoutbreaking in vitro compared with actin or microtubules, and also toincrease cell elasticity in vivo. Therefore, we hypothesized that thepresence of vimentin could support the anisotropic mechanicalstrain of single-cell migration. To study this, we fluorescentlylabeled vimentin with an mEmerald tag using TALEN genomeediting. We observed vimentin architecture in migrating humanforeskin fibroblasts and found that network organization variedfrom long, linear bundles, or “fibers,” to shorter fragments with amesh-like organization. We developed image analysis tools employ-ing steerable filtering and iterative graph matching to characterizethe fibers embedded in the surrounding mesh. Vimentin fibers werealigned with fibroblast branching and migration direction. The pres-ence of the vimentin network was correlated with 10-fold slowerlocal actin retrograde flow rates, as well as spatial homogenizationof actin-based forces transmitted to the substrate. Vimentin fiberscoaligned with and were required for the anisotropic orientation oftraction stresses. These results indicate that the vimentin networkacts as a load-bearing superstructure capable of integrating andreorienting actin-based forces. We propose that vimentin’s role incell motility is to govern the alignment of traction stresses thatpermit single-cell migration.
vimentin | fiber orientation | traction stress | intermediate filaments |mesenchymal migration
Vimentin is a marker for cancer metastasis and is required forsingle-cell mesenchymal migration (1). Vimentin is known to
interact with both actin and microtubules (MTs) (2, 3), two cy-toskeleton polymers that have been well studied in the context ofmigration. Actin assembly and contraction provide the propulsiveand traction forces required for forward translocation of cells (4–6), while at the same time, MTs determine cell orientation (7).Vimentin has been proposed to structurally integrate these net-works (8, 9); however, how this occurs in the process of cell mi-gration when the interacting cytoskeletons are under permanentturnover and reorganization is unknown, owing mainly to the lackof appropriate live cell imaging studies and image analysis toolsfor inferring underlying mechanisms.Previous work demonstrated that injection of dominant-negative
nonpolymeric vimentin into a fibroblast resulted in local lamellipodformation (10), a region of rapid actin flow. Recently, vimentinexpression was shown to affect the position of the lamellar arc, andalso to be dependent on lamellar actin-binding proteins for distri-bution (11). Moreover, our recent study in epithelial cells un-dergoing an epithelial-to-mesenchymal transition (EMT) found thatvimentin tightly integrated with MTs in a positive feedback loop, inwhich MTs support the assembly of a vimentin network, which inturn templates MT growth to confer persistence in cell polarity (12).Taken together, the foregoing studies suggest that vimentin’s abilityto interact with both MTs and actin could reinforce cell polarity anddirected migration.
Similar to other intermediate filaments, vimentin structure isessential for the mechanical integrity of cells and tissues. In vitrorheology measurements have demonstrated that in contrast toMTs and actin, the vimentin network is both easily deformableand able to withstand high strains without breaking (13). Vimentinalso increases cytoplasmic elasticity in vivo (14). Interestingly,whereas MTs and actin polymerize unidirectionally, vimentin fil-aments polymerize both longitudinally and laterally without po-larity (15). This capacity for variable spatial regulation of vimentindensity suggests that the architecture of the network could be akey determinant of its function.To test this hypothesis, we sought to monitor the structure of
the vimentin network in a native mesenchymal environment. Weused transcription activator-like effector nuclease (TALEN)-basedgenome modification to express fluorescent-labeled vimentin innonimmortalized human foreskin fibroblasts (hFFs). We foundthat the vimentin network exhibited both longer continuous re-gions, resembling fibers, as well as shorter, fragmented, mesh-likeregions. We have developed image analysis tools to perform ori-entation analysis on the fibers, and found that these fibers werealigned with fibroblast branching and migration direction. Thevimentin network also was spatially correlated with the slowestactin retrograde flow, and regulated the homogeneity of tractionforce. Finally, we found that vimentin fibers were required fortraction stress orientation. Individual fibers were locally alignedwith traction stresses, and vimentin level determined the degree ofanisotropy in traction stress orientation. These results indicate thatthe presence of vimentin governs the orientation of traction forcesin mesenchymal cells.
Significance
Vimentin is a marker for the epithelial-to-mesenchymal transi-tion, which is thought to lead to cancer metastasis. Withoutvimentin, planar cells exhibit polarity defects and cannotmigrateas single cells. The mechanisms by which vimentin influences cellmigration remain mostly unknown, however. Generally, in-termediate filaments regulate the mechanical integrity of cellsand tissues; thus, we hypothesized that vimentin could be in-volved in supporting the directionally aligned force transmissionrequired for single-cell migration. We have developed imagingand analysis methods to probe the role of vimentin in mesen-chymal cell migration that will have broad utility in intermediatefilament research. In summary, our results demonstrate thatvimentin governs the alignment of the cell traction forcesneeded for directed single-cell mesenchymal migration.
Author contributions: N.C. and G.D. designed research; N.C. performed research; N.C.,C.J.B., E.G., A.M., and A.G. contributed new reagents; C.J.B. genome edited and charac-terized cell lines; L.D. contributed new algorithms; N.C., L.D., and S.J.H. analyzed data;and N.C., T.J.M., and G.D. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1614610114/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1614610114 PNAS | May 16, 2017 | vol. 114 | no. 20 | 5195–5200
CELL
BIOLO
GY
Dow
nloa
ded
by g
uest
on
Apr
il 15
, 202
0

ResultsIdentification of Vimentin Fibers. We sought to accurately charac-terize the structure of the vimentin network in living cells whileminimizing as much as possible perturbation owing to the over-expression and tagging of vimentin. We introduced the mEmeraldgene at the N terminus of the endogenous vimentin locus in non-immortalized human foreskin fibroblasts (hFFs) using TALENs (12,16) (Methods). The fraction of mEmerald-vimentin was 4 ± 4% ofthe total vimentin in edited cells (Fig. S1A). TALEN-edited hFFsmigrated at the same speed as control cells (Fig. S1B), and no dif-ferences in morphology or vimentin distribution were detected be-tween wild-type (WT) and TALEN-edited hFFs (Fig. S2).We then imaged the mEmerald-vimentin network organization
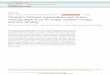
in hFFs at high resolution using spinning disk confocal microscopy(Fig. 1A). We observed that polymer density and organizationvaried from short, mesh-like randomly oriented fragments tolonger, linear bundles (Fig. 1B, left to right). To assess this com-plex organization, we developed image analysis tools to distinguishthe more bundled, fibrous regions from the remaining networkmesh. In a first step, we used a previously published algorithm (12)to identify the structure of the network. In brief, from the rawimage (Fig. 1C), we thresholded the background and applied a
multiscale steerable filter (17) to detect curvilinear features (12)(Methods). The resulting filter response map (Fig. 1D) describedthe confidence for each pixel in belonging to the vimentin net-work. We next applied nonmaximum suppression (12, 18) toskeletonize the vimentin network. This was followed by a graph-based connection algorithm that assembled, based on their prox-imity and end-to-end orientation, curvilinear skeleton segments intoa complete network (12), the result of which is a map of vimentinfragments of various lengths (Fig. 1E).In a second step, we distinguished the longer, bundled fragments
from the shorter fragments. To establish a length threshold forvimentin fragments, we sought to subtly perturb the vimentin net-work organization and classify resistant structures as fibers. No spe-cific reagent against vimentin network assembly has been identified;however, it is well established that vimentin assembly requires intactsurrounding actin and MT cytoskeletons (2, 9). Thus, we hypothe-sized that acute (5 min) disruption of filamentous actin or MTswould affect the finer components of the vimentin network as well(Fig. S3 A–C). Indeed, after actin depolymerization by cytochalasinD, we observed a shift in the vimentin network organization toward amore fibrous state (Fig. S3 A and B). When we applied a lengththreshold of 4 μm, which matches the length scale of actin stress fi-bers, the ratio of fiber to remaining network increased twofold for
fiber and mesh overlay
detected
fiber
0 180˚
fibrousfibrous/meshmesh
fragment mapraw image
fiber orientation
steerable filter response
A B
C D E
F G H
Fig. 1. Characterization of the vimentin network by computational image processing. (A) Z-stack maximum intensity projection of mEmerald-vimentindistribution within an hFF cell. (B) Vimentin mesh (Left and Middle images) and fibrous architecture (Middle and Right images). All images are z-stackmaximum intensity projections of mEmerald-vimentin in three representative cells. (Insets) 2× zoom of the areas of detail. (C) Raw spinning disk confocalimage of mEmerald-vimentin in an hFF cell. (D) Multiscale steerable filtering was applied to the image in C to enhance curvilinear features. (E–G) Extraction ofvimentin fibers. (E) Nonmaximum suppression was applied to D, followed by iterative graph matching of skeletonized curvilinear features in close proximityand orientation to extract a map of network fragments of various lengths. (F) Fragments from E meeting an experimentally determined length threshold(4 μm) were classified as vimentin fibers (magenta). All high- confidence pixels from D not classified as fibers were classified as mesh (gray). (G) Inset ofF showing the raw image (Top) and analyzed pixel map (Bottom) of fibrous and mesh-like vimentin. (H) Orientation of vimentin fibers in the image frame ofreference. (Scale bar: 10 μm.)
5196 | www.pnas.org/cgi/doi/10.1073/pnas.1614610114 Costigliola et al.
Dow
nloa
ded
by g
uest
on
Apr
il 15
, 202
0

this condition relative to the untreated condition (Fig. S3D),reflecting what we observed. Thus, we applied a 4-μm threshold forfiber identification in all analyses (Fig. 1 F and G, magenta pixels).All high-confidence pixels from the steerable filter map not classifiedas fibers (i.e., fragments below the fiber length threshold, as well asthose so short and with an isotropic orientation that they wereeliminated by the nonmaximum suppression and graph-based con-nection algorithms) were classified as mesh (Fig. 1 F and G, graypixels). For the pixels belonging to fibers, we also determined thelocal fiber orientation (12) (Fig. 1H).We tested the ability of our orientation algorithms to measure
changes in detected vimentin fibers in response to partial MTdepolymerization, which we previously described as disruptingvimentin network polarity (12). Indeed, we found that MT dis-ruption reduced the coalignment between the vimentin fiberswithin a cell (Fig. S3 A, C, and E), demonstrating that the fibersare not organized randomly.
Vimentin Network Geometry in Migrating Fibroblasts. We next ap-plied our imaging and analysis tools to measure vimentin networkorganization in migrating fibroblasts. During random migration,fibroblasts exhibit multiple protrusion branches (Movie S1). Ex-amination of individual branch retraction events showed that thevimentin fibers apparently remained intact, folding inward withslight compaction during cell retraction (Movie S2). Although wecould not track individual fibers, we estimated the stability of in-dividual fibers by tracking the fiber/mesh ratio over time. Al-though the amount of fibers varied among cells (Fig. S4A), thefiber/mesh ratio remained fairly constant for an individual cellover the migration period (Fig. S4 B and C).We then asked how the fibers were oriented in relation to cell
shape and the direction of movement (Fig. 2A). To this end, wefirst segmented the cell footprint and applied skeletonization tothe resulting mask (Methods and Fig. 2B, Left). We then de-termined the number and orientation of skeletonized branches ina cell within a given frame. We also evaluated displacement of thecell by tracking the position of the center of the mask from frameto frame (Fig. 2B, Right). Vectors connecting the mask centers in
consecutive frames defined the instantaneous direction of cellmovement (Methods). We found that vimentin fibers aligned withfibroblast branches (Fig. 2C, Left and Movie S3), whereas the meanfiber orientation tended to align with the direction of cell movement(Fig. 2C, Right). These results show that on the time scale of cellmigration, the vimentin network is composed of apparently stablefibers embedded in a surrounding mesh, and these fibers align withfibroblast protrusion and the overall direction of cell movement.
Vimentin Network Position Relative to Actin Flow Speed. We nextsought to test our hypothesis that vimentin architecture couldregulate force transmission during single-cell migration. To dothis, we first measured interaction of the vimentin network withactin flow, which results from actomyosin contraction as the primaryforce generator during planar mesenchymal cell migration (19, 20).To observe colocalization between vimentin and actin, we tran-siently expressed an actin SNAP-tag and labeled it with SNAP-CellTMR (New England BioLabs) in hFFs expressing mEmerald-vimentin (Fig. 3A). Actin flow speed was measured using particleimage velocimetry methods (21, 22). We then compared in-stantaneous actin flow speeds in regions containing no vimentinwith those in regions containing vimentin mesh or fibers (Fig. 3B–D). Compared with actin flow in regions without vimentin, actinflow speeds were significantly reduced when colocalized withvimentin mesh (Fig. 3D), and even more reduced when colocalizedwith vimentin fibers (Fig. 3D and Movies S4 and S5). The re-lationship between actin flow speed and vimentin did not dependon the relative position within the cell; actin flows across the cellperiphery and cell interior were fastest in the absence of vimentinand slowest when colocalized with vimentin fibers (Fig. S5A).However, we found no significant correlation between the directionof actin flow and the orientation of vimentin fibers (Fig. S5B).
Vimentin Network Effects on Traction Force Magnitude. Given thespatial correlation between the presence of vimentin and re-duced actin flow speed, we hypothesized that the elastic andrelatively stable vimentin network could absorb and redistributeforces generated by actomyosin contractility. To assess forcetransmission in the presence of vimentin, we measured thetraction force distribution on 8.5-kPa bead-coated silicone gelsubstrates (23) in TALEN-modified hFFs expressing either
t=370 m
0
18
0˚
t=500 mt=0 m t=160m
Pe
rce
nta
ge
Fib
ers
(%
)
Fiber orientation
rel. branch orientation
0.20
0
0.10
90˚
0.20
0
0.10
90˚Mean fiber orientation
rel. cell movement
Θ
B C
A
Fig. 2. Vimentin network geometry during mesenchymal migration.(A) Vimentin fiber orientation inmigrating hFFs. The arrow points to the same cellover time. Note that the image contrast of this time lapse was adjusted to moreclearly visualize peripheral vimentin fibers (Methods). (B) hFFs expressing mEmerald-vimentinwere stainedwith a 642-nmemittingmembrane dye tomeasure cell shapeand position. The outline of cells was masked and then skeletonized (Methods). Thecell position was determined by the center of the cell mask. The orientation of cellmovement was tracked from frame to frame. (C, Left) The angles between thebranches of the skeletonized cell shape and vimentin fibers in the same region ofthe cell were compared for each branch in each frame. Shown are all individualmeasurements over all time-lapse sequences (8-h time lapse; n = 60 cells). Data aremean values from four separate experiments. The measured distribution deviatesfrom a uniform distribution with P < 10−5 (Kolmogorov–Smirnov test). (C, Right) Themean vimentin fiber orientation angle within a cell was also compared with theorientation of whole cell movement. The measured distribution deviates from auniform distribution with P < 10−5 (Kolmogorov–Smirnov test). (Scale bar: 10 μm.)
A
D
B C
Fig. 3. Vimentin network relative to actin flow speed. TALEN-modified hFFsexpressing mEmerald-vimentin and transiently expressing actin SNAP-tagwere incubated with SNAP-TMR. Both wavelengths were imaged every 5 s.Actin behavior was analyzed using qFSM software (22). (A) SNAP-TMR actindistribution in hFFs. Zoom-in of inset is the raw vimentin image overlaid withthe cell outline. (Scale bars: 5 μm.) (B) Detected vimentin fibers overlaid withactin flow vectors (white). (C) Zoom-in of boxed regions in B. Vimentin fibersare shown in red. Each white pixel represents the position of an identifiedactin speckle. Vector length represents the time interval over which a specklewas tracked. (D) Actin flow speed vs. colocalized vimentin polymer organiza-tion (none, mesh, fiber) for n = 7 cells. Shown from left to right: no vimentin,M = 5,046 actin flow tracks; mesh, M = 14,130 flow tracks; and filamentousvimentin, M = 418 flow tracks.
Costigliola et al. PNAS | May 16, 2017 | vol. 114 | no. 20 | 5197
CELL
BIOLO
GY
Dow
nloa
ded
by g
uest
on
Apr
il 15
, 202
0

control or vimentin knockdown lentivirus. Traction force distri-butions appeared to be more homogeneous with control cells thanwith knockdown cells (Fig. 4A andB). Measurements of traction forcemaxima in the cell center indicated that with control cells, tractionforces were threefold more homogeneously distributed than withknockdown cells, whereas no difference was observed at the cell pe-riphery, which tends to be a vimentin-depleted region (Fig. 4C). Withboth control and knockdown cells, vimentin position was anti-correlated with traction force maxima (Fig. 4D andE). Based on thesedata, we concluded that the presence and position of vimentin result insignificantly more homogeneous force transmission to the substrate.
Vimentin Network Effects on Traction Stress Orientation. For di-rected migration, mesenchymal cells must regulate not only themagnitude of transmitted forces, but also their directionality. Wefound that vimentin fibers aligned along the direction of fibroblastprotrusion and migration (Fig. 2), and thus asked whether thevimentin network itself affected the directionality of tractionstresses. We first compared vimentin fiber orientation with that oftraction stresses (Fig. 5 A and B). In contrast to the absence ofalignment between vimentin fibers and actin flow, we found a highdegree of alignment between the orientation of cell traction stressesand that of local fibrous vimentin (Fig. 5C). We then checkedwhether knockdown of vimentin altered local traction stress orien-tation. To this end, we measured the SD of traction stress orien-tation within 2.5-μm-radius windows throughout the cell. We foundthat knockdown cells had nearly randomly oriented traction stressesrelative to control cells (Fig. 5D; see Methods for a definition of thealignment index). This result demonstrates that the presence ofvimentin is required for traction stress alignment.Our results show that the vimentin network colocalizes with regions
of slowest actin flow, redistributes local actin network-based forces, andis required for traction stress alignment.We conclude that the vimentinnetwork acts as a load-bearing superstructure capable of redistributingand determining the orientation of actomyosoin-generated forces (Fig.5E). In the absence of vimentin, actomyosin-generated forces arewholly transmitted to the substrate via discrete adhesions throughoutthe cell–substrate interface; however, in the presence of vimentin fi-bers, actomyosin-generated forces are partially absorbed by the inter-acting vimentin superstructure and redirected to adhesions in theperipheral band low in vimentin concentration. As a result, the tractionforce field is aligned with the colocalized vimentin fibers.
DiscussionEMT and subsequent single-cell migration are marked by theexpression of vimentin. The principal manner in which intermediatefilaments have been shown to influence cell function is by providingstructural integrity to cells; however, how or why vimentin is requiredfor normal mesenchymal migration has remained unclear. Here, weprovide evidence that the structural organization of the vimentinnetwork governs the alignment of traction stresses required forsingle-cell migration.We found that vimentin fibers co-oriented with fibroblast
branching, as well as the overall direction of cell movement (Fig. 2).This is similar to the alignment of MTs with the direction of mi-gration (7), suggesting some convergence of function between thesetwo cytoskeleton components. Recently we showed that vimentinand MT orientations reinforced one another, as well as overall cellpolarity, during migration (12). Thus, we hypothesized that becauseof vimentin’s known interaction with both MTs and actin, vimentincould function to integrate the polarizing effect of MTs withactomyosin-based forces that drive planar mesenchymal migration.Indeed, we measured significantly slower actin flow in the
presence of vimentin, particularly vimentin fibers, indicating
max
min
A
E
20
No
. pe
rip
he
ral
ma
xim
a
0 0ctl kd
40
ctl
**
BTr
act
ion
(P
a)
F 1000
3002500
0
Vim. Intensity
(control)
0
1000
Vim. Intensity
(knock down)
Inte
rior
Pe
riph
ery
cou
nt
1000
0
0
1000
C
kd
II
III
No
. in
teri
or
ma
xim
a
D
Fig. 4. Vimentin network effects on traction forces. (A, Left) Pseudocolorimage of traction force exerted by an hFF expressing control lentivirus. (A,Right) Traction force exerted by an hFF expressing shRNA targeting vimen-tin. Traction force range (minimum to maximum) was adjusted to display thedistribution effectively. In the left panel, the range corresponds to 0–0.8 kPa;in the right panel, the range corresponds to 0–1.0 kPa. (Scale bar: 5 μm.)(C, Left) Mean number per cell of traction force peaks in the interior(area >7 μm from the cell edge; Methods) of control and vimentin kd cells.**P = 0.0115, t test. (C, Right) Mean number of traction peaks in the 7-μm-wide peripheral band. Control cells, n = 20; kd cells, n = 19. Error bars rep-resent SEM. (D and E) Zoom-in views of boxes indicated in A, showingvimentin distribution (Left) and traction force distribution (Right). (D) I; (E) II.
(F) Interior (Top) and peripheral (Bottom) traction stress vs. colocalizedvimentin intensity for representative control (Left) and kd (Right) cells. Thecounts of traction stresses for given magnitude and vimentin intensity arepresented on a pseudocolor scale, with a range of 0–3000.
5198 | www.pnas.org/cgi/doi/10.1073/pnas.1614610114 Costigliola et al.
Dow
nloa
ded
by g
uest
on
Apr
il 15
, 202
0

interaction between these two cytoskeleton systems in fibro-blasts. Although it is difficult to determine the cause and effectof protein colocalization, recent work (11) demonstrated thatvimentin and actomyosin coregulate their distributions throughactin-vimentin linkers, such as plectin (8). Vimentin and actin net-works may potentially interact via bulk effects as well; in fact,vimentin has been shown to regulate actin network stiffness byproviding steric constraints in vitro (24, 25).We next investigated whether vimentin, as an elastic network
colocalizing with slower actin flow, affected the distribution of forcesexerted by the actin cytoskeleton on the substrate. Actin retrogradeflow and myosin contractility on the actin cytoskeleton are known toexert stress on the extracellular substrate via discrete adhesions (26).Although it was previously reported that traction stresses in regionsof slower actin flow are more homogenously distributed (and in-dependent of adhesion size) (27), why this is so was not clear. Weused traction force microscopy to map the points in the cell whereintracellularly generated forces are coupled to the extracellularsubstrate. We found that the highest traction stresses occurredoutside the footprint of the vimentin network, whereas forces insidethe footprint were spatially more homogeneous. These results leadto the proposal that vimentin serves as a cell-internal load-bearingsuperstructure that redistributes local actin-generated forces.One prediction of the vimentin superstructure model is that an-
isotropic architecture, such as fiber formation, would set the ori-entation of the traction stresses and through it the direction of cellmigration. To test this prediction, we analyzed the spatial correla-tion of the direction of vimentin fibers and traction stresses andindeed found traction stresses to be aligned with vimentin fibers(Fig. 5). Moreover, the local coalignment of traction stresses wasdependent on vimentin expression. Thus, our data and analysestogether identify a role for the vimentin network as a secondarysuperstructure within the cell that reorients actin-based forces and isessential for traction stress alignment. In view of the mutual struc-tural guidance between vimentin and MTs (12), it will be interestingin future studies to examine vimentin’s role as a potential mediatorbetween MT organization and traction force transmission.
MethodsTALEN Assembly. The N-terminal addition of EGFP or mEmerald to vimentinhas been shown to yield functional fusion proteins (10, 28). To stably expressmEmerald-vimentin, we used a TALEN-based genome editing approach. Thetarget site selection and the assembly of these TALENs have been describedearlier (12, 29).
Cells. The hFFs were obtained from American Type Culture Collection andgrown in high-glucose DMEM containing 25 mM D-glucose, 4 mM L-gluta-mine, and 110 mg/mL sodium pyruvate (Gibco, Life Technologies) supple-mented with 10% FBS (Gibco), 15 mM Hepes (Gibco), and penicillin/streptomycin (Gibco). For Western blot analysis, mouse anti-vimentin (cloneV9; Sigma-Aldrich), rabbit anti-vimentin against a C-terminal peptide (C-20;Santa Cruz Biotechnology), and mouse and rabbit anti-GAPDH (GT239 andGTX100118, respectively; GeneTex) antibodies were used. For immunofluo-rescence, chicken polyclonal to vimentin ab24525 and rabbit monoclonal[EP1332Y] to α-tubulin (Abcam) were used.
Imaging. All images were collected with a Yokogawa CSU-X1 spinning diskconfocal system on a Nikon Ti inverted microscope equipped with a PerfectFocus System for continuousmaintenance of focus. Cells were fixed for 15minat 37 °C with 1% glutaraldehyde and then permeabilized with 0.5% TritonX-100 for 5 min. Actin was labeled with Alexa Fluor 647 phalloidin (Invi-trogen). For migration assays, six-well 1.5 MatTek dishes coated with 10 μg/mLfibronectin were used for coimaging of control shRNA and vimentin shRNAhFF lines. Cells were plated at 4 h before imaging and then incubated for5 min at 37 °C with a CellMask Deep Red plasma membrane stain (ThermoFisher Scientific) at 1 h before imaging. mEmerald-vimentin was excitedwith a 488-nm, 100-mW solid-state laser with an ET525/50m emission filter(Chroma Technology). The Deep Red membrane stain was excited with a642-nm 100-mW laser with an ET700/75m emission filter (Chroma Tech-nology). Images were collected every 5 min, using an exposure time of 50 msfor mEmerald-vimentin and 200 ms for the Deep Red membrane stain. Imageswere collected with a Nikon Apo 40x 0.95 dry objective lens.
Image Representation. For fine vimentin structures to be visible in the figurepanels, we adjusted the contrast of raw vimentin images using ImageJsoftware. Raw images first underwent intensity inversion followed by
Tra
ctio
n v
ect
or
cou
nt
x 1
0^
4
A CB
Angle of stress vector 90˚
Tra
ctio
n s
tre
ss v
ect
ors
D
E
Loca
l alig
nm
en
t o
f
tra
ctio
n v
ect
ors 0.6
0.4
0.2
0
Ctl(vim
- sp
ace)
Ctl(vim
+ sp
ace)
Θ
vim k
d(vim
+space
)
vim k
d(vim
-space
)
Vimentin FVimentin F
Actomyosin FActomyosin F
Adhesion FAdhesion F
Network based
force redistribution
Fig. 5. Effect of vimentin fibers on traction stress orientation. (A, Top) Segmented vimentin fibers color-coded for orientation and traction stress vectororientation for control cell shown in Fig. 4A. (A, Bottom) Zoom-in of box indicated in the top panel. (Scale bars: 5 μm.) (B and C) Illustration (B) and mea-surement of (C) angular differences between vimentin fibers and colocalized traction force vectors for n = 12 cells. The measured distribution deviates from auniform distribution with P < 10−5 (Kolmogorov–Smirnov test). (D) Local alignment of traction stress vectors within a circular window of 2.5-μm radiusmeasured in control and vimentin kd cells in regions with and without detectible vimentin. Control cells, n = 12; kd cells, n = 9. (E) Model of the vimentinnetwork as a load-bearing superstructure that redirects actomyosin forces to the peripheral adhesions and aligns the traction force field with the orientationof the vimentin fibers.
Costigliola et al. PNAS | May 16, 2017 | vol. 114 | no. 20 | 5199
CELL
BIOLO
GY
Dow
nloa
ded
by g
uest
on
Apr
il 15
, 202
0

contrast adjustment, such that local structure could be visualized without ob-scuring any surrounding features in the region shown. For time-lapse images suchas those that appear in Fig. 2, the same contrast settings were appliedthroughout.
shRNA Constructs and Reagents. Human vimentin expression was inhibitedusing the target sequence 5′ACGTACGTCAGCAATATGA-3′. The control shRNAexpression vector contained a 5′-ATGTACTGCGCGTGGAGA-3′ sequence. Theloop sequence between the sense and antisense sequences was 5′-TTCAA-GAGA-3′. These sequences were cloned into pSilencer 5.1 H1-retroviral vector(Life Technologies) in accordance with the manufacturer’s instructions.
Traction Force Imaging and Analysis. Substrates were prepared as describedpreviously (23). Dishes were coated with 10 μg/mL fibronectin and 100 μg/mL1-ethyl-3-(3-dimethylaminopropyl) carbodimide in PBS for 1 h at 37 °C. Cellswere seeded at 4 h before imaging and incubated with Deep Red plasmamembrane stain. Red-labeled bead positions were imaged using a 561-nm,200-mW laser with an ET620/60m emission filter with an exposure time of300 ms. Beads were imaged before and after cell removal via 15 min of 2.5%Trypsin at 37 °C. Images were collected with a Nikon Plan Apo VC 1.4 NA oilimmersion objective lens. Traction force data were analyzed as describedpreviously (30).
We observed a difference in interior traction force distributions betweencontrol and vimentin kd cells. To quantify these differences, we segmentedislands with high traction by thresholding the image with a combination ofOtsu (31) and Rosin (32) thresholding. Specifically, given the threshold valuesdetermined by the Otsu algorithm TO (in which the underlying assumption isa bimodal traction distribution) and by the Rosin algorithm TR (in which theunderlying assumption is a unimodal traction distribution with high-forceoutliers), an empirical threshold level reflecting the mixture of these theo-retical distributions was computed as 2/3 TO + 1/3 TR. We considered tractionislands with an area >0.2 μm2. To quantify the level of spatial traction ho-mogeneity, we counted the number of high-traction islands in either a pe-ripheral band 7 μm from the cell edge or the cell interior.
For traction stress alignment, the traction stress isotropywas defined as thecircular SD (S) of traction stress orientation within a window with a radius of2.5 μm. The local “alignment” of the traction stresses was defined as thecomplement of the mean value of S for a given window (SD of uniformorientation distribution - jSj). The distribution shown is for all windows inall cells.
Actin Flow Imaging and Analysis. Cells were transfected with SNAP-tag actinusing the Neon Transfection System (Invitrogen) at 24 h before the
experiment. At 1.5 h before seeding, cells were incubatedwith SNAP-tag TMRligand (New England BioLabs) following the manufacturer’s protocol. At 1 hbefore imaging, DMEM imaging medium containing 10% FBS, 1% oxyfluor,10 μM DL-lactate, 15 mM Hepes, and no phenol red was added to the cells. Alayer of mineral oil was also added to prevent evaporation. Images werecollected with a Nikon Plan Apo 60× objective. Actin flow data were ana-lyzed as described previously (21, 22).
Vimentin Network Structure Analysis. We observed that vimentin polymerdensity and organization varied from short mesh-like, randomly orientedfragments to longer, linear bundles. To assess this complex organization, wefirst used a previously published algorithm designed to extract filamentousfeatures (12). In brief, we used a steerable filter (17) to enhance the raw imagedata curvilinear features and then applied a nonmaximum suppression operatorthat accounts for the local orientation of a putative curvilinear feature. Com-bined, these two procedures generated a map of 1-pixel-wide vimentin frag-ments. We next clustered fragments likely belonging to the same fiber based onco-orientation and spatial proximity using an iterative graph-matching algo-rithm. The graph matching includes the rescue of low-confidence fragments thatare structurally well linked to high-confidence fragments. The output of thereconstruction is a network of vimentin fragments, each presented by an or-dered chain of pixels and the local fragment orientation.
In a second step, we applied an experimentally determined lengththreshold of 4 μm to extract long “fibers” from the fragment map. All high-confidence pixels in the steerable filter response map minus the pixels be-longing to long fibers were classified as “mesh.”
Cell-Shape Tracking. To track cell shape over time, a binarymask of the cell wasskeletonized using medial axis transformation as implemented in the MATLABfunction bwmorph. Skeleton branches were tracked over time to identify cellprotrusions and their directionality. Instantaneous cell speed and directionalitywere determined as the magnitude and orientation of the displacement of thecenter of the mass of the binary mask between consecutive frames.
ACKNOWLEDGMENTS. We thank Jennifer Waters and staff of the NikonImaging Center at Harvard Medical School for help with light microscopy;Robert Goldman and Saleem Mahammad (Northwestern University MedicalSchool) for help with constructs; and Jessica Tytell (formerly of the G.D.laboratory), Hunter Elliott (Harvard Image and Data Analysis Core), andJagesh Shah (Harvard Medical School) for valuable discussions. This study wassupported by National Institutes of Health PPG Grant P01 GM096971 (to G.D.)and 2R01 GM039565-28 (to T.J.M.). C.J.B. was supported by fellowships fromthe Swiss National Science Foundation and the Novartis Research Foundation.
1. Mendez MG, Kojima S, Goldman RD (2010) Vimentin induces changes in cell shape,motility, and adhesion during the epithelial to mesenchymal transition. FASEB J 24:1838–1851.
2. Huber F, Boire A, López MP, Koenderink GH (2015) Cytoskeletal crosstalk: When threedifferent personalities team up. Curr Opin Cell Biol 32:39–47.
3. Hookway C, et al. (2015) Microtubule-dependent transport and dynamics of vimentinintermediate filaments. Mol Biol Cell 26:1675–1686.
4. Lin CH, Forscher P (1995) Growth cone advance is inversely proportional to retrogradeF-actin flow. Neuron 14:763–771.
5. Vallotton P, Danuser G, Bohnet S, Meister JJ, Verkhovsky AB (2005) Tracking retro-grade flow in keratocytes: News from the front. Mol Biol Cell 16:1223–1231.
6. Theriot JA, Mitchison TJ (1991) Actin microfilament dynamics in locomoting cells.Nature 352:126–131.
7. Etienne-Manneville S (2013) Microtubules in cell migration. Annu Rev Cell Dev Biol 29:471–499.
8. Svitkina TM, Verkhovsky AB, Borisy GG (1996) Plectin sidearms mediate interaction ofintermediate filaments with microtubules and other components of the cytoskeleton.J Cell Biol 135:991–1007.
9. Goldman RD, Khuon S, Chou YH, Opal P, Steinert PM (1996) The function of in-termediate filaments in cell shape and cytoskeletal integrity. J Cell Biol 134:971–983.
10. Helfand BT, et al. (2011) Vimentin organization modulates the formation of la-mellipodia. Mol Biol Cell 22:1274–1289.
11. Jiu Y, et al. (2015) Bidirectional interplay between vimentin intermediate filamentsand contractile actin stress fibers. Cell Reports 11:1511–1518.
12. Gan Z, et al. (2016) Vimentin enhances directed cell migration by stabilizingmicrotubule-mediated cell polarity. Cell Systems 3:252–263.
13. Janmey PA, Euteneuer U, Traub P, Schliwa M (1991) Viscoelastic properties of vimentincompared with other filamentous biopolymer networks. J Cell Biol 113:155–160.
14. Guo M, et al. (2013) The role of vimentin intermediate filaments in cortical and cy-toplasmic mechanics. Biophys J 105:1562–1568.
15. Nöding B, Herrmann H, Köster S (2014) Direct observation of subunit exchange alongmature vimentin intermediate filaments. Biophys J 107:2923–2931.
16. Cermak T, et al. (2011) Efficient design and assembly of custom TALEN and other TALeffector-based constructs for DNA targeting. Nucleic Acids Res 39:e82.
17. Jacob M, Unser M (2004) Design of steerable filters for feature detection using canny-like criteria. IEEE Trans Pattern Anal Mach Intell 26:1007–1019.
18. Canny J (1986) A computational approach to edge detection. IEEE Trans Pattern AnalMach Intell 8:679–698.
19. Gardel ML, Schneider IC, Aratyn-Schaus Y, Waterman CM (2010) Mechanical integration ofactin and adhesion dynamics in cell migration. Annu Rev Cell Dev Biol 26:315–333.
20. Theriot JA, Mitchison TJ, Tilney LG, Portnoy DA (1992) The rate of actin-based motilityof intracellular Listeria monocytogenes equals the rate of actin polymerization.Nature 357:257–260.
21. Lim J, Danuser G (2009) Live cell imaging of F-actin dynamics via fluorescent specklemicroscopy (FSM). J Vis Exp 30:1325.
22. Mendoza MC, Besson S, Danuser G (2012) Quantitative fluorescent speckle micros-copy (QFSM) to measure actin dynamics. Curr Protoc Cytom 62:2.18:2.18.1–2.18.26.
23. Gutierrez E, et al. (2011) High refractive index silicone gels for simultaneous total internalreflection fluorescence and traction force microscopy of adherent cells. PLoS One 6:e23807.
24. Jensen MH, Morris EJ, Goldman RD, Weitz DA (2014) Emergent properties of com-posite semiflexible biopolymer networks. BioArchitecture 4:138–143.
25. Esue O, Carson AA, Tseng Y, Wirtz D (2006) A direct interaction between actin and vi-mentin filaments mediated by the tail domain of vimentin. J Biol Chem 281:30393–30399.
26. Gardel ML, et al. (2008) Traction stress in focal adhesions correlates biphasically withactin retrograde flow speed. J Cell Biol 183:999–1005.
27. Beningo KA, Dembo M, Kaverina I, Small JV, Wang YL (2001) Nascent focal adhesionsare responsible for the generation of strong propulsive forces in migrating fibro-blasts. J Cell Biol 153:881–888.
28. Yoon M, Moir RD, Prahlad V, Goldman RD (1998) Motile properties of vimentin in-termediate filament networks in living cells. J Cell Biol 143:147–157.
29. Christian ML, et al. (2012) Targeting G with TAL effectors: A comparison of activities ofTALENs constructed with NN and NK repeat variable di-residues. PLoS One 7:e45383.
30. Han SJ, Oak Y, Groisman A, Danuser G (2015) Traction microscopy to identify forcemodulation in subresolution adhesions. Nat Methods 12:653–656.
31. Otsu N (1979) A threshold selection method from gray-level histograms. IEEE TransSyst Man Cybern 9:62–66.
32. Rosin PL (2001) Unimodal thresholding. Pattern Recognit 34:2083–2096.
5200 | www.pnas.org/cgi/doi/10.1073/pnas.1614610114 Costigliola et al.
Dow
nloa
ded
by g
uest
on
Apr
il 15
, 202
0